Early error detection is generic, but subsequent adaption to errors is not:
|
|
|
- Matilda Collins
- 5 years ago
- Views:
Transcription
1 Early error detection is generic, but subsequent adaption to errors is not: evidence from ERPs Monica Dhar a and Gilles Pourtois a a Department of Experimental Clinical, and Health Psychology, Ghent University, Belgium Corresponding author: Monica Dhar Department of Experimental Clinical and Health Psychology Ghent University Henri Dunantlaan Gent Belgium Phone: Fax: monica.dhar@ugent.be 1
2 Abstract The aim of the present study was to investigate whether error detection and subsequent regulatory processes could be influenced by pre-familiarisation with task-relevant stimulus features. To this end, nineteen healthy adults performed a speeded Go/NoGo task with compound targets, involving two concurrent stimulus attributes, which were either prefamiliarised or not, while high-density EEG was recorded. During the speeded Go/NoGo task, response errors clearly elicited an error-related negativity (ERN) and error positivity (Pe), but these error-related components were not modulated by familiarisation. By comparison, post-error adaptive processes were found to depend on familiarisation, as distinct topographic ERP effects were evidenced for familiarised vs. non-familiarised stimuli. Moreover, post-error slowing was abolished in the condition comprising familiarised attributes. These results suggest that prefamiliarisation with a stimulus property leaves unaffected error detection mechanisms, while altering subsequent adaptive processes. Whereas error detection mechanisms may be generic, the automatic adaptive processes consecutive to error detection may be malleable, and influenced by pre-familiarisation of stimulus features. Keywords: ERP, ERN, Pe, ACC, post-error slowing 2
3 1. Introduction The concept of cognitive control relates to our ability to monitor actions, such as errors, and to judge how (in)appropriate these are depending on contextual demands or personal goals. Upon detection of such an event, an increase in control enables us to adapt to the changing demands of the environment, and thus to regulate our behaviour. Error processing in particular has often been used to study cognitive control brain mechanisms and two distinct components have been described to be involved in error processing: an evaluative component, comprising the detection of an unexpected outcome and a regulative component involved in exerting top-down attentional control (Botvinick, Braver, Barch, Carter, & Cohen, 2001), whereby on detection of an error, the evaluative system swiftly signals the control system to heighten control as required. Event-related potential (ERP) studies have provided insight into the processes involved in the commission of errors and cognitive control (Falkenstein, Hoormann, Christ, & Hohnsbein, 2000). In the response-locked averaged ERP, a fronto-central negativity emerges at a latency of about ms when an error is committed that has been described as the error-related negativity (ERN; Gehring, Goss, Coles, Meyer, & Donchin, 1993) or error negativity (Ne; Falkenstein, Hohnsbein, Hoormanan, & Blanke, 1991). This component is followed by a more posterior positive deflection with a latency of around ms following errors, the error positivity (Pe). The ERN primarily indexes an early (cognitive) mismatch process between the intended or desired and actual response (Coles, Scheffers, & Holroyd, 2001; Falkenstein et al., 1991; Nieuwenhuis, Ridderinkhof, Blom, Band, & Kok, 2001). Alternative theoretical accounts suggested that the ERN reflects either mechanisms of reinforcement learning implicating dopaminergic midbrain structures (Holroyd & Coles, 2002; Nieuwenhuis, Holroyd, Mol, & Coles, 2004) or conflict monitoring processes (Carter et al., 1998; van Veen, Cohen, Botvinick, 3
4 Stenger, & Carter, 2001; Yeung, Botvinick, & Cohen, 2004). Hence, the ERN might reflect the early detection of a mismatch between motor representations, remaining unavailable to conscious awareness. By contrast, the Pe component may reflect a more elaborate (perhaps conscious) stage of error detection, such as previously suggested (Nieuwenhuis et al., 2001; Ridderinkhof, Ramautar, & Wijnen, 2009). Therefore, these components appear to reflect qualitatively different stages of evaluative error processing. The dorsal Anterior Cingulate Cortex (dacc) has been identified as the main neural generator of the ERN (Debener et al., 2005; Dehaene, Posner, & Tucker, 1994; Miltner et al., 2003; van Veen & Carter, 2002). This structure receives dopamine input from the basal ganglia that have an evaluative function and assist in action selection by allocating attention to behaviourally salient events (Redgrave, Gurney, & Reynolds, 2008). According to the reinforcement learning theory (Holroyd & Coles, 2002), the basal ganglia generate predictions for success or failure of an action. A response error typically leads to a phasic decrease of the mesencephalic dopamine signal that is readily transmitted to the dorsal ACC, which in turn elicits the ERN. Since the ERN may be generated in the absence of error awareness (Nieuwenhuis et al., 2001), early error detection brain mechanisms may not be all that sensitive to the magnitude or severity of response errors. These early action monitoring systems would primarily detect the presence of a mismatch between the actual and desired motor outcome, without coding the amount of perceived discrepancy between the two. Yet, this system may demonstrate some flexibility, as the ERN amplitude has been shown to be sensitive to the speedaccuracy trade-off, with larger amplitudes occurring when accuracy is emphasised over speed (Falkenstein et al., 2000; Gehring et al., 1993). ERN amplitude was also found to be modulated by the saliency of errors, i.e. larger for more salient errors (Hajcak, Moser, Yeung, & Simons, 4
5 2005), and error magnitude, in that it was larger for more severe errors (Bernstein, Scheffers, & Coles, 1995). Behavioural adaptation after errors has typically been observed in the form of a systematic slowing of response latencies for correct trials immediately following an error (Rabbitt, 1966). However, there is some disagreement as to the mechanism causing post-error slowing. Some researchers have taken post-error slowing to reflect enhanced cognitive control, allowing for increased accuracy on the trial immediately following an error as a result of an augmented response threshold (Botvinick et al., 2001; Brewer & Smith, 1984). Jentzsch and Dudschig (2009) postulated that post-error adaptation depends on the amount of time available for behavioural adjustments before the next trial. When the response-stimulus interval (RSI) is short, little time is available for central processing, causing a bottleneck and consequently posterror slowing, whereas when there is enough time to adjust behaviour before the next trial, posterror adaptation is more effective and responses are slow because they are more controlled. Many researchers observed that post-error slowing was unrelated to ERN amplitude (Hajcak, McDonald, & Simons, 2003; Luu, Flaisch, & Tucker, 2000; Nieuwenhuis et al., 2001), although some did find a relationship between ERN and post-error slowing, with larger ERN amplitudes coinciding with longer post-error response times (Debener et al., 2005; Gehring et al., 1993). However, contrary to expectation, greater slowing is not always accompanied by greater posterror accuracy (Hajcak & Simons, 2008; Hajcak, et al., 2003). An alternative account was proposed by Notebaert and colleagues (2009). In a recent study, they manipulated error frequency and studied the effect on post-error slowing. Response slowing was found not only following errors when they were infrequent, but also following correct trials when these were the infrequent stimulus. They concluded that post-error slowing was the result of an orienting 5
6 response to infrequent events (Barcelo, Escera, Corral, & Perianez, 2006; Bernstein, 1969; Sokolov, 1963). Few ERP studies have delved into the neural processing associated with regulatory functions subsequent to errors. However, recently Hikosaka and Isoda (2010) discussed two mechanisms by which individuals are able to rapidly switch their behaviour to adapt to a new context, namely proactive and retroactive behavioural switching. Proactive switching, controlled by the pre-supplementary motor area, refers to the exertion of top-down control to prevent errors. Conversely, retroactive switching is a reactive automatic form of switching that is mediated by the ACC. Building on this framework, Ullsperger and King (2010) suggested that the proactiveretroactive framework would not only apply to switching of behaviour but to all adjustments in cognitive control. However, the electrophysiological correlates of the regulative processes upon commission of an error, such as post-error slowing, have not been studied in great detail. Still little is known about the ERP correlates of behavioural adaptation following errors, in comparison to a wealth of studies that have already shed light on brain systems implicated in early error detection mechanisms (ERN and Pe components). In this study, we were interested in the potential for modulation in evaluative and regulatory error processing systems. To address the question whether error detection and adaptation mechanisms after errors are equally flexible or not, we designed a simple experiment wherein we familiarised participants with a stimulus attribute in order to test how this factor could in turn modulate the size and expression of these error evaluation and regulation mechanisms. Familiarisation has previously been found to modulate top-down or executive control processes, suggesting that these monitoring systems may be differentially engaged or activated depending on the familiarity or relevance of the stimuli. For example, topographical 6
7 modulation of P3 has been found after familiarisation with a working memory task (Segalowitz, Wintink, & Cudmore, 2001). Similarly, cognitive control mechanisms engaged during the monitoring of response errors (and/or subsequent adaptation/slowing down) may also be flexible, and differentially influenced by how familiar or relevant the stimulus is, although this prediction has not been tested yet. To this end, we used a modified version of a speeded Go/NoGo task (Vocat, Pourtois, & Vuilleumier, 2008). In the present study, participants were pre-familiarised with either colour information or face information. In effect, a single stimulus feature was artificially made more important, hence potentially giving this stimulus feature a greater advantage for selection (Jagadeesh, Chelazzi, Mishkin, & Desimone, 2001). This process of familiarisation is automatic and does not require conscious effort (Davidson & Glisky, 2002). Subsequently, the participants performed two Go/NoGo tasks in which compound target stimuli (coloured faces) were presented encompassing both stimulus features (colour and emotional expression) simultaneously. In the colour Go/NoGo participants detected a predefined colour and in the face Go/NoGo they detected a certain facial expression. On a task as simple as a Go/NoGo task, participants generally commit very few response errors (i.e. false alarms on NoGo trials). Therefore, to ensure that errors would occur frequently, time pressure was introduced by means of a response deadline procedure enabling the collection of many response errors within a short period of time, without disrupting basic error monitoring functions and cognitive control (see Vocat et al., 2008). We tested whether the evaluative component of action monitoring, as reflected by the ERN and Pe components, would be modulated by stimulus familiarisation, or whether they would be stable, in line with a generic function of the ERN (Barch et al., 2001; Braver, Barch, Gray, Molfese, & Snyder, 2001; Miltner, Braun, & Coles, 1997). Moreover, we examined 7
8 whether we could detect a modulation of post-error adjustments in behaviour and in neural processing as a function of the familiarisation procedure. 2. Materials and methods 2.1 Participants Nineteen healthy right-handed university students (15 women) with a mean age of 21.9 (SD = 2.08) participated in the experiment, for which they were required to give written informed consent. All reported normal or corrected-to-normal vision and none had a history of brainrelated illness. The study was approved by the local ethics committee. 2.2 Design and stimuli We primarily sought to have compound stimuli, providing information concurrently on two orthogonal (lower-level) categorical dimensions, namely face expression and colour, for which there was no overtraining. This allowed us to use the exact same visual stimuli during our Go/NoGo task, with the visual processing of one of the two attributes/dimensions being reliably familiarised, while the other was not (counterbalanced across participants). Results of pilot testing enabled us to establish that facial expression and colour provided two distinct relevant visual attributes that could easily be merged and combined to create compound stimuli. Moreover, it was also important to have two dimensions being relatively orthogonal to one another, such that enhancing familiarisation with the processing of one attribute did not 8
9 immediately interfere with the visual processing of the other dimension. Only one of these two dimensions (either colour or emotional facial expression, counterbalanced across participants) was made more familiar to participants by the means of dedicated familiarisation blocks. These compound stimuli were then used during a Go/NoGo task. This task involved the inhibition of a prepotent response tendency toward either the pre-familiarised or non-familiarised stimulus dimension, enabling us to compare cognitive control effects for these two conditions. Familiarisation procedure. Preceding each Go/NoGo block of 100 trials, the familiarisation task, consisting of 20 trials, was always presented. The group was divided in two, so that half of the participants were administered four blocks of the colour-choice and the other half four blocks of the face-choice task. Each trial consisted of two stimuli presented in the left and right visual field. The trials were presented, counterbalancing the position of the target (left or right). As mentioned above, colours and facial expressions (by Ekman & Friesen, 1976) were employed as these could be easily contrasted (blue-turquoise and neutral-fearful). The participants were instructed to respond to the target (blue/fearful) stimulus with the left hand when it appeared on the left-hand side and with the right hand when it appeared to the right. After a delay of 500 ms, correct responses were followed by a green patch in the centre of the screen (positive feedback) and incorrect responses by a red patch (negative feedback), presented for 1000 ms. The interval from feedback offset to cue onset was set at 1000 ms. Speeded Go/NoGo task. Two versions of this task, a colour and a face task, were designed. A trial sequence of the colour task consisted of a centrally presented black and white face in an oval frame presented for a duration of 500 ms followed immediately by a blue face on 60% of the trials (Go) and a turquoise face on 40% of the trials (NoGo) for a duration of 1000 ms or until a response was given. Participants were required to respond as quickly and accurately as possible 9
10 to the target stimulus by pressing the spacebar with their right hand. Presentation of fearful and neutral expressions was balanced across conditions. Correspondingly, a trial sequence of the face task began with a centrally presented coloured blue or turquoise oval patch presented for 500 ms, followed by a fearful face on 60% of the trials (Go) and a neutral face on 40% of the trials for 1000 ms or until a response was given. Blue or turquoise faces were presented evenly across conditions. Fearful and neutral expressions from six identities from the Ekman and Friesen series (3 men and 3 women) were used. These stimuli were cropped from the hairline. This experimental procedure ensured that imperative Go stimuli (as well as NoGo stimuli) were actually identical between the two versions of the task. Time pressure was implemented by means of a response deadline. Feedback on task performance was presented on a trial-by-trial basis and consisted of a negative feedback (a small red patch) following correct but slow Go responses, as well as incorrect NoGo responses, whereas a positive feedback (a small green patch) was presented following correct Go responses falling within the time limit and correct inhibitions. For each participant, this limit was initially set to 300 ms on the very first trial and was subsequently adjusted and updated (higher or lower) for each trial using an algorithm that averaged the present reaction time (RT) with the block average RT to determine the threshold for the next trial. This procedure has been utilised extensively in previous studies (Aarts & Pourtois, in press; Koban, Pourtois, Vocat, & Vuilleumier, 2010; Vocat et al., 2008; Pourtois et al., 2010). If the current RT was slower than the current limit, the participant received negative feedback in the form of a red patch. If the RT was faster than the current limit, the participant received positive feedback (green patch). This procedure ensured that many false alarms could be obtained on NoGo trials despite fluctuations in speed on a trial-by-trial basis, because this arbitrary cut-off for correct responses was updated 10
11 and adjusted online after each trial, and it inevitably encouraged participants to be fast. Feedback stimuli appeared with a delay of 500 ms after the response and were presented centrally for a duration of 500 ms followed by a pause of 1000 ms before the next trial started. Four blocks of the Go/NoGo task were administered two colour blocks and two face blocks which were alternated counterbalancing the order of colour and face blocks across participants. Errors on the Go/NoGo tasks were defined as incorrect responses (false alarms). Fast hits were hits (i.e. correct responses on go trials) that fell within the arbitrary (and updated) time limit on a particular trial. In this case participants received positive feedback. Slow hits were also correct responses on go trials, but that were slower than the arbitrary time limit for a particular trial. In this case, participants received negative feedback. Ratings. After each block, participants were asked to rate, on an 11-point Likert scale, the avoidability of errors (1 = highly avoidable to 11 = highly unavoidable), task difficulty (1 = very easy to 11 = very difficult), and how annoying they found each block (1 = not at all annoying to 11 = very annoying). Mean RTs of the familiarisation were submitted to repeated measures analyses of variance (ANOVA) with the within-subjects factor block (with four levels). Mean RTs for hits (fast and slow) and errors on the Go/NoGo task were submitted to repeated measures ANOVAs with the within-subjects factors Familiarisation (with two levels: familiarised vs. nonfamiliarised) and Accuracy (with two levels: correct vs. incorrect). Accuracy on the Go/NoGo task and subjective rating scores of the familiarised and non-familiarised condition were submitted to pairwise comparisons, using Student paired t-tests. 11
12 2.3 ERP recording Continuous EEG was acquired at 512 Hz through a 128-channel Biosemi ActiveTwo system (Biosemi, Amsterdam, Netherlands) referenced to the CMS-DRL ground (which functions as a feedback loop driving the average potential across the montage as close as possible to the amplifier zero). Data were recalculated offline against the average reference. Vertical EOG was recorded from infraorbital and supraorbital electrodes placed in line with the pupil of the left eye. Horizontal EOG was not recorded as all stimuli were centrally presented. Response-locked averages (ERP waveforms) were calculated by following a standard sequence of data transformations applied to the raw EEG data (Picton et al., 2000). First, a -200 to 500 ms window from response onset was segmented. Subsequently, a baseline correction was performed using the pre-response interval of 200 ms and the Gratton and colleagues algorithm (Gratton, Coles, & Donchin, 1983) was used to correct vertical eye movements. For each participant, an amplitude threshold for artefact rejection was chosen that allowed 1/3 of the segments to be rejected and 2/3 to be kept. This method takes into account individual differences in amplitude and ensures that a similar number of trials are used across participants to compute the individual average waveforms, despite these amplitude variations. Epochs with an amplitude above or below this individually determined threshold were considered artefacts and were therefore rejected (M = -76.1/+76.1 mv, SD = 6.83). Bad or excessively noisy channels were interpolated using spherical splines. Individual epochs were averaged, and a 30 Hz low-pass filter was applied. We also computed ERP waveforms using a 0.5 Hz high pass digital filter applied to the raw EEG data, as well as waveforms without applying a 30 Hz low-pass digital filter. In both cases, very similar waveforms (amplitude, latency and morphology of the ERP 12
13 components) were obtained, ruling out the possibility that the filter settings used (i.e. 30 Hz lowpass filter applied to the segmented data) created any substantial distortion or temporal drift in the ERP data. 2.4 Mean amplitudes Following standard practice, the ERN was defined as a conspicuous fronto-central negativity (electrodes FCz and Cz) peaking early (0-60 ms) following response onset. The Pe was first described to be maximal at centro-parietal sites (Falkenstein et al., 1991). Yet, here the Pe was found to have a more central scalp distribution (electrode Cz) and was defined as the positive component ( ms post-response onset) immediately following the ERN. Mean amplitudes of the CRN/ERN and Pe for hits and errors were submitted to repeated measures analyses of variance (ANOVAs) with the within-subjects factors Familiarisation (with two levels: familiarised vs. non-familiarised) and Accuracy (with two levels: correct vs. incorrect). 2.5 Topographic analyses In order to capture more global ERP differences between familiarised and non-familiarised stimuli during the detection of errors and subsequent adaptation, a detailed topographic mapping analysis of the ERP data was next performed (Michel, Seeck, & Lantis, 1999; Michel et al., 2001; Murray, Brunet, & Michel, 2008; Pourtois, 2010; Pourtois, Dan, Grandjean, Sander, Vuilleumier, 2005; Pourtois, Delplanque, Michel, Vuilleumier, 2008; Pourtois, Thut, Peralta, Michel, Vuilleumier, 2005). To precisely characterise topographic modulations over time and 13
14 conditions, we used a spatial cluster analysis. This pattern analysis efficiently summarises ERP data by a limited number of field configurations, previously referred to as functional microstates (Lehmann & Skrandies, 1980; Michel et al., 1999). These methods have been detailed elsewhere (Murray et al., 2008; Pourtois et al., 2008) and we provide only the essentials here. The rationale of the microstate segmentation is to isolate periods of temporal stability (and by extension changes) in the manner that the global electric field is distributed over the scalp surface and over time points, by using a formal statistical approach applied to the whole topography information rather than to values from single electrodes. This stems from the fact that different map topographies reflect different configurations of electric sources in the brain, that is, different neural networks. The spatio-temporal segmentation algorithm is derived from a k-mean spatial cluster analysis (Pascual-Marqui, Michel, & Lehmann, 1995) and identifies the most dominant scalp topographies appearing in the group-averaged ERPs of each condition and over time. The optimal number of topographic maps explaining the whole data set is determined objectively by cross validation. Finally, to statistically assess the validity of the hypothesis that emerges from the clustering algorithm, a back-fitting is performed. The template maps that were identified by the clustering algorithm applied to the group-averaged ERPs are fitted back to the individual participant ERP data to provide estimates of their representation across time and conditions. This back-fitting is performed by calculating the spatial correlation between singleparticipant ERPs and template maps. Here, we performed two distinct topographic pattern analyses. (i) First, we analysed response-locked ERPs and compared error processing for familiarised and non-familiarised stimuli. (ii) Next, we analysed post-error correct hits, and again compared the processing of familiarised and non-familiarised stimuli. For each analysis, the topographic segmentation was 14
15 applied on group-averaged data from -100 ms until 500 ms after response onset. Map duration was entered in repeated-measures analyses of variance (ANOVA) with two within-subject factors, familiarisation (familiarised vs. non-familiarised) and map configuration (i.e. the dominant electric field distributions previously identified by the spatial cluster analysis). These analyses were carried out using CARTOOL software (Version 3.34; developed by D. Brunet, Functional Brain Mapping Laboratory, Geneva, Switzerland). 2.6 Source localisation Finally, to estimate the likely neural sources underlying the electrical field configurations identified by the previous analyses, we used a specific distributed linear inverse solution, namely standardised low-resolution brain electromagnetic tomography (sloreta, Pascual-Marqui, 2002). sloreta is based on the neurophysiological assumption of coherent coactivation of neighbouring cortical areas (known to have highly synchronised activity, see Silva, Amitai, & Connors, 1991) and, accordingly, it computes the smoothest of all possible activity distributions (i.e. no a priori assumption is made on the number and locations of the sources). Mathematical validation of this distributed source localisation technique has been recently demonstrated (Sekihara, Sahani, & Nagarajan, 2005). sloreta solutions are computed within a three-shell spherical head model co-registered to the MNI152 template (Mazziotta et al., 2001). The source locations were therefore given as (x, y, z) coordinates (x from left to right; y from posterior to anterior; z from inferior to superior). sloreta estimates the 3-dimensional intracerebral current density distribution in 6239 voxels (5 mm resolution). 15
16 3. Results 3.1 Behavioural results and subjective ratings For calculation of mean RTs, trials for which RTs fell below or above 3 SDs from each participant s mean were considered invalid and were thus removed. These trials were also rejected from the ERP analyses. Accuracy on the familiarisation was high with participants reaching a mean accuracy of 95.2% (SD = 0.05). Mean RTs from block 1 to block 4 were: 719 ms (SD = 336), 521 ms (SD = 182), 481 ms (SD = 153), and 475 ms (SD = 163). For RT, a main effect was found for block (F(3,16) = 25.53, p <.001), consistent with an improved performance as a function of time. This result suggests that the familiarisation procedure was efficient to induce learning, and presumably to strengthen the mental representation of the pre-selected stimulus feature. Analysis of the subjective ratings indicated that familiarisation had no effect on how annoying the task was perceived to be (t(18) = -0.9, p = ns). Mean ratings were 8.74 (SD = 1.96) in the familiarised condition and 9.13 (SD = 1.76) in the non-familiarised condition. However, errors were judged to be reliably less avoidable in the non-familiarised condition (M = 9.21, SD = 1.56) than in the familiarised condition (M = 8.32, SD = 1.59; t(18) = -2.36, p =.03), confirming a differential influence of familiarisation on the processing of the pre-familiarised vs. non-familiarised stimuli during the Go/NoGo task. In addition, there was a marginally significant effect of familiarisation on ratings of task difficulty (t(18) = 2.06, p =.054), with the prefamiliarised condition (M = 4.97, SD = 2.8) being judged as slightly more difficult compared to the non-familiarised condition (M = 4.29, SD = 2.77). 16
17 Mean accuracy and mean hit and error RTs for the familiarised and non-familiarised condition during the Go/NoGo task are presented in Table 1. No difference between conditions was found for mean accuracy (t(18) = 0.13, p = ns) with the mean number of false alarms being 26 in both the familiarised and the non-familiarised condition, and number of misses being marginal (familiarised: 1.68 misses and non-familiarised: 3.16 misses). For RT, a significant interaction was found between familiarisation and accuracy (F(1,18) = 6.54, p =.02), indicating that error RT was shorter for pre-familiarised stimuli compared to unfamiliar stimuli (t(18) = , p =.003), whereas there was no significant RT difference for hits in the two conditions (t(18) = -1.0, p = ns). These results suggest that participants committed more impulsive response errors with the pre-familiarised, relative to the non-familiarised stimulus attribute. This effect underscores the differential impact of the pre-familiarised vs. non-familiarised stimulus feature during the Go/NoGo task. We also found that the magnitude of the post-error slowing effect, defined as the difference between RT on correct after correct trials (RT cc ) and RT on correct after error trials (RT ec ), did not differ between the two conditions, since there was no interaction between familiarisation and accuracy (F(1,18) = 2.2, p = ns). However, a possible confound may have arisen in this analysis, due to the mixed feedback on correct trials prior to hits, as in the RT cc condition the feedback could be either positive (for fast hits) or negative (for slow hits). So, to obtain a better comparison with RT ec, for the RT cc condition fast hits were omitted from the analysis in order to control for feedback after hits and therefore hits could be preceded by only negative feedback, as was also the case for post-error hits. This analysis yielded a significant interaction between familiarisation and accuracy (F(1,18) = 6.59, p =.019), the post-error slowing effect being significant for the non-familiarised condition (t(18) = -3.68, p =.002), but 17
18 not for the familiarised condition (t(18) = -0.30, p = ns). Mean familiarised RT cc was 325 ms (SD = 45) and RT ec was 322 ms (SD = 68). Mean non-familiarised RT cc was 311 ms (SD = 38) and RT ec was 336 ms (SD = 44). In other words, we found that the post-error slowing adaptation effect was reliably blunted in the condition with pre-familiarised stimulus features, further confirming a differential impact of the familiarisation procedure on the regulative component of action monitoring. Fig. 1 depicts the post-error slowing effect separately for the familiarised and the non-familiarised condition as computed in this latter analysis. [Insert Figure 1 and Table 1] 3.2 ERP results Errors elicited conspicuous ERN and Pe components, relative to correct hits. However, the familiarisation did not change the amplitude of these error-related ERP components, as verified by standard peak analyses. Fig. 2 shows the response-locked ERPs separately for the familiarised and non-familiarised condition at electrodes FCz (2a and b) and Cz (2c and d). Mean ERN amplitude was larger for errors, relative to correct hits, for electrode Cz (F(1,18) = 10.7, p =.005), but not for FCz (F(1,18) = 2.96, p = ns). This effect of accuracy was not modulated by familiarisation, either for electrode FCz (F(1,18) = 0.02, p = ns) or Cz (F(1,18) = 0.19, p = ns). These results suggest that the early detection of response errors (ERN) was immune to the relative familiarity of these response errors. Similar to the ERN, the amplitude of the Pe component was not influenced by the familiarisation procedure (electrode Cz: F(1,18) = 1.69, p = ns), although, as expected, this positive component had a much larger amplitude for errors compared to hits (electrode Cz: F(1,18) = 23.37, p <.001). 18
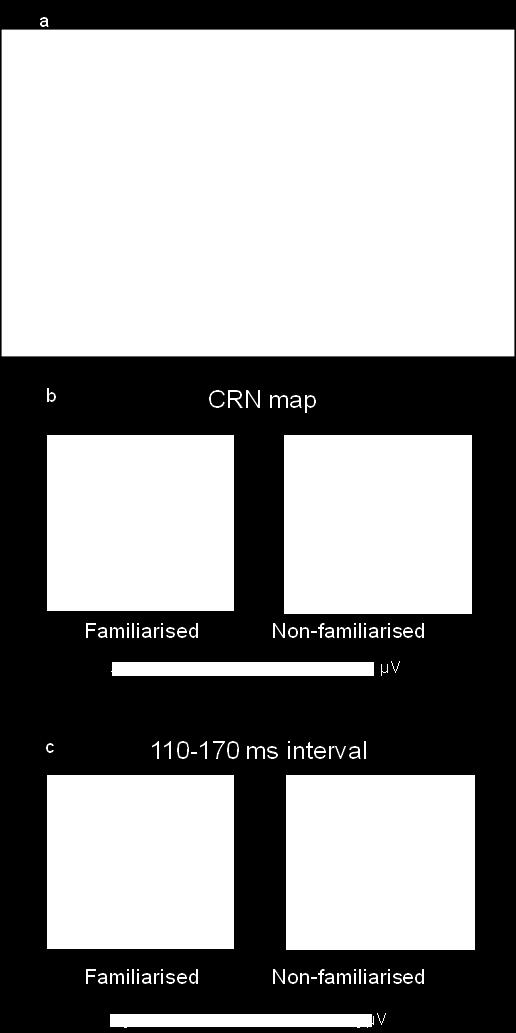
19 Given the blunted post-error slowing effect seen with the pre-familiarised stimulus attribute, an additional analysis was performed to investigate whether there was any significant effect of familiarisation on the correct-related negativity (CRN) (Coles et al., 2001; Vidal, Hasbroucq, Grapperon, & Bonnet, 2000) recorded for correct hits immediately following errors. Fig. 3a depicts the response-locked grand average ERPs for post-error hits and post-slow hits at electrode FCz. Similarly to the analysis of the post-error RT data (see here above), post-error hits were therefore compared to post-slow hits (separately in the familiarised and non-familiarised condition), ensuring that the immediately preceding feedback was balanced in these two conditions. Included are the topographic maps of the CRN in the 0-60 ms interval (Fig. 3b), showing that the CRN in the familiarised and non-familiarised condition shared a similar topography. We found that the amplitude of the CRN for hits following errors was smaller for the familiarised attribute, compared to the non-familiarised attribute, although this effect did not reach statistical significance (FCz; t(18) = 1.9, p =.075; Cz: t(18) = 1.2, p = ns). Fig. 3a also suggests some difference between the two post-error hits conditions, but at a later time interval following the CRN time-course (i.e. an additional negativity following the CRN in the interval from approximately 110 to 170 ms, demarcated in Fig. 3a by vertical bars, was seen only for the familiarised attribute). However, for these two pre-selected electrode positions, the difference between the conditions was not significant in this interval on correct trials after errors (FCz: t(18) = -1.03, p = ns; Cz: t(18) = -1.21, p = ns). Yet, the topographical maps (i.e. the spatial configuration of the electric field, as computed when taking all 128 channels into account) for the familiarised and non-familiarised condition corresponding to this interval turned out to be different, as displayed in Fig. 3c. A frontal negativity, somewhat lateralised to the right, and a central positivity could be distinguished in the map for the familiarised condition. For the non- 19
20 familiarised condition, again a central positivity could be observed, yet the frontal negativity was somewhat left-lateralised. To verify the presence of a significant change in the distribution of the electric field across conditions, we therefore used a topographic mapping technique to better characterise any reliable electrophysiological differences between these two conditions for ERPs time-locked to the onset of the response, and corresponding here to the immediate post-error processing. The second goal of this alternative data analysis was to verify whether the topography of early-error related ERP components (ERN and Pe, as measured at a few electrode positions) were actually not altered as a function of the familiarisation procedure, a question that could not be fully resolved using only the classical peak analyses reported here above. Using these alternative topographic analyses, we aimed to test whether familiarisation, rather than merely changing local amplitude at a few electrode positions, might also disrupt the electric field configuration of ERPs generated during the processing of errors, as well as the adaptation/corrective processes following their occurrences (post-error slowing). [Insert Figures 2 and 3] 3.3 Topographic results We first performed a topographic analysis on the response-locked ERPs corresponding to errors and hits separately for the familiarised and non-familiarised condition, using a broad temporal window encompassing both the ERN and Pe component (from -100 until 500 ms after response onset). A solution with 10 maps was found to explain 98.7% of the variance. However, this analysis did not reveal any reliable topographic differences between familiarised and non- 20
21 familiarised attributes (ERN: t(18) = -1.43, p = ns; CRN: t(18) = 0.69, p = ns; Pe: t(18) = 0.98, p = ns). Thus, the ERN, CRN, and Pe components all had the same scalp distribution for these two conditions, further corroborating the results of the mean amplitude analyses described above. We then performed a similar topographic analysis, but on the response-locked ERPs corresponding to hits immediately following errors. This analysis again encompassed the prominent ERP deflections found following response onset (from -100 until 500 ms postresponse onset). A solution with 10 maps was found to explain 97.3% of the variance. Interestingly, we found that familiarisation actually affected the sequence of dominant scalp maps. During the ms time interval following response onset, the topographic analysis disclosed a significant change of the electric field configuration as a function of familiarisation. The dominant map during this interval in the familiarised condition showed a qualitatively different distribution of the electric field compared to the dominant map found during the same time interval in the non-familiarised condition (Fig. 3c). These two maps were then fitted back to the individual data. The result of this analysis is shown in Fig. 4, depicting the duration of these two maps in the familiarised and non-familiarised condition. A significant map x familiarisation interaction was found (F(1,18) = 6.35, p =.021), indicating that in each condition a different topographic map was predominant. This was confirmed by directly testing the difference between the conditions for the duration of the dominant map. The dominant map in the familiarised condition had a significantly longer duration in this condition (t(18) = 2.52, p =.021), whereas the dominant map in the non-familiarised condition was significantly longer lasting in the non-familiarised condition (t(18) = -2.52, p =.021). These results suggest that familiarising participants with a stimulus attribute had a reliable influence on the expression of regulatory brain processes at stake early on following response onset. Because this topographic 21
22 change necessarily denotes a change in the underlying configuration of intracranial generators, we next used sloreta to corroborate this assumption and gain insight into the putative neural generators of these two different topographic maps. [Insert Figures 4] 3.4 Source localisation results We used sloreta (Fig. 5) to shed light on the configuration of intracranial generators underlying the dominant topographies revealed by the previous analysis. For the distinctive scalp map identified for the familiarised attribute (post-error hits), sloreta disclosed a solution comprising Brodmann area 6 of the medial frontal lobe (MFL; maximum: X = -5, Y = -20, Z = 70), as depicted in Fig. 5a. This area corresponds to the supplementary motor area (SMA), which is known to be responsible for initiating motor responses, including from memory (Tanji, 1994). Noteworthy is the fact that this medial frontal cortex activation also included more ventral sites, including the dorsal ACC Brodmann area 24 (X = 5, Y = -20, Z = 45). For the scalp map identified for unfamiliar stimuli, and being reliably different from pre-familiarised stimuli, sloreta revealed main sources within the medial frontal cortex (Fig. 5b), also corresponding to Brodmann area 24 within the ACC (maximum: X = 3, Y = -24, Z = 39), an area associated with behavioural adjustments (Kerns et al., 2004). These results suggest a shift in the network that was recruited during the period after an error was committed, for pre-familiarised stimuli. Whereas the dorsal ACC seems to be shared in both conditions, the involvement of more dorsal regions, including the SMA, is specific to the condition with familiarised attributes. Next, a 22
23 paired t-test was performed in sloreta comparing the source reconstructions of the familiarised to the non-familiarised condition in the interval from 110 to 170 ms post-response onset (post-error conditions). Based on the descriptive sloreta results presented here above, we expected that more dorsal frontal regions (including the SMA) would be activated during the monitoring of correct post-error actions performed with familiarised stimuli, whereas more ventral medial frontal regions (including the dorsal ACC) would be shared across the two conditions. The results of the t-test are presented in Fig. 5c. The scale indicates the t-value ranging from to In the SMA/Brodmann area 6 (maximum effect found at X = 10, Y = -30, Z = 55) there was a significantly greater activation in the familiarised condition compared to the non-familiarised condition (t(18) = 2.3, p <.05). The reversed contrast did not yield any significant suprathreshold node, indicating that although both conditions shared activation of the dorsal ACC, the SMA activation was specific to the familiarised condition. [Insert Figure 5] 4. Discussion In the current study we trained participants with a single stimulus feature, and evaluated potential effects on performance and cognitive control during an unrelated Go/NoGo task. More specifically, we aimed to test whether the evaluative or regulative component of cognitive control may react in a flexible way, according to slight changes in stimulus representation, artificially induced here using a dedicated familiarisation procedure. The main results of this study show that the regulative, but not the evaluative component of cognitive control was 23
24 affected by familiarisation. The latter process may be somewhat immune to effects of the familiarisation, whereas the former seems more malleable by influences of cognitive or motivational factors, further emphasising the distinct contributions of these two processes during action monitoring and cognitive control (Botvinick et al., 2001). The familiarisation procedure was quite successful to induce a substantial learning, reflected by faster RTs during the course of the experiment. Strikingly, we found that this procedure actually had a differential impact on the processing of familiarised vs. nonfamiliarised stimulus attributes during the main (and completely unrelated) Go/NoGo task. The classical post-error slowing effect was abolished for familiarised, relative to non-familiarised attributes. It is possible that learning a stimulus feature during the familiarisation task caused interference when performing the Go/NoGo task when the learned feature was irrelevant to the task (Nelson, Reuter-Lorenz, Sylvester, Jonides, & Smith, 2003). Although error rates were equal in the two familiarisation conditions, participants rated errors to non-familiarised stimuli to be less avoidable than errors to familiarised stimuli, indicating a differential effect of the familiarisation procedure on perceived performance. Moreover, participants made more impulsive response errors during the Go/NoGo task with stimuli that had been pre-familiarised, as evidenced by shorter error RTs in this condition. This effect might indicate a decreased inhibition or impulse control over the experimentally familiarised stimulus feature (Greene, 1999). Yet, a similar number of errors were committed for familiarised and non-familiarised attributes, therefore allowing for a neat comparison of ERP measures in the two conditions of the Go/NoGo task, without extra noise being introduced by uneven trial numbers. Altogether, these behavioural results demonstrated an effect of the familiarisation on performance during the Go/NoGo task. 24
25 Importantly, despite differential effects of familiarisation on the regulative component of cognitive control and subjective ratings, there was no modulation of error detection brain mechanisms, as neither the ERN nor the Pe was modified. More specifically, neither the amplitude nor the topography of these two error-related ERP components was influenced by prefamiliarising participants with a specific stimulus feature. Imaging studies have repeatedly shown a pattern of activation in medial prefrontal cortices during error commission that does not appear to vary depending on the task that is performed (e.g. Carter et al., 1998; Dehaene et al., 1994; Menon, Adleman, White, Glover, & Reiss, 2001; Ullsperger & von Cramon, 2001), supporting the idea that the ERN may reflect a singular error detection mechanism (Hester, Fassbender, & Garavan, 2004; Miltner et al., 1997). The results of the present study are in line with the assumption that early error detection brain mechanisms are somehow generic. Note that a relatively large CRN was observed in response to hits in this study. This amplification of the CRN is likely to be due to the nature of the speeded task used here, more specifically to the prompt response deadline (Gehring et al., 1993; Luu et al., 2000; Pailing & Segalowitz, 2004; Vidal, Burle, Bonnet, Grapperon, & Hasbroucq, 2003). Although the ERN and Pe were not influenced by familiarisation, a selective impact was found on early adaptation processes consecutive to the commission of errors, as observed for the post-error slowing and ERP activity to post-error hits in the interval shortly after the CRN. In the non-familiarised condition the typical post-error slowing was still present, whereas in the prefamiliarised condition the normal adaptation after an error was abolished. It seems that familiarisation increased early adaptation to errors only for the pre-familiarised condition, as participants were able to adjust their behaviour rapidly. Also, the amplitude of the CRN on correct trials following errors was marginally smaller in this condition. Consistent with these 25
26 findings, the source localisation results revealed distinct sources of activity for early adaptation effects on correct trials after errors. In the familiarised condition, post-error hits were mainly processed in the SMA, a pattern of activation that was associated with decreased adaptation in the familiarised condition, i.e. a suppression of post-error slowing. In the non-familiarised condition, the post-error activity was greatest in the ACC and this activation was accompanied by typical post-error slowing. The dissociation in the locus of maximal activity may have a functional significance. Behavioural adjustments after errors have been described to take two different forms, so-called retroactive adaptation governed by the ACC and proactive adaptation handled by the pre-sma (Hikosaka & Isoda, 2010; Ullsperger & King, 2010). Proactive adaptation refers to the active enhancement of cognitive control processes and implies an attentional component that reduces interference from irrelevant contextual information. Retroactive adaptation occurs as the result of a negative outcome and is considered to be a general reactive form of adjustment. These two functionally distinct modes of control may act to produce a balance between effort and efficiency (Ullsperger & King, 2010). Taking into account the diminished post-error slowing in addition to impulsive errors and SMA activation for posterror hits for the pre-familiarised stimuli, the results may be suggestive of a more automatic style of adaptation, possibly facilitated by the familiarity of the relevant stimulus attribute in this condition. For the non-familiarised stimuli, greater post-error slowing along with slower error RTs and ACC activity (although not significantly stronger than in the familiarised condition) was observed, possibly indicative of a more controlled style of error adaptation. This conclusion would be in line with the findings of Hester et al. (2004), who observed that increased pre-sma activation on a Go/NoGo task was associated with fast responding, while increased ACC activation was associated with slow responding, which was more attentive and controlled. 26
27 Alternatively, because the dorsal ACC is usually considered to be the main generator of the ERN as well as the CRN component (see Roger, Bénar, Vidal, Hasbroucq, & Burle, 2010), while the SMA may be related to proactive behavioural adjustments (Hikosaka & Isoda, 2010; Ullsperger & King, 2010), our new ERP results ( ms post-response onset) suggest that a prolonged CRN/action monitoring effect may be at stake in the non-familiarised condition. By contrast, in the familiarised condition, there is likely a rapid transition following response onset from dorsal ACC (CRN) to more dorsal frontal regions (including the SMA), consistent with the involvement of a differential proactive behavioural control mechanism when monitoring the adequacy of actions performed with pre-familiarised stimulus attributes. Our results raise the question how it is possible that transient familiarisation had an effect on adaptive processing but not error detection itself, if we assume that one is a necessary precursor of the other (Botvinick et al., 2001). One possible explanation would be that the two functions operate independently of each other. In line with this reasoning some investigators have found that the ERN and measures of post-error adaptation are not directly related (Castellar, Kühn, Fias, & Notebaert, 2010; Dudschig & Jentzsch, 2009; Hajcak et al., 2003; Nieuwenhuis et al., 2001), while others found evidence to the contrary (Gehring et el., 1993). Further research will have to be conducted to shed more light on this matter in order to establish under which circumstances adaptive and regulative components of cognitive control are separate processes. In conclusion, error detection was found to be unaffected by familiarisation, consistent with the notion that this mechanism is generic, whereas post-error adaptation was altered, with dissociable regions being recruited depending on the familiarisation condition, suggesting that there is some malleability in adaptive processing brought about by top-down processes. 27
28 Acknowledgements This work is supported by grants from the European Research Council (Starting Grant #200758) and Ghent University (BOF Grant #05Z01708). 28
29 References Aarts, K. & Pourtois, G. (in press). Anxiety does not only increase, but also alters early error monitoring functions. Cognitive Affective & Behavioral Neuroscience. Barcelo, F., Escera, C., Corral, M. J., & Perianez, J. A. (2006). Task switching and novelty processing activate a common neural network for cognitive control. Journal of Cognitive Neuroscience, 18(10), Barch, D. M., Braver, T. S., Akbudak, E., Conturo, T., Ollinger, J., & Snyder, A. (2001). Anterior cingulate cortex and response conflict: Effects of response modality and processing domain. Cerebral Cortex, 11(9), Bernstein, A. S. (1969). To what does the orienting response respond. Psychophysiology, 6(3), Bernstein, P. S., Scheffers, M. K., & Coles, M. G. H. (1995). Where did I go wrong - A psychological analysis of error-detection. Journal of Experimental Psychology-Human Perception and Performance, 21(6), Botvinick, M. M., Braver, T. S., Barch, D. M., Carter, C. S., & Cohen, J. D. (2001). Conflict monitoring and cognitive control. Psychological Review, 108(3), Braver, T. S., Barch, D. M., Gray, J. R., Molfese, D. L., & Snyder, A. (2001). Anterior cingulate cortex and response conflict: Effects of frequency, inhibition and errors. Cerebral Cortex, 11(9),
30 Brewer, N. & Smith, G. A. (1984). How normal and retarded individuals monitor and regulate speed and accuracy of responding in serial choice tasks. Journal of Experimental Psychology-General, 113(1), Carter, C. S., Braver, T. S., Barch, D. M., Botvinick, M. M., Noll, D., & Cohen, J. D. (1998). Anterior cingulate cortex, error detection, and the online monitoring of performance. Science, 280(5364), Castellar, E. N., Kühn, S., Fias, W., & Notebaert, W. (2010). Outcome expectancy and not accuracy determines post-error slowing: ERP support Cognitive Affective and Behavioral Neurosciences, 10, Coles, M. G. H., Scheffers, M. K., & Holroyd, C. B. (2001). Why is there an ERN/Ne on correct trials? Response representations, stimulus-related components, and the theory of errorprocessing. Biological Psychology, 56(3), Davidson, P. S. R. & Glisky, E. L. (2002). Neuropsychological correlates of recollection and familiarity in normal aging. Cognitive Affective & Behavioral Neuroscience, 2(2), Debener, S., Ullsperger, M., Siegel, M., Fiehler, K., von Cramon, D. Y., & Engel, A. K. (2005). Trial-by-trial coupling of concurrent electroencephalogram and functional magnetic resonance imaging identifies the dynamics of performance monitoring. Journal of Neuroscience, 25(50), Dehaene, S., Posner, M. I., & Tucker, D. M. (1994). Localization of a neural system for errordetection and compensation. Psychological Science, 5(5),
31 Dudschig, C. & Jentzsch, I. (2009). Speeding before and slowing after errors: Is it all just strategy? Brain Research, 1296, Ekman, P. & Friesen, V. (1976). Pictures of facial affect. Consulting Psychologists Publishing, Palo Alto, California. Falkenstein, M., Hohnsbein, J., Hoormann, J., & Blanke, L. (1991). Effects of crossmodal divided attention on late ERP components. 2. Error processing in choice reaction tasks. Electroencephalography and Clinical Neurophysiology, 78(6), Falkenstein, M., Hoormann, J., Christ, S., & Hohnsbein, J. (2000). ERP components on reaction errors and their functional significance: a tutorial. Biological Psychology, 51(2-3), Gehring, W. J., Goss, B., Coles, M. G. H., Meyer, D. E., & Donchin, E. (1993). A neural system for error-detection and compensation. Psychological Science, 4(6), Gratton, G., Coles, M. G. H., & Donchin, E. (1983). A new method for off-line removal of ocular artifact. Electroencephalography and Clinical Neurophysiology, 55(4), Greene, R. L. (1999). The role of familiarity in recognition. Psychonomic Bulletin & Review, 6(2), Hajcak, G., McDonald, N., & Simons, R. F. (2003). To err is autonomic: Error-related brain potentials, ANS activity, and post-error compensatory behavior. Psychophysiology, 40(6), Hajcak, G., Moser, J. S., Yeung, N., & Simons, R. F. (2005). On the ERN and the significance of 31 errors. Psychophysiology, 42(2),
32 Hajcak, G. & Simons, R. F. (2008). Oops!.. I did it again: An ERP and behavioral study of double-errors. Brain and Cognition, 68(1), Hester, R., Fassbender, C., & Garavan, H. (2004). Individual differences in error processing: A review and reanalysis of three event-related fmri studies using the GO/NOGO task. Cerebral Cortex, 14(9), Hikosaka, O. & Isoda, M. (2010). Switching from automatic to controlled behavior: cortico-basal ganglia mechanisms. Trends in Cognitive Sciences, 14(4), Holroyd, C. B., & Coles, M. G. H. (2002). The neural basis. of human error processing: Reinforcement learning, dopamine, and the error-related negativity. Psychological Review, 109(4), Jagadeesh, B., Chelazzi, L., Mishkin, M., & Desimone, R. (2001). Learning increases stimulus salience in anterior inferior temporal cortex of the macaque. Journal of Neurophysiology, 86(1), Jentzsch, I. & Dudschig, C. (2009). Why do we slow down after an error? Mechanisms underlying the effects of posterror slowing. Quarterly Journal of Experimental Psychology, 62(2), Kerns, J. G., Cohen, J. D., MacDonald, A. W., Cho, R. Y., Stenger, V. A., & Carter, C. S. (2004). Anterior Cingulate conflict monitoring and adjustments in control. Science, 303(5660),
33 Koban, L., Pourtois, G., Vocat, R., & Vuilleumier, P. (2010). When your errors make me lose or win: Event-related potentials to observed errors of cooperators and competitors. Social Neuroscience, 5(4), Lehmann, D. & Skrandies, W. (1980). Reference-free identification of components of checkerboard-evoked multichannel potential fields. Electroencephalography and Clinical Neurophysiology, 48(6), Luu, P., Flaisch, T., & Tucker, D. M. (2000). Medial frontal cortex in action monitoring. Journal of Neuroscience, 20, Mazziotta, J., Toga, A., Evans, A., Fox, P., Lancaster, J., Zilles, K., et al. (2001). A probabilistic atlas and reference system for the human brain: International Consortium for Brain Mapping (ICBM). Philosophical Transactions of the Royal Society B-Biological Sciences, 356(1412), Menon, V., Adleman, N. E., White, C. D., Glover, G. H., & Reiss, A. L. (2001). Error-related brain activation during a Go/NoGo response inhibition task. Human Brain Mapping, 12(3), Michel, C. M., Seeck, M., & Landis, T. (1999). Spatiotemporal dynamics of human cognition. News in Physiological Sciences, 14, Michel, C. M., Thut, G., Morand, S., Khateb, A., Pegna, A. J., de Peralta, R. G., et al. (2001). Electric source imaging of human brain functions. Brain Research Reviews, 36(2-3),
34 Miltner, W. H. R., Braun, C. H., & Coles, M. G. H. (1997). Event-related brain potentials following incorrect feedback in a time-estimation task: Evidence for a "generic" neural system for error detection. Journal of Cognitive Neuroscience, 9(6), Miltner, W. H. R., Lemke, U., Weiss, T., Holroyd, C., Scheffers, M. K., & Coles, M. G. H. (2003). Implementation of error-processing in the human anterior cingulate cortex: a source analysis of the magnetic equivalent of the error-related negativity. Biological Psychology, 64(1-2), Murray, M. M., Brunet, D., & Michel, C. M. (2008). Topographic ERP analyses: A step-by-step tutorial review. Brain Topography, 20(4), Nelson, J. K., Reuter-Lorenz, P. A., Sylvester, C. Y. C., Jonides, J., & Smith, E. E. (2003). Dissociable neural mechanisms underlying response-based and familiarity-based conflict in working memory. Proceedings of the National Academy of Sciences of the United States of America, 100(19), Nieuwenhuis, S., Holroyd, C. B., Mol, N., & Coles, M. G. H. (2004). Reinforcement-related brain potentials from medial frontal cortex: origins and functional significance. Neuroscience and Biobehavioral Reviews, 28(4), Nieuwenhuis, S., Ridderinkhof, K. R., Blom, J., Band, G. P. H., & Kok, A. (2001). Error-related brain potentials are differentially related to awareness of response errors: Evidence from an antisaccade task. Psychophysiology, 38(5), Notebaert, W., Houtman, F., Van Opstal, F., Gevers, W., Fias, W., & Verguts, T. (2009). Posterror slowing: An orienting account. Cognition, 111(2),
35 Pailing, P. E. & Segalowitz, S. J. (2004). The effects of uncertainty in error monitoring on associated ERPs. Brain and Cognition, 56(2), Pascual-Marqui, R. D. (2002). Standardized low-resolution brain electromagnetic tomography (sloreta): Technical details. Methods and Findings in Experimental and Clinical Pharmacology, 24, Pascual-Marqui, R. D., Michel, C. M., & Lehmann, D. (1995). Segmentation of brain electricalactivity into microstates - model estimation and validation. IEEE Transactions on Biomedical Engineering, 42(7), Picton, T. W., Bentin, S., Berg, P., Donchin, E., Hillyard, S. A., Johnson, R., et al. (2000). Guidelines for using human event-related potentials to study cognition: Recording standards and publication criteria. Psychophysiology, 37(2), Pourtois, G. (2010). Early Error Detection Predicted by Reduced Pre-response Control Process: An ERP Topographic Mapping Study. Brain Topography ( DOI /s Pourtois, G., Dan, E. S., Grandjean, D., Sander, D., & Vuilleumier, P. (2005). Enhanced extrastriate visual response to bandpass spatial frequency filtered fearful faces: Time course and topographic evoked-potentials mapping. Human Brain Mapping, 26(1), Pourtois, G., Delplanque, S., Michel, C., & Vuilleumier, P. (2008). Beyond conventional eventrelated brain potential (ERP): Exploring the time-course of visual emotion processing using topographic and principal component analyses. Brain Topography, 20(4),
36 Pourtois, G., Thut, G., de Peralta, R. G., Michel, C., & Vuilleumier, P. (2005). Two electrophysiological stages of spatial orienting towards fearful faces: Early temporoparietal activation preceding gain control in extrastriate visual cortex. Neuroimage, 26(1), Pourtois, G., Vocat, R., N'diaye, K., Spinelli, L., Seeck, M., & Vuilleumier, P. (2010). Errors recruit both cognitive and emotional monitoring systems: Simultaneous intracranial recordings in the dorsal anterior cingulate gyrus and amygdala combined with fmri. Neuropsychologia, 48, Rabbit, P. M. A. (1966). Errors and error correction in choice reaction tasks. Journal of Abnormal Psychology, 71, Redgrave, P., Gurney, K., & Reynolds, J. (2008). What is reinforced by phasic dopamine signals? Brain Research Reviews, 58(2), Ridderinkhof, K. R., Ramautar, J. R., & Wijnen, J. G. (2009). To P-E or not to P-E: A P3-like ERP component reflecting the processing of response errors. Psychophysiology, 46(3), Roger, C., Bénar, C., Vidal, F., Hasbroucq, T. and Burle, B. (2010). Rostral Cingulate Zone and correct response monitoring: ICA and source localization evidences for the unicity of correct- and error negativities. Neuroimage, 51, Segalowitz, S. J., Wintink, A. J., & Cudmore, L. J. (2001). P3 topographical change with task familiarization and task complexity. Cognitive Brain Research, 12,
37 Sekihara, K., Sahani, M., & Nagarajan, S. S. (2005). Localization bias and spatial resolution of adaptive and non-adaptive spatial filters for MEG source reconstruction. Neuroimage, 25(4), Silva, L. R., Amitai, Y., & Connors, B. W. (1991). Intrinsic oscillations of neocortex generated by layer-5 pyramidal neurons. Science, 251(4992), Sokolov, E. N. (1963). Higher nervous functions - orienting reflex. Annual Review of Physiology, 25, Tanji, J. (1994). The supplementary motor area in the cerebral cortex. Neuroscience Research, 19(3), Ullsperger, M. & King, J. A. (2010). Proactive and reactive recruitment of cognitive control: Comment on Hikosaka and Isoda. Trends in Cognitive Sciences, 14(5), Ullsperger, M. & von Cramon, D. Y. (2001). Subprocesses of performance monitoring: A dissociation of error processing and response competition revealed by event-related fmri and ERPs. Neuroimage, 14(6), van Veen, V. & Carter, C. S. (2002). The timing of action-monitoring processes in the anterior cingulate cortex. Journal of Cognitive Neuroscience, 14(4), van Veen, V., Cohen, J. D., Botvinick, M. M., Stenger, V. A., & Carter, C. S. (2001). Anterior cingulate cortex, conflict monitoring, and levels of processing. Neuroimage, 14(6), Vidal, F., Burle, B., Bonnet, M., Grapperon, J., & Hasbroucq, T. (2003). Error negativity on 37 correct trials: a reexamination of available data. Biological Psychology, 64(3),
38 Vidal, F., Hasbroucq, T., Grapperon, J., & Bonnet, M. (2000). Is the 'error negativity' specific to errors? Biological Psychology, 51(2-3), Vocat, R., Pourtois, G., & Vuilleumier, P. (2008). Unavoidable errors: A spatio-temporal analysis of time-course and neural sources of evoked potentials associated with error processing in a speeded task. Neuropsychologia, 46(10), Yeung, N., Botvinick, M. M., & Cohen, J. D. (2004). The neural basis of error detection: Conflict monitoring and the error-related negativity. Psychological Review, 111(4),
39 Captions Fig. 1: Post-error slowing (RT) effect for the familiarised and non-familiarised condition. Error bars represent the S.E.M. Fig. 2: Response-locked grand average ERPs at FCz (a and b) and Cz (c and d) for the familiarised and non-familiarised hits vs. errors. Topographic maps corresponding to the ERN/CRN time interval (0-60 ms) and Pe time interval ( ms) are presented. Fig. 3: Response-locked grand average ERPs for post-error hits and post-slow hits (post-fast hits were not included in this analysis) in the familiarised and non-familiarised condition at FCz (a), topographic maps of the CRN in the 0-60 ms interval (b) and topographic maps during the ms interval (c) for post-error hits are presented for the familiarised and non-familiarised condition. Vertical bars in Fig. 3a demarcate the ms interval. Fig.4: Results of the fitting procedure (see methods). The duration (in time frames) of the two dominant maps (see results) is depicted for the familiarised and non-familiarised condition, with error bars representing S.E.M. Fig.5: From left to right: horizontal, sagittal, and coronal views of the source localisation obtained using sloreta. Main generators of the dominant topographic map found for the familiarised (a) and non-familiarised (b) condition (see results section) were found to involve non-overlapping regions of the medial frontal cortex. These generators were clearly located more ventral and deeper within the dorsal ACC for the non-familiarised condition, relative to the familiarised condition. A: anterior; P: posterior; L: left; R: right. Results of the t-test between the conditions familiarised non familiarised (c). Positive 39
40 T-values indicate significantly greater activity in the familiarised compared to nonfamiliarised condition in dorsal frontal cortex, including the SMA (p <.05). Figure 1. Figure 2. 40

41 Figure 3. 41

42 Figure 4. Figure 5. 42
Anxiety not only increases, but also alters early error-monitoring functions
 Cognitive, Affective, & Behavioral Neuroscience 2010, 10 (4), 479-492 doi:10.3758/cabn.10.4.479 Anxiety not only increases, but also alters early error-monitoring functions KRISTIEN AARTS AND GILLES POURTOIS
Cognitive, Affective, & Behavioral Neuroscience 2010, 10 (4), 479-492 doi:10.3758/cabn.10.4.479 Anxiety not only increases, but also alters early error-monitoring functions KRISTIEN AARTS AND GILLES POURTOIS
Cognition. Post-error slowing: An orienting account. Wim Notebaert a, *, Femke Houtman a, Filip Van Opstal a, Wim Gevers b, Wim Fias a, Tom Verguts a
 Cognition 111 (2009) 275 279 Contents lists available at ScienceDirect Cognition journal homepage: www.elsevier.com/locate/cognit Brief article Post-error slowing: An orienting account Wim Notebaert a,
Cognition 111 (2009) 275 279 Contents lists available at ScienceDirect Cognition journal homepage: www.elsevier.com/locate/cognit Brief article Post-error slowing: An orienting account Wim Notebaert a,
Running head: AFFECTIVE EVALUATION OF ACTIONS
 Running head: AFFECTIVE EVALUATION OF ACTIONS Erroneous and correct actions have a different affective valence: Evidence from ERPs Kristien Aarts, Jan De Houwer, and Gilles Pourtois Department of Experimental-Clinical
Running head: AFFECTIVE EVALUATION OF ACTIONS Erroneous and correct actions have a different affective valence: Evidence from ERPs Kristien Aarts, Jan De Houwer, and Gilles Pourtois Department of Experimental-Clinical
Cardiac and electro-cortical responses to performance feedback reflect different aspects of feedback processing
 Cardiac and electro-cortical responses to performance feedback reflect different aspects of feedback processing Frederik M. van der Veen 1,2, Sander Nieuwenhuis 3, Eveline A.M Crone 2 & Maurits W. van
Cardiac and electro-cortical responses to performance feedback reflect different aspects of feedback processing Frederik M. van der Veen 1,2, Sander Nieuwenhuis 3, Eveline A.M Crone 2 & Maurits W. van
1 Performance monitoring and anxiety Running head: Performance monitoring and anxiety
 1 Running head: Anxiety disrupts the evaluative component of performance monitoring: An ERP study Kristien Aarts and Gilles Pourtois Department of Experimental Clinical and Health Psychology, Ghent University,
1 Running head: Anxiety disrupts the evaluative component of performance monitoring: An ERP study Kristien Aarts and Gilles Pourtois Department of Experimental Clinical and Health Psychology, Ghent University,
Error-preceding brain activity: Robustness, temporal dynamics, and boundary conditions $
 Biological Psychology 70 (2005) 67 78 www.elsevier.com/locate/biopsycho Error-preceding brain activity: Robustness, temporal dynamics, and boundary conditions $ Greg Hajcak a, *, Sander Nieuwenhuis b,
Biological Psychology 70 (2005) 67 78 www.elsevier.com/locate/biopsycho Error-preceding brain activity: Robustness, temporal dynamics, and boundary conditions $ Greg Hajcak a, *, Sander Nieuwenhuis b,
Reward prediction error signals associated with a modified time estimation task
 Psychophysiology, 44 (2007), 913 917. Blackwell Publishing Inc. Printed in the USA. Copyright r 2007 Society for Psychophysiological Research DOI: 10.1111/j.1469-8986.2007.00561.x BRIEF REPORT Reward prediction
Psychophysiology, 44 (2007), 913 917. Blackwell Publishing Inc. Printed in the USA. Copyright r 2007 Society for Psychophysiological Research DOI: 10.1111/j.1469-8986.2007.00561.x BRIEF REPORT Reward prediction
The Impact of Cognitive Deficits on Conflict Monitoring Predictable Dissociations Between the Error-Related Negativity and N2
 PSYCHOLOGICAL SCIENCE Research Article The Impact of Cognitive Deficits on Conflict Monitoring Predictable Dissociations Between the Error-Related Negativity and N2 Nick Yeung 1 and Jonathan D. Cohen 2,3
PSYCHOLOGICAL SCIENCE Research Article The Impact of Cognitive Deficits on Conflict Monitoring Predictable Dissociations Between the Error-Related Negativity and N2 Nick Yeung 1 and Jonathan D. Cohen 2,3
Electrophysiological correlates of error correction
 Psychophysiology, 42 (2005), 72 82. Blackwell Publishing Inc. Printed in the USA. Copyright r 2005 Society for Psychophysiological Research DOI: 10.1111/j.1469-8986.2005.00265.x Electrophysiological correlates
Psychophysiology, 42 (2005), 72 82. Blackwell Publishing Inc. Printed in the USA. Copyright r 2005 Society for Psychophysiological Research DOI: 10.1111/j.1469-8986.2005.00265.x Electrophysiological correlates
Figure 1. Source localization results for the No Go N2 component. (a) Dipole modeling
 Dipole modeling") Supplementary materials 1 Figure 1. Source localization results for the No Go N2 component. (a) Dipole modeling analyses placed the source of the No Go N2 component in the dorsal ACC, near the ACC source
Supplementary materials 1 Figure 1. Source localization results for the No Go N2 component. (a) Dipole modeling analyses placed the source of the No Go N2 component in the dorsal ACC, near the ACC source
Neuropsychologia 50 (2012) Contents lists available at SciVerse ScienceDirect. Neuropsychologia
 Contents lists available at SciVerse ScienceDirect. Neuropsychologia") Neuropsychologia 50 (2012) 1286 1296 Contents lists available at SciVerse ScienceDirect Neuropsychologia j ourna l ho me pag e: ww w.elsevier.com/locate/neuropsychologia Anxiety disrupts the evaluative
Neuropsychologia 50 (2012) 1286 1296 Contents lists available at SciVerse ScienceDirect Neuropsychologia j ourna l ho me pag e: ww w.elsevier.com/locate/neuropsychologia Anxiety disrupts the evaluative
Brainpotentialsassociatedwithoutcome expectation and outcome evaluation
 COGNITIVE NEUROSCIENCE AND NEUROPSYCHOLOGY Brainpotentialsassociatedwithoutcome expectation and outcome evaluation Rongjun Yu a and Xiaolin Zhou a,b,c a Department of Psychology, Peking University, b State
COGNITIVE NEUROSCIENCE AND NEUROPSYCHOLOGY Brainpotentialsassociatedwithoutcome expectation and outcome evaluation Rongjun Yu a and Xiaolin Zhou a,b,c a Department of Psychology, Peking University, b State
GOAL RELEVANCE INFLUENCES PERFORMANCE MONITORING AT THE LEVEL OF THE FRN AND P3 COMPONENTS
 GOAL RELEVANCE INFLUENCES PERFORMANCE MONITORING AT THE LEVEL OF THE FRN AND P3 COMPONENTS Wioleta Walentowska (1,), Agnes Moors (,3), Katharina Paul (), and Gilles Pourtois () (1) Psychophysiology Laboratory,
GOAL RELEVANCE INFLUENCES PERFORMANCE MONITORING AT THE LEVEL OF THE FRN AND P3 COMPONENTS Wioleta Walentowska (1,), Agnes Moors (,3), Katharina Paul (), and Gilles Pourtois () (1) Psychophysiology Laboratory,
Neuropsychologia 49 (2011) Contents lists available at ScienceDirect. Neuropsychologia
 Contents lists available at ScienceDirect. Neuropsychologia") Neuropsychologia 49 (2011) 405 415 Contents lists available at ScienceDirect Neuropsychologia journal homepage: www.elsevier.com/locate/neuropsychologia Dissociable correlates of response conflict and
Neuropsychologia 49 (2011) 405 415 Contents lists available at ScienceDirect Neuropsychologia journal homepage: www.elsevier.com/locate/neuropsychologia Dissociable correlates of response conflict and
When things look wrong: Theta activity in rule violation
 Neuropsychologia 45 (2007) 3122 3126 Note When things look wrong: Theta activity in rule violation Gabriel Tzur a,b, Andrea Berger a,b, a Department of Behavioral Sciences, Ben-Gurion University of the
Neuropsychologia 45 (2007) 3122 3126 Note When things look wrong: Theta activity in rule violation Gabriel Tzur a,b, Andrea Berger a,b, a Department of Behavioral Sciences, Ben-Gurion University of the
Biological Psychology
 Biological Psychology 87 (2011) 25 34 Contents lists available at ScienceDirect Biological Psychology journal homepage: www.elsevier.com/locate/biopsycho Dissociated roles of the anterior cingulate cortex
Biological Psychology 87 (2011) 25 34 Contents lists available at ScienceDirect Biological Psychology journal homepage: www.elsevier.com/locate/biopsycho Dissociated roles of the anterior cingulate cortex
Sensitivity of Electrophysiological Activity from Medial Frontal Cortex to Utilitarian and Performance Feedback
 Sensitivity of Electrophysiological Activity from Medial Frontal Cortex to Utilitarian and Performance Feedback Sander Nieuwenhuis 1, Nick Yeung 1, Clay B. Holroyd 1, Aaron Schurger 1 and Jonathan D. Cohen
Sensitivity of Electrophysiological Activity from Medial Frontal Cortex to Utilitarian and Performance Feedback Sander Nieuwenhuis 1, Nick Yeung 1, Clay B. Holroyd 1, Aaron Schurger 1 and Jonathan D. Cohen
Acta Psychologica. Conflict and error adaptation in the Simon task. Wim Notebaert, Tom Verguts. Contents lists available at ScienceDirect
 Acta Psychologica 136 (2011) 212 216 Contents lists available at ScienceDirect Acta Psychologica journal homepage: www. e lsevier. com/ l ocate/ a ctpsy Conflict and error adaptation in the Simon task
Acta Psychologica 136 (2011) 212 216 Contents lists available at ScienceDirect Acta Psychologica journal homepage: www. e lsevier. com/ l ocate/ a ctpsy Conflict and error adaptation in the Simon task
Outcome expectancy and not accuracy determines posterror slowing: ERP support
 Cognitive, Affective, & Behavioral Neuroscience 2010, 10 (2), 270-278 doi:10.3758/cabn.10.2.270 Outcome expectancy and not accuracy determines posterror slowing: ERP support ELENA NÚÑEZ CASTELLAR Ghent
Cognitive, Affective, & Behavioral Neuroscience 2010, 10 (2), 270-278 doi:10.3758/cabn.10.2.270 Outcome expectancy and not accuracy determines posterror slowing: ERP support ELENA NÚÑEZ CASTELLAR Ghent
Learning-related changes in reward expectancy are reflected in the feedback-related negativity
 European Journal of Neuroscience, Vol. 27, pp. 1823 1835, 2008 doi:10.1111/j.1460-9568.2008.06138.x Learning-related changes in reward expectancy are reflected in the feedback-related negativity Christian
European Journal of Neuroscience, Vol. 27, pp. 1823 1835, 2008 doi:10.1111/j.1460-9568.2008.06138.x Learning-related changes in reward expectancy are reflected in the feedback-related negativity Christian
Electrophysiological Analysis of Error Monitoring in Schizophrenia
 Journal of Abnormal Psychology Copyright 2006 by the American Psychological Association 2006, Vol. 115, No. 2, 239 250 0021-843X/06/$12.00 DOI: 10.1037/0021-843X.115.2.239 Electrophysiological Analysis
Journal of Abnormal Psychology Copyright 2006 by the American Psychological Association 2006, Vol. 115, No. 2, 239 250 0021-843X/06/$12.00 DOI: 10.1037/0021-843X.115.2.239 Electrophysiological Analysis
The Neural Basis of Error Detection: Conflict Monitoring and the Error-Related Negativity
 Psychological Review Copyright 2004 by the American Psychological Association 2004, Vol. 111, No. 4, 931 959 0033-295X/04/$12.00 DOI: 10.1037/0033-295X.111.4.931 The Neural Basis of Error Detection: Conflict
Psychological Review Copyright 2004 by the American Psychological Association 2004, Vol. 111, No. 4, 931 959 0033-295X/04/$12.00 DOI: 10.1037/0033-295X.111.4.931 The Neural Basis of Error Detection: Conflict
Alterations in Error-Related Brain Activity and Post-Error Behavior Over Time
 Illinois Wesleyan University From the SelectedWorks of Jason R. Themanson, Ph.D Summer 2012 Alterations in Error-Related Brain Activity and Post-Error Behavior Over Time Jason R. Themanson, Illinois Wesleyan
Illinois Wesleyan University From the SelectedWorks of Jason R. Themanson, Ph.D Summer 2012 Alterations in Error-Related Brain Activity and Post-Error Behavior Over Time Jason R. Themanson, Illinois Wesleyan
Interactions of Focal Cortical Lesions With Error Processing: Evidence From Event-Related Brain Potentials
 Neuropsychology Copyright 2002 by the American Psychological Association, Inc. 2002, Vol. 16, No. 4, 548 561 0894-4105/02/$5.00 DOI: 10.1037//0894-4105.16.4.548 Interactions of Focal Cortical Lesions With
Neuropsychology Copyright 2002 by the American Psychological Association, Inc. 2002, Vol. 16, No. 4, 548 561 0894-4105/02/$5.00 DOI: 10.1037//0894-4105.16.4.548 Interactions of Focal Cortical Lesions With
Error-related negativity predicts academic performance
 Psychophysiology, 47 (2010), 192 196. Wiley Periodicals, Inc. Printed in the USA. Copyright r 2009 Society for Psychophysiological Research DOI: 10.1111/j.1469-8986.2009.00877.x BRIEF REPORT Error-related
Psychophysiology, 47 (2010), 192 196. Wiley Periodicals, Inc. Printed in the USA. Copyright r 2009 Society for Psychophysiological Research DOI: 10.1111/j.1469-8986.2009.00877.x BRIEF REPORT Error-related
Neural Correlates of Human Cognitive Function:
 Neural Correlates of Human Cognitive Function: A Comparison of Electrophysiological and Other Neuroimaging Approaches Leun J. Otten Institute of Cognitive Neuroscience & Department of Psychology University
Neural Correlates of Human Cognitive Function: A Comparison of Electrophysiological and Other Neuroimaging Approaches Leun J. Otten Institute of Cognitive Neuroscience & Department of Psychology University
Working memory capacity and task goals modulate error-related ERPs
 Received: 26 November Accepted: November 16 DOI.1111/psyp.128 ORIGINAL ARTICLE Working memory capacity and task goals modulate error-related ERPs James R. Coleman Jason M. Watson David L. Strayer Department
Received: 26 November Accepted: November 16 DOI.1111/psyp.128 ORIGINAL ARTICLE Working memory capacity and task goals modulate error-related ERPs James R. Coleman Jason M. Watson David L. Strayer Department
Supplementary materials for: Executive control processes underlying multi- item working memory
 Supplementary materials for: Executive control processes underlying multi- item working memory Antonio H. Lara & Jonathan D. Wallis Supplementary Figure 1 Supplementary Figure 1. Behavioral measures of
Supplementary materials for: Executive control processes underlying multi- item working memory Antonio H. Lara & Jonathan D. Wallis Supplementary Figure 1 Supplementary Figure 1. Behavioral measures of
IS THE HIGH PROBABILITY OF TYPE II ERROR AN ISSUE IN ERROR AWARENESS ERP STUDIES?
 IS THE HIGH PROBABILITY OF TYPE II ERROR AN ISSUE IN ERROR AWARENESS ERP STUDIES? Master Degree Project in Cognitive Neuroscience One year Advanced level 30 ECTS Spring term 2016 Boushra Dalile Supervisor(s):
IS THE HIGH PROBABILITY OF TYPE II ERROR AN ISSUE IN ERROR AWARENESS ERP STUDIES? Master Degree Project in Cognitive Neuroscience One year Advanced level 30 ECTS Spring term 2016 Boushra Dalile Supervisor(s):
DATA MANAGEMENT & TYPES OF ANALYSES OFTEN USED. Dennis L. Molfese University of Nebraska - Lincoln
 DATA MANAGEMENT & TYPES OF ANALYSES OFTEN USED Dennis L. Molfese University of Nebraska - Lincoln 1 DATA MANAGEMENT Backups Storage Identification Analyses 2 Data Analysis Pre-processing Statistical Analysis
DATA MANAGEMENT & TYPES OF ANALYSES OFTEN USED Dennis L. Molfese University of Nebraska - Lincoln 1 DATA MANAGEMENT Backups Storage Identification Analyses 2 Data Analysis Pre-processing Statistical Analysis
ERP Correlates of Feedback and Reward Processing in the Presence and Absence of Response Choice
 Cerebral Cortex May 2005;15:535-544 doi:10.1093/cercor/bhh153 Advance Access publication August 18, 2004 ERP Correlates of Feedback and Reward Processing in the Presence and Absence of Response Choice
Cerebral Cortex May 2005;15:535-544 doi:10.1093/cercor/bhh153 Advance Access publication August 18, 2004 ERP Correlates of Feedback and Reward Processing in the Presence and Absence of Response Choice
Mental representation of number in different numerical forms
 Submitted to Current Biology Mental representation of number in different numerical forms Anna Plodowski, Rachel Swainson, Georgina M. Jackson, Chris Rorden and Stephen R. Jackson School of Psychology
Submitted to Current Biology Mental representation of number in different numerical forms Anna Plodowski, Rachel Swainson, Georgina M. Jackson, Chris Rorden and Stephen R. Jackson School of Psychology
Response-selection Conflict Contributes to Inhibition of Return
 Response-selection Conflict Contributes to Inhibition of Return David J. Prime and Pierre Jolicoeur Abstract & Here we examined the relationship between inhibition of return (IOR) and response-selection
Response-selection Conflict Contributes to Inhibition of Return David J. Prime and Pierre Jolicoeur Abstract & Here we examined the relationship between inhibition of return (IOR) and response-selection
Attention Response Functions: Characterizing Brain Areas Using fmri Activation during Parametric Variations of Attentional Load
 Attention Response Functions: Characterizing Brain Areas Using fmri Activation during Parametric Variations of Attentional Load Intro Examine attention response functions Compare an attention-demanding
Attention Response Functions: Characterizing Brain Areas Using fmri Activation during Parametric Variations of Attentional Load Intro Examine attention response functions Compare an attention-demanding
Perceptual properties of feedback stimuli influence the feedback-related negativity in the flanker gambling task
 Psychophysiology, 51 (014), 78 788. Wiley Periodicals, Inc. Printed in the USA. Copyright 014 Society for Psychophysiological Research DOI: 10.1111/psyp.116 Perceptual properties of feedback stimuli influence
Psychophysiology, 51 (014), 78 788. Wiley Periodicals, Inc. Printed in the USA. Copyright 014 Society for Psychophysiological Research DOI: 10.1111/psyp.116 Perceptual properties of feedback stimuli influence
Supplementary Materials for. Error Related Brain Activity Reveals Self-Centric Motivation: Culture Matters. Shinobu Kitayama and Jiyoung Park
 1 Supplementary Materials for Error Related Brain Activity Reveals Self-Centric Motivation: Culture Matters Shinobu Kitayama and Jiyoung Park University of Michigan *To whom correspondence should be addressed.
1 Supplementary Materials for Error Related Brain Activity Reveals Self-Centric Motivation: Culture Matters Shinobu Kitayama and Jiyoung Park University of Michigan *To whom correspondence should be addressed.
BRAIN COMPUTER INTERFACEs (BCIs), as any other
, as any other") IEEE TRANSACTIONS ON BIOMEDICAL ENGINEERING, VOL. 55, NO. 3, MARCH 2008 923 Error-Related EEG Potentials Generated During Simulated Brain Computer Interaction Pierre W. Ferrez* and José del R. Millán*
IEEE TRANSACTIONS ON BIOMEDICAL ENGINEERING, VOL. 55, NO. 3, MARCH 2008 923 Error-Related EEG Potentials Generated During Simulated Brain Computer Interaction Pierre W. Ferrez* and José del R. Millán*
What Matters in the Cued Task-Switching Paradigm: Tasks or Cues? Ulrich Mayr. University of Oregon
 What Matters in the Cued Task-Switching Paradigm: Tasks or Cues? Ulrich Mayr University of Oregon Running head: Cue-specific versus task-specific switch costs Ulrich Mayr Department of Psychology University
What Matters in the Cued Task-Switching Paradigm: Tasks or Cues? Ulrich Mayr University of Oregon Running head: Cue-specific versus task-specific switch costs Ulrich Mayr Department of Psychology University
Electrophysiological correlates of anterior cingulate function in a go/no-go task: Effects of response conflict and trial type frequency
 Cognitive, Affective, & Behavioral Neuroscience 2003, 3 (1), 17-26 Electrophysiological correlates of anterior cingulate function in a go/no-go task: Effects of response conflict and trial type frequency
Cognitive, Affective, & Behavioral Neuroscience 2003, 3 (1), 17-26 Electrophysiological correlates of anterior cingulate function in a go/no-go task: Effects of response conflict and trial type frequency
This presentation is the intellectual property of the author. Contact them for permission to reprint and/or distribute.
 Modified Combinatorial Nomenclature Montage, Review, and Analysis of High Density EEG Terrence D. Lagerlund, M.D., Ph.D. CP1208045-16 Disclosure Relevant financial relationships None Off-label/investigational
Modified Combinatorial Nomenclature Montage, Review, and Analysis of High Density EEG Terrence D. Lagerlund, M.D., Ph.D. CP1208045-16 Disclosure Relevant financial relationships None Off-label/investigational
Dissociable neural correlates for familiarity and recollection during the encoding and retrieval of pictures
 Cognitive Brain Research 18 (2004) 255 272 Research report Dissociable neural correlates for familiarity and recollection during the encoding and retrieval of pictures Audrey Duarte a, *, Charan Ranganath
Cognitive Brain Research 18 (2004) 255 272 Research report Dissociable neural correlates for familiarity and recollection during the encoding and retrieval of pictures Audrey Duarte a, *, Charan Ranganath
Biological Psychology
 Biological Psychology 114 (2016) 108 116 Contents lists available at ScienceDirect Biological Psychology journal homepage: www.elsevier.com/locate/biopsycho Impacts of motivational valence on the error-related
Biological Psychology 114 (2016) 108 116 Contents lists available at ScienceDirect Biological Psychology journal homepage: www.elsevier.com/locate/biopsycho Impacts of motivational valence on the error-related
An investigation of the effect of preparation on response execution and inhibition in the go/nogo task
 University of Wollongong Research Online University of Wollongong Thesis Collection 1954-2016 University of Wollongong Thesis Collections 2005 An investigation of the effect of preparation on response
University of Wollongong Research Online University of Wollongong Thesis Collection 1954-2016 University of Wollongong Thesis Collections 2005 An investigation of the effect of preparation on response
What matters in the cued task-switching paradigm: Tasks or cues?
 Journal Psychonomic Bulletin & Review 2006,?? 13 (?), (5),???-??? 794-799 What matters in the cued task-switching paradigm: Tasks or cues? ULRICH MAYR University of Oregon, Eugene, Oregon Schneider and
Journal Psychonomic Bulletin & Review 2006,?? 13 (?), (5),???-??? 794-799 What matters in the cued task-switching paradigm: Tasks or cues? ULRICH MAYR University of Oregon, Eugene, Oregon Schneider and
Functional lesions and human action monitoring: combining repetitive transcranial magnetic stimulation and event-related brain potentials
 Clinical Neurophysiology 115 (2004) 145 153 www.elsevier.com/locate/clinph Functional lesions and human action monitoring: combining repetitive transcranial magnetic stimulation and event-related brain
Clinical Neurophysiology 115 (2004) 145 153 www.elsevier.com/locate/clinph Functional lesions and human action monitoring: combining repetitive transcranial magnetic stimulation and event-related brain
Why should I care? Challenging free will attenuates neural reaction to errors
 doi:10.1093/scan/nsu068 SCAN (2015) 10, 262^268 Why should I care? Challenging free will attenuates neural reaction to errors Davide Rigoni, 1 Gilles Pourtois, 2 and Marcel Brass 1,3 1 Department of Experimental
doi:10.1093/scan/nsu068 SCAN (2015) 10, 262^268 Why should I care? Challenging free will attenuates neural reaction to errors Davide Rigoni, 1 Gilles Pourtois, 2 and Marcel Brass 1,3 1 Department of Experimental
Event-Related Potentials Recorded during Human-Computer Interaction
 Proceedings of the First International Conference on Complex Medical Engineering (CME2005) May 15-18, 2005, Takamatsu, Japan (Organized Session No. 20). Paper No. 150, pp. 715-719. Event-Related Potentials
Proceedings of the First International Conference on Complex Medical Engineering (CME2005) May 15-18, 2005, Takamatsu, Japan (Organized Session No. 20). Paper No. 150, pp. 715-719. Event-Related Potentials
To Bet or Not to Bet? The Error Negativity or Error-related Negativity Associated with Risk-taking Choices
 To Bet or Not to Bet? The Error Negativity or Error-related Negativity Associated with Risk-taking Choices Rongjun Yu 1 and Xiaolin Zhou 1,2,3 Abstract & The functional significance of error-related negativity
To Bet or Not to Bet? The Error Negativity or Error-related Negativity Associated with Risk-taking Choices Rongjun Yu 1 and Xiaolin Zhou 1,2,3 Abstract & The functional significance of error-related negativity
Aerobic Fitness and Cognitive Development: Event-Related Brain Potential and Task Performance Indices of Executive Control in Preadolescent Children
 Illinois Wesleyan University From the SelectedWorks of Jason R. Themanson, Ph.D 2009 Aerobic Fitness and Cognitive Development: Event-Related Brain Potential and Task Performance Indices of Executive Control
Illinois Wesleyan University From the SelectedWorks of Jason R. Themanson, Ph.D 2009 Aerobic Fitness and Cognitive Development: Event-Related Brain Potential and Task Performance Indices of Executive Control
Performance monitoring and the causal attribution of errors
 Cogn Affect Behav Neurosci (2011) 11:309 320 DOI 10.3758/s13415-011-0033-2 Performance monitoring and the causal attribution of errors Marco Steinhauser & Andrea Kiesel Published online: 14 May 2011 #
Cogn Affect Behav Neurosci (2011) 11:309 320 DOI 10.3758/s13415-011-0033-2 Performance monitoring and the causal attribution of errors Marco Steinhauser & Andrea Kiesel Published online: 14 May 2011 #
A Computational Model of Anterior Cingulate Function in Speeded Response Tasks: Effects of Frequency, Sequence, and Conflict
 A Computational Model of Anterior Cingulate Function in Speeded Response Tasks: Effects of Frequency, Sequence, and Conflict Andrew D. Jones 1, Raymond Cho 2,3, Leigh E. Nystrom 2, Jonathan D. Cohen 2,3,
A Computational Model of Anterior Cingulate Function in Speeded Response Tasks: Effects of Frequency, Sequence, and Conflict Andrew D. Jones 1, Raymond Cho 2,3, Leigh E. Nystrom 2, Jonathan D. Cohen 2,3,
Flexible Conflict Management: Conflict Avoidance and Conflict Adjustment in Reactive. Cognitive Control
 1 RUNNING HEAD: AVOIDANCE OF AND ADJUSTMENT TO CONFLICT Flexible Conflict Management: Conflict Avoidance and Conflict Adjustment in Reactive Cognitive Control David Dignath, Andrea Kiesel, & Andreas B.
1 RUNNING HEAD: AVOIDANCE OF AND ADJUSTMENT TO CONFLICT Flexible Conflict Management: Conflict Avoidance and Conflict Adjustment in Reactive Cognitive Control David Dignath, Andrea Kiesel, & Andreas B.
Learning to Become an Expert: Reinforcement Learning and the Acquisition of Perceptual Expertise
 Learning to Become an Expert: Reinforcement Learning and the Acquisition of Perceptual Expertise Olav E. Krigolson 1,2, Lara J. Pierce 1, Clay B. Holroyd 1, and James W. Tanaka 1 Abstract & To elucidate
Learning to Become an Expert: Reinforcement Learning and the Acquisition of Perceptual Expertise Olav E. Krigolson 1,2, Lara J. Pierce 1, Clay B. Holroyd 1, and James W. Tanaka 1 Abstract & To elucidate
Conflict monitoring and feature overlap: Two sources of sequential modulations
 Psychonomic Bulletin & Review 2007, 14 (4), 742-748 Conflict monitoring and feature overlap: Two sources of sequential modulations Çağlar Akçay and Eliot Hazeltine University of Iowa, Iowa City, Iowa Recent
Psychonomic Bulletin & Review 2007, 14 (4), 742-748 Conflict monitoring and feature overlap: Two sources of sequential modulations Çağlar Akçay and Eliot Hazeltine University of Iowa, Iowa City, Iowa Recent
SUPPLEMENTARY INFORMATION. Table 1 Patient characteristics Preoperative. language testing
 Categorical Speech Representation in the Human Superior Temporal Gyrus Edward F. Chang, Jochem W. Rieger, Keith D. Johnson, Mitchel S. Berger, Nicholas M. Barbaro, Robert T. Knight SUPPLEMENTARY INFORMATION
Categorical Speech Representation in the Human Superior Temporal Gyrus Edward F. Chang, Jochem W. Rieger, Keith D. Johnson, Mitchel S. Berger, Nicholas M. Barbaro, Robert T. Knight SUPPLEMENTARY INFORMATION
Processed by HBI: Russia/Switzerland/USA
 1 CONTENTS I Personal and clinical data II Conclusion. III Recommendations for therapy IV Report. 1. Procedures of EEG recording and analysis 2. Search for paroxysms 3. Eyes Open background EEG rhythms
1 CONTENTS I Personal and clinical data II Conclusion. III Recommendations for therapy IV Report. 1. Procedures of EEG recording and analysis 2. Search for paroxysms 3. Eyes Open background EEG rhythms
Neural Correlates of Complex Tone Processing and Hemispheric Asymmetry
 International Journal of Undergraduate Research and Creative Activities Volume 5 Article 3 June 2013 Neural Correlates of Complex Tone Processing and Hemispheric Asymmetry Whitney R. Arthur Central Washington
International Journal of Undergraduate Research and Creative Activities Volume 5 Article 3 June 2013 Neural Correlates of Complex Tone Processing and Hemispheric Asymmetry Whitney R. Arthur Central Washington
Resistance to forgetting associated with hippocampus-mediated. reactivation during new learning
 Resistance to Forgetting 1 Resistance to forgetting associated with hippocampus-mediated reactivation during new learning Brice A. Kuhl, Arpeet T. Shah, Sarah DuBrow, & Anthony D. Wagner Resistance to
Resistance to Forgetting 1 Resistance to forgetting associated with hippocampus-mediated reactivation during new learning Brice A. Kuhl, Arpeet T. Shah, Sarah DuBrow, & Anthony D. Wagner Resistance to
Stimulus and Response-Related Conflict Processing in the Human Brain
 Stimulus and Response-Related Conflict Processing in the Human Brain A dissertation submitted in partial fulfillment of the requirements for the degree of Doctor of Philosophy at George Mason University
Stimulus and Response-Related Conflict Processing in the Human Brain A dissertation submitted in partial fulfillment of the requirements for the degree of Doctor of Philosophy at George Mason University
Beware misleading cues: Perceptual similarity modulates the N2/P3 complex
 Psychophysiology, 43 (2006), 253 260. Blackwell Publishing Inc. Printed in the USA. Copyright r 2006 Society for Psychophysiological Research DOI: 10.1111/j.1469-8986.2006.00409.x Beware misleading cues:
Psychophysiology, 43 (2006), 253 260. Blackwell Publishing Inc. Printed in the USA. Copyright r 2006 Society for Psychophysiological Research DOI: 10.1111/j.1469-8986.2006.00409.x Beware misleading cues:
The feedback correct-related positivity: Sensitivity of the event-related brain potential to unexpected positive feedback
 Psychophysiology, 45 (2008), ** **. Blackwell Publishing Inc. Printed in the USA. Copyright r 2008 Society for Psychophysiological Research DOI: 10.1111/j.1469-8986.2008.00668.x The feedback correct-related
Psychophysiology, 45 (2008), ** **. Blackwell Publishing Inc. Printed in the USA. Copyright r 2008 Society for Psychophysiological Research DOI: 10.1111/j.1469-8986.2008.00668.x The feedback correct-related
Distinct Frontal Networks Are Involved in Adapting to Internally and Externally Signaled Errors
 Cerebral Cortex March 2013;23:703 713 doi:10.1093/cercor/bhs056 Advance Access publication March 16, 2012 Distinct Frontal Networks Are Involved in Adapting to Internally and Externally Signaled Errors
Cerebral Cortex March 2013;23:703 713 doi:10.1093/cercor/bhs056 Advance Access publication March 16, 2012 Distinct Frontal Networks Are Involved in Adapting to Internally and Externally Signaled Errors
Patricia L. Davies Department of Occupational Therapy Colorado State University
 DEVELOPMENTAL NEUROPSYCHOLOGY, 25(3), 355 376 Copyright 2004, Lawrence Erlbaum Associates, Inc. Development of Response-Monitoring ERPs in 7- to 25-Year-Olds Patricia L. Davies Department of Occupational
DEVELOPMENTAL NEUROPSYCHOLOGY, 25(3), 355 376 Copyright 2004, Lawrence Erlbaum Associates, Inc. Development of Response-Monitoring ERPs in 7- to 25-Year-Olds Patricia L. Davies Department of Occupational
Supplementary Motor Area exerts Proactive and Reactive Control of Arm Movements
 Supplementary Material Supplementary Motor Area exerts Proactive and Reactive Control of Arm Movements Xiaomo Chen, Katherine Wilson Scangos 2 and Veit Stuphorn,2 Department of Psychological and Brain
Supplementary Material Supplementary Motor Area exerts Proactive and Reactive Control of Arm Movements Xiaomo Chen, Katherine Wilson Scangos 2 and Veit Stuphorn,2 Department of Psychological and Brain
Reliability of error-related brain activity
 BRAIN RESEARCH 1284 (2009) 89 99 available at www.sciencedirect.com www.elsevier.com/locate/brainres Research Report Reliability of error-related brain activity Doreen M. Olvet, Greg Hajcak Department
BRAIN RESEARCH 1284 (2009) 89 99 available at www.sciencedirect.com www.elsevier.com/locate/brainres Research Report Reliability of error-related brain activity Doreen M. Olvet, Greg Hajcak Department
NeuroImage 50 (2010) Contents lists available at ScienceDirect. NeuroImage. journal homepage:
 Contents lists available at ScienceDirect. NeuroImage. journal homepage:") NeuroImage 50 (2010) 329 339 Contents lists available at ScienceDirect NeuroImage journal homepage: www.elsevier.com/locate/ynimg Switching associations between facial identity and emotional expression:
NeuroImage 50 (2010) 329 339 Contents lists available at ScienceDirect NeuroImage journal homepage: www.elsevier.com/locate/ynimg Switching associations between facial identity and emotional expression:
FINAL PROGRESS REPORT
 (1) Foreword (optional) (2) Table of Contents (if report is more than 10 pages) (3) List of Appendixes, Illustrations and Tables (if applicable) (4) Statement of the problem studied FINAL PROGRESS REPORT
(1) Foreword (optional) (2) Table of Contents (if report is more than 10 pages) (3) List of Appendixes, Illustrations and Tables (if applicable) (4) Statement of the problem studied FINAL PROGRESS REPORT
Journal of Psychophysiology An International Journal
 ISSN-L 0269-8803 ISSN-Print 0269-8803 ISSN-Online 2151-2124 Official Publication of the Federation of European Psychophysiology Societies Journal of Psychophysiology An International Journal 3/13 www.hogrefe.com/journals/jop
ISSN-L 0269-8803 ISSN-Print 0269-8803 ISSN-Online 2151-2124 Official Publication of the Federation of European Psychophysiology Societies Journal of Psychophysiology An International Journal 3/13 www.hogrefe.com/journals/jop
Manuscript under review for Psychological Science. Direct Electrophysiological Measurement of Attentional Templates in Visual Working Memory
 Direct Electrophysiological Measurement of Attentional Templates in Visual Working Memory Journal: Psychological Science Manuscript ID: PSCI-0-0.R Manuscript Type: Short report Date Submitted by the Author:
Direct Electrophysiological Measurement of Attentional Templates in Visual Working Memory Journal: Psychological Science Manuscript ID: PSCI-0-0.R Manuscript Type: Short report Date Submitted by the Author:
The error-related potential and BCIs
 The error-related potential and BCIs Sandra Rousseau 1, Christian Jutten 2 and Marco Congedo 2 Gipsa-lab - DIS 11 rue des Mathématiques BP 46 38402 Saint Martin d Héres Cedex - France Abstract. The error-related
The error-related potential and BCIs Sandra Rousseau 1, Christian Jutten 2 and Marco Congedo 2 Gipsa-lab - DIS 11 rue des Mathématiques BP 46 38402 Saint Martin d Héres Cedex - France Abstract. The error-related
Time Course of Error Detection and Correction in Humans: Neurophysiological Evidence
 The Journal of Neuroscience, November 15, 2002, 22(22):9990 9996 Time Course of Error Detection and Correction in Humans: Neurophysiological Evidence Antoni Rodríguez-Fornells, Arthur R. Kurzbuch, and
The Journal of Neuroscience, November 15, 2002, 22(22):9990 9996 Time Course of Error Detection and Correction in Humans: Neurophysiological Evidence Antoni Rodríguez-Fornells, Arthur R. Kurzbuch, and
An ERP Examination of the Different Effects of Sleep Deprivation on Exogenously Cued and Endogenously Cued Attention
 Sleep Deprivation and Selective Attention An ERP Examination of the Different Effects of Sleep Deprivation on Exogenously Cued and Endogenously Cued Attention Logan T. Trujillo, PhD 1 ; Steve Kornguth,
Sleep Deprivation and Selective Attention An ERP Examination of the Different Effects of Sleep Deprivation on Exogenously Cued and Endogenously Cued Attention Logan T. Trujillo, PhD 1 ; Steve Kornguth,
Drink alcohol and dim the lights: The impact of cognitive deficits on medial frontal cortex function
 Cognitive, Affective, & Behavioral Neuroscience 2007, 7 (4), 347-355 Drink alcohol and dim the lights: The impact of cognitive deficits on medial frontal cortex function Nick Yeung University of Oxford,
Cognitive, Affective, & Behavioral Neuroscience 2007, 7 (4), 347-355 Drink alcohol and dim the lights: The impact of cognitive deficits on medial frontal cortex function Nick Yeung University of Oxford,
Balancing Cognitive Demands: Control Adjustments in the Stop-Signal Paradigm
 Journal of Experimental Psychology: Learning, Memory, and Cognition 2011, Vol. 37, No. 2, 392 404 2010 American Psychological Association 0278-7393/10/$12.00 DOI: 10.1037/a0021800 Balancing Cognitive Demands:
Journal of Experimental Psychology: Learning, Memory, and Cognition 2011, Vol. 37, No. 2, 392 404 2010 American Psychological Association 0278-7393/10/$12.00 DOI: 10.1037/a0021800 Balancing Cognitive Demands:
Visual Context Dan O Shea Prof. Fei Fei Li, COS 598B
 Visual Context Dan O Shea Prof. Fei Fei Li, COS 598B Cortical Analysis of Visual Context Moshe Bar, Elissa Aminoff. 2003. Neuron, Volume 38, Issue 2, Pages 347 358. Visual objects in context Moshe Bar.
Visual Context Dan O Shea Prof. Fei Fei Li, COS 598B Cortical Analysis of Visual Context Moshe Bar, Elissa Aminoff. 2003. Neuron, Volume 38, Issue 2, Pages 347 358. Visual objects in context Moshe Bar.
Submitted report on Sufi recordings at AAPB 2013 in Portland. Not for general distribution. Thomas F. Collura, Ph.D. July, 2013
 Submitted report on Sufi recordings at AAPB 2013 in Portland Not for general distribution. Thomas F. Collura, Ph.D. July, 2013 Summary of EEG findings The intent of the EEG monitoring was to see which
Submitted report on Sufi recordings at AAPB 2013 in Portland Not for general distribution. Thomas F. Collura, Ph.D. July, 2013 Summary of EEG findings The intent of the EEG monitoring was to see which
The impact of numeration on visual attention during a psychophysical task; An ERP study
 The impact of numeration on visual attention during a psychophysical task; An ERP study Armita Faghani Jadidi, Raheleh Davoodi, Mohammad Hassan Moradi Department of Biomedical Engineering Amirkabir University
The impact of numeration on visual attention during a psychophysical task; An ERP study Armita Faghani Jadidi, Raheleh Davoodi, Mohammad Hassan Moradi Department of Biomedical Engineering Amirkabir University
Context Specificity of Post-Error and Post-Conflict Cognitive Control Adjustments
 Context Specificity of Post-Error and Post-Conflict Cognitive Control Adjustments Sarah E. Forster 1, Raymond Y. Cho 2,3,4 * 1 Department of Psychological and Brain Sciences, Indiana University, Bloomington,
Context Specificity of Post-Error and Post-Conflict Cognitive Control Adjustments Sarah E. Forster 1, Raymond Y. Cho 2,3,4 * 1 Department of Psychological and Brain Sciences, Indiana University, Bloomington,
Psychological Science
 Psychological Science http://pss.sagepub.com/ Beyond Fear : Rapid Spatial Orienting Toward Positive Emotional Stimuli Tobias Brosch, David Sander, Gilles Pourtois and Klaus R. Scherer Psychological Science
Psychological Science http://pss.sagepub.com/ Beyond Fear : Rapid Spatial Orienting Toward Positive Emotional Stimuli Tobias Brosch, David Sander, Gilles Pourtois and Klaus R. Scherer Psychological Science
Ingroup categorization and response conflict: Interactive effects of target race, flanker compatibility, and infrequency on N2 amplitude
 Psychophysiology, 47 (2010), 596 601. Wiley Periodicals, Inc. Printed in the USA. Copyright r 2010 Society for Psychophysiological Research DOI: 10.1111/j.1469-8986.2010.00963.x BRIEF REPORT Ingroup categorization
Psychophysiology, 47 (2010), 596 601. Wiley Periodicals, Inc. Printed in the USA. Copyright r 2010 Society for Psychophysiological Research DOI: 10.1111/j.1469-8986.2010.00963.x BRIEF REPORT Ingroup categorization
Running Head: VISUAL WORKING MEMORY LOAD AND ATTENTIONAL CONTROL
 Running Head: VISUAL WORKING MEMORY LOAD AND ATTENTIONAL CONTROL Visual working memory load disrupts template-guided attentional selection during visual search Nick Berggren* and Martin Eimer Department
Running Head: VISUAL WORKING MEMORY LOAD AND ATTENTIONAL CONTROL Visual working memory load disrupts template-guided attentional selection during visual search Nick Berggren* and Martin Eimer Department
Relating cognitive and affective theories of the errorrelated
 3 Performance Monitoring, Decision Making, and Cognitive Control Monterosso J, Ehrman R, Napier K, O'Brien CP, Childress AR (2001) Three Decision-Making Tasks in Cocaine- Dependent Patients: Do they measure
3 Performance Monitoring, Decision Making, and Cognitive Control Monterosso J, Ehrman R, Napier K, O'Brien CP, Childress AR (2001) Three Decision-Making Tasks in Cocaine- Dependent Patients: Do they measure
Independence of Visual Awareness from the Scope of Attention: an Electrophysiological Study
 Cerebral Cortex March 2006;16:415-424 doi:10.1093/cercor/bhi121 Advance Access publication June 15, 2005 Independence of Visual Awareness from the Scope of Attention: an Electrophysiological Study Mika
Cerebral Cortex March 2006;16:415-424 doi:10.1093/cercor/bhi121 Advance Access publication June 15, 2005 Independence of Visual Awareness from the Scope of Attention: an Electrophysiological Study Mika
ERP correlates of retrieval orientation: Direct versus indirect memory tasks
 available at www.sciencedirect.com www.elsevier.com/locate/brainres Research Report ERP correlates of retrieval orientation: Direct versus indirect memory tasks Michael Hornberger a, Michael D. Rugg b,
available at www.sciencedirect.com www.elsevier.com/locate/brainres Research Report ERP correlates of retrieval orientation: Direct versus indirect memory tasks Michael Hornberger a, Michael D. Rugg b,
ANALYZING EVENT-RELATED POTENTIALS
 Adavanced Lifespan Neurocognitive Development: EEG signal processing for lifespan research Dr. Manosusos Klados Liesa Ilg ANALYZING EVENT-RELATED POTENTIALS Chair for Lifespan Developmental Neuroscience
Adavanced Lifespan Neurocognitive Development: EEG signal processing for lifespan research Dr. Manosusos Klados Liesa Ilg ANALYZING EVENT-RELATED POTENTIALS Chair for Lifespan Developmental Neuroscience
Address for correspondence: School of Psychological Sciences, Zochonis Building, University of Manchester, Oxford Road, M139PL, Manchester, UK
 BBS-D-15-00891_ Mather_ Talmi & Barnacle Emotionally arousing context modulates the ERP correlates of neutral picture processing: An ERP test of the Glutamate Amplifies Noradrenergic Effects (GANE) model
BBS-D-15-00891_ Mather_ Talmi & Barnacle Emotionally arousing context modulates the ERP correlates of neutral picture processing: An ERP test of the Glutamate Amplifies Noradrenergic Effects (GANE) model
ARTICLE IN PRESS. Biological Psychology xxx (2014) xxx xxx. Contents lists available at ScienceDirect. Biological Psychology
 xxx xxx. Contents lists available at ScienceDirect. Biological Psychology") Biological Psychology xxx (2014) xxx xxx Contents lists available at ScienceDirect Biological Psychology jo ur nal home p age: www.elsevier.com/locate/biopsycho 1 2 3 Q1 4 Q2 Mindset induction effects
Biological Psychology xxx (2014) xxx xxx Contents lists available at ScienceDirect Biological Psychology jo ur nal home p age: www.elsevier.com/locate/biopsycho 1 2 3 Q1 4 Q2 Mindset induction effects
State sadness reduces neural sensitivity to nonrewards versus rewards Dan Foti and Greg Hajcak
 Neurophysiology, basic and clinical 143 State sadness reduces neural sensitivity to nonrewards versus rewards Dan Foti and Greg Hajcak Both behavioral and neural evidence suggests that depression is associated
Neurophysiology, basic and clinical 143 State sadness reduces neural sensitivity to nonrewards versus rewards Dan Foti and Greg Hajcak Both behavioral and neural evidence suggests that depression is associated
Master Thesis: Can the negative deflections found with EEG on frontocentral electrodes be explained by one underlying process?
 Master Thesis: Can the negative deflections found with EEG on frontocentral electrodes be explained by one underlying process? Name: Gerben Klein Studentnr: 3029182 Master: Neuroscience and Cognition,
Master Thesis: Can the negative deflections found with EEG on frontocentral electrodes be explained by one underlying process? Name: Gerben Klein Studentnr: 3029182 Master: Neuroscience and Cognition,
Description of the Spectro-temporal unfolding of temporal orienting of attention.
 Description of the Spectro-temporal unfolding of temporal orienting of attention. All behaviors unfold over time; therefore, our ability to perceive and adapt our behavior according to the temporal constraints
Description of the Spectro-temporal unfolding of temporal orienting of attention. All behaviors unfold over time; therefore, our ability to perceive and adapt our behavior according to the temporal constraints
SUPPLEMENTARY INFORMATION. Supplementary Figure 1
 SUPPLEMENTARY INFORMATION Supplementary Figure 1 The supralinear events evoked in CA3 pyramidal cells fulfill the criteria for NMDA spikes, exhibiting a threshold, sensitivity to NMDAR blockade, and all-or-none
SUPPLEMENTARY INFORMATION Supplementary Figure 1 The supralinear events evoked in CA3 pyramidal cells fulfill the criteria for NMDA spikes, exhibiting a threshold, sensitivity to NMDAR blockade, and all-or-none
The mind s eye, looking inward? In search of executive control in internal attention shifting
 Psychophysiology, 40 (2003), 572 585. Blackwell Publishing Inc. Printed in the USA. Copyright r 2003 Society for Psychophysiological Research The mind s eye, looking inward? In search of executive control
Psychophysiology, 40 (2003), 572 585. Blackwell Publishing Inc. Printed in the USA. Copyright r 2003 Society for Psychophysiological Research The mind s eye, looking inward? In search of executive control
2. Which of the following is not an element of McGuire s chain of cognitive responses model? a. Attention b. Comprehension c. Yielding d.
 Chapter 10: Cognitive Processing of Attitudes 1. McGuire s (1969) model of information processing can be best described as which of the following? a. Sequential b. Parallel c. Automatic 2. Which of the
Chapter 10: Cognitive Processing of Attitudes 1. McGuire s (1969) model of information processing can be best described as which of the following? a. Sequential b. Parallel c. Automatic 2. Which of the
The Function and Organization of Lateral Prefrontal Cortex: A Test of Competing Hypotheses
 The Function and Organization of Lateral Prefrontal Cortex: A Test of Competing Hypotheses Jeremy R. Reynolds 1 *, Randall C. O Reilly 2, Jonathan D. Cohen 3, Todd S. Braver 4 1 Department of Psychology,
The Function and Organization of Lateral Prefrontal Cortex: A Test of Competing Hypotheses Jeremy R. Reynolds 1 *, Randall C. O Reilly 2, Jonathan D. Cohen 3, Todd S. Braver 4 1 Department of Psychology,
A model of dual control mechanisms through anterior cingulate and prefrontal cortex interactions
 Neurocomputing 69 (2006) 1322 1326 www.elsevier.com/locate/neucom A model of dual control mechanisms through anterior cingulate and prefrontal cortex interactions Nicola De Pisapia, Todd S. Braver Cognitive
Neurocomputing 69 (2006) 1322 1326 www.elsevier.com/locate/neucom A model of dual control mechanisms through anterior cingulate and prefrontal cortex interactions Nicola De Pisapia, Todd S. Braver Cognitive
Outline of Talk. Introduction to EEG and Event Related Potentials. Key points. My path to EEG
 Outline of Talk Introduction to EEG and Event Related Potentials Shafali Spurling Jeste Assistant Professor in Psychiatry and Neurology UCLA Center for Autism Research and Treatment Basic definitions and
Outline of Talk Introduction to EEG and Event Related Potentials Shafali Spurling Jeste Assistant Professor in Psychiatry and Neurology UCLA Center for Autism Research and Treatment Basic definitions and
The Influence of Motivational Salience on Attention Selection: An ERP Investigation
 University of South Florida Scholar Commons Graduate Theses and Dissertations Graduate School 6-30-2016 The Influence of Motivational Salience on Attention Selection: An ERP Investigation Constanza De
University of South Florida Scholar Commons Graduate Theses and Dissertations Graduate School 6-30-2016 The Influence of Motivational Salience on Attention Selection: An ERP Investigation Constanza De
Selective Attention. Modes of Control. Domains of Selection
 The New Yorker (2/7/5) Selective Attention Perception and awareness are necessarily selective (cell phone while driving): attention gates access to awareness Selective attention is deployed via two modes
The New Yorker (2/7/5) Selective Attention Perception and awareness are necessarily selective (cell phone while driving): attention gates access to awareness Selective attention is deployed via two modes
Development of action monitoring through adolescence into adulthood: ERP and source localization
 Developmental Science 10:6 (2007), pp 874 891 DOI: 10.1111/j.1467-7687.2007.00639.x PAPER Blackwell Publishing Ltd Development of action monitoring through adolescence into adulthood: ERP and source localization
Developmental Science 10:6 (2007), pp 874 891 DOI: 10.1111/j.1467-7687.2007.00639.x PAPER Blackwell Publishing Ltd Development of action monitoring through adolescence into adulthood: ERP and source localization