* Corresponding author: Tel.: ; Fax:
|
|
|
- Leon Malone
- 6 years ago
- Views:
Transcription
1 JVI Accepted Manuscript Posted Online 8 February 2017 J. Virol. doi: /jvi Copyright 2017 American Society for Microbiology. All Rights Reserved HIV/AIDS vaccine candidates based on replication-competent recombinant poxvirus NYVAC-C-KC expressing trimeric gp140 and Gagderived VLPs or lacking the viral molecule B19 that inhibits type I interferon activate relevant HIV-1-specific B and T cell immune functions in non-human primates Juan García-Arriaza a, Beatriz Perdiguero a, Jonathan L. Heeney b, Michael S. Seaman c, David C. Montefiori d, Nicole L. Yates d, Georgia D. Tomaras d, Guido Ferrari d, Kathryn E. Foulds e, Mario Roederer e, Steven G. Self f, Bhavesh Borate f Raphael Gottardo f, Sanjay Phogat g, Jim Tartaglia g, Susan W. Barnett h, Brian Burke h, Anthony D. Cristillo i, Deborah E. Weiss i, Carter Lee j, Karen V. Kibler k, Bertram L. Jacobs k, Ralf Wagner l, Song Ding m, Giuseppe Pantaleo n, and Mariano Esteban a * a Department of Molecular and Cellular Biology, Centro Nacional de Biotecnología, Consejo Superior de Investigaciones Científicas (CSIC), Madrid, Spain. b Department of Veterinary Medicine, University of Cambridge, Cambridge, United Kingdom. c Center for Virology and Vaccine Research, Beth Israel Deaconess Medical Center, Boston, Massachusetts, USA. d Duke University Medical Center, Durham, North Carolina, USA. e Vaccine Research Center, National Institute of Allergy and Infectious Diseases (NIAID), National Institutes of Health (NIH), Bethesda, Maryland, USA. f Statistical Center for HIV/AIDS Research and Prevention, Fred Hutchinson Cancer Research Center, Seattle, Washington, USA. g Sanofi Pasteur, Swiftwater, Pennsylvania, USA. h Novartis Vaccines and Diagnostics, Inc., Cambridge, Massachusetts, USA. i Advanced BioScience Laboratories, Inc., Rockville, Maryland, USA. j Global Solutions on Infectious Diseases, South San Francisco, California, USA. k Biodesign Institute, Arizona State University, Tempe, Arizona, USA. l University of Regensburg, Regensburg, Germany. m EuroVacc Foundation, Lausanne, Switzerland. n Division of Immunology and Allergy, Department of Medicine, Centre Hospitalier Universitaire Vaudois, University of Lausanne, Lausanne, Switzerland. 1
2 * Corresponding author: Tel.: ; Fax: mesteban@cnb.csic.es Running title: HIV immunogenicity in macaques of replicating NYVACs Key words: HIV-1, poxvirus, NYVAC, B19R, non-human primates, immunogenicity, cellular responses, humoral responses. Abstract word count: 250 Manuscript word count:
3 ABSTRACT The non-replicating attenuated poxvirus vector NYVAC expressing clade C(CN54) HIV- 1 Env(gp120), Gag-Pol-Nef antigens (NYVAC-C) showed in phase I clinical trials limited immunogenicity. To enhance the capacity of the NYVAC vector to trigger broad humoral responses and a more balanced activation of CD4 + and CD8 + T cells, here we compared the HIV-1-specific immunogenicity elicited in non-human primates immunized with two replicating NYVAC vectors that have been modified by the insertion of K1L and C7L vaccinia viral host-range genes and express clade C(ZM96) trimeric HIV-1 gp140 protein or a Gag(ZM96)-Pol-Nef(CN54) polyprotein as Gagderived virus-like particles (termed NYVAC-C-KC). Additionally, one NYVAC-C-KC vector was generated by deleting the viral gene B19R, an inhibitor of type I interferon response (NYVAC-C-KC-ΔB19R). An immunization protocol mimicking the RV144 phase III clinical trial was used. Two groups of macaques received two doses of the corresponding NYVAC-C-KC vectors (weeks 0 and 4), and booster doses with NYVAC-C-KC vectors plus clade C HIV-1 gp120 protein (weeks 12 and 24). The two replicating NYVAC-C-KC vectors induced an enhanced and similar HIV-1-specific CD4 + and CD8 + T cell responses, similar levels of binding IgG antibodies, low levels of IgA antibodies, high levels of antibody-dependent cellular cytotoxicity responses and HIV-1-neutralizing antibodies. Small differences within the NYVAC-C-KC-ΔB19R group were seen in the magnitude of CD4 + and CD8 + T cells, induction of some cytokines and in the neutralization of some HIV-1 isolates. Thus, replication-competent NYVAC-C-KC vectors acquired relevant immunological properties as vaccine candidates against HIV/AIDS, and the viral B19 molecule exerts some control of immune functions. 3
4 IMPORTANCE It is of special importance to find a safe and effective HIV/AIDS vaccine that can induce strong and broad T cell and humoral immune responses correlating with HIV-1 protection. Here we developed novel replicating poxvirus NYVAC-based HIV/AIDS vaccine candidates expressing clade C HIV-1 antigens, with one of them lacking the vaccinia B19 protein, an inhibitor of type I interferon response. Immunization of nonhuman primates with these novel NYVAC-C-KC vectors and the protein component gp120 elicited a high level of T cell and humoral immune responses, with the vector containing the deletion in B19R inducing a trend to higher magnitude of CD4 + and CD8 + T cells and neutralization of some HIV-1 strains. These poxvirus vectors could be considered as HIV/AIDS vaccine candidates based on their activation of potential immune correlates of protection. INTRODUCTION According to the UNAIDS, about 36.7 million of adults and children were living with HIV worldwide at the end of 2015; however, the number of people newly infected continues to fall, being in % lower than in 2001 ( These data are the result of the global implementation of preventive and therapeutic strategies. Nevertheless, the development of a vaccine remains among the best hopes for controlling the HIV/AIDS pandemic. To date, the phase III clinical trial RV144 is the only HIV-1 vaccine efficacy trial that has demonstrated a modest level of protection (31.2%) against HIV-1 infection in humans (1). The RV144 study combined a recombinant canarypox virus vector (ALVAC) expressing HIV-1 antigens from clades B and E as a prime with recombinant HIV-1 gp120 proteins from clades B and CRF01_AE as a boost. Further studies evaluating potential immune correlates of protection have shown that CD4 + T cells, IgG antibodies to the V1/V2 and V3 loops of HIV-1 gp120, IgG3 antibodies to gp120, together with ADCC responses, correlated with decreased risk of HIV-1 infection, whereas IgA antibodies to the envelope correlated with a decreased vaccine efficacy in the vaccine group (2-7). These clinical findings provided for the first time evidence that an HIV/AIDS vaccine can prevent HIV-1 infection and highlighted that poxvirus vectors should be considered as one of the future HIV/AIDS vaccine candidate vectors. Among the poxviruses, the highly attenuated vaccinia virus (VACV) strain NYVAC has emerged as a potential HIV/AIDS vaccine candidate (8, 9). Importantly, several NYVAC-based recombinants expressing HIV-1 antigens from clades B or C have been evaluated as HIV/AIDS vaccine candidates in preclinical (10-28) and clinical (20, 29-34) studies with encouraging results of HIV-1-specific T cell and humoral 4
5 immune responses. However, new strategies have been implemented to improve poxvirus vector immunogenicity (35). Among these strategies, we have previously reported that deletion of immunomodulatory VACV genes such as B19R and/or B8R, that block interferon (IFN) type I and type II pathways (17) or insertion of the host range VACV C7L gene into NYVAC recombinant vectors expressing clade C (CN54) HIV-1 Env (gp120), and Gag-Pol-Nef antigens as a polyprotein (NYVAC-C) significantly improved the magnitude and quality of the HIV-1-specific immune responses in mice (23). Furthermore, deletion of B19R and/or B8R genes in NYVAC-C triggered an upregulation of innate immune pathways in infected human monocytes with a robust expression of type I IFNs and IFN-stimulated genes (ISGs), a strong activation of the inflammasome, and upregulation of the expression of IL-1β and proinflammatory cytokines (12). Moreover, restoration of replication competence in human cells of NYVAC-C by re-incorporation of the K1L and C7L VACV host range genes (NYVAC-C- KC) with or without removal of the immunomodulatory viral molecule B19, enhanced cross-presentation and proliferation of HIV-1-specific memory CD8 + T cells in vitro (26). These recombinants selectively activated IFN-induced genes and genes involved in antigen processing and presentation as determined by microarray analysis of infected human dendritic cells (DCs) (19, 26). At the same time, these constructs maintained limited virus spread in tissues and an attenuated phenotype (26). Additionally, further improved NYVAC recombinants expressing HIV-1 immunogens, such as HIV-1 clade C (ZM96) trimeric soluble gp140 or Gag(ZM96)-Pol-Nef(CN54) as Gag-derived VLPs have been shown an enhanced HIV-1-specific immunogenicity profile in mice (24) and non-human primates (NHPs) (10, 13). Clinical trials with homologous NYVAC vectors expressing HIV-1 antigens (gp120 Env, and the polyprotein Gag-Pol-Nef) have shown a limited immunogenicity profile with preference for CD4 + T cell activation, markedly enhanced when priming was performed with a DNA vector expressing the same HIV-1 antigens (29-33). Thus, in order to optimize the immunization protocol with NYVAC vectors expressing HIV-1 antigens, various approaches have been evaluated in NHPs,either comparing NYVAC versus ALVAC (13), or combining NYVAC with DNA vectors (10), peptides (22) and with dendritic cell targets (28), and have all demonstrated promising results. Here, as part of the Poxvirus T cell Vaccine Discovery Consortium (PTVDC) from the Collaboration for AIDS Vaccine Discovery (CAVD) of Bill and Melinda Gates Foundation, we extended our previous studies with NYVAC recombinants (19, 26) and evaluated in NHP novel NYVAC recombinants. Hence, we combined in a single recombinant NYVAC vector a set of strategies: restoration of replication competence, expression of novel HIV-1 immunogens (trimeric gp140 and Gag-Pol-Nef as Gag- 5
6 derived VLPs), and deletion of B19R immunomodulatory gene (NYVAC recombinants termed NYVAC-C-KC and NYVAC-C-KC-ΔB19R). Thus, NYVAC-C-KC and NYVAC-C- KC-ΔB19R were compared in immunized NHP to evaluate the HIV-1-specific immunogenicity profile induced by these novel NYVAC recombinants, when applied in a prime/boost approach following similar delivery of the immunogens, poxvirus and protein, as for the protocol used in the RV144 phase III clinical trial. The aim was to define the type of HIV-1-specific T cell and humoral immune responses induced by these vectors as a function of immunological markers that have been correlated with HIV-1 immune efficacy. The results showed that replicating NYVAC-C-KC vectors together with booster of the purified gp120 protein component induced enhanced activation of HIV-1-specific CD4 + and CD8 + T cell immune responses, together with a strong induction of HIV-1-specific humoral immune responses. These results demonstrate that replicating NYVAC-C-KC vectors triggered relevant HIV-1-specific immunological properties as potential correlates of protection, with VACV B19 protein exerting some control of immune functions, and supporting the use of these novel NYVAC-C-KC recombinants as HIV/AIDS vaccine candidates. 6
7 MATERIALS AND METHODS Replication-competent recombinants NYVAC-C-KC and NYVAC-C-KC-ΔB19R expressing clade C HIV-1 antigens. VACV K1L and C7L genes were inserted into poxvirus recombinants NYVAC-gp140 and NYVAC-Gag-Pol-Nef, previously described in (24), to generate respectively the replication-competent recombinants NYVAC-KCgp140 and NYVAC-KC-Gag-Pol-Nef, following the same methodology previously described (19). Thus, the replication-competent NYVAC-C-KC vector consists of two NYVAC-KC viruses that express different clade C HIV-1 antigens under the same synthetic early/late poxvirus promoter (36), one (NYVAC-KC-gp140) expressing Env gp140 from strain 96ZM651 and the other (NYVAC-KC-Gag-Pol-Nef) expressing Gag from strain 96ZM651 and Pol and Nef from strain CN54. A description of the HIV-1 gp140 and Gag-Pol-Nef sequences included in the NYVAC-KC recombinants was previously reported (10, 24). Next, the VACV B19R gene (B18R in WR strain) was deleted from the replication-competent recombinants NYVAC-KC-gp140 and NYVAC- KC-Gag-Pol-Nef, to generate respectively the replication-competent recombinants NYVAC-KC-gp140-ΔB19R and NYVAC-KC-Gag-Pol-Nef-ΔB19R, following the same methodology previously described (19). Once generated, the resulting replicationcompetent NYVAC-KC-gp140, NYVAC-KC-Gag-Pol-Nef, NYVAC-KC-gp140-ΔB19R and NYVAC-KC-Gag-Pol-Nef-ΔB19R recombinant viruses were expanded in large cultures of primary chicken embryo fibroblast (CEF) cells followed by virus purification through two 36% (wt/vol) sucrose cushions. Titers were determined by plaque immunostaining in BSC-40 cells. Expression of HIV-1 antigens by Western blotting and analysis of plaque size were done as previously described (24). Infection of human macrophages (THP-1 cells) and RNA analysis of IFN-β, MIP-1α, IL-8, and IL-1β by quantitative real-time PCR was done as previously described (37, 38). For simplicity of terminology, we refer to the combined mixed inoculation of NYVAC-KC-gp140 plus NYVAC-KC-Gag-Pol-Nef as NYVAC-C-KC and that of NYVAC-KC-gp140-ΔB19R plus NYVAC-KC-Gag-Pol-Nef-ΔB19R as NYVAC-C-KC-ΔB19R. HIV-1 proteins. For the immunizations performed in this study we have used a bivalent HIV-1 gp120 protein containing a mixture of TV1 gp120 and 1086 gp120, both from clade C. These proteins were expressed from stably transfected Chinese hamster ovary (CHO) cell lines, purified and characterized as previously described (39). Non-human primates. Animals used in this study (designated AUP513) were outbred adult male Indian rhesus macaques (Macaca mulatta) which were housed, fed, given environmental enrichment, and handled at the animal facility of Advanced BioScience 7
8 Laboratories (ABL), Inc. (Rockville, MD). The study protocol and animal care are in accordance with the standards of the Association for the Assessment and Accreditation of Laboratory Animal Care International (AAALAC International). The study was approved by the ABL, Inc., Institutional Animal Care and Use Committee in accordance with international guidelines. All procedures were carried out under anesthesia (ketamine administered at 10 mg/kg of body weight) by trained personnel under the supervision of veterinary staff, and all efforts were made to ameliorate animal welfare and to minimize animal suffering. During the study, macaques were observed for general behavior, clinical symptoms, and local reactions at the injection sites twice daily the week after immunizations. When animals were sedated for immunizations or sample collections, body weight and temperature were measured. The age of the animals ranged between 2.5 and 2.8 years, with a mean of 2.6 years and the weight range was between 3.1 to 5.7 kg, with a mean of 3.8 kg. At selected time points, a physical examination was performed by a veterinarian, and clinical chemistry and hematology parameters were measured. All rhesus macaques were negative for tuberculosis, simian retrovirus (SRV), simian T cell leukemia virus (STLV-1), herpesvirus B, simian immunodeficiency virus (SIV), measles and poxvirus immunogens prior to the study, and have also negative fecal culture for salmonella, shigella, campylobacter and yersinia. Furthermore, animals were immunologically naïve for the vaccine components. A total of 16 animals were divided into two groups of eight animals each. Vaccines and immunization schedule. Two immunizations groups of eight rhesus macaques were included in the study protocol (termed AUP513), which was also previously reported (13). Group 1 received two immunizations with NYVAC-C-KC (containing a 1:1 mixture of NYVAC-KC-gp140 and NYVAC-KC-Gag-Pol-Nef) at weeks 0 and 4 and was boosted with two immunizations of NYVAC-C-KC plus bivalent HIV-1 gp120 proteins from clade C (containing a 1:1 mixture of TV1 gp120 and 1086 gp120) at weeks 12 and 24. Group 2 received two immunizations with NYVAC-C-KC-ΔB19R (containing a 1:1 mixture of NYVAC-KC-gp140-ΔB19R and NYVAC-KC-Gag-Pol-Nef- ΔB19R) at weeks 0 and 4 and was boosted with two immunizations of NYVAC-C-KC- ΔB19R plus bivalent HIV-1 gp120 proteins from clade C (TV1 and 1086 gp120) at weeks 12 and 24. All immunizations with the replication-competent NYVAC-C-KC vectors and proteins were given intramuscularly (i.m.) in the deltoid muscle of the upper right arm for the replication-competent NYVAC-C-KC vectors and in the opposite site, the upper left arm, for the gp120 proteins. A dose of 1 x 10 8 PFU of each replication-competent NYVAC-C-KC vector (NYVAC-KC-gp140 plus NYVAC-KC-Gag- 8
9 Pol-Nef in the case of NYVAC-C-KC, and NYVAC-KC-gp140-ΔB19R plus NYVAC-KC- Gag-Pol-Nef-ΔB19R for NYVAC-C-KC-ΔB19R; 2 x 10 8 PFU of total virus in 1.0 ml of Tris-buffered saline) and 50 µg of each of the TV1 and 1086 clade C HIV-1 gp120 proteins (with adjuvant MF59; 100 µg of total protein in 1.0 ml) was used for each immunization. At weeks 0, 6, 14, 26 and 36 (at the beginning of the study, two weeks after the second, third, and fourth immunizations, and at the end of the study, respectively), peripheral blood mononuclear cells (PBMCs) and serum samples were obtained from each immunized animal, and HIV-1-specific T cell and humoral immune responses were analyzed. Blood samples were processed following current procedures (40). Peptides. Overlapping peptides (15-mers with 11 amino acids overlapping) spanning the Env, Gag, Pol, and Nef HIV-1 clade C regions were matched to the inserts expressed by NYVAC-C-KC and NYVAC-C-KC-ΔB19R. Peptides used in the IFN-y ELISpot and intracellular cytokine staining (ICS) assays for T cell stimulation were grouped in nine different peptide pools (Env-1, Env-2, Env-3, Pol-1, Pol-2, Gag-1, Gag/Pol, Gag-2/Pol and Nef), with about 60 peptides per pool. IFN-γ ELISpot assay. The overall strength of HIV-1-specific T cell immune responses induced was analyzed by IFN-γ ELISpot analysis from freshly isolated PBMCs obtained from each immunized rhesus macaque, as previously described (10). Briefly, PBMCs were stimulated in triplicate with HIV-1 Env, Gag, Pol, and Nef peptide pools at 1 µg/ml, or with phytohemagglutinin (PHA) (2.5 µg/ml) as a positive control, while addition of medium only served as a negative control, following the protocol described previously (10). The spot forming units (SFUs) were counted as a measure of the magnitude of HIV-1-specific T cells. ICS assay. The HIV-1-specific CD4 + and CD8 + T cell immune responses induced were analyzed by polychromatic ICS from PBMCs obtained from each immunized rhesus macaque, as previously described (13, 40). In short, cryopreserved PBMCs were thawed and rested overnight in R10 medium (RPMI 1640 [BioWhittaker, Walkersville, MD], 10% FBS, 2 mm L-glutamine, 100 U/ml penicillin G, 100 μg/ml streptomycin) with 50 U/ml Benzonase (Novagen, Madison, WI) in a 37ºC, 5% CO 2 incubator. Next day, 1 x 10 6 to 3 x 10 6 cells were stimulated in 96-well plates with the corresponding HIV-1 Env, Gag, Pol, and Nef peptide pools (2 μg/ml) in the presence of brefeldin A (10 μg/ml; BD Biosciences, San Jose, CA) for 6 h. Negative controls received an equal concentration of dimethyl sulfoxide (DMSO) instead of peptides. For staining, cells 9
10 were fixed, permeabilized, and stained with different antibodies. The following fluorescence-labeled antibodies were used: CD3-Cy7-APC (clone SP34.2; BD Biosciences), CD4-BV421 (clone OKT4; BioLegend), CD8-BV570 (clone RPA-T8; BioLegend), IFN-γ-APC (clone B27; BD Biosciences), IL-2-PE (clone MQ1-17H12; BD Biosciences) and TNF-α-FITC (clone Mab11; BD Biosciences). An Aqua LIVE/DEAD kit (Invitrogen, Carlsbad, CA) was used to exclude dead cells. All antibodies were previously titrated to determine the optimal concentration. Samples were acquired on an LSR II flow cytometer and analyzed using FlowJo version 9.8 (Treestar, Inc., Ashland, OR). HIV-1-specific binding antibody assay. HIV-1-specific binding IgG and IgA antibodies to HIV-1 gp120/gp140 proteins, and murine leukemia virus (MuLV) gp70- scaffolded V1/V2 were measured using a binding antibody multiplex assay (BAMA) in serum and rectal weck elutions from each immunized rhesus macaque, as previously described (6, 7, 41). HIV-1 antigens used to analyze the levels of IgG or IgA binding antibodies included: (i) consensus gp140 proteins of clade A (A1.con.env CF), clade B (B.con.env CF), clade C (C.con.env CF); and group M (Con S gp140 CF), (ii) primary Env variants 1086 gp120 (clade C), TV1 gp120 (clade C), JRFL gp140 (clade B), and MSA4076 gp140 (clade A1; also known as OOMSA); and (iii) MuLV gp70-scaffolded V1/V2 (from clade B; gp70_b.casea2 V1/V2). Different plasma serial dilutions were made, and the primary readout was the mean fluorescence intensity (MFI). For IgA, results are expressed as the MFI minus the background MFI at dilution 1/80. For IgG, MFI data were transformed to values defining the area under the curve (AUC), which resemble the integral of the curve for MFI plotted against serial dilutions. Moreover, the specific activity of rectal mucosal IgG binding antibodies was calculated by dividing the antibody titer by the total IgG concentration. Values 3-fold above the baseline visit were considered positive, and the cutoff was established using serum-negative samples. All assays were performed under Good Clinical Laboratory Practices (GCLP)-compliant conditions. Antibody-dependent cellular cytotoxicity (ADCC) assay. The ADCC activity against recombinant gp120 proteins from isolates 1086 or TV1 (clade C) was analyzed in serum from each immunized rhesus macaque according to the ADCC-GranToxiLux (GTL) procedure, as previously described (42, 43). The results of the GTL assay were considered positive if the percentage of Granzyme B activity after background subtraction was 8% for the infected target cells (44). The log 10 titer of the ADCC antibodies present in the plasma was calculated by interpolating the log reciprocal of 10
11 the last plasma dilution that yielded positive Granzyme B activity ( 8%). The GTL- ADCC assay was performed under GCLP-compliant guidelines. Neutralizing antibodies against HIV-1. Neutralizing antibodies (NAb) against HIV-1 were analyzed in serum from each immunized rhesus macaque using the TZM-bl cell assay and the more sensitive A3R5 cell assay, as previously described (45-48). For the TZM-bl assay pseudoviruses carrying the following Envs were used: BaL.26 (tier 1B, clade B), Ce1086 (tier 2, clade C), Ce1176 (tier 2, clade C), Ce2010 (tier 2, clade C), Du151.2 (tier 2, clade C), MN.3 (tier 1A, clade B), MW (tier 1A, clade C), SF162.LS (tier 1A, clade B), TH023.6 (tier 1 Clade AE), TV1.21 (tier 2, clade C), and MuLV (as a negative control). For the A3R5 cell assay, we used viral infectious molecular clones (IMCs) which encoded luciferase on the genome (and expressed in the cells only upon infection) and carrying the ectodomains of env genes of the following isolates: Ce1086 (tier 2, clade C), Ce1176 (tier 2, clade C), Ce2010 (tier 2, clade C), Du151.2 (tier 2, clade C), and TV1.21 (tier 2, clade C). For both assays, the neutralization titers are represented as the log 10 of the serum dilution at which relative luminescence units (RLU) were reduced by 50% (ID50) compared to virus control wells after subtraction of background RLU in cell control wells. Both assays were done under GCLP-compliant conditions. Statistical procedures. The Wilcoxon rank sum test was used to compare differences between groups. P values of <0.05 were considered significant. All values used for analyzing proportionate representation of responses were background subtracted. Box plots were used to summarize the distribution of various immune responses, where the midline of the box indicates the median, the ends of the box denote the 25th and 75th percentiles, and whiskers extended to the extreme data points that are no more than 1.5 times the interquartile range (IQR) or, if no values meet this criterion, to the data extremes. Spearman s rank-based correlations were used to compare association of CD4 + and CD8 + T cell responses with antibody responses from BAMA, GTL ADCC and NAb assays at week 26 between NYVAC-C-KC and NYVAC-C-KC-ΔB19R vectors. Specifically, DMSO-adjusted percent positivity for each cytokine and pooled antigen combination from the CD4 + and CD8 + T cell responses was assessed for correlation with: 1) AUC against gp120.tv1 antigen for the IgG subclass in the BAMA assay, 2) titer against the gp120.tv1 antigen in the ADCC GTL assay, and 3) titer against the MW isolate in the TZM.bl NAb assay. Significance of the correlations was derived using a one-sided p-value with the alternative that true correlation is greater than 0. 11
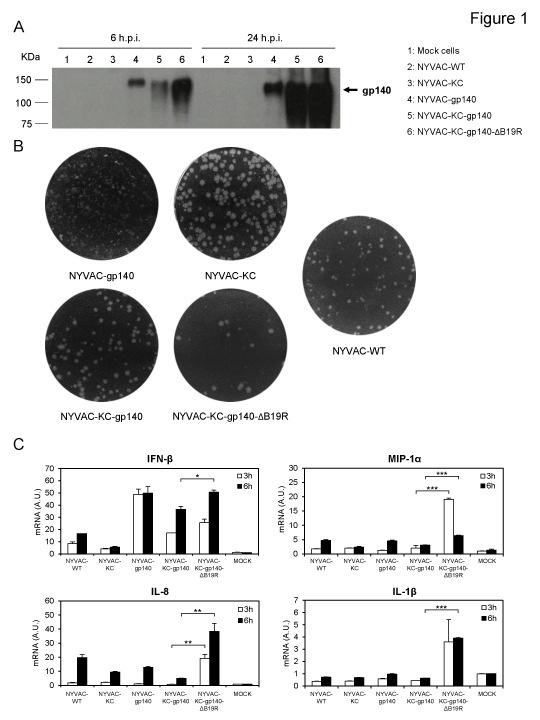
12 RESULTS Enhanced expression, plaque size and innate immune profile of NYVAC-C-KC and NYVAC-C-KC-ΔB19R vectors. We have previously described the generation and characterization of non-replicating NYVAC vectors expressing clade C HIV-1 trimeric gp140 or Gag-Pol-Nef as a polyprotein processed into Gag-derived VLPs and their immune behavior in mice (24) and in NHPs (13). Since these vectors do not replicate in human cells, it was important to define if novel replication-competent NYVAC-KC vectors could be more immunogenic as a function of higher levels of antigen expression during infection. Analysis by Western blot of the expression of HIV-1 gp140 in human HeLa cells is shown in Fig. 1A. Clearly, higher levels of expression at late times post infection were observed in the NYVAC-C-KC vectors as opposed to parental NYVAC-C. These differences were also noticeable after analysis of the virus plaque size phenotype in cultured BSC-40 cells. The NYVAC-C-KC vectors have larger plaque size phenotype, with or without B19R, than parental NYVAC-WT or NYVAC-gp140, consistent with a higher replication capacity of NYVAC-KC vectors over parental NYVAC (Fig. 1B). Analysis by real time PCR of the innate immune response elicited in human macrophages (THP-1) infected with the NYVAC-C-KC vectors, showed that compared to NYVAC-KC-gp140, NYVAC-KC-gp140-ΔB19R triggered a significant upregulation of the mrna levels of type I IFN (IFN-β), MIP-1α, IL-8, and IL-1β (Fig. 1C), indicating some differential innate immune responses between these vaccines. Immunological responses induced in NHP after prime/boost immunization with NYVAC-C-KC or NYVAC-C-KC-ΔB19R vectors. To define the HIV-1-specific immune responses induced by these two novel optimized replication-competent NYVAC vectors expressing HIV-1 clade C immunogens and containing a deletion in VACV B19R gene, and to further evaluate whether these responses could be relevant in the control of HIV-1 replication, we analyzed a number of immune parameters that have been previously described as potential indicators for correlates of protection (2-7). The immunization protocol in NHPs is depicted in Figure 2 and follows a similar immunization scheme as described in the RV144 efficacy clinical trial (1), allowing a head-to-head comparison of the immunogenicity induced by both NYVAC-C-KC vectors. Figure 2A summarizes the 2 immunization groups included in the study (see Materials and Methods for details). Rhesus macaques were divided into two groups of eight animals each: group 1 received in the upper right arm an intramuscular (i.m.) administration of 2x10 8 pfu of NYVAC-C-KC (1:1 mixture of 1x10 8 pfu of NYVAC-KCgp140 and 1x10 8 pfu of NYVAC-KC-Gag-Pol-Nef); group 2 received a similar 12
13 administration protocol but with NYVAC-C-KC-ΔB19R (1:1 mixture of 1x10 8 pfu of NYVAC-KC-gp140-ΔB19R and 1x10 8 pfu of NYVAC-KC-Gag-Pol-Nef-ΔB19R). The first two administrations with the NYVAC-C-KC vectors were given at weeks 0 and 4; thereafter animals received at weeks 12 and 24 i.m booster doses of the corresponding NYVAC-C-KC vectors in the upper right arm, while in the left arm animals received booster immunization with a bivalent clade C gp120 protein (1:1 mixture of 50 µg of 1086 gp120 and 50 µg of TV1 gp120 proteins adjuvanted with MF59). Immunological monitoring of HIV-1-specific T and B cell immune responses induced by both groups was performed from PBMCs, serum or rectal mucosal samples isolated at weeks 0, 6, 14, 26, and 36 (at the beginning of the study, 2 weeks after the second, third, and fourth immunizations, and at the end of the study, respectively), following similar analytical approaches as previously described for NHP (13) (Fig. 2B). HIV-1-specific T cell immune responses. The total magnitude of HIV-1-specific T cell immune responses induced by NYVAC-C-KC and NYVAC-C-KC-ΔB19R vectors was measured at weeks 0, 6, 14, 26, and 36 by IFN-γ ELISpot assay. The results showed that the mean SFU values induced by both immunization groups were low until week 14, and clearly peaked at week 26 (two weeks after the completion of the prime/boost immunization protocol) (Fig. 3A). As expected, the SFU values decline at week 36. There were no significant statistical differences in the levels of IFN-γ positive T cells between the two groups at any timepoint. However, at week 26 of immunization NYVAC-C-KC elicited a trend to higher numbers of SFUs; while at week 36 of immunization NYVAC-C-KC-ΔB19R induced a trend to higher responses. Furthermore, we also analyzed the magnitude of HIV-1-specific T cell immune responses induced by NYVAC-C-KC and NYVAC-C-KC-ΔB19R vectors by ICS assay. Thus, we measured at weeks 6, 14, 26, and 36 the percentage of HIV-1-specific CD4 + and CD8 + T cells secreting IFN-γ, IL-2 and/or TNF-α after stimulation of PBMCs obtained from each immunized rhesus macaque with peptide pools that spanned the HIV-1 Env, Gag, Pol, and Nef antigens present in the inserts. As observed in Fig. 3B and 3C, the total magnitude of HIV-1-specific CD4 + and CD8 + T cell responses peaked at week 26, with a decline at week 36, similar to the results obtained with the IFN-γ ELISpot assay. There were no statistically significant differences in the magnitude of HIV-1-specific CD4 + T cell responses induced by both immunization groups at any timepoint (Fig. 3B). However, at weeks 14 and 36 of immunization NYVAC-C-KC- ΔB19R induced a trend to higher HIV-1-specific CD4 + T cell responses; while at week 26 of immunization NYVAC-C-KC induced a trend to higher responses. On the other 13
14 hand, the analysis of the magnitude of the total HIV-1-specific CD8 + T cell responses (Fig. 3C), showed that immunization with NYVAC-C-KC-ΔB19R induced a trend toward increased levels over NYVAC-C-KC at all time points, but the differences were not significant. Moreover, the analysis of the cytokine responses generated by both immunization groups revealed that in agreement with the total magnitude of HIV-1- specific CD4 + T cell responses, immunization with NYVAC-C-KC-ΔB19R induced at week 36 a trend to higher magnitude of HIV-1-specific CD4 + T cells producing TNF-α (Fig. 4B), or IL-2 (Fig. 4C), but the differences were not significant. However, at week 26 of immunization with NYVAC-C-KC a trend to higher magnitude of HIV-1-specific CD4 + T cells producing IFN-γ (Fig. 4A), TNF-α (Fig. 4B), or IL-2 (Fig. 4C) was observed. In the case of HIV-1-specific CD8 + T cells, and in agreement with the total values of CD8 + T cells in Figure 3C, immunization with NYVAC-C-KC-ΔB19R induced at most of the time points a trend toward higher magnitude of HIV-1-specific CD8 + T cells producing IFN-γ (Fig. 4D), TNF-α (Fig. 4E), or IL-2 (Fig. 4F), although there were no statistical differences between groups. Binding antibody responses. Since the RV144 phase III clinical trial showed that IgG antibodies against the V1/V2 and V3 regions of HIV-1 gp120 correlated with decreased risk of HIV-1 infection (2-7), we next analyzed the HIV-1 specific humoral immune responses elicited after immunization with the two NYVAC-C-KC vectors. Thus, we quantified in individual serum samples obtained from each immunized rhesus macaque the total binding of IgG antibody levels against clade C HIV-1 gp140, gp120, and MuLVgp70-scaffolded V1/V2 proteins at weeks 6, 14, 26, and 36 (Fig. 5). Results showed that there was strong IgG binding antibody levels to the different HIV-1 envelopes tested in all animals when the NYVAC-C-KC vectors were immunized together with the gp120 protein component, although there were no statistical significant differences between the two groups. In these immunization groups, the maximum IgG antibody levels were detected at week 26 (two weeks after the second booster dose with NYVAC-C-KC/gp120 protein), and by week 36 IgG antibody levels declined but still were maintained at levels superior to two doses of NYVAC-C-KC vectors (week 6). Immunization with NYVAC-C-KC elicited a trend to higher levels of binding IgG antibodies against consensus HIV-1 gp140 protein of group M (Fig. 5A), gp120 from isolate TV1 (Fig. 5B), gp120 from isolate 1086 (Fig. 5C), and the MuLV gp70-scaffolded V1/V2 protein (Fig. 5D), although there were no statistical differences between groups. 14
15 Similarly to the results in Figure 5, cross-clade binding IgG antibody levels induced by both immunization groups were low by week 6 but increased markedly by week 26, and waned with time at week 36 but remained to levels similar or higher to week 6 (data not shown). Overall, both immunization groups induced similar high levels of cross-clade binding IgG antibodies against consensus HIV-1 gp140 of clade A, clade B, clade A gp140 (00MSA) and clade B gp140 (JRFL), with no statistically significant differences between the groups. A trend to higher cross-clade binding IgG antibodies for NYVAC-C-KC was noted at week 14. In addition, rectal IgG binding responses against group M HIV-1 gp140 consensus, and clade C gp120 from isolates 1086 and TV1 were detected in both immunization groups, with a peak of response at week 26, and a decline at week 36 (Fig. 5E), although differences were not significant. The rates of responders at week 26 were higher in the NYVAC-C-KC-ΔB19R immunization group, while at week 36 was higher in the NYVAC-C-KC immunization group. Because the RV144 phase III clinical trial showed that high levels of binding plasma IgA antibodies to HIV-1 Env correlated directly with an increased risk of infection (4, 5), we next analyzed the binding IgA antibodies induced after immunization with the two NYVAC-C-KC vectors in serum and in rectal mucosal (data not shown). The results in serum showed that both immunization groups induced very low levels (barely above the background for most time-points) of binding IgA antibodies against the consensus HIV-1 gp140 protein of clade C, gp120 from isolates TV1, and gp120, and the MuLV gp70-scaffolded protein. Furthermore, similar negligible levels of binding IgA antibodies to consensus HIV-1 gp140 proteins of clade A, B, group M, clade A gp140 (00MSA) and clade B gp140 (JRFL) were observed from the two vaccinated groups. There were no rectal IgA binding antibodies (data not shown). ADCC responses. Since ADCC can mediate non-neutralizing antibody effector functions dependent on Fc receptor engagement, and has been suggested as a correlate of protection in the RV144 phase III clinical trial (4, 49), we next analyzed the ability of NYVAC-C-KC and NYVAC-C-KC-ΔB19R to induce ADCC responses against HIV-1 gp120 proteins from isolates TV1 and 1086 at weeks 0, 26, and 36 in individual serum samples obtained from each immunized rhesus macaque (Fig. 6A). The results showed that both immunization groups elicited similar titers of ADCC at week 26 against HIV-1 gp120 from isolates TV1 and 1086, with a decline at week 36. The titers against target cells coated with the HIV-1 gp120 protein of TV1 isolate were greater than those obtained with HIV-1 gp120 of 1086 isolate. Although there were no statistical differences between plasma samples from the two NYVAC-C-KC vectors, 15
16 immunization with NYVAC-C-KC-ΔB19R induced a trend to higher ADCC titers against HIV-1 gp120 protein of TV1 isolate at week 36 than NYVAC-C-KC. On the other hand, NYVAC-C-KC elicited higher ADCC responses against HIV-1 gp Neutralizing antibody responses. The induction of HIV-1 NAb is of special importance to control HIV-1 (50). Thus, we next analyzed the capacity of NYVAC-C-KC and NYVAC-C-KC-ΔB19R to elicit NAb against a panel of different Tier 1 and 2 HIV-1 viruses at weeks 26 and 36 in individual serum samples obtained from each immunized rhesus macaque (Fig. 6B). The results showed that neutralization titers induced by both immunization groups were higher at week 26 than at week 36, with overall no significant differences between both groups in the neutralization against the different Tier 1 and 2 HIV-1 isolates. However, some minor differences were observed. Using the more sensitive A3R5 assay, results showed that NYVAC-C-KC-ΔB19R induced a trend to higher titers of NAb against HIV-1 gp120 TV1 (Tier 2, clade C), and Ce1086 (Tier 2, clade C), at weeks 26 and 36, although there were no statistical differences compared to NYVAC-C-KC. The TZM.bl assay showed that both immunization groups induced similar levels of NAb against a wide range of Tier 1 HIV- 1 isolates. Correlation between T cell and antibody responses. Next, to define if there is a correlation between the two vectors in the elicitation of CD4/CD8 T cell responses versus the induced antibody responses, we performed a statistical correlation analysis at week 26. Figure 7 shows correlation coefficients between T cell responses (ICS assay from both CD4/CD8) and antibody responses (BAMA, ADCC GTL and NAb assays). In general, CD4 + T cell responses showed a higher number of positive correlations with antibody responses as compared to CD8 + T cell responses. Within CD4 + T cell responses, NYVAC-C-KC-ΔB19R showed a more favorable trend for a correlation with antibody responses as compared to NYVAC-C-KC. Most of the correlations fail to be significant (< 0.05) due to low sample size, except between CD4 + and ADCC and NAb within the NYVAC-C-KC-ΔB19R group. DISCUSSION After the modest efficacy obtained in the RV144 phase III clinical trial the scientific community has focused on the generation and optimization of HIV/AIDS vaccine candidates to afford protection to a large percentage of vaccinees. Poxviruses, and in particular the highly attenuated VACV strains MVA and NYVAC, have being widely used as HIV/AIDS vaccine candidates and are included as 16
17 components of some of the clinical trials planned within the next 5 years on the basis of lessons learned from recent trials (8, 9, 51). However, despite the safety and immunogenicity profiles exhibited by these attenuated VACV strains, more efficient vectors that enhance the magnitude, breadth, polyfunctionality, and durability of the immune responses to exogenously expressed antigens are desirable. Different strategies have been followed for poxviruses to achieve that purpose (35). One of them is the deletion of VACV immunomodulatory genes, that are still present in the vector genome, and whose gene products may be predicted to interfere with the optimal induction of cellular and humoral responses. In this sense, we have previously reported enhanced immunogenicity in mice with MVA and NYVAC-based recombinants by single or multiple deletions of VACV immunomodulatory genes such as B8R and/or B19R (17), A46R (25), C12L (52), C6L and/or K7R (37, 38), A41L and/or B16R (53), N2L (54) and F1L (55). Another strategy is the generation of new vectors with replication competence in human cells in order to increase the timing and levels of expression of the heterologous antigen in the host. To date, some replicationcompetent recombinant VACV-based vaccines have been used for various infectious diseases, demonstrating that they are able to elicit potent humoral and cellular mediated immune responses, and to confer long-lasting protection while maintaining a safety phenotype (23, 56-58). In this study, we aimed to define best-in-class protocols of immunization following lessons learned from the RV144 efficacy clinical trial with poxvirus vectors. We have evaluated and compared the immunogenicity in NHP of two replicationcompetent NYVAC-C recombinants generated by restoration of replication competence in human cells after the re-incorporation of the K1L and C7L host range genes (NYVAC-C-KC) or by combination of restoration of replication competence with removal of the immunomodulatory viral molecule B19 (NYVAC-C-KC-ΔB19R) that blocks type I IFN response. Through extensive analysis of HIV-1 specific immune responses, including the measurement of several immune markers that have been associated in the RV144 phase III clinical trial with immune efficacy (2-7), we observed that both NYVAC-C-KC and NYVAC-C-KC-ΔB19R viruses were able to induce a broad spectrum of HIV-1-specific B and T cell responses against the HIV-1 antigens. Thus, the vaccine-induced T cell immune responses were high in magnitude in both immunization groups, and the deletion of the immunomodulatory gene B19R on NYVAC-C-KC recombinant improved to some extent the magnitude of the HIV-1- specific CD4 + and CD8 + T cell immune responses. Moreover, a low level of IgA binding antibodies, high level of binding IgG antibodies in serum and rectal mucosal samples, ADCC responses and NAb against different tier 1 and 2 HIV-1 isolates were elicited by 17
18 NYVAC-C-KC and NYVAC-C-KC-ΔB19R, with some minor differences between the two groups. Interestingly, at the peak of response (week 26), the higher levels of HIV- 1-specific CD4 + and CD8 + T cells elicited by NYVAC-C-KC and NYVAC-C-KC-ΔB19R coincides with similar high levels of antibodies to Env as well as to high HIV neutralization titers and of ADCC at the same time, suggesting a correlation between enhanced CD4 + /CD8 + T cell responses and B cell responses, pointing that CD4 + and/or CD8 + T cells might influence the B cell response. In fact, a correlation analysis (Fig. 7) showed that percentages of HIV-1-specific CD4 + T cell responses induced by the NYVAC-C-KC-ΔB19R vector have a more favourable trend for a correlation with antibody responses than for NYVAC-C-KC. However, the correlation between CD8 + T cell and B cell responses was less pronounced than for CD4 + T cells. How does the B19R gene operate? It has been shown that B19 is a type I IFNbinding protein that acts both in solution and when bound to glycosaminoglycans to inhibit the action of a wide range of IFNs from different species, sequestering and suppressing efficiently the function of type I IFNs (59, 60). In cells infected with a NYVAC vector lacking B19, we have shown in human DCs enhanced IFN-alpha production, maturation and expression of IFN-induced pathways and IFN-regulated transcription factors as well as multiple inflammatory cytokines (12). Immunization of mice with NYVAC lacking B19R and expressing Env, Gag, Pol, and Nef clade C (CN54) HIV-1 antigens, improved the magnitude and quality of HIV-1-specific CD8 + T cell adaptive immune responses and impacted their memory phase, changing the contraction, the memory differentiation and the magnitude and functionality profile, while for B cell responses B19 had no apparent effect on antibody levels (17). The immunomodulatory effects of B19 were more pronounced in mice than in NHP. Here in NHP we noted at some time points after boost immunization the influence of B19 by a trend to slightly higher levels of T cells and of some cytokines triggered by NYVAC-C- KC-ΔB19R over NYVAC-C-KC. The overall effect of B19 on Env-specific binding IgG antibodies, IgA, NAb and of ADCC was not different between the NYVAC-C-KC vectors. Thus, it seems that B19 main effect is on T cell responses, and the role of B19R blocking type I IFN could explain the small differences in immunogenicity, in particular on T cell responses, observed in macaques inoculated with NYVAC-C-KC versus NYVAC-C-KC-ΔB19R. In our previous study in NHP we demonstrated that non-replicating NYVAC vector expressing the same HIV-1 antigens and the same protocol as in this current study was superior to the ALVAC vector used in the RV144 phase III clinical trial (13). Comparison of the immunogenicity profiles triggered by the NYVAC-C-KC vectors (this 18
19 study) with non-replicating NYVAC vector (13) using the same standardized immune assays revealed an overall higher magnitude of HIV-1-specific T cell and humoral immune responses induced by the NYVAC-C-KC vectors. This is likely due to more antigen expression from the NYVAC-C-KC vectors (Fig. 1), as the virus replicates more efficiently in NHP than NYVAC lacking the VACV host range genes K1L and C7L. These results correlated with our previous in vitro findings, which demonstrated that NYVAC-KC vectors acquired specific biological characteristics over non-replicating NYVAC, giving replication-competency in human keratinocytes and dermal fibroblasts, activation of selective host cell signal transduction pathways, and higher virus spread in tissues while maintaining a highly attenuated phenotype (19, 26). While antigen exposure has been defined recently as a determinant of HIV-1 broadly neutralizing antibody (bnab) induction (61), the question remains as to how best to induce bnabs by vaccination approaches. Using the protocols described here we have been able to trigger a broad spectrum of T and B cell immune responses. A balanced of T and B cell immune responses has been correlated with levels of protection by HIV-vaccines in NHP models of HIV infection (62). A number of HIV/SIV vaccine protection studies in NHPs have revealed a role for humoral responses as potential correlates of immunity (63-65), broadly including the types of immune responses elicited by this vaccine regimen. However, it remains to be defined whether the responses elicited by NYVAC-C-KC vectors confer in NHP protection against a SHIV virus challenge and/or are able to induce bnabs or other potentially protective immune responses in a prophylactic phase I clinical trial. The high immunological profiles induced by the NYVAC-C-KC vectors, including induction of CD4 + and CD8 + T cell immune responses, high levels of anti-env binding antibodies to both gp140 and V1/V2 sites, enhanced ADCC responses and neutralizing capacity, together with low levels of IgA antibodies, are all potential correlates of protection, suggesting that these viral vectors could be considered as improved HIV/AIDS vaccine candidates. ACKNOWLEDGMENTS This investigation was supported by the PTVDC/CAVD Program with support from the Bill and Melinda Gates Foundation (BMGF). Humoral immune monitoring data were supported by the BMGF CAVIMC grant and the NIH/NIAID Duke Center for AIDS Research (CFAR; 5P30 AI064518). Novartis Vaccines received support for this work under contract number HHSN C from DAIDS, NIAID, NIH. We thank Dr. Marcella Sarzotti-Kelsoe for quality assurance oversight, William T. Williams, Robert Howington, R. Glenn Overman, Cristina Sánchez and Victoria 19
20 Jiménez for technical assistance, Sheetal Sawant for BAMA data management and Hua-Xin Liao and Bart Haynes for envelope and V1/V2 protein reagents. 20
21 REFERENCES 1. Rerks-Ngarm S, Pitisuttithum P, Nitayaphan S, Kaewkungwal J, Chiu J, Paris R, Premsri N, Namwat C, de Souza M, Adams E, Benenson M, Gurunathan S, Tartaglia J, McNeil JG, Francis DP, Stablein D, Birx DL, Chunsuttiwat S, Khamboonruang C, Thongcharoen P, Robb ML, Michael NL, Kunasol P, Kim JH Vaccination with ALVAC and AIDSVAX to prevent HIV-1 infection in Thailand. N Engl J Med 361: Chung AW, Ghebremichael M, Robinson H, Brown E, Choi I, Lane S, Dugast AS, Schoen MK, Rolland M, Suscovich TJ, Mahan AE, Liao L, Streeck H, Andrews C, Rerks-Ngarm S, Nitayaphan S, de Souza MS, Kaewkungwal J, Pitisuttithum P, Francis D, Michael NL, Kim JH, Bailey- Kellogg C, Ackerman ME, Alter G Polyfunctional Fc-effector profiles mediated by IgG subclass selection distinguish RV144 and VAX003 vaccines. Sci Transl Med 6:228ra Gottardo R, Bailer RT, Korber BT, Gnanakaran S, Phillips J, Shen X, Tomaras GD, Turk E, Imholte G, Eckler L, Wenschuh H, Zerweck J, Greene K, Gao H, Berman PW, Francis D, Sinangil F, Lee C, Nitayaphan S, Rerks- Ngarm S, Kaewkungwal J, Pitisuttithum P, Tartaglia J, Robb ML, Michael NL, Kim JH, Zolla-Pazner S, Haynes BF, Mascola JR, Self S, Gilbert P, Montefiori DC Plasma IgG to linear epitopes in the V2 and V3 regions of HIV-1 gp120 correlate with a reduced risk of infection in the RV144 vaccine efficacy trial. PLoS One 8:e Haynes BF, Gilbert PB, McElrath MJ, Zolla-Pazner S, Tomaras GD, Alam SM, Evans DT, Montefiori DC, Karnasuta C, Sutthent R, Liao HX, DeVico AL, Lewis GK, Williams C, Pinter A, Fong Y, Janes H, DeCamp A, Huang Y, Rao M, Billings E, Karasavvas N, Robb ML, Ngauy V, de Souza MS, Paris R, Ferrari G, Bailer RT, Soderberg KA, Andrews C, Berman PW, Frahm N, De Rosa SC, Alpert MD, Yates NL, Shen X, Koup RA, Pitisuttithum P, Kaewkungwal J, Nitayaphan S, Rerks-Ngarm S, Michael NL, Kim JH Immune-correlates analysis of an HIV-1 vaccine efficacy trial. N Engl J Med 366: Tomaras GD, Ferrari G, Shen X, Alam SM, Liao HX, Pollara J, Bonsignori M, Moody MA, Fong Y, Chen X, Poling B, Nicholson CO, Zhang R, Lu X, Parks R, Kaewkungwal J, Nitayaphan S, Pitisuttithum P, Rerks-Ngarm S, Gilbert PB, Kim JH, Michael NL, Montefiori DC, Haynes BF Vaccineinduced plasma IgA specific for the C1 region of the HIV-1 envelope blocks binding and effector function of IgG. Proc Natl Acad Sci U S A 110: Yates NL, Liao HX, Fong Y, decamp A, Vandergrift NA, Williams WT, Alam SM, Ferrari G, Yang ZY, Seaton KE, Berman PW, Alpert MD, Evans DT, O'Connell RJ, Francis D, Sinangil F, Lee C, Nitayaphan S, Rerks-Ngarm S, Kaewkungwal J, Pitisuttithum P, Tartaglia J, Pinter A, Zolla-Pazner S, Gilbert PB, Nabel GJ, Michael NL, Kim JH, Montefiori DC, Haynes BF, Tomaras GD Vaccine-induced Env V1-V2 IgG3 correlates with lower HIV-1 infection risk and declines soon after vaccination. Sci Transl Med 6:228ra Zolla-Pazner S, decamp A, Gilbert PB, Williams C, Yates NL, Williams WT, Howington R, Fong Y, Morris DE, Soderberg KA, Irene C, Reichman C, Pinter A, Parks R, Pitisuttithum P, Kaewkungwal J, Rerks-Ngarm S, Nitayaphan S, Andrews C, O'Connell RJ, Yang ZY, Nabel GJ, Kim JH, Michael NL, Montefiori DC, Liao HX, Haynes BF, Tomaras GD Vaccine-induced IgG antibodies to V1V2 regions of multiple HIV-1 subtypes correlate with decreased risk of HIV-1 infection. PLoS One 9:e
22 Gomez CE, Perdiguero B, Garcia-Arriaza J, Esteban M Poxvirus vectors as HIV/AIDS vaccines in humans. Hum Vaccin Immunother 8: Pantaleo G, Esteban M, Jacobs B, Tartaglia J Poxvirus vector-based HIV vaccines. Curr Opin HIV AIDS 5: Asbach B, Kliche A, Kostler J, Perdiguero B, Esteban M, Jacobs BL, Montefiori DC, LaBranche CC, Yates NL, Tomaras GD, Ferrari G, Foulds KE, Roederer M, Landucci G, Forthal DN, Seaman MS, Hawkins N, Self SG, Sato A, Gottardo R, Phogat S, Tartaglia J, Barnett SW, Burke B, Cristillo AD, Weiss DE, Francis J, Galmin L, Ding S, Heeney JL, Pantaleo G, Wagner R Potential To Streamline Heterologous DNA Prime and NYVAC/Protein Boost HIV Vaccine Regimens in Rhesus Macaques by Employing Improved Antigens. J Virol 90: Corbett M, Bogers WM, Heeney JL, Gerber S, Genin C, Didierlaurent A, Oostermeijer H, Dubbes R, Braskamp G, Lerondel S, Gomez CE, Esteban M, Wagner R, Kondova I, Mooij P, Balla-Jhagjhoorsingh S, Beenhakker N, Koopman G, van der Burg S, Kraehenbuhl JP, Le Pape A Aerosol immunization with NYVAC and MVA vectored vaccines is safe, simple, and immunogenic. Proc Natl Acad Sci U S A 105: Delaloye J, Filali-Mouhim A, Cameron MJ, Haddad EK, Harari A, Goulet JP, Gomez CE, Perdiguero B, Esteban M, Pantaleo G, Roger T, Sekaly RP, Calandra T Interleukin-1- and type I interferon-dependent enhanced immunogenicity of an NYVAC-HIV-1 Env-Gag-Pol-Nef vaccine vector with dual deletions of type I and type II interferon-binding proteins. J Virol 89: Garcia-Arriaza J, Perdiguero B, Heeney J, Seaman M, Montefiori DC, Labranche C, Yates NL, Shen X, Tomaras GD, Ferrari G, Foulds KE, McDermott A, Kao SF, Roederer M, Hawkins N, Self S, Yao J, Farrell P, Phogat S, Tartaglia J, Barnett SW, Burke B, Cristillo A, Weiss D, Lee C, Kibler K, Jacobs B, Asbach B, Wagner R, Ding S, Pantaleo G, Esteban M Head-to-Head Comparison of Poxvirus NYVAC and ALVAC Vectors Expressing Identical HIV-1 Clade C Immunogens in Prime-Boost Combination with Env Protein in Nonhuman Primates. J Virol 89: Gomez CE, Najera JL, Jimenez EP, Jimenez V, Wagner R, Graf M, Frachette MJ, Liljestrom P, Pantaleo G, Esteban M Head-to-head comparison on the immunogenicity of two HIV/AIDS vaccine candidates based on the attenuated poxvirus strains MVA and NYVAC co-expressing in a single locus the HIV-1BX08 gp120 and HIV-1(IIIB) Gag-Pol-Nef proteins of clade B. Vaccine 25: Gomez CE, Najera JL, Jimenez V, Bieler K, Wild J, Kostic L, Heidari S, Chen M, Frachette MJ, Pantaleo G, Wolf H, Liljestrom P, Wagner R, Esteban M Generation and immunogenicity of novel HIV/AIDS vaccine candidates targeting HIV-1 Env/Gag-Pol-Nef antigens of clade C. Vaccine 25: Gomez CE, Najera JL, Sanchez R, Jimenez V, Esteban M Multimeric soluble CD40 ligand (scd40l) efficiently enhances HIV specific cellular immune responses during DNA prime and boost with attenuated poxvirus vectors MVA and NYVAC expressing HIV antigens. Vaccine 27: Gomez CE, Perdiguero B, Najera JL, Sorzano CO, Jimenez V, Gonzalez- Sanz R, Esteban M Removal of vaccinia virus genes that block interferon type I and II pathways improves adaptive and memory responses of the HIV/AIDS vaccine candidate NYVAC-C in mice. J Virol 86: Hulot SL, Korber B, Giorgi EE, Vandergrift N, Saunders KO, Balachandran H, Mach LV, Lifton MA, Pantaleo G, Tartaglia J, Phogat S, Jacobs B, Kibler K, Perdiguero B, Gomez CE, Esteban M, Rosati M, Felber BK, Pavlakis GN, Parks R, Lloyd K, Sutherland L, Scearce R, Letvin NL, Seaman MS, Alam 22
23 SM, Montefiori D, Liao HX, Haynes BF, Santra S Comparison of Immunogenicity in Rhesus Macaques of Transmitted-Founder, HIV-1 Group M Consensus, and Trivalent Mosaic Envelope Vaccines Formulated as a DNA Prime, NYVAC, and Envelope Protein Boost. J Virol 89: Kibler KV, Gomez CE, Perdiguero B, Wong S, Huynh T, Holechek S, Arndt W, Jimenez V, Gonzalez-Sanz R, Denzler K, Haddad EK, Wagner R, Sekaly RP, Tartaglia J, Pantaleo G, Jacobs BL, Esteban M Improved NYVAC-based vaccine vectors. PLoS One 6:e Mooij P, Balla-Jhagjhoorsingh SS, Beenhakker N, van Haaften P, Baak I, Nieuwenhuis IG, Heidari S, Wolf H, Frachette MJ, Bieler K, Sheppard N, Harari A, Bart PA, Liljestrom P, Wagner R, Pantaleo G, Heeney JL Comparison of human and rhesus macaque T-cell responses elicited by boosting with NYVAC encoding human immunodeficiency virus type 1 clade C immunogens. J Virol 83: Mooij P, Balla-Jhagjhoorsingh SS, Koopman G, Beenhakker N, van Haaften P, Baak I, Nieuwenhuis IG, Kondova I, Wagner R, Wolf H, Gomez CE, Najera JL, Jimenez V, Esteban M, Heeney JL Differential CD4+ versus CD8+ T-cell responses elicited by different poxvirus-based human immunodeficiency virus type 1 vaccine candidates provide comparable efficacies in primates. J Virol 82: Mooij P, Koopman G, Drijfhout JW, Nieuwenhuis IG, Beenhakker N, Koestler J, Bogers WM, Wagner R, Esteban M, Pantaleo G, Heeney JL, Jacobs BL, Melief CJ Synthetic long peptide booster immunization in rhesus macaques primed with replication-competent NYVAC-C-KC induces a balanced CD4/CD8 T-cell and antibody response against the conserved regions of HIV-1. J Gen Virol 96: Najera JL, Gomez CE, Garcia-Arriaza J, Sorzano CO, Esteban M Insertion of vaccinia virus C7L host range gene into NYVAC-B genome potentiates immune responses against HIV-1 antigens. PLoS One 5:e Perdiguero B, Gomez CE, Cepeda V, Sanchez-Sampedro L, Garcia-Arriaza J, Mejias-Perez E, Jimenez V, Sanchez C, Sorzano CO, Oliveros JC, Delaloye J, Roger T, Calandra T, Asbach B, Wagner R, Kibler KV, Jacobs BL, Pantaleo G, Esteban M Virological and Immunological Characterization of Novel NYVAC-Based HIV/AIDS Vaccine Candidates Expressing Clade C Trimeric Soluble gp140(zm96) and Gag(ZM96)-Pol- Nef(CN54) as Virus-Like Particles. J Virol 89: Perdiguero B, Gomez CE, Di Pilato M, Sorzano CO, Delaloye J, Roger T, Calandra T, Pantaleo G, Esteban M Deletion of the vaccinia virus gene A46R, encoding for an inhibitor of TLR signalling, is an effective approach to enhance the immunogenicity in mice of the HIV/AIDS vaccine candidate NYVAC-C. PLoS One 8:e Quakkelaar ED, Redeker A, Haddad EK, Harari A, McCaughey SM, Duhen T, Filali-Mouhim A, Goulet JP, Loof NM, Ossendorp F, Perdiguero B, Heinen P, Gomez CE, Kibler KV, Koelle DM, Sekaly RP, Sallusto F, Lanzavecchia A, Pantaleo G, Esteban M, Tartaglia J, Jacobs BL, Melief CJ Improved innate and adaptive immunostimulation by genetically modified HIV-1 protein expressing NYVAC vectors. PLoS One 6:e Wild J, Bieler K, Kostler J, Frachette MJ, Jeffs S, Vieira S, Esteban M, Liljestrom P, Pantaleo G, Wolf H, Wagner R Preclinical evaluation of the immunogenicity of C-type HIV-1-based DNA and NYVAC vaccines in the Balb/C mouse model. Viral Immunol 22: Zurawski G, Zurawski S, Flamar AL, Richert L, Wagner R, Tomaras GD, Montefiori DC, Roederer M, Ferrari G, Lacabaratz C, Bonnabau H, Klucar P, Wang Z, Foulds KE, Kao SF, Yates NL, LaBranche C, Jacobs BL, Kibler K, Asbach B, Kliche A, Salazar A, Reed S, Self S, Gottardo R, Galmin L, 23
24 Weiss D, Cristillo A, Thiebaut R, Pantaleo G, Levy Y Targeting HIV-1 Env gp140 to LOX-1 Elicits Immune Responses in Rhesus Macaques. PLoS One 11:e Bart PA, Goodall R, Barber T, Harari A, Guimaraes-Walker A, Khonkarly M, Sheppard NC, Bangala Y, Frachette MJ, Wagner R, Liljestrom P, Kraehenbuhl JP, Girard M, Goudsmit J, Esteban M, Heeney J, Sattentau Q, McCormack S, Babiker A, Pantaleo G, Weber J EV01: a phase I trial in healthy HIV negative volunteers to evaluate a clade C HIV vaccine, NYVAC- C undertaken by the EuroVacc Consortium. Vaccine 26: Bart PA, Huang Y, Karuna ST, Chappuis S, Gaillard J, Kochar N, Shen X, Allen MA, Ding S, Hural J, Liao HX, Haynes BF, Graham BS, Gilbert PB, McElrath MJ, Montefiori DC, Tomaras GD, Pantaleo G, Frahm N HIVspecific humoral responses benefit from stronger prime in phase Ib clinical trial. J Clin Invest 124: Harari A, Bart PA, Stohr W, Tapia G, Garcia M, Medjitna-Rais E, Burnet S, Cellerai C, Erlwein O, Barber T, Moog C, Liljestrom P, Wagner R, Wolf H, Kraehenbuhl JP, Esteban M, Heeney J, Frachette MJ, Tartaglia J, McCormack S, Babiker A, Weber J, Pantaleo G An HIV-1 clade C DNA prime, NYVAC boost vaccine regimen induces reliable, polyfunctional, and long-lasting T cell responses. J Exp Med 205: Harari A, Rozot V, Cavassini M, Enders FB, Vigano S, Tapia G, Castro E, Burnet S, Lange J, Moog C, Garin D, Costagliola D, Autran B, Pantaleo G, Bart PA NYVAC immunization induces polyfunctional HIV-specific T-cell responses in chronically-infected, ART-treated HIV patients. Eur J Immunol 42: McCormack S, Stohr W, Barber T, Bart PA, Harari A, Moog C, Ciuffreda D, Cellerai C, Cowen M, Gamboni R, Burnet S, Legg K, Brodnicki E, Wolf H, Wagner R, Heeney J, Frachette MJ, Tartaglia J, Babiker A, Pantaleo G, Weber J EV02: a Phase I trial to compare the safety and immunogenicity of HIV DNA-C prime-nyvac-c boost to NYVAC-C alone. Vaccine 26: Perreau M, Welles HC, Harari A, Hall O, Martin R, Maillard M, Dorta G, Bart PA, Kremer EJ, Tartaglia J, Wagner R, Esteban M, Levy Y, Pantaleo G DNA/NYVAC vaccine regimen induces HIV-specific CD4 and CD8 T-cell responses in intestinal mucosa. J Virol 85: Garcia-Arriaza J, Esteban M Enhancing poxvirus vectors vaccine immunogenicity. Hum Vaccin Immunother Chakrabarti S, Sisler JR, Moss B Compact, synthetic, vaccinia virus early/late promoter for protein expression. Biotechniques 23: Garcia-Arriaza J, Arnaez P, Gomez CE, Sorzano CO, Esteban M Improving Adaptive and Memory Immune Responses of an HIV/AIDS Vaccine Candidate MVA-B by Deletion of Vaccinia Virus Genes (C6L and K7R) Blocking Interferon Signaling Pathways. PLoS One 8:e Garcia-Arriaza J, Najera JL, Gomez CE, Tewabe N, Sorzano CO, Calandra T, Roger T, Esteban M A candidate HIV/AIDS vaccine (MVA-B) lacking vaccinia virus gene C6L enhances memory HIV-1-specific T-cell responses. PLoS One 6:e Srivastava IK, Kan E, Sun Y, Sharma VA, Cisto J, Burke B, Lian Y, Hilt S, Biron Z, Hartog K, Stamatatos L, Diaz-Avalos R, Cheng RH, Ulmer JB, Barnett SW Comparative evaluation of trimeric envelope glycoproteins derived from subtype C and B HIV-1 R5 isolates. Virology 372: Foulds KE, Donaldson M, Roederer M OMIP-005: Quality and phenotype of antigen-responsive rhesus macaque T cells. Cytometry A 81:
25 Tomaras GD, Yates NL, Liu P, Qin L, Fouda GG, Chavez LL, Decamp AC, Parks RJ, Ashley VC, Lucas JT, Cohen M, Eron J, Hicks CB, Liao HX, Self SG, Landucci G, Forthal DN, Weinhold KJ, Keele BF, Hahn BH, Greenberg ML, Morris L, Karim SS, Blattner WA, Montefiori DC, Shaw GM, Perelson AS, Haynes BF Initial B-cell responses to transmitted human immunodeficiency virus type 1: virion-binding immunoglobulin M (IgM) and IgG antibodies followed by plasma anti-gp41 antibodies with ineffective control of initial viremia. J Virol 82: Lambotte O, Pollara J, Boufassa F, Moog C, Venet A, Haynes BF, Delfraissy JF, Saez-Cirion A, Ferrari G High antibody-dependent cellular cytotoxicity responses are correlated with strong CD8 T cell viral suppressive activity but not with B57 status in HIV-1 elite controllers. PLoS One 8:e Pollara J, Hart L, Brewer F, Pickeral J, Packard BZ, Hoxie JA, Komoriya A, Ochsenbauer C, Kappes JC, Roederer M, Huang Y, Weinhold KJ, Tomaras GD, Haynes BF, Montefiori DC, Ferrari G High-throughput quantitative analysis of HIV-1 and SIV-specific ADCC-mediating antibody responses. Cytometry A 79: Edmonds TG, Ding H, Yuan X, Wei Q, Smith KS, Conway JA, Wieczorek L, Brown B, Polonis V, West JT, Montefiori DC, Kappes JC, Ochsenbauer C Replication competent molecular clones of HIV-1 expressing Renilla luciferase facilitate the analysis of antibody inhibition in PBMC. Virology 408: McLinden RJ, Labranche CC, Chenine AL, Polonis VR, Eller MA, Wieczorek L, Ochsenbauer C, Kappes JC, Perfetto S, Montefiori DC, Michael NL, Kim JH Detection of HIV-1 neutralizing antibodies in a human CD4(+)/CXCR4(+)/CCR5(+) T-lymphoblastoid cell assay system. PLoS One 8:e Montefiori DC Measuring HIV neutralization in a luciferase reporter gene assay. Methods Mol Biol 485: Sarzotti-Kelsoe M, Daniell X, Todd CA, Bilska M, Martelli A, LaBranche C, Perez LG, Ochsenbauer C, Kappes JC, Rountree W, Denny TN, Montefiori DC Optimization and validation of a neutralizing antibody assay for HIV-1 in A3R5 cells. J Immunol Methods 409: Todd CA, Greene KM, Yu X, Ozaki DA, Gao H, Huang Y, Wang M, Li G, Brown R, Wood B, D'Souza MP, Gilbert P, Montefiori DC, Sarzotti-Kelsoe M Development and implementation of an international proficiency testing program for a neutralizing antibody assay for HIV-1 in TZM-bl cells. J Immunol Methods 375: Bonsignori M, Pollara J, Moody MA, Alpert MD, Chen X, Hwang KK, Gilbert PB, Huang Y, Gurley TC, Kozink DM, Marshall DJ, Whitesides JF, Tsao CY, Kaewkungwal J, Nitayaphan S, Pitisuttithum P, Rerks-Ngarm S, Kim JH, Michael NL, Tomaras GD, Montefiori DC, Lewis GK, DeVico A, Evans DT, Ferrari G, Liao HX, Haynes BF Antibody-dependent cellular cytotoxicity-mediating antibodies from an HIV-1 vaccine efficacy trial target multiple epitopes and preferentially use the VH1 gene family. J Virol 86: Tomaras GD, Haynes BF Advancing Toward HIV-1 Vaccine Efficacy through the Intersections of Immune Correlates. Vaccines (Basel) 2: Excler JL, Tomaras GD, Russell ND Novel directions in HIV-1 vaccines revealed from clinical trials. Curr Opin HIV AIDS 8: Falivene J, Del Medico Zajac MP, Pascutti MF, Rodriguez AM, Maeto C, Perdiguero B, Gomez CE, Esteban M, Calamante G, Gherardi MM Improving the MVA vaccine potential by deleting the viral gene coding for the IL-18 binding protein. PLoS One 7:e
26 Garcia-Arriaza J, Najera JL, Gomez CE, Sorzano CO, Esteban M Immunogenic profiling in mice of a HIV/AIDS vaccine candidate (MVA-B) expressing four HIV-1 antigens and potentiation by specific gene deletions. PLoS One 5:e Garcia-Arriaza J, Gomez CE, Sorzano CO, Esteban M Deletion of the vaccinia virus N2L gene encoding an inhibitor of IRF3 improves the immunogenicity of modified vaccinia virus Ankara expressing HIV-1 antigens. J Virol 88: Perdiguero B, Gomez CE, Najera JL, Sorzano CO, Delaloye J, Gonzalez- Sanz R, Jimenez V, Roger T, Calandra T, Pantaleo G, Esteban M Deletion of the viral anti-apoptotic gene F1L in the HIV/AIDS vaccine candidate MVA-C enhances immune responses against HIV-1 antigens. PLoS One 7:e Dai K, Liu Y, Liu M, Xu J, Huang W, Huang X, Liu L, Wan Y, Hao Y, Shao Y Pathogenicity and immunogenicity of recombinant Tiantan Vaccinia Virus with deleted C12L and A53R genes. Vaccine 26: Huang X, Lu B, Yu W, Fang Q, Liu L, Zhuang K, Shen T, Wang H, Tian P, Zhang L, Chen Z A novel replication-competent vaccinia vector MVTT is superior to MVA for inducing high levels of neutralizing antibody via mucosal vaccination. PLoS One 4:e Vijaysri S, Jentarra G, Heck MC, Mercer AA, McInnes CJ, Jacobs BL Vaccinia viruses with mutations in the E3L gene as potential replicationcompetent, attenuated vaccines: intra-nasal vaccination. Vaccine 26: Colamonici OR, Domanski P, Sweitzer SM, Larner A, Buller RM Vaccinia virus B18R gene encodes a type I interferon-binding protein that blocks interferon alpha transmembrane signaling. J Biol Chem 270: Symons JA, Alcami A, Smith GL Vaccinia virus encodes a soluble type I interferon receptor of novel structure and broad species specificity. Cell 81: Rusert P, Kouyos RD, Kadelka C, Ebner H, Schanz M, Huber M, Braun DL, Hoze N, Scherrer A, Magnus C, Weber J, Uhr T, Cippa V, Thorball CW, Kuster H, Cavassini M, Bernasconi E, Hoffmann M, Calmy A, Battegay M, Rauch A, Yerly S, Aubert V, Klimkait T, Boni J, Fellay J, Regoes RR, Gunthard HF, Trkola A, Swiss HIVCS Determinants of HIV-1 broadly neutralizing antibody induction. Nat Med doi: /nm Fouts TR, Bagley K, Prado IJ, Bobb KL, Schwartz JA, Xu R, Zagursky RJ, Egan MA, Eldridge JH, LaBranche CC, Montefiori DC, Le Buanec H, Zagury D, Pal R, Pavlakis GN, Felber BK, Franchini G, Gordon S, Vaccari M, Lewis GK, DeVico AL, Gallo RC Balance of cellular and humoral immunity determines the level of protection by HIV vaccines in rhesus macaque models of HIV infection. Proc Natl Acad Sci U S A 112:E Barouch DH, Alter G, Broge T, Linde C, Ackerman ME, Brown EP, Borducchi EN, Smith KM, Nkolola JP, Liu J, Shields J, Parenteau L, Whitney JB, Abbink P, Ng'ang'a DM, Seaman MS, Lavine CL, Perry JR, Li W, Colantonio AD, Lewis MG, Chen B, Wenschuh H, Reimer U, Piatak M, Lifson JD, Handley SA, Virgin HW, Koutsoukos M, Lorin C, Voss G, Weijtens M, Pau MG, Schuitemaker H Protective efficacy of adenovirus/protein vaccines against SIV challenges in rhesus monkeys. Science 349: Barouch DH, Stephenson KE, Borducchi EN, Smith K, Stanley K, McNally AG, Liu J, Abbink P, Maxfield LF, Seaman MS, Dugast AS, Alter G, Ferguson M, Li W, Earl PL, Moss B, Giorgi EE, Szinger JJ, Eller LA, Billings EA, Rao M, Tovanabutra S, Sanders-Buell E, Weijtens M, Pau MG, Schuitemaker H, Robb ML, Kim JH, Korber BT, Michael NL
27 Protective efficacy of a global HIV-1 mosaic vaccine against heterologous SHIV challenges in rhesus monkeys. Cell 155: Roederer M, Keele BF, Schmidt SD, Mason RD, Welles HC, Fischer W, Labranche C, Foulds KE, Louder MK, Yang ZY, Todd JP, Buzby AP, Mach LV, Shen L, Seaton KE, Ward BM, Bailer RT, Gottardo R, Gu W, Ferrari G, Alam SM, Denny TN, Montefiori DC, Tomaras GD, Korber BT, Nason MC, Seder RA, Koup RA, Letvin NL, Rao SS, Nabel GJ, Mascola JR Immunological and virological mechanisms of vaccine-mediated protection against SIV and HIV. Nature 505: FIGURE LEGENDS Figure 1. Enhanced expression, plaque size phenotype and innate immune profile of NYVAC-C-KC vectors. (A) Expression of HIV-1 gp140 by Western blotting. Monolayers of HeLa cells were mock-infected or infected at 5 PFU/cell with NYVAC- WT, NYVAC-KC, NYVAC-gp140, NYVAC-KC-gp140 or NYVAC-KC-gp140-ΔB19R. At 6 or 24h post infection, cells were lysed in Laemmli buffer, and cell extracts were fractionated by 8% SDS-PAGE and analyzed by Western blotting using the rabbit polyclonal anti-gp120 antibody. (B) Analysis of the plaque size. Monkey BSC-40 cells were infected with serial dilutions of NYVAC-WT, NYVAC-KC, NYVAC-gp140, NYVAC- KC-gp140 or NYVAC-KC-gp140-ΔB19R. At 48h post infection, cells were fixed and stained with crystal violet to visualize virus plaques. (C) Analysis of the innate immune profile in infected human THP-1 macrophages. Human THP-1 macrophages were mock-infected or infected at 5 PFU/cell with NYVAC-WT, NYVAC-KC, NYVAC-gp140, NYVAC-KC-gp140 or NYVAC-KC-gp140-ΔB19R. At 3 and 6 hours post-infection, RNA was extracted and the mrna levels of IFN-β, MIP-1α, IL-8, IL-1β, and HPRT were analyzed by RT-PCR. Results were expressed as the ratio of the gene of interest to HPRT mrna levels. A.U: arbitrary units. p values indicate significantly higher responses comparing NYVAC-KC-gp140 to NYVAC-KC-gp140-ΔB19R at the same hour (*, p<0.05; **, p<0.005; ***, p<0.001). Figure 2. Immunization schedule for non-human primates. (A) Immunization groups included in the AUP513 study. Eight NHPs (rhesus macaques) were immunized in each group at weeks 0 and 4 with the corresponding replication-competent NYVAC- C-KC poxvirus vector (NYVAC-C-KC or NYVAC-C-KC-ΔB19R) and at weeks 12 and 24 with a combination of replication-competent NYVAC-C-KC poxvirus vector plus a clade C HIV-1 gp120 protein, as detailed in Materials and Methods. The composition of NYVAC-C-KC, NYVAC-C-KC-ΔB19R, and the bivalent clade C HIV-1 gp120 protein is detailed in Materials and Methods. (B) Chronological diagram showing the immunization schedule and the immunogenicity endpoints used in this study. At weeks 27
28 , 4, 12 and 24, animals were immunized (see panel A). A dose of 1 x 10 8 PFU of each recombinant replication-competent poxvirus vector (NYVAC-KC-gp140 plus NYVAC- KC-Gag-Pol-Nef for the NYVAC-C-KC vector; and NYVAC-KC-gp140-ΔB19R plus NYVAC-KC-Gag-Pol-Nef-ΔB19R for the NYVAC-C-KC-ΔB19R vector; 2 x 10 8 PFU of total virus) and 50 µg of each clade C HIV-1 gp120 protein (TV1 gp120 plus 1086 gp120; 100 µg of total protein) were used in each immunization. Bivalent clade C gp120 protein was administered together with MF59 adjuvant. At weeks 0, 6, 14, 26, and 36 (at the beginning of the study, 2 weeks after the second, third, and fourth immunizations, and at the end of the study, respectively), PBMCs, serum and rectal mucosal samples were obtained from each immunized animal, and HIV-1-specific T cell and humoral immune responses were analyzed. Figure 3. Magnitude of HIV-1-specific T cell immune responses. PBMCs were collected at weeks 0, 6, 14, 26, and 36 from each rhesus macaque (n=8 per group) immunized with NYVAC-C-KC or NYVAC-C-KC-ΔB19R, and the total magnitude of HIV-1-specific T cell immune responses triggered by the different immunization groups were measured by ELISpot (A) or ICS (B and C) assays following stimulation of PBMCs with HIV-1 Env, Gag, Pol and Nef peptide pools. The total magnitude of SFUs per million of cells is shown (A), with values representing the number of IFN-γ positive T cells against Env, Gag, Pol, and Nef peptide pools. The total magnitudes of HIV-1- specific CD4 + (B) and CD8 + (C) T cell responses elicited by the different immunization groups are shown, with the values representing the sums of the percentages of T cells producing IFN-γ and/or TNF-α and/or IL-2 against Env, Gag, Pol, and Nef peptide pools. Values from unstimulated controls were subtracted in all cases. Each dot represents the value for one immunized macaque. Box plots represent the distribution of data values, with the line inside the box indicating the median value. p values and the number of responding rhesus macaques are indicated. Figure 4. Magnitude of HIV-1-specific CD4 + and CD8 + T cells producing cytokines. The overall magnitudes of HIV-1-specific CD4 + (panels A to C) and CD8 + (panels D to F) T cells elicited by the different immunization groups and producing IFNγ (A and D), TNF-α (B and E), or IL-2 (C and F) are shown. PBMCs were collected at weeks 6, 14, 26, and 36 from each rhesus macaque (n=8 per group) immunized with NYVAC-C-KC or NYVAC-C-KC-ΔB19R. HIV-1-specific CD4 + and CD8 + T cell immune responses triggered by both immunization groups were measured by ICS assay following stimulation of PBMCs with HIV-1 Env, Gag, Pol, and Nef peptide pools. The values represent the sums of the percentages of T cells producing IFN-γ (A and D) or 28
29 TNF-α (B and E) or IL-2 (C and F) against Env, Gag, Pol, and Nef peptide pools. Values from unstimulated controls were subtracted in all cases. Each dot represents the value for one immunized macaque. Box plots represent the distribution of data values, with the line inside the box indicating the median value. p values and the number of responding rhesus macaques are indicated. Figure 5. Binding IgG antibody responses. Total plasma binding IgG antibody levels against HIV-1 gp140 group M consensus (scon) (A), gp120 from isolate TV1 (B), gp120 from isolate 1086 (C), and MuLV gp70-scaffolded V1/V2 proteins (D) induced by the different immunization groups. (E) Rectal binding IgG antibody levels against HIV-1 gp140 group M consensus (scon), and gp120 from isolates 1086 and TV1 induced by the different immunization groups. Individual serum or rectal mucosal samples were obtained at weeks 6, 14, 26, and 36 from each rhesus macaque (n=8 per group) immunized with NYVAC-C-KC or NYVAC-C-KC-ΔB19R. Binding IgG antibodies were measured by BAMA (A to D), or by analyzing the binding magnitude normalized to total rhesus IgG (specific activity) (E), as indicated in Materials and Methods. The magnitudes of the total binding IgG antibody responses are expressed as the AUC from serial dilutions of plasma. Each dot represents the value for one immunized macaque. Box plots represent the distribution of data values, with the line inside the box indicating the median value. p values and the number of responding rhesus macaques are indicated. Figure 6. ADCC and NAb responses against HIV-1. (A) ADCC activity induced by the different immunization groups. Individual plasma samples were obtained at weeks 0, 26, and 36 from each rhesus macaque (n=8 per group) immunized with NYVAC-C- KC or NYVAC-C-KC-ΔB19R. ADCC activity against gp120 TV1 and 1086 proteins was measured as indicated in Materials and Methods. (B) Neutralization titers induced by the different immunization groups. Individual serum samples were obtained at weeks 26, and 36 from each rhesus macaque (n=8 per group) immunized with NYVAC-C-KC or NYVAC-C-KC-ΔB19R. NAb against different tier 1 and 2 HIV-1 strains were measured using the TZM-bl or A3R5 assays, as described in Materials and Methods. Each dot represents the value for one immunized macaque. Box plots represent the distribution of data values, with the line inside the box indicating the median value. p values and the number of responding rhesus macaques are indicated. Figure 7. Correlation between T cell and antibody responses. (A) Correlation plot summarizing Spearman s rank-based correlations of CD4 + T cell responses with B-cell 29
30 assays for NYVAC-C-KC and NYVAC-C-KC-ΔB19R at week 26, and ordered by overall median correlation for each cytokine and pooled antigen combination. (B) Correlation plot summarizing Spearman s rank-based correlations of CD8 + T cell responses with B-cell assays for NYVAC-C-KC and NYVAC-C-KC-ΔB19R at week 26, and ordered by overall median correlation for each cytokine and pooled antigen combination. CD4 + and CD8 + T cell responses were assessed for correlation with the gp120.tv1 antigen in the BAMA and ADCC GTL assays, and the MW isolate in the TZM.bl NAb assay. In each case, reported p-values are calculated using a onesided test with alternative that the true correlation coefficient is greater than 0. 30
31
Higher priming doses enhance HIV-specific humoral but not cellular. responses in a randomized, double-blind phase Ib clinical trial of
 Higher priming doses enhance HIV-specific humoral but not cellular responses in a randomized, double-blind phase Ib clinical trial of preventive HIV-1 vaccines Pierre-Alexandre Bart a,b, Yunda Huang c,
Higher priming doses enhance HIV-specific humoral but not cellular responses in a randomized, double-blind phase Ib clinical trial of preventive HIV-1 vaccines Pierre-Alexandre Bart a,b, Yunda Huang c,
Supplementary Figure 1. ALVAC-protein vaccines and macaque immunization. (A) Maximum likelihood
 Maximum likelihood") Supplementary Figure 1. ALVAC-protein vaccines and macaque immunization. (A) Maximum likelihood tree illustrating CRF01_AE gp120 protein sequence relationships between 107 Envs sampled in the RV144 trial
Supplementary Figure 1. ALVAC-protein vaccines and macaque immunization. (A) Maximum likelihood tree illustrating CRF01_AE gp120 protein sequence relationships between 107 Envs sampled in the RV144 trial
Streamlining Heterologous DNA-Prime and NYVAC/Protein-Boost HIV Vaccine Regimens in
 JVI Accepted Manuscript Posted Online 10 February 2016 J. Virol. doi:10.1128/jvi.03135-15 Copyright 2016, American Society for Microbiology. All Rights Reserved. 1 2 Streamlining Heterologous DNA-Prime
JVI Accepted Manuscript Posted Online 10 February 2016 J. Virol. doi:10.1128/jvi.03135-15 Copyright 2016, American Society for Microbiology. All Rights Reserved. 1 2 Streamlining Heterologous DNA-Prime
Employing Improved Antigens Allows Streamlining of Heterologous DNA-Prime and NYVAC/Protein-Boost HIV Vaccine Regimens in Rhesus Macaques
 Title Employing Improved Antigens Allows Streamlining of Heterologous DNA-Prime and NYVAC/Protein-Boost HIV Vaccine Regimens in Rhesus Macaques Running title: Improved HIV Antigens and Delivery Schedules
Title Employing Improved Antigens Allows Streamlining of Heterologous DNA-Prime and NYVAC/Protein-Boost HIV Vaccine Regimens in Rhesus Macaques Running title: Improved HIV Antigens and Delivery Schedules
HVTN Laboratory Program: Immunogenicity and Research Assays
 HVTN Laboratory Program: Immunogenicity and Research Assays Erica Andersen-Nissen, PhD Director, Cape Town HVTN Immunology Laboratory Considerations for a Pan-African HIV Vaccine Development Agenda Kigali,
HVTN Laboratory Program: Immunogenicity and Research Assays Erica Andersen-Nissen, PhD Director, Cape Town HVTN Immunology Laboratory Considerations for a Pan-African HIV Vaccine Development Agenda Kigali,
HIV: RV 144 prime boost HIV vaccine efficacy study
 HIV: RV 144 prime boost HIV vaccine efficacy study Nelson L. Michael, M.D., Ph.D Colonel, Medical Corps, U.S. Army Director US Military HIV Research Program (MHRP) Walter Reed Army Institute of Research
HIV: RV 144 prime boost HIV vaccine efficacy study Nelson L. Michael, M.D., Ph.D Colonel, Medical Corps, U.S. Army Director US Military HIV Research Program (MHRP) Walter Reed Army Institute of Research
HVTN P5 Vaccine Trials
 HVTN P5 Vaccine Trials Erica Andersen-Nissen, PhD Director, Cape Town HVTN Immunology Laboratory Considerations for a Pan-African HIV Vaccine Development Agenda Kigali, Rwanda 16-17 March 2015 HVTN Mission
HVTN P5 Vaccine Trials Erica Andersen-Nissen, PhD Director, Cape Town HVTN Immunology Laboratory Considerations for a Pan-African HIV Vaccine Development Agenda Kigali, Rwanda 16-17 March 2015 HVTN Mission
Immunogenicity of ALVAC HIV (vcp1521) and AIDSVAX B/E Prime Boost Vaccination in RV144, Thai Phase III HIV Vaccine Trial
 and AIDSVAX B/E Prime Boost Vaccination in RV144, Thai Phase III HIV Vaccine Trial") Immunogenicity of ALVAC HIV (vcp1521) and AIDSVAX B/E Prime Boost Vaccination in RV144, Thai Phase III HIV Vaccine Trial M. de Souza, R. Trichavaroj, A. Schuetz, W. Chuenarom, Y. Phuang ngern, S. Jongrakthaitae,
Immunogenicity of ALVAC HIV (vcp1521) and AIDSVAX B/E Prime Boost Vaccination in RV144, Thai Phase III HIV Vaccine Trial M. de Souza, R. Trichavaroj, A. Schuetz, W. Chuenarom, Y. Phuang ngern, S. Jongrakthaitae,
Professor Andrew McMichael
 BHIVA AUTUMN CONFERENCE 2011 Including CHIVA Parallel Sessions Professor Andrew McMichael University of Oxford 17 18 November 2011, Queen Elizabeth II Conference Centre, London BHIVA AUTUMN CONFERENCE
BHIVA AUTUMN CONFERENCE 2011 Including CHIVA Parallel Sessions Professor Andrew McMichael University of Oxford 17 18 November 2011, Queen Elizabeth II Conference Centre, London BHIVA AUTUMN CONFERENCE
AIDSVaccine2010 Atlanta, Georgia Willy Bogers. NIH HIVRad Grant nr 5P01AI066287
 HIV-1 envelope-cd4 receptor complexes elicit broad T- and B- cell immune responses as well as cross-reactive neutralizing antibodies in Rhesus macaques NIH HIVRad Grant nr 5P01AI066287 AIDSVaccine2010
HIV-1 envelope-cd4 receptor complexes elicit broad T- and B- cell immune responses as well as cross-reactive neutralizing antibodies in Rhesus macaques NIH HIVRad Grant nr 5P01AI066287 AIDSVaccine2010
GOVX-B11: A Clade B HIV Vaccine for the Developed World
 GeoVax Labs, Inc. 19 Lake Park Drive Suite 3 Atlanta, GA 3 (678) 384-72 GOVX-B11: A Clade B HIV Vaccine for the Developed World Executive summary: GOVX-B11 is a Clade B HIV vaccine targeted for use in
GeoVax Labs, Inc. 19 Lake Park Drive Suite 3 Atlanta, GA 3 (678) 384-72 GOVX-B11: A Clade B HIV Vaccine for the Developed World Executive summary: GOVX-B11 is a Clade B HIV vaccine targeted for use in
HIV-specific humoral responses benefit from stronger prime in phase Ib clinical trial
 The Journal of Clinical Investigation Clinical Medicine HIV-specific humoral responses benefit from stronger prime in phase Ib clinical trial Pierre-Alexandre Bart, 1,2,11 Yunda Huang, 3,11 Shelly T. Karuna,
The Journal of Clinical Investigation Clinical Medicine HIV-specific humoral responses benefit from stronger prime in phase Ib clinical trial Pierre-Alexandre Bart, 1,2,11 Yunda Huang, 3,11 Shelly T. Karuna,
The Kitchen Sink. Glenda Gray AIDS vaccine at the cross roads: how to adapt to a new preven9on age? Monday 7 th October, 2013, Barcelona
 The Kitchen Sink Glenda Gray AIDS vaccine at the cross roads: how to adapt to a new preven9on age? Monday 7 th October, 2013, Barcelona The only important slide of my presentation. All efficacy trial in
The Kitchen Sink Glenda Gray AIDS vaccine at the cross roads: how to adapt to a new preven9on age? Monday 7 th October, 2013, Barcelona The only important slide of my presentation. All efficacy trial in
on April 26, 2018 by guest
 JVI Accepted Manuscript Posted Online 20 July 2016 J. Virol. doi:10.1128/jvi.00853-16 Copyright 2016, American Society for Microbiology. All Rights Reserved. 1 2 3 4 5 6 7 8 9 10 11 Induction of heterologous
JVI Accepted Manuscript Posted Online 20 July 2016 J. Virol. doi:10.1128/jvi.00853-16 Copyright 2016, American Society for Microbiology. All Rights Reserved. 1 2 3 4 5 6 7 8 9 10 11 Induction of heterologous
Novel Vaccine Products for Planned Phase I Immunogenicity Studies in Infants
 Office of AIDS Research Novel Vaccine Products for Planned Phase I Immunogenicity Studies in Infants L. Jean Patterson, PhD Office of AIDS Research, NIH February 7, 2017 Office of AIDS Research OAR Responsibilities
Office of AIDS Research Novel Vaccine Products for Planned Phase I Immunogenicity Studies in Infants L. Jean Patterson, PhD Office of AIDS Research, NIH February 7, 2017 Office of AIDS Research OAR Responsibilities
HIV and Challenges of Vaccine Development
 Dale and Betty Bumpers Vaccine Research Center National Institute of Allergy and Infectious Diseases National Institutes of Health HIV and Challenges of Vaccine Development Richard A. Koup, MD INTEREST
Dale and Betty Bumpers Vaccine Research Center National Institute of Allergy and Infectious Diseases National Institutes of Health HIV and Challenges of Vaccine Development Richard A. Koup, MD INTEREST
4/14/2016. HIV Vaccines and Immunoprotection: Where Are We? Learning Objectives. After attending this presentation, participants will be able to:
 HIV Vaccines and Immunoprotection: Where Are We? Mark J. Mulligan, MD, FIDSA Distinguished Professor of Medicine Emory University School of Medicine Atlanta, Georgia FINAL: 04/01/16 Atlanta, Georgia: Friday,
HIV Vaccines and Immunoprotection: Where Are We? Mark J. Mulligan, MD, FIDSA Distinguished Professor of Medicine Emory University School of Medicine Atlanta, Georgia FINAL: 04/01/16 Atlanta, Georgia: Friday,
Recombinant Baculovirus Derived HIV-1 Virus-Like Particles Elicit Potent Neutralizing Antibody Responses
 Recombinant Baculovirus Derived HIV-1 Virus-Like Particles Elicit Potent Neutralizing Antibody Responses Weimin Liu University of Alabama at Birmingham Introduction and Rationale Virus-like particles (VLPs)
Recombinant Baculovirus Derived HIV-1 Virus-Like Particles Elicit Potent Neutralizing Antibody Responses Weimin Liu University of Alabama at Birmingham Introduction and Rationale Virus-like particles (VLPs)
Why are validated immunogenicity assays important for HIV vaccine development?
 Why are validated immunogenicity assays important for HIV vaccine development? There is a need to compare immunogenicity of products in the pipeline, when similar or different in class when developed by
Why are validated immunogenicity assays important for HIV vaccine development? There is a need to compare immunogenicity of products in the pipeline, when similar or different in class when developed by
ALVAC -HIV and AIDSVAX B/E Prime-Boost HIV-1 Preventive Vaccine Regimen. Results of the Thai HIV Vaccine Trial, RV144
 ALVAC -HIV and AIDSVAX B/E Prime-Boost HIV-1 Preventive Vaccine Regimen Results of the Thai HIV Vaccine Trial, RV144 SupachaiRerks-Ngarm, PunneePittisutthithum, SorachaiNitayaphan, JaranitKaewkungwal,
ALVAC -HIV and AIDSVAX B/E Prime-Boost HIV-1 Preventive Vaccine Regimen Results of the Thai HIV Vaccine Trial, RV144 SupachaiRerks-Ngarm, PunneePittisutthithum, SorachaiNitayaphan, JaranitKaewkungwal,
Supporting Information
 Supporting Information Sui et al..7/pnas.997 Pre-CLP CM9 LA9 SL Tat# Pol Vif % Tetramer + CD + CD + Vac+IL- +IL- Vac Fig. S. Frequencies of six different CD + CD + Mamu-A*-tetramer + cells were measured
Supporting Information Sui et al..7/pnas.997 Pre-CLP CM9 LA9 SL Tat# Pol Vif % Tetramer + CD + CD + Vac+IL- +IL- Vac Fig. S. Frequencies of six different CD + CD + Mamu-A*-tetramer + cells were measured
Boosts Following Priming with gp120 DNA
 Neutralizing Antibody Responses Induced with V3-scaffold Protein Boosts Following Priming with gp120 DNA Susan Zolla-Pazner NYU School of Medicine Problems with Whole Env Immunogens Poor induction of Abs
Neutralizing Antibody Responses Induced with V3-scaffold Protein Boosts Following Priming with gp120 DNA Susan Zolla-Pazner NYU School of Medicine Problems with Whole Env Immunogens Poor induction of Abs
staining and flow cytometry
 Detection of influenza virus-specific T cell responses by intracellular by cytokine intracellular staining cytokine staining and flow cytometry Detection of influenza virus-specific T cell responses and
Detection of influenza virus-specific T cell responses by intracellular by cytokine intracellular staining cytokine staining and flow cytometry Detection of influenza virus-specific T cell responses and
Progress in HIV Vaccine Development Magdalena Sobieszczyk, MD, MPH. Division of Infectious Diseases Columbia University Medical Center
 Progress in HIV Vaccine Development Magdalena Sobieszczyk, MD, MPH Division of Infectious Diseases Columbia University Medical Center 1 Outline A short history of HIV vaccine design and development Describe
Progress in HIV Vaccine Development Magdalena Sobieszczyk, MD, MPH Division of Infectious Diseases Columbia University Medical Center 1 Outline A short history of HIV vaccine design and development Describe
Replicating measles-shiv vaccine induces long term preservation of central memory CD4 cells in the gut of macaques challenged with SHIV89.
 Replicating measles-shiv vaccine induces long term preservation of central memory CD4 cells in the gut of macaques challenged with SHIV89.6P Frédéric Tangy Viral Genomics and Vaccination Laboratory Measles
Replicating measles-shiv vaccine induces long term preservation of central memory CD4 cells in the gut of macaques challenged with SHIV89.6P Frédéric Tangy Viral Genomics and Vaccination Laboratory Measles
A Path to an HIV Vaccine: GSID Consortium Activities. Faruk Sinangil, PhD 4th Annual CAVD Meeting Miami, FL December 1-4, 2009
 A Path to an HIV Vaccine: GSID Consortium Activities Faruk Sinangil, PhD 4th Annual CAVD Meeting Miami, FL December 1-4, 2009 Project Goals Acquire and disseminate information that will contribute to the
A Path to an HIV Vaccine: GSID Consortium Activities Faruk Sinangil, PhD 4th Annual CAVD Meeting Miami, FL December 1-4, 2009 Project Goals Acquire and disseminate information that will contribute to the
Novel Heterologous Prime-Boost Vaccine Strategies for HIV. Dan Barouch April 18, 2012
 Novel Heterologous Prime-Boost Vaccine Strategies for HIV Dan Barouch April 18, 2012 Desired Features of a Next Generation HIV-1 Vaccine Candidate The RV1 study suggests that an HIV-1 vaccine is possible
Novel Heterologous Prime-Boost Vaccine Strategies for HIV Dan Barouch April 18, 2012 Desired Features of a Next Generation HIV-1 Vaccine Candidate The RV1 study suggests that an HIV-1 vaccine is possible
Antibody Dependent Cellular Cytotxic activity: Past and Future. Guido Ferrari, M.D. Duke University Medical Center
 Antibody Dependent Cellular Cytotxic activity: Past and Future Guido Ferrari, M.D. Duke University Medical Center Mechanism of Antibody Dependent Cellular Cytotoxicity (ADCC) ADCC Effector Cells (NK, monocytes/macrophages,
Antibody Dependent Cellular Cytotxic activity: Past and Future Guido Ferrari, M.D. Duke University Medical Center Mechanism of Antibody Dependent Cellular Cytotoxicity (ADCC) ADCC Effector Cells (NK, monocytes/macrophages,
An HIV-1 clade C DNA prime, NYVAC boost vaccine regimen induces reliable, polyfunctional, and long-lasting T cell responses
 ARTICLE An HIV-1 clade C DNA prime, NYVAC boost vaccine regimen induces reliable, polyfunctional, and long-lasting T cell responses The Journal of Experimental Medicine CORRESPONDENCE Giuseppe Pantaleo:
ARTICLE An HIV-1 clade C DNA prime, NYVAC boost vaccine regimen induces reliable, polyfunctional, and long-lasting T cell responses The Journal of Experimental Medicine CORRESPONDENCE Giuseppe Pantaleo:
HIV/AIDS: vaccines and alternate strategies for treatment and prevention
 Ann. N.Y. Acad. Sci. ISSN 0077-8923 ANNALS OF THE NEW YORK ACADEMY OF SCIENCES Online Meeting Report HIV/AIDS: vaccines and alternate strategies for treatment and prevention Yegor Voronin 1 and Sanjay
Ann. N.Y. Acad. Sci. ISSN 0077-8923 ANNALS OF THE NEW YORK ACADEMY OF SCIENCES Online Meeting Report HIV/AIDS: vaccines and alternate strategies for treatment and prevention Yegor Voronin 1 and Sanjay
HIV 1 Preventive Vaccine Regimen. Community based Trial in Thailand V 144. for the MOPH TAVEG Collaboration
 V ALVAC HIV and AIDSVAX B/E Prime Boost HIV 1 Preventive Vaccine Regimen Final Results of the Phase III Community based Trial in Thailand Supachai Rerks Ngarm, Punnee Pittisutthithum, Sorachai Nitayaphan,
V ALVAC HIV and AIDSVAX B/E Prime Boost HIV 1 Preventive Vaccine Regimen Final Results of the Phase III Community based Trial in Thailand Supachai Rerks Ngarm, Punnee Pittisutthithum, Sorachai Nitayaphan,
FUNDERS UPDATE- BMGF. Third HIV Env Manufacturing Workshop Sponsored by DAIDS-NIAID-NIH and the HIV Global Vaccine Enterprise July 20-21, 2017
 FUNDERS UPDATE- BMGF Third HIV Env Manufacturing Workshop Sponsored by DAIDS-NIAID-NIH and the HIV Global Vaccine Enterprise July 20-21, 2017 Susan Barnett Senior Program Officer, Product and Clinical
FUNDERS UPDATE- BMGF Third HIV Env Manufacturing Workshop Sponsored by DAIDS-NIAID-NIH and the HIV Global Vaccine Enterprise July 20-21, 2017 Susan Barnett Senior Program Officer, Product and Clinical
Trial Objectives immunogenicity safety
 Optimal priming of poxvirus vector (NYVAC)-based HIV vaccine regimens requires 3 DNA injections. Long term Results of the EV03/ANRS Vac20 Phase I/II Trial. Yves Levy, Kim Ellefsen, Wolfgang Stöehr, Pierre-Alexandre
Optimal priming of poxvirus vector (NYVAC)-based HIV vaccine regimens requires 3 DNA injections. Long term Results of the EV03/ANRS Vac20 Phase I/II Trial. Yves Levy, Kim Ellefsen, Wolfgang Stöehr, Pierre-Alexandre
Advancing Toward HIV-1 Vaccine Efficacy through the Intersections of Immune Correlates
 Vaccines 2014, 2, 15-35; doi:10.3390/vaccines2010015 Review OPEN ACCESS vaccines ISSN 2076-393X www.mdpi.com/journal/vaccines Advancing Toward HIV-1 Vaccine Efficacy through the Intersections of Immune
Vaccines 2014, 2, 15-35; doi:10.3390/vaccines2010015 Review OPEN ACCESS vaccines ISSN 2076-393X www.mdpi.com/journal/vaccines Advancing Toward HIV-1 Vaccine Efficacy through the Intersections of Immune
NIH Public Access Author Manuscript Nature. Author manuscript; available in PMC 2009 July 1.
 NIH Public Access Author Manuscript Published in final edited form as: Nature. 2009 January 1; 457(7225): 87 91. doi:10.1038/nature07469. Immune Control of an SIV Challenge by a T Cell-Based Vaccine in
NIH Public Access Author Manuscript Published in final edited form as: Nature. 2009 January 1; 457(7225): 87 91. doi:10.1038/nature07469. Immune Control of an SIV Challenge by a T Cell-Based Vaccine in
Antibody-Dependent Cell-Mediated Cytotoxicity in Simian Immunodeficiency Virus-Infected Rhesus Monkeys
 JOURNAL OF VIROLOGY, July 2011, p. 6906 6912 Vol. 85, No. 14 0022-538X/11/$12.00 doi:10.1128/jvi.00326-11 Copyright 2011, American Society for Microbiology. All Rights Reserved. Antibody-Dependent Cell-Mediated
JOURNAL OF VIROLOGY, July 2011, p. 6906 6912 Vol. 85, No. 14 0022-538X/11/$12.00 doi:10.1128/jvi.00326-11 Copyright 2011, American Society for Microbiology. All Rights Reserved. Antibody-Dependent Cell-Mediated
Preventive and therapeutic HIV vaccines. Markus Bickel Infektiologikum Frankfurt
 Preventive and therapeutic HIV vaccines Markus Bickel Infektiologikum Frankfurt No conflicts to declare Disclosures Background FAQ: by patients and colleagues Publication about promising results, especially
Preventive and therapeutic HIV vaccines Markus Bickel Infektiologikum Frankfurt No conflicts to declare Disclosures Background FAQ: by patients and colleagues Publication about promising results, especially
Commercially available HLA Class II tetramers (Beckman Coulter) conjugated to
 conjugated to") Class II tetramer staining Commercially available HLA Class II tetramers (Beckman Coulter) conjugated to PE were combined with dominant HIV epitopes (DRB1*0101-DRFYKTLRAEQASQEV, DRB1*0301- PEKEVLVWKFDSRLAFHH,
Class II tetramer staining Commercially available HLA Class II tetramers (Beckman Coulter) conjugated to PE were combined with dominant HIV epitopes (DRB1*0101-DRFYKTLRAEQASQEV, DRB1*0301- PEKEVLVWKFDSRLAFHH,
A global approach to HIV-1 vaccine development
 A global approach to HIV-1 vaccine development The Harvard community has made this article openly available. Please share how this access benefits you. Your story matters. Citation Published Version Accessed
A global approach to HIV-1 vaccine development The Harvard community has made this article openly available. Please share how this access benefits you. Your story matters. Citation Published Version Accessed
Rapid perforin upregulation directly ex vivo by CD8 + T cells is a defining characteristic of HIV elite controllers
 Rapid perforin upregulation directly ex vivo by CD8 + T cells is a defining characteristic of HIV elite controllers Adam R. Hersperger Department of Microbiology University of Pennsylvania Evidence for
Rapid perforin upregulation directly ex vivo by CD8 + T cells is a defining characteristic of HIV elite controllers Adam R. Hersperger Department of Microbiology University of Pennsylvania Evidence for
Innate and Cellular Immunology Control of Infection by Cell-mediated Immunity
 Innate & adaptive Immunity Innate and Cellular Immunology Control of Infection by Cell-mediated Immunity Helen Horton PhD Seattle Biomedical Research Institute Depts of Global Health & Medicine, UW Cellular
Innate & adaptive Immunity Innate and Cellular Immunology Control of Infection by Cell-mediated Immunity Helen Horton PhD Seattle Biomedical Research Institute Depts of Global Health & Medicine, UW Cellular
NIH Public Access Author Manuscript Nat Med. Author manuscript; available in PMC 2010 September 1.
 NIH Public Access Author Manuscript Published in final edited form as: Nat Med. 2010 March ; 16(3): 319 323. doi:10.1038/nm.2089. Mosaic HIV-1 Vaccines Expand the Breadth and Depth of Cellular Immune Responses
NIH Public Access Author Manuscript Published in final edited form as: Nat Med. 2010 March ; 16(3): 319 323. doi:10.1038/nm.2089. Mosaic HIV-1 Vaccines Expand the Breadth and Depth of Cellular Immune Responses
NK mediated Antibody Dependent Cellular Cytotoxicity in HIV infections
 NK mediated Antibody Dependent Cellular Cytotoxicity in HIV infections Amy Chung Dr. Ivan Stratov Prof. Stephen Kent ADCC process consists of Target cell QuickTime and a TIFF (Uncompressed) FcγR decompressor
NK mediated Antibody Dependent Cellular Cytotoxicity in HIV infections Amy Chung Dr. Ivan Stratov Prof. Stephen Kent ADCC process consists of Target cell QuickTime and a TIFF (Uncompressed) FcγR decompressor
Virus Panels for Assessing Vaccine-Elicited Neutralizing Antibodies
 Virus Panels for Assessing Vaccine-Elicited Neutralizing Antibodies Michael Seaman, Ph.D. Center for Virology and Vaccine Research Beth Israel Deaconess Medical Center Harvard Medical School J. Virol.
Virus Panels for Assessing Vaccine-Elicited Neutralizing Antibodies Michael Seaman, Ph.D. Center for Virology and Vaccine Research Beth Israel Deaconess Medical Center Harvard Medical School J. Virol.
The Harvard community has made this article openly available. Please share how this access benefits you. Your story matters
 Antibody Responses After Analytic Treatment Interruption in Human Immunodeficiency Virus-1- Infected Individuals on Early Initiated Antiretroviral Therapy The Harvard community has made this article openly
Antibody Responses After Analytic Treatment Interruption in Human Immunodeficiency Virus-1- Infected Individuals on Early Initiated Antiretroviral Therapy The Harvard community has made this article openly
HIV Vaccine. Sunee Sirivichayakul, Ph.D. Faculty of Medicine Chulalongkorn University. August 22, 2014
 HIV Vaccine Sunee Sirivichayakul, Ph.D. Faculty of Medicine Chulalongkorn University August 22, 2014 Immunity Natural immunity Active natural immunity e.g., infection Passive natural immunity e.g., trans-placental
HIV Vaccine Sunee Sirivichayakul, Ph.D. Faculty of Medicine Chulalongkorn University August 22, 2014 Immunity Natural immunity Active natural immunity e.g., infection Passive natural immunity e.g., trans-placental
EMERGING ISSUES IN THE HUMORAL IMMUNE RESPONSE TO HIV. (Summary of the recommendations from an Enterprise Working Group)
") AIDS Vaccine 07, Seattle, August 20-23, 2007 EMERGING ISSUES IN THE HUMORAL IMMUNE RESPONSE TO HIV (Summary of the recommendations from an Enterprise Working Group) The Working Group Reston, Virginia,
AIDS Vaccine 07, Seattle, August 20-23, 2007 EMERGING ISSUES IN THE HUMORAL IMMUNE RESPONSE TO HIV (Summary of the recommendations from an Enterprise Working Group) The Working Group Reston, Virginia,
The humoral immune responses to IBV proteins.
 The humoral immune responses to IBV proteins. E. Dan Heller and Rosa Meir The Hebrew University of Jerusalem, Israel COST FA1207 meeting WG2 + WG3, Budapest, Jan. 2015 1 IBV encodes four major structural
The humoral immune responses to IBV proteins. E. Dan Heller and Rosa Meir The Hebrew University of Jerusalem, Israel COST FA1207 meeting WG2 + WG3, Budapest, Jan. 2015 1 IBV encodes four major structural
Supplementary appendix
 Supplementary appendix This appendix formed part of the original submission and has been peer reviewed. We post it as supplied by the authors. Supplement to: Barouch DH, Tomaka FL Wegmann F, et al. Evaluation
Supplementary appendix This appendix formed part of the original submission and has been peer reviewed. We post it as supplied by the authors. Supplement to: Barouch DH, Tomaka FL Wegmann F, et al. Evaluation
DATA SHEET. Biological Activity: Vif Binding assays pending.
 Proteins APOBEC >> All DATA SHEET Reagent: Recombinant Human APOBEC3F(E. coli) Catalog Number: 11097 Lot Number: 3 098121 Release Category: A Provided: 1mg/ml. Mass/vial: 50µg Diluent: PBS, 0.1% Sarcosyl.
Proteins APOBEC >> All DATA SHEET Reagent: Recombinant Human APOBEC3F(E. coli) Catalog Number: 11097 Lot Number: 3 098121 Release Category: A Provided: 1mg/ml. Mass/vial: 50µg Diluent: PBS, 0.1% Sarcosyl.
2288 GJ Rijswijk, The Netherlands. Albinusdreef 2, 2333 ZA Leiden, The Netherlands
 Journal of General Virology (215), 96, 1478 1483 DOI 1.199/vir..74 Short Communication Correspondence Petra Mooij mooij@bprc.nl Synthetic long peptide booster immunization in rhesus macaques primed with
Journal of General Virology (215), 96, 1478 1483 DOI 1.199/vir..74 Short Communication Correspondence Petra Mooij mooij@bprc.nl Synthetic long peptide booster immunization in rhesus macaques primed with
HIV Vaccine. 15 September 2016 นพ.นคร เปรมศร ผ อ านวยการส าน กงาน โครงการศ กษาว คซ นเอดส ทดลอง
 HIV Vaccine 15 September 2016 นพ.นคร เปรมศร ผ อ านวยการส าน กงาน โครงการศ กษาว คซ นเอดส ทดลอง GLOBAL STATISTICS 2015 17 million people were accessing antiretroviral therapy 36.7 million people globally
HIV Vaccine 15 September 2016 นพ.นคร เปรมศร ผ อ านวยการส าน กงาน โครงการศ กษาว คซ นเอดส ทดลอง GLOBAL STATISTICS 2015 17 million people were accessing antiretroviral therapy 36.7 million people globally
Development of a Universal T Cell Vaccine. Tomáš Hanke Weatherall Institute of Molecular Medicine University of Oxford United Kingdom
 Development of a Universal T Cell Vaccine Tomáš Hanke Weatherall Institute of Molecular Medicine University of Oxford United Kingdom Development of HIV-1 vaccines Induction of cell-mediated responses Immunogens
Development of a Universal T Cell Vaccine Tomáš Hanke Weatherall Institute of Molecular Medicine University of Oxford United Kingdom Development of HIV-1 vaccines Induction of cell-mediated responses Immunogens
HIV-1 vaccine-induced immunity in the test-of-concept Step Study: a case cohort analysis
 HIV-1 vaccine-induced immunity in the test-of-concept Step Study: a case cohort analysis M Juliana McElrath, Stephen C De Rosa, Zoe Moodie, Sheri Dubey, Lisa Kierstead, Holly Janes, Olivier D Defawe, Donald
HIV-1 vaccine-induced immunity in the test-of-concept Step Study: a case cohort analysis M Juliana McElrath, Stephen C De Rosa, Zoe Moodie, Sheri Dubey, Lisa Kierstead, Holly Janes, Olivier D Defawe, Donald
Activation of NK Cells by ADCC Antibodies and HIV Disease Progression
 RAPID COMMUNICATION Activation of NK Cells by ADCC Antibodies and HIV Disease Progression Amy W. Chung, BSc, PhD,* Marjon Navis, PhD,* Gamze Isitman, BSc,* Leia Wren, BSc,* Julie Silvers, RN, Janaki Amin,
RAPID COMMUNICATION Activation of NK Cells by ADCC Antibodies and HIV Disease Progression Amy W. Chung, BSc, PhD,* Marjon Navis, PhD,* Gamze Isitman, BSc,* Leia Wren, BSc,* Julie Silvers, RN, Janaki Amin,
Current State of HIV Vaccine Development
 Current State of HIV Vaccine Development Sandhya Vasan, MD US Military HIV Research Program Henry M. Jackson Foundation Armed Forces Research Institute of Medical Sciences, Bangkok, Thailand AVAC HVAD
Current State of HIV Vaccine Development Sandhya Vasan, MD US Military HIV Research Program Henry M. Jackson Foundation Armed Forces Research Institute of Medical Sciences, Bangkok, Thailand AVAC HVAD
Are we targeting the right HIV determinants?
 QuickTime et un décompresseur TIFF (non compressé) sont requis pour visionner cette image. AIDS Vaccine 2009 October 22 nd 2009 - Paris Are we targeting the right HIV determinants? Françoise BARRÉ-SINOUSSI
QuickTime et un décompresseur TIFF (non compressé) sont requis pour visionner cette image. AIDS Vaccine 2009 October 22 nd 2009 - Paris Are we targeting the right HIV determinants? Françoise BARRÉ-SINOUSSI
IL-12 and GM-CSF in DNA/MVA Immunizations against HIV-1 CRF12_BF Nef Induced T-Cell Responses With an Enhanced Magnitude, Breadth and Quality
 IL-12 and GM-CSF in DNA/MVA Immunizations against HIV-1 CRF12_BF Nef Induced T-Cell Responses With an Enhanced Magnitude, Breadth and Quality Ana María Rodríguez, María Fernanda Pascutti., Cynthia Maeto.,
IL-12 and GM-CSF in DNA/MVA Immunizations against HIV-1 CRF12_BF Nef Induced T-Cell Responses With an Enhanced Magnitude, Breadth and Quality Ana María Rodríguez, María Fernanda Pascutti., Cynthia Maeto.,
Correlates of Immunity: RV144 - Lessons Learned
 Correlates of Immunity: RV144 - Lessons Learned US Military HIV Research Program (MHRP) Nelson L. Michael, MD, PhD AIDS Vaccine 2010, Atlanta GA 01 October 2010 Outline RV 144 Correlates Discovery Towards
Correlates of Immunity: RV144 - Lessons Learned US Military HIV Research Program (MHRP) Nelson L. Michael, MD, PhD AIDS Vaccine 2010, Atlanta GA 01 October 2010 Outline RV 144 Correlates Discovery Towards
Identification of Mutation(s) in. Associated with Neutralization Resistance. Miah Blomquist
 in. Associated with Neutralization Resistance. Miah Blomquist") Identification of Mutation(s) in the HIV 1 gp41 Subunit Associated with Neutralization Resistance Miah Blomquist What is HIV 1? HIV-1 is an epidemic that affects over 34 million people worldwide. HIV-1
Identification of Mutation(s) in the HIV 1 gp41 Subunit Associated with Neutralization Resistance Miah Blomquist What is HIV 1? HIV-1 is an epidemic that affects over 34 million people worldwide. HIV-1
Induction of Innate Immune Responses in HVTN 071: a Trial using the MRKAd5 gag/pol/nef Vaccine from the Step Study
 Induction of Innate Immune Responses in HVTN 71: a Trial using the MRKAd5 gag/pol/nef Vaccine from the Step Study Erica Andersen-Nissen Vaccine and Infectious Disease Institute Fred Hutchinson Cancer Research
Induction of Innate Immune Responses in HVTN 71: a Trial using the MRKAd5 gag/pol/nef Vaccine from the Step Study Erica Andersen-Nissen Vaccine and Infectious Disease Institute Fred Hutchinson Cancer Research
Biomedical Engineering for Global Health. Lecture 10 HIV/AIDS vaccine development
 Biomedical Engineering for Global Health Lecture 10 HIV/AIDS vaccine development Review of lecture 9 How do vaccines work? Types ofvaccines: Review of lecture 9 Are vaccines effective? -Edward Jenner s
Biomedical Engineering for Global Health Lecture 10 HIV/AIDS vaccine development Review of lecture 9 How do vaccines work? Types ofvaccines: Review of lecture 9 Are vaccines effective? -Edward Jenner s
Fostering Clinical Development for HIV-1 Vaccine
 W Fostering Clinical Development for HIV-1 Vaccine Ravimiarenduse alane seminar 9. oktoobril Tallinnas Mart Ustav, PhD CSO, SVP 1 FIT Biotech Founded in 1995 Operations in Tampere, Finland and Tartu, Estonia
W Fostering Clinical Development for HIV-1 Vaccine Ravimiarenduse alane seminar 9. oktoobril Tallinnas Mart Ustav, PhD CSO, SVP 1 FIT Biotech Founded in 1995 Operations in Tampere, Finland and Tartu, Estonia
Alternate Antibody-Based Therapeutic Strategies To Purge the HIV Cell Reservoir
 Alternate Antibody-Based Therapeutic Strategies To Purge the HIV Cell Reservoir Giuseppe Pantaleo, M.D. Professor of Medicine Head, Division of Immunology and Allergy Executive Director, Swiss Vaccine
Alternate Antibody-Based Therapeutic Strategies To Purge the HIV Cell Reservoir Giuseppe Pantaleo, M.D. Professor of Medicine Head, Division of Immunology and Allergy Executive Director, Swiss Vaccine
New Preventive Technology: Providing New Options to Stop the Spread of HIV/AIDS. Dublin, Ireland. June 24, AIDS Vaccines.
 New Preventive Technology: Providing New Options to Stop the Spread of HIV/AIDS Dublin, Ireland June 24, 2004 AIDS Vaccines - An R&D Briefing This paper has been prepared by the International AIDS Vaccine
New Preventive Technology: Providing New Options to Stop the Spread of HIV/AIDS Dublin, Ireland June 24, 2004 AIDS Vaccines - An R&D Briefing This paper has been prepared by the International AIDS Vaccine
Current Strategies in HIV-1 Vaccine Development Using Replication-Defective Adenovirus as a Case Study
 Note: I have added some clarifying comments to the slides -- please click on Comments under View to see them. Current Strategies in HIV-1 Vaccine Development Using Replication-Defective Adenovirus as a
Note: I have added some clarifying comments to the slides -- please click on Comments under View to see them. Current Strategies in HIV-1 Vaccine Development Using Replication-Defective Adenovirus as a
Application of systems biology to identify predictors of HIV vaccine immunogenicity
 Application of systems biology to identify predictors of HIV vaccine immunogenicity Daniel Zak 1, Erica Andersen-Nissen 2, Haesun Park 3, Scott Hansen 3, Karl Mullen 3, Kristi Hamilton 1, Kathleen Kennedy
Application of systems biology to identify predictors of HIV vaccine immunogenicity Daniel Zak 1, Erica Andersen-Nissen 2, Haesun Park 3, Scott Hansen 3, Karl Mullen 3, Kristi Hamilton 1, Kathleen Kennedy
Principle of the FluoroSpot assay. Anti-tag mab-green. Streptavidin-Red. Detection mab-tag. Detection mab-biotin. Analyte. Analyte.
 FluoroSpot 1 The principle objective of the FluoroSpot assay is the simultaneous measurement of dual cytokine secretion at the single cell level. This is accomplished by using a mixture of monoclonal antibodies
FluoroSpot 1 The principle objective of the FluoroSpot assay is the simultaneous measurement of dual cytokine secretion at the single cell level. This is accomplished by using a mixture of monoclonal antibodies
Advancing HIV Vaccines into Efficacy studies. Glenda Gray WHO PD VAC 7-9 September 2015 Geneva, Switzerland
 Advancing HIV Vaccines into Efficacy studies Glenda Gray WHO PD VAC 7-9 September 2015 Geneva, Switzerland What we need to end AIDS: controlling viremia New classes of Drugs Point of care viral load &
Advancing HIV Vaccines into Efficacy studies Glenda Gray WHO PD VAC 7-9 September 2015 Geneva, Switzerland What we need to end AIDS: controlling viremia New classes of Drugs Point of care viral load &
Developing a safe and effective vaccine against human immunodeficiency
 RESEARCH ARTICLE crossmark Mucosal Immunization with Newcastle Disease Virus Vector Coexpressing HIV-1 Env and Gag Proteins Elicits Potent Serum, Mucosal, and Cellular Immune Responses That Protect against
RESEARCH ARTICLE crossmark Mucosal Immunization with Newcastle Disease Virus Vector Coexpressing HIV-1 Env and Gag Proteins Elicits Potent Serum, Mucosal, and Cellular Immune Responses That Protect against
HIV Vaccines: Basic Science
 Dale and Betty Bumpers Vaccine Research Center National Institute of Allergy and Infectious Diseases National Institutes of Health HIV Vaccines: Basic Science Richard A. Koup, MD 6 th INTEREST Workshop
Dale and Betty Bumpers Vaccine Research Center National Institute of Allergy and Infectious Diseases National Institutes of Health HIV Vaccines: Basic Science Richard A. Koup, MD 6 th INTEREST Workshop
Module 8: Introduction to Immune Correlates of Protection. Instructors: Ivan Chan, Peter Gilbert, Paul T. Edlefsen, Ying Huang
 Module 8: Evaluating Immune Correlates of Protection Instructors: Ivan Chan, Peter Gilbert, Paul T. Edlefsen, Ying Huang Session 7: RV144 Thai Trial Case Study (HIV Vaccine Efficacy Trial) Summer Institute
Module 8: Evaluating Immune Correlates of Protection Instructors: Ivan Chan, Peter Gilbert, Paul T. Edlefsen, Ying Huang Session 7: RV144 Thai Trial Case Study (HIV Vaccine Efficacy Trial) Summer Institute
Global Panel of HIV-1 Env Reference Strains for Standardized Assessments of Vaccine- Elicited Neutralizing Antibodies
 JVI Accepts, published online ahead of print on 18 December 2013 J. Virol. doi:10.1128/jvi.02853-13 Copyright 2013, American Society for Microbiology. All Rights Reserved. 1 2 Global Panel of HIV-1 Env
JVI Accepts, published online ahead of print on 18 December 2013 J. Virol. doi:10.1128/jvi.02853-13 Copyright 2013, American Society for Microbiology. All Rights Reserved. 1 2 Global Panel of HIV-1 Env
Supplementary Fig. 1: Ex vivo tetramer enrichment with anti-c-myc beads
 Supplementary Fig. 1: Ex vivo tetramer enrichment with anti-c-myc beads Representative example of comparative ex vivo tetramer enrichment performed in three independent experiments with either conventional
Supplementary Fig. 1: Ex vivo tetramer enrichment with anti-c-myc beads Representative example of comparative ex vivo tetramer enrichment performed in three independent experiments with either conventional
Update on influenza vaccination using microneedle delivery
 Mark Prausnitz serves as a consultant and is an inventor on patents licensed to companies developing products related to this presentation. This potential conflict of interest is being managed by Georgia
Mark Prausnitz serves as a consultant and is an inventor on patents licensed to companies developing products related to this presentation. This potential conflict of interest is being managed by Georgia
Supporting Information
 Supporting Information Walker et al. 1.173/pnas.111753118 - JR-CSF 1 3 1 4 1 5 1 6 1 7 1 8 blank beads protein A beads JR-FL - 1 3 1 4 1 5 1 6 1 7 1 8 - MGRM-C26 1 3 1 4 1 5 1 6 1 7 1 8 reciprocal serum
Supporting Information Walker et al. 1.173/pnas.111753118 - JR-CSF 1 3 1 4 1 5 1 6 1 7 1 8 blank beads protein A beads JR-FL - 1 3 1 4 1 5 1 6 1 7 1 8 - MGRM-C26 1 3 1 4 1 5 1 6 1 7 1 8 reciprocal serum
Development of a VP6 subunit rotavirus vaccine A dual role of VP6 as a vaccine antigen and an adjuvant
 30 August 2018, Minsk 13TH INTERNATIONAL ROTAVIRUS SYMPOSIUM Development of a VP6 subunit rotavirus vaccine A dual role of VP6 as a vaccine antigen and an adjuvant Dr. Vesna Blazevic Head of Laboratory
30 August 2018, Minsk 13TH INTERNATIONAL ROTAVIRUS SYMPOSIUM Development of a VP6 subunit rotavirus vaccine A dual role of VP6 as a vaccine antigen and an adjuvant Dr. Vesna Blazevic Head of Laboratory
DNA and Protein Vaccination Confers Protection Upon Mucosal Challenge with Heterologous SIVsmE660 (OA 10.04)
") DNA and Protein Vaccination Confers Protection Upon Mucosal Challenge with Heterologous SIVsmE660 (OA 10.04) Rashmi Jalah Human Retrovirus Pathogenesis Section, Vaccine Branch, CCR, National Cancer Institute
DNA and Protein Vaccination Confers Protection Upon Mucosal Challenge with Heterologous SIVsmE660 (OA 10.04) Rashmi Jalah Human Retrovirus Pathogenesis Section, Vaccine Branch, CCR, National Cancer Institute
Progress on new vaccine strategies against chronic viral infections
 Progress on new vaccine strategies against chronic viral infections Jay A. Berzofsky,, Masaki Terabe, Igor M. Belyakov J Clin Invest. 2004;114(4):450-462. https://doi.org/10.1172/jci22674. Review Among
Progress on new vaccine strategies against chronic viral infections Jay A. Berzofsky,, Masaki Terabe, Igor M. Belyakov J Clin Invest. 2004;114(4):450-462. https://doi.org/10.1172/jci22674. Review Among
Gene Vaccine Dr. Sina Soleimani
 Gene Vaccine Dr. Sina Soleimani Human Viral Vaccines Quality Control Laboratory (HVVQC) Titles 1. A short Introduction of Vaccine History 2. First Lineage of Vaccines 3. Second Lineage of Vaccines 3. New
Gene Vaccine Dr. Sina Soleimani Human Viral Vaccines Quality Control Laboratory (HVVQC) Titles 1. A short Introduction of Vaccine History 2. First Lineage of Vaccines 3. Second Lineage of Vaccines 3. New
Professor Jonathan Weber
 THIRD JOINT CONFERENCE OF BHIVA AND BASHH 2014 Professor Jonathan Weber Imperial College, London 1-4 April 2014, Arena and Convention Centre Liverpool THIRD JOINT CONFERENCE OF BHIVA AND BASHH 2014 Professor
THIRD JOINT CONFERENCE OF BHIVA AND BASHH 2014 Professor Jonathan Weber Imperial College, London 1-4 April 2014, Arena and Convention Centre Liverpool THIRD JOINT CONFERENCE OF BHIVA AND BASHH 2014 Professor
RAISON D ETRE OF THE IMMUNE SYSTEM:
 RAISON D ETRE OF THE IMMUNE SYSTEM: To Distinguish Self from Non-Self Thereby Protecting Us From Our Hostile Environment. Innate Immunity Acquired Immunity Innate immunity: (Antigen nonspecific) defense
RAISON D ETRE OF THE IMMUNE SYSTEM: To Distinguish Self from Non-Self Thereby Protecting Us From Our Hostile Environment. Innate Immunity Acquired Immunity Innate immunity: (Antigen nonspecific) defense
Nature Medicine: doi: /nm.2109
 HIV 1 Infects Multipotent Progenitor Cells Causing Cell Death and Establishing Latent Cellular Reservoirs Christoph C. Carter, Adewunmi Onafuwa Nuga, Lucy A. M c Namara, James Riddell IV, Dale Bixby, Michael
HIV 1 Infects Multipotent Progenitor Cells Causing Cell Death and Establishing Latent Cellular Reservoirs Christoph C. Carter, Adewunmi Onafuwa Nuga, Lucy A. M c Namara, James Riddell IV, Dale Bixby, Michael
Optimizing Intracellular Flow Cytometry:
 Optimizing Intracellular Flow Cytometry: Simultaneous Detection of Cytokines and Transcription Factors An encore presentation by Jurg Rohrer, PhD, BD Biosciences 10.26.10 Outline Introduction Cytokines
Optimizing Intracellular Flow Cytometry: Simultaneous Detection of Cytokines and Transcription Factors An encore presentation by Jurg Rohrer, PhD, BD Biosciences 10.26.10 Outline Introduction Cytokines
Module 8: Evaluating Immune Correlates of Protection
 Module 8: Evaluating Immune Correlates of Protection Instructors: Ivan Chan, Peter Gilbert, Paul T. Edlefsen, Ying Huang Session 7: RV144 Thai Trial Case Study (HIV Vaccine Efficacy Trial) Summer Institute
Module 8: Evaluating Immune Correlates of Protection Instructors: Ivan Chan, Peter Gilbert, Paul T. Edlefsen, Ying Huang Session 7: RV144 Thai Trial Case Study (HIV Vaccine Efficacy Trial) Summer Institute
A centralized gene-based HIV-1 vaccine elicits broad cross-clade cellular immune responses in rhesus monkeys
 A centralized gene-based HIV-1 vaccine elicits broad cross-clade cellular immune responses in rhesus monkeys Sampa Santra, Bette T. Korber, Mark Muldoon, Dan H. Barouch, Gary J. Nabel, Feng Gao, Beatrice
A centralized gene-based HIV-1 vaccine elicits broad cross-clade cellular immune responses in rhesus monkeys Sampa Santra, Bette T. Korber, Mark Muldoon, Dan H. Barouch, Gary J. Nabel, Feng Gao, Beatrice
Superior Control of HIV-1 Replication by CD8+ T Cells Targeting Conserved Epitopes: Implications for HIV Vaccine Design
 Superior Control of HIV-1 Replication by CD8+ T Cells Targeting Conserved Epitopes: Implications for HIV Vaccine Design Pratima Kunwar 1,7, Natalie Hawkins 2, Warren L. Dinges 1,3, Yi Liu 5, Erin E. Gabriel
Superior Control of HIV-1 Replication by CD8+ T Cells Targeting Conserved Epitopes: Implications for HIV Vaccine Design Pratima Kunwar 1,7, Natalie Hawkins 2, Warren L. Dinges 1,3, Yi Liu 5, Erin E. Gabriel
Mosaic vaccines elicit CD8 + T lymphocyte responses that confer enhanced immune coverage of diverse HIV strains in monkeys
 Mosaic vaccines elicit CD + T lymphocyte responses that fer enhanced immune coverage of diverse HIV strains in monkeys 21 Nature America, Inc. All rights reserved. Sampa Santra 1, Hua-Xin Liao 2, Ruijin
Mosaic vaccines elicit CD + T lymphocyte responses that fer enhanced immune coverage of diverse HIV strains in monkeys 21 Nature America, Inc. All rights reserved. Sampa Santra 1, Hua-Xin Liao 2, Ruijin
SUPPLEMENTARY INFORMATION
 Complete but curtailed T-cell response to very-low-affinity antigen Dietmar Zehn, Sarah Y. Lee & Michael J. Bevan Supp. Fig. 1: TCR chain usage among endogenous K b /Ova reactive T cells. C57BL/6 mice
Complete but curtailed T-cell response to very-low-affinity antigen Dietmar Zehn, Sarah Y. Lee & Michael J. Bevan Supp. Fig. 1: TCR chain usage among endogenous K b /Ova reactive T cells. C57BL/6 mice
Isolation of a Broadly Neutralizing Antibody with Low Somatic Mutation from a Chronically Infected HIV-1 Patient
 Isolation of a Broadly Neutralizing Antibody with Low Somatic Mutation from a Chronically Infected HIV-1 Patient Amanda Fabra García, Carolina Beltrán Pavez, Alberto Merino Mansilla, Cristina Xufré, Isabel
Isolation of a Broadly Neutralizing Antibody with Low Somatic Mutation from a Chronically Infected HIV-1 Patient Amanda Fabra García, Carolina Beltrán Pavez, Alberto Merino Mansilla, Cristina Xufré, Isabel
24 26 January 2013, Hong Kong SAR, CHINA. TITLE from VIEW and SLIDE MASTER February 27, 2013
 The first WHO integrated meeting on development and clinical trials of influenza vaccines that induce broadly protective and long-lasting immune responses 24 26 January 2013, Hong Kong SAR, CHINA 1 TITLE
The first WHO integrated meeting on development and clinical trials of influenza vaccines that induce broadly protective and long-lasting immune responses 24 26 January 2013, Hong Kong SAR, CHINA 1 TITLE
Generation of Robust Antibody Responses to HIV-1 MPER Antigens in Mice Reconstituted with Cultured B cells
 Generation of Robust Antibody Responses to HIV-1 MPER Antigens in Mice Reconstituted with Cultured B cells T. Matt Holl 1, Masayuki Kuraoka 1, Dongmei Liao 1, Laurent Verkoczy 2, M. Anthony Moody 2, Munir
Generation of Robust Antibody Responses to HIV-1 MPER Antigens in Mice Reconstituted with Cultured B cells T. Matt Holl 1, Masayuki Kuraoka 1, Dongmei Liao 1, Laurent Verkoczy 2, M. Anthony Moody 2, Munir
Blocking antibodies and peptides. Rat anti-mouse PD-1 (29F.1A12, rat IgG2a, k), PD-
, PD-") Supplementary Methods Blocking antibodies and peptides. Rat anti-mouse PD-1 (29F.1A12, rat IgG2a, k), PD- L1 (10F.9G2, rat IgG2b, k), and PD-L2 (3.2, mouse IgG1) have been described (24). Anti-CTLA-4 (clone
Supplementary Methods Blocking antibodies and peptides. Rat anti-mouse PD-1 (29F.1A12, rat IgG2a, k), PD- L1 (10F.9G2, rat IgG2b, k), and PD-L2 (3.2, mouse IgG1) have been described (24). Anti-CTLA-4 (clone
Planned Vaccine Trials to Follow-up on RV 144. Merlin Robb, MD Deputy Director for Clinical Research MHRP
 Planned Vaccine Trials to Follow-up on RV 144 Merlin Robb, MD Deputy Director for Clinical Research MHRP Planned studies are mutually reinforcing and will amplify public health impact and regional relevance
Planned Vaccine Trials to Follow-up on RV 144 Merlin Robb, MD Deputy Director for Clinical Research MHRP Planned studies are mutually reinforcing and will amplify public health impact and regional relevance
DEBATE ON HIV ENVELOPE AS A T CELL IMMUNOGEN HAS BEEN GAG-GED
 DEBATE ON HIV ENVELOPE AS A T CELL IMMUNOGEN HAS BEEN GAG-GED Viv Peut Kent Laboratory, University of Melbourne, Australia WHY ENVELOPE? Env subject to both humoral and cellular immune responses Perhaps
DEBATE ON HIV ENVELOPE AS A T CELL IMMUNOGEN HAS BEEN GAG-GED Viv Peut Kent Laboratory, University of Melbourne, Australia WHY ENVELOPE? Env subject to both humoral and cellular immune responses Perhaps
Fayth K. Yoshimura, Ph.D. September 7, of 7 HIV - BASIC PROPERTIES
 1 of 7 I. Viral Origin. A. Retrovirus - animal lentiviruses. HIV - BASIC PROPERTIES 1. HIV is a member of the Retrovirus family and more specifically it is a member of the Lentivirus genus of this family.
1 of 7 I. Viral Origin. A. Retrovirus - animal lentiviruses. HIV - BASIC PROPERTIES 1. HIV is a member of the Retrovirus family and more specifically it is a member of the Lentivirus genus of this family.
CD25-PE (BD Biosciences) and labeled with anti-pe-microbeads (Miltenyi Biotec) for depletion of CD25 +
 and labeled with anti-pe-microbeads (Miltenyi Biotec) for depletion of CD25 +") Supplements Supplemental Materials and Methods Depletion of CD25 + T-cells from PBMC. Fresh or HD precultured PBMC were stained with the conjugate CD25-PE (BD Biosciences) and labeled with anti-pe-microbeads
Supplements Supplemental Materials and Methods Depletion of CD25 + T-cells from PBMC. Fresh or HD precultured PBMC were stained with the conjugate CD25-PE (BD Biosciences) and labeled with anti-pe-microbeads
CD4 + T-cell-inducing HIV vaccines may have an impact on viral control
 CD4 + T-cell-inducing HIV vaccines may have an impact on viral control Clues from cytokines produced by CD4 + T-cells from HIVinfected patients and vaccinated seronegative volunteers Eva Van Braeckel 1,
CD4 + T-cell-inducing HIV vaccines may have an impact on viral control Clues from cytokines produced by CD4 + T-cells from HIVinfected patients and vaccinated seronegative volunteers Eva Van Braeckel 1,
Improved Innate and Adaptive Immunostimulation by Genetically Modified HIV-1 Protein Expressing NYVAC Vectors
 Improved Innate and Adaptive Immunostimulation by Genetically Modified HIV-1 Protein Expressing NYVAC Vectors Esther D. Quakkelaar 1, Anke Redeker 1, Elias K. Haddad 2, Alexandre Harari 3,4, Stella Mayo
Improved Innate and Adaptive Immunostimulation by Genetically Modified HIV-1 Protein Expressing NYVAC Vectors Esther D. Quakkelaar 1, Anke Redeker 1, Elias K. Haddad 2, Alexandre Harari 3,4, Stella Mayo
Received 25 January 2011/Accepted 22 April 2011
 JOURNAL OF VIROLOGY, July 2011, p. 7029 7036 Vol. 85, No. 14 0022-538X/11/$12.00 doi:10.1128/jvi.00171-11 Copyright 2011, American Society for Microbiology. All Rights Reserved. An HIV-1 gp120 Envelope
JOURNAL OF VIROLOGY, July 2011, p. 7029 7036 Vol. 85, No. 14 0022-538X/11/$12.00 doi:10.1128/jvi.00171-11 Copyright 2011, American Society for Microbiology. All Rights Reserved. An HIV-1 gp120 Envelope