Title Page. Viral Exploitation of Host SOCS Protein Functions. Lisa Nowoslawski Akhtar and Etty N. Benveniste
|
|
|
- Elmer Eaton
- 6 years ago
- Views:
Transcription
1 JVI Accepts, published online ahead of print on 17 November 2010 J. Virol. doi: /jvi Copyright 2010, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved. 1 Title Page Viral Exploitation of Host SOCS Protein Functions 7 8 Lisa Nowoslawski Akhtar and Etty N. Benveniste 9 10 Department of Cell Biology, University of Alabama at Birmingham, Birmingham, AL, USA Corresponding Author: Dr. Etty (Tika) Benveniste University of Alabama at Birmingham, Department of Cell Biology, Tinsley Harrison Tower Room 926, 1900 University Blvd., Birmingham, Alabama , USA Phone: (205) ; Fax: (205) ; tika@uab.edu
2 21 22 Abstract Over the past decade a family of host proteins known as Suppressors Of Cytokine Signaling 23 (SOCS) have emerged as frequent targets of viral exploitation. Under physiologic circumstances, SOCS proteins negatively regulate inflammatory signaling pathways by facilitating ubiquitination and proteosomal degradation of pathway machinery. Their expression is tightly regulated to prevent excessive inflammation while maintaining protective anti- 27 pathogenic responses. Numerous viruses, however, have developed mechanisms to induce robust host SOCS protein expression following infection, essentially hijacking SOCS function to promote virus survival. To date, SOCS proteins have been shown to inhibit protective antiviral signaling pathways, allowing viruses to evade the host immune response, and to ubiquitinate viral proteins, facilitating intracellular viral trafficking and progeny virus assembly. Importantly, manipulation of SOCS proteins not only facilitates progression of the viral life cycle, but also powerfully shapes the presentation of viral disease. SOCS proteins can define host susceptibility to infection, contribute to peripheral disease manifestations such as immune dysfunction and cancer, and even modify the efficacy of therapeutic interventions. Looking toward the future, it is clear that a better understanding of the role of SOCS proteins in viral diseases will be essential in our struggle to modulate and even eliminate the pathogenic effects of viruses on the host. 39 2
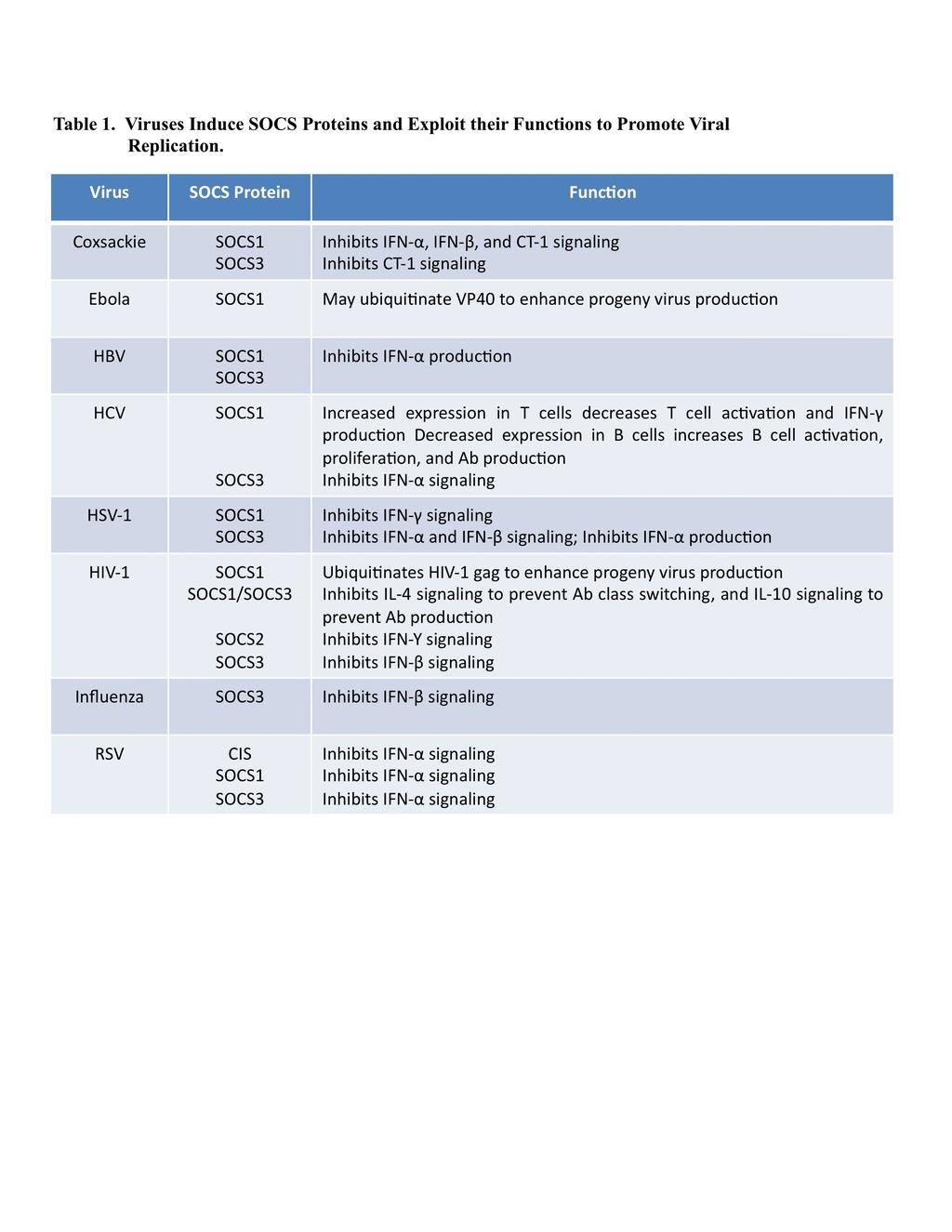
3 Introduction Viruses possess a compact genome which is only sufficient to encode the most essential viral proteins. Therefore, viruses must rely on the host cell to supply a number of additional proteins that are required for completion of the viral life cycle, in order to establish a productive infection. For example, viruses may require the use of host cell surface receptors to enter a cell, DNA polymerases to replicate the viral genome, RNA polymerases to transcribe viral genes, and translational machinery to produce viral proteins, as well as various other cellular proteins to enhance intracellular trafficking of viral components and to evade immune detection. Determining the requirement for these cellular factors contributes not only to our basic 48 knowledge of the viral life cycle, but also to our understanding of viral disease. Viral dependency on a particular cellular protein can define host range and cellular tropism, contribute to the development of virus-associated pathology, or even offer new therapeutic strategies. Considering the current obstacles to successful treatment of many viral illnesses, including human immunodeficiency virus type 1 (HIV-1), identification of new targets can not be overlooked. The SOCS family has recently been identified as a group of host proteins that can be exploited for viral benefit. Members of the SOCS family are induced upon infection by a number of different viruses including HIV-1, hepatitis C virus (HCV), hepatitis B virus (HBV), herpes simplex virus type 1 (HSV-1), respiratory syncytial virus (RSV), Ebola virus, influenza A virus, and coxsackie virus; and subsequently contribute to viral replication and pathogenesis. This review will focus on the virally exploited functions of SOCS proteins, as well as the consequences of these functions for viral disease and therapy. 61 3
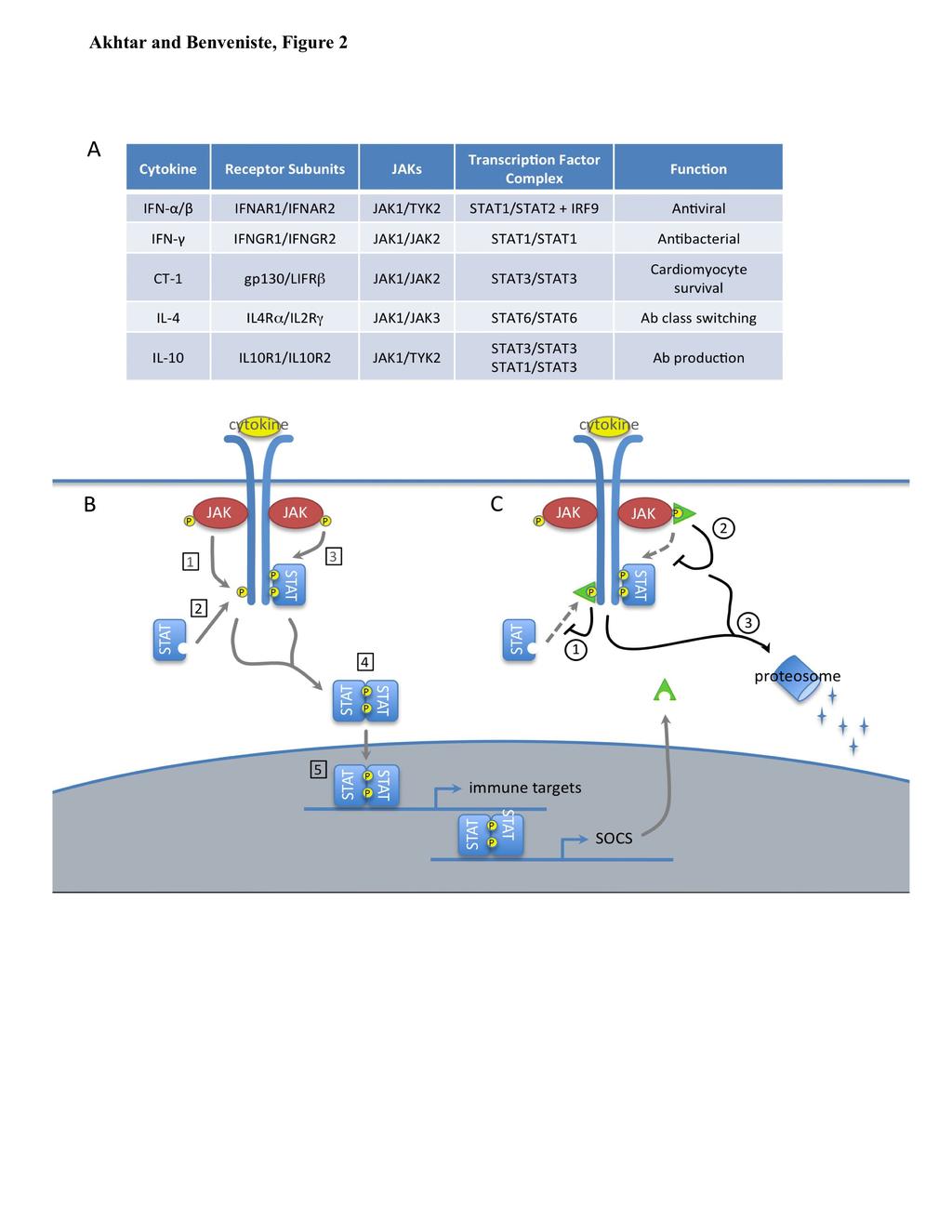
4 SOCS Protein Structure and Function The SOCS family of proteins contains eight members: cytokine-inducible SH2-domaincontaining protein (CIS) and SOCS1-SOCS7. Each contain a central SH2 domain, a C-terminal SOCS box, and an N-terminal domain of varying length and composition (Figure 1) (78). The SH2 domain determines the target of each SOCS protein by binding specific phosphorylated tyrosine residues on its preferred substrate. Once bound, the SOCS box can interact with a complex containing Elongin B and C, Cullin5, and RING-box-2, to form an E3 ubiquitin ligase. By bringing the SH2-bound substrate into close proximity with ubiquitinating machinery, SOCS proteins facilitate the ubiquitination of target proteins, marking them for degradation via the proteosome (51). Certain SOCS proteins also harbor an additional effector domain in their N- terminus. SOCS1 and SOCS3 contain a kinase inhibitory region (KIR) that functions as a pseudosubstrate to inhibit kinase activity. Collectively, this multifaceted structure provides for a wide, and rapidly growing, range of biological effects. The most well described function of SOCS proteins is the negative regulation of the JAK/STAT pathway (Figure 2). In this pathway, cytokine stimulation of cell surface receptors activates receptor-associated tyrosine kinases known as Janus Kinases (JAKs), which subsequently phosphorylate the receptor cytoplasmic domain and lead to the recruitment of 79 Signal Transducers and Activators of Transcription (STATs). Recruited STATs are then activated by JAK phosphorylation, allowing them to dimerize and enter the nucleus to induce the transcription of target genes. SOCS proteins regulate this pathway in a family member-specific manner, by binding specific substrates within the JAK/STAT signaling receptor complex and terminating pathway activation in one of three ways: 1) SOCS box-mediated ubiquitination and degradation of bound receptor components, 2) competition with recruited STAT proteins for 4
5 shared phosphotyrosine residues, or 3) KIR-mediated inhibition of JAK activity (78). SOCS1 and SOCS3 are thought to bind phosphotyrosine residues within the activation loop of the JAK and/or nearby residues on the receptor cytoplasmic domain, and inhibit signal transduction either 88 through their KIR or SOCS box. By contrast, CIS and SOCS2 are thought to bind 89 phosphotyrosine residues on the cytoplasmic domain of the receptor and compete with, or 90 sterically hinder, the binding of recruited STATs. Certain SOCS proteins have also been reported to regulate additional signal transduction pathways, although these functions have been less well described. SOCS1 can inhibit the NF- B pathway by binding the NF- B subunit p65 (55), or one of the upstream signaling components MAL (36) or IRAK (14,42), and targeting 94 them for ubiquitination and degradation by the proteosome. SOCS1 can also induce the proteosomal degradation of ASK1 (25), an upstream activator of JNK and p38, thereby inhibiting these arms of the MAPK pathway. SOCS3 can inhibit both the NF- B and JNK/p38 pathways by binding the upstream signaling molecule TRAF6 and preventing its association with, and activation of, TAK1 (20). Together, these SOCS protein functions limit excessive pathway activation and shape complex signaling responses. As a critical regulator of multiple signal transduction pathways, SOCS protein expression is carefully regulated in a cell type- and stimulus-specific manner (32). SOCS proteins are most classically induced by cytokine stimulation of the JAK/STAT pathway (Figure 2B), creating a negative feedback loop to limit excessive inflammation (Figure 2C). However, their expression can also be induced following activation of the NF- B or MAPK pathways, and in response to a variety of stimuli including LPS, TNF-, isoproterenol and statins. In most cases, transcriptional activation is the sole mechanism for enhancing SOCS levels, although TNF- has been shown to 107 promote SOCS3 mrna stability (15). Additionally, the short half-life of SOCS proteins 5
6 (typically 1-2 h) can be altered by phosphorylation (21) or ubiquitination (58), and by association with the serine/threonine kinase PIM1 (9,48) or other SOCS proteins. By tightly controlling SOCS protein levels, these mechanisms provide for optimal pathway activation in a variety of cellular contexts. Viruses, however, can independently induce SOCS proteins and exploit their functions to promote viral replication (Table 1). Enhanced SOCS expression can inhibit the normal function of JAK/STAT-regulated pathways, most notably those critical for initiating innate and adaptive 115 antiviral immune responses. SOCS box activity can also be utilized to ubiquitinate viral proteins, thereby facilitating the intracellular trafficking of viral proteins required for progeny virus particle production. These hijacked host functions provide significant benefit to viral pathogens, and are critical for understanding both viral replication and disease Virally Exploited Functions of SOCS Proteins JAK/STAT Pathway Inhibition Provides for Viral Immune Evasion Viral infection results in a robust and multifaceted host immune response. The rapid, nonspecific, innate immune response is primarily driven by the virally induced expression of type I interferons (IFNs): IFN- and IFN-. Type I IFNs exert their effects by signaling through the JAK/STAT pathway to induce the transcription of multiple antiviral targets (65). Following IFN stimulation of the cell-surface receptor subunits IFN- / receptor (IFNAR) 1 and IFNAR2, receptor-associated JAKs (JAK1 and TYK2) recruit and phosphorylate cytoplasmic STAT1 and STAT2. The STAT1-STAT2 heterodimer associates with IFN-regulatory factor (IRF) 9 to form the IFN-stimulated gene factor 3 (ISGF3), which enters the nucleus and binds IFN-stimulated regulatory elements (ISRE) in the promoters of hundreds of antiviral IFN-stimulated genes 6
7 (ISGs) to induce their expression (Figure 2A). Some of the most well studied antiviral ISGs include the double stranded RNA-dependent protein kinase (PKR), which inhibits viral protein translation, 2,5 -oligoadenylate synthetase (OAS), which results in viral RNA cleavage, and the Mx proteins, which interfere with viral transcription. Collectively, ISGs have been shown to inhibit every stage of viral replication, from viral entry and uncoating to assembly and release, providing the host with formidable protection against viral infection. Following the innate immune response is the slower, antigen-specific, adaptive immune response. Adaptive immunity to viral infection requires the balanced effects of both cytotoxic T lymphocytes (CTLs or CD8+ T cells) and B cell-generated antibodies, the actions of which are orchestrated by CD4+ T cells (67). CTLs target virally-infected cells after recognizing viral antigens presented on the cell surface in the context of MHC class I molecules. CTLs proliferate in response to antigen recognition and CD4+ T cell help, and act to eradicate virus from these cells either through a lytic mechanism involving perforins and granzymes, or through a non-lytic mechanism involving production of the antiviral cytokine IFN-. By contrast, B cell-generated 145 antibodies typically target cell-free virus. B cells that recognize virus-specific antigens proliferate in response to antigen recognition and CD4+ T cell help, and generate antibodies capable of binding specific viral epitopes. These virus-specific antibodies can neutralize or promote aggregation of cell-free virus, preventing further infection of cells, or target virally infected cells for destruction by complement- or natural killer cell-dependent mechanisms. The functions of these two arms of the adaptive immune response complement each other, and are both necessary for effective viral control. When functioning properly, the innate and adaptive immune systems prevent productive viral infection. In response, viral evolution has favored the acquisition of a number of different 7
8 immune evasion mechanisms (67). Viruses can prevent IFN production, IFN signaling, or the function of antiviral ISGs; limit the production of CTLs and their ability to recognize virally infected targets; or rapidly mutate epitopes recognized by neutralizing antibodies. Often viruses use multiple methods of immune evasion, highlighting the importance of this function for survival Innate Immunity The obvious potential of SOCS proteins to inhibit the innate immune response to viral infection was recognized only shortly after the initial descriptions of SOCS functions. Through the use of overexpression studies, Song and Shuai determined that SOCS1 and SOCS3, but not SOCS2, could attenuate both type I (IFN- ) and type II IFN (IFN- ) signaling through the JAK/STAT pathway (63). They showed that exogenously expressed SOCS1 and SOCS3 were sufficient to inhibit not only IFN-induced phosphorylation and nuclear translocation of STAT1, but also the antiviral effect of IFN on vesicular stomatitis virus infection. Much later, the observation that SOCS1 knockout mice were extremely resistant to Semliki Forest virus infection provided confirmation that endogenously expressed SOCS proteins play an important role in regulating innate antiviral immunity. Hertzog and colleagues reported that SOCS1- deficiency resulted in decreased viral load and increased host survival following viral infection, which was reversed in the presence of IFNAR1-deficiency or upon treatment with IFN- / neutralizing antibodies, indicating both that type I IFNs are critical to the endogenous antiviral response and that SOCS1 functions to inhibit them (16). Further studies indicated that SOCS1 exerted these effects, at least in part, by directly binding IFNAR1 to inhibit STAT1 phosphorylation and the expression of the antiviral target gene OAS. Collectively, these studies 8
9 provided evidence that SOCS proteins are capable of dampening the antiviral effects of IFNs by disrupting JAK/STAT signaling within the host cell. It was not long before it was recognized that viruses could hijack host SOCS proteins to manipulate antiviral IFN signaling to their advantage. HCV, a small, positive-sense, singlestranded (ss) RNA virus that infects hepatocytes of the liver to cause liver inflammation (hepatitis), was the first virus reported to independently induce SOCS expression. Notably, HCV was already well known for its effective suppression of the host antiviral immune response, as evidenced by its propensity for establishing a chronic, lifelong infection in approximately 80% of cases (5). But the contribution of SOCS proteins to HCV s immune evasion arsenal was not described until 2003 by Bode et al. Their early studies showed that overexpression of the HCV core protein induced SOCS3 expression in HepG2 cells, which correlated with an inhibition of IFN- -induced STAT1 activation, nuclear translocation, and DNA binding (6). The authors further showed that HCV core protein was sufficient to enhance the replication of an experimental influenza virus engineered to lack its own IFN antagonist, suggesting that HCV core protein-induced SOCS3 was capable of inhibiting the antiviral type I IFN response through disruption of the JAK/STAT pathway. Later studies confirmed that SOCS3 expression was also increased in HCV-infected HepG2 cells, and in the peripheral lymphocytes of patients infected with HCV as compared to healthy controls (49). While a number of subsequent studies gave further support to the role of HCV-induced SOCS3 in viral persistence (29,50), not all reports are 196 consistent. Limited studies indicate that SOCS1, but not SOCS3 expression, is increased in the hepatic tissue of patients with chronic HCV infection (26). Another reports that in OR6 cells and JFH1-infected Huh7.5.1 cells, SOCS3 expression inhibits HCV replication in an mtor- dependent manner (60). While the factors contributing to these discrepancies are not yet clear, 9
10 the bulk of the literature continues to suggest that enhanced expression of SOCS proteins during HCV infection dramatically impairs innate antiviral signaling, which contributes not only to the chronicity of HCV infection, but also the decreased response of patients to exogenous IFN therapy, which will be described further below. HIV-1, a virus which infects CD4+ T cells and monocytes/macrophages thereby causing devastating host immunodeficiency, has also been shown to induce SOCS proteins in order to 206 suppress antiviral innate immunity. HIV-1 is a relatively large retrovirus with a complex immune evasion strategy, including the production of multiple viral accessory proteins (Vif, Nef, Vpu, Vpr) designed specifically to thwart innate immunity (35). Studies performed in our lab have determined that HIV-1 Tat, a regulatory protein whose primary function is to enhance viral transcription by recruiting transcription factors to the HIV-1 promoter, also contributes to HIV-1 immune evasion by inducing SOCS3 expression (2). HIV-1 Tat-induced SOCS3 attenuated IFN- signaling in macrophages both upstream, at the level of STAT1 and STAT2 activation, and downstream, at the level of PKR and ISG20 antiviral gene expression. In vitro, SOCS3 expression was sufficient to overcome the inhibitory effect of IFN- and enhance HIV-1 replication in macrophages, while in vivo, SOCS3 expression in the central nervous system (CNS) of simian immunodeficiency virus (SIV)-infected macaques correlated with increases in CNS SIV replication and the onset of CNS disease. These studies suggest that HIV-1 Tatinduced SOCS3 disrupts protective type I IFN signaling within macrophages, allowing for enhanced viral replication and viral pathogenesis. HIV-1 Tat has also been shown to attenuate type II IFN signaling in human monocytes in a SOCS2-dependent manner (10). This has significant implications for protection from bacterial and parasitic infections that are common in the context of HIV-1 infection, and will be described further below. 10
11 HSV-1 is a large DNA virus that chronically infects its hosts through evasion of antiviral immunity and latency (46). HSV-1 initially infects and replicates within keratinocytes of the skin, and then subsequently establishes a more long term, latent infection within adjacent 226 neurons, where it is hidden from immune surveillance. Periodically, HSV-1 will become 227 activated by external stimuli and productively re-infect nearby keratinocytes, resulting in skin 228 damaging lesions. Interestingly, SOCS proteins have been shown to contribute to HSV immune evasion in multiple ways. SOCS3 is induced by HSV-1 in a number of cell lines, through a mechanism that involves the viral tegument proteins UL41 and UL13, and STAT3 (76,77). In cells which are capable of expressing SOCS3, HSV-1 is sufficient to inhibit IFN- induced STAT1 phosphorylation and downstream OAS production, and establish a rapidly propagating infection (76). However, in cells that do not express SOCS3 in response to HSV-1, or when SOCS3 expression is inhibited with STAT3 or SOCS3 antagonists, viral replication is attenuated. This attenuation is rescued following treatment with IFN- / neutralizing antibodies, indicating that HSV-1-induced SOCS3 is sufficient to inhibit antiviral type I IFN signaling, thereby enhancing viral replication (76). Notably, in addition to targeting IFN signaling, HSV-1- induced SOCS3 has also been shown to inhibit IFN- production. During a typical antiviral response, IFN- is initially expressed and induces the JAK/STAT-dependent production of IRF7. IRF7 then induces the expression of IFN- to amplify the innate immune response (59). Interestingly, viruses which are sufficient to induce SOCS3 also inhibit production of the IRF7 protein and downstream IFN- transcription, while viruses deficient in the SOCS3-inducing tegument proteins UL41 and UL13 do not (77). Collectively, these studies show that HSV-1- induced SOCS3 can inhibit both the production and signal transduction of antiviral type I IFNs by disrupting the JAK/STAT pathway. In addition to type I IFNs, type II IFNs are also critical 11
12 antiviral agents during HSV-1 infection which promote the establishment of latency (13). Importantly, type II IFNs are not sufficient to perform this function during the initial period of viral replication, or periodic outbreaks, in keratinocytes. HSV-1 has been shown to induce the expression of SOCS1 specifically in HEL-30 keratinocytes, but not other cells such as L929 fibroblasts (18). HSV-1-induced SOCS1 expression correlates with inhibition of IFN- -induced STAT1 activation and enhanced viral replication in keratinocytes. However, the antiviral effect of IFN- on HSV-1 replication is restored in the presence of a SOCS1 antagonist, indicating that the selective expression of HSV-1-induced SOCS1 in keratinocytes inhibits the antiviral and latency-promoting effects of type II IFNs. Notably, induction of either SOCS1 or SOCS3 by HSV-1 is cell type-specific. In the examples described above, the innate immune evasion provided by SOCS expression within a particular cell type was critical for defining the susceptibility of that cell to HSV-1 infection. Because the ability to evade innate immunity is required for establishing a productive viral infection, SOCS expression within a particular cell type has significant implications for the permissiveness of a wide range of cells to viral infection. This common theme will be explored in further examples below. Innate immune defense mechanisms present within cardiomyocytes of the heart are not driven by IFNs alone, but also by cardiotropin (CT)-1 (62), an IL-6 family member that signals through the JAK/STAT pathway following activation of the gp130 receptor subunit (Figure 2A). Coxsackie virus, a positive-sense ssrna enterovirus, has a propensity for infecting cardiomyocytes, often leading to severe inflammation of the heart tissue (myocarditis), heart damage (cardiomyopathy), and heart failure. Studies performed by Knowlton and colleagues, which show that coxsackie virus can induce the expression of SOCS1 and SOCS3, have implicated both of these SOCS proteins in the ability of coxsackie virus to evade antiviral 12
13 immune responses within the cardiomyocyte and establish a productive infection. Early studies focused on the role of SOCS1 in this process (75). SOCS1 was found to prevent cardiomyocyte protection by IFN-, IFN-, and CT-1 when overexpressed in vitro, and to attenuate STAT1 and STAT3 activation, as well as enhance viral replication, heart damage, and mortality when overexpressed in a cardiomyocyte-specific transgenic mouse in vivo. Furthermore, injection of a dominant negative (dn) SOCS1 construct into the heart prevented virus-induced heart damage. These studies suggested that coxsackie virus-induced SOCS1 could inhibit antiviral innate immunity initiated through either the IFN or gp130 receptors, thereby enhancing replication- 277 induced damage of cardiomyocytes. Later, studies focusing on SOCS3 provided a more complete understanding of innate immune protection in these cells (73). SOCS3 overexpression in a cardiomyocyte-specific transgenic mouse resulted in increased viral load, heart damage, and mortality following coxsackie virus infection. These results, which were similar to its SOCS1 counterpart, suggested that SOCS3 was also capable of dramatically inhibiting innate immunity. However, SOCS3 overexpression in vitro was only sufficient to prevent STAT3 phosphorylation and cardiomyocyte protection by CT-1, and not IFN- or IFN-, suggesting that CT-1 signaling through the gp130 receptor, rather than IFN signaling, is the critical mediator of innate immunity in cardiomyocytes. The authors further showed that CT-1 protects cardiomyocytes without suppressing viral replication, by preventing coxsackie virus-induced dystrophin cleavage and disruption of the cell membrane in a STAT3-dependent manner. These data indicate that SOCS1 and SOCS3 contribute to coxsackie virus immune evasion by attenuating JAK/STAT signaling through the gp130 receptor, and that virally inducted SOCS proteins are sufficient to inhibit even this alternative innate immune mechanism. 13
14 Additional viruses follow this well established pattern and induce various SOCS proteins for the purpose of inhibiting host innate immune defenses. Influenza A virus (47,52), RSV (24,79), and HBV (30,72) (Table 1), are just a few examples of viruses that benefit in this manner by exploiting SOCS protein function. It is likely that the discovery of others will be soon to follow Adaptive Immunity SOCS-mediated inhibition of the JAK/STAT pathway can also be used by viruses to evade the adaptive immune response. As described above, antibody production by B cells is a critical component of the comprehensive immune response to viral infection. For antibodies to be fully effective, they must be capable of neutralizing virus at portal sites of entry, including the respiratory, intestinal, and genital mucosa, in addition to the systemic circulation. To achieve this type of response, B cells must be competent to undergo class switch recombination to produce antibodies containing the multifunctional IgA, IgG, or IgE heavy chain, rather than the IgM heavy chain which has only limited potential. This switch requires T cell-dependent activation of B cells through the NF- B and JAK/STAT pathways (66). In the context of HIV-1 infection, however, the HIV-1 accessory protein Nef is secreted by infected cells and accumulates within B cells to upregulate inhibitors of these pathways (53). Nef-induced SOCS1 and SOCS3 inhibit IL-4-induced STAT6 activation (Figure 2A), and Nef-induced I B inhibits CD40L-induced NF- B activation, effectively preventing class switch recombination in B cells. Antibody production is also dependent on JAK/STAT signaling via IL-10 activation of STAT1 and STAT3 (Figure 2A). HIV-1 Nef-induced SOCS proteins are also sufficient to inhibit this pathway, thereby inhibiting both the quantity and efficacy of B cell generated antibodies. In 14
15 light of studies indicating that robust neutralizing antibody responses may be responsible for the dramatically slowed disease progression found in a select group of HIV-1 infected individuals, termed long-term non-progressors (7), this role of SOCS proteins is troubling. HCV-induced alterations in SOCS proteins also have a dramatic effect on lymphocytes of the adaptive immune system. Stimulation of CD8+ T cells with HCV core protein results in an increase in SOCS1 protein (74). This is accompanied by a decrease in STAT1 activation, markers of general T cell activation, and JAK/STAT-mediated IFN- production; a response which greatly impedes the ability of CTLs to effectively clear virus, and promotes chronic HCV infection. By contrast, HCV core protein decreases the levels of SOCS1 in B cells (39,74). This correlates with increased STAT1 activation and markers of general B cell activation, as well as increased B cell proliferation and antibody (IgM and IgG) production. While it may seem counterintuitive for a viral protein to promote enhancement of an immune cell compartment, the clonal B cell proliferation and antibody production that result are relatively ineffective for viral neutralization. Furthermore, this clonal expansion of B cell populations can also lead to important complications of chronic HCV infection, which are described further below SOCS Box-mediated Ubiquitination Promotes Intracellular Viral Trafficking To achieve successful viral replication, culminating in the production of progeny virus particles, viruses must move considerable distances within the host cell. Viruses must traverse the host cell membrane, transport their genomes to locations within the cytoplasm or nucleus for replication and transcription, and coordinate the movement of both their genome and translated viral proteins to specific locations at the plasma membrane for viral particle assembly, before finally budding from the cell surface. To achieve these complex movements, viruses hijack the 15
16 intrinsic transport machinery of the host cell, using the intracellular scaffold of actin microfilaments and microtubules, as well as associated kinesin and dynein motor complexes, to traffic their components throughout the cell and thereby facilitate their own propagation (41). While early studies suggested that viral components could interact directly with host transport machinery, more recent studies have indicated that additional host proteins are required to facilitate these interactions. Yamamoto and colleagues were the first to report that SOCS proteins play a role in promoting intracellular trafficking of viral proteins, and therefore viral egress from the host cell. They showed that HIV-1 infection could induce the expression of SOCS1 in T cells, which led to enhanced HIV-1 replication and virion production in the absence of enhanced HIV-1 transcription (56). Further analysis indicated that SOCS1 achieved this effect by enhancing microtubule stability and facilitating microtubule-dependent trafficking of the HIV-1 structural protein Gag to the plasma membrane for viral particle assembly (43). SOCS1 directly binds Gag via its SH2 domain, and ubiquitinates this viral substrate via its SOCS box, thereby promoting association of Gag with microtubules and enhancing its stability and trafficking. HIV-1 infection studies performed in the absence of SOCS1 yielded decreased Gag ubiquitination and Gag protein levels, as well as a decrease in subsequent particle release (43). Overexpression of SOCS3 in T cells was not sufficient to reproduce these results, indicating that the effects of SOCS1 on trafficking of viral proteins are family member-specific. Importantly, these reports describe the first SOCS-mediated enhancements of viral replication independent of immune evasion or JAK/STAT pathway inhibition. Although HIV-1 is the only virus for which this process has been well described, recent studies with Ebola virus have suggested that this may represent a more common mechanism. 16
17 360 Ebola virus is a negative-sense ssrna virus that leads to the development of Ebola hemorrhagic 361 fever. This virus disrupts both vascular integrity and effective coagulation by infecting endothelial cells and hepatocytes, respectively, resulting in the rapid leakage of blood through vessel walls and ultimately hypovolemic shock. Glycoproteins on the Ebola particle surface can activate signaling downstream of Toll-like receptor (TLR) 4, leading to the production of SOCS1 (45). Because TLR4 stimulation has previously been shown to increase Ebola virus production, and because ubiquitination of the Ebola structural protein VP40 has been shown to enhance virus budding, Harty and colleagues hypothesize that SOCS1 may ubiquitinate VP40, and thereby facilitate viral particle production. The authors cite preliminary results (not presented in the manuscript) which suggest that SOCS1 does ubiquitinate VP40, resulting in a modest increase in viral egress. Further studies will be needed to evaluate whether SOCS1-mediated ubiquitination plays a critical role in promoting intracellular trafficking, and therefore viral particle production, for Ebola or other viruses Effects of SOCS Expression on Viral Disease Susceptibility to Viral Infection When the host innate immune response is fully functional, a virus can not establish a productive infection. Therefore, to achieve successful infection in a particular host or cell type, viruses must be able to at least partially circumvent the IFN response in that environment. Differences in the ability of a given virus to overcome this response in various hosts or cell types 380 can contribute to differences in susceptibility to viral infection. Two of the most current examples of this phenomenon involve HIV-1. While HIV-1 readily infects human T cells and monocytes/macrophages, it is not capable of infecting cells of the rhesus macaque. Macaque 17
18 infection is prevented by TRIM5, an IFN-induced antiviral protein present in the macaque cell cytoplasm that recognizes a motif in the HIV-1 capsid and interferes with proper uncoating (68). Human cells contain a slightly different version of TRIM5 which is not capable of inhibiting HIV-1, whereas SIV is less sensitive to the macaque TRIM protein, thereby allowing each retrovirus to infect its preferred host without innate immune interference. The efficacy of apolipoprotein B mrna-editing enzyme catalytic polypeptide-like 3G (APOBEC3G), another IFN-inducible antiviral protein, can determine the cell type-specificity of HIV-1 infection (11). APOBEC3G can inhibit HIV-1 replication in three ways: 1) inhibition of reverse transcription, 2) induction of G-to-A hypermutation, and 3) inhibition of integration. However, the ability of APOBEC3G to perform these functions is highly dependent on its assembly state in a particular cell, with arrangement in low-molecular-mass (LMM) ribonucleoprotein complexes being permissive for full inhibitory function, and arrangement in high-molecular-mass (HMM) complexes being non-permissive. Consequently, resting CD4+ T cells in the peripheral blood which contain LMM complexes can not be productively infected, while the same cells in lymphoid tissues containing HMM complexes are readily infected by HIV-1 (12,31). Similarly, LMM complexes are converted to HMM complexes during monocyte differentiation to macrophages, which parallels an increase in HIV-1 infectivity (12). Clearly the ability of a virus to overcome host IFN responses is a critical predictor of permissiveness to infection. In this regard, the cell type-specificity of SOCS expression can determine susceptibility to infection. RSV, a negative sense, ssrna virus, is an interesting example of this phenomenon. RSV causes a severe respiratory tract infection in its host, targeting both pulmonary epithelial cells and macrophages. However, differences in the ability of RSV to productively infect cells of the monocyte/macrophage lineage have been tied to SOCS expression. While RSV can 18
19 induce the expression of SOCS1, SOCS3 and CIS, and establish a successful infection in macrophages (PMA-differentiated U937 cells), it is not capable of SOCS expression or infection in monocytes (undifferentiated U937 cells) (79). A subsequent report showed that knockdown of any of these SOCS proteins in cells susceptible to infection resulted in an increase in STAT1/2 phosphorylation and expression of the IFN-induced antiviral target gene OAS, as well as a correlating decrease in RSV replication (24). These studies suggest that SOCS expression contributes to the differences in permissiveness to RSV infection that accompany the cellular differentiation state of macrophages. HIV-1-induced SOCS expression has also been shown to play a role in permitting infection. Wahl and colleagues observed that tonsil mucosal associated lymphoid tissues are much more susceptible to HIV-1 infection than peripheral blood mononuclear cells (PBMCs), in part due to the increased levels of SOCS1 and SOCS3 present in these tissues (40). Increased SOCS expression in tonsil tissues, both constitutively and in response to HIV-1 infection, correlated with decreased STAT1 activation in response to IFN-, and a decreased T H 1 response 420 in the presence of IFN-. These studies suggest that SOCS expression may contribute to determining susceptibility to HIV-1 infection by inhibiting both the innate and adaptive immune responses to infection. In addition, studies from our lab show that HIV-1 Tat-induced SOCS3 expression is selective for cells of macrophage lineage in comparison to other CNS cells examined, such as astrocytes and neurons (2). The mechanism behind this specificity requires further study, but may involve differential surface receptor expression or pathway activation between cell types. It is intriguing to note, however, that the cell types in which Tat is able to induce SOCS3, and thereby overcome the innate antiviral response, are synonomous with those able to support a productive infection of HIV-1 within the brain (i.e. macrophages and 19
20 microglia). Although further studies are required in this area, it is enticing to speculate that Tat- induced SOCS3 may contribute to determining permissiveness for HIV-1 infection in the brain Virally Associated Diseases SOCS proteins are induced by viruses to promote the successful completion of their own 434 life cycle. However, aberrant SOCS expression can also powerfully shape the overall presentation of viral disease by contributing to the peripheral manifestations of viral infection. For example, while the direct result of HIV-1 replication is CD4+ T cell death, the most well recognized manifestation of HIV-1 disease may be infection with opportunistic pathogens such as mycobacteria, protozoa, and fungus. These pathogens are cleared in the immunocompetent host primarily by T cell-mediated IFN- signaling. While decreased T cell numbers in the HIV- 1-infected host certainly hinder proper clearance, it has been shown that even exogenously supplied IFN- is not fully effective (4), suggesting that nonproductive signaling is also at fault. SOCS1 and SOCS3 expression during Mycobacterium avium infection has been shown to correlate with decreased STAT1 activation and failed clearance following IFN- treatment (69). It seems likely that HIV-1-induced SOCS1 and SOCS3 proteins function similarly during coinfection to provide a safe haven for M. avium by inhibiting IFN- signaling. Furthermore, HIV- 1 Tat-induced SOCS2 has also been reported to inhibit IFN- signaling in monocytes (10). Cheng et al. recently showed that HIV-1 Tat is sufficient to dampen both STAT1 activation and downstream target gene expression in response to IFN-, but that this protective immune response is restored following knockdown of SOCS2. Although the authors of this study did not evaluate the effect of Tat-induced SOCS2 on pathogen clearance, these data suggest that multiple 20
21 HIV-1-induced SOCS proteins may promote secondary infections by inhibiting the antipathogenic effects of IFN-. Patients with chronic HCV infection experience multiple associated diseases, several of which have been linked to SOCS protein expression. A propensity toward glucose intolerance and type II diabetes mellitus has been documented (37,38), which is thought to result from reduced expression of insulin receptor substrates (IRS) 1 and 2 during chronic HCV disease (27). White and colleagues have shown that overexpressed SOCS1 and SOCS3 are capable of binding both IRS proteins, and targeting them for ubiquitination and proteasomal degradation in a SOCS box-dependent manner (54). Furthermore, liver-specific overexpression of SOCS1 in a mouse model in vivo also reduced IRS1 and 2 protein levels, and subsequently led to insulin resistance (54). A later report provided evidence that overexpression of the HCV core protein, either in vitro or in vivo, could promote a similar decrease in IRS1 and 2 expression, but only following the induction of SOCS3 (27). These data suggest that HCV-induced SOCS3 can target critical IRS proteins for proteasomal degradation, thereby contributing to the glucose intolerance characteristic of HCV infection. Recent studies demonstrate that HBV-induced SOCS3 can also mediate the ubiquitination of IRS1 and thereby inhibit insulin signaling (28). Although limited, these results suggest that SOCS-induced dysfunction of insulin signaling may be a broader phenomenon. While most reports have focused on HCV-induced increases in SOCS protein expression, several have shown that HCV-related decreases in SOCS proteins within specific cell types or tissue regions may also be associated with disease. A major extrahepatic manifestation of HCV infection affecting 30-50% of patients is mixed cryoglobulinaemia, which is caused by rampant overproduction of monoclonal or polycolonal IgM and IgG antibodies by B cells (8). Another 21
22 associated disease results directly from aberrant B cell proliferation, known as non-hodgkin lymphoma (70). As previously mentioned, SOCS1 is decreased in B cells taken from HCVinfected patients (39) or those incubated with HCV core protein in vitro (74). Decreased SOCS1 expression correlates with increased STAT1 activation, and with increases in both B cell proliferation and IgM/IgG production. Therefore, HCV-induced decreases in SOCS1 expression in B cells may promote B cell dysregulation and clonal expansion, leading to the lymphoproliferative disorders characteristic of HCV infection. Patients with chronic HCV infection also display a propensity toward hepatocellular carcinoma (HCC) (33). Interestingly, while HCV infection has been shown in a number of studies to increase SOCS1 and SOCS3 expression in hepatocytes to promote viral replication, HCV-induced decreases in these SOCS proteins tend to correlate with hepatocarcinogenesis. A report by Yoshimura and colleagues provides some resolution to this conflict by establishing that SOCS3 levels are increased in the livers of patients with HCV infection when compared to normal controls, but that within HCV-infected livers, hepatocytes in regional areas of HCC contain drastically reduced levels of SOCS3 compared to non-hcc areas (44). The lower levels of SOCS3 in HCC liver regions correlate with increased STAT3 activation, which may lead to enhanced expression of anti-apoptotic molecules such as Bcl-2 and Bcl-XL and promote cancer development. Further studies will be required to determine the cause of differential regulation of SOCS proteins by HCV in different regions of the liver Therapeutic Potential Because the exploitation of host SOCS protein functions provides such a potent viral advantage, these proteins also possess considerable therapeutic potential. One of the most 22
23 straightforward therapeutic applications of SOCS proteins has been shown in the context of chronic HCV infection. The most common treatment for HCV infection is the exogenous administration of IFN- (17,19), given to augment the endogenous antiviral host response. However, a significant proportion of patients, particularly those with genotype 1 HCV infection, do not respond to therapy (17,19). Multiple studies have shown that HCV-induced SOCS3 expression, which is known to inhibit endogenous host IFN responses, can also be used as an independent predictor of therapeutic response to IFN- treatment (29,49,50). SOCS3 expression is elevated in non-responders when compared to responders both in peripheral blood mononuclear cells (49) and hepatic tissue (29). These data also provide evidence that differences in HCV-induced SOCS3 expression may be dependent on both viral and host factors. Patients with genotype 1 HCV infection tend to have higher levels of SOCS3 expression, providing a rationale for their propensity toward lack of therapeutic response (29,49). In addition, patients with a particular SOCS3 genotype (-4874 AA) also express elevated SOCS3 levels and consequently have a poorer response to therapy (50). Not all studies agree, however, as one report suggests that elevated SOCS1, and not SOCS3, in the liver of HCV-infected patients provides a better prediction of response to viral therapy (26). Regardless, analyzing the levels of SOCS protein expression in the HCV-infected patient may allow for early prediction of therapeutic response, and therefore more appropriate treatment decisions. This correlation may also provide the information necessary for developing a more effective treatment in the future. One therapeutic strategy to consider would be suppression of SOCS protein levels or function during viral infection. The effectiveness of this approach in dampening viral replication has been explored in a small number of studies. A dnsocs1 construct containing a point mutation in the KIR (F59D), previously shown to cause the destabilization of both SOCS1 and 23
24 SOCS3 (23), was evaluated in the treatment of coxsackie virus-infection (75). In an in vitro reporter assay, dnsocs1 enhanced heart protective CT-1-induced STAT3 activity, and 522 overcame STAT3 inhibition in response to SOCS1 overexpression, in cardiomyocytes. In addition, it was able to sustain STAT1 and STAT3 phosphorylation in response to the innate antiviral agents IFN- and CT-1, respectively. In vivo, injection of dnsocs1 into the hearts of coxsackie virus-infected mice attenuated both virus replication and cardiomyocyte damage. These studies suggest that dnsocs1, by reducing levels of coxsackie virus-induced SOCS1 and SOCS3, allows endogenous antiviral JAK/STAT signaling to protect the heart from viral infection. Another SOCS1 antagonist, a peptide mimic of the phosphorylated JAK2 activation loop (pjak2), exhibits even broader antiviral activity. By binding the KIR of SOCS1 and preventing its function, pjak2 has been shown to reverse the inhibition of SOCS1 overexpression on IL-6-induced STAT3 phosphorylation, and to enhance reporter assay activity at the promoter of antiviral target genes in response to IFN- (71). Keratinocytes, which are highly susceptible to HSV-1 infection due to virus-induced SOCS1 expression, are protected from HSV-1-induced death following pretreatment with pjak2 (18). Treatment with pjak2 alone enhances keratinocyte survival only modestly, while co-treatment with IFN- results in complete protection. These data indicate that pjak2 is sufficient to attenuate SOCS1 function, thereby allowing either endogenous or exogenous JAK/STAT signaling pathways to exert their antiviral effects, and promote keratinocyte survival. In a similar manner, pretreatment of mice with pjak2 also provides protection against lethal vaccinia and encephalomyocarditis virus infection (1). While these examples are limited, and preliminary, they suggest that manipulation of SOCS proteins is possible during viral infection, and may provide a potent mechanism for inhibiting viral replication. 24
25 543 Enhanced expression of SOCS proteins during viral infection may actually be favorable 544 for one therapeutic strategy. Oncolytic HSVs (ohsvs) are robust anticancer therapeutics engineered to infect and replicate within tumor cells, thereby reducing tumor burden through direct cell lysis. A recent study has shown that the ability of the ohsv vector G207 to replicate within target cells, and thereby have a therapeutic effect, is dependent on its induction of SOCS1 (34). ohsv induced SOCS1 and inhibited STAT1 activation in cells permissive for replication, but not in cells where replication was poor. SOCS1 knockdown in ohsv-permissive cells resulted in a >10 fold decrease in viral replication. Therefore, ohsv-induced expression of SOCS1 is critical for susceptibility of cells to viral replication and, consequently, therapeutic effect. Exogenous overexpression of SOCS1 by therapeutic viral vectors has been proposed in order to inhibit the host innate immune response (57). This type of strategy may enhance the range and efficacy of viral vectors in the future. Manipulation of SOCS proteins may even be used to prevent viral infection entirely. Currently utilized vaccine strategies have been unable to produce a meaningful immune response to vaccination with HIV-1 antigens (3). One obstacle is that endogenous SOCS1 expression in host dendritic cells (DCs) inhibits effective antigen presentation to T and B cells, thereby limiting immune activation (22,61). In a series of experiments comparing mice immunized with either normal or SOCS1-depleted DCs carrying the HIV-1 envelope protein gp120, Chen and colleagues showed that DC function is enhanced in the absence of SOCS1, allowing for a more robust cellular and humoral response to HIV-1 antigens (64). Immunization with SOCS1- depleted DCs resulted in more robust CTL activity against gp120-pulsed target cells, along with increased numbers of IFN- - and perforin-positive CD8+ T cells. Gp120-specific CD4+ T cells were also present in greater numbers in vivo, and exhibited increased proliferation, and increased 25
26 production of both T H 1 and T H 2 polarizing cytokines following interaction with gp120-pulsed DCs ex vivo. The HIV-1-specific humoral response was also enhanced. The number of gp120- specific IgG producing B cells was increased, as were markers of their activation state, and their production of gp120-specific antibodies. In addition, mice immunized with SOCS1-depleted DCs generated a much longer lasting immune response, exhibiting drastically superior gp120- specific CTL and antibody responses six months after immunization as compared to mice immunized with SOCS1-containing DCs. Together, these data indicate that SOCS1 expression in DCs critically regulates anti-hiv immunity, and that an enhanced, but balanced, cellular and humoral response can be generated against HIV-1 antigens in the absence of SOCS1. In light of these results, the authors further examined whether the administration of SOCS1 sirna expressor DNA (psuper-socs1-sirna) could enhance the potency of a co-administered HIV-1 DNA vaccine. Co-administration of SOCS1 sirna increased gp120-specific antibody titers, as well as CTL and CD4+ T cell responses, suggesting that limiting SOCS1 expression may be an effective adjunct to current vaccine development strategies Conclusions Virus-induced SOCS proteins represent a powerful tool for viral survival. They can mediate inhibition of the JAK/STAT pathway, allowing viruses to evade the host immune response, or viral protein ubiquitination, facilitating protein trafficking and progeny virus 585 assembly. Virally enhanced expression of SOCS proteins also has a profound role in determining the presentation of viral disease, due its peripheral effects on the host organism. Understanding the role of SOCS proteins in the context of viral diseases will provide us not only 26
27 with a better understanding of the complexities of virus-host interactions, but also with potentially new therapeutic targets
28 Acknowledgments This work was funded in part by Public Health Service grants NS and NS from NINDS, a grant from the National Multiple Sclerosis Society (RG 3892-A-12), and a Collaborative MS Research Center Award from the NMSS (CA 1059-A-13). L.N.A. is supported by the UAB Medical Scientist Training Program and by Public Health Service grants T32-AI and F30-NS
29 References 1. Ahmed, C. M., R. Dabelic, J. P. Martin, L. D. Jager, S. M. Haider, and H. M. Johnson Enhancement of antiviral immunity by small molecule antagonist of suppressor of cytokine signaling. J. Immunol. 185: Akhtar, L. N., H. Qin, M. T. Muldowney, L. L. Yanagisawa, O. Kutsch, J. E. Clements, and E. N. Benveniste Suppressor of Cytokine Signaling 3 inhibits antiviral IFN-{beta} signaling to enhance HIV-1 replication in macrophages. J. Immunol. 185: Bansal, G. P., A. Malaspina, and J. Flores Future paths for HIV vaccine research: Exploiting results from recent clinical trials and current scientific advances. Curr. Opin. Mol. Ther. 12: Biggs, B. A., M. Hewish, S. Kent, K. Hayes, and S. M. Crowe HIV-1 infection of human macrophages impairs phagocytosis and killing of Toxoplasma gondii. J. Immunol. 154: Bode, J. G., E. D. Brenndorfer, and D. Haussinger Subversion of innate host antiviral strategies by the hepatitis C virus. Arch. Biochem. Biophys. 462: Bode, J. G., S. Ludwig, C. Ehrhardt, U. Albrecht, A. Erhardt, F. Schaper, P. C. Heinrich, and D. Haussinger IFN-alpha antagonistic activity of HCV core protein involves induction of suppressor of cytokine signaling-3. FASEB J. 17: Cecilia, D., C. Kleeberger, A. Munoz, J. V. Giorgi, and S. Zolla-Pazner A longitudinal study of neutralizing antibodies and disease progression in HIV-1-infected subjects. J. Infect. Dis. 179: Charles, E. D. and L. B. Dustin Hepatitis C virus-induced cryoglobulinemia. Kidney Int. 76: Chen, X. P., J. A. Losman, S. Cowan, E. Donahue, S. Fay, B. Q. Vuong, M. C. Nawijn, D. Capece, V. L. Cohan, and P. Rothman Pim serine/threonine kinases regulate the stability of Socs-1 protein. Proc. Natl. Acad. Sci. U. S. A 99: Cheng, S. M., J. C. Li, S. S. Lin, D. C. Lee, L. Liu, Z. Chen, and A. S. Lau HIV-1 transactivator protein induction of suppressor of cytokine signaling-2 contributes to dysregulation of IFN{gamma} signaling. Blood 113: Chiu, Y. L. and W. C. Greene The APOBEC3 cytidine deaminases: an innate defensive network opposing exogenous retroviruses and endogenous retroelements. Annu. Rev. Immunol. 26:
30 Chiu, Y. L., V. B. Soros, J. F. Kreisberg, K. Stopak, W. Yonemoto, and W. C. Greene Cellular APOBEC3G restricts HIV-1 infection in resting CD4+ T cells. Nature 435: Decman, V., P. R. Kinchington, S. A. Harvey, and R. L. Hendricks Gamma interferon can block herpes simplex virus type 1 reactivation from latency, even in the presence of late gene expression. J. Virol. 79: Dimitriou, I. D., L. Clemenza, A. J. Scotter, G. Chen, F. M. Guerra, and R. Rottapel Putting out the fire: coordinated suppression of the innate and adaptive immune systems by SOCS1 and SOCS3 proteins. Immunol. Rev. 224: Ehlting, C., W. S. Lai, F. Schaper, E. D. Brenndorfer, R. J. Matthes, P. C. Heinrich, S. Ludwig, P. J. Blackshear, M. Gaestel, D. Haussinger, and J. G. Bode Regulation of suppressor of cytokine signaling 3 (SOCS3) mrna stability by TNF-alpha involves activation of the MKK6/p38MAPK/MK2 cascade. J. Immunol. 178: Fenner, J. E., R. Starr, A. L. Cornish, J. G. Zhang, D. Metcalf, R. D. Schreiber, K. Sheehan, D. J. Hilton, W. S. Alexander, and P. J. Hertzog Suppressor of cytokine signaling 1 regulates the immune response to infection by a unique inhibition of type I interferon activity. Nat. Immunol. 7: Forde, K. A. and K. R. Reddy Hepatitis C virus infection and immunomodulatory therapies. Clin. Liver Dis. 13: Frey, K. G., C. M. Ahmed, R. Dabelic, L. D. Jager, E. N. Noon-Song, S. M. Haider, H. M. Johnson, and N. J. Bigley HSV-1-induced SOCS-1 expression in keratinocytes: use of a SOCS-1 antagonist to block a novel mechanism of viral immune evasion. J. Immunol. 183: Fried, M. W., M. L. Shiffman, K. R. Reddy, C. Smith, G. Marinos, F. L. Goncales, Jr., D. Haussinger, M. Diago, G. Carosi, D. Dhumeaux, A. Craxi, A. Lin, J. Hoffman, and J. Yu Peginterferon alfa-2a plus ribavirin for chronic hepatitis C virus infection. N. Engl. J. Med. 347: Frobose, H., S. G. Ronn, P. E. Heding, H. Mendoza, P. Cohen, T. Mandrup-Poulsen, and N. Billestrup Suppressor of Cytokine Signaling-3 inhibits interleukin-1 signaling by targeting the TRAF-6/TAK1 complex. Mol. Endocrinol. 20: Haan, S., P. Ferguson, U. Sommer, M. Hiremath, D. W. McVicar, P. C. Heinrich, J. A. Johnston, and N. A. Cacalano Tyrosine phosphorylation disrupts elongin interaction and accelerates SOCS3 degradation. J. Biol. Chem. 278: Hanada, T., K. Tanaka, Y. Matsumura, M. Yamauchi, H. Nishinakamura, H. Aburatani, R. Mashima, M. Kubo, T. Kobayashi, and A. Yoshimura Induction of hyper Th1 cell-type immune responses by dendritic cells lacking the suppressor of cytokine signaling-1 gene. J. Immunol. 174:
31 Hanada, T., T. Yoshida, I. Kinjyo, S. Minoguchi, H. Yasukawa, S. Kato, H. Mimata, Y. Nomura, Y. Seki, M. Kubo, and A. Yoshimura A mutant form of JAB/SOCS1 augments the cytokine-induced JAK/STAT pathway by accelerating degradation of wild-type JAB/CIS family proteins through the SOCS-box. J. Biol. Chem. 276: Hashimoto, K., K. Ishibashi, K. Ishioka, D. Zhao, M. Sato, S. Ohara, Y. Abe, Y. Kawasaki, Y. Sato, S. Yokota, N. Fujii, R. S. Peebles, Jr., M. Hosoya, and T. Suzutani RSV replication is attenuated by counteracting expression of the suppressor of cytokine signaling (SOCS) molecules. Virology 391: He, Y., W. Zhang, R. Zhang, H. Zhang, and W. Min SOCS1 inhibits tumor necrosis factor-induced activation of ASK1-JNK inflammatory signaling by mediating ASK1 degradation. J. Biol. Chem. 281: Imanaka, K., S. Tamura, K. Fukui, N. Ito, S. Kiso, Y. Imai, T. Naka, T. Kishimoto, S. Kawata, and Y. Shinomura Enhanced expression of suppressor of cytokine signalling-1 in the liver of chronic hepatitis C: possible involvement in resistance to interferon therapy. J. Viral Hepat. 12: Kawaguchi, T., T. Yoshida, M. Harada, T. Hisamoto, Y. Nagao, T. Ide, E. Taniguchi, H. Kumemura, S. Hanada, M. Maeyama, S. Baba, H. Koga, R. Kumashiro, T. Ueno, H. Ogata, A. Yoshimura, and M. Sata Hepatitis C virus down-regulates insulin receptor substrates 1 and 2 through up-regulation of suppressor of cytokine signaling 3. Am. J. Pathol. 165: Kim, K., K. H. Kim, and J. Cheong Hepatitis B virus X protein impairs hepatic insulin signaling through degradation of IRS1 and induction of SOCS3. PLoS. One. 5:e Kim, K. A., W. Lin, A. W. Tai, R. X. Shao, E. Weinberg, C. B. De Sa Borges, A. K. Bhan, H. Zheng, Y. Kamegaya, and R. T. Chung Hepatic SOCS3 expression is strongly associated with non-response to therapy and race in HCV and HCV/HIV infection. J. Hepatol. 50: Koeberlein, B., H. A. zur, N. Bektas, H. Zentgraf, R. Chin, L. T. Nguyen, R. Kandolf, J. Torresi, and C. T. Bock Hepatitis B virus overexpresses suppressor of cytokine signaling-3 (SOCS3) thereby contributing to severity of inflammation in the liver. Virus Res. 148: Kreisberg, J. F., W. Yonemoto, and W. C. Greene Endogenous factors enhance HIV infection of tissue naive CD4 T cells by stimulating high molecular mass APOBEC3G complex formation. J. Exp. Med. 203: Kubo, M., T. Hanada, and A. Yoshimura Suppressors of cytokine signaling and immunity. Nat. Immunol. 4:
32 Levrero, M Viral hepatitis and liver cancer: the case of hepatitis C. Oncogene 25: Mahller, Y. Y., B. Sakthivel, W. H. Baird, B. J. Aronow, Y. H. Hsu, T. P. Cripe, and R. Mehrian-Shai Molecular analysis of human cancer cells infected by an oncolytic HSV-1 reveals multiple upregulated cellular genes and a role for SOCS1 in virus replication. Cancer Gene Ther. 15: Malim, M. H. and M. Emerman HIV-1 accessory proteins--ensuring viral survival in a hostile environment. Cell Host. Microbe 3: Mansell, A., R. Smith, S. L. Doyle, P. Gray, J. E. Fenner, P. J. Crack, S. E. Nicholson, D. J. Hilton, L. A. O'Neill, and P. J. Hertzog Suppressor of cytokine signaling 1 negatively regulates Toll-like receptor signaling by mediating Mal degradation. Nat. Immunol. 7: Mehta, S. H., F. L. Brancati, S. A. Strathdee, J. S. Pankow, D. Netski, J. Coresh, M. Szklo, and D. L. Thomas Hepatitis C virus infection and incident of type 2 diabetes. Hepatology 38: Mehta, S. H., F. L. Brancati, M. S. Sulkowski, S. A. Strathdee, M. Szklo, and D. L. Thomas Prevalence of type 2 diabetes mellitus among persons with hepatitis C virus infection in the United States. Ann. Intern. Med. 133: Moorman, J., Z. P. Dong, L. Ni, C. Zhang, T. Borthwick, and Z. Q. Yao Abnormal B-cell activation associated with TALL-1 over-expression and SOCS-1 suppression during chronic hepatitis C virus infection. Immunology 128: Moutsopoulos, N. M., N. Vazquez, T. Greenwell-Wild, I. Ecevit, J. Horn, J. Orenstein, and S. M. Wahl Regulation of the tonsil cytokine milieu favors HIV susceptibility. J. Leukoc. Biol. 80: Naghavi, M. H. and S. P. Goff Retroviral proteins that interact with the host cell cytoskeleton. Curr. Opin. Immunol. 19: Nakagawa, R., T. Naka, H. Tsutsui, M. Fujimoto, A. Kimura, T. Abe, E. Seki, S. Sato, O. Takeuchi, K. Takeda, S. Akira, K. Yamanishi, I. Kawase, K. Nakanishi, and T. Kishimoto SOCS-1 participates in negative regulation of LPS responses. Immunity. 17: Nishi, M., A. Ryo, N. Tsurutani, K. Ohba, T. Sawasaki, R. Morishita, K. Perrem, I. Aoki, Y. Morikawa, and N. Yamamoto Requirement for microtubule integrity in the SOCS1-mediated intracellular dynamics of HIV-1 Gag. FEBS Lett. 583: Ogata, H., T. Kobayashi, T. Chinen, H. Takaki, T. Sanada, Y. Minoda, K. Koga, G. Takaesu, Y. Maehara, M. Iida, and A. Yoshimura Deletion of the SOCS3 gene in liver parenchymal cells promotes hepatitis-induced hepatocarcinogenesis. Gastroenterology 131:
33 Okumura, A., P. M. Pitha, A. Yoshimura, and R. N. Harty Interaction between Ebola virus glycoprotein and host toll-like receptor 4 leads to induction of proinflammatory cytokines and SOCS1. J. Virol. 84: Paladino, P. and K. L. Mossman Mechanisms employed by herpes simplex virus 1 to inhibit the interferon response. J. Interferon Cytokine Res. 29: Pauli, E. K., M. Schmolke, T. Wolff, D. Viemann, J. Roth, J. G. Bode, and S. Ludwig Influenza A virus inhibits type I IFN signaling via NF-kappaB-dependent induction of SOCS-3 expression. PLoS. Pathog. 4:e Peltola, K. J., K. Paukku, T. L. Aho, M. Ruuska, O. Silvennoinen, and P. J. Koskinen Pim-1 kinase inhibits STAT5-dependent transcription via its interactions with SOCS1 and SOCS3. Blood 103: Persico, M., M. Capasso, E. Persico, M. Svelto, R. Russo, D. Spano, L. Croce, M. La, V, F. Moschella, F. Masutti, R. Torella, C. Tiribelli, and A. Iolascon Suppressor of cytokine signaling 3 (SOCS3) expression and hepatitis C virus-related chronic hepatitis: Insulin resistance and response to antiviral therapy. Hepatology 46: Persico, M., M. Capasso, R. Russo, E. Persico, L. Croce, C. Tiribelli, and A. Iolascon Elevated expression and polymorphisms of SOCS3 influence patient response to antiviral therapy in chronic hepatitis C. Gut 57: Piessevaux, J., D. Lavens, F. Peelman, and J. Tavernier The many faces of the SOCS box. Cytokine Growth Factor Rev. 19: Pothlichet, J., M. Chignard, and M. Si-Tahar Cutting edge: innate immune response triggered by influenza A virus is negatively regulated by SOCS1 and SOCS3 through a RIG-I/IFNAR1-dependent pathway. J. Immunol. 180: Qiao, X., B. He, A. Chiu, D. M. Knowles, A. Chadburn, and A. Cerutti Human immunodeficiency virus 1 Nef suppresses CD40-dependent immunoglobulin class switching in bystander B cells. Nat. Immunol. 7: Rui, L., M. Yuan, D. Frantz, S. Shoelson, and M. F. White SOCS-1 and SOCS- 3 block insulin signaling by ubiquitin-mediated degradation of IRS1 and IRS2. J. Biol. Chem. 277: Ryo, A., F. Suizu, Y. Yoshida, K. Perrem, Y. C. Liou, G. Wulf, R. Rottapel, S. Yamaoka, and K. P. Lu Regulation of NF-kappaB signaling by Pin1-dependent prolyl isomerization and ubiquitin-mediated proteolysis of p65/rela. Mol. Cell 12: Ryo, A., N. Tsurutani, K. Ohba, R. Kimura, J. Komano, M. Nishi, H. Soeda, S. Hattori, K. Perrem, M. Yamamoto, J. Chiba, J. Mimaya, K. Yoshimura, S. Matsushita, M. Honda, A. Yoshimura, T. Sawasaki, I. Aoki, Y. Morikawa, and N. 33
34 Yamamoto SOCS1 is an inducible host factor during HIV-1 infection and regulates the intracellular trafficking and stability of HIV-1 Gag. Proc. Natl. Acad. Sci. U. S. A 105: Sakurai, H., K. Tashiro, K. Kawabata, T. Yamaguchi, F. Sakurai, S. Nakagawa, and H. Mizuguchi Adenoviral expression of suppressor of cytokine signaling-1 reduces adenovirus vector-induced innate immune responses. J. Immunol. 180: Sasaki, A., K. Inagaki-Ohara, T. Yoshida, A. Yamanaka, M. Sasaki, H. Yasukawa, A. E. Koromilas, and A. Yoshimura The N-terminal truncated isoform of SOCS3 translated from an alternative initiation AUG codon under stress conditions is stable due to the lack of a major ubiquitination site, Lys-6. J. Biol. Chem. 278: Sato, M., H. Suemori, N. Hata, M. Asagiri, K. Ogasawara, K. Nakao, T. Nakaya, M. Katsuki, S. Noguchi, N. Tanaka, and T. Taniguchi Distinct and essential roles of transcription factors IRF-3 and IRF-7 in response to viruses for IFN-alpha/beta gene induction. Immunity. 13: Shao, R. X., L. Zhang, L. F. Peng, E. Sun, W. J. Chung, J. Y. Jang, W. L. Tsai, G. Hyppolite, and R. T. Chung Suppressor of cytokine signaling 3 suppresses hepatitis C virus replication in an mtor-dependent manner. J. Virol. 84: Shen, L., K. Evel-Kabler, R. Strube, and S. Y. Chen Silencing of SOCS1 enhances antigen presentation by dendritic cells and antigen-specific anti-tumor immunity. Nat. Biotechnol. 22: Sheng, Z., D. Pennica, W. I. Wood, and K. R. Chien Cardiotrophin-1 displays early expression in the murine heart tube and promotes cardiac myocyte survival. Development 122: Song, M. M. and K. Shuai The suppressor of cytokine signaling (SOCS) 1 and SOCS3 but not SOCS2 proteins inhibit interferon-mediated antiviral and antiproliferative activities. J. Biol. Chem. 273: Song, X. T., K. Evel-Kabler, L. Rollins, M. Aldrich, F. Gao, X. F. Huang, and S. Y. Chen An alternative and effective HIV vaccination approach based on inhibition of antigen presentation attenuators in dendritic cells. PLoS. Med. 3:e Stark, G. R., I. M. Kerr, B. R. Williams, R. H. Silverman, and R. D. Schreiber How cells respond to interferons. Annu. Rev. Biochem. 67: Stavnezer, J., J. E. Guikema, and C. E. Schrader Mechanism and regulation of class switch recombination. Annu. Rev. Immunol. 26: Strauss, J. H. and E. G. Strauss Host defense against viral infection and viral counterdefenses, p In J. H. Strauss and E. G. Strauss (ed.), Viruses and human disease. Academic Press, San Diego. 34
35 Stremlau, M., C. M. Owens, M. J. Perron, M. Kiessling, P. Autissier, and J. Sodroski The cytoplasmic body component TRIM5alpha restricts HIV-1 infection in Old World monkeys. Nature 427: Vazquez, N., T. Greenwell-Wild, S. Rekka, J. M. Orenstein, and S. M. Wahl Mycobacterium avium-induced SOCS contributes to resistance to IFN-gamma-mediated mycobactericidal activity in human macrophages. J. Leukoc. Biol. 80: Viswanatha, D. S. and A. Dogan Hepatitis C virus and lymphoma. J. Clin. Pathol. 60: Waiboci, L. W., C. M. Ahmed, M. G. Mujtaba, L. O. Flowers, J. P. Martin, M. I. Haider, and H. M. Johnson Both the suppressor of cytokine signaling 1 (SOCS- 1) kinase inhibitory region and SOCS-1 mimetic bind to JAK2 autophosphorylation site: implications for the development of a SOCS-1 antagonist. J. Immunol. 178: Xu, Y., Y. Hu, B. Shi, X. Zhang, J. Wang, Z. Zhang, F. Shen, Q. Zhang, S. Sun, and Z. Yuan HBsAg inhibits TLR9-mediated activation and IFN-alpha production in plasmacytoid dendritic cells. Mol. Immunol. 46: Yajima, T., H. Yasukawa, E. S. Jeon, D. Xiong, A. Dorner, M. Iwatate, M. Nara, H. Zhou, D. Summers-Torres, M. Hoshijima, K. R. Chien, A. Yoshimura, and K. U. Knowlton Innate defense mechanism against virus infection within the cardiac myocyte requiring gp130-stat3 signaling. Circulation 114: Yao, Z. Q., D. Prayther, C. Trabue, Z. P. Dong, and J. Moorman Differential regulation of SOCS-1 signalling in B and T lymphocytes by hepatitis C virus core protein. Immunology 125: Yasukawa, H., T. Yajima, H. Duplain, M. Iwatate, M. Kido, M. Hoshijima, M. D. Weitzman, T. Nakamura, S. Woodard, D. Xiong, A. Yoshimura, K. R. Chien, and K. U. Knowlton The suppressor of cytokine signaling-1 (SOCS1) is a novel therapeutic target for enterovirus-induced cardiac injury. J. Clin. Invest 111: Yokota, S., N. Yokosawa, T. Okabayashi, T. Suzutani, and N. Fujii Induction of suppressor of cytokine signaling-3 by herpes simplex virus type 1 confers efficient viral replication. Virology 338: Yokota, S., N. Yokosawa, T. Okabayashi, T. Suzutani, S. Miura, K. Jimbow, and N. Fujii Induction of suppressor of cytokine signaling-3 by herpes simplex virus type 1 contributes to inhibition of the interferon signaling pathway. J. Virol. 78: Yoshimura, A., T. Naka, and M. Kubo SOCS proteins, cytokine signalling and immune regulation. Nat. Rev. Immunol. 7: Zhao, D. C., T. Yan, L. Li, S. You, and C. Zhang Respiratory syncytial virus inhibits interferon-alpha-inducible signaling in macrophage-like U937 cells. J. Infect. 54:
36 Figure Legends Figure 1. SOCS Protein Structure. All SOCS proteins contain a central SH2 domain and a C- terminal SOCS box. The SH2 domain determines the target of each SOCS protein by binding specific phosphorylated (P) tyrosine residues on its preferred substrate (commonly JAK proteins). The SOCS box interacts with ubiquitinating machinery including Elongin B (EB), Elongin C (EC), Cullin5, RING-box-2 (Rbx2), and an E2 ubiquitin conjugating enzyme. By bringing the bound substrate into proximity with ubiquitinating machinery, the SOCS protein facilitates ubiquitination (U) of target proteins, thereby marking them for degradation by the proteosome. Certain SOCS proteins (SOCS1 and SOCS3) also contain an N-terminal kinase inhibitory region (KIR). The KIR is thought to function as a pseudosubstrate to inhibit the kinase activity of proteins that are bound by, or in close proximity to, the SOCS protein. Figure 2. SOCS Proteins Negatively Regulate the JAK/STAT Pathway. (A) Composition of selected JAK/STAT pathways. (B) Function of JAK/STAT pathway. Cytokine stimulation of cell surface receptors activates receptor-associated JAK proteins by phosphorylation (P). Activated JAKs phosphorylate receptor cytoplasmic domains (1) which leads to the recruitment of cytoplasmic STAT proteins (2). Recruited STATs are activated by JAK phosphorylation (3), allowing them to dimerize (4), and enter the nucleus as a transcription factor complex to induce the expression of target genes (5). Gene targets often include both immune effectors and SOCS proteins. (C) SOCS proteins. SOCS proteins negatively regulate this pathway by binding specific substrates within the JAK/STAT receptor complex and terminating pathway activation in the following ways: competition with recruited STAT proteins for shared phosphotyrosine residues (1), kinase inhibitory region (KIR)-mediated inhibition of JAK activity (2), or SOCS box-mediated ubiquitination and degradation of bound receptor components (3). 36
37
38
39
Fayth K. Yoshimura, Ph.D. September 7, of 7 HIV - BASIC PROPERTIES
 1 of 7 I. Viral Origin. A. Retrovirus - animal lentiviruses. HIV - BASIC PROPERTIES 1. HIV is a member of the Retrovirus family and more specifically it is a member of the Lentivirus genus of this family.
1 of 7 I. Viral Origin. A. Retrovirus - animal lentiviruses. HIV - BASIC PROPERTIES 1. HIV is a member of the Retrovirus family and more specifically it is a member of the Lentivirus genus of this family.
Medical Virology Immunology. Dr. Sameer Naji, MB, BCh, PhD (UK) Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University
 Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University") Medical Virology Immunology Dr. Sameer Naji, MB, BCh, PhD (UK) Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University Human blood cells Phases of immune responses Microbe Naïve
Medical Virology Immunology Dr. Sameer Naji, MB, BCh, PhD (UK) Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University Human blood cells Phases of immune responses Microbe Naïve
Human Immunodeficiency Virus
 Human Immunodeficiency Virus Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Viruses and hosts Lentivirus from Latin lentis (slow), for slow progression of disease
Human Immunodeficiency Virus Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Viruses and hosts Lentivirus from Latin lentis (slow), for slow progression of disease
AGAINST VIRAL INFECTIONS. Identify the types of immunity involve in the mechanisms of protection against viral infections.
 LECTURE: 02 Title: THE IMMUNOLOGICAL PROTECTIVE MECHANISMS AGAINST VIRAL INFECTIONS LEARNING OBJECTIVES: The student should be able to: Identify the types of immunity involve in the mechanisms of protection
LECTURE: 02 Title: THE IMMUNOLOGICAL PROTECTIVE MECHANISMS AGAINST VIRAL INFECTIONS LEARNING OBJECTIVES: The student should be able to: Identify the types of immunity involve in the mechanisms of protection
Herpesviruses. Virion. Genome. Genes and proteins. Viruses and hosts. Diseases. Distinctive characteristics
 Herpesviruses Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Virion Enveloped icosahedral capsid (T=16), diameter 125 nm Diameter of enveloped virion 200 nm Capsid
Herpesviruses Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Virion Enveloped icosahedral capsid (T=16), diameter 125 nm Diameter of enveloped virion 200 nm Capsid
Lecture 11. Immunology and disease: parasite antigenic diversity
 Lecture 11 Immunology and disease: parasite antigenic diversity RNAi interference video and tutorial (you are responsible for this material, so check it out.) http://www.pbs.org/wgbh/nova/sciencenow/3210/02.html
Lecture 11 Immunology and disease: parasite antigenic diversity RNAi interference video and tutorial (you are responsible for this material, so check it out.) http://www.pbs.org/wgbh/nova/sciencenow/3210/02.html
Intrinsic cellular defenses against virus infection
 Intrinsic cellular defenses against virus infection Detection of virus infection Host cell response to virus infection Interferons: structure and synthesis Induction of antiviral activity Viral defenses
Intrinsic cellular defenses against virus infection Detection of virus infection Host cell response to virus infection Interferons: structure and synthesis Induction of antiviral activity Viral defenses
Antiviral Chemotherapy
 Viruses are intimate intracellular parasites and their destruction may cause destruction of infected cells. Many virus infections were considered to be self-limited. Most of the damage to cells in virus
Viruses are intimate intracellular parasites and their destruction may cause destruction of infected cells. Many virus infections were considered to be self-limited. Most of the damage to cells in virus
CELL BIOLOGY - CLUTCH CH THE IMMUNE SYSTEM.
 !! www.clutchprep.com CONCEPT: OVERVIEW OF HOST DEFENSES The human body contains three lines of against infectious agents (pathogens) 1. Mechanical and chemical boundaries (part of the innate immune system)
!! www.clutchprep.com CONCEPT: OVERVIEW OF HOST DEFENSES The human body contains three lines of against infectious agents (pathogens) 1. Mechanical and chemical boundaries (part of the innate immune system)
CYTOKINE RECEPTORS AND SIGNAL TRANSDUCTION
 CYTOKINE RECEPTORS AND SIGNAL TRANSDUCTION What is Cytokine? Secreted popypeptide (protein) involved in cell-to-cell signaling. Acts in paracrine or autocrine fashion through specific cellular receptors.
CYTOKINE RECEPTORS AND SIGNAL TRANSDUCTION What is Cytokine? Secreted popypeptide (protein) involved in cell-to-cell signaling. Acts in paracrine or autocrine fashion through specific cellular receptors.
Structure and Function of Antigen Recognition Molecules
 MICR2209 Structure and Function of Antigen Recognition Molecules Dr Allison Imrie allison.imrie@uwa.edu.au 1 Synopsis: In this lecture we will examine the major receptors used by cells of the innate and
MICR2209 Structure and Function of Antigen Recognition Molecules Dr Allison Imrie allison.imrie@uwa.edu.au 1 Synopsis: In this lecture we will examine the major receptors used by cells of the innate and
The Adaptive Immune Responses
 The Adaptive Immune Responses The two arms of the immune responses are; 1) the cell mediated, and 2) the humoral responses. In this chapter we will discuss the two responses in detail and we will start
The Adaptive Immune Responses The two arms of the immune responses are; 1) the cell mediated, and 2) the humoral responses. In this chapter we will discuss the two responses in detail and we will start
How HIV Causes Disease Prof. Bruce D. Walker
 How HIV Causes Disease Howard Hughes Medical Institute Massachusetts General Hospital Harvard Medical School 1 The global AIDS crisis 60 million infections 20 million deaths 2 3 The screen versions of
How HIV Causes Disease Howard Hughes Medical Institute Massachusetts General Hospital Harvard Medical School 1 The global AIDS crisis 60 million infections 20 million deaths 2 3 The screen versions of
NTD Vaccine Design Toolkit and Training Workshop Providence, RI January 05, 2011 Cytokines Leslie P. Cousens, PhD EpiVax, Inc.
 NTD Vaccine Design Toolkit and Training Workshop Providence, RI January 05, 2011 Cytokines Leslie P. Cousens, PhD EpiVax, Inc. Cytokines Properties of Cytokines Cytokines are proteins with specific roles
NTD Vaccine Design Toolkit and Training Workshop Providence, RI January 05, 2011 Cytokines Leslie P. Cousens, PhD EpiVax, Inc. Cytokines Properties of Cytokines Cytokines are proteins with specific roles
General information. Cell mediated immunity. 455 LSA, Tuesday 11 to noon. Anytime after class.
 General information Cell mediated immunity 455 LSA, Tuesday 11 to noon Anytime after class T-cell precursors Thymus Naive T-cells (CD8 or CD4) email: lcoscoy@berkeley.edu edu Use MCB150 as subject line
General information Cell mediated immunity 455 LSA, Tuesday 11 to noon Anytime after class T-cell precursors Thymus Naive T-cells (CD8 or CD4) email: lcoscoy@berkeley.edu edu Use MCB150 as subject line
All animals have innate immunity, a defense active immediately upon infection Vertebrates also have adaptive immunity
 1 2 3 4 5 6 7 8 9 The Immune System All animals have innate immunity, a defense active immediately upon infection Vertebrates also have adaptive immunity Figure 43.2 In innate immunity, recognition and
1 2 3 4 5 6 7 8 9 The Immune System All animals have innate immunity, a defense active immediately upon infection Vertebrates also have adaptive immunity Figure 43.2 In innate immunity, recognition and
Immunity to Viruses. Patricia Fitzgerald-Bocarsly September 25, 2008
 Immunity to Viruses Patricia Fitzgerald-Bocarsly September 25, 2008 The Immune System Deals with a Huge Range of Pathogens Roitt, 2003 Immune Responses to Viruses Viruses are dependent on the host cell
Immunity to Viruses Patricia Fitzgerald-Bocarsly September 25, 2008 The Immune System Deals with a Huge Range of Pathogens Roitt, 2003 Immune Responses to Viruses Viruses are dependent on the host cell
Chapter 23 Immunity Exam Study Questions
 Chapter 23 Immunity Exam Study Questions 1. Define 1) Immunity 2) Neutrophils 3) Macrophage 4) Epitopes 5) Interferon 6) Complement system 7) Histamine 8) Mast cells 9) Antigen 10) Antigens receptors 11)
Chapter 23 Immunity Exam Study Questions 1. Define 1) Immunity 2) Neutrophils 3) Macrophage 4) Epitopes 5) Interferon 6) Complement system 7) Histamine 8) Mast cells 9) Antigen 10) Antigens receptors 11)
Anti-infectious Immunity
 Anti-infectious Immunity innate immunity barrier structures Secretory molecules Phagocytes NK cells Anatomical barriers 1. Skin and mucosa barrier 2.hemo-Spinal Fluid barrier 3. placental barrier Phagocytic
Anti-infectious Immunity innate immunity barrier structures Secretory molecules Phagocytes NK cells Anatomical barriers 1. Skin and mucosa barrier 2.hemo-Spinal Fluid barrier 3. placental barrier Phagocytic
Hepatitis C Virus and Cytokine Responses
 Hepatitis C Virus and Cytokine Responses Eui-Cheol Shin, M.D., Ph.D. Laboratory of Immunology & Infectious Diseases (LIID), Graduate School of Medical Science & Engineering (GSMSE), KAIST Daejeon, Korea
Hepatitis C Virus and Cytokine Responses Eui-Cheol Shin, M.D., Ph.D. Laboratory of Immunology & Infectious Diseases (LIID), Graduate School of Medical Science & Engineering (GSMSE), KAIST Daejeon, Korea
T Cell Activation. Patricia Fitzgerald-Bocarsly March 18, 2009
 T Cell Activation Patricia Fitzgerald-Bocarsly March 18, 2009 Phases of Adaptive Immune Responses Phases of T cell responses IL-2 acts as an autocrine growth factor Fig. 11-11 Clonal Expansion of T cells
T Cell Activation Patricia Fitzgerald-Bocarsly March 18, 2009 Phases of Adaptive Immune Responses Phases of T cell responses IL-2 acts as an autocrine growth factor Fig. 11-11 Clonal Expansion of T cells
Third line of Defense
 Chapter 15 Specific Immunity and Immunization Topics -3 rd of Defense - B cells - T cells - Specific Immunities Third line of Defense Specific immunity is a complex interaction of immune cells (leukocytes)
Chapter 15 Specific Immunity and Immunization Topics -3 rd of Defense - B cells - T cells - Specific Immunities Third line of Defense Specific immunity is a complex interaction of immune cells (leukocytes)
VIRUSES. Biology Applications Control. David R. Harper. Garland Science Taylor & Francis Group NEW YORK AND LONDON
 VIRUSES Biology Applications Control David R. Harper GS Garland Science Taylor & Francis Group NEW YORK AND LONDON vii Chapter 1 Virus Structure and 2.2 VIRUS MORPHOLOGY 26 Infection 1 2.3 VIRAL CLASSIFICATION
VIRUSES Biology Applications Control David R. Harper GS Garland Science Taylor & Francis Group NEW YORK AND LONDON vii Chapter 1 Virus Structure and 2.2 VIRUS MORPHOLOGY 26 Infection 1 2.3 VIRAL CLASSIFICATION
Page 32 AP Biology: 2013 Exam Review CONCEPT 6 REGULATION
 Page 32 AP Biology: 2013 Exam Review CONCEPT 6 REGULATION 1. Feedback a. Negative feedback mechanisms maintain dynamic homeostasis for a particular condition (variable) by regulating physiological processes,
Page 32 AP Biology: 2013 Exam Review CONCEPT 6 REGULATION 1. Feedback a. Negative feedback mechanisms maintain dynamic homeostasis for a particular condition (variable) by regulating physiological processes,
MedChem 401~ Retroviridae. Retroviridae
 MedChem 401~ Retroviridae Retroviruses plus-sense RNA genome (!8-10 kb) protein capsid lipid envelop envelope glycoproteins reverse transcriptase enzyme integrase enzyme protease enzyme Retroviridae The
MedChem 401~ Retroviridae Retroviruses plus-sense RNA genome (!8-10 kb) protein capsid lipid envelop envelope glycoproteins reverse transcriptase enzyme integrase enzyme protease enzyme Retroviridae The
Adaptive Immunity: Humoral Immune Responses
 MICR2209 Adaptive Immunity: Humoral Immune Responses Dr Allison Imrie 1 Synopsis: In this lecture we will review the different mechanisms which constitute the humoral immune response, and examine the antibody
MICR2209 Adaptive Immunity: Humoral Immune Responses Dr Allison Imrie 1 Synopsis: In this lecture we will review the different mechanisms which constitute the humoral immune response, and examine the antibody
, virus identified as the causative agent and ELISA test produced which showed the extent of the epidemic
 1 Two attributes make AIDS unique among infectious diseases: it is uniformly fatal, and most of its devastating symptoms are not due to the causative agent Male to Male sex is the highest risk group in
1 Two attributes make AIDS unique among infectious diseases: it is uniformly fatal, and most of its devastating symptoms are not due to the causative agent Male to Male sex is the highest risk group in
The Immune System. These are classified as the Innate and Adaptive Immune Responses. Innate Immunity
 The Immune System Biological mechanisms that defend an organism must be 1. triggered by a stimulus upon injury or pathogen attack 2. able to counteract the injury or invasion 3. able to recognise foreign
The Immune System Biological mechanisms that defend an organism must be 1. triggered by a stimulus upon injury or pathogen attack 2. able to counteract the injury or invasion 3. able to recognise foreign
Viral Evasion Strategies
 Viral Evasion Strategies Lecture 15 Virology W3310/4310 Spring 2013 Don t look back, something might be gaining on you! -SATCHEL PAIGE Host vs. Virus What one does to the other? Evolution of strategies
Viral Evasion Strategies Lecture 15 Virology W3310/4310 Spring 2013 Don t look back, something might be gaining on you! -SATCHEL PAIGE Host vs. Virus What one does to the other? Evolution of strategies
Helminth worm, Schistosomiasis Trypanosomes, sleeping sickness Pneumocystis carinii. Ringworm fungus HIV Influenza
 Helminth worm, Schistosomiasis Trypanosomes, sleeping sickness Pneumocystis carinii Ringworm fungus HIV Influenza Candida Staph aureus Mycobacterium tuberculosis Listeria Salmonella Streptococcus Levels
Helminth worm, Schistosomiasis Trypanosomes, sleeping sickness Pneumocystis carinii Ringworm fungus HIV Influenza Candida Staph aureus Mycobacterium tuberculosis Listeria Salmonella Streptococcus Levels
Antiviral Chemotherapy
 12 Antiviral Chemotherapy Why antiviral drugs? Vaccines have provided considerable success in preventing viral diseases; However, they have modest or often no therapeutic effect for individuals who are
12 Antiviral Chemotherapy Why antiviral drugs? Vaccines have provided considerable success in preventing viral diseases; However, they have modest or often no therapeutic effect for individuals who are
Chapter 10 (pages ): Differentiation and Functions of CD4+ Effector T Cells Prepared by Kristen Dazy, MD, Scripps Clinic Medical Group
: Differentiation and Functions of CD4+ Effector T Cells Prepared by Kristen Dazy, MD, Scripps Clinic Medical Group") FIT Board Review Corner September 2015 Welcome to the FIT Board Review Corner, prepared by Andrew Nickels, MD, and Sarah Spriet, DO, senior and junior representatives of ACAAI's Fellows-In-Training (FITs)
FIT Board Review Corner September 2015 Welcome to the FIT Board Review Corner, prepared by Andrew Nickels, MD, and Sarah Spriet, DO, senior and junior representatives of ACAAI's Fellows-In-Training (FITs)
7.012 Quiz 3 Answers
 MIT Biology Department 7.012: Introductory Biology - Fall 2004 Instructors: Professor Eric Lander, Professor Robert A. Weinberg, Dr. Claudette Gardel Friday 11/12/04 7.012 Quiz 3 Answers A > 85 B 72-84
MIT Biology Department 7.012: Introductory Biology - Fall 2004 Instructors: Professor Eric Lander, Professor Robert A. Weinberg, Dr. Claudette Gardel Friday 11/12/04 7.012 Quiz 3 Answers A > 85 B 72-84
T-cell activation T cells migrate to secondary lymphoid tissues where they interact with antigen, antigen-presenting cells, and other lymphocytes:
 Interactions between innate immunity & adaptive immunity What happens to T cells after they leave the thymus? Naïve T cells exit the thymus and enter the bloodstream. If they remain in the bloodstream,
Interactions between innate immunity & adaptive immunity What happens to T cells after they leave the thymus? Naïve T cells exit the thymus and enter the bloodstream. If they remain in the bloodstream,
T-cell activation T cells migrate to secondary lymphoid tissues where they interact with antigen, antigen-presenting cells, and other lymphocytes:
 Interactions between innate immunity & adaptive immunity What happens to T cells after they leave the thymus? Naïve T cells exit the thymus and enter the bloodstream. If they remain in the bloodstream,
Interactions between innate immunity & adaptive immunity What happens to T cells after they leave the thymus? Naïve T cells exit the thymus and enter the bloodstream. If they remain in the bloodstream,
ACTIVATION AND EFFECTOR FUNCTIONS OF CELL-MEDIATED IMMUNITY AND NK CELLS. Choompone Sakonwasun, MD (Hons), FRCPT
, FRCPT") ACTIVATION AND EFFECTOR FUNCTIONS OF CELL-MEDIATED IMMUNITY AND NK CELLS Choompone Sakonwasun, MD (Hons), FRCPT Types of Adaptive Immunity Types of T Cell-mediated Immune Reactions CTLs = cytotoxic T lymphocytes
ACTIVATION AND EFFECTOR FUNCTIONS OF CELL-MEDIATED IMMUNITY AND NK CELLS Choompone Sakonwasun, MD (Hons), FRCPT Types of Adaptive Immunity Types of T Cell-mediated Immune Reactions CTLs = cytotoxic T lymphocytes
Immunity and Infection. Chapter 17
 Immunity and Infection Chapter 17 The Chain of Infection Transmitted through a chain of infection (six links) Pathogen: Disease causing microorganism Reservoir: Natural environment of the pathogen Portal
Immunity and Infection Chapter 17 The Chain of Infection Transmitted through a chain of infection (six links) Pathogen: Disease causing microorganism Reservoir: Natural environment of the pathogen Portal
T Cell Effector Mechanisms I: B cell Help & DTH
 T Cell Effector Mechanisms I: B cell Help & DTH Ned Braunstein, MD The Major T Cell Subsets p56 lck + T cells γ δ ε ζ ζ p56 lck CD8+ T cells γ δ ε ζ ζ Cα Cβ Vα Vβ CD3 CD8 Cα Cβ Vα Vβ CD3 MHC II peptide
T Cell Effector Mechanisms I: B cell Help & DTH Ned Braunstein, MD The Major T Cell Subsets p56 lck + T cells γ δ ε ζ ζ p56 lck CD8+ T cells γ δ ε ζ ζ Cα Cβ Vα Vβ CD3 CD8 Cα Cβ Vα Vβ CD3 MHC II peptide
HIV INFECTION: An Overview
 HIV INFECTION: An Overview UNIVERSITY OF PAPUA NEW GUINEA SCHOOL OF MEDICINE AND HEALTH SCIENCES DIVISION OF BASIC MEDICAL SCIENCES DISCIPLINE OF BIOCHEMISTRY & MOLECULAR BIOLOGY PBL MBBS II SEMINAR VJ
HIV INFECTION: An Overview UNIVERSITY OF PAPUA NEW GUINEA SCHOOL OF MEDICINE AND HEALTH SCIENCES DIVISION OF BASIC MEDICAL SCIENCES DISCIPLINE OF BIOCHEMISTRY & MOLECULAR BIOLOGY PBL MBBS II SEMINAR VJ
Lecture 2: Virology. I. Background
 Lecture 2: Virology I. Background A. Properties 1. Simple biological systems a. Aggregates of nucleic acids and protein 2. Non-living a. Cannot reproduce or carry out metabolic activities outside of a
Lecture 2: Virology I. Background A. Properties 1. Simple biological systems a. Aggregates of nucleic acids and protein 2. Non-living a. Cannot reproduce or carry out metabolic activities outside of a
Polyomaviridae. Spring
 Polyomaviridae Spring 2002 331 Antibody Prevalence for BK & JC Viruses Spring 2002 332 Polyoma Viruses General characteristics Papovaviridae: PA - papilloma; PO - polyoma; VA - vacuolating agent a. 45nm
Polyomaviridae Spring 2002 331 Antibody Prevalence for BK & JC Viruses Spring 2002 332 Polyoma Viruses General characteristics Papovaviridae: PA - papilloma; PO - polyoma; VA - vacuolating agent a. 45nm
The Immune System All animals have innate immunity, a defense active immediately
 The Immune System All animals have innate immunity, a defense active immediately upon infection Vertebrates also have adaptive immunity Figure 43.2 INNATE IMMUNITY (all animals) Recognition of traits shared
The Immune System All animals have innate immunity, a defense active immediately upon infection Vertebrates also have adaptive immunity Figure 43.2 INNATE IMMUNITY (all animals) Recognition of traits shared
Oncolytic virus strategy
 Oncolytic viruses Oncolytic virus strategy normal tumor NO replication replication survival lysis Oncolytic virus strategy Mechanisms of tumor selectivity of several, some of them naturally, oncolytic
Oncolytic viruses Oncolytic virus strategy normal tumor NO replication replication survival lysis Oncolytic virus strategy Mechanisms of tumor selectivity of several, some of them naturally, oncolytic
Immunology - Lecture 2 Adaptive Immune System 1
 Immunology - Lecture 2 Adaptive Immune System 1 Book chapters: Molecules of the Adaptive Immunity 6 Adaptive Cells and Organs 7 Generation of Immune Diversity Lymphocyte Antigen Receptors - 8 CD markers
Immunology - Lecture 2 Adaptive Immune System 1 Book chapters: Molecules of the Adaptive Immunity 6 Adaptive Cells and Organs 7 Generation of Immune Diversity Lymphocyte Antigen Receptors - 8 CD markers
Scott Abrams, Ph.D. Professor of Oncology, x4375 Kuby Immunology SEVENTH EDITION
 Scott Abrams, Ph.D. Professor of Oncology, x4375 scott.abrams@roswellpark.org Kuby Immunology SEVENTH EDITION CHAPTER 13 Effector Responses: Cell- and Antibody-Mediated Immunity Copyright 2013 by W. H.
Scott Abrams, Ph.D. Professor of Oncology, x4375 scott.abrams@roswellpark.org Kuby Immunology SEVENTH EDITION CHAPTER 13 Effector Responses: Cell- and Antibody-Mediated Immunity Copyright 2013 by W. H.
Cytokines modulate the functional activities of individual cells and tissues both under normal and pathologic conditions Interleukins,
 Cytokines http://highered.mcgraw-hill.com/sites/0072507470/student_view0/chapter22/animation the_immune_response.html Cytokines modulate the functional activities of individual cells and tissues both under
Cytokines http://highered.mcgraw-hill.com/sites/0072507470/student_view0/chapter22/animation the_immune_response.html Cytokines modulate the functional activities of individual cells and tissues both under
Immune system. Aims. Immune system. Lymphatic organs. Inflammation. Natural immune system. Adaptive immune system
 Aims Immune system Lymphatic organs Inflammation Natural immune system Adaptive immune system Major histocompatibility complex (MHC) Disorders of the immune system 1 2 Immune system Lymphoid organs Immune
Aims Immune system Lymphatic organs Inflammation Natural immune system Adaptive immune system Major histocompatibility complex (MHC) Disorders of the immune system 1 2 Immune system Lymphoid organs Immune
RAISON D ETRE OF THE IMMUNE SYSTEM:
 RAISON D ETRE OF THE IMMUNE SYSTEM: To Distinguish Self from Non-Self Thereby Protecting Us From Our Hostile Environment. Innate Immunity Acquired Immunity Innate immunity: (Antigen nonspecific) defense
RAISON D ETRE OF THE IMMUNE SYSTEM: To Distinguish Self from Non-Self Thereby Protecting Us From Our Hostile Environment. Innate Immunity Acquired Immunity Innate immunity: (Antigen nonspecific) defense
Mary ET Boyle, Ph. D. Department of Cognitive Science UCSD
 ? Mary ET Boyle, Ph. D. Department of Cognitive Science UCSD Christian S Lobsiger & Don W Cleveland (2007) Nature Neuroscience 10, 1355-1360 Astrocytes: interlinked gatekeepers of glutamate astrocytes
? Mary ET Boyle, Ph. D. Department of Cognitive Science UCSD Christian S Lobsiger & Don W Cleveland (2007) Nature Neuroscience 10, 1355-1360 Astrocytes: interlinked gatekeepers of glutamate astrocytes
Immunology Basics Relevant to Cancer Immunotherapy: T Cell Activation, Costimulation, and Effector T Cells
 Immunology Basics Relevant to Cancer Immunotherapy: T Cell Activation, Costimulation, and Effector T Cells Andrew H. Lichtman, M.D. Ph.D. Department of Pathology Brigham and Women s Hospital and Harvard
Immunology Basics Relevant to Cancer Immunotherapy: T Cell Activation, Costimulation, and Effector T Cells Andrew H. Lichtman, M.D. Ph.D. Department of Pathology Brigham and Women s Hospital and Harvard
Chapter 11 CYTOKINES
 Chapter 11 CYTOKINES group of low molecular weight regulatory proteins secreted by leukocytes as well as a variety of other cells in the body (8~30kD) regulate the intensity and duration of the immune
Chapter 11 CYTOKINES group of low molecular weight regulatory proteins secreted by leukocytes as well as a variety of other cells in the body (8~30kD) regulate the intensity and duration of the immune
Adaptive Immunity: Specific Defenses of the Host
 17 Adaptive Immunity: Specific Defenses of the Host SLOs Differentiate between innate and adaptive immunity, and humoral and cellular immunity. Define antigen, epitope, and hapten. Explain the function
17 Adaptive Immunity: Specific Defenses of the Host SLOs Differentiate between innate and adaptive immunity, and humoral and cellular immunity. Define antigen, epitope, and hapten. Explain the function
Physiology Unit 3. ADAPTIVE IMMUNITY The Specific Immune Response
 Physiology Unit 3 ADAPTIVE IMMUNITY The Specific Immune Response In Physiology Today The Adaptive Arm of the Immune System Specific Immune Response Internal defense against a specific pathogen Acquired
Physiology Unit 3 ADAPTIVE IMMUNITY The Specific Immune Response In Physiology Today The Adaptive Arm of the Immune System Specific Immune Response Internal defense against a specific pathogen Acquired
Antiviral Drugs Lecture 5
 Antiviral Drugs Lecture 5 Antimicrobial Chemotherapy (MLAB 366) 1 Dr. Mohamed A. El-Sakhawy 2 Introduction Viruses are microscopic organisms that can infect all living cells. They are parasitic and multiply
Antiviral Drugs Lecture 5 Antimicrobial Chemotherapy (MLAB 366) 1 Dr. Mohamed A. El-Sakhawy 2 Introduction Viruses are microscopic organisms that can infect all living cells. They are parasitic and multiply
HIV Immunopathogenesis. Modeling the Immune System May 2, 2007
 HIV Immunopathogenesis Modeling the Immune System May 2, 2007 Question 1 : Explain how HIV infects the host Zafer Iscan Yuanjian Wang Zufferey Abhishek Garg How does HIV infect the host? HIV infection
HIV Immunopathogenesis Modeling the Immune System May 2, 2007 Question 1 : Explain how HIV infects the host Zafer Iscan Yuanjian Wang Zufferey Abhishek Garg How does HIV infect the host? HIV infection
Section Lectures: Immunology/Virology Time: 9:00 am 10:00 am LRC 105 A & B
 Section Director: Cliff Bellone, Ph.D. Office: Doisy Hall - R 405 Phone: 577-8449 E-Mail: bellonec@slu.edu Lecturers: James Swierkosz, Ph.D. Office: Medical School Rm. 412 Phone: 577-8430 E-Mail: swierkoszje@slu.edu
Section Director: Cliff Bellone, Ph.D. Office: Doisy Hall - R 405 Phone: 577-8449 E-Mail: bellonec@slu.edu Lecturers: James Swierkosz, Ph.D. Office: Medical School Rm. 412 Phone: 577-8430 E-Mail: swierkoszje@slu.edu
Macrophage Activation & Cytokine Release. Dendritic Cells & Antigen Presentation. Neutrophils & Innate Defense
 Macrophage Activation & Cytokine Release Dendritic Cells & Antigen Presentation Neutrophils & Innate Defense Neutrophils Polymorphonuclear cells (PMNs) are recruited to the site of infection where they
Macrophage Activation & Cytokine Release Dendritic Cells & Antigen Presentation Neutrophils & Innate Defense Neutrophils Polymorphonuclear cells (PMNs) are recruited to the site of infection where they
Cytokines (II) Dr. Aws Alshamsan Department of Pharmaceu5cs Office: AA87 Tel:
 Dr. Aws Alshamsan Department of Pharmaceu5cs Office: AA87 Tel:") Cytokines (II) Dr. Aws Alshamsan Department of Pharmaceu5cs Office: AA87 Tel: 4677363 aalshamsan@ksu.edu.sa Learning Objectives By the end of this lecture you will be able to: 1 Understand the physiological
Cytokines (II) Dr. Aws Alshamsan Department of Pharmaceu5cs Office: AA87 Tel: 4677363 aalshamsan@ksu.edu.sa Learning Objectives By the end of this lecture you will be able to: 1 Understand the physiological
There are 2 major lines of defense: Non-specific (Innate Immunity) and. Specific. (Adaptive Immunity) Photo of macrophage cell
 and. Specific. (Adaptive Immunity) Photo of macrophage cell") There are 2 major lines of defense: Non-specific (Innate Immunity) and Specific (Adaptive Immunity) Photo of macrophage cell Development of the Immune System ery pl neu mφ nk CD8 + CTL CD4 + thy TH1 mye
There are 2 major lines of defense: Non-specific (Innate Immunity) and Specific (Adaptive Immunity) Photo of macrophage cell Development of the Immune System ery pl neu mφ nk CD8 + CTL CD4 + thy TH1 mye
Interleukin-6; pathogenesis and treatment of autoimmune inflammatory diseases
 54 Review Article Interleukin-6; pathogenesis and treatment of autoimmune inflammatory diseases Toshio Tanaka 1, 2), Masashi Narazaki 3), Kazuya Masuda 4) and Tadamitsu Kishimoto 4, ) 1) Department of
54 Review Article Interleukin-6; pathogenesis and treatment of autoimmune inflammatory diseases Toshio Tanaka 1, 2), Masashi Narazaki 3), Kazuya Masuda 4) and Tadamitsu Kishimoto 4, ) 1) Department of
Under the Radar Screen: How Bugs Trick Our Immune Defenses
 Under the Radar Screen: How Bugs Trick Our Immune Defenses Session 7: Cytokines Marie-Eve Paquet and Gijsbert Grotenbreg Whitehead Institute for Biomedical Research HHV-8 Discovered in the 1980 s at the
Under the Radar Screen: How Bugs Trick Our Immune Defenses Session 7: Cytokines Marie-Eve Paquet and Gijsbert Grotenbreg Whitehead Institute for Biomedical Research HHV-8 Discovered in the 1980 s at the
Unit 5 The Human Immune Response to Infection
 Unit 5 The Human Immune Response to Infection Unit 5-page 1 FOM Chapter 21 Resistance and the Immune System: Innate Immunity Preview: In Chapter 21, we will learn about the branch of the immune system
Unit 5 The Human Immune Response to Infection Unit 5-page 1 FOM Chapter 21 Resistance and the Immune System: Innate Immunity Preview: In Chapter 21, we will learn about the branch of the immune system
LYMPHOCYTES & IMMUNOGLOBULINS. Dr Mere Kende, Lecturer SMHS
 LYMPHOCYTES & IMMUNOGLOBULINS Dr Mere Kende, Lecturer SMHS Immunity Immune- protection against dangers of non-self/invader eg organism 3 components of immune system 1 st line: skin/mucosa/cilia/hair/saliva/fatty
LYMPHOCYTES & IMMUNOGLOBULINS Dr Mere Kende, Lecturer SMHS Immunity Immune- protection against dangers of non-self/invader eg organism 3 components of immune system 1 st line: skin/mucosa/cilia/hair/saliva/fatty
BIT 120. Copy of Cancer/HIV Lecture
 BIT 120 Copy of Cancer/HIV Lecture Cancer DEFINITION Any abnormal growth of cells that has malignant potential i.e.. Leukemia Uncontrolled mitosis in WBC Genetic disease caused by an accumulation of mutations
BIT 120 Copy of Cancer/HIV Lecture Cancer DEFINITION Any abnormal growth of cells that has malignant potential i.e.. Leukemia Uncontrolled mitosis in WBC Genetic disease caused by an accumulation of mutations
Antigen Presentation and T Lymphocyte Activation. Abul K. Abbas UCSF. FOCiS
 1 Antigen Presentation and T Lymphocyte Activation Abul K. Abbas UCSF FOCiS 2 Lecture outline Dendritic cells and antigen presentation The role of the MHC T cell activation Costimulation, the B7:CD28 family
1 Antigen Presentation and T Lymphocyte Activation Abul K. Abbas UCSF FOCiS 2 Lecture outline Dendritic cells and antigen presentation The role of the MHC T cell activation Costimulation, the B7:CD28 family
Third line of Defense. Topic 8 Specific Immunity (adaptive) (18) 3 rd Line = Prophylaxis via Immunization!
 (18) 3 rd Line = Prophylaxis via Immunization!") Topic 8 Specific Immunity (adaptive) (18) Topics - 3 rd Line of Defense - B cells - T cells - Specific Immunities 1 3 rd Line = Prophylaxis via Immunization! (a) A painting of Edward Jenner depicts a cow
Topic 8 Specific Immunity (adaptive) (18) Topics - 3 rd Line of Defense - B cells - T cells - Specific Immunities 1 3 rd Line = Prophylaxis via Immunization! (a) A painting of Edward Jenner depicts a cow
Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics
 Hepadnaviruses Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Hepatitis viruses A group of unrelated pathogens termed hepatitis viruses cause the vast majority
Hepadnaviruses Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Hepatitis viruses A group of unrelated pathogens termed hepatitis viruses cause the vast majority
11/15/2011. Outline. Structural Features and Characteristics. The Good the Bad and the Ugly. Viral Genomes. Structural Features and Characteristics
 Chapter 19 - Viruses Outline I. Viruses A. Structure of viruses B. Common Characteristics of Viruses C. Viral replication D. HIV II. Prions The Good the Bad and the Ugly Viruses fit into the bad category
Chapter 19 - Viruses Outline I. Viruses A. Structure of viruses B. Common Characteristics of Viruses C. Viral replication D. HIV II. Prions The Good the Bad and the Ugly Viruses fit into the bad category
Examples of questions for Cellular Immunology/Cellular Biology and Immunology
 Examples of questions for Cellular Immunology/Cellular Biology and Immunology Each student gets a set of 6 questions, so that each set contains different types of questions and that the set of questions
Examples of questions for Cellular Immunology/Cellular Biology and Immunology Each student gets a set of 6 questions, so that each set contains different types of questions and that the set of questions
The Adaptive Immune Response. B-cells
 The Adaptive Immune Response B-cells The innate immune system provides immediate protection. The adaptive response takes time to develop and is antigen specific. Activation of B and T lymphocytes Naive
The Adaptive Immune Response B-cells The innate immune system provides immediate protection. The adaptive response takes time to develop and is antigen specific. Activation of B and T lymphocytes Naive
The T cell receptor for MHC-associated peptide antigens
 1 The T cell receptor for MHC-associated peptide antigens T lymphocytes have a dual specificity: they recognize polymporphic residues of self MHC molecules, and they also recognize residues of peptide
1 The T cell receptor for MHC-associated peptide antigens T lymphocytes have a dual specificity: they recognize polymporphic residues of self MHC molecules, and they also recognize residues of peptide
HIV & AIDS: Overview
 HIV & AIDS: Overview UNIVERSITY OF PAPUA NEW GUINEA SCHOOL OF MEDICINE AND HEALTH SCIENCES DIVISION OF BASIC MEDICAL SCIENCES DISCIPLINE OF BIOCHEMISTRY & MOLECULAR BIOLOGY PBL SEMINAR VJ TEMPLE 1 What
HIV & AIDS: Overview UNIVERSITY OF PAPUA NEW GUINEA SCHOOL OF MEDICINE AND HEALTH SCIENCES DIVISION OF BASIC MEDICAL SCIENCES DISCIPLINE OF BIOCHEMISTRY & MOLECULAR BIOLOGY PBL SEMINAR VJ TEMPLE 1 What
Scott Abrams, Ph.D. Professor of Oncology, x4375 Kuby Immunology SEVENTH EDITION
 Scott Abrams, Ph.D. Professor of Oncology, x4375 scott.abrams@roswellpark.org Kuby Immunology SEVENTH EDITION CHAPTER 11 T-Cell Activation, Differentiation, and Memory Copyright 2013 by W. H. Freeman and
Scott Abrams, Ph.D. Professor of Oncology, x4375 scott.abrams@roswellpark.org Kuby Immunology SEVENTH EDITION CHAPTER 11 T-Cell Activation, Differentiation, and Memory Copyright 2013 by W. H. Freeman and
19 Viruses BIOLOGY. Outline. Structural Features and Characteristics. The Good the Bad and the Ugly. Structural Features and Characteristics
 9 Viruses CAMPBELL BIOLOGY TENTH EDITION Reece Urry Cain Wasserman Minorsky Jackson Outline I. Viruses A. Structure of viruses B. Common Characteristics of Viruses C. Viral replication D. HIV Lecture Presentation
9 Viruses CAMPBELL BIOLOGY TENTH EDITION Reece Urry Cain Wasserman Minorsky Jackson Outline I. Viruses A. Structure of viruses B. Common Characteristics of Viruses C. Viral replication D. HIV Lecture Presentation
EBV infection B cells and lymphomagenesis. Sridhar Chaganti
 EBV infection B cells and lymphomagenesis Sridhar Chaganti How EBV infects B-cells How viral genes influence the infected B cell Differences and similarities between in vitro and in vivo infection How
EBV infection B cells and lymphomagenesis Sridhar Chaganti How EBV infects B-cells How viral genes influence the infected B cell Differences and similarities between in vitro and in vivo infection How
RAISON D ETRE OF THE IMMUNE SYSTEM:
 RAISON D ETRE OF THE IMMUNE SYSTEM: To Distinguish Self from Non-Self Thereby Protecting Us From Our Hostile Environment. Innate Immunity Adaptive Immunity Innate immunity: (Antigen - nonspecific) defense
RAISON D ETRE OF THE IMMUNE SYSTEM: To Distinguish Self from Non-Self Thereby Protecting Us From Our Hostile Environment. Innate Immunity Adaptive Immunity Innate immunity: (Antigen - nonspecific) defense
Chapter 24 The Immune System
 Chapter 24 The Immune System The Immune System Layered defense system The skin and chemical barriers The innate and adaptive immune systems Immunity The body s ability to recognize and destroy specific
Chapter 24 The Immune System The Immune System Layered defense system The skin and chemical barriers The innate and adaptive immune systems Immunity The body s ability to recognize and destroy specific
STRUCTURE, GENERAL CHARACTERISTICS AND REPRODUCTION OF VIRUSES
 STRUCTURE, GENERAL CHARACTERISTICS AND REPRODUCTION OF VIRUSES Introduction Viruses are noncellular genetic elements that use a living cell for their replication and have an extracellular state. Viruses
STRUCTURE, GENERAL CHARACTERISTICS AND REPRODUCTION OF VIRUSES Introduction Viruses are noncellular genetic elements that use a living cell for their replication and have an extracellular state. Viruses
Chapter 6. Antigen Presentation to T lymphocytes
 Chapter 6 Antigen Presentation to T lymphocytes Generation of T-cell Receptor Ligands T cells only recognize Ags displayed on cell surfaces These Ags may be derived from pathogens that replicate within
Chapter 6 Antigen Presentation to T lymphocytes Generation of T-cell Receptor Ligands T cells only recognize Ags displayed on cell surfaces These Ags may be derived from pathogens that replicate within
Persistent Infections
 Persistent Infections Lecture 17 Biology 3310/4310 Virology Spring 2017 Paralyze resistance with persistence WOODY HAYES Acute vs persistent infections Acute infection - rapid and self-limiting Persistent
Persistent Infections Lecture 17 Biology 3310/4310 Virology Spring 2017 Paralyze resistance with persistence WOODY HAYES Acute vs persistent infections Acute infection - rapid and self-limiting Persistent
Medical Virology. Herpesviruses, Orthomyxoviruses, and Retro virus. - Herpesviruses Structure & Composition: Herpesviruses
 Medical Virology Lecture 2 Asst. Prof. Dr. Dalya Basil Herpesviruses, Orthomyxoviruses, and Retro virus - Herpesviruses Structure & Composition: Herpesviruses Enveloped DNA viruses. All herpesviruses have
Medical Virology Lecture 2 Asst. Prof. Dr. Dalya Basil Herpesviruses, Orthomyxoviruses, and Retro virus - Herpesviruses Structure & Composition: Herpesviruses Enveloped DNA viruses. All herpesviruses have
Chapter 13: Cytokines
 Chapter 13: Cytokines Definition: secreted, low-molecular-weight proteins that regulate the nature, intensity and duration of the immune response by exerting a variety of effects on lymphocytes and/or
Chapter 13: Cytokines Definition: secreted, low-molecular-weight proteins that regulate the nature, intensity and duration of the immune response by exerting a variety of effects on lymphocytes and/or
Antigen Presentation to T lymphocytes
 Antigen Presentation to T lymphocytes Immunology 441 Lectures 6 & 7 Chapter 6 October 10 & 12, 2016 Jessica Hamerman jhamerman@benaroyaresearch.org Office hours by arrangement Antibodies and T cell receptors
Antigen Presentation to T lymphocytes Immunology 441 Lectures 6 & 7 Chapter 6 October 10 & 12, 2016 Jessica Hamerman jhamerman@benaroyaresearch.org Office hours by arrangement Antibodies and T cell receptors
Putting it Together. Stephen Canfield Secondary Lymphoid System. Tonsil Anterior Cervical LN s
 Putting it Together Stephen Canfield smc12@columbia.edu Secondary Lymphoid System Tonsil Anterior Cervical LN s Axillary LN s Mediastinal/Retroperitoneal LN s Thoracic Duct Appendix Spleen Inguinal LN
Putting it Together Stephen Canfield smc12@columbia.edu Secondary Lymphoid System Tonsil Anterior Cervical LN s Axillary LN s Mediastinal/Retroperitoneal LN s Thoracic Duct Appendix Spleen Inguinal LN
Pathogens and the immune system
 Pathogens and the immune system Veronica Leautaud, Ph.D. vl2@ rice.edu Keck Hall 224 / 232-lab Lecture 8 BIOE 301-Bioengineering and World Health Review of lecture 7 Science Science is the human activity
Pathogens and the immune system Veronica Leautaud, Ph.D. vl2@ rice.edu Keck Hall 224 / 232-lab Lecture 8 BIOE 301-Bioengineering and World Health Review of lecture 7 Science Science is the human activity
Dr. Ahmed K. Ali. Outcomes of the virus infection for the host
 Lec. 9 Dr. Ahmed K. Ali Outcomes of the virus infection for the host In the previous few chapters we have looked at aspects of the virus replication cycle that culminate in the exit of infective progeny
Lec. 9 Dr. Ahmed K. Ali Outcomes of the virus infection for the host In the previous few chapters we have looked at aspects of the virus replication cycle that culminate in the exit of infective progeny
Basis and Clinical Applications of Interferon
 Interferon Therapy Basis and Clinical Applications of Interferon JMAJ 47(1): 7 12, 2004 Jiro IMANISHI Professor, Kyoto Prefectural University of Medicine Abstract: Interferon (IFN) is an antiviral substance
Interferon Therapy Basis and Clinical Applications of Interferon JMAJ 47(1): 7 12, 2004 Jiro IMANISHI Professor, Kyoto Prefectural University of Medicine Abstract: Interferon (IFN) is an antiviral substance
October 26, Lecture Readings. Vesicular Trafficking, Secretory Pathway, HIV Assembly and Exit from Cell
 October 26, 2006 Vesicular Trafficking, Secretory Pathway, HIV Assembly and Exit from Cell 1. Secretory pathway a. Formation of coated vesicles b. SNAREs and vesicle targeting 2. Membrane fusion a. SNAREs
October 26, 2006 Vesicular Trafficking, Secretory Pathway, HIV Assembly and Exit from Cell 1. Secretory pathway a. Formation of coated vesicles b. SNAREs and vesicle targeting 2. Membrane fusion a. SNAREs
محاضرة مناعت مدرس المادة :ا.م. هدى عبدالهادي علي النصراوي Immunity to Infectious Diseases
 محاضرة مناعت مدرس المادة :ا.م. هدى عبدالهادي علي النصراوي Immunity to Infectious Diseases Immunity to infection depends on a combination of innate mechanisms (phagocytosis, complement, etc.) and antigen
محاضرة مناعت مدرس المادة :ا.م. هدى عبدالهادي علي النصراوي Immunity to Infectious Diseases Immunity to infection depends on a combination of innate mechanisms (phagocytosis, complement, etc.) and antigen
AP Biology. Viral diseases Polio. Chapter 18. Smallpox. Influenza: 1918 epidemic. Emerging viruses. A sense of size
 Hepatitis Viral diseases Polio Chapter 18. Measles Viral Genetics Influenza: 1918 epidemic 30-40 million deaths world-wide Chicken pox Smallpox Eradicated in 1976 vaccinations ceased in 1980 at risk population?
Hepatitis Viral diseases Polio Chapter 18. Measles Viral Genetics Influenza: 1918 epidemic 30-40 million deaths world-wide Chicken pox Smallpox Eradicated in 1976 vaccinations ceased in 1980 at risk population?
Adaptive Immunity. Lecture 14 Biology W3310/4310 Virology Spring Life is simple, but we insist on making it complicated CONFUCIUS
 Adaptive Immunity Lecture 14 Biology W3310/4310 Virology Spring 2016 Life is simple, but we insist on making it complicated CONFUCIUS Host defenses Intrinsic - Always present in the uninfected cell - Apoptosis,
Adaptive Immunity Lecture 14 Biology W3310/4310 Virology Spring 2016 Life is simple, but we insist on making it complicated CONFUCIUS Host defenses Intrinsic - Always present in the uninfected cell - Apoptosis,
C. Incorrect! MHC class I molecules are not involved in the process of bridging in ADCC.
 Immunology - Problem Drill 13: T- Cell Mediated Immunity Question No. 1 of 10 1. During Antibody-dependent cell mediated cytotoxicity (ADCC), the antibody acts like a bridge between the specific antigen
Immunology - Problem Drill 13: T- Cell Mediated Immunity Question No. 1 of 10 1. During Antibody-dependent cell mediated cytotoxicity (ADCC), the antibody acts like a bridge between the specific antigen
Shiv Pillai Ragon Institute, Massachusetts General Hospital Harvard Medical School
 CTLs, Natural Killers and NKTs 1 Shiv Pillai Ragon Institute, Massachusetts General Hospital Harvard Medical School CTL inducing tumor apoptosis 3 Lecture outline CD8 + Cytotoxic T lymphocytes (CTL) Activation/differentiation
CTLs, Natural Killers and NKTs 1 Shiv Pillai Ragon Institute, Massachusetts General Hospital Harvard Medical School CTL inducing tumor apoptosis 3 Lecture outline CD8 + Cytotoxic T lymphocytes (CTL) Activation/differentiation
Mucosal Immune System
 Exam Format 100 points - 60 pts mandatory; 40 points where 4, 10 point questions will be chosen Some open-ended questions, some short answer. Kuby question Cytokines Terminology How do cytokines achieve
Exam Format 100 points - 60 pts mandatory; 40 points where 4, 10 point questions will be chosen Some open-ended questions, some short answer. Kuby question Cytokines Terminology How do cytokines achieve
1. Overview of Adaptive Immunity
 Chapter 17A: Adaptive Immunity Part I 1. Overview of Adaptive Immunity 2. T and B Cell Production 3. Antigens & Antigen Presentation 4. Helper T cells 1. Overview of Adaptive Immunity The Nature of Adaptive
Chapter 17A: Adaptive Immunity Part I 1. Overview of Adaptive Immunity 2. T and B Cell Production 3. Antigens & Antigen Presentation 4. Helper T cells 1. Overview of Adaptive Immunity The Nature of Adaptive
Question 1. Kupffer cells, microglial cells and osteoclasts are all examples of what type of immune system cell?
 Abbas Chapter 2: Sarah Spriet February 8, 2015 Question 1. Kupffer cells, microglial cells and osteoclasts are all examples of what type of immune system cell? a. Dendritic cells b. Macrophages c. Monocytes
Abbas Chapter 2: Sarah Spriet February 8, 2015 Question 1. Kupffer cells, microglial cells and osteoclasts are all examples of what type of immune system cell? a. Dendritic cells b. Macrophages c. Monocytes
Virus and Prokaryotic Gene Regulation - 1
 Virus and Prokaryotic Gene Regulation - 1 We have discussed the molecular structure of DNA and its function in DNA duplication and in transcription and protein synthesis. We now turn to how cells regulate
Virus and Prokaryotic Gene Regulation - 1 We have discussed the molecular structure of DNA and its function in DNA duplication and in transcription and protein synthesis. We now turn to how cells regulate
chapter 17: specific/adaptable defenses of the host: the immune response
 chapter 17: specific/adaptable defenses of the host: the immune response defense against infection & illness body defenses innate/ non-specific adaptable/ specific epithelium, fever, inflammation, complement,
chapter 17: specific/adaptable defenses of the host: the immune response defense against infection & illness body defenses innate/ non-specific adaptable/ specific epithelium, fever, inflammation, complement,
Section 6. Junaid Malek, M.D.
 Section 6 Junaid Malek, M.D. The Golgi and gp160 gp160 transported from ER to the Golgi in coated vesicles These coated vesicles fuse to the cis portion of the Golgi and deposit their cargo in the cisternae
Section 6 Junaid Malek, M.D. The Golgi and gp160 gp160 transported from ER to the Golgi in coated vesicles These coated vesicles fuse to the cis portion of the Golgi and deposit their cargo in the cisternae
This is a free sample of content from The Hepatitis B and Delta Viruses. Click here for more information on how to buy the book.
 Index A Acute liver failure (ALF), viral hepatitis, 7 Adalimab, hepatitis B management in immunosuppression patients, 279 Alemtuzumab, hepatitis B management in immunosuppression patients, 280 ALF. See
Index A Acute liver failure (ALF), viral hepatitis, 7 Adalimab, hepatitis B management in immunosuppression patients, 279 Alemtuzumab, hepatitis B management in immunosuppression patients, 280 ALF. See