CHARACTERIZATION OF IMMUNE RESPONSES TO WOLBACHIA IN INDIVIDUALS WITH LYMPHATIC FILARIASIS GEORGE ALBERT PUNKOSDY
|
|
|
- Dominick Morgan
- 6 years ago
- Views:
Transcription
1 CHARACTERIZATION OF IMMUNE RESPONSES TO WOLBACHIA IN INDIVIDUALS WITH LYMPHATIC FILARIASIS by GEORGE ALBERT PUNKOSDY (Under the Direction of Patrick J. Lammie) ABSTRACT Lymphatic filariasis is a parasitic disease caused by infection with the filarial nematodes Wuchereria bancrofti, Brugia malayi, and Brugia timori. For some time, researchers have known that these worms harbor endosymbiotic bacterium belonging to the genus Wolbachia; however, it is not known what effect Wolbachia have on the development of the filarial disease. In order to test this hypothesis, the following studies were designed to determine whether individuals with lymphatic filariasis mount immune responses to Wolbachia. First, it was demonstrated in Brugia malayi-infected rhesus monkeys that antibodies to a major Wolbachia surface protein (WSP) were associated with the development of lymphedema and worm death. Similar results were also obtained using cross sectional serum samples from individuals living in Leogane, Haiti, an area endemic for lymphatic filariasis. In these studies, individuals with lymphedema or hydrocele had significantly higher levels of antibodies to WSP than infectionand gender-matched individuals without the chronic manifestations of the disease. In order to investigate the fate of Wolbachia following worm death, the in situ distribution of Wolbachia was assessed in granulomatous nodules collected from individuals in Recife, Brazil that developed following adult worm death. In 4/17 of these nodules, WSP staining was observed
2 not only inside the filarial worms but also in the surrounding inflammation. In one case, Wolbachia antigen staining was observed inside human macrophages/giant cells that make up the granuloma. However, there were no differences in the histological characteristics of nodules where Wolbachia antigens staining was observed outside the worm compared to nodules where Wolbachia antigen staining was only observed inside the worm. Finally, in order to investigate whether individuals with lymphatic filariasis mount inflammatory immune responses to WSP, cytokine/chemokine responses were assayed in PBMC cultures stimulated with swsp. In these studies, it was observed that the majority of cell cultures from individuals living in Leogane, Haiti produced the monocyte chemoattractants MCP-1 and MIP-1β in response to swsp. Although levels of MIP-1β were similar among the different groups of Haitians, cell cultures from individuals with lymphedema produced significantly more MCP-1 than did cell cultures from individuals who were microfilaremic. INDEX WORDS: Lymphatic filariasis, Wolbachia, Human, Pathogenesis, Lymphedema, Hydrocele, Wolbachia surface protein (WSP), Antibody response, Monocyte chemoattractant protein (MCP)-1, Macrophage inflammatory protein (MIP)-1β
3 CHARACTERIZATION OF IMMUNE RESPONSES TO WOLBACHIA IN INDIVIDUALS WITH LYMPHATIC FILARIASIS by GEORGE ALBERT PUNKOSDY B.S., The University of Georgia, 1998 A Dissertation Submitted to the Graduate Faculty of The University of Georgia in Partial Fulfillment of the Requirements for the Degree DOCTOR OF PHILOSOPHY ATHENS, GEORGIA 2004
4 2004 George Albert Punkosdy All Rights Reserved
5 CHARACTERIZATION OF IMMUNE RESPONSES TO WOLBACHIA IN INDIVIDUALS WITH LYMPHATIC FILARIASIS by GEORGE ALBERT PUNKOSDY Major Professor: Patrick Lammie Committee: Donald Champagne Daniel Colley Duncan Krause Rick Tarleton Electronic Version Approved: Maureen Grasso Dean of the Graduate School The University of Georgia December 2004
6 ACKNOWLEDGEMENTS I would like to express my sincere gratitude to my major professor, Dr. Patrick Lammie, for giving me the opportunity to work on this project and allowing me the freedom to pursue my own interests and develop as a scientist. I could not have asked to work for a better professional or personal role model. I would like to thank my committee, Dr. Don Champagne, Dr. Dan Colley, Dr. Duncan Krause, and Dr. Rick Tarleton for their guidance and support. During my time in graduate school I have had the opportunity to work with many truly amazing people. In particular, I would like to thank Dr. David Addiss, Dr. Mark Eberhard, and Dr. Jeannette Guarner for their helpful suggestions and support. I would like to thank Dr. Gerusa Dreyer, whose passion for science and medicine has served as an inspiration to me. Special thanks to all of the past and present members of the lab who have provided such an enjoyable working environment. I would like to especially thank Delynn Moss for sharing an office, as well as countless football stories, with me. To Susan Wilson, thank you for your support both inside and outside the lab during this process. I would like to express my appreciation to the individuals in Haiti and Brazil living with lymphatic filariasis who provided samples for my experiments. Certainly, without their desire to understand more about this disease, none of this work would have been possible. iv
7 TABLE OF CONTENTS Page ACKNOWLEDGEMENTS... iv LIST OF TABLES... viii LIST OF FIGURES... ix CHAPTER 1 INTRODUCTION...1 Background and Epidemiology...1 Life Cycle...2 Dynamics of Infection...4 Immune Responses in Lymphatic Filariasis...7 Development of Disease...10 Bacterial Involvement in Lymphedema Development...12 Wolbachia Bacteria of Filarial Worms...13 Statement of Purpose DETECTION OF SERUM IgG ANTIBODIES SPECIFIC FOR WOLBACHIA SURFACE PROTEIN IN RHESUS MONKEYS INFECTED WITH BRUGIA MALAYI...24 Abstract...25 Introduction...25 Materials and Methods...26 v
8 Results...29 Discussion CHARACTERIZATION OF ANTIBODY RESPONSES TO WOLBACHIA SURFACE PROTEIN IN HUMANS WITH LYMPHATIC FILARIASIS...37 Abstract...38 Introduction...39 Materials and Methods...41 Results...44 Discussion IMMUNOLOCALIZATION OF WOLBACHIA IN BIOPSY SPECIMENS COLLECTED FROM PATIENTS IN RECIFE, BRAZIL WITH BANCROFTIAN FILARIASIS...66 Abstract...66 Introduction...67 Materials and Methods...69 Results...71 Discussion HUMAN PERIPHERAL BLOOD MONONCULEAR CELLS PRODUCE THE MONOCYTE CHEMOATTRACTANTS MCP-1 AND MIP-1β IN RESPONSE TO WOLBACHIA SURFACE PROTEIN...87 Abstract...87 Introduction...88 Materials and Methods...90 vi
9 Results...92 Discussion CONCLUSIONS REFERENCES vii
10 LIST OF TABLES Page Table 1.1: Listing of filarial species positive and negative for Wolbachia...19 Table 2.1: Summary of infection outcome for rhesus monkeys in each of the four infection groups...34 Table 3.1: Comparison of anti-wsp antibody responses among the groups...56 Table 3.2: Association between anti-wsp antibody responses and clinical findings in men with hydrocele...57 Table 4.1: Summary of histological results for specimens examined Table 5.1: Demographic and parasitologic characteristics of the different groups viii
11 LIST OF FIGURES Page Figure 1.1: Localization of Wolbachia in adult female filarial worm...21 Figure 1.2: Immunolocalization of Wolbachia in the embryonic stages of development of B. pahangi worms...22 Figure 2.1: Representative composite graphs showing the course of infection and antibody responses of rhesus monkeys in the bolus + trickle group...35 Figure 3.1: Composite graph showing a temporal association between anti-wsp IgG responses and the onset of lymphedema...58 Figure 3.2: Anti-WSP IgG levels are associated with the presence of lymphedema...59 Figure 3.3: Correlation between anti-wsp IgG levels and lymphedema duration among anti- WSP + women with lymphedema...61 Figure 3.4: Anti-WSP levels are associated with the presence of hydrocele...62 Figure 3.5: Linear epitopes of WSP recognized by anti-wsp + individuals with lymphedema or hydrocele, asymptomatic Ag + Mf + individuals, asymptomatic Ag Mf individuals, and North Americans...64 Figure 4.1: Histological characteristics of W. bancrofti granuloma...81 Figure 4.2: Immunolocalization of Wolbachia in adult B. pahangi worms...82 Figure 4.3: Immunolocalization of Wolbachia in inflammatory nodules...83 ix
12 Figure 4.4: Comparison of the inflammatory characteristics of similarly aged nodules where Wolbachia staining was only seen inside the filarial worm and where Wolbachia staining was seen in the surrounding inflammation...85 Figure 5.1: IL-10 produced by unstimulated PBMC cultures from Haitian individuals with lymphedema (LE), asymptomatic individuals who were Mf (+), and North Americans (NA) Figure 5.2: Net production of IL-2, IL-10, and IL-4 and IFN-γ in response to BpAg in PBMC cultures from Haitian individuals with lymphedema (LE), asymptomatic individuals who were Mf (+), and North Americans (NA) Figure 5.3: Net production of MCP-1 and MIP-1β in response to BpAg and swsp in PBMC cultures from Haitian individuals with lymphedema (LE), asymptomatic individuals who were Mf (+), and North Americans (NA) x
13 CHAPTER 1 INTRODUCTION Background and Epidemiology Lymphatic filariasis is an infectious parasitic disease that has existed as a public health problem in human populations for thousands of years. Today, at least 100 million people living in more than 80 countries are actively infected by the lymphatic-dwelling filarial nematodes that cause lymphatic filariasis, and another one billion people live in areas in which active transmission of infection is occurring and are at risk of becoming infected (Michael et al., 1996). Countries affected by lymphatic filariasis form a belt around the tropical regions of the world and include areas in Africa, Southeast Asia, the Pacific, the Caribbean, and South America. Although most cases of lymphatic filariasis are asymptomatic, the disease is still a major cause of morbidity in these regions (WHO, 1995). Approximately 25 million people suffer from lymphedema/elephantiasis and hydrocele; the chronic manifestations of lymphatic filariasis. These disfiguring manifestations cause a significant decrease in quality of life and often result in social ostracization of affected individuals. Although it is hard to quantitate the economic impact that this disease has on the lives of affected individuals, in Orissa, India, it has been estimated that patients with chronic filarial disease lose > 2 months of work per year because of the disease, and treatment costs account for ~ 7% of their annual income (Babu et al., 2002). Still, many people can not afford more expensive procedures to reduce disease burden, such as hydrocele surgery, and are forced to alter their lifestyle to cope with the disease. 1
14 In 1993, the International Task Force for Disease Eradication listed lymphatic filariasis as one of six potentially eradicable diseases (CDC, 1993), and in 1997 the World Health Assembly passed a resolution calling for the elimination of lymphatic filariasis as a public health problem by the year 2020 (WHO, 1997). Since this time, a global alliance has been formed and guidelines established for mass drug administrations in areas endemic for lymphatic filariasis. In 2002 alone, > 59 million people worldwide received either diethylcarbamazine (DEC) or ivermectin plus albendazole for the treatment of lymphatic filariasis as part of the elimination program. However, despite these ongoing control programs, research is still needed in order to further understand the parasite's biology and the mechanism of disease development. Life Cycle The three species of filarial nematodes that cause lymphatic filariasis in humans are Wuchereria bancrofti, Brugia malayi, and Brugia timori (Nematoda; Onchocercidae). W. bancrofti accounts for approximately 90% of all infections while B. malayi and B. timori collectively make up the other ~10%; mostly in Southeast Asia and the Pacific. The parasite s life cycle consists of dioecious male and female adult worms, the microfilaria stage, and four larval stages (L1-L4). The third larval stage (L3) is the infectious stage and is transmitted to humans via a mosquito intermediate host. Following the bite of an infective mosquito, L3 are deposited on the skin and actively penetrate through the bite wound of the mosquito. The L3 enter the lymphatic vessels under the skin and begin their afferent migration through the lymphatic vessels and lymph nodes. Between 9 and 14 days post infection the L3 molt to form fourth stage larvae (L4), and approximately 30 days post infection the L4 molt into adult male and female worms. During the time in which the larvae develop into adult worms in the human, 2
15 the parasites undergo a dramatic period of growth (from ~1.4 mm in the L3 stage to ~4 cm as adult males and ~8-10 cm as adult females). Adult filarial worms can reside within the lumen of the lymphatic vessels anywhere in the body; however, W. bancrofti adults are typically found in the lymphatic vessels of the lower extremities in females and the lymphatic vessels of the spermatic cord in males. Although the life span of adult worms is not precisely known, it is estimated that adult females can remain reproductively active on the order of 5 years (Vanamail et al., 1996). Reproduction of the parasite requires insemination of the female with sperm from the male. The embryos develop over a period of 3 weeks in the uterus of the female and are released as fully-formed sheathed microfilaria. Each gravid female can release millions of microfilaria over her lifetime. The microfilaria migrate from the lymphatic vessels into the peripheral circulation, and their density in the blood fluctuates dramatically over a 24-hour period. In most areas of the world, W. bancrofti microfilaria are nocturnally periodic with the peak parasitemia occurring between 10PM and 2AM, the peak biting times of the mosquitoes typically serving as vectors. In order for the filarial worms to infect their intermediate host, a female mosquito must ingest microfilaria during a blood meal on a microfilaremic individual. After ingestion, the microfilaria exsheath, penetrate the midgut of the mosquito, and migrate toward the thoracic muscles. Here the microfilaria transform into first stage larvae (L1). During the next two weeks, the parasites undergo two more molts to form the L3 stage. The L3 migrate to the feeding structures in the head of the mosquito and are released when a mosquito takes a subsequent bloodmeal to complete the parasite s life cycle. At least four different genera of mosquitoes (Aedes, Anopheles, Culex, and Mansonia) have been identified as vectors for filarial worms, and 3
16 the principal vector in any region depends on such factors as sanitation, natural breeding sites for mosquitoes, and a mosquito s vectorial capacity (Bartholomay and Christensen, 2002). Diagnosis of lymphatic filariasis depends on the detection of the parasite or parasite antigens in the blood. The classical gold-standard technique has been the detection of microfilaria in a peripheral blood sample drawn during the time of peak parasitemia (usually at night). This technique is now considered quite insensitive, and when used alone can underestimate the true prevalence of infection in an area. Recently, a highly sensitive and specific test that detects the presence of an adult worm antigen in the blood of actively infected individuals has been developed to diagnose W. bancrofti infection (Weil et al., 1987; More and Copeman, 1990). This test is useful because antigen levels remain constant throughout the day, and it has the added advantage of being able to detect single-sex and non-fecund infections in which microfilaria are not produced. Dynamics of Infection Although lymphatic filariasis is endemic in most of the tropical regions of the world, the prevalence of filarial infection in different regions can vary dramatically. To illustrate this point, consider the two populations in which many of the following studies take place. In Recife, Brazil, the overall prevalence of filarial infection is < 1% and infection is concentrated in small endemic foci. In comparison, filarial infection in Leogane, Haiti, is much more widespread and > 50% of the population is actively infected. While these examples represent extremes, actual prevalence values in any region can lie anywhere along this continuum of high and low values. Differences in infection prevalences between any two populations are likely to be influenced by many factors; however, entomologic determinants that lead to differences in exposure intensity 4
17 and/or transmission efficiency may be particularly important. The intensity of exposure to filarial worms in an area can be measured in terms of the annual transmission potential (ATP), the number of L3 that a person living in an endemic area is predicted to be exposed to per year. As expected, the ATP can vary greatly between areas of high and low infection prevalence, and several studies have shown that microfilaria and antigen prevalences as well as disease severity positively correlate with ATP (Kazura et al., 1997; Michael et al., 2001). In addition, studies have shown that individual vector species vary in their efficiency to transmit the parasites to humans and this has important consequences for the establishment of infections in a community (Southgate, 1992; Burkot et al., 2002). To add an additional level of complexity to the dynamics of filarial infection, there is a considerable amount of evidence to suggest that, within a given population, individual susceptibility to filarial infection is not uniform. In all populations endemic for lymphatic filariasis there are certain individuals, termed "endemic normals", who seem to never develop patent filarial infections and show no clinical signs or symptoms of filariasis (Kazura, 2000). One could hypothesize that the reason some individuals do not develop infection may be related to differential exposure to infective larvae. While vector-feeding patterns in some areas may show some degree of non-randomness (D. Goodman, unpublished data), these results are not sufficient to fully explain the patterns of infection seen in a community. In fact, in studies where antibodies to L3 have been measured as a marker of exposure, it has been shown that exposure is ubiquitous among all individuals living in areas of endemicity and that individuals are exposed throughout their life to infective larvae (Bailey et al., 1995). So, if exposure is uniform, what causes some individuals to develop infection with filarial worms and other to remain resistant to infection? 5
18 One possible explanation is that the acquisition of filarial infection is related to the cumulative amount of exposure received over an individual's lifetime. Consistent with this hypothesis, data from many areas demonstrate that filarial infection prevalence increases as a function of age, beginning in childhood. Although transplacental infection is not thought to occur (Eberhard et al., 1993), it is widely recognized that filarial infection can be acquired early in life, and active infections have been documented in individuals as young as 2-years of age (Lammie et al., 1998). Filarial infection increases with age throughout childhood and infection prevalence in children < 20 years of age is proportional to that in the adult population (Witt and Ottesen, 2001). Among adults, it is predicted that filarial infection reaches equilibrium. Although there are very limited data from some areas to suggest that infection prevalence actually decreases in older adult age groups (Das et al., 1990), these findings seem to be the exception rather than the rule. In most areas of the world, age-specific prevalence curves of antigenemia and/or microfilaremia show that filarial infection tends to either plateau or steadily increase with age throughout adulthood (Lammie et al., 1994; Chanteau et al., 1995; Michael et al., 2001). This relationship between infection prevalence and age suggests that protective immunity is not acquired through multiple exposures to infective larvae; instead it is more likely that multiple exposures are required in order for an individual to develop an active infection. Another factor that may play a role in individual susceptibility/resistance to filarial infection is exposure to filarial antigens in the in utero environment. In studies in Haiti and Kenya, children born to infected mothers were found to have a nearly three- to four-fold greater risk of developing filarial infection than individuals born to uninfected mothers (Lammie et al., 1991; Malhotra et al., 2003). These studies also found that children born to infected mothers were less immunologically responsive to filarial antigens than those of children born to 6
19 uninfected mothers, thus suggesting that in utero exposure to filarial antigens may induce an immunological environment conducive to the development of the filarial parasite. The mechanism by which in utero exposure may modulate responses to filarial antigens later in life is largely unclear; however, cellular anergy and idiotypic mechanisms may be involved. Given the current understanding of the dynamics of filarial infection, it is not readily apparent how one s susceptibility to infection leads to the development of disease. Hydrocele and lymphedema are thought to share some of the same pathologic mechanisms; however, there is a drastic difference between infection prevalence in these two groups. Men with hydrocele form a heterogeneous group with the percentage of men actively infected closely paralleling the prevalence of infection seen in the community (Addiss et al., 1995). In contrast, in many areas of the world there is a clear dissociation between active infection and the presence of lymphedema. Most patients with lymphedema show no obvious signs of current filarial infection, and it is unclear whether these patients were ever microfilaremic. In a longitudinal follow-up study performed in Sri Lanka, amicrofilaremic individuals were significantly more likely to develop lymphangitis and lymphedema than microfilaremic individuals (Dissanayake, 2001). This finding does not dispute the fact that filarial nematodes are involved in the development of lymphedema since the incidence of lymphatic obstructive disease is higher in endemic areas than in non-endemic areas. Instead, there are likely to be many factors that collectively contribute to the development of lymphedema. Immune Responses in Lymphatic Filariasis Another factor that may be important in determining infection outcome is the type of immune response one mounts to filarial antigens. Because of the variety of clinical and 7
20 parasitological outcomes of infection seen in endemic areas, lymphatic filariasis is often considered to be a spectral disease. It is well documented that there is an association between infection outcome and host immune responsiveness (Ottesen, 1984). As a result, groups are often defined not only by their infection status but also by characteristic immune responses to filarial antigens. At one end of the spectrum are individuals who are actively infected by filarial worms. By definition, these individuals are microfilaremic and/or antigen-positive, and are characterized by down-regulated filarial-specific immune responses. Peripheral blood mononuclear cells (PBMC) from these individuals show very little proliferative response to crude filarial antigens, and cytokine responses are skewed toward a Th-2 biased immune response (Mahanty et al., 1996; Ravichandran et al., 1997). This hyporesponsiveness of PBMC to filarial antigens among chronically infected individuals is likely to occur in an IL-10 dependent manner as anti-il-10 neutralizing antibodies have been shown to reverse the proliferative defect and restore the ability of these cells to produce IFN-γ (King et al., 1993; Mahanty et al., 1997). This immuno-suppressive environment may be established with the immune systems first encounter of filarial antigens, namely the interaction with antigen presenting cells (APC). Studies have shown that dendritic cells cultured with live microfilaria or L3 are defective in their ability to induce CD4+ T cell proliferation (Semnani et al., 2003; Semnani et al., 2004). While it is not entirely clear what other factors may cause such a downregulation in immune responsiveness, some evidence suggests the parasite itself may actively contribute through the induction of apoptosis of CD4+ T-cells (Jenson et al., 2002) and the expression of down-regulatory cytokine-like molecules (Gomez-Escobar et al., 1998). Although these individuals do have defects in filarial-specific T cell responses, B cell responses seem to remain intact. Typically, chronically infected individuals do have high serum levels of anti- 8
21 filarial IgG antibodies; however, anti-filarial antibodies in these individuals are typically of the IgG4 subclass (Kwan-Lim et al., 1990). Interestingly, individuals with chronic filarial infection are also largely asymptomatic and show few signs of pathology suggesting that disease development may be, at least in part, immune-mediated. The next group along the spectrum of immune responses seen in lymphatic filariasis is made up by asymptomatic/amicrofilaremic individuals who fail to develop infection despite exposure to infective larvae. Immune responses in these antigen-negative individuals are characterized by intense proliferative responses and IL-2 production in response to filarial antigens (Dimock et al., 1996). In addition, these individuals preferentially mount antibody responses of the IgG1, IgG2, and IgG3 subclasses. These studies suggest that Th-1-like immune responses may confer some degree of protection from filarial infection, while individuals that mount Th-2-like immune responses are more likely to develop chronic infection with filarial worms. Consistent with the idea that pathology is at least to some extent immune-mediated; patients with lymphedema mount the most intense anti-filarial immune responses of any group in an endemic area (Lammie et al., 1993). An interesting paradox is that in many areas the vast majority of patients with lymphedema are also uninfected by filarial worms (amicrofilaremic and antigen-negative). As expected from the simple Th-1/Th-2 dichotomy of immune responses seen in uninfected and infected individuals, patients with lymphedema display Th-1 biased immune responses to filarial antigens. In addition to displaying high levels of PBMC proliferation, lymphedema patients also show significantly higher levels of IFN-γ production than do microfilaremic controls (de Almeida et al., 1998). Serum levels of antifilarial IgG1, IgG2, and IgG3 are significantly higher among lymphedema patients than among sex-matched individuals 9
22 without disease (Baird et al., 2002). Furthermore, T-cells from individuals with lymphedema show greater levels of transendothelial migration than do T-cells of microfilaremic individuals, and this process is dependent on endothelial expression of VLA-4/ICAM-1 (Plier et al., 1997). Taken together, these data suggest that while Th-1 responses confer some degree of protection, too much of an inflammatory response is also associated with disease development. Development of Disease Virtually all individuals living in areas endemic for lymphatic filariasis develop some degree of lymphatic pathology. Even asymptomatic microfilaremic individuals who display down-regulated anti-filarial immune responses experience subclinical lymphangiectasia around living adult worms (Dissanayake et al., 1995; Freedman et al., 1995; Noroes et al., 1996). These changes in the lymphatic architecture are likely to be a parasite-induced phenomenon and not immune-mediated since similar responses are seen in SCID mice infected with B. malayi (Nelson et al., 1991). Inflammation is typically not seen around the dilated lymphatic vessels harboring parasites and the endothelial lining remains intact as long as the worms are alive. Nevertheless, some people, for reasons that are not entirely clear, go on to develop chronic disease. The pathology of chronic filarial disease shows a shift from subclinical lymphangiectasia to inflammatory-mediated destruction of lymphatic vessels. A key factor in the development of both hydrocele and lymphedema seems to be the death of the adult worm. Once filarial worms die, by whatever mechanism, there is an intense granulomatous inflammatory response around the dead worm characterized by infiltrating neutrophils, eosinophils, and mononuclear cells (Dreyer et al., 2000). Eventually, this inflammatory reaction, termed filarial adenolymphangitis (FADL), can lead to complete obstruction of the lymphatic vessel. This type of inflammatory 10
23 response is often accompanied by retrograde lymphangitis, fever, and headache. Occasionally, acute episodes of lymphedema and hydrocele are seen following FADL attacks; however, these symptoms usually spontaneously resolve. Current thinking is that FADL attacks trigger the initial pathologic events that eventually lead to the development of chronic disease and set the stage for more severe pathology. Perhaps the greatest difference in the development of pathology in lymphedema versus hydrocele is how lymphedema progresses in the absence of filarial infection. As opposed to hydrocele, in which the adult worm is almost entirely responsible for pathology (Dreyer et al., 2000), many factors collectively contribute to the development of lymphedema. These factors include genetic predisposition, continuous exposure to infective larvae, as well as secondary bacterial infections. Pedigree analysis of families in Haiti has shown that cases of lymphedema cluster in certain high-risk families (T.Cuenco, 2001). Persons in these families were significantly more likely to have lymphedema than would be expected based on the prevalence of lymphedema in the population. However, the genes involved in lymphedema development are not known. Recent reports have shown that heterozygous mutations in the VEGFR-3 or FOXC2 genes result in primary and hereditary lymphedema (Karkkainen et al., 2000; Finegold et al., 2001), and a possible link between these genes and the development of filarial lymphedema is under investigation. Even though lymphedema patients remain free of infection by filarial worms, they are still constantly exposed through the bites of infective mosquitoes. The fate of these L3 is not known since they do not appear to develop into adult worms; however, it is conceivable that they may somehow modulate the immune response in patients with lymphedema. In laboratory models of filarial infection, injection of L3 antigen extracts into jirds (Meriones unguiculatus) 11
24 was followed by enhanced cellular responsiveness and granulomatous reactions that peaked 7 to 14 days post injection (Rao et al., 1996). Brugia pahangi-infected dogs also mounted heightened proliferative responses 4-6 weeks post infection (Schreuer and Hammerberg, 1993). Furthermore, primates experimentally infected with Loa loa initially mounted Th-1-like immune responses that were characterized by the secretion of IL-2 and IFN-γ before immune responses were down-regulated by adult worms (Leroy et al., 1997). These findings suggest that during the earliest periods of infection, all hosts may display inflammatory responses to filarial antigens similar to immune responses seen in patients with lymphedema. Although it has been hypothesized that the parasite may actually induce these responses to aid its development (Ravindran, 2001), these immune responses, if not down-regulated, can contribute to the inflammatory environment seen in individuals who develop disease. Bacterial Involvement in Lymphedema Development Although the connection between bacterial infections and lymphedema development was initially made many years ago, the importance of this association was not fully appreciated until recently. Lymphatic dysfunction due to damaged lymphatic vessels results in impaired lymph flow and the accumulation of lymph in inflamed areas. This stagnant environment is conducive to bacterial growth; leading to the hypothesis that lymphedema patients may be more susceptible to bacterial infection than individuals without disease. In fact, studies have shown that women with lymphedema show heightened immune reactivity to common bacterial antigens such as streptolysin O (Baird et al., 2002), and secondary bacterial infections trigger acute adenolymphangitis attacks that are clinically distinct from FADL (Dreyer et al., 1999). Acute attacks of bacterial origin typically manifest as a diffuse cutaneous inflammatory response in 12
25 which the skin can be hot to the touch. Systemic manifestations such as fevers and chills are usually more severe than those seen in FADL, and attacks can cause a person to be bedridden for up to a week. It is thought that bacterial organisms gain entry to the host through skin lesions that form as lymphedema progresses, and streptococci have been cultured from the blood of individuals during an acute attack (Olszewski et al., 1999). Further evidence that these attacks are of bacterial origin is that hygienic practices that reduce normal skin flora drastically reduce the number of acute attacks that a person experiences (Shenoy et al., 1998; Dreyer et al., 1999). Recurrent attacks of adenolymphangitis are thought to be a common cause of lymphedema and elephantiasis; however, further work must be done to fully understand how bacteria contribute to disease development. This includes understanding the triggers that lead to heightened immune reactivity to bacterial antigens in persons with lymphedema. Wolbachia Bacteria of Filarial Worms One bacterial organism that has received much attention lately is an endosymbiont of the filarial nematodes that cause lymphatic filariasis. Ultrastructural analysis has shown this bacterium to be an obligate intracellular organism that lives harmoniously within filarial worms. The bacterium resides within a host vacuole along the length of the larval and adult lateral chords in both male and female worms and in the oogonia and oocytes of the female worm (Figure 1.1) (Kozek, 1977; Kozek and Marroquin, 1977; Taylor et al., 1999). It is gram-negative and exists in three distinct morphological forms similar to Chlamydia spp.: (1) a large bacillary form filled with granular material, (2) a spheroidal form with a dense central inclusion, and (3) another spheroidal form containing ribosome-like particles (Kozek, 1977). Furthermore, the presence of 13
26 this bacterial symbiont is widespread among filarial worms with the majority of species examined to date testing positive (Table 1.1). Although this symbiotic bacterium was initially discovered in filarial worms more than twenty years ago (McLaren et al., 1975), only recently have we begun to understand its interaction with the worm host. Phylogenetic analyses using 16S rrna and ftsz genes have shown this bacterium belongs to the genus Wolbachia, a rickettsia-like α-proteobacterium that also live symbiotically within ~20% of arthropod species (Sironi et al., 1995). In arthropods, Wolbachia is maternally transmitted to offspring via the cytoplasm of infected eggs and cause several reproductive abnormalities including feminization of genetic males, induction of parthenogenetic behavior, and expression of cytoplasmic incompatibility which ensures that infected females produce viable offspring only with males infected with the same Wolbachia strain or uninfected males (Braig et al., 1998). Wolbachia of filarial worms is also found in developing oocytes and share the same mode of transmission (Figure 1.2); however, it is not known whether Wolbachia of filarial worms is capable of influencing nematode reproduction in similar ways as it does in arthropods. Wolbachia of arthropods and nematodes form a monophyletic group that branches into four lineages (A-D) (Bandi et al., 1998). Groups A and B represent Wolbachia of arthropods. Group C contains Wolbachia of tissue-dwelling filarial nematodes (Onchocerca and Dirofilaria spp.), and Group D contains Wolbachia of lymphatic-dwelling filarial nematodes (Wuchereria, Brugia, and Litomosoides spp.). With the possible exception of Group C Wolbachia, evolutionary distances between groups are greater than the distances within groups, and the phylogenetic relationships of Wolbachia in filarial nematodes parallel the reported evolution of the filarial nematodes themselves (Xie et al., 1994; Casiraghi et al., 2001). This observation 14
27 suggests that the relationship between Wolbachia and filarial nematodes is ancestral and not the result of a recent horizontal transmission of Wolbachia from insects to filariae. These results lead to the hypothesis that the co-evolution between worm and bacterial species has resulted in a mutualistic association in which each partner benefits from the presence of the other. However, studies designed to further characterize the nature of this symbiotic relationship are complicated by the fact that all attempts to generate aposymbiotic worms have been unsuccessful. The initial hint that filarial nematodes depend on the presence of Wolbachia for their own development came when laboratory maintained jirds were prophylactically treated with tetracycline to prevent staphylococcal dermatitis (Bosshardt et al., 1993). Here, researchers found that prophylactic tetracycline treatment resulted in a 97% decrease in adult worm recovery after infection with B. pahangi L3. Similarly, if tetracycline therapy was initiated 27 days after infection, then the mean microfilaremia of treated animals was significantly lower than that of untreated controls. More recent studies have shown that indeed Wolbachia is susceptible to tetracycline and the death of the bacterium results in filarial infertility (Bandi et al., 1999; Hoerauf et al., 1999). A study by Casiraghi et al. (2002) suggests that tetracycline therapy affects worm viability by interfering with the moulting process in B. pahangi; however, these results do not explain how antibiotics could affect the microfilaria or adult stages since neither of these stages molt within the definitive host. One possible way in which antibiotics could affect worm viability that has not been addressed is that the death of the bacteria releases toxic substances within the worm that cause damage to worm tissues. Several groups are currently investigating the efficacy of antibiotic treatment for human filariasis, and these studies have the potential to provide novel chemotherapeutic approaches to the control of filariasis (Hoerauf et al., 2000). 15
28 Given the evidence that secondary bacterial infections contribute to the development of filarial lymphedema, it is also important to consider whether the presence of Wolbachia in filarial worms influences the outcome of filarial infection. There are several reasons to think that Wolbachia may be important in understanding disease caused by filarial worms. For example, there are a few cases already reported in which a parasitic worm is known to contain symbiotic bacteria that result in host pathogenicity. The trematode Nanophyetus salmincola is known to transmit Neorickettsia helminthoeca, an obligate intracellular bacterium of the family Rickettsiaceae, to canines using a fish intermediate host and resulting in salmon poisoning disease (Cordy and Gorham, 1950). Also, entomopathogenic nematodes are known to carry enteric gram-negative γ-proteobacteria that secrete toxins resulting in death of the host insect (Forst et al., 1997). These results emphasize that while symbiotic organisms may exist commensally in one host, bacteria or bacterial products can cause severe damage when released into the definitive host of the worm. Further evidence that Wolbachia may be of importance in understanding lymphatic filariasis comes from the possible involvement of Wolbachia in systemic inflammatory reactions following filarial chemotherapy. Common systemic side effects experienced by patients following treatment with the filaricidal drugs DEC and ivermectin include fever, headache, myalgia, and malaise and are thought to be the result of release of proinflammatory cytokines such as IL-1, IL-6, and TNF-α (Zheng et al., 1991; Yazdanbakhsh et al., 1992; Turner et al., 1994). A recent study by Keiser et al. (2002) showed that Wolbachia DNA could be detected in serum samples of patients infected with O. volvulus following treatment with either DEC or ivermectin and that peak DNA levels positively correlated with both serum TNF-α levels and the patient s clinical reaction score to treatment. Considering that CpG motifs in bacterial DNA are 16
29 a potent stimulator of innate inflammatory responses, these results suggest that Wolbachia may be a mediator of inflammatory responses seen following treatment. Similarly, a Wolbachia lipopolysaccharide (LPS)-like molecule in filarial worm extracts has been described that can stimulate inflammatory responses in C3H/HeN mice and macrophage cell lines (Brattig et al., 2000; Taylor et al., 2000). This Wolbachia LPS-like molecule has also been implicated in neutrophil infiltration and stromal haze when worm extracts were injected into the eye of LPS responsive mice (Saint Andre et al., 2002). While it is hypothesized that these LPS-like responses are involved in inflammatory responses following chemotherapy, definitive evidence is still lacking, and these results from laboratory models must be interpreted cautiously since the filarial parasite of humans that is associated with the most severe adverse reactions following treatment, Loa loa, does not contain Wolbachia (Buttner et al., 2003; Grobusch et al., 2003; McGarry et al., 2003). Nevertheless, these results are still consistent with the idea that Wolbachia antigens can stimulate host immune responses if released from filarial worms and may be a potential trigger for the development of filarial disease. Statement of Purpose The development of chronic filarial disease is a complex process that is the result of many interrelated factors. In addition to genetic and environmental factors, filarial infection and secondary bacterial infections are of significant importance, at least for lymphedema development. The key to fully understanding the mechanism of disease development is to understand how these and potentially many more unknown factors collectively contribute to the disease process. In particular, a better understanding of the factors that trigger the pathologic process could potentially explain why certain individuals develop chronic disease while others 17
30 do not. While it is accepted that the initial events in the development of disease are immunemediated destruction of lymphatic vessels, it is still unclear what causes the intense immune responses seen in persons with disease. Immune responses in these individuals exhibit a shift from down regulated Th-2 responses to inflammatory Th-1 biased immune responses. Because of the characteristic immune responses of actively infected individuals, it is likely that most worm antigens stimulate Th-2 biased immune responses, and antigens released following death of adult worms trigger this shift to a Th-1 response. Many bacterial infections trigger Th-1-like immune responses, and for this reason it is important to consider Wolbachia as a potential trigger for the initiation of the filarial disease process. As previously mentioned, Wolbachia DNA and LPS-like molecules may stimulate inflammatory responses of the innate immune system. While these responses may be of importance in understanding systemic inflammatory events following treatment, Wolbachia-specific immune responses of antigen specific B- and T-cells are more likely to contribute to the chronic immune activation seen in patients with disease. In order to investigate Wolbachia as a potential trigger for the development of chronic filarial disease, it is important to determine whether individuals living in areas endemic for lymphatic filariasis mount humoral and cell-mediated immune responses to Wolbachia antigens. If Wolbachia is involved in the development of disease, then Wolbachia-specific immune responses should be more common among individuals with lymphedema or hydrocele than among individuals without chronic disease. In addition, anti-wolbachia immune responses should be temporally related to disease development, and Wolbachia bacteria should be associated with inflammatory responses in vitro and in vivo. The following studies are designed to test these hypotheses. 18
31 Table 1.1. Listing of filarial species positive and negative for Wolbachia. 19 Parasite a Definitive Host 16S rdna Accession Number Reference(s) Wolbachia positive Brugia malayi Human AF Bandi et al., 1998; Taylor et al., 1999 Brugia pahangi Cat AF Bandi et al., 1998; Taylor et al., 1999 Brugia timori Human AF Fischer et al., 2002 Dipetalonema gracile Monkey AJ Casiraghi et al., 2004 Dirofilaria immitis Dog Z49261 Sironi et al., 1995 Dirofilaria repens Dog AJ Bandi et al., 1998 Dirofilaria tenuis Raccoon - b Punkosdy, unpublished Litosoma westi Gopher AJ Casiraghi et al., 2004 Litomosoides brasiliensis Bat AJ Casiraghi et al., 2004 Litomosoides galizai Rice Rat AJ Casiraghi et al., 2004 Litomosoides hamletti Bat AJ Casiraghi et al., 2004 Litomosoides sigmodontis Mouse, Rat AF Bandi et al., 1998 Mansonella ozzardi Human AJ Casiraghi et al., 2001b Onchocerca gibsoni Cattle AJ Bandi et al., 1998 Onchocerca gutturosa Cattle AJ Bandi et al., 1998 Onchocerca jakutensis Deer - b Plenge-Bonig et al., 1995 Onchocerca lupi Dog AJ Egyed et al., 2002 Onchocerca ochengi Cattle AJ Bandi et al., 1998 Onchocerca volvulus Human AF Henkle-Duhrsen et al., 1998 Wuchereria bancrofti Human AF Bandi et al., 1998; Taylor et al., 1999 Wolbachia negative Acanthoceilonema reconditum Dog - Casiraghi et al., 2004 Acanthoceilonema vitae Mouse - Bandi et al., 1998 Filaria martis Marten - Casiraghi et al., 2004 Foleyella furcata Chameleon - Casiraghi et al., 2004 Litomosoides yutajensis Bat - Casiraghi et al., 2004
32 Table1.1. (continued) Parasite a Definitive Host 16S rdna Accession Number Reference(s) Wolbachia negative (continued) Loa loa Human - Buttner et al., 2003; Grobusch et al., 2003; McGarry et al., 2003 Ochoterenella sp. Toad - Casiraghi et al., 2004 Onchocerca flexuosa Deer - Plenge-Bonig et al., 1995; Henkle-Duhrsen et al., 1998 Setaria equina Horse - Chirgwin et al., 2002 Setaria labiatopapillosa Cattle - Casiraghi et al., 2004 Setaria tundra Deer - Casiraghi et al., 2004 a Conflicting data exist for the presense/absense of Wolbachia in Mansonella perstans. Grobusch et al. (2003) report that Wolbachia is absent; however, 16S rdna sequence data have been deposited in Genbank (Accession number AY278355). 20 b Rickettsia-like endosymbionts have been observed in D. tenuis and O. jakutensis by immunostaining and electron microscopy, respectively; however, no molecular data exists.


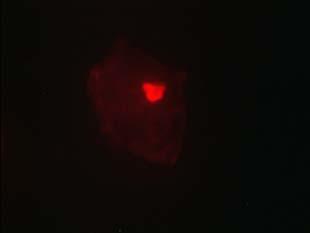
33 Figure 1.1. Localization of Wolbachia in adult female filarial worm. Specimen contains adult female Onchocerca volvulus worm stained with anti-wsp monoclonal antibody (red) and counterstained with hematoxylin (purple). 21
34 Figure 1.2. Immunolocalization of Wolbachia in the embryonic stages of development of B. pahangi worms. Each vertical row represents images of the same embryo that are either unstained (A, D, G, and J), stained with propidium iodide (B, E, and H), or stained with anti- WSP monoclonal antibody (C, F, I, and K). A-F. Localization of Wolbachia in very early embryos. G-I. Localization of Wolbachia in a morula stage embryo. Note the concentration of Wolbachia at one pole of the organism. J-K. Localization of Wolbachia in a fully developed embryo just before hatching. 22

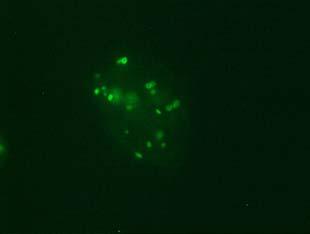
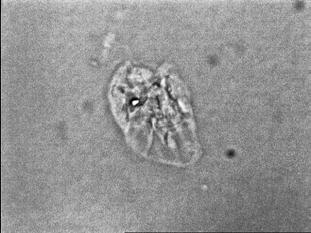
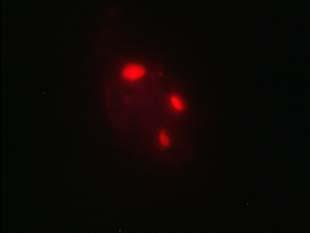
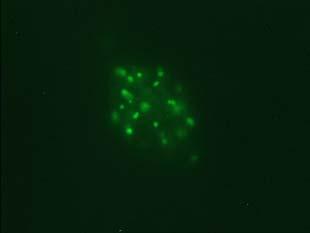
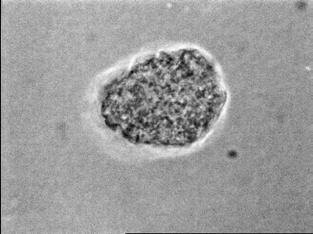
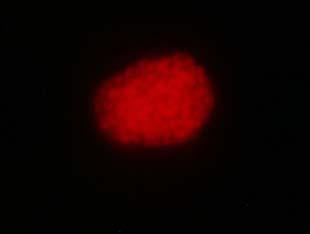
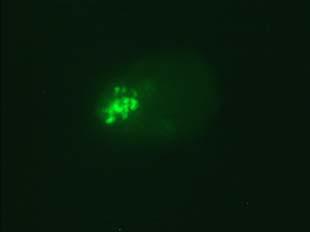
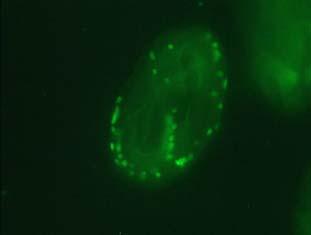
35 A B C D E F G H I J K 23
36 CHAPTER 2 DETECTION OF SERUM IgG ANTIBODIES SPECIFIC FOR WOLBACHIA SURFACE PROTEIN IN RHESUS MONKEYS INFECTED WITH BRUGIA MALAYI 1 1 Punkosdy, G.A., V.A. Dennis, B.L. Lasater, G. Tzertzinis, J.M. Foster, and P.J. Lammie The Journal of Infectious Diseases. 184: Reprinted here with permission of publisher. 24
37 Abstract The mechanism of lymphedema development in individuals with lymphatic filariasis is presently poorly understood. To investigate whether Wolbachia, symbiotic bacteria living within filarial nematodes, may be involved in disease progression, Wolbachia-specific immune responses were assayed in a group of Brugia malayi-infected rhesus monkeys. Serum IgG antibodies specific for a major Wolbachia surface protein (WSP) were detected in 2 of 12 infected monkeys. It is interesting that both of these monkeys developed lymphedema after becoming amicrofilaremic. WSP-specific antibody responses were temporally associated with increases in antifilarial IgG1 antibodies as well as lymphedema development. These findings suggest that Wolbachia may be important in understanding disease caused by filarial worms. Introduction Lymphatic filariasis is a debilitating parasitic disease affecting millions of people living throughout the tropics. Of these people, ~25 million exhibit disfiguring manifestations of lymphedema or elephantiasis. Although our understanding of the disease mechanism is incomplete, it is thought that lymphatic damage caused by adult worms, host immune responses, and secondary bacterial infections are all likely to be involved in disease progression (Freedman, 1998; Dreyer and Piessens, 2000). Recently, additional interest in this area has been sparked by the possibility that Wolbachia, intracellular symbiotic bacteria living within filarial worms, may play a role in pathogenesis. Although this idea was first proposed > 20 years ago (Kozek, 1977), only recently has experimental evidence in support of this hypothesis been generated (Bazzocchi et al., 2000; Brattig et al., 2000; Taylor et al., 2000). 25
38 If Wolbachia is involved in pathogenesis, then infected hosts may display Wolbachiaspecific immune responses. Studies designed to relate Wolbachia-specific immune responses to the natural history of infection and disease in humans are complicated by the fact that longitudinal specimens are difficult to obtain. As an alternative, Brugia malayi-infected rhesus monkeys are an excellent laboratory model for filariasis. In the present study, we utilized a recombinant form of a major Wolbachia surface protein (WSP) (Bazzocchi et al., 2000b) and serum samples from a previous longitudinal study involving Brugia malayi-infected rhesus monkeys (Dennis et al., 1998; Giambartolomei et al., 1998) to characterize host antibody responses to Wolbachia in lymphatic filariasis. Materials and Methods Animal infection. Fourteen male rhesus monkeys (Macaca mulatta) were used in this study. Animals were divided into 4 groups and infected as follows: 5 monkeys were infected with a bolus of 200 B. malayi third-stage larvae (L3) in RPMI 1640 medium (bolus infected group), as described elsewhere (Giambartolomei et al., 1998). Two monkeys were infected by repeated inoculations of 25 B. malayi L3 at ~1-month intervals over a period of 48 months (41 total trickle infections; trickle infection group). Five monkeys were initially infected with a bolus of 200 B. malayi L3 and then, after 96 weeks, were challenged by 41 inoculations of 25 B. malayi L3 at 1-month intervals (bolus + trickle infection group). Two control monkeys received injections of only RPMI 1640 medium. All injections were made subcutaneously in the lower right leg. Microfilaremia was monitored every 2 weeks via blood drawn at night. Serum samples were collected before infection and at ~ 4-week intervals after infection. All serum 26
39 samples were labeled alphanumerically and assayed by an investigator blinded to the infection status of the monkeys. WSP expression and purification. Brugia malayi genomic DNA extractions were performed by grinding a pool of adult worms in DNAzol (Gibco BRL), according to the manufacturer s instructions. Polymerase chain reaction primers were designed to amplify and directionally clone the entire coding sequence of the Wolbachia wsp gene minus the predicted N- terminal signal sequence (European Molecular Biology Laboratory accession no. AJ252061) (Bazzocchi et al., 2000b) into the Kpn1 and Pst1 restriction sites of the pqe41 expression vector (Qiagen). The forward primer was engineered to contain a thrombin cleavage site (shown underlined), so that WSP could be cleaved from its fusion partner (forward, 5 CGG GTA CCC CTG GTT CCG CGT GGA TCC CCT GTT GGT CCA ATA GCT G 3 ; reverse, 5 CAA CTG CAG TTA GAA ATT AAA CGC TAT TCC 3 ). Plasmids containing inserts were transformed into Escherichia coli JM109 competent cells (Promega), and a positive clone was selected by growth on Luria-Bertani plates containing carbenicillin. The identity of the resulting positive clone was confirmed by automated DNA sequencing. Expression of the recombinant WSP fusion protein was induced by the addition of isopropyl-β-d-thiogalactopyranoside to a final concentration of 1 mm. The recombinant fusion protein was purified using a nickelnitriloacetic acid (Ni-NTA) column in the presence of 8 M urea, according to the manufacturer s instructions (Qiagen). WSP protein was then cleaved from the dihydrofolate reductase (DHFR) fusion partner by overnight incubation with thrombin at room temperature, and pure WSP protein was isolated by passing the cleaved protein over a Ni-NTA column again to bind the DHFR fraction plus any uncleaved protein. Expression of recombinant WSP was monitored by SDS-PAGE and immunoblotting with a cross-reactive rabbit anti-wsp polyclonal antibody 27
40 raised against WSP from arthropod Wolbachia (a gift from S. O Neill, Yale University) (Dobson et al., 1999). Protein concentration was determined by using the bicinchoninic acid protein microassay (Pierce Biotechnology). ELISA. Filarial specific IgG1 antibody titers were determined as described elsewhere (Hitch et al., 1991). In brief, 96-well plates were coated with B. malayi adult worm antigen (2 µg/ml) diluted in 0.1 M NaHCO 3 by overnight incubation at 4 o C. Plates were then blocked with 0.3% PBST (0.1 M PBS + 0.3% Tween-20) for 1 h at 4 o C. Serum samples (1:50 in 0.05% PBST) were then added in duplicate. A standard curve consisting of 2-fold serial dilutions (1:10 to 1:1280) of a human serum sample with a known antifilarial IgG1 concentration was included on every plate. After washing, plates were incubated with a biotinylated mouse anti-human IgG1 secondary antibody (1:1000; Zymed) and streptavidin/alkaline phosphatase (1:500; Gibco BRL), with another washing step between. Plates were then developed by the addition of 0.1% p- nitrophenylphosphate in 3 mm MgCl 2 and 10% diethanolamine at ph ~ 9.8. Plate absorbance was read with a UVmax microplate reader (405 nm; Molecular Devices), and antibody levels were determined by comparison to the standard curve. Anti-WSP IgG antibodies were determined similarly, the only differences being in the concentration of secondary antibody and incubation times. First, 96-well plates were coated with WSP (0.5 µg/ml). Following overnight blocking, serum samples diluted in 0.3% PBST (1:50) were added in duplicate and were incubated overnight at 4 o C. A standard curve consisting of 2- fold serial dilutions (1:10 to 1:1280) of serum from a human with a high anti-wsp antibody titer was also included on every plate. The next day, plates were washed, and 50 µl of a mouse antihuman IgG secondary antibody (1:500; Zymed) was added for 2 h. Subsequent steps were performed as above. 28
41 Results All twelve monkeys that were given subcutaneous injections of infective larvae developed patent B. malayi infections. Following a week prepatent period, all monkeys in the bolus infection group, both monkeys in the trickle infection group, and 2 monkeys of the bolus + trickle group became microfilaremic and remained so throughout the entire study (Table 2.1). The other 3 monkeys in the bolus + trickle group (F-660, F-712, and F-585) became amicrofilaremic 15, 26, and 27 weeks after the bolus infection, respectively. One of these monkeys (F-585) became microfilaremic again following the initiation of trickle infections and remained microfilaremic. In addition, these same three monkeys (F-660, F-712, and F-585) developed >one episode of unilateral pitting lymphedema of the entire lower right leg and foot (site of L3 inoculation). Assays for WSP-specific IgG demonstrated detectable humoral responses in serum samples from only 2 of the 12 infected monkeys. Of interest, these 2 WSP-responding monkeys were also the same monkeys that developed lymphedema after becoming amicrofilaremic (F-660 and F-712). For monkey F-660, we saw an initial anti-wsp peak of 878 arbitrary units (U) around week 20 postinfection (PI) (Figure 2.1A). This corresponded to the point at which this monkey became amicrofilaremic and was immediately followed by an episode of lymphedema. Furthermore, this period of anti-wsp reactivity was closely associated with a peak of antifilarial IgG1 antibodies 25 weeks PI (Figure 2.1A). Following initiation of trickle infections at 96 weeks, another increase in antifilarial IgG1 (10.7 µg/ml) and anti-wsp IgG (1016 U) occurred that peaked at weeks 109 and 115 PI, respectively (13 and 19 weeks after trickle infection). Monkey F-660 later showed a third broader peak in anti-wsp IgG (978 U) around week 140 PI (46 weeks after trickle infection), coincident with a second episode of lymphedema from weeks 29
42 132 to 176 PI (38-82 weeks after trickle infection). Unlike the previous two peaks, this peak did not appear to be accompanied by an increase in antifilarial IgG1. Monkey F-712 experienced a similar course of infection with 3 episodes of anti-wsp reactivity (Figure 2.1B). The first anti-wsp episode occurred at 25 weeks PI, 1 week before this monkey became amicrofilaremic. In contrast to the initial anti-wsp response in monkey F- 660, the initial response in monkey F-712 was not accompanied by an episode of lymphedema. This monkey did, however, experience 2 other anti-wsp IgG responses that were temporally associated with lymphedema and peaked at weeks 165 and 214 PI (69 and 118 after trickle infection). All three peaks of WSP antibody reactivity were also associated with increases in antifilarial antibodies. In the other 10 infected monkeys, no anti-wsp reactivity was detected above background at any point during infection, even among monkeys that became amicrofilaremic. For example, monkey F-585 showed no anti-wsp response when it became amicrofilaremic 27 weeks after the initial infection (Figure 2.1C). This monkey also experienced an episode of lymphedema from weeks 134 to 182 PI (38-86 after trickle infection) that was not accompanied by anti-wsp reactivity, but this monkey remained microfilaremic throughout this time. Monkey F-661 also showed no evidence of anti-wsp reactivity throughout the entire study (Figure 2.1D). This monkey remained microfilaremic except for a period during weeks PI ( weeks after trickle infection) and, like monkey F-585, exhibited a low antifilarial IgG1 response after the initiation of the trickle infection. In their failure to develop WSP-specific antibody responses, these 2 monkeys were representative of all remaining B. malayi infected monkeys. In addition, control monkeys did not show anti-wsp or antifilarial antibody reactivity (data not shown). 30
43 Discussion We have demonstrated that a small proportion of B. malayi-infected rhesus monkeys exhibit IgG responses to a WSP. It is interesting to note that the 2 monkeys in which Wolbachiaspecific immune responses were detected both developed lymphedema after becoming amicrofilaremic. These results imply that WSP-specific antibody responses may be a useful marker for either disease development or worm death. Because Wolbachia bacteria are embedded within filarial worms, Wolbachia antigens will come into contact with components of the mammalian immune system only if bacterial products are somehow released from filarial worms. One plausible mechanism by which Wolbachia antigens could be released from filarial worms would be the release of bacteria or bacterial products after worm death. Because the mammalian host is home to several stages of the parasite life cycle, it is important to consider whether monkeys display Wolbachia-specific antibody responses after the death of microfilaria, L3 infective larvae, and/or adult worms. Results from this study suggest that infected monkeys do not mount a detectable anti-wsp IgG response after death of either L3 or microfilaria alone. Ten monkeys in this study were initially infected by injection of a large bolus of 200 infective larvae, many of which died and did not establish infection. In no instance, however, did we detect an anti-wsp IgG response immediately after infection. Similarly, transitions from microfilaremia to amicrofilaremia that were not accompanied by elevated antifilarial IgG1 levels (Figure 2.1C and D) were not associated with anti-wsp responses. This absence of WSP reactivity suggests that death of L3 or microfilaria through attrition is not sufficient to induce anti-wsp IgG responses. These results are in contrast to results showing that Dirofilaria immitis-infected cats universally mount antibody responses to WSP (Bazzocchi et al., 2000). While the explanation for these differences 31
44 is not entirely clear, perhaps the mechanism by which WSP is released differs between lymphatic- and non-lymphatic dwelling filarial worms. Alternatively, WSP responses in monkeys and humans (author's unpublished data) may be down-regulated in a Th2-predominant immune environment which accompanies active infection (King et al., 1993). In 2 of 3 monkeys that became amicrofilaremic after the bolus infection, the transition from microfilaremia to amicrofilaremia was accompanied by an anti-wsp IgG response (Figure 2.1A and B). In addition, monkey F-660 showed a second similar episode of anti-wsp reactivity ~19 weeks after the initiation of the trickle infections (Figure 2.1A). Although we have no direct way of assessing adult worm death in these monkeys, it is possible that, in addition to clearance of microfilaria, each of these 3 episodes was associated with the immunologically mediated death of the adult worms. Evidence in support of this conclusion comes from the observation that each episode of WSP reactivity was accompanied by an increase in antifilarial IgG1 antibodies (Figure 2.1A and B) and that both monkeys exhibited elevated T cell responses to adult filarial antigen (Giambartolomei et al., 1998). In this report, we also demonstrate an association between the development of lymphedema and WSP reactivity. Four of 5 observed episodes of lymphedema were temporally associated with increases in anti-wsp IgG production (Figure 2.1A and B). The single episode of lymphedema not associated with anti-wsp reactivity occurred in a monkey that was microfilaremic (Figure 2.1C). The explanation for the relationship between WSP reactivity and lymphedema is not clear, which reflects the uncertainty about whether the pathogenesis of filarial lymphedema is immune mediated or related to bacterial infections (Freedman, 1998). On the one hand, patients with lymphedema, in many settings, are predominantly filarial antigen negative, which implies a relationship between disease status and antifilarial immune status 32
45 (Lammie et al., 1993; Addiss et al., 1995). Perhaps the development of lymphedema in monkeys is associated with immune-mediated killing of adult worms, and WSP responses are only coincidentally associated with these events. On the other hand, opportunistic bacterial infections significantly contribute to acute attacks of adenolymphangitis and disease progression (Dreyer et al., 1999). As an alternative explanation, Wolbachia-specific antibody responses may be a marker or trigger of heightened antibacterial responses. In either case, further studies are needed to determine whether Wolbachia contributes to lymphedema development directly by stimulating B and T cell-dependent inflammation through antigen-specific pathways or indirectly by stimulating effector cells that cross-react with other bacterial antigens. 33
46 Table 2.1. Summary of infection outcome for rhesus monkeys in each of the 4 infection groups. Infection Group Infection Outcome a n Edema Wolbachia Surface Protein Reactivity Bolus Mf (+) 5 No No Trickle Mf (+) 2 No No Bolus + trickle Mf (+) 2 No No Mf (+) 1 Yes No Mf (+) Mf ( ) 2 Yes Yes Uninfected Mf ( ) 2 No No a Mf (+), microfilaremic; Mf ( ), amicrofilaremic. 34
47 Figure 2.1. Representative composite graphs showing the course of infection and antibody responses of rhesus monkeys in the bolus + trickle group. Bar graph in each panel represents the period of time that each monkey remained microfilaremic (black bar) and/or experienced lymphedema (gray bar). Line graphs represent anti-wolbachia surface protein (WSP) IgG (solid line) and antifilarial IgG1 (dashed line) antibody responses. Anti-WSP IgG values are given as arbitrary units on the left axis, and antifilarial IgG1 values are given as µg/ml equivalents of human antibody levels on the right axis. All animals were given the bolus infection at week 0, and the trickle infections were initiated at week 96. A, Monkey F-660; B, monkey F-712; C, monkey F-585; D, monkey F-661. Mf (+), microfilaremic. 35
48 A F-660 B F Mf (+) Edema 1 Mf (+) Edema units µg/ml units µg/ml C weeks F weeks D F-661 Mf (+) Edema Mf (+) Edema units µg/ml units µg/ml weeks weeks
49 CHAPTER 3 CHARACTERIZATION OF ANTIBODY RESPONSES TO WOLBACHIA SURFACE PROTEIN IN HUMANS WITH LYMPHATIC FILARIASIS 1 1 Punkosdy, G.A., D.G. Addiss, and P.J. Lammie Infection and Immunity. 71: Reprinted here with permission of publisher. 37
50 Abstract Symbiotic Wolbachia organisms of filarial nematodes have received much attention as possible chemotherapy targets and disease-causing organisms. In order to further investigate the association between anti-wolbachia immune responses and chronic filarial disease in humans, antibody responses to Wolbachia surface protein (WSP) were assayed in serum samples collected from 232 individuals living in Leogane, Haiti, an area where Wuchereria bancrofti infection is endemic, and from 67 North Americans with no history of lymphatic filariasis. As opposed to antifilarial antibody responses, which were largely influenced by the patient's infection status, the prevalence and levels of anti-wsp immunoglobulin G (IgG) antibodies among individuals with lymphedema or hydrocele were significantly greater than those in gender- and infection-matched individuals without disease. In at least one case, the anti-wsp IgG response was coincident with the onset of lymphedema development, and among anti-wsppositive women with lymphedema, anti-wsp IgG levels were negatively correlated with the duration of lymphedema. The presence of anti-wsp IgG was also associated with the severity of inguinal adenopathy among men with hydrocele. In addition to the presence of anti-wsp antibodies among Haitians, 15 of 67 (22%) serum samples collected from individuals from North America, where filariasis is not endemic, were also positive for anti-wsp antibodies. In comparison to those from Haitians, anti-wsp antibodies from North Americans primarily recognized a distinct region of WSP located within the highly conserved second transmembrane domain. The results of this study demonstrate that anti-wsp antibody responses are associated with the presence of chronic filarial morbidity and not filarial infection status in humans and suggest that WSP should be further studied as a potential trigger for the development of filarial disease. 38
51 Introduction Bancroftian filariasis is a mosquito transmitted parasitic disease of humans that has been considered to be potentially eradicable due to the inefficiency of transmission of the filarial parasites to humans and the fact that there are no zoonotic reservoir hosts of the parasite. The goals of the current global lymphatic filariasis elimination program are to (i) reduce microfilaremia levels, by using filaricidal drugs, to a level that is too low to sustain transmission of filarial parasites to humans and (ii) reduce the morbidity associated with chronic filarial disease (Cox, 2000). However, in order to achieve these goals, research efforts are still needed to develop better filaricidal drugs (especially macrofilaricides) and a better understanding of the etiology of chronic filarial disease. One aspect of the biology of filarial nematodes that may be exploited in the effort to advance the elimination program is the presence of a rickettsia-like endosymbiont belonging to the genus Wolbachia found inside many filarids. Recent studies of symbiotic Wolbachia organisms suggest that these bacteria may be potentially important as both chemotherapeutic targets and disease causing organisms. In animal models of filarial infection, treatment with antibiotics that specifically target Wolbachia decreases microfilaria loads, inhibits development of larval worms, and renders adult female worms infertile (Bosshardt et al., 1993; Bandi et al., 1999; Hoerauf et al., 1999; Rao and Well, 2002). In addition, high doses of antibiotics have been shown to have adulticidal effects in Onchocerca volvulus and Brugia malayi (Langworthy et al., 2000; Rao and Well, 2002). Other studies have shown that inflammatory responses induced by Wolbachia endotoxin may be responsible for the systemic adverse reactions following treatment with microfilaricidal drugs (Brattig et al., 2000; Taylor et al., 2000; Keiser et al., 2002). These results imply that therapy that eliminates Wolbachia may reduce the adverse reactions associated with current treatment 39
52 regimens. Human trials in Ghana are currently exploring the efficacy of using doxycycline as a possible treatment for human onchocerciasis (Hoerauf et al., 2000; Hoerauf et al., 2001). While the lengthy course of antibiotic therapy and the possibility of inducing antibiotic resistance may make anti-wolbachia treatment impractical as a public health measure, such therapy may be beneficial to patients on an individual basis (i.e., treatment for infected individuals returning from areas where filariasis is endemic). In addition to the possible role of Wolbachia as a chemotherapy target, evidence suggests that Wolbachia antigens can stimulate host immune responses that may be associated with the development of filarial disease. In a laboratory model of onchocerciasis, Wolbachia endotoxin has been shown to mediate neutrophil infiltration and stromal haze when a worm extract including Wolbachia antigens was injected into the eyes of mice (Saint Andre et al., 2002). Furthermore, we have shown that B. malayi-infected rhesus monkeys mount antibody responses to Wolbachia surface protein (WSP) that are temporally associated with the death of filarial worms and lymphedema development (Punkosdy et al., 2001). Although these studies suggest that Wolbachia may be important in understanding human disease caused by filarial worms, no studies to date have reported Wolbachia-specific immune responses among human populations with lymphatic filariasis. In the present study, we have assayed antibody responses to WSP in a cohort of Haitian individuals living in an area where Wuchereria bancrofti infection is endemic. The results reported here compare anti-wsp and antifilarial antibody responses among individuals with morbidity to those of individuals without morbidity to determine whether the presence of disease, as opposed simply to infection, is associated with anti-wsp antibody responses. 40
53 Materials and Methods Study population. Banked serum samples from 232 adult individuals living in Leogane, Haiti, an area where W. bancrofti infection is endemic, were selected based on serum availability for a retrospective analysis of antibody responses to WSP. In addition, 10 longitudinally collected serum specimens from one individual were available to assay anti-wsp antibody levels before and after the onset of lymphedema. Serum samples were collected over a 10-year period ranging from 1989 to 1999 and stored frozen at 20º C until use. All serum samples were collected before the initiation of the ongoing mass drug administration that is part of the lymphatic filariasis elimination program in this area. Individuals in this study were selected to represent the major parasitologic and clinical outcomes of infection seen in Leogane, Haiti. Infection status at the time of blood drawing was determined by the presence of microfilaremia and/or filarial antigenemia. Microfilaremia was assessed by filtering 1 ml of whole nocturnal blood through a Nuclepore filter, and microfilaremia was recorded as number of microfilariae (Mf) per milliliter of blood. Antigenemia was assessed by the commercially available ICT card test or antigen enzyme-linked immunosorbent assay (ELISA). Clinical disease status was determined by physical examination at the time the serum sample was collected. The two major clinical outcomes of infection seen in this area are hydrocele in men and lymphedema of the leg, primarily in women. In addition, serum samples from 67 North Americans with no history of filariasis were selected and assayed for anti-wsp antibody responses. All serum samples from human subjects were collected under protocols approved by the institutional review boards of the Centers for Disease Control and Prevention (CDC) and the University of Georgia. ELISA. Antifilarial immunoglobulin G1 (IgG1) and IgG4 antibody levels were determined by ELISA with a crude adult Brugia pahangi antigen extract as previously described 41
54 (Hitch et al., 1991). Levels of serum antibody to a recombinant WSP antigen were also determined by ELISA. The wsp gene from Wolbachia of B. malayi was cloned into the expression vector pqe41 (Qiagen, Valencia, Calif.), and the recombinant WSP protein was expressed and purified as previously described (Punkosdy et al., 2001). The wsp genes from Wolbachia of B. malayi (EMBL accession number AJ252062) and W. bancrofti (EMBL accession number AJ252180) share 97% identity at the nucleotide level and 98% identity at the amino acid level (Bazzocchi et al., 2000b). Ninety-six-well microtiter plates were coated with WSP (0.5 µg/ml) diluted in 0.1 M NaHCO 3 by overnight incubation at 4º C. Following a blocking period with 0.1 M phosphate buffered saline + 0.3% Tween 20 (0.3% PBST), human serum samples diluted 1:50 in 0.3% PBST were added to the wells in duplicate and incubated overnight at 4º C. The next day, plates were washed four times with 0.3% PBST and then incubated with a biotinylated mouse anti-human IgG secondary antibody (1:500; Zymed, South San Francisco, Calif.) for 2 h at room temperature. Following another wash step, plates were incubated with streptavidin-alkaline phosphatase (1:500; GibcoBRL, Grand Island, N.Y.) for 1 h at room temperature and subsequently developed by the addition of 0.1% p- nitrophenylphosphate-3 mm MgCl 2-10% diethanolamine (ph ~9.8). The optical density (OD) of each well was determined with a Molecular Devices UVmax microplate reader. Plates were allowed to develop to an optical density at 405 nm of 1 for the highest point on the standard curve (see below). Each plate contained a standard curve consisting of twofold serial dilutions (1:20 to 1:2,560) of a human serum sample determined to have a high antibody titer to WSP (data not shown). The highest point on the standard curve was assigned a value of 1,280 arbitrary units, and unit values for unknown serum samples were determined by comparison to the standard 42
55 curve. Determinations of duplicate serum samples with a coefficient of variation 15% were repeated. In addition to the standard curve, each plate contained three Haitian serum samples that were determined to be negative by Western blotting (data not shown). A cutoff for determining a positive anti-wsp response was determined independently for each plate by the mean unit values of the negative controls plus three standard deviations. Epitope mapping. Twenty-six biotinylated peptides were chemically synthesized to cover the entire predicted amino acid sequence of the B. malayi WSP protein (minus the N- terminal signal sequence). Peptides were synthesized as 18-mers that overlapped by nine amino acids. Peptides were solubilized according to the manufacturer s instructions, and 100 ng of each peptide (diluted in 0.05% PBST) was added to an individual well of streptavidin-coated microtiter plates in duplicate and incubated overnight at 4º C. The next day, microtiter plates were washed and blocked with a 1% casein-0.3% PBST solution. Serum samples that were determined to be positive for anti-wsp IgG antibodies by ELISA (n = 77) were assayed to determine which linear epitopes of the WSP protein were recognized. Because most IgG antibodies to WSP were of the IgG1 subclass (data not shown), we assayed IgG1 responses to the overlapping peptides. Human serum samples diluted 1:50 in 1% casein-0.3% PBST were added to each plate and incubated overnight at 4º C. Serum antibodies that recognized WSP peptides were detected by the addition of a secondary mouse anti-human IgG1 antibody (1:2000; provided by V. Tsang, CDC) followed by an alkaline phosphatase-labeled rabbit anti-mouse IgG antibody (1:1000; Zymed). Plates were then developed by the addition of 0.1% p- nitrophenylphosphate-3 mm MgCl 2-10% diethanolamine (ph ~9.8) as described above. Statistical analysis. Statistical analyses to assess differences in anti-wsp antibody responses among the different groups were performed with EpiInfo version 6.03 software 43
56 (CDC). The χ 2 test was used to compare difference in the seroprevalence of antibodies between groups, and the nonparametric Kruskal-Wallis H test was used to compare differences in median antibody levels between groups. A significant difference was defined as a P value of < Results As part of our previous studies using B. malayi-infected rhesus monkeys, we saw a temporal association between the development of anti-wsp antibody responses and lymphedema development (Punkosdy et al., 2001). In our studies in Leogane, we have few sets of longitudinal serum specimens from incident lymphedema cases. As an initial approach to investigate the potential association between antibody responses to WSP and chronic filarial disease in humans, we assayed longitudinal serum samples collected from one individual that were collected before and after the onset of disease. At the time the first serum sample was collected, this individual was a 54-year-old male living in Leogane, Haiti, with a microfilaremia of 100 Mf/ml of blood. This individual was enrolled in a treatment study and treated with diethylcarbamazine. After treatment, he became amicrofilaremic, and approximately 1 year after treatment, he developed unilateral lymphedema of the leg. All serum samples collected before the onset of lymphedema were negative for anti-wsp IgG; however, there was a significant but transient increase in anti-wsp IgG coincident with the onset of lymphedema (Figure 3.1). Antifilarial IgG1 antibody levels began to increase after the development of lymphedema and peaked at approximately 2 years after lymphedema development. There was no evidence of antifilarial IgG4 antibodies in any of the serum samples from this patient (Figure 3.1). This result prompted us to conduct a retrospective analysis of antibody responses to WSP in patients living 44
57 in an area where lymphatic filariasis is endemic to further investigate the association between anti-wsp IgG and filarial disease. We selected 232 serum samples collected from individuals living in Leogane, Haiti, and 67 serum samples collected from North Americans with no history of lymphatic filariasis to assay for the presence of anti-wsp IgG. Demographic and parasitologic characteristics of the study groups and anti-wsp seroprevalence data are shown in Table 3.1. Individuals with lymphedema or hydrocele were significantly more likely to be seropositive for anti-wsp IgG (anti-wsp + ) than asymptomatic antigen-positive and microfilaria-positive (Ag + Mf + ) (P = and P = 0.034, respectively), asymptomatic Ag Mf (P = and P = 0.004, respectively) and North American (P = and P = 0.004, respectively) individuals. Antibodies to WSP were also detected in 10 of 18 (56%) asymptomatic individuals who were Ag + Mf. Despite the small number of individuals in this group, they were also significantly more likely to be anti-wsp + than asymptomatic Ag + Mf + (P = 0.034), asymptomatic Ag Mf (P = 0.006) and North American (P = 0.006) individuals. However, median anti-wsp antibody levels did not differ significantly between any of these groups (data not shown). Due to the uncertainty as to whether individuals who were Ag + Mf harbored microfilaria at levels too low to be detected, had recently cleared infections with adult worms, or had single-sex filarial infections and the lack of statistically significant associations, these individuals were excluded from further analysis. There were no statistically significant differences in gender (41 versus 36% male; P = 0.50), median age (35 versus 31 years old; P = 0.11), or antigen or microfilaria prevalence (41 versus 43% Ag + Mf + ; P = 0.70) between anti-wsp + and anti-wsp individuals from Haiti, respectively. However, anti- WSP + individuals did have a slightly higher median antifilarial IgG1 level than anti-wsp individuals (2.2 versus 1.7 µg/ml; P = 0.048). Because of the differences in the pathogenesis of 45
58 filarial lymphedema and hydrocele, disease specific anti-wsp antibody results were analyzed separately for these two outcomes. Anti-WSP antibody responses among lymphedema patients. Of 44 individuals with lymphedema, 42 (95%) were female, 2 (5%) were Ag + Mf +, and 2 (5%) were Ag + Mf. Because the vast majority of patients with lymphedema were Ag Mf females, anti-wsp antibody results of these 40 women were compared to those of women in the other groups who had no evidence of lymphedema. The median anti-wsp antibody level was significantly higher for women with lymphedema than for asymptomatic Ag + Mf + women (P = 0.011), asymptomatic Ag Mf women (P = 0.009), and North American individuals (P = ) (Figure 3.2A). Therefore, women with lymphedema had significantly higher levels of serum anti-wsp IgG than gender- and infection-matched individuals without lymphedema. Although the median ages of these groups of women differed, there was no correlation between anti-wsp IgG levels and age. Women with lymphedema also had a significantly higher median antifilarial IgG1 level than asymptomatic Ag + Mf + women (P = ) and asymptomatic Ag Mf women (P = ) (Figure 3.2B). In contrast, antifilarial IgG4 levels were highest amongst asymptomatic Ag + Mf + women (Figure 3.2C). Among the 20 women in this study with lymphedema who were anti-wsp +, sufficient data regarding the duration of time since the onset of lymphedema were available for 12 (60%). Anti-WSP antibody levels were inversely correlated with lymphedema duration (P = 0.02) (Figure 3.3). Many studies have suggested a role for secondary bacterial infections in the progression of lymphedema development, thus raising the question of the possible association between host immune responsiveness to WSP and other bacterial antigens. As part of a previous study in our lab (Baird et al., 2002), immune responses of patients with lymphedema to various bacterial 46
59 antigens were assayed. Among 25 Ag Mf women with lymphedema who were included in both this study and the previous study and for whom antibody data for WSP and other bacterial antigens were available, there were no differences in median IgG responses to Pseudomonas exotoxin (41 versus 17 arbitrary units; P = 0.28), Staphylococcus enterotoxin B (202 versus 329 arbitrary units; P = 0.19), Streptococcus group A antigen (78 versus 62 arbitrary units; P = 0.55), or streptolysin O (131 versus 142 arbitrary units; P = 0.83) between individuals who were anti- WSP + and anti-wsp, respectively. In addition, there was no association between WSP positivity and the period of time since a patient s last acute attack of adenolymphangitis (7 versus 5.5 months; P = 0.86) or the number of acute attacks experienced in the previous 18 months (1.7 ± 1.5 versus 2.1 ± 1.6; P = 0.54). Anti-WSP antibody responses among hydrocele patients. Unlike patients with lymphedema, men with hydrocele form a heterogeneous group in which the percentage of men actively infected with filarial worms parallels the prevalence of microfilaremia seen in the community. Of 57 men in this study with hydrocele, 22 were Ag + Mf +, 23 were Ag Mf, and 12 were Ag + Mf. Median anti-wsp antibody levels of men with hydrocele were significantly greater than those of infection-matched men without hydrocele (Figure 3.4A). There were no differences in the median ages of any of the groups shown in Figure 3.4; therefore, men with hydrocele had significantly higher levels of anti-wsp IgG than age-, and infection-matched men without hydrocele. Consistent with a previous report (Addiss et al., 1995), antifilarial antibody responses among men in this study with hydrocele were influenced by infection status. Men with hydrocele who were Ag + Mf + had a significantly lower median antifilarial IgG1 response (P = 0.002) (Figure 3.4B) and a significantly higher median antifilarial IgG4 response (P = 0.001) 47
60 (Figure 3.4C) than men with hydrocele who were Ag Mf. Although not statistically significant, the same trend was seen in men without hydrocele. Sufficient data were available for 28 of 57 (49%) men with hydrocele to make comparisons between anti-wsp antibody responses and clinical observations. Among these 28 men, WSP-seropositivity was associated with the degree of inguinal adenopathy (P = 0.036) (Table 3.2). Although not statistically significant, a similar association was also seen between the percentage of men who were anti-wsp + and the presence of inguinal lymph node tenderness (P = 0.071). In addition, men who were anti-wsp + had a greater median hydrocele volume (241 versus 79 ml); however, this difference was not statistically significant (P = 0.21). Epitope mapping. Because 22% of serum samples collected from North Americans with no history of lymphatic filariasis were anti-wsp +, we attempted to determine whether there was anything unique about these responses that could possibly explain immune reactivity to WSP in areas where the infection is not endemic. Our approach was to determine whether individuals living in an area where lymphatic filariasis is endemic recognized different regions of the WSP protein than North Americans, by using 26 linear peptides that span the entire region of our recombinant WSP. Of 77 individuals in this study who were anti-wsp +, 52 (68%) had detectable serum IgG1 antibodies to at least one of the 26 WSP peptides, and positive individuals responded to an average of 2.3 ± 1.2 peptides (mean ± standard deviation). There were no differences in the percentage of individuals positive for at least one peptide or the average number of peptides recognized between any of the groups. The decreased sensitivity of the epitope mapping compared to the ELISA with recombinant WSP may be attributable to the specificity of this assay for IgG1 as opposed to total IgG or to the recognition of conformationally determined epitopes in the ELISA with recombinant WSP. In fact, when 48
61 serum samples were preabsorbed using individual peptides and then assayed for antibodies to recombinant WSP, we found that antibodies to the linear epitopes only accounted for only a fraction of the total antibodies (data not shown). However, there were still differences in antibodies to the linear peptides between Haitian and North American individuals. The percentage of individuals in each group who were positive for a particular peptide is shown in Figure 3.5. Anti-WSP + individuals with lymphedema or hydrocele primarily recognized peptides located at either the N- or C-terminal regions of the protein. Lymphedema patients responded primarily to peptides 1, 3, 5, and 23, and hydrocele patients to peptides 1 and 24 (Figure 3.5A and B). The majority of anti-wsp + asymptomatic microfilaremic individuals also responded to peptides 1 and 24; however, 38% of these individuals also responded to peptide 15 (Figure 3.5C). Twenty-nine percent of anti-wsp + normal individuals from the area of endemicity responded to peptides 1, 21, and 25, located at the N- and C-terminal regions of the protein. In addition, 43% and 57% of these individuals also responded to peptides 7 and 15, respectively (Figure 3.5D). In comparison, anti-wsp + North Americans showed a strikingly different pattern of reactivity. These individuals primarily responded to peptide 15, and they showed little recognition of peptides located at the N- and C-terminal regions of the WSP protein (Figure 3.5E). Predictions of structural motifs within WSP have repeatedly suggested the presence of two transmembrane domains (amino acids 112 to 127 and 137 to150) (Braig et al., 1998; Jiggins et al., 2002). The first 13 amino acids of peptide 15 (TPYVGVGLGVAYI) lie within the second transmembrane domain predicted by Jiggins et al. (2002). 49
62 Discussion The chronic manifestations of severe lymphatic filariasis are dominated by the clinical syndromes of lymphedema and hydrocele. Although both disease manifestations are characterized by accumulation of fluid in the affected part of the body, there is considerable debate about the underlying etiology and the pathologic mechanisms for these two clinical manifestations. While both manifestations are likely to share some aspects of their pathophysiologic processes, lymphedema is thought to be the outcome of a complicated interplay of parasitologic factors, host genetic factors, and secondary bacterial infections (Lammie et al., 2002). In contrast, the pathology of hydrocele has been considered to be almost entirely caused by the adult worm (Dreyer et al., 2000). Additionally, host inflammatory responses stimulated by parasite antigens are thought to contribute to the development of disease (Freedman, 1998); however, immune responses to adult worm extracts in these two groups are strongly associated with the patient s infection status. Although patients with lymphedema display the greatest levels of antifilarial immunity (Baird et al., 2002), similar types of responses are also seen in asymptomatic Ag Mf individuals (Dimock et al., 1996). In addition, antifilarial immune responses among men with hydrocele are more closely associated with the patient s infection status than the presence of disease (Addiss et al., 1995). As a result, specific parasite factors that trigger the inflammatory responses associated with disease development in these individuals have not yet been identified. In this report, we show that patients with either lymphedema or hydrocele were significantly more likely to mount antibody responses to WSP than gender- and infection-matched individuals without disease. In fact, men with hydrocele had remarkably similar seroprevalence and intensity of anti-wsp IgG responses independent of microfilaria or antigen status (Figure 3.4A). These results demonstrate that anti-wsp antibody responses are 50
63 associated with the presence of chronic filarial disease and not simply filarial infection status in humans. Further support for the association between anti-wsp antibody responses and filarial disease is provided by the associations between anti-wsp IgG and clinical markers of disease noted in this study. Consistent with our previous results in B. malayi-infected rhesus monkeys (Punkosdy et al., 2001), in the one case where longitudinal responses were assayed before and after the onset of disease, a transient peak in anti-wsp IgG was seen at the onset of lymphedema (Figure 3.1). Similarly, in cross-sectional data, there was an inverse correlation between anti- WSP IgG levels and lymphedema duration among anti-wsp + individuals (Figure 3.3). Also, among men with hydrocele, anti-wsp + men were more likely to have moderate degrees of inguinal adenopathy and inguinal lymph node tenderness than anti-wsp men. Because the lymphatic vessels of the spermatic cord (the primary site of adult W. bancrofti in men) drain into the abdominal lymph nodes rather than the inguinal lymph nodes, this observation is unlikely to represent a hydrocele-specific response. Therefore, similar complaints may be expected among men without hydrocele following worm death. Unfortunately, data concerning inguinal adenopathy and lymph node tenderness were only available for men with hydrocele. Nonetheless, the temporal association between anti-wsp antibody responses and filarial disease development and the association with inguinal adenopathy and tenderness suggest that Wolbachia should be further studied as a potential trigger for development of filarial disease. One other factor that has repeatedly been suggested to play an important role in the progression of filarial disease is lymphatic damage caused by secondary bacterial infections. Recurrent bacterial infections that manifest as acute dermatolymphangioadenitis have been shown to contribute to the development of chronic lymphedema (Dreyer et al., 1999). In 51
64 addition, patients with lymphedema have been shown to display heightened immune reactivity to bacterial antigens (especially streptolysin O) compared to infection-matched individuals without disease (Baird et al., 2002). This raises the question of the possible association between anti- Wolbachia immune responses and immune responses directed at other bacteria. In this study, we found there was no difference in levels of serum antibody to various bacterial antigens between anti-wsp + and anti-wsp women with lymphedema. Therefore, we believe that anti-wsp reactivity is not caused by cross-reaction with other bacterial infections that occur commonly among persons with lymphedema. Instead, it is likely that Wolbachia bacteria are recognized by the host immune system during the initial pathologic events following worm death and that secondary bacterial infections contribute to the progression of disease development only after these events lead to lymphatic stasis and an inability to clear invading organisms. To the extent that filarial pathology is associated with a shift in host response from a Th2- to a Th1-type immune response, an interesting hypothesis is that immune reactivity to Wolbachia may trigger this shift in host response to a Th-1 like immune response to both filarial and nonfilarial antigens. This heightened inflammatory reactivity to bacteria antigens then may be associated with increased lymphatic damage and skin pathology seen as disease progresses. Despite the associations between antibody reactivity to WSP and severe filarial disease noted in this study, only 45% of lymphedema patients and 47% of hydrocele patients were anti- WSP +. In the absence of longitudinal data, it is unclear whether any of the anti-wsp individuals had ever mounted antibody responses to WSP. Longitudinal data from humans (Figure 3.1) and monkeys (Punkosdy et al., 2001) demonstrate the importance of longitudinal data in analyzing antibody responses to WSP. In both cases, peaks in anti-wsp IgG were temporally associated with the onset of clinically apparent disease, and detectable levels of anti- 52
65 WSP IgG were transient. However, longitudinal specimens from humans who develop filarial disease are difficult to obtain, and as a result, it may be impossible to determine the true patterns of antibody responses to WSP using only cross-sectional data. An alternative approach may be to assay for cell-mediated immune responses to WSP. A critical component of the cell-mediated immune system is the production of memory T cells that can be stimulated with antigens in vitro to mount immune responses similar to that which they would mount in vivo. Considering the importance of T cells in the production of an effective antibody response (i.e., switching of the constant region of the heavy chain to produce IgG isotype antibodies), individuals who mount antibody responses to WSP would be expected to also display cell-mediated immunity to WSP. These experiments would have the added benefit of determining whether WSP could stimulate inflammatory-type immune responses that may serve as a potential trigger for the development of disease. In addition to detecting anti-wsp IgG in serum samples from Haitian individuals living in an area where lymphatic filariasis is endemic, we found that 22% of serum samples from North Americans with no history of lymphatic filariasis were also anti-wsp +. Based on this observation, there are at least three hypotheses that may explain why some North Americans are anti-wsp +. The first hypothesis, and perhaps the most difficult to test, is that some degree of cross-reactivity between WSP and unidentified bacterial antigens exists in human populations where the infection is not endemic. We consider this hypothesis unlikely given the lack of an association between WSP and other bacterial antigens and the observation that WSP epitopes recognized by North Americans in this study do not have significant homology to non- Wolbachia antigens when compared by BLAST search (data not shown). However, carefully controlled experiments would be needed to rule this hypothesis out. Alternatively, exposure to 53
66 other Wolbachia-containing filarial nematodes may elicit an anti-wsp antibody response. Dirofilaria and Mansonella spp. are endemic in many regions of North America, and anti-wsp antibody responses have been reported in Dirofilaria immitis-infected cats and humans (Bazzocchi et al., 2000; Simon et al., 2003). Finally, a third hypothesis is that Wolbachia of filarial worms does not represent the only means of human exposure to Wolbachia antigen(s). In this study, we found that the WSP epitopes primarily recognized by Haitians with lymphedema and hydrocele were concentrated at the amino- and carboxy-terminal ends of the protein, while the first 13 amino acids of the epitope that was primarily recognized by North Americans were located in the second transmembrane domain predicted by Jiggins et al. (2002). Interestingly, mathematical predictions based on the ratio of synonymous and nonsynonymous amino acid substitutions suggest that the transmembrane regions of WSP are not under positive selective pressure in either arthropod or nematode Wolbachia (Jiggins et al., 2002). These results suggest that the transmembrane regions of WSP are likely to have the greatest degree of sequence conservation between different strains of Wolbachia. In addition to filarial worms, Wolbachia bacteria also reside in a number of other invertebrates distributed throughout the world that are known to have contact with humans (Jeyaprakash and Hoy, 2000; Werren and Windsor, 2000). Although it has been reported that human exposure to Wolbachia antigens from arthropods does not occur (Zimmer, 2001), this hypothesis has not been empirically tested. In light of the observation that anti-wsp IgG can be detected in human subjects where the infection is not endemic, this hypothesis deserves further consideration. The recognition of WSP by the human immune system and the association between antibody responses to WSP and chronic filarial disease raise the question of whether Wolbachia may play a causative role in the development of filarial disease. Because Wolbachia bacteria are 54
67 located inside the filarial worm, it is likely that Wolbachia antigens will only come into contact with components of the host immune system only if they are released following worm death. Interestingly, a critical factor in the development of both lymphedema and hydrocele seems to be the death of the adult worm. Therefore, release of Wolbachia following worm death would put these bacteria in an environment in which Wolbachia-specific immune responses may trigger the initial events in the development of chronic filarial disease. Further studies to determine whether Wolbachia may play a causative role in the development of filarial disease should focus on the localized immune responses to Wolbachia following worm death. Analysis of histologic specimens collected following worm death may help determine whether Wolbachia is released following worm death and whether Wolbachia organisms or their antigens come into contact with components of the human immune system. In addition, an examination of cell-mediated immunity to Wolbachia antigens may help determine whether Wolbachia may contribute to the localized inflammation that is associated with disease. 55
68 Table 3.1. Comparison of anti-wsp antibody responses among the groups. Population Group n Median Age (yr) Age Range (yr) % M/F a WSP +b No. % Haitian Symptomatic Lymphedema / Hydrocele / Asymptomatic Ag + Mf / Ag Mf / Ag + Mf / North American Asymptomatic, Ag Mf 67 Unk c Unk Unk a M, male; F, female. b P = , by χ 2 test for significant differences among groups. c Unk, unknown.
69 Table 3.2. Association between anti-wsp antibody responses and clinical findings in men with hydrocele. Clinical Finding a n WSP + No. % Inguinal Adenopathy No Mild Moderate Inguinal Tenderness No Yes a P = and P = by χ 2 test for overall differences between groups for inguinal adenopathy and inguinal lymph node tenderness, respectively. 57
70 Ag (+)/ Mf (+) Edema Arbitrary Units ug/ml Weeks 0 Figure 3.1. Composite graph showing a temporal association between anti-wsp IgG responses and the onset of lymphedema. Longitudinal serum samples were collected from a 54-year-old male before and after the onset of lymphedema. This individual was treated with diethylcarbamazine at week zero and developed unilateral lymphedema of the leg approximately 1 year posttreatment. Horizontal bars represent the period of time during which this individual was Ag + Mf + (black bar) and experienced lymphedema (gray bar). Line graphs represent anti- WSP IgG levels (squares), given as arbitrary units on the left axis, and antifilarial IgG1 (circles) and antifilarial IgG4 (triangles) levels, given in micrograms per milliliter on the right axis. 58
71 Figure 3.2. Anti-WSP IgG levels are associated with the presence of lymphedema. Box and whisker plots show anti-wsp IgG (A), antifilarial IgG1 (B), and antifilarial IgG4 (C) antibody data from women in this study and North Americans. Horizontal lines represent the 25 th, 50 th, and 75 th percentiles of anti-wsp IgG responses, given as arbitrary units, and antifilarial responses, given in micrograms per milliliter. Vertical lines represent the nonoutlier minimum and maximum reponses for each group, and circles represent outliers. 59
72 350 A Arbitrary Units Lymphedema Ag (+)/Mf (+) Ag (-)/Mf (-) N American B ug/ml Lymphedema Ag (+)/Mf (+) Ag (-)/Mf (-) N American C ug/ml Lymphedema Ag (+)/Mf (+) Ag (-)/Mf (-) N American 60
73 Arbitrary Units Lymphedema Duration (yrs) Figure 3.3. Correlation between anti-wsp IgG levels and lymphedema duration among anti- WSP + women with lymphedema. Correlation (r = 0.66) was determined by linear regression analysis. 61
74 Figure 3.4. Anti-WSP IgG levels are associated with the presence of hydrocele. Box and whisker plots show anti-wsp (A), antifilarial IgG1 (B), and antifilarial IgG4 (C) antibody data from men in this study stratified by infection status. Individuals with hydrocele are shown on the left, and individuals without hydrocele are shown on the right. Horizontal lines represent the 25 th, 50 th, and 75 th percentiles of anti-wsp IgG responses, given as arbitrary units, and antifilarial responses, given in micrograms per milliliter. Vertical lines represent the nonoutlier minimum and maximum responses for each group, and circles represent outliers. 62
75 A Hydrocele Asymptomatic Arbitrary Units Ag (+)/Mf (+) Ag (-)/Mf (-) Ag (+)/Mf (+) Ag (-)/Mf (-) 25 B ug/ml Ag (+)/Mf (+) Ag (-)/Mf (-) Ag (+)/Mf (+) Ag (-)/Mf (-) 2 C 1.5 ug/ml Ag (+)/Mf (+) Ag (-)/Mf (-) Ag (+)/Mf (+) Ag (-)/Mf (-) 63
76 Figure 3.5. Linear epitopes of WSP recognized by anti-wsp + individuals with lymphedema (A) (n = 12) or hydrocele (B) (n = 16), asymptomatic Ag + Mf + individuals (C) (n = 8), asymptomatic Ag Mf individuals (D) (n = 7), and North Americans (E) (n = 9). Results are shown as the percentage of individuals recognizing at least one peptide who are positive for each of the 26 overlapping WSP peptides. 64
77 A Lymphedema B Hydrocele % Positive % Positive Peptides Peptides C Asymptomatic Ag (+)/Mf (+) D Asymptomatic Ag (-)/Mf (-) % Positive 40 % Positive Peptides Peptides E N American % Positive Peptides 65
78 CHAPTER 4 IMMUNOLOCALIZATION OF WOLBACHIA IN BIOPSY SPECIMENS COLLECTED FROM PATIENTS IN RECIFE, BRAZIL WITH BANCROFTIAN FILARIASIS Abstract Recent studies have suggested that endosymbiotic Wolbachia of filarial worms is released following worm death and contributes to the development of filarial pathology through the induction of inflammatory immune responses. In order to study the fate of Wolbachia in the localized environment following adult worm death, the in situ distribution of Wolbachia antigens in 19 histology specimens collected from Brazilian patients with lymphatic filariasis was assessed using a monoclonal antibody developed against Wolbachia surface protein (anti-wsp). Specimens studied included 14 granulomatous nodules with dead parasites and 5 lymphatic vessel segments biopsied from adjacent living worm nests. No Wolbachia antigen staining was observed in any of the lymphatic vessel segments. In the granulomatous nodules, Wolbachia antigen staining was observed outside the worms in only 4/14 (29%) of the specimens, usually in small amounts and in close association with the worm, and in one case (7%) Wolbachia antigen staining was identified inside inflammatory cells. However, there were no correlations between patient age, nodule age, or any histological characteristics of nodules that showed Wolbachia staining outside the worm and nodules that did not. These results suggest that the worm, and not Wolbachia, is probably the major antigenic source for the chronic granulomatous reaction associated with worm death in patients with lymphatic filariasis. 66
79 Introduction Lymphatic filariasis affects more that 100 million people and is one of the leading causes of morbidity worldwide. Although the pathogenesis of lymphatic filariasis is not completely understood, several studies have shown that distinct histological pictures and anti-filarial immune responses are associated with the presence of live and dead parasites and that these responses may influence the course of disease development. In virtually all individuals harboring live filarial parasites, the presence of the adult worm is accompanied by some degree of subclinical lymphangiectasia (Noroes et al., 1996). However, anti-filarial immune responses in these individuals are down-regulated and the lymphatic dilation is not associated with a host inflammatory response (Maizels et al., 2000; Figueredo-Silva et al., 2002). In contrast, the death of the adult worm triggers a shift in host immune responsiveness and leads to a characteristic tissue reaction around the dead worm. Clinically, adult worm death usually manifests in the form of a granulomatous nodule that is characterized by an inflammatory reaction composed primarily of epitheloid macrophages and multinucleated giant cells accompanied by eosinophils, lymphocytes, and plasma cells (Dreyer et al., 2000). The process of nodule formation is thought to progress from an inflammatory phase which is associated with the destruction of the filarial worm to a remodeling phase in which the inflammatory reaction is resolved, parasite antigens are sequestered, and the architecture of the lymphatic vessel is restored (albeit with some residual damage caused by the previous lymphangiectasia and the inflammatory response during the granulomatous reaction) (Figueredo-Silva et al., 2002). However, the exact mechanisms by which these events may occur are not entirely clear. Presumably, factors that are released by the dying worms play a critical role in the development of the inflammatory reaction; however, further work is needed in order to determine exactly what these factors are. 67
80 One potential candidate for a factor that may trigger the inflammatory reaction and the altered immunologic reactivity associated with worm death is Wolbachia, a rickettsia-like endosymbiont that resides within most species of filarial worms. There is a growing body of evidence suggesting that Wolbachia of filarial nematodes may be recognized by the mammalian immune system and that Wolbachia may influence the progression of the disease caused by these worms. Rhesus monkeys and humans with chronic manifestations of lymphatic filariasis display heightened antibody reactivity to Wolbachia Surface Protein (WSP) compared to infectionmatched individuals without clinically apparent disease (Punkosdy et al., 2001; Punkosdy et al., 2003). In animal models of onchocerciasis, a Wolbachia lipopolysaccharide (LPS)-like molecule has been shown to mediate neutrophil infiltration into the cornea (Brattig et al., 2001; Saint Andre et al., 2002). In addition, Wolbachia DNA levels in serum samples collected from individuals following treatment with microfilaricidal drugs have been shown to correlate with the systemic adverse reactions associated with treatment (Keiser et al., 2002). A more recent study has suggested an important role for the Toll-like receptor-2 (TLR-2) and TLR-4 in immune recognition of Wolbachia (Brattig et al., 2004). Taken together, these data suggest that immune responses directed at Wolbachia may play an important role in the development of filarial disease; however, further studies are needed to determine the fate of Wolbachia following worm death and the exact components of the host immune system that interact with Wolbachia. In this study, we have assessed the in situ distribution of Wolbachia antigens in histology specimens collected from Brazilian patients with lymphatic filariasis in order to determine whether Wolbachia contributes to the inflammatory reactions associated with worm death. 68
81 Materials and Methods Biopsy specimens. Nineteen biopsy specimens that were previously collected from 15 men infected with Wuchereria bancrofti and who resided in Recife, Brazil, were selected for the present study. All biopsy specimens were obtained during routine surgical procedures carried out at Nucleo de Ensino Pesquisa e Assistencia em Filariose (NEPAF), Universidade Federal de Pernambuco, Recife, Brazil, and all patients provided informed consent before undergoing surgical procedures. Tissue sections from formalin-fixed, paraffin-embedded specimens without personal identifying information were sent to the lab at CDC and processed to determine the in situ distribution of Wolbachia. All experiments using materials from human subjects were performed under protocols approved by the institutional review boards of Universidade Federal de Pernambuco, Centers for Disease Control and Prevention, and the University of Georgia. A chart review was conducted for these patients and the data collected included: age, clinical presentation (intrascrotal nodule or lymphangitis), mode of diagnosis (detection of microfilaria in a nocturnal blood sample and/or ultrasound), and treatment received. Nodule age was estimated based on the time interval between nodule formation (determined by physical exam and patient history) and surgery. Because of the known adulticidal effects of DEC and the association between DEC-treatment and the development of intrascrotal nodules (Noroes et al., 1997), nodules that were detected within 7 days of DEC treatment were considered to be DECrelated. One tissue section from each specimen was stained with hematoxylin and eosin and read by a pathologist (JG) who was blinded with respect to the clinical data. The histological features studied included: 1. degree of inflammation in the nodule recorded as abundant or mild, 2. inflammatory cell type present in the nodule (macrophages, giant cells, eosinophils), 3. presence 69
82 or absence of fibrosis (eosinophilic fibrillar material, collagen, and fibroblasts in the nodule), 4. necrosis defined as eosinophilic amorphous material in the nodule was quantified as marked (large collections of necrotic tissue) or mild (only a small rim around the worm). Production of anti-wsp monoclonal antibody. Two male BALB/c mice were immunized three times at approximate 1 month intervals by intraperitoneal injection with 5 µg of recombinant WSP antigen using TiterMax (Norcross, GA) as an adjuvant. Serum antibody levels to WSP were periodically monitored by ELISA as previously described (Punkosdy et al., 2001). The mouse with a higher titer of antibodies was given a final injection of 2.5 µg WSP in PBS in the tail vein and was sacrificed 3 days later. Its spleen cells were harvested, fused to SP2/O myeloma cells, and cultured at 37 o C and 5% CO 2 in Ultraculture media (Biowhittaker, Walkersville, MD) supplemented with glutamine, HAT, and 10% fetal bovine serum. Cell culture supernatants from individual hybridoma clones were assayed for antibodies to WSP by ELISA, and a clone with a high titer of antibodies specific for WSP was selected. The isotype of antibody produced by this clone was determined to be IgG2a using a mouse isotyping kit according to the manufacturer s instructions (Zymed, South San Francisco, CA). Proteins from 500 ml hybridoma cell culture supernatant were precipitated by slowly adding an equal volume of a saturated (NH 4 ) 2 SO 4 solution. The resulting pellet was resuspended in Protein A MAPS II binding buffer (Biorad, Hercules, CA) and dialyzed overnight. Monoclonal antibodies were purified by passage over a Protein A column and elution with Protein A MAPS II elution buffer according to the manufacturer s instructions. Purified antibodies were dialyzed into PBS and stored frozen at -80 o C. Immunohistochemistry. Five µm tissue sections were deparaffinized using xylene and ethanol and then rehydrated in PBS. Slides were blocked with 0.05% PBST (0.1M PBS % 70
83 Tween-20) plus 20% normal sheep serum (Equitech, Kerrville, TX). Tissue sections were incubated for 60 minutes with the anti-wsp monoclonal antibody. Optimal dilution of the antibody (1:250) in 0.05% PBST plus 20% normal sheep serum had been determined by previous experiments on positive control tissue. After being incubated with the primary antibody, slides were washed and biotinylated swine anti-mouse secondary antibody, streptavidin-alkaline phosphatase, and napthol fast red chromogenic substrate (LSAB2 Universal alkaline phosphatase system, DAKO Corporation, Carpinteria, CA) were sequentially added. Sections were then counterstained with Mayer s hematoxylin (Fisher Scientific, Pittsburgh, PA) and mounted using Faramount aqueous mounting medium (DAKO). The positive controls were formalin-fixed adult male and female Brugia pahangi worms (obtained from the Filariasis Repository at the University of Georgia, Athens). This antibody did not show cross reactivity with formalin-fixed, paraffin-embedded cultures of Staphylococcus aureus, Staphylococcus epidermidis, Streptococcus pyogenes, Rickettsia spp., Ehrlichia spp. and Candida albicans (clinical isolates at CDC). A negative control for each section consisted of the sequential section incubated with an unrelated isotype control antibody instead of the anti-wsp antibody. Comparisons and correlations between the observed Wolbachia staining patterns in the nodules with histologic and clinical data were performed to determine whether we could identify any factors that could be associated with the release of Wolbachia. Results Fifteen adult males (median age = 22 years, range years) who had previously undergone surgery at NEPAF, Universidade Federal de Pernambuco were included in this study. All were diagnosed with Wuchereria bancrofti infection based on clinical presentation of 71
84 intrascrotal nodule or lymphangitis, detection of microfilaria in a nocturnal blood sample, and/or ultrasound. At the time of their original enrollment, 10/15 (67%) were microfilaremic (107 mf/ml, range ) and 9/11 (82%) had intrascrotal worm nests detectable by ultrasound. Twelve of the fifteen individuals (80%) received anti-filarial treatment at the time of their original enrollment with the following drugs or combinations of drugs: diethylcarbamazine (DEC) (n = 8), ivermectin (n = 2), DEC + ivermectin (n = 1), and ivermectin + albendazole (n = 1). The dose of each drug received varied depending on the protocol of the original study; however, all doses were in the therapeutic range as defined by established protocols. Two individuals had undergone previous surgery for hydrocele three and sixteen years before the current procedure, and one individual had a history of retrograde lymphangitis and transitory acute edema of the leg. Two individuals presented with excess intrascrotal fluid at the time of the current surgery. A total of 19 biopsy specimens were collected from 15 patients and examined in this study (one patient had 4 specimens removed, another had 2, and the rest had 1). Of these specimens, 14 (74%) were granulomatous nodules containing adult W. bancrofti worms, and 5 (26%) were lymphatic vessels from adjacent living worm nests. All 14 nodules were intrascrotal and only one was associated with patient discomfort. Thirteen (93%) of these nodules were palpated on physical exam before surgery, and one was found during surgery for hydrocele. Nodules were estimated to range in age from 8 days to > 8 months and 4 were considered to be DEC-related (Table 4.1). Of the 5 lymphatic vessel specimens examined, 4 (80%) were from the intrascrotal lymphatics, and 1 (20%) was from the scrotal wall lymphatics. One patient was described as having a mixed reaction where a nodule containing dead parasites and a lymphatic vessel segment adjacent to living worms were collected from the same site. 72
85 Histology. All lymphatic nodules examined displayed a distinctive granulomatous inflammatory reaction around the parasites similar to that described in previous reports (Wartman, 1944; Michael, 1945; Jungmann et al., 1992; Figueredo-Silva et al., 2002). All nodules were characterized by a mononuclear cell infiltrate with the dominant cell type being activated macrophages with an epitheloid appearance (Figure 4.1) and giant cells were present in 6/14 (43%) nodules. Neutrophils were generally absent in all nodules, and eosinophils were present in 8/14 (57%). Fibrosis was present in 6/14 (43%) nodules. All nodules also contained necrotic areas. In 11/14 (79%) cases, necrosis was considered marked while in the other 3 (21%) necrosis was considered to be mild. In contrast, all lymphatic vessel specimens contained normal human tissue with no granulomatous inflammatory reaction. In fact, a mild mononuclear cell infiltrate was only seen in one specimen, and interestingly, this was the specimen that was collected from the individual who had a nodule with dead and living worms. Fibrosis was observed in 3/5 (60%) lymphatic vessel specimens, and no specimens contained increased areas of necrotic tissue. Adult W. bancrofti worm(s) were present in all 14 nodules examined. In 13/14 (93%) nodules, we were able to determine the sex of the worm(s). All of these nodules contained at least one female worm, and male worm(s) were present in 4 (31%). In 13/14 (93%) nodules, all worms were considered to be dead based on our analysis of worm morphology. In the nodule taken from the mixed reaction, in addition to an area containing at least one dead male and female worm, there was also a section that contained a live, healthy male worm. In all but this one case where the worm was determined to be alive, the tissue reaction extended to the cuticle of the worm(s) such that little or no lymph channel remained. In general, worms were degenerated and fragmented and the internal anatomy was largely deteriorated. In addition, 73
86 worms in 7/14 (50%) nodules were calcified. No worms were present in any of the lymphatic vessel specimens examined. Immunohistochemistry. In order to demonstrate the ability of our anti-wsp monoclonal antibody to detect Wolbachia in formalin fixed tissue specimens, we initially stained formalin fixed B. pahangi worms that were obtained from infected gerbils (Figure 4.2). Consistent with previous reports, Wolbachia bacteria were detected in the lateral chord(s) of both sexes and in developing embryos within the female uterus. The localization of Wolbachia using immunohistochemical stains for the 14 nodules examined in this study revealed 3 basic outcomes: (1) Wolbachia staining inside the worm (n = 8), (2) Wolbachia staining inside the worm and surrounding inflammation (n = 4), and (3) no staining inside the worm or surrounding inflammation (n = 2). Representative examples of groups 1 and 2 and shown in Figure 4.3. For worms that remained relatively intact, the Wolbachia staining patterns were similar to those observed in B. pahangi controls (figure 4.3A). As the worms appeared to be more degenerated it became more difficult to determine the exact tissue in the worm that was staining positive (Figure 4.3C and D). In some cases, a diffuse granular Wolbachia stain was seen throughout the remaining structure of the worm (Figure 4.3D and F). In general, as long as the cuticle of the worm was intact, Wolbachia staining was restricted to inside the worm. In the 4 cases where Wolbachia was found outside the worm the cuticle was separated from the worm, and only small packets of granular staining in the inflammatory infiltrate surrounding the worm could be observed (Figure 4.3E, F and G). There was only one case where granular staining was observed inside human macrophages/giant cells (Figure 4.3H). The specificity of the staining patterns observed was verified by the absence of 74
87 staining of consecutive tissue sections incubated with the isotype control antibody. We did not observe any staining for Wolbachia in any of the lymphatic vessel specimens. We also compared the observed Wolbachia staining patterns in the lymphatic nodules with our histologic and clinical results to determine whether we could identify any factors that could be associated with the release of Wolbachia. Comparing groups 1 and 2, there was no difference in mean age (25.3 ± 5.7 versus 25 ± 7.3 years, respectively); however, group 1 did tend to have higher microfilaria levels (1664 ± 1234 versus 501 ± 940, respectively). Our limited data suggests that the release of Wolbachia is independent of both granuloma age and whether nodules were DEC-related. While all three of the nodules in group 2 for which granuloma age could be estimated were < 2 months old, it is important to note that there were an equal number of nodules in group 1 that were in this same age range. In addition, 2/4 (50%) nodules that developed within 7 days of DEC-treatment had Wolbachia staining in the surrounding inflammation. In the case where we had 4 nodules collected from one individual, there was also a great deal of variation with respect to Wolbachia staining. We observed Wolbachia in the surrounding inflammation in only 1 of 4 nodules from this patient. Finally, there were no differences in inflammatory characteristics between nodules from groups 1 and 2. Figure 4.4 compares the inflammatory characteristics of similarly aged nodules from an individual in group 1 and an individual in group 2 (nodule age = 11 and 10 days, respectively). Both showed the typical granulomatous inflammatory reaction surrounding the worm (Figure 4.4A and B) including giant cells (Figure 4.4C and D). In addition, both nodules showed similar numbers of eosinophils (Figure 4.4E and F). 75
88 Discussion Evidence that Wolbachia stimulate inflammatory immune responses has led to the hypothesis that Wolbachia play a role in the pathogenesis of lymphatic filariasis and onchocerciasis (Taylor, 2003). Implicit in this argument is the assumption that Wolbachia bacteria and/or antigens are released following the death of the filarial worm and recognized by the human immune system. Support for this hypothesis has been generated in reports showing that Wolbachia DNA (Keiser et al., 2002) and anti-wolbachia antibodies (Punkosdy et al., 2001) can be detected in serum samples from infected individuals following worm death. However, it is still unclear how these responses relate to the development of filarial disease. Given the importance of the death of the adult filarial worm in the pathogenesis of lymphatic filariasis, we conducted the present study to analyze the earliest stages of the host response to Wolbachia. In our study, only 30% of the filarial nodules showed Wolbachia antigen staining outside the worms, usually in small amounts and in close association with the worm. Rarely, was Wolbachia antigen staining identified inside inflammatory cells. The fact that we did not observe Wolbachia antigen staining in human tissue of all nodules, all of which contained dead adult worms, suggests that the release of Wolbachia is influenced by additional factors. One factor that we considered might account for the observed patterns of Wolbachia release was whether nodule formation was related to anti-filarial treatment or not. The only currently available anti-filarial drug that has been shown to have adulticidal activity is DEC. Treatment of men with DEC is associated with the development of intrascrotal nodules that typically appear within one week of treatment and are caused by the death of the adult worm (Noroes et al., 1997). We observed Wolbachia staining in the surrounding human tissue in 50% of the nodules in this study that were considered DEC-related. However, we also observed 76
89 Wolbachia staining in 18% of the nodules that were not DEC-related ("spontaneous"). As a result, we believe that the release of Wolbachia in the nodules we examined appears to be independent of whether nodule formation was DEC-related or not and, instead, may be associated with the earlier events in nodule formation. Following the release of Wolbachia after worm death, by whatever mechanism, it is important to consider the fate of Wolbachia in human tissue. One possibility is that Wolbachia could, like many other rickettsial species, invade human cells and establish an infection. While we do not have any ultrastructural data to characterize Wolbachia morphology within human cells or determine where along the endocytic pathway the bacteria may accumulate, we do not believe that Wolbachia replicate in human cells. We did not observe overwhelming amounts of Wolbachia antigen staining in the human tissue surrounding dead worms or any staining in either of the nodules examined that were > 4 months of age. In addition, our attempts to maintain Wolbachia in culture in vitro using various cell lines have been unsuccessful thus far. Instead, we favor an alternative explanation that the release of Wolbachia is localized and that the bacteria are degraded after they come into contact with human immune cells and are presented to components of the adaptive immune system. This explanation is consistent with previous studies showing immune recognition of Wolbachia (Taylor et al., 2000; Brattig et al., 2001; Cross et al., 2001; Punkosdy et al., 2001; Punkosdy et al., 2003). The recognition of Wolbachia by the human immune system raises the question as to whether Wolbachia influences the development of the inflammatory reaction in the localized environment in which adult worm death takes place. While we can not definitively rule out the possibility that the release of Wolbachia in the earliest stages of nodule development influences the course of nodule formation, we did not observe any differences in inflammatory 77
90 characteristics between nodules where we could detect Wolbachia outside the filarial worm and nodules where we could not. This is consistent with a previous report demonstrating that there were no changes in the inflammatory characteristics of lymphatic lesions in B. pahangi-infected gerbils following treatment with tetracycline to clear Wolbachia infections (Chirgwin et al., 2003). The distinctive granulomatous response associated with adult worm death in lymphatic filariasis is characteristic of a chronic inflammatory reaction in response to a persistent antigenic source. This type of reaction is also seen in other chronic infections with more distantly related tissue-dwelling helminths that do not harbor Wolbachia, such as schistosomiasis, suggesting that the worm itself is a potent stimulator of chronic inflammation. In contrast, inflammatory reactions mounted in response to most bacterial species result an acute inflammatory reaction characterized by a polymorphonuclear cell response. Exceptions are bacteria that can exist in a chronic infectious state, such as Mycobacterium tuberculosis, Treponema pallidum, and Chlamydia spp., and in each of these infections, bacteria can usually be found at the site of the chronic inflammatory reaction (Beatty et al., 1994; Orme, 1998; Salazar et al., 2002). The fact that we did not see a persistent presence of Wolbachia in the inflammatory nodules examined suggests that the worm, and not Wolbachia, is probably the major antigenic source for the chronic granulomatous reaction. So, if Wolbachia is not contributing to the chronic inflammatory response associated with worm death, what are the consequences of anti-wolbachia immune responses and what effect do they have on the development of filarial disease? The absence of infection with adult filarial worms among individuals living in populations endemic for lymphatic filariasis is associated with a shift from a down-regulated Th2 immune response to an inflammatory Th1 response in peripheral blood lymphocytes (Maizels et al., 2000). Perhaps Wolbachia, through the immune 78
91 recognition of Th1 inducing LPS-like and CPG molecules, may contribute to this shift in peripheral immune responses. In addition, the development of disease in lymphatic filariasis, especially lymphedema, is associated with heightened immune reactivity to various bacterial antigens. While it has not been experimentally demonstrated, perhaps immune responses to Wolbachia in patients with lymphatic filariasis result in cross-reactivity to other bacteria, or vice versa, contributing to the development of disease through damage induced by secondary bacterial infections. Future studies designed to test these hypotheses may help to further define the potential role that Wolbachia play in the pathogenesis of lymphatic filariasis. 79
92 Table 4.1. Summary of histological results for specimens examined. 80 Granuloma Age Inflammation Parasite Degree Cell Type b Fibrosis Sex c Degeneration d Calcified DEC-related Wolbachia e > 8 days, < 2 months Mild M, E, GC Yes F 2 No No 2 > 9 days, < 4 months Abundant M, E No M/F 1-2 Yes No 1 10 days Abundant M, E, GC No F 3 No Yes 2 11 days Abundant M, E No F 3 Yes Yes 1 15 days Mild M, E Yes F 2 Yes No 3 26 days Adundant M, E No M/F 2 No Yes 1 34 days Mild M, E No F 2 No Yes 2 > 1 month, < 2 months Abundant M, GC No U 3 No No 1 4 months a Abundant M, GC Yes M/F 3 No No 1 8 months Mild M Yes F 3 Yes No 3 Unknown Mild M, GC Yes F 2 No No 1 Unknown a Abundant M, E Yes F 3 Yes No 2 Unknown a Abundant M, GC No F 3 Yes No 1 Unknown a Adundant M No M/F 2-3 Yes No 1 a Nodules were collected at the same time from a single patient. b M, macrophage; E, eosinophils; GC, giant cells. c M, male; F, female; U, unidentifiable. d (1), worm was intact and internal structures recognizable; (2), worm was degenerated but internal structures were still recognizable; (3), worm was degenerated and internal structures were no longer recognizable. e (1), Wolbachia staining was seen inside the worm; (2), Wolbachia staining was observed in the worm and surrounding inflammation; (3), no Wolbachia staining inside the worm or surrounding inflammation.

93 P N E F L Figure 4.1. Histological characteristics of W. bancrofti granuloma. Hematoxylin and eosin stain of an inflammatory nodule showing all of the inflammatory characteristics observed (E, epitheloid macrophages; F, fibrosis; L, lymphocytes; N, necrosis; P, parasite). 81

.")

 Female worm showing Wolbachia staining in the lateral chord")
94 A B C D Figure 4.2. Immunolocalization of Wolbachia in adult B. pahangi worms. Specimens are stained with anti-wsp monoclonal antibody (red) and counterstained with hematoxylin (purple). (A) Female worm showing Wolbachia staining in the lateral chords. (B) Male worm showing Wolbachia staining in the lateral chord. (C) Female worm showing Wolbachia staining in the lateral chord and uterus. (D) Higher magnification of a female worm to show granular staining pattern of Wolbachia in the lateral chord. 82
95 Figure 4.3. Immunolocalization of Wolbachia in inflammatory nodules. Images are representative of specimens with Wolbachia staining only in the filarial worm (A-D) and specimens with Wolbachia staining in the surrounding human tissue (E-H). Specimens are stained with anti-wsp monoclonal antibody (red) and counterstained with hematoxylin (purple). (A) Intact worm showing Wolbachia staining in the lateral chord (40X). (B) Calcified female worm with Wolbachia staining detectable in the lateral chord and uterus (40X). (C) Degenerated worm surrounded by necrotic tissue (40X). Wolbachia staining can be seen inside the body cavity of the worm. (D) Extremely degenerated parasites surrounded by host macrophages/giant cells (63X). Note that Wolbachia staining is only seen inside the remaining structure of the worm. (E) Dead parasite with cuticle separated from the body of the worm. Wolbachia staining can be seen outside the worm (40X). (F) Dead worm with Wolbachia staining in surrounding human tissue (40X). (G) Extremely degenerated parasite with Wolbachia staining located throughout the parasite and in the surrounding inflammation (40X). (H) Human macrophages/giant cells located in close proximity to a degenerated worm (63X). Wolbachia staining can be seen inside the human cells. 83

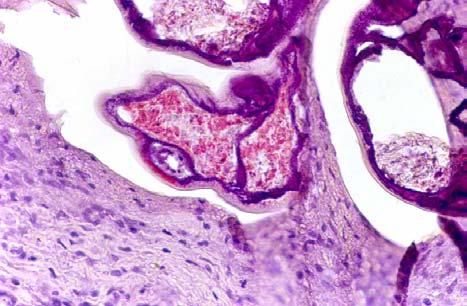
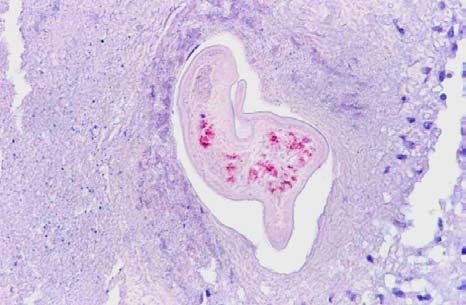
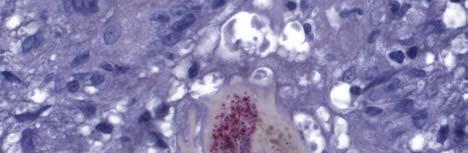
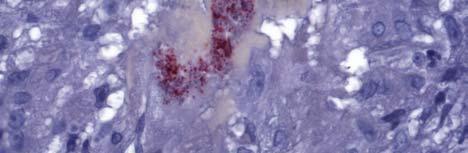
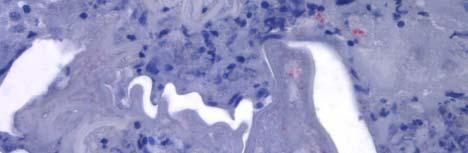
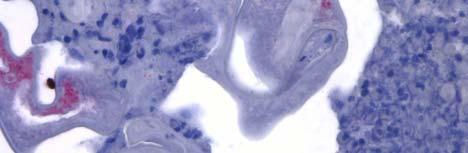
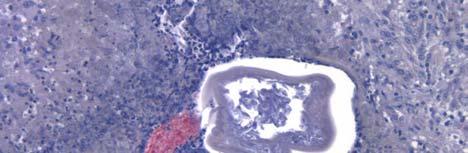
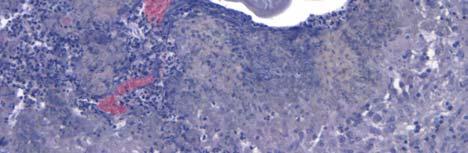
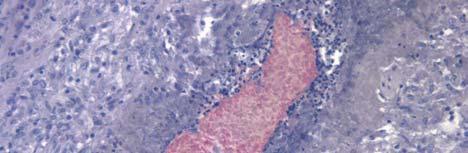

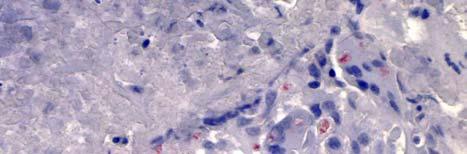
96 A B C D E F G H 84
97 Figure 4.4. Comparison of the inflammatory characteristics of similarly aged nodules where Wolbachia staining was only seen inside the filarial worm (A, C, and E) and where Wolbachia staining was seen in the surrounding inflammation (B, D, and F). Both nodules displayed a granulomatous inflammatory reaction (A and B) including giant cells (C and D). In addition, both nodules had similar numbers of eosinophils (E and F). 85

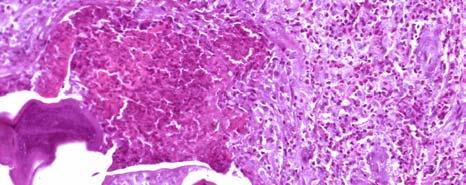
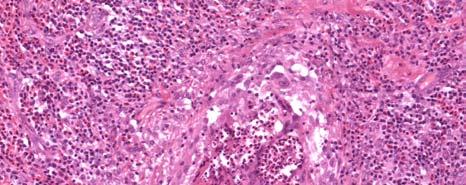
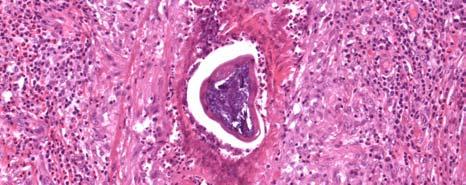
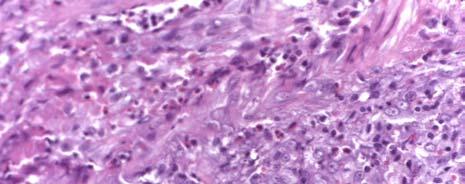
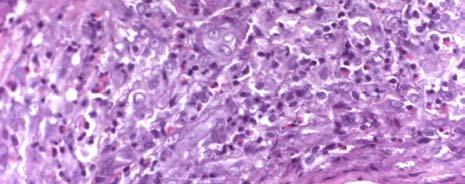
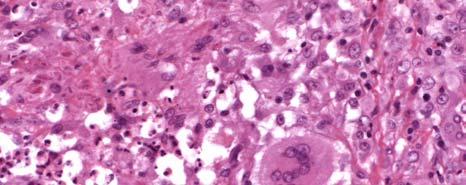
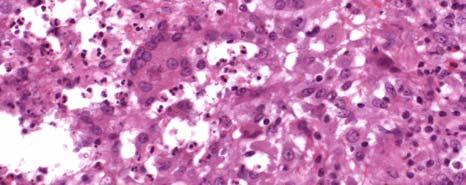
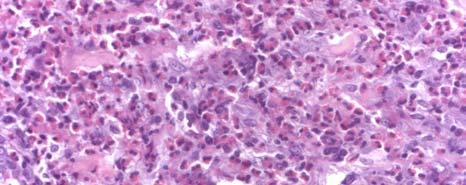
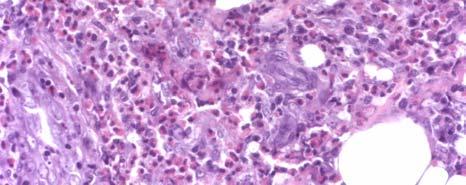
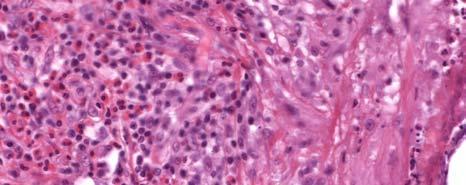
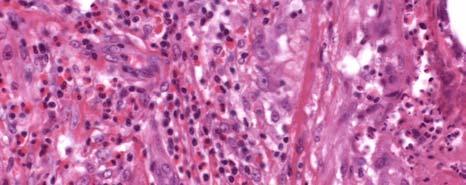
98 A B C D E F 86
PARASITOLOGY CASE HISTORY #13 (BLOOD PARASITES) (Lynne S. Garcia)
 (Lynne S. Garcia)") PARASITOLOGY CASE HISTORY #13 (BLOOD PARASITES) (Lynne S. Garcia) An epidemiologic survey was undertaken in a small town in Myanmar (Burma) endemic for lymphatic filariasis. Blood specimens were collected
PARASITOLOGY CASE HISTORY #13 (BLOOD PARASITES) (Lynne S. Garcia) An epidemiologic survey was undertaken in a small town in Myanmar (Burma) endemic for lymphatic filariasis. Blood specimens were collected
Lymphatic Filariasis
 REFERENCES CONTENT ALERTS Characterization of Antibody Responses to Wolbachia Surface Protein in Humans with Lymphatic Filariasis George A. Punkosdy, David G. Addiss and Patrick J. Lammie Infect. Immun.
REFERENCES CONTENT ALERTS Characterization of Antibody Responses to Wolbachia Surface Protein in Humans with Lymphatic Filariasis George A. Punkosdy, David G. Addiss and Patrick J. Lammie Infect. Immun.
Helminths in tropical regions
 Helminths in tropical regions Schistosoma spp. Blood flukes Schistosomiasis is one of the most widespread parasitic infections in humans Humans are the principal hosts for: Schistosoma mansoni, Schistosoma
Helminths in tropical regions Schistosoma spp. Blood flukes Schistosomiasis is one of the most widespread parasitic infections in humans Humans are the principal hosts for: Schistosoma mansoni, Schistosoma
DIAGNOSTIC SLIDE SESSION CASE 10
 DIAGNOSTIC SLIDE SESSION CASE 10 B.K. Kleinschmidt-DeMasters, MD Disclosures: I am not a trainee Caterina made me do this CASE 2016-10: : The patient is a 5-year-old girl with Down syndrome, obstructive
DIAGNOSTIC SLIDE SESSION CASE 10 B.K. Kleinschmidt-DeMasters, MD Disclosures: I am not a trainee Caterina made me do this CASE 2016-10: : The patient is a 5-year-old girl with Down syndrome, obstructive
Wuchereria Morphology 10 cm 250 : m
 Wucheria bancrofti Brugia malayi Lymphatic filariasis Lymphatic Filariasis 119 million infected Elephantiasis Manifestation of lymphatic filariasis Morphology I Adult: White and thread-like. Two rings
Wucheria bancrofti Brugia malayi Lymphatic filariasis Lymphatic Filariasis 119 million infected Elephantiasis Manifestation of lymphatic filariasis Morphology I Adult: White and thread-like. Two rings
Lecture 5: Dr. Jabar Etaby
 Lecture 5: Dr. Jabar Etaby 1 2 Onchocerca volvulus (Blinding filariasis; river blindness) Microfilaria of Onchocerca volvulus, from skin snip from a patient seen in Guatemala. Wet preparation 3 Some important
Lecture 5: Dr. Jabar Etaby 1 2 Onchocerca volvulus (Blinding filariasis; river blindness) Microfilaria of Onchocerca volvulus, from skin snip from a patient seen in Guatemala. Wet preparation 3 Some important
IMMUNO-EPIDEMIOLOGY OF BANCROFTIAN FILARIASIS : A 14-YEAR FOLLOW-UP STUDY IN ODISHA, INDIA
 IMMUNO-EPIDEMIOLOGY OF BANCROFTIAN FILARIASIS : A 14-YEAR FOLLOW-UP STUDY IN ODISHA, INDIA NN Mandal, KG Achary, SK Kar and MS Bal Division of Immunology, Regional Medical Research Centre, Indian Council
IMMUNO-EPIDEMIOLOGY OF BANCROFTIAN FILARIASIS : A 14-YEAR FOLLOW-UP STUDY IN ODISHA, INDIA NN Mandal, KG Achary, SK Kar and MS Bal Division of Immunology, Regional Medical Research Centre, Indian Council
I. Wuchereria bancrofti
 Parasites that affect the Musculoskeletal system (continued) Filarial Worms - Nematodes. - Tissue parasites. - Require an intermediate host, which is usually an insect. - Do not lay eggs like other worms,
Parasites that affect the Musculoskeletal system (continued) Filarial Worms - Nematodes. - Tissue parasites. - Require an intermediate host, which is usually an insect. - Do not lay eggs like other worms,
Ascaridida. Ascaridoidea* Heterakoidea* Cosmocercoidea Subuluroidea Seuratoidea
 Ascaridida Ascaridoidea* Heterakoidea* Cosmocercoidea Subuluroidea Seuratoidea Ascaridoidea Parasites in intestine of all classes of vertebrates. Aquatic cycles use invertebrate and vertebrate (fish) intermediate
Ascaridida Ascaridoidea* Heterakoidea* Cosmocercoidea Subuluroidea Seuratoidea Ascaridoidea Parasites in intestine of all classes of vertebrates. Aquatic cycles use invertebrate and vertebrate (fish) intermediate
Downloaded from:
 O Hara, GA; Elliott, AM (2016) HIV and Helminths - Not All Worms Created Equal? Trends in parasitology. ISSN 1471-4922 DOI: https://doi.org/10.1016/j.pt.2016 Downloaded from: http://researchonline.lshtm.ac.uk/3327115/
O Hara, GA; Elliott, AM (2016) HIV and Helminths - Not All Worms Created Equal? Trends in parasitology. ISSN 1471-4922 DOI: https://doi.org/10.1016/j.pt.2016 Downloaded from: http://researchonline.lshtm.ac.uk/3327115/
Epidemiology and immunopathology of bancroftian filariasis
 Microbes and Infection, 1, 1999, 1015 1022 1999 Éditions scientifiques et médicales Elsevier SAS. All rights reserved Review Epidemiology and immunopathology of bancroftian filariasis Adriana B. de Almeida*,
Microbes and Infection, 1, 1999, 1015 1022 1999 Éditions scientifiques et médicales Elsevier SAS. All rights reserved Review Epidemiology and immunopathology of bancroftian filariasis Adriana B. de Almeida*,
How the Innate Immune System Profiles Pathogens
 How the Innate Immune System Profiles Pathogens Receptors on macrophages, neutrophils, dendritic cells for bacteria and viruses Broad specificity - Two main groups of bacteria: gram positive, gram-negative
How the Innate Immune System Profiles Pathogens Receptors on macrophages, neutrophils, dendritic cells for bacteria and viruses Broad specificity - Two main groups of bacteria: gram positive, gram-negative
PBS Class #2 Introduction to the Immune System part II Suggested reading: Abbas, pgs , 27-30
 PBS 803 - Class #2 Introduction to the Immune System part II Suggested reading: Abbas, pgs. 15-25, 27-30 Learning Objectives Compare and contrast the maturation of B and T lymphocytes Compare and contrast
PBS 803 - Class #2 Introduction to the Immune System part II Suggested reading: Abbas, pgs. 15-25, 27-30 Learning Objectives Compare and contrast the maturation of B and T lymphocytes Compare and contrast
SEA-CD-275. Frequently asked questions
 SEA-CD-275 Frequently asked questions on LYMPHATIC FILARIASIS (ELEPHANTIASIS) World Health Organization 2013 All rights reserved. Requests for publications, or for permission to reproduce or translate
SEA-CD-275 Frequently asked questions on LYMPHATIC FILARIASIS (ELEPHANTIASIS) World Health Organization 2013 All rights reserved. Requests for publications, or for permission to reproduce or translate
Parasitic Protozoa, Helminths, and Arthropod Vectors
 PowerPoint Lecture Slides for MICROBIOLOGY ROBERT W. BAUMAN Chapter 23 Parasitic Protozoa, Helminths, and Arthropod Vectors Helminthic Parasites of Humans Helminths are macroscopic, multicellular, eukaryotic
PowerPoint Lecture Slides for MICROBIOLOGY ROBERT W. BAUMAN Chapter 23 Parasitic Protozoa, Helminths, and Arthropod Vectors Helminthic Parasites of Humans Helminths are macroscopic, multicellular, eukaryotic
Early tissue migration of and host response to Brugia pahangi in gerbils
 Louisiana State University LSU Digital Commons LSU Master's Theses Graduate School 2004 Early tissue migration of and host response to Brugia pahangi in gerbils Kristina Houpe Porthouse Louisiana State
Louisiana State University LSU Digital Commons LSU Master's Theses Graduate School 2004 Early tissue migration of and host response to Brugia pahangi in gerbils Kristina Houpe Porthouse Louisiana State
The Filarial Dance Sign in Scrotal Filarial Infection
 The Filarial Dance Sign in Scrotal Filarial Infection Poster No.: C-0232 Congress: ECR 2015 Type: Authors: Keywords: DOI: Educational Exhibit P. P. Suthar 1, N. Patel 2 ; 1 Vadodara, Gu/IN, 2 Vadodara,
The Filarial Dance Sign in Scrotal Filarial Infection Poster No.: C-0232 Congress: ECR 2015 Type: Authors: Keywords: DOI: Educational Exhibit P. P. Suthar 1, N. Patel 2 ; 1 Vadodara, Gu/IN, 2 Vadodara,
Evaluating Vaccine Candidates for Filariasis. Christopher Paul Morris
 Evaluating Vaccine Candidates for Filariasis by Christopher Paul Morris Dissertation submitted to the Faculty of the Emerging Infectious Disease Graduate Program Uniformed Services University of the Health
Evaluating Vaccine Candidates for Filariasis by Christopher Paul Morris Dissertation submitted to the Faculty of the Emerging Infectious Disease Graduate Program Uniformed Services University of the Health
44th Myanmar Health Research Congress
 44th Myanmar Health Research Congress Early Detection of Lymphatic Disturbance in Adolescent Infected with Lymphatic Filariasis Jan Douglass 1, Susan Gordon 1, Patricia Graves 1, Ben Dickson 1, Dr Ni Ni
44th Myanmar Health Research Congress Early Detection of Lymphatic Disturbance in Adolescent Infected with Lymphatic Filariasis Jan Douglass 1, Susan Gordon 1, Patricia Graves 1, Ben Dickson 1, Dr Ni Ni
The impact of density-dependent processes on the eradicability of parasitic diseases
 The impact of density-dependent processes on the eradicability of parasitic diseases Hans Peter Duerr Martin Eichner Klaus Dietz Department of Medical Biometry University of Tübingen Oberwolfach 2004 1/12
The impact of density-dependent processes on the eradicability of parasitic diseases Hans Peter Duerr Martin Eichner Klaus Dietz Department of Medical Biometry University of Tübingen Oberwolfach 2004 1/12
POSTGRADUATE INSTITUTE OF MEDICINE UNIVERSITY OF COLOMBO MD (MEDICAL PARASITOLOGY) EXAMINATION JANUARY, 2001 PAPER 1
 EXAMINATION JANUARY, 2001 PAPER 1") JANUARY, 2001 Date: 15 th January 2001 Time: 2.00 p.m. -500 p.m. PAPER 1 Answer all five (5) questions Answer each question in a separate book 1. Discuss the underlying principles relating to the use of
JANUARY, 2001 Date: 15 th January 2001 Time: 2.00 p.m. -500 p.m. PAPER 1 Answer all five (5) questions Answer each question in a separate book 1. Discuss the underlying principles relating to the use of
Clinical and Laboratory Aspects of Filariasis
 CLINICAL MICROBIOLOGY REVIEWS, Jan. 1989, p. 39-50 Vol. 2, No. 1 0893-8512/89/010039-12$02.00/0 Copyright 1989, American Society for Microbiology Clinical and Laboratory Aspects of Filariasis JAYASRI NANDURI
CLINICAL MICROBIOLOGY REVIEWS, Jan. 1989, p. 39-50 Vol. 2, No. 1 0893-8512/89/010039-12$02.00/0 Copyright 1989, American Society for Microbiology Clinical and Laboratory Aspects of Filariasis JAYASRI NANDURI
Changes in Cytokine, Filarial Antigen, and DNA Levels Associated With Adverse Events Following Treatment of Lymphatic Filariasis
 The Journal of Infectious Diseases MAJOR ARTICLE Changes in Cytokine, Filarial Antigen, and DNA Levels Associated With Adverse Events Following Treatment of Lymphatic Filariasis Britt J. Andersen, 1 Jessica
The Journal of Infectious Diseases MAJOR ARTICLE Changes in Cytokine, Filarial Antigen, and DNA Levels Associated With Adverse Events Following Treatment of Lymphatic Filariasis Britt J. Andersen, 1 Jessica
ACTIVATION OF T LYMPHOCYTES AND CELL MEDIATED IMMUNITY
 ACTIVATION OF T LYMPHOCYTES AND CELL MEDIATED IMMUNITY The recognition of specific antigen by naïve T cell induces its own activation and effector phases. T helper cells recognize peptide antigens through
ACTIVATION OF T LYMPHOCYTES AND CELL MEDIATED IMMUNITY The recognition of specific antigen by naïve T cell induces its own activation and effector phases. T helper cells recognize peptide antigens through
The role of endosymbiotic Wolbachia bacteria in filarial disease
 Blackwell Science, LtdOxford, UKCMICellular Microbiology1462-5814Blackwell Publishing Ltd, 2003February 20046297104Review ArticleA. G. Hise, I. Gillette-Ferguson and E. PearlmanWolbachia endobacteria in
Blackwell Science, LtdOxford, UKCMICellular Microbiology1462-5814Blackwell Publishing Ltd, 2003February 20046297104Review ArticleA. G. Hise, I. Gillette-Ferguson and E. PearlmanWolbachia endobacteria in
Blood Smears Only 5 February Sample Preparation and Quality Control 13B A
 NEW YORK STATE Parasitology Proficiency Testing Program Blood Smears Only 5 February 2013 The purpose of the New York State Proficiency Testing Program in the category of Parasitology Blood Smears Only
NEW YORK STATE Parasitology Proficiency Testing Program Blood Smears Only 5 February 2013 The purpose of the New York State Proficiency Testing Program in the category of Parasitology Blood Smears Only
Effect of aggressive prolonged diethylcarbamazine therapy on circulating antigen levels in bancroftian lariasis
 Tropical Medicine and International Health volume 6 no 1 pp 37±41 january 2001 Effect of aggressive prolonged diethylcarbamazine therapy on circulating antigen levels in bancroftian lariasis David O. Freedman
Tropical Medicine and International Health volume 6 no 1 pp 37±41 january 2001 Effect of aggressive prolonged diethylcarbamazine therapy on circulating antigen levels in bancroftian lariasis David O. Freedman
Cover Page. The handle holds various files of this Leiden University dissertation.
 Cover Page The handle http://hdl.handle.net/1887/20942 holds various files of this Leiden University dissertation. Author: Wammes, Linda Judith Title: Immune regulation during parasitic infections : from
Cover Page The handle http://hdl.handle.net/1887/20942 holds various files of this Leiden University dissertation. Author: Wammes, Linda Judith Title: Immune regulation during parasitic infections : from
Wolbachia-Induced Neutrophil Activation in a Mouse Model of Ocular Onchocerciasis (River Blindness)
") INFECTION AND IMMUNITY, Oct. 2004, p. 5687 5692 Vol. 72, No. 10 0019-9567/04/$08.00 0 DOI: 10.1128/IAI.72.10.5687 5692.2004 Copyright 2004, American Society for Microbiology. All Rights Reserved. Wolbachia-Induced
INFECTION AND IMMUNITY, Oct. 2004, p. 5687 5692 Vol. 72, No. 10 0019-9567/04/$08.00 0 DOI: 10.1128/IAI.72.10.5687 5692.2004 Copyright 2004, American Society for Microbiology. All Rights Reserved. Wolbachia-Induced
An Operational Research on Annual Mass Drug Administration (MDA) For Elimination of Lymphatic Filariasis in Medak District, Telangana
 For Elimination of Lymphatic Filariasis in Medak District, Telangana") Original Research Article An Operational Research on Annual Mass Drug Administration (MDA) For Elimination of Lymphatic Filariasis in Medak District, Telangana P. Samuel Rajkumar 1*, Tukaram Kishanrao
Original Research Article An Operational Research on Annual Mass Drug Administration (MDA) For Elimination of Lymphatic Filariasis in Medak District, Telangana P. Samuel Rajkumar 1*, Tukaram Kishanrao
Introduction Parasitology. Parasitology Department Medical Faculty of Universitas Sumatera Utara
 Introduction Parasitology Parasitology Department Medical Faculty of Universitas Sumatera Utara Parasite Definition: Dependence of one living organism on another 2 CLASSIFICATION (based on habitat) t)
Introduction Parasitology Parasitology Department Medical Faculty of Universitas Sumatera Utara Parasite Definition: Dependence of one living organism on another 2 CLASSIFICATION (based on habitat) t)
MASS TREATMENT FOR LYMPHATIC FILARIASIS MASS TREATMENT TO ELIMINATE FILARIASIS IN PAPUA NEW GUINEA
 MASS TREATMENT FOR LYMPHATIC FILARIASIS MASS TREATMENT TO ELIMINATE FILARIASIS IN PAPUA NEW GUINEA MOSES J. BOCKARIE, PH.D., DANIEL J. TISCH, M.P.H., WILL KASTENS, B.SC., NEAL D.E. ALEXANDER, PH.D., ZACHARY
MASS TREATMENT FOR LYMPHATIC FILARIASIS MASS TREATMENT TO ELIMINATE FILARIASIS IN PAPUA NEW GUINEA MOSES J. BOCKARIE, PH.D., DANIEL J. TISCH, M.P.H., WILL KASTENS, B.SC., NEAL D.E. ALEXANDER, PH.D., ZACHARY
Guillermo H. Giambartolomei a,1, Barbara L. Lasater a, François Villinger b, Vida A. Dennis a, * Short communication
 Acta Tropica 78 (2001) 67 71 www.parasitology-online.com Short communication Diminished expression of the costimulatory ligand CD80 (B7.1) gene correlates with antigen-specific cellular unresponsiveness
Acta Tropica 78 (2001) 67 71 www.parasitology-online.com Short communication Diminished expression of the costimulatory ligand CD80 (B7.1) gene correlates with antigen-specific cellular unresponsiveness
Adaptive immune responses: T cell-mediated immunity
 MICR2209 Adaptive immune responses: T cell-mediated immunity Dr Allison Imrie allison.imrie@uwa.edu.au 1 Synopsis: In this lecture we will discuss the T-cell mediated immune response, how it is activated,
MICR2209 Adaptive immune responses: T cell-mediated immunity Dr Allison Imrie allison.imrie@uwa.edu.au 1 Synopsis: In this lecture we will discuss the T-cell mediated immune response, how it is activated,
محاضرة مناعت مدرس المادة :ا.م. هدى عبدالهادي علي النصراوي Immunity to Infectious Diseases
 محاضرة مناعت مدرس المادة :ا.م. هدى عبدالهادي علي النصراوي Immunity to Infectious Diseases Immunity to infection depends on a combination of innate mechanisms (phagocytosis, complement, etc.) and antigen
محاضرة مناعت مدرس المادة :ا.م. هدى عبدالهادي علي النصراوي Immunity to Infectious Diseases Immunity to infection depends on a combination of innate mechanisms (phagocytosis, complement, etc.) and antigen
Filarial susceptibility and effects of Wolbachia in Aedes pseudoscutellaris mosquitoes
 Medical and Veterinary Entomology (2005) 19, 60 65 Filarial susceptibility and effects of Wolbachia in Aedes pseudoscutellaris mosquitoes T. J. DUTTON and S. P. SINKINS Vector Research Group, Liverpool
Medical and Veterinary Entomology (2005) 19, 60 65 Filarial susceptibility and effects of Wolbachia in Aedes pseudoscutellaris mosquitoes T. J. DUTTON and S. P. SINKINS Vector Research Group, Liverpool
Chapter 22: The Lymphatic System and Immunity
 Bio40C schedule Lecture Immune system Lab Quiz 2 this week; bring a scantron! Study guide on my website (see lab assignments) Extra credit Critical thinking questions at end of chapters 5 pts/chapter Due
Bio40C schedule Lecture Immune system Lab Quiz 2 this week; bring a scantron! Study guide on my website (see lab assignments) Extra credit Critical thinking questions at end of chapters 5 pts/chapter Due
Evaluation of the ICT whole blood antigen card test to detect infection due to nocturnally periodic Wuchereria bancrofti in South India
 TMIH559 Tropical Medicine and International Health volume 5 no 5 pp 359 363 may 2000 Evaluation of the ICT whole blood antigen card test to detect infection due to nocturnally periodic Wuchereria bancrofti
TMIH559 Tropical Medicine and International Health volume 5 no 5 pp 359 363 may 2000 Evaluation of the ICT whole blood antigen card test to detect infection due to nocturnally periodic Wuchereria bancrofti
Lymphoid System: cells of the immune system. Answer Sheet
 Lymphoid System: cells of the immune system Answer Sheet Q1 Which areas of the lymph node have most CD3 staining? A1 Most CD3 staining is present in the paracortex (T cell areas). This is towards the outside
Lymphoid System: cells of the immune system Answer Sheet Q1 Which areas of the lymph node have most CD3 staining? A1 Most CD3 staining is present in the paracortex (T cell areas). This is towards the outside
Disease-causing organisms
 1 of 41 2 of 41 Disease-causing organisms Organisms that cause disease are called pathogens. What are the four major types of pathogen? bacteria fungi protozoa virus 3 of 41 How do pathogens cause illness?
1 of 41 2 of 41 Disease-causing organisms Organisms that cause disease are called pathogens. What are the four major types of pathogen? bacteria fungi protozoa virus 3 of 41 How do pathogens cause illness?
Malaria. Population at Risk. Infectious Disease epidemiology BMTRY 713 (Lecture 23) Epidemiology of Malaria. April 6, Selassie AW (DPHS) 1
 Epidemiology of Malaria. April 6, Selassie AW (DPHS) 1") Infectious Disease Epidemiology BMTRY 713 (A. Selassie, DrPH) Lecture 23 Vector-Borne Disease (Part II) Epidemiology of Malaria Learning Objectives 1. Overview of malaria Global perspectives 2. Identify
Infectious Disease Epidemiology BMTRY 713 (A. Selassie, DrPH) Lecture 23 Vector-Borne Disease (Part II) Epidemiology of Malaria Learning Objectives 1. Overview of malaria Global perspectives 2. Identify
Blood Smears Only 07 February Sample Preparation and Quality Control 12B A
 NEW YORK STATE Parasitology Proficiency Testing Program Blood Smears Only 07 February 2012 The purpose of the New York State Proficiency Testing Program in the category of Parasitology Blood Smears Only
NEW YORK STATE Parasitology Proficiency Testing Program Blood Smears Only 07 February 2012 The purpose of the New York State Proficiency Testing Program in the category of Parasitology Blood Smears Only
Dengue Virus-Danger from Deadly Little Dragon
 Molecular Medicine Dengue Virus-Danger from Deadly Little Dragon Dr.G.MATHAN Assistant Professor Department of Biomedical Science Bharathidasan University Tiruchirappalli, Tamil Nadu Vector (A carrier)
Molecular Medicine Dengue Virus-Danger from Deadly Little Dragon Dr.G.MATHAN Assistant Professor Department of Biomedical Science Bharathidasan University Tiruchirappalli, Tamil Nadu Vector (A carrier)
Killing filarial nematode parasites: role of treatment options and host immune response
 Kwarteng et al. Infectious Diseases of Poverty (2016) 5:86 DOI 10.1186/s40249-016-0183-0 SCOPING REVIEW Killing filarial nematode parasites: role of treatment options and host immune response Alexander
Kwarteng et al. Infectious Diseases of Poverty (2016) 5:86 DOI 10.1186/s40249-016-0183-0 SCOPING REVIEW Killing filarial nematode parasites: role of treatment options and host immune response Alexander
PARASITOLOGY INTRODUCTION
 PARASITOLOGY INTRODUCTION DEFINITION Parasite means : one who eats at the table of another. Differs from bacteria and viruses in their complex life cycles, intermediate hosts and chronicity. Evolved to
PARASITOLOGY INTRODUCTION DEFINITION Parasite means : one who eats at the table of another. Differs from bacteria and viruses in their complex life cycles, intermediate hosts and chronicity. Evolved to
The Adaptive Immune Response. B-cells
 The Adaptive Immune Response B-cells The innate immune system provides immediate protection. The adaptive response takes time to develop and is antigen specific. Activation of B and T lymphocytes Naive
The Adaptive Immune Response B-cells The innate immune system provides immediate protection. The adaptive response takes time to develop and is antigen specific. Activation of B and T lymphocytes Naive
I. Critical Vocabulary
 I. Critical Vocabulary A. Immune System: a set of glands, tissues, cells, and dissolved proteins that combine to defend against non-self entities B. Antigen: any non-self chemical that triggers an immune
I. Critical Vocabulary A. Immune System: a set of glands, tissues, cells, and dissolved proteins that combine to defend against non-self entities B. Antigen: any non-self chemical that triggers an immune
Downloaded from
 Class IX: Biology Chapter: Why do we fall ill Chapter Notes Key learnings: 1) Our body s well-being is dependent on the proper functioning of its cells and tissues. 2) All our body parts and activities
Class IX: Biology Chapter: Why do we fall ill Chapter Notes Key learnings: 1) Our body s well-being is dependent on the proper functioning of its cells and tissues. 2) All our body parts and activities
IN THIS ISSUE: LYMPHATIC FILARIASIS: ELIMINATING ONE OF HUMANITY S MOST DEVASTATING DISEASES
 ACTION AGAINST WORMS JULY 2010 ISSUE 14 Lymphatic filariasis patient, Debaraj Behera, in Dhalapathar, Orissa, India, 2009. IN THIS ISSUE: Lymphatic filariasis: eliminating one of humanity s most devastating
ACTION AGAINST WORMS JULY 2010 ISSUE 14 Lymphatic filariasis patient, Debaraj Behera, in Dhalapathar, Orissa, India, 2009. IN THIS ISSUE: Lymphatic filariasis: eliminating one of humanity s most devastating
Microfilaria persistent foci during post MDA and the risk assessment of resurgence in India
 Mehta et al. Tropical Medicine and Health (2018) 46:25 https://doi.org/10.1186/s41182-018-0107-8 Tropical Medicine and Health RESEARCH Microfilaria persistent foci during post MDA and the risk assessment
Mehta et al. Tropical Medicine and Health (2018) 46:25 https://doi.org/10.1186/s41182-018-0107-8 Tropical Medicine and Health RESEARCH Microfilaria persistent foci during post MDA and the risk assessment
1. Overview of Adaptive Immunity
 Chapter 17A: Adaptive Immunity Part I 1. Overview of Adaptive Immunity 2. T and B Cell Production 3. Antigens & Antigen Presentation 4. Helper T cells 1. Overview of Adaptive Immunity The Nature of Adaptive
Chapter 17A: Adaptive Immunity Part I 1. Overview of Adaptive Immunity 2. T and B Cell Production 3. Antigens & Antigen Presentation 4. Helper T cells 1. Overview of Adaptive Immunity The Nature of Adaptive
Blood Smears Only 19 May Sample Preparation and Quality Control
 NEW YORK STATE Parasitology Proficiency Testing Program Blood Smears Only 9 May 205 The purpose of the New York State Proficiency Testing Program in the category of Parasitology - Blood Smears Only is
NEW YORK STATE Parasitology Proficiency Testing Program Blood Smears Only 9 May 205 The purpose of the New York State Proficiency Testing Program in the category of Parasitology - Blood Smears Only is
INTRODUCTION. Lymphatic filariasis (LF), a deforming and debilitating disease transmitted by
, a deforming and debilitating disease transmitted by") INTRODUCTION Lymphatic filariasis (LF), a deforming and debilitating disease transmitted by '. mosquitoes, causes elephantiasis and male genital damage and is a major social and economic scourge in the
INTRODUCTION Lymphatic filariasis (LF), a deforming and debilitating disease transmitted by '. mosquitoes, causes elephantiasis and male genital damage and is a major social and economic scourge in the
Multicentre laboratory evaluation of Brugia Rapid dipstick test for detection of brugian filariasis
 Tropical Medicine and International Health volume 8 no 10 pp 895 900 october 2003 Multicentre laboratory evaluation of Brugia Rapid dipstick test for detection of brugian filariasis N. Rahmah 1, R. K.
Tropical Medicine and International Health volume 8 no 10 pp 895 900 october 2003 Multicentre laboratory evaluation of Brugia Rapid dipstick test for detection of brugian filariasis N. Rahmah 1, R. K.
Yoshinori Mitsui 1* and Mitsumasa Miura 1. Received 4 June, 2010 Accepted 4 August, 2010 Published online 28 August, 2010
 Tropical Medicine and Health Vol. 38 No. 3, 2010, pp. 97-101 Copyright 2010 by The Japanese Society of Tropical Medicine Original article Development of a simple and convenient feeding device to infect
Tropical Medicine and Health Vol. 38 No. 3, 2010, pp. 97-101 Copyright 2010 by The Japanese Society of Tropical Medicine Original article Development of a simple and convenient feeding device to infect
INTRODUCTION. 1 Onchocerciasis
 TABLE OF CONTENTS TABLE OF CONTENTS...0 INTRODUCTION...1 1 Onchocerciasis...1 2 Wolbachia endosymbiotic bacteria...4 3 Pathogenesis and host immune responses in onchocerca keratitis...5 4 Aim of this Study...9
TABLE OF CONTENTS TABLE OF CONTENTS...0 INTRODUCTION...1 1 Onchocerciasis...1 2 Wolbachia endosymbiotic bacteria...4 3 Pathogenesis and host immune responses in onchocerca keratitis...5 4 Aim of this Study...9
The Adaptive Immune Responses
 The Adaptive Immune Responses The two arms of the immune responses are; 1) the cell mediated, and 2) the humoral responses. In this chapter we will discuss the two responses in detail and we will start
The Adaptive Immune Responses The two arms of the immune responses are; 1) the cell mediated, and 2) the humoral responses. In this chapter we will discuss the two responses in detail and we will start
 Malaria parasites Malaria parasites are micro-organisms that belong to the genus Plasmodium. There are more than 100 species of Plasmodium, which can infect many animal species such as reptiles, birds,
Malaria parasites Malaria parasites are micro-organisms that belong to the genus Plasmodium. There are more than 100 species of Plasmodium, which can infect many animal species such as reptiles, birds,
I. Lines of Defense Pathogen: Table 1: Types of Immune Mechanisms. Table 2: Innate Immunity: First Lines of Defense
 I. Lines of Defense Pathogen: Table 1: Types of Immune Mechanisms Table 2: Innate Immunity: First Lines of Defense Innate Immunity involves nonspecific physical & chemical barriers that are adapted for
I. Lines of Defense Pathogen: Table 1: Types of Immune Mechanisms Table 2: Innate Immunity: First Lines of Defense Innate Immunity involves nonspecific physical & chemical barriers that are adapted for
Immunodeficiency. (2 of 2)
") Immunodeficiency (2 of 2) Acquired (secondary) immunodeficiencies More common Many causes such as therapy, cancer, sarcoidosis, malnutrition, infection & renal disease The most common of which is therapy-related
Immunodeficiency (2 of 2) Acquired (secondary) immunodeficiencies More common Many causes such as therapy, cancer, sarcoidosis, malnutrition, infection & renal disease The most common of which is therapy-related
Subramanian Swaminathan
 Modelling Lymphatic Filariasis Transmission and Control Subramanian Swaminathan This thesis was realized with support from a WHO/TDR Research training grant, and with financial support from the Stichting
Modelling Lymphatic Filariasis Transmission and Control Subramanian Swaminathan This thesis was realized with support from a WHO/TDR Research training grant, and with financial support from the Stichting
Unusual presentation of Filariasis in a tertiary care hospital in Western Rajasthan: A case report
 ISSN: 2319-7706 Volume 4 Number 1 (2015) pp. 685-689 http://www.ijcmas.com Original Research Article Unusual presentation of Filariasis in a tertiary care hospital in Western Rajasthan: A case report Swati
ISSN: 2319-7706 Volume 4 Number 1 (2015) pp. 685-689 http://www.ijcmas.com Original Research Article Unusual presentation of Filariasis in a tertiary care hospital in Western Rajasthan: A case report Swati
GUIDELINES FOR CLINICAL MANAGEMENT OF HUMAN DIROFILARIA INFECTIONS
 GUIDELINES FOR CLINICAL MANAGEMENT OF HUMAN DIROFILARIA INFECTIONS Prepared for and approved by the Executive Board of ESDA Background Zoonoses are infectious diseases of animals that can be transmitted
GUIDELINES FOR CLINICAL MANAGEMENT OF HUMAN DIROFILARIA INFECTIONS Prepared for and approved by the Executive Board of ESDA Background Zoonoses are infectious diseases of animals that can be transmitted
Question 1. Kupffer cells, microglial cells and osteoclasts are all examples of what type of immune system cell?
 Abbas Chapter 2: Sarah Spriet February 8, 2015 Question 1. Kupffer cells, microglial cells and osteoclasts are all examples of what type of immune system cell? a. Dendritic cells b. Macrophages c. Monocytes
Abbas Chapter 2: Sarah Spriet February 8, 2015 Question 1. Kupffer cells, microglial cells and osteoclasts are all examples of what type of immune system cell? a. Dendritic cells b. Macrophages c. Monocytes
Lymphatic filariasis: new insights and prospects for control Thomas B. Nutman
 Lymphatic filariasis: new insights and prospects for control Thomas B Nutman Although lymphatic filariasis remains among the major causes of disability among the tropical infectious diseases, dramatic
Lymphatic filariasis: new insights and prospects for control Thomas B Nutman Although lymphatic filariasis remains among the major causes of disability among the tropical infectious diseases, dramatic
Parasitology Questions. Choose the best correct answer in the following statements
 Parasitology Questions Choose the best correct answer in the following statements ١. A.. is the larval stage of a fluke that is covered with cilia and swims about seeking out a snail to serve as an intermediate
Parasitology Questions Choose the best correct answer in the following statements ١. A.. is the larval stage of a fluke that is covered with cilia and swims about seeking out a snail to serve as an intermediate
Influence of Maternal Filariasis on Childhood Infection and Immunity to Wuchereria bancrofti in Kenya
 INFECTION AND IMMUNITY, Sept. 2003, p. 5231 5237 Vol. 71, No. 9 0019-9567/03/$08.00 0 DOI: 10.1128/IAI.71.9.5231 5237.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved. Influence
INFECTION AND IMMUNITY, Sept. 2003, p. 5231 5237 Vol. 71, No. 9 0019-9567/03/$08.00 0 DOI: 10.1128/IAI.71.9.5231 5237.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved. Influence
There are 2 major lines of defense: Non-specific (Innate Immunity) and. Specific. (Adaptive Immunity) Photo of macrophage cell
 and. Specific. (Adaptive Immunity) Photo of macrophage cell") There are 2 major lines of defense: Non-specific (Innate Immunity) and Specific (Adaptive Immunity) Photo of macrophage cell Development of the Immune System ery pl neu mφ nk CD8 + CTL CD4 + thy TH1 mye
There are 2 major lines of defense: Non-specific (Innate Immunity) and Specific (Adaptive Immunity) Photo of macrophage cell Development of the Immune System ery pl neu mφ nk CD8 + CTL CD4 + thy TH1 mye
Sensitivity and Specificity of ELISA in Detection of Microfilariae
 ORIGINAL ARTICLE Sensitivity and Specificity of in Detection of Microfilariae Lakshmi Jyothi 1, MVR Reddy 2 1. Associate Professor of Microbiology, Medicity Institute of Medical Sciences, Hyderabad. 2.
ORIGINAL ARTICLE Sensitivity and Specificity of in Detection of Microfilariae Lakshmi Jyothi 1, MVR Reddy 2 1. Associate Professor of Microbiology, Medicity Institute of Medical Sciences, Hyderabad. 2.
Immune System AP SBI4UP
 Immune System AP SBI4UP TYPES OF IMMUNITY INNATE IMMUNITY ACQUIRED IMMUNITY EXTERNAL DEFENCES INTERNAL DEFENCES HUMORAL RESPONSE Skin Phagocytic Cells CELL- MEDIATED RESPONSE Mucus layer Antimicrobial
Immune System AP SBI4UP TYPES OF IMMUNITY INNATE IMMUNITY ACQUIRED IMMUNITY EXTERNAL DEFENCES INTERNAL DEFENCES HUMORAL RESPONSE Skin Phagocytic Cells CELL- MEDIATED RESPONSE Mucus layer Antimicrobial
Cytokines modulate the functional activities of individual cells and tissues both under normal and pathologic conditions Interleukins,
 Cytokines http://highered.mcgraw-hill.com/sites/0072507470/student_view0/chapter22/animation the_immune_response.html Cytokines modulate the functional activities of individual cells and tissues both under
Cytokines http://highered.mcgraw-hill.com/sites/0072507470/student_view0/chapter22/animation the_immune_response.html Cytokines modulate the functional activities of individual cells and tissues both under
Class 9 th Why do we fall ill?
 Class 9 th Why do we fall ill? Health: health is a state of physical, mental and social well being. The health of all individuals is dependent on their physical environment, social environment, and their
Class 9 th Why do we fall ill? Health: health is a state of physical, mental and social well being. The health of all individuals is dependent on their physical environment, social environment, and their
Q Fever What men and women on the land need to know
 Q Fever What men and women on the land need to know Dr. Stephen Graves Director, Australian Rickettsial Reference Laboratory Director, Division of Microbiology, Pathology North (Hunter) NSW Health Pathology,
Q Fever What men and women on the land need to know Dr. Stephen Graves Director, Australian Rickettsial Reference Laboratory Director, Division of Microbiology, Pathology North (Hunter) NSW Health Pathology,
Defense mechanism against pathogens
 Defense mechanism against pathogens Immune System What is immune system? Cells and organs within an animal s body that contribute to immune defenses against pathogens ( ) Bacteria -Major entry points ;open
Defense mechanism against pathogens Immune System What is immune system? Cells and organs within an animal s body that contribute to immune defenses against pathogens ( ) Bacteria -Major entry points ;open
Immunology Lecture 4. Clinical Relevance of the Immune System
 Immunology Lecture 4 The Well Patient: How innate and adaptive immune responses maintain health - 13, pg 169-181, 191-195. Immune Deficiency - 15 Autoimmunity - 16 Transplantation - 17, pg 260-270 Tumor
Immunology Lecture 4 The Well Patient: How innate and adaptive immune responses maintain health - 13, pg 169-181, 191-195. Immune Deficiency - 15 Autoimmunity - 16 Transplantation - 17, pg 260-270 Tumor
11/25/2017. THE IMMUNE SYSTEM Chapter 43 IMMUNITY INNATE IMMUNITY EXAMPLE IN INSECTS BARRIER DEFENSES INNATE IMMUNITY OF VERTEBRATES
 THE IMMUNE SYSTEM Chapter 43 IMMUNITY INNATE IMMUNITY EXAMPLE IN INSECTS Exoskeleton made of chitin forms the first barrier to pathogens Digestive system is protected by a chitin-based barrier and lysozyme,
THE IMMUNE SYSTEM Chapter 43 IMMUNITY INNATE IMMUNITY EXAMPLE IN INSECTS Exoskeleton made of chitin forms the first barrier to pathogens Digestive system is protected by a chitin-based barrier and lysozyme,
Immunological Aspects of Parasitic Diseases in Immunocompromised Individuals. Taniawati Supali. Department of Parasitology
 Immunological Aspects of Parasitic Diseases in Immunocompromised Individuals Taniawati Supali Department of Parasitology 1 Defense mechanism in human Th17 (? ) Acute Chronic Th1 Th 2 Intracellular Treg
Immunological Aspects of Parasitic Diseases in Immunocompromised Individuals Taniawati Supali Department of Parasitology 1 Defense mechanism in human Th17 (? ) Acute Chronic Th1 Th 2 Intracellular Treg
Fluid movement in capillaries. Not all fluid is reclaimed at the venous end of the capillaries; that is the job of the lymphatic system
 Capillary exchange Fluid movement in capillaries Not all fluid is reclaimed at the venous end of the capillaries; that is the job of the lymphatic system Lymphatic vessels Lymphatic capillaries permeate
Capillary exchange Fluid movement in capillaries Not all fluid is reclaimed at the venous end of the capillaries; that is the job of the lymphatic system Lymphatic vessels Lymphatic capillaries permeate
HIGH PREVALENCE OF BRUGIA TIMORI INFECTION IN THE HIGHLAND OF ALOR ISLAND, INDONESIA
 Am. J. Trop. Med. Hyg., 66(5), 2002, pp. 560 565 Copyright 2002 by The American Society of Tropical Medicine and Hygiene HIGH PREVALENCE OF BRUGIA TIMORI INFECTION IN THE HIGHLAND OF ALOR ISLAND, INDONESIA
Am. J. Trop. Med. Hyg., 66(5), 2002, pp. 560 565 Copyright 2002 by The American Society of Tropical Medicine and Hygiene HIGH PREVALENCE OF BRUGIA TIMORI INFECTION IN THE HIGHLAND OF ALOR ISLAND, INDONESIA
Transmission Intensity Affects Both Antigen-Specific and Nonspecific T-Cell Proliferative Responses in Loa loa Infection
 INFECTION AND IMMUNITY, Mar. 2002, p. 1475 1480 Vol. 70, No. 3 0019-9567/02/$04.00 0 DOI: 10.1128/IAI.70.3.1475 1480.2002 Copyright 2002, American Society for Microbiology. All Rights Reserved. Transmission
INFECTION AND IMMUNITY, Mar. 2002, p. 1475 1480 Vol. 70, No. 3 0019-9567/02/$04.00 0 DOI: 10.1128/IAI.70.3.1475 1480.2002 Copyright 2002, American Society for Microbiology. All Rights Reserved. Transmission
Impact of a Community-Based Lymphedema Management Program on Episodes of Adenolymphangitis (ADLA) and Lymphedema Progression - Odisha State, India
 and Lymphedema Progression - Odisha State, India") Impact of a Community-Based Lymphedema Management Program on Episodes of Adenolymphangitis (ADLA) and Lymphedema Progression - Odisha State, India Katherine E. Mues, Emory University Michael Deming, Emory
Impact of a Community-Based Lymphedema Management Program on Episodes of Adenolymphangitis (ADLA) and Lymphedema Progression - Odisha State, India Katherine E. Mues, Emory University Michael Deming, Emory
Disease Transmission Methods
 Disease Transmission Methods In epidemiology, transmission simply means any method by which an infectious agent is spread from one host to another. Knowing the type of pathogen often, but not always, identifies
Disease Transmission Methods In epidemiology, transmission simply means any method by which an infectious agent is spread from one host to another. Knowing the type of pathogen often, but not always, identifies
Immunity. Acquired immunity differs from innate immunity in specificity & memory from 1 st exposure
 Immunity (1) Non specific (innate) immunity (2) Specific (acquired) immunity Characters: (1) Non specific: does not need special recognition of the foreign cell. (2) Innate: does not need previous exposure.
Immunity (1) Non specific (innate) immunity (2) Specific (acquired) immunity Characters: (1) Non specific: does not need special recognition of the foreign cell. (2) Innate: does not need previous exposure.
Alberta Health Public Health Notifiable Disease Management Guidelines July 2012
 July 2012 Typhus - Louseborne Revision Dates Case Definition Reporting Requirements Remainder of the Guideline (i.e., Etiology to References sections inclusive) July 2012 July 2012 October 2005 Case Definition
July 2012 Typhus - Louseborne Revision Dates Case Definition Reporting Requirements Remainder of the Guideline (i.e., Etiology to References sections inclusive) July 2012 July 2012 October 2005 Case Definition
2013 Disease Detectives
 2013 Disease Detectives Since the catastrophic earthquake that hit Haiti in January of 2010, there have been an alarming number of cases of Cholera, spread by the Vibrio cholera bacterium, reported within
2013 Disease Detectives Since the catastrophic earthquake that hit Haiti in January of 2010, there have been an alarming number of cases of Cholera, spread by the Vibrio cholera bacterium, reported within
1.Nematodes. Parasitology/Helminths
 1.Nematodes Parasitology/Helminths Helminths all helminths are relatively large (> 1 mm long); some are very large (> 1 m long). all have well-developed organ systems and most are active feeders. the body
1.Nematodes Parasitology/Helminths Helminths all helminths are relatively large (> 1 mm long); some are very large (> 1 m long). all have well-developed organ systems and most are active feeders. the body
Where are we heading?
 Unit 4: Where are we heading? Unit 4: Introduction Unit 1: Why should we care about infectious diseases? Unit 2: What does it mean to have an infectious disease? Unit 3: When does a microbe become a pathogen?
Unit 4: Where are we heading? Unit 4: Introduction Unit 1: Why should we care about infectious diseases? Unit 2: What does it mean to have an infectious disease? Unit 3: When does a microbe become a pathogen?
Infection : a disease or condition caused by a microorganism Microorganisms are the tiniest living organisms on earth that
 Chapter 13: Infectious Diseases The Process of Infection Infection : a disease or condition caused by a microorganism Microorganisms are the tiniest living organisms on earth that eat, reproduce, and die
Chapter 13: Infectious Diseases The Process of Infection Infection : a disease or condition caused by a microorganism Microorganisms are the tiniest living organisms on earth that eat, reproduce, and die
M. Haarbrink, 1 G. K. Abadi, 4 W. A. Buurman, 2 M. A. Dentener, 3 A. J. Terhell, 1 and M. Yazdanbakhsh 1
 564 Strong Association of Interleukin-6 and Lipopolysaccharide-Binding Protein with Severity of Adverse Reactions after Diethylcarbamazine Treatment of Microfilaremic Patients M. Haarbrink, 1 G. K. Abadi,
564 Strong Association of Interleukin-6 and Lipopolysaccharide-Binding Protein with Severity of Adverse Reactions after Diethylcarbamazine Treatment of Microfilaremic Patients M. Haarbrink, 1 G. K. Abadi,
Repellent Soap. The Jojoo Mosquito. Africa s innovative solution to Malaria prevention. Sapphire Trading Company Ltd
 The Jojoo Mosquito Repellent Soap Africa s innovative solution to Malaria prevention Sapphire Trading Company Ltd P.O.Box: 45938-00100 Nairobi, Kenya. Tel: +254 735 397 267 +254 733 540 868 +254 700 550
The Jojoo Mosquito Repellent Soap Africa s innovative solution to Malaria prevention Sapphire Trading Company Ltd P.O.Box: 45938-00100 Nairobi, Kenya. Tel: +254 735 397 267 +254 733 540 868 +254 700 550
Topics in Parasitology BLY Vertebrate Immune System
 Topics in Parasitology BLY 533-2008 Vertebrate Immune System V. Vertebrate Immune System A. Non-specific defenses against pathogens 1. Skin - physical barrier a. Tough armor protein KERATIN b. Surface
Topics in Parasitology BLY 533-2008 Vertebrate Immune System V. Vertebrate Immune System A. Non-specific defenses against pathogens 1. Skin - physical barrier a. Tough armor protein KERATIN b. Surface
The Struggle with Infectious Disease. Lecture 5
 The Struggle with Infectious Disease Lecture 5 Tropical Diseases Life and infectious disease began in the rift valley/rain forest of Eastern Africa Peoples migrated North to temperate regions Tropical
The Struggle with Infectious Disease Lecture 5 Tropical Diseases Life and infectious disease began in the rift valley/rain forest of Eastern Africa Peoples migrated North to temperate regions Tropical
Pathogenicity of Infectious Diseases
 Pathogenicity of Infectious Diseases Pathogenicity of Infectious Diseases HOST DISEASE TRIAD PATHOGEN ENVIRONMENT OTHER MICROBES Microbial Interactions KOCH'S POSTULATES Four criteria that were established
Pathogenicity of Infectious Diseases Pathogenicity of Infectious Diseases HOST DISEASE TRIAD PATHOGEN ENVIRONMENT OTHER MICROBES Microbial Interactions KOCH'S POSTULATES Four criteria that were established
Epidemiology and entomology of the Zika virus outbreak
 Epidemiology and entomology of the Zika virus outbreak M A T T H E W B A Y L I S I N S T I T U T E O F I N F E C T I O N A N D G L O B A L H E A L T H U N I V E R S I T Y O F L I V E R P O O L Zika in
Epidemiology and entomology of the Zika virus outbreak M A T T H E W B A Y L I S I N S T I T U T E O F I N F E C T I O N A N D G L O B A L H E A L T H U N I V E R S I T Y O F L I V E R P O O L Zika in
Chapter 11 Risk, Toxicology, and Human Health
 Chapter 11 Risk, Toxicology, and Human Health Risk Expressed in terms of probability: how likely it is that some event will occur. Risk = (Exposure)(harm) Risk assessment (identifying, occurrence, assessing)
Chapter 11 Risk, Toxicology, and Human Health Risk Expressed in terms of probability: how likely it is that some event will occur. Risk = (Exposure)(harm) Risk assessment (identifying, occurrence, assessing)
Comparative Studies on the Biology and Filarial Susceptibility of Selected Blood-feeding and Autogenous Aedes togoi Sub-colonies
 Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 98(4): 481-485, June 2003 481 Comparative Studies on the Biology and Filarial Susceptibility of Selected Blood-feeding and Autogenous Aedes togoi Sub-colonies
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 98(4): 481-485, June 2003 481 Comparative Studies on the Biology and Filarial Susceptibility of Selected Blood-feeding and Autogenous Aedes togoi Sub-colonies
PDF EYE WORM DISEASE OF AFRICA
 15 January, 2018 PDF EYE WORM DISEASE OF AFRICA Document Filetype: PDF 470.56 KB 0 PDF EYE WORM DISEASE OF AFRICA Lymphatic filariasis affects over 120 million people in the tropics and sub-tropics of
15 January, 2018 PDF EYE WORM DISEASE OF AFRICA Document Filetype: PDF 470.56 KB 0 PDF EYE WORM DISEASE OF AFRICA Lymphatic filariasis affects over 120 million people in the tropics and sub-tropics of
Chapter 2 Maternal Helminth Infections
 Chapter 2 Maternal Helminth Infections Kathrin Straubinger and Clarissa Prazeres da Costa In this chapter we will discuss the impact of the most prevalent human helminth diseases on pregnancy. In the first
Chapter 2 Maternal Helminth Infections Kathrin Straubinger and Clarissa Prazeres da Costa In this chapter we will discuss the impact of the most prevalent human helminth diseases on pregnancy. In the first
Histopathologic Improvement with Lymphedema Management, Léogâne, Haiti
 Histopathologic Improvement with Lymphedema Management, Léogâne, Haiti Susan F. Wilson,* Jeannette Guarner,* Alix L. Valme, Jacky Louis-Charles, Tara L. Jones,* and David G. Addiss* In countries where
Histopathologic Improvement with Lymphedema Management, Léogâne, Haiti Susan F. Wilson,* Jeannette Guarner,* Alix L. Valme, Jacky Louis-Charles, Tara L. Jones,* and David G. Addiss* In countries where
Study Guide 23, 24 & 47
 Study Guide 23, 24 & 47 STUDY GUIDE SECTION 23-3 Bacteria and Humans Name Period Date 1. One bacterial disease that is transmitted by contaminated drinking water is a. Lyme disease b. gonorrhea c. tuberculosis
Study Guide 23, 24 & 47 STUDY GUIDE SECTION 23-3 Bacteria and Humans Name Period Date 1. One bacterial disease that is transmitted by contaminated drinking water is a. Lyme disease b. gonorrhea c. tuberculosis