Role of Ubiquitin in PIV5 Particle Formation
|
|
|
- Judith Patterson
- 6 years ago
- Views:
Transcription
1 JVI Accepts, published online ahead of print on 18 January 2012 J. Virol. doi: /jvi Copyright 2012, American Society for Microbiology. All Rights Reserved Role of Ubiquitin in PIV5 Particle Formation Megan S. Harrison 1#, Phuong Tieu Schmitt 1, Zifei Pei 1, and Anthony P. Schmitt 1,2* Department of Veterinary and Biomedical Sciences 1, and Center for Molecular Immunology and Infectious Disease 2, The Pennsylvania State University, University Park, PA Running Title: PIV5 M protein ubiquitination *Corresponding Author: Department of Veterinary and Biomedical Sciences The Pennsylvania State University 115 Henning Building University Park, PA Tel: Fax: aps13@psu.edu # Present address: Department of Cell Biology Yale University School of Medicine New Haven, CT 06510
2 ABSTRACT Ubiquitin is important for the budding of many retroviruses and other enveloped viruses, but the precise role for ubiquitin in virus budding remains unclear. Here, we characterized ubiquitination of the matrix (M) protein of a paramyxovirus, parainfluenza virus 5 (PIV5). PIV5 M protein (but not PIV5 nucleocapsid protein) was found to be targeted for monoubiquitination in transfected mammalian cells. Major sites of ubiquitin attachment identified by mass spectrometry analysis were lysine residues at amino acid positions 79/80, 130, and 247. Cumulative mutation of lysine residues 79, 80, and 130 to arginines led to an altered pattern of M protein ubiquitination and impaired VLP production function. However, cumulative mutation of lysine residues 79, 80, 130, and 247 to arginines restored M protein ubiquitination and VLP production, suggesting that ubiquitin is attached to alternative sites on M protein when the primary ones have been removed. Additional lysine residues were targeted for mutagenesis based on the UbiPred algorithm. M protein with seven lysine residues changed to arginines exhibited altered ubiquitination and poor VLP production function. Recombinant virus was generated encoding seven-lysine-mutant M protein, and this virus exhibited six-fold reduced maximum titer, with the defect mainly attributed to the budding of noninfectious particles. The recombinant virus was assembly-deficient, judged by redistribution of viral M and hemagglutinin-neuraminidase proteins in infected cells. Similar assembly defects were observed for wt virus after proteasome-inhibitor treatment. Collectively, these findings suggest that monoubiquitination of PIV5 M protein is important for proper virus assembly and for the budding of infectious particles. 2
3 INTRODUCTION Parainfluenza virus 5 (PIV5, formerly SV5) is a paramyxovirus belonging to the Rubulavirus genus, a genus which also includes mumps virus, Tioman virus, Menangle virus, and human parainfluenza virus types 2 and 4 (21). Paramyxoviruses carry their genetic information on nonsegmented, single-stranded, negative-sense RNA genomes, which are encapsidated by nucleocapsid (N or NP) proteins, forming helical ribonucleoproteins (RNPs). These RNPs act as the templates for viral transcription and genome replication mediated by viral RNA-dependent RNA polymerases, which are composed of large (L) and phosphoprotein (P) subunits. RNPs are ultimately packaged into virions, which are membrane-enveloped, roughly spherical, and variable in size with an average diameter of approximately 150 nm (21). The interiors of the virion envelopes are lined with matrix (M) proteins, which are the most abundant proteins in virions and play dynamic roles in coordinating virus assembly and budding (reviewed in reference 9). Transmembrane glycoproteins stud the virion envelopes and chiefly function to mediate viral attachment to target cells, as well as fusion of viral and target cell membranes to allow entry. The cytoplasmic tails of the PIV5 glycoproteins, hemagglutinin-neuraminidase (HN) and fusion (F), are also required for efficient virus assembly (44, 58). Additional PIV5 components include the SH and V proteins, which function within infected cells to block apoptosis and interferon signaling (12, 17). During enveloped virus particle formation, viral components must assemble at selected sites on cellular membranes, causing the membrane to become deformed outward. Ultimately, virions detach from the membranes after fission events, which in many cases are aided by cellular components. The viral envelopes consist of lipids derived from cellular membranes at the sites from which budding has occurred (reviewed in references 9, 45, 51). For some paramyxoviruses, expression of M protein alone is sufficient to induce the release of virus-like particles (VLPs) from transfected cells (reviewed in reference 9). However, PIV5- like particles are produced efficiently only when PIV5 M protein is expressed together with PIV5 NP protein and one of the PIV5 glycoproteins, either HN or F (47). 3
4 Ubiquitin has diverse roles in regulating eukaryotic cellular processes (8). In addition to the well-established function for polyubiquitination in targeting proteins for proteolysis by the 26S proteasome (15), monoubiquitination can influence a variety of cellular processes including endosomal sorting, endocytosis, and nuclear export (8, 16). Ubiquitin has been implicated in the process of enveloped virus budding, with the role of ubiquitin most thoroughly considered within the framework of retrovirus budding. Retrovirus particles contain high levels of free ubiquitin (27, 32, 41, 55), and retroviral Gag proteins are often modified through ubiquitin attachment (7, 32, 34, 50). Treatment of cells with proteasome inhibitors, which deplete free cellular pools of ubiquitin, can inhibit both Gag monoubiquitination and retrovirus budding (3, 36, 48, 50, 56). The precise role for ubiquitin in virus budding is incompletely understood, however. Gag protein ubiquitination has been postulated to facilitate recognition of Gag proteins by endosomal sorting complex required for transport (ESCRT) proteins, which are involved in the budding of several enveloped viruses (reviewed in references 27, 28). Evidence for the involvement of Gag ubiquitination in retrovirus budding comes from experiments using Gag proteins directly fused to ubiquitin, which in some cases allows budding to occur even in the presence of proteasome inhibitors (36) or in the absence of functional late domains (18, 62). In addition, removal of lysine residues from the human immunodeficiency virus-1 (HIV-1), Rous sarcoma virus (RSV), and human T-cell leukemia virus type 1 (HTLV-1) Gag proteins results in particle release defects (6, 14, 22, 49). Although these observations suggest that ubiquitin attachment to Gag proteins can facilitate particle release, they do not exclude the possibility that ubiquitin could also function for virus budding independent of its attachment to viral proteins. Indeed, mutation of the single lysine residue in an engineered prototypic foamy virus (PFV) Gag protein did not affect VLP release (63). It has recently been proposed that although the presence of ubiquitin at virus assembly sites is important for the recruitment of ubiquitinbinding ESCRT proteins, some flexibility exists in terms of whether that ubiquitin is conjugated to viral proteins, or alternatively is conjugated to host proteins which are also present at virus assembly sites (62). Additional evidence links ubiquitin to the budding of some negative-strand RNA viruses. Ubiquitination of the vesicular stomatitis virus (VSV) and Ebola virus matrix proteins have been demonstrated in vitro (10, 11). These matrix proteins harbor PPxY-type late domains 4
5 which direct binding to Nedd4-like ubiquitin ligases (10, 11, 52, 61). ISG15 expression inhibits the ubiquitination of Ebola virus matrix protein and impairs particle release (25, 30). Proteasome inhibitor treatments have been found to inhibit the budding of paramyxoviruses, including PIV5, Nipah virus, and Sendai virus (46, 57, 59), although for Sendai virus this inhibition was cell-type dependent (59). In the case of Nipah virus, proteasome inhibitor treatment caused nuclear retention of M protein, together with impairments in the release of Nipah virions and VLPs (57). A single conserved lysine residue within a bipartite nuclear localization signal was shown to be important for regulating Nipah virus M protein nuclear import, ubiquitination, and membrane association (57). Possible ubiquitination of measles virus M protein has been observed as well (40), but a functional role for measles virus M protein ubiquitination has not yet been reported. Here, we demonstrate that PIV5 M protein is a target for ubiquitin conjugation. Primary ubiquitin acceptor sites were identified by mass spectrometry (MS) analysis, and through use of targeted lysine mutagenesis, we provide evidence in support of a role for ubiquitin in the production of infectious PIV5 virions. MATERIALS AND METHODS Plasmids and lysine mutagenesis. Plasmids pcaggs-piv5 M, pcaggs-piv5 NP, and pcaggs-piv5 HN have been described before (47). cdnas encoding PIV5 M proteins with lysine to arginine substitutions were generated by PCR mutagenesis of the wildtype (wt) sequence, and subcloned into the eukaryotic expression vector pcaggs (29). cdna encoding tandem-tagged M-HS protein was generated by PCR, resulting in a modified M protein with the sequence HHHHHHWSHPQFEK appended to its C-terminal end. Plasmid pmt123 encoding HA-UB (53) was a kind gift of Cecile Pickart. Additional HA-UB expression plasmids, prk5-ha-ubiquitin-wt (prk5-ha-ubwt) and prk5-ha-ubiquitin-ko (prk5-ha- UBKO) (Addgene plasmids and 17603) were obtained from Addgene (Cambridge, MA). prk5-ha-ubko encodes HA-UB in which all lysine residues have been changed to arginine to prevent polyubiquitin chain formation (24). prk5-ha-ubwt is the analogous vector encoding wt ubiquitin. PIV5 infectious clone psv5 M.NS (46) was modified to generate plasmid psv5-m.k4,5,8,14,19,21,26r, used for recombinant virus rescue. Nomenclature for lysine mutants is based on the numbering of lysine residues within PIV5 M 5
6 protein from the N-terminal end to the C-terminal end. Amino acid positions are as follows: K3, a.a. position 36; K4, 79; K5, 80; K8, 130; K11, 155; K14, 192; K19, 247; K21, 287; K26, 325; K32, 366. Detection of ubiquitinated viral proteins. For detection of M protein ubiquitination, 293T cells in 10-cm-diameter dishes were transfected with pcaggs plasmids corresponding to PIV5 M-HS protein (or derivatives), 2 μg/dish, together with pmt123 plasmid encoding HA- UB, or prk5 plasmids encoding HA-UBWT or HA-UBKO, 2 μg/dish. 16 h post transfection (p.t.), the culture medium was replaced with Dulbecco's modified Eagle medium (DMEM) (Invitrogen, Carlsbad, CA) containing 10% fetal bovine serum (FBS). 40 h p.t., cells were harvested and lysates prepared in StrepTactin Lysis Buffer (100 mm Tris, ph 8; 150 mm NaCl; 1 mm EDTA; 1% Triton X-100; 0.1% SDS). For experiments with proteasome inhibitor treatments, 293T cells in 10-cm-diameter dishes were transfected with pcaggs plasmids corresponding to PIV5 M-HS protein (or derivatives), 0.8 μg/dish, together with prk5 HA-UBKO, 0.5 μg/dish. 14 h p.t., the culture medium was replaced with DMEM containing 10% FBS and MG-132 (Cayman Chemical, Ann Arbor, MI). 16 h p.t., the culture medium was replaced with fresh DMEM containing 10% FBS and MG h p.t., cells were harvested and lysates prepared in StrepTactin Lysis Buffer. Clarified lysates were incubated with StrepTactin MacroPrep resin (IBA BioTAGnology, Gottingen, Germany) for 2 h. The resin was then washed six times with StrepTactin Lysis Buffer and suspended in sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS- PAGE) loading buffer containing 2.5% (wt/vol) dithiothreitol. Samples were analyzed by SDS-PAGE using 10% gels. Immunoblotting was performed using the monoclonal antibody M-f (42) specific to PIV5 M protein (a kind gift of Richard Randall, St. Andrews University, St. Andrews, Scotland) and the antibody 12CA5 specific to the HA epitope tag (Abcam, Cambridge, MA). Detection was with a Fuji FLA-7000 laser scanner (FujiFilm Medical Systems, Stamford, CT). Alternatively, for detection of ubiquitination by radiolabeling and immunoprecipitation, 293T cells in 6-cm-diameter dishes were transfected with pcaggs plasmids corresponding to PIV5 M (or derivatives), 1 μg/dish unless indicated otherwise, or PIV5 NP protein, 6
7 ng/dish) together with pmt123 HA-UB, 1 μg/dish. At 24 h p.t., the culture medium was replaced with DMEM containing 1/10th the normal amounts of methionine and cysteine and 40 μci of [ 35 S] Promix/ml (Perkin Elmer, Waltham, MA). After an additional 18 h, cells were harvested, lysates were prepared in RIPA buffer (10 mm Tris, ph 7.4; 1% deoxycholate; 1% Triton X-100; 0.1% sodium dodecyl sulfate [SDS]) containing 0.15 M NaCl, 50 mm iodoacetamide, and 1 mm phenylmethylsulfonyl fluoride, and M proteins or NP proteins were immunoprecipitated with M-f and NP-a monoclonal antibodies respectively, as described (47). Proteins were resolved by SDS-PAGE on 10% gels and radiolabeled proteins were detected using a Fuji FLA-7000 laser scanner. Detection of free ubiquitin in purified virion preparations. MDBK cells in 10-cmdiameter dishes were infected with PIV5 at a multiplicity of infection (MOI) of 1 PFU/cell. Three days post-infection (p.i.), released virions were purified from media fractions by centrifugation through 20% sucrose cushions and flotation on sucrose gradients as described previously (37). Purified virions were suspended in PBS solution and digested with subtilisin A (Sigma-Aldrich, St. Louis, MO) (final concentration 0.2 µg/ml for 5 min. at 37 C). In some cases, virions were solubilized with Triton X-100 (American Bioanalytical, Natick, MA) (0.2% final concentration) prior to subtilisin digestion. Samples were mixed with equal volumes of SDS-PAGE loading buffer and fractionated by SDS-PAGE on 17.5% gels. Gels were stained with Coomassie Brilliant Blue (American Bioanalytical), or used for immunoblotting with antiubiquitin antibody (clone VU-1, LifeSensors, Malvern, PA). M protein affinity purification and mass spectrometry. 293T cells in groups of four 10-cm-diameter dishes were transfected with pcaggs plasmid encoding M-HS, 2 μg/dish, with or without pmt123 HA-UB, 2 μg/dish. 16 h p.t., the culture medium was replaced with DMEM containing 10% FBS. After an additional 24 h, cells were harvested and lysed using StrepTactin Lysis Buffer. Lysates were clarified by centrifugation and like samples were pooled. M proteins were purified from the lysates using an ÄKTAprime plus FPLC system (GE Healthcare Amersham Biosciences AB, Uppsala, Sweden) equipped with a 1 ml StrepTrap-HP column, with proteins eluted using StrepTactin Lysis Buffer containing 2.5 mm desthiobiotin (Sigma-Aldrich). Eluted proteins were concentrated to 1/4 the original volume 7
8 using U-Tube concentrators, (Novagen, Madison, WI), and the concentrated samples were resolved by SDS-PAGE using a 10% gel and stained with Coomassie Brilliant Blue. On the same gel, unconcentrated protein samples were also loaded for staining with SYPRO Orange (Bio-Rad Laboratories) and for immunoblotting with anti-m protein and anti-ha tag antibodies. A band corresponding to monoubiquitinated M protein species 2 was excised from the Coomassie-stained portion of the gel and submitted to the Taplin Mass Spectrometry Facility (Harvard Medical School, Boston, MA) for ubiquitination analysis by multidimensional liquid chromatography and tandem mass spectrometry (LC/LC-MS/MS), which was performed essentially as described (19). Measurements of VLP production. To generate VLPs, 293T cells in 6-cm-diameter dishes were transfected with pcaggs plasmids corresponding to PIV5 M (or derivatives), 0.4 μg/dish; PIV5 NP, 50 ng/dish; and PIV5 HN, 1 μg/dish. At 24 h p.t., the culture medium was replaced with DMEM containing 1/10 the normal amounts of methionine and cysteine and 40 μci of [ 35 S] Promix/ml. After an additional 18 h, cell and media fractions were harvested. Cell lysates were prepared with RIPA buffer and viral M, NP, and HN proteins were immunoprecipitated with M-f, NP-a, and HN1b monoclonal antibodies as described previously (47). VLPs were purified from media fractions by centrifugation through 20% sucrose cushions followed by flotation on sucrose gradients as described (37). Purified VLPs were loaded directly onto SDS gels and proteins were detected using a Fuji FLA-7000 laser scanner. Relative VLP production was calculated as the amount of M protein detected in purified VLPs divided by the amount of M protein detected in the corresponding RIPA cell lysate fraction, normalized to the value obtained with wt M protein. Recombinant virus generation. PIV5 was recovered from cdna as described (13, 38, 58). Briefly, BSR-T7 cells grown to 80% confluency in 6-cm-diameter dishes were transfected with variants of the infectious clone psv5 M.NS (46), together with helper plasmids encoding PIV5 NP, P, and L proteins as described (58). The cells were incubated in DMEM supplemented with 2% FBS at 37 C for 4 days. The culture media was harvested and used to infect BHK-21F cells for virus plaque-purification as described (35). Plaquepurified viruses were passaged in MDBK cells to generate virus stocks. Total RNA was 8
9 isolated from infected MDBK cells and used as template for reverse transcription and PCR amplification for sequencing of the entire PIV5 genome as described (13). DNA sequencing was performed by the Pennsylvania State University Genomics Core Facility. Virus growth curve analysis. Growth curve analysis was performed as described previously (38, 44). Briefly, MDBK cells grown in 24-well plates were infected with recombinant viruses at a MOI of 0.01 PFU/cell. After adsorption, cells were washed with PBS and cultures were incubated at 37 C in DMEM supplemented with 2% FBS for 0, 12, 24, 48, 72, 96, 120, or 144 h. Culture supernatants were collected at each time point and virus titers were measured by plaque assay on BHK-21F cells as described (35). Measurements of virion production and virion infectivity. MDBK cells in 6-cmdiameter dishes were infected with recombinant viruses at a MOI of 1.0 PFU/cell. At 24 h p.i., the culture medium was replaced with DMEM containing 1/10 the normal amounts of methionine and cysteine and 40 μci of [ 35 S] Promix/ml. After an additional 16 h, the media fraction was clarified by low speed centrifugation, and a 0.05 ml aliquot (3.3% of the total volume) was removed for titer determination by plaque assay. The remainder of the media fraction was used for virion purification, which was performed according to the same procedure described above for VLP purification. Cell fractions were used for lysate preparation and immunoprecipitation of viral proteins as described above. Purified virions and immunoprecipitated proteins were fractionated by SDS-PAGE on 10% gels and detected using a Fuji FLA-7000 laser scanner. Relative virion production was calculated as the amount of M protein detected in purified virions divided by the amount of M protein detected in the corresponding RIPA cell lysate fraction, normalized to the value obtained with wt PIV5. PFU:particle ratios were calculated as the total number of PFU detected in the culture media divided by the quantity of M protein detected in the corresponding purified virions, normalized to the value obtained with wt rpiv5. Immunofluorescence microscopy. CV-1 cells grown on glass coverslips to 50% confluency were infected with recombinant viruses, rpiv5 or rpiv5-m.k4,5,8,14,19,21,26r, at a MOI of 1.0 PFU/cell. At 18 h p.i., culture media was replaced with DMEM supplemented 9
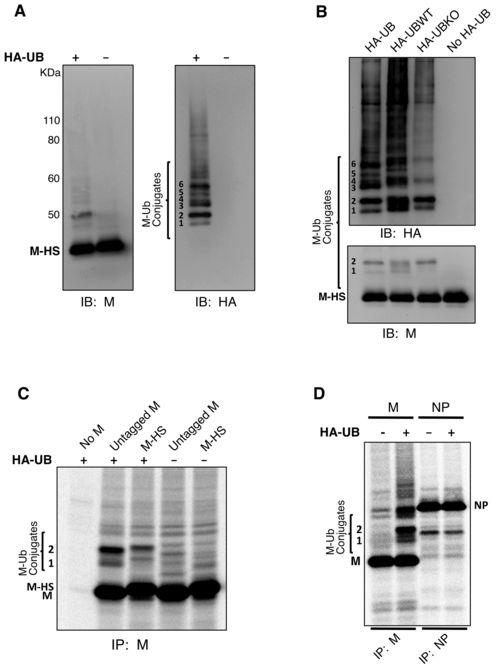
10 with 2% FBS containing MG-132 at concentrations of 0, 10, or 25 μm. At 24 h p.i., monolayers were fixed with 4% methanol-free formaldehyde in PBS for 15 min. Cells were incubated for 30 min. at room temperature in a solution containing 1% BSA and 0.1% cold water fish gelatin in PBS, then incubated for 2 h with the HN protein-specific mab HN1b (IgG2a isotype). Cells were washed three times with PBS and permeabilized by incubating for 30 min. at room temperature in a solution containing 1% BSA, 0.1% cold water fish gelatin, and 0.1% saponin, then incubated for 2 h with the M protein-specific mab M-h (IgG3 isotype). Cells were washed three times with PBS and incubated for 1 h with a mixture of IgG2a-specific Alexa Fluor 594 and IgG3-specific Alexa Fluor 488 goat anti-mouse secondary antibodies (Invitrogen). Cells were washed an additional three times with PBS and nuclei were stained with Hoechst (Invitrogen). Coverslips were mounted on microscope slides and cells were visualized with a Zeiss AxioImager M1 fluorescence microscope (Zeiss, Inc., Thornwood, N.Y.). Images were captured using a Hamamatsu Orca R2 CCD camera. RESULTS PIV5 M protein is a target for ubiquitin conjugation. Several retroviral Gag proteins, as well as Ebola VP40 and VSV M proteins, can act as targets for ubiquitin attachment (7, 10, 11, 32, 34, 50). To determine whether PIV5 M protein can be targeted for ubiquitination in transfected mammalian cells, M protein that harbors tandem C-terminal 6xHis and Strep (II) tags (M-HS) was expressed together with HA-tagged ubiquitin (HA-UB) in 293T cells and pulled down from cell lysates using affinity resin specific to the Strep (II) tag. Several distinct bands migrating above M protein, consistent with M protein-ubiquitin conjugates, were detected on immunoblots probed with anti-m or anti-ha antibodies (Fig. 1A). These bands could not be detected in lysates from cells transfected to express HA-UB alone (data not shown). Six distinct ubiquitinated M protein species were observed and numbered 1-6 from the lowest to the highest apparent molecular weight (Fig. 1A). The most prominent of these are species 2 and species 6. Species 2 has an apparent molecular weight of 50 kda, and this is roughly consistent with the addition of a single, 8.5 kda HA-ubiquitin molecule to M-HS protein, which migrates with an apparent molecular weight of 42 kda. Species 6 has an 10
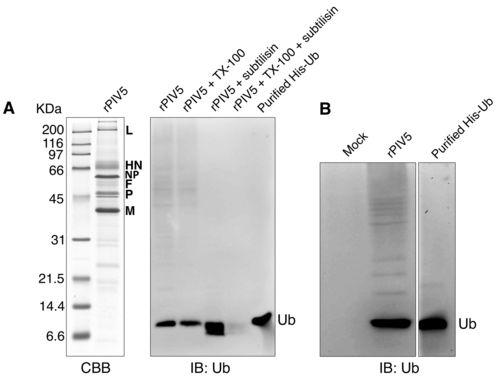
11 apparent molecular weight of 58 kda, consistent with the addition of two HA-ubiquitin molecules to M-HS. To further characterize these modified M proteins, a lysine-deficient HA-tagged ubiquitin (HA-UBKO) (24) that does not allow polyubiquitin chain formation was expressed. HA-UBKO expression substantially reduced the formation of ubiquitinated M protein species 3, 4, 5, and 6, but had little effect on species 1 and 2 (Fig. 1B), thereby defining species 1 and 2 as likely monoubiquitinated forms of M protein. Reduced levels of some higher-molecular weight forms of ubiquitinated M protein could still be detected upon HA-UBKO expression (Fig. 1B). These may represent M protein that is multiply monoubiquitinated. To confirm that ubiquitin conjugation occurs not only with tandem-tagged M protein but also with unmodified M protein, M proteins were expressed together with HA-UB, and after radiolabeling of cells the M proteins were collected from cell lysates by immunoprecipitation (Fig. 1C). Bands consistent with monoubiquitinated M protein species 1 and 2 were detected in both cases, with small differences in migration attributable to the mass conferred by the tandem affinity tags (Fig. 1C). To rule out the possibility that HA-UB overexpression might nonspecifically lead to ubiquitination of any viral protein, HA-UB was expressed together with the PIV5 NP protein. In this case, no evidence for ubiquitination could be detected, in contrast to results obtained with PIV5 M protein (Fig. 1D). Of note, HA-UB expression, which has the potential to enhance polyubiquitination and induce protein degradation, did not reduce the levels of either M protein or NP protein in cell lysates (Fig. 1D). Detection of free ubiquitin in PIV5 virions. To investigate the possibility that free ubiquitin might be present within PIV5 virions, sucrose gradient-purified virions produced from infected MDBK cells (in the absence of HA-UB expression) were analyzed by immunoblotting with anti-ubiquitin antibodies (Fig. 2A, B). Free ubiquitin was readily detected in the purified virion preparations, and this ubiquitin was resistant to digestion with the protease subtilisin (Fig. 2A). Treatment of virion preparations with nonionic detergent (0.2% Triton X-100) caused the ubiquitin to become sensitive to subtilisin digestion. These findings suggest that free ubiquitin is contained within virus particles and is protected from protease digestion by virion membranes. To further establish that ubiquitin is contained within virus particles, as 11
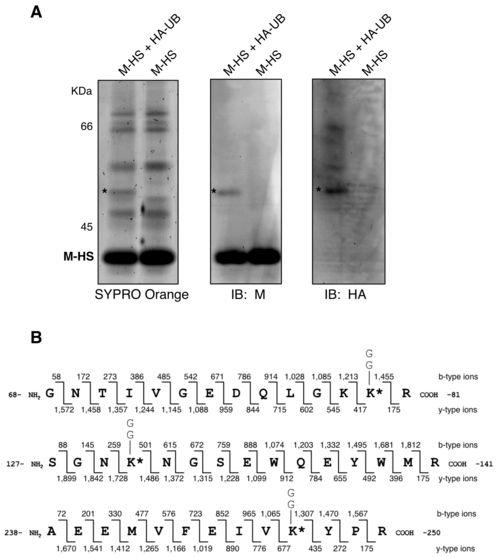
12 opposed to exosomes or microvesicles that can co-purify with virions on sucrose gradients (2, 5, 31), virions were purified as before and in parallel, supernatants from mock-infected cells were subjected to the same purification procedure. Once again, free ubiquitin was readily detected in purified virions, but no ubiquitin was detected in samples prepared from uninfected cells (Fig. 2B). These findings indicate that free ubiquitin is contained within PIV5 virions, consistent with earlier observations made with HIV-1 and other enveloped viruses (27, 32, 41, 55). Identification of M protein lysine residues targeted for ubiquitin attachment. To define primary sites of ubiquitin attachment to PIV5 M protein, 293T cells were transfected to produce M-HS together with HA-UB. M-HS was affinity-purified from cell lysates by FPLC via the Strep (II) tag, and the resulting proteins were fractionated by SDS-PAGE for total protein staining and immunoblotting (Fig. 3A). A band consistent with monoubiquitinated M protein species 2 was detected by whole protein staining and by immunoblotting with either anti-m protein or anti-ha antibodies. This band was excised and subjected to in-gel trypsinization and multidimensional liquid chromatography and tandem mass spectrometry (LC/LC-MS/MS) for ubiquitination analysis, which is based on detection of di-glycine signatures derived from the C-terminus of ubiquitin that are left on ubiquitinated amino acids following trypsinization, leading to Da mass increases (19, 20). Three peptides harboring ubiquitinated lysine residues were identified (Fig. 3B). One peptide spanning amino acid (a. a.) residues was ubiquitinated, with conjugation most likely occurring at position 80 (lysine 5), although conjugation at the neighboring position 79 (lysine 4) could not be completely ruled out. A second peptide spanning a. a. residues was ubiquitinated at position 130 (lysine 8). A third peptide spanning a. a. residues was ubiquitinated at position 247 (lysine 19). These lysine residues will be referred to throughout this study as K4, K5, K8, and K19. Based on this analysis of M protein ubiquitination, M protein species 2 likely represents a heterogeneous population of monoubiquitinated M proteins. The relative frequency of ubiquitination at each particular site could not be determined using this approach. In addition, it should be noted that out of 33 total lysines contained within PIV5 M protein, 21 were covered by one or more of the MS-identified peptides, while the remaining 12 (K1, K3, K11, K12, K18, K20, K22, K26, K28, K29, K30, and K33) were not covered by any of these 12
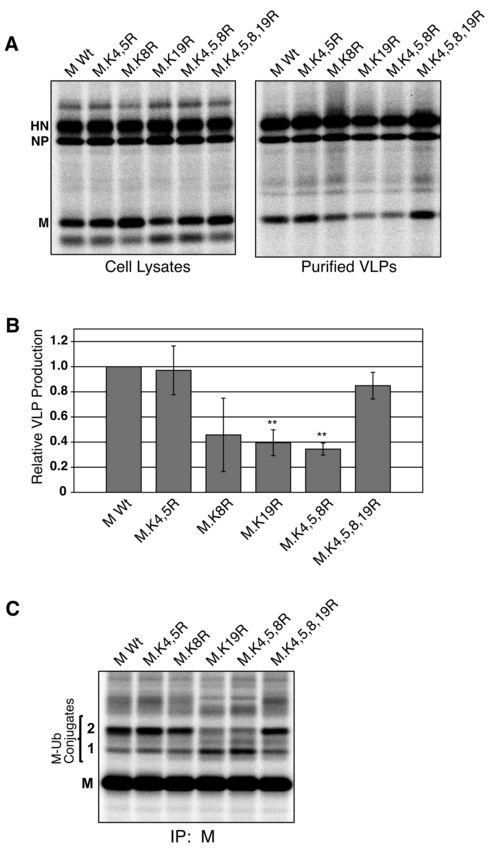
13 peptides. However, of these 12 lysines, 7 (all but K11, K20, K28, K29, and K33) were used as cleavage sites during the trypsinization process, and consequently are unlikely to have been ubiquitinated, as trypsin cannot cleave at ubiquitinated lysine residues (20, 39). Mutagenesis of MS-identified ubiquitin acceptor sites in PIV5 M protein alters VLP production function. To assess the importance of PIV5 M protein ubiquitination for M protein function, MS-identified lysine residues were mutated individually and in combination to arginine residues, and the resulting lysine-altered M proteins were tested for the ability to direct VLP production. Combined lysine mutations were made successively from the N- terminus of M protein. Mutant M proteins were co-expressed with PIV5 NP and HN proteins in 293T cells by transient transfection. VLPs were collected from the culture medium, purified using sucrose gradients, and quantified after SDS-PAGE (Fig. 4A, B). In addition, to evaluate the impact of lysine alterations on M protein ubiquitination, mutant M proteins were coexpressed together with HA-UB, followed by radiolabeling and M protein immunoprecipitation (Fig. 4C). Mutation of K4 and K5 (M.K4,5R) had little effect either on M protein ubiquitination, judged by detection of ubiquitinated species 1 and 2, or on the efficiency of VLP production (Fig. 4). Similarly, little change in ubiquitination was observed upon mutation of K8, although VLP production was variable in this case. In contrast, mutation of K19 led to a clear change in M protein ubiquitination marked by a decrease in ubiquitinated species 2 and an increase in ubiquitinated species 1 (Fig. 4C). This change in ubiquitination was accompanied by a reproducible and statistically significant impairment in VLP production function, to a level that was about 40% of the wt M protein (Fig. 4A,B). Very similar results were obtained with M protein mutant M.K4,5,8R (K4, K5, and K8 all changed to arginine), with changes in M protein ubiquitination correlated with poor VLP production function. Hence, normal ubiquitination appears to be important for one or more M protein functions necessary for VLP release (e.g., trafficking, assembly at membranes, or particle budding). Unexpectedly, mutation of K4, K5, K8, and K19 together (M.K4,5,8,19R) led to M protein ubiquitination that was indistinguishable from that observed with wt M protein (Fig. 4C). This restoration in M protein ubiquitination was accompanied by a return to wt levels of VLP production (Fig. 4A, B). The restoration of M protein ubiquitination and VLP production function upon cumulative removal of all MS-identified ubiquitin acceptor sites supports the possibility that ubiquitin is attached to 13
14 alternative sites on M protein when the primary ones have been removed, and that only a subset of these secondary acceptor sites are capable of supporting M protein budding function upon ubiquitin attachment. Cumulative mutagenesis of seven lysine residues in PIV5 M protein further disrupts VLP production. In an effort to more completely affect M protein ubiquitination, additional lysine residues were targeted for mutation based on the ubiquitination prediction algorithm UbiPred (54). Lysines 3, 4, 5, 11, 14, 19, 21, 26, and 32 were identified by UbiPred as potential sites of M protein ubiquitination, with threshold scores above 0.5. Of these, K4, K5, and K19 are MS-identified ubiquitin acceptor sites (Fig. 3B). K14, K21, and K32 are present within MS-identified peptides, but were not identified by MS as ubiquitin acceptor sites. K3, K11, and K26 were not covered by any of the MS-identified peptides. UbiPredidentified lysines were mutated individually to arginine and VLP production and M protein ubiquitination were assessed as before. For most of these mutants, there was no noticeable defect in either VLP production or ubiquitination (data not shown). M.K11R and M.K32R mutations each led to disruption of VLP production function to levels less than 15% of wt M protein, but had no detectable effects on M protein monoubiquitination (data not shown). It is possible that these mutations disrupt M protein function for reasons unrelated to ubiquitination, and they were not included in subsequent cumulative mutagenesis experiments for this reason. M protein lysines 4, 5, 8, 14, 19, 21, and 26 were targeted for cumulative mutation to arginine, with mutations made successively from the N-terminus of the protein. Simultaneous mutation of the first four of these lysines (M.K4,5,8,14R) did not have a statistically significant impact on VLP production function (Fig. 5A, B). However, simultaneous mutation of five, six, or seven lysine residues led to a progressive worsening of VLP production function, down to approximately 10% of normal levels when all seven lysine residues were removed (Fig. 5A, B). Several of these lysine to arginine mutations were also introduced into M protein individually, resulting in smaller effects on VLP production, although K26R mutation reduced VLP production to 30% of the wt level (Fig. 5C). Thus, the most severe defects in VLP production were observed only upon cumulative mutation of lysine residues. Cumulative mutagenesis had no substantial effects on M protein stability or M protein membrane-binding 14
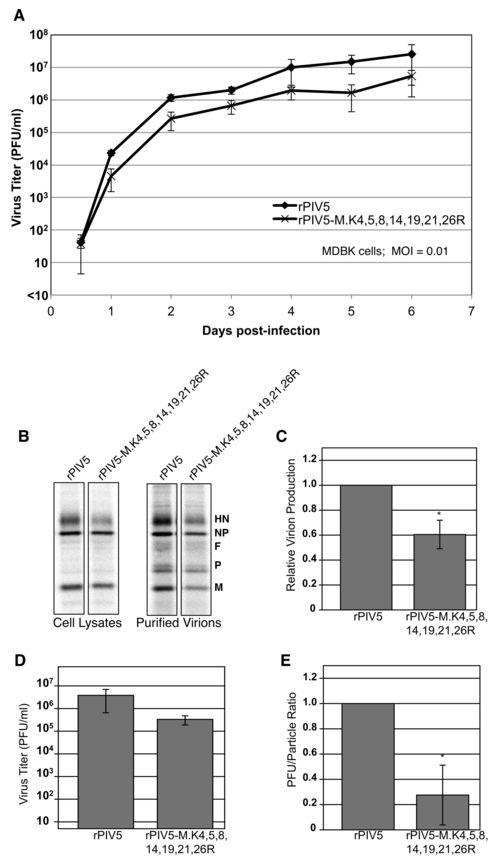
15 function, however (data not shown), indicating that these impairments are likely to be specific to the virus assembly and/or budding functions of M protein. The same cumulative M protein mutants that were defective for VLP production (those with five or more lysines removed) also exhibited visible alterations in the M protein ubiquitination pattern (Fig. 5D), with a decrease in the intensity of species 2 and an increase in the intensity of species 1 as observed before with M.K4,5,8R (Fig. 4D). Further confirmation of altered ubiquitination was obtained for the seven-lysine-removed mutant M.K4,5,8,14,19,21,26R. This mutant M protein was appended with tandem 6xHis and Strep (II) affinity tags, and expressed in cells together with HA-UB. Analysis of affinity-isolated M proteins on anti-ha immunoblots confirmed a shift in the ubiquitination profile for the mutant M, marked by a decline in the intensity of M protein species 2 and an elevation in the intensity of species 1 (Fig. 5E). A recombinant virus encoding seven-lysine-mutant M protein exhibits altered multiplication kinetics and impaired infectious virion production. PIV5 encoding cumulative lysine-to-arginine mutant M protein M.K4,5,8,14,19,21,26R in place of the wt M protein was recovered by reverse genetics using methods described previously (13, 58). Virus was propagated by passaging in MDBK cells and the viral titers were measured by plaque assay on BHK-21F cells. The complete viral genome was sequenced to confirm the presence of the intended lysine-to-arginine mutations and the absence of any additional mutations, and this genome sequence was found to be stable for at least three passages in cell culture. Growth characteristics were assessed using a multi-step growth curve (MOI = 0.01 PFU/cell) in MDBK cells (Fig. 6A). The maximum titer achieved by rpiv5- M.K4,5,8,14,19,21,26R was about 5 x 10 6 PFU/ml, a reduction of approximately 6-fold compared to wt virus. Hence, seven-lysine-altered virus exhibits impaired multiplication kinetics in cultured mammalian cells. To determine if seven-lysine-altered virus exhibits particle production defects, MDBK cells were infected (MOI = 1.0 PFU/cell), and particles were purified from the culture supernatants after radiolabeling of cells. Viral protein synthesis was not significantly impaired in mutant virus-infected cells, judged by the levels of HN, NP, and M proteins detected by immunoprecipitation from infected cell lysates (Fig. 6B, left panel). Particle production, 15
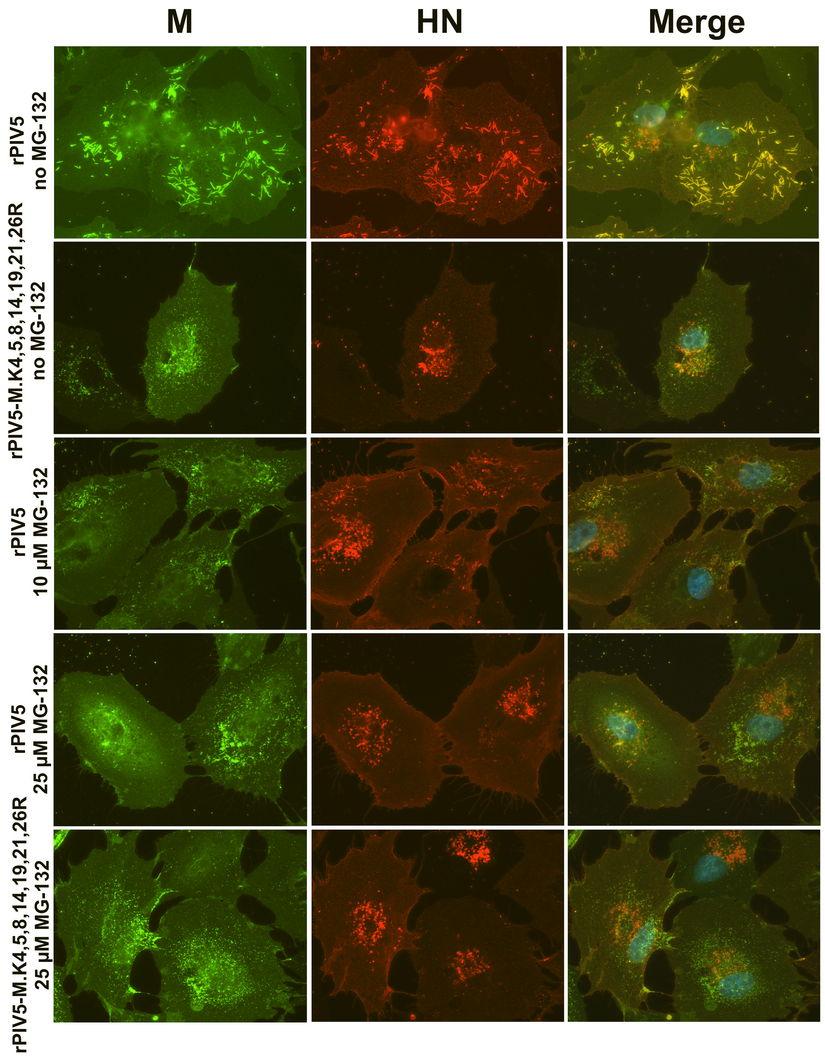
16 however, was moderately impaired for the mutant virus, declining to about 60% of the level observed for wt virus (Fig. 6B, C). There were no obvious differences in polypeptide composition between mutant virus and wt virus (Fig. 6B, right panel). Viral titers were determined by plaque assay from small aliquots of culture supernatants, which had been reserved prior to virion purification. Mutant virus titer was reduced by approximately 10-fold as compared to wt virus titer (Fig. 6D). PFU/particle ratios were calculated to estimate viral infectivity, and the mutant virus PFU/particle ratio was found to be nearly 5-fold lower than that of wt virus (Fig. 6E). Overall, these results indicate that seven-lysine-mutant virus is defective, but that the nature of the defect is somewhat different from what was expected based on VLP production experiments (Fig. 5A). Instead of a severe reduction in particle release, the mutant virus exhibits only a minor defect in particle release. The larger defect for this virus is that the majority of particles that are released are noninfectious. Recombinant PIV5 encoding seven-lysine-mutant M protein is assembly deficient. The localizations of viral M and HN proteins in cells infected with wt rpiv5 and rpiv5-m.k4,5,8,14,19,21,26r were examined by immunofluorescence microscopy. In cells infected with wt rpiv5, surface HN protein and intracellular M protein were found strongly colocalized at filament-shaped clusters (Fig. 7), which are presumed to represent sites of active virus budding (38, 43, 44, 58). Some M protein is also found outside of these clusters, somewhat diffused within the cytoplasm (38). In cells infected with rpiv5- M.K4,5,8,14,19,21,26R, a clear redistribution of M and HN proteins was observed. M and HN proteins were localized in discrete punctate clusters, as well as diffused on the cell surface (Fig. 7). Filament-shaped structures were rarely detected in these cells, either by M or HN protein staining. This redistribution of M and HN proteins appears to be similar to that which was observed previously for PIV5 harboring glycoproteins with truncated cytoplasmic tails (44, 58). These findings indicate that cumulative mutations to lysine residues in PIV5 M protein results in a virus assembly defect. Proteasome inhibitor treatment impairs PIV5 assembly. PIV5 particle production was previously shown to be inhibited by proteasome inhibitor treatments (46). Here, VLPproducing cells expressing either wt M-HS or seven-lysine-mutant M-HS were treated with 16
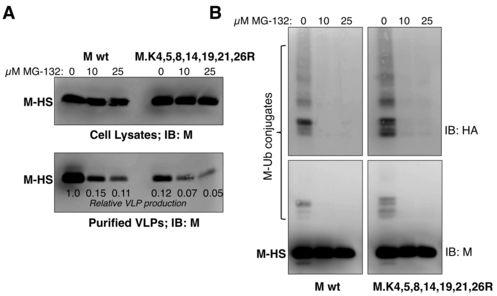
17 the proteasome inhibitor MG-132. This reduced VLP production driven by the wt M-HS protein by 5- to 10-fold (Fig. 8A), consistent with previous observations (46). VLP production driven by seven-lysine-mutant M protein was poor in the absence of MG-132, and was further reduced in the presence of MG-132. However, in this case the magnitude of the reduction caused by MG-132 treatment was substantially smaller (1.7- to 2.5-fold). This finding is consistent with the possibility that proteasome inhibitor treatment and M protein lysine removal inhibit VLP production through related mechanisms. To determine the effect of MG-132 treatment on PIV5 M protein monoubiquitination, cells were transfected to express either wt M-HS or seven-lysine-mutant M-HS, together with HA- UBKO, and the transfected cells were treated with increasing amounts of MG-132. Ubiquitination of M-HS could no longer be detected after treatment of cells with 10 µm MG- 132 (Fig. 8B). MG-132 treatment also prevented the ubiquitination of seven-lysine-mutant M protein. To more clearly define the defect in PIV5 replication caused by proteasome inhibitor treatment, PIV5-infected cells treated with increasing concentrations of MG-132 were examined by fluorescence microscopy (Fig. 7). MG-132 treatment impaired the assembly of PIV5 M and HN proteins into filament-shaped clusters. Fewer of these assembly structures were observed, and M protein was instead visible in punctate clusters, similar to the localization observed for rpiv5-m.k4,5,8,14,19,21,26r in the absence of proteasome inhibitor treatment. MG-132 treatment of rpiv5-m.k4,5,8,14,19,21,26r-infected cells did not further alter the localizations of viral proteins (Fig. 7). Overall, these findings suggest that similar impairments to PIV5 assembly can be induced either through preventing M protein ubiquitination (via proteasome inhibitor treatment) or through altering the sites at which ubiquitin can be conjugated to M protein (through lysine removal). DISCUSSION 17
18 Several enveloped viruses require ubiquitin-binding ESCRT components for budding, which has led to the notion that ubiquitination of viral and/or host factors at virus assembly sites could be necessary for the efficient recruitment of these components (27, 55, 62). Here, we demonstrate that PIV5 M protein is a target for ubiquitination and, using MS-analysis, identify lysine residues that act as the preferred monoubiquitination sites. Cumulative mutagenesis of M protein lysine residues (both MS-identified and predicted ubiquitination sites) led to a reduction in PIV5 M protein monoubiquitination and impaired VLP production. Recombinant PIV5 encoding lysine-mutant M protein exhibited reductions both in the quantity of particles produced and the infectivity of those particles. Infection with this virus also led to mis-localization of M protein within the infected cells, consistent with altered virus assembly, rather than a late budding defect. A similar mis-localization of M protein was observed in cells infected with wt PIV5 and treated with the proteasome inhibitor MG-132. These findings suggest that, while PIV5 budding is ESCRT-dependent (46), PIV5 M protein ubiquitination likely contributes to M protein trafficking and assembly events either instead of or in addition to the budding steps that are facilitated by ESCRT proteins. Several Gag and matrix proteins from retroviruses, filoviruses, rhabdoviruses, and paramyxoviruses have been shown to be monoubiquitinated or multiply monoubiquitinated (10, 11, 27, 30, 57). In addition, monoubiquitination of influenza virus NP protein was recently reported (23). We report here that PIV5 M protein, but not PIV5 NP protein, is a target for ubiquitin conjugation in cells transfected to produce viral proteins together with HA- UB. The most abundant form of ubiquitinated M protein is a monoubiquitinated species, designated here as monoubiquitinated M protein species 2. MS-based ubiquitination analysis of this species revealed it to be composed of a heterogeneous population of PIV5 M proteins monoubiquitinated on at least three independent sites: K4/K5, K8, and K19. The mechanism by which ubiquitin is conjugated to these specific lysine residues within PIV5 M protein has not yet been defined, and warrants further investigation. Matrix proteins of some negativestrand RNA viruses are ubiquitin-modified through the action of Nedd4-like ubiquitin ligases, which recognize the PPxY-type late domains that are contained within these matrix proteins (10, 11, 52, 61). PIV5 M protein lacks PPxY-type late domain sequences, however, and it is not yet clear which cellular ubiquitin-ligase(s) are responsible for ubiquitin attachment in this case. 18
19 One of the first findings indicating a role for ubiquitin in retrovirus budding was the detection of high levels of free ubiquitin in the virions of retroviruses, including avian leukosis virus, RSV, HIV-1, simian immunodeficiency virus (SIV), and murine leukemia virus (MLV) (33, 34, 41). Here, we report that free ubiquitin co-purified with PIV5 virions and was most likely enclosed within virion membranes. The precise source of the free ubiquitin that is found in virions remains unverified, though it is postulated that deubiquitination of ubiquitinated viral or cellular proteins during virus release by ubiquitin hydrolases would account for the presence of the unconjugated ubiquitin in virions (41). Free ubiquitin incorporation could also result from ubiquitin binding by viral or cellular proteins that are involved in the viral budding process. Removal of lysine residue acceptor sites from viral proteins through lysine to arginine mutagenesis is a key strategy that has been used to investigate the consequences of Gag or matrix protein ubiquitination for virus budding (6, 14, 22, 49). A number of potential complications are inherent to this approach and must be carefully navigated, however. One is the possibility that lysines might provide important functions that are unrelated to ubiquitin attachment. Lysine residues act as targets for a variety of post-translational modifications in addition to ubiquitination, including acetylation (60) and SUMOylation (1). In addition, lysine residues are often key residues in nuclear localization motifs (26) and are sometimes important for protein-protein interactions that dictate intracellular localization, such as ER localization specified by di-lysine motifs (4). In this study, focus was placed on lysine residues which, when removed, induced changes both to the M protein ubiquitination profile and to M protein VLP production function. Some lysine to arginine mutations, such as K11R and K32R, led to severe VLP production defects without visibly affecting the M protein ubiquitination pattern. These mutations were excluded from cumulative mutagenesis experiments, as the phenotypes observed could be completely unrelated to ubiquitination. However, even in these cases it is possible that M protein ubiquitination was altered, but in a more subtle way that did not upset the relative intensities of the various ubiquitinated species that could be resolved on SDS gels. Hence, further study will be needed to determine why removal of these particular lysines from PIV5 M protein led to VLP production defects. A second complication inherent to the lysine mutagenesis approach is the flexibility of ubiquitin ligases during the selection of ubiquitin attachment sites, which frequently allows for 19
20 the use of secondary attachment sites when the primary ones have been removed. For example, budding function of RSV Gag protein is impaired only when all five lysine residues in the MA portion of the protein, near the PPPY late domain, have been removed. If any one of these five lysine residues is present, budding function is restored (49). Likewise, removal of individual lysine residues from HIV-1 Gag protein did not severely restrict budding function; only cumulative replacement of all lysine residues within NC and SP2 or in NC and p6 substantially impaired the budding of particles (6). Guided by these previous studies, we placed our experimental emphasis here on cumulative lysine-to-arginine mutagenesis of the PIV5 M protein, and indeed we observed the most severe effects on particle formation when 7 lysine residues were simultaneously changed to arginine residues. Although less emphasis here was placed on the corresponding single mutant M proteins, some of these could also be informative and are worthy of further study, in particular the K26R mutation which alone caused a reduction in VLP production to about 30% of normal levels. Even within the framework of cumulative M protein mutagenesis, interpretation of results was not completely straightforward in some instances. In particular, removal of all four of the MS-identified lysine residues from M protein (K4,5,8,19R) did not impair VLP production function, whereas removal of just three of those lysine residues (K4,5,8R) substantially impaired VLP production. This suggests that the relationship between M protein ubiquitination and M protein VLP production function is complex, with ubiquitin attachment redirected to alternative sites when the primary ones have been removed, and only a subset of these sites able to contribute to the VLP production function upon ubiquitin attachment. Additional complexities were observed in M proteins altered at K3. K3 was among the nine lysine residues identified by the UbiPred algorithm as potential ubiquitin attachment sites, and ubiquitin attachment at K3 likely accounts for the aberrantly-migrating ubiquitinated M protein species 1 (data not shown). M proteins with cumulative removal of lysine residues were prepared both with K3 intact (Fig. 5) and K3 removed (data not shown). M proteins with K3 removed were generally more functional for VLP production than M proteins with K3 intact, suggesting that ubiquitin attachment at K3 contributes little to the virus assembly process. Removal of primary lysine acceptor sites in the context of an intact K3 may favor a nonproductive shift in ubiquitination towards K3, as evidenced by changes in the intensities of ubiquitinated species 1 and 2 (Fig. 20
21 C-D). K3 removal may further redirect the shift in ubiquitination towards other, alternative lysine residues, to the benefit of particle release. Recently, Wang et al. investigated the importance of a lysine residue at position 258 of the Nipah virus M protein, which is conserved among paramyxoviruses. Substitution with alanine or arginine at this position resulted in defects to VLP production, M protein nuclear localization, M protein ubiquitination, and M protein membrane binding (57). The analogous residue within PIV5 M protein is at amino acid position 259 (K20). We have found that mutation of K20 to alanine or arginine severely impairs PIV5 VLP production, but does not alter the pattern of M protein ubiquitination and does not obviously affect the nuclearcytoplasmic distribution of the protein (data not shown). It is possible that the function provided by this lysine residue differs somewhat among the paramyxovirus M proteins. For retroviruses such as HIV-1, RSV, and HTLV-1, particle release defects can be induced through cumulative lysine to arginine mutagenesis of Gag proteins, or through proteasome inhibitor treatments that deplete free ubiquitin levels (3, 6, 14, 22, 36, 48-50, 56). Here, we observed defects in the earlier steps of PIV5 assembly when its M protein was modified through cumulative lysine to arginine mutation, or upon treatment of infected cells with a proteasome inhibitor. While late defects in retrovirus budding could in some cases be restored through direct fusion of ubiquitin to Gag (18, 36, 62), we were unable to restore PIV5 assembly through fusion of ubiquitin to PIV5 M protein (data not shown). This may be due to difficulties in maintaining M protein folding in the context of ubiquitin fusion, however, as even wt M protein fused to the N- or C-terminal end of ubiquitin was nonfunctional for VLP production. Recombinant PIV5 encoding seven-lysine-mutant M protein exhibited impaired growth kinetics, reduced particle release, and poor particle infectivity. Microscopy analysis of cells infected with this virus revealed a redistribution of M protein consistent with a defect in virus assembly, and a similar redistribution was observed in cells infected with wt virus and treated with the proteasome inhibitor MG-132. We interpret these results to suggest that PIV5 M protein monoubiquitination may be important for the trafficking of the protein to budding sites at the plasma membrane. Dysregulation of the assembly process may lead to altered particles and reduced infectivity, although the polypeptide composition of released virions was not noticeably altered. Further investigation is required to more precisely define this ubiquitin-related defect in virion infectivity. 21
22 ACKNOWLEDGMENTS We thank Cecile Pickart for cdna corresponding to HA-Ubiquitin, and we thank the laboratory of Dr. Kouacou Konan for assistance with fluorescence microscopy. This work was supported by research grant AI from the National Institute of Allergy and Infectious Diseases to A.P.S. and predoctoral fellowship award U from the American Heart Association to M.S.H. REFERENCES 1. Anckar, J., and L. Sistonen SUMO: getting it on. Biochem. Soc. Trans. 35: Bess, J. W., Jr., R. J. Gorelick, W. J. Bosche, L. E. Henderson, and L. O. Arthur Microvesicles are a source of contaminating cellular proteins found in purified HIV-1 preparations. Virology 230: Bouamr, F., J. A. Melillo, M. Q. Wang, K. Nagashima, M. de Los Santos, A. Rein, and S. P. Goff PPPYVEPTAP motif is the late domain of human T-cell leukemia virus type 1 Gag and mediates its functional interaction with cellular proteins Nedd4 and Tsg101. J. Virol. 77: Cosson, P., and F. Letourneur Coatomer interaction with di-lysine endoplasmic reticulum retention motifs. Science 263: Gluschankof, P., I. Mondor, H. R. Gelderblom, and Q. J. Sattentau Cell membrane vesicles are a major contaminant of gradient-enriched human immunodeficiency virus type-1 preparations. Virology 230: Gottwein, E., S. Jager, A. Habermann, and H. G. Krausslich Cumulative mutations of ubiquitin acceptor sites in human immunodeficiency virus type 1 Gag cause a late budding defect. J. Virol. 80: Gottwein, E., and H. G. Krausslich Analysis of human immunodeficiency virus type 1 Gag ubiquitination. J. Virol. 79: Haglund, K., and I. Dikic Ubiquitylation and cell signaling. EMBO J. 24:
23 Harrison, M. S., T. Sakaguchi, and A. P. Schmitt Paramyxovirus assembly and budding: building particles that transmit infections. Int. J. Biochem. Cell Biol. 42: Harty, R. N., M. E. Brown, J. P. McGettigan, G. Wang, H. R. Jayakar, J. M. Huibregtse, M. A. Whitt, and M. J. Schnell Rhabdoviruses and the cellular ubiquitin-proteasome system: a budding interaction. J. Virol. 75: Harty, R. N., M. E. Brown, G. Wang, J. Huibregtse, and F. P. Hayes A PPxY motif within the VP40 protein of Ebola virus interacts physically and functionally with a ubiquitin ligase: implications for filovirus budding. Proc. Natl. Acad. Sci. USA 97: He, B., G. Y. Lin, J. E. Durbin, R. K. Durbin, and R. A. Lamb The SH integral membrane protein of the paramyxovirus simian virus 5 is required to block apoptosis in MDBK cells. J. Virol. 75: He, B., R. G. Paterson, C. D. Ward, and R. A. Lamb Recovery of infectious SV5 from cloned DNA and expression of a foreign gene. Virology 237: Heidecker, G., P. A. Lloyd, F. Soheilian, K. Nagashima, and D. Derse The role of WWP1-Gag interaction and Gag ubiquitination in assembly and release of human T-cell leukemia virus type 1. J. Virol. 81: Hershko, A., A. Ciechanover, and A. Varshavsky The ubiquitin system. Nat. Med. 6: Hicke, L., and R. Dunn Regulation of membrane protein transport by ubiquitin and ubiquitin-binding proteins. Annu. Rev. Cell Dev. Biol. 19: Horvath, C. M Weapons of STAT destruction. Interferon evasion by paramyxovirus V protein. Eur. J. Biochem. 271: Joshi, A., U. Munshi, S. D. Ablan, K. Nagashima, and E. O. Freed Functional replacement of a retroviral late domain by ubiquitin fusion. Traffic 9: Kaiser, P., and J. Wohlschlegel Identification of ubiquitination sites and determination of ubiquitin-chain architectures by mass spectrometry, p In R. J. Deshaies (ed.), Ubiquitin and Protein Degradation, Part B, vol Elsevier Academic Press, San Diego. 20. Kirkpatrick, D. S., C. Denison, and S. P. Gygi Weighing in on ubiquitin: the expanding role of mass-spectrometry-based proteomics. Nat. Cell Biol. 7: Lamb, R. A., and G. D. Parks Paramyxoviridae : the viruses and their replication, p In D. M. Knipe and P. M. Howley (ed.), Fields Virology, Fifth ed. Lippincott, Williams and Wilkins, Philadelphia. 22. Le Blanc, I., A. R. Rosenberg, and M. C. Dokhelar Multiple functions for the basic amino acids of the human T-cell leukemia virus type 1 matrix protein in viral transmission. J. Virol. 73:
24 Liao, T. L., C. Y. Wu, W. C. Su, K. S. Jeng, and M. M. Lai Ubiquitination and deubiquitination of NP protein regulates influenza A virus RNA replication. EMBO J. 29: Lim, K. L., K. C. Chew, J. M. Tan, C. Wang, K. K. Chung, Y. Zhang, Y. Tanaka, W. Smith, S. Engelender, C. A. Ross, V. L. Dawson, and T. M. Dawson Parkin mediates nonclassical, proteasomal-independent ubiquitination of synphilin-1: implications for Lewy body formation. J. Neurosci. 25: Malakhova, O. A., and D.-E. Zhang ISG15 Inhibits Nedd4 Ubiquitin E3 Activity and Enhances the Innate Antiviral Response. J. Biol. Chem. 283: Marfori, M., A. Mynott, J. J. Ellis, A. M. Mehdi, N. F. Saunders, P. M. Curmi, J. K. Forwood, M. Boden, and B. Kobe Molecular basis for specificity of nuclear import and prediction of nuclear localization. Biochim. Biophys. Acta. 27. Martin-Serrano, J The role of ubiquitin in retroviral egress. Traffic 8: Morita, E., and W. I. Sundquist Retrovirus budding. Annu. Rev. Cell Dev. Biol. 20: Niwa, H., K. Yamamura, and J. Miyazaki Efficient selection for high-expression transfectants with a novel eukaryotic vector. Gene 108: Okumura, A., P. M. Pitha, and R. N. Harty ISG15 inhibits Ebola VP40 VLP budding in an L-domain-dependent manner by blocking Nedd4 ligase activity. Proc. Natl. Acad. Sci. USA 105: Olver, C., and M. Vidal Proteomic analysis of secreted exosomes. Subcell. Biochem. 43: Ott, D. E., L. V. Coren, E. N. Chertova, T. D. Gagliardi, and U. Schubert Ubiquitination of HIV-1 and MuLV Gag. Virology 278: Ott, D. E., L. V. Coren, T. D. Copeland, B. P. Kane, D. G. Johnson, R. C. Sowder, 2nd, Y. Yoshinaka, S. Oroszlan, L. O. Arthur, and L. E. Henderson Ubiquitin is covalently attached to the p6gag proteins of human immunodeficiency virus type 1 and simian immunodeficiency virus and to the p12gag protein of Moloney murine leukemia virus. J. Virol. 72: Ott, D. E., L. V. Coren, R. C. Sowder, 2nd, J. Adams, and U. Schubert Retroviruses have differing requirements for proteasome function in the budding process. J. Virol. 77: Paterson, R. G., and R. A. Lamb The molecular biology of influenza viruses and paramyxoviruses, p In A. Davidson and R. M. Elliot (ed.), Molecular Virology: A Practical Approach. IRL Oxford University Press, Oxford. 36. Patnaik, A., V. Chau, and J. W. Wills Ubiquitin is part of the retrovirus budding machinery. Proc. Natl. Acad. Sci. USA 97:
25 Pei, Z., Y. Bai, and A. P. Schmitt PIV5 M protein interaction with host protein angiomotin-like 1. Virology 397: Pei, Z., M. S. Harrison, and A. P. Schmitt Parainfluenza virus 5 M protein interaction with host protein negatively affects virus particle formation. J. Virol. 85: Peng, J., D. Schwartz, J. E. Elias, C. C. Thoreen, D. Cheng, G. Marsischky, J. Roelofs, D. Finley, and S. P. Gygi A proteomics approach to understanding protein ubiquitination. Nat. Biotechnol. 21: Pohl, C., W. P. Duprex, G. Krohne, B. K. Rima, and S. Schneider-Schaulies Measles virus M and F proteins associate with detergent-resistant membrane fractions and promote formation of virus-like particles. J. Gen. Virol. 88: Putterman, D., R. B. Pepinsky, and V. M. Vogt Ubiquitin in avian leukosis virus particles. Virology 176: Randall, R. E., D. F. Young, K. K. Goswami, and W. C. Russell Isolation and characterization of monoclonal antibodies to simian virus 5 and their use in revealing antigenic differences between human, canine and simian isolates. J. Gen. Virol. 68: Ravid, D., G. P. Leser, and R. A. Lamb A role for caveolin 1 in assembly and budding of the paramyxovirus parainfluenza virus 5. J. Virol. 84: Schmitt, A. P., B. He, and R. A. Lamb Involvement of the cytoplasmic domain of the hemagglutinin-neuraminidase protein in assembly of the paramyxovirus simian virus 5. J. Virol. 73: Schmitt, A. P., and R. A. Lamb Escaping from the Cell: Assembly and Budding of Negative-Strand RNA Viruses. Curr. Top. Microbiol. Immunol. 283: Schmitt, A. P., G. P. Leser, E. Morita, W. I. Sundquist, and R. A. Lamb Evidence for a new viral late domain core sequence, FPIV, necessary for budding of a paramyxovirus. J. Virol. 79: Schmitt, A. P., G. P. Leser, D. L. Waning, and R. A. Lamb Requirements for budding of paramyxovirus simian virus 5 virus-like particles. J. Virol. 76: Schubert, U., D. E. Ott, E. N. Chertova, R. Welker, U. Tessmer, M. F. Princiotta, J. R. Bennink, H. G. Krausslich, and J. W. Yewdell Proteasome inhibition interferes with gag polyprotein processing, release, and maturation of HIV-1 and HIV-2. Proc. Natl. Acad. Sci. USA 97: Spidel, J. L., R. C. Craven, C. B. Wilson, A. Patnaik, H. Wang, L. M. Mansky, and J. W. Wills Lysines close to the Rous sarcoma virus late domain critical for budding. J. Virol. 78: Strack, B., A. Calistri, M. A. Accola, G. Palu, and H. G. Göttlinger A role for ubiquitin ligase recruitment in retrovirus release. Proc. Natl. Acad. Sci. USA 97:
26 Takimoto, T., and A. Portner Molecular mechanism of paramyxovirus budding. Virus Res. 106: Timmins, J., G. Schoehn, S. Ricard-Blum, S. Scianimanico, T. Vernet, R. W. Ruigrok, and W. Weissenhorn Ebola virus matrix protein VP40 interaction with human cellular factors Tsg101 and Nedd4. J. Mol. Biol. 326: Treier, M., L. M. Staszewski, and D. Bohmann Ubiquitin-dependent c-jun degradation in vivo is mediated by the delta domain. Cell 78: Tung, C. W., and S. Y. Ho Computational identification of ubiquitylation sites from protein sequences. BMC Bioinformatics 9: Vogt, V. M Ubiquitin in retrovirus assembly: actor or bystander? Proc. Natl. Acad. Sci. USA 97: Wang, H., N. J. Machesky, and L. M. Mansky Both the PPPY and PTAP motifs are involved in human T-cell leukemia virus type 1 particle release. J. Virol. 78: Wang, Y. E., A. Park, M. Lake, M. Pentecost, B. Torres, T. E. Yun, M. C. Wolf, M. R. Holbrook, A. N. Freiberg, and B. Lee Ubiquitin-regulated nuclear-cytoplasmic trafficking of the nipah virus matrix protein is important for viral budding. PLoS Pathog. 6:e Waning, D. L., A. P. Schmitt, G. P. Leser, and R. A. Lamb Roles for the cytoplasmic tails of the fusion and hemagglutinin-neuraminidase proteins in budding of the paramyxovirus simian virus 5. J. Virol. 76: Watanabe, H., Y. Tanaka, Y. Shimazu, F. Sugahara, M. Kuwayama, A. Hiramatsu, K. Kiyotani, T. Yoshida, and T. Sakaguchi Cell-specific inhibition of paramyxovirus maturation by proteasome inhibitors. Microbiol. Immunol. 49: Yang, X. J., and E. Seto Lysine acetylation: codified crosstalk with other posttranslational modifications. Mol. Cell 31: Yasuda, J., M. Nakao, Y. Kawaoka, and H. Shida Nedd4 regulates egress of Ebola virus-like particles from host cells. J. Virol. 77: Zhadina, M., and P. D. Bieniasz Functional interchangeability of late domains, late domain cofactors and ubiquitin in viral budding. PLoS Pathog. 6:e Zhadina, M., M. O. McClure, M. C. Johnson, and P. D. Bieniasz Ubiquitin-dependent virus particle budding without viral protein ubiquitination. Proc. Natl. Acad. Sci. USA 104: FIGURE LEGENDS 26
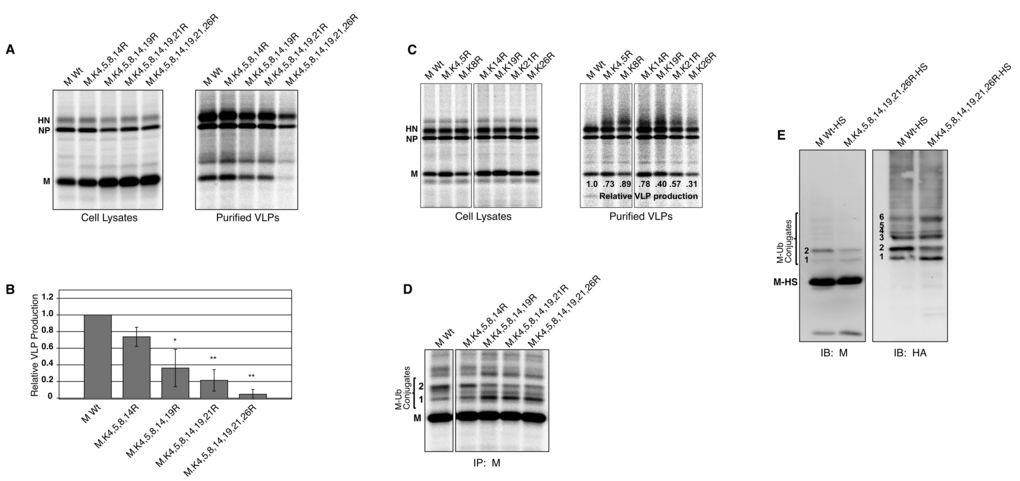
27 Figure 1. Detection of ubiquitinated PIV5 M protein. (A) 293T cells were transfected to express the affinity-tagged PIV5 M-HS protein together with HA-UB, as indicated. Cells were harvested, lysates prepared, and M-HS protein was pulled down using StrepTactin MacroPrep resin. Polypeptides were resolved on a 10% SDS gel and transferred to a membrane, which was then cut into two portions for immunoblotting with antibodies specific to either PIV5 M protein or HA epitope tag, as indicated. Six M-ubiquitin conjugates are numbered from the smallest to the largest apparent molecular weight. (B) 293T cells were transfected to express M-HS protein together with HA-UB, HA-UBWT, or HA-UBKO, as indicated. HA-UBKO lacks lysines necessary for polyubiquitin chain formation. Cell lysates were prepared, M-HS protein was pulled down using affinity resin, and immunoblotting was performed as in (A). (C) 293T cells were transfected to express either untagged PIV5 M protein or PIV5 M-HS together with HA-UB as indicated. After metabolic labeling of cells, M proteins were immunoprecipitated from cell lysates, resolved by SDS-PAGE, and detected using a phosphorimager. Two M protein derivatives having migration consistent with monoubiquitination are indicated. (D) 293T cells were transfected to express PIV5 M protein (200 ng plasmid DNA / dish) or PIV5 NP protein together with HA-UB, as indicated. After metabolic labeling of cells, M and NP proteins were collected from cell lysates by immunoprecipitation, resolved by SDS-PAGE, and detected using a phosphorimager. Two bands which likely correspond to monoubiquitinated M proteins are indicated. Figure 2. Detection of free ubiquitin in purified PIV5 virions. (A) MDBK cells were infected with PIV5 and the virions released into the supernatant were purified on a sucrose gradient. Purified virions were treated with subtilisin in the presence or absence of Triton X- 100, as indicated, and polypeptides were fractionated by SDS-PAGE. Part of the gel was stained with Coomassie blue to visualize viral proteins (left panel). The remainder of the gel was subjected to immunoblot analysis using antibody specific to ubiquitin (right panel). Histagged, purified ubiquitin was included as a size marker. (B) MDBK cells were either PIV5- infected or mock-infected, and the culture supernatants were subjected to sucrose gradient purification. Polypeptides were fractionated by SDS-PAGE, and free ubiquitin was detected by immunoblotting. 27
28 Figure 3. Affinity purification of PIV5 M protein and identification of ubiquitin acceptor sites by MS. (A) 293T cells were transfected to express M-HS together with HA-UB as indicated. Cell lysates were prepared, and M-HS protein was purified using a ÄKTAprime FPLC system equipped with a StrepTrap HP column. Eluted fractions were resolved on a 10% SDS gel, which was then divided into three parts for Sypro Orange total protein staining (left panel), immunoblotting with anti-m antibody (middle panel), and immunoblotting with anti-ha antibody (right panel). Asterisks denote the migration of a polypeptide that was excised from a Coomassie blue-stained portion of the gel for MS-based ubiquitination analysis. (B) Depictions of tandem MS spectra for three peptides of PIV5 M protein potentially harboring the di-glycine ubiquitination signature on lysine residues as indicated. The masses of the fragments identified in each spectrum are represented by b-type ions, where the charge is on the N-terminus of the fragment, and y-type ions, where the charge is on the C-terminus of the fragment. The mass differences can be used to deduce the amino acid sequence as it compares to that of PIV5 M protein. Each peptide contains sites that are known to be occupied by lysine residues, though instead of a Da mass difference associated with a lysine residue, the mass difference is ~ 242 Da (denoted by *), which is consistent with the di-glycine remnant of ubiquitin. Figure 4. Mutagenesis of MS-identified ubiquitin acceptor sites in PIV5 M protein alters VLP production function. (A) 293T cells were transfected to produce the indicated PIV5 M proteins together with PIV5 NP and HN proteins. Altered M proteins were named according to the lysine residues (numbered 1 to 33 from the N-terminal end) that have been changed to arginine. After metabolic labeling of cells, VLPs from culture supernatants were harvested and purified by sucrose gradient flotation. Viral proteins from cell lysates were collected by immunoprecipitation. Proteins were fractionated by SDS-PAGE and visualized using a phosphorimager. (B) Three independent experiments were performed as described for panel A, and relative VLP production was calculated as the amount of M protein detected in VLPs divided by the amount of M protein detected in the corresponding cell lysate fraction, normalized to the value obtained with wt M protein. Error bars indicate standard deviations. Differences from the values obtained with wt M protein were assessed for statistical significance using two-tailed Student's t test, with P values <0.01 denoted by **. (C) 293T 28
29 cells were transfected to express the indicated PIV5 M proteins together with HA-UB. After metabolic labeling of cells, M proteins were immunoprecipitated from cell lysates, resolved by SDS-PAGE, and detected using a phosphorimager. Two M protein derivatives having migration consistent with monoubiquitination are indicated. Figure 5. Cumulative mutagenesis of seven lysine residues in PIV5 M protein disrupts VLP production. (A) 293T cells were transfected to produce the indicated PIV5 M proteins with cumulative lysine to arginine mutations together with PIV5 NP and HN proteins, and VLP production was measured as described in the legend to Fig. 4. (B) Relative VLP production was calculated from three independent experiments performed as in panel A. Error bars indicate standard deviations. Differences from the value obtained with wt M protein were assessed for statistical significance using two-tailed Student s t test, with P values <0.05 denoted by * and P values <0.01 denoted by **. (C) 293T cells were transfected to produce the indicated PIV5 M proteins with single lysine to arginine mutations together with PIV5 NP and HN proteins, and VLP production was measured and quantified as described in the legend to Fig. 4. (D) 293T cells were transfected to express the indicated PIV5 M proteins with cumulative lysine to arginine mutations together with HA-UB. After metabolic labeling of cells, M proteins were immunoprecipitated from cell lysates, resolved by SDS-PAGE, and detected using a phosphorimager. Two M protein derivatives having migration consistent with monoubiquitination are indicated. (E) 293T cells were transfected to express HA-UB together with either wt M-HS protein or seven lysine mutant M-HS protein, as indicated. Cells were harvested, lysates prepared, and M protein was pulled down using StrepTactin MacroPrep resin. Polypeptides were resolved on 10% SDS gels for immunoblotting, using antibodies specific to the PIV5 M protein and the HA epitope tag, as indicated. Numbers indicate the migrations of M protein derivatives that are likely ubiquitin-modified. Figure 6. Recombinant PIV5 harboring seven-lysine-mutant M protein exhibits altered multiplication kinetics and impaired infectious virion production. (A) MDBK cells were infected with the indicated recombinant viruses at a MOI. of 0.01 PFU/cell. Culture media was harvested at the indicated times, and virus titers were measured by plaque assay on 29
30 BHK-21F cells. Values represent averages of three independent experiments, with error bars indicating standard deviations. (B) MDBK cells were infected with the indicated recombinant viruses at a MOI of 1.0 PFU/cell. After metabolic labeling of cells, virions from culture supernatants were harvested and purified by sucrose gradient flotation. Viral proteins from cell lysates were collected by immunoprecipitation. Proteins were fractionated by SDS-PAGE and visualized using a phosphorimager. (C) Relative virion production was calculated from three independent experiments performed as in panel B. Virion production was calculated as described for VLP production in the legend to Fig. 4, and normalized to the value obtained with wt rpiv5. (D) Virus titers, measured by plaque assay on BHK-21F cells, were determined from three independent experiments performed as in panel B. Titers were determined from small aliquots of media harvested from the MDBK cells, prior to virion purification. (E) PFU to particle ratios were determined from three independent experiments performed as in panel B. The ratio was calculated as the total number of PFU detected in the culture medium divided by the quantity of M protein detected in purified virions, normalized to the value obtained with wt rpiv5. For panels B-D, error bars indicate standard deviations. Differences from the values obtained with rpiv5 were assessed for statistical significance using two-tailed Student's t test, with P values <0.05 denoted by *. Figure 7. Redistribution of M and HN proteins in virus-infected cells. CV-1 cells grown on glass coverslips were infected with recombinant PIV5 at a MOI of 1.0 PFU/cell. 18 h p.i., cells were incubated with the indicated quantities of MG h p.i., cells were fixed with formaldehyde, and incubated with mab specific to PIV5 HN protein (IgG2a isotype). Cells were then fixed a second time, permeabilized with 0.1% saponin, and incubated with mab specific to PIV5 M protein (IgG3 isotype). Isotype-specific secondary antibodies were used (IgG2a-specific Alexa Fluor 594 and IgG3-specific Alexa Fluor 488). Hoechst dye was used to stain nuclei. Fluorescence was visualized with a Zeiss AxioImager M1 fluorescence microscope. Figure 8. Impairments in VLP production and M protein ubiquitination induced by proteasome inhibitor treatment. (A) 293T cells were transfected to express M-HS or M-HS K.4,5,8,14,19,21,26R, together with the PIV5 NP and HN proteins for VLP production. 30
31 Transfected cells were treated with the indicated amounts of MG-132 at 14 h p.t. Cell and media fractions were harvested at 34 h p.t. VLPs were purified from media fractions by centrifugation through sucrose cushions and flotation on sucrose gradients. Cell lysate and purified VLP fractions were analyzed by immunoblotting with antibodies specific to PIV5 M protein. Relative VLP production was calculated as the amount of M protein detected in VLPs divided by the amount of M protein detected in the corresponding cell lysate fraction, normalized to the value obtained with wt M protein in the absence of MG-132. (B) 293T cells were transfected to express M-HS or M-HS K.4,5,8,14,19,21,26R, together with HA-UBKO. Transfected cells were treated with the indicated amounts of MG-132 at 14 h p.t., and harvested at 34 h p.t. Lysates were prepared and M protein was pulled down using StrepTactin MacroPrep resin. Polypeptides were resolved on 10% SDS gels for immunoblotting, using antibodies specific to the PIV5 M protein and the HA epitope tag, as indicated. 31
32
33
34
35
36
37
38
39
HIV-1 Virus-like Particle Budding Assay Nathan H Vande Burgt, Luis J Cocka * and Paul Bates
 HIV-1 Virus-like Particle Budding Assay Nathan H Vande Burgt, Luis J Cocka * and Paul Bates Department of Microbiology, Perelman School of Medicine at the University of Pennsylvania, Philadelphia, USA
HIV-1 Virus-like Particle Budding Assay Nathan H Vande Burgt, Luis J Cocka * and Paul Bates Department of Microbiology, Perelman School of Medicine at the University of Pennsylvania, Philadelphia, USA
Recombinant Protein Expression Retroviral system
 Recombinant Protein Expression Retroviral system Viruses Contains genome DNA or RNA Genome encased in a protein coat or capsid. Some viruses have membrane covering protein coat enveloped virus Ø Essential
Recombinant Protein Expression Retroviral system Viruses Contains genome DNA or RNA Genome encased in a protein coat or capsid. Some viruses have membrane covering protein coat enveloped virus Ø Essential
Supplementary information. MARCH8 inhibits HIV-1 infection by reducing virion incorporation of envelope glycoproteins
 Supplementary information inhibits HIV-1 infection by reducing virion incorporation of envelope glycoproteins Takuya Tada, Yanzhao Zhang, Takayoshi Koyama, Minoru Tobiume, Yasuko Tsunetsugu-Yokota, Shoji
Supplementary information inhibits HIV-1 infection by reducing virion incorporation of envelope glycoproteins Takuya Tada, Yanzhao Zhang, Takayoshi Koyama, Minoru Tobiume, Yasuko Tsunetsugu-Yokota, Shoji
Structural vs. nonstructural proteins
 Why would you want to study proteins associated with viruses or virus infection? Receptors Mechanism of uncoating How is gene expression carried out, exclusively by viral enzymes? Gene expression phases?
Why would you want to study proteins associated with viruses or virus infection? Receptors Mechanism of uncoating How is gene expression carried out, exclusively by viral enzymes? Gene expression phases?
Shin-Hee Kim, Yongqi Yan, and Siba K. Samal*
 JOURNAL OF VIROLOGY, Oct. 2009, p. 10250 10255 Vol. 83, No. 19 0022-538X/09/$08.00 0 doi:10.1128/jvi.01038-09 Copyright 2009, American Society for Microbiology. All Rights Reserved. Role of the Cytoplasmic
JOURNAL OF VIROLOGY, Oct. 2009, p. 10250 10255 Vol. 83, No. 19 0022-538X/09/$08.00 0 doi:10.1128/jvi.01038-09 Copyright 2009, American Society for Microbiology. All Rights Reserved. Role of the Cytoplasmic
Fayth K. Yoshimura, Ph.D. September 7, of 7 RETROVIRUSES. 2. HTLV-II causes hairy T-cell leukemia
 1 of 7 I. Diseases Caused by Retroviruses RETROVIRUSES A. Human retroviruses that cause cancers 1. HTLV-I causes adult T-cell leukemia and tropical spastic paraparesis 2. HTLV-II causes hairy T-cell leukemia
1 of 7 I. Diseases Caused by Retroviruses RETROVIRUSES A. Human retroviruses that cause cancers 1. HTLV-I causes adult T-cell leukemia and tropical spastic paraparesis 2. HTLV-II causes hairy T-cell leukemia
Mumps Virus Matrix, Fusion, and Nucleocapsid Proteins Cooperate for Efficient Production of Virus-Like Particles
 JVI Accepts, published online ahead of print on 13 May 2009 J. Virol. doi:10.1128/jvi.00421-09 Copyright 2009, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.
JVI Accepts, published online ahead of print on 13 May 2009 J. Virol. doi:10.1128/jvi.00421-09 Copyright 2009, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.
Plasmid-Driven Formation of Influenza Virus-Like Particles
 JOURNAL OF VIROLOGY, Jan. 2000, p. 547 551 Vol. 74, No. 1 0022-538X/00/$04.00 0 Copyright 2000, American Society for Microbiology. All Rights Reserved. Plasmid-Driven Formation of Influenza Virus-Like
JOURNAL OF VIROLOGY, Jan. 2000, p. 547 551 Vol. 74, No. 1 0022-538X/00/$04.00 0 Copyright 2000, American Society for Microbiology. All Rights Reserved. Plasmid-Driven Formation of Influenza Virus-Like
SUPPLEMENTAL INFORMATION
 SUPPLEMENTAL INFORMATION EXPERIMENTAL PROCEDURES Tryptic digestion protection experiments - PCSK9 with Ab-3D5 (1:1 molar ratio) in 50 mm Tris, ph 8.0, 150 mm NaCl was incubated overnight at 4 o C. The
SUPPLEMENTAL INFORMATION EXPERIMENTAL PROCEDURES Tryptic digestion protection experiments - PCSK9 with Ab-3D5 (1:1 molar ratio) in 50 mm Tris, ph 8.0, 150 mm NaCl was incubated overnight at 4 o C. The
SUPPLEMENTARY INFORMATION
 Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
Received 3 September 2002/Accepted 15 January 2003
 JOURNAL OF VIROLOGY, Apr. 2003, p. 4646 4657 Vol. 77, No. 8 0022-538X/03/$08.00 0 DOI: 10.1128/JVI.77.8.4646 4657.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved. Ability of
JOURNAL OF VIROLOGY, Apr. 2003, p. 4646 4657 Vol. 77, No. 8 0022-538X/03/$08.00 0 DOI: 10.1128/JVI.77.8.4646 4657.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved. Ability of
Supplementary material: Materials and suppliers
 Supplementary material: Materials and suppliers Electrophoresis consumables including tris-glycine, acrylamide, SDS buffer and Coomassie Brilliant Blue G-2 dye (CBB) were purchased from Ameresco (Solon,
Supplementary material: Materials and suppliers Electrophoresis consumables including tris-glycine, acrylamide, SDS buffer and Coomassie Brilliant Blue G-2 dye (CBB) were purchased from Ameresco (Solon,
Influenza viruses. Virion. Genome. Genes and proteins. Viruses and hosts. Diseases. Distinctive characteristics
 Influenza viruses Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Virion Enveloped particles, quasi-spherical or filamentous Diameter 80-120 nm Envelope is derived
Influenza viruses Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Virion Enveloped particles, quasi-spherical or filamentous Diameter 80-120 nm Envelope is derived
Julianne Edwards. Retroviruses. Spring 2010
 Retroviruses Spring 2010 A retrovirus can simply be referred to as an infectious particle which replicates backwards even though there are many different types of retroviruses. More specifically, a retrovirus
Retroviruses Spring 2010 A retrovirus can simply be referred to as an infectious particle which replicates backwards even though there are many different types of retroviruses. More specifically, a retrovirus
Materials and Methods , The two-hybrid principle.
 The enzymatic activity of an unknown protein which cleaves the phosphodiester bond between the tyrosine residue of a viral protein and the 5 terminus of the picornavirus RNA Introduction Every day there
The enzymatic activity of an unknown protein which cleaves the phosphodiester bond between the tyrosine residue of a viral protein and the 5 terminus of the picornavirus RNA Introduction Every day there
Appendix. Table of Contents
 Appendix Table of Contents Appendix Figures Figure S1: Gp78 is not required for the degradation of mcherry-cl1 in Hela Cells. Figure S2: Indel formation in the MARCH6 sgrna targeted HeLa clones. Figure
Appendix Table of Contents Appendix Figures Figure S1: Gp78 is not required for the degradation of mcherry-cl1 in Hela Cells. Figure S2: Indel formation in the MARCH6 sgrna targeted HeLa clones. Figure
Supplementary Information
 Supplementary Information Supplementary Figure 1. CD4 + T cell activation and lack of apoptosis after crosslinking with anti-cd3 + anti-cd28 + anti-cd160. (a) Flow cytometry of anti-cd160 (5D.10A11) binding
Supplementary Information Supplementary Figure 1. CD4 + T cell activation and lack of apoptosis after crosslinking with anti-cd3 + anti-cd28 + anti-cd160. (a) Flow cytometry of anti-cd160 (5D.10A11) binding
MCB130 Midterm. GSI s Name:
 1. Peroxisomes are small, membrane-enclosed organelles that function in the degradation of fatty acids and in the degradation of H 2 O 2. Peroxisomes are not part of the secretory pathway and peroxisomal
1. Peroxisomes are small, membrane-enclosed organelles that function in the degradation of fatty acids and in the degradation of H 2 O 2. Peroxisomes are not part of the secretory pathway and peroxisomal
Section 6. Junaid Malek, M.D.
 Section 6 Junaid Malek, M.D. The Golgi and gp160 gp160 transported from ER to the Golgi in coated vesicles These coated vesicles fuse to the cis portion of the Golgi and deposit their cargo in the cisternae
Section 6 Junaid Malek, M.D. The Golgi and gp160 gp160 transported from ER to the Golgi in coated vesicles These coated vesicles fuse to the cis portion of the Golgi and deposit their cargo in the cisternae
Study of different types of ubiquitination
 Study of different types of ubiquitination Rudi Beyaert (rudi.beyaert@irc.vib-ugent.be) VIB UGent Center for Inflammation Research Ghent, Belgium VIB Training Novel Proteomics Tools: Identifying PTMs October
Study of different types of ubiquitination Rudi Beyaert (rudi.beyaert@irc.vib-ugent.be) VIB UGent Center for Inflammation Research Ghent, Belgium VIB Training Novel Proteomics Tools: Identifying PTMs October
7.012 Problem Set 6 Solutions
 Name Section 7.012 Problem Set 6 Solutions Question 1 The viral family Orthomyxoviridae contains the influenza A, B and C viruses. These viruses have a (-)ss RNA genome surrounded by a capsid composed
Name Section 7.012 Problem Set 6 Solutions Question 1 The viral family Orthomyxoviridae contains the influenza A, B and C viruses. These viruses have a (-)ss RNA genome surrounded by a capsid composed
Ubiquitination and deubiquitination of NP protein regulates influenza A virus RNA replication
 Manuscript EMBO-2010-74756 Ubiquitination and deubiquitination of NP protein regulates influenza A virus RNA replication Tsai-Ling Liao, Chung-Yi Wu, Wen-Chi Su, King-Song Jeng and Michael Lai Corresponding
Manuscript EMBO-2010-74756 Ubiquitination and deubiquitination of NP protein regulates influenza A virus RNA replication Tsai-Ling Liao, Chung-Yi Wu, Wen-Chi Su, King-Song Jeng and Michael Lai Corresponding
VIROLOGY. Engineering Viral Genomes: Retrovirus Vectors
 VIROLOGY Engineering Viral Genomes: Retrovirus Vectors Viral vectors Retrovirus replicative cycle Most mammalian retroviruses use trna PRO, trna Lys3, trna Lys1,2 The partially unfolded trna is annealed
VIROLOGY Engineering Viral Genomes: Retrovirus Vectors Viral vectors Retrovirus replicative cycle Most mammalian retroviruses use trna PRO, trna Lys3, trna Lys1,2 The partially unfolded trna is annealed
Sarah A. Kopecky and Douglas S. Lyles*
 JOURNAL OF VIROLOGY, Apr. 2003, p. 4658 4669 Vol. 77, No. 8 0022-538X/03/$08.00 0 DOI: 10.1128/JVI.77.8.4658 4669.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved. Contrasting
JOURNAL OF VIROLOGY, Apr. 2003, p. 4658 4669 Vol. 77, No. 8 0022-538X/03/$08.00 0 DOI: 10.1128/JVI.77.8.4658 4669.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved. Contrasting
Supplemental Figure 1 ELISA scheme to measure plasma total, mature and furin-cleaved
 1 Supplemental Figure Legends Supplemental Figure 1 ELISA scheme to measure plasma total, mature and furin-cleaved PCSK9 concentrations. 4 Plasma mature and furin-cleaved PCSK9s were measured by a sandwich
1 Supplemental Figure Legends Supplemental Figure 1 ELISA scheme to measure plasma total, mature and furin-cleaved PCSK9 concentrations. 4 Plasma mature and furin-cleaved PCSK9s were measured by a sandwich
A Single Amino Acid Residue Change in the P Protein of Parainfluenza Virus 5 Elevates Viral Gene Expression
 JOURNAL OF VIROLOGY, Sept. 2008, p. 9123 9133 Vol. 82, No. 18 0022-538X/08/$08.00 0 doi:10.1128/jvi.00289-08 Copyright 2008, American Society for Microbiology. All Rights Reserved. A Single Amino Acid
JOURNAL OF VIROLOGY, Sept. 2008, p. 9123 9133 Vol. 82, No. 18 0022-538X/08/$08.00 0 doi:10.1128/jvi.00289-08 Copyright 2008, American Society for Microbiology. All Rights Reserved. A Single Amino Acid
Ali Alabbadi. Bann. Bann. Dr. Belal
 31 Ali Alabbadi Bann Bann Dr. Belal Topics to be discussed in this sheet: Particles-to-PFU Single-step and multi-step growth cycles Multiplicity of infection (MOI) Physical measurements of virus particles
31 Ali Alabbadi Bann Bann Dr. Belal Topics to be discussed in this sheet: Particles-to-PFU Single-step and multi-step growth cycles Multiplicity of infection (MOI) Physical measurements of virus particles
~Lentivirus production~
 ~Lentivirus production~ May 30, 2008 RNAi core R&D group member Lentivirus Production Session Lentivirus!!! Is it health threatening to lab technician? What s so good about this RNAi library? How to produce
~Lentivirus production~ May 30, 2008 RNAi core R&D group member Lentivirus Production Session Lentivirus!!! Is it health threatening to lab technician? What s so good about this RNAi library? How to produce
Size nm m m
 1 Viral size and organization Size 20-250nm 0.000000002m-0.000000025m Virion structure Capsid Core Acellular obligate intracellular parasites Lack organelles, metabolic activities, and reproduction Replicated
1 Viral size and organization Size 20-250nm 0.000000002m-0.000000025m Virion structure Capsid Core Acellular obligate intracellular parasites Lack organelles, metabolic activities, and reproduction Replicated
Retroviruses. ---The name retrovirus comes from the enzyme, reverse transcriptase.
 Retroviruses ---The name retrovirus comes from the enzyme, reverse transcriptase. ---Reverse transcriptase (RT) converts the RNA genome present in the virus particle into DNA. ---RT discovered in 1970.
Retroviruses ---The name retrovirus comes from the enzyme, reverse transcriptase. ---Reverse transcriptase (RT) converts the RNA genome present in the virus particle into DNA. ---RT discovered in 1970.
Figure S1. Schematic presentation of genomic replication of idsiv after transfection and infection. After transfection of idsiv plasmid DNA into 293T
 Figure S1. Schematic presentation of genomic replication of idsiv after transfection and infection. After transfection of idsiv plasmid DNA into 293T cells, the RNA genomes with all modifications are generated
Figure S1. Schematic presentation of genomic replication of idsiv after transfection and infection. After transfection of idsiv plasmid DNA into 293T cells, the RNA genomes with all modifications are generated
Viral reproductive cycle
 Lecture 29: Viruses Lecture outline 11/11/05 Types of viruses Bacteriophage Lytic and lysogenic life cycles viruses viruses Influenza Prions Mad cow disease 0.5 µm Figure 18.4 Viral structure of capsid
Lecture 29: Viruses Lecture outline 11/11/05 Types of viruses Bacteriophage Lytic and lysogenic life cycles viruses viruses Influenza Prions Mad cow disease 0.5 µm Figure 18.4 Viral structure of capsid
VIRAL TITER COUNTS. The best methods of measuring infectious lentiviral titer
 VIRAL TITER COUNTS The best methods of measuring infectious lentiviral titer FLUORESCENCE CYCLES qpcr of Viral RNA SUMMARY Viral vectors are now routinely used for gene transduction in a wide variety of
VIRAL TITER COUNTS The best methods of measuring infectious lentiviral titer FLUORESCENCE CYCLES qpcr of Viral RNA SUMMARY Viral vectors are now routinely used for gene transduction in a wide variety of
19/06/2013. Viruses are not organisms (do not belong to any kingdom). Viruses are not made of cells, have no cytoplasm, and no membranes.
. Viruses are not made of cells, have no cytoplasm, and no membranes.") VIRUSES Many diseases of plants and animals are caused by bacteria or viruses that invade the body. Bacteria and viruses are NOT similar kinds of micro-organisms. Bacteria are classified as living organisms,
VIRUSES Many diseases of plants and animals are caused by bacteria or viruses that invade the body. Bacteria and viruses are NOT similar kinds of micro-organisms. Bacteria are classified as living organisms,
TFEB-mediated increase in peripheral lysosomes regulates. Store Operated Calcium Entry
 TFEB-mediated increase in peripheral lysosomes regulates Store Operated Calcium Entry Luigi Sbano, Massimo Bonora, Saverio Marchi, Federica Baldassari, Diego L. Medina, Andrea Ballabio, Carlotta Giorgi
TFEB-mediated increase in peripheral lysosomes regulates Store Operated Calcium Entry Luigi Sbano, Massimo Bonora, Saverio Marchi, Federica Baldassari, Diego L. Medina, Andrea Ballabio, Carlotta Giorgi
Fayth K. Yoshimura, Ph.D. September 7, of 7 HIV - BASIC PROPERTIES
 1 of 7 I. Viral Origin. A. Retrovirus - animal lentiviruses. HIV - BASIC PROPERTIES 1. HIV is a member of the Retrovirus family and more specifically it is a member of the Lentivirus genus of this family.
1 of 7 I. Viral Origin. A. Retrovirus - animal lentiviruses. HIV - BASIC PROPERTIES 1. HIV is a member of the Retrovirus family and more specifically it is a member of the Lentivirus genus of this family.
Tel: ; Fax: ;
 Tel.: +98 216 696 9291; Fax: +98 216 696 9291; E-mail: mrasadeghi@pasteur.ac.ir Tel: +98 916 113 7679; Fax: +98 613 333 6380; E-mail: abakhshi_e@ajums.ac.ir A Soluble Chromatin-bound MOI 0 1 5 0 1 5 HDAC2
Tel.: +98 216 696 9291; Fax: +98 216 696 9291; E-mail: mrasadeghi@pasteur.ac.ir Tel: +98 916 113 7679; Fax: +98 613 333 6380; E-mail: abakhshi_e@ajums.ac.ir A Soluble Chromatin-bound MOI 0 1 5 0 1 5 HDAC2
Viral structure م.م رنا مشعل
 Viral structure م.م رنا مشعل Viruses must reproduce (replicate) within cells, because they cannot generate energy or synthesize proteins. Because they can reproduce only within cells, viruses are obligate
Viral structure م.م رنا مشعل Viruses must reproduce (replicate) within cells, because they cannot generate energy or synthesize proteins. Because they can reproduce only within cells, viruses are obligate
Dr. Ahmed K. Ali Attachment and entry of viruses into cells
 Lec. 6 Dr. Ahmed K. Ali Attachment and entry of viruses into cells The aim of a virus is to replicate itself, and in order to achieve this aim it needs to enter a host cell, make copies of itself and
Lec. 6 Dr. Ahmed K. Ali Attachment and entry of viruses into cells The aim of a virus is to replicate itself, and in order to achieve this aim it needs to enter a host cell, make copies of itself and
Supplementary Figure 1. SC35M polymerase activity in the presence of Bat or SC35M NP encoded from the phw2000 rescue plasmid.
 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 Supplementary Figure 1. SC35M polymerase activity in the presence of Bat or SC35M NP encoded from the phw2000 rescue plasmid. HEK293T
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 Supplementary Figure 1. SC35M polymerase activity in the presence of Bat or SC35M NP encoded from the phw2000 rescue plasmid. HEK293T
Coronaviruses. Virion. Genome. Genes and proteins. Viruses and hosts. Diseases. Distinctive characteristics
 Coronaviruses Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Virion Spherical enveloped particles studded with clubbed spikes Diameter 120-160 nm Coiled helical
Coronaviruses Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Virion Spherical enveloped particles studded with clubbed spikes Diameter 120-160 nm Coiled helical
There are approximately 30,000 proteasomes in a typical human cell Each proteasome is approximately 700 kda in size The proteasome is made up of 3
 Proteasomes Proteasomes Proteasomes are responsible for degrading proteins that have been damaged, assembled improperly, or that are of no profitable use to the cell. The unwanted protein is literally
Proteasomes Proteasomes Proteasomes are responsible for degrading proteins that have been damaged, assembled improperly, or that are of no profitable use to the cell. The unwanted protein is literally
Supplementary Material
 Supplementary Material Nuclear import of purified HIV-1 Integrase. Integrase remains associated to the RTC throughout the infection process until provirus integration occurs and is therefore one likely
Supplementary Material Nuclear import of purified HIV-1 Integrase. Integrase remains associated to the RTC throughout the infection process until provirus integration occurs and is therefore one likely
Virus Entry/Uncoating
 Virus Entry/Uncoating Delivery of genome to inside of a cell Genome must be available for first step of replication The Problem--barriers to infection Virion Barriers: Non-enveloped viruses capsid Enveloped
Virus Entry/Uncoating Delivery of genome to inside of a cell Genome must be available for first step of replication The Problem--barriers to infection Virion Barriers: Non-enveloped viruses capsid Enveloped
Chapter 6- An Introduction to Viruses*
 Chapter 6- An Introduction to Viruses* *Lecture notes are to be used as a study guide only and do not represent the comprehensive information you will need to know for the exams. 6.1 Overview of Viruses
Chapter 6- An Introduction to Viruses* *Lecture notes are to be used as a study guide only and do not represent the comprehensive information you will need to know for the exams. 6.1 Overview of Viruses
TSH Receptor Monoclonal Antibody (49) Catalog Number MA3-218 Product data sheet
 Catalog Number MA3-218 Product data sheet") Website: thermofisher.com Customer Service (US): 1 800 955 6288 ext. 1 Technical Support (US): 1 800 955 6288 ext. 441 TSH Receptor Monoclonal Antibody (49) Catalog Number MA3-218 Product data sheet Details
Website: thermofisher.com Customer Service (US): 1 800 955 6288 ext. 1 Technical Support (US): 1 800 955 6288 ext. 441 TSH Receptor Monoclonal Antibody (49) Catalog Number MA3-218 Product data sheet Details
Name Section Problem Set 6
 Name Section 7.012 Problem Set 6 Question 1 The viral family Orthomyxoviridae contains the influenza A, B and C viruses. These viruses have a (-)ss RNA genome surrounded by a capsid composed of lipids
Name Section 7.012 Problem Set 6 Question 1 The viral family Orthomyxoviridae contains the influenza A, B and C viruses. These viruses have a (-)ss RNA genome surrounded by a capsid composed of lipids
Lecture 2: Virology. I. Background
 Lecture 2: Virology I. Background A. Properties 1. Simple biological systems a. Aggregates of nucleic acids and protein 2. Non-living a. Cannot reproduce or carry out metabolic activities outside of a
Lecture 2: Virology I. Background A. Properties 1. Simple biological systems a. Aggregates of nucleic acids and protein 2. Non-living a. Cannot reproduce or carry out metabolic activities outside of a
supplementary information
 Figure S1 Nucleotide binding status of RagA mutants. Wild type and mutant forms of MycRagA was transfected into HEK293 cells and the transfected cells were labeled with 32 Pphosphate. MycRagA was immunoprecipitated
Figure S1 Nucleotide binding status of RagA mutants. Wild type and mutant forms of MycRagA was transfected into HEK293 cells and the transfected cells were labeled with 32 Pphosphate. MycRagA was immunoprecipitated
Essential Medium, containing 10% fetal bovine serum, 100 U/ml penicillin and 100 µg/ml streptomycin. Huvec were cultured in
 Supplemental data Methods Cell culture media formulations A-431 and U-87 MG cells were maintained in Dulbecco s Modified Eagle s Medium. FaDu cells were cultured in Eagle's Minimum Essential Medium, containing
Supplemental data Methods Cell culture media formulations A-431 and U-87 MG cells were maintained in Dulbecco s Modified Eagle s Medium. FaDu cells were cultured in Eagle's Minimum Essential Medium, containing
Chapter 18. Viral Genetics. AP Biology
 Chapter 18. Viral Genetics 2003-2004 1 A sense of size Comparing eukaryote bacterium virus 2 What is a virus? Is it alive? DNA or RNA enclosed in a protein coat Viruses are not cells Extremely tiny electron
Chapter 18. Viral Genetics 2003-2004 1 A sense of size Comparing eukaryote bacterium virus 2 What is a virus? Is it alive? DNA or RNA enclosed in a protein coat Viruses are not cells Extremely tiny electron
7.012 Quiz 3 Answers
 MIT Biology Department 7.012: Introductory Biology - Fall 2004 Instructors: Professor Eric Lander, Professor Robert A. Weinberg, Dr. Claudette Gardel Friday 11/12/04 7.012 Quiz 3 Answers A > 85 B 72-84
MIT Biology Department 7.012: Introductory Biology - Fall 2004 Instructors: Professor Eric Lander, Professor Robert A. Weinberg, Dr. Claudette Gardel Friday 11/12/04 7.012 Quiz 3 Answers A > 85 B 72-84
NOTES. Matthew S. Walters, Christos A. Kyratsous, and Saul J. Silverstein*
 JOURNAL OF VIROLOGY, July 2010, p. 6861 6865 Vol. 84, No. 13 0022-538X/10/$12.00 doi:10.1128/jvi.00335-10 Copyright 2010, American Society for Microbiology. All Rights Reserved. NOTES The RING Finger Domain
JOURNAL OF VIROLOGY, July 2010, p. 6861 6865 Vol. 84, No. 13 0022-538X/10/$12.00 doi:10.1128/jvi.00335-10 Copyright 2010, American Society for Microbiology. All Rights Reserved. NOTES The RING Finger Domain
October 26, Lecture Readings. Vesicular Trafficking, Secretory Pathway, HIV Assembly and Exit from Cell
 October 26, 2006 Vesicular Trafficking, Secretory Pathway, HIV Assembly and Exit from Cell 1. Secretory pathway a. Formation of coated vesicles b. SNAREs and vesicle targeting 2. Membrane fusion a. SNAREs
October 26, 2006 Vesicular Trafficking, Secretory Pathway, HIV Assembly and Exit from Cell 1. Secretory pathway a. Formation of coated vesicles b. SNAREs and vesicle targeting 2. Membrane fusion a. SNAREs
Identification of Mutation(s) in. Associated with Neutralization Resistance. Miah Blomquist
 in. Associated with Neutralization Resistance. Miah Blomquist") Identification of Mutation(s) in the HIV 1 gp41 Subunit Associated with Neutralization Resistance Miah Blomquist What is HIV 1? HIV-1 is an epidemic that affects over 34 million people worldwide. HIV-1
Identification of Mutation(s) in the HIV 1 gp41 Subunit Associated with Neutralization Resistance Miah Blomquist What is HIV 1? HIV-1 is an epidemic that affects over 34 million people worldwide. HIV-1
Chapter 19: Viruses. 1. Viral Structure & Reproduction. 2. Bacteriophages. 3. Animal Viruses. 4. Viroids & Prions
 Chapter 19: Viruses 1. Viral Structure & Reproduction 2. Bacteriophages 3. Animal Viruses 4. Viroids & Prions 1. Viral Structure & Reproduction Chapter Reading pp. 393-396 What exactly is a Virus? Viruses
Chapter 19: Viruses 1. Viral Structure & Reproduction 2. Bacteriophages 3. Animal Viruses 4. Viroids & Prions 1. Viral Structure & Reproduction Chapter Reading pp. 393-396 What exactly is a Virus? Viruses
Schwarz et al. Activity-Dependent Ubiquitination of GluA1 Mediates a Distinct AMPAR Endocytosis
 Schwarz et al Activity-Dependent Ubiquitination of GluA1 Mediates a Distinct AMPAR Endocytosis and Sorting Pathway Supplemental Data Supplemental Fie 1: AMPARs undergo activity-mediated ubiquitination
Schwarz et al Activity-Dependent Ubiquitination of GluA1 Mediates a Distinct AMPAR Endocytosis and Sorting Pathway Supplemental Data Supplemental Fie 1: AMPARs undergo activity-mediated ubiquitination
Influenza A H1N1 HA ELISA Pair Set
 Influenza A H1N1 HA ELISA Pair Set for H1N1 ( A/Puerto Rico/8/1934 ) HA Catalog Number : SEK11684 To achieve the best assay results, this manual must be read carefully before using this product and the
Influenza A H1N1 HA ELISA Pair Set for H1N1 ( A/Puerto Rico/8/1934 ) HA Catalog Number : SEK11684 To achieve the best assay results, this manual must be read carefully before using this product and the
SUPPLEMENT. Materials and methods
 SUPPLEMENT Materials and methods Cell culture and reagents Cell media and reagents were from Invitrogen unless otherwise indicated. Antibiotics and Tet-certified serum were from Clontech. In experiments
SUPPLEMENT Materials and methods Cell culture and reagents Cell media and reagents were from Invitrogen unless otherwise indicated. Antibiotics and Tet-certified serum were from Clontech. In experiments
LESSON 4.4 WORKBOOK. How viruses make us sick: Viral Replication
 DEFINITIONS OF TERMS Eukaryotic: Non-bacterial cell type (bacteria are prokaryotes).. LESSON 4.4 WORKBOOK How viruses make us sick: Viral Replication This lesson extends the principles we learned in Unit
DEFINITIONS OF TERMS Eukaryotic: Non-bacterial cell type (bacteria are prokaryotes).. LESSON 4.4 WORKBOOK How viruses make us sick: Viral Replication This lesson extends the principles we learned in Unit
Chapter 19: Viruses. 1. Viral Structure & Reproduction. What exactly is a Virus? 11/7/ Viral Structure & Reproduction. 2.
 Chapter 19: Viruses 1. Viral Structure & Reproduction 2. Bacteriophages 3. Animal Viruses 4. Viroids & Prions 1. Viral Structure & Reproduction Chapter Reading pp. 393-396 What exactly is a Virus? Viruses
Chapter 19: Viruses 1. Viral Structure & Reproduction 2. Bacteriophages 3. Animal Viruses 4. Viroids & Prions 1. Viral Structure & Reproduction Chapter Reading pp. 393-396 What exactly is a Virus? Viruses
Improve Protein Analysis with the New, Mass Spectrometry- Compatible ProteasMAX Surfactant
 Improve Protein Analysis with the New, Mass Spectrometry- Compatible Surfactant ABSTRACT Incomplete solubilization and digestion and poor peptide recovery are frequent limitations in protein sample preparation
Improve Protein Analysis with the New, Mass Spectrometry- Compatible Surfactant ABSTRACT Incomplete solubilization and digestion and poor peptide recovery are frequent limitations in protein sample preparation
Protein Trafficking in the Secretory and Endocytic Pathways
 Protein Trafficking in the Secretory and Endocytic Pathways The compartmentalization of eukaryotic cells has considerable functional advantages for the cell, but requires elaborate mechanisms to ensure
Protein Trafficking in the Secretory and Endocytic Pathways The compartmentalization of eukaryotic cells has considerable functional advantages for the cell, but requires elaborate mechanisms to ensure
Overview of virus life cycle
 Overview of virus life cycle cell recognition and internalization release from cells progeny virus assembly membrane breaching nucleus capsid disassembly and genome release replication and translation
Overview of virus life cycle cell recognition and internalization release from cells progeny virus assembly membrane breaching nucleus capsid disassembly and genome release replication and translation
Viral Vectors In The Research Laboratory: Just How Safe Are They? Dawn P. Wooley, Ph.D., SM(NRM), RBP, CBSP
, RBP, CBSP") Viral Vectors In The Research Laboratory: Just How Safe Are They? Dawn P. Wooley, Ph.D., SM(NRM), RBP, CBSP 1 Learning Objectives Recognize hazards associated with viral vectors in research and animal
Viral Vectors In The Research Laboratory: Just How Safe Are They? Dawn P. Wooley, Ph.D., SM(NRM), RBP, CBSP 1 Learning Objectives Recognize hazards associated with viral vectors in research and animal
Encapsidation of Sendai Virus Genome RNAs by Purified
 JOURNAL OF VIROLOGY, Mar. 1988, p. 834-838 22-538X/88/3834-5$2./ Copyright C) 1988, American Society for Microbiology Vol. 62, No. 3 Encapsidation of Sendai Virus Genome RNAs by Purified NP Protein during
JOURNAL OF VIROLOGY, Mar. 1988, p. 834-838 22-538X/88/3834-5$2./ Copyright C) 1988, American Society for Microbiology Vol. 62, No. 3 Encapsidation of Sendai Virus Genome RNAs by Purified NP Protein during
Quantifying Lipid Contents in Enveloped Virus Particles with Plasmonic Nanoparticles
 Quantifying Lipid Contents in Enveloped Virus Particles with Plasmonic Nanoparticles Amin Feizpour Reinhard Lab Department of Chemistry and the Photonics Center, Boston University, Boston, MA May 2014
Quantifying Lipid Contents in Enveloped Virus Particles with Plasmonic Nanoparticles Amin Feizpour Reinhard Lab Department of Chemistry and the Photonics Center, Boston University, Boston, MA May 2014
Nature Medicine: doi: /nm.4322
 1 2 3 4 5 6 7 8 9 10 11 Supplementary Figure 1. Predicted RNA structure of 3 UTR and sequence alignment of deleted nucleotides. (a) Predicted RNA secondary structure of ZIKV 3 UTR. The stem-loop structure
1 2 3 4 5 6 7 8 9 10 11 Supplementary Figure 1. Predicted RNA structure of 3 UTR and sequence alignment of deleted nucleotides. (a) Predicted RNA secondary structure of ZIKV 3 UTR. The stem-loop structure
Lecture 11. Immunology and disease: parasite antigenic diversity
 Lecture 11 Immunology and disease: parasite antigenic diversity RNAi interference video and tutorial (you are responsible for this material, so check it out.) http://www.pbs.org/wgbh/nova/sciencenow/3210/02.html
Lecture 11 Immunology and disease: parasite antigenic diversity RNAi interference video and tutorial (you are responsible for this material, so check it out.) http://www.pbs.org/wgbh/nova/sciencenow/3210/02.html
Temperature-Sensitive Mutants Isolated from Hamster and
 JOURNAL OF VIROLOGY, Nov. 1975, p. 1332-1336 Copyright i 1975 American Society for Microbiology Vol. 16, No. 5 Printed in U.S.A. Temperature-Sensitive Mutants Isolated from Hamster and Canine Cell Lines
JOURNAL OF VIROLOGY, Nov. 1975, p. 1332-1336 Copyright i 1975 American Society for Microbiology Vol. 16, No. 5 Printed in U.S.A. Temperature-Sensitive Mutants Isolated from Hamster and Canine Cell Lines
supplementary information
 DOI: 10.1038/ncb2153 Figure S1 Ectopic expression of HAUSP up-regulates REST protein. (a) Immunoblotting showed that ectopic expression of HAUSP increased REST protein levels in ENStemA NPCs. (b) Immunofluorescent
DOI: 10.1038/ncb2153 Figure S1 Ectopic expression of HAUSP up-regulates REST protein. (a) Immunoblotting showed that ectopic expression of HAUSP increased REST protein levels in ENStemA NPCs. (b) Immunofluorescent
Supplementary materials
 Supplementary materials Chemical library from ChemBridge 50,240 structurally diverse small molecule compounds dissolved in DMSO Hits Controls: No virus added μ Primary screening at 20 g/ml of compounds
Supplementary materials Chemical library from ChemBridge 50,240 structurally diverse small molecule compounds dissolved in DMSO Hits Controls: No virus added μ Primary screening at 20 g/ml of compounds
Last time we talked about the few steps in viral replication cycle and the un-coating stage:
 Zeina Al-Momani Last time we talked about the few steps in viral replication cycle and the un-coating stage: Un-coating: is a general term for the events which occur after penetration, we talked about
Zeina Al-Momani Last time we talked about the few steps in viral replication cycle and the un-coating stage: Un-coating: is a general term for the events which occur after penetration, we talked about
AP Biology. Viral diseases Polio. Chapter 18. Smallpox. Influenza: 1918 epidemic. Emerging viruses. A sense of size
 Hepatitis Viral diseases Polio Chapter 18. Measles Viral Genetics Influenza: 1918 epidemic 30-40 million deaths world-wide Chicken pox Smallpox Eradicated in 1976 vaccinations ceased in 1980 at risk population?
Hepatitis Viral diseases Polio Chapter 18. Measles Viral Genetics Influenza: 1918 epidemic 30-40 million deaths world-wide Chicken pox Smallpox Eradicated in 1976 vaccinations ceased in 1980 at risk population?
Supplementary data Supplementary Figure 1 Supplementary Figure 2
 Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
Lecture Readings. Vesicular Trafficking, Secretory Pathway, HIV Assembly and Exit from Cell
 October 26, 2006 1 Vesicular Trafficking, Secretory Pathway, HIV Assembly and Exit from Cell 1. Secretory pathway a. Formation of coated vesicles b. SNAREs and vesicle targeting 2. Membrane fusion a. SNAREs
October 26, 2006 1 Vesicular Trafficking, Secretory Pathway, HIV Assembly and Exit from Cell 1. Secretory pathway a. Formation of coated vesicles b. SNAREs and vesicle targeting 2. Membrane fusion a. SNAREs
VIRUSES. 1. Describe the structure of a virus by completing the following chart.
 AP BIOLOGY MOLECULAR GENETICS ACTIVITY #3 NAME DATE HOUR VIRUSES 1. Describe the structure of a virus by completing the following chart. Viral Part Description of Part 2. Some viruses have an envelope
AP BIOLOGY MOLECULAR GENETICS ACTIVITY #3 NAME DATE HOUR VIRUSES 1. Describe the structure of a virus by completing the following chart. Viral Part Description of Part 2. Some viruses have an envelope
04_polarity. The formation of synaptic vesicles
 Brefeldin prevents assembly of the coats required for budding Nocodazole disrupts microtubules Constitutive: coatomer-coated Selected: clathrin-coated The formation of synaptic vesicles Nerve cells (and
Brefeldin prevents assembly of the coats required for budding Nocodazole disrupts microtubules Constitutive: coatomer-coated Selected: clathrin-coated The formation of synaptic vesicles Nerve cells (and
p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO
 Supplementary Information p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO Yuri Shibata, Masaaki Oyama, Hiroko Kozuka-Hata, Xiao Han, Yuetsu Tanaka,
Supplementary Information p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO Yuri Shibata, Masaaki Oyama, Hiroko Kozuka-Hata, Xiao Han, Yuetsu Tanaka,
Application of μmacs Streptavidin MicroBeads for the analysis of HIV-1 directly from patient plasma
 Excerpt from MACS&more Vol 8 1/2004 Application of μmacs Streptavidin MicroBeads for the analysis of HIV-1 directly from patient plasma L. Davis Lupo and Salvatore T. Butera HIV and Retrovirology Branch,
Excerpt from MACS&more Vol 8 1/2004 Application of μmacs Streptavidin MicroBeads for the analysis of HIV-1 directly from patient plasma L. Davis Lupo and Salvatore T. Butera HIV and Retrovirology Branch,
Hepatitis B Antiviral Drug Development Multi-Marker Screening Assay
 Hepatitis B Antiviral Drug Development Multi-Marker Screening Assay Background ImQuest BioSciences has developed and qualified a single-plate method to expedite the screening of antiviral agents against
Hepatitis B Antiviral Drug Development Multi-Marker Screening Assay Background ImQuest BioSciences has developed and qualified a single-plate method to expedite the screening of antiviral agents against
The Interferon-Induced Gene ISG15 Blocks Retrovirus Release from Cells Late in the Budding Process
 JOURNAL OF VIROLOGY, May 2010, p. 4725 4736 Vol. 84, No. 9 0022-538X/10/$12.00 doi:10.1128/jvi.02478-09 Copyright 2010, American Society for Microbiology. All Rights Reserved. The Interferon-Induced Gene
JOURNAL OF VIROLOGY, May 2010, p. 4725 4736 Vol. 84, No. 9 0022-538X/10/$12.00 doi:10.1128/jvi.02478-09 Copyright 2010, American Society for Microbiology. All Rights Reserved. The Interferon-Induced Gene
19 Viruses BIOLOGY. Outline. Structural Features and Characteristics. The Good the Bad and the Ugly. Structural Features and Characteristics
 9 Viruses CAMPBELL BIOLOGY TENTH EDITION Reece Urry Cain Wasserman Minorsky Jackson Outline I. Viruses A. Structure of viruses B. Common Characteristics of Viruses C. Viral replication D. HIV Lecture Presentation
9 Viruses CAMPBELL BIOLOGY TENTH EDITION Reece Urry Cain Wasserman Minorsky Jackson Outline I. Viruses A. Structure of viruses B. Common Characteristics of Viruses C. Viral replication D. HIV Lecture Presentation
Supplementary Figure 1 Role of Raf-1 in TLR2-Dectin-1-mediated cytokine expression
 Supplementary Figure 1 Supplementary Figure 1 Role of Raf-1 in TLR2-Dectin-1-mediated cytokine expression. Quantitative real-time PCR of indicated mrnas in DCs stimulated with TLR2-Dectin-1 agonist zymosan
Supplementary Figure 1 Supplementary Figure 1 Role of Raf-1 in TLR2-Dectin-1-mediated cytokine expression. Quantitative real-time PCR of indicated mrnas in DCs stimulated with TLR2-Dectin-1 agonist zymosan
Supplementary Figure 1. Prevalence of U539C and G540A nucleotide and E172K amino acid substitutions among H9N2 viruses. Full-length H9N2 NS
 Supplementary Figure 1. Prevalence of U539C and G540A nucleotide and E172K amino acid substitutions among H9N2 viruses. Full-length H9N2 NS nucleotide sequences (a, b) or amino acid sequences (c) from
Supplementary Figure 1. Prevalence of U539C and G540A nucleotide and E172K amino acid substitutions among H9N2 viruses. Full-length H9N2 NS nucleotide sequences (a, b) or amino acid sequences (c) from
Late Domain-Independent Rescue of a Release-Deficient Moloney Murine Leukemia Virus by the Ubiquitin Ligase Itch
 JOURNAL OF VIROLOGY, Jan. 2010, p. 704 715 Vol. 84, No. 2 0022-538X/10/$12.00 doi:10.1128/jvi.01319-09 Late Domain-Independent Rescue of a Release-Deficient Moloney Murine Leukemia Virus by the Ubiquitin
JOURNAL OF VIROLOGY, Jan. 2010, p. 704 715 Vol. 84, No. 2 0022-538X/10/$12.00 doi:10.1128/jvi.01319-09 Late Domain-Independent Rescue of a Release-Deficient Moloney Murine Leukemia Virus by the Ubiquitin
Role of ESCRT-I in Retroviral Budding
 JOURNAL OF VIROLOGY, Apr. 2003, p. 4794 4804 Vol. 77, No. 8 0022-538X/03/$08.00 0 DOI: 10.1128/JVI.77.8.4794 4804.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved. Role of ESCRT-I
JOURNAL OF VIROLOGY, Apr. 2003, p. 4794 4804 Vol. 77, No. 8 0022-538X/03/$08.00 0 DOI: 10.1128/JVI.77.8.4794 4804.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved. Role of ESCRT-I
Influenza or flu is a
 Clinical and Research Area Infectious Diseases Influenza Virus Types A and B Influenza or flu is a respiratory illness that is caused by influenza viruses. Influenza viruses type A and type B cause seasonal
Clinical and Research Area Infectious Diseases Influenza Virus Types A and B Influenza or flu is a respiratory illness that is caused by influenza viruses. Influenza viruses type A and type B cause seasonal
STRUCTURE, GENERAL CHARACTERISTICS AND REPRODUCTION OF VIRUSES
 STRUCTURE, GENERAL CHARACTERISTICS AND REPRODUCTION OF VIRUSES Introduction Viruses are noncellular genetic elements that use a living cell for their replication and have an extracellular state. Viruses
STRUCTURE, GENERAL CHARACTERISTICS AND REPRODUCTION OF VIRUSES Introduction Viruses are noncellular genetic elements that use a living cell for their replication and have an extracellular state. Viruses
SUPPLEMENTARY INFORMATION
 sirna pool: Control Tetherin -HA-GFP HA-Tetherin -Tubulin Supplementary Figure S1. Knockdown of HA-tagged tetherin expression by tetherin specific sirnas. HeLa cells were cotransfected with plasmids expressing
sirna pool: Control Tetherin -HA-GFP HA-Tetherin -Tubulin Supplementary Figure S1. Knockdown of HA-tagged tetherin expression by tetherin specific sirnas. HeLa cells were cotransfected with plasmids expressing
Influenza virus exploits tunneling nanotubes for cell-to-cell spread
 Supplementary Information Influenza virus exploits tunneling nanotubes for cell-to-cell spread Amrita Kumar 1, Jin Hyang Kim 1, Priya Ranjan 1, Maureen G. Metcalfe 2, Weiping Cao 1, Margarita Mishina 1,
Supplementary Information Influenza virus exploits tunneling nanotubes for cell-to-cell spread Amrita Kumar 1, Jin Hyang Kim 1, Priya Ranjan 1, Maureen G. Metcalfe 2, Weiping Cao 1, Margarita Mishina 1,
2) What is the difference between a non-enveloped virion and an enveloped virion? (4 pts)
 What is the difference between a non-enveloped virion and an enveloped virion? (4 pts)") Micro 260 SFCC Spring 2010 Name: All diagrams and drawings shall be hand drawn (do not photo-copied from a publication then cut and pasted into work sheet). Do not copy other student s answers. Para phase
Micro 260 SFCC Spring 2010 Name: All diagrams and drawings shall be hand drawn (do not photo-copied from a publication then cut and pasted into work sheet). Do not copy other student s answers. Para phase
Paramyxovirus Glycoprotein Incorporation, Assembly and Budding: A Three Way Dance for Infectious Particle Production
 University of Kentucky UKnowledge Molecular and Cellular Biochemistry Faculty Publications Molecular and Cellular Biochemistry 8-2014 Paramyxovirus Glycoprotein Incorporation, Assembly and Budding: A Three
University of Kentucky UKnowledge Molecular and Cellular Biochemistry Faculty Publications Molecular and Cellular Biochemistry 8-2014 Paramyxovirus Glycoprotein Incorporation, Assembly and Budding: A Three
SUPPLEMENTARY INFORMATION. Supplementary Figures S1-S9. Supplementary Methods
 SUPPLEMENTARY INFORMATION SUMO1 modification of PTEN regulates tumorigenesis by controlling its association with the plasma membrane Jian Huang 1,2#, Jie Yan 1,2#, Jian Zhang 3#, Shiguo Zhu 1, Yanli Wang
SUPPLEMENTARY INFORMATION SUMO1 modification of PTEN regulates tumorigenesis by controlling its association with the plasma membrane Jian Huang 1,2#, Jie Yan 1,2#, Jian Zhang 3#, Shiguo Zhu 1, Yanli Wang
A mechanism for glycoconjugate vaccine activation of the adaptive immune system and its implications for vaccine design
 A mechanism for glycoconjugate vaccine activation of the adaptive immune system and its implications for vaccine design Fikri Y. Avci 1,2, Xiangming Li 3, Moriya Tsuji 3, Dennis L. Kasper 1,2* Supplementary
A mechanism for glycoconjugate vaccine activation of the adaptive immune system and its implications for vaccine design Fikri Y. Avci 1,2, Xiangming Li 3, Moriya Tsuji 3, Dennis L. Kasper 1,2* Supplementary
PTM Discovery Method for Automated Identification and Sequencing of Phosphopeptides Using the Q TRAP LC/MS/MS System
 Application Note LC/MS PTM Discovery Method for Automated Identification and Sequencing of Phosphopeptides Using the Q TRAP LC/MS/MS System Purpose This application note describes an automated workflow
Application Note LC/MS PTM Discovery Method for Automated Identification and Sequencing of Phosphopeptides Using the Q TRAP LC/MS/MS System Purpose This application note describes an automated workflow
Relative activity (%) SC35M
 SC35M") a 125 Bat (H17N) b 125 A/WSN (H1N1) Relative activity (%) 0 75 50 25 Relative activity (%) 0 75 50 25 0 Pos. Neg. PA PB1 Pos. Neg. NP PA PB1 PB2 0 Pos. Neg. NP PA PB1 PB2 SC35M Bat Supplementary Figure
a 125 Bat (H17N) b 125 A/WSN (H1N1) Relative activity (%) 0 75 50 25 Relative activity (%) 0 75 50 25 0 Pos. Neg. PA PB1 Pos. Neg. NP PA PB1 PB2 0 Pos. Neg. NP PA PB1 PB2 SC35M Bat Supplementary Figure
LESSON 1.4 WORKBOOK. Viral sizes and structures. Workbook Lesson 1.4
 Eukaryotes organisms that contain a membrane bound nucleus and organelles. Prokaryotes organisms that lack a nucleus or other membrane-bound organelles. Viruses small, non-cellular (lacking a cell), infectious
Eukaryotes organisms that contain a membrane bound nucleus and organelles. Prokaryotes organisms that lack a nucleus or other membrane-bound organelles. Viruses small, non-cellular (lacking a cell), infectious
Gladstone Institutes, University of California (UCSF), San Francisco, USA
, San Francisco, USA") Fluorescence-linked Antigen Quantification (FLAQ) Assay for Fast Quantification of HIV-1 p24 Gag Marianne Gesner, Mekhala Maiti, Robert Grant and Marielle Cavrois * Gladstone Institutes, University of
Fluorescence-linked Antigen Quantification (FLAQ) Assay for Fast Quantification of HIV-1 p24 Gag Marianne Gesner, Mekhala Maiti, Robert Grant and Marielle Cavrois * Gladstone Institutes, University of