CD8 T cell dependent and independent immunity against Plasmodium following vaccination
|
|
|
- Holly Floyd
- 6 years ago
- Views:
Transcription

 thesis, University of Iowa, 2016. http://ir.uiowa.edu/etd/3073. Follow this and additional works at: http://ir.uiowa.edu/etd Part of the Microbiology Commons")
1 University of Iowa Iowa Research Online Theses and Dissertations Spring 2016 CD8 T cell dependent and independent immunity against Plasmodium following vaccination Katherine Lee Doll Kanne University of Iowa Copyright 2016 Katherine L. Doll Kanne This dissertation is available at Iowa Research Online: Recommended Citation Doll Kanne, Katherine Lee. "CD8 T cell dependent and independent immunity against Plasmodium following vaccination." PhD (Doctor of Philosophy) thesis, University of Iowa, Follow this and additional works at: Part of the Microbiology Commons
2 CD8 T CELL DEPENDENT AND INDPENDENT IMMUNITY AGAINST PLASMODIUM FOLLOWING VACCINATION by Katherine Lee Doll Kanne A thesis submitted in partial fulfillment of the requirements for the Doctor of Philosophy degree in Microbiology in the Graduate College of The University of Iowa May 2016 Thesis Supervisor: Professor John T. Harty
3 Graduate College The University of Iowa Iowa City, Iowa CERTIFICATE OF APPROVAL This is to certify that the Ph.D. thesis of PH.D. THESIS Katherine L. Doll Kanne has been approved by the Examining Committee for the thesis requirement for the Doctor of Philosophy degree in Microbiology at the May 2016 graduation. Thesis Committee: John T. Harty, Thesis Supervisor Stanley Perlman Kevin L. Legge Hai-Hui Xue Katherine N. Gibson-Corley
4 To my family and friends, for their endless encouragement and support. ii
5 ACKNOWLEDGEMENTS I would like to thank Dr. John T. Harty for his guidance and mentorship. I have appreciated your infinite depths of scientific knowledge. Thank you for encouraging me to learn new things, accept new challenges, and to build the confidence to come to my own conclusions. Your passion for science is evident in your body of work, and in your ability to be a great mentor to many graduate students and post-doctoral scholars. I look forward to carrying on your scientific legency in my future endevours. I would also like to thank my committee members, Drs. Stanley Perlman, Kevin Legge, Hai-Hui Xue, and Katherine Gibson-Corley for your time, input, and guidance over the years. Thank you to the great scientists I have worked and trained alongside in the Harty laboratory during my graduate career. We have had many great scientific discussions and heated debates that have helped me think critically about the literature. My experience in the laboratory would not been as enjoyable without the many Harty lab members: Drs. Noah Butler, Nathan Schmidt, Jeffrey Nolz, Martin Richer, Bram Slutter, Natalija Budimir, Scott Anthony, Stina Urban, Gabe Starbeck-Miller, and Marie Kim. Also thanks to Lecia (Pewe) Epping, Lisa Hancox and Cathryn Varga for your assistance throughout the years. A special thank you to Allison Christiaansen of the Varga Laboratory for being helpful through the ups and downs of our graduate school years. I greatly appreciated all of your help. I would like to thank my family and friends. My parents, Richard and Diane Doll: To my father for his unending love and support, and to my mother who I am sure would iii
6 be proud of my achievements. My sisters, Jacinda and Natalie: for your positivity and laughter. To my extended family and friends, thank you for your encouragement and support. And lastly, but importantly, to my husband Bryon: for your love, support and understanding during the challenging graduate school journey. iv
7 ABSTRACT Infection with Plasmodium species leads to nearly 400,000 deaths a year despite widespread use of mosquito bed nets, insecticides, and anti-malarial drugs. To date, there is not a licensed vaccine capable of providing complete protection from Plasmodium infection to vaccinees. Whole parasite vaccination of humans and rodents can achieve complete protection in vaccinees, but the dose of sporozoites, number of administrations, and production concerns in generating these types of vaccines will likely prevent these approaches from achieving worldwide use. However, the protective immunological responses against Plasmodium parasites engendered by these vaccination approaches can be studied and aid in the development of advanced subunit vaccines against Plasmodium. Using rodent models of malaria to elucidate the features of protective immunity engendered by whole parasite vaccination, it has been repeatedly shown that CD8 T cell responses directed against liver-stage parasite antigens can provide complete protection with some contribution by CD4 T cells and antibody responses depending on the model system studied. However, the quantatitive and qualitative requirements for CD8 T cell immunity against Plasmodium remain largely undefined. To enhance our understanding of how to generate protective immunity against Plasmodium, I have utilized rodent models of malaria to study the superior protection afforded from single-dose vaccination with virulent sporozoites administered under prophylatic chloroquine-cover, referred to as chemoprophylaxis and sporozoites (CPS) vaccination, compared to the well-studied approach of administering radiation-attenuated Plasmodium sporozoites (RAS). RAS vaccination has long been considered the gold v
8 standard in vaccination due the ability of RAS vaccination to engender complete protection following sporozoite challenge of vaccinated humans and rodents. However, CPS vaccination is arguably a superior vaccination approach since it can achieve protection through less vaccine administrations relative to RAS vaccination, but the immunological basis of this enhanced CPS vaccine-induced immune response was unclear. In my study, I utilized a stringent host/parasite model to find that C57Bl/6 mice administered CPS vaccination with P. yoelii sporozoites elicit substantially higher parasite-specific CD8 T cell responses than RAS vaccination, but CPS-induced CD8 T cells were not necessary for protection following liver-stage sporozoite or blood-stage parasite challenge. CPS vaccination resulted in a low grade, transient parasitemia shortly following cessation of chloroquine treatment, which led to the generation of potent antibody responses to blood-stage parasites; this blood-stage parasite-specific antibody response correlated with sterilizing protection in sporozoite and blood-stage challenged CPS-vaccinated mice. Therefore, my data provide a mechanistic basis for enhanced protective immunity elicited by single-dose CPS vaccination in a rodent model that is independent of CD8 T cells. The other portion of my work examines how CD8 T cell specificity impacts protective capacity against Plasmodium. I show that robust CD8 T cell responses of similar phenotype are mounted following prime-boost immunization against three novel Plasmodium berghei protein-derived epitopes in addition to a previously described protective, immunodominant epitope. I show that only CD8 T cells specific to sporozoite surface-expressed protein-derived epitopes, but not the intracellular protein-derived epitopes, are efficiently recognized by sporozoite-infected hepatocytes in vi
9 vitro and protective sterilizing immunity in vivo. These results suggest that antigenic targets must be efficiently presented by infected hepatocytes for CD8 T cells to eliminate liver-stage Plasmodium infection and proteins expressed on the surface of sporozoites may be good target antigens for protective CD8 T cells. Collectively, my work highlights the ability to generate protective immunity against Plasmodium infections that is CD8 T cell dependent or independent, whether achieved through potent blood-stage-specific antibody responses, or via numerically large monospecific CD8 T cell responses that target parasite antigens that are efficiently presented during liver-stage infection. These studies are relevant in understanding how to efficiently engender protective immunity against Plasmodium, and could aid in the advancement of subunit vaccination approaches that generate immunity through the priming of responses from multiple arms of the immune response, targeting both the liver- and blood-stages of Plasmodium. vii
10 PUBLIC ABSTRACT Plasmodium species are the causative agent of malaria, a disease that leads to approximately 400,000 deaths a year. Infection prevention methods have drastically reduced the mortality rate. However, these prevention methods are not enough to lead to complete control and thus vaccination remains the best strategy to eliminate malarial disease, and lead to worldwide eradication of the Plasmodium parasite. Currently, no licensed vaccine is available that provides complete protection in vaccinated individuals. In order to develop an effective vaccine, a better understanding of what constitutes a protective anti-plasmodium immune response is essential. To this end, rodent models of malaria have been utilized to mechanistically define the requirements of protective immunity. Previous work has shown that whole parasite vaccination protects individuals due to a cell population that specializes in killing parasite-infected cells, called the CD8 T cell. However, using a stringent rodent model of whole parasite vaccination, I found that CD8 T cells have no measurable role in protection, and in fact protection was likely due to antibody responses against the parasite. Another part of my study sought to better define an important feature of CD8 T cells, termed their specificity. I found in my examination of four specificities that only CD8 T cells that target surface-expresed parasite antigens were protective. Collectively, my studies have helped further our understanding of how to generate protective immune responses against the Plasmodium parasite and may aid in the development of an advanced vaccine that can protect humans against Plasmodium infections, and thus malarial disease. viii
11 TABLE OF CONTENTS LIST OF TABLES... xiii LIST OF FIGURES... xiv LIST OF ABBREVIATIONS... xvii CHAPTER I: INTRODUCTION...1 Natural and unnatural immunity against Plasmodium...1 Global impact and lifecycle of Plasmodium infections...1 Naturally-acquired immunity against Plasmodium does not develop...5 Generation of protective cellular immunity by whole sporozoite vaccination...6 CD8 T cell responses...9 Radiation attenuated sporozoites...9 Genetically attenuated parasites...11 Chemoprophylaxis and sporozoites...13 CD4 T cell responses...14 Antibody responses...16 Subunit vaccination approaches against Plasmodium have limited efficacy...19 Quantity and Quality of the CD8 T cell response against Plasmodium...23 Quantity of the CD8 T cell response to engender protection...23 Qualitative features dicating protection against Plasmodium...25 Rationale and objectives for current studies...27 Thesis objectives...28 CHAPTER II: CD8 T CELL INDEPENDENT IMMUNITY AGAINST SINGLE DOSE CHEMOPROPHLYAXIS AND SPOROZOITE IMMUNIZATION AGAINST PLASMODIUM YOELII...33 Abstract...33 Introduction...34 Materials and Methods...37 Mice and immunizations...37 Plasmodium parasites...38 ix
12 Detection of parasite-specific CD8 and CD4 T cells in blood...39 Listeria monocytogenes infections...39 Adoptive transfer of Thy1.1/1.2 + CS 280 TCR transgenic CD8 T cells...40 T cell depletion...40 Blood-stage reactive and anti-msp1 antibody detection...41 Liver parasite burden...41 Detection of persisting parasites under CQ treatment...42 Statistical Analysis...42 Results...43 CPS is associated with significantly larger anti-plasmodial CD8 and CD4 T cell responses and enhanced protection from challenge compared to RAS vaccination...43 CPS vaccination induces CD8 T cells against a broader spectrum of liver-stage antigens compared to RAS vaccination...47 CPS-induced CD8 T cells are not required for protection from sporozoite challenge...49 CPS-induced protection is associated with transient blood-stage parasite exposure...50 CPS-induced protection is associated with induction of blood-stage reactive antibodies...52 CPS confers protection against blood-stage challenge...55 Discussion...55 CHAPTER III: DISCRIMINATING PROTECTIVE FROM NON-PROTECTIVE PLASMODIUM-SPECIFIC CD8 T CELL RESPONSES...86 Abstract...86 Introduction...87 Material and Methods...91 Mouse strains...91 Generation of recombinant Listeria expressing Plasmodium CD8 T cell epitopes...91 DC-LM vaccination to generate antigen-specific CD8 T cells...92 Plasmodium parasites and sterilizing protection challenges...92 Quantification and analysis of Plasmodium-specific CD8 T cell populations...93 In vivo cytolytic assay...93 x
13 Functional avidity analysis...94 Liver parasite burden...94 Primary hepatocyte culture...95 Statistical analysis...97 Results...97 Radiation-attenuated sporozoite (RAS) vaccination induces CD8 T cells against identified and unidentified antigens...97 DC-LM vaccination induces large, long-lasting Plasmodium-specific CD8 T cell responses displaying different capacity for sterilizing protection...99 DC-LM vaccination generated CD8 T cells with similar phenotype and functionality Antigen sensitivity of DC-LM generated CSP , GAP , S , and TRAP specific CD8 T cells is similar DC-LM generated GAP and S specific CD8 T cells do not reduce liver parasite burden DC-LM generated GAP and S specific CD8 T cells recognize infected hepatocytes less efficiently than CSP , and TRAP specific CD8 T cells Discussion CHAPTER IV: FUTURE PERSPECTIVES Elucidating immune mediators of protective immunity against Plasmodium infection through WSV platforms CPS vaccination requires CD4 T cells and antibodies for protection in the stringent P. yoelii/c57bl/6 model Cross-stage reactive antibodies may be critical for CPS vaccine-induced protection Generation of immunity against heterologous Plasmodium strains CPS vaccination could be used as a model to elucidate the induction of experimental cerebral malaria Advancing subunit vaccination strategies through enhancing quantity and quality of protective CD8 T cell responses against liver-stage infection Localization of antigen targeted by an anti-plasmodium CD8 T cell response may dictate protective capacity Quantitative and qualitative features of CD8 T cell-mediated immunity as learned from novel CD8 T cell specificities xi
14 Beyond specificity: localization of the CD8 T cell response may matter Conclusion REFERENCES xii
15 LIST OF TABLES Table 1. Single-dose chemoprophylaxis and sporozoite (CPS) vaccination provides sterilizing protection from P. yoelii sporozoite challenge in C57Bl/6 mice...72 Table 2. Infection with P. berghei ANKA sporozoites primes CD8 T cell responses to several identified peptide epitopes Table 3. DC-LM generated CSP and TRAP specific CD8 T cells can provide sterilizing protection from sporozoite challenge xiii
16 LIST OF FIGURES Figure 1. Plasmodium infection of hepatocytes and red blood cells defines the two distinct stages of the Plasmodium life cycle Figure 2. CD8 T cells and antibodies target Plasmodium antigens during the course of liver-stage and blood-stage infection Figure 3. CPS induces larger effector and memory CD8 T cell responses than RAS vaccination in C57Bl/6 mice Figure 4. CPS induces larger effector and memory CD4 T cell responses than RAS vaccination in C57Bl/6 mice Figure 5. Chloroquine administration does not alter the kinetics or magnitude of the CD8 T cell response in C57Bl/6 mice 7 days after administration of attenuated recombinant Listeria monocytogenes expressing OVA Figure 6. I.V. administration of CPS or RAS induces larger effector and memory CD8 T cell response than S.C. administration in C57Bl/6 mice Figure 7. I.V. administration of CPS or RAS induces larger effector and memory CD4 T cell responses than S.C. administration in C57Bl/6 mice Figure 8. CPS vaccination induces larger magnitude CD8 T cell responses targeting a broader spectrum of liver-stage antigens than RAS vaccination Figure 9. CPS-induced CD8 T cells are not required for protection from homologous sporozoite challenge Figure 10. Parasites persist in the spleen and blood during CQ treatment and control of persistent parasites is dependent on CD4 T cells Figure 11. CPS vaccination with 25 days of CQ cover abrogates breakthrough parasitemia as well as protection Figure 12. CPS with 10 days of CQ cover correlates with higher anti-parasite antibody responses Figure 13. T cells are not required for protection from blood-stage parasite challenge in single administration of CPS mice xiv
17 Figure 14. Radiation-attenuated sporozoite vaccination primes CSP , GAP , S , and TRAP specific CD8 T cell responses Figure 15. Dendritic cell prime, recombinant L. monocytogenes boost (DC-LM) vaccination generates large Plasmodium-specific memory CD8 T cell responses in C57Bl/6 mice Figure 16. DC-LM TRAP vaccination of BALB.b mice generates a sterilely protective response in the majority of vaccinated mice Figure 17. DC-LM CSP, GAP50, S20, or TRAP vaccination of CB6.F1 mice leads to easily detectable epitope-specific memory CD8 T cell responses Figure 18. DC-LM GAP50 or S20 vaccinated BALB.b x BALB/c F1 mice have larger magnitude epitope-specific responses compared to DC-LM CSP or TRAP vaccinated mice Figure 19. Protective and non-protective memory CD8 T cells exhibit a similar surface phenotype in PBL Figure 20. Splenic-derived protective and non-protective memory CD8 T cells exhibit a similar surface phenotype Figure 21. Liver-derived protective and non-protective memory CD8 T cells exhibit a similar surface phenotype Figure 22. Protective and non-protective DC-LM generated splenic CD8 T cell responses are similar in cytokine expression following peptide stimulation Figure 23. Protective and non-protective DC-LM generated CD8 T cell responses are similar in capacity to kill peptide-pulsed targets in vivo Figure 24. Antigen sensitivity of protective and non-protective DC-LM generated CD8 T cell specificities is similar Figure 25. Non-protective GAP and S specific CD8 T cell responses do not significantly reduce liver parasite burden Figure 26. Efficient recognition of infected hepatocytes correlates with epitopespecific protection Figure 27. Whole sporozoite vaccination induce anti-plasmodium CD8 T cells and antibodies mediate protection against Plasmodium infections xv
18 Figure 28. Proposed model defining a differentiating factor between antigens targeted by protective and non-protective anti-plasmodium CD8 T cell responses xvi
19 LIST OF ABBREVIATIONS AICDA AMA-1 ANOVA APC CD8 CFUs CPS CSP CTV CQ activation-induced cytidine deaminase apical membrane antigen-1 analysis of variance antigen-presenting cell co-receptor for the T cell receptor on cytotoxic T cells colony forming units chemoprophylaxis and sporozoites circumsporozoite protein cell trace violet chloroquine CXCR6 chemokine (C-X-C motif) receptor 6 DC DMEM dendritic cell dulbecco modified eagle medium EC 50 effective concentration 50 ECM ELISA FBS FITC experimental cerebral malaria enzyme-linked immunosorbent assay fetal bovine serum fluorescein isothiocyanate GAP50 glideosome associate protein 50 GAPDH GFP GLURP HBsAg ICS IFNγ glyceraldehyde 3-phosphate dehydrogenase green fluorescent protein glutamate rich protein hepatitis B virus antigen intracellular cytokine stain interferon gamma xvii
20 IgG IL-2 I.M. I.P. I.V. IVM LM LPS LSA-1 mab MHC MOI MSP-1 NO - OT-I OVA pdc Pb PBL PBS Pcc P/S PV Py RBC rlm immunoglobulin G interleukin-2 intramuscular intraperitoneal intravenous intravital microscropy Listeria monocytogenes lipopolysaccaride liver-stage antigen-1 monoclonal antibody major histocompatibility complex multiplicity of infection merozoite specific protein-1 nitric oxide CD8 T cell receptor transgenic cells specific for ovalbumin ovalbumin plasmocytoid dendritic cell Plasmodium berghei ANKA peripheral blood leukocytes phosphate buffered saline Plasmodium chabaudi chabaudi penicillin/streptomycin parasitophorous vacuole Plasmodium yoelii 17XNL red blood cell recombinant Listeria monocytogenes xviii
21 qpcr qrt-pcr RAS RPM rrna quantitative polymerase chain reaction quantitative reverse transcription polymerase chain reaction radiation attenuated sporozoites rotations per minute ribosomal ribonucleic acid S20 sporozoite-specific protein 20 S.D. S.E.M. standard deviation standard error of the mean SSP2 sporozoite surface protein 2 TCR Tg TNFα TRAP WSV T cell receptor transgenic tumor necrosis factor alpha thrombosponin related adhesion protein whole sporozoite vaccination xix
22 CHAPTER I: INTRODUCTION Natural and unnatural immunity against Plasmodium Global impact and lifecycle of Plasmodium infections Infections with mosquito-borne protozoan parasites of the genus Plasmodium lead to an estimated 2.5 million cases, and 400,000 deaths a year (1, 2). These high rates of infection and death leads to detrimental health and socioeconomic impact in over 100 countries. The use of bed nets, insecticides, and other vector control measures, in concert with anti-malaria drugs, have had a dramatic impact as evident by a reduction in the mortality rate from more than a million deaths a year in the 1990s. Thus, it is clear that interventions have had a positive impact on reducing infection rates and subsequent malaria disease in malaria-endemic areas. However, Plasmodium is increasingly becoming resistant to drug interventions (3-6), highlighting the need for additional control measures, such as targeting the multiple life-cycle stages of the Plasmodium parasite through vaccination measures (7), in order to control the parasite and lead to the worldwide goal of eradication (8, 9). The Plasmodium parasite has a complex multi-host, multi-phase life cycle (10). Similar to other organisms in the Phylum Apicomplexa, the Plasmodium parasite has a portion of its lifecycle within an insect vector, in this case the mosquito, and another within a vertebrate host such as a mammal or bird. Transmission of human Plasmodium species to date is restricted to only the Anopheles genus of mosquitoes (11, 12). When it 1
23 comes to the vertebrate host, Plasmodium species are generally restricted to specific hosts such that human Plasmodium species (P. falciparum, P. vivax, P. ovale, P. malariae, and P. knowlesi) infect humans (or non-human primates), and rodent strains (P. yoelii, P. berghei, and P. chaubaudi) are infectious to rodents, but are not infectious to human subjects (13, 14). It is likely these groups of Plasmodium species diverged from each other thousands of years ago (15), but despite this, large overlap in sequence homology exists between classic human and rodent Plasmodium species (15-17). Consequently, these conserved similarities among Plasmodium species have allowed studies utilizing rodent models of malaria to reveal important mechanistic details about immunity against Plasmodium infections that can be extended to human infections (18, 19). The life cycle of the Plasmodium parasite can be further divided into two distinct stages within the vertebrate host: liver-stage and the blood-stage (Figure 1) (20). Infection of vertebrate hosts begins when a mosquito harboring Plasmodium sporozoites in the mosquito salivary glands injects parasites during a blood meal. Plasmodiuminfected mosquitoes may inject approximately sporozoites into the skin during a blood meal (21-26). Early imaging studies indicated injected sporozoites may remain at the injection site for a few hours before approximately 60% of them travel via gliding motility to a nearby blood vessel to access the blood stream and traffic to the liver (27-29). Of interest, recent studies have shown that some parasites never leave the site of inoculation, and instead undergo abortive differentiation in the skin (30, 31). However, a portion of sporozoites do reach the liver and interact with the resident macrophage, Kupffer cells, or the liver endothelium via heparin sulfate glycans expressed on the cell 2
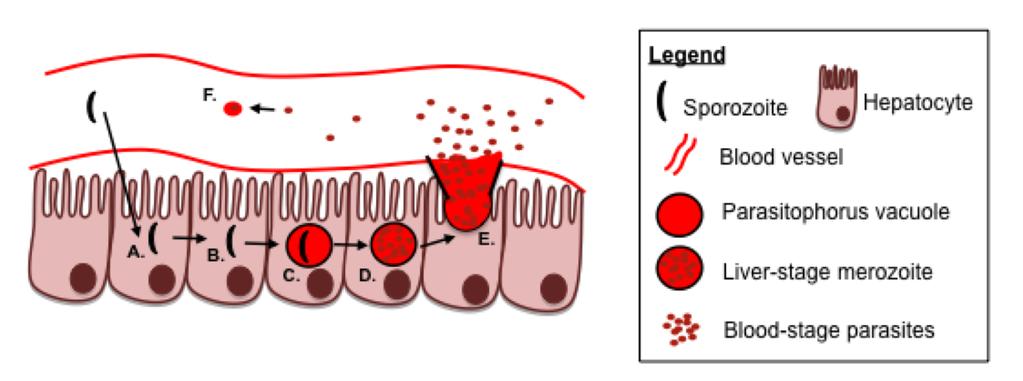
24 surface (32-34). Sporozoites travel through Kupffer or endothelial cells into the liver parenchyma in order to infect the underlying hepatocytes and initiate liver-stage infection (Figure 1) (34, 35). Interestingly, sporozoites will not necessarily initiate a productive infection within the first hepatocyte they enter (Figure 1A), but may undergo an event called cell traversal to traffic through several hepatocytes (Figure 1B)(36, 37). Eventually the sporozoite will stop traversing hepatocytes and instead form a moving junction with a target hepatocyte (36, 38-40), developing a parasitophorous vacuole (PV) around itself to initiate infection (Figure 1C). The PV shields and thus prevents access of liver-stage antigens to host cell proteolytic machinery, however the parasite does target proteins to the PV (41), and may possibly export proteins into the cytoplasm and nucleus of the infected cell (42, 43). The initial interaction to infect a hepatocyte instead of traversing is depends on the sporozoite surface protein circumsporozoite protein (CSP) to attach to heparin sulfate molecules on the hepatocyte (32, 44-46). Additionally the sporozoite surface protein thrombospondin related-adhesion protein (TRAP) is also required for internalization of the parasite into hepatocytes, but it is not required for attachment (47, 48). Within the PV, the sporozoite differentiates into a liver-stage merozoite, undergoing multiple rounds of replication (35, 39, 49, 50). During the process of differentiation and replication in the PV within the hepatocyte, the parasite will develop a merosome structure, which is a PV with nearly 7,000 or more merozoite parasites inside (51). In the process of developing from a single sporozoite into thousands of merozoites, the parasite expresses novel liver-stage antigens; this will have an important impact as later in the development of immune responses 3
25 against Plasmodium antigens (Figure 1D). Eventually the merozoite filled merosome buds from the infected hepatocyte into the liver sinusoid then travels to the lung before releasing the merozoite contents into the circulation (Figure 1E)(52). The liver-stage of infection lasts approximately seven days in a human host, and two day within the rodent host. Throughout the course of the liver-stage infection, the host does not experience symptomatic infection. Therefore, because of the initial low parasite density, and the lack of symptoms of malarial disease, the liver-stage of infection is considered an attractive life-stage to target via therapeutics and vaccination. Following release from the liver into the blood, the parasites will infect red blood cells to initiate the blood-stage of the Plasmodium lifecycle (Figure 1F). In contrast to the liver-stage of infection, the blood-stage is characterized by high parasite density. Each merozoite, from among thousands of merozoites released from an infected hepatocyte, could potentially infect a single red blood cell, which leads to a large amplification from the initial sporozoite infection. Within an infected red blood cell, the merozoite will differentiate and replicate producing approximately parasites merozoites per infected red blood cell over a 1-3 day time frame (53). Eventually bloodstage merozoites will rupture the red blood cell and go on to infect additional red blood cells, continuing to amplify the blood-stage infection. This amplification leads to an increase in measurable blood-stage parasitemia (% of infected red blood cells). Eventually, the infected host will have a high enough density of parasites infecting RBCs that blood-stage infection is detectable by Giemsa-stained blood smear; at this point the infected host has a patent infection. As parasitemia increases in the host, symptoms of 4
26 malarial disease begin such as fever, anemia, fatigue, sweats, and in severe cases coma, or respiratory distress (1, 14, 54). Through an unknown mechanism, some blood-stage parasites will differentiate into the sexual stages of the parasites called gametocytes (55). The life cycle of the Plasmodium parasite transitions from the mammalian host back to the mosquito vector when a mosquito successfully takes up female and male Plasmodium gametocytes from an infected mammalian host during a blood meal. In the mosquito vector, a complex series of mosquito-specific events eventually leads to the generation of sporozoites within the mosquito host (56). Naturally-acquired immunity against Plasmodium does not develop Humans living within malaria-endemic regions can be continuously infected with Plasmodium throughout their life, and never develop complete, natural immunity (57, 58). This is in contrast with several viral and bacterial infections where the host develops sufficient memory immune responses to the pathogen such that future exposures do not lead to disease (59, 60). Repetitive exposure to Plasmodium infections over time leads to less severe clinical disease in adults, whereas individuals with less disease history, primarily children <5 years old, are the most susceptible to severe disease and death from Plasmodium infections (1, 14). Induction of humoral immunity does occur, as evidenced by antibody titers directed against defined liver-stage and blood-stage antigens (61-63), but for still unknown reasons these responses are insufficient for complete immunity. The lack of naturally-acquired immunity against Plasmodium has long spurred an interest in understanding how to make unnatural immunity via whole sporozoite or subunit 5
27 vaccination strategies such that sterilizing protection (lack of detectable blood-stage parasites) is achieved in human subjects following experimental sporozoite challenge or natural infection (64-66). Generation of protective cellular immunity by whole sporozoite vaccination The first published evidence that attenuated whole sporozoite vaccination (WSV) could provide protection in an animal model was shown in birds. Immunization with UV light damaged P. gallinaceum sporozoites protected birds from mortality following sporozoite challenge (67, 68). Shortly thereafter, attenuated WSV studies were extended into rodents. Sterile protection from Plasmodium challenge (defined as absence of bloodstage infection following sporozoite challenge) was initially demonstrated in the 1960s when mice were immunized with the bites from mosquitoes harboring infectious P. berghei parasites that were attenuated due to irradiation of the infected mosquito vector (69). Irradiation of the mosquito harboring Plasmodium damages the parasite DNA, thus preventing the ability of the parasite to complete liver-stage differentiation. Thus, the parasite can infect hepatocytes, but cannot develop merozoites to initiate symptomatic blood-stage infection (69, 70). Importantly, this approach termed radiation-attenuated sporozoite (RAS) vaccination, also provided sterilizing immunity when tested in human subjects (71, 72). Of note, sterilizing immunity in humans required the bites of more than 1,000 irradiated P. falciparum-infected mosquitoes. Because of the ability of RAS vaccination to protect humans, this WSV approach is commonly considered the old standard of malaria vaccination. However, the future application of this vaccination 6
28 approach depends on overcoming concerns with the safety of administering an irradiated whole parasite vaccine, as well as the logistics of field application (i.e. high parasite dose requirements, multiple administrations of vaccine, etc.) (70, 73, 74). In regard to safety, RAS vaccination requires the sporozoites be sufficiently irradiated to prevent completion of liver-stage infection, but not over irradiated to lose immunogenicity and thus the capacity to induce immune-mediated protection (75). Additionally, these sporozoites are hand dissected from laboratory-reared mosquitoes, subjected to assays to insure sterility, and then cryopreserved for long-term storage all factors that complicate the scalability of production (70). Further, application to the field requires knowledge of total dose required for protection, timing of booster immunizations, and preservation of the vaccine in a field lacking a cold-chain network. Extensive work has been done to address these concerns over the last several years (70). Currently, the RAS vaccination approach is being aggressively developed and tested in safety and clinical trials in hopes that it can be eventually distributed in the field (70, 76-80). An approach that seeks to overcome some of the safety concerns inherent to RAS vaccination is the use of genetically attenuated parasite (GAP) vaccination. In this approach, gene-specific deletions are created that do not affect mosquito or blood-stage replication, but specifically prevent the production of proteins essential for liver-stage development as a means of attenuating the sporozoite without losing immunogenicity. Because of the deletion of a gene(s) in the GAP approach, it is arguably a more controlled attenuation than the irradiation used for attenuation in RAS vaccination. Further, vaccination with some GAPs may elicit more potent immunity than RAS due to 7
29 the longer development period the parasite can undergo in the hepatocyte. Indeed, this has been demonstrated to be the case in a BALB/c mouse model of RAS versus lateliver-stage-arresting GAP vaccination (81). Successful GAP vaccination requires the deletion of an essential gene for complete liver-stage development to attenuate the parasite and prevent the progression to blood-stage infection. Should this attenuation fail, clinical disease may occur after vaccination. In the first human clinical trial of GAP vaccination, one of six patients experienced blood-stage infection (breakthrough parasitemia) following a high dose vaccination with P. faliciparum p52 - /p36 - parasites (82). Importantly, immunization with GAP parasites deleted in the homologs of these genes in P. yoelii led to no breakthrough in mouse studies using a P. yoelii (83). Unfortunately, this occurrence highlights the major disadvantage of GAP vaccination approaches that are primarily tested using rodent Plasmodium before moving into P. falciparum for evaluation in human subject studies. GAP vaccination overcomes safety concerns of irradiating sporozoites inherent to the RAS approach, but its success requires absolute attenuation of complete liver-stage development through gene deletion. An alternative approach to RAS or GAP vaccination is chemoprophylaxis and sporozoites (CPS) vaccination (84), whereby the subject is administered virulent sporozoites via mosquito bite or needle injection while concurrently receiving antimalarial drugs that prevent blood-stage infection (63, 84-86). To date, human subject studies have focused on the use of chloroquine drug administration. Chloroquine does not alter the liver-stage infection, but blood-stage infection is halted after a single round of infection due to the blood-stage parasite-specific action of the drug (87). CPS appears to 8
30 be the most potent WSV approach based on rodent and human challenge studies (84, 85, 88). Although concerns about chloroquine resistance of Plasmodium will prevent the direct application of this specific approach to the field (6, 89), it does provide a platform to understand the mechanistic requirements of immunity required to provide complete, sterilizing protection (i.e., no blood-stage infection) against sporozoite challenge. The application of WSV approaches in rodent models of malaria have led to a mechanistic understanding of the immunological responses that contribute to protection in rodents. Lessons learned in rodent models have helped direct efforts to understand how protection is mediated in human subjects receiving WSV. Rodent and human studies have both shown important roles for CD8 T cells, CD4 T cells and antibodies in WSV-induced protection (Figure 2). To date, subunit vaccination of humans has not led to sterilizing immunity in the majority of vaccinees (90-93). Thus, studies of the immune response generated following the gold standard for vaccination (RAS) or other WSV approaches in rodent models of malaria continue to enhance our understanding of the underlying immune responses that contribute to protection. CD8 T cell responses Radiation attenuated sporozoites The first direct evidence of the role of CD8 T cells in vaccine-induced protection against Plasmodium was demonstrated using mouse models of RAS vaccination. Two independent studies showed that antibody-mediated depletion of CD8 T cells prior to 9
31 challenge abrogated protection in RAS-vaccinated mice, whether P. berghei or P. yoelii RAS was evaluated (94, 95). Not long after these studies, adoptive transfer studies using a CSP-specific CD8 T cell clone generated from RAS-vaccinated mice conferred sterilizing protection to naive mice against challenge with sporozoites (96, 97). Many additional studies using various sporozoite doses, number of immunizations, mouse strain, and parasite species combinations have further identified a critical role for CD8 T cell-mediated protection following RAS vaccination of mice (76, ). Thus, the rodent-ras vaccination model has greatly aided in the understanding of correlates of protective immunity against Plasmodium as it allows comparisons between mouse strains and rodent Plasmodium species for the magnitude, phenotype, and functionality of the CD8 T cell response required for protection. For instance, the quantity or functionality of the protective CD8 T cell response can differ based on the rodent model as demonstrated in studies examining the protection of P. berghei ANKA RAS-vaccinated C57Bl/6 mice from challenge with homologous parasites, which required memory CD8 T cells compromising 11% or greater of circulating CD8 T cells (99). In contrast, a threshold of ~4 % of the circulating CD8 T cells is required to achieve robust sterilizing immunity in BALB/c mice immunized with P. berghei ANKA RAS (99). Thus, larger CD8 T cell responses are required to protect C57Bl/6 mice from P. berghei ANKA than BALB/c mice. In the case of C57Bl/6 mice, protection following RAS vaccination was provided by CD8 T cells expressing IFNγ and TNFα, whereas only IFNγ expression was detected from CD8 T cells from BALB/c mice (99). It remains unknown if these differences in 10
32 cytokine production impact the threshold of memory CD8 T cells required for protection after RAS vaccination. Radiation-attenuated sporozoite vaccination of human subjects has been successful in providing protection from sporozoite challenge (71, 72, 78). Despite the plethora of direct evidence for the contribution of Plasmodium-specific CD8 T cells in protection from challenge in mouse models of RAS vaccination (76, 94, 95, 98, 99), direct evidence is lacking in human studies because of the inability to conduct CD8 T cell depletion studies. However, studies have shown that RAS vaccination of human subjects elicits peripheral blood T cell responses producing IFNγ, TNFα, and IL-2 as measured following ex vivo sporozoite stimulation (76, 78). Moreover, examination of the RAS vaccination induced CD8 T cell response reveals a dose-dependent increase in the frequency of Plasmodium-specific IFNγ + CD8 T cells that correlates with protection of human subjects from challenge (78). These studies indicate that liver-stage-specific, cytokine-producing CD8 T cell responses are induced following RAS vaccination of human subjects and may contribute to RAS vaccine-induced protection. Genetically attenuated parasites Similar to RAS vaccination, CD8 T cells play a dominant role in GAP-induced protection from sporozoite challenge (81, ). Various rodent Plasmodium GAP with targeted deletions that affect different phases of the liver-stage lifecycle have been tested in rodent vaccination studies (41, 83, 101, 102, ). The main immunological 11
33 appeal of the GAP vaccination strategy relative to RAS is the capacity to stop the infection late in the liver stage, thereby potentially increasing the antigenic targets for CD8 T cell recognition (Figure 2B,C). Vaccination of mice with late-liver-stage-arresting GAP led to a significant increase in the CD8 T cell response at both the effector phase and memory phase compared to vaccination with an early-liver-stage-arresting GAP sporozoites or RAS (81). Further, vaccination with late liver-stage-arresting GAP required fewer immunizations to achieve protection compared to early-liver-stagearresting GAP or RAS vaccination (81). Genetically attenuated parasite vaccination approaches using later-liver-stage-arresting parasites could be a potent approach for human vaccination, potentially reducing the dose/number of immunizations required to elicit CD8 T cell-mediated immunity in human subjects. To date, only one human clinical trial using an early-arresting GAP vaccine has been published (82). This vaccination utilized a GAP deficient in two genes, p52, and p36, which arrests the parasite early in liver-stage development (83). Peripheral blood Plasmodium-specific CD8 T cell responses producing primarily IFNγ were detected following ex vivo stimulation with whole sporozoites in all the human subjects at 90 days after the second immunization (82). Whether the GAP vaccination-induced IFNγ producing CD8 T cells correlate with protection in these human subjects is unknown as protection from challenge was not assessed in this phase I study. Taken together, the induction of IFNγ-producing Plasmodium-specific CD8 T cell responses following human vaccination with GAP suggests that GAP vaccination may elicit protective immune responses in human subjects and thus warrants further investigation. 12
34 Chemoprophylaxis and sporozoites Administration of virulent sporozoites concurrent with the antimalarial drug chloroquine (CPS) appears to be the most potent WSV approach to date (81, 88), perhaps due to the additional target antigens provided with complete liver-stage development and abortive blood-stage infection (110). In fact, CPS can elicit protective immune responses against P. yoelii 265BY in BALB/c mice with one vaccine administration whereas two or more adminsitrations of RAS were required for sterilizing protection (88). Further, GAP and RAS vaccination required two or three vaccination administrations, respectively, to engender sterilizing immunity against homologous P. yoelii 17XNL sporozoite challenge in a BALB/c mouse model (81, 88). Taken together, CPS is able to efficiently induce potent immunity with fewer administrations than RAS or GAP. The prime mediators of this protective immunity in CPS versus RAS vaccinated BALB/c mice administered virulent P. yoelii 265BY sporozoites were CD4 and CD8 T cells. Antibody-mediated depletion of CD4 or CD8 T cells just prior to challenge increased the liver parasite burden, suggesting a role for CD4 and CD8 T cells against liver-stage parasites (88). More importantly, human subjects vaccinated through a CPS approach are protected from sporozoite challenge, and this protection is associated with anti-plasmodium T cell responses (85, 86). Human subjects receiving CPS developed peripheral blood T cells that produced IFNγ, TNFα, and IL-2 cytokines following ex vivo stimulation with bloodstage parasites or sporozoites (85, 86). In one study, a longitudinal phenotypic analysis of blood-derived CD4 and CD8 T cells from CPS-vaccinated donors indicated proliferation and cytotoxic profiles (111), but to my knowledge no human CPS 13
35 vaccination study have functionally evaluated CD4 and CD8 T cell subsets individually. Thus, the basis of the durable immunity mediated by CPS vaccination (86) correlates with T cell responses, but it remains unclear what specific T cell subsets best correlate with this protection. Collectively, human subjects receiving CPS mounted T cell responses specific to sporozoite or blood-stage parasites, but the CD8 T cell portion of this response is not well defined. Human studies, as well as mouse models, have supported the role of liver-stagedirected CD8 T cell responses in protection using WSV approaches. Evidence for the protective capacity of CD8 T cells in mouse studies has largely been determined through the use of antibody-mediated depletion of CD8 T cells just prior to challenge, through the use of knockout mouse models, as well as cellular transfer experiments (94-96, 112). Although each of these approaches has caveats, the relatively consistent result of CD8 T cell dependence for protection in mouse models of WSV, particularly after RAS and GAP vaccination, supports a critical role for CD8 T cells in protection from sporozoite challenge. Taken together, mouse and human studies of WSV have demonstrated the induction of Plasmodium-specific CD8 T cell responses, which directly contribute to protection in several rodent studies, and are correlated with protection in human studies. CD4 T cell responses While substantial data support the role of WSV-induced CD8 T cells in protection against sporozoite challenge, the protective role of CD4 T cells is less clear. Studies using 14
36 a mouse model of malaria suggest that a role of CD4 T cells in protection against sporozoite challenge may depend on the mouse strain used for evaluation (98). In one such study, RAS vaccination was administered to seven inbred mouse strains and the contribution of CD4 T cells in protection was evaluated by performing CD4 T cellspecific antibody-mediated depletion prior to challenge. In contrast to CD8 T cells, which were required for protection in all tested mouse strains, CD4 T cells were required for protection in only three out of seven mouse strains (C57Bl/6; B6,129; B10.D2)(98). Interestingly, two prominently used mouse strains for Plasmodium studies, the BALB/c and C57Bl/6, substantially differed in their requirement for CD4 T cells in protection. Namely, protection in RAS-vaccinated BALB/c mice was CD4 T cell independent, while CD4 T cells played an important role in protection of RAS-vaccinated C57Bl/6 mice against sporozoite challenge (98). Thus, so far studies indicate the role of CD4 T cells in WSV-induced protection is unclear, and may be mouse strain dependent; this in contrast to the requirement for CD8 T cells in WSV-induced protection in tested mouse strains thus far. Vaccination of human subjects with WSV induces detectable Plasmodiumspecific CD4 T cells responses in the peripheral blood. It has been shown that these CD4 T cells are capable of producing cytokines upon ex vivo stimulation with blood-stage parasites or sporozoites (76, 78, 82, 85). Furthermore, the magnitude of the cytokine producing RAS-induced CD4 T cell response directly correlated with the administered vaccine dose (78). Although testing the direct involvement of CD4 T cells in protection against sporozoite infection in humans is not possible, it is clear that CD4 T cell 15
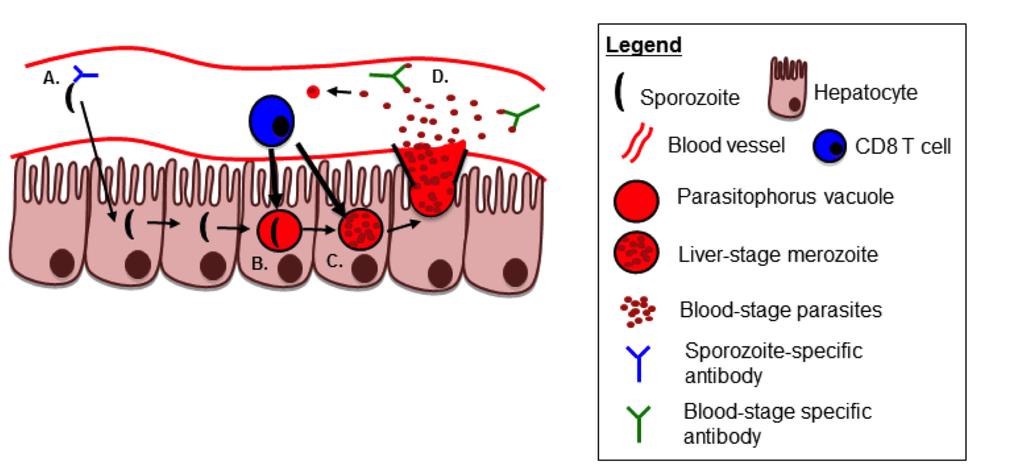
37 responses are elicited in human subjects by WSV vaccination and their magnitude depends on vaccine dose and correlates with observed protection. Antibody responses Anti-Plasmodium antibodies are capable of inhibiting parasite infection and contributing to parasite clearance of blood-stage infections (28, ), but their role in protection following WSV remains largely undefined. Before the initial studies of RAS vaccination demonstrating a prominent role for liver-stage-directed CD8 T cells in protection against challenge, mouse studies of RAS vaccination revealed the induction of antibodies against CSP, suggesting an important role of humoral immunity in protection (116, 117). However, RAS vaccination of C57Bl/6N x BALB/c AnN F1 mice can elicit protection independently of B cells with as few as two immunizations (100), which indicates that antibodies are not necessary for RAS-induced protection. Taken together, WSV approaches can elicit anti-sporozoite antibody responses, but protection from challenge may be independent of B cells/ antibodies through CD8 T cells (100). To date, anti-csp antibodies are arguably the most studied antibody response following human WSV (63, 78, 82, 85). Anti-CSP antibody responses have been shown to inhibit sporozoite infection of hepatocytes in vitro and in vivo (28, 115), and monoclonal CSP-specific antibodies can confer sterilizing protection against sporozoite challenge in mice (Figure 2A)(117, 118). Antibodies directed against CSP have been detected in many human studies of WSV (63, 78, 82, 85). Following RAS vaccination 16
38 of human subjects, anti-csp titers correlated with the immunization dose, with protected individuals having higher antibody titers compared to non-protected individuals (78). In human GAP and CPS vaccination studies, anti-csp antibody responses were detected in the majority of volunteers (82, 84, 85). However, only one of six patients who received CPS had detectable CSP antibodies in a follow-up study (28 months later) (86), suggesting that CPS-induced anti-csp antibody responses are short-lived. The contribution of anti-csp antibodies to protection of human subjects following WSV may not be clear, but in RAS, GAP, and CPS approaches, it is clear that human subjects can make anti-csp antibody responses. Antibodies directed against other liver-stage targets besides CSP (i.e., LSA-1 and SSP2) have been detected in human subjects following WSV (63, 119). The protective capacity of these non-csp-specific antibodies is currently unknown. However, total WSV-induced antibodies in human subjects may contribute to protection through inhibiting sporozoite infection of hepatocytes. For example, plasma collected three months following two-dose immunization of human subjects with GAP vaccination inhibited P. falciparum invasion of hepatocytes in vitro (120). This inhibition of invasion was similar to plasma collected from human subjects who received 4 5 doses of RAS sporozoites (78). Thus, both RAS and GAP vaccination of human subjects induce antibodies, which can inhibit hepatocyte invasion in vitro. Human CPS studies have shown the persistence of blood-stage parasites following vaccine administration (85), suggesting that blood-stage-specific immune responses could contribute to protection. These results highlight the continued need to consider the anti-parasitic humoral immune response, which could contribute to 17
39 complete, sterilizing immunity in addition to or independently of anti-plasmodium CD8 T cell responses. Despite the prominent role of liver-stage-directed CD8 T cells in protection following WSV, and lack of robust evidence for T cell-independent antibodymediated protection, further studies of antibody responses may help in the development of the most potent strategy for human vaccination. Human subjects receiving WSV can make antibody responses directed against liver-stage parasite antigens, which may aid in protection through inhibition of sporozoite infection. Blood-stage-specific antibodies can be detected in human subjects receiving WSV, but only if the subject was exposed to blood-stage parasites (i.e. CPS)(Figure 2D). In human RAS studies, where the subject is never exposed to blood-stage infection, antibodies directed against crude blood-stage parasite lysate or against known bloodstage antigens were not detected (76, 78). Likewise, GAP vaccination of human subjects did not elicit anti-merozoite surface protein-1 (MSP-1) antibodies, a known blood-stage antigen, except in the one patient that had detectable blood-stage infection from vaccination breakthrough (82). In contrast to RAS and GAP vaccination, CPS vaccination induces antibodies directed against crude blood-stage parasite lysate, which is not surprising given the known exposure to blood-stage parasites during vaccination (85). Antibodies against specific blood-stage antigens such as MSP-1, AMA-1 or GLURP were not detected in some CPS studies (82, 85, 86), but were detected in one human CPS vaccination with a similar study design (121). Taken together, these results indicate that CPS vaccination, wherein subjects are exposed to some blood-stage infection (84, 85) can lead to induction of blood-stage specific antibodies. However, to 18
40 date these antibody response are insufficient to sterilely protect CPS vaccinated human subjects from blood-stage parasite challenge (84). However, it is unknown whether CPSinduced blood-stage-specific immunity is capable of controlling infection and contributing to reduction of disease, since the humans undergoing blood-stage challenge were treated with anti-malarial drugs immediately upon detection of blood-stage parasite. Thus, exposure of human subjects to blood-stage parasites during CPS elicits bloodstage-specific antibody responses, but it is unknown whether these antibodies contribute to the total protective immune response. Collectively, WSV approaches have demonstrated success when applied to human subjects, and the use of rodent models of malaria have provided information regarding what features of the WSV-induced immune response likely provide protective outcomes. However, the application of WSV approaches in the field is complicated by many factors (i.e. high parasite dose, laboratory-reared mosquitoes, multiple administrations in an environment largely lacking medical facilities for proper administration, etc) (70). Thus, there is a need to better understand the features of what dictates a protective CD8 T cell response (i.e. the quanitity and quality of the response) and application of WSV approaches in rodent models of malaria will be essential to achieve this understanding. Subunit vaccination approaches against Plasmodium have limited efficacy The most advanced subunit vaccination, Mosquirix, was recently licensed in the European Union (122). Mosquirix may be more commonly known as RTS,S, as 19
41 designated during its pre-clinical development and clinical trials. RTS,S is a subunit vaccine composed of particles of hepatitis B S antigen (HBsAg) and a CSP polypeptide comprised of 19 NANP repeats and the carboxyl-terminal fused to the amino-terminal of the S antigen (123, 124) administered with the AS02 adjuvant (125). Although this subunit vaccine was recently licensed for use, it has very low overall efficiency and vaccination-induced protection wanes over time (126). For example, phase IIb trials of Mozambique children demonstrated a 58% reduction in severe disease during the first six months (126). In studies from phase III clinical trials in the targeted age population of 1-4 year olds and infants, RTS,S vaccination lead to 56% and 65%, respectively in severe disease reduction (127, 128). Although these results indicate a degree of protection due to the absence of severe disease in a portion of vaccinees, the results are still suboptimal as blood-stage infection still occurs in these children. Thus, the RTS,S vaccine will not likely lead to long-term eradication of malarial disease, a goal set forth by the World Health Organization and the Bill and Melinda Gates Foundation to achieve by 2030 (1, 8, 9). Therefore, it remains important to continue to study WSV and subunit vaccination approaches to ultimately develop more efficacious subunit vaccines to achieve this goal. To this end, several prime-boost subunit vaccination platforms are being studied to overcome the limitations of the RTS,S vaccine. The more successful of these vaccination platforms employ viral vector heterologous prime-boost platforms wherein the subject is exposed to an antigen in the context of a one viral vector, and then later exposed to the same antigen but in the context of an entirely new viral vector (129). This heterologous approach is necessary as initial exposure to the vector during the priming of 20
42 the response generates immune responses specific to the viral vector leading to enhanced clearance and less optimal boosting with the desired antigen in a homologous vector (130, 131). Several antigens are being targeted for use in viral vectored heterologous primeboost vaccines, most notably the CSP and TRAP liver-stage antigens since responses generated against these two antigens can confer protection or reduce parasite burden in rodent models (96, 97, ). Additionally, some viral vectored subunit vaccines are targeting blood-stage expressed antigens ( ). Inclusion of liver-stage antigens CSP and/or TRAP into subunit vaccine formulations for human vaccination have led to some degree of sterilizing protection in human subjects. In regards to viral vector heterologous prime boost vaccines using CSP antigen, sterilizing protection of vaccinated malaria-naïve individuals following a controlled challenge has been observed in 7% of subjects (1/15 subjects)(90). In contrast, evaluation of sterilizing immunity developed following prime-boost vaccination with a TRAP polyepitope string, ME-TRAP, achieved 13% (2/15) sterilizing immunity in malaria-naïve human subjects (90). Studies evaluating efficacy in a malaria-endemic population in Kenya using this same vaccine demonstrated a calculated 67% reduction in infection in vaccinated subjects during the eight weeks of monitoring (140). However, this study did not examine protection following a controlled challenge but instead evaluated risk reduction of infection in an edemic area. Collectively, these results demonstrate an advancement in viral vectored prime-boost subunit vaccines to provide sterilizing immunity relative to RTS,S, but 21
43 additionally highlight the need to further develop subunit vaccines to achieve better efficacy. Subunit vaccination approaches likely have been met with limited success due to two limiting factors: 1) limited ability to induce numerically large protective T cell and antibody responses in vaccinees and 2) the lack of defined antigens which can be targeted by protective cellular immune responses. The magnitude of the CD8 T cells induced by subunit vaccination correlates with protection against malaria, whereas antibodies do not (91, 141). Thus, inducing numerically larger CD8 T cells responses may enhance the efficacy of subunit vaccines. Further enhancement of subunit vaccines may be achieved through targeting other Plasmodium antigens than CSP and TRAP. The identification of additional antigens through screening methods for incorporation in subunit vaccines is a difficult task since the Plasmodium genome contains an estimated 5,000 open reading frames (142, 143), but more importantly an estimated 1991 proteins are expressed by the sporozoite as it leaves the salivary glands of an infected mosquito (144). Therefore, a need remains to identify additional antigenic targets to incorporate into advance subunit vaccination approaches, and/or enhance the quantity or quality of the current induced responses with CSP and TRAP antigens. In order to achieve this goal, lessons learned from other infectious disease systems and from the gold standard in malaria vaccination will need to be employed. 22
44 Quantity and Quality of the CD8 T cell response against Plasmodium The importance of CD8 T cells in mediating pathogen clearance is widely appreciated in many disease models. The work done in other infectious disease models has helped define our understanding of the generation and maintenance of protective CD8 T cell responses. Particularly, these models have shown the importance of quantity (the total numbers) (134, 145, 146), and that the quality of the CD8 T cell response impacts its protective capacity (147, 148). These studies have lead to an understanding of how to generate and maintain protective CD8 T cell responses against various infectious pathogens, which provides a framework to apply these lessons to the development of advanced subunit vaccination approaches to protect human subjects against malarial disease. Quantity of the CD8 T cell response to engender protection The number of antigen-specific CD8 T cells correlates with protective outcomes in infectious disease (145, 146), and Plasmodium infection is no exception. Through rodent models of malaria, it has been shown that sterilizing protection from sporozoite challenge requires numerically large CD8 T cell responses, on the order of > times larger than required for some bacterial or viral infections (145, 146). One clear demonstration of this extreme numerical requirement was through the generation of large CSP specific CD8 T cell response by administration of peptide-loaded mature 23
45 dendritic cells followed by boosting with recombinant L. monocytogenes expressing the peptide epitope (DC-LM vaccination) in BALB/c mice. The majority of DC-LM vaccinated BALB/c mice were sterilely protected from sporozoite challenge if the frequency of CSP 252 -specific CD8 T cells exceeded ~1% of all PBL (134). Further studies addressed the numerical requirement of CD8 T cells to provide sterilizing immunity using RAS vaccination of BALB/c mice. Interestingly, diversifying the CD8 T cell response by targeting multiple unknown, non-csp specificities with RAS vaccination (149, 150) did not drastically reduce the numerical requirement for CD8 T cell-mediated protection (99). Taken together, these results illustrate the extreme numerical requirement for CD8 T cell-mediated sterilizing immunity. The magnitude of the CD8 T cell response in peripheral blood of human subjects correlates with efficacy in several WSV and subunit vaccination studies (78, 85, 90, 151). Additionally, this correlation also extends to observations in non-human primates (152). Thus, to date there is clear evidence that the quantity of the CD8 T cell response against Plasmodium impacts protection in rodents, non-human primates, and humans. This correlation may be helpful to evaluate antigen candidates for vaccines, or predict the efficacy of a subunit vaccination approach before human clinical trials. However, a limiting factor in regards to human studies is that CD8 T cell responses can only be measured in the PBL, which may not be an accurate predictor of the quantity of the response in the spleen or liver. Thus, responses in the liver may be more relevant to protection than a circulating CD8 T cell population (76). In sum, the quantity of the CD8 24
46 T cell response against Plasmodium antigens is an important factor contributing to sterilizing protection. Qualitative features dicating protection against Plasmodium In addition to the quantity of an anti-plasmodium CD8 T cell response, the quality (phenotype, functionality, specificity, location, etc.) of the response also can impact protection from sporozoite challenge (153). While it is known that large magnitude CD8 T cell responses generated through vaccination are required to provide sterilizing immunity to sporozoite challenge of rodents, and likely also to human subjects (134), less is known about the qualitative features that are important for CD8 T cell-mediated clearance of parasite-infected hepatocytes. In regards to phenotype, CD8 T cells exhibiting an effector memory phenotype (CD62L lo ) have correlated with better protection relative to central memory phenotype (CD62L hi ) CD8 T cells in two independent studies (99, 154). Importantly, these studies used distinct vaccination methods (RAS versus heterologous prime-boost) to arrive at a similar conclusion. In regards to functionality, such as cytokine production and effector capacity, it has been repeatedly reported that expression of IFNγ by CD8 T cells is required for a protective response (94, 155, 156). Of interest, the cytolytic mechanism by which CD8 T cells mediate clearance of infection differs depending on the rodent Plasmodium infection (155). For instance, when P. berghei CSP-specific CD8 T cells were induced using a prime-boost subunit vaccine approach in wild-type or various knockout mouse backgrounds, it was demonstrated that CSP-specific CD8 T cells depend on IFNγ and 25
47 TNFα to mediate protection from challenge (155). In contrast, in a similar prime-boost vaccination model inducing P. yoelii CSP-specific CD8 T cells, it was shown that CSPspecific CD8 T cells largely mediate protection using perforin-mediated cytolysis following challenge. Collectively, qualitative details regarding the phenotype and functionality of Plasmodium-specific CD8 T cells have identified some important features that relate to protection but much remains to be learned to enhance anti- Plasmodial immunity following vaccination. Very little is known about how the specificity of the CD8 T cell response against Plasmodium impacts protection. The lack of defined CD8 T cell epitopes from Plasmodium has prevented the field from addressing this question in rodent models of malaria. However, using recombinant proteins or polyepitope strings, it is clear that CSP and TRAP proteins are targeted by protective CD8 T cell responses in rodents and humans (90, 96, 97, 134, 136, 151). But in the absence of any other defined epitopes than CSP in P. berghei and CSP in P. yoelii (96, 97), this question remained experimentally unaddressed. The Plasmodium genome contains approximately 5,000 open reading frames (142, 143), which complicates the identification of potential antigenic targets of protective CD8 T cells. However, many attempts have been made to identify CD8 T cell epitopes. Recently, three new CD8 T cell epitopes from Plasmodium berghei ANKA (Pb) have been described (136, 157). Thus, in addition to the wellstudied CSP in P. berghei, now an epitope derived from the well-described sporozoite surface protein TRAP (158) and the well-described intracellular protein GAP50 (159) have been described. Additionally, an epitope derived from a protein of 26
48 unknown function, identified as sporozoite-specific protein 20 (S20) in a sporozoite proteomic screen from P. yoelii sporozoites (160), was described. The putative S20 protein lacks a signal peptide sequence, thus suggesting an intracellular localization (160). Collectively, these novel epitopes in addition to the CSP 252 epitope can help address the question of whether specificity matters. If the specificity of the CD8 T cell response does matter, it will be important to screen potential antigen targets before inclusion in subunit vaccines in order to develop a high efficacy subunit vaccine for vaccination of humans against Plasmodium. Rationale and objectives for current studies Plasmodium infections lead to approximately 400,000 deaths worldwide annually. Although immunity can be engender with WSV approaches, the feasibility of these approaches for widescale vaccination provides challenges that may not be overcome. To lead to the eventual eradication of malaria, significant advances in subunit vaccinationmediated protection must be developed. These advances require a better understanding of the quantitiative and qualitative features of protective CD8 T cell responses targeting Plasmodium-infected hepatocytes and may additionally require a better understanding of humoral immunity targeting blood-stage parasites. To this end, I have studied the immunological correlates of protection against Plasmodium sporozoite infection following WSV and in a heterologous prime-boost platform to elucidate features of protective cellular immune responses to lead to the advancement of subunit vaccination of human subjects against Plasmodium. 27
49 Thesis objectives 1. Determine the immune mediators of protection against Plasmodium induced by WSV utilizing virulent sporozoites concurrently administered with CQ drug cover (CPS). 2. Determine the contribution to protection mediated by recently described CD8 T cell epitopes against liver-stage Plasmodium infection. 28
50 Figure 1. Plasmodium infection of hepatocytes and red blood cells defines the two distinct stages of the Plasmodium life cycle Plasmodium sporozoites are injected into the host during an infected mosquito bloodmeal. Sporozoites will traffic in the skin until they access the circulation and then quickly travel to the liver. Sporozoites will penetrate through the liver endothelial layer and enter into hepatocytes (A). Following entry into a hepatocyte, sporozoites may traverse through several neighboring hepatocytes (B) before infecting a hepatocyte, forming a PV around itself in the process (C). The PV-contained sporozoite will undergo early liver stage differentiation, followed by late liver-stage differentiation and replication to form thousands of merozoites contained within the PV (D). Merozoites are released from the PV into the circulation (E), enabling accessibility to red blood cells, which upon infection of red blood cells, begins the blood-stage of infection (F). 29
51 30
52 Figure 2. CD8 T cells and antibodies target Plasmodium antigens during the course of liver-stage and blood-stage infection Plasmodium infection can be inhibited by anti-parasitic CD8 T cells and antibodies. Sporozoites within the circulation can be targeted by anti-sporozoite antibodies (A), inhibiting sporozoite infection of hepatocytes. CD8 T cells targeting early liver-stageexpressed antigens (B) or late liver-stage-expressed antigens (C) can kill infected hepatocytes, preventing the release of PV-contained merozoites from being released from the liver into the circulation. Merozoites that are released into the circulation can be targeted by anti-blood-stage specific antibody responses, preventing red blood cell infection. 31
53 32
54 CHAPTER II: CD8 T CELL INDEPENDENT IMMUNITY AGAINST SINGLE DOSE CHEMOPROPHLYAXIS AND SPOROZOITE IMMUNIZATION AGAINST PLASMODIUM YOELII Abstract Sporozoite vaccination of both humans and rodents elicits potent anti-malarial immunity, but the dose of sporozoites and the number of immunizations required vary with vaccination approaches. Here I examine the immunological basis for superior protection afforded from single-dose vaccination with virulent sporozoites administered under prophylatic chloroquine-cover, referred to as chemoprophylaxis and sporozoites (CPS) vaccination, compared to the well-studied approach of administering radiationattenuated Plasmodium sporozoites (RAS). Earlier rodent studies utilizing CPS and RAS vaccination suggested a major role of CD8 T cells in reducing liver parasite burden after sporozoite challenge in a BALB/c mouse model. Consistent with this, I find that in C57Bl/6 mice CPS elicits substantially higher parasite-specific CD8 T cell responses than RAS vaccination and enhances immunity against P. yoelii infection. However, I show CPS-induced CD8 T cells are not necessary for protection following liver-stage sporozoite or blood-stage parasite challenge. Mechanistically, I found protection afforded from single-dose CPS is associated with low grade, transient parasitemia shortly following cessation of chloroquine treatment and generation of potent antibody responses to blood-stage parasites. Collectively, our data show the mechanistic basis for enhanced protective immunity against P. yoelli elicited by CPS in highly susceptible C57Bl/6 mice 33
55 is independent of CD8 T cells but rather require a humoral response. These studies may be relevant in understanding the potent immunity observed with CPS in humans. Introduction Plasmodium infection exacts a significant toll on human public health with more than 375,000 malaria-related deaths reported in 2010 (161). Anti-malarial vaccination represents an attractive intervention to break the cycle of disease transmission. Wholeparasite based approaches, specifically vaccination with radiation-attenuated sporozoites (RAS), have proven capable of generating immunity in humans (72). Despite this success, RAS induced protection appears to require immunization with very large numbers of parasites (>1000 bites from mosquitoes harboring RAS (72)) and needle delivered RAS requires 4-5 high-dose I.V. administrations to achieve protection (77, 78). Another approach first described in rodents (chemoprophylaxis and sporozoites, CPS) (88, ) also elicits protection against subsequent sporozoite exposure in human subjects (84, 85). In this approach, human subjects receive mosquito bite inoculation of virulent P. falciparum sporozoites while concurrently undergoing chloroquine (CQ) chemoprophylaxis (84, 85). CQ administration interrupts the capacity of blood-stage parasites to detoxify heme by preventing the polymerization of heme into hemazoin in red blood cells (89). Eventually, buildup of heme is toxic to the parasite and the red blood cell. Importantly, this CPS approach required fewer mosquito bites (~36-45 bites over 3 exposures) to elicit full protective immunity (84, 85). Thus, in humans CPS appears to induce much more potent immunity compared to RAS vaccination. 34
56 Protection afforded from whole-sporozoite vaccinations, such as CPS and RAS, is reported to involve liver-stage directed CD8 T cells (81, 88, 99, 165). For example, in a rodent model of CPS whereby BALB/c mice were given a single dose of 10 5 virulent P. yoelii 265BY sporozoites followed by 10 consecutive days of CQ chemoprophylaxis, reduction in liver parasite burden after challenge 15 days later involved CD8 T cells, IFN-γ and NO - as the primary immune effectors (88). Similarly, CPS-induced protection in humans correlates with CD8 and CD4 T cells producing effector cytokines (85). In rodent models of RAS immunization, protection is critically linked to CD8 T cells exhibiting activity against the liver-stage of infection (98). Collectively, these results highlight that CD8 T cell-mediated liver-stage protection can be achieved following whole-sporozoite vaccination approaches, such as CPS or RAS. Although protection in rodents and humans receiving attenuated whole-sporozoite vaccination is associated with CD8 T cells against liver-stage antigens, it remains unclear how a single dose of CPS can afford immunity in rodents whereas multiple, high-doses of RAS are required (88). These two whole-sporozoite vaccination approaches differ in that RAS vaccination results in only transient, non-replicative infection of hepatocytes, whereas CPS using chloroquine (CQ) allows for productive infection of hepatocytes, release of merozoites and infection of red blood cells (RBC). Due to the blood-stage specific inhibitory effects of CQ (87, 164), merozoites are unable to undergo further rounds of replication in RBC. Thus, critical differences in antigen load, and antigen targets may lead to differences in the protective T cell response and/or humoral 35
57 responses, which may underlie the exceedingly potent immunity induced by CPS compared to RAS. CPS vaccination of BALB/c mice showed that CD4 T cells, in addition to CD8 T cells, contribute to protection from sporozoite challenge (88). Therefore, it is possible that superior protection from CPS vaccination relative to RAS may be due to the development of anti-parasitic responses by immune mediators besides CD8 T cells. In one study, the administration of blood-stage parasites concurrently with CQ in BALB/c mice lead to the development of cross-stage reactive (liver- and blood-stage reactive) antibodies (166). Although administration of blood-stage parasites concurrent with CQ differs from sporozoites plus CQ, both immunization approaches result in exposure to blood-stage parasite in a controlled matter. Thus, the generation of cross-stage reactive antibodies may result from both sporozoite and blood-stage parasite immunizations concurrent with CQ. Therefore, it is possible that anti-plasmodium CD4 T cell and cross-stage reactive antibodies are contributing to superior protection afforded by CPS vaccination compared to RAS vaccination. Although the widespread prevalence of CQ-resistant P. falciparum and P. vivax complicates direct clinical application of the CPS vaccination approach (6, 89, 167), protection elicited by CPS platforms in human subjects further underscores the potential for whole sporozoite approaches to elucidate the cellular and immunologic requirements for successful anti-malarial vaccination. At a minimum, experimental CPS may directly aid identification of both host and parasite-specific factors that determine high levels of protective anti-plasmodial immunity. Thus, understanding the immunological 36
58 mechanisms that underlie enhanced immunity following low-dose CPS would fill a critically important knowledge gap. Here, I analyzed the immunological basis of superior immunity induced by CPS compared to RAS vaccination in a stringent parasite-host model. Materials and Methods Mice and immunizations Female 6-8 week old C57BL/6 and BALB/c mice were purchased from the National Cancer Institute (Frederick, MD) and housed at the University of Iowa animal care unit at the appropriate biosafety level. C57BL/6 µs-aid -/- mice deficient in the immunoglobin heavy-chain µ-chain secretory domain and activation-induced cystidine deaminase (168), were a kind gift from F. Lund (University of Alabama, Birmingham) and were bred at the University of Iowa. BALB/c Thy1.1/1.2 + CS 280 T cell receptor transgenic mice (169) were a kind gift from F. Zavala (Johns Hopkins University, Baltimore, MA). The Institution Animal Care and Use Committee approved animal experiments. P. yoelii 17XNL (Py) sporozoites were isolated from the salivary glands of infected A. stephensi mosquitoes obtained from New York University insectary and radiation attenuated by exposure to 200 Gy (20,000 rads). Mice were vaccinated I.V. or S.C. with 10,000 radiation-attenuated or virulent sporozoites. Mice vaccinated with virulent sporozoites (CPS vaccination) were given 10, 25, or 30 daily I.P. injections of 37
59 100 µl (8mg/mL) chloroquine (CQ) diphosphate salt (Sigma, St. Louis, MO) in PBS from d0-d9, d0-d24 or d0-d29, respectively. Plasmodium parasites Naïve and immunized mice were challenged I.V. >60 days post-immunization with the indicated number of virulent Py sporozoites or 10 6 Py-infected red blood cells. Pyinfected red blood cells were prepared by harvesting blood from Py-blood-stage-infected donor mouse. Giemsa stain of thin blood smear from donor blood was used to determine parasite density (%parasitemia) and hemocytometer was used to determine #RBC/unit volume. Py-blood-stage-infected blood was mixed with 2 volumes of freezing solution (9 parts Alzevers Solution (Sigma): 1 part Glycerol (Sigma)). Vials of quantified donor blood in freezing solution were flash frozen in liquid nitrogen, and stored in liquid nitrogen until thawed, and resuspended in PBS to achieve 10 6 Py-infected RBCs/200uL. Low-grade, transient, parasitemia following CQ drug cessation was quantified by evaluating Giemsa stain of thin blood smear from day by examining at least 20 fields containing at least 150 red blood cells. Naïve mice used for challenge studies received ten daily injections of chloroquine >50 days prior to challenge to control for potential residual drug activity. Patent parasitemia following challenge was evaluated by Giemsa stain of thin blood smear from day 4-10 post-challenge. Protection is defined as the absence of detectable blood-stage parasites at all time-points examined. At least 20 fields containing at least 150 red blood cells were examined for each mouse designated as exhibiting protection. Mice exhibiting less than 5% parasitemia at all time-points 38
60 examined, and never for more than four consecutive days of detectable parasitemia were classified as resistant. Detection of parasite-specific CD8 and CD4 T cells in blood Vaccine-induced CD8 T cell populations were identified by staining cells isolated from spleen or peripheral blood with anti-cd8α (53-6.7; Biolegend, San Diego, CA) and anti- CD11a (M17/4; Biolegend) antibodies. Antigen-specific CD8 T cells were described as as CD8α lo and CD11a hi (170, 171). Vaccine-induced CD4 T cell populations were identified by staining cells isolated from spleen or peripheral blood with anti-cd4 (H129.12, Biolegend), anti-cd11a, and anti-cd49d (R1-2, Biolegend) (172). Antigenspecific CD4 T cells were described as CD4 + CD11a + CD49d +. Cells were analyzed using a BD FACSCanto or BD LSR Fortessa and data were analyzed using FLOWJO Software (Tree Star, Inc, Ashland, OR). Animals were pre-bled prior to vaccination to establish individual background of circulating CD8α lo CD11a hi or CD4 + CD11a + CD49d + cell frequencies. Listeria monocytogenes infections Mice were administered 1-2 x 10 7 CFUs of attenuated recombinant LM expressing ovalbumin (OVA)(173). Mice were given 7 daily I.P. injections of CQ. LM-OVA infection induced CD8 T cell populations were identified on day 7 post infection by 39
61 staining single-cell suspension of splenocytes with anti-cd8α antibody and MHC class I OVA tetramers. Tetramer preparation was performed in house. Adoptive transfer of Thy1.1/1.2 + CS 280 TCR transgenic CD8 T cells Thy1.1/1.2 + CS 280 T cell receptor (TCR) transgenic (Tg) CD8 T cells, specific for the CS epitope of P. yoelii circumsporozoite protein (174), were obtained from the PBL or spleen of naïve donors and the number of input Tg cells was determined by staining for CD8α and Thy1.1/1.2 + (clones , 0X-7, Biolegend, respectively) as previously described (175). Approximately 10 3 Thy1.1/1.2 + CS 280 TCR Tg CD8 T cells were transferred into naïve Thy1.2 + BALB/c mice at day -1. The following day, recipient mice were administered CPS or RAS vaccination. Six days later the frequency and total number of splenic RAS- or CPS-induced CD8α lo CD11a hi CS 280 TCR Tg CD8 T cells was determined. T cell depletion Mice were depleted of CD8 T cells or CD4 T cells by two I.P. injections of 50µg or 200µg anti-cd8 (2.43; BioXCell, West Lebanon, NH) or 200µg anti-cd4 (GK1.5; BioXCell). Depletion was confirmed in tissues for preliminary experiments or prior to challenge by staining PBLs with anti-cd8 and anti-cd4 (clones and H129.12, respectively). 40
62 Blood-stage reactive and anti-msp1 antibody detection Mice were administered 10,000 Py sporozoites with 10 or 25 days of daily I.P. CQ injections, and serum was collected >40 days post immunization. Dilutions of serum (prepared as described (176)) were allowed to react to Py-parasitized red blood cell lysate (prepared as described (177)) or recombinant MSP1 protein (BEI Resources, Manassas, VA) immobilized onto Maxisorb Immunoplates (Nunc-Immuno, Rockford, IL). Total IgG antibodies were detected with horseradish peroxidase-conjugated goat anti-mouse IgG (Jackson ImmunoResearch, Pike Grove, PA) and tetramethylbenzidine substrate (Sigma). Results are presented as absorbance (450 nm) as a function of serum dilution. Liver parasite burden Mice were administered 10,000 Py sporozoites with or without 10 days of daily I.P. CQ injections. Naïve mice were administered 10 days of daily I.P. CQ. Mice were challenged >60 days after immunization with 40,000 Py sporozoites. Liver RNA was extracted at 44 hours post challenge using TRIzol (Invitrogen) and one-step quantitative real-time reverse transcription PCR analysis for P. yoelii 18S rrna was performed on 2µg of liver RNA. P. yoelii 18S rrna was detected using 5 - GGGGATTGGTTTTGACGTTTTTGCG-3 (forward primer), 5 - AAGCATTAAATAAAGCGAATACATCCTTAT-3 (reverse primer), and 5 -FAM- CAATTGGTTTACCTTTTGCTCTTT-TAMRA-3 (probe), which generates a 133-bp 41
63 fragment (178). Data were normalized to GAPDH control Ct value and infected naïve control Ct values by calculating 2^(-ΔΔCt). Data plotted as % parasite burden +/- S.E.M. with values normalized relative to infected naïve controls. Detection of persisting parasites under CQ treatment Mice were administered 10,000 Py sporozoites with or without 10 days of daily I.P. CQ injections. Naïve mice were administered 10 days of daily I.P. CQ. Mice were euthanized and spleens were excised on day 9 post-immunization (on last day of CQ treatment). Splenic RNA was extracted using TRIzol (Invitrogen) for quantitative realtime PCR analysis for P. yoelii 18S rrna using primers described above. 18s rrna copy numbers was estimated using a standard curve generated from known amounts of plasmid copy containing the identified 18s DNA insert. Statistical Analysis Data were analyzed using Prism4 or Prism6 software. Specific tests of statistical significance are detailed in figure legends and table footnotes. 42
64 Results CPS is associated with significantly larger anti-plasmodial CD8 and CD4 T cell responses and enhanced protection from challenge compared to RAS vaccination Whole sporozoite vaccination (WSV) models in rodents have been used to study the requirements for protective liver-stage immunity against stringent sporozoite challenge (83, 104, 108, ). For example, RAS-vaccinated C57Bl/6 mice are more difficult to protect than RAS-vaccinated BALB/c against Plasmodium yoelii 17XNL (Py) infection, with the former requiring additional sporozoite immunizations to elicit similar protection (98, 99). Similarly, CPS vaccination was more potent in inducing protection than RAS in BALB/c mice (88). Because several studies have shown that protection mediated by WSV approaches is dependent on CD8 T cell responses against liver-stage antigens (94-98, 183), I sought to evaluate whether RAS versus CPS vaccination of C57Bl/6 mice led to quantitative differences in WSV-induced CD8 T cell responses. Due to the lack of defined H-2 b -restricted P. yoelii CD8 T cell epitopes at the initiation of this study, I examined the activation of the CD8 T cell response using a CD8 T cell surrogate activation marker approach, developed in our laboratory (170, 171), which allowed for the measurement of the magnitude and kinetics of the total CD8 T cell response to vaccination in the peripheral blood of individual mice without a priori knowledge of MHC-restriction, antigen-specificity, or precise epitopes. This method relies on the stable changes in surface marker expression of CD8α and CD11a on CD8 T cells in response to T cell receptor mediated activation following infection or immunization in vivo (171). Thus, naïve CD8 T cells are CD8α hi intermediate (int), CD11a 43
65 whereas antigen-specific effector and memory CD8 T cells are CD8α lo, CD11a hi (170, 171). Naïve C57Bl/6 mice have low frequencies of CD8α lo, CD11a hi T cells at d0, prior to vaccination (Figure 3A-B). Following CPS or RAS vaccination wherein mice receive I.V. administration of 10 4 Py sporozoites with concurrent CQ drug cover (CPS) or radiation attenuated sporozoites (RAS), the total vaccine-induced CD8 T cell response in CPS vaccinated mice measured from PBL, was significantly larger in frequency at both effector (p < 0.001, d6) and memory (p < 0.05, d47) time points compared to RAS vaccination (Figure 3B). Additionally, I also measured the magnitude of the total vaccine-induced CD4 T cell response following RAS versus CPS vaccination through CD4 T cell surrogate activation markers, which relies on the stable change in upregulated surface marker expression of CD49d and CD11a on CD4 T cells in response to T cell receptor mediated activation following infection or immunization in vivo (172). Similar to CD8 T cells, CPS vaccinated mice had significantly larger CD4 T cell responses from PBL at both effector (p < 0.001, d6) and memory (p < 0.05, d47) time points compared to RAS vaccinated mice (Figure 4B). Together, these data indicate that CPS vaccination of C57Bl/6 mice results in larger total activated CD8 and CD4 T cell responses at effector and memory time points relative to RAS vaccination. Importantly, CQ is known to inhibit the acidification of endosomes (8, 9), which may potentially influence antigen presentation following vaccination of mice with either CPS or RAS. However, in parallel experiments I determined that daily administration of CQ to RAS vaccinated mice altered neither the magnitude nor kinetics of the total CD8 T cell response (Figure 5A-B) nor the total CD4 T cell response (Figure 5C-D) as measured 44
66 by surrogate activation markers. Further, I evaluated whether CQ administration alters the magnitude of an antigen-specific CD8 T cell response in another infectious disease system by infecting naïve C57Bl/6 mice with attenuated recombinant Listeria monocytogenes expressing ovalbumin concurrent with daily injections of CQ. Analysis of tetramer staining at the peak of the OVA-specific CD8 T cell response (d7) showed no significant differences in the magnitude of the OVA-specific CD8 T cell response in mice administered CQ treatment (Figure 5E). Taken together, these data indicate that CPS vaccination induces larger magnitude Plasmodium-specific CD8 T and CD4 T cell responses compared to RAS vaccination, and the use of CQ does not impact the magnitude nor kinetics of these responses. Vaccination of human subjects is typically administered using subcutaneous (S.C.) or intramuscular (I.M.) routes, and not via I.V. administration. However, to date human subjects administered RAS vaccination via S.C. or I.M. routes were not protected following sporozoite challenge but were protected if the vaccination were administered I.V. or via the bites of hundreds of infected mosquitoes (71, 72, 76, 78). To address whether the route of administration alters the magnitude of the CD8 and CD4 T cell response, and thus affect protective immunity, mice were I.V. or S.C. administered either RAS or CPS vaccination and the kinetics of the total activated CD8 and CD4 T cell response were tracked longitudinally in the PBL via surrogate activation markers. Administration of CPS vaccination via the I.V. route lead to significantly increased CD8 T cells responses at the peak of the effector responses (p < 0.001, d6-8), but not at an early memory time point (d38)(figure 6A-B). In contrast, administration of RAS 45
67 vaccination via I.V. versus S.C. route lead to significantly enhanced CD8 T cell responses at the peak effector response (p < or 0.01 for d6 or d8, respectively), as well as an early memory time point (p < 0.01, d38) (Figure 6C-D). Taken together, these data indicate that I.V. administration leads to larger effector CD8 T cell responses following CPS or RAS vaccination, however this difference is only maintained into memory time points in mice receiving RAS vaccination. Analysis of the total activated CD4 T cell response following I.V. versus S.C. CPS vaccination lead to similar results as shown with the total activated CD8 T cell response in that the peak effector CD4 T cell responses (d6-8) were significantly larger in I.V. administered mice (p < 0.001), but not enhanced at an early memory time point (d47)(figure 7A-B). Mice administered I.V. RAS vaccination had significantly greater total activated CD4 T cell responses in the PBL at effector time points (p < 0.01, d6-8), and early memory time point (p < 0.01, d47)(figure 7C-D). Taken together, these results indicate that I.V. administrations of CPS and RAS vaccination lead to enhanced CD8 and CD4 T cell responses at effector time points relative to RAS vaccination, but significant differences in the magnitude of the T cell responses occurs at memory time points only in mice administered RAS vaccination. To understand whether the magnitude Plasmodium-specific CD8 T cell responses correlates with enhanced protective immunity in a stringent parasite-host model, naive C57Bl/6 mice subjected to I.V. or S.C. administration of CPS or RAS vaccination were challenged >60 days after immunization by I.V. injection of 10 3 virulent Py sporozoites (~300x the ID 50 ). Challenged mice were evaluated from 4-10 days post challenge for 46
68 evidence of blood-stage parasites by Giemsa-stained blood smear. All naive control mice, whether previously treated with CQ or not, exhibited blood-stage parasitemia, a single I.V. administration of CPS under 10 days of CQ treatment induced protection (no evidence of blood-stage parasites) in 72% of mice, whereas mice receiving a single I.V. administration of RAS vaccination exhibited no protection (Table I). Parallel results were observed for mice immunized via the S.C. route in that S.C. administration of CPS vaccination afforded protected in 47% of mice, whereas S.C. administration of RAS vaccination lead to 0% of mice protection (Table I). Thus, single administration CPS, whether I.V. or S.C. delivered, can induce sterilizing protection against sporozoite challenge at a memory time point even in a stringent C57Bl/6-Py model, and this protection correlates with enhanced CD8 and CD4 T cell responses relative to RAS vaccinated mice (Figures 6-7). CPS vaccination induces CD8 T cells against a broader spectrum of liver-stage antigens compared to RAS vaccination Superior protection following single administration CPS vaccination correlates with larger CD8 and CD4 T cell responses compared to RAS vaccination. Because CD8 T cells targeting liver-stage antigens have been shown to be important mediators of protection against liver-stage infection, I sought to determine if the enhanced magnitude of CD8 T cells and protection due to CPS vaccination was due to the induction of a CD8 T cell response targeting a broader spectrum of liver-stage antigens compared to RAS vaccination. To address this, I utilized the BALB/c Thy1.1/1.2 + CD8 T cell CS 280 T cell 47
69 receptor (TCR) transgenic (Tg) mouse, which allowed me to track a H-2K d -restricted CD8 T cell response. This mouse strain only produces CD8 T cells that recognize the CS epitope contained with the early liver-stage expressed circumsporozoite protein (CSP) of P. yoelii sporozoites (169). Naïve Thy1.2 + BALB/c recipient mice were administered 10 3 Thy1.1/1.2 + CS 280 TCR Tg cells I.V. and one day later administered either CPS or RAS vaccination (Figure 8A). The total splenic activated CD8 T cell response was measured six days later in vaccinated recipient mice. As observed in C57Bl/6 hosts, the magnitude of the total activated CD8 T cell response in CPS vaccinated BALB/c mice was significantly larger than RAS vaccinated mice (p < 0.01, Figure 8B). Analysis of splenic Thy1.1/1.2 + CS 280 TCR Tg cells showed no difference in total number (Figure 8C), indicating that CPS and RAS vaccinations induce similar numbers of CS 280 TCR Tg cells, which target an early liver-stage expressed protein that should be equivalently expressed in viable sporozoites and RAS. Enumeration of total number splenic activated CD8 T cells via surrogate activation markers corroborate frequency data in that CPS vaccination induces significantly larger activated CD8 T cell responses in the spleen (p < 0.01, Figure 8D), suggesting the differences observed in Figure 8B and 8D are due to CD8 T cells targeting antigens besides the early liver-stage, immunodominant CSP antigen. Taken together, these data suggest that larger magnitude CD8 T cell responses following CPS vaccination are due to induction of CD8 T cells targeting a broader spectrum of antigens. 48
70 CPS-induced CD8 T cells are not required for protection from sporozoite challenge It has been argued that CD8 T cells mediate CPS-induced protection in humans and BALB/c mice (85, 88, 184). I evaluated the requirement for CPS-induced CD8 T cells in mediating protection against a sporozoite (liver-stage) challenge in C57Bl/6 mice by antibody-mediated depletion of CD8 T cells at a memory time point prior to challenge. Groups of mice were given a single administration of CPS and >60 days later were administered 50µg of anti-cd8 depleting antibody twice over three days. Successful depletion was verified by analyzing blood, spleen and liver. This treatment resulted in >98.5% CD8 T cell depletion in all tissues examined and provided a guide to determine the concentration of anti-cd8 depleting antibody required for complete CD8 T cell depletion in relevant tissues (Figure 9A). A different group of mice received a single administration of CPS and >60 days later were administered 200µg of anti-cd8 antibody one day prior and one day after sporozoite challenge to ensure depletion of CD8 T cells in peripheral tissues. CPS vaccinated mice that were CD8 T cell depleted or received control rat IgG treatment were challenged with 10 3 Py sporozoites. In contrast to previous studies (81, 99), CD8 T cell depletion did not decrease the frequency of protected mice, compared to non-depleted control mice (Figure 9B). However, CPS mice that were not protected (~25% in each group) exhibited substantial control of infection compared to naïve control mice that develop parasitemia lasting >25 days. These CPS mice were categorized as resistant if parasitemia did not exceed 5% or was cleared below the level of detection (Giemsa-stained blood smear negative) by 10 days after challenge. Thus, in a stringent host-parasite model of protection, CD8 T cells induced by 49
71 single administration CPS are not required to provide protection or substantial resistance against sporozoite challenge. To determine if a liver-stage specific protective role for CPS-induced CD8 T cells could be documented, I measured Py-specific 18s rrna in the liver after a high dose sporozoite challenge in CD8-depleted and non-depleted CPS mice. Mice were given a single CPS administration and depleted of CD8 T cells or given rat IgG control prior to challenge with 40,000 Py sporozoites (Figure 9C). Parasite burdens were modestly reduced in CPS mice with this high-dose challenge, however, depletion of CD8 T cells did not significantly increase liver Py-specific 18s rrna compared to non-depleted controls (Figure 9C). Collectively, these data demonstrate that single administration CPS-induced CD8 T cells are not required to achieve immunity against P. yoelii sporozoite challenge of C57Bl/6 mice. CPS-induced protection is associated with transient blood-stage parasite exposure CPS allows for complete liver-stage development and merozoite entry into red blood cells before infection is truncated by the blood-stage-specific inhibitory effects of CQ (87, 88, 164). In a study where human subjects were administered CPS vaccination, Plasmodium-specific nucleic acids, but not patent parasitemia, were detectable for several days in the blood of all ten patients following the first vaccination (85). Further, when patients received second and third CPS administrations, 6 and 3 patients had detectable Plasmodium-specific nucleic acid in the blood, respectively (85). Thus, it is possible that 50
72 CQ chemoprophylaxis allows for a sustained subpatent (not detectable by blood smear) blood-stage infection, which may potentiate the generation of blood-stage-specific cellular and humoral immune responses that contribute to protection. I evaluated single administration CPS in C57Bl/6 mice for evidence of sustained subpatent blood-stage infection by testing for Py-specific 18s rrna in the spleen on the last day (d9) of CQ chemoprophylaxis (178). Despite CQ treatment, Py-specific 18s rrna was above the limit of detection in 60% of the CPS mice (Figure 10A). The presence of Py-specific RNA suggests that metabolically active, viable blood-stage parasites may persist for the duration of the 10 day CQ chemoprophylaxis. CD4 T cells are a critical cellular component necessary for control of blood-stage parasite replication and persistence (185, 186). To further address a role for persisting blood-stage parasites, naïve mice undergoing single administration CPS were immunocompromised by CD4 T cell depletion from day 0 through day 9, post sporozoite infection (Figure 10B), and subsequently monitored for the appearance of blood-stage parasites by Giemsa-stained blood smear. Following cessation of CQ chemoprophylaxis, CD4 T cell depleted CPS mice had uncontrolled blood-stage infection (Figure 10C). Strikingly, all CPS immunocompetent (no depletion) mice also exhibited detectable, albeit very low levels of blood-stage parasitemia following CQ cessation (Figure 10C). Higher resolution analyses revealed that approximately 3-5 days following cessation of CQ chemoprophylaxis all mice exhibited patent (albeit low-grade) parasitemia (Figure 10D). The duration of low-grade, transient, parasitemia was between 3-6 days, and ranged from slightly above the limit of detection (greater or equal to 1 parasite out of 51
73 3000 red blood cells, 0.033% parasitemia) to 5% parasitemia (Figure 10E). By day 7 after cessation of CQ, all CPS mice cleared the infection without requirement for further intervention. Taken together, these results indicate that after single administration CPS, viable blood-stage parasites persist in C57Bl/6 mice following ten consecutive days of CQ chemoprophylaxis. CPS-induced protection is associated with induction of blood-stage reactive antibodies Low-grade, transient parasitemia after CQ cessation in single administration CPS mice may increase the duration or amount of exposure to blood-stage antigens and contribute to protective immunity. I next tested if abrogating low-grade patent parasitemia decreased protection in single administration CPS mice. Mice received CPS with 10, 15, 20, or 25 days of CQ chemoprophylaxis. I observed a correlation in the reduction of mice exhibiting breakthrough parasitemia following CQ cessation as the duration of CQ treatment increased (Figure 11A). Importantly, all 10 day CQ mice exhibited low-grade, transient, parasitemia after CQ withdrawal, however 25 day treated CQ mice did not exhibit patency after cessation of drug treatment (Figure 11A). Both 10 day and 25 days CQ treatment groups were challenged >60 days after start of immunization (>35 days after end of CQ treatment). CPS mice exhibiting parasitemia after CQ cessation exhibited greater than 65% sterile protection (Figure 11B). Conversely, single administration CPS mice treated for 25 days with CQ, which did not exhibit low-grade, transient, parasitemia after CQ cessation, were not sterilely protected. 52
74 Nevertheless, these mice still exhibited control of infection and were thus, relatively resistant to challenge (Figure 11B). It is well established that antibodies are elicited against asexual blood-stage parasites during Py blood-stage infection (187, 188). Since CPS-induced protection does not require CD8 T cells, but is associated with multi-day exposure to blood-stage infection (control of which depends on CD4 T cells (Figure 10C)), I hypothesized that CPS may elicit antibodies reactive against blood-stage antigens that overcomes specific requirements for CD8 T cells in protection. To evaluate this, I determined levels of total IgG against parasitized red blood cell lysate in serum from CPS mice experiencing lowgrade, transient parasitemia compared to no parasitemia CPS mice. As expected, both vaccination groups had detectable levels of total IgG against blood-stage antigens. However, serum from mice that had experienced low-grade, transient parasitemia after CPS contained significantly higher titers of blood-stage parasite-specific IgG (p < 0.01, Figure 12A). Additionally, I determined levels of total IgG against a defined blood-stage antigen, merozoite surface protein 1 (MSP1), in serum from CPS mice experiencing lowgrade, transient parasitemia (CPS-10d CQ) compared to no parasitemia CPS mice (CPS- 25d CQ). Similar to total IgG responses against parasitized red blood cell lysate, both vaccination groups had detectable levels of total IgG against MSP1 (Figure 12B). Although 10 day CQ treated mice had higher detectable responses, the difference between 10 day and 25 day CQ treatment mice was not statistically significant. Next, I determined if CPS-induced antibodies were required for control of blood-stage parasites. To address this question, I utilized µs-aicda -/- mice on a C57Bl/6 background, as these 53
75 mice lack the ability to secrete functional antibodies, but still maintain B cells (168). Naïve µs-aicda -/- mice and wild-type C57Bl/6 mice were vaccinated with 10 4 virulent Py sporozoites with 10 days of CQ chemoprophylaxis. After cessation of CQ chemoprophylaxis, µs-aicda -/- mice exhibited uncontrolled parasitemia and all eventually succumbed to hyperparasitemia (days 35-60). Thus, I administered days of CQ to µs-aicda -/- mice in an attempt to prevent the occurrence of parasitemia. In 7 out of 15 of the µs-aicda -/- mice administered CPS with days of CQ, parasitemia still occurred after CQ cessation, and these mice all eventually succumbed to hyperparasitemia (Figure 12C). These results demonstrate the ability of Py blood-stage parasites to persist for a long duration under CQ chemoprophylaxis specifically in the absence of secreted antibodies. Further, as an extension for the requirement for CD4 T cells to control low-grade, transient parasitemia (Figure 10C), antibodies appear to be necessary to control the low-grade, transient parasitemia that occurs after CQ treatment cessation. I challenged the 8 surviving vaccinated µs-aicda -/- mice with 10 3 virulent Py sporozoites. All 8 challenged CPS µs-aicda -/- mice developed patent parasitemia and 7 out of 8 succumbed following the challenge. Taken together, these data suggest CPS vaccination elicits antibodies reactive against blood-stage antigens, which are critical for the control of low-grade, transient parasitemia after CQ cessation, and protection following sporozoite challenge. 54
76 CPS confers protection against blood-stage challenge Our results demonstrate that CPS induces antibodies reactive against blood-stage antigens and does not require CD8 T cells for protection against liver-stage, sporozoite challenge in the Py model of infection in C57Bl/6 mice. To further define the role of antibodies in CPS-induced protection, mice were administered 10 4 sporozoites with 10 days of CQ chemoprophylaxis, to permit low-grade, transient parasitemia after CQ cessation and induction of higher blood-stage reactive antibody titers. Greater than 60 days later, some mice were depleted of both CD8 and CD4 T cells prior to Py challenge. T cell depleted and control mice were then challenged with 10 6 Py parasitized red blood cells, which resulted in patent parasitemia in all non-depleted, naïve mice. Strikingly, T cell depletion did not reduce the fraction of CPS mice exhibiting immunity or resistance to this blood-stage challenge compared to control IgG treated CPS mice (Figure 13). Taken together, these data indicate that neither CD4 nor CD8 T cells are required for protection against blood-stage challenge in this model of single administration CPS in C57Bl/6 mice. Collectively, our results suggest that CPS-induced antibodies are the primary mediator of protection acting primarily to blood-stage infection. Discussion CPS vaccination elicits superior protection compared to RAS vaccination in human and rodents (72, 85, 88). However, the precise mechanisms of this superior protection are unknown. To address this knowledge gap, I used a stringent parasite/mouse model to best understand the requirements to achieve sterililizing 55
77 protection (no evidence of blood-stage parasites via blood smear after sporozoite challenge). The C57Bl/6-Py 17XNL model is a stringent model to evaluate protection since this parasite/host model is harder to protect following sporozoite challenge compared to BALB/c mice challenged with Py or P. berghei (81, 98, 99). Unexpectedly, I found evidence for patent blood-stage infection after cessation of CQ chemoprophylaxis in the CPS mice, permitting not only protection from sporozoite challenge, but also protection from blood-stage parasite challenge. Whole-sporozoite vaccine-induced, liver-stage directed CD8 T cells are required for protection in multiple rodent models. This requirement applies to both RAS (98, 99, 165) and genetically attenuated parasite (GAP) vaccination (81). Thus, as expected, CPS also induces CD8 T cells that are capable of reducing the liver parasite burden. Indeed, in a BALB/c-P. yoelii 265BY model of CPS, CD8 T cells, IFN-γ and NO - were the primary immune effectors of protection (88, 165). Similarly, in a C57Bl/6 model using P. berghei ANKA sporozoites, protection from challenge was correlated with the amount of IFN-γ produced by memory CD8 T cells (165). In accordance with rodent studies, CPS-induced protection in humans correlates with the presence of T cells producing effector cytokines (85). Collectively, these results highlight that significant CD8 T cellmediated liver-stage protection can be achieved following whole-sporozoite vaccination approaches, such as CPS, GAP or RAS. Previously, it was shown that protection against sporozoite challenge correlates with larger vaccine-induced effector and memory CD8 T cell responses (81). The larger magnitude of CD8 T cell response was a result of additional liver-stage antigens 56
78 expressed during genetically attenuated parasite (GAP) vaccination compared to RAS. Surprisingly, I found that despite significantly larger effector and memory CD8 T cells induced from I.V. delivered CPS compared to RAS vaccination (p < 0.01), CD8 T cells were not required for protection from sporozoite challenge in the C57Bl/6 model of Py 17XNL CPS. RAS and GAP vaccination require multiple immunizations to achieve CD8 T cell mediated protection from challenge in the C57Bl/6-Py17XNL model (81). For instance, in Py fabb/f -/- GAP vaccination, wherein the parasite is blocked late in liverstage differentiation, a single immunization with 2 x 10 4 attenuated sporozoites failed to protect C57Bl/6 mice from sporozoite challenge (81). These results highlight the requirement for numerically boosting RAS- and GAP-induced CD8 T cell responses to provide protective immunity against sporozoite challenge. In the C57Bl/6-Py 17XNL CPS model, a single immunization dose elicited protective immunity, but this was dependent on low-grade, transient parasitemia after cessation of CQ treatment. Thus, an explanation for the larger CD8 T cell response from CPS may be exposure to blood-stage antigens. Several studies suggest that CD8 T cells are induced from exposure to bloodstage parasites (18, 189). The specificity of the CPS-induced effector and memory CD8 T cell responses is unknown since both liver- and blood-stage antigens are expressed during immunization. However, our data indicate in the C57Bl/6-Py 17XNL model of CPS, CD8 T cells are not required for immunity against liver-stage or blood-stage challenge. Therefore, our results suggest other component(s) of the immune response elicited by CPS may be primarily responsible for immunity. It is possible the CPSinduced CD4 T cells could contribute to reducing liver-stage parasites (88). Indeed, both 57
79 I.V. and S.C. CPS vaccination induced larger magnitude CD8 and CD4 T cell responses than RAS vaccination. Although CD4 T cells were required during the early stage of vaccination, likely to help in the induction of antibody responses, neither memory CD4 nor CD8 T cells were required for protection against blood-stage challenge, highlighting that the potent blood-stage immunity I observe is independent of memory T cells. Sporozoites administered with CQ chemoprophylaxis are competent to undergo a full program of amplification and differentiation in the liver and are released from the liver to undergo one round of infection in the blood as blood-stage merozoites. Due to the inhibitory effects of CQ, replication and subsequent infection is inhibited by CQinduced toxicity to the RBC and parasite (87, 164). In the C57Bl/6-Py 17XNL CPS model, I found that CQ chemoprophylaxis does not completely eliminate the parasites during the 10 day course of treatment. Unexpectedly, I show subpatent parasites persist during CQ chemoprophylaxis, which can replicate to patent levels once CQ treatment ends. Further, multiple days (3-6 days) of patent, low-grade parasitemia were required for the induction of immunity. Moreover, depletion of CD8 and CD4 T cells prior to blood-stage challenge did not abrogate CPS-induced protection. Low-grade, transient parasitemia during CPS with CQ chemoprophylaxis is not limited to rodent studies, but has also been observed in human CPS studies (84, 85). Indeed, in these two studies, PCR analysis of the blood revealed the majority of patients had subpatent infection following the first administration of CPS. Interestingly, in one study, 2 of the 15 human subjects undergoing CPS were reported to have a positive thick blood smear on day 7 and received atovaquone/proguanil treatment before continuation in the study (84). The 58
80 mechanism of blood-stage parasite persistence during CQ chemoprophylaxis in humans and rodents is unknown. CQ interrupts the capacity of merozoites to detoxify heme by preventing the polymerization of heme into hemazoin in red blood cells (89). Recently one study found evidence that rodent blood-stage parasites can persist in plasmocytoid dendritic cells (pdcs) (190). Persistence in a non-red blood cell is a potential mechanism whereby Plasmodia may avoid the impact of CQ chemoprophylaxis. It remains to be determined whether human Plasmodium blood-stage parasites can similarly persist in pdcs, but such persistence could provide an explanation for patent parasitemia observed following the cessation of CQ treatment in rodents. In the C57Bl/6-Py 17XNL model of CPS, I demonstrate the requirement of lowgrade, transient parasitemia in induction of protective blood-stage directed antibody responses to provide protection. Since my initial description of transient parasitemia in a rodent CPS vaccination model, transient parasitemia has been observed and correlated with sterilizing immunity in another rodent model of malaria (C57Bl/6-P. chabaudi AS) (191), providing further support for a mechanism wherein parasites can persist under CQ cover in rodents thereby potentiating immunity. Althought the role of subpatent, or lowgrade, transient parasitemia, in the human studies was not further investigated, it is possible low-grade blood-stage infections could elicit blood-stage immunity contributing to the potent immunity induced by CPS vaccination in human subjects. Indeed, Pombo et al. demonstrated the induction of blood-stage immunity from low-grade blood-stage infection through the vaccination of human subjects with three administrations of ~30 P. falciparum-infected red blood cells followed by drug treatment 8-14 days later (185). 59
81 This vaccination strategy resulted in complete protection in 3 of the 4 human subjects when challenged with ~30 P. falciparum-infected red blood cells. The delayed treatment, compared to immediate treatment in human studies of CPS (84, 85), could allow sufficient replication and host exposure to blood-stage specific antigens during subpatent parasitemia to induce blood-stage immunity. However, the authors could not link protection to measureable IgG responses against blood-stage antigens. Rather, they correlated protection with induction of a Th1 response (185). Conversely, when the human subjects receiving CPS vaccination were challenged with ~2,000 viable P. falciparum-infected erythrocytes I.V., the subjects were not protected and did not differ from naïve controls in the pre-patent period as detected by thick blood smear or PCR (84). From these results, the authors concluded that CPS in human subjects did not elicit functional blood-stage immunity. However, it is important to note the human model precludes the ability to determine if vaccine-induced blood-stage immunity can control parasitemia, because these patients must be treated when blood-stage parasites are first detected to prevent symptoms of malaria. Indeed, I found that the pre-patent period did not dramatically differ in mice containing antibodies reactive to blood-stage antigens compared to naïve mice. Nevertheless, blood-stage specific antibodies ultimately contribute to demonstrable control and earlier clearance of parasitemia in a stringent rodent model of malaria (18). Thus, it is possible the restricted analysis of antibodies produced against only one liver-stage (CSP) and two blood-stage antigens (AMA-1 and GLURP)(84), combined with the inability to directly test whether blood-stage immunity 60
82 can control parasitemia, may explain the lack of support for blood-stage immunity in humans receiving CPS. Interestingly, another human CPS vaccination study found slightly conflicting results in regards to induction of blood-stage-specific antibodies. Using a similar CPS vaccination strategy as before (84), but using a dose escalation setup instead of a standard 15 mosquito bites, human volunteers were subjected to 5, 10, or 15 bites by P. falciparum-infected mosquitoes under CQ chemoprophalysis on three occasions (121). Vaccinated subjects generated memory B cell and antibody responses against preerythrocytic (i.e. sporozoite and/or liver-stage antigens) and cross-stage antigens (liverand blood-stage specific) such as MSP-1, AMA-1, and GLURP. Although these humoral responses did not correlate with sterile protection in humans, these responses did correlate with the magnitude of the immunizing parasite dose (121). Therefore, CPS vaccination of human subjects does generate antibody responses against antigens expressed during blood-stage infection but it remains difficult to determine the contribution of these antibody responses to protection following challenge. Based on ethical constraints in human studies, rodent models are essential to understand the immunological mechanism underlying immunity elicited by wholesporozoite and blood-stage parasite vaccinations. Indeed, the scalability of production, potential for drug resistance, and requirement of multiple immunizations complicates the application of whole-parasite vaccination approaches in the field, whereas a subunit vaccination approach could circumvent all these issues. The rodent models of RAS and CPS vaccination have predicted a strategy that provides protection in humans (69, 71, 61
83 72). Therefore, continued studies in rodent models, and the analysis of the targeted antigens responsible for cellular and antibody-dependent immunity from sporozoite and blood-stage challenges, may reveal potent components for eliciting protection which can be translated for use in a human subunit vaccine. In conclusion, our data provide insight into the cellular and immunological basis for enhanced protective immunity following vaccination with virulent sporozoites under CQ chemoprophylaxis. Our data reveal CPS-induced immunity is not solely mediated by memory CD8 T cells, but is also mediated by T-cell independent blood-stage immune responses induced following exposure of the host to low-grade blood-stage parasitemia. In sum, these data further strengthen the rationale for mechanistic studies focused on identifying the cellular and humoral antigenic targets of whole parasite vaccine-induced, protective anti-plasmodial immunity. 62
84 Figure 3. CPS induces larger effector and memory CD8 T cell responses than RAS vaccination in C57Bl/6 mice C57Bl/6 mice were immunized I.V. with 10 4 Py either as RAS or CPS with 10 days CQ chemoprophylaxis. (A), Representative CD8 T cell responses in blood at the indicated days post vaccination first gated on CD8 + T cells. Numbers indicate the frequency of CD8 T cells exhibiting an antigen-experienced (CD8α lo, CD11a hi ) phenotype. (B), Cumulative and kinetic analysis of CPS versus RAS vaccine-induced CD8 T cell responses. Data are Mean +/- S.D. of 20 mice per group and are representative of two independent experiments. Statistical analysis employed Student s T-test comparing CPS to RAS at the indicated days. ***, p < ; *, p <
85 64
86 Figure 4. CPS induces larger effector and memory CD4 T cell responses than RAS vaccination in C57Bl/6 mice C57Bl/6 mice were immunized I.V. with 10 4 Py either as RAS or CPS with 10 days CQ chemoprophylaxis. (A), Representative CD4 T cell responses in blood at the indicated days post vaccination. Plots shown in (A) are first gated on CD4 + cells. Numbers indicate the frequency of CD4 T cells exhibiting an antigen-experienced (CD49d +, CD11a + ) phenotype. (B), Cumulative and kinetic analysis of CPS versus RAS vaccineinduced CD4 T cell responses. Data are Mean +/- S.D. for 20 mice per group, and are representative of two independent experiments. Statistical analysis employed Student s T-test comparing CPS to RAS at the indicated days. ***, p < ; *, p <
87 66
88 Figure 5. Chloroquine administration does not alter the kinetics or magnitude of the CD8 T cell response in C57Bl/6 mice 7 days after administration of attenuated recombinant Listeria monocytogenes expressing OVA (A-D) C57Bl/6 mice were immunized I.V. with 10 4 Py RAS with or without daily injections of CQ chemoprophylaxis. (A), Representative splenic CD8 T cell responses exhibiting an antigen-experienced (CD8α lo, CD11a hi ) phenotype at day 6 post immunization. (B), Cumulative and kinetic analysis of RAS +/- CQ vaccine-induced CD8 T cell responses. Data are Mean +/- S.D. for 5 mice per group, and are representative of two independent experiments (N=10 total mice per group across two experiments). (C), Representative splenic CD4 T cell responses exhibiting an antigen-experienced (CD49d +, CD11a + ) phenotype. (B), Cumulative and kinetic analysis of RAS +/- CQ vaccineinduced CD4 T cell responses. Data are Mean +/- S.D. for 5 mice per group, and are representative of two independent experiments (N=10 total mice per group across two experiments. (E), Cumulative data of splenic OVA-specific CD8 T cell responses of C57Bl/6 mice 7 days after administration of attenuated recombinant Listeria monocytogenes expressing OVA +/- CQ. Data are Mean +/- S.D. for five mice per group for one experiment. n.s. indicates not significant as analyzed by Student s T-test. 67
89 68
90 Figure 6. I.V. administration of CPS or RAS induces larger effector and memory CD8 T cell response than S.C. administration in C57Bl/6 mice C57Bl/6 mice were I.V. or S.C. immunized with 10 4 Py either as RAS or CPS with 10 days CQ chemoprophylaxis. (A and C), Representative CD8 T cell responses in blood at the indicated days post CPS (A) or RAS (C) vaccination. Numbers indicate the frequency of PBL CD8 T cells exhibiting an antigenexperienced (CD8α lo, CD11a hi ) phenotype. (B and D), Cumulative and kinetic analysis of I.V. versus S.C. CPS vaccination (B) or RAS vaccination (D). Data are Mean +/- S.E.M. for 20 mice per group from two combined independent experiments. Statistical analysis employed Student s T-test. ***, p < 0.001; **, p < n.s. indicates not significant. 69
91 70
92 Figure 7. I.V. administration of CPS or RAS induces larger effector and memory CD4 T cell responses than S.C. administration in C57Bl/6 mice C57Bl/6 mice were I.V. or S.C. immunized with 10 4 Py either as RAS or CPS with 10 days CQ chemoprophylaxis. (A and C), Plots are first gated on CD4 + cells. Representative CD4 T cell responses in blood at the indicated days post CPS (A) or RAS (C) vaccination. Numbers indicate the frequency of CD4 T cells exhibiting an antigen-experienced (CD49d +, CD11a + ) phenotype. (B and D), Cumulative and kinetic analysis of I.V. versus S.C. CPS vaccination (B) or RAS vaccination (D). Data are Mean +/- S.E.M. for 20 mice per group from two combined independent experiments. Statistical analysis employed Student s T-test. ***, p <0.001; **, p < n.s. indicates not significant. 71
93 72
94 Table 1. Single-dose chemoprophylaxis and sporozoite (CPS) vaccination provides sterilizing protection from P. yoelii sporozoite challenge in C57Bl/6 mice Attenuated Sporozoite Vaccination a Day challenged post-vaccination % Sterilizing protection b Naïve + CQ c N/A 0% (0/20) 10,000 I.V. RAS >60 days 0% (0/20) 10,000 I.V. CPS >60 days 72% (13/18) 10,000 S.C. RAS >60 days 0% (0/20) 10,000 S.C. CPS >60 days 47% (9/19) a Age-matched C57Bl/6 female mice were vaccinated I.V. or S.C. with 10 4 P. yoelii RAS or with virulent P. yoelii sporozoites plus once daily I.P. injection of CQ for 10 days, (ITV). b Sterilizing protection is defined as complete absence of detectable blood-stage parasitemia when assessed on days 3, 5, 7, 9, and 10 post-challenge with 10 3 P. yoelii sporozoites. c Naive C57Bl/6 female mice were administered CQ similarly as CPS group. 73
95 Figure 8. CPS vaccination induces larger magnitude CD8 T cell responses targeting a broader spectrum of liver-stage antigens than RAS vaccination (A), Schematic of experimental design. Naïve Thy1.2 + BALB/c recipient mice were I.V. administered 10 3 Thy1.1/1.2 + CS 280 TCR Tg cells from naïve Thy1.1/1.2 + CS 280 TCR transgenic BALB/c mice. Recipient mice were vaccinated one day later with CPS or RAS vaccination. (B), Frequency of splenic CD8 T cells exhibiting an antigen-experienced (CD8α lo, CD11a hi ) six days following vaccination first gated on CD8 + T cells. (C), Enumeration of Thy1.1+ CS 280 TCR transgenic cells in recipient mice. (D), Total number of splenic activated CD8 T cell response was measured six days after vaccination. Data are Mean +/- S.D. for 5 mice per group, and are representative of two independent experiments (N=10 total mice per each group across two experiments). Statistical analysis employed Student s T-test. **, p < n.s. indicates not significant. 74
96 75
97 Figure 9. CPS-induced CD8 T cells are not required for protection from homologous sporozoite challenge (A) CD8 T cell depletion in blood, spleen and liver of CPS mice (day 60 post immunization) given control Ig or anti-cd8 antibody (50 ug, day 57 and 59 post CPS). Statistical analysis employed Student s T-test. N=5 mice per group. Experiment performed once. ***, p < 0.001; **, p < (B), Percent of CPS (day 60) or naïve C57Bl/6 mice exhibiting protection (black shading), resistance (grey shading, defined in Materials and Methods), or no protection (open) to sporozoite challenge with or without depletion of CD8 T cells. Data are pooled from four independent experiments. N indicates total number of mice represented in indicated group of graph. P = for protection between rat IgG and anti-cd8 groups as assessed by Chi-square analysis. (C), Liver parasite burden 44 hours after 40,000 Py sporozoite challenge in CPS (day >50) or naïve mice depleted of CD8 T cells or treated with control Ig. Pyspecific 18s ribosomal RNA transcripts were quantified via qrt-pcr. Data are normalized to GAPDH transcripts and infected naive control Ct values (set at 100%) +/- S.E.M. Data are pooled from two independent experiments. N indicates total number of mice represented in indicated group of graph. Statistical analysis employed One-way ANOVA comparison to Naïve group. n.s. indicates not significant compared to Naïve control. 76
98 77
99 Figure 10. Parasites persist in the spleen and blood during CQ treatment and control of persistent parasites is dependent on CD4 T cells (A), Py-specific ribosomal RNA copy number in the spleen of I.V Py sporozoite-infected control (day 9) or CPS (day 9) C57Bl/6 mice. Dots represent individual mice and are pooled from two independent experiments. (B), CD4 T cell depletion in blood of CPS vaccination mice. Arrows indicate days 200µg of CD4 T cell depleting antibody was administered. Each line indicates measured %CD4 + T cells of Thy1.2 + cells in PBL from individual mice, N=5 total mice per group. (C), Parasitemia in CD4 T cell depleted or non depleted CPS mice from (B) after cessation of CQ. (D), Higher resolution of parasitemia frequency and kinetics in nondepleted group shown in (C). Error bars are Mean +/- S.D. of five mice from one experiment. (E), Frequency and kinetics of parasitemia of individual mice from (D); N=5. 78
100 79
101 Figure 11. CPS vaccination with 25 days of CQ cover abrogates breakthrough parasitemia as well as protection (A), Percentage of CPS vaccinated C57Bl/6 mice with breakthrough parasitemia following 10, 15, 20, or 25 days of CQ cover. Data are Mean +/- S.D. from three independent experiments with 9 total mice per group. One-way ANOVA analysis with multiple comparsions indicates p < 0.05 for every comparsion between groups. (B), Percent of C57Bl/6 CPS mice, given 10 days or 25 days of CQ chemoprophylaxis, or RAS vaccinated mice exhibiting protection (black shading), resistance (grey shading), or no protection (no shading) after I.V. challenge with 10 3 virulent Py sporozoites >60 days after vaccination. Data are pooled from four independent experiments. N indicates total number of mice in represented in indicated group. p < by Chi- Square analysis comparing CPS-10d CQ to CPS-25d CQ for protection. 80
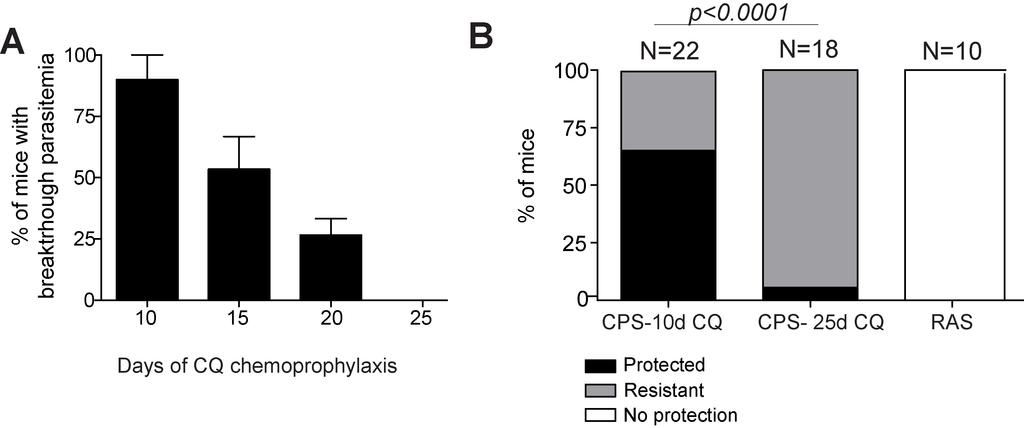
102 81
103 Figure 12. CPS with 10 days of CQ cover correlates with higher antiparasite antibody responses (A), Optical density measurement of total serum IgG recognizing parasitized red blood cell lysates from naïve or CPS C57Bl/6 mice (CPS-10d CQ or CPS- 25d CQ). The CPS-25d group did not exhibit patent parasitemia after CQ cessation. Results are representative of two independent experiments. Error bars indicate Mean +/- S.E.M., N=10 mice total per group. Statistical analysis employed Student s T-test. **, p < 0.01; *, p < (B), similar to (A) but measuring total IgG against MSP1 recombinant protein. Error bars indicate Mean +/- S.D. No significant differences between 10d and 25d groups, N=5 total mice per group. Experiment performed once. (C), Survival curve following 10 3 sporozoite challenge of previously CPS vaccinated 10 day C57Bl/6 wild-type mice compared to CPS µs-aicda -/ day CQ mice. p = based on Log-rank Mantel Cox test. 82
104 83
105 Figure 13. T cells are not required for protection from blood-stage parasite challenge in single administration of CPS mice Percentages of CPS or naïve C57Bl/6 mice exhibiting protection (black shading), resistance (gray shading), or no protection (no shading) with or without depletion of CD8 and CD4 T cells prior to I.V. challenge with 10 6 blood-stage Py parasites. N indicates total number of mice represented in indicated group. Data are pooled from three independent experiments. Statistical analysis employed Chi-Square analysis of CPS Rat IgG to CPS anti- CD4/CD8, p =
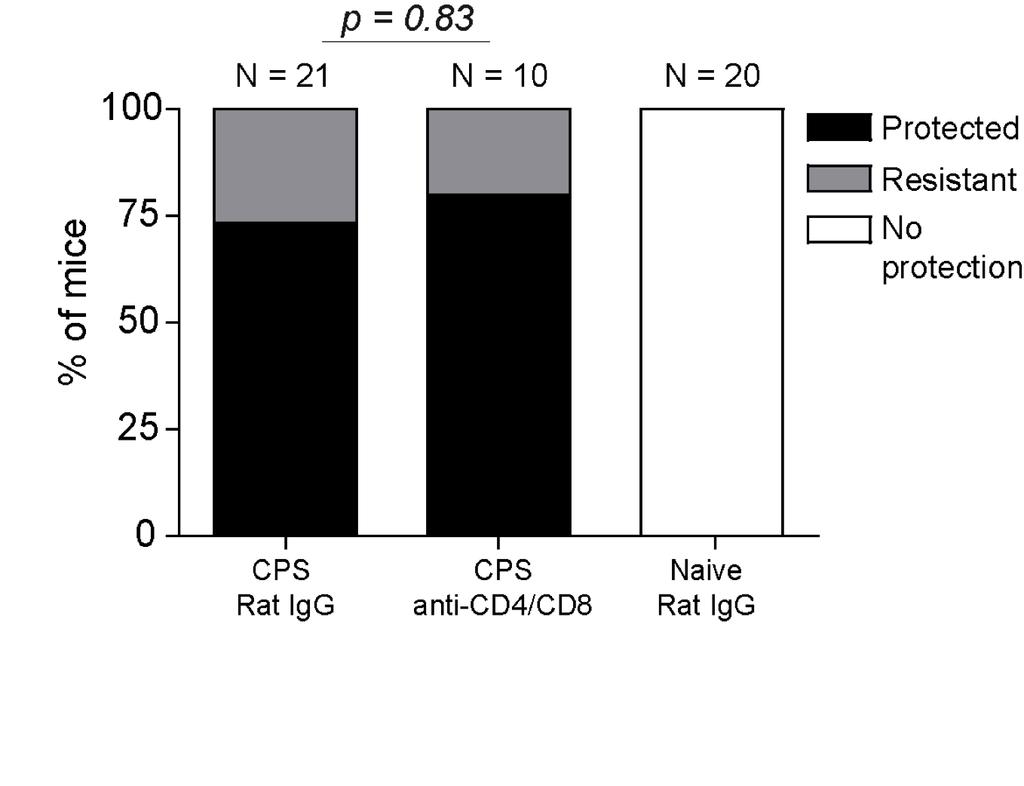
106 85
107 CHAPTER III: DISCRIMINATING PROTECTIVE FROM NON-PROTECTIVE PLASMODIUM-SPECIFIC CD8 T CELL RESPONSES Abstract Despite decades of research, malaria remains a global health crisis. Current subunit vaccine approaches do not provide efficient long-term, sterilizing immunity against Plasmodium infections in humans. Conversely, whole parasite vaccinations with their larger array of target antigens have conferred long lasting sterilizing protection to humans. Similar studies in rodent models of malaria reveal that CD8 T cells play a critical role in liver-stage immunity after whole parasite vaccination. However, it is unknown whether all CD8 T cell specificities elicited by whole parasite vaccination contribute to protection, an issue of great relevance for enhanced subunit vaccination. Here we show that robust CD8 T cell responses of similar phenotype are mounted following prime-boost immunization against Plasmodium berghei GAP , S , TRAP or CSP protein-derived epitopes in mice, but only CSP and TRAP specific CD8 T cells provide sterilizing immunity and reduce liver parasite burden following sporozoite challenge. Further, CD8 T cells specific to sporozoite surface-expressed CSP and TRAP proteins, but not the intracellular GAP50 and S20 proteins, efficiently recognize sporozoite-infected hepatocytes in vitro. These results suggest that 1) protection-relevant antigenic targets, regardless of their immunogenic potential, must be efficiently presented by infected hepatocytes for CD8 T cells to 86
108 eliminate liver-stage Plasmodium infection and 2) proteins expressed on the surface of sporozoites may be better target antigens for protective CD8 T cells. Introduction Malarial disease leads to approximately 400,000 deaths every year despite the widespread use of interventions such as bed nets and insecticides, and continues to exert significant health and socioeconomic impact on a third of the world s population. Recently, the first malaria vaccine, Mosquirix (RTS,S), was licensed for use by the European Union (122). Mosquirix is a recombinant protein-based subunit vaccine, which induces humoral and CD4 T cell responses against the circumsporozoite protein (CSP) of P. falciparum (92). Unfortunately, this subunit vaccine does not reach ideal rates of efficacy and protection wanes over time (92, 192). Conversely, administration of whole radiation-attenuated sporozoites (RAS) can lead to complete, sterilizing immunity in humans and rodents (69, 71, 72, 99). Mechanistic studies in rodents revealed that RASinduced protection is dependent upon CD8 T cells, likely against a spectrum of Plasmodium antigens (94, 95, 98, 99). While effective, RAS vaccination has some complications in safety and application in the field due to the requirement of a large parasite dose, need for aseptic, laboratory-reared mosquitoes, and the lack of immunogenicity unless administered via mosquito bite or intravascular injection (78, 79). In parallel, efforts are underway to evaluate viral vectored subunit vaccines, expressing one of a few potential target antigens, that would ideally elicit CD8 T cell responses to liver-stage antigens (131, 193). However, controlled human challenge trials have not 87
109 revealed robust sterilizing immunity after viral vectored subunit immunizations (91, 140, 194). One possible path forward for subunit vaccines would be immunizations with a combination of target antigens identified from RAS immunized hosts and there are ongoing efforts in such malaria antigen-discovery. However, it is unknown which antigens would serve as the best targets for protective CD8 T cells. In this regard, RAS vaccination of humans and rodents can serve as a platform for new CD8 T cell antigen discovery for inclusion in subunit vaccines. However, because RAS vaccination induces CD8 T cell responses against a potentially large spectrum of parasite antigens, it remains unclear whether all of the RAS-induced antigen-specific CD8 T cells contribute to protective immunity, or, if only a subset of parasite antigens recognized by the RAS-induced CD8 T cell response are targets of protective immunity. Resolving this question is important in order to design subunit vaccines composed only of antigens targeted by CD8 T cells capable of providing protection. Plasmodium sporozoites delivered via mosquito bite or intradermal injections prime CD8 T cell responses against a broad spectrum of antigens, largely within the skin draining lymph nodes via cross-presentation mediated by dendritic cells ( ). Plasmodium-specific memory CD8 T cells target parasite-infected hepatocytes to provide liver-stage immunity (199, 200). A recent study suggests that parasite antigen needs to be accessible to the endogenous pathway for MHC class I presentation by infected hepatocytes to allow recognition by protective CD8 T cells (197). Consistent with this notion, access of antigen to host cell endogenous MHC class I presentation machinery in the cytosol is a critical factor affecting the efficiency by which CD8 T cells can 88
110 recognized infected cells (201). Thus, some parasite antigens may be more easily accessible to the host cell cytosol for presentation compared to others. Recently, the importance of antigen secretion for CD8 T cell-mediated protection in Plasmodium was suggested from studies of mice containing large numbers of OT-I T cell receptor (TCR) transgenic (Tg) cells, specific for an epitope from ovalbumin (OVA) that were immunized with Pb RAS-expressing secreted or non-secreted OVA. Despite similar OT- I responses in each group, homologous challenge resulted in better control of parasites expressing secreted compared to non-secreted OVA (202). However, these studies relied on a model antigen in mice containing supraphysiologic numbers of TCR Tg cells and did not address whether endogenous liver-stage antigens similarly engender protection by CD8 T cells. Further, the individual contribution to protection mediated by CD8 T cells targeting the secreted antigen remains unclear as these mice contained additional CD8 T cells targeting the entire spectrum of Plasmodium antigens due to homologous parasite immunization and challenge. Thus, it remains to be determined whether compartmentalization of antigens within the complex Plasmodium parasite alters the efficiency of direct MHC class I antigen presentation, and therefore the protective capacity of the CD8 T cell response. The Plasmodium genome contains approximately 5,000 open reading frames (142, 143), complicating the systematic identification of potential antigenic targets of protective CD8 T cells. Recently, three new CD8 T cell epitopes from Plasmodium berghei ANKA (Pb) have been described (136, 157). These three new epitopes are derived from a well-described sporozoite surface protein (TRAP)(158), a well-described 89
111 intracellular protein (GAP50)(159), and a protein of unknown function (S20) lacking a signal peptide sequence, thus suggesting an intracellular localization (160). Initial characterization of the protective capacity of TRAP and S specific CD8 T cells against liver-stage infection relied on a model of RAS immunization and elimination of specific CD8 T cell responses (tolerization) by high dose peptide-epitope injection (136). These studies provided some evidence that eliminating the TRAP , but not S specific CD8 T cells compromised control of liver parasite burden in difficultto-protect C57Bl/6 mice (98, 203). However, this approach could miss contributions by S specific CD8 T cells, due to the presence of an excess of other protective CD8 T cells. Additionally, this study evaluated TRAP as the sole protective antigen after viral-vectored immunization. Here, TRAP-immunization resulted in reduced liverparasite burden after challenge, but no detectable sterilizing immunity. Thus, the potential of TRAP or S20 to serve as antigens for sterilizing immunity remains to be determined. To date, no studies have evaluated CD8 T cells specific for GAP50 in liver-stage immunity. To evaluate candidate antigens for inclusion in subunit vaccines, the protective capacity of the CD8 T cell response needs to be systematically evaluated in isolation of other anti-plasmodium responses, and by sufficient magnitude CD8 T cell responses for sterilizing immunity, if the antigen is permissive (134). Using these three new epitopes, in addition to the well-described circumsporozoite (CSP) epitope (97), derived from the abundant surface protein CSP (142, 204), I examined how the specificity of an anti-plasmodium CD8 T cell response relates to their capacity to provide sterilizing liver-stage immunity. 90
112 Material and Methods Mouse strains Female 6-8 week old BALB/cJ and CB6.F1 (C57BL/6 x BALB/c F1) mice were purchased from the National Cancer Institute/Charles River (Frederick, MD). BALB.b mice were purchased from Jackson Laboratory (Frederick, MD) and bred for use in experiments. BALB.b mice were bred to BALB/cJ mice to generate BALB.b x BALB/cJ F1 (H-2 bxd ) mice. Females 6-12 week old were used in experiments. All mice were housed at the University of Iowa animal care unit at the appropriate biosafety level. The University of Iowa Institutional Animal Care and Use Committee approved animal experiments. Generation of recombinant Listeria expressing Plasmodium CD8 T cell epitopes Attenuated (ΔactA/ΔinlB) recombinant Listeria monocytogenes (rlm) expressing P. berghei ANKA CD8 T cell epitopes were generated as described previously (205, 206). Briefly, I generated LM codon-optimized constructs containing amino acid coding sequences of the defined GAP , S , or TRAP epitopes, with a 3 flanking amino acids on each end to encourage natural processing. I used the following sequences: GAP50 (ESKSQLLNAKYLKQF); S20 (KDNVYSFLYLFGGD); TRAP (NLTSALLNVDNLIQK). rlm expressing CSP (NDDSYIPSAEKILEF) has been described (134). Residues in bold represent the minimum epitope. 91
113 DC-LM vaccination to generate antigen-specific CD8 T cells Groups of mice were individually primed with I.V. administration of 5 x 10 5 Flt3L-induced, mature dendritic cells (DC) coated with CSP , GAP , S , or TRAP peptides as previously described (134). Seven days later mice were boosted by I.V. infection with 1-2 x 10 7 CFUs of rlm expressing the same epitope. rlm strains were grown and prepared for injection as previously described (134, 173). Plasmodium parasites and sterilizing protection challenges P. berghei ANKA clone 234 (Pb) sporozoites were isolated from the salivary glands of infected A. stephensi mosquitoes obtained from an in-house insectary as previously described (134). P. berghei ANKA-GFP expressing (Pb-GFP) sporozoites were isolated from the salivary glands of infected A. stephensi mosquitoes purchased from the insectary of New York University (NY, NY). For some experiments, sporozoites were attenuated by radiation (radiation-attenuated sporozoites, RAS) with 200 Gy by cesium irradiation. In-house insectary mosquitoes were used for RAS and sterilizing immunity experiments. GFP-expressing parasites were used for primary hepatocyte culture experiments and liver parasite burden experiments. Naïve and DC-LM immunized mice (>60 days post LM-boost) were challenged I.V. with 10 3 Pb sporozoites. Sterilizing protection from sporozoite challenge is defined as the absence of detectable parasites by Giemsa-stained blood smear on days 4-10 after challenge. At least 20 fields containing at least 100 red blood cells (RBCs) per field were examined at each time point to assess parasitemia. 92
114 Quantification and analysis of Plasmodium-specific CD8 T cell populations DC-LM vaccination induced CD8 T cell populations were identified by staining peripheral blood or single-cell suspension of splenocytes or liver-derived lymphocytes with anti-cd8α FITC (clone 5H10-1) and one of the following respective MHC class I tetramers: K d -CS , D b -GAP , K b -S , D b -TRAP prepared in house using published protocols (207, 208). Further phenotypic analysis was performed by staining with the following antibodies: CD62L (clone MEL-14), KLRG1 (clone 2F1/KLRG1), CD127 (clone A7R34), CD122 (clone 5F4), and CD25 (clone PC61.5) in appropriate combinations of fluorochromes. Alternatively, Plasmodium-specific CD8 T cell populations were identified by intracellular cytokine staining (ICS) with anti-cd8α and anti-ifn-γ (clone XMG1.2, ebioscience, San Diego, CA), TNFα (clone MP6-XT22), and IL-2 (clone JES6-5H4, ebioscience) following cognate peptide stimulation ex vivo in the presence of Brefeldin A as previously described (203). All antibodies were purchased from Biolegend (San Diego, CA), unless otherwise specified. Cells were analyzed using a BD LSR Fortessa and data were analyzed using FLOWJO Software (Tree Star, Inc, Ashland, OR). In vivo cytolytic assay Splenocytes from naïve H-2 bxd donors were left uncoated or coated with 1 mm CSP , GAP , S , or TRAP peptide for 1 hour at 37 C then washed with PBS. Peptide-pulsed cells were labeled with 2.5 mm CellTrace Violet (CTV, Life Technologies, Carlsbad, CA) and uncoated cells were labeled with 0.25 mm CTV. A 93
115 mixture of labeled splenocytes (2 x 10 6 CTV hi and 2 x 10 6 CTV lo ) were injected I.V. into naïve or DC-LM vaccinated mice. Killing of peptide-pulsed CTV hi -labeled cells was detected in the spleen 4 hours after donor cell injection by flow cytometric analysis. Percentage killing was calculated as: 100-(100*[(% CTV hi / % CTV lo )/(% CTV hi in naive mice/% CTV lo in naive mice)]). Functional avidity analysis Splenocytes from DC-LM vaccinated BALB.b x BALB/c F1 mice >60 post LM boost were incubated for 5 hours with the indicated concentration of stimulating peptide in the presence of Brefeldin A. Following stimulation, cells were surface stained for anti- CD8α and anti-ifnγ as described above. To calculate the percentage of the maximum response, each sample was normalized to the response generated by stimulation with 500nM of peptide (the maximum response). Liver parasite burden Mice were challenged 14 days after LM boost with 5 x10 5 Pb sporozoites I.V. Liver RNA was extracted at 42 hours post challenge using TRIzol (Invitrogen, Carlsbad, CA) followed by DNase digestion/clean-up with RNasy Kit (Qiagen, Hilden, Germany). One-step quantitative real-time reverse transcription PCR analysis for P. berghei 18S rrna was performed on 2µg of liver RNA using Fast Virus One-step qpcr (Applied Biosystems, Foster City, CA). P. berghei 18S rrna was detected using 5-94
116 CGCAAGCGAGAAAGTTAAAAGAA-3 (forward primer), 5 - GAGTCAAATTAAGCCGCAAGCT -3 (reverse primer), and 5 - FAM- TGACGGAAGGGCACCACCAGG-TAMRA-3 (probe), which generates a 71-bp fragment (203). Data were normalized for input RNA with GAPDH control (Rodent GAPDH Control Reagents, Life Technologies). Data plotted as +/- S.E.M. of Log10 of parasite-specific 18s rrna transcripts based on 18s rrna standards run in parallel. Primary hepatocyte culture Primary hepatocytes were isolated from naïve H-2 bxd mice as described (209). Mice were anesthetized with ketamine, and using aseptic technique, angiocatherization into the portal vein was performed. Steady-state perfusion of liver was performed first with 1x PBS (5mL/min for 5 minutes), then Liver Perfusion Medium (5mL/min for 3 minutes, Gibco, ThermoFisher Scientific, Carlsbad, CA), and finally Liver Digest Medium (3mL/minute for 5-8 minutes, Gibco). Digested liver was excised and a single cell suspension made, suspended in wash solution composed of DMEM (Gibco), 10% FBS, and 1x antibiotic/mycotic solution (containing penicillin, streptomycin and amphotericin B, Sigma-Aldrich, St. Louis, MO). Non-hepatocytes and debris were then removed with a 50% Percoll (GE Healthcare, Little Chalfont, United Kingdom) gradient. Remaining cells were counted and resuspended in DMEM with 10% FBS and 1x antibiotic/mycotic. 2 x 10 4 cells were plated on collagen-coated 96-well plates in 100 µl volume and incubated overnight in 37 C, 5% CO 2 incubator. Primary hepatocyte cultures were infected with 2 x 10 4 PbANKA-GFP or CTV-labeled sporozoites per 96 95
117 well (MOI ~1), and the plate was centrifuged for 10 minutes at 2400 RPM without braking. Cultures were furthered incubated for 20 or 40 hours at 37 C, 5% CO 2 to allow infection and liver-stage parasite development. Infection was confirmed by analyzing GFP + or CTV + hepatocytes by inverted fluorescent microscopy (when cultures infected with GFP+ sporozoites) or flow cytometry (when cultures were infected with 5mM CTVlabeled sporozoites). Wells were washed twice with 1x PBS and then exposed to enriched splenic-derived CD8 T cells from DC-LM vaccinated BALB.b x BALB/cJ F1 mice in DMEM supplemented with 10% FBS and 1x antibiotic/mycotic medium. CD8 T cell enrichment from DC-LM vaccinated mouse spleens was performed using EasySep CD8 T cell isolation kit (StemCell Technologies, Vancouver, Canada). 8 x 10 4 antigen-specific CD8 T cells with 1x Brefeldin A (Biolegend) were added to 96-wells based on calculations of 90% CD8 T cell enrichment, and pre-harvest peripheral blood CD8 + tetramer + staining of donor cell population. CD8 T cells were also added to non-infected hepatocyte culture wells with or without 500nM exogenous peptide to serve as positive and negative controls, respectively. CD8 T cells added onto hepatocyte cultures were incubated for an additional 6 hours at 37 C, 5% CO 2. IFNγ + CD8 T cells were detected following intracellular cytokine staining and flow cytometry to determine the proportion of IFNγ-producers in well containing sporozoite-infected hepatocytes (for 20 or 40 hours) relative to wells stimulated with exogenous peptide. 96
118 Statistical analysis Data were analyzed using GraphPad Prism6 software (GraphPad Software, La Jolla, CA) to determine statistical significance as indicated in figure legends. Results Radiation-attenuated sporozoite (RAS) vaccination induces CD8 T cells against identified and unidentified antigens RAS vaccination provides sterilizing immunity, defined as the absence of detectable blood stage parasites in humans and rodents challenged with viable sporozoites by mosquito bite or needle stick (69, 71, 72, 78). Studies using mouse models have shown that RAS-induced protection is largely mediated by Plasmodiumspecific CD8 T cell responses (94, 95, 98, 99). Recently, three novel Pb epitopes have been described through studies utilizing C57Bl/6 (H-2 b ) mice (136, 157). To verify the authenticity of these new responses and to characterize what portion of the RAS-induced CD8 T cell response is comprised of CD8 T cells specific for these three novel epitopes relative to the well-characterized CSP epitope (97) (Table II), BALB.b x BALB/c F1 mice were I.V. administered 10 4 Pb-RAS and the total antigen-specific CD8 T cell response, as well as individual antigen-specific CD8 T cell responses were measured in the spleen 6 days later, at the peak of the RAS-induced CD8 T cell response for this immunizing dose (99). BALB.b x BALB/c F1 mice were utilized to allow the tracking of all four responses (CSP , GAP , S , and TRAP ) within one mouse strain due to the varying MHC Class I restriction of the different peptides (Table II). The 97
119 total activated CD8 T cell response was measured using a surrogate activation marker approach where responding Plasmodium-specific CD8 T cells up-regulate CD11a and down-regulate CD8α (171) following RAS vaccination (81, 99, 210, 211). Frequencies of CD8α lo CD11a hi CD8 T cells were measured from the spleen prior to and at 6 days post RAS vaccination, revealing that ~10-15% of CD8 T cells were activated by vaccination (Figure 14A). Within the total activated CD8 T cell response at day 6 following RAS vaccination, CSP , GAP , S , and TRAP specific CD8 T cells were readily and reproducibly detectable after peptide-stimulated detection of IFNγ (Figure 14B), whereas specific responses were not detected above background from the CD8α hi CD11a int naïve populations (Figure 14C). These four CD8 T cell responses individually averaged from ~2-6% of the RAS-induced CD8 T cell response and collectively accounted for approximately 15% of the total RAS-induced CD8 T cell response (Figure 14D). These results verify the authenticity of these novel responses, and indicate that the epitopes recognized by the majority of Pb-specific CD8 T cell response remain to be identified. In sum, these data demonstrate that the total CD8 T cell response following RAS vaccination in BALB/c x BALB.b F1 H-2 bxd mice is composed of multiple identified and as yet unidentified antigen-specific CD8 T cell responses. 98
120 DC-LM vaccination induces large, long-lasting Plasmodium-specific CD8 T cell responses displaying different capacity for sterilizing protection Vaccination-induced sterile immunity is defined by prevention of detectable blood-stage infection after sporozoite challenge and thus subsequent malarial disease. This protection can be achieved in Pb infection models by inducing a memory CD8 T cell response against the CSP epitope (97, 134, 135). However, the numerical requirements for protection by this single-specificity memory CD8 T cell response were substantial, requiring >8% of the circulating CD8 T cell compartment for sterilizing immunity, even in relatively resistant BALB/c mice (99). The superior protection afforded by RAS vaccination compared to current RTS,S or viral vectored subunit vaccination approaches might be due to an increase in the number of targeted antigens in the RAS-induced CD8 T cell repertoire (99, 149, 150). However, despite this almost certain broadening of the anti-parasite CD8 T cell repertoire, RAS vaccination does not meaningfully reduce the numerical requirement for CD8 T cell-mediated protection in rodent models (99). These results suggest that not all Plasmodium-specific CD8 T cell responses contribute to protective immunity. To test the protective capacity of memory CD8 T cells individually targeting the three distinct novel antigens, I generated large magnitude, single-specificity anti-plasmodium CD8 T cell responses in C57Bl/6 mice using an accelerated prime-boost vaccination strategy (173). Single-specificity CD8 T cell responses were primed in naïve C57Bl/6 mice by immunization with mature dendritic cells (DC) coated with one of three of the novel peptide epitopes (Table II) and then boosted by administration of an attenuated recombinant Listeria monocytogenes 99
121 (LM) expressing the same peptide (DC-LM vaccination). DC-LM vaccination induced single-specificity anti-plasmodium memory (>60 days post LM boost) CD8 T cell responses that constituted ~4-18% of the total CD8 T cell compartment in the blood, depending on the epitope specificity, with the GAP specific CD8 T cells exhibiting the largest response, and TRAP specific CD8 T cells exhibiting a significantly (p < 0.05) smaller response than GAP and S specific CD8 T cells (Figure 15A,B). Infected mosquitoes deliver ~ sporozoites to the host during a blood meal (21). To compare the capacity of DC-LM generated Plasmodium-specific CD8 T cells to provide sterile protection following sporozoite infection, vaccinated and control C57Bl/6 mice were challenged with a near physiological challenge dose (10 3 virulent Pb sporozoites administered I.V.) that reproducibly infects all naïve control mice (99, 134). Infected mice were monitored for survival, as Pb ANKA injection of C57Bl/6 mice leads to experimental cerebral malaria (ECM) in the majority of mice in 6-14 days after infection (212, 213), or mice eventually succumb to hyperparasitemia 2-3 weeks later (214). Challenged unvaccinated mice succumbed to ECM within 8 days of challenge (Figure 15C). All DC-LM vaccinated C57Bl/6 mice also succumbed to ECM, but with delayed kinetics relative to the unvaccinated control. DC-LM vaccinated S20 mice all succumbed to ECM by day 9 post challenge (p < 0.01 relative to unvaccinated) whereas DC-LM vaccinated GAP50 and TRAP mice completely succumbed by day 10 (p < 0.01 and p < relative to unvaccinated, respectively). The extended survival of DC-LM TRAP vaccinated mice relative to unvaccinated controls extend recent studies showing 100
122 the ability of TRAP specific CD8 T cells to reduce liver parasite burden (but not provide sterilizing immunity) in susceptible C57Bl/6 mice (136). These data indicate that despite large magnitude memory CD8 T cells responses specific to Plasmodium liverstage expressed antigen in C57Bl/6 mice (Figure 15A,B), not all liver-stage parasites were targeted for elimination to prevent blood-stage infection and development of ECM. RAS-vaccinated C57Bl/6 mice are more difficult to protect than BALB/c mice (98), indicating that mouse strains can have varying degrees of stringency to achieve sterilizing protection. Because I was unable to observe sterilizing protection nor survival in any DC-LM vaccinated C57Bl/6 mice, I tested whether DC-LM generated singlespecificity CD8 T cell responses could afford sterile protection in a less stringent BALB.b model, which are mice with BALB/c mouse background genes congenic for H- 2 b expression. To test the protective capacity of memory CD8 T cells individually targeting one of the three novel antigens, BALB.b mice were DC-LM GAP50, S20, or TRAP vaccinated. DC-LM vaccination induced single-specificity anti-plasmodium memory (>60 days post LM boost) CD8 T cell responses that constituted 20-55% of the total CD8 T cell compartment in the blood, depending on the epitope specificity, with the GAP specific CD8 T cell again exhibiting a significantly larger response than S or TRAP specific responses (p < 0.001, Figure 16A,B). To assess if DC-LM generated Plasmodium-specific CD8 T cells could lead to protection in BALB.b mice, DC-LM vaccinated BALB.b mice were challenged with 10 3 Pb sporozoites and assessed over 10 days by Giemsa-stained blood smear for evidence of blood-stage parasites. Sterilizing immunity was defined as the absence of detectable 101
123 parasitemia at all days of analysis. Despite having the largest magnitude anti-plasmodium CD8 T cell response relative to the other DC-LM vaccinated groups, DC-LM GAP50 vaccination lead to no sterilizing protection in any of the sporozoite challenge mice (0/16). In contrast, mice administered DC-LM TRAP vaccination, which exhibited the lowest magnitude CD8 T cell response, had significantly greater protection following challenge, with 82% (14/17) of the mice sterilely protected (p < relative to GAP50 or S20)(Figure 16B). Approximately 11% (2/17) of DC-LM S20 vaccinated BALB.b mice were sterilely protected from sporozoite challenge, but DC-LM S20 vaccination was not statistically different than DC-LM GAP50 vaccination (p = 0.49)(Figure 16B). Taken together, these data from a less stringent mouse model allowed for the determination of the potential for TRAP specific CD8 T cells to mediate a substantial degree of sterilizing protection following sporozoite challenge. Furthermore, these data indicate that anti-plasmodium CD8 T cell specificities are not equivalent at mediating protection following sporozoite challenge as lower frequencies of TRAP specific memory CD8 T cells could mediate sterilizing protection, whereas relatively larger magnitude GAP specific memory CD8 T cell responses did not provide sterilizing protection to any mice. From a less stringent mouse model (BALB.b), I determined that TRAP specific CD8 T cells could mediate sterilizing protection from sporozoite challenge, whereas GAP and S specific CD8 T cells could not. In order to determine a mechanistic basis for the differences in the protective capacity of these novel CD8 T cell specificities, I next tested if TRAP specific CD8 T cells could mediate 102
124 sterilizing protection in an H-2 bxd mouse strain, which would allow for comparisons of the novel CD8 T cell specifcities with the well-described, protective CSP H-2K d - restricted CD8 T cell specificity (Table II). CB6.F1 (C57Bl/6 x BALB/c F1) mice and BALB.b x BALB/c F1 mice (both H-2 bxd ) were DC-LM CSP, GAP50, S20, or TRAP vaccinated. DC-LM vaccination induced single-specificity anti-plasmodium memory (>60 days post LM boost) CD8 T cell responses that constituted 10-20% of the total CD8 T cell compartment in the blood of CB6.F1 mice (Figure 17A,B) or 10-40% in BALB.b x BALB/c F1 mice (Figure 18A,B). In both H-2 bxd mice strains, GAP and S specific CD8 T cell responses were larger in magnitude than CSP , and TRAPspecific CD8 T cell responses (Figures 17B and 18B) with the differences being most prominent in the BALB.b x BALB/c F1 mouse background (p < 0.01 for any comparison between CSP or TRAP relative to GAP50 or S20). As expected based on prior studies examining protection of DC-LM generated CSP specific CD8 T cells in relatively resistant BALB/c mice (134, 155), CSP specific CD8 T cell responses were capable of providing sterilizing protection following sporozoite challenge in a portion of the CB6.F1 and BALB.b x BALB/c F1 mice (14/31; 45%) (Table III). Similarly, TRAP specific CD8 T cells also provided sterilizing protection in some CB6.F1 and BALB.b x BALB/c F1 mice following sporozoite challenge (6/24; 25%). However, GAP and S specific CD8 T cells did not provide sterilizing protection to any vaccinated CB6.F1 or BALB.b x BALB/c F1 mice (Table III). This result occurred despite the GAP and S immunized mice having larger magnitude CD8 T cell responses compared to 103
125 CSP and TRAP specific CD8 T cell responses following DC-LM vaccination (Figures 17B and 18B). Taken together, these data demonstrate that DC-LM vaccination can generate large memory Plasmodium-specific CD8 T cell responses, which have different capacities to prevent blood-stage infection following sporozoite challenge based on their antigen-specificity in two different H2 bxd F1 mouse strains. DC-LM vaccination generated CD8 T cells with similar phenotype and functionality The quantities and qualitative characteristics of memory CD8 T cells can impact their capacity to provide immunity against specific pathogens (99, 134, 147, 215, 216). To determine if DC-LM generated CD8 T cell populations specific for the four defined epitopes displayed qualitative differences that could explain their differential ability to provide sterilizing immunity, I assessed CD8 T cell expression of select memory makers, cytokine production, and cytolytic capacity in BALB.b x BALB/c F1 mice. Memory CD8 T cells specific for CSP , GAP , S , and TRAP were identified by MHC class I tetramer analyses of cells from blood (Figure 19), spleen (Figure 20), and liver (Figure 21) expressed similar amounts of cell surface CD62L, KLRG1, and CD127 within each organ. Based on statistical analysis, some modest differences were observed between groups, but these differences did not reveal trends between protective CSP , and TRAP specific CD8 T cells, and non-protective GAP and S specific CD8 T cells. Thus, differences in memory CD8 T cell phenotypes may not account for epitope-specific differences in protection by these Plasmodium-specific CD8 T cells. 104
126 Cytokine production by memory CD8 T cells, specifically IFNγ, can contribute to sterilizing immunity against Plasmodium sporozoite infection (94, 98, 155). To assess cytokine production, splenic memory CD8 T cells were identified by IFNγ production after in vitro peptide stimulation, (Figure 22A) and polyfunctionality was assessed by detection of TNFα and IL-2 production by IFNγ + cells (Figure 22B). Again, although modest differences in polyfunctionality were observed between epitope-specific CD8 T cell populations, the majority of cells of each specificity produced both IFNγ and TNFα or IFNγ, TNFα, and IL-2 (Figure 22C). Thus, differences in cytokine production likely do not account for epitope-specific differences in protection by these Plasmodium epitope-specific CD8 T cells. Additionally, cytolysis has been shown to play a major role in memory CD8 T cell protection against Plasmodium sporozoite infection (155, 200, 217). To assess cytolysis, I performed epitope-specific in vivo killing assays in DC-LM vaccinated or naïve mice (Figure 23). DC-LM generated CD8 T cell populations were capable of rapid (4 hour) in vivo killing of specific peptide coated targets, although TRAP specific CD8 T cell mediated killing was decreased relative to the other specificities (Figure 23A, B). However, this decrease was likely due to the lower frequency of TRAP specific CD8 T cells in the analyzed mice (Figure 23C). In fact, in vivo killing by TRAP specific CD8 T cells was proportional to the frequency of circulating memory CD8 T cells (Pearson r = 0.982)(Figure 23D). Thus, differences in cytolytic capacity likely do not account for epitope-specific differences in protection by these Plasmodium-specific CD8 T cells. 105
127 Antigen sensitivity of DC-LM generated CSP , GAP , S , and TRAP specific CD8 T cells is similar Differences in the sensitivity of DC-LM generated CD8 T cells to respond to cognate antigen may impact the ability of a CD8 T cell specificity to detect and kill infected cells expressing cognate peptide expressed on surface MHC Class I molecules. To determine if differences in antigen sensitivity could account for the lack of protection of DC-LM vaccinated GAP50 and S20 mice following sporozoite challenge, I measured the ability of DC-LM generated CSP , GAP , S , and TRAP specific splenic memory CD8 T cells to produce IFNγ following ex vivo stimulation with titrating doses of their respective peptide (Figure 24A). I observed no significant difference between the CD8 T cell specificities in sensitivity to their respective antigen (Figure 24B). Therefore, differences in antigen sensitivity likely do not account for differences in protection by these Plasmodium-specific CD8 T cells. DC-LM generated GAP and S specific CD8 T cells do not reduce liver parasite burden Failure of epitope-specific CD8 T cells to provide sterilizing immunity does not rule out the potential for this population to provide partial control of liver-stage infection. To test if DC-LM generated CD8 T cells can reduce parasite burden in the liver after sporozoite challenge, I vaccinated BALB.b x BALB/c F1 mice and measured Pb-specific 106
128 18s rrna transcripts in the liver 42 hours after I.V. sporozoite challenge, a time point representing a late-liver stage infection just prior to release of parasites into the blood. In order to enhance the possibility of observing partial control of liver parasites by GAP and S specific CD8 T cells, DC-LM vaccinated mice were challenged with a high dose (4 x 10 4 ) of parasites 14 days post LM boost, an early memory time point with near maximal CD8 T cell numbers after boosting. Also, to control for possible alterations in the liver environment due to prior LM infection and subsequent high magnitude single-specificity CD8 T cell response, control mice were vaccinated similarly with DCs coated with an irrelevant epitope from ovalbumin (OVA , SIINFEKL), and boosted with rlm expressing the OVA epitope (173). Forty-two hours after sporozoite challenge, mice with CSP or TRAP specific CD8 T cells had significantly (p < ) reduced liver parasite burden (>100-fold) compared to mice with OVA-specific CD8 T cells (Figure 25). CSP and TRAP specific CD8 T cell responses reduced the PCR signal to near background levels of the assay (indicated by the unchallenged naïve group), a result in line with their ability to provide sterilizing immunity (Table III). In contrast, DC-LM generated GAP and S specific CD8 T cells did not reduce the liver parasite burden relative to mice containing OVA-specific CD8 T cells (Figure 25). This result indicates that GAP and S specific CD8 T cells do not provide detectable control of liver-stage infection, and these antigens would not be useful target antigens for subunit vaccination. Taken together, these data indicate that despite similar magnitude, phenotype and 107
129 functional qualities, control of liver-stage Plasmodium by mono-specific memory CD8 T cells depends on the precise epitope specificity. DC-LM generated GAP and S specific CD8 T cells recognize infected hepatocytes less efficiently than CSP , and TRAP specific CD8 T cells Despite having similar CD8 T cell intrinsic features (i.e. phenotype, cytokine production and in vivo killing capacity) to CSP and TRAP specific CD8 T cells, GAP and S specific CD8 T cells did not reduce liver parasite burden of sporozoite challenged DC-LM vaccinated mice (Figure 25). Thus, I hypothesized GAP and S specific CD8 T cell responses fail to reduce liver parasite burden due to inefficient recognition of Plasmodium-infected hepatocytes. To test this, I generated in vitro primary hepatocyte cultures from H-2 bxd mice and confirmed that the primary hepatocytes were permissive to Pb sporozoite infection (Figure 26A). CD8 T cells were enriched from spleens of DC-LM CSP , GAP , S , or TRAP vaccinated mice by depletion of B cells and CD4 T cells, and were added to sporozoite-infected hepatocyte cultures at various time points post infection and then analyzed for the percent of CD8 T cells producing IFNγ as a measure of hepatocyte antigen presentation. Data were normalized to the maximum percentage of IFNγ producing cells for each specificity after stimulation with exogenous cognate peptide added to the hepatocyte culture. Representative data for CSP and GAP specific CD8 T cells (Figure 26B) indicate that a substantial fraction (~20-108
130 23%) of CSP specific CD8 T cells produced IFNγ after culture with 20 hourinfected hepatocytes when only a small fraction (1-2%) of GAP specific CD8 T cells produced IFNγ (Figure 26B). Similarly, incubation of TRAP specific CD8 T cells with 20 hour-infected hepatocytes led to a detectable frequency of IFNγ producers (~10%)(Figure 26C). In contrast, incubation of GAP or S specific CD8 T cells with 20 hour-infected hepatocytes yielded only 1% IFNγ producers (Figure 26C). In mouse models of malaria, liver-stage infection lasts approximately hours, therefore it is possible that a 20 hour-infection did not allow enough time for GAP50 or S20 antigens to be processed and presented by infected hepatocytes. To address this, CSP , GAP , S , or TRAP specific CD8 T cells were added to 40 hour-infected hepatocytes. Although CSP and TRAP expression is limited to sporozoites and they would be considered early antigens, a detectable frequency of CSP and TRAP specific CD8 T cells (~8% and 12%, respectively) still produced IFNγ when added to 40 hour infected hepatocytes, likely due to persistence of surface peptide-mhc complexes. IFNγ production by GAP and S specific CD8 T cells remained low (~1% and 3%, respectively)(figure 26D). These data indicate that after sporozoite infection of hepatocytes the CSP and TRAP epitopes are more efficiently recognized by CD8 T cells than GAP and S epitopes, highlighting the utility of this assay approach in identifying and selecting antigenic targets for further characterization and inclusion in subunit vaccines. 109
131 Discussion My results reveal that detection of a liver-stage antigen-specific CD8 T cell response after immunization with whole parasite vaccines, such as RAS, does not ensure the specific liver-stage antigen will serve as a target of protective immunity after subunit vaccination. This dichotomy between CD8 T cell priming and protective immunity was not CD8 T cell intrinsic, but rather based on the efficiency by which subunit vaccineinduced CD8 T cells could recognize infected hepatocytes, the primary target cell for protective liver-stage immunity (200). These data, which may have important implications for subunit vaccine design, were made possible by the description of novel CD8 T cell epitopes through the work of others (136, 157). My results demonstrate that, despite the capacity to generate similar functional capacity and extreme large memory CD8 T cell populations, some epitopes (GAP and S ) do not serve as targets of protective CD8 T cells, which provides strong evidence that the specificity of the CD8 T cell response is a critical issue for generating protective immunity by subunit immunization. Because the Plasmodium parasite encodes ~5,000 open reading frames (142, 143), the identification of protective antigens is difficult to determine systematically. Therefore, it is important to understand the features of protective antigens, and apply this knowledge to the rational design of subunit vaccines. For example, our results suggest that expression on the surface of sporozoites (ie. CSP and TRAP) may be a shared characteristic of antigens that can serve as targets of CD8 T cells that control liver-stage Plasmodium infection. 110
132 Previous studies evaluated TRAP and S specific CD8 T cells for the capacity to contribute to protective liver-stage immunity (136). TRAP specific CD8 T cells clearly contribute to control of liver parasite burden in RAS immunized mice and were capable of reducing parasite burden as the sole mediators of immunity in viral vector immunization studies. However, TRAP specific CD8 T cells failed to provide sterilizing immunity, perhaps due to the analyses in C57Bl/6 mice that are notoriously difficult to protect from Pb. No evidence of protection by S specific CD8 T cells could be detected. Of note, the S specific CD8 T cell response was lower than the TRAP specific CD8 T cell response in RAS immunized mice and this also manifested as a substantial reduction in S specific in vivo cytolysis (136). Thus, another potential explanation is that insufficient S specific CD8 T cells were engendered by RAS immunization to meaningfully assess their contribution to protection in the presence of other protective CD8 T cell specificities. Conclusive demonstration that an antigen can serve as a target of protective CD8 T cells requires a model where the functional qualities and numbers of CD8 T cells meet or exceed those of other specificities, such as CSP, that are able to provide protective immunity (134). Finally, I examined protection by CD8 T cells specific for GAP50, an antigen expressed at both the sporozoite and blood-stage of infection (157). Although GAP specific CD8 T cells have been associated with experimental cerebral malaria (ECM) in C57Bl/6 mice, the H-2 bxd mice used in our study are not susceptible to ECM. Thus, a GAP specific CD8 T cell response is not inherently linked to ECM and the potential of these CD8 T cells to provide liver-stage immunity remained unclear. Here I examined a system 111
133 where single-specificity CD8 T cell numbers were substantially larger in magnitude than induced by RAS vaccination (Figure 3B)(136), and I used H-2 bxd mice that are easier to protect compared to C57Bl/6 mice (98, 203). Additionally, the use of H-2 bxd mice permitted us to compare TRAP , S and GAP specific memory CD8 T cell function, phenotype and protective capacity with the gold standard CSP protective epitope. These key differences of our model help discriminate between protective and non-protective CD8 T cell specificities. The efficiency of antigen presentation can alter the protective capacity of CD8 T cell responses. MHC class I antigen presentation to CD8 T cells can occur through either the endogenous direct MHC class I pathway, or alternatively through cross-presentation (218). Delivery of sporozoite antigen by mosquito bite infection or intradermal injection primes CD8 T cell responses to Plasmodium antigen largely due to cross-presentation by professional APCs in secondary lymphoid organs outside of the liver (196, 198). However, in order to clear infected hepatocytes, CD8 T cells have to recognize their cognate peptide presented by MHC class I molecules on the surface of infected hepatocytes. Antigen presentation by infected hepatocytes requires cytosolic availability of the target antigen and its subsequent degradation by the proteasome, to produce peptides to be presented in the complex with MHC class I. These peptides may be generated by direct or cross-presentation by hepatocytes (219). However, cross presentation likely involves uptake of antigen from non-viable parasites whereas direct presentation ensues from productively infected hepatocytes. Therefore, due to efficiency of presentation, antigens from viable parasites presented via the endogenous MHC I 112
134 pathway likely serve as optimal targets of protective CD8 T cell responses. Collectively, our data demonstrate that the efficiency of antigen presentation by infected hepatocytes for CD8 T cell recognition is an important factor to identify target antigens to induce sterilizing immunity (Figure 26). Given the potentially large number of candidate antigens that could be identified from Plasmodium immunized or infected subjects, additional assays to further trim down vaccine relevant candidate antigens seem essential. Antigen localization in the parasite may be an important factor in determining the efficiency of antigen presentation and subsequent CD8 T cell recognition. Antigens that are secreted, or easily accessible to the cytosol of the infected cell, are optimal for processing and presentation via the endogenous, direct pathway of MHC class I presentation. The importance of antigen localization, or compartmentalization, in antigen presentation has been described in infection settings where CD8 T cells are major contributors to pathogen clearance (201, ). As previously mentioned, a recent study demonstrated that immunization of mice containing 2 x 10 5 OT-I TCR Tg cells with Pb RAS-expressing secreted OVA induced better liver-stage parasite control than Pb RAS-expressing non-secreted OVA, when each group was challenged with the homologous recombinant parasite. However, interpretation of this study remains complex because the immunized mice also contained the full spectrum of Plasmodium-specific CD8 T cells, not just those specific for the recombinant antigen, and each group was challenged with a different recombinant parasite (202). In contrast, our results evaluate the capacity of single epitope-specific CD8 T cells, specific for four distinct Plasmodiumderived antigens, to recognize sporozoite-infected hepatocytes and provide protection in 113
135 vivo. Here I have shown that Plasmodium sporozoite surface protein-derived epitopes (ex: CSP and TRAP ) are more efficiently presented to CD8 T cells after Pbinfection of hepatocytes relative to intracellular protein-derived epitopes (ex: GAP and S )(Figure 26). Interestingly, these results directly correlate with the capacity of epitope-specific CD8 T cells to reduce liver parasite burden (Figure 25), and more importantly the capability of providing sterilizing immunity (Table III). Thus, I have described an assay that readily identifies antigenic targets that are not efficiently recognized by CD8 T cells in infected hepatocytes and therefore are likely not protective CD8 T cells for liver-stage infection. Because surface proteins of sporozoites can serve as protective antigens, surface proteomics screens may identify additional candidate antigens for evaluation. However, due to the small number of epitopes screened in our study (n=4) it is not possible to strongly predict that all CD8 T cell targeted antigens derived from sporozoite surface proteins will serve as protective antigens. As new CD8 T cell epitopes are described, these epitopes could be screened in a similar manner as described here. In sum, the assay I describe here can be readily applied to rodent models of malaria and may aid in identification of protective antigens. In turn, this information may provide additional basic information on features of protective antigens that may be applied to subunit vaccine design for P. falciparum. It should be noted that protective CD8 T cell target antigens are likely not restricted to surface proteins of sporozoites. For example, immunization with late arresting genetically attenuated sporozoites (GAP) induced a more robust CD8 T cell response and better protection than RAS or early-arresting GAP (81). These data suggest 114
136 that antigens expressed during the replication of parasites within hepatocytes can also be targets of protective immunity, if they can be efficiently presented by hepatocytes via MHC class I. Finally, antigen abundance may be critical and, at least for some antigens, may be developmentally regulated. For example, the GAP50 protein is essential for parasite motility and invasion of both hepatocytes and RBCs (159). However, this protein would likely not play a major role during liver-stage replication and may not be expressed at high levels during the replication and differentiation of parasites within hepatocytes. Thus, multiple factors (i.e. antigen localization, abundance, timing in infection, etc) likely collectively contribute to defining a protective antigen. Identification of additional CD8 T cell epitopes, particularly liver-stage antigens, will be critical to further elucidate the mechanism(s) that define protective antigen. In summary, I describe a dichotomy between the priming of, and protectionmediated by CD8 T cells as well as an assay to screen candidate antigens to lead to the successful design of a CD8 T cell-mediated subunit vaccine against malaria. Our data indicate that not all Plasmodium-specific CD8 T cells primed by whole parasite vaccination can provide sterilizing immunity following sporozoite challenge. Antigen localization, and/or relative protein abundance, may be important criteria in the determination of a protective antigen due to accessibility to endogenous MHC presentation machinery that likely affects the efficiency of peptide-mhc presentation in hepatocytes and subsequent CD8 T cell recognition. Since current vaccination strategies are suboptimal (92, 192, 223), or have complications that may limit their use in the field 115
137 (70), it remains important to characterize protective CD8 T cell responses against Plasmodium to aid in the design of efficacious subunit vaccines. 116
138 Table 2. Infection with P. berghei ANKA sporozoites primes CD8 T cell responses to several identified peptide epitopes Antigen 1 Epitope Restriction Element Peptide Reference Circumsporozoite protein (CSP), abundant surface protein (PBANKA_040320). Glideosome associated protein 50 (GAP50), intracellular protein required for gliding motility (PBANKA_081900). Sporozoite-specific gene 20 (S20), hypothetical protein (PBANKA_142920). Thrombospondin related adhesion protein (TRAP), surface protein (PBANKA_134980). CSP H-2K d SYIPSAEKI Romero et al GAP H-2D b SQLLNAKYL Howland et al S H-2K b VNYSFLYLF Hafalla et al TRAP H-2D b SALLNVDNL Hafalla et al Relevant data pertaining to four described P. berghei ANKA CD8 T cell epitopes. Accession numbers for PlasmoDB database are listed, as well as the amino acid sequences and restriction element of the described epitopes. 117
139 Figure 14. Radiation-attenuated sporozoite vaccination primes CSP , GAP , S , and TRAP specific CD8 T cell responses BALB.b x BALB/c F1 mice were immunized I.V. with 10 4 Pb-RAS and splenocytes were harvested at day 0 or 6 days to analyze CD8 T cell responses. (A), Flow plots analyzing antigen-experienced (CD8α lo, CD11a hi ) CD8 T cell phenotype at day 0 or day 6 post RAS vaccination. (B-C), Representative flow plots of IFNγ-producing CD8α lo, CD11a hi T cells (B) and CD8α hi, CD11a int T cells (C) following peptide stimulation 6 days after RAS vaccination. (D), Cumulative data from two independent experiments, N=6 total mice. Error bars indicate mean +/- S.E.M. with no statistical differences between groups by one-way ANOVA analysis. 118
140 119
141 Figure 15. Dendritic cell prime, recombinant L. monocytogenes boost (DC- LM) vaccination generates large Plasmodium-specific memory CD8 T cell responses in C57Bl/6 mice Naïve C57Bl/6 mice were primed with 5 x 10 5 LPS-matured DCs coated with a single Plasmodium peptide (GAP , S , or TRAP ). Mice were boosted seven days following prime with 1-2 x 10 7 CFUs of rlm expressing the same Plasmodium peptide used in priming. Peripheral blood of DC-LM vaccinated mice was analyzed at least >60 days post boost for frequency of Plasmodium-specific CD8 T cell response by MHC class I tetramer staining. (A), Flow plots illustrating representative Plasmodium-specific memory CD8 T cell responses. (B), Cumulative data from memory (>60 days post LM boost) Plasmodium-specific CD8 T cell responses. Combined data from two independent experiments, N=5-10 total mice per group. Error bars indicate mean +/- S.E.M. Statistical comparison via one-way ANOVA. *** indicates p <0.001, ** indicates p <0.01, * indicates p <0.05. The number of mice (protected/total) after 10 3 Pb sporozoite challenge is indicated. (C), DC-LM vaccinated and unvaccinated control mice were challenged with 10 3 Pb sporozoites and monitored for survival. Data from two combined experiments, N=5-10 mice per group. Statistical comparison via Log Rank Mantel Cox survival analysis of DC-LM vaccinated groups to unvaccinated control. *** indicates p <0.001, ** indicates p <0.01, * indicates p <
142 121
143 Figure 16. DC-LM TRAP vaccination of BALB.b mice generates a sterilely protective response in the majority of vaccinated mice BALB.b mice were primed with 5 x 10 5 LPS-matured DCs coated with a single Plasmodium peptide (GAP , S , or TRAP ). Mice were boosted seven days following prime with 1-2 x 10 7 CFUs of rlm expressing the same Plasmodium peptide used in priming. Peripheral blood of DC-LM vaccinated mice was analyzed >60 days post boost for frequency of Plasmodium-specific CD8 T cell response by MHC class I tetramer staining. (A), Flow plots illustrating representative Plasmodium-specific memory CD8 T cell responses. (B), Cumulative data from memory Plasmodium-specific CD8 T cell responses. Combined data from two independent experiments, N=16 or 17 mice per group. Error bars indicate mean +/- S.E.M. The number of mice (protected/total) after 10 3 Pb sporozoite challenge is indicated. Statistical comparison via one-way ANOVA. Top: Comparison of CD8 T cell responses. Bottom: Comparison of protection. *** indicates p < 0.001, * indicates p <
144 123
145 Figure 17. DC-LM CSP, GAP50, S20, or TRAP vaccination of CB6.F1 mice leads to easily detectable epitope-specific memory CD8 T cell responses CB6.F1 (C57Bl/6 x BALB/c F1) mice were primed with 5 x 10 5 DCs coated with a single Plasmodium peptide (CSP , GAP , S , or TRAP ). Mice were boosted seven days following prime with 1-2 x 10 7 CFUs of rlm expressing the same Plasmodium peptide used in priming. Peripheral blood of DC-LM vaccinated mice was analyzed >60 days post boost for frequency of Plasmodium-specific CD8 T cell response by MHC class I tetramer staining. (A), Flow plots illustrating representative Plasmodiumspecific memory CD8 T cell responses. (B), Cumulative data from memory Plasmodium-specific CD8 T cell responses. Combined data from three independent experiments, N=10 or 17 mice per group. Error bars indicate mean +/- S.E.M. Statistical comparison via one-way ANOVA. ** indicates p <0.01, * indicates p <0.05. Groups with significant differences are not individually indicated. N indicates number of mice in group. 124
146 125
147 Figure 18. DC-LM GAP50 or S20 vaccinated BALB.b x BALB/c F1 mice have larger magnitude epitope-specific responses compared to DC-LM CSP or TRAP vaccinated mice BALB.b x BALB/c F1 mice were primed with 5 x 10 5 DCs coated with a single Plasmodium peptide (CSP , GAP , S , or TRAP ). Mice were boosted seven days following prime with 1-2 x 10 7 CFUs of rlm expressing the same Plasmodium peptide used in priming. Peripheral blood of DC-LM vaccinated mice was analyzed >60 days post boost for frequency of Plasmodium-specific CD8 T cell response by MHC class I tetramer staining. (A), Flow plots illustrating representative Plasmodium-specific memory CD8 T cell responses. (B), Cumulative data from memory Plasmodium-specific CD8 T cell responses. Combined data from two independent experiments, N=10 total mice per group. Error bars indicate mean +/- S.E.M. Statistical comparison via one-way ANOVA. *** indicates p < 0.001, ** indicates p < 0.01, * indicates p < 0.05, n.s.= not-significant. 126
148 127
149 Table 3. DC-LM generated CSP and TRAP specific CD8 T cells can provide sterilizing protection from sporozoite challenge DC-LM vaccination 1 %Protected #Protected/ Statistical analysis Challenged CSP % 14/31 p = GAP % 0/23 n.s. S % 0/20 n.s. TRAP % 6/24 p = Unvaccinated 0% 0/18 N/A 1 BALB.b x BALB/c F1 and CB6.F1 mice were DC-LM vaccinated. DC-LM vaccinated mice (>60 days post LM boost) and unvaccinated mice were challenged with 10 3 virulent P. berghei ANKA sporozoites I.V. and assessed for protection from blood-stage infection by Giemsa-stained blood smear for 10 days. Individual DC-LM vaccinated groups analyzed relative to Unvaccinated group via Chi-square test with Fisher s exact test, two-tailed at 95% confidence level. n.s. indicates not significant. N/A indicates not applicable. 128
150 Figure 19. Protective and non-protective memory CD8 T cells exhibit a similar surface phenotype in PBL BALB.b x BALB/c F1 mice were DC-LM vaccinated, and >60 days post boost, CD8 T cell responses from peripheral blood were analyzed for expression of cell surface markers CD62L, KLRG1, and CD127. (A), Representative flow plots gated on CD8 + tetramer + cells. Grey indicates isotype control, and black line indicates specified cell surface marker. (B), Cumulative data illustrating frequency of CD8 + tetramer + populations positive for specified cell surface marker. Data in (B) are combined from 2-3 independent experiments. N=7 mice per group for CD62L and KLRG1 analysis from three independent experiments. N=4 per group for CD127 analysis from two independent experiments. Error bars are mean +/- S.E.M. Statistical comparison via one-way ANOVA. * indicates p < Groups without significant differences are not individually indicated. 129
151 130
152 Figure 20. Splenic-derived protective and non-protective memory CD8 T cells exhibit a similar surface phenotype BALB.b x BALB/c F1 mice were DC-LM vaccinated, and >60 days post boost, CD8 T cell responses from spleen were analyzed for expression of cell surface markers CD62L, KLRG1, and CD127. (A), Representative flow plots gated on CD8 + tetramer + cells. Grey indicates isotype control, and black line indicates specified cell surface marker. (B), Cumulative data illustrating frequency of CD8 + tetramer + populations positive for specified cell surface marker. Data in (B) are combined from 2-3 independent experiments. N=7 mice per group for CD62L and KLRG1 analysis from three independent experiments. N=4 per group for CD127 analysis from two independent experiments. Error bars are mean +/- S.E.M. Statistical comparison via oneway ANOVA. *** indicates p < 0.001, ** indicates p < 0.01, * indicates p < Groups without significant differences are not individually indicated. 131
153 132
154 Figure 21. Liver-derived protective and non-protective memory CD8 T cells exhibit a similar surface phenotype BALB.b x BALB/c F1 mice were DC-LM vaccinated, and >60 days post boost, CD8 T cell responses from liver were analyzed for expression of cell surface markers CD62L, KLRG1, and CD127. (A), Representative flow plots gated on CD8 + tetramer + splenocytes. Grey indicates isotype control, and black line indicates specified cell surface marker. (B), Cumulative data illustrating frequency of CD8 + tetramer + populations positive for specified cell surface marker. Data in (B) are combined from 2-3 independent experiments. N=7 mice per group for CD62L and KLRG1 analysis from three independent experiments. N=4 per group for CD127 analysis from two independent experiments. Error bars are mean +/- S.E.M. Statistical comparison via oneway ANOVA. * indicates p < Groups without significant differences are not individually indicated. 133
155 134
156 Figure 22. Protective and non-protective DC-LM generated splenic CD8 T cell responses are similar in cytokine expression following peptide stimulation BALB.b x BALB/c F1 (H-2 bxd ) mice were DC-LM vaccinated, and >60 days post boost, CD8 T cell responses from spleen were analyzed for expression of IFNγ, TNFα and IL-2 following cognate peptide stimulation. (A), Representative flow plots first gated on CD8 + splenocytes, then IFNγ +. (B), IFNγ + cells gated in (A) were analyzed for expression of TNFα and IL-2. (C), Cumulative frequency of single (IFNγ + ), double (IFNγ +, and TNFα + ), or triple (IFNγ +, TNFα +, and IL-2 + ) cytokine producers. Data in (C) are combined results of three independent experiments, N=6-7 total per group. Error bars are mean +/- S.E.M. One-way ANOVA analysis with multiple comparisons on double cytokine producers: CSP vs. GAP50, p < 0.05; GAP50 vs. TRAP, p < Comparison of triple cytokine producers: CSP vs. GAP50, p < 0.05; GAP50 vs. S20, p < 0.05; GAP50 vs. TRAP, p <
157 136
158 Figure 23. Protective and non-protective DC-LM generated CD8 T cell responses are similar in capacity to kill peptide-pulsed targets in vivo Naïve and DC-LM vaccinated BALB.b x BALB/c F1 (H-2 bxd ) mice were I.V. administered a mixture of 2 x 10 6 CTV hi CSP , GAP , S , or TRAP peptide-pulsed and 2 x 10 6 CTV lo unpulsed splenocytes from donor naïve mice. (A), Representative histograms of CTV staining of splenocytes harvested from naïve and DC-LM vaccinated recipient mice 4 hours post 1:1 administration of CTV hi and CTV lo donor cells. (B), Cumulative specific killing of CTV-labeled donor cells. Each dot is an individual mouse. (C), Percentage of CD8α + tetramer + cells in peripheral blood of DC-LM vaccinated mice prior to receiving CTV-labeled donor splenocytes, each dot is an individual mouse. (D), Percentage specific killing versus frequency tetramer + of CD8α + TRAP tetramer + cells from (B) and (C) respectively. Data are combined results of two independent experiments, N=4-5 total mice per group, error bars indicate mean +/- S.E.M. One-way ANOVA analysis of (A) indicates p < for every comparison against TRAP group. Analysis of correlation between tetramer frequency and % cytotoxicity of TRAP group in (D): Pearson r = 0.982, p <
159 138
160 Figure 24. Antigen sensitivity of protective and non-protective DC-LM generated CD8 T cell specificities is similar (A), The percentage of IFNγ + CD8 + T cells at >60 days post the indicated DC- LM vaccination from BALB.b x BALB/c F1 mice was determined after ex vivo stimulation with titrated concentrations of the respective peptide. Data are Mean +/- S.E.M. normalized to the percentage of IFNγ + cells at peptide saturation (500 nm). (B), Summary of the half maximal effective concentration (EC 50 ) indicating half way point between the baseline and maximum stimulation of IFNγ production by DC-LM generated single specificity responses. Data are combined results of two independent experiments, N=4 total mice per group, error bars indicate mean +/- S.E.M. No significant differences among groups in (B) as determined by one-way ANOVA comparison of the means. 139
161 140
162 Figure 25. Non-protective GAP and S specific CD8 T cell responses do not significantly reduce liver parasite burden BALB.b x BALB/c F1 (H-2 bxd ) DC-LM vaccinated mice (>14 days post LM boost with the indicated specificity) were challenged I.V. with 4 x 10 4 Pb sporozoites. Pb-specific 18s ribosomal RNA transcripts were quantified via qrt-pcr from total RNA extracted from livers of 42 hours post sporozoite challenged mice. Liver RNA from unvaccinated, unchallenged mice ( Naïve group) served as negative control. Data shown are pooled from two independent experiments, N = 4-6 mice per group. Error bars indicate mean +/- S.E.M. Statistical comparison via one-way ANOVA comparison to mice DC-LM vaccinated with irrelevant OVA epitope. *** indicates p < 0.001, ** indicates p < N.S.= not significant. LOD indicates limit of detection. 141
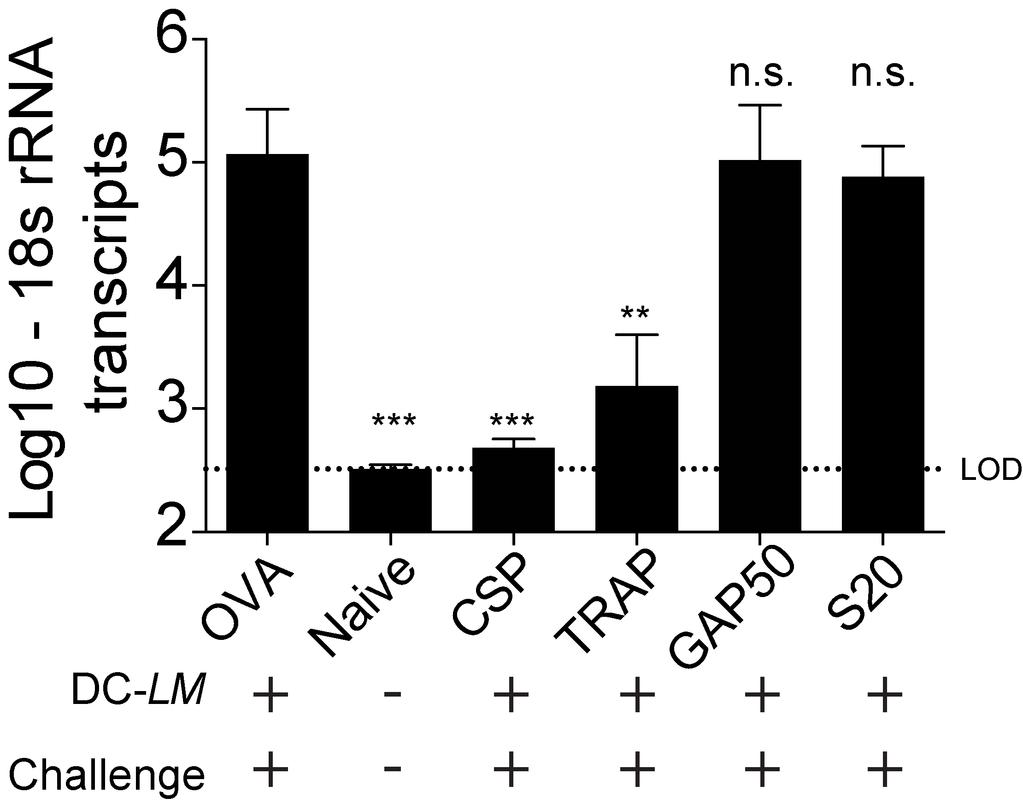
163 142
164 Figure 26. Efficient recognition of infected hepatocytes correlates with epitope-specific protection (A), CTV-labeled sporozoite infection of primary hepatocytes is dose dependent. Percentage of infected hepatocytes at 24 hours post infection with the indicated multiplicity of infection (MOI) was determined by analyzing CTV + cells by flow cytometry. Splenic CD8 T cells enriched from DC-LM vaccinated BALB.b x BALB/c F1 (H-2 bxd ) mice were added to primary hepatocyte cultures previously co-incubated with Pb sporozoites (MOI ~1) for 20 or 40 hours. (B), Representative IFNγ production by CD8 T cells following 6 hour incubation with uninfected hepatocytes, uninfected hepatocytes + exogenous cognate peptide, or 20 hour Pb-infected hepatocytes. (C), Cumulative data of IFNγ producing, Plasmodium-specific CD8 T cells added at 20 hour post-infection with sporozoites. (D), Similar to (C) but CD8 T cells were added at 40 hour post-infection with sporozoites. Data are pooled from seven experiments with each specificity, per infection time, tested at least twice. Error bars are mean +/- S.E.M. in (C) and (D). One-way ANOVA comparison of means significant differences (p < 0.05): in (C) CSP vs. GAP, CSP vs. S20, CSP vs. TRAP; TRAP vs. GAP50, TRAP vs. S20; in (D) TRAP vs. GAP
165 144
166 CHAPTER IV: FUTURE PERSPECTIVES Elucidating immune mediators of protective immunity against Plasmodium infection through WSV platforms CPS vaccination requires CD4 T cells and antibodies for protection in the stringent P. yoelii/c57bl/6 model Whole sporozoite vaccination (WSV) platforms have been used to understand the immunological requirements of protection against Plasmodium infections. These platforms have successfully provided sterilizing immunity in rodents, non-human primates, and humans (69, 72, 76, 78, 81, 99). Application of WSV in rodent models of malaria has shown that protection is largely afforded due to CD8 T cells targeting sporozoite and liver-stage expressed antigens (Figure 27A)(81, 94, 95, 98, 99). However, CD8 T cells are not the only contributors to protection. Some studies have shown that CD4 T cells and antibodies also contribute to protection (88, 98, 211). In Chapter II, I support these findings, demonstrating that CPS vaccine-induced CD4 T cells and antibodies are required for the clearance of blood-stage parasites that persist during CQ chemoprophlayxis. Specifically, I show that antibodies generated during the low-grade, transient patent parasitemia after CQ cessation, correlate with sterilizing immunity following a sporozoite challenge of CPS vaccinated mice (Figures 11 and 12). This antibody response is essential for sterilizing immunity, as CPS-vaccinated µs-aicda -/- mice (which maintain B cells, but do not secrete antibodies) cannot control parasitemia 145
167 following a sporozoite challenge and eventually succumb to hyperparasitemia. Moreover, CPS vaccinated wild-type C57Bl/6 mice, depleted of CD4 and CD8 T cells by antibody-mediated depletion, maintained sterilizing immunity following challenge with blood-stage parasites (Figure 13). These data indicate that CPS-induced memory immune responses could mediate protection against blood-stage parasites independentally of CD4 and CD8 T cells. Thus, in a stringent parasite/host model of CPS vaccination, I found a role for CD4 T cells and antibodies, with little to no detectable contribution by CPS-induced CD8 T cell response (Figures 9 and 13). Based on my data, and the current literature on CD4 T cell helper responses (reviewed in (224, 225)), it is likely that CD4 T cells are essential for the generation of potent anti-blood-stage antibody responses during the exposure to low-grade, transient blood-stage parasitemia, but CD4 T cells are not required after these antibodies are generated when CPS-vaccinated mice are challenged at a memory time point (Figure 13). Cross-stage reactive antibodies may be critical for CPS vaccine-induced protection While the role of liver-stage antigen-specific CD8 T cells in other WSV platforms (RAS and GAP) is well-defined, in the CPS vaccination model, mice are exposed to blood-stage antigens that may lead to the development of potent blood-stage or crossstage (targets liver-stage and blood-stage antigens) immunity via antibodies that are sufficient to mediate protection in the absence of a CD8 or CD4 T cell response. Indeed, evidence of the induction of cross-stage reactive antibodies following controlled bloodstage parasite exposure has been described before. BALB/c mice vaccinated with Py 146
168 265BY blood-stage parasites with 10 days of CQ chemoprophylaxis experienced patent self-limiting parasitemia that lasted up to 20 days following initial vaccine administration (88). Fifteeen days following resolution of patent parasitemia, vaccinated mice were challenged with homologous sporozoites and assessed for sterilizing immunity. Protection from sporozoite challenge was assessed by Giemsa-stained blood smear, which shown 100% of vaccinated mice were protected from sporozoite challenge in one experiment. Further, the authors reported a 95% reduction in liver parasite burden in vaccinated mice relative to controls, which is a more direct measurement of the liverstage component of cross-stage immunity than measuring sterilizing immunity (88). The authors concluded that cross-stage reactivity against liver-stage antigens could be induced by vaccination with blood-stage parasites concurrently with CQ chemoprophylaxis. I hypothesize that CPS vaccination using sporozoites similarly leads to development of cross-stage reactive antibodies (Figure 27B). I showed that antibody titers against bloodstage parasite lysate and a blood-stage specific protein (MSP-1) were increased in mice exposed to breakthrough parasitemia that occurred under the 10 day CQ regimen relative to the absence of breakthrough parasitemia under the 25 day CQ treatment (Figure 12). I predict that ELISA analysis of antibodies against intact sporozoites would show that CPS vaccination induces cross-reactive antibodies specific to sporozoites. If so, it is possible that these anti-sporozoite antibodies could block early hepatocyte infection events (115, 120, 226), and therefore provide a degree of liver-stage protection. Alternatively, antibodies generated by exposure to blood-stage parasites may be cross-reactive with antigens expressed by the liver-stage parasite. Resolving this would likely require in 147
169 vitro analysis of the inhibition of parasite development in the presence/absence of CPSinduced antibodies. Generation of immunity against heterologous Plasmodium strains WSV approaches have aided in our understanding of the immunological requirements of mediating sterilizing immunity. However, WSV studies of rodents and humans have primarily addressed vaccine-induced protection against the immunizing, homologous Plasmodium strain, and little has been done to address the protection afforded against heterologous Plasmodium infection. Some studies have indicated little to no detectable heterologous immunity using rodent models of malaria. For example, in one study CBA/Ca mice were immunized via I.P. administration of rodent-specific Plasmodium chabaudi (Pcc) strain AJ sporozoites or blood-stage parasites while concurrently receiving an anti-blood-stage parasite-specific drug (mefloquine). Pcc AJvaccinated mice showed no delay in time to patency (measured as first day blood-stage parasites are seen by Giemsa-stained blood smear) following sporozoite challenge with Pcc substrain CB (227, 228). Results were similar if vaccination was performed with Pcc CB, followed by challenge with Pcc AJ. In contrast, studies utilizing other rodent Plasmodium strains in WSV approaches have shown partial cross-species protection (81, 109, 182, 229). For example, three administrations of genetically attenuated parasite (GAP) Py fabb/f - into BALB/c mice confers sterilizing protection from heterologous challenge with wild-type Pb sporozoites in 100% of mice (5/5 mice tested)(81). Further, five administrations of Py RAS confers sterilizing protection in 54% of Pb challenged 148
170 BALB/c mice (182). The conflicting nature of the Py/Pb experiments relative to the Pcc experiments may be due to many factors such as different rodent Plasmodium species utilized in the studies, type of whole parasite attenuation, immunization schedule and number of administrations, route of administration (I.P. versus I.V), or mouse strains. These differences can only be resolved through extensive experimentation, testing these variables (i.e. types of WSV, immunization schedule, route of immunization, etc.) headto-head to best understand the protocols that engender the best heterologous immunity in each mouse strain/parasite species scenario. I have demonstrated how route of immunization and type of WSV approach impacts the magnitude of the total Plasmodium-specific CD8 and CD4 T cell responses (Figures 6 and 7) and sterilizing immunity (Table I). However, the sterilizing protection was likely enhanced in CPS vaccinated mice due to development of anti-blood-stage specific antibody responses as a result of low-grade, transient parasitemia during immunization. Importantly, this is not an observation exclusive to the C57Bl/6-Py rodent malaria model as a similar observation was subsequently observed using P. chabaudi parasites (191). Thus, multiple factors may impact the protective immune responses engendered following WSV, which can impact the degree of protection against heterologous Plasmodium challenge. Because many substrains of Plasmodium falciparum and Plasmodium vivax can be prevalent in the endemic areas ( ), these conflicting studies collectively highlight the need to better understand how to engender cross-strain immunological responses that transcend beyond the homologous, immunizing Plasmodium strain. 149
171 To better understand the optimal parameters to engender sterilizing protection against heterologous challenge, I could test CPS vaccine-induced protection against heterologous strains by utilizing rodent Plasmodium strains Py and Pb as these two strains have distinct disease manifestations in C57Bl/6 mice, differ in the kinetics and magnitude of parasitemia, and allow for the use of well-established and novel CD8 T cell epitopes to track antigen-specific CD8 T cell responses (96, 97, 136, 157, 234). CPS vaccination may be the optimal WSV approach to understand the varying immunological requirements of heterologous Plasmodium infections as CPS vaccination confers enhanced protection in rodent models relative to RAS or GAP vaccination with fewer vaccine administrations (88, 211). Specifically I could examine to what extent CPS-Py vaccination protects against challenge with Pb sporozoites. I hypothesize based on the current literature (81, 182), this single administration CPS vaccination strategy would lead to a measureable delay in time to patency, but would not provide sterilizing immunity. Adjustments to the dose of immunizing parasites, or the number of administrations are additional experiments I could perform to better understand the immunological requirements leading to sterilizing protection from heterologous sporozoite challenge. I predict that additional CPS administrations would provide sterilizing heterologous protection, which would be dependent on CD8 T cells to reduce the liver parasite burden. Experiments similar in design to those described in Chapter II would be able to resolve these predictions. 150
172 CPS vaccination could be used as a model to elucidate the induction of experimental cerebral malaria Blood-stage infection of naïve C57Bl/6 mice leads to experimental cerebral malaria (ECM) in the majority of the challenged mice. ECM, a lethal condition, is characterized as the breakdown of the blood-brain barrier, the sequestration of bloodstage parasites to the brain endothelial layer, and requires CD8 T cells and the production of IFNγ (reviewed in (235)). There are still many facets of ECM that are unclear, such as the timing of these events in relation to infection exposure, as well as if the CD8 T cells found in the brain of mice experiencing ECM symptoms are Plasmodium-specific or are non-specific. Because of the evidence of Py GAP vaccination affording protection against Pb sporozoite challenge (81), it would be interesting to know if CPS-Py vaccination would afford protection from ECM in Pb-challenged C57Bl/6 mice. If so, CPS-Py vaccination could serve as a model to understand how magnitude and duration of Pb blood-stage parasitemia leads to the induction of ECM, as well as characterize the specificity of the CD8 T cell responses that are helping drive the disease. Differences in the total activated CD8 T cell response, parasite sequestration in the brain, the kinetics of the blood-brain barrier breakdown could be compared between Pb sporozoite challenge of CPS-Py vaccinated mice versus naïve C57Bl/6 mice. More importantly, the recent description of novel H-2 b -restricted CD8 T cell epitopes derived from Pb antigens provides an opportunity to directly address the role of CPS-induced CD8 T cells in protection from challenge, or whether parasite-specific CD8 T cells play a role in the induction of ECM (136, 157, 234). Taken together, many avenues exist to further define 151
173 the immunological requirements of protection against heterologous challenges. Since vaccination of human subjects via WSV approaches have largely ignored testing of protection against heterologous challenge, experiments in rodent models of malaria could help direct the advancement of WSV or subunit approaches to achieve strain-transceding, heterologous protection against Plasmodium infections. Advancing subunit vaccination strategies through enhancing quantity and quality of protective CD8 T cell responses against liver-stage infection Localization of antigen targeted by an anti-plasmodium CD8 T cell response may dictate protective capacity Plasmodium-specific CD8 T cells targeting liver-stage expressed antigens can provide sterilizing immunity in humans and rodents (69, 71, 72, 76, 78, 98). Recently, new CD8 T cell epitopes, derived from liver-stage proteins expressed by rodent Plasmodium, have been described enhancing our ability to study Plasmodium-specific CD8 T cell responses (136, 157, 210, 234). For example, these novel epitopes have allowed for studies addressing how the specificity of the CD8 T cell response may impact protective capacity. In Chapter III, I have shown that not all Plasmodium-specific CD8 T cell responses generated following WSV infection contribute to protection against subsequent sporozoite exposure (Table III). Importantly, only two of four tested CD8 T cell specificities (CSP and TRAP specific CD8 T cells) were capable of mediating protection, and these two specificities target antigens expressed on the surface 152
174 of the Plasmodium sporozoite. These findings provide evidence that localization of the antigen during liver-stage infection may impact the ability of infected hepatocytes to present cognate peptide on surface MHC class I molecules. I hypothesized that many CD8 T cell specificities are primed following sporozoite infection, but only CD8 T cell specificities that target antigens that are surface expressed by the parasite, or easily released from the parasite parasitophorous vacuole (PV) during liver-stage infection are readily accessible to the hepatocyte peptide presentation machine for presentation by MHC class I molecules (Figure 28). Differences in the efficiency of peptide presentation would impact the ability of antigen-specific CD8 T cells to recognize infected cells. To experimentally support this, I examined the ability of DC-LM vaccine-generated CD8 T cells to recognize cognate antigen presented by sporozoite-infected hepatocytes. I found that CSP and TRAP specific CD8 T cells, the two CD8 T cell specificities capable of mediating protection in my system, produced IFNγ following recognition of their cognate antigen presented on sporozoite-infected hepatocytes. In contrast, the frequency of GAP and S specific CD8 T cells recognizing their cognate antigen was lower than CSP- and TRAP-specific CD8 T cells (Figure 26). Collectively, these data indicate that surface expression of parasite antigens may impact antigen presentation efficacy, and subsequently the capacity to be targeted by CD8 T cell responses (Figure 28). In order to lend more support to the antigen localization hypothesis beyond my data described in Chapter III, additional CD8 T cell epitopes will need to be described and screened for protective capacity. Prime-boost vaccination strategies will likely be 153
175 essential to determine the protective capacity of these new CD8 T cell epitopes through examining numerically large, individual CD8 T cell specificities in the absence of any other anti-plasmodial response. Over two decades passed between the description of the CSP epitope and the novel GAP , S , and TRAP Pb-derived CD8 T cell epitopes, thus it remains possible it may a considerable amount of time until sufficient quantity of Pb-derived CD8 T cell epitopes are described to enable further studies to potentially support this hypothesis. Therefore, an alternative way to address this quesetion would be through the generation of transgenic parasites. To this end, Pb transgenic parasites could be designed that express the GAP epitope sequence followed by the signal sequence of TRAP under control of a promoter that is expressed throughout the parasite life cycle; this transgenic parasite design (named PbGAP50sec) would theoretically enable for the GAP CD8 T cell epitope sequence to be surface expressed on the sporozoite similarly to TRAP protein, potentially enabling the eventual secretion or access of this peptide sequence to the host cell cytosol and proteolytic machinery. The construct would be designed to express hemagglutinin (HA) to test for construct expression throughout the liver-stage and blood-stages of infection. DC-LM GAP50 and OVA vaccinated mice would be challenged with high dose (>10 4 ) or low dose (10 3 ) transgenic PbGAP50sec parasites and assessed for liver parasite burden at 42 hours post infection in high dose challenged mice, or alternatively assessed for sterilizing protection via Giemsa stained blood smear in the low dose challenged group of mice. The high/low dose regiment tests a high dose commonly utilized in rodent WSV studies (88, 211), whereas the low dose is the dose required to sufficiently infect all naïve mice 154
176 (134, 236). If the location of the protein-derived epitope affects the antigen processing efficiency and eventual CD8 T cell recognition, I hypothesize this experiment would show a decrease in liver parasite burden in DC-LM GAP50 mice relative to DC-LM OVA vaccinated mice because DC-LM vaccine-induced GAP specific CD8 T cells would more efficiently detect and kill infected hepatocytes. Further, I predict DC-LM GAP50 vaccinated mice receiving low dose PbGAP50sec challenge would have delayed time to blood-stage patency relative to DC-LM OVA vaccinated mice. If these experiments provide the predicted results, this experimental approach could be extended to test the S20 epitope in a similar matter. Quantitative and qualitative features of CD8 T cell-mediated immunity as learned from novel CD8 T cell specificities To date, the majority of our understanding of the quantitative and qualitative features of protective CD8 T cell responses have tracked CD8 T cells directed against the immunodominant CSP epitopes: CSP in P. berghei and CSP in P. yoelii (96, 97, 134, 149, 155, 174, 196, 197, 217). However, the recent description of novel CD8 T cell epitopes enables the expansion of these studies particularly with the ability to track two distinct protective CD8 T cell specificities (CSP and TRAP ) and two non-protective CD8 T cell specificities (GAP and S ). Now additional data can be gathered regarding the quantitative and qualitative features of CD8 T cellmediated immunity against Plasmodium. 155
177 To date, protection in humans following WSV or prime-boost subunit vaccination has correlated with the magnitude of the Plasmodium-specific CD8 T cell response and the number of vaccine administrations (76, 78, 91). However, it is unclear what is the most optimal dose, dosing schedule, and number of vaccine administrations that provides the most durable, protective CD8 T cell response against Plasmodium. Using rodent models of malaria and four distinct CD8 T cell specificities, I can address these questions. Firstly, I would address how the total CD8 T cell response and the four distinct Plasmodium-specific CD8 T cell-specific responses are affected by the number of vaccine administrations. CB6.F1 (H-2 bxd ) mice would be RAS vaccinated with high-dose (2 x 10 4 ) or low dose (1 x 10 3 ) Pb sporozoites over 1, 2, 3, 4, or 5 administrations, with 3- week intervals between administrations. The 3-week interval allows for an accelerated vaccination schedule compared to waiting for memory time points to boost (>60 days), while also allowing for the responses to contract from the peak effector responses before another RAS vaccine administration. RAS vaccination would be used since GAP specific CD8 T cells can be induced by blood-stage infections (157), and I want to exclusively evaluate liver-stage induced immunity. I would compare the frequency and total CD8 T cell response in the spleen and the liver using surrogate activation markers and MHC class I tetramer straining. These experiments would help determine if multiple administrations do enhance the total CD8 T cell response, which would be expected by the current Plasmodium literature (78, 81, 99, 211), and if higher sporozoite doses correlate with higher magnitude CD8 T cell responses important to know for WSV approaches as generating aspetically-harvested, laboratory-reared sporozoites is a critical 156
178 limiting factor to widescale vaccination (70, 80). However, more importantly, these studies could help determine if there are differences in the capacity of protective CD8 T cell responses to be boosted relative to non-protective CD8 T cells. This is important to know because measuring the total CD8 T cell response would measure the increases in non-protective responses as well, which would not contribute to the overall protection induced by WSV approaches. Indeed, there is evidence that differential boosting of Plasmodium-specific CD8 T cell responses does occur. CPS-vaccination of BALB/c mice with P. yoelii lead to the description of the L CD8 T cell epitope derived from a parasite ribosomal protein (210). The authors shown that despite multiple administrations of CPS vaccination, the L specific CD8 T cell response did not numerically increase. However, the L specific CD8 T cell response did numerially increase using a DC-LM heterologous prime-boost approach. In contrast, the CSP specific CD8 T cell response numerically increased following CPS administrations or DC-LM prime-boost. The CSP specific CD8 T cell response is known to mediate protection (155, 183, 203), but to date the protective capacity of the L specific CD8 T cell response has not been published. Thus, a better characterization of the induction, boosting, and maintenance of protective and non-protective anti-plasmodium CD8 T cell responses could lead to understanding of how to specifically enhance only the CD8 T cell responses that meaningfully contribute to protective immunity. It is important to understand how to generate protective immunity in a manner that utilizes the least amount of time and resources to aid in achieving the goal of efficacious, widescale vaccination of humans against Plasmodium. 157
179 Beyond specificity: localization of the CD8 T cell response may matter Tissue-resident CD8 T cell populations have become an area of increasing interest in the fields of muscosal immunology ( ), as well as in the field of malaria (240, 241). Increased technological advances in cellular imaging, particularly intravital microscopy (IVM)(242, 243), have the potential to reveal information regarding the location of CD8 T cell responses following vaccination, or challenge. Multiple studies have provided evidence that protective Plasmodium-specific CD8 T cells target and kill liver-stage-infected hepatocytes (103, 199, 200, 244). More recently, IVM was applied to study the mechanism in which CD8 T cells target Plasmodium-infected hepatocytes. Using GFP-expressing sporozoites to infect mice, a loss of GFP fluorescence was observed in hepatocytes surrounded by clusters of in vitro generated CSP-specific effector CD8 T cells, suggesting the direct killing of Plasmodium-infected hepatocytes via a mechanism requiring multiple antigen-specific CD8 T cells (200). These observations help provide an explanation regarding the large numerical CD8 T cell requirement for protection since several CD8 T cells were associated with killing of a single infected hepatocyte (134). However, it is still unclear where protective Plasmodium-specific CD8 T cells are localized following WSV vaccination, and the movements of these cells upon challenge. There is an increasing interest in the role of tissue-resident memory CD8 T cell populations generated following infection or vaccination in multiple models (reviewed elsewhere ( )). However, to date it remains unclear how the resident memory populations identified in the skin, brain, and 158
180 mucosal tissues compare phenotypically and functionally to populations of Plasmodiumspecific CD8 T cells localized in the liver of vaccinated mice. It will be important to thoroughly describe Plasmodium-specific CD8 T cells within the liver, particularly in regards to whether these cells are resident within the parenchymal tissue, or alternatively, whether they are associated with the endothelial barrier of the liver sinusoids. Recent IVM data suggests that hepatitis B virus-specific in vitro generated effector CD8 T cells in the liver are localized in the vasculature and do not require migration into liver parenchymal tissue to kill infected hepatocytes (245). While this is a different infection setting, it is possible that Plasmodium-specific CD8 T cells in the liver exhibit a similar location in the vasculature during killing of parasite-infected hepatocytes. This result has important implications as most studies of liver-resident memory CD8 T cells in Plasmodium have utilized perfusion techniques to eliminate circulating cells, but it is possible perfusion techniques may dissociate important resident CD8 T cell populations, closely associated with the sinusoids, from analysis. To date, liver resident memory populations generated by WSV have been described as CD8 T cells that remain in the liver following perfusion (240, 241), but their contribution to protective immunity is unclear. CXCR6 expression has also been described as an important molecule involved in CD8 T cell liver homing and residence (246). Expression of chemokine receptor CXCR6 may be a marker to identify these liver resident Plasmodium-specific CD8 T cell populations (240, 241). For example, CXCR6 expression on CD8 T cells was shown to be required for long-term maintenance of Plasmodium-specific CD8 T cell populations in the liver. This was shown by an 159
181 experimental design initially requiring transfer of either CXCR6 +/+ or CXCR6 -/- CD OT-I CD8 TCR Tg CD8 T cells into wild-type BALB/c recipient mice, followed by RAS immunization with P. berghei CS 5M, a P. berghei strain which has the H-2K d -restricted CSP epitope (SYPSAEKI) altered to the OVA epitope (SIINFEKL) by targeted substitutions (197). Immunization with Pb CS 5M -RAS thus induces OVA-specific CD8 T cell responses that can be tracked by OVA-specific CD8 T cell tetramer staining or alternatively through staining for an allelically disparate marker (CD45.1 in this case). Wild-type BALB/c mice receiving CXCR6 +/+ OT-I cells prior to Pb CS 5M -RAS immunization maintained OVA-specific liver CD8 T cell responses into memory timepoints, but this response was not maintained in livers of mice receiving CXCR6 -/- OT-I cells, indicating a requirement for CXCR6 expression for maintance of Plasmodium-specific liver CD8 T cells. Further studies need to define these liverresident CD8 T cells beyond the requirement for CXCR6 such as their protective capacity and how they differ from circulating CD8 T cell populations. Additionally, it will be important to characterize any differences from the more strict definitions of resident T cell populations in other tissues and model systems ( ). It remains important to determine the true location of these populations, the signals that draw and maintain these CD8 T cell populations, and the relative time they remain associated with the liver sinusoids. Comparisons of Plasmodium-specific CD8 T cell populations in the liver with more strictly defined resident memory CD8 T cell populations will be helpful in determining if unique features exist, their importance in vaccine-induced protection and 160
182 how to create or modify liver-localized CD8 T cell responses against Plasmodium to foster better liver-stage immunity. Recently, the Harty laboratory has acquired essential tools to elucidate the localization of Plasmodium-specific CD8 T cell populations in the liver, such as the generation of mcherry-expressing TRAP TCR retrogenic mice (247), CXCR6 GFP knockin mice (CXCR6 GFP/+ )(248), and a two photon microscope to enable intravital imaging of cellular events (249). I hypothesize that liver-associated memory Plasmodium-specific CD8 T cells are generated following WSV approaches or infection, and these CD8 T cells express CXCR6 and localize to the sinusoids, not the parachyma in the liver. To test this, I would adoptively transfer TRAP specific CD8 T cells from a retrogenic donor into naïve CB6.F1 receipient mice. CB6.F1 receipient mice would then be vaccinated with a RAS vaccination regimen that induces the largest number of Plasmodium-specific CD8 T cells, as determined by experiments previously discussed in this future perspectives chapter. I would use intravital microscopy to image CXCR6 +, TRAP specific CD8 TCR retrogenic cells in the liver, particularly noting the localization of these cells in RAS-vaccinated mice receiving wild-type sporozoite challenge compared to non-challenged, RAS-vaccinated mice. These studies will aid in the understanding of the localization, and movement of these liver-associated CD8 T cell memory populations. These types of experiments can be used to determine how to foster better liver-stage immunity against Plasmodium. 161
183 Conclusion Our understanding of the immunological requirements to engender sterillzing immunity against Plasmodium has improved to the point that both WSV and subunit approaches can achieve sterilizing immunity in vaccinated human subjects (72, 78, 85, 90, 131, 194). However, there are severe limitations to each of these vaccination approaches that, at least to date, will likely prevent their use to achieve the goal of worldwide eradication of Plasmodium. With the advent of novel CD8 T cell epitopes to track Plasmodium-specific CD8 T cell responses against rodent Pb antigens, in addition to development of CD8 TCR retrogenic mice and continued enhancement of live imaging microcsopy tools, our understanding of the CD8 T cell requirements for protection will improve, which should enhance the efficacy of subunit vaccination approaches in the future. However, it remains possible subunit vaccination approaches that exclusively rely on CD8 T cells to mediate protection may never achieve high efficacy. Therefore, it is plausible that a successful subunit vaccine approach will need to include anti-plasmodial CD4 T cells and antibody responses to best achieve cross-stage, and crossstrain/heterologous protection. Regardless of which arm(s) of the immune response are required to ultimately achieve this end goal, due to their relative accurate ability to predict protective outcomes in vaccinated human subjects, rodent models of malaria will continue to be critical to understand the immunological requirements of protection against Plasmodium infections. 162
184 Figure 27. Whole sporozoite vaccination induce anti-plasmodium CD8 T cells and antibodies mediate protection against Plasmodium infections WSV strategies (i.e. RAS and GAP vaccination) expose the host to sporozoite antigens, inducing anti-sporozoite antibody responses capable of partial inhibition of sporozoiteinfection of hepatocytes and the induction of anti-liver-stage directed CD8 T cell responses (A). In contrast, while CPS vaccination also induces these responses, protection in a C57Bl/6-P. yoelii model appears to be dependent on antibody responses direct against blood-stage antigens (B). 163
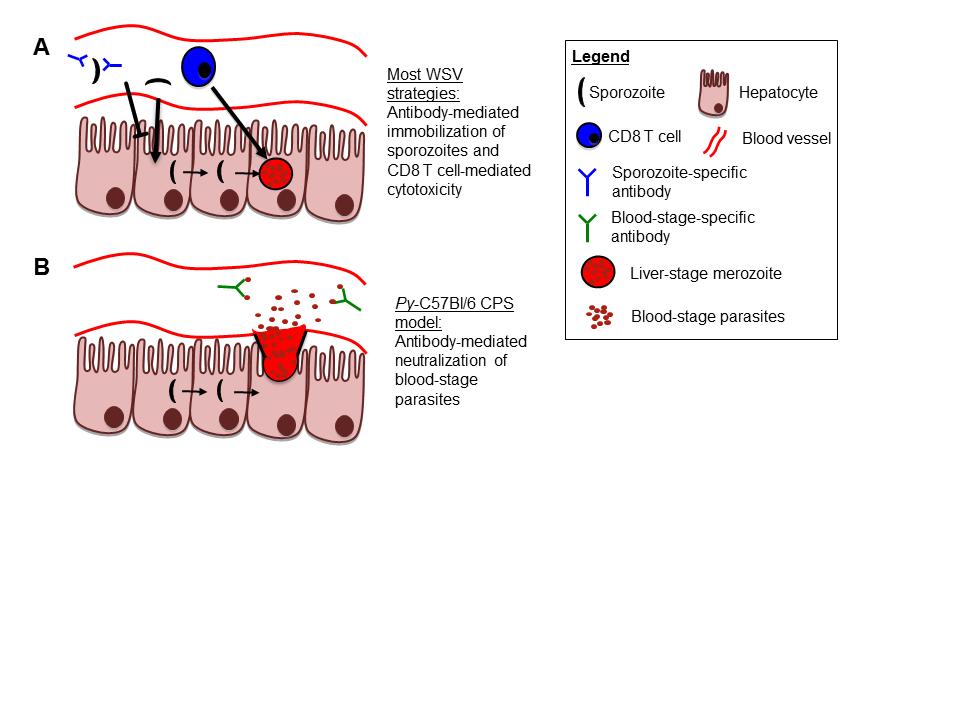
185 164
186 Figure 28. Proposed model defining a differentiating factor between antigens targeted by protective and non-protective anti-plasmodium CD8 T cell responses Non-protective CD8 T cells (i.e. GAP and S specific CD8 T cell responses) may target parasite antigens that are not easily accessible to the host cell cytosol to access proteolytic machinery for peptide processing and presentation on hepatocyte surface by MHC class I complexes (A). Protective CD8 T cell responses (i.e. CSP and TRAP specific CD8 T cells) may target parasite antigens that are easily accessible to host cell cytosol, allowing efficient processing and presentation on MHC class I molecules, and subsequent antigen-specific CD8 T cell recognition and killing (which may be IFNγ dependent). 165
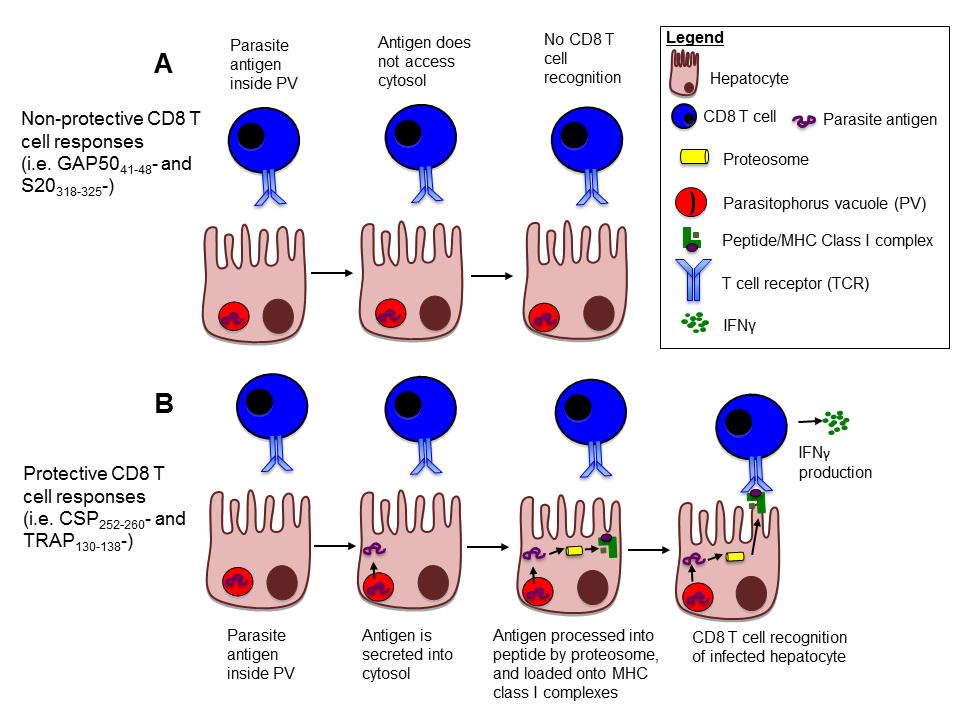
187 166
Malaria parasite vaccine development Strategies & Targets
 Malaria parasite vaccine development Strategies & Targets Tulane University Ahmed Aly Most malaria disease deaths are among children and pregnant women A child or a pregnant woman dies of malaria nearly
Malaria parasite vaccine development Strategies & Targets Tulane University Ahmed Aly Most malaria disease deaths are among children and pregnant women A child or a pregnant woman dies of malaria nearly
Therapeutic PD L1 and LAG 3 blockade rapidly clears established blood stage Plasmodium infection
 Supplementary Information Therapeutic PD L1 and LAG 3 blockade rapidly clears established blood stage Plasmodium infection Noah S. Butler, Jacqueline Moebius, Lecia L. Pewe, Boubacar Traore, Ogobara K.
Supplementary Information Therapeutic PD L1 and LAG 3 blockade rapidly clears established blood stage Plasmodium infection Noah S. Butler, Jacqueline Moebius, Lecia L. Pewe, Boubacar Traore, Ogobara K.
 Malaria parasites Malaria parasites are micro-organisms that belong to the genus Plasmodium. There are more than 100 species of Plasmodium, which can infect many animal species such as reptiles, birds,
Malaria parasites Malaria parasites are micro-organisms that belong to the genus Plasmodium. There are more than 100 species of Plasmodium, which can infect many animal species such as reptiles, birds,
Malaria vaccines where are we now?
 Malaria vaccines where are we now? Lars Hviid (lhviid@sund.ku.dk) Centre for Medical Parasitology Department of Immunology and Microbiology Slide 1 Background: A very interested but disinterested real
Malaria vaccines where are we now? Lars Hviid (lhviid@sund.ku.dk) Centre for Medical Parasitology Department of Immunology and Microbiology Slide 1 Background: A very interested but disinterested real
Ph.D. Thesis: Protective immune response in P.falciparum malaria 2011 CHAPTER I: Introduction. S.D. Lourembam 16
 CHAPTER I: Introduction S.D. Lourembam 16 1. INTRODUCTION Malaria remains a major global health problem with 300 to 500 million clinical infections and more than a million deaths reported each year. In
CHAPTER I: Introduction S.D. Lourembam 16 1. INTRODUCTION Malaria remains a major global health problem with 300 to 500 million clinical infections and more than a million deaths reported each year. In
Malaria. Population at Risk. Infectious Disease epidemiology BMTRY 713 (Lecture 23) Epidemiology of Malaria. April 6, Selassie AW (DPHS) 1
 Epidemiology of Malaria. April 6, Selassie AW (DPHS) 1") Infectious Disease Epidemiology BMTRY 713 (A. Selassie, DrPH) Lecture 23 Vector-Borne Disease (Part II) Epidemiology of Malaria Learning Objectives 1. Overview of malaria Global perspectives 2. Identify
Infectious Disease Epidemiology BMTRY 713 (A. Selassie, DrPH) Lecture 23 Vector-Borne Disease (Part II) Epidemiology of Malaria Learning Objectives 1. Overview of malaria Global perspectives 2. Identify
Invest in the future, defeat malaria
 Invest in the future, defeat malaria Malaria is caused by parasites from the genus Plasmodium, which are spread to people by infected mosquitoes. There are five species of Plasmodium that can infect humans.
Invest in the future, defeat malaria Malaria is caused by parasites from the genus Plasmodium, which are spread to people by infected mosquitoes. There are five species of Plasmodium that can infect humans.
Naive and memory CD8 T cell responses after antigen stimulation in vivo
 University of Iowa Iowa Research Online Theses and Dissertations Summer 2011 Naive and memory CD8 T cell responses after antigen stimulation in vivo Matthew David Martin University of Iowa Copyright 2011
University of Iowa Iowa Research Online Theses and Dissertations Summer 2011 Naive and memory CD8 T cell responses after antigen stimulation in vivo Matthew David Martin University of Iowa Copyright 2011
5/11/2009. Information About Malaria. What is Malaria? How many people think Malaria is caused by mosquitoes? Evolution & History
 Information About Malaria Evolution & History Etiology, Host Specific Parasites & Symptoms & Pathology (Human) Treatments (Human) What is Malaria? How many people think Malaria is caused by mosquitoes?
Information About Malaria Evolution & History Etiology, Host Specific Parasites & Symptoms & Pathology (Human) Treatments (Human) What is Malaria? How many people think Malaria is caused by mosquitoes?
Unit 5 The Human Immune Response to Infection
 Unit 5 The Human Immune Response to Infection Unit 5-page 1 FOM Chapter 21 Resistance and the Immune System: Innate Immunity Preview: In Chapter 21, we will learn about the branch of the immune system
Unit 5 The Human Immune Response to Infection Unit 5-page 1 FOM Chapter 21 Resistance and the Immune System: Innate Immunity Preview: In Chapter 21, we will learn about the branch of the immune system
Third line of Defense
 Chapter 15 Specific Immunity and Immunization Topics -3 rd of Defense - B cells - T cells - Specific Immunities Third line of Defense Specific immunity is a complex interaction of immune cells (leukocytes)
Chapter 15 Specific Immunity and Immunization Topics -3 rd of Defense - B cells - T cells - Specific Immunities Third line of Defense Specific immunity is a complex interaction of immune cells (leukocytes)
Ultrastructural Studies on Plasmodium vivax
 Characterization of Human Malaria Parasites Ultrastructural Studies on Plasmodium vivax For the first time a detailed ultrastructural study was carried out on P. vivax. Fine structural analysis of growth
Characterization of Human Malaria Parasites Ultrastructural Studies on Plasmodium vivax For the first time a detailed ultrastructural study was carried out on P. vivax. Fine structural analysis of growth
by Peter Dumoulin A dissertation submitted to Johns Hopkins University in conformity with the requirements for the degree of Doctor of Philosophy
 STUDIES ON EXOERYTHROCYTIC DEVELOPMENT OF PLASMODIUM FALCIPARUM IN VITRO: DETECTION AND ISOLATION OF PLASMODIUM LIVER STAGES AND ANALYSIS OF CIRCUMSPOROZOITE PROTEIN ANTIGEN PROCESSING by Peter Dumoulin
STUDIES ON EXOERYTHROCYTIC DEVELOPMENT OF PLASMODIUM FALCIPARUM IN VITRO: DETECTION AND ISOLATION OF PLASMODIUM LIVER STAGES AND ANALYSIS OF CIRCUMSPOROZOITE PROTEIN ANTIGEN PROCESSING by Peter Dumoulin
Jing Huang, 1 Tiffany Tsao, 1 Min Zhang, 2 and Moriya Tsuji Introduction
 Mediators of Inflammation Volume 214, Article ID 728939, 6 pages http://dx.doi.org/1.1155/214/728939 Research Article Circumsporozoite Protein-Specific K d -Restricted CD8+ T Cells Mediate Protective Antimalaria
Mediators of Inflammation Volume 214, Article ID 728939, 6 pages http://dx.doi.org/1.1155/214/728939 Research Article Circumsporozoite Protein-Specific K d -Restricted CD8+ T Cells Mediate Protective Antimalaria
SUPPLEMENTARY INFORMATION
 Complete but curtailed T-cell response to very-low-affinity antigen Dietmar Zehn, Sarah Y. Lee & Michael J. Bevan Supp. Fig. 1: TCR chain usage among endogenous K b /Ova reactive T cells. C57BL/6 mice
Complete but curtailed T-cell response to very-low-affinity antigen Dietmar Zehn, Sarah Y. Lee & Michael J. Bevan Supp. Fig. 1: TCR chain usage among endogenous K b /Ova reactive T cells. C57BL/6 mice
Primer DNA sequence (5 to 3 )
") Supplemental Table 1 Oligonucleotide primers used in the study Primer DNA sequence (5 to 3 ) Plasmodium falciparum 18s forward Plasmodium falciparum 18s reverse Plasmodium falciparum MSP1 forward Plasmodium
Supplemental Table 1 Oligonucleotide primers used in the study Primer DNA sequence (5 to 3 ) Plasmodium falciparum 18s forward Plasmodium falciparum 18s reverse Plasmodium falciparum MSP1 forward Plasmodium
Third line of Defense. Topic 8 Specific Immunity (adaptive) (18) 3 rd Line = Prophylaxis via Immunization!
 (18) 3 rd Line = Prophylaxis via Immunization!") Topic 8 Specific Immunity (adaptive) (18) Topics - 3 rd Line of Defense - B cells - T cells - Specific Immunities 1 3 rd Line = Prophylaxis via Immunization! (a) A painting of Edward Jenner depicts a cow
Topic 8 Specific Immunity (adaptive) (18) Topics - 3 rd Line of Defense - B cells - T cells - Specific Immunities 1 3 rd Line = Prophylaxis via Immunization! (a) A painting of Edward Jenner depicts a cow
The Immune System. These are classified as the Innate and Adaptive Immune Responses. Innate Immunity
 The Immune System Biological mechanisms that defend an organism must be 1. triggered by a stimulus upon injury or pathogen attack 2. able to counteract the injury or invasion 3. able to recognise foreign
The Immune System Biological mechanisms that defend an organism must be 1. triggered by a stimulus upon injury or pathogen attack 2. able to counteract the injury or invasion 3. able to recognise foreign
I. Lines of Defense Pathogen: Table 1: Types of Immune Mechanisms. Table 2: Innate Immunity: First Lines of Defense
 I. Lines of Defense Pathogen: Table 1: Types of Immune Mechanisms Table 2: Innate Immunity: First Lines of Defense Innate Immunity involves nonspecific physical & chemical barriers that are adapted for
I. Lines of Defense Pathogen: Table 1: Types of Immune Mechanisms Table 2: Innate Immunity: First Lines of Defense Innate Immunity involves nonspecific physical & chemical barriers that are adapted for
I. Critical Vocabulary
 I. Critical Vocabulary A. Immune System: a set of glands, tissues, cells, and dissolved proteins that combine to defend against non-self entities B. Antigen: any non-self chemical that triggers an immune
I. Critical Vocabulary A. Immune System: a set of glands, tissues, cells, and dissolved proteins that combine to defend against non-self entities B. Antigen: any non-self chemical that triggers an immune
Immune response. This overview figure summarizes simply how our body responds to foreign molecules that enter to it.
 Immune response This overview figure summarizes simply how our body responds to foreign molecules that enter to it. It s highly recommended to watch Dr Najeeb s lecture that s titled T Helper cells and
Immune response This overview figure summarizes simply how our body responds to foreign molecules that enter to it. It s highly recommended to watch Dr Najeeb s lecture that s titled T Helper cells and
PARASITOLOGY CASE HISTORY #14 (BLOOD PARASITES) (Lynne S. Garcia)
 (Lynne S. Garcia)") PARASITOLOGY CASE HISTORY #14 (BLOOD PARASITES) (Lynne S. Garcia) A 37-year-old woman, who had traveled to New Guinea for several weeks, presented to the medical clinic with fever, chills, and rigors within
PARASITOLOGY CASE HISTORY #14 (BLOOD PARASITES) (Lynne S. Garcia) A 37-year-old woman, who had traveled to New Guinea for several weeks, presented to the medical clinic with fever, chills, and rigors within
MALARIA CONTROL FROM THE INDIVIDUAL TO THE COMMUNITY
 MALARIA CONTROL FROM THE INDIVIDUAL TO THE COMMUNITY Calvin L. Wilson MD Clinical Professor of Family Medicine and Public Health University of Colorado Anschutz OBJECTIVES 1. Understand the unique characteristics
MALARIA CONTROL FROM THE INDIVIDUAL TO THE COMMUNITY Calvin L. Wilson MD Clinical Professor of Family Medicine and Public Health University of Colorado Anschutz OBJECTIVES 1. Understand the unique characteristics
INTERVENTION MODEL OF MALARIA
 INTERVENTION MODEL OF MALARIA TAYLOR MCCLANAHAN Abstract. Every year up to about 300 million people are infected by malaria, an infectious disease caused by Plasmodium species parasites. Consequently,
INTERVENTION MODEL OF MALARIA TAYLOR MCCLANAHAN Abstract. Every year up to about 300 million people are infected by malaria, an infectious disease caused by Plasmodium species parasites. Consequently,
EXPERIMENTAL MODELS OF CEREBRAL MALARIA
 EXPERIMENTAL MODELS OF HASTINGS OZWARA, PhD MALARIA PROGRAMME INSTITUTE OF PRIMATE RESEARCH P. O BOX 24481, KAREN - 00502 NAIROBI TEL 254 20 882571/4 FAX 254 20 882546 E-MAIL: OZWARA@IPR.OR.KE INTRODUCTION
EXPERIMENTAL MODELS OF HASTINGS OZWARA, PhD MALARIA PROGRAMME INSTITUTE OF PRIMATE RESEARCH P. O BOX 24481, KAREN - 00502 NAIROBI TEL 254 20 882571/4 FAX 254 20 882546 E-MAIL: OZWARA@IPR.OR.KE INTRODUCTION
University of Veterinary and Animal Sciences, Bikaner), V.P.O. Bajor, Dist. Sikar, Rajasthan, India
, V.P.O. Bajor, Dist. Sikar, Rajasthan, India") REVIEW ARTICLE www.ijapc.com e-issn 2350-0204 Malaria, A Widely Prevalent Mosquito-Borne Infection in Humans and Recommended Herbal Therapy Subha Ganguly 1*, Satarupa Roy 2 1 Associate Department of Veterinary
REVIEW ARTICLE www.ijapc.com e-issn 2350-0204 Malaria, A Widely Prevalent Mosquito-Borne Infection in Humans and Recommended Herbal Therapy Subha Ganguly 1*, Satarupa Roy 2 1 Associate Department of Veterinary
Malaria and vaccination: an update. Robert Sauerwein Dept. of Medical Microbiology Radboud University Medical Center, Nijmegen, the Netherlands
 Malaria and vaccination: an update ) Robert Sauerwein Dept. of Medical Microbiology Radboud University Medical Center, Nijmegen, the Netherlands Malaria Control Drugs for treatment and/or prophylaxis Artemisinin-based
Malaria and vaccination: an update ) Robert Sauerwein Dept. of Medical Microbiology Radboud University Medical Center, Nijmegen, the Netherlands Malaria Control Drugs for treatment and/or prophylaxis Artemisinin-based
Chapter 22: The Lymphatic System and Immunity
 Bio40C schedule Lecture Immune system Lab Quiz 2 this week; bring a scantron! Study guide on my website (see lab assignments) Extra credit Critical thinking questions at end of chapters 5 pts/chapter Due
Bio40C schedule Lecture Immune system Lab Quiz 2 this week; bring a scantron! Study guide on my website (see lab assignments) Extra credit Critical thinking questions at end of chapters 5 pts/chapter Due
The Malarias: Plasmodium falciparum Plasmodium vivax Plasmodium malariae Plasmodium ovale. Distribution of Plasmodium falciparum
 The Malarias: Plasmodium falciparum Plasmodium vivax Plasmodium malariae Plasmodium ovale Distribution of Plasmodium falciparum 1 Distribution Of Plasmodium vivax 2 Global Risk By Country-Proportionality
The Malarias: Plasmodium falciparum Plasmodium vivax Plasmodium malariae Plasmodium ovale Distribution of Plasmodium falciparum 1 Distribution Of Plasmodium vivax 2 Global Risk By Country-Proportionality
The Malarias: Plasmodium falciparum Plasmodium vivax Plasmodium malariae Plasmodium ovale. Watersheds of the African Continent
 The Malarias: Plasmodium falciparum Plasmodium vivax Plasmodium malariae Plasmodium ovale Global Risk By Country-Proportionality Plot P. falciparum P. vivax Distribution of Plasmodium falciparum 3 million
The Malarias: Plasmodium falciparum Plasmodium vivax Plasmodium malariae Plasmodium ovale Global Risk By Country-Proportionality Plot P. falciparum P. vivax Distribution of Plasmodium falciparum 3 million
Maria M. Mota Ana Rodriguez Editors. Malaria. Immune Response to Infection and Vaccination
 Maria M. Mota Ana Rodriguez Editors Malaria Immune Response to Infection and Vaccination Malaria Maria M. Mota Ana Rodriguez Editors Malaria Immune Response to Infection and Vaccination Editors Maria M.
Maria M. Mota Ana Rodriguez Editors Malaria Immune Response to Infection and Vaccination Malaria Maria M. Mota Ana Rodriguez Editors Malaria Immune Response to Infection and Vaccination Editors Maria M.
GOVX-B11: A Clade B HIV Vaccine for the Developed World
 GeoVax Labs, Inc. 19 Lake Park Drive Suite 3 Atlanta, GA 3 (678) 384-72 GOVX-B11: A Clade B HIV Vaccine for the Developed World Executive summary: GOVX-B11 is a Clade B HIV vaccine targeted for use in
GeoVax Labs, Inc. 19 Lake Park Drive Suite 3 Atlanta, GA 3 (678) 384-72 GOVX-B11: A Clade B HIV Vaccine for the Developed World Executive summary: GOVX-B11 is a Clade B HIV vaccine targeted for use in
Scott Abrams, Ph.D. Professor of Oncology, x4375 Kuby Immunology SEVENTH EDITION
 Scott Abrams, Ph.D. Professor of Oncology, x4375 scott.abrams@roswellpark.org Kuby Immunology SEVENTH EDITION CHAPTER 11 T-Cell Activation, Differentiation, and Memory Copyright 2013 by W. H. Freeman and
Scott Abrams, Ph.D. Professor of Oncology, x4375 scott.abrams@roswellpark.org Kuby Immunology SEVENTH EDITION CHAPTER 11 T-Cell Activation, Differentiation, and Memory Copyright 2013 by W. H. Freeman and
A COMPARATIVE STUDY OF QUININE V/S ARTESUNATE IN SEVERE MALARIA PATIENTS IN NORTHWESTERN RAJASTHAN, INDIA
 A COMPARATIVE STUDY OF QUININE V/S ARTESUNATE IN SEVERE MALARIA PATIENTS IN NORTHWESTERN RAJASTHAN, INDIA K. C. Nayak, Rakesh Meena, *Surendra Kumar, B. K. Gupta, V. B. Singh and Varun Kulkarni Department
A COMPARATIVE STUDY OF QUININE V/S ARTESUNATE IN SEVERE MALARIA PATIENTS IN NORTHWESTERN RAJASTHAN, INDIA K. C. Nayak, Rakesh Meena, *Surendra Kumar, B. K. Gupta, V. B. Singh and Varun Kulkarni Department
Malaria DR. AFNAN YOUNIS
 Malaria DR. AFNAN YOUNIS Objectives: Epidemiology of malaria Clinical picture Mode of transmission Risk factors Prevention and control Malaria is a life-threatening disease caused by Plasmodium parasites
Malaria DR. AFNAN YOUNIS Objectives: Epidemiology of malaria Clinical picture Mode of transmission Risk factors Prevention and control Malaria is a life-threatening disease caused by Plasmodium parasites
Fabio T M Costa, PhD. University of Campinas (UNICAMP); Campinas, SP, Brazil. Supported by:
; Campinas, SP, Brazil. Supported by:") Fabio T M Costa, PhD University of Campinas (UNICAMP); Campinas, SP, Brazil Supported by: P. falciparum associated pathologies In falciparum malaria cytoadhesion of mature forms are related to fatalities!
Fabio T M Costa, PhD University of Campinas (UNICAMP); Campinas, SP, Brazil Supported by: P. falciparum associated pathologies In falciparum malaria cytoadhesion of mature forms are related to fatalities!
Protozoan Infections of the Circulatory System *
 OpenStax-CNX module: m64867 1 Protozoan Infections of the Circulatory System * Geo Lin-Cereghino Based on Parasitic Infections of the Circulatory and Lymphatic Systems by OpenStax This work is produced
OpenStax-CNX module: m64867 1 Protozoan Infections of the Circulatory System * Geo Lin-Cereghino Based on Parasitic Infections of the Circulatory and Lymphatic Systems by OpenStax This work is produced
Hepatitis virus immunity. Mar 9, 2005 Rehermann and Nascimbeni review Crispe review
 Hepatitis virus immunity Mar 9, 2005 Rehermann and Nascimbeni review Crispe review HBV & HCV infection outcomes Both viruses cause immune-mediated active and chronic hepatitis HBV Vertical transmission
Hepatitis virus immunity Mar 9, 2005 Rehermann and Nascimbeni review Crispe review HBV & HCV infection outcomes Both viruses cause immune-mediated active and chronic hepatitis HBV Vertical transmission
Fluid movement in capillaries. Not all fluid is reclaimed at the venous end of the capillaries; that is the job of the lymphatic system
 Capillary exchange Fluid movement in capillaries Not all fluid is reclaimed at the venous end of the capillaries; that is the job of the lymphatic system Lymphatic vessels Lymphatic capillaries permeate
Capillary exchange Fluid movement in capillaries Not all fluid is reclaimed at the venous end of the capillaries; that is the job of the lymphatic system Lymphatic vessels Lymphatic capillaries permeate
Ad35.CS.01 RTS,S/AS01 prime boost second generation malaria vaccine candidate
 Ad35.CS.01 /AS01 prime boost second generation malaria vaccine candidate Johan Vekemans MD, PhD Malaria vaccines Global Vaccine Development GSK Biologicals NIH 2012 1 The need for a malaria vaccine: targets
Ad35.CS.01 /AS01 prime boost second generation malaria vaccine candidate Johan Vekemans MD, PhD Malaria vaccines Global Vaccine Development GSK Biologicals NIH 2012 1 The need for a malaria vaccine: targets
Research Activities AMA-1 CSP
 Research Activities Project 1 - CD1d-binding NKT Stimulating Glycolipids as an Adjuvant for Malaria/HIV Vaccines We aimed to identify a unique glycolipid which can exert potent stimulatory activity on
Research Activities Project 1 - CD1d-binding NKT Stimulating Glycolipids as an Adjuvant for Malaria/HIV Vaccines We aimed to identify a unique glycolipid which can exert potent stimulatory activity on
Question 1. Kupffer cells, microglial cells and osteoclasts are all examples of what type of immune system cell?
 Abbas Chapter 2: Sarah Spriet February 8, 2015 Question 1. Kupffer cells, microglial cells and osteoclasts are all examples of what type of immune system cell? a. Dendritic cells b. Macrophages c. Monocytes
Abbas Chapter 2: Sarah Spriet February 8, 2015 Question 1. Kupffer cells, microglial cells and osteoclasts are all examples of what type of immune system cell? a. Dendritic cells b. Macrophages c. Monocytes
Malaria. Edwin J. Asturias, MD
 Malaria Edwin J. Asturias, MD Associate Professor of Pediatrics and Epidemiology Director for Latin America Center for Global Health, Colorado School of Public Health Global Health and Disasters Course
Malaria Edwin J. Asturias, MD Associate Professor of Pediatrics and Epidemiology Director for Latin America Center for Global Health, Colorado School of Public Health Global Health and Disasters Course
Scott Abrams, Ph.D. Professor of Oncology, x4375 Kuby Immunology SEVENTH EDITION
 Scott Abrams, Ph.D. Professor of Oncology, x4375 scott.abrams@roswellpark.org Kuby Immunology SEVENTH EDITION CHAPTER 13 Effector Responses: Cell- and Antibody-Mediated Immunity Copyright 2013 by W. H.
Scott Abrams, Ph.D. Professor of Oncology, x4375 scott.abrams@roswellpark.org Kuby Immunology SEVENTH EDITION CHAPTER 13 Effector Responses: Cell- and Antibody-Mediated Immunity Copyright 2013 by W. H.
Malaria. An Overview of Life-cycle, Morphology and Clinical Picture
 Malaria An Overview of Life-cycle, Morphology and Clinical Picture Malaria Malaria is the most important of all tropical parasitic disease,causes death and debility and is endemic throughout the tropics
Malaria An Overview of Life-cycle, Morphology and Clinical Picture Malaria Malaria is the most important of all tropical parasitic disease,causes death and debility and is endemic throughout the tropics
NKTR-255: Accessing The Immunotherapeutic Potential Of IL-15 for NK Cell Therapies
 NKTR-255: Accessing The Immunotherapeutic Potential Of IL-15 for NK Cell Therapies Saul Kivimäe Senior Scientist, Research Biology Nektar Therapeutics NK Cell-Based Cancer Immunotherapy, September 26-27,
NKTR-255: Accessing The Immunotherapeutic Potential Of IL-15 for NK Cell Therapies Saul Kivimäe Senior Scientist, Research Biology Nektar Therapeutics NK Cell-Based Cancer Immunotherapy, September 26-27,
Cell Mediated Immunity CELL MEDIATED IMMUNITY. Basic Elements of Cell Mediated Immunity (CMI) Antibody-dependent cell-mediated cytotoxicity (ADCC)
 Antibody-dependent cell-mediated cytotoxicity (ADCC)") Chapter 16 CELL MEDIATED IMMUNITY Cell Mediated Immunity Also known as Cellular Immunity or CMI The effector phase T cells Specificity for immune recognition reactions TH provide cytokines CTLs do the
Chapter 16 CELL MEDIATED IMMUNITY Cell Mediated Immunity Also known as Cellular Immunity or CMI The effector phase T cells Specificity for immune recognition reactions TH provide cytokines CTLs do the
Malaria Life Cycle Life Cycle
 Malaria Malaria Life Cycle Life Cycle Oocyst Zygote Sporogony Sporozoites Mosquito Salivary Gland Gametocytes Exoerythrocytic (hepatic) cycle Hypnozoites (for P. vivax and P. ovale) Erythrocytic Cycle
Malaria Malaria Life Cycle Life Cycle Oocyst Zygote Sporogony Sporozoites Mosquito Salivary Gland Gametocytes Exoerythrocytic (hepatic) cycle Hypnozoites (for P. vivax and P. ovale) Erythrocytic Cycle
Kato, N. et al, Nature, 2016, Accelerated Article Preview. Celeste Alverez Current Literature 9/10/ /10/2016 Celeste Wipf Group 1
 Kato, N. et al, Nature, 2016, Accelerated Article Preview Celeste Alverez Current Literature 9/10/2016 9/10/2016 Celeste Alverez @ Wipf Group 1 Malaria Caused by Plasmodium parasites Carried by 30-40 species
Kato, N. et al, Nature, 2016, Accelerated Article Preview Celeste Alverez Current Literature 9/10/2016 9/10/2016 Celeste Alverez @ Wipf Group 1 Malaria Caused by Plasmodium parasites Carried by 30-40 species
3. Lymphocyte proliferation (fig. 15.4): Clones of responder cells and memory cells are derived from B cells and T cells.
: Clones of responder cells and memory cells are derived from B cells and T cells.") Chapter 15 Adaptive, Specific Immunity and Immunization* *Lecture notes are to be used as a study guide only and do not represent the comprehensive information you will need to know for the exams. Specific
Chapter 15 Adaptive, Specific Immunity and Immunization* *Lecture notes are to be used as a study guide only and do not represent the comprehensive information you will need to know for the exams. Specific
Running head: VECTOR-BORNE DISEASE 1
 Running head: VECTOR-BORNE DISEASE 1 Vector-Borne Disease: Malaria in Sub-Sahara Africa Maritza Morejon GCH 360- Environmental Health George Mason University VECTOR-BORNE DISEASE 2 Introduction Malaria
Running head: VECTOR-BORNE DISEASE 1 Vector-Borne Disease: Malaria in Sub-Sahara Africa Maritza Morejon GCH 360- Environmental Health George Mason University VECTOR-BORNE DISEASE 2 Introduction Malaria
Significance of the MHC
 CHAPTER 7 Major Histocompatibility Complex (MHC) What is is MHC? HLA H-2 Minor histocompatibility antigens Peter Gorer & George Sneell (1940) Significance of the MHC role in immune response role in organ
CHAPTER 7 Major Histocompatibility Complex (MHC) What is is MHC? HLA H-2 Minor histocompatibility antigens Peter Gorer & George Sneell (1940) Significance of the MHC role in immune response role in organ
Counting Malaria Out: Progress of Efforts in the Americas and More Stories of Innovations. Counting Malaria Out in the Americas
 Counting Malaria Out: Progress of Efforts in the Americas and More Stories of Innovations Malaria: A Review Malaria is a disease caused by a parasite of the genus Plasmodium. More than 150 species of Plasmodium
Counting Malaria Out: Progress of Efforts in the Americas and More Stories of Innovations Malaria: A Review Malaria is a disease caused by a parasite of the genus Plasmodium. More than 150 species of Plasmodium
Gene Vaccine Dr. Sina Soleimani
 Gene Vaccine Dr. Sina Soleimani Human Viral Vaccines Quality Control Laboratory (HVVQC) Titles 1. A short Introduction of Vaccine History 2. First Lineage of Vaccines 3. Second Lineage of Vaccines 3. New
Gene Vaccine Dr. Sina Soleimani Human Viral Vaccines Quality Control Laboratory (HVVQC) Titles 1. A short Introduction of Vaccine History 2. First Lineage of Vaccines 3. Second Lineage of Vaccines 3. New
Immune System AP SBI4UP
 Immune System AP SBI4UP TYPES OF IMMUNITY INNATE IMMUNITY ACQUIRED IMMUNITY EXTERNAL DEFENCES INTERNAL DEFENCES HUMORAL RESPONSE Skin Phagocytic Cells CELL- MEDIATED RESPONSE Mucus layer Antimicrobial
Immune System AP SBI4UP TYPES OF IMMUNITY INNATE IMMUNITY ACQUIRED IMMUNITY EXTERNAL DEFENCES INTERNAL DEFENCES HUMORAL RESPONSE Skin Phagocytic Cells CELL- MEDIATED RESPONSE Mucus layer Antimicrobial
Medical Virology Immunology. Dr. Sameer Naji, MB, BCh, PhD (UK) Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University
 Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University") Medical Virology Immunology Dr. Sameer Naji, MB, BCh, PhD (UK) Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University Human blood cells Phases of immune responses Microbe Naïve
Medical Virology Immunology Dr. Sameer Naji, MB, BCh, PhD (UK) Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University Human blood cells Phases of immune responses Microbe Naïve
40% (90% (500 BC)
") MALARIA causative agent = Plasmodium species 40% of world s population lives in endemic areas 3-500 million clinical cases per year 1.5-2.7 million deaths (90% Africa) known since antiquity early medical
MALARIA causative agent = Plasmodium species 40% of world s population lives in endemic areas 3-500 million clinical cases per year 1.5-2.7 million deaths (90% Africa) known since antiquity early medical
NKTR-255: Accessing IL-15 Therapeutic Potential through Robust and Sustained Engagement of Innate and Adaptive Immunity
 NKTR-255: Accessing IL-15 Therapeutic Potential through Robust and Sustained Engagement of Innate and Adaptive Immunity Peiwen Kuo Scientist, Research Biology Nektar Therapeutics August 31 st, 2018 Emerging
NKTR-255: Accessing IL-15 Therapeutic Potential through Robust and Sustained Engagement of Innate and Adaptive Immunity Peiwen Kuo Scientist, Research Biology Nektar Therapeutics August 31 st, 2018 Emerging
D-LAB HEALTH SP 725. Jose Gomez-Marquez
 SP 725 Jose Gomez-Marquez 1 Vaccine Preventable Diseases Causes of 2.5 million child deaths out of 10.5 million child deaths globally, 2002 Source: WHO Wkly Epidemiol Rec. (2006) 81:189-196. 2 Rationale
SP 725 Jose Gomez-Marquez 1 Vaccine Preventable Diseases Causes of 2.5 million child deaths out of 10.5 million child deaths globally, 2002 Source: WHO Wkly Epidemiol Rec. (2006) 81:189-196. 2 Rationale
Chapter 24 The Immune System
 Chapter 24 The Immune System The Immune System Layered defense system The skin and chemical barriers The innate and adaptive immune systems Immunity The body s ability to recognize and destroy specific
Chapter 24 The Immune System The Immune System Layered defense system The skin and chemical barriers The innate and adaptive immune systems Immunity The body s ability to recognize and destroy specific
The most effective vaccination protocol against malaria, in
 Complete, long-lasting protection against malaria of mice primed and boosted with two distinct viral vectors expressing the same plasmodial antigen Oscar Bruña-Romero, Gloria González-Aseguinolaza, Julius
Complete, long-lasting protection against malaria of mice primed and boosted with two distinct viral vectors expressing the same plasmodial antigen Oscar Bruña-Romero, Gloria González-Aseguinolaza, Julius
Prof. Ibtesam Kamel Afifi Professor of Medical Microbiology & Immunology
 By Prof. Ibtesam Kamel Afifi Professor of Medical Microbiology & Immunology Lecture objectives: At the end of the lecture you should be able to: Enumerate features that characterize acquired immune response
By Prof. Ibtesam Kamel Afifi Professor of Medical Microbiology & Immunology Lecture objectives: At the end of the lecture you should be able to: Enumerate features that characterize acquired immune response
Examples of questions for Cellular Immunology/Cellular Biology and Immunology
 Examples of questions for Cellular Immunology/Cellular Biology and Immunology Each student gets a set of 6 questions, so that each set contains different types of questions and that the set of questions
Examples of questions for Cellular Immunology/Cellular Biology and Immunology Each student gets a set of 6 questions, so that each set contains different types of questions and that the set of questions
Topics in Parasitology BLY Vertebrate Immune System
 Topics in Parasitology BLY 533-2008 Vertebrate Immune System V. Vertebrate Immune System A. Non-specific defenses against pathogens 1. Skin - physical barrier a. Tough armor protein KERATIN b. Surface
Topics in Parasitology BLY 533-2008 Vertebrate Immune System V. Vertebrate Immune System A. Non-specific defenses against pathogens 1. Skin - physical barrier a. Tough armor protein KERATIN b. Surface
Chapter 15 Adaptive, Specific Immunity and Immunization
 Chapter 15 Adaptive, Specific Immunity and Immunization Adaptive Immunity: The third line of defense Third line of defense acquired and specific. Dual System of B and T lymphocytes- Immunocompetence Antigen
Chapter 15 Adaptive, Specific Immunity and Immunization Adaptive Immunity: The third line of defense Third line of defense acquired and specific. Dual System of B and T lymphocytes- Immunocompetence Antigen
Principles of Adaptive Immunity
 Principles of Adaptive Immunity Chapter 3 Parham Hans de Haard 17 th of May 2010 Agenda Recognition molecules of adaptive immune system Features adaptive immune system Immunoglobulins and T-cell receptors
Principles of Adaptive Immunity Chapter 3 Parham Hans de Haard 17 th of May 2010 Agenda Recognition molecules of adaptive immune system Features adaptive immune system Immunoglobulins and T-cell receptors
Adaptive immune responses: T cell-mediated immunity
 MICR2209 Adaptive immune responses: T cell-mediated immunity Dr Allison Imrie allison.imrie@uwa.edu.au 1 Synopsis: In this lecture we will discuss the T-cell mediated immune response, how it is activated,
MICR2209 Adaptive immune responses: T cell-mediated immunity Dr Allison Imrie allison.imrie@uwa.edu.au 1 Synopsis: In this lecture we will discuss the T-cell mediated immune response, how it is activated,
IOM Immunization Safety Review 11/12/2001. Immunological Competition and the Infant Immune Response to Vaccines
 IOM Immunization Safety Review 11/12/2001 Immunological Competition and the Infant Immune Response to Vaccines Richard Insel University of Rochester Goals Neonatal and Infant Immune System Broad Effects
IOM Immunization Safety Review 11/12/2001 Immunological Competition and the Infant Immune Response to Vaccines Richard Insel University of Rochester Goals Neonatal and Infant Immune System Broad Effects
MALARIA P. FALCIPARUM / P. VIVAX
 MALARIA P. FALCIPARUM / P. VIVAX 1. EXPLANATION OF THE TEST: Malaria is a serious, sometimes fatal, parasitic disease characterized by fever, chills, and anemia and is caused by a parasite that is transmitted
MALARIA P. FALCIPARUM / P. VIVAX 1. EXPLANATION OF THE TEST: Malaria is a serious, sometimes fatal, parasitic disease characterized by fever, chills, and anemia and is caused by a parasite that is transmitted
Adaptive Immunity: Humoral Immune Responses
 MICR2209 Adaptive Immunity: Humoral Immune Responses Dr Allison Imrie 1 Synopsis: In this lecture we will review the different mechanisms which constitute the humoral immune response, and examine the antibody
MICR2209 Adaptive Immunity: Humoral Immune Responses Dr Allison Imrie 1 Synopsis: In this lecture we will review the different mechanisms which constitute the humoral immune response, and examine the antibody
Structure and Function of Antigen Recognition Molecules
 MICR2209 Structure and Function of Antigen Recognition Molecules Dr Allison Imrie allison.imrie@uwa.edu.au 1 Synopsis: In this lecture we will examine the major receptors used by cells of the innate and
MICR2209 Structure and Function of Antigen Recognition Molecules Dr Allison Imrie allison.imrie@uwa.edu.au 1 Synopsis: In this lecture we will examine the major receptors used by cells of the innate and
COURSE: Medical Microbiology, MBIM 650/720 - Fall TOPIC: Antigen Processing, MHC Restriction, & Role of Thymus Lecture 12
 COURSE: Medical Microbiology, MBIM 650/720 - Fall 2008 TOPIC: Antigen Processing, MHC Restriction, & Role of Thymus Lecture 12 FACULTY: Dr. Mayer Office: Bldg. #1, Rm B32 Phone: 733-3281 Email: MAYER@MED.SC.EDU
COURSE: Medical Microbiology, MBIM 650/720 - Fall 2008 TOPIC: Antigen Processing, MHC Restriction, & Role of Thymus Lecture 12 FACULTY: Dr. Mayer Office: Bldg. #1, Rm B32 Phone: 733-3281 Email: MAYER@MED.SC.EDU
Cover Page. The handle holds various files of this Leiden University dissertation
 Cover Page The handle http://hdl.handle.net/1887/35908 holds various files of this Leiden University dissertation Author: Soema, Peter Title: Formulation of influenza T cell peptides : in search of a universal
Cover Page The handle http://hdl.handle.net/1887/35908 holds various files of this Leiden University dissertation Author: Soema, Peter Title: Formulation of influenza T cell peptides : in search of a universal
Theiler s Murine Encephalomyelitis Virus-Induced CNS Autoimmunity
 Theiler s Murine Encephalomyelitis Virus-Induced CNS Autoimmunity Virus-induced molecular mimicry is part of a mouse model of multiple sclerosis that is providing insights about the disease in humans Julie
Theiler s Murine Encephalomyelitis Virus-Induced CNS Autoimmunity Virus-induced molecular mimicry is part of a mouse model of multiple sclerosis that is providing insights about the disease in humans Julie
PBS Class #2 Introduction to the Immune System part II Suggested reading: Abbas, pgs , 27-30
 PBS 803 - Class #2 Introduction to the Immune System part II Suggested reading: Abbas, pgs. 15-25, 27-30 Learning Objectives Compare and contrast the maturation of B and T lymphocytes Compare and contrast
PBS 803 - Class #2 Introduction to the Immune System part II Suggested reading: Abbas, pgs. 15-25, 27-30 Learning Objectives Compare and contrast the maturation of B and T lymphocytes Compare and contrast
Identification of vaccine candidate antigens for a Babesia bovis transmission blocking vaccine
 Identification of vaccine candidate antigens for a Babesia bovis transmission blocking vaccine Carlos E. Suarez Animal Disease Research Unit ARS-USDA Department of Veterinary Microbiology and Pathology
Identification of vaccine candidate antigens for a Babesia bovis transmission blocking vaccine Carlos E. Suarez Animal Disease Research Unit ARS-USDA Department of Veterinary Microbiology and Pathology
TITLE: MODULATION OF T CELL TOLERANCE IN A MURINE MODEL FOR IMMUNOTHERAPY OF PROSTATIC ADENOCARCINOMA
 AD Award Number: DAMD17-01-1-0085 TITLE: MODULATION OF T CELL TOLERANCE IN A MURINE MODEL FOR IMMUNOTHERAPY OF PROSTATIC ADENOCARCINOMA PRINCIPAL INVESTIGATOR: ARTHUR A HURWITZ, Ph.d. CONTRACTING ORGANIZATION:
AD Award Number: DAMD17-01-1-0085 TITLE: MODULATION OF T CELL TOLERANCE IN A MURINE MODEL FOR IMMUNOTHERAPY OF PROSTATIC ADENOCARCINOMA PRINCIPAL INVESTIGATOR: ARTHUR A HURWITZ, Ph.d. CONTRACTING ORGANIZATION:
Lecture 11. Immunology and disease: parasite antigenic diversity
 Lecture 11 Immunology and disease: parasite antigenic diversity RNAi interference video and tutorial (you are responsible for this material, so check it out.) http://www.pbs.org/wgbh/nova/sciencenow/3210/02.html
Lecture 11 Immunology and disease: parasite antigenic diversity RNAi interference video and tutorial (you are responsible for this material, so check it out.) http://www.pbs.org/wgbh/nova/sciencenow/3210/02.html
Chapter 1. Full file at
 Chapter 1 1. Which is the best definition of immunity? Answer: B A. The state of having been exposed to a pathogen repeatedly B. The state of being resistant to reinfection with a pathogen C. When an individual
Chapter 1 1. Which is the best definition of immunity? Answer: B A. The state of having been exposed to a pathogen repeatedly B. The state of being resistant to reinfection with a pathogen C. When an individual
A Hopeful Beginning for Malaria Vaccines
 Transcript Details This is a transcript of an educational program accessible on the ReachMD network. Details about the program and additional media formats for the program are accessible by visiting: https://reachmd.com/programs/focus-on-global-medicine/a-hopeful-beginning-for-malariavaccines/4043/
Transcript Details This is a transcript of an educational program accessible on the ReachMD network. Details about the program and additional media formats for the program are accessible by visiting: https://reachmd.com/programs/focus-on-global-medicine/a-hopeful-beginning-for-malariavaccines/4043/
Biotechnology-Based Vaccines. Dr. Aws Alshamsan Department of Pharmaceutics Office: AA87 Tel:
 Biotechnology-Based Vaccines Dr. Aws Alshamsan Department of Pharmaceutics Office: AA87 Tel: 4677363 aalshamsan@ksu.edu.sa Objectives of this lecture By the end of this lecture you will be able to: 1.
Biotechnology-Based Vaccines Dr. Aws Alshamsan Department of Pharmaceutics Office: AA87 Tel: 4677363 aalshamsan@ksu.edu.sa Objectives of this lecture By the end of this lecture you will be able to: 1.
ACTIVATION AND EFFECTOR FUNCTIONS OF CELL-MEDIATED IMMUNITY AND NK CELLS. Choompone Sakonwasun, MD (Hons), FRCPT
, FRCPT") ACTIVATION AND EFFECTOR FUNCTIONS OF CELL-MEDIATED IMMUNITY AND NK CELLS Choompone Sakonwasun, MD (Hons), FRCPT Types of Adaptive Immunity Types of T Cell-mediated Immune Reactions CTLs = cytotoxic T lymphocytes
ACTIVATION AND EFFECTOR FUNCTIONS OF CELL-MEDIATED IMMUNITY AND NK CELLS Choompone Sakonwasun, MD (Hons), FRCPT Types of Adaptive Immunity Types of T Cell-mediated Immune Reactions CTLs = cytotoxic T lymphocytes
Adaptive Immunity: Specific Defenses of the Host
 17 Adaptive Immunity: Specific Defenses of the Host SLOs Differentiate between innate and adaptive immunity, and humoral and cellular immunity. Define antigen, epitope, and hapten. Explain the function
17 Adaptive Immunity: Specific Defenses of the Host SLOs Differentiate between innate and adaptive immunity, and humoral and cellular immunity. Define antigen, epitope, and hapten. Explain the function
Nature Immunology: doi: /ni Supplementary Figure 1. Gene expression profile of CD4 + T cells and CTL responses in Bcl6-deficient mice.
 Supplementary Figure 1 Gene expression profile of CD4 + T cells and CTL responses in Bcl6-deficient mice. (a) Gene expression profile in the resting CD4 + T cells were analyzed by an Affymetrix microarray
Supplementary Figure 1 Gene expression profile of CD4 + T cells and CTL responses in Bcl6-deficient mice. (a) Gene expression profile in the resting CD4 + T cells were analyzed by an Affymetrix microarray
Of mice and men: practical approaches to malaria vaccine discovery and development
 Of mice and men: practical approaches to malaria vaccine discovery and development --> The consequences of the screen employed to identify a parasite molecule as a «major vaccine candidate «Or the problem
Of mice and men: practical approaches to malaria vaccine discovery and development --> The consequences of the screen employed to identify a parasite molecule as a «major vaccine candidate «Or the problem
Memory CD4 T Cells Enhance Primary CD8 T-Cell Responses
 INFECTION AND IMMUNITY, July 2007, p. 3556 3560 Vol. 75, No. 7 0019-9567/07/$08.00 0 doi:10.1128/iai.00086-07 Copyright 2007, American Society for Microbiology. All Rights Reserved. Memory CD4 T Cells
INFECTION AND IMMUNITY, July 2007, p. 3556 3560 Vol. 75, No. 7 0019-9567/07/$08.00 0 doi:10.1128/iai.00086-07 Copyright 2007, American Society for Microbiology. All Rights Reserved. Memory CD4 T Cells
NTD Vaccine Design Toolkit and Training Workshop Providence, RI January 05, 2011 Cytokines Leslie P. Cousens, PhD EpiVax, Inc.
 NTD Vaccine Design Toolkit and Training Workshop Providence, RI January 05, 2011 Cytokines Leslie P. Cousens, PhD EpiVax, Inc. Cytokines Properties of Cytokines Cytokines are proteins with specific roles
NTD Vaccine Design Toolkit and Training Workshop Providence, RI January 05, 2011 Cytokines Leslie P. Cousens, PhD EpiVax, Inc. Cytokines Properties of Cytokines Cytokines are proteins with specific roles
Lecture 2: Virology. I. Background
 Lecture 2: Virology I. Background A. Properties 1. Simple biological systems a. Aggregates of nucleic acids and protein 2. Non-living a. Cannot reproduce or carry out metabolic activities outside of a
Lecture 2: Virology I. Background A. Properties 1. Simple biological systems a. Aggregates of nucleic acids and protein 2. Non-living a. Cannot reproduce or carry out metabolic activities outside of a
answer Marks Guidance 1 (a) 2 max Mark the first answer on each prompt line. ACCEPT ora throughout nucleus / nuclei ; 1 ACCEPT DNA not free
 2 max Mark the first answer on each prompt line. ACCEPT ora throughout nucleus / nuclei ; 1 ACCEPT DNA not free") Question answer Marks Guidance 1 (a) max Mark the first answer on each prompt line. ACCEPT ora throughout 1 6 7 nucleus / nuclei ; other named organelle / membrane bound organelles ; linear chromosomes
Question answer Marks Guidance 1 (a) max Mark the first answer on each prompt line. ACCEPT ora throughout 1 6 7 nucleus / nuclei ; other named organelle / membrane bound organelles ; linear chromosomes
van der Ventel, Michelle (2012) The role of IL-4Ra signalling in gene deficient mice during asexual-stage Plasmodium chabaudi AS infection.
 The role of IL-4Ra signalling in gene deficient mice during asexual-stage Plasmodium chabaudi AS infection.") van der Ventel, Michelle (2012) The role of IL-4Ra signalling in gene deficient mice during asexual-stage Plasmodium chabaudi AS infection. PhD thesis http://theses.gla.ac.uk/3438/ Copyright and moral
van der Ventel, Michelle (2012) The role of IL-4Ra signalling in gene deficient mice during asexual-stage Plasmodium chabaudi AS infection. PhD thesis http://theses.gla.ac.uk/3438/ Copyright and moral
Are booster immunisations needed for lifelong hepatitis B immunity?
 Are booster immunisations needed for lifelong hepatitis B immunity? European Consensus Group on Hepatitis B immunity, following meeting in Florence in October 1998 To date there are no data to support
Are booster immunisations needed for lifelong hepatitis B immunity? European Consensus Group on Hepatitis B immunity, following meeting in Florence in October 1998 To date there are no data to support
Chapter 1. Chapter 1 Concepts. MCMP422 Immunology and Biologics Immunology is important personally and professionally!
 MCMP422 Immunology and Biologics Immunology is important personally and professionally! Learn the language - use the glossary and index RNR - Reading, Note taking, Reviewing All materials in Chapters 1-3
MCMP422 Immunology and Biologics Immunology is important personally and professionally! Learn the language - use the glossary and index RNR - Reading, Note taking, Reviewing All materials in Chapters 1-3
ESCMID Online Lecture Library. by author
 4-8 June 2012 T. van Gool, MD, PhD A. Bart, PhD Academic Medical Center Amsterdam F. Derouin, MD, PhD Hôpital Saint Louis Paris T. Kortbeek, MD RIVM, Bilthoven How good is training in Clinical Parasitology?
4-8 June 2012 T. van Gool, MD, PhD A. Bart, PhD Academic Medical Center Amsterdam F. Derouin, MD, PhD Hôpital Saint Louis Paris T. Kortbeek, MD RIVM, Bilthoven How good is training in Clinical Parasitology?
Memory NK cells during mousepox infection. Min Fang, Ph.D, Professor Institute of Microbiology, Chinese Academy of Science
 Memory NK cells during mousepox infection Min Fang, Ph.D, Professor Institute of Microbiology, Chinese Academy of Science Infectious Diseases are a Major Cause of Death Worldwide May 14 th 1796 Prevalence
Memory NK cells during mousepox infection Min Fang, Ph.D, Professor Institute of Microbiology, Chinese Academy of Science Infectious Diseases are a Major Cause of Death Worldwide May 14 th 1796 Prevalence
Determinants of Immunogenicity and Tolerance. Abul K. Abbas, MD Department of Pathology University of California San Francisco
 Determinants of Immunogenicity and Tolerance Abul K. Abbas, MD Department of Pathology University of California San Francisco EIP Symposium Feb 2016 Why do some people respond to therapeutic proteins?
Determinants of Immunogenicity and Tolerance Abul K. Abbas, MD Department of Pathology University of California San Francisco EIP Symposium Feb 2016 Why do some people respond to therapeutic proteins?
11/25/2017. THE IMMUNE SYSTEM Chapter 43 IMMUNITY INNATE IMMUNITY EXAMPLE IN INSECTS BARRIER DEFENSES INNATE IMMUNITY OF VERTEBRATES
 THE IMMUNE SYSTEM Chapter 43 IMMUNITY INNATE IMMUNITY EXAMPLE IN INSECTS Exoskeleton made of chitin forms the first barrier to pathogens Digestive system is protected by a chitin-based barrier and lysozyme,
THE IMMUNE SYSTEM Chapter 43 IMMUNITY INNATE IMMUNITY EXAMPLE IN INSECTS Exoskeleton made of chitin forms the first barrier to pathogens Digestive system is protected by a chitin-based barrier and lysozyme,
Fayth K. Yoshimura, Ph.D. September 7, of 7 HIV - BASIC PROPERTIES
 1 of 7 I. Viral Origin. A. Retrovirus - animal lentiviruses. HIV - BASIC PROPERTIES 1. HIV is a member of the Retrovirus family and more specifically it is a member of the Lentivirus genus of this family.
1 of 7 I. Viral Origin. A. Retrovirus - animal lentiviruses. HIV - BASIC PROPERTIES 1. HIV is a member of the Retrovirus family and more specifically it is a member of the Lentivirus genus of this family.
Chapter 13 Lymphatic and Immune Systems
 The Chapter 13 Lymphatic and Immune Systems 1 The Lymphatic Vessels Lymphoid Organs Three functions contribute to homeostasis 1. Return excess tissue fluid to the bloodstream 2. Help defend the body against
The Chapter 13 Lymphatic and Immune Systems 1 The Lymphatic Vessels Lymphoid Organs Three functions contribute to homeostasis 1. Return excess tissue fluid to the bloodstream 2. Help defend the body against
Acquired Immunity 2. - Vaccines & Immunological Memory - Wataru Ise. WPI Immunology Frontier Research Center (IFReC) Osaka University.
 Osaka University.") Acquired Immunity 2 - Vaccines & Immunological Memory - Wataru Ise WPI Immunology Frontier Research Center (IFReC) Osaka University Outline 1. What is vaccine (vaccination)? 2. What is immunological memory?
Acquired Immunity 2 - Vaccines & Immunological Memory - Wataru Ise WPI Immunology Frontier Research Center (IFReC) Osaka University Outline 1. What is vaccine (vaccination)? 2. What is immunological memory?