The structural organization of substrate loading in iterative polyketide synthases
|
|
|
- Kathleen Knight
- 6 years ago
- Views:
Transcription
1 SUPPLEMENTARY INFORMATION Articles In the format provided by the authors and unedited. The structural organization of substrate loading in iterative polyketide synthases Dominik A. Herbst 1, Callie R. Huitt-Roehl 2, Roman P. Jakob 1, Jacob M. Kravetz 2, Philip A. Storm 2, Jamie R. Alley 2, Craig A. Townsend 2 and Timm Maier 1 * 1 Department of Biozentrum, University of Basel, Basel, Switzerland. 2 Department of Chemistry, Johns Hopkins University, Baltimore, MD, USA. * timm.maier@unibas.ch Nature Chemical Biology Nature America Inc., part of Springer Nature. All rights reserved.
2 Supplementary Table 1 X-ray and cryo-em data collection and refinement statistics. a CTB1 SAT-KS-MAT Data collection Space group C2221 Cell dimensions a, b, c (Å) , 230.2, ( ) 90.0, 90.0, 90.0 Resolution (Å) Rmerge (%)* 21.2 (193.8) I/ I* (1.03) CC1/2* 99.5 (43.3) Completeness (%)* 99.2 (91.9) Redundancy* 8.5 (5.6) Refinement Resolution (Å) Rwork/ Rfree 0.21 / 0.24 No. atoms 39,201 Protein 38,682 Ligand/ion 167 Water 352 B-factors Protein (Å 2 ) Ligand/ion (Å 2 ) Water (Å 2 ) R.m.s deviations Bond lengths (Å) Bond angles (º) b CTB1 SAT -KS-MAT =ACP2 Data collection and processing Nominal magnification 105,000x Voltage (kev) 300 Electron exposure (e- Å -2 ) initial / final 90 / 41 Defocus range(μm) Pixel size (Å) Symmetry imposed C1 Movies 1,728 Frames per movie 60 Initial particles images 122,089 Final particle images 25,107 Refinements Initial model used Model resolution (Å) FSC threshold ab initio Model resolution range(å) 4-10 Map sharpening B-factor (Å 2 ) -350 Model composition Non-hydrogen atoms 19,628 Protein residues 2,591 Ligands 0 R.m.s deviations Bond lengths (Å) Bond angles (º) Validation MolProbity score 1.57 All-atom clashscore 6.09 Rotamer outliers (%) 0.00 Ramachandran plot Favored (%) Allowed (%) 3.41 Disallowed (%) 0.12 (a) Crystallographic data collection and refinement statistics. The resolution cutoff was determined by CC1/2 criterion (Karplus, P. A. & Diederichs, K. Science 336, (2012)). *, Highest resolution shell is shown in parentheses. (b) Cryo-electron microscopic data collection and refinement statistics.
3 Supplementary Table 2 Structural comparison and interface analysis of CTB1 SAT-KS- MAT. a Structure 1 Structure 2 Cα r.m.s.d. [Å] Aligned residues CTB1 SAT CTB1 SAT (2 nd chain) Closest* in PDB (3G87) no multienzyme human FAS MAT (3HHD) CazM SAT (4RPM) loading domain PksE AT (5DZ7) procine FAS MAT (2VZ9) PKS13 AT (3TZX) PksC AT (5DZ6) CTB1 MAT DEBS AT5 (2HG4) CurL AT (4MZ0) DEBS AT3, (2QO3) DisD AT (3RGI) VinK (5CZD) AVES1 AT (4RL1) loading domain ZmaA (4QBU) DYNE8 AT (4AMP) MAS-like AT (5BP1) OzmQ partial AT (4OQJ) CTB1 KS CTB1 KS (2 nd chain) PksL KS (5ENY) C0ZGQ5 KS (4Z37) CurL KS (4MZ0) MgsF KS (4TKT) PksJ KS (4NA3) human FAS KS (3HHD) procine FAS KS (2VZ9) DEBS KS3, (2QO3) DEBS KS5 (2HG4) MgsE KS (4QYR) OzmH KS0 (4OPF) MAS-like KS (5BP1) OzmN KS (4WKY) RhiE KS (4KC5) Closest* - FabF KS (4JB6) no multienzyme OzmQ (4OQJ) CTB1 MAT CTB1 MAT (2 nd chain) CurL AT (4MZ0) MAS-like AT (5BP1) PKS13 AT (3TZY) human FAS MAT (3HHD) procine FAS MAT (2VZ9) DYNE8 AT (4AMP) DEBS AT3, (2QO3) DEBS AT5 (2HG4) Closest* in PDB: AVES1 AT (4RL1) loading domain ZmaA (4QBU) DisD AT (3RGI) PksC AT (5DZ6) PksE AT (5DZ7) VinK (5CZD) OzmQ partial AT (4OQJ) CazM SAT (4RPM) loading domain
4 b Interface 1 Interface 2 Min area [Å 2 ] Max area [Å 2 ] Hydrogen bonds Salt bridges SAT KS-LD-MAT-post MAT linker SAT KS SAT MAT SAT SAT SAT dimer KS dimer SAT dimer KS-LD-MAT-post MAT linker dimer SAT SAT-KS linker KS KS KS LD KS post-mat linker LD post-mat linker MAT post-mat linker (a) Cα r.m.s. deviations obtained for structural comparison of crystallized CTB1 domains with their closest structural neighbors in the PDB and of multienzyme PKSs and FASs (to the best of our knowledge). For structures with several protomers only the best matches are reported. *,Qscore based,, trans-at PKS. (b) Interfaces in the crystal structures of CTB1 SAT-KS-MAT. Interface with the second protomer.
5 Supplementary Table 3 Plasmids used in this study Plasmid Protein Vector Tag MW (g/mol) (M -1 cm -1 ) pectb1-nka6 SAT-KS-MAT pet-24a C-His pectb1-skm-c119a-s1010a SAT -KS-MAT pet-24a C-His p28ctb1-acp1 ACP1 pet-28a N-His N/A p28ctb1-acp2 ACP2 pet-28a N-His pectb1-skm-r461a SAT-KS-MAT-R461A pet-24a C-His pectb1-skm-r461e SAT-KS-MAT-R461E pet-24a C-His pectb1-skm-r658a SAT-KS-MAT-R658A pet-24a C-His pectb1-skm-r658e SAT-KS-MAT-R658E pet-24a C-His pectb1-skm-r879a SAT-KS-MAT-R879A pet-24a C-His pectb1-skm-r879e SAT-KS-MAT-R879E pet-24a C-His pectb1-pt PT pet-24a C-His pectb1-te TE pet-28a N-His Supplementary Table 4 Primers used in this study. Primer Sequence 5-3 CTB1-ACP1-5-A ATTACATATGGCAACCCAAGTGACTCCGCAA CTB1-ACP1-3-C ATTAGCGGCCGCGATCTCGTTAGGGGATGGATCAGT CTB1-ACP2-5 ATTACATATGGATCCATCCCCTAACGAGAT CTB1-ACP2-stop-3 TAATGCGGCCGCCTATTCCGTTGACCCAGAGAACC T7 TAATACGACTCACTATAGGG T7-term GCTAGTTATTGCTCAGCGG CTB1-C119A-5 TGCATTACCGGCGTTGCAACCGGCGCA CTB1-C119A-3 CGTCAATGCGCCGGTTGCAACGCCGGT CTB1-S1010A-5 GGTAGTTGGCCACGCGTTGGGCGAGTATGC CTB1-S1010A-3 GCATACTCGCCCAACGCGTGGCCAACTACC CTB1-R461A-5 ACATGAGTCCGGCGGAAGCGCCGC CTB1-R461A-3 GCGGCGCTTCCGCCGGACTCATGT CTB1-R461E-5 ACATGAGTCCGGAAGAAGCGCCGC CTB1-R461E-3 GCGGCGCTTCTTCCGGACTCATGT CTB1-R658A-5 GCCTCTATCACTGCGCCTCATGCCGGAG CTB1-R658A-3 CTCCGGCATGAGGCGCAGTGATAGAGGC CTB1-R658E-5 GCCTCTATCACTGAACCTCATGCCGGAG CTB1-R658E-3 CTCCGGCATGAGGTTCAGTGATAGAGGC CTB1-R879A-5 GAGTGCACCATGCGCACAGAGCCGTAGC CTB1-R879A-3 GCTACGGCTCTGTGCGCATGGTGCACTC CTB1-R879E-5 GAGTGCACCATGAACACAGAGCCGTAGC CTB1-R879E-3 GCTACGGCTCTGTGTTCATGGTGCACTC

, KS (b), and MAT (c)")
6 Supplementary Figure 1 Stereo electron density quality assessment maps. 2Fo-Fc electron density maps at 1.0 σ of the SAT (a), KS (b), and MAT (c) active site regions are shown.
ATs as described by Herbst et al. 2016 (Nature 531, 533-537).")
7 Supplementary Figure 2 Comparison of PKS and FAS condensing region structures. (a) Overview of all superposed condensing region structures on the CTB1 KS domain. Structural differences are visualized by hinge-bending motions of the (M)ATs as described by Herbst et al (Nature 531, ). Colored loop representation is shown for CTB1, all other structures are shown grey transparent. Equivalent cylindrical helix of the (M)AT domains are shown to indicate structural differences relative to CTB1 (light red) for PikAIII (dark red, b), human FAS
8 (hfas, dark blue, PDB:3hhd, c), porcine FAS (pfas, PDB:2vz9, violet, d), CurL (cyan, PDB:4mz0, e), DEBS module 3 (green, PDB:2qo3, f), DEBS module 5 (yellow, PDB:2hg4, g) and a MAS-like PKS (orange, PDB:5bp1, h). (b-h) Inset of (a) in cartoon representation showing the individual superpositions. The model corresponding to PikAIII (b) was fitted according to Dutta et al (Nature 510, ) (EMDB:5649).

,")
9 Supplementary Figure 3 SAT interfaces in the loading/condensing region of CTB1. (a) Overview of interfaces with insets showing the location of the SAT/SAT (b,c), SAT/MAT (d,e),
10 and SAT/KS (f,g) interfaces. All interfaces are shown in a top view. Transparent surfaces are colored according to interfacing residues of the domain in the back. All interfaces are shown from both sides corresponding to a 180 rotation between the left and right panel. Residue numbers correspond to interfacing residues in the front shown in the same color. Cα positions are shown as spheres. Salt bridges (yellow) and hydrogen bonds (cyan) are indicated as dotted lines.
 Uniprot number.")
11 Supplementary Figure 4 Integration of loading domains in PKS. (a), Sequence alignment of 34 SAT-KS linker sequences of NR-PKS. All sequences are labelled as protein name (organism abbreviation) Uniprot number. (b) Superposition of the CazM SAT (black) onto the SAT domain of CTB1 SAT-KS-MAT (light grey) shows a similar location of the SAT C-terminal linker ending (dark red/light red for CazM/CTB1), which points towards the N-terminus of the KS in CTB1. (c) Superposition of the loading domain of the AVES1 avermectin modpks (black) onto the SAT domain of CTB1 SAT-KS-MAT reveals differences in C-terminal linker organization. (d) Superposition of the AVES1 loading domain onto CTB1 MAT shows that the
12 AVES1 post-loading domain linker contains an α-helix that matches the linker architecture of MAT in CTB1 (and AT domains in modpks).
 Superpositions onto previously known structures (as listed in parenthesis) of the SAT (a, pink, with CazM SAT in white), MAT (b, red, with CurL AT in wheat), and KS (c, orange, with CurL KS in")
13 Supplementary Figure 5 The individual active sites of SAT, MAT and KS are structurally conserved. (a-c) Superpositions onto previously known structures (as listed in parenthesis) of the SAT (a, pink, with CazM SAT in white), MAT (b, red, with CurL AT in wheat), and KS (c, orange, with CurL KS in light orange) domains are shown in cartoon representation. Active site residues are shown in stick representation with residue numbers corresponding to CTB1. Electron density maps for CTB1 active site residues are shown with a blue (2Fo-Fc at 1.0 σ) and green (Fo-Fc at 3.0 σ) mesh. Unidentified difference density around the active site residues C119 (SAT) and C553 (KS) indicated partial acylation or oxidation. (d) A narrow and hydrophobic active site tunnel (white) connects the KS active sites.

14
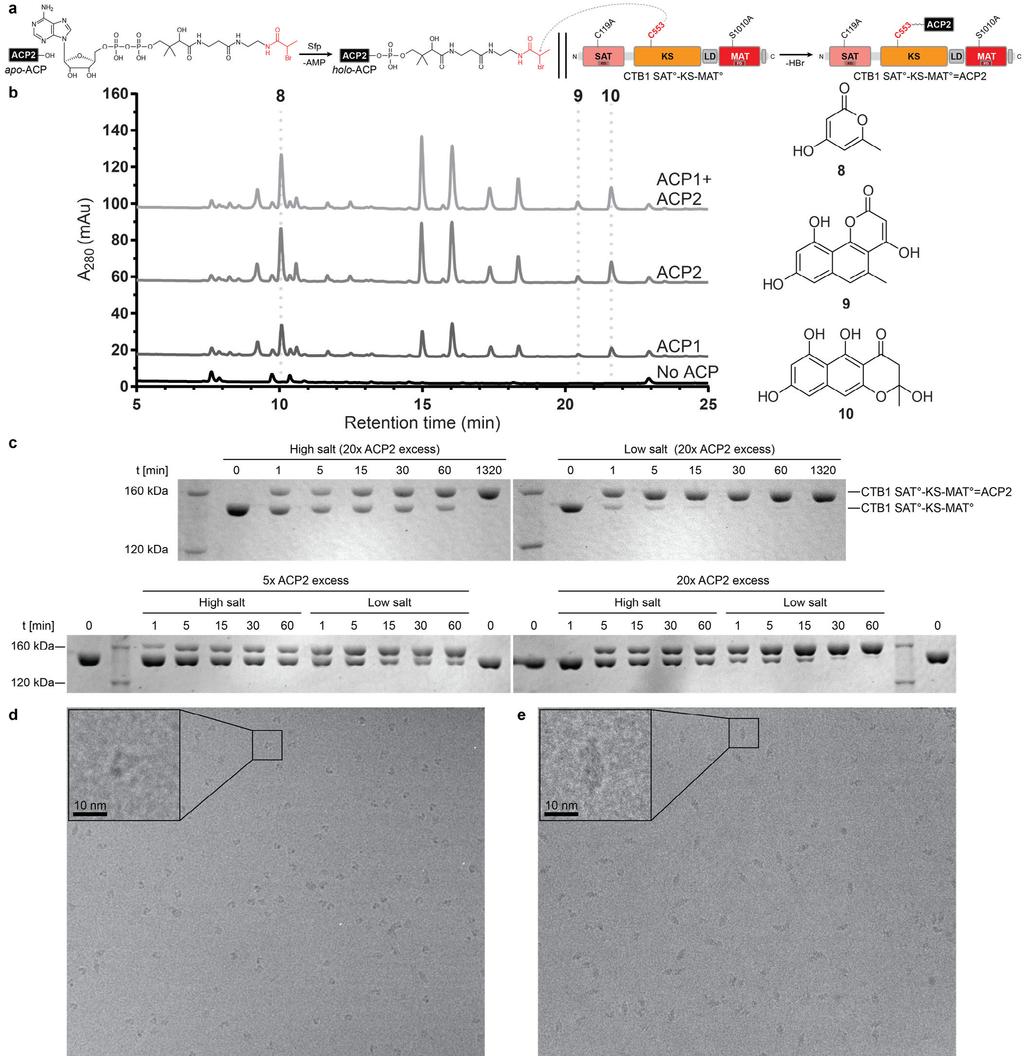
15 Supplementary Figure 6 ACP activity, crosslinking and cryo-em sample preparation. (a) Scheme for ACP2 loading with a dephospho-amino-coa analog (black), containing the α- bromopropionyl crosslinker (red), and site specific crosslinking to the KS domain of CTB1 SAT -KS-MAT. (b) HPLC chromatograms (280 nm) of minimal in vitro CTB1 reconstitution reactions with SAT-KS-MAT and ACP1 (10 μm), ACP2 (10 μm), or both (5 μm each). Curves are off-set for visualization. Previously characterized CTB1 derailment products 8, 9, and 10, and negative controls lacking ACP are indicated. No increase in activity is observed when ACP1 and ACP2 are combined, suggesting they are biosynthetically equivalent and interchangeable. These experiments were repeated twice with similar results. (c) Denaturing SDS-PAGE analysis with Coomassie staining of crosslinking time course in high (50 mm Tris ph 7.5, 250 mm NaCl) and low (50 mm Tris ph 7.5) salt buffer as well as for two different CTB1 SAT -KS- MAT :ACP2 ratios. Crosslinking proceeds faster in low salt buffer and at higher excess of ACP2. Gels are cropped. These experiments were repeated more than three times from independent protein preparations with similar results. (d,e) Drift corrected and dose filtered cryo- EM micrographs reveal clearly recognizable shapes for monomeric (d) and dimeric (e) particles. More than 10 grids have been prepared independently from more than three protein preparations with similar results. (f) Denaturating SDS-PAGE analysis with Coomassie staining of the sample used for cryo-em grid preparation and data collection before SEC and after grid preparation.

16
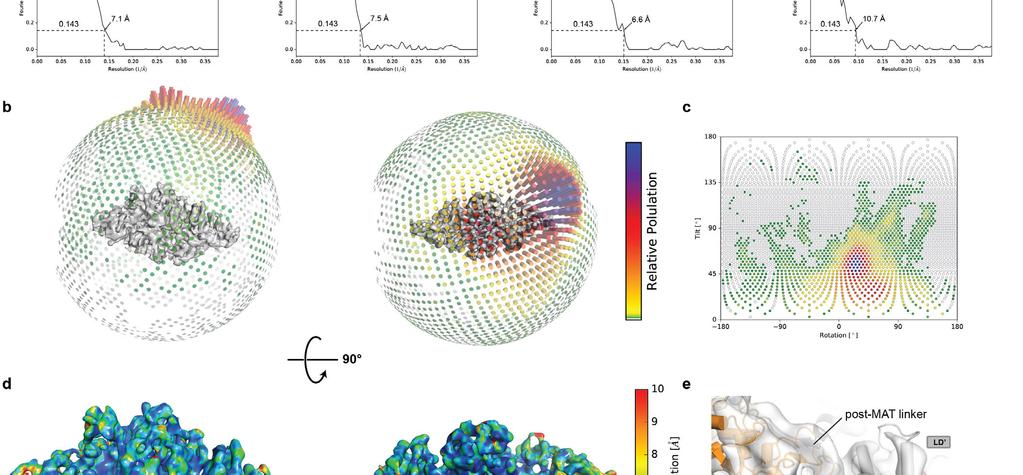
17 Supplementary Figure 7 Cryo-EM data processing scheme. (a) 2D- and 3D-classification and sorting scheme for reconstructions of CTB1 SAT -KS-MAT =ACP2 dimers as well as monomers. 3D-class distributions are indicated below the models. FSC plots (corrected for effects of the mask) used for determining resolution based on the criterion (Rosenthal, P. B. & Henderson, R. J. Mol. Biol. 333, (2003)) is shown below the final models. A single data set has been collected from one pre-screened protein and grid preparation. (b, c) 3D (b) and 2D (c) angular distribution plot based on the alignment used for the final reconstruction. (d) Local resolution map of the final reconstruction at 7.1 Å resolution shows a resolution below 5 Å in the central KS region, decreasing towards the lateral MAT domains to around 10 Å. (e) The final map reveals features of secondary structure elements as well as linkers. All maps are contoured at 7.1σ.
. The potential ACP2 binding region on the KS surface is less ordered on the side lacking ACP2 density.")
18 Supplementary Figure 8 Conformational variability in the cryo-em structure of CTB1 SAT 0 -KS-MAT 0 =ACP2. (a, b) KS-based C2 symmetry differences map of the final reconstruction at 7.1 Å resolution, contoured at 7.1σ (green: positive, red: negative). The potential ACP2 binding region on the KS surface is less ordered on the side lacking ACP2 density. (b) An unmasked map of the final reconstruction contoured at 3σ and colored by distance to the atomic model shows enhanced noise around the second ACP binding site.
19 Supplementary Figure 9 CTB1 mutant activity in deconstruction experiments and ACP2 interface validation. Representative HPLC chromatograms (280 nm) of full in vitro CTB1 reconstitution reactions (SAT-KS-MAT + ACP2 + PT + TE) with SAT-KS-MAT interface mutants are shown. Curves are off-set for visualization. Nor-toralactone production is indicated by a peak at a retention time of 28.6 min. These experiments were repeated three times with similar results.
20
21 Supplementary Figure 10 CTB1 SAT-KS-MAT alignment of regions interfacing ACP2 and phylogeny. All sequences are labelled as protein name (organism abbreviation) Uniprot number. (a) Sequence alignment of regions relevant for ACP2 interaction. *, indicates interface mutants that reduce activity. (b) Phylogenetic analysis of NR-PKS indicates three main clades with one clustering around CTB1. Distance unit is given as accepted amino-acid substitutions per site and indicated by scale bar.
22
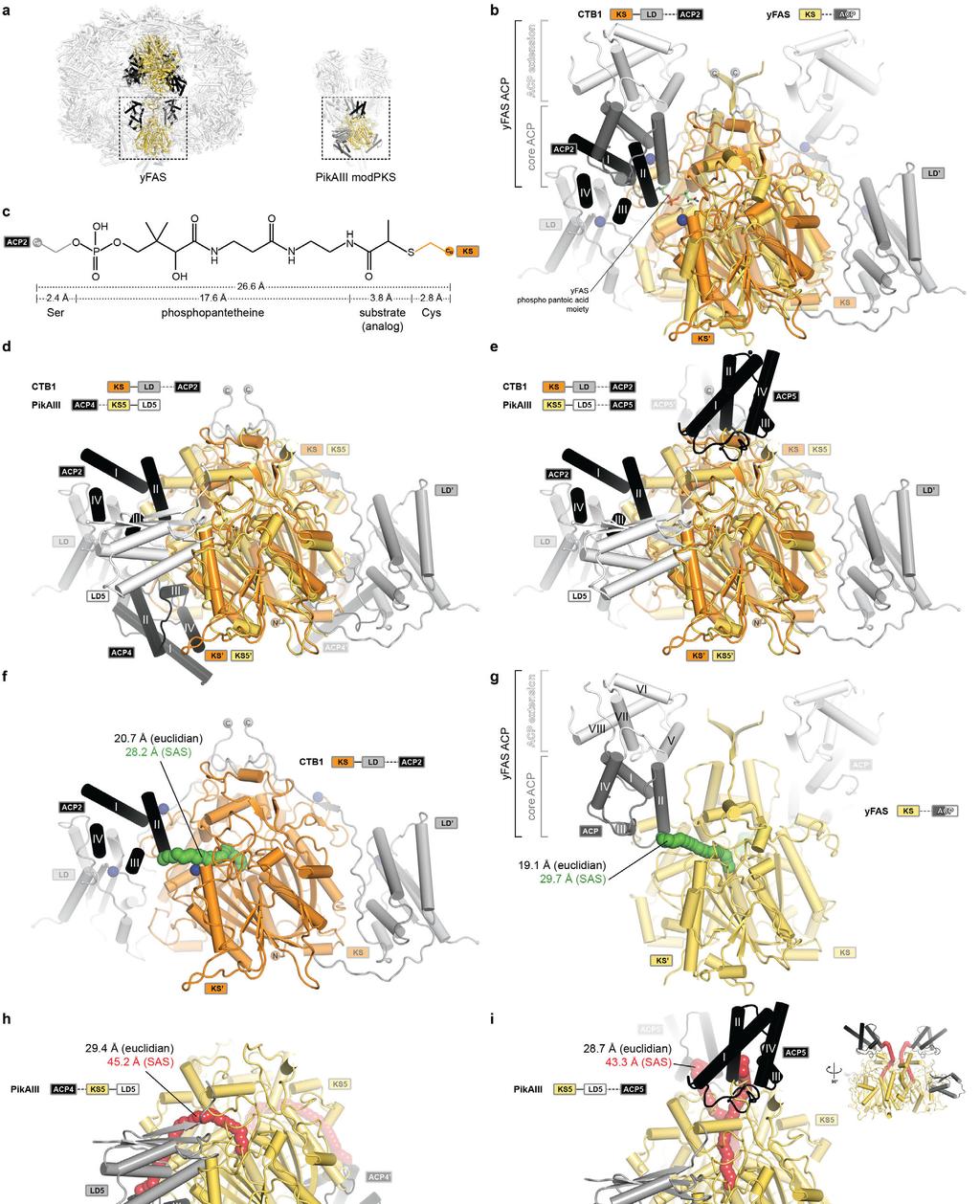
23 Supplementary Figure 11 Comparison of KS-ACP interactions in PKS and FAS. (a) ACP interactions with multienzyme KS domains have been observed in the hetero-dodecameric yfas and PikAIII modpks. Superposition of the architecturally distinct yfas-ks with CTB1 KS- LD =ACP2. (b) The divergent ACP of yfas consists of a C-terminal four-helix bundle extension (white) and the conserved four-helix bundle at the N-terminus, which interacts with the KS in a similar position as observed for ACP2 in CTB1. ACP-interfacing residues in CTB1 are indicated by blue spheres (see Fig. 3) (c) Derivation of the length of a fully extended crosslink between ACP2 and KS in CTB1. The defined length of the Ppant cofactor bridges a distance of approx. 27 Å between the ACP-Ser and the KS-Cys Cα atoms for functional ACP interaction with active sites. (d, e) Superpositions of PikAIII with ACP4 (d) and ACP5 (e) on CTB1 KS- LD=ACP2 show the relative locations of the ACP binding interfaces. The LD5 arrangement in PikAIII is inverse to CTB1. (f-i) Cα-Cα solvent accessible surface (SAS) distances between the ACP anchor and the KS active site cysteine in CTB1 (f), yfas (g), and PikAIII with ACP4 (h) and ACP5 (i) are indicated. Solvent accessible surface paths are indicated by tubes of spheres and colored according to the agreement (green) or disagreement (red) with the length of the Ppant cofactor. The path for ACP5 to KS5 in PikAIII (i) is only accessible in the absence of the post-(m)at linker, which was not resolved in the cryo-em structure. An additional miniaturized-sideview is indicated. Corresponding euclidian distances are given in the panels. Domain colors are indicated in the panels; only KS, LD, and ACP domains are shown (b-i).
 and black (SAT -KS-MAT =ACP2).")
24 Supplementary Figure 12 Structural comparison between CTB1 SAT-KS-MAT and SAT -KS-AT =ACP2. (a) Superposition on the central KS dimer. Transparent cartoon representation of CTB1 is shown in white (SAT-KS-MAT) and black (SAT -KS-MAT =ACP2). Selected helices are shown in light (SAT-KS-MAT) and dark (SAT -KS-MAT =ACP2) domain
25 colors. (b) Schematic representation of relative domain motions. Distances are derived by domain-wise Cα-r.m.s.d. calculations. (c, d) Structural differences at the MAT-SAT interface in the cryo-em reconstruction are shown for the unsharpened map. While the side with visualized ACP2 (c) reveals a gap, the side with unresolved ACP2 shows a connected interface (d).
26 Supplementary Video 1 ACP2 binding to the CTB1 loading/condensing region. Structures of the CTB1 loading/condensing region with and without crosslinked ACP2 are morphed. In the ACP2-bound structure, ACP is visualized at a single binding-site in an overall structure with increased asymmetry. ACP2 is located in a cleft between the KS dimer and one LD. Around ACP2, LD and MAT are moved upwards, and the interface between the neighboring MAT and SAT domains is loosened. The SAT is translocated towards the KS and the second SAT is pushed against the KS and MAT in the opposite half of the dimer.
(B D) Three views of the final refined 2Fo-Fc electron density map of the Vpr (red)-ung2 (green) interacting region, contoured at 1.4σ.
 Three views of the final refined 2Fo-Fc electron density map of the Vpr (red)-ung2 (green) interacting region, contoured at 1.4σ.") Supplementary Figure 1 Overall structure of the DDB1 DCAF1 Vpr UNG2 complex. (A) The final refined 2Fo-Fc electron density map, contoured at 1.4σ of Vpr, illustrating well-defined side chains. (B D) Three
Supplementary Figure 1 Overall structure of the DDB1 DCAF1 Vpr UNG2 complex. (A) The final refined 2Fo-Fc electron density map, contoured at 1.4σ of Vpr, illustrating well-defined side chains. (B D) Three
Supplementary Materials for
 www.sciencemag.org/cgi/content/full/science.aal4326/dc1 Supplementary Materials for Structure of a eukaryotic voltage-gated sodium channel at near-atomic resolution Huaizong Shen, Qiang Zhou, Xiaojing
www.sciencemag.org/cgi/content/full/science.aal4326/dc1 Supplementary Materials for Structure of a eukaryotic voltage-gated sodium channel at near-atomic resolution Huaizong Shen, Qiang Zhou, Xiaojing
Supplementary Figure 1 (previous page). EM analysis of full-length GCGR. (a) Exemplary tilt pair images of the GCGR mab23 complex acquired for Random
. EM analysis of full-length GCGR. (a) Exemplary tilt pair images of the GCGR mab23 complex acquired for Random") S1 Supplementary Figure 1 (previous page). EM analysis of full-length GCGR. (a) Exemplary tilt pair images of the GCGR mab23 complex acquired for Random Conical Tilt (RCT) reconstruction (left: -50,right:
S1 Supplementary Figure 1 (previous page). EM analysis of full-length GCGR. (a) Exemplary tilt pair images of the GCGR mab23 complex acquired for Random Conical Tilt (RCT) reconstruction (left: -50,right:
Supplementary Information Janssen et al.
 Supplementary Information Janssen et al. Insights into complement convertase formation based on the structure of the factor B CVF complex Bert J.C. Janssen 1, Lucio Gomes 1, Roman I. Koning 2, Dmitri I.
Supplementary Information Janssen et al. Insights into complement convertase formation based on the structure of the factor B CVF complex Bert J.C. Janssen 1, Lucio Gomes 1, Roman I. Koning 2, Dmitri I.
Supplementary Table 1. Data collection and refinement statistics (molecular replacement).
.") Supplementary Table 1. Data collection and refinement statistics (molecular replacement). Data set statistics HLA A*0201- ALWGPDPAAA PPI TCR PPI TCR/A2- ALWGPDPAAA PPI TCR/A2- ALWGPDPAAA Space Group P2
Supplementary Table 1. Data collection and refinement statistics (molecular replacement). Data set statistics HLA A*0201- ALWGPDPAAA PPI TCR PPI TCR/A2- ALWGPDPAAA PPI TCR/A2- ALWGPDPAAA Space Group P2
Table S1. X-ray data collection and refinement statistics
 Table S1. X-ray data collection and refinement statistics Data collection H7.167 Fab-Sh2/H7 complex Beamline SSRL 12-2 Wavelength (Å) 0.97950 Space group I2 1 3 Unit cell parameters (Å, º) a = b = c=207.3,
Table S1. X-ray data collection and refinement statistics Data collection H7.167 Fab-Sh2/H7 complex Beamline SSRL 12-2 Wavelength (Å) 0.97950 Space group I2 1 3 Unit cell parameters (Å, º) a = b = c=207.3,
Detergent solubilised 5 TMD binds pregnanolone at the Q245 neurosteroid potentiation site.
 Supplementary Figure 1 Detergent solubilised 5 TMD binds pregnanolone at the Q245 neurosteroid potentiation site. (a) Gel filtration profiles of purified 5 TMD samples at 100 nm, heated beforehand for
Supplementary Figure 1 Detergent solubilised 5 TMD binds pregnanolone at the Q245 neurosteroid potentiation site. (a) Gel filtration profiles of purified 5 TMD samples at 100 nm, heated beforehand for
Transient β-hairpin Formation in α-synuclein Monomer Revealed by Coarse-grained Molecular Dynamics Simulation
 Transient β-hairpin Formation in α-synuclein Monomer Revealed by Coarse-grained Molecular Dynamics Simulation Hang Yu, 1, 2, a) Wei Han, 1, 3, b) Wen Ma, 1, 2 1, 2, 3, c) and Klaus Schulten 1) Beckman
Transient β-hairpin Formation in α-synuclein Monomer Revealed by Coarse-grained Molecular Dynamics Simulation Hang Yu, 1, 2, a) Wei Han, 1, 3, b) Wen Ma, 1, 2 1, 2, 3, c) and Klaus Schulten 1) Beckman
Supplementary Materials for
 advances.sciencemag.org/cgi/content/full/2/4/e1500980/dc1 Supplementary Materials for The crystal structure of human dopamine -hydroxylase at 2.9 Å resolution Trine V. Vendelboe, Pernille Harris, Yuguang
advances.sciencemag.org/cgi/content/full/2/4/e1500980/dc1 Supplementary Materials for The crystal structure of human dopamine -hydroxylase at 2.9 Å resolution Trine V. Vendelboe, Pernille Harris, Yuguang
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION doi:10.1038/nature22394 Supplementary Table 1 Observed intermolecular interactions within the GLP-1:GLP-1R TMD interface. Superscripts refer to the Wootten residue numbering system
SUPPLEMENTARY INFORMATION doi:10.1038/nature22394 Supplementary Table 1 Observed intermolecular interactions within the GLP-1:GLP-1R TMD interface. Superscripts refer to the Wootten residue numbering system
Supplementary Materials for
 advances.sciencemag.org/cgi/content/full/4/3/eaaq0762/dc1 Supplementary Materials for Structures of monomeric and oligomeric forms of the Toxoplasma gondii perforin-like protein 1 Tao Ni, Sophie I. Williams,
advances.sciencemag.org/cgi/content/full/4/3/eaaq0762/dc1 Supplementary Materials for Structures of monomeric and oligomeric forms of the Toxoplasma gondii perforin-like protein 1 Tao Ni, Sophie I. Williams,
Supplementary Figure 1 Preparation, crystallization and structure determination of EpEX. (a), Purified EpEX and EpEX analyzed on homogenous 12.
, Purified EpEX and EpEX analyzed on homogenous 12.") Supplementary Figure 1 Preparation, crystallization and structure determination of EpEX. (a), Purified EpEX and EpEX analyzed on homogenous 12.5 % SDS-PAGE gel under reducing and non-reducing conditions.
Supplementary Figure 1 Preparation, crystallization and structure determination of EpEX. (a), Purified EpEX and EpEX analyzed on homogenous 12.5 % SDS-PAGE gel under reducing and non-reducing conditions.
Supplementary Materials for
 advances.sciencemag.org/cgi/content/full/2/9/e1600292/dc1 Supplementary Materials for Native phasing of x-ray free-electron laser data for a G protein coupled receptor Alexander Batyuk, Lorenzo Galli,
advances.sciencemag.org/cgi/content/full/2/9/e1600292/dc1 Supplementary Materials for Native phasing of x-ray free-electron laser data for a G protein coupled receptor Alexander Batyuk, Lorenzo Galli,
Supporting Information
 Supporting Information Guan et al. 10.1073/pnas.1217609110 Fig. S1. Three patterns of reactivity for CD4-induced (CD4i) mabs. The following representative ELISAs show three patterns of reactivity for CD4i
Supporting Information Guan et al. 10.1073/pnas.1217609110 Fig. S1. Three patterns of reactivity for CD4-induced (CD4i) mabs. The following representative ELISAs show three patterns of reactivity for CD4i
Supporting Information
 Supporting Information Mechanism of inactivation of -aminobutyric acid aminotransferase by (1S,3S)-3-amino-4-difluoromethylenyl-1- cyclopentanoic acid (CPP-115) Hyunbeom Lee, 1, Emma H. Doud, 1,2 Rui Wu,
Supporting Information Mechanism of inactivation of -aminobutyric acid aminotransferase by (1S,3S)-3-amino-4-difluoromethylenyl-1- cyclopentanoic acid (CPP-115) Hyunbeom Lee, 1, Emma H. Doud, 1,2 Rui Wu,
FCC2 5CY7 FCC1 5CY8. Actinonin 5CVQ
 Table S1. Data collection and refinement statistics Data collection Apo 5E5D MA 5CPD MAS 5CP0 Actinonin 5CVQ FCC1 5CY8 FCC2 5CY7 FCC 5CWY FCC4 5CVK FCC5 5CWX FCC6 5CVP X-ray source 17A-KEK PAL4A PAL4A
Table S1. Data collection and refinement statistics Data collection Apo 5E5D MA 5CPD MAS 5CP0 Actinonin 5CVQ FCC1 5CY8 FCC2 5CY7 FCC 5CWY FCC4 5CVK FCC5 5CWX FCC6 5CVP X-ray source 17A-KEK PAL4A PAL4A
Table S1: Kinetic parameters of drug and substrate binding to wild type and HIV-1 protease variants. Data adapted from Ref. 6 in main text.
 Dynamical Network of HIV-1 Protease Mutants Reveals the Mechanism of Drug Resistance and Unhindered Activity Rajeswari Appadurai and Sanjib Senapati* BJM School of Biosciences and Department of Biotechnology,
Dynamical Network of HIV-1 Protease Mutants Reveals the Mechanism of Drug Resistance and Unhindered Activity Rajeswari Appadurai and Sanjib Senapati* BJM School of Biosciences and Department of Biotechnology,
Nature Structural & Molecular Biology: doi: /nsmb Supplementary Figure 1
 Supplementary Figure 1 Design of isolated protein and RNC constructs, and homogeneity of purified RNCs. (a) Schematic depicting the design and nomenclature used for all the isolated proteins and RNCs used
Supplementary Figure 1 Design of isolated protein and RNC constructs, and homogeneity of purified RNCs. (a) Schematic depicting the design and nomenclature used for all the isolated proteins and RNCs used
Modeling holo-acp:dh and holo-acp:kr complexes of modular polyketide synthases: a docking and molecular dynamics study
 Anand and Mohanty BMC Structural Biology 2012, 12:10 RESEARCH ARTICLE Open Access Modeling holo-acp:dh and holo-acp:kr complexes of modular polyketide synthases: a docking and molecular dynamics study
Anand and Mohanty BMC Structural Biology 2012, 12:10 RESEARCH ARTICLE Open Access Modeling holo-acp:dh and holo-acp:kr complexes of modular polyketide synthases: a docking and molecular dynamics study
SUPPLEMENTARY INFORMATION FOR. (R)-Profens Are Substrate-Selective Inhibitors of Endocannabinoid Oxygenation. by COX-2
-Profens Are Substrate-Selective Inhibitors of Endocannabinoid Oxygenation. by COX-2") SUPPLEMENTARY INFORMATION FOR (R)-Profens Are Substrate-Selective Inhibitors of Endocannabinoid Oxygenation by COX-2 Kelsey C. Duggan, Daniel J. Hermanson, Joel Musee, Jeffery J. Prusakiewicz, Jami L.
SUPPLEMENTARY INFORMATION FOR (R)-Profens Are Substrate-Selective Inhibitors of Endocannabinoid Oxygenation by COX-2 Kelsey C. Duggan, Daniel J. Hermanson, Joel Musee, Jeffery J. Prusakiewicz, Jami L.
Atypical Natural Killer T-cell receptor recognition of CD1d-lipid antigens supplementary Information.
 Atypical Natural Killer T-cell receptor recognition of CD1d-lipid antigens supplementary Information. Supplementary Figure 1. Phenotypic analysis of TRBV25-1 + and TRBV25-1 - CD1d-α-GalCerreactive cells.
Atypical Natural Killer T-cell receptor recognition of CD1d-lipid antigens supplementary Information. Supplementary Figure 1. Phenotypic analysis of TRBV25-1 + and TRBV25-1 - CD1d-α-GalCerreactive cells.
Supporting Information
 Supporting Information McCullough et al. 10.1073/pnas.0801567105 A α10 α8 α9 N α7 α6 α5 C β2 β1 α4 α3 α2 α1 C B N C Fig. S1. ALIX Bro1 in complex with the C-terminal CHMP4A helix. (A) Ribbon diagram showing
Supporting Information McCullough et al. 10.1073/pnas.0801567105 A α10 α8 α9 N α7 α6 α5 C β2 β1 α4 α3 α2 α1 C B N C Fig. S1. ALIX Bro1 in complex with the C-terminal CHMP4A helix. (A) Ribbon diagram showing
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature10913 Supplementary Figure 1 2F o -F c electron density maps of cognate and near-cognate trna Leu 2 in the A site of the 70S ribosome. The maps are contoured at 1.2 sigma and some of
doi:10.1038/nature10913 Supplementary Figure 1 2F o -F c electron density maps of cognate and near-cognate trna Leu 2 in the A site of the 70S ribosome. The maps are contoured at 1.2 sigma and some of
Structure of the measles virus hemagglutinin bound to the CD46 receptor. César Santiago, María L. Celma, Thilo Stehle and José M.
 Supporting Figures and Table for Structure of the measles virus hemagglutinin bound to the CD46 receptor César Santiago, María L. Celma, Thilo Stehle and José M. Casasnovas This PDF file includes: Supplementary
Supporting Figures and Table for Structure of the measles virus hemagglutinin bound to the CD46 receptor César Santiago, María L. Celma, Thilo Stehle and José M. Casasnovas This PDF file includes: Supplementary
Supporting Information Identification of Amino Acids with Sensitive Nanoporous MoS 2 : Towards Machine Learning-Based Prediction
 Supporting Information Identification of Amino Acids with Sensitive Nanoporous MoS 2 : Towards Machine Learning-Based Prediction Amir Barati Farimani, Mohammad Heiranian, Narayana R. Aluru 1 Department
Supporting Information Identification of Amino Acids with Sensitive Nanoporous MoS 2 : Towards Machine Learning-Based Prediction Amir Barati Farimani, Mohammad Heiranian, Narayana R. Aluru 1 Department
Phenylketonuria (PKU) Structure of Phenylalanine Hydroxylase. Biol 405 Molecular Medicine
 Structure of Phenylalanine Hydroxylase. Biol 405 Molecular Medicine") Phenylketonuria (PKU) Structure of Phenylalanine Hydroxylase Biol 405 Molecular Medicine 1998 Crystal structure of phenylalanine hydroxylase solved. The polypeptide consists of three regions: Regulatory
Phenylketonuria (PKU) Structure of Phenylalanine Hydroxylase Biol 405 Molecular Medicine 1998 Crystal structure of phenylalanine hydroxylase solved. The polypeptide consists of three regions: Regulatory
List of Figures. List of Tables
 Supporting Information for: Signaling Domain of Sonic Hedgehog as Cannibalistic Calcium-Regulated Zinc-Peptidase Rocio Rebollido-Rios 1, Shyam Bandari 3, Christoph Wilms 1, Stanislav Jakuschev 1, Andrea
Supporting Information for: Signaling Domain of Sonic Hedgehog as Cannibalistic Calcium-Regulated Zinc-Peptidase Rocio Rebollido-Rios 1, Shyam Bandari 3, Christoph Wilms 1, Stanislav Jakuschev 1, Andrea
Introduction to proteins and protein structure
 Introduction to proteins and protein structure The questions and answers below constitute an introduction to the fundamental principles of protein structure. They are all available at [link]. What are
Introduction to proteins and protein structure The questions and answers below constitute an introduction to the fundamental principles of protein structure. They are all available at [link]. What are
Nature Structural & Molecular Biology: doi: /nsmb Supplementary Figure 1
 Supplementary Figure 1 The UBL and RING1 interface remains associated in the complex structures of Parkin and pub. a) Asymmetric Unit of crystal structure of UBLR0RBR and pub complex showing UBL (green),
Supplementary Figure 1 The UBL and RING1 interface remains associated in the complex structures of Parkin and pub. a) Asymmetric Unit of crystal structure of UBLR0RBR and pub complex showing UBL (green),
Unveiling transient protein-protein interactions that modulate inhibition of alpha-synuclein aggregation
 Supplementary information Unveiling transient protein-protein interactions that modulate inhibition of alpha-synuclein aggregation by beta-synuclein, a pre-synaptic protein that co-localizes with alpha-synuclein.
Supplementary information Unveiling transient protein-protein interactions that modulate inhibition of alpha-synuclein aggregation by beta-synuclein, a pre-synaptic protein that co-localizes with alpha-synuclein.
Structural biology of viruses
 Structural biology of viruses Biophysical Chemistry 1, Fall 2010 Coat proteins DNA/RNA packaging Reading assignment: Chap. 15 Virus particles self-assemble from coat monomers Virus Structure and Function
Structural biology of viruses Biophysical Chemistry 1, Fall 2010 Coat proteins DNA/RNA packaging Reading assignment: Chap. 15 Virus particles self-assemble from coat monomers Virus Structure and Function
SDS-Assisted Protein Transport Through Solid-State Nanopores
 Supplementary Information for: SDS-Assisted Protein Transport Through Solid-State Nanopores Laura Restrepo-Pérez 1, Shalini John 2, Aleksei Aksimentiev 2 *, Chirlmin Joo 1 *, Cees Dekker 1 * 1 Department
Supplementary Information for: SDS-Assisted Protein Transport Through Solid-State Nanopores Laura Restrepo-Pérez 1, Shalini John 2, Aleksei Aksimentiev 2 *, Chirlmin Joo 1 *, Cees Dekker 1 * 1 Department
Data are contained in multiple tabs in Excel spreadsheets and in CSV files.
 Contents Overview Curves Methods Measuring enzymatic activity (figure 2) Enzyme characterisation (Figure S1, S2) Enzyme kinetics (Table 3) Effect of ph on activity (figure 3B) Effect of metals and inhibitors
Contents Overview Curves Methods Measuring enzymatic activity (figure 2) Enzyme characterisation (Figure S1, S2) Enzyme kinetics (Table 3) Effect of ph on activity (figure 3B) Effect of metals and inhibitors
Structural analysis of fungus-derived FAD glucose dehydrogenase
 Structural analysis of fungus-derived FAD glucose dehydrogenase Hiromi Yoshida 1, Genki Sakai 2, Kazushige Mori 3, Katsuhiro Kojima 3, Shigehiro Kamitori 1, and Koji Sode 2,3,* 1 Life Science Research
Structural analysis of fungus-derived FAD glucose dehydrogenase Hiromi Yoshida 1, Genki Sakai 2, Kazushige Mori 3, Katsuhiro Kojima 3, Shigehiro Kamitori 1, and Koji Sode 2,3,* 1 Life Science Research
SUPPLEMENTARY INFORMATION. Computational Assay of H7N9 Influenza Neuraminidase Reveals R292K Mutation Reduces Drug Binding Affinity
 SUPPLEMENTARY INFORMATION Computational Assay of H7N9 Influenza Neuraminidase Reveals R292K Mutation Reduces Drug Binding Affinity Christopher Woods 1, Maturos Malaisree 1, Ben Long 2, Simon McIntosh-Smith
SUPPLEMENTARY INFORMATION Computational Assay of H7N9 Influenza Neuraminidase Reveals R292K Mutation Reduces Drug Binding Affinity Christopher Woods 1, Maturos Malaisree 1, Ben Long 2, Simon McIntosh-Smith
STRUCTURE OF A UBIQUITIN-LOADED HECT LIGASE REVEALS THE MOLECULAR BASIS FOR CATALYTIC PRIMING SUPPLEMENTARY INFORMATION
 STRUCTURE OF A UBIQUITINLOADED ECT LIGASE REVEALS TE MOLECULAR BASIS FOR CATALYTIC PRIMING Elena Maspero 1, Eleonora Valentini 1, Sara Mari 1, Valentina Cecatiello 2, Paolo Soffientini 1, Sebastiano Pasqualato
STRUCTURE OF A UBIQUITINLOADED ECT LIGASE REVEALS TE MOLECULAR BASIS FOR CATALYTIC PRIMING Elena Maspero 1, Eleonora Valentini 1, Sara Mari 1, Valentina Cecatiello 2, Paolo Soffientini 1, Sebastiano Pasqualato
Nature Structural & Molecular Biology: doi: /nsmb.1933
 The structural basis of open channel block in a prokaryotic pentameric ligand-gated ion channel Ricarda J. C. Hilf, Carlo Bertozzi, Iwan Zimmermann, Alwin Reiter, Dirk Trauner and Raimund Dutzler a GLIC
The structural basis of open channel block in a prokaryotic pentameric ligand-gated ion channel Ricarda J. C. Hilf, Carlo Bertozzi, Iwan Zimmermann, Alwin Reiter, Dirk Trauner and Raimund Dutzler a GLIC
Supplementary Material
 Supplementary Material Materials and methods Enzyme assay The enzymatic activity of -glucosidase toward salicin was measured with the Miller method (Miller, 1959) using glucose as the standard. A total
Supplementary Material Materials and methods Enzyme assay The enzymatic activity of -glucosidase toward salicin was measured with the Miller method (Miller, 1959) using glucose as the standard. A total
SUPPLEMENTAL MATERIAL. UNC119 is required for G protein trafficking in sensory neurons
 1 SUPPLEMENTAL MATERIAL UNC119 is required for G protein trafficking in sensory neurons Houbin Zhang, Ryan N. Constantine, Sergey Vorobiev, Yang Chen, Jayaraman Seetharaman, Yuanpeng Janet Huang, Rong
1 SUPPLEMENTAL MATERIAL UNC119 is required for G protein trafficking in sensory neurons Houbin Zhang, Ryan N. Constantine, Sergey Vorobiev, Yang Chen, Jayaraman Seetharaman, Yuanpeng Janet Huang, Rong
Chapter 6. X-ray structure analysis of D30N tethered HIV-1 protease. dimer/saquinavir complex
 Chapter 6 X-ray structure analysis of D30N tethered HIV-1 protease dimer/saquinavir complex 6.1 Introduction: The arrival of HIV protease inhibitors (PIs) in late 1995 marked the beginning of an important
Chapter 6 X-ray structure analysis of D30N tethered HIV-1 protease dimer/saquinavir complex 6.1 Introduction: The arrival of HIV protease inhibitors (PIs) in late 1995 marked the beginning of an important
Supplementary Information for Structural basis for the inhibition of Polo-like kinase 1
 Supplementary Information for Structural basis for inhibition of Polo-like kinase 1 Jun Xu, Chen Shen, Tao Wang & Junmin Quan Correspondence authors: Tao Wang, taowang@pkusz.edu.cn; Junmin Quan, quanjm@szpku.edu.cn
Supplementary Information for Structural basis for inhibition of Polo-like kinase 1 Jun Xu, Chen Shen, Tao Wang & Junmin Quan Correspondence authors: Tao Wang, taowang@pkusz.edu.cn; Junmin Quan, quanjm@szpku.edu.cn
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION TITLE: Structural Basis of Signal Sequence Surveillance and Selection by the SRP-SR Complex AUTHORS and AFFILIATIONS Ottilie von Loeffelholz 1,2, Kèvin Knoops 1,2,6, Aileen Ariosa
SUPPLEMENTARY INFORMATION TITLE: Structural Basis of Signal Sequence Surveillance and Selection by the SRP-SR Complex AUTHORS and AFFILIATIONS Ottilie von Loeffelholz 1,2, Kèvin Knoops 1,2,6, Aileen Ariosa
Supplementary Information
 Supplementary Information Using the pimeloyl-coa synthetase adenylation fold to synthesise fatty acid thioesters Menglu Wang 1a ; Lucile Moynié 2a, Peter J. Harrison 1, Van Kelly 1, Andrew Piper 1, James
Supplementary Information Using the pimeloyl-coa synthetase adenylation fold to synthesise fatty acid thioesters Menglu Wang 1a ; Lucile Moynié 2a, Peter J. Harrison 1, Van Kelly 1, Andrew Piper 1, James
Thermal shift binding experiments were carried out using Thermofluor 384 ELS system. Protein
 Supplementary Methods Thermal shift assays Thermal shift binding experiments were carried out using Thermofluor 384 ELS system. Protein unfolding was examined by monitoring the fluorescence of ANS (1-anilinonaphthalene-8-
Supplementary Methods Thermal shift assays Thermal shift binding experiments were carried out using Thermofluor 384 ELS system. Protein unfolding was examined by monitoring the fluorescence of ANS (1-anilinonaphthalene-8-
SUPPLEMENTARY INFORMATION
 doi: 10.1038/nature07422 SUPPLEMENTARY INFRMATIN K S(P) R S I M Q(L4) R M 7 6 Sp Q I K R 5 4 3 2 1 L(L0) L L S E 0 +1 +2 +3 Figure S1a Difference electron density (mfo DFc) for the peptide (Qpeptide),
doi: 10.1038/nature07422 SUPPLEMENTARY INFRMATIN K S(P) R S I M Q(L4) R M 7 6 Sp Q I K R 5 4 3 2 1 L(L0) L L S E 0 +1 +2 +3 Figure S1a Difference electron density (mfo DFc) for the peptide (Qpeptide),
HDL surface lipids mediate CETP binding as revealed by electron microscopy and molecular dynamics simulation
 HDL surface lipids mediate CETP binding as revealed by electron microscopy and molecular dynamics simulation Meng Zhang 1, River Charles 1, Huimin Tong 1, Lei Zhang 1, Mili Patel 2, Francis Wang 1, Matthew
HDL surface lipids mediate CETP binding as revealed by electron microscopy and molecular dynamics simulation Meng Zhang 1, River Charles 1, Huimin Tong 1, Lei Zhang 1, Mili Patel 2, Francis Wang 1, Matthew
obtained for the simulations of the E2 conformation of SERCA in a pure POPC lipid bilayer (blue) and in a
 and in a") Supplementary Figure S1. Distribution of atoms along the bilayer normal. Normalized density profiles obtained for the simulations of the E2 conformation of SERCA in a pure POPC lipid bilayer (blue) and
Supplementary Figure S1. Distribution of atoms along the bilayer normal. Normalized density profiles obtained for the simulations of the E2 conformation of SERCA in a pure POPC lipid bilayer (blue) and
Structural Characterization of Prion-like Conformational Changes of the Neuronal Isoform of Aplysia CPEB
 Structural Characterization of Prion-like Conformational Changes of the Neuronal Isoform of Aplysia CPEB Bindu L. Raveendra, 1,5 Ansgar B. Siemer, 2,6 Sathyanarayanan V. Puthanveettil, 1,3,7 Wayne A. Hendrickson,
Structural Characterization of Prion-like Conformational Changes of the Neuronal Isoform of Aplysia CPEB Bindu L. Raveendra, 1,5 Ansgar B. Siemer, 2,6 Sathyanarayanan V. Puthanveettil, 1,3,7 Wayne A. Hendrickson,
Supplementary Information
 Supplementary Information Two common structural motifs for TCR recognition by staphylococcal enterotoxins Karin Erica Johanna Rödström 1, Paulina Regenthal 1, Christopher Bahl 2, Alex Ford 2, David Baker
Supplementary Information Two common structural motifs for TCR recognition by staphylococcal enterotoxins Karin Erica Johanna Rödström 1, Paulina Regenthal 1, Christopher Bahl 2, Alex Ford 2, David Baker
SUPPLEMENTARY INFORMATION (SI) FIGURES AND TABLES
 FIGURES AND TABLES") SUPPLEMENTARY INFORMATION (SI) FIGURES AND TABLES 1 Title: Discovery of a junctional epitope antibody that stabilizes IL-6 and gp80 protein:protein interaction and modulates its downstream signaling Authors:
SUPPLEMENTARY INFORMATION (SI) FIGURES AND TABLES 1 Title: Discovery of a junctional epitope antibody that stabilizes IL-6 and gp80 protein:protein interaction and modulates its downstream signaling Authors:
Figure S1. HP1α localizes to centromeres in mitosis and interacts with INCENP. (A&B) HeLa
 HeLa") SUPPLEMENTARY FIGURES Figure S1. HP1α localizes to centromeres in mitosis and interacts with INCENP. (A&B) HeLa tet-on cells that stably express HP1α-CFP, HP1β-CFP, or HP1γ-CFP were monitored with livecell
SUPPLEMENTARY FIGURES Figure S1. HP1α localizes to centromeres in mitosis and interacts with INCENP. (A&B) HeLa tet-on cells that stably express HP1α-CFP, HP1β-CFP, or HP1γ-CFP were monitored with livecell
Arginine side chain interactions and the role of arginine as a mobile charge carrier in voltage sensitive ion channels. Supplementary Information
 Arginine side chain interactions and the role of arginine as a mobile charge carrier in voltage sensitive ion channels Craig T. Armstrong, Philip E. Mason, J. L. Ross Anderson and Christopher E. Dempsey
Arginine side chain interactions and the role of arginine as a mobile charge carrier in voltage sensitive ion channels Craig T. Armstrong, Philip E. Mason, J. L. Ross Anderson and Christopher E. Dempsey
CS612 - Algorithms in Bioinformatics
 Spring 2016 Protein Structure February 7, 2016 Introduction to Protein Structure A protein is a linear chain of organic molecular building blocks called amino acids. Introduction to Protein Structure Amine
Spring 2016 Protein Structure February 7, 2016 Introduction to Protein Structure A protein is a linear chain of organic molecular building blocks called amino acids. Introduction to Protein Structure Amine
Rapid Characterization of Allosteric Networks. with Ensemble Normal Mode Analysis
 Supporting Information Rapid Characterization of Allosteric Networks with Ensemble Normal Mode Analysis Xin-Qiu Yao 1, Lars Skjærven 2, and Barry J. Grant 1* 1 Department of Computational Medicine and
Supporting Information Rapid Characterization of Allosteric Networks with Ensemble Normal Mode Analysis Xin-Qiu Yao 1, Lars Skjærven 2, and Barry J. Grant 1* 1 Department of Computational Medicine and
ATP-independent reversal of a membrane protein aggregate by a chloroplast SRP
 ATP-independent reversal of a membrane protein aggregate by a chloroplast SRP Peera Jaru-Ampornpan 1, Kuang Shen 1,3, Vinh Q. Lam 1,3, Mona Ali 2, Sebastian Doniach 2, Tony Z. Jia 1, Shu-ou Shan 1 Supplementary
ATP-independent reversal of a membrane protein aggregate by a chloroplast SRP Peera Jaru-Ampornpan 1, Kuang Shen 1,3, Vinh Q. Lam 1,3, Mona Ali 2, Sebastian Doniach 2, Tony Z. Jia 1, Shu-ou Shan 1 Supplementary
Bioinformatics for molecular biology
 Bioinformatics for molecular biology Structural bioinformatics tools, predictors, and 3D modeling Structural Biology Review Dr Research Scientist Department of Microbiology, Oslo University Hospital -
Bioinformatics for molecular biology Structural bioinformatics tools, predictors, and 3D modeling Structural Biology Review Dr Research Scientist Department of Microbiology, Oslo University Hospital -
Nature Structural and Molecular Biology: doi: /nsmb Supplementary Figure 1
 Supplementary Figure 1 Mutational analysis of the SA2-Scc1 interaction in vitro and in human cells. (a) Autoradiograph (top) and Coomassie stained gel (bottom) of 35 S-labeled Myc-SA2 proteins (input)
Supplementary Figure 1 Mutational analysis of the SA2-Scc1 interaction in vitro and in human cells. (a) Autoradiograph (top) and Coomassie stained gel (bottom) of 35 S-labeled Myc-SA2 proteins (input)
Supplementary Materials. High affinity binding of phosphatidylinositol-4-phosphate. by Legionella pneumophila DrrA
 Supplementary Materials High affinity binding of phosphatidylinositol-4-phosphate by Legionella pneumophila DrrA Running title: Molecular basis of PtdIns(4)P-binding by DrrA Stefan Schoebel, Wulf Blankenfeldt,
Supplementary Materials High affinity binding of phosphatidylinositol-4-phosphate by Legionella pneumophila DrrA Running title: Molecular basis of PtdIns(4)P-binding by DrrA Stefan Schoebel, Wulf Blankenfeldt,
This PDF file includes: Supplementary Figures 1 to 6 Supplementary Tables 1 to 2 Supplementary Methods Supplementary References
 Structure of the catalytic core of p300 and implications for chromatin targeting and HAT regulation Manuela Delvecchio, Jonathan Gaucher, Carmen Aguilar-Gurrieri, Esther Ortega, Daniel Panne This PDF file
Structure of the catalytic core of p300 and implications for chromatin targeting and HAT regulation Manuela Delvecchio, Jonathan Gaucher, Carmen Aguilar-Gurrieri, Esther Ortega, Daniel Panne This PDF file
This exam consists of two parts. Part I is multiple choice. Each of these 25 questions is worth 2 points.
 MBB 407/511 Molecular Biology and Biochemistry First Examination - October 1, 2002 Name Social Security Number This exam consists of two parts. Part I is multiple choice. Each of these 25 questions is
MBB 407/511 Molecular Biology and Biochemistry First Examination - October 1, 2002 Name Social Security Number This exam consists of two parts. Part I is multiple choice. Each of these 25 questions is
Supplemental Information. An Atlas of b-glucuronidases. in the Human Intestinal Microbiome
 Structure, Volume 2 Supplemental Information An Atlas of b-glucuronidases in the Human Intestinal Microbiome Rebecca M. Pollet, Emma H. D'Agostino, William G. Walton, Yongmei Xu, Michael S. Little, Kristen
Structure, Volume 2 Supplemental Information An Atlas of b-glucuronidases in the Human Intestinal Microbiome Rebecca M. Pollet, Emma H. D'Agostino, William G. Walton, Yongmei Xu, Michael S. Little, Kristen
Reviewers' Comments: Reviewer #2: Remarks to the Author:
 Reviewers' Comments: Reviewer #1: Remarks to the Author: This manuscript presents cryo-em studies of a giant virus Paramecium bursaria chlorella virus 1 (PBCV-1), which is the first cryo-em structure of
Reviewers' Comments: Reviewer #1: Remarks to the Author: This manuscript presents cryo-em studies of a giant virus Paramecium bursaria chlorella virus 1 (PBCV-1), which is the first cryo-em structure of
The mechanism of patellamide macrocyclization revealed by study of the Prochloron sp PatG macrocyclase domain
 The mechanism of patellamide macrocyclization revealed by study of the Prochloron sp PatG macrocyclase domain Jesko Koehnke 1,2, Andrew Bent 1,2, Wael E. Houssen 2,3,4, David Zollman 1, Falk Morawitz 1,
The mechanism of patellamide macrocyclization revealed by study of the Prochloron sp PatG macrocyclase domain Jesko Koehnke 1,2, Andrew Bent 1,2, Wael E. Houssen 2,3,4, David Zollman 1, Falk Morawitz 1,
Tivadar Orban, Beata Jastrzebska, Sayan Gupta, Benlian Wang, Masaru Miyagi, Mark R. Chance, and Krzysztof Palczewski
 Structure, Volume Supplemental Information Conformational Dynamics of Activation for the Pentameric Complex of Dimeric G Protein-Coupled Receptor and Heterotrimeric G Protein Tivadar Orban, Beata Jastrzebska,
Structure, Volume Supplemental Information Conformational Dynamics of Activation for the Pentameric Complex of Dimeric G Protein-Coupled Receptor and Heterotrimeric G Protein Tivadar Orban, Beata Jastrzebska,
SUPPORTING INFORMATION FOR. A Computational Approach to Enzyme Design: Using Docking and MM- GBSA Scoring
 SUPPRTING INFRMATIN FR A Computational Approach to Enzyme Design: Predicting ω- Aminotransferase Catalytic Activity Using Docking and MM- GBSA Scoring Sarah Sirin, 1 Rajesh Kumar, 2 Carlos Martinez, 2
SUPPRTING INFRMATIN FR A Computational Approach to Enzyme Design: Predicting ω- Aminotransferase Catalytic Activity Using Docking and MM- GBSA Scoring Sarah Sirin, 1 Rajesh Kumar, 2 Carlos Martinez, 2
SUPPLEMENTARY INFORMATION
 Supplementary Discussion The HGGG sequence motif forms an oxyanion hole in HLSs. In the case of GID1, the third Gly residue of the sequence motif is replaced with Ser (Ser123 in OsGID1), i.e., the sequence
Supplementary Discussion The HGGG sequence motif forms an oxyanion hole in HLSs. In the case of GID1, the third Gly residue of the sequence motif is replaced with Ser (Ser123 in OsGID1), i.e., the sequence
Supplementary Figure-1. SDS PAGE analysis of purified designed carbonic anhydrase enzymes. M1-M4 shown in lanes 1-4, respectively, with molecular
 Supplementary Figure-1. SDS PAGE analysis of purified designed carbonic anhydrase enzymes. M1-M4 shown in lanes 1-4, respectively, with molecular weight markers (M). Supplementary Figure-2. Overlay of
Supplementary Figure-1. SDS PAGE analysis of purified designed carbonic anhydrase enzymes. M1-M4 shown in lanes 1-4, respectively, with molecular weight markers (M). Supplementary Figure-2. Overlay of
Model for the activation of flaviviral fusion proteins 6
 Model for the activation of flaviviral fusion proteins 6 6.1 Introduction The envelope protein E of a flavivirus is composed of a soluble ectodomain and a C-terminal, membrane-associated domain, the so-called
Model for the activation of flaviviral fusion proteins 6 6.1 Introduction The envelope protein E of a flavivirus is composed of a soluble ectodomain and a C-terminal, membrane-associated domain, the so-called
172R 172K TAM-2/172R TAM-2/172K. AZT concentration [nm] AZT concentration [nm] MgCl 2 2.5K 2.5K 5K 2.5K 5K 2.5K K 5K 2.5K 5K 2.5K 50 2.
![172R 172K TAM-2/172R TAM-2/172K. AZT concentration [nm] AZT concentration [nm] MgCl 2 2.5K 2.5K 5K 2.5K 5K 2.5K K 5K 2.5K 5K 2.5K 50 2.](/thumbs/82/85138523.jpg "172R 172K TAM-2/172R TAM-2/172K. AZT concentration [nm] AZT concentration [nm] MgCl 2 2.5K 2.5K 5K 2.5K 5K 2.5K K 5K 2.5K 5K 2.5K 50 2.") 5 5 5 5 A MgCl 2 172R 172K TAM-2/172R TAM-2/172K AZT concentration [nm] B 172R 172K TAM-2/172R TAM-2/172K AZT concentration [nm] ATP + ATP - Supplemental Figure 1. Primer extension of HIV-1 RT polymorphisms
5 5 5 5 A MgCl 2 172R 172K TAM-2/172R TAM-2/172K AZT concentration [nm] B 172R 172K TAM-2/172R TAM-2/172K AZT concentration [nm] ATP + ATP - Supplemental Figure 1. Primer extension of HIV-1 RT polymorphisms
Lateral opening in the intact β-barrel assembly machinery captured by cryo-em
 Lateral opening in the intact β-barrel assembly machinery captured by cryo-em Iadanza et al. Supplementary Figure 1. Purification of the full BamABCDE complex and complexes containing BamA mutants. (a,d,g)
Lateral opening in the intact β-barrel assembly machinery captured by cryo-em Iadanza et al. Supplementary Figure 1. Purification of the full BamABCDE complex and complexes containing BamA mutants. (a,d,g)
Amino Acids. Review I: Protein Structure. Amino Acids: Structures. Amino Acids (contd.) Rajan Munshi
 Rajan Munshi") Review I: Protein Structure Rajan Munshi BBSI @ Pitt 2005 Department of Computational Biology University of Pittsburgh School of Medicine May 24, 2005 Amino Acids Building blocks of proteins 20 amino acids
Review I: Protein Structure Rajan Munshi BBSI @ Pitt 2005 Department of Computational Biology University of Pittsburgh School of Medicine May 24, 2005 Amino Acids Building blocks of proteins 20 amino acids
HOMEWORK II and Swiss-PDB Viewer Tutorial DUE 9/26/03 62 points total. The ph at which a peptide has no net charge is its isoelectric point.
 BIOCHEMISTRY I HOMEWORK II and Swiss-PDB Viewer Tutorial DUE 9/26/03 62 points total 1). 8 points total T or F (2 points each; if false, briefly state why it is false) The ph at which a peptide has no
BIOCHEMISTRY I HOMEWORK II and Swiss-PDB Viewer Tutorial DUE 9/26/03 62 points total 1). 8 points total T or F (2 points each; if false, briefly state why it is false) The ph at which a peptide has no
BIO 311C Spring Lecture 15 Friday 26 Feb. 1
 BIO 311C Spring 2010 Lecture 15 Friday 26 Feb. 1 Illustration of a Polypeptide amino acids peptide bonds Review Polypeptide (chain) See textbook, Fig 5.21, p. 82 for a more clear illustration Folding and
BIO 311C Spring 2010 Lecture 15 Friday 26 Feb. 1 Illustration of a Polypeptide amino acids peptide bonds Review Polypeptide (chain) See textbook, Fig 5.21, p. 82 for a more clear illustration Folding and
Insights into the Giardia intestinalis Enolase and Human Plasminogen interaction
 Electronic Supplementary Material (ESI) for Molecular BioSystems. This journal is The Royal Society of Chemistry 2017 Supplementary Information Insights into the Giardia intestinalis Enolase and Human
Electronic Supplementary Material (ESI) for Molecular BioSystems. This journal is The Royal Society of Chemistry 2017 Supplementary Information Insights into the Giardia intestinalis Enolase and Human
Supplementary Figure 1 Overall structure of the transmembrane pore of FraC. a, Crystal packing of the pore along the y-z, b, the z-x, and c, the y-x
 Supplementary Figure 1 Overall structure of the transmembrane pore of FraC. a, Crystal packing of the pore along the y-z, b, the z-x, and c, the y-x planes. The N-terminal and the β-core regions are depicted
Supplementary Figure 1 Overall structure of the transmembrane pore of FraC. a, Crystal packing of the pore along the y-z, b, the z-x, and c, the y-x planes. The N-terminal and the β-core regions are depicted
The role of Ca² + ions in the complex assembling of protein Z and Z-dependent protease inhibitor: A structure and dynamics investigation
 www.bioinformation.net Hypothesis Volume 8(9) The role of Ca² + ions in the complex assembling of protein Z and Z-dependent protease inhibitor: A structure and dynamics investigation Zahra Karimi 1 *,
www.bioinformation.net Hypothesis Volume 8(9) The role of Ca² + ions in the complex assembling of protein Z and Z-dependent protease inhibitor: A structure and dynamics investigation Zahra Karimi 1 *,
Supplementary Information
 Supplementary Information Archaeal Elp3 catalyzes trna wobble uridine modification at C5 via a radical mechanism Kiruthika Selvadurai, Pei Wang, Joseph Seimetz & Raven H Huang* Department of Biochemistry,
Supplementary Information Archaeal Elp3 catalyzes trna wobble uridine modification at C5 via a radical mechanism Kiruthika Selvadurai, Pei Wang, Joseph Seimetz & Raven H Huang* Department of Biochemistry,
Supplementary Figure 1. Heavy chain sequences of 2G1 and 8M2 aligned with V H 1-69
 Supplementary Figure 1. Heavy chain sequences of 2G1 and 8M2 aligned with V H 1-69 germline gene sequence. 2G1 and 8M2 acquired 14 and 18 mutations from the germline gene sequence, respectively. In 2G1,
Supplementary Figure 1. Heavy chain sequences of 2G1 and 8M2 aligned with V H 1-69 germline gene sequence. 2G1 and 8M2 acquired 14 and 18 mutations from the germline gene sequence, respectively. In 2G1,
Supplementary materials
 Supplementary materials Chemical library from ChemBridge 50,240 structurally diverse small molecule compounds dissolved in DMSO Hits Controls: No virus added μ Primary screening at 20 g/ml of compounds
Supplementary materials Chemical library from ChemBridge 50,240 structurally diverse small molecule compounds dissolved in DMSO Hits Controls: No virus added μ Primary screening at 20 g/ml of compounds
Danish Research Institute of Translational Neuroscience DANDRITE, Nordic-EMBL Partnership
 Supplementary Information for Tuning of the Na,K-ATPase by the beta subunit Florian Hilbers 1,2,3, Wojciech Kopec 4, Toke Jost Isaksen 3,5, Thomas Hellesøe Holm 3,5, Karin Lykke- Hartmann 3,5,6, Poul Nissen
Supplementary Information for Tuning of the Na,K-ATPase by the beta subunit Florian Hilbers 1,2,3, Wojciech Kopec 4, Toke Jost Isaksen 3,5, Thomas Hellesøe Holm 3,5, Karin Lykke- Hartmann 3,5,6, Poul Nissen
<Supplemental information>
 The Structural Basis of Endosomal Anchoring of KIF16B Kinesin Nichole R. Blatner, Michael I. Wilson, Cai Lei, Wanjin Hong, Diana Murray, Roger L. Williams, and Wonhwa Cho Protein
The Structural Basis of Endosomal Anchoring of KIF16B Kinesin Nichole R. Blatner, Michael I. Wilson, Cai Lei, Wanjin Hong, Diana Murray, Roger L. Williams, and Wonhwa Cho Protein
Supplementary Information A Hydrophobic Barrier Deep Within the Inner Pore of the TWIK-1 K2P Potassium Channel Aryal et al.
 Supplementary Information A Hydrophobic Barrier Deep Within the Inner Pore of the TWIK-1 K2P Potassium Channel Aryal et al. Supplementary Figure 1 TWIK-1 stability during MD simulations in a phospholipid
Supplementary Information A Hydrophobic Barrier Deep Within the Inner Pore of the TWIK-1 K2P Potassium Channel Aryal et al. Supplementary Figure 1 TWIK-1 stability during MD simulations in a phospholipid
Molecular Graphics Perspective of Protein Structure and Function
 Molecular Graphics Perspective of Protein Structure and Function VMD Highlights > 20,000 registered Users Platforms: Unix (16 builds) Windows MacOS X Display of large biomolecules and simulation trajectories
Molecular Graphics Perspective of Protein Structure and Function VMD Highlights > 20,000 registered Users Platforms: Unix (16 builds) Windows MacOS X Display of large biomolecules and simulation trajectories
KDM2A. Reactions. containing. Reactions
 Supplementary Figure 1 KDM2A catalyses only lysine demethylation.. MALDI-TOF MS of demethylation of the shown variant histone peptides as catalysed by recombinant KDM2A. Reactions containing enzyme are
Supplementary Figure 1 KDM2A catalyses only lysine demethylation.. MALDI-TOF MS of demethylation of the shown variant histone peptides as catalysed by recombinant KDM2A. Reactions containing enzyme are
BIRKBECK COLLEGE (University of London)
") BIRKBECK COLLEGE (University of London) SCHOOL OF BIOLOGICAL SCIENCES M.Sc. EXAMINATION FOR INTERNAL STUDENTS ON: Postgraduate Certificate in Principles of Protein Structure MSc Structural Molecular Biology
BIRKBECK COLLEGE (University of London) SCHOOL OF BIOLOGICAL SCIENCES M.Sc. EXAMINATION FOR INTERNAL STUDENTS ON: Postgraduate Certificate in Principles of Protein Structure MSc Structural Molecular Biology
Supplementary Figure 1. Sample preparation schematic. First (Stage I), square islands of MoO 3 are prepared by either photolithography followed by
, square islands of MoO 3 are prepared by either photolithography followed by") Supplementary Figure 1. Sample preparation schematic. First (Stage I), square islands of MoO 3 are prepared by either photolithography followed by thermal evaporation and liftoff or by a process where
Supplementary Figure 1. Sample preparation schematic. First (Stage I), square islands of MoO 3 are prepared by either photolithography followed by thermal evaporation and liftoff or by a process where
The N-terminal loop of IRAK-4 death domain regulates ordered assembly of the Myddosome signalling scaffold
 The N-terminal loop of IRAK-4 death domain regulates ordered assembly of the Myddosome signalling scaffold Anthony C.G. Dossang * 1, Precious G. Motshwene * 1, Yang Yang* 1, Martyn F. Symmons*, Clare E.
The N-terminal loop of IRAK-4 death domain regulates ordered assembly of the Myddosome signalling scaffold Anthony C.G. Dossang * 1, Precious G. Motshwene * 1, Yang Yang* 1, Martyn F. Symmons*, Clare E.
Supplementary Information
 Supplementary Information Structural Basis of Sialidase in Complex with Geranylated Flavonoids as Potent Natural Inhibitors Youngjin Lee, a,b Young Bae Ryu, d Hyung-Seop Youn, a,b Jung Keun Cho, e Young
Supplementary Information Structural Basis of Sialidase in Complex with Geranylated Flavonoids as Potent Natural Inhibitors Youngjin Lee, a,b Young Bae Ryu, d Hyung-Seop Youn, a,b Jung Keun Cho, e Young
Biological Mass Spectrometry. April 30, 2014
 Biological Mass Spectrometry April 30, 2014 Mass Spectrometry Has become the method of choice for precise protein and nucleic acid mass determination in a very wide mass range peptide and nucleotide sequencing
Biological Mass Spectrometry April 30, 2014 Mass Spectrometry Has become the method of choice for precise protein and nucleic acid mass determination in a very wide mass range peptide and nucleotide sequencing
Protein Primer, Lumry I, Chapter 10, Enzyme structure, middle
 Protein Primer, Lumry I, Chapter 10, Enzyme structure, 5-15-03 10-1 Chapter 10. Enzyme structure A major feature of the trypsin family of proteases is the palindromic pattern taken by the B factors of
Protein Primer, Lumry I, Chapter 10, Enzyme structure, 5-15-03 10-1 Chapter 10. Enzyme structure A major feature of the trypsin family of proteases is the palindromic pattern taken by the B factors of
Supplementary Figures
 Supplementary Figures Supplementary Figure 1. (a) Uncropped version of Fig. 2a. RM indicates that the translation was done in the absence of rough mcirosomes. (b) LepB construct containing the GGPG-L6RL6-
Supplementary Figures Supplementary Figure 1. (a) Uncropped version of Fig. 2a. RM indicates that the translation was done in the absence of rough mcirosomes. (b) LepB construct containing the GGPG-L6RL6-
Electron Microscopy Imaging Reveals Unique Higher Order Structures of Adalimumab-TNFα and Infliximab-TNFα Complexes
 Electron Microscopy Imaging Reveals Unique Higher Order Structures of Adalimumab-TNFα and Infliximab-TNFα Complexes Siew Leong Chan Protein Analytics, AbbVie Bioresearch Center Worcester MA Acknowledgement
Electron Microscopy Imaging Reveals Unique Higher Order Structures of Adalimumab-TNFα and Infliximab-TNFα Complexes Siew Leong Chan Protein Analytics, AbbVie Bioresearch Center Worcester MA Acknowledgement
Cbl ubiquitin ligase: Lord of the RINGs
 Cbl ubiquitin ligase: Lord of the RINGs Not just quite interesting - really interesting! A cell must be able to degrade proteins when their activity is no longer required. Many eukaryotic proteins are
Cbl ubiquitin ligase: Lord of the RINGs Not just quite interesting - really interesting! A cell must be able to degrade proteins when their activity is no longer required. Many eukaryotic proteins are
1. Arresting cancer by energy starvation
 1. Arresting cancer by energy starvation Cancer, a top killer, may be stopped from its deadly progression with the latest discovery by NUS researchers. They have found how a compound that is undergoing
1. Arresting cancer by energy starvation Cancer, a top killer, may be stopped from its deadly progression with the latest discovery by NUS researchers. They have found how a compound that is undergoing
Adaptable Lipid Matrix Promotes Protein Protein Association in Membranes
 Supporting information Adaptable Lipid Matrix Promotes Protein Protein Association in Membranes Andrey S. Kuznetsov, Anton A. Polyansky,, Markus Fleck, Pavel E. Volynsky, and Roman G. Efremov *,, M. M.
Supporting information Adaptable Lipid Matrix Promotes Protein Protein Association in Membranes Andrey S. Kuznetsov, Anton A. Polyansky,, Markus Fleck, Pavel E. Volynsky, and Roman G. Efremov *,, M. M.
Proteins? Protein function. Protein folding. Protein folding diseases. Protein interactions. Macromolecular assemblies. The end product of Genes
 Proteins? Protein function Protein folding Protein folding diseases Protein interactions Macromolecular assemblies The end product of Genes Protein Unfolding DOD Acid Catalysis DOD HDOD + N H N D C N C
Proteins? Protein function Protein folding Protein folding diseases Protein interactions Macromolecular assemblies The end product of Genes Protein Unfolding DOD Acid Catalysis DOD HDOD + N H N D C N C
The antiparasitic drug ivermectin is a novel FXR ligand that regulates metabolism
 Supplementary Information The antiparasitic drug ivermectin is a novel FXR ligand that regulates metabolism Address correspondence to Yong Li (yongli@xmu.edu.cn, Tel: 86-592-218151) GW464 CDCA Supplementary
Supplementary Information The antiparasitic drug ivermectin is a novel FXR ligand that regulates metabolism Address correspondence to Yong Li (yongli@xmu.edu.cn, Tel: 86-592-218151) GW464 CDCA Supplementary
Chapter 3. Structure of Enzymes. Enzyme Engineering
 Chapter 3. Structure of Enzymes Enzyme Engineering 3.1 Introduction With purified protein, Determining M r of the protein Determining composition of amino acids and the primary structure Determining the
Chapter 3. Structure of Enzymes Enzyme Engineering 3.1 Introduction With purified protein, Determining M r of the protein Determining composition of amino acids and the primary structure Determining the
Interactions of Polyethylenimines with Zwitterionic and. Anionic Lipid Membranes
 Interactions of Polyethylenimines with Zwitterionic and Anionic Lipid Membranes Urszula Kwolek, Dorota Jamróz, Małgorzata Janiczek, Maria Nowakowska, Paweł Wydro, Mariusz Kepczynski Faculty of Chemistry,
Interactions of Polyethylenimines with Zwitterionic and Anionic Lipid Membranes Urszula Kwolek, Dorota Jamróz, Małgorzata Janiczek, Maria Nowakowska, Paweł Wydro, Mariusz Kepczynski Faculty of Chemistry,
Chapter VI. Increased affinity in the complex of yeast cytochrome c and cytochrome c peroxidase
 Chapter VI Increased affinity in the complex of yeast cytochrome c and cytochrome c peroxidase Chapter VI Abstract We report the study of T12A Cyt c CcP complex using isothermal titration calorimetry (ITC),
Chapter VI Increased affinity in the complex of yeast cytochrome c and cytochrome c peroxidase Chapter VI Abstract We report the study of T12A Cyt c CcP complex using isothermal titration calorimetry (ITC),