For Publication in Journal of Virology
|
|
|
- Isabella Harvey
- 6 years ago
- Views:
Transcription
1 JVI Accepts, published online ahead of print on 23 August 2006 J. Virol. doi: /jvi Copyright 2006, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved. For Publication in Journal of Virology Dated Latency Associated Nuclear Antigen of KSHV Recruits Uracil DNA Glycosylase 2 at the Terminal Repeats and is Important for Latent Persistence of the Virus. Subhash C. Verma, Bharat G. Bajaj, Qiliang Cai, Huaxin Si, Todd Seelhammer and Erle S. Robertson* 201E Johnson Pavilion, 3610 Hamilton Walk, Department of Microbiology and Tumor Virology Program of the Abramson Comprehensive Cancer Center University of Pennsylvania, School of Medicine Philadelphia PA Corresponding author: Tel: Fax: E mail address: erle@mail.med.upenn.edu * Corresponding author 1
2 Abstract: Latency Associated Nuclear Antigen (LANA) of KSHV is expressed in all forms of KSHV mediated tumors and is important for TR mediated replication and persistence of the virus. LANA does not exhibit any enzymatic activity by itself but is critical for replication and maintenance of the viral genome. To identify LANA binding proteins, we used the identified LANA Binding Sequence-1 (LBS-1) DNA affinity column and have determined the identity of a number of proteins associated with LANA. One of the identified proteins was Uracil DNA Glycosylase 2 (UNG2). UNG2 is important for removing uracil residues yielded after either misincorporation of dutp during replication or deamination of cytosine. Specificity of LANA and UNG2 interaction was confirmed by using a scrambled DNA sequence affinity column. Interaction of LANA and UNG2 was further confirmed by in-vitro binding and coimmunoprecipitation assays. Co-localization of these proteins was also detected in PELs as well as in co-transfected KSHV negative cell line. UNG2 binds to the carboxyl terminus of LANA and retains its enzymatic activity in the complex. However, no major effect on TR mediated DNA replication was observed using a UNG2 deficient (UNG-/-) cell line. Infection of UNG-/- and wild type MEFs with KSHV did not reveal any difference however UNG-/- produced a significantly reduced number of virion particles after induction. Interestingly, depletion of UNG2 in PELs using shrna reduced the viral genome copies and produced infection deficient virus. Keywords: KSHV, LANA, UNG2, Latent replication, post-replicative repair. 2
3 Introduction: Kaposi s sarcoma associated herpesvirus (KSHV) also designated as human herpes virus 8 (HHV8) is the biologic agent of Kaposi sarcoma (KS), Primary Effusion Lymphoma (PEL) and Multicentric Castleman s Disease (MCD) (12-14, 19, 65, 73). KSHV persists indefinitely in these infected cells as an episomes with the expression of limited number of genes (20, 35, 39, 66, 75). Among these genes, the Latency Associated Nuclear Antigen (LANA) express from a polycistronic mrna and is detected in all forms of KSHV tumors (18, 36). Cells infected with KSHV shows anchorage independent growth and increased telomerase activity suggesting that KSHV induces cell immortalization (22, 37, 70). Also, KSHV infection induces chromosomal instability, an important event during tumorigenesis (52). These functions were found to be regulated by the major latent protein, LANA (37, 64, 70). Additionally, LANA can also down regulate apoptotic pathways, p53 and prb and induces cell immortalization in combination with Hras (23, 60). LANA also modulates the distribution of GSK-3β, a negative regulator of Wnt signaling pathway, in a cell cycle dependent manner and induces cells to enter into S-phase (10, 24). LANA, which was initially detected by the sera from a KSHV infected patient in an immunofluorescence assay, is a large nuclear protein and typically detected in a punctate nuclear pattern in KSHV infected cells (34, 46, 61). KSHV genome was detected at the sites of LANA on chromosome spreads of KSHV infected cells suggesting a role for LANA in KSHV genome tethering (4, 16). Later studies mapped the domains of LANA important for tethering to the host chromosomes (5, 6, 17). LANA associates with human chromatin through the amino terminal domain and remains attached during all the phases of cell cycle (7, 54, 63). LANA tethers KSHV genome to the host chromosomes through binding at the terminal repeats (TRs) of KSHV 3
4 genome (62). The terminal repeats (TRs) are 801 bp long, high GC region of the KSHV genome and are present as multi-copy tandem repeats (44, 62). Each copy of TR contains two LANA binding sequence (LBS1/2, high/low affinity) separated by 22bp DNA sequence (25). LANA binds to LBS through amino acids of the carboxyl terminal domain (17, 38). Deletion mutants of this region concluded that the amino acids are likely to be the DNA contacting residues of LANA (38). TR also supports replication of a plasmid in a LANA dependent manner (28, 32). A single copy of the TR element is able to support replication but the mechanism of replication is not fully understood (25, 28, 71). Sequence analysis and deletion mutation of TR mapped the minimal sequence essential for replication to a 29- to 32-bp-long GC-rich sequence upstream to the LANA binding sequence 1 (LBS1) and LANA binding sequence 2 (LBS2) (33). Although LANA is critical for replication of TR containing plasmids but it does not have any enzymatic activity required for replication therefore suggests for a recruitment of necessary cellular proteins for replication at TR. The mechanism of replication mediated by the TR is now beginning to be resolved and the involvement of a number of cellular proteins is being investigated (67, 71). Among the proteins identified so far includes Human Origin Recognition Complexes (hsorcs) which interacts with LANA at the TR (47, 71). EBNA1, functional homolog of LANA also interacts with ORCs and is essential for the replication of EBV genome (reviewed in 8). Besides replication LANA is critical for the maintenance of KSHV episomal DNA. LANA depleted cells, either by using recombinant virus (Bac36 LANA) or shrna for LANA, failed to maintain KSHV episomes for long term suggesting its role in persistence and segregation (26, 74). Therefore, to understand the role of LANA in the process of replication and segregation, we wanted to identify LANA interacting proteins when LANA is bound to its 4
5 cognate sequence. Analysis of LANA interacting proteins revealed a number of cellular proteins important for episomal persistence. Uracil DNA Glycosylase 2 was identified as one of the LANA interacting proteins. Uracil DNA Glycosylase 2 (UNG2) is encoded by the UDG gene which spans approximately 13.8kb and is located at chromosome 12q24.1 (42). This gene encodes for the mitochondrial Uracil DNA Glycosylase (UNG1) as well as Nuclear Uracil DNA Glycosylase (UNG2), both share identical catalytic domain but differ at N-terminal amino acids (49). The major role of UNG2 is the removal of dutps from nuclear DNA yielded after either misincorporation during replication process or due to deamination of cytosine residues (43). UNG2 is located in the nucleoplasm and at replication foci. The mechanism of its translocation and association with replication foci is not well understood (41). However, it is proposed that UNG2 is transported along with proliferating cell nuclear antigen (PCNA), similar to the DNA ligase I and DNA Pol iota, part of post-replicative repair enzyme (41). UNG2 also associates with PCNA through its PCNA interacting motif (Q 4 xxlxxff 11 ) (41). The N-terminal domain of UNG2 also interacts with replication protein A (RP-A) at the replication foci (51). Interaction of UNG2 with PCNA and RP-A is important for base excision repair (BER) in the replication foci (43, 51). In this report we demonstrate that LANA binds to UNG2 through its carboxyl terminal domain adjacent to the DNA binding domain. We also show the interaction of these two proteins in immunofluorescence assay. The LANA-UNG2 complex possessed glycosylase activity therefore suggesting the involvement of UNG2 in the removal of any misincorporated uracil residues during replication process. KSHV infected cells with depleted UNG2 levels using 5
6 shrna showed reduced copy number of the viral genome thus suggested that UNG2 is an important protein for the persistence of the viral genome. 6
7 Materials and Methods: Cells, Plasmids and antibodies: BC-3 and BCBL-1, KSHV positive primary effusion lymphoma (PELs) cell lines, BJAB and DG75, KSHV negative cell lines were cultured in RPMI supplemented with 10% fetal bovine serum, 2mM L-glutamine and penicillin-streptomycin (5U/ml and 5µg/ml, respectively). Human Embryonic Kidney (HEK) 293 and 293T cells were cultured in Dulbecco s modified Eagle medium supplemented with 10% fetal bovine serum, 2mM L-glutamine, penicillinstreptomycin (5U/ml and 5µg/ml, respectively). Primary MEF cells Ung / and Ung +/+ (F11.1) were gift from Prof. Hans E. Krokan (Department of Cancer Research and Molecular Medicine, Norwegian University of Science and Technology, Medical Faculty, N-7489 Trondheim, Norway) and were established from transformed clones arising spontaneously after repeated passage in culture (2). MEFs were cultured in Dulbecco's modified Eagle's medium high glucose with glutamine/ham's F12 with sodium pyruvate (1/1), 10% fetal bovine serum, 1x nonessential amino acid solution and penicillin/streptomycin. pbspurotr containing entire (801bp) TR unit was generated by cloning TR at the NotI site of pbs SKII+ (Stratagene Inc. La Jolla, CA) described previously (71). Expression vectors of LANA full length and truncated mutants tagged with myc epitope at its carboxyl-terminus are described previously (70). DsRed-LANA is described previously (64). UNG2 fused to GFP, pung2egfp and rabbit UNG2 specific antibody (PU101) was a kind gift from Prof. Hans E. Krokan (Department of Cancer Research and Molecular Medicine, Norwegian University of Science and Technology, Medical Faculty, N Trondheim, Norway). cdna encoding the UNG2 ORF (open reading frame) was PCR amplified using pung2egfp as template and cloned into pgex2tk and pcdna3.1ha to make GST fusion protein and HA tagged protein, respectively. The PCR amplified product was 7
8 cloned into restriction enzyme sites BamHI and EcoRI of pcdna3.1ha and pgex2tk, respectively. Rabbit anti-lana polyclonal antibody was a kind gift from Prof. Bala Chandran (Rosalind Franklin University of Medicine and Science, North Chicago, Illinois). RTA was detected using mouse monoclonal antibody described previously (45). DNA affinity column: Preparation of DNA affinity column is described previously (71). Briefly, oligo containing LANA binding sequence (17bp, italics and underlined) containing additional 3 nucleotide on 5 and 6 nucleotide at 3 (italics) (GATCCGCCTCCCGCCCGGGCATGGGGCCGCGG) with overlapping BamHI sites (bold) was synthesized and annealed with its complementary strand. Multimerized ds DNA was ligated to the CNBr activated Sepharose beads and packed onto a Bio-Rad column. Nuclear extracts (NE) from BC-3 (KSHV positive) and BJAB (KSHV negative) cells were prepared as described earlier and bound to the LBS affinity column as described previously (71). The eluted fractions were aliquoted and frozen at -80 o C. Control DNA column was prepared by synthesizing similar length DNA sequence with scrambled LANA binding sites (5 -GATCCGCCATCGTAGATCTGTACTGTACGCGG-3 ). Similar amount of total nuclear extract was bound to the DNA columns with multimerized LBS and scrambled LBS. Proteins eluted after extensive washing of the column were resolved on SDS-PAGE and subjected for either western blot or sequencing of the band after gel excision. MALDI-TOFF and LCQ (MS) analysis: Distinct protein bands from KSHV positive cells nuclear extracts (BC-3) were excised from the Coomassie G-250 stained gel (Fig. 1A). These bands were subjected for MALDI-TOP 8
9 and LCQ proteomic analysis at the proteomics core facility at the University Of Pennsylvania School of Medicine. Proteins for each band with LCQ score over 20 were reported. Western Blot Analysis: An aliquot of fraction eluted with 500mM NaCl containing elution buffer was resolved on SDS-PAGE followed by detection with UNG2 specific antibodies (rabbit polyclonal antibody, PU101). The signals were detected using Alexa Flour 680 and 800 (Molecular Probes and Rockland, Carlsbad, CA and Gilbertsville, PA, respectively) conjugated antibodies with Odyssey Infra-Red scanning technology (LiCor, Linclon, NE). Myc and HA tagged proteins were detected using 9E10 and 12CA5 hybridoma, respectively. In-vitro binding of UNG2 to LANA: Full length (1-1162aa) or different truncation mutants of LANA was in-vitro translated using coupled in-vitro transcription/translation system (TNT) (Promega Inc. Madison, WI) according to the manufacturer s instructions in the presence of [ 35 S] Methionine/Cysteine (Perkin Elmer Inc., Boston, MA. pgex-ung2 expressing GST-fused UNG2 protein was expressed in E. coli as described previously (71). Approximately 10µg of GST-UNG2 protein was used for binding with in-vitro translated LANA and its derivatives. In-vitro translated products were precleared with Glutathione Sepharose beads in binding buffer (1X phosphate buffered saline, 0.1% NP-40, 0.5mM dithiothreitol [DTT], 10% glycerol, 1mM PMSF, 2µg of aprotinin per ml) for 30min. Precleared LANA was then incubated with either GST or GST-UNG2 fusion proteins in binding buffer. Binding was performed overnight at 4 o C with constant rotation followed by collecting the beads through centrifugation. The beads were washed 3 times with 1ml of binding buffer followed by resuspending them in SDS-lysis buffer and resolving them on SDS-PAGE. 9
10 The bound fraction was analyzed after drying the gel and exposing to phosphor Imager plate (Molecular Dynamics, Inc.). Myc tagged LANA expressed in 293T cells were also subjected for binding to GST- UNG2. 293T cells expressing myc tagged LANA were lysed in RIPA buffer (50 mm Tris, ph 7.5, 150 mm NaCl, 0.5% NP-40, 1 mm EDTA, ph 8.0) with protease inhibitors (1 mm PMSF, 10 µg/ml pepstatin, 10 µg/ml leupeptin, and 10 µg/ml aprotinin) and centrifuged at 15,000rpm, 4 o C to remove cell debris. GST-UNG2 fusion protein was added to cell lysate and subjected for overnight binding in RIPA buffer. Bound complex was washed five times to remove loosely bound proteins and resolved on SDS-PAGE. LANA was detected using anti-myc in western blot. Immunolocalization of LANA and UNG2: HEK 293 cells were co-transfected with pa3m LANA and pung2egfp. BC-3 cells were only transfected with pung2egfp and endogenous LANA was detected using rabbit anti- LANA serum. 24h post-transfection cells were spread on glass slides and followed by fixing them in acetone/methanol (1:1) for 10 minutes at -20 o C. Slides were air dried and incubated with 20% normal goat serum in 1XPBS to block the non-specific binding sites. LANA was detected using rabbit anti-lana polyclonal serum at room temperature in a humidified chamber followed by washing three times for 5 minutes in PBS. LANA signal was detected using Alexa Flour 594 (Molecular Probes, Carlsbad, CA). UNG2 was visualized by fluorescence from the EGFP fusion protein. Slides were then washed four times with 1XPBS, mounted with Paramount G and visualized using an Olympus confocal laser scanning microscope. Detection of UNG activity: To detect uracil DNA glycosylase activity of UNG2 when it is bound to LANA, we used a previously described (11) PCR based assay as depicted in Fig. 4A. The use of dutp instead of dttp in a PCR amplification results in an amplicon which contains uracil residues. Incubation of 10
11 the amplicon with UNG2 results in the excision of uracil residues from du containing PCR products. Heat treatment and alkaline ph conditions results in the degradation of the abasic polynucleotide of the du-containing PCR product which is further blocked from subsequent PCR re-amplification. Any DNA fragment can be used regardless of its origin for UNG assay, the only important criteria is that it should be amplifiable in PCR. In this assay we used the KSHV K1 gene which was used as the DNA template and amplified with primers S-K1 (5'- ATGTTCCTGTATGTTGTCTGCAG-3 ) and AS-K1 (5'- TCAGTACCAATCCACTGGTTGC- 3 ) either with dutp (deoxy uridine triphosphate) or with dttp (deoxy thymidine triphosphate) in the mixtures of deoxynucleoside triphosphates leading to a PCR product of 870 bp, designated the (du) DNA template or (dt) DNA template, respectively. Subsequent PCR amplification using du DNA or dt DNA templates treated with UNG2-LANA was then performed up to 30 amplification cycles with identical conditions. Briefly, 1 µl of templates DNA either du or dt was assayed in a final volume of 50 µl in the presence of 1.5 mm MgCl2, 200nM of each amplification primer, 200nM each deoxynucleoside triphosphate, and 1.5 U of Taq polymeras. PCR products were resolved on a 1% agarose gel. UNG2 was co-immunoprecipitated along with LANA from 293T cells as the source of UNG activity. LANA by itself was also used to test for any UNG activity after immunoprecipitation. Co-Immunoprecipitation of LANA and UNG2: Myc tagged LANA and HA tagged UNG2 were transfected either separately or together into 293T cells. Cells were harvested 36h post-transfection and lysed in RIPA buffer (50 mm Tris, ph 7.5, 150 mm NaCl, 0.5% NP-40, 1 mm EDTA, ph 8.0) with protease inhibitors (1 mm PMSF, 10 µg/ml pepstatin, 10 µg/ml leupeptin, and 10 µg/ml aprotinin). Lysates were 11
12 centrifuged to remove cell debris and precleared. Lysates were then incubated with anti-myc antibody (myc ascites) overnight at 4 o C with rotation followed by incubation with Protein A+G Sepharose beads at 4 o C for 1h. The resulting immunoprecipitates were collected by centrifugation at 2000Xg for 2 min at 4 C and the pellets were washed four times with 1 ml of ice-cold RIPA butter. Half of the immune complex was re-suspended in 30 µl of 2X SDS protein sample buffer (62.5 mm Tris, ph 6.8, 40 mm DTT, 2% SDS, 0.025% bromophenol blue, and 10% glycerol) for western blot. UNG2 was detected using anti-ha antibody followed reprobing the membrane with anti-myc antibody for LANA detection. Short-term replication assay: pbspuroa3 (3 copies of TR) was co-transfected into MEF either UNG-/- or UNG+/+ (UNG+/+) with LANA expression vector. 96h post-transfection DNA was extracted using modified Hirt s procedure described previously (31). Extracted DNA was digested with EcoRI (to linearize) or EcoRI+DpnI with sufficient enzyme overnight. Digested DNA was resolved on a 0.8% agarose gel and transferred to a nylon membrane. DpnI resistant copies of TR plasmid were detected by hybridization with a 32 P-labeled TR probe using Phosphor Imager plates (Molecular Dynamics Inc.). BrdU labeling and immnunoprecipitation of replicated DNA: BrdU (5-Bromo-2-deoxy Uridine, Thymidine analog) was administered 72h posttransfection in MEFs and pulsed for overnight to label the replicating DNA. DNA was extracted as described previously and digested with DpnI overnight followed by immunoprecipitation of BrdU labeled DNA from 90% of the digested product using anti-brdu mouse monoclonal antibody (Santa Cruz Biotechnology, Santa Cruz, CA) (29). The remaining 10% of the digested DNA was used as input. 1 µl of the above immunoprecipitated DNA was used for the detection 12
13 of BrdU incorporated and DpnI resistant copies of the pbspuroa3 after replication using primers described previously (71). Purification of KSHV virion: Virions were purified and virion proteins were fractioned and detected as described previously (45, 71). UNG2 was detected using rabbit anti-ung2 antibody in proteins from pelleted as well as gradient purified virion. Infection of MEF cells with KSHV virus and induction for virus production: MEF, UNG-/- and UNG+/+ cells grown at 50-70% confluency in 100mm diameter tissues culture dishes was infected with same amount of concentrated virus from identical number of BCBL-1 cells. Virus produced from approximately 20 Million BCBL-1 cells were used for infecting single 100mm culture dish in the presence of polybrene (Sigma, Saint Louis, MO). 2 days after infection cells were passaged and replated after thorough washing to remove any virion particles adhering to the cell surface. Efficiency of BCBL-1 infection in these infected MEFs was determined using immunofluorescence assay with rabbit anti-lana antibody. Equal number of MEFs, UNG-/- and UNG+/+ (infected with BCBL-1 virus) were plated in 100mm culture dish for induction to produce virus. Cells were induced with 20ng of TPA/ml and 1.5mM sodium butyrate (Sigma, Saint Louis, MO) for 5 days followed by collecting the supernatant and clearing by centrifugation at 2,000 rpm for 15 min to remove cells and cell debris. Then, the supernatant was filtered through 0.45-µm-pore-size filters and virions were pelleted at 20,000 rpm for 2 h. Virion DNA was isolated by disrupting the virus at 60 o C for 2h in lysis buffer (10mM Tris, ph 8.5/ 1mM EDTA, ph8.0/ 1% Sarcosyl/0.1mg/ml proteinase K). Lysates were phenol extracted followed by CHCl3:IAA extraction. Relative number of produced virion was quantified by real-time qpcr using K1 gene amplification described previously (72). 13
14 shrna Treatment and quantitation of KSHV episomal copies: UNG2-specific shrnas was described previously (56) and the sequence is as follows: sense, 5 -AUCGGCCAGAAGACGCUCUdTdT-3, corresponding to the first exon (nucleotides ) of the UNG genomic region. Firefly luciferase gene shrna was used as a control shrna (BD Clonetech, Mountain View, CA). shrna for UNG2 (5 - GATCCGATCGGCCAGAAGACGCTCTTTCAAGAGAAGAGCGTCTTCTGGCCGATCTTTTT TGATATCG-3 ) was cloned into psirenretrop (BD Clontech) after annealing with its complementary sequence. 15µg of either psirenretrop UNG2shRNA or psirenretrop Luc shrna was transfected into 10 million BC-3 and BCBL-1 cells. Transfected cells were selected with puromycin (3µg/ml) for three weeks. Puromycin resistant colonies were assayed for UNG2 protein in western blot assay using anti-ung2 specific antibodies. Total DNA from these selected colonies were extracted by lysing cells in 5mM EDTA, 1% sarcosyl, followed by proteinase K digestion (1). Relative numbers of KSHV episomal copies were calculated by real-time qpcr amplification of the K1 gene. 14
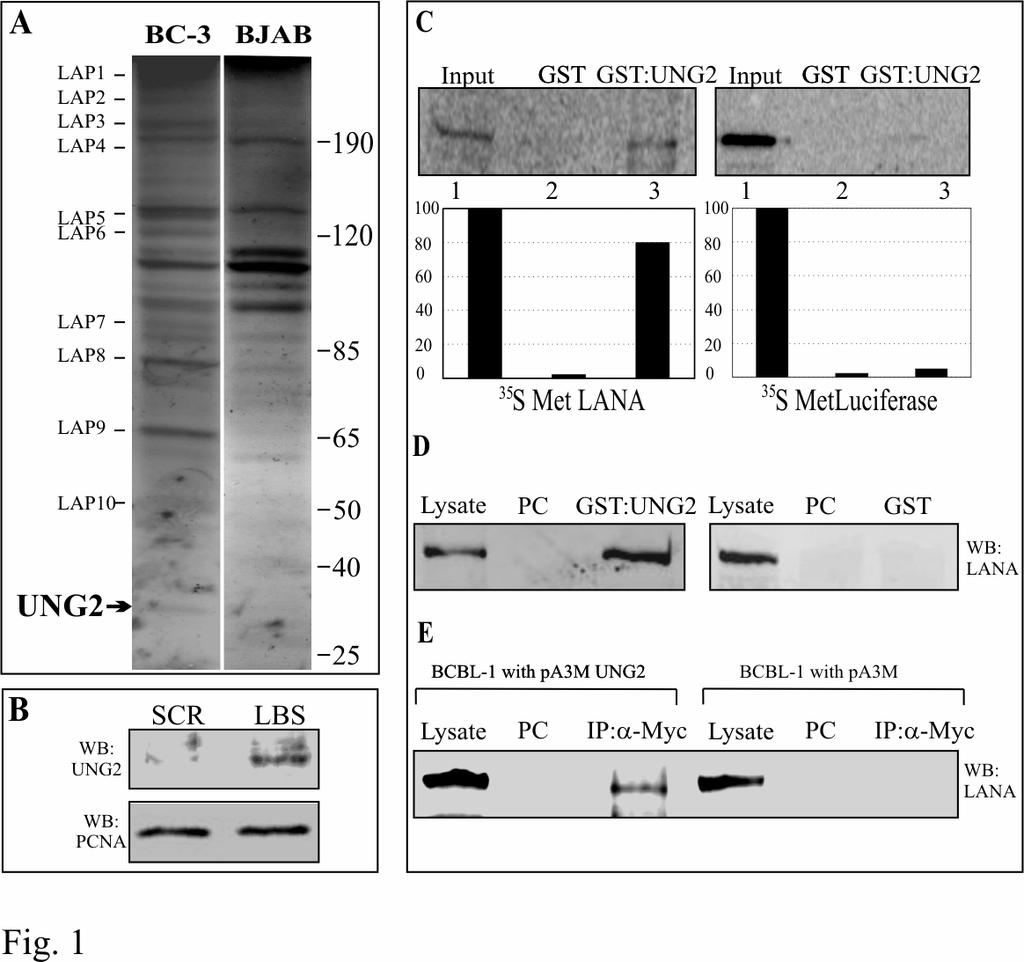
15 Results: Host cellular protein binds to the multimerized LANA Binding Sequence (LBS) affinity column: Multiple studies have demonstrated a role for LANA in KSHV genome replication (28, 32, 71). However, LANA does not have any enzymatic activity associated with DNA replication including DNA unwinding or ATPase activities, required for DNA replication. LANA is also important for persistence of the KSHV genome as well as replication of TR containing plasmids (4, 16). LANA deleted recombinant KSHV and shrna depleting LANA transcript reduced KSHV genomic copies demonstrating that tethering is important for long term persistence (26, 74). LANA has been shown to bind to cellular proteins to support tethering and replication of KSHV genome (7, 16, 40, 63, 71). In order to identify cellular proteins associated with LANA when LANA is bound to its cognate sequence we performed LBS affinity binding assay (71). Fractions from BC-3 and BJAB (KSHV positive and negative, respectively) eluted at 500mM NaCl were resolved on a 10% SDS-PAGE. Coomassie staining of the gel detected unique bands in eluates from BC-3 LBS affinity column (Fig 1A). There were also a number of unique bands in BJAB NE, however we decided to identify the bands from BC-3 NE as the immediate focus was to identify the LANA associated proteins. These unique bands from BC-3 NE were designated as LAP1-11 (LANA Associating Proteins). Identification of LANA associating proteins: Coomassie stained bands were excised from the gel and submitted for identification using LC-MS/MS analysis at the Proteomics core of the University of Pennsylvania. The Mascot software, which uses a statistical scoring algorithm for the identification of protein bands was used (53). The MS spectra were also manually checked to ensure that each identification were of 15
16 high confidence. Matching of the theoretical molecular weight of the identified bands also supported the MS result and the level of confidence. The identities of these bands are listed in Table 1. LAP11 was identified as Uracil DNA Glycosylase (UNG2, Accession number NP_550433). Uracil DNA glycosylase (UNG2) has been shown to remove uracil residues from the replicating DNA through a Base Excision Repair (BER) pathway (41). UNG2 preferentially binds to a DNA affinity column conjugated with LANA binding sequence: In order to confirm the specificity of LANA and UNG2 binding to the LBS affinity column we incubated BC-3 nuclear extract with the columns of LBS and scrambled LBS (SCR LBS) DNA under similar condition. After thorough washing of the column proteins were eluted from both the columns and resolved on 10% SDS-PAGE. Western blot analysis for the detection of UNG2 in eluates from these two columns revealed that UNG2 predominantly associated with LBS column compared to SCR LBS column although a faint signal was seen in SCR eluate (Fig 1B). LANA was not detected in SCR LBS column confirming inability of LANA to bind to the scrambled sequence (data not shown). UNG2 forms complexes with LANA in-vitro and in human cells expressing LANA: We confirmed the above binding results of UNG2 with LANA using in-vitro binding assays with GST-UNG2 as bait. In-vitro translated LANA incubated with GST-UNG2 fusion protein showed precipitation of LANA (Fig. 1C, lane 3) suggesting that these proteins can interact in vitro. GST alone showed no obvious/visible precipitation of LANA (Fig. 1C lane 2) and GST-UNG2 did not show binding to the control protein, luciferase (Fig 1C, lane 6) suggesting that the interaction of GST-UNG2 with LANA is likely to be specific in the cells. 16
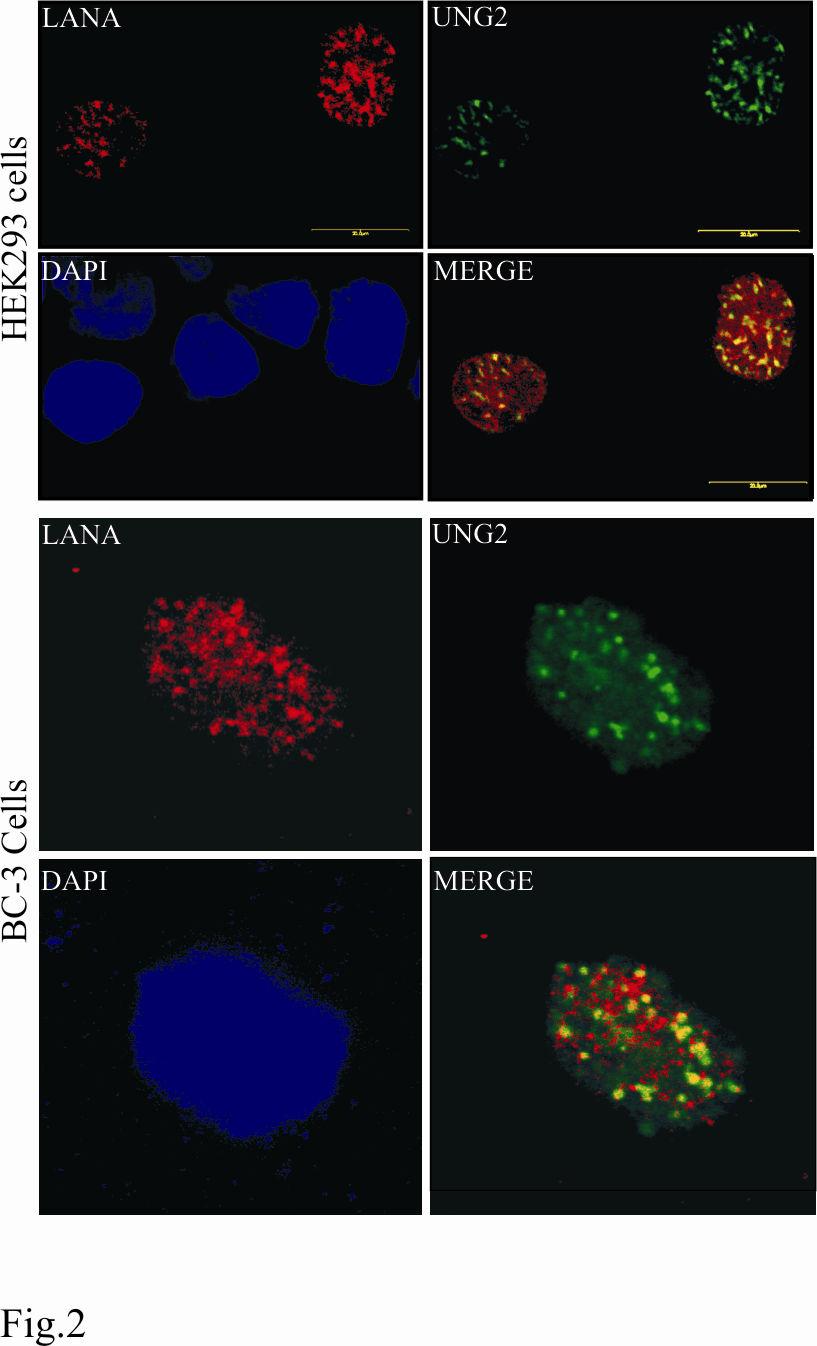
17 We also used LANA heterologously expressed in HEK293T cells to test for its ability to bind GST-UNG2. 293T lysates containing LANA was precleared with Glutathione Sepharose beads and incubated with either GST-UNG2 of GST (Fig 1D). Bound LANA was detected using anti-myc antibody in GST-UNG2 lane (Fig. 1D, lane 3) but not in the GST lane (Fig. 1D, lane 6) corroborated the above results showing binding of these two proteins. KSHV infected cells express LANA during latent infection therefore we wanted to determine whether endogenous LANA expressed from KSHV genome is also capable of interacting with UNG2. PELs (BCBL-1) transfected with either pa3m (myc vector) or pa3mung2 (myc-ung2) were lysed and immunoprecipitated for UNG2 using myc antibody. Detection of LANA in pa3mung2 transfected BCBL-1 cells (Fig 1E, lane 3) was due to coimmunoprecipitation of LANA with UNG2 supporting the association of these two proteins in KSHV positive cells. Absence of LANA in myc IP lane transfected with pa3m supports the specificity of the association in this assay (Fig 1E, lane 6). UNG2 co-localizes with LANA in transiently transfected and KSHV infected cells: HEK293 cells were transfected with pdsred-lana and pegfpung2 and plated on glass slides. 12h post-transfection cells were stained with DAPI and analyzed by confocal laser microscopy. Detection of LANA and UNG2 showed a punctate nuclear pattern and that the majority of the UNG2 signal was colocalized with RFP-LANA again corroborating the above results that these two proteins interact in vivo as they were in same nuclear compartments (Fig.2). We further detected colocalization of LANA and UNG2 in the KSHV infected PEL cell lines, BC-3 (Fig.2). We transfected pegfpung2 in BC-3 cells and detected LANA using rabbit anti-lana antibody followed by staining with goat anti-rabbit Alexa Flour 594 (red). The 17
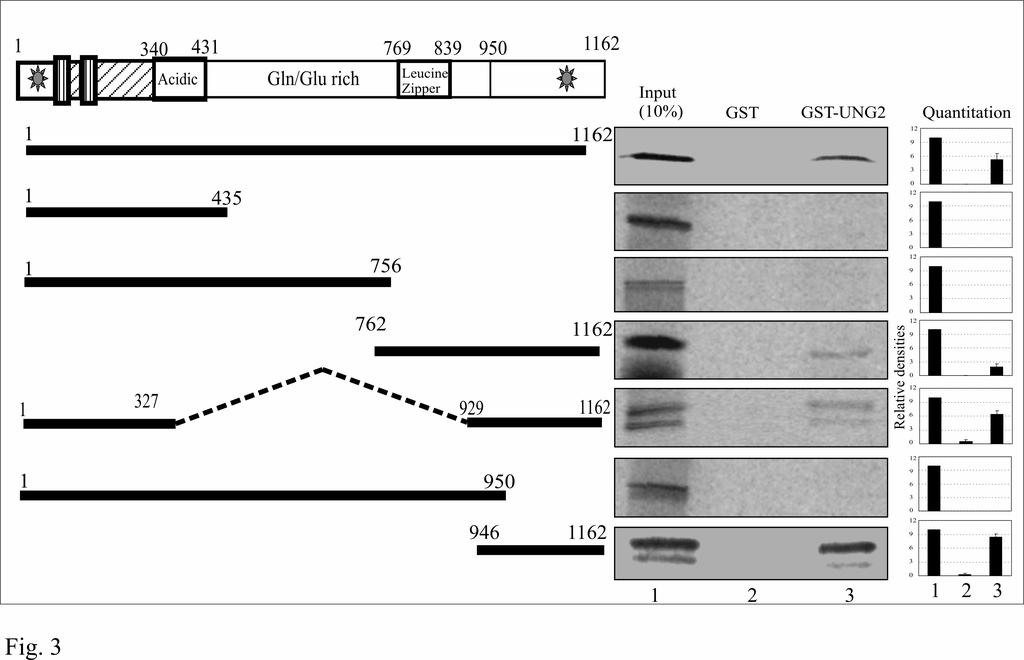
18 merged image of these two proteins showed distinct colocalization of LANA and UNG2 in BC-3 cells. UNG2 binds to the C-terminus of LANA: Different domains of LANA have been shown to be involved for various functions including genome tethering and transcriptional modulation (73). Therefore we wanted to determine the domain of LANA important for the recruitment of UNG2. Mutants of LANA deleted for specific domains were in vitro translated and assayed for binding to GST-UNG2 using 10% of the total translated product as input (Fig. 3). As indicated above, GST did not show any significant binding to the LANA deletion mutants. Additionally, mutants of LANA lacking the C-terminus did not show any binding to GST-UNG2 (Fig. 3, LANA 1-435aa, 1-756, and 1-950aa). LANA deletion mutants which contained the extreme C-terminus ( ) showed significant binding to UNG2 indicating that UNG2 can bind to LANA in close proximity to the DNA binding domain of LANA ( aa) (38). Cellular UNG2 retains its activity when complexes with LANA: We then wanted to determine whether UNG2 when complexed with LANA retains its enzymatic activity? We used an indirect PCR based assay (11) for the detection of UNG2 activity as described in materials and methods section and schematic (see Fig. 4A). If the template DNA containing du is incubated with purified UNG2 and heated at alkaline ph, UNG2 excises the uracil residue from the template and thus generate nick throughout the entire length. This prevents re-amplification of the template DNA. In contrast to the above, DNA containing dt remains intact and can be re-amplified. We used purified UNG2 expressed in E. coli (data not shown) as well as immunoprecipitated UNG2 expressed in HEK293 cells for the test of UNG assay. Results of UNG2 assay on dt and du containing K1 gene amplicons are shown in figure 18
19 4C. Immunoprecipitates from HEK293 cells transfected with vector alone control did not affect re-amplification of K1 gene in both the dt and du containing templates. As expected immunoprecipitates from HA-UNG2 expressing cells blocked re-amplification of du containing templates but not the dt containing templates demonstrating the presence of UNG activity in the complex. In order to assay the uracil DNA glycosylase activity of UNG2 in complex with LANA, we used LANA co-immunoprecipitated UNG2 using anti-myc antibody from LANA-myc and HA-UNG2 transfected HEK293 cells as the source of UNG2. We confirmed coimmunoprecipitation of UNG2 using anti-ha western blot (Fig 4B, lanes 3 and 4). Immunoprecipitates (IP) from the LANA-myc and HA-UNG2 transfected separately were also analyzed by western blot using anti-myc and anti-ha antibodies (Fig. 4B, lanes 1, 2 and 5, 6 respectively). For UNG assay dt and du containing K1 amplicons were incubated with the IP complex from LANA-myc, LANA-myc+HA-UNG2 and HA-UNG2 only transfected cells. Templates from these combinations were subjected to re-amplification and the results are shown in figure 4D. Immunoprecipitates from LANA-myc transfected cells did not block reamplification of the target DNA suggesting that LANA by its own does not have any uracil DNA glycosylase activity (Fig. 4D lanes 1 and 2). Template containing du residues followed by incubation with immunoprecipitate from LANA-myc + HA-UNG2 transfected cells, blocked reamplification suggesting the presence of uracil DNA glycosylase activity in the immunoprecipitated complex (Fig. 4D, lanes 3 and 4). Immunoprecipitates with anti-myc antibody from HA-UNG2 transfected cells did not block re-amplification of K1 template thus ruled out the possibility of non-specific precipitation of UNG2 with anti-myc antibody (Fig. 4D, lanes 5 and 6). This suggested that the UNG2 activity associated with LANA-UNG2 complex is 19
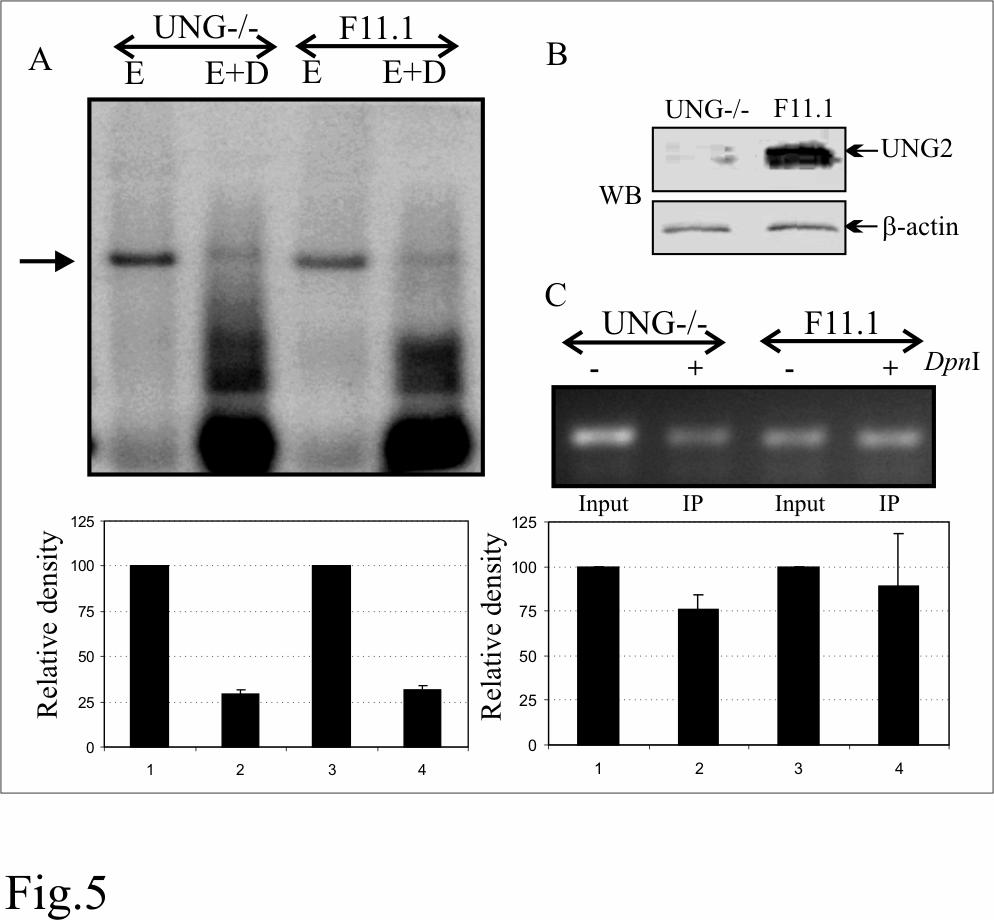
20 mostly due to the UNG2 bound to LANA. Importantly, this assay is not a quantitative assay rather a qualitative assay and detects the presence or absence of DNA glycosylase activity. The reduction in re-amplification of du containing templates treated with anti-myc immunoprecipitates from LANA-myc and HA-UNG2 could be because of the unstable nature of DNA containing RNA base, uracil. Additionally, endogenous UNG2 immunoprecipitated with LANA-myc may also have contributed to the UNG2 activity. Replication of TR containing plasmid was similar in UNG-/- and UNG+/+ (F11.1) Mouse Embryonic Fibroblasts (MEFs): Since UNG2 binds to the carboxyl terminus of LANA which is important for replication, we wanted to determine whether UNG2 is a capable of modulating TR mediated replication? We performed a DpnI sensitivity assay to determine the replication of TR containing plasmids in UNG deficient MEFs. DpnI assay relies on the fact that DpnI digests DNA only if the Adenine (A) of its recognition site (GATC) is methylated. Since eukaryotic cells do not possess dam methylase, plasmid replicated in these cells lacks methylation at Adenine and thus becomes resistant to the DpnI digestion. We used TR containing plasmid DNA prepared in E. coli (dam+) for the replication assay by transfecting into MEFs and determining the DpnI resistant copies in southern blot as well as in PCR. pbspuroa3 (Three TR containing plasmid) was co-transfected with LANA expressing vector into UNG deficient (UNG-/-) and wild type MEF (UNG+/+). Episomal DNA extracted using modified Hirt s procedure was digested with either EcoRI (to linearize) or EcoRI and DpnI overnight and transferred to gene screen membrane followed by detection with 32 P labeled TR probe. DpnI resistant bands were quantified and the relative density is plotted normalizing with input as 10%. Relative densities of DpnI resistant band in UNG-/- and UNG+/+ cells line did not show much difference suggesting that UNG2 does not 20
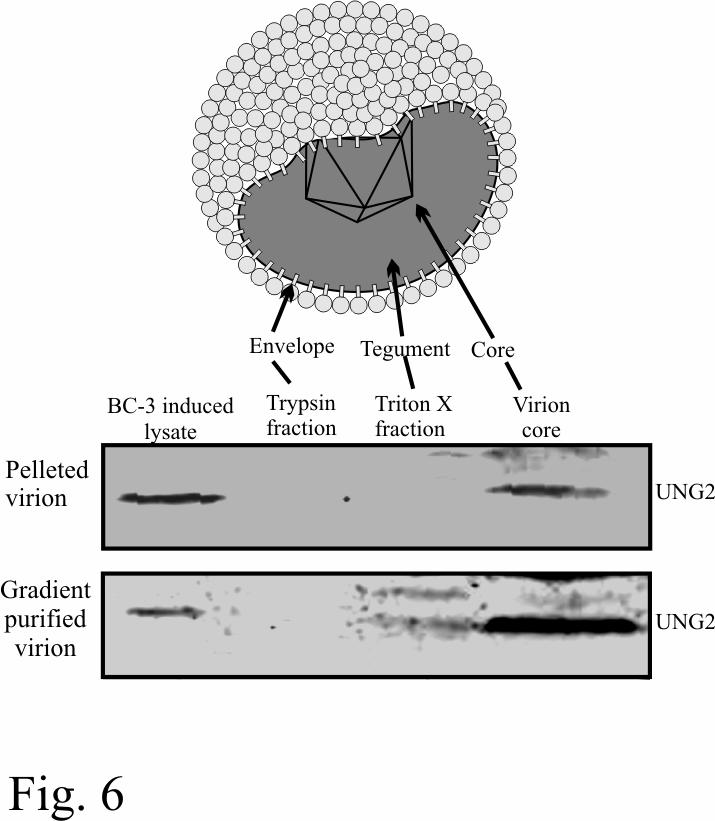
21 directly affect DNA replication (Fig. 5A). Western blot shows the absence of UNG2 in UNG-/- MEF (Fig. 5B). We further supported the above results using BrdU incorporation in newly replicated DNA. UNG-/- and UNG+/+ cells co-transfected with pbspuroa3 and pa3mlana were pulsed with BrdU for 12h followed by extraction of episomal DNA. 90% of the DpnI digested Hirt DNA was subjected for immunoprecipitation with anti-brdu antibody. A region on the vector backbone from the input as well as immunoprecipitated DNA was amplified and the relative densities of the amplicons were determined using 10% as input. Relative densities of the amplicons in UNG-/- and UNG+/+ did not show much difference and thus supported the above result (Fig. 5C). UNG2 packages into virion particle: We and others have previously shown that virions carry cellular as well as viral proteins most probably for rapidly establishing latent infection following a short burst of lytic reactivation (39, 45). Therefore, we wanted to determine whether cellular UNG2 is packaged into virion particles. We determined the presence of UNG2 in pelleted as well as gradient purified virion in a western blot assay. Protein preparation used previously for the detection of RTA and LANA ORCs was used for the detection of UNG2. Specifically, western blot analysis showed the presence of UNG2 predominantly in the core of KSHV virion particles although there was some signal associated with the tegument (Fig. 6). KSHV virions produced from BCBL-1 cells are capable of infecting UNG-/- MEF cells: KSHV virions produced after TPA and sodium butyrate induction were concentrated and purified for infection assays. Equal amounts of virion containing suspension was added to the UNG-/- and UNG+/+ MEFs. Infected cells were stained with anti-lana antibody after one 21
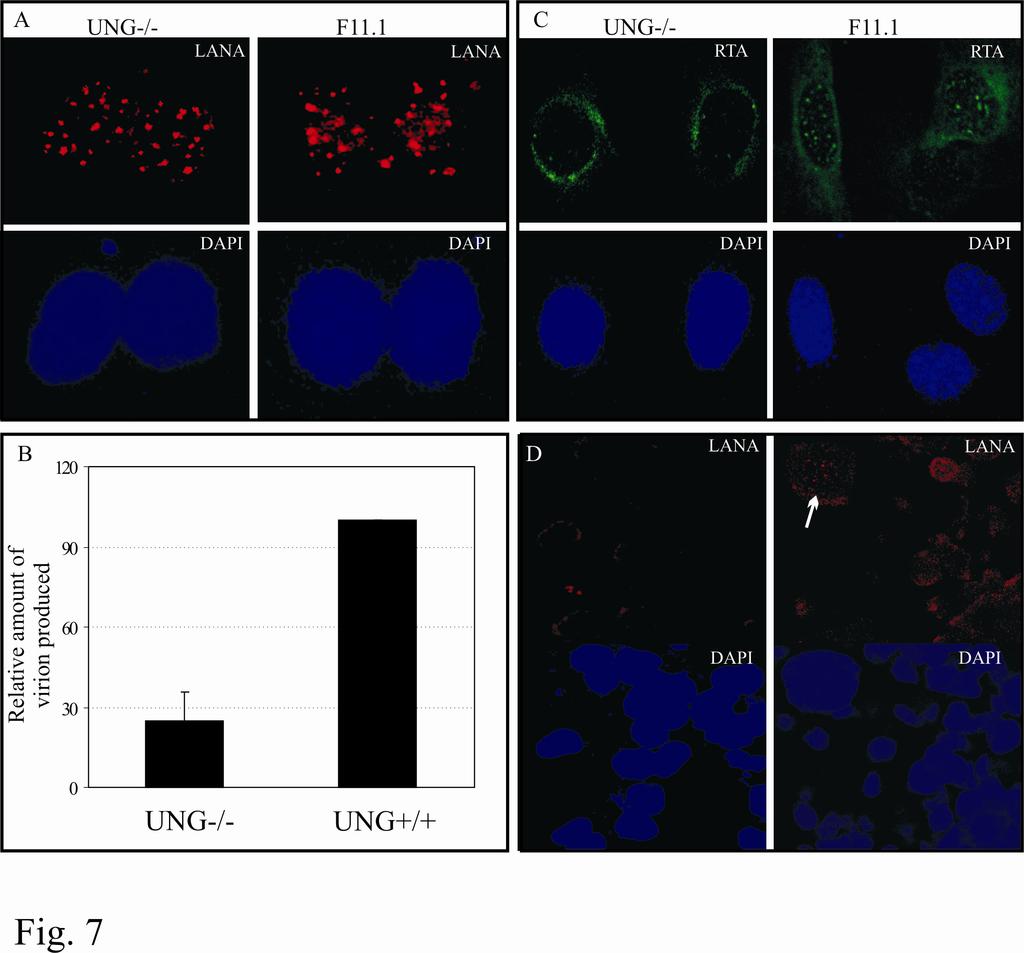
22 passage to detect the infection and establishment of latent infection. LANA staining was indistinguishable in UNG-/- and UNG+/+ cells and a representation of staining is shown in figure 7A. Since UNG2 is a base excision repair enzyme responsible for removal of any misincorporated uracil residues it may not have reflected difference in initial infection. Therefore we passaged the MEFs (UNG-/-, UNG+/+) infected with KSHV for several rounds in order to accumulate mutations due to misincorporation of uracil and deamination of cytosine in the genome. If the inserted mutation occurs in any regulatory gene that would most likely affect the production of virions after induction. In fact, the results of our virion quantitation data showed approximately 3 fold reduction in the number of virions produced from UNG-/- cells when compared with UNG+/+ cells (Fig. 7B). We further wanted to determine the levels of RTA in these MEFs as RTA is essential activating expression of the KSHV lytic genes (69). As expected, we found reduced levels of RTA (Fig. 7C) in UNG-/- MEFs explaining in part reason for reduction in the number of virions produced. Next we examined the infectivity of the virions produced and purified from these UNG-/- and UNG+/+ MEFs. We used equivalent amounts of virion particles (quantified using virion DNA in qpcr) from UNG-/- and UNG+/+ cells for the infection of HEK293 cells. Infectivity was detected by immunostaining for LANA in these cells. LANA staining suggested that that infectivity of the virus progeny produced from the MEFs were highly retarded with very little or no infectivity of UNG-/- produced virion. However, there were few infected cells, detected by LANA staining, with the virions produced from UNG+/+ (Fig. 7D). PELs treated with UNG2 shrna showed reduced number of episomal copies: BC-3 and BCBL-1 cells selected with UNG2 shrna for three weeks were assayed for UNG2 expression levels. UNG2 shrna treated PELs, BC-3 and BCBL-1 showed efficient 22
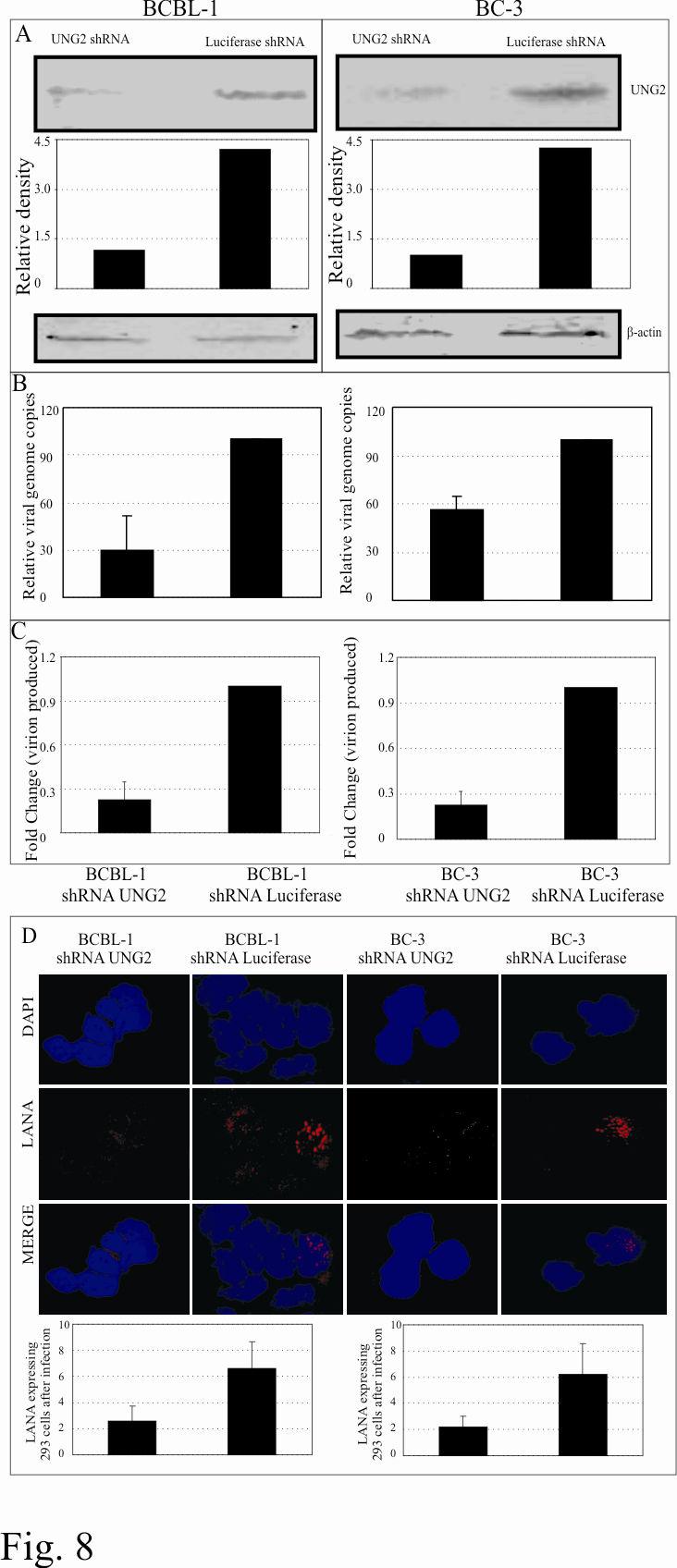
23 depletion of the UNG2 close to 85% reduction compared to the control shrna treated cells (luciferase shrna) (Fig. 8A). We further determined the latently persisting copies of episomal DNA in these shrna treated cells using real-time quantitative PCR assay. The relative number of KSHV episomal copies were significantly reduced in UNG2 shrna treated PELs (Fig. 8B). Upon induction of these cells with TPA and sodium butyrate, the number of virions produced were further reduced which may be a consequence of increased accumulation of mutations in the KSHV genome (Fig. 8C). This result corroborates with the data obtained from UNG2 deficient MEFs. In order to detect the infectivity of the virion produced from UNG2 shrna treated PELs, we infected HEK293 cells with equivalent numbers of virions as quantified by qpcr and detected LANA expression which is a hallmark of latent infection. LANA staining suggested that virions produced from UNG2shRNA treated cells were also able to infect HEK293 cells, however with significantly reduced efficiency compared to the control shrna treated cells (Fig. 8D). Bar diagram represents the number of LANA positive cells in a total of approximately 100 cells in the optical field. Representations of LANA staining cells are shown in figure 8D. 23
24 Discussion: Latency Associated Nuclear Antigen (LANA) of KSHV is critical for tethering of the viral episomes and replication (4, 16). The mechanism of LANA mediated replication is not fully understood but the evidence so far suggests that LANA can recruit components of the cellular replication machinery at the latent replication origin, the TR element (47, 67, 71). Our LBS affinity column data strengthens this hypothesis that host cellular replication machinery is involved in the replication of KSHV episomes (71). Most of the proteins which were identified by the affinity column contribute to DNA replication. These proteins include DNA dependent protein kinase catalytic subunit (DNA-PKcs), DNA topoisomerase 2β, ATP-dependent DNA helicase II (Ku80) and uracil-dna glycosylase isoform 2 (UNG2). DNA dependent protein kinase catalytic subunit of ~460kDa is a serine/threonine kinase and belongs to the PI (phosphatidyl)-3 kinase (p110) family (30). Ku80 is an ATP dependent DNA helicase which forms a dimeric regulatory component with K70 proteins (21, 27). DNA-PKcs, Ku80 and Ku70 forms a heterotrimeric enzyme (DNA-PK) which was previously shown to be involved in phosphorylation of DNA bound proteins and transcription factors including Sp1, p53 and the carboxy-terminal domain of RNA polymerase II, DNA damage and the DNA-activated protein kinase (3). This suggests that DNA-PKcs may play a role in regulating transcription, replication, and recombination as well as DNA repair (50). DNA topoisomerase II beta plays a critical role in controlling DNA topology which is important for transcription, replication and recombination (reviewed in 9). Interestingly, LANA has previously been shown to be involved in the replication as well as KSHV episome segregation during cell division. However, the mechanism of the 24
25 segregation is not understood yet. Association of these proteins with LANA provides clues as to how LANA may function for replication and genome segregation. The role of Uracil DNA Glycosylase (UNG2) has been well studied in base excision repair pathway which specifically removes the RNA base uracil from DNA (41, 43). Incorporation of uracil can occur in DNA either by misincorporation of dutp during replication process or by deamination of cytosine residues. Deamination of cytosine generates G:U mispairs which leads to an A:T transition unless repaired before the next rounds of replication (43). In the genomic DNA the frequency of cytosine deamination is in the order of per genome per day (41). Uracil misincorporation in the genome is due to the direct incorporation of dutp during DNA replication. Incorporation of dutp in DNA during replication is prevented by a specific enzyme dutpase which converts dutp into dump thus lowering the ratio of dutp:dttp and therefore reduces the chance of dutp incorporation (reviewed in (15, 41). Damaged bases may be miscoding, cytotoxic or both (43). To counteract these effects cells have several defense mechanisms either to eliminate the defected cells or repair the damage. These damages then cause a cell cycle arrest allowing repair to take place before cells proceed to the next round of replication. Thus maintaining the integrity of the cellular genome (41, 43). Large DNA human herpesvirus encodes for dutpase and uracil DNA glycosylase (reviewed in 15, 41). However it is unclear why these viruses like to encode genes for these repair enzymes if the host cells contain these genes (reviewed in 15, 41). It is postulated that viruses might need these gene to prevent incorporation and/or retention of uracil in viral DNA under certain physiological conditions, including viral infection and terminal differentiation which had reduced UNG2 and dutpase activities (55, 58, 68). Viruses deficient for dutpase and UNG2 are reported to have decreased replication and virulence (58). Vaccinia virus 25
26 deficient for UNG2 showed impaired replication and virus viability (48, 68). UNG2 encoded by Herpes simplex virus is dispensable for viral replication in immortalized cells but has been shown to be important for efficient viral reactivation and latency in the murine nervous system (57, 59). Recombinant HSV-1 lacking UNG2 has been shown to accumulate mutations over time in culture (59). Cytomegalovirus (CMV) also encodes for dutpase and UNG2 and mutants lacking these enzymes have shown to have defects in DNA replication (55). These studies have suggested that UNG2 is an important enzyme for maintaining the integrity of the viral genome. KSHV like other herpesvirus encodes for a dutpase enzyme but the expression of this gene was only detected during lytic cycle of the virus (data not shown). Therefore, dutpase encoded by KSHV most likely helps in reducing the ratio of dutp:dttp in reactivated cells thus prevents misincorporation of uracil in newly synthesized viral DNA before been packaged into virion particles. Most uracil lesions of the DNA are repaired by a mechanism called Base Excision Repair (BER) which is carried out by the DNA glycosylases (41). Uracil DNA glucosylase hydrolyses the glycosydic bond between the target base, uracil and deoxyribose generating a apurinic/apyrimidinic (AP) site in DNA and releasing a free base (41, 43). These AP sites are repaired by AP endonuclease, deoxyribophosphodiesterase and filled in by DNA polymerase β followed by ligation with DNA ligase III (41). Additionally, human UNG2 has been shown to interact with replication protein A (RPA), important for short patch repair pathways in association with DNA polymerase and proliferating cell nuclear antigen (PCNA) (41). Detection of human UNG2 as a LANA associating protein proposed the hypothesis that LANA may also have a role in maintaining the integrity of KSHV episomal DNA during latent infection by recruiting UNG2 in close proximity to replication foci. It is well established that 26
27 LANA is critical for TR mediated replication of plasmid (28, 32). It is not entirely clear how LANA facilitates replication of TR plasmid but growing evidence suggests that LANA binds to the Origin Recognition Complexes (ORCs) at the TR (71). Binding of MCMs to the TRs are also confirmed in indirect binding assays (Chromatin Immunoprecipitation Assays) (71). Since TR containing plasmids cannot replicate without LANA, either LANA recruits cellular replication machinery to the TR or stabilizes the association of these proteins at the replicator element of the TR. Understanding the role of LANA in KSHV replication is an important area of research and is currently ongoing in our lab. Interaction of UNG2 with carboxyl terminus of LANA, which is also the DNA binding domain (16, 38), suggests that UNG2 remains in close proximity to the TR DNA (Fig. 9). Thus, it maybe involved in removal of any incorporated uracil residues during post-replication. In-vitro assays have previously shown that uracil gets rapidly removed from replicatively incorporated dutp residues in isolated nuclei (51). Since UNG2 retained its glycosylase activity when complexed with LANA demonstrated by PCR based assay indicates that LANA may recruit UNG2 for the repair of any misincorporated uracil residues in TR DNA post-replication. UNG2 is not required for latent DNA replication as UNG2 deficient mouse embryonic fibroblasts (UNG-/-) did not show any significant differences in the replication ability of TR containing plasmids when compared with wild type MEF. LANA was not detected in virion tegument (71) but we detected UNG2 in the purified virion suggesting that it may be important during infection or for maintaining viral genome integrity. In order to determine its role we infected UNG-/- and UNG+/+ cells with BCBL-1 virions which showed comparable levels of infection. However, the virions produced from UNG2 deficient MEF after induction was significantly reduced most likely due to the defect in uracil repair pathway and generation of 27
28 defective copies of latently persisting episomal DNA. Detection of lower levels of RTA in UNG- /- MEF explains partly the reason for a lower number of virions produced from these KSHV infected MEFs. Off note reduction in the number of latently persisting KSHV episomal copies in UNG-/- cells may be just because of the lack of UNG2 and not primarily due to the interaction with LANA. Therefore the strategies to disrupt the interaction of UNG2 with LANA are underway in our lab.. The KSHV positive PEL cells lines, BCBL-1 and BC-3 with reduced levels of UNG2 targeted by shrna showed reduced copies of episomally persisting KSHV genome. Induction of these UNG2 shrna cells produced 3-4 fold lower numbers of KSHV virions compared to cells expressing a control shrna. Importantly, the cells selected with the shrna specific for UNG2 was unable to grow for extended periods in cultures. This may be due to either a loss of UNG2 function or growth retardation because of lower KSHV episomal copies or a combination of the two effects. Infection of 293 cells with the virions produced from shrna treated cells was less infectious as determined on the basis of LANA staining in 293 cells infected with equal number of virions produced from either UNG2 or luciferase (control) shrna treated PELs. In conclusion, the data presented here suggests that most likely LANA brings UNG2 in close proximity to its episomal DNA through interaction with its DNA binding domain (C-terminus) and therefore helps in removing any misincorporated uracil during post-replication repair. Cells lacking UNG2 fails to remove these misincorporated uracil and the successive accumulation of these mutations lead to the generation of defective episomal copies and thus production of defective virions (Fig 9). Therefore, targeting UNG2 function could potentially be used as a useful target for blocking KSHV persistence in the infected cells. 28
29 Acknowledgements: This work was supported by Public Health service grant from NCI CA and CA091792, NIDCR DE01436 and DE (ESR). ESR is a scholar of the Leukemia and Lymphoma Society of America. We thank Prof. Hans E. Krokan and Prof. Geir Slupphaug, Norwegian University of Science and Technology, Norway for providing reagents and helpful suggestions. We also thank Dr. Chao-Xing Yuan and the Proteomics core facility at the University of Pennsylvania, School of Medicine for the identification of protein bands. 29
30 References: 1. Aiyar, A., C. Tyree, and B. Sugden The plasmid replicon of EBV consists of multiple cis-acting elements that facilitate DNA synthesis by the cell and a viral maintenance element. Embo J 17: Andersen, S., T. Heine, R. Sneve, I. Konig, H. E. Krokan, B. Epe, and H. Nilsen Incorporation of dump into DNA is a major source of spontaneous DNA damage, while excision of uracil is not required for cytotoxicity of fluoropyrimidines in mouse embryonic fibroblasts. Carcinogenesis 26: Anderson, C. W DNA damage and the DNA-activated protein kinase. Trends Biochem Sci 18: Ballestas, M. E., P. A. Chatis, and K. M. Kaye Efficient persistence of extrachromosomal KSHV DNA mediated by latency-associated nuclear antigen. Science 284: Ballestas, M. E., and K. M. Kaye Kaposi's sarcoma-associated herpesvirus latency-associated nuclear antigen 1 mediates episome persistence through cis-acting terminal repeat (TR) sequence and specifically binds TR DNA. J Virol 75: Barbera, A. J., M. E. Ballestas, and K. M. Kaye The Kaposi's sarcomaassociated herpesvirus latency-associated nuclear antigen 1 N terminus is essential for chromosome association, DNA replication, and episome persistence. J Virol 78: Barbera, A. J., J. V. Chodaparambil, B. Kelley-Clarke, V. Joukov, J. C. Walter, K. Luger, and K. M. Kaye The nucleosomal surface as a docking station for Kaposi's sarcoma herpesvirus LANA. Science 311: Bell, S. P., and A. Dutta DNA replication in eukaryotic cells. Annu Rev Biochem 71: Berger, J. M., and J. C. Wang Recent developments in DNA topoisomerase II structure and mechanism. Curr Opin Struct Biol 6: Boshoff, C Kaposi virus scores cancer coup. Nat Med 9: Bouhamdan, M., S. Benichou, F. Rey, J. M. Navarro, I. Agostini, B. Spire, J. Camonis, G. Slupphaug, R. Vigne, R. Benarous, and J. Sire Human immunodeficiency virus type 1 Vpr protein binds to the uracil DNA glycosylase DNA repair enzyme. J Virol 70: Cesarman, E., Y. Chang, P. S. Moore, J. W. Said, and D. M. Knowles Kaposi's sarcoma-associated herpesvirus-like DNA sequences in AIDS-related body-cavity-based lymphomas. N Engl J Med 332: Chang, Y Kaposi's sarcoma and Kaposi's sarcoma associated herpesvirus (human herpesvirus 8): where are we now? J Natl Cancer Inst 89: Chang, Y., E. Cesarman, M. S. Pessin, F. Lee, J. Culpepper, D. M. Knowles, and P. S. Moore Identification of herpesvirus-like DNA sequences in AIDS-associated Kaposi's sarcoma. Science 266: Chen, R., H. Wang, and L. M. Mansky Roles of uracil-dna glycosylase and dutpase in virus replication. J Gen Virol 83:
31 16. Cotter, M. A., 2nd, and E. S. Robertson The latency-associated nuclear antigen tethers the Kaposi's sarcoma-associated herpesvirus genome to host chromosomes in body cavity-based lymphoma cells. Virology 264: Cotter, M. A., 2nd, C. Subramanian, and E. S. Robertson The Kaposi's sarcoma-associated herpesvirus latency-associated nuclear antigen binds to specific sequences at the left end of the viral genome through its carboxy-terminus. Virology 291: Dittmer, D., M. Lagunoff, R. Renne, K. Staskus, A. Haase, and D. Ganem A cluster of latently expressed genes in Kaposi's sarcoma-associated herpesvirus. J Virol 72: Dupin, N., T. L. Diss, P. Kellam, M. Tulliez, M. Q. Du, D. Sicard, R. A. Weiss, P. G. Isaacson, and C. Boshoff HHV-8 is associated with a plasmablastic variant of Castleman disease that is linked to HHV-8-positive plasmablastic lymphoma. Blood 95: Dupin, N., C. Fisher, P. Kellam, S. Ariad, M. Tulliez, N. Franck, E. van Marck, D. Salmon, I. Gorin, J. P. Escande, R. A. Weiss, K. Alitalo, and C. Boshoff Distribution of human herpesvirus-8 latently infected cells in Kaposi's sarcoma, multicentric Castleman's disease, and primary effusion lymphoma. Proc Natl Acad Sci U S A 96: Dvir, A., S. R. Peterson, M. W. Knuth, H. Lu, and W. S. Dynan Ku autoantigen is the regulatory component of a template-associated protein kinase that phosphorylates RNA polymerase II. Proc Natl Acad Sci U S A 89: Flore, O., S. Rafii, S. Ely, J. J. O'Leary, E. M. Hyjek, and E. Cesarman Transformation of primary human endothelial cells by Kaposi's sarcoma-associated herpesvirus. Nature 394: Friborg, J., Jr., W. Kong, M. O. Hottiger, and G. J. Nabel p53 inhibition by the LANA protein of KSHV protects against cell death. Nature 402: Fujimuro, M., F. Y. Wu, C. ApRhys, H. Kajumbula, D. B. Young, G. S. Hayward, and S. D. Hayward A novel viral mechanism for dysregulation of beta-catenin in Kaposi's sarcoma-associated herpesvirus latency. Nat Med 9: Garber, A. C., J. Hu, and R. Renne Latency-associated nuclear antigen (LANA) cooperatively binds to two sites within the terminal repeat, and both sites contribute to the ability of LANA to suppress transcription and to facilitate DNA replication. J Biol Chem 277: Godfrey, A., J. Anderson, A. Papanastasiou, Y. Takeuchi, and C. Boshoff Inhibiting primary effusion lymphoma by lentiviral vectors encoding short hairpin RNA. Blood 105: Gottlieb, T. M., and S. P. Jackson The DNA-dependent protein kinase: requirement for DNA ends and association with Ku antigen. Cell 72: Grundhoff, A., and D. Ganem The latency-associated nuclear antigen of Kaposi's sarcoma-associated herpesvirus permits replication of terminal repeat-containing plasmids. J Virol 77: Hansen, R. S., T. K. Canfield, M. M. Lamb, S. M. Gartler, and C. D. Laird Association of fragile X syndrome with delayed replication of the FMR1 gene. Cell 73:
32 30. Hartley, K. O., D. Gell, G. C. Smith, H. Zhang, N. Divecha, M. A. Connelly, A. Admon, S. P. Lees-Miller, C. W. Anderson, and S. P. Jackson DNA-dependent protein kinase catalytic subunit: a relative of phosphatidylinositol 3-kinase and the ataxia telangiectasia gene product. Cell 82: Hirt, B Selective extraction of polyoma DNA from infected mouse cell cultures. J Mol Biol 26: Hu, J., A. C. Garber, and R. Renne The latency-associated nuclear antigen of Kaposi's sarcoma-associated herpesvirus supports latent DNA replication in dividing cells. J Virol 76: Hu, J., and R. Renne Characterization of the minimal replicator of Kaposi's sarcoma-associated herpesvirus latent origin. J Virol 79: Kedes, D. H., M. Lagunoff, R. Renne, and D. Ganem Identification of the gene encoding the major latency-associated nuclear antigen of the Kaposi's sarcoma-associated herpesvirus. J Clin Invest 100: Kedes, D. H., E. Operskalski, M. Busch, R. Kohn, J. Flood, and D. Ganem The seroepidemiology of human herpesvirus 8 (Kaposi's sarcoma-associated herpesvirus): distribution of infection in KS risk groups and evidence for sexual transmission. Nat Med 2: Kellam, P., C. Boshoff, D. Whitby, S. Matthews, R. A. Weiss, and S. J. Talbot Identification of a major latent nuclear antigen, LNA-1, in the human herpesvirus 8 genome. J Hum Virol 1: Knight, J. S., M. A. Cotter, 2nd, and E. S. Robertson The latency-associated nuclear antigen of Kaposi's sarcoma-associated herpesvirus transactivates the telomerase reverse transcriptase promoter. J Biol Chem 276: Komatsu, T., M. E. Ballestas, A. J. Barbera, B. Kelley-Clarke, and K. M. Kaye KSHV LANA1 binds DNA as an oligomer and residues N-terminal to the oligomerization domain are essential for DNA binding, replication, and episome persistence. Virology 319: Krishnan, H. H., P. P. Naranatt, M. S. Smith, L. Zeng, C. Bloomer, and B. Chandran Concurrent expression of latent and a limited number of lytic genes with immune modulation and antiapoptotic function by Kaposi's sarcoma-associated herpesvirus early during infection of primary endothelial and fibroblast cells and subsequent decline of lytic gene expression. J Virol 78: Krithivas, A., M. Fujimuro, M. Weidner, D. B. Young, and S. D. Hayward Protein interactions targeting the latency-associated nuclear antigen of Kaposi's sarcomaassociated herpesvirus to cell chromosomes. J Virol 76: Krokan, H. E., F. Drablos, and G. Slupphaug Uracil in DNA--occurrence, consequences and repair. Oncogene 21: Krokan, H. E., M. Otterlei, H. Nilsen, B. Kavli, F. Skorpen, S. Andersen, C. Skjelbred, M. Akbari, P. A. Aas, and G. Slupphaug Properties and functions of human uracil-dna glycosylase from the UNG gene. Prog Nucleic Acid Res Mol Biol 68: Krokan, H. E., R. Standal, and G. Slupphaug DNA glycosylases in the base excision repair of DNA. Biochem J 325 (Pt 1): Lagunoff, M., and D. Ganem The structure and coding organization of the genomic termini of Kaposi's sarcoma-associated herpesvirus. Virology 236:
33 45. Lan, K., D. A. Kuppers, S. C. Verma, N. Sharma, M. Murakami, and E. S. Robertson Induction of Kaposi's sarcoma-associated herpesvirus latencyassociated nuclear antigen by the lytic transactivator RTA: a novel mechanism for establishment of latency. J Virol 79: Lennette, E. T., D. J. Blackbourn, and J. A. Levy Antibodies to human herpesvirus type 8 in the general population and in Kaposi's sarcoma patients. Lancet 348: Lim, C., H. Sohn, D. Lee, Y. Gwack, and J. Choe Functional dissection of latency-associated nuclear antigen 1 of Kaposi's sarcoma-associated herpesvirus involved in latent DNA replication and transcription of terminal repeats of the viral genome. J Virol 76: Millns, A. K., M. S. Carpenter, and A. M. DeLange The vaccinia virus-encoded uracil DNA glycosylase has an essential role in viral DNA replication. Virology 198: Nilsen, H., M. Otterlei, T. Haug, K. Solum, T. A. Nagelhus, F. Skorpen, and H. E. Krokan Nuclear and mitochondrial uracil-dna glycosylases are generated by alternative splicing and transcription from different positions in the UNG gene. Nucleic Acids Res 25: Odegaard, E., C. R. Yang, and D. A. Boothman DNA-dependent protein kinase does not play a role in adaptive survival responses to ionizing radiation. Environ Health Perspect 106 Suppl 1: Otterlei, M., E. Warbrick, T. A. Nagelhus, T. Haug, G. Slupphaug, M. Akbari, P. A. Aas, K. Steinsbekk, O. Bakke, and H. E. Krokan Post-replicative base excision repair in replication foci. Embo J 18: Pan, H., F. Zhou, and S. J. Gao Kaposi's sarcoma-associated herpesvirus induction of chromosome instability in primary human endothelial cells. Cancer Res 64: Perkins, D. N., D. J. Pappin, D. M. Creasy, and J. S. Cottrell Probability-based protein identification by searching sequence databases using mass spectrometry data. Electrophoresis 20: Piolot, T., M. Tramier, M. Coppey, J. C. Nicolas, and V. Marechal Close but distinct regions of human herpesvirus 8 latency-associated nuclear antigen 1 are responsible for nuclear targeting and binding to human mitotic chromosomes. J Virol 75: Prichard, M. N., G. M. Duke, and E. S. Mocarski Human cytomegalovirus uracil DNA glycosylase is required for the normal temporal regulation of both DNA synthesis and viral replication. J Virol 70: Priet, S., N. Gros, J. M. Navarro, J. Boretto, B. Canard, G. Querat, and J. Sire HIV-1-associated uracil DNA glycosylase activity controls dutp misincorporation in viral DNA and is essential to the HIV-1 life cycle. Mol Cell 17: Pyles, R. B., N. M. Sawtell, and R. L. Thompson Herpes simplex virus type 1 dutpase mutants are attenuated for neurovirulence, neuroinvasiveness, and reactivation from latency. J Virol 66: Pyles, R. B., and R. L. Thompson Evidence that the herpes simplex virus type 1 uracil DNA glycosylase is required for efficient viral replication and latency in the murine nervous system. J Virol 68:
34 59. Pyles, R. B., and R. L. Thompson Mutations in accessory DNA replicating functions alter the relative mutation frequency of herpes simplex virus type 1 strains in cultured murine cells. J Virol 68: Radkov, S. A., P. Kellam, and C. Boshoff The latent nuclear antigen of Kaposi sarcoma-associated herpesvirus targets the retinoblastoma-e2f pathway and with the oncogene Hras transforms primary rat cells. Nat Med 6: Rainbow, L., G. M. Platt, G. R. Simpson, R. Sarid, S. J. Gao, H. Stoiber, C. S. Herrington, P. S. Moore, and T. F. Schulz The 222- to 234-kilodalton latent nuclear protein (LNA) of Kaposi's sarcoma-associated herpesvirus (human herpesvirus 8) is encoded by orf73 and is a component of the latency-associated nuclear antigen. J Virol 71: Russo, J. J., R. A. Bohenzky, M. C. Chien, J. Chen, M. Yan, D. Maddalena, J. P. Parry, D. Peruzzi, I. S. Edelman, Y. Chang, and P. S. Moore Nucleotide sequence of the Kaposi sarcoma-associated herpesvirus (HHV8). Proc Natl Acad Sci U S A 93: Shinohara, H., M. Fukushi, M. Higuchi, M. Oie, O. Hoshi, T. Ushiki, J. Hayashi, and M. Fujii Chromosome binding site of latency-associated nuclear antigen of Kaposi's sarcoma-associated herpesvirus is essential for persistent episome maintenance and is functionally replaced by histone H1. J Virol 76: Si, H., and E. S. Robertson Kaposi's sarcoma-associated herpesvirus-encoded latency-associated nuclear antigen induces chromosomal instability through inhibition of p53 function. J Virol 80: Soulier, J., L. Grollet, E. Oksenhendler, P. Cacoub, D. Cazals-Hatem, P. Babinet, M. F. d'agay, J. P. Clauvel, M. Raphael, L. Degos, and et al Kaposi's sarcomaassociated herpesvirus-like DNA sequences in multicentric Castleman's disease. Blood 86: Staskus, K. A., W. Zhong, K. Gebhard, B. Herndier, H. Wang, R. Renne, J. Beneke, J. Pudney, D. J. Anderson, D. Ganem, and A. T. Haase Kaposi's sarcomaassociated herpesvirus gene expression in endothelial (spindle) tumor cells. J Virol 71: Stedman, W., Z. Deng, F. Lu, and P. M. Lieberman ORC, MCM, and histone hyperacetylation at the Kaposi's sarcoma-associated herpesvirus latent replication origin. J Virol 78: Stuart, D. T., C. Upton, M. A. Higman, E. G. Niles, and G. McFadden A poxvirus-encoded uracil DNA glycosylase is essential for virus viability. J Virol 67: Sun, R., S. F. Lin, L. Gradoville, Y. Yuan, F. Zhu, and G. Miller A viral gene that activates lytic cycle expression of Kaposi's sarcoma-associated herpesvirus. Proc Natl Acad Sci U S A 95: Verma, S. C., S. Borah, and E. S. Robertson Latency-associated nuclear antigen of Kaposi's sarcoma-associated herpesvirus up-regulates transcription of human telomerase reverse transcriptase promoter through interaction with transcription factor Sp1. J Virol 78: Verma, S. C., T. Choudhuri, R. Kaul, and E. S. Robertson Latency-associated nuclear antigen (LANA) of Kaposi's sarcoma-associated herpesvirus interacts with origin 34
35 recognition complexes at the LANA binding sequence within the terminal repeats. J Virol 80: Verma, S. C., K. Lan, T. Choudhuri, and E. S. Robertson Kaposi's sarcomaassociated herpesvirus-encoded latency-associated nuclear antigen modulates K1 expression through its cis-acting elements within the terminal repeats. J Virol 80: Verma, S. C., and E. S. Robertson Molecular biology and pathogenesis of Kaposi sarcoma-associated herpesvirus. FEMS Microbiol Lett 222: Ye, F. C., F. C. Zhou, S. M. Yoo, J. P. Xie, P. J. Browning, and S. J. Gao Disruption of Kaposi's sarcoma-associated herpesvirus latent nuclear antigen leads to abortive episome persistence. J Virol 78: Zhong, W., H. Wang, B. Herndier, and D. Ganem Restricted expression of Kaposi sarcoma-associated herpesvirus (human herpesvirus 8) genes in Kaposi sarcoma. Proc Natl Acad Sci U S A 93:
36 Figure Legends: Figure 1: LANA bound to its cognate sequence in affinity column interacts with host cell proteins. A. Proteins eluted with buffer containing 500mM NaCl from LANA Binding Sequence (LBS) affinity column were resolved on 10%SDS-PAGE and stained with coomassie brilliant blue. Bands specific to BC-3 cells (indicated as LAP, LANA Associating Proteins) were excised and sequenced. LAP12 was identified as uracil DNA glycosylase (UNG2). B. UNG2 elutes from the affinity column containing LBS DNA but not from the scrambled DNA (SCR LBS) column. Equal amounts of BC-3 nuclear extract (NE) was incubated with either LBS or scrambled LBS DNA affinity column followed by thoroughly washing to remove any loosely bound protein. Protein eluted at 500mM NaCl containing buffer was subjected for the detection of UNG2 in western blot. Proliferating Cell Nuclear Antigen (PCNA) shows equal amounts of NE was taken in binding reaction. C. In-vitro translated LANA specifically binds to GST-UNG2. 35 S methionine labeled LANA was incubated with either GST or GST-UNG2 and the bound fractions were quantified. Control protein (luciferase) did not bind to GST-UNG2. D. Lysates from 293T cells expressing LANA-myc was incubated with either GST or GST-UNG2 and the bound fraction was analyzed by western blot using anti-myc antibody. LANA was precipitated using GST-UNG2 but not in GST lane. PC; preclear with Glutathione Sepharose beads. E. LANA co-immunoprecipitated with UNG2 from BCBL-1 cells. pa3mung2 or pa3m vector only were transfected into BCBL-1 cells and immunoprecipitated using myc ascites. Western detection using anti-lana antibody showed co-immunoprecipitation of LANA from 36
37 pa3mung2 transfected cells but not from vector only transfected cells. PC; preclear with protein A and protein G beads. Figure 2: UNG2 colocalizes with LANA. HEK293 cells transfected with pdsred-lana (red) and pegfpung2 (green) showed colocalization (yellow) of these two proteins detected in cotransfected cells (merge panel). Cells stained with DAPI shows nuclear staining (blue). UNG2 colocalizes with LANA in BC-3 cells. BC-3 cells transfected with pegfpung2 were stained with anti-lana antibody followed by detection with AlexaFlour 594 (red) showed nuclear punctate staining of LANA. UNG2 detected as GFP fusion protein showed colocalization of these two proteins. Figure 3: UNG2 binds to the carboxyl terminus of LANA. Indicated LANA deletion mutants were in-vitro translated using transcription/translation kit in the presence of 35 S methionine and subjected for binding with either GST or GST-UNG2. LANA deletion mutants containing the carboxyl terminus ( aa, 1-327^ , and ) bound to GST-UNG2. Bound fractions were quantified and the relative binding with each LANA deletion mutants were calculated based on three independent experiments are shown as a bar graph. Figure 4: UNG2 retains its glycosylase activity when complexes with LANA. A. Schematic for the detection of UNG2 activity in PCR assay. Amplification of any gene using specific set of primers in the presence of du incorporates uracil in the DNA. Treatment of this amplicon with UNG2 cleaves the glycosydic bond and generates apurinic/pyrimidinic (AP) sites which blocks next rounds of amplification. dt containing amplicon remains unaffected with UNG2 therefore serves as template for the next rounds of amplification. B. Co-immunoprecipitation of UNG2 with LANA using anti-myc antibody. UNG2 co-immunoprecipitates with LANA (lane 4) detected in anti-ha WB. HA-UNG2 cotransfected with pa3m (vector only) was not detected in 37
38 myc IP lane suggesting for the specificity of co-ip (lane 6). C. Immunoprecipitated UNG2 blocked reamplification of target DNA. KSHV K1 gene amplified using specific primer in the presence of dt or du and treated with UNG2 blocked reamplification of du containing amplicon (lane 4). Lane 5 shows specific amplification of the K1 gene using purified DNA as template. D. Immunoprecipitated LANA did not block reamplification of du containing templates after UNG assay (lane 2). UNG2 co-immunoprecipiated with LANA blocked reamplification of du conataining templates (lane 4). Lanes 5 and 6 is the reamplification product using dt and du containing DNA as template after UNG assay with co-ip complex from pa3m vector and HAUNG2 transfected cells. Lanes 7 and 8 are the reamplication products from equal amounts of template containing dt and du used in the UNG assay. Reduced amounts of re-amplification in du containing templates were most probably due to instability of RNA base, uracil containing DNA. Figure 5: TR containing plasmid (pbspuroa3) replicates in UNG-/- MEF with almost similar efficiency to the wild type MEF (UNG+/+). A. pbspuroa3 containing three copies of TRs was co-transfected with pa3mlana into UNG-/- and F11.1 (UNG+/+) cells. Plasmid extracted after 96h post-transfection were digested with either EcoRI or EcoRI and DpnI followed by southern detection of DpnI resistant copies indicated replication of plasmids. Densitometric analysis of DpnI resistant band from three experiments indicates the presence of almost similar number of replicated copies in these two MEFs. B. Western detection of UNG2 in MEFs, UNG- /- and UNG+/+. β-actin panel to show equal protein loading. C. Quantitation of replicated copies of the TR containing plasmids using BrdU labeling. pbspuroa3 and pa3mlana co-transfected UNG-/- and UNG+/+ cells were pulsed with BrdU followed by plasmid extraction. 90% of the DpnI digested Hirt DNA was immunoprecipitated with anti-brdu antibody followed by 38
39 purification of bound DNA. Equal volume of the purified DNA from UNG-/- and UNG+/+ were used as template for the amplification of on the vector backbone. Densitometric analysis of amplified bands from three independent experiments is shown as bar diagram. Figure 6: KSHV virion packages cellular UNG2. BC-3 virion produced from 500million cells were pelleted as well as purified in sucrose gradient centrifugation. Different fractions of the virions were extracted as described previously (71) followed by western detection of UNG2. Figure 7: KSHV virion infects UNG2 deficient MEF (UNG-/-) with almost similar efficiency as wild type (UNG+/+). A. Representation of immunolocalization of LANA in KSHV infected MEFs. B. Quantitation of KSHV virion produced from infected UNG-/- and UNG+/+ MEFs showed approx. 30% reduction in the number of virions produced from equal number of infected MEFs. C. Detection of RTA in KSHV infected UNG-/- and UNG+/+ cells after induction with TPA and sodium butyrate. D. LANA immunostaining in 293 cells infected with KSHV virions produced from UNG-/- and UNG+/+ cells. Infection of 293 cell was greatly reduced with the viruses produced from MEFs with almost no LANA detection in 293 cells infected with virus from UNG-/- and only few cells with UNG+/+ produced virus. Figure 8: UNG2 shrna treated PELs showed reduced number of episomal copies and produced less virion after reactivation. A. Detection of UNG2 in shrna treated BCBL-1 and BC-3 cells showed significant knockdown of UNG2 levels due to UNG2 shrna treated cells but not with control luciferase shrna. β-actin blots show equal protein loading. B. UNG2 shrna treated BCBL-1 and BC-3 cells showed 40-70% decrease in viral genome copy number during latency. Viral copies were calculated in realtime quantitative PCR using K1 gene as target for amplification. C. Relative number of virions produced after TPA and sodium butyrate treatment of BCBL-1 and BC-3 cells selected with UNG2 and Luc shrna. Virions were quantified by 39
40 isolating DNA from purified virion and quantification using K1 gene amplification in realtime qpcr. D. Immuno staining of LANA in 293 cells infected with virions produced from UNG2 and Luc shrna treated cells. Equal numbers of virions from UNG2 and Luc shrna selected cells were used for infection of 293 cells. Average number of LANA positive cells per optical field was counted and presented as bar graph. Based on LANA staining virions produced from UNG2 shrna PELs showed 3-4 fold reduced infectivity. Figure 9: Proposed model of LANA and UNG2 interaction at KSHV terminal repeats. LANA is a large (1162aa) nuclear protein important for tethering of the viral genome to the host chromosome (4, 16). 3D structure of LANA was predicted using Robetta, free protein structure prediction server ( which shows distinct amino and carboxyl terminal domains. Amino terminal domain of LANA shows distinct chromosome binding sequence (CBS, amino acids 5-22). DNA binding domain (DBD) of LANA which lies between amino acids (38) are marked in blue (LANA-C). The interaction of LANA with UNG2 was mapped to the carboxyl terminus ( aa). The interaction of LANA and UNG2 is most likely important for bringing UNG2 in close proximity to the TR DNA which most likely important for removing any uracil residues incorporated during replication process. Incorporation of uracil if un-repaired may lead to the production of defective virus. PELs with depleted UNG2 using shrna showed reduced KSHV genomic copies may be due to the accumulation of mutations caused by the incorporation of un-repaired uracil residues during successive rounds of replication. 40
41 Table 1: Identification of the protein bands from nuclear extracts of BC-3 cells detected in LBS affinity column. Band Protein Identity Accession LAP1 DNA-dependent protein kinase catalytic subunit (DNA-PK catalytic subunit) (DNA-PKcs) Number P78527 LAP2 Spectrin, beta, non-erythrocytic 1 isoform 1 NP_ LAP3 Clathrin heavy chain 1 NP_ LAP4 DNA topoisomerase 2-beta Q02880 LAP5 Cyclin dependent kinase like protein 5 NP_ LAP6 Splicing factor 3b, subunit 1b NP_ LAP7 EBNA2 coactivator (p100) NP_ LAP8 ATP-dependent DNA helicase II (Ku80) NP_ LAP9 Heat shock protein hsp70 NP_ LAP10 Actin binding protein ABP620 BAA83821 LAP11 Uracil-DNA glycosylase isoform 2 (UNG2) NP_
42 42
43 43
44 44
45 45
46 46
47 47
48 48
49 49
Recombinant Protein Expression Retroviral system
 Recombinant Protein Expression Retroviral system Viruses Contains genome DNA or RNA Genome encased in a protein coat or capsid. Some viruses have membrane covering protein coat enveloped virus Ø Essential
Recombinant Protein Expression Retroviral system Viruses Contains genome DNA or RNA Genome encased in a protein coat or capsid. Some viruses have membrane covering protein coat enveloped virus Ø Essential
Egr-1 regulates RTA transcription through a cooperative involvement of transcriptional regulators
 /, 2017, Vol. 8, (No. 53), pp: 91425-91444 Egr-1 regulates RTA transcription through a cooperative involvement of transcriptional regulators Roni Sarkar 1 and Subhash C. Verma 1 1 Department of Microbiology
/, 2017, Vol. 8, (No. 53), pp: 91425-91444 Egr-1 regulates RTA transcription through a cooperative involvement of transcriptional regulators Roni Sarkar 1 and Subhash C. Verma 1 1 Department of Microbiology
Supplementary Information
 Supplementary Information Supplementary Figure 1. CD4 + T cell activation and lack of apoptosis after crosslinking with anti-cd3 + anti-cd28 + anti-cd160. (a) Flow cytometry of anti-cd160 (5D.10A11) binding
Supplementary Information Supplementary Figure 1. CD4 + T cell activation and lack of apoptosis after crosslinking with anti-cd3 + anti-cd28 + anti-cd160. (a) Flow cytometry of anti-cd160 (5D.10A11) binding
SUPPLEMENTARY INFORMATION
 Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
Protocol for Gene Transfection & Western Blotting
 The schedule and the manual of basic techniques for cell culture Advanced Protocol for Gene Transfection & Western Blotting Schedule Day 1 26/07/2008 Transfection Day 3 28/07/2008 Cell lysis Immunoprecipitation
The schedule and the manual of basic techniques for cell culture Advanced Protocol for Gene Transfection & Western Blotting Schedule Day 1 26/07/2008 Transfection Day 3 28/07/2008 Cell lysis Immunoprecipitation
EBV essential antigen EBNA3C attenuates H2AX expression
 JVI Accepts, published online ahead of print on 15 January 2014 J. Virol. doi:10.1128/jvi.03568-13 Copyright 2014, American Society for Microbiology. All Rights Reserved. 1 EBV essential antigen EBNA3C
JVI Accepts, published online ahead of print on 15 January 2014 J. Virol. doi:10.1128/jvi.03568-13 Copyright 2014, American Society for Microbiology. All Rights Reserved. 1 EBV essential antigen EBNA3C
Supplementary Figure 1.TRIM33 binds β-catenin in the nucleus. a & b, Co-IP of endogenous TRIM33 with β-catenin in HT-29 cells (a) and HEK 293T cells
 and HEK 293T cells") Supplementary Figure 1.TRIM33 binds β-catenin in the nucleus. a & b, Co-IP of endogenous TRIM33 with β-catenin in HT-29 cells (a) and HEK 293T cells (b). TRIM33 was immunoprecipitated, and the amount of
Supplementary Figure 1.TRIM33 binds β-catenin in the nucleus. a & b, Co-IP of endogenous TRIM33 with β-catenin in HT-29 cells (a) and HEK 293T cells (b). TRIM33 was immunoprecipitated, and the amount of
Received 6 May 2003/Accepted 4 September 2003
 JOURNAL OF VIROLOGY, Dec. 2003, p. 12494 12506 Vol. 77, No. 23 0022-538X/03/$08.00 0 DOI: 10.1128/JVI.77.23.12494 12506.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved. ORF73
JOURNAL OF VIROLOGY, Dec. 2003, p. 12494 12506 Vol. 77, No. 23 0022-538X/03/$08.00 0 DOI: 10.1128/JVI.77.23.12494 12506.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved. ORF73
MicroRNA sponges: competitive inhibitors of small RNAs in mammalian cells
 MicroRNA sponges: competitive inhibitors of small RNAs in mammalian cells Margaret S Ebert, Joel R Neilson & Phillip A Sharp Supplementary figures and text: Supplementary Figure 1. Effect of sponges on
MicroRNA sponges: competitive inhibitors of small RNAs in mammalian cells Margaret S Ebert, Joel R Neilson & Phillip A Sharp Supplementary figures and text: Supplementary Figure 1. Effect of sponges on
Materials and Methods , The two-hybrid principle.
 The enzymatic activity of an unknown protein which cleaves the phosphodiester bond between the tyrosine residue of a viral protein and the 5 terminus of the picornavirus RNA Introduction Every day there
The enzymatic activity of an unknown protein which cleaves the phosphodiester bond between the tyrosine residue of a viral protein and the 5 terminus of the picornavirus RNA Introduction Every day there
HIV-1 Virus-like Particle Budding Assay Nathan H Vande Burgt, Luis J Cocka * and Paul Bates
 HIV-1 Virus-like Particle Budding Assay Nathan H Vande Burgt, Luis J Cocka * and Paul Bates Department of Microbiology, Perelman School of Medicine at the University of Pennsylvania, Philadelphia, USA
HIV-1 Virus-like Particle Budding Assay Nathan H Vande Burgt, Luis J Cocka * and Paul Bates Department of Microbiology, Perelman School of Medicine at the University of Pennsylvania, Philadelphia, USA
Polyomaviridae. Spring
 Polyomaviridae Spring 2002 331 Antibody Prevalence for BK & JC Viruses Spring 2002 332 Polyoma Viruses General characteristics Papovaviridae: PA - papilloma; PO - polyoma; VA - vacuolating agent a. 45nm
Polyomaviridae Spring 2002 331 Antibody Prevalence for BK & JC Viruses Spring 2002 332 Polyoma Viruses General characteristics Papovaviridae: PA - papilloma; PO - polyoma; VA - vacuolating agent a. 45nm
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION doi:1.138/nature9814 a A SHARPIN FL B SHARPIN ΔNZF C SHARPIN T38L, F39V b His-SHARPIN FL -1xUb -2xUb -4xUb α-his c Linear 4xUb -SHARPIN FL -SHARPIN TF_LV -SHARPINΔNZF -SHARPIN
SUPPLEMENTARY INFORMATION doi:1.138/nature9814 a A SHARPIN FL B SHARPIN ΔNZF C SHARPIN T38L, F39V b His-SHARPIN FL -1xUb -2xUb -4xUb α-his c Linear 4xUb -SHARPIN FL -SHARPIN TF_LV -SHARPINΔNZF -SHARPIN
Supplementary data Supplementary Figure 1 Supplementary Figure 2
 Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
Structural vs. nonstructural proteins
 Why would you want to study proteins associated with viruses or virus infection? Receptors Mechanism of uncoating How is gene expression carried out, exclusively by viral enzymes? Gene expression phases?
Why would you want to study proteins associated with viruses or virus infection? Receptors Mechanism of uncoating How is gene expression carried out, exclusively by viral enzymes? Gene expression phases?
T H E J O U R N A L O F C E L L B I O L O G Y
 T H E J O U R N A L O F C E L L B I O L O G Y Supplemental material Krenn et al., http://www.jcb.org/cgi/content/full/jcb.201110013/dc1 Figure S1. Levels of expressed proteins and demonstration that C-terminal
T H E J O U R N A L O F C E L L B I O L O G Y Supplemental material Krenn et al., http://www.jcb.org/cgi/content/full/jcb.201110013/dc1 Figure S1. Levels of expressed proteins and demonstration that C-terminal
SUPPLEMENTARY INFORMATION. Supplementary Figures S1-S9. Supplementary Methods
 SUPPLEMENTARY INFORMATION SUMO1 modification of PTEN regulates tumorigenesis by controlling its association with the plasma membrane Jian Huang 1,2#, Jie Yan 1,2#, Jian Zhang 3#, Shiguo Zhu 1, Yanli Wang
SUPPLEMENTARY INFORMATION SUMO1 modification of PTEN regulates tumorigenesis by controlling its association with the plasma membrane Jian Huang 1,2#, Jie Yan 1,2#, Jian Zhang 3#, Shiguo Zhu 1, Yanli Wang
Kaposi s Sarcoma-Associated Herpesvirus-Encoded Latency-Associated Nuclear Antigen Induces Chromosomal Instability through Inhibition of p53 Function
 JOURNAL OF VIROLOGY, Jan. 2006, p. 697 709 Vol. 80, No. 2 0022-538X/06/$08.00 0 doi:10.1128/jvi.80.2.697 709.2006 Copyright 2006, American Society for Microbiology. All Rights Reserved. Kaposi s Sarcoma-Associated
JOURNAL OF VIROLOGY, Jan. 2006, p. 697 709 Vol. 80, No. 2 0022-538X/06/$08.00 0 doi:10.1128/jvi.80.2.697 709.2006 Copyright 2006, American Society for Microbiology. All Rights Reserved. Kaposi s Sarcoma-Associated
The Schedule and the Manual of Basic Techniques for Cell Culture
 The Schedule and the Manual of Basic Techniques for Cell Culture 1 Materials Calcium Phosphate Transfection Kit: Invitrogen Cat.No.K2780-01 Falcon tube (Cat No.35-2054:12 x 75 mm, 5 ml tube) Cell: 293
The Schedule and the Manual of Basic Techniques for Cell Culture 1 Materials Calcium Phosphate Transfection Kit: Invitrogen Cat.No.K2780-01 Falcon tube (Cat No.35-2054:12 x 75 mm, 5 ml tube) Cell: 293
p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO
 Supplementary Information p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO Yuri Shibata, Masaaki Oyama, Hiroko Kozuka-Hata, Xiao Han, Yuetsu Tanaka,
Supplementary Information p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO Yuri Shibata, Masaaki Oyama, Hiroko Kozuka-Hata, Xiao Han, Yuetsu Tanaka,
(Stratagene, La Jolla, CA) (Supplemental Fig. 1A). A 5.4-kb EcoRI fragment
 (Supplemental Fig. 1A). A 5.4-kb EcoRI fragment") SUPPLEMENTAL INFORMATION Supplemental Methods Generation of RyR2-S2808D Mice Murine genomic RyR2 clones were isolated from a 129/SvEvTacfBR λ-phage library (Stratagene, La Jolla, CA) (Supplemental Fig.
SUPPLEMENTAL INFORMATION Supplemental Methods Generation of RyR2-S2808D Mice Murine genomic RyR2 clones were isolated from a 129/SvEvTacfBR λ-phage library (Stratagene, La Jolla, CA) (Supplemental Fig.
Supplementary Material
 Supplementary Material Nuclear import of purified HIV-1 Integrase. Integrase remains associated to the RTC throughout the infection process until provirus integration occurs and is therefore one likely
Supplementary Material Nuclear import of purified HIV-1 Integrase. Integrase remains associated to the RTC throughout the infection process until provirus integration occurs and is therefore one likely
Received 28 June 2004/Accepted 29 September 2004
 JOURNAL OF VIROLOGY, Mar. 2005, p. 3127 3138 Vol. 79, No. 5 0022-538X/05/$08.00 0 doi:10.1128/jvi.79.5.3127 3138.2005 Copyright 2005, American Society for Microbiology. All Rights Reserved. The Latency-Associated
JOURNAL OF VIROLOGY, Mar. 2005, p. 3127 3138 Vol. 79, No. 5 0022-538X/05/$08.00 0 doi:10.1128/jvi.79.5.3127 3138.2005 Copyright 2005, American Society for Microbiology. All Rights Reserved. The Latency-Associated
Herpesviruses. Virion. Genome. Genes and proteins. Viruses and hosts. Diseases. Distinctive characteristics
 Herpesviruses Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Virion Enveloped icosahedral capsid (T=16), diameter 125 nm Diameter of enveloped virion 200 nm Capsid
Herpesviruses Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Virion Enveloped icosahedral capsid (T=16), diameter 125 nm Diameter of enveloped virion 200 nm Capsid
Chromatin IP (Isw2) Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles.
 Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles.") Chromatin IP (Isw2) 7/01 Toshi last update: 06/15 Reagents Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles. 2.5 M glycine. TBS:
Chromatin IP (Isw2) 7/01 Toshi last update: 06/15 Reagents Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles. 2.5 M glycine. TBS:
Received 25 November 2003/Accepted 7 April 2004
 JOURNAL OF VIROLOGY, Aug. 2004, p. 8615 8629 Vol. 78, No. 16 0022-538X/04/$08.00 0 DOI: 10.1128/JVI.78.16.8615 8629.2004 Copyright 2004, American Society for Microbiology. All Rights Reserved. Kaposi s
JOURNAL OF VIROLOGY, Aug. 2004, p. 8615 8629 Vol. 78, No. 16 0022-538X/04/$08.00 0 DOI: 10.1128/JVI.78.16.8615 8629.2004 Copyright 2004, American Society for Microbiology. All Rights Reserved. Kaposi s
(a) Schematic diagram of the FS mutation of UVRAG in exon 8 containing the highly instable
 Schematic diagram of the FS mutation of UVRAG in exon 8 containing the highly instable") Supplementary Figure 1. Frameshift (FS) mutation in UVRAG. (a) Schematic diagram of the FS mutation of UVRAG in exon 8 containing the highly instable A 10 DNA repeat, generating a premature stop codon
Supplementary Figure 1. Frameshift (FS) mutation in UVRAG. (a) Schematic diagram of the FS mutation of UVRAG in exon 8 containing the highly instable A 10 DNA repeat, generating a premature stop codon
Supplemental Materials and Methods Plasmids and viruses Quantitative Reverse Transcription PCR Generation of molecular standard for quantitative PCR
 Supplemental Materials and Methods Plasmids and viruses To generate pseudotyped viruses, the previously described recombinant plasmids pnl4-3-δnef-gfp or pnl4-3-δ6-drgfp and a vector expressing HIV-1 X4
Supplemental Materials and Methods Plasmids and viruses To generate pseudotyped viruses, the previously described recombinant plasmids pnl4-3-δnef-gfp or pnl4-3-δ6-drgfp and a vector expressing HIV-1 X4
Supplementary Information. Supplementary Figure 1
 Supplementary Information Supplementary Figure 1 1 Supplementary Figure 1. Functional assay of the hcas9-2a-mcherry construct (a) Gene correction of a mutant EGFP reporter cell line mediated by hcas9 or
Supplementary Information Supplementary Figure 1 1 Supplementary Figure 1. Functional assay of the hcas9-2a-mcherry construct (a) Gene correction of a mutant EGFP reporter cell line mediated by hcas9 or
Nature Methods: doi: /nmeth Supplementary Figure 1
 Supplementary Figure 1 Subtiligase-catalyzed ligations with ubiquitin thioesters and 10-mer biotinylated peptides. (a) General scheme for ligations between ubiquitin thioesters and 10-mer, biotinylated
Supplementary Figure 1 Subtiligase-catalyzed ligations with ubiquitin thioesters and 10-mer biotinylated peptides. (a) General scheme for ligations between ubiquitin thioesters and 10-mer, biotinylated
Supplementary Information POLO-LIKE KINASE 1 FACILITATES LOSS OF PTEN-INDUCED PROSTATE CANCER FORMATION
 Supplementary Information POLO-LIKE KINASE 1 FACILITATES LOSS OF PTEN-INDUCED PROSTATE CANCER FORMATION X. Shawn Liu 1, 3, Bing Song 2, 3, Bennett D. Elzey 3, 4, Timothy L. Ratliff 3, 4, Stephen F. Konieczny
Supplementary Information POLO-LIKE KINASE 1 FACILITATES LOSS OF PTEN-INDUCED PROSTATE CANCER FORMATION X. Shawn Liu 1, 3, Bing Song 2, 3, Bennett D. Elzey 3, 4, Timothy L. Ratliff 3, 4, Stephen F. Konieczny
Supplementary Figure 1 Role of Raf-1 in TLR2-Dectin-1-mediated cytokine expression
 Supplementary Figure 1 Supplementary Figure 1 Role of Raf-1 in TLR2-Dectin-1-mediated cytokine expression. Quantitative real-time PCR of indicated mrnas in DCs stimulated with TLR2-Dectin-1 agonist zymosan
Supplementary Figure 1 Supplementary Figure 1 Role of Raf-1 in TLR2-Dectin-1-mediated cytokine expression. Quantitative real-time PCR of indicated mrnas in DCs stimulated with TLR2-Dectin-1 agonist zymosan
Supplementary information. MARCH8 inhibits HIV-1 infection by reducing virion incorporation of envelope glycoproteins
 Supplementary information inhibits HIV-1 infection by reducing virion incorporation of envelope glycoproteins Takuya Tada, Yanzhao Zhang, Takayoshi Koyama, Minoru Tobiume, Yasuko Tsunetsugu-Yokota, Shoji
Supplementary information inhibits HIV-1 infection by reducing virion incorporation of envelope glycoproteins Takuya Tada, Yanzhao Zhang, Takayoshi Koyama, Minoru Tobiume, Yasuko Tsunetsugu-Yokota, Shoji
7.012 Quiz 3 Answers
 MIT Biology Department 7.012: Introductory Biology - Fall 2004 Instructors: Professor Eric Lander, Professor Robert A. Weinberg, Dr. Claudette Gardel Friday 11/12/04 7.012 Quiz 3 Answers A > 85 B 72-84
MIT Biology Department 7.012: Introductory Biology - Fall 2004 Instructors: Professor Eric Lander, Professor Robert A. Weinberg, Dr. Claudette Gardel Friday 11/12/04 7.012 Quiz 3 Answers A > 85 B 72-84
Soft Agar Assay. For each cell pool, 100,000 cells were resuspended in 0.35% (w/v)
") SUPPLEMENTARY MATERIAL AND METHODS Soft Agar Assay. For each cell pool, 100,000 cells were resuspended in 0.35% (w/v) top agar (LONZA, SeaKem LE Agarose cat.5004) and plated onto 0.5% (w/v) basal agar.
SUPPLEMENTARY MATERIAL AND METHODS Soft Agar Assay. For each cell pool, 100,000 cells were resuspended in 0.35% (w/v) top agar (LONZA, SeaKem LE Agarose cat.5004) and plated onto 0.5% (w/v) basal agar.
Tel: ; Fax: ;
 Tel.: +98 216 696 9291; Fax: +98 216 696 9291; E-mail: mrasadeghi@pasteur.ac.ir Tel: +98 916 113 7679; Fax: +98 613 333 6380; E-mail: abakhshi_e@ajums.ac.ir A Soluble Chromatin-bound MOI 0 1 5 0 1 5 HDAC2
Tel.: +98 216 696 9291; Fax: +98 216 696 9291; E-mail: mrasadeghi@pasteur.ac.ir Tel: +98 916 113 7679; Fax: +98 613 333 6380; E-mail: abakhshi_e@ajums.ac.ir A Soluble Chromatin-bound MOI 0 1 5 0 1 5 HDAC2
Antibodies: LB1 buffer For 50 ml For 10ml For 30 ml Final 1 M HEPES, ph 2.5 ml 0.5 ml 1.5 ml 50mM. 5 M NaCl 1.4 ml 280 µl 0.
 Experiment: Date: Tissue: Purpose: ChIP-Seq Antibodies: 11x cross-link buffer: Regent Stock Solution Final Vol for 10 ml of 11xstock concentration 5 M NaCl 0.1M 0.2 ml 0.5 M EDTA 1 mm 20 ul 0.5 M EGTA,
Experiment: Date: Tissue: Purpose: ChIP-Seq Antibodies: 11x cross-link buffer: Regent Stock Solution Final Vol for 10 ml of 11xstock concentration 5 M NaCl 0.1M 0.2 ml 0.5 M EDTA 1 mm 20 ul 0.5 M EGTA,
Nature Medicine: doi: /nm.4322
 1 2 3 4 5 6 7 8 9 10 11 Supplementary Figure 1. Predicted RNA structure of 3 UTR and sequence alignment of deleted nucleotides. (a) Predicted RNA secondary structure of ZIKV 3 UTR. The stem-loop structure
1 2 3 4 5 6 7 8 9 10 11 Supplementary Figure 1. Predicted RNA structure of 3 UTR and sequence alignment of deleted nucleotides. (a) Predicted RNA secondary structure of ZIKV 3 UTR. The stem-loop structure
HEK293FT cells were transiently transfected with reporters, N3-ICD construct and
 Supplementary Information Luciferase reporter assay HEK293FT cells were transiently transfected with reporters, N3-ICD construct and increased amounts of wild type or kinase inactive EGFR. Transfections
Supplementary Information Luciferase reporter assay HEK293FT cells were transiently transfected with reporters, N3-ICD construct and increased amounts of wild type or kinase inactive EGFR. Transfections
For the 5 GATC-overhang two-oligo adaptors set up the following reactions in 96-well plate format:
 Supplementary Protocol 1. Adaptor preparation: For the 5 GATC-overhang two-oligo adaptors set up the following reactions in 96-well plate format: Per reaction X96 10X NEBuffer 2 10 µl 10 µl x 96 5 -GATC
Supplementary Protocol 1. Adaptor preparation: For the 5 GATC-overhang two-oligo adaptors set up the following reactions in 96-well plate format: Per reaction X96 10X NEBuffer 2 10 µl 10 µl x 96 5 -GATC
RayBio KinaseSTAR TM Akt Activity Assay Kit
 Activity Assay Kit User Manual Version 1.0 March 13, 2015 RayBio KinaseSTAR TM Akt Activity Kit Protocol (Cat#: 68AT-Akt-S40) RayBiotech, Inc. We Provide You With Excellent Support And Service Tel:(Toll
Activity Assay Kit User Manual Version 1.0 March 13, 2015 RayBio KinaseSTAR TM Akt Activity Kit Protocol (Cat#: 68AT-Akt-S40) RayBiotech, Inc. We Provide You With Excellent Support And Service Tel:(Toll
Conditional and reversible disruption of essential herpesvirus protein functions
 nature methods Conditional and reversible disruption of essential herpesvirus protein functions Mandy Glaß, Andreas Busche, Karen Wagner, Martin Messerle & Eva Maria Borst Supplementary figures and text:
nature methods Conditional and reversible disruption of essential herpesvirus protein functions Mandy Glaß, Andreas Busche, Karen Wagner, Martin Messerle & Eva Maria Borst Supplementary figures and text:
Impact of hyper-o-glcnacylation on apoptosis and NF-κB activity SUPPLEMENTARY METHODS
 SUPPLEMENTARY METHODS 3D culture and cell proliferation- MiaPaCa-2 cell culture in 3D was performed as described previously (1). Briefly, 8-well glass chamber slides were evenly coated with 50 µl/well
SUPPLEMENTARY METHODS 3D culture and cell proliferation- MiaPaCa-2 cell culture in 3D was performed as described previously (1). Briefly, 8-well glass chamber slides were evenly coated with 50 µl/well
Supplementary Figure 1. AdipoR1 silencing and overexpression controls. (a) Representative blots (upper and lower panels) showing the AdipoR1 protein
 Representative blots (upper and lower panels) showing the AdipoR1 protein") Supplementary Figure 1. AdipoR1 silencing and overexpression controls. (a) Representative blots (upper and lower panels) showing the AdipoR1 protein content relative to GAPDH in two independent experiments.
Supplementary Figure 1. AdipoR1 silencing and overexpression controls. (a) Representative blots (upper and lower panels) showing the AdipoR1 protein content relative to GAPDH in two independent experiments.
Supplementary Figure 1. SC35M polymerase activity in the presence of Bat or SC35M NP encoded from the phw2000 rescue plasmid.
 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 Supplementary Figure 1. SC35M polymerase activity in the presence of Bat or SC35M NP encoded from the phw2000 rescue plasmid. HEK293T
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 Supplementary Figure 1. SC35M polymerase activity in the presence of Bat or SC35M NP encoded from the phw2000 rescue plasmid. HEK293T
Supporting Online Material Material and Methods References Supplemental Figures S1, S2, and S3
 Supporting Online Material Material and Methods References Supplemental Figures S1, S2, and S3 Sarbassov et al. 1 Material and Methods Materials Reagents were obtained from the following sources: protein
Supporting Online Material Material and Methods References Supplemental Figures S1, S2, and S3 Sarbassov et al. 1 Material and Methods Materials Reagents were obtained from the following sources: protein
Supplementary Figure S1. Venn diagram analysis of mrna microarray data and mirna target analysis. (a) Western blot analysis of T lymphoblasts (CLS)
 Western blot analysis of T lymphoblasts (CLS)") Supplementary Figure S1. Venn diagram analysis of mrna microarray data and mirna target analysis. (a) Western blot analysis of T lymphoblasts (CLS) and their exosomes (EXO) in resting (REST) and activated
Supplementary Figure S1. Venn diagram analysis of mrna microarray data and mirna target analysis. (a) Western blot analysis of T lymphoblasts (CLS) and their exosomes (EXO) in resting (REST) and activated
MEK1 Assay Kit 1 Catalog # Lot # 16875
 MEK1 Assay Kit 1 Kit Components Assay Dilution Buffer (ADB), Catalog # 20-108. Three vials, each containing 1.0ml of assay dilution buffer (20mM MOPS, ph 7.2, 25mM ß-glycerol phosphate, 5mM EGTA, 1mM sodium
MEK1 Assay Kit 1 Kit Components Assay Dilution Buffer (ADB), Catalog # 20-108. Three vials, each containing 1.0ml of assay dilution buffer (20mM MOPS, ph 7.2, 25mM ß-glycerol phosphate, 5mM EGTA, 1mM sodium
Electron micrograph of phosphotungstanic acid-stained exosomes derived from murine
 1 SUPPLEMENTARY INFORMATION SUPPLEMENTARY FIGURES Supplementary Figure 1. Physical properties of murine DC-derived exosomes. a, Electron micrograph of phosphotungstanic acid-stained exosomes derived from
1 SUPPLEMENTARY INFORMATION SUPPLEMENTARY FIGURES Supplementary Figure 1. Physical properties of murine DC-derived exosomes. a, Electron micrograph of phosphotungstanic acid-stained exosomes derived from
Supplementary information
 Supplementary information Human Cytomegalovirus MicroRNA mir-us4-1 Inhibits CD8 + T Cell Response by Targeting ERAP1 Sungchul Kim, Sanghyun Lee, Jinwook Shin, Youngkyun Kim, Irini Evnouchidou, Donghyun
Supplementary information Human Cytomegalovirus MicroRNA mir-us4-1 Inhibits CD8 + T Cell Response by Targeting ERAP1 Sungchul Kim, Sanghyun Lee, Jinwook Shin, Youngkyun Kim, Irini Evnouchidou, Donghyun
~Lentivirus production~
 ~Lentivirus production~ May 30, 2008 RNAi core R&D group member Lentivirus Production Session Lentivirus!!! Is it health threatening to lab technician? What s so good about this RNAi library? How to produce
~Lentivirus production~ May 30, 2008 RNAi core R&D group member Lentivirus Production Session Lentivirus!!! Is it health threatening to lab technician? What s so good about this RNAi library? How to produce
Supplementary Figure 1. Normal T lymphocyte populations in Dapk -/- mice. (a) Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk
 Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk") Supplementary Figure 1. Normal T lymphocyte populations in Dapk -/- mice. (a) Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk -/- mice were stained for expression of CD4 and CD8.
Supplementary Figure 1. Normal T lymphocyte populations in Dapk -/- mice. (a) Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk -/- mice were stained for expression of CD4 and CD8.
Data Sheet TIGIT / NFAT Reporter - Jurkat Cell Line Catalog #60538
 Data Sheet TIGIT / NFAT Reporter - Jurkat Cell Line Catalog #60538 Background: TIGIT is a co-inhibitory receptor that is highly expressed in Natural Killer (NK) cells, activated CD4+, CD8+ and regulatory
Data Sheet TIGIT / NFAT Reporter - Jurkat Cell Line Catalog #60538 Background: TIGIT is a co-inhibitory receptor that is highly expressed in Natural Killer (NK) cells, activated CD4+, CD8+ and regulatory
Construction of a hepatocellular carcinoma cell line that stably expresses stathmin with a Ser25 phosphorylation site mutation
 Construction of a hepatocellular carcinoma cell line that stably expresses stathmin with a Ser25 phosphorylation site mutation J. Du 1, Z.H. Tao 2, J. Li 2, Y.K. Liu 3 and L. Gan 2 1 Department of Chemistry,
Construction of a hepatocellular carcinoma cell line that stably expresses stathmin with a Ser25 phosphorylation site mutation J. Du 1, Z.H. Tao 2, J. Li 2, Y.K. Liu 3 and L. Gan 2 1 Department of Chemistry,
Proteomic profiling of small-molecule inhibitors reveals dispensability of MTH1 for cancer cell survival
 Supplementary Information for Proteomic profiling of small-molecule inhibitors reveals dispensability of MTH1 for cancer cell survival Tatsuro Kawamura 1, Makoto Kawatani 1, Makoto Muroi, Yasumitsu Kondoh,
Supplementary Information for Proteomic profiling of small-molecule inhibitors reveals dispensability of MTH1 for cancer cell survival Tatsuro Kawamura 1, Makoto Kawatani 1, Makoto Muroi, Yasumitsu Kondoh,
Life Sciences 1A Midterm Exam 2. November 13, 2006
 Name: TF: Section Time Life Sciences 1A Midterm Exam 2 November 13, 2006 Please write legibly in the space provided below each question. You may not use calculators on this exam. We prefer that you use
Name: TF: Section Time Life Sciences 1A Midterm Exam 2 November 13, 2006 Please write legibly in the space provided below each question. You may not use calculators on this exam. We prefer that you use
MTC-TT and TPC-1 cell lines were cultured in RPMI medium (Gibco, Breda, The Netherlands)
") Supplemental data Materials and Methods Cell culture MTC-TT and TPC-1 cell lines were cultured in RPMI medium (Gibco, Breda, The Netherlands) supplemented with 15% or 10% (for TPC-1) fetal bovine serum
Supplemental data Materials and Methods Cell culture MTC-TT and TPC-1 cell lines were cultured in RPMI medium (Gibco, Breda, The Netherlands) supplemented with 15% or 10% (for TPC-1) fetal bovine serum
Inefficient establishment of KSHV latency suggests an additional role for continued lytic replication in Kaposi sarcoma pathogenesis
 Inefficient establishment of KSHV latency suggests an additional role for continued lytic replication in Kaposi sarcoma pathogenesis See the related Commentary beginning on page 21. Adam Grundhoff and
Inefficient establishment of KSHV latency suggests an additional role for continued lytic replication in Kaposi sarcoma pathogenesis See the related Commentary beginning on page 21. Adam Grundhoff and
Supplementary Figure 1. MAT IIα is Acetylated at Lysine 81.
 IP: Flag a Mascot PTM Modified Mass Error Position Gene Names Score Score Sequence m/z [ppm] 81 MAT2A;AMS2;MATA2 35.6 137.28 _AAVDYQK(ac)VVR_ 595.83-2.28 b Pre-immu After-immu Flag- WT K81R WT K81R / Flag
IP: Flag a Mascot PTM Modified Mass Error Position Gene Names Score Score Sequence m/z [ppm] 81 MAT2A;AMS2;MATA2 35.6 137.28 _AAVDYQK(ac)VVR_ 595.83-2.28 b Pre-immu After-immu Flag- WT K81R WT K81R / Flag
HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation
 SUPPLEMENTARY INFORMATION Materials and Methods Human cell lines and culture conditions HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation in exon 20 of BRCA1
SUPPLEMENTARY INFORMATION Materials and Methods Human cell lines and culture conditions HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation in exon 20 of BRCA1
T H E J O U R N A L O F C E L L B I O L O G Y
 T H E J O U R N A L O F C E L L B I O L O G Y Supplemental material Stelter et al., http://www.jcb.org/cgi/content/full/jcb.201105042/dc1 S1 Figure S1. Dyn2 recruitment to edid-labeled FG domain Nups.
T H E J O U R N A L O F C E L L B I O L O G Y Supplemental material Stelter et al., http://www.jcb.org/cgi/content/full/jcb.201105042/dc1 S1 Figure S1. Dyn2 recruitment to edid-labeled FG domain Nups.
SUPPLEMENTARY INFORMATION
 Figure S1 Induction of non-apoptotic death of SV40-transformed and primary DKO MEFs, and DKO thymocytes. (A-F) STS-induced non-apoptotic death of DKO MEF. (A, B) Reduced viability of DKO MEFs after exposure
Figure S1 Induction of non-apoptotic death of SV40-transformed and primary DKO MEFs, and DKO thymocytes. (A-F) STS-induced non-apoptotic death of DKO MEF. (A, B) Reduced viability of DKO MEFs after exposure
Chapter 3. Expression of α5-megfp in Mouse Cortical Neurons. on the β subunit. Signal sequences in the M3-M4 loop of β nachrs bind protein factors to
 22 Chapter 3 Expression of α5-megfp in Mouse Cortical Neurons Subcellular localization of the neuronal nachr subtypes α4β2 and α4β4 depends on the β subunit. Signal sequences in the M3-M4 loop of β nachrs
22 Chapter 3 Expression of α5-megfp in Mouse Cortical Neurons Subcellular localization of the neuronal nachr subtypes α4β2 and α4β4 depends on the β subunit. Signal sequences in the M3-M4 loop of β nachrs
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature12652 Supplementary Figure 1. PRDM16 interacts with endogenous EHMT1 in brown adipocytes. Immunoprecipitation of PRDM16 complex by flag antibody (M2) followed by Western blot analysis
doi:10.1038/nature12652 Supplementary Figure 1. PRDM16 interacts with endogenous EHMT1 in brown adipocytes. Immunoprecipitation of PRDM16 complex by flag antibody (M2) followed by Western blot analysis
Cell Lysis Buffer. Catalog number: AR0103
 Cell Lysis Buffer Catalog number: AR0103 Boster s Cell Lysis Buffer is a ready-to-use Western blot related reagent solution used for efficient extraction of total soluble protein in nondenatured state
Cell Lysis Buffer Catalog number: AR0103 Boster s Cell Lysis Buffer is a ready-to-use Western blot related reagent solution used for efficient extraction of total soluble protein in nondenatured state
TITLE: The Role of hcdc4 as a Tumor Suppressor Gene in Genomic Instability Underlying Prostate Cancer
 AD Award Number: TITLE: The Role of hcdc4 as a Tumor Suppressor Gene in Genomic Instability Underlying Prostate Cancer PRINCIPAL INVESTIGATOR: Audrey van Drogen, Ph.D. CONTRACTING ORGANIZATION: Sidney
AD Award Number: TITLE: The Role of hcdc4 as a Tumor Suppressor Gene in Genomic Instability Underlying Prostate Cancer PRINCIPAL INVESTIGATOR: Audrey van Drogen, Ph.D. CONTRACTING ORGANIZATION: Sidney
Mechanisms of alternative splicing regulation
 Mechanisms of alternative splicing regulation The number of mechanisms that are known to be involved in splicing regulation approximates the number of splicing decisions that have been analyzed in detail.
Mechanisms of alternative splicing regulation The number of mechanisms that are known to be involved in splicing regulation approximates the number of splicing decisions that have been analyzed in detail.
SUPPLEMENTARY INFORMATION
 Supplementary Discussion The cell cycle machinery and the DNA damage response network are highly interconnected and co-regulated in assuring faithful duplication and partition of genetic materials into
Supplementary Discussion The cell cycle machinery and the DNA damage response network are highly interconnected and co-regulated in assuring faithful duplication and partition of genetic materials into
SUPPLEMENTAL INFORMATION
 SUPPLEMENTAL INFORMATION EXPERIMENTAL PROCEDURES Tryptic digestion protection experiments - PCSK9 with Ab-3D5 (1:1 molar ratio) in 50 mm Tris, ph 8.0, 150 mm NaCl was incubated overnight at 4 o C. The
SUPPLEMENTAL INFORMATION EXPERIMENTAL PROCEDURES Tryptic digestion protection experiments - PCSK9 with Ab-3D5 (1:1 molar ratio) in 50 mm Tris, ph 8.0, 150 mm NaCl was incubated overnight at 4 o C. The
Chromatin Immunoprecipitation (ChIPs) Protocol (Mirmira Lab)
 Protocol (Mirmira Lab)") Chromatin Immunoprecipitation (ChIPs) Protocol (Mirmira Lab) Updated 12/3/02 Reagents: ChIP sonication Buffer (1% Triton X-100, 0.1% Deoxycholate, 50 mm Tris 8.1, 150 mm NaCl, 5 mm EDTA): 10 ml 10 % Triton
Chromatin Immunoprecipitation (ChIPs) Protocol (Mirmira Lab) Updated 12/3/02 Reagents: ChIP sonication Buffer (1% Triton X-100, 0.1% Deoxycholate, 50 mm Tris 8.1, 150 mm NaCl, 5 mm EDTA): 10 ml 10 % Triton
Auxin-Inducible Degron (AID) System Total Set
 System Total Set") Catalog No. BRS-APC011A Auxin-Inducible Degron (AID) System Total Set This AID System Charactoristics Principle of Auxin-Inducible Degron (AID ) System AID plasmid protein 1. Transfection of AID plasmid
Catalog No. BRS-APC011A Auxin-Inducible Degron (AID) System Total Set This AID System Charactoristics Principle of Auxin-Inducible Degron (AID ) System AID plasmid protein 1. Transfection of AID plasmid
Kaposi's Sarcoma-Associated Herpesvirus Induces ATM and H2AX. DNA Damage Response Early During de novo Infection of Primary
 JVI Accepts, published online ahead of print on 18 December 2013 J. Virol. doi:10.1128/jvi.03126-13 Copyright 2013, American Society for Microbiology. All Rights Reserved. JVI03126-13 Revised Manuscript
JVI Accepts, published online ahead of print on 18 December 2013 J. Virol. doi:10.1128/jvi.03126-13 Copyright 2013, American Society for Microbiology. All Rights Reserved. JVI03126-13 Revised Manuscript
Supplementary Figure 1. PD-L1 is glycosylated in cancer cells. (a) Western blot analysis of PD-L1 in breast cancer cells. (b) Western blot analysis
 Western blot analysis of PD-L1 in breast cancer cells. (b) Western blot analysis") Supplementary Figure 1. PD-L1 is glycosylated in cancer cells. (a) Western blot analysis of PD-L1 in breast cancer cells. (b) Western blot analysis of PD-L1 in ovarian cancer cells. (c) Western blot analysis
Supplementary Figure 1. PD-L1 is glycosylated in cancer cells. (a) Western blot analysis of PD-L1 in breast cancer cells. (b) Western blot analysis of PD-L1 in ovarian cancer cells. (c) Western blot analysis
Mammalian Membrane Protein Extraction Kit
 Mammalian Membrane Protein Extraction Kit Catalog number: AR0155 Boster s Mammalian Membrane Protein Extraction Kit is a simple, rapid and reproducible method to prepare cellular protein fractions highly
Mammalian Membrane Protein Extraction Kit Catalog number: AR0155 Boster s Mammalian Membrane Protein Extraction Kit is a simple, rapid and reproducible method to prepare cellular protein fractions highly
SUPPLEMENTARY INFORMATION
 Supplementary Table 1. Cell sphingolipids and S1P bound to endogenous TRAF2. Sphingolipid Cell pmol/mg TRAF2 immunoprecipitate pmol/mg Sphingomyelin 4200 ± 250 Not detected Monohexosylceramide 311 ± 18
Supplementary Table 1. Cell sphingolipids and S1P bound to endogenous TRAF2. Sphingolipid Cell pmol/mg TRAF2 immunoprecipitate pmol/mg Sphingomyelin 4200 ± 250 Not detected Monohexosylceramide 311 ± 18
Supplements. Figure S1. B Phalloidin Alexa488
 Supplements A, DMSO, PP2, PP3 Crk-myc Figure S1. (A) Src kinase activity is necessary for recruitment of Crk to Nephrin cytoplasmic domain. Human podocytes expressing /7-NephrinCD () were treated with
Supplements A, DMSO, PP2, PP3 Crk-myc Figure S1. (A) Src kinase activity is necessary for recruitment of Crk to Nephrin cytoplasmic domain. Human podocytes expressing /7-NephrinCD () were treated with
Islet viability assay and Glucose Stimulated Insulin Secretion assay RT-PCR and Western Blot
 Islet viability assay and Glucose Stimulated Insulin Secretion assay Islet cell viability was determined by colorimetric (3-(4,5-dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide assay using CellTiter
Islet viability assay and Glucose Stimulated Insulin Secretion assay Islet cell viability was determined by colorimetric (3-(4,5-dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide assay using CellTiter
Autoregulation of DNA Binding and Protein Stability of Kaposi s Sarcoma-Associated Herpesvirus ORF50 Protein
 JOURNAL OF VIROLOGY, Oct. 2004, p. 10657 10673 Vol. 78, No. 19 0022-538X/04/$08.00 0 DOI: 10.1128/JVI.78.19.10657 10673.2004 Copyright 2004, American Society for Microbiology. All Rights Reserved. Autoregulation
JOURNAL OF VIROLOGY, Oct. 2004, p. 10657 10673 Vol. 78, No. 19 0022-538X/04/$08.00 0 DOI: 10.1128/JVI.78.19.10657 10673.2004 Copyright 2004, American Society for Microbiology. All Rights Reserved. Autoregulation
Use of the red fluorescent protein as a marker of Kaposi s sarcoma-associated herpesvirus lytic gene expression
 Virology 325 (2004) 225 240 www.elsevier.com/locate/yviro Use of the red fluorescent protein as a marker of Kaposi s sarcoma-associated herpesvirus lytic gene expression Jeffrey Vieira* and Patricia M.
Virology 325 (2004) 225 240 www.elsevier.com/locate/yviro Use of the red fluorescent protein as a marker of Kaposi s sarcoma-associated herpesvirus lytic gene expression Jeffrey Vieira* and Patricia M.
Instructions for Use. APO-AB Annexin V-Biotin Apoptosis Detection Kit 100 tests
 3URGXFW,QIRUPDWLRQ Sigma TACS Annexin V Apoptosis Detection Kits Instructions for Use APO-AB Annexin V-Biotin Apoptosis Detection Kit 100 tests For Research Use Only. Not for use in diagnostic procedures.
3URGXFW,QIRUPDWLRQ Sigma TACS Annexin V Apoptosis Detection Kits Instructions for Use APO-AB Annexin V-Biotin Apoptosis Detection Kit 100 tests For Research Use Only. Not for use in diagnostic procedures.
Luminescent platforms for monitoring changes in the solubility of amylin and huntingtin in living cells
 Electronic Supplementary Material (ESI) for Molecular BioSystems. This journal is The Royal Society of Chemistry 2016 Contents Supporting Information Luminescent platforms for monitoring changes in the
Electronic Supplementary Material (ESI) for Molecular BioSystems. This journal is The Royal Society of Chemistry 2016 Contents Supporting Information Luminescent platforms for monitoring changes in the
RAW264.7 cells stably expressing control shrna (Con) or GSK3b-specific shrna (sh-
 or GSK3b-specific shrna (sh-") 1 a b Supplementary Figure 1. Effects of GSK3b knockdown on poly I:C-induced cytokine production. RAW264.7 cells stably expressing control shrna (Con) or GSK3b-specific shrna (sh- GSK3b) were stimulated
1 a b Supplementary Figure 1. Effects of GSK3b knockdown on poly I:C-induced cytokine production. RAW264.7 cells stably expressing control shrna (Con) or GSK3b-specific shrna (sh- GSK3b) were stimulated
Protocol for purification of recombinant protein from 300 ml yeast culture
 Protocol for purification of recombinant protein from 300 ml yeast culture Equipment and reagents needed: Zirconia beads (0.5 mm diameter from BSP, Germany) Paint Shaker (at 4 C) Tube rotator for 15 ml
Protocol for purification of recombinant protein from 300 ml yeast culture Equipment and reagents needed: Zirconia beads (0.5 mm diameter from BSP, Germany) Paint Shaker (at 4 C) Tube rotator for 15 ml
A Hepatocyte Growth Factor Receptor (Met) Insulin Receptor hybrid governs hepatic glucose metabolism SUPPLEMENTARY FIGURES, LEGENDS AND METHODS
 Insulin Receptor hybrid governs hepatic glucose metabolism SUPPLEMENTARY FIGURES, LEGENDS AND METHODS") A Hepatocyte Growth Factor Receptor (Met) Insulin Receptor hybrid governs hepatic glucose metabolism Arlee Fafalios, Jihong Ma, Xinping Tan, John Stoops, Jianhua Luo, Marie C. DeFrances and Reza Zarnegar
A Hepatocyte Growth Factor Receptor (Met) Insulin Receptor hybrid governs hepatic glucose metabolism Arlee Fafalios, Jihong Ma, Xinping Tan, John Stoops, Jianhua Luo, Marie C. DeFrances and Reza Zarnegar
Product Manual. Omni-Array Sense Strand mrna Amplification Kit, 2 ng to 100 ng Version Catalog No.: Reactions
 Genetic Tools and Reagents Universal mrna amplification, sense strand amplification, antisense amplification, cdna synthesis, micro arrays, gene expression, human, mouse, rat, guinea pig, cloning Omni-Array
Genetic Tools and Reagents Universal mrna amplification, sense strand amplification, antisense amplification, cdna synthesis, micro arrays, gene expression, human, mouse, rat, guinea pig, cloning Omni-Array
Supplementary Fig. S1. Schematic diagram of minigenome segments.
 open reading frame 1565 (segment 5) 47 (-) 3 5 (+) 76 101 125 149 173 197 221 246 287 open reading frame 890 (segment 8) 60 (-) 3 5 (+) 172 Supplementary Fig. S1. Schematic diagram of minigenome segments.
open reading frame 1565 (segment 5) 47 (-) 3 5 (+) 76 101 125 149 173 197 221 246 287 open reading frame 890 (segment 8) 60 (-) 3 5 (+) 172 Supplementary Fig. S1. Schematic diagram of minigenome segments.
Supplementary material: Materials and suppliers
 Supplementary material: Materials and suppliers Electrophoresis consumables including tris-glycine, acrylamide, SDS buffer and Coomassie Brilliant Blue G-2 dye (CBB) were purchased from Ameresco (Solon,
Supplementary material: Materials and suppliers Electrophoresis consumables including tris-glycine, acrylamide, SDS buffer and Coomassie Brilliant Blue G-2 dye (CBB) were purchased from Ameresco (Solon,
Inhibition of trna 3 Lys -Primed Reverse Transcription by Human APOBEC3G during Human Immunodeficiency Virus Type 1 Replication
 JOURNAL OF VIROLOGY, Dec. 2006, p. 11710 11722 Vol. 80, No. 23 0022-538X/06/$08.00 0 doi:10.1128/jvi.01038-06 Copyright 2006, American Society for Microbiology. All Rights Reserved. Inhibition of trna
JOURNAL OF VIROLOGY, Dec. 2006, p. 11710 11722 Vol. 80, No. 23 0022-538X/06/$08.00 0 doi:10.1128/jvi.01038-06 Copyright 2006, American Society for Microbiology. All Rights Reserved. Inhibition of trna
Sestrin2 and BNIP3 (Bcl-2/adenovirus E1B 19kDa-interacting. protein3) regulate autophagy and mitophagy in renal tubular cells in. acute kidney injury
 regulate autophagy and mitophagy in renal tubular cells in. acute kidney injury") Sestrin2 and BNIP3 (Bcl-2/adenovirus E1B 19kDa-interacting protein3) regulate autophagy and mitophagy in renal tubular cells in acute kidney injury by Masayuki Ishihara 1, Madoka Urushido 2, Kazu Hamada
Sestrin2 and BNIP3 (Bcl-2/adenovirus E1B 19kDa-interacting protein3) regulate autophagy and mitophagy in renal tubular cells in acute kidney injury by Masayuki Ishihara 1, Madoka Urushido 2, Kazu Hamada
Essential Medium, containing 10% fetal bovine serum, 100 U/ml penicillin and 100 µg/ml streptomycin. Huvec were cultured in
 Supplemental data Methods Cell culture media formulations A-431 and U-87 MG cells were maintained in Dulbecco s Modified Eagle s Medium. FaDu cells were cultured in Eagle's Minimum Essential Medium, containing
Supplemental data Methods Cell culture media formulations A-431 and U-87 MG cells were maintained in Dulbecco s Modified Eagle s Medium. FaDu cells were cultured in Eagle's Minimum Essential Medium, containing
Transcription Program of Murine Gammaherpesvirus 68
 JOURNAL OF VIROLOGY, Oct. 2003, p. 10488 10503 Vol. 77, No. 19 0022-538X/03/$08.00 0 DOI: 10.1128/JVI.77.19.10488 10503.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved. Transcription
JOURNAL OF VIROLOGY, Oct. 2003, p. 10488 10503 Vol. 77, No. 19 0022-538X/03/$08.00 0 DOI: 10.1128/JVI.77.19.10488 10503.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved. Transcription
Graveley Lab shrna knockdown followed by RNA-seq Biosample Preparation and Characterization Document
 Graveley Lab shrna knockdown followed by RNA-seq Biosample Preparation and Characterization Document Wet Lab: Sara Olson and Lijun Zhan Computational Lab: Xintao Wei and Michael Duff PI: Brenton Graveley
Graveley Lab shrna knockdown followed by RNA-seq Biosample Preparation and Characterization Document Wet Lab: Sara Olson and Lijun Zhan Computational Lab: Xintao Wei and Michael Duff PI: Brenton Graveley
SUPPLEMENT. Materials and methods
 SUPPLEMENT Materials and methods Cell culture and reagents Cell media and reagents were from Invitrogen unless otherwise indicated. Antibiotics and Tet-certified serum were from Clontech. In experiments
SUPPLEMENT Materials and methods Cell culture and reagents Cell media and reagents were from Invitrogen unless otherwise indicated. Antibiotics and Tet-certified serum were from Clontech. In experiments
Serum Amyloid A3 Gene Expression in Adipocytes is an Indicator. of the Interaction with Macrophages
 Serum Amyloid A3 Gene Expression in Adipocytes is an Indicator of the Interaction with Macrophages Yohei Sanada, Takafumi Yamamoto, Rika Satake, Akiko Yamashita, Sumire Kanai, Norihisa Kato, Fons AJ van
Serum Amyloid A3 Gene Expression in Adipocytes is an Indicator of the Interaction with Macrophages Yohei Sanada, Takafumi Yamamoto, Rika Satake, Akiko Yamashita, Sumire Kanai, Norihisa Kato, Fons AJ van
Activation of p90 Ribosomal S6 Kinase by ORF45 of Kaposi s Sarcoma-Associated Herpesvirus and Its Role in Viral Lytic Replication
 JOURNAL OF VIROLOGY, Feb. 2008, p. 1838 1850 Vol. 82, No. 4 0022-538X/08/$08.00 0 doi:10.1128/jvi.02119-07 Copyright 2008, American Society for Microbiology. All Rights Reserved. Activation of p90 Ribosomal
JOURNAL OF VIROLOGY, Feb. 2008, p. 1838 1850 Vol. 82, No. 4 0022-538X/08/$08.00 0 doi:10.1128/jvi.02119-07 Copyright 2008, American Society for Microbiology. All Rights Reserved. Activation of p90 Ribosomal
hexahistidine tagged GRP78 devoid of the KDEL motif (GRP78-His) on SDS-PAGE. This
 on SDS-PAGE. This") SUPPLEMENTAL FIGURE LEGEND Fig. S1. Generation and characterization of. (A) Coomassie staining of soluble hexahistidine tagged GRP78 devoid of the KDEL motif (GRP78-His) on SDS-PAGE. This protein was expressed
SUPPLEMENTAL FIGURE LEGEND Fig. S1. Generation and characterization of. (A) Coomassie staining of soluble hexahistidine tagged GRP78 devoid of the KDEL motif (GRP78-His) on SDS-PAGE. This protein was expressed
For the rapid, sensitive and accurate quantification of Ras in various samples
 ab128504 Ras Assay Kit Instructions for Use For the rapid, sensitive and accurate quantification of Ras in various samples This product is for research use only and is not intended for diagnostic use.
ab128504 Ras Assay Kit Instructions for Use For the rapid, sensitive and accurate quantification of Ras in various samples This product is for research use only and is not intended for diagnostic use.
Supplementary Appendix
 Supplementary Appendix This appendix has been provided by the authors to give readers additional information about their work. Supplement to: Choi YL, Soda M, Yamashita Y, et al. EML4-ALK mutations in
Supplementary Appendix This appendix has been provided by the authors to give readers additional information about their work. Supplement to: Choi YL, Soda M, Yamashita Y, et al. EML4-ALK mutations in
Kaposi s Sarcoma-Associated Herpesvirus Transactivator RTA Promotes Degradation of the Repressors To RegulateViral Lytic Replication
 University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Virology Papers Virology, Nebraska Center for 4-2008 Kaposi s Sarcoma-Associated Herpesvirus Transactivator RTA Promotes
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Virology Papers Virology, Nebraska Center for 4-2008 Kaposi s Sarcoma-Associated Herpesvirus Transactivator RTA Promotes