Functional Properties of the Human Cytomegalovirus IE86 Protein Required for. Transcriptional Regulation and Virus Replication. Julie A.
|
|
|
- Alexina Georgiana Parker
- 6 years ago
- Views:
Transcription
1 JVI Accepts, published online ahead of print on 16 June 2010 J. Virol. doi: /jvi Copyright 2010, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved. Harris, et al./page Functional Properties of the Human Cytomegalovirus IE86 Protein Required for Transcriptional Regulation and Virus Replication Siabhon M. Harris 1, Brady Bullock 1, Elizabeth Westgard 1, Hua Zhu 2, Richard M. Stenberg 1 and Julie A. Kerry 1 Department of Microbiology and Molecular Cell Biology, Eastern Virginia Medical School, Norfolk, VA and Department of Microbiology and Molecular Genetics, New Jersey Medical School, University of Medicine and Dentistry of New Jersey, Newark, NJ 2 Running Title: HCMV IE86 Functional Analysis Manuscript word count: 5,445 Abstract word count: 229 words Corresponding Author: Julie Anne Kerry, Ph.D. Eastern Virginia Medical School 700 West Olney Rd, Lewis Hall Norfolk, VA Phone: (757) Fax: (757) kerryja@evms.edu
2 Harris, et al./page ABSTRACT The human cytomegalovirus (HCMV) IE86 protein is essential for HCMV replication due to its ability to transactivate critical viral early promoters. In the current study, we performed a comprehensive mutational analysis between amino acids 535 and 545 of IE86 and assessed the impact of these mutations on IE86-mediated transcriptional activation. Using transient assays and complementing analysis in recombinant HCMV clones, we show that single amino acid mutations differentially impair the ability of IE86 to mediate transactivation of essential early gene promoters. The conserved tyrosine at amino acid 544 is critical for activation of the UL54 promoter in vitro and in the context of the viral genome. In contrast, mutation of the proline at position 535 disrupted activation of the UL54 promoter in transient assays, but displayed similar activity to wild type IE86 when assessed in the genomic context. To examine the underlying mechanism of this differential effect, GST pulldown assays were performed, revealing that Y544 is critical for binding to the TATA binding protein (TBP), suggesting that this interaction is likely necessary for the ability of IE86 to activate the UL54 promoter. In contrast, mutation of either P535 or Y544 disrupted activation of the UL promoter both in vitro and in vivo, suggesting that interaction with TBP is not sufficient for IE86-mediated activation of this early promoter. Together, these studies demonstrate that IE86 activates early promoters by distinct mechanisms.
3 Harris, et al./page 3 1 INTRODUCTION Human cytomegalovirus (HCMV), a member of the herpesviridae, is a ubiquitous and opportunistic pathogen that under conditions of immune immaturity or suppression is a major cause of disease and mortality (40). The first subset of viral genes to be expressed upon HCMV infection are the immediate early (IE) gene products (40). The major IE (MIE) proteins IE72 and IE86 are produced as a result of differential splicing from the UL gene region (40). Expression of these gene products is controlled by the MIE promoter (MIEP), which in turn is regulated by a complex interplay between viral and cellular proteins (38, 60). For example, the IE72 and pp71 viral proteins enhance activation of the MIEP leading to increased MIE gene products (11, 33, 62). In contrast, IE86 negatively regulates expression of the MIE gene products through direct binding to the cis repression sequence (CRS), located between the TATA box and the transcription start signal of the MIEP (10, 32, 47, 48). It is thought that IE86 inhibits transcription from the MIEP by blocking RNA polymerase recruitment and preventing assembly of the preinitiation complex (29). The 579 amino acid IE86 protein encoded by the IE2 transcript is a potent transactivator of viral and cellular genes and is essential for virus replication (18, 37, 61). The essential nature of the IE86 protein is due in part to a requirement for IE86 to activate the promoters of viral genes required for replication, including the UL112/113 replication proteins and the UL54 DNA polymerase (18, 24, 42, 53, 57, 59). Early promoter activation by IE86 is thought to be mediated by direct binding of IE86 to DNA and/or protein interactions with cellular and viral transcription factors (15, 61). For example, IE86 interacts in vitro with components of the basal transcription
4 machinery and transcriptional regulators such as TBP, SP1, and CREB (8, 16, 28, 51, 61, 64). It has been suggested that IE86 mediates transcriptional activation at a step following TBP recruitment to the promoter (25). However, these analyses relied on artificial recruitment of TBP to a generic promoter construct. In a series of elegant studies, Lukac et al demonstrated that IE86 activation required a functional TATA box and determined that IE86 had TAF-like functions (35, 36). These studies suggest that the interaction of IE86 with TBP and other basal promoter factors may be critical for transcriptional activation by this protein. In addition, IE86 interacts with histone modifiers such as P/CAF and HDAC3 suggesting that the modulation of chromatin modification facilitates transcriptional activation by IE86 (6, 44, 49). Indeed, HAT recruitment was found to be critical for the activation of viral early promoters (7). The IE86 protein also regulates early gene transcription by direct binding to DNA. For example, the promoter of the UL gene is regulated by an ATF/CREB binding site and an IE86 binding site (2, 53, 54). Deletion of the IE86 binding site causes a reduction of IE86-mediated transactivation of this promoter during infection (52, 53). The IE86 protein possesses a number of additional functions that are likely important for efficient virus replication, including the regulation of cell cycle progression, inhibition of host DNA synthesis and modulation of cytokine expression (61). As a result of the essential role of IE86 in virus replication, numerous studies have been performed to identify functional domains of the IE86 protein and assess the biological significance of these regions (reviewed in (61)). However, most of these analyses relied on the use of large-scale deletions within the IE86 protein raising the possibility that alterations in protein structure result that complicate interpretation of the data. Despite this caveat, a core domain in the carboxy-terminal region of IE86 from amino acid 450 to 544 has been identified
5 that overlaps regions important for DNA binding, dimerization and interaction with the basal transcription factors TBP and TFIIB (3, 61). Mutations within this domain disrupt both transactivation and autoregulation by IE86, suggesting that these two properties are linked (3). In support of this, research in our laboratory showed that insertion of 4 amino acids adjacent to the alanine residue at position 540 of IE86 caused an 8-fold decrease in activation of the UL54 promoter in transient assays with a corresponding 10-fold increase in MIEP activation (59). Similarly, studies by White et al. examined mutations or deletions in IE86 between amino acids 356 to 359, 427 to 435, and 505 to 511 within HCMV bacterial artificial chromosome (BAC) constructs (63). The resultant viruses were non-viable, correlating with a lack of activation of the UL44 and UL early genes during infection as well as a loss of repression of the MIEP (63). However, this study lacked corresponding data regarding the functional properties of IE86 that were impacted by these specific mutations. A more recent study showed that simultaneous mutation of amino acids 535 and 537 within IE86 results in a loss of early gene activation but the ability of IE86 to autoregulate the MIE promoter was retained (45), suggesting that in some cases, these properties are distinguishable. Further, this analysis demonstrated that the ability of IE86 to associate with viral early promoters correlated with transcriptional activation (45). However, this study did not directly assess the effects of single amino acid mutations on transcriptional regulation. Thus, it is possible that more than one functional property of IE86 was disrupted by the mutation. In the current work, we have performed a mutational analysis of ten individual amino acids within the core domain to assess the impact on transcriptional activation of viral early promoters involved in virus replication. These studies reveal that in all cases, activation correlated directly with autoregulation by IE86. In addition, assessment of two specific mutants in the context of the viral genome demonstrated differential
6 activation of the UL54 and UL early promoters, suggesting that more than one mechanism is involved in the regulation of these essential early genes by IE86. Further analysis showed that the interaction of IE86 with TBP appears to be critical for the activation of the UL54 early promoter
7 7 93 MATERIALS AND METHODS Cells and Plasmids Primary human foreskin fibroblasts (HFF) were obtained from Clonetics Corporation and maintained as previously described (55). HeLa cells were obtained from ATCC and grown in Iscove s Medium with 10% fetal bovine serum and 1% penicillin and streptomycin. The psvh plasmid used for expression of the MIE gene products (13) was modified by deletion of a Kpn I restriction enzyme site to generate psvhk. The pmcrs86 plasmid that contains the IE86 cdna sequence under the control of the MIE promoter lacking the cis repression sequence was kindly provided by J. A. Nelson (Oregon Health Sciences University). The cdna construct was corrected for IE86 mutations by the replacement of a Sma I-Bsu 36I fragment from psvhk as previously described (4). Mutagenesis of the IE86 coding sequence in the psvhk and pmcrs86 constructs was achieved using the Quikchange site-directed PCR mutagenesis kit XL (Stratagene). Primer sequences for individual mutations are shown in Table 1. All constructs were confirmed by DNA sequencing using the BigDye Terminator v3.1 Cycle Sequencing kit and analyzed on an ABI Primer 3130 Genetic Analyzer (Applied Biosystems). The psvod plasmid used as an empty vector control has been previously described (39). The reporter plasmids pmiep-luc and pul54-luc have also been described elsewhere (23, 55). The pul luc reporter plasmid was generated by PCR amplification of p729cat (57) kindly provided by D. H. Spector (University of California, San Diego) and subsequent cloning of the Hind III to Bgl II promoter fragment into the pgl3-basic plasmid (Promega).
8 Luciferase Assays and Western Blot analysis HFF cells were transfected with the indicated plasmids using either Lipofectamine 2000 (Invitrogen) or the DEAE-dextran technique (13, 43). At 48 hours after transfection, cell extracts were harvested and assessed for luciferase activity as described (55). Significance was determined by p<0.05 using a two-tailed Student s T-test. Western blot analysis of the cell extracts was performed using either the polyclonal peptide antibody 1218 that recognizes the IE86 protein (58) or the MAB810 monoclonal antibody (Millipore) essentially as previously described (12). In some cases, IRDye labeled secondary antibodies (IRDye 800CW goat antimouse IgG (Licor)) were used and the proteins detected using the Odyssey Infrared Imaging System (Licor). Protein loading was controlled for by western analysis using the actin antibody (A1978, Sigma). RNA Extraction and Real Time RT-PCR HFFs were transfected with 10 µg of total plasmid DNA by the DEAE-dextran method (13). Samples were harvested 48 hours post-transfection using the RNeasy Kit protocol (Qiagen). Cells transfected with 10 µg of BAC constructs by the calcium phosphate method (27) were harvested at the indicated times and RNA isolated using Trizol reagent (Invitrogen). In either case, samples were treated with DNAse I (Qiagen) and cdna generated using 1-5 µg of total RNA primed with 250 ng of random hexamers (Superscript, Invitrogen). Real time PCR was then performed using the iq SYBR green supermix (Invitrogen) using a Biorad icycler system and analyzed using the PFAFFL mathematical model (46). Primers used to amplify the IE2, UL and GAPDH genes have been previously described (63). The UL54 gene was amplified using the following primers: forward: 5'-CAGCCGCTGCAGAACCTCTTTC; reverse
9 '-CTGCTCGTTGGTGTAAGGCG. HCMV BAC mutagenesis BAC constructs were generated essentially as previously described (14, 41). To minimize the possibility of revertants, mutagenesis of IE86 was repeated using the following primer sequences (mutations in IE86 are underlined, forward primer shown): P535A-3, GTG GGT TCA TGC TGG CGA TCT ACG AGA CGG, and Y544A-2, CCA CGA AGG CCG CGG CCG TGG GGC. Deletion of the HCMV UL exon 5 region coding for IE86 specific sequences was accomplished as previously described (14, 41) using the following primers containing a 20 bp overhang with homology to the C-terminus of IE86 between amino acids 449 and 579: 5 CCA TGG CCC TCT CCA CTC CCT TCC TCA TGG AGC ACA CCA TGG CTC TTG TTG GCT AGT GCG TA 3 and 5 CAC TAT GTA CAA GAG TCC ATG TCT CTC TTT CCA GTT TTT CTC TGC CAG TGT TAC AAC CAA 3. This PCR fragment was gel-purified and transformed into electro- and recombination-competent DY380 cells containing the HCMV Towne-BAC to generate the HCMV Exon5 BAC. Positive clones containing successful deletion of the amino acids 449 to 579 and replacement of this region with the kanamycin resistance marker were confirmed by growth on LB agar plates containing kanamycin and chloramphenicol, PCR analysis and restriction enzyme digestion. To generate pie2-zeo for homologous recombination with the HCMV Exon5 BAC, the Zeocin gene was first amplified from pcmv/zeo (Invitrogen) using BamHI-LoxP-Zeo-LoxP- BglII forward and reverse primers: 5 -AGA TCT ATA ACT TCG TAT AGC ATA CAT TAT 160 ACG AAG TTA TGG AAC GGA CCG TGT TGA C-3 and 5 -GGA TCC ATA ACT TCG 161 TAT AGC ATA CAT TAT ACG AAG TTA TCA AGT TTC GAG GTC GAG GTG-3,
10 respectively and cloned into pcr4-topo (Invitrogen). Subsequently, wild type or mutant IE86 fragments were amplified from pmcrs86 using PstI-435-IE2-579-BamHI primers: 5 -CTG CAG AAC CTG GCC CTC TCC ACT C-3 and 5 -GGA TCC ACT TAC TGA GAC TTG TTC CTC AGG TCC-3 and the resultant fragment cloned into the pzeo-topo construct between the Pst I and Bam HI sites. The IE2-Zeo region was then amplified from pie2-zeo using the following primers: 5 CAA CCT GGC CCT CTC ACT CCC TTC CTC ATG GAG CAC ACC ATG CCC GTG ACA CAT 3 and 5 CGG GGA ATC ACT ATG TAC AAG AGT CCA TGT CTC TCT TTC CAG TTT TTC AGA TCT ATA ACT TCG TAT AAT G 3. This PCR fragment was gel-purified and transformed into electro- and recombination-competent DY380 cells containing the HCMV Exon5 BAC. Positive clones were confirmed by growth on LB agar plates containing zeocin and chloramphenicol, PCR analysis, southern blot analysis and restriction enzyme digestion. Southern blot analysis was performed as previously described (9). The following primers were used for PCR confirmation of HCMV BAC constructs: DS IE2 Primer: 5 GAT GTC TCG CAG GGT GGG TAG ATG 3, US IE2 Primer: 5 GCA TGT TCC GCA ACA CCA ATC G 3, Internal IE2 Primer 1: 5 AAG ACC TGG ACA CCC TGA GCC TG 3, and Internal IE2 Primer 2: 5 CAG GCT CAG GGT GTC CAG GTC TTC 3. Transfection of BAC DNA and Growth Curve Analysis Large scale BAC DNA purifications were performed using Clontech s Nucleobond BAC 100 kit according to the manufacturer s instructions. For BAC transfections in HFFs, 10 µg of BAC DNA and 1 µg of pcmvpp71 DNA (33) were transfected into HFF by the calcium
11 phosphate method (27). A plasmid expressing Cre Recombinase (1 µg) was also transfected with the BACs in DNA and virus replication assays (65). Media was changed every 4-5 days until plaque outgrowth occurred. Cells were monitored for 3 or more weeks for plaque formation. Reconstituted virus was harvested from the supernatant of BAC transfected cells when % of cells displayed cytopathic effects (CPE). Viral DNA replication was assessed by harvesting BAC-transfected cells at various time points using Trizol reagent (Invitrogen). Virus replication assays were performed as described in Lorz, et al (34). Briefly, 20 µl of viral supernatant was incubated with 160 µl of 10 mg/ml proteinase K for 1 hour at 56 C. DNA was denatured by incubation at 95 C for 5 minutes. Real time PCR analysis was performed as described above using equal amounts of total DNA (100 ng). The primer sequences used for amplification of gb have been previously described (45). GST-Pulldown Assays Wild type and mutant IE86 proteins were generated using the TnT Coupled Reticulocyte Lysate System (Promega). A construct designed to express the TATA-binding protein (TBP) as a GST-fusion protein was generated by amplifying the TBP sequence from cdna generated from HFFs using the following primers: htbp-f 5'- ATGGATCAGAACAACAGCCTGCC and htbp-r 5'- GCCATTACGTCGTCTTCCTGAATC. The resultant fragment was subcloned as an Eco RI fragment into pgex-6p and the fidelity of the construct confirmed by DNA sequencing. GST-fusion proteins were isolated from E. coli BL21 cultures by sonication in Buffer A (20 mm HEPES, ph 7.2, 100 mm KCl, 0.2 mm EDTA, 20% Glycerol, 1% Triton X-100) followed by purification using Glutathione sepharose (GE Healthcare). Binding assays were performed in Binding Buffer (25 mm HEPES, ph 7.5, 12.5 mm magnesium chloride, 20% glycerol, 0.1% NP-
12 , 150 mm potassium chloride, 0.15mg/ml bovine serum albumin, 1 mm DTT) for 2 hours at 4 0 C. Complexes were washed 6 times in NETN buffer (20 mm Tris, ph 8.0, 100 mm NaCl, 0.5% NP-40, 0.1 mm EDTA) and the bound proteins resolved by SDS-PAGE on 12.5% acrylamide gels. Images were visualized using a Typhoon 9410 Variable Mode Imager. Protein quantification was performed using ImageQuant TL Software
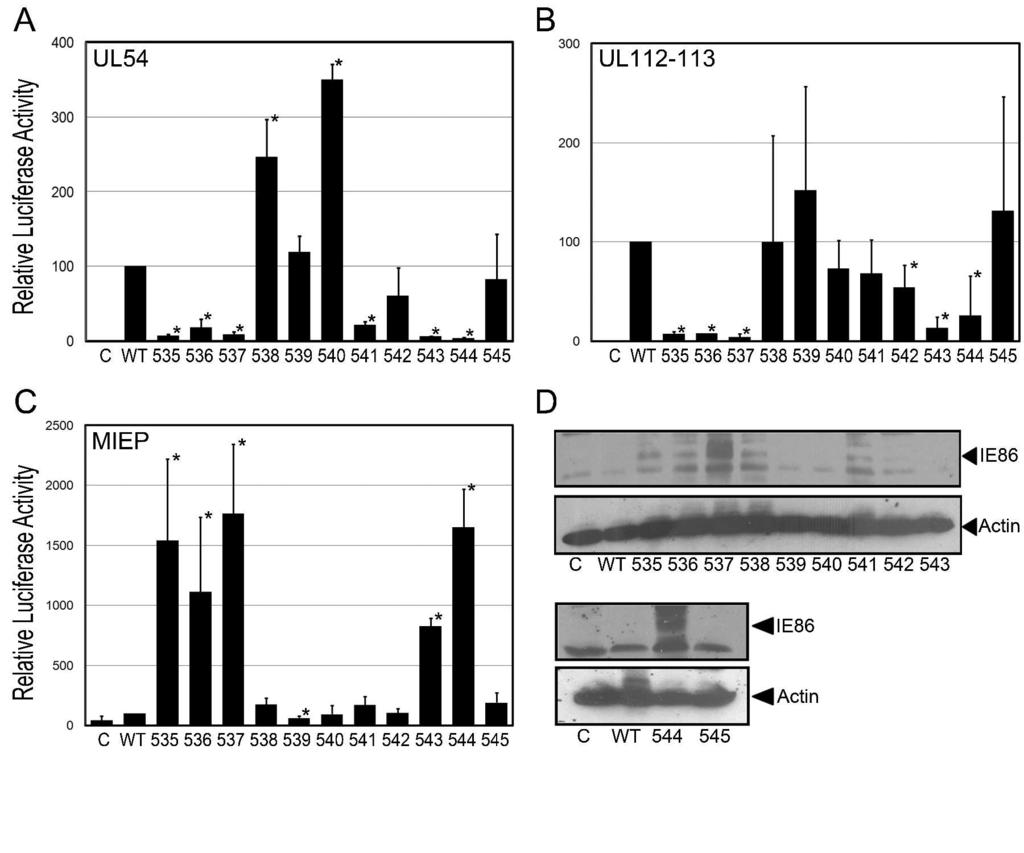
13 RESULTS Mutational Analysis of the IE86 Core Region Previous studies in our laboratory determined that insertion of 4 amino acids at position 540 in the carboxy-terminus of IE86 resulted in the loss of both MIE promoter repression and early promoter activation (59), suggesting that this region was critical for IE86 functions. Later studies identified a "core" domain within IE86 from amino acid 450 to 544 that is essential for the ability of IE86 to regulate gene expression (3). Consistent with this notion, this region of IE86 is highly conserved across cytomegaloviruses (45). To more closely address the role of this domain in IE86-mediated transcriptional regulation, we performed a comprehensive mutational analysis between amino acids 535 and 545. Each residue within this region was individually mutated to either an alanine, or valine, and the ability of the IE86 mutant in the context of the entire IE gene region to regulate two essential viral early promoters was assessed (Figure 1 A and B). This analysis revealed that mutations of amino acids 535, 536, 537, 543 and 544 resulted in a significant decrease in the ability of IE86 to activate the UL and UL54 promoters when compared to the wild type construct. These results are consistent with the role of the core region of IE86 in the transcriptional regulation of viral early genes (3). Interestingly, mutation of the threonine residue at position 541 demonstrated a significant decrease in UL54 promoter activation but had minimal effects on the ability to regulate the UL promoter. This finding demonstrates that a single amino acid mutation may have differential effects on early gene promoter activation, suggesting that individual early promoters may be regulated by distinct mechanisms.
14 The IE86 protein regulates its own expression by binding to the CRS of the MIE promoter (22, 32, 47). To assess the extent of correlation between IE86 early promoter activation and autoregulatory functions, we determined the effects of the IE86 mutations on MIEP activity (Fig. 1C). This analysis revealed that mutations of amino acids 535, 536, 537, 543 or 544 resulted in a loss in IE86 auto-repressive properties. Indeed, these mutants exhibited a significantly greater enhancement of MIEP promoter activity than might be expected from a simple loss of repression, suggesting that mutation of amino acids 535, 536, 537, 543 or 544 leads to MIEP activation by IE86. These results also indicate that mutations which disrupt transactivation of the UL and UL54 early promoters likely disrupt the ability of IE86 to autoregulate the MIEP. To confirm the effects of these mutations on IE86 auto-repression, we assessed IE86 protein levels in the transfected cells using an antibody that specifically recognizes the IE86 protein (Fig. 1D). This analysis shows that the wild type IE86 protein expressed in the context of the entire IE gene region is barely detectable, consistent with previous analysis using this antibody (58). In contrast, the majority of mutants that exhibited enhanced MIEP activation also expressed higher levels of the IE86 protein. In particular, mutation of either amino acid 537 or 544 resulted in dramatically increased expression of IE86. However, in some cases the level of IE86 protein did not correlate well with the observed transcriptional activity. For example, mutation of amino acid 543 results in enhanced activation of the MIEP, but minimal increases in IE86 protein levels. These findings suggest that other IE proteins expressed by the psvh construct may influence the transcriptional activity of IE86 (58). In order to determine effects specific for the IE86 protein, we assessed the IE86 mutants in the pmcrs86 vector that expresses the IE86 cdna under the control of a mutated MIE
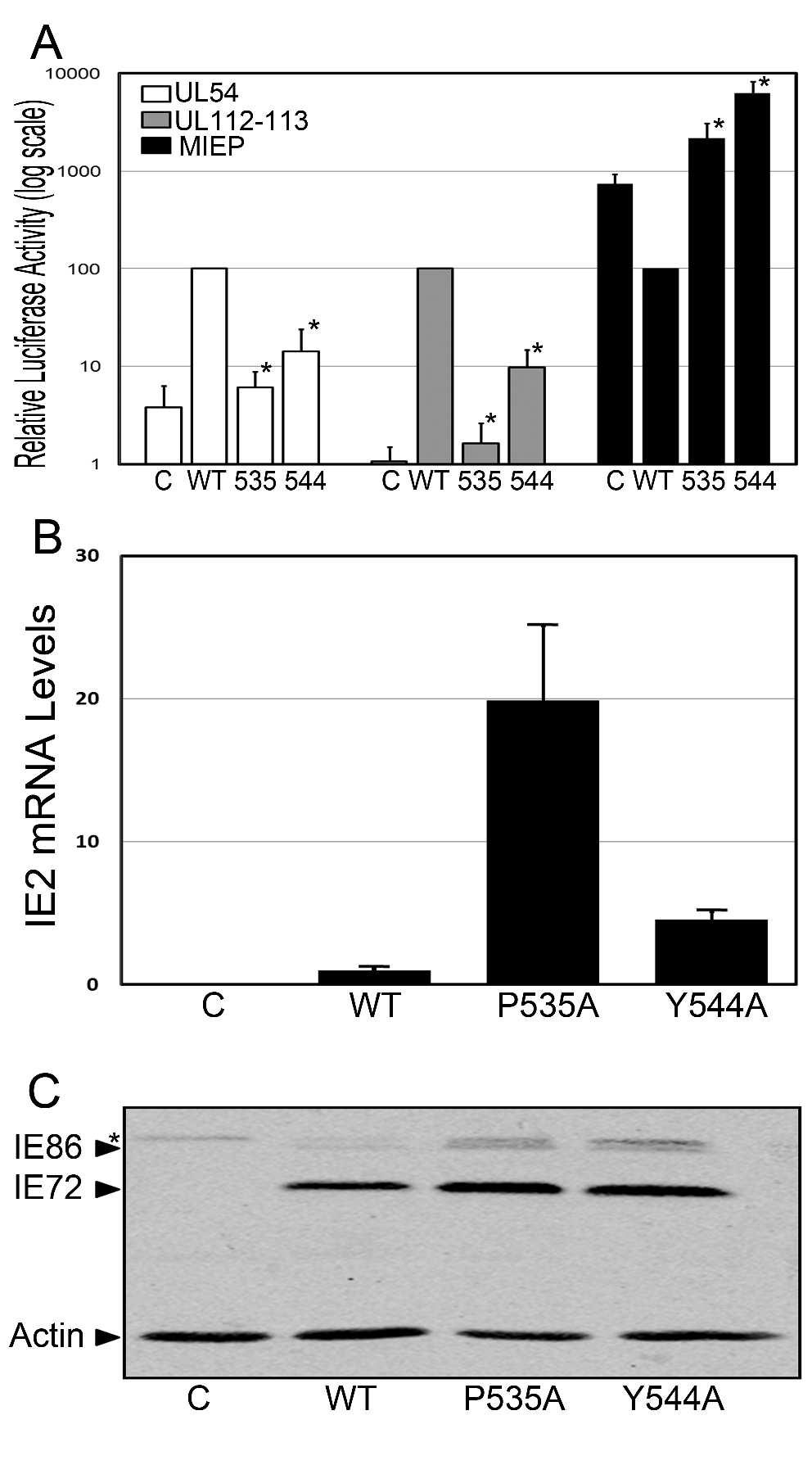
15 promoter designed to prevent autoregulation of the transfected plasmid (Figure 2). To refine our analysis, these studies were performed with two representative mutants, P535A and Y544A that correspond to highly conserved residues (45). Consistent with our examination in the context of the entire genomic region, mutations in IE86 at amino acids 535 and 544 impaired transcriptional regulation UL54 and UL promoters in the absence of other viral gene products (Fig. 2A). These findings suggest that the changes in early promoter regulation were due to the mutations in the IE86 protein. Significantly, analysis of the ability of the P535A and Y544A mutants to regulate the MIE promoter revealed not just a lack of repression, but an enhancement of promoter activity, suggesting that multiple mechanisms likely contribute to the regulation of the MIEP by the IE86 protein. To determine whether the mutations introduced affected IE86 protein stability, we assessed the accumulation of IE2 mrna and IE86 protein levels in transfected cells by real time RT-PCR and western blot analysis, respectively. We observed a 20-fold enhancement of IE2 transcript levels over that observed with the wild type construct in cells transfected with the IE86 P535A mutant, and a 5-fold enhancement with the IE86 Y544A mutant (Fig. 2B). These findings are consistent with the lack of auto-repression observed with these two mutants (Fig. 2A). Western blot analysis revealed that both mutants displayed enhanced IE86 protein levels when compared to the wild type construct (Fig. 2C). However, despite a 4-fold higher level of the IE2 P535A mutant transcript when compared to the IE2 Y544A mutant, we observed similar levels of IE86 protein, suggesting that the presence of a proline residue at position 535 may be important to maintain IE86 protein structure and/or stability. However, we cannot rule out the
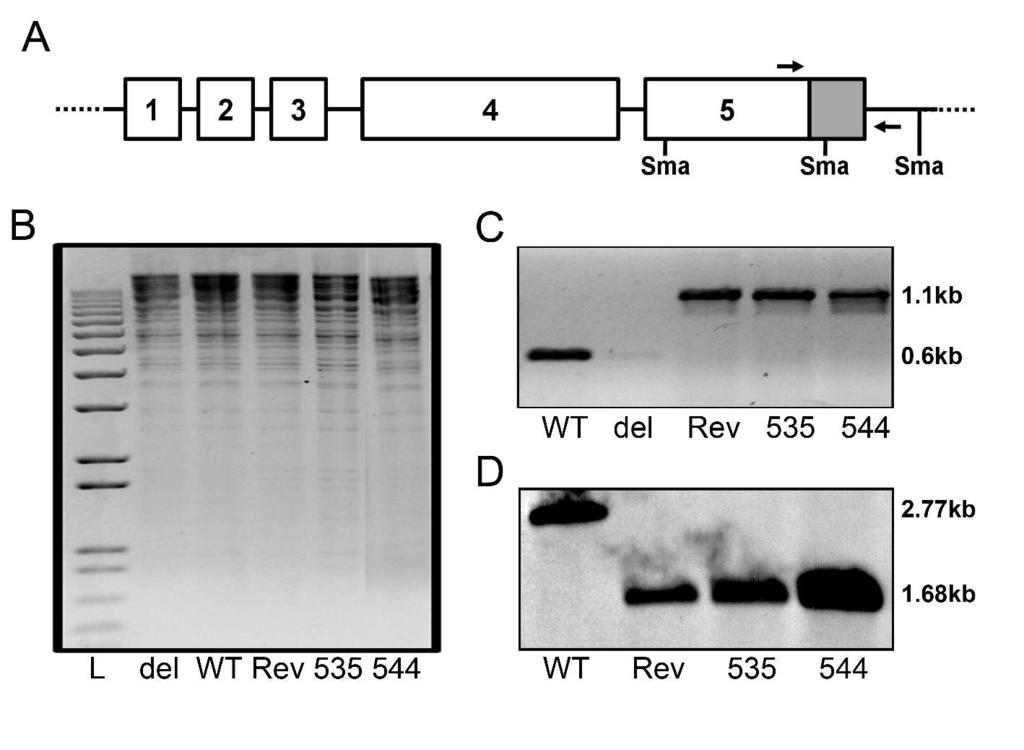
16 possibility that other post-transcriptional mechanisms such as translational efficiency contribute to the altered ratio of IE86 transcript and protein levels Assessment of IE86 mutants in the context of the viral genome To more closely investigate the transcriptional regulatory properties of IE86 that are essential for the activation of early genes, we assessed the effect of mutations at amino acids 535 and 544 on transcriptional regulation in the context of the viral genome. To accomplish this, we generated equivalent mutations in the HCMV Towne BAC (37). The constructs were confirmed by PCR, southern blot analysis and restriction enzyme digestion (Figure 3). During the generation of these constructs, additional sequences not present into the original HCMV Towne BAC were introduced. To ensure that these sequences did not interfere with viral growth, we compared viral DNA synthesis after transfection of the WT BAC and a WT revertant BAC construct (WT-Rev) into primary fibroblasts using primers that amplify the gb region of the genome (Fig. 4A). These studies showed identical levels of viral DNA synthesis for the WT and WT-Rev BAC constructs. We also performed single-step growth curve analysis of virus recovered after BAC transfections (34) and confirmed that there was no significant difference in replication between these two viruses (Fig. 4B). These results demonstrate that the additional sequences inserted into the BAC constructs did not affect the replication of the HCMV BACderived virus. Our analysis using transient transfection assays suggested that mutations at either amino acid 535 or 544 inhibited the ability of IE86 to activate the UL54 and UL promoters (Fig. 1). Since the products of both of these genes are essential for virus replication, we assessed
17 the capability of BAC constructs containing these mutations to yield viable virus. Following transfection of the BAC DNA into HFFs, cultures were monitored and maintained for up to 6 weeks with no indication of virus replication. These experiments were performed multiple times and efficient delivery of the BAC DNA into the cells was confirmed by PCR. These results confirm that the mutations in IE86 at amino acids 535 and 544 prevents virus replication. To determine the stage at which virus replication was blocked, UL54 and UL mrna transcript levels in BAC transfected cells were assessed by real time RT-PCR (Figure 5). Surprisingly, cells transfected with the BAC containing the mutation of proline 535 contained similar levels of the UL54 transcript to that observed with the WT-Rev construct at days 1 and 4 post-transfection (Fig. 5A), suggesting that the P535A mutation within IE86 has little affect on the ability of the UL54 promoter to be activated in this context. This result was in marked contrast to our in vitro analysis showing that IE86 containing the P535A substitution is unable to activate the UL54 promoter in transient assays (Fig. 1). At later times after transfection, the levels of UL54 transcripts in cells transfected with the IE86 P535A mutant BAC declined significantly when compared to WT-Rev, consistent with the null phenotype of this virus. Analysis of the substitution of the tyrosine at position 544 in IE86 revealed that this mutation abrogated activation of the UL54 promoter (Fig. 5A), in line with the activity of this mutant when assessed in transient assays (Fig. 1). These studies therefore revealed distinct differences in the ability of recombinant viruses containing mutations in IE86 at amino acid 535 and 544 to regulate the UL54 early gene, despite similar phenotypes when assessed in transient transfection studies. When UL transcript levels were assessed, we observed minimal activation of this gene with either mutant at any time after transfection of the BAC construct (Fig. 5B). These
18 data suggest that both mutations disrupt a functional property of IE86 that is essential for UL promoter activation. Further, they suggest that efficient expression of the UL gene region is at least one determinant of the outcome of HCMV infection We next determined the effects of mutations in IE86 at amino acid 535 and 544 on the ability to auto-repress the MIEP. Following transfection of the wildtype reverent BAC, IE2 transcript levels remain stable at 1 and 4 days post transfection, then begin to increase at 7 and 10 days post infection due to the progression of virus infection (Fig. 5C). Similar to results observed in transient transfection assays (Fig. 1C), we initially observed a lack of MIE promoter repression in cells transfected with BAC DNA containing IE86 mutations at amino acids 535 or 544, resulting in a 65- and 4-fold increase in IE2 mrna levels, respectively (Fig. 5C). Subsequently, the level of IE2 transcripts gradually declined, consistent with the block in virus replication observed with these two mutations. To confirm that the increase in IE2 transcripts correlated with IE86 protein levels, we performed western blot analysis on cells harvested at 2 days post transfection (Fig. 5D). This analysis showed enhanced levels of the IE86 protein in cells transfected with the BAC DNA containing IE86 mutations at amino acids 535 or 544 and is consistent with the in vitro analysis of these mutants (Figs. 1 & 2) Assessment of IE86 functional properties The IE86 protein regulates viral gene expression by binding to viral promoters and/or interactions with cellular and viral transcription factors (61). Previous studies suggest that IE86 may be recruited to the UL54 promoter due to interactions with cellular factors, such as TBP (23, 24). To determine if the mutations in the C-terminus of IE86 disrupt the interaction of IE86 with
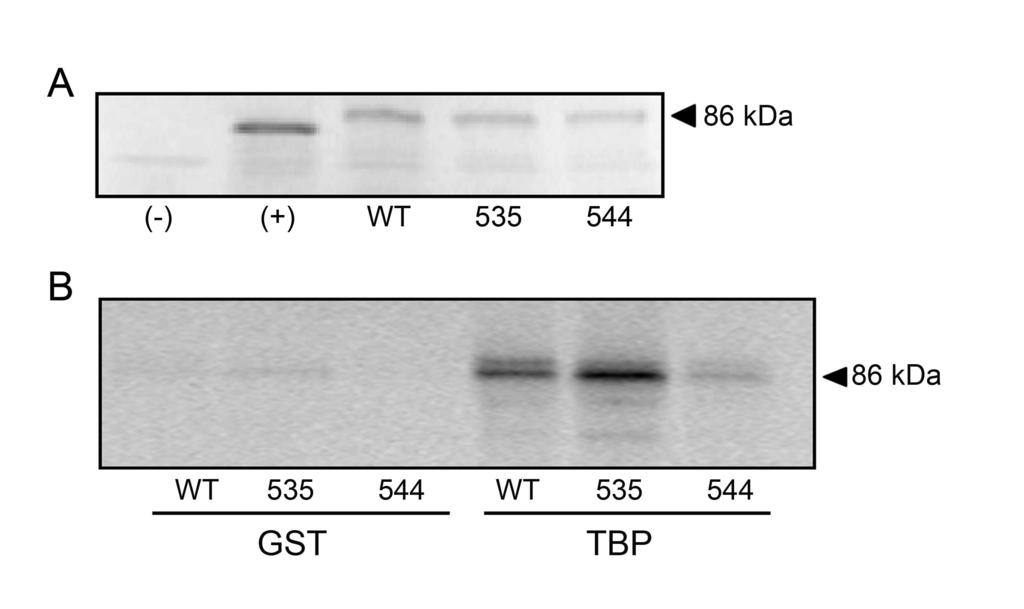
19 TBP, we performed GST pulldown analysis (Figure 6). Consistent with previous studies (8), our experiments show that wild type IE86 interacts efficiently with TBP (Fig. 6B), as does the IE86 protein containing the P535A substitution. In contrast, IE86 containing a mutation of tyrosine 544 exhibited markedly reduced binding with TBP (Fig. 6B). These studies indicate that IE86 interaction with TBP correlates with the ability to activate the UL54 promoter in the context of the viral genome. In addition, these findings suggest that the interaction between IE86 and TBP is not sufficient for the activation of the UL promoter as the P535A mutant is unable to activate this promoter in the context of the viral genome.
20 DISCUSSION Several studies have investigated the functional domains of the HCMV IE86 protein that are important for transcriptional regulation of viral early genes (reviewed in (61)). Many of these studies approached this challenge using multiple amino acid mutations or large deletions that could have confounding effects on IE86 protein structure. In the current study, we have performed a comprehensive mutational analysis of amino acids 535 to 545 of the IE86 sequence. These studies revealed that two clusters of amino acids within this region (aa and aa ) were particularly important for the ability of IE86 to activate viral early promoters as well as to regulate the MIEP. Strikingly, these residues correspond to sequences that are highly conserved in IE86 homologs from other species (45) with amino acids 535, 536, 537 and 543 found to be invariant. Surprisingly, we noted that mutation of other highly conserved residues within this domain of IE86 have minimal effects on transcriptional regulation in transient assays. For example, glutamic or aspartic acid residues at amino acid position 538 are retained across species (45). However, in our study, mutation of this residue to an alanine had no significant effect on the ability of IE86 to activate the UL54 or UL promoters. Similarly, amino acids 542 and 545 are highly conserved in primate cytomegaloviruses, but were found not to be important for the regulation of these early promoters. One possibility is that these residues are important for other IE86 functions not assessed in transient assays. Another important observation from this initial analysis was the finding that mutation of amino acid 541 differentially affected transcriptional activation of the two early promoters that we assessed. This finding is consistent with previous analysis of an equivalent IE86 mutant (4), and may be due to effects on IE86 post-translational modifications.
21 We considered the possibility that mutation of these highly conserved residues results in altered folding or conformation of IE86, accounting for the reduced activities. Indeed, when we compared the P535A and Y544A mutants, we observed four-fold higher levels of the IE2 P535A transcript, but similar steady-state protein levels. This suggests that substitution of an alanine for the proline at position 535 may result in decreased stability of IE86, consistent with an altered conformation. However, analysis of the mutants in the context of the IE86 cdna construct revealed enhancement of MIEP activity. This suggests that these IE86 mutants are capable of activating the MIEP, possible via effects on chromatin modifying enzymes (6, 21, 26, 44, 49) and is inconsistent with major changes in the conformation of the IE86 protein. The ability of the IE86 mutants to enhance the MIEP is also indicative of a loss of the autoregulatory functions of this protein. Indeed, we consistently observed correlation between loss of IE86 autoregulation and transactivation of essential early gene promoters. In addition, assessment of the IE86 P535A and Y544A mutants in the context of the viral genome revealed a similar lack of MIEP repression. These results support other studies suggesting that IE86 domains involved in transcriptional activation and repression overlap (3, 59, 63). In contrast, a recent study evaluating a double mutation at amino acid 535 and 537 in the context of the HCMV BAC demonstrated that these two effects were separable (45). This discrepancy could be explained by the temporal differences at which RNA expression was assessed. Another possible explanation is that the tyrosine 537 mutation may partially compensate for the proline 535 mutation with respect to autorepression due to localized effects on protein conformation. Analysis in the context of the viral genome revealed that the proline at amino acid position 535 and the tyrosine residue at position 544 were essential for the generation of viable
22 HCMV. This finding is consistent with reduced activation of essential viral early gene promoters observed in transient assays. Surprisingly, assessment of the IE86 P535A mutant revealed similar levels of UL54 mrna to WT-Rev following transfection of the BAC clones. This implies that other viral proteins are important for activation of the UL54 promoter, and/or sufficient functions of IE86 are retained to enable activation of this promoter in this context. At later times after transfection we observed a decrease in UL54 transcript levels that likely reflects a lack of progression of the infection due to a failure to activate other essential viral early genes such as the UL gene region. The levels of UL transcripts were approximately 10-fold lower than wild type at one day post transfection, and were essentially undetectable by day four. A similar result was obtained when UL transcript levels were assessed in cells transfected with the BAC containing the IE86 Y544A mutant, and is in line with results of transient assays showing that both mutations disrupt IE86 activation of this promoter. Previous studies showed that the UL promoter is controlled by ATF/CREB and IE86 binding sites (50). It is possible that these mutations disrupt the ability of IE86 to bind DNA, resulting in lower levels of Ul transcripts. This is also consistent with the lack of autoregulatory activity observed with both mutants. The phenotype of the HCMV clone containing the Y544A substitution within IE86 is consistent with this mutation disrupting a function of IE86 that is critical for the activation of viral early promoters, such as the ability of IE86 to associate with the basal transcriptional machinery (8, 16). Indeed, our analysis revealed that mutation of amino acid 544 causes a dramatic reduction in TBP binding. This finding is consistent with studies showing that IE86 has TAF-like functions and adds further support to the notion that IE86-TBP interactions are critical
23 for the transcriptional activation of viral early promoters (25, 35, 36). Although the TBP interaction domain of IE86 does not map to this region (8), other studies suggest that dimerization of IE86 is important for IE86-TBP interactions (56). Since amino acid 544 overlaps the dimerization domain of IE86 (1), it is possible that this mutation inhibits IE86 dimerization. However, it should also be noted that these studies were performed in vitro, and may not reflect interactions that occur in the context of virus infection. For example, it is known that IE86 phosphorylation and sumoylation have significant effects on IE86-mediated transcriptional activation (5, 17, 19, 20, 30, 31). We are currently examining the recruitment of viral and cellular factors to viral early promoters in the context of viral infection in order to further address this question. In summary, our current analysis has revealed two important findings: (1) IE86- mediated activation of viral essential early promoters occurs via distinct mechanisms; and (2) the interaction between IE86 and TBP is critical for the activation of the UL54 early promoter.
24 ACKNOWLEDGMENTS The authors would like to acknowledge Dr. Zhen Zhang for his invaluable technical advice in the development of the BAC clones and Dr. Lisa Bolin for her intellectual and technical contributions. This study was supported by Public Health Service grant AI38372 (J.A.K.) from the National Institutes of Health. Downloaded from on May 3, 2018 by guest
25 Harris, et al./page 25 1 REFERENCES Ahn, J. H., C. J. Chiou, and G. S. Hayward Evaluation and mapping of the DNA binding and oligomerization domains of the IE2 regulatory protein of human cytomegalovirus using yeast one and two hybrid interaction assays. Gene 210: Arlt, H., D. Lang, S. Gebert, and T. Stamminger Identification of binding sites for the 86-kilodalton IE2 protein of human cytomegalovirus within an IE2-responsive viral early promoter. J. Virol. 68: Asmar, J., L. Wiebusch, M. Truss, and C. Hagemeier The putative zinc finger of the human cytomegalovirus IE2 86-kilodalton protein is dispensable for DNA binding and autorepression, thereby demarcating a concise core domain in the C terminus of the protein. J Virol 78: Barrasa, M. I., N. Harel, Y. Yu, and J. C. Alwine Strain variations in single amino acids of the 86-kilodalton human cytomegalovirus major immediate-early protein (IE2) affect its functional and biochemical properties: implications of dynamic protein conformation. J Virol 77: Berndt, A., H. Hofmann-Winkler, N. Tavalai, G. Hahn, and T. Stamminger Importance of covalent and noncovalent SUMO interactions with the major human cytomegalovirus transactivator IE2p86 for viral infection. J Virol 83: Bryant, L. A., P. Mixon, M. Davidson, A. J. Bannister, T. Kouzarides, and J. H. Sinclair The Human Cytomegalovirus 86-Kilodalton Major Immediate-Early Protein Interacts Physically and Functionally with Histone Acetyltransferase P/CAF. J. Virol. 74: Bryant, L. A., P. Mixon, M. Davidson, A. J. Bannister, T. Kouzarides, and J. H. Sinclair The human cytomegalovirus 86-kilodalton major immediate-early protein interacts physically and functionally with histone acetyltransferase P/CAF. J Virol 74: Caswell, R., C. Hagemeier, C. J. Chiou, G. Hayward, T. Kouzarides, and J. Sinclair The human cytomegalovirus 86K immediate early (IE) 2 protein requires the basic region of the TATA-box binding protein (TBP) for binding, and interacts with TBP and transcription factor TFIIB via regions of IE2 required for transcriptional regulation. J Gen Virol 74: Chau, N. H., C. D. Vanson, and J. A. Kerry Transcriptional regulation of the human cytomegalovirus US11 early gene. J Virol 73: Cherrington, J. M., E. L. Khoury, and E. S. Mocarski Human cytomegalovirus ie2 negatively regulates alpha gene expression via a short target sequence near the transcription start site. J Virol 65: Cherrington, J. M., and E. S. Mocarski Human cytomegalovirus ie1 transactivates the alpha promoter-enhancer via an 18-base-pair repeat element. J Virol 63: Ciocco-Schmitt, G. M., Z. Karabekian, E. W. Godfrey, R. M. Stenberg, A. E. Campbell, and J. A. Kerry Identification and characterization of novel murine cytomegalovirus M (e1) gene products. Virology 294:
26 Depto, A. S., and R. M. Stenberg Regulated expression of the human cytomegalovirus pp65 gene: octamer sequence in the promoter is required for activation by viral gene products. J Virol 63: Dunn, W., C. Chou, H. Li, R. Hai, D. Patterson, V. Stolc, H. Zhu, and F. Liu Functional profiling of a human cytomegalovirus genome. Proc Natl Acad Sci U S A 100: Fortunato, E. A., and D. H. Spector Regulation of human cytomegalovirus gene expression. Adv Virus Res 54: Hagemeier, C., S. Walker, R. Caswell, T. Kouzarides, and J. Sinclair The human cytomegalovirus 80-kilodalton but not the 72-kilodalton immediate-early protein transactivates heterologous promoters in a TATA box-dependent mechanism and interacts directly with TFIID. J. Virol. 66: Harel, N. Y., and J. C. Alwine Phosphorylation of the human cytomegalovirus 86-kilodalton immediate- early protein IE2. J Virol 72: Heider, J. A., W. A. Bresnahan, and T. E. Shenk Construction of a rationally designed human cytomegalovirus variant encoding a temperature-sensitive immediate-early 2 protein. PNAS 99: Heider, J. A., Y. Yu, T. Shenk, and J. C. Alwine Characterization of a human cytomegalovirus with phosphorylation site mutations in the immediate-early 2 protein. J Virol 76: Hofmann, H., S. Floss, and T. Stamminger Covalent modification of the transactivator protein IE2-p86 of human cytomegalovirus by conjugation to the ubiquitinhomologous proteins SUMO-1 and hsmt3b. J Virol 74: Hsu, C. H., M. D. Chang, K. Y. Tai, Y. T. Yang, P. S. Wang, C. J. Chen, Y. H. Wang, S. C. Lee, C. W. Wu, and L. J. Juan HCMV IE2-mediated inhibition of HAT activity downregulates p53 function. EMBO J 23: Jupp, R., S. Hoffmann, A. Depto, R. M. Stenberg, P. Ghazal, and J. A. Nelson Direct interaction of the human cytomegalovirus IE86 protein with the cis repression signal does not preclude TBP from binding to the TATA box. J. Virol. 67: Kerry, J. A., M. A. Priddy, T. Y. Jervey, C. P. Kohler, T. L. Staley, C. D. Vanson, T. R. Jones, A. C. Iskenderian, D. G. Anders, and R. M. Stenberg Multiple regulatory events influence human cytomegalovirus DNA polymerase (UL54) expression during viral infection. J Virol 70: Kerry, J. A., M. A. Priddy, and R. M. Stenberg Identification of sequence elements in the human cytomegalovirus DNA polymerase gene promoter required for activation by viral gene products. J Virol 68: Kim, J. M., Y. Hong, K. T. Jeang, and S. Kim Transactivation activity of the human cytomegalovirus IE2 protein occurs at steps subsequent to TATA box-binding protein recruitment. J Gen Virol 81: Klucher, K. M., M. Sommer, J. T. Kadonaga, and D. H. Spector In vivo and in vitro analysis of transcriptional activation mediated by the human cytomegalovirus major immediate-early proteins. Mol Cell Biol 13: Kohler, C. P., J. A. Kerry, M. Carter, V. P. Muzithras, T. R. Jones, and R. M. Stenberg Use of recombinant virus to assess human cytomegalovirus early and late promoters in the context of the viral genome. J Virol 68:
27 Lang, D., S. Gebert, H. Arlt, and T. Stamminger Functional interaction between the human cytomegalovirus 86-kilodalton IE2 protein and the cellular transcription factor CREB. J. Virol. 69: Lee, G., J. Wu, P. Luu, P. Ghazal, and O. Flores Inhibition of the association of RNA polymerase II with the preinitiation complex by a viral transcriptional repressor. PNAS 93: Lee, H. R., and J. H. Ahn Sumoylation of the major immediate-early IE2 protein of human cytomegalovirus Towne strain is not required for virus growth in cultured human fibroblasts. J Gen Virol 85: Lee, J. M., H. J. Kang, H. R. Lee, C. Y. Choi, W. J. Jang, and J. H. Ahn PIAS1 enhances SUMO-1 modification and the transactivation activity of the major immediateearly IE2 protein of human cytomegalovirus. FEBS Lett 555: Liu, B., T. W. Hermiston, and M. F. Stinski A cis-acting element in the major immediate-early (IE) promoter of human cytomegalovirus is required for negative regulation by IE2. J Virol 65: Liu, B., and M. F. Stinski Human cytomegalovirus contains a tegument protein that enhances transcription from promoters with upstream ATF and AP-1 cis-acting elements. J Virol 66: Lorz, K., H. Hofmann, A. Berndt, N. Tavalai, R. Mueller, U. Schlotzer-Schrehardt, and T. Stamminger Deletion of open reading frame UL26 from the human cytomegalovirus genome results in reduced viral growth, which involves impaired stability of viral particles. J Virol 80: Lukac, D. M., N. Y. Harel, N. Tanese, and J. C. Alwine TAF-like functions of human cytomegalovirus immediate-early proteins. J Virol 71: Lukac, D. M., J. R. Manuppello, and J. C. Alwine Transcriptional activation by the human cytomegalovirus immediate-early proteins: requirements for simple promoter structures and interactions with multiple components of the transcription complex. J Virol 68: Marchini, A., H. Liu, and H. Zhu Human Cytomegalovirus with IE-2 (UL122) Deleted Fails To Express Early Lytic Genes. J. Virol. 75: Meier, J. L., and M. F. Stinski Regulation of human cytomegalovirus immediateearly gene expression. Intervirology 39: Mellon, P., V. Parker, Y. Gluzman, and T. Maniatis Identification of DNA sequences required for transcription of the human alpha 1-globin gene in a new SV40 host-vector system. Cell 27: Mocarski, E. S., Jr., and Courcelle, C. T Cytomegaloviruses and Their Replication, p In D. M. Knipe, and Howley, P. M. (ed.), Fields Virology, Fourth Edition ed, vol. 2. Lippincott, Williams and Wilkins, Philadelphia. 41. Netterwald, J., S. Yang, W. Wang, S. Ghanny, M. Cody, P. Soteropoulos, B. Tian, W. Dunn, F. Liu, and H. Zhu Two gamma interferon-activated site-like elements in the human cytomegalovirus major immediate-early promoter/enhancer are important for viral replication. J Virol 79: Pari, G. S., and D. G. Anders Eleven loci encoding trans-acting factors are required for transient complementation of human cytomegalovirus orilyt-dependent DNA replication. J Virol 67:
28 Pari, G. S., and Y. Xu Gene transfer into mammalian cells using calcium phosphate and DEAE-dextran. Methods Mol Biol 245: Park, J. J., Y. E. Kim, H. T. Pham, E. T. Kim, Y. H. Chung, and J. H. Ahn Functional interaction of the human cytomegalovirus IE2 protein with histone deacetylase 2 in infected human fibroblasts. J Gen Virol 88: Petrik, D. T., K. P. Schmitt, and M. F. Stinski The autoregulatory and transactivating functions of the human cytomegalovirus IE86 protein use independent mechanisms for promoter binding. J Virol 81: Pfaffl, M. W A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res 29:e Pizzorno, M. C., and G. S. Hayward The IE2 gene products of human cytomegalovirus specifically down-regulate expression from the major immediate-early promoter through a target sequence located near the cap site. J Virol 64: Pizzorno, M. C., P. O'Hare, L. Sha, R. L. LaFemina, and G. S. Hayward trans-activation and autoregulation of gene expression by the immediate-early region 2 gene products of human cytomegalovirus. J Virol 62: Reeves, M., J. Murphy, R. Greaves, J. Fairley, A. Brehm, and J. Sinclair Autorepression of the human cytomegalovirus major immediate-early promoter/enhancer at late times of infection is mediated by the recruitment of chromatin remodeling enzymes by IE86. J Virol 80: Rodems, S. M., C. L. Clark, and D. H. Spector Separate DNA Elements Containing ATF/CREB and IE86 Binding Sites Differentially Regulate the Human Cytomegalovirus UL Promoter at Early and Late Times in the Infection. J. Virol. 72: Schwartz, R., B. Helmich, and D. H. Spector CREB and CREB-binding proteins play an important role in the IE2 86- kilodalton protein-mediated transactivation of the human cytomegalovirus 2.2-kilobase RNA promoter. J. Virol. 70: Schwartz, R., B. Helmich, and D. H. Spector CREB and CREB-binding proteins play an important role in the IE2 86-kilodalton protein-mediated transactivation of the human cytomegalovirus 2.2-kilobase RNA promoter. J Virol 70: Schwartz, R., M. H. Sommer, A. Scully, and D. H. Spector Site-specific binding of the human cytomegalovirus IE2 86-kilodalton protein to an early gene promoter. J. Virol. 68: Scully, A. L., M. H. Sommer, R. Schwartz, and D. H. Spector The human cytomegalovirus IE2 86-kilodalton protein interacts with an early gene promoter via site-specific DNA binding and protein-protein associations. J Virol 69: Shen, W., E. Westgard, L. Huang, M. D. Ward, J. L. Osborn, N. H. Chau, L. Collins, B. Marcum, M. A. Koach, J. Bibbs, O. J. Semmes, and J. A. Kerry Nuclear trafficking of the human cytomegalovirus pp71 (ppul82) tegument protein. Virology 376: Sommer, M. H., A. L. Scully, and D. H. Spector Transactivation by the human cytomegalovirus IE2 86-kilodalton protein requires a domain that binds to both the TATA boxbinding protein and the retinoblastoma protein. J Virol 68: Staprans, S. I., D. K. Rabert, and D. H. Spector Identification of sequence requirements and trans-acting functions necessary for regulated expression of a human cytomegalovirus early gene. J Virol 62:
29 Stenberg, R. M., A. S. Depto, J. Fortney, and J. A. Nelson Regulated expression of early and late RNAs and proteins from the human cytomegalovirus immediateearly gene region. J Virol 63: Stenberg, R. M., J. Fortney, S. W. Barlow, B. P. Magrane, J. A. Nelson, and P. Ghazal Promoter-specific trans activation and repression by human cytomegalovirus immediate-early proteins involves common and unique protein domains. J Virol 64: Stinski, M. F., and H. Isomura Role of the cytomegalovirus major immediate early enhancer in acute infection and reactivation from latency. Med Microbiol Immunol 197: Stinski, M. F., and D. T. Petrik Functional roles of the human cytomegalovirus essential IE86 protein. Curr Top Microbiol Immunol 325: Stinski, M. F., and T. J. Roehr Activation of the major immediate early gene of human cytomegalovirus by cis-acting elements in the promoter-regulatory sequence and by virus-specific trans-acting components. J Virol 55: White, E. A., C. L. Clark, V. Sanchez, and D. H. Spector Small Internal Deletions in the Human Cytomegalovirus IE2 Gene Result in Nonviable Recombinant Viruses with Differential Defects in Viral Gene Expression. J. Virol. 78: Wu, J., J. O'Neill, and M. S. Barbosa Transcription factor Sp1 mediates cellspecific trans-activation of the human cytomegalovirus DNA polymerase gene promoter by immediate-early protein IE86 in glioblastoma U373MG cells. J Virol 72: Yu, D., G. A. Smith, L. W. Enquist, and T. Shenk Construction of a selfexcisable bacterial artificial chromosome containing the human cytomegalovirus genome and mutagenesis of the diploid TRL/IRL13 gene. J Virol 76: Downloaded from on May 3, 2018 by guest
30 Table 1. Primers used for site-directed PCR mutatgenesis a Primer Primer Sequence Mutation P535A-2 5 GTGGGTTCATGCTGGCTATCTACGAGACGGC 3 Pro 535 Ala I536A 5 GGTTCATGCTGCCTGCCTACGAGACGGCCA 3 Ile 536 Ala Y537A 5 CATGCTGCCTATCGCCGAGACGGCCACG 3 Try 537 Ala E538A 5 GTTCATGCTGCCTATCTACGCGACGGCCACGAAGG 3 Asn 538 Ala T539A 5 CCTATCTACGAGGCGGCCACGAAGGC 3 Try 539 Ala A540V 5 TATCTACGAGACGGTCACGAAGGCCTACGC 3 Ala 540 Val T541A 5 CTATCTACGAGACGGCCGCGAAGGCCTA 3 Try 541 Ala K542A 5 TATCTACGAGACGGCCACGGCGGCCTACGC 3 Asn 542 Ala A543V 5 GGCCACGAAGGTCTACGCCGTGGG 3 Ala 543 Val Y544A 5 CCACGAAGGCCGCCGCCGTGGGGC 3 Try 544 Ala A545V 5 CGAAGGCCTACGTCGTGGGGCAGTTTGA 3 Ala 545 Ala a Only forward primer sequence shown for each primer set and mutations in IE2 are underlined Downloaded from on May 3, 2018 by guest
31 FIGURE LEGENDS Figure 1. Effect of IE86 mutations on UL54, UL and MIE promoter regulation. The pul54-luc (A), pul luc (B) or the pmiep-luc (C) reporter constructs were cotransfected into primary fibroblasts with the psvhk vector that expresses either wild type IE86 or the indicated mutant in the context of the MIE gene region. Cell extracts were assessed for luciferase activity 48 hours after transfection. Data is the average and standard deviation (SD) of a minimum of two experiments performed in duplicate, normalized to promoter activity in the presence of wild type IE86. (D) Total protein was harvested at 48 hours post transfection and separated on a 10 % SDS-PAGE. Western blot analysis was performed using a peptide antibody that recognizes the IE86 protein. An antibody to actin was included to normalize for protein loading. Symbols: * significantly different from wild type, p<0.05. "C" - level of promoter activity in the presence of the psv0d empty vector control plasmid. Figure 2. Effect of IE86 mutations on promoter activation, IE2 mrna and IE86 protein expression. (A) The pmiep-luc, pul54-luc or UL Luc reporter constructs were cotransfected into primary fibroblasts with pmcrs86 vector expressing either wildtype IE86 or the indicated mutant. Cell extracts were assessed for luciferase activity 48 hours after transfection. Data is the average and standard deviation (SD) of a minimum of two experiments performed in duplicate, normalized to promoter activity in the presence of wild type IE86 and presented on a log-scale to assist in the comparison of promoter activation. Symbols: * significantly different from wild type, p<0.05. "C" - level of promoter activity in the in the presence of the psv0d empty vector control plasmid. (B & C) Primary fibroblasts were transfected with the psvhk
32 vector expressing either wild type IE86 or the indicated mutant. (B) Total RNA was harvested at 48 hours post transfection and real time RT-PCR analysis performed to determine IE2 mrna levels. Analyses were performed in duplicate and relative expression normalized to GAPDH mrna levels was expressed as mean ± SD. (C) Total protein was harvested at 48 hours post transfection and separated on a 12.5% SDS-PAGE. Western blot analysis was performed using an antibody that recognizes IE72 and IE86. An antibody to actin was included to normalize for protein loading. Symbol: * non-specific band. Figure 3. Analysis of mutant HCMV BACs. (A) Schematic of the MIE gene region of the mutant BACs. The Zeocin resistance cassette containing a Sma I site is represented by the gray box. The location of primer sequences used to confirm appropriate recombination are also indicated by arrows. (B) Restriction enzyme analysis of HCMV BACs was performed using Sal I and Hind III. Following digestion, the fragmented DNA was run on a 0.8% agarose gel. (C) PCR analysis of HCMV BACs was performed using the indicated primers and the gel fragments separated on a 1.2% agarose gel. The HCMV WT BAC results in an approximately 0.6 kb fragment, with the Zeocin resistance cassette increasing the size of this fragment to 1.1 kb. (D) Southern blot analysis of HCMV BACs digested with Sma I showing the presence of the additional Sma I site in the WT-Rev, P535A and Y544A BAC constructs due to the presence of the Zeocin resistance cassette. Figure 4. Replication kinetics of HCMV WT and WT-Rev virus. WT-Rev and mutant HCMV BACs were transfected into primary fibroblasts by calcium phosphate transfections. (A) DNA replication was assessed by isolating genomic DNA at the indicated times following transfection
33 and assessment of gb DNA levels by real time PCR. (B) Growth kinetics were assessed by isolating genomic DNA from the culture supernatant at the indicated times following transfection and assessment of gb DNA levels by real time PCR. Figure 5. Regulation of viral gene expression in BAC transfected cells. Total RNA was isolated from BAC transfected cells at the indicated times post-transfection and analyzed by real-time RT-PCR using primers specific for the UL54 (A), UL (B) and IE2 (C) transcript levels. Data is expressed as mean ± SD from two independent experiments performed in duplicate. Symbol: * significantly different from wild type, p<0.05. (D) Total protein was harvested at 48 hours post transfection and separated on a 10 % SDS-PAGE. Western blot analysis was performed using a peptide antibody that recognizes the IE86 protein. An antibody to actin was included to normalize for protein loading. Figure 6. GST-pulldown analysis of IE86 mutant proteins. (A) SDS-PAGE analysis of wild type and the indicated IE86 mutant proteins generated by in vitro transcription/translation reactions. Positive (+) and negative (-) control IVTT reactions were included. (B) Wild type or mutant IE86 proteins were incubated with bacterially expressed GST or GST-TBP. GST complexes were then purified by glutathione sepharose beads and the bound proteins separated by SDS-PAGE and the 35 S-labeled IE86 proteins visualized on a Typhoon imager.
34
35
36
37
38
39
c Tuj1(-) apoptotic live 1 DIV 2 DIV 1 DIV 2 DIV Tuj1(+) Tuj1/GFP/DAPI Tuj1 DAPI GFP
 apoptotic live 1 DIV 2 DIV 1 DIV 2 DIV Tuj1(+) Tuj1/GFP/DAPI Tuj1 DAPI GFP") Supplementary Figure 1 Establishment of the gain- and loss-of-function experiments and cell survival assays. a Relative expression of mature mir-484 30 20 10 0 **** **** NCP mir- 484P NCP mir- 484P b Relative
Supplementary Figure 1 Establishment of the gain- and loss-of-function experiments and cell survival assays. a Relative expression of mature mir-484 30 20 10 0 **** **** NCP mir- 484P NCP mir- 484P b Relative
Supplementary Appendix
 Supplementary Appendix This appendix has been provided by the authors to give readers additional information about their work. Supplement to: Sherman SI, Wirth LJ, Droz J-P, et al. Motesanib diphosphate
Supplementary Appendix This appendix has been provided by the authors to give readers additional information about their work. Supplement to: Sherman SI, Wirth LJ, Droz J-P, et al. Motesanib diphosphate
Supplemental Data. Shin et al. Plant Cell. (2012) /tpc YFP N
 /tpc YFP N") MYC YFP N PIF5 YFP C N-TIC TIC Supplemental Data. Shin et al. Plant Cell. ()..5/tpc..95 Supplemental Figure. TIC interacts with MYC in the nucleus. Bimolecular fluorescence complementation assay using
MYC YFP N PIF5 YFP C N-TIC TIC Supplemental Data. Shin et al. Plant Cell. ()..5/tpc..95 Supplemental Figure. TIC interacts with MYC in the nucleus. Bimolecular fluorescence complementation assay using
Supplementary Table 3. 3 UTR primer sequences. Primer sequences used to amplify and clone the 3 UTR of each indicated gene are listed.
 Supplemental Figure 1. DLKI-DIO3 mirna/mrna complementarity. Complementarity between the indicated DLK1-DIO3 cluster mirnas and the UTR of SOX2, SOX9, HIF1A, ZEB1, ZEB2, STAT3 and CDH1with mirsvr and PhastCons
Supplemental Figure 1. DLKI-DIO3 mirna/mrna complementarity. Complementarity between the indicated DLK1-DIO3 cluster mirnas and the UTR of SOX2, SOX9, HIF1A, ZEB1, ZEB2, STAT3 and CDH1with mirsvr and PhastCons
Figure S1. Analysis of genomic and cdna sequences of the targeted regions in WT-KI and
 Figure S1. Analysis of genomic and sequences of the targeted regions in and indicated mutant KI cells, with WT and corresponding mutant sequences underlined. (A) cells; (B) K21E-KI cells; (C) D33A-KI cells;
Figure S1. Analysis of genomic and sequences of the targeted regions in and indicated mutant KI cells, with WT and corresponding mutant sequences underlined. (A) cells; (B) K21E-KI cells; (C) D33A-KI cells;
Supplementary Document
 Supplementary Document 1. Supplementary Table legends 2. Supplementary Figure legends 3. Supplementary Tables 4. Supplementary Figures 5. Supplementary References 1. Supplementary Table legends Suppl.
Supplementary Document 1. Supplementary Table legends 2. Supplementary Figure legends 3. Supplementary Tables 4. Supplementary Figures 5. Supplementary References 1. Supplementary Table legends Suppl.
Nature Structural & Molecular Biology: doi: /nsmb Supplementary Figure 1
 Supplementary Figure 1 U1 inhibition causes a shift of RNA-seq reads from exons to introns. (a) Evidence for the high purity of 4-shU-labeled RNAs used for RNA-seq. HeLa cells transfected with control
Supplementary Figure 1 U1 inhibition causes a shift of RNA-seq reads from exons to introns. (a) Evidence for the high purity of 4-shU-labeled RNAs used for RNA-seq. HeLa cells transfected with control
Supplementary Figure 1 a
 Supplementary Figure a Normalized expression/tbp (A.U.).6... Trip-br transcripts Trans Trans Trans b..5. Trip-br Ctrl LPS Normalized expression/tbp (A.U.) c Trip-br transcripts. adipocytes.... Trans Trans
Supplementary Figure a Normalized expression/tbp (A.U.).6... Trip-br transcripts Trans Trans Trans b..5. Trip-br Ctrl LPS Normalized expression/tbp (A.U.) c Trip-br transcripts. adipocytes.... Trans Trans
Supplementary Figure 1. ROS induces rapid Sod1 nuclear localization in a dosagedependent manner. WT yeast cells (SZy1051) were treated with 4NQO at
 were treated with 4NQO at") Supplementary Figure 1. ROS induces rapid Sod1 nuclear localization in a dosagedependent manner. WT yeast cells (SZy1051) were treated with 4NQO at different concentrations for 30 min and analyzed for
Supplementary Figure 1. ROS induces rapid Sod1 nuclear localization in a dosagedependent manner. WT yeast cells (SZy1051) were treated with 4NQO at different concentrations for 30 min and analyzed for
Supplementary Materials
 Supplementary Materials 1 Supplementary Table 1. List of primers used for quantitative PCR analysis. Gene name Gene symbol Accession IDs Sequence range Product Primer sequences size (bp) β-actin Actb gi
Supplementary Materials 1 Supplementary Table 1. List of primers used for quantitative PCR analysis. Gene name Gene symbol Accession IDs Sequence range Product Primer sequences size (bp) β-actin Actb gi
a) Primary cultures derived from the pancreas of an 11-week-old Pdx1-Cre; K-MADM-p53
 Primary cultures derived from the pancreas of an 11-week-old Pdx1-Cre; K-MADM-p53") 1 2 3 4 5 6 7 8 9 10 Supplementary Figure 1. Induction of p53 LOH by MADM. a) Primary cultures derived from the pancreas of an 11-week-old Pdx1-Cre; K-MADM-p53 mouse revealed increased p53 KO/KO (green,
1 2 3 4 5 6 7 8 9 10 Supplementary Figure 1. Induction of p53 LOH by MADM. a) Primary cultures derived from the pancreas of an 11-week-old Pdx1-Cre; K-MADM-p53 mouse revealed increased p53 KO/KO (green,
The Autoregulatory and Transactivating Functions of the Human Cytomegalovirus IE86 Protein Use Independent Mechanisms for Promoter Binding
 JOURNAL OF VIROLOGY, June 2007, p. 5807 5818 Vol. 81, No. 11 0022-538X/07/$08.00 0 doi:10.1128/jvi.02437-06 Copyright 2007, American Society for Microbiology. All Rights Reserved. The Autoregulatory and
JOURNAL OF VIROLOGY, June 2007, p. 5807 5818 Vol. 81, No. 11 0022-538X/07/$08.00 0 doi:10.1128/jvi.02437-06 Copyright 2007, American Society for Microbiology. All Rights Reserved. The Autoregulatory and
Supplementary Figure 1 MicroRNA expression in human synovial fibroblasts from different locations. MicroRNA, which were identified by RNAseq as most
 Supplementary Figure 1 MicroRNA expression in human synovial fibroblasts from different locations. MicroRNA, which were identified by RNAseq as most differentially expressed between human synovial fibroblasts
Supplementary Figure 1 MicroRNA expression in human synovial fibroblasts from different locations. MicroRNA, which were identified by RNAseq as most differentially expressed between human synovial fibroblasts
CD31 5'-AGA GAC GGT CTT GTC GCA GT-3' 5 ' -TAC TGG GCT TCG AGA GCA GT-3'
 Table S1. The primer sets used for real-time RT-PCR analysis. Gene Forward Reverse VEGF PDGFB TGF-β MCP-1 5'-GTT GCA GCA TGA ATC TGA GG-3' 5'-GGA GAC TCT TCG AGG AGC ACT T-3' 5'-GAA TCA GGC ATC GAG AGA
Table S1. The primer sets used for real-time RT-PCR analysis. Gene Forward Reverse VEGF PDGFB TGF-β MCP-1 5'-GTT GCA GCA TGA ATC TGA GG-3' 5'-GGA GAC TCT TCG AGG AGC ACT T-3' 5'-GAA TCA GGC ATC GAG AGA
Culture Density (OD600) 0.1. Culture Density (OD600) Culture Density (OD600) Culture Density (OD600) Culture Density (OD600)
 0.1. Culture Density (OD600) Culture Density (OD600) Culture Density (OD600) Culture Density (OD600)") A. B. C. D. E. PA JSRI JSRI 2 PA DSAM DSAM 2 DSAM 3 PA LNAP LNAP 2 LNAP 3 PAO Fcor Fcor 2 Fcor 3 PAO Wtho Wtho 2 Wtho 3 Wtho 4 DTSB Low Iron 2 4 6 8 2 4 6 8 2 22 DTSB Low Iron 2 4 6 8 2 4 6 8 2 22 DTSB
A. B. C. D. E. PA JSRI JSRI 2 PA DSAM DSAM 2 DSAM 3 PA LNAP LNAP 2 LNAP 3 PAO Fcor Fcor 2 Fcor 3 PAO Wtho Wtho 2 Wtho 3 Wtho 4 DTSB Low Iron 2 4 6 8 2 4 6 8 2 22 DTSB Low Iron 2 4 6 8 2 4 6 8 2 22 DTSB
Abbreviations: P- paraffin-embedded section; C, cryosection; Bio-SA, biotin-streptavidin-conjugated fluorescein amplification.
 Supplementary Table 1. Sequence of primers for real time PCR. Gene Forward primer Reverse primer S25 5 -GTG GTC CAC ACT ACT CTC TGA GTT TC-3 5 - GAC TTT CCG GCA TCC TTC TTC-3 Mafa cds 5 -CTT CAG CAA GGA
Supplementary Table 1. Sequence of primers for real time PCR. Gene Forward primer Reverse primer S25 5 -GTG GTC CAC ACT ACT CTC TGA GTT TC-3 5 - GAC TTT CCG GCA TCC TTC TTC-3 Mafa cds 5 -CTT CAG CAA GGA
Plasmids Western blot analysis and immunostaining Flow Cytometry Cell surface biotinylation RNA isolation and cdna synthesis
 Plasmids psuper-retro-s100a10 shrna1 was constructed by cloning the dsdna oligo 5 -GAT CCC CGT GGG CTT CCA GAG CTT CTT TCA AGA GAA GAA GCT CTG GAA GCC CAC TTT TTA-3 and 5 -AGC TTA AAA AGT GGG CTT CCA GAG
Plasmids psuper-retro-s100a10 shrna1 was constructed by cloning the dsdna oligo 5 -GAT CCC CGT GGG CTT CCA GAG CTT CTT TCA AGA GAA GAA GCT CTG GAA GCC CAC TTT TTA-3 and 5 -AGC TTA AAA AGT GGG CTT CCA GAG
Supplementary Figures
 Supplementary Figures Supplementary Figure 1. H3F3B expression in lung cancer. a. Comparison of H3F3B expression in relapsed and non-relapsed lung cancer patients. b. Prognosis of two groups of lung cancer
Supplementary Figures Supplementary Figure 1. H3F3B expression in lung cancer. a. Comparison of H3F3B expression in relapsed and non-relapsed lung cancer patients. b. Prognosis of two groups of lung cancer
A smart acid nanosystem for ultrasensitive. live cell mrna imaging by the target-triggered intracellular self-assembly
 Electronic Supplementary Material (ESI) for Chemical Science. This journal is The Royal Society of Chemistry 2017 A smart ZnO@polydopamine-nucleic acid nanosystem for ultrasensitive live cell mrna imaging
Electronic Supplementary Material (ESI) for Chemical Science. This journal is The Royal Society of Chemistry 2017 A smart ZnO@polydopamine-nucleic acid nanosystem for ultrasensitive live cell mrna imaging
Table S1. Oligonucleotides used for the in-house RT-PCR assays targeting the M, H7 or N9. Assay (s) Target Name Sequence (5 3 ) Comments
 Target Name Sequence (5 3 ) Comments") SUPPLEMENTAL INFORMATION 2 3 Table S. Oligonucleotides used for the in-house RT-PCR assays targeting the M, H7 or N9 genes. Assay (s) Target Name Sequence (5 3 ) Comments CDC M InfA Forward (NS), CDC M
SUPPLEMENTAL INFORMATION 2 3 Table S. Oligonucleotides used for the in-house RT-PCR assays targeting the M, H7 or N9 genes. Assay (s) Target Name Sequence (5 3 ) Comments CDC M InfA Forward (NS), CDC M
Supplementary Table 2. Conserved regulatory elements in the promoters of CD36.
 Supplementary Table 1. RT-qPCR primers for CD3, PPARg and CEBP. Assay Forward Primer Reverse Primer 1A CAT TTG TGG CCT TGT GCT CTT TGA TGA GTC ACA GAA AGA ATC AAT TC 1B AGG AAA TGA ACT GAT GAG TCA CAG
Supplementary Table 1. RT-qPCR primers for CD3, PPARg and CEBP. Assay Forward Primer Reverse Primer 1A CAT TTG TGG CCT TGT GCT CTT TGA TGA GTC ACA GAA AGA ATC AAT TC 1B AGG AAA TGA ACT GAT GAG TCA CAG
Beta Thalassemia Case Study Introduction to Bioinformatics
 Beta Thalassemia Case Study Sami Khuri Department of Computer Science San José State University San José, California, USA sami.khuri@sjsu.edu www.cs.sjsu.edu/faculty/khuri Outline v Hemoglobin v Alpha
Beta Thalassemia Case Study Sami Khuri Department of Computer Science San José State University San José, California, USA sami.khuri@sjsu.edu www.cs.sjsu.edu/faculty/khuri Outline v Hemoglobin v Alpha
BIOLOGY 621 Identification of the Snorks
 Name: Date: Block: BIOLOGY 621 Identification of the Snorks INTRODUCTION: In this simulation activity, you will examine the DNA sequence of a fictitious organism - the Snork. Snorks were discovered on
Name: Date: Block: BIOLOGY 621 Identification of the Snorks INTRODUCTION: In this simulation activity, you will examine the DNA sequence of a fictitious organism - the Snork. Snorks were discovered on
 www.lessonplansinc.com Topic: Protein Synthesis - Sentence Activity Summary: Students will simulate transcription and translation by building a sentence/polypeptide from words/amino acids. Goals & Objectives:
www.lessonplansinc.com Topic: Protein Synthesis - Sentence Activity Summary: Students will simulate transcription and translation by building a sentence/polypeptide from words/amino acids. Goals & Objectives:
Toluidin-Staining of mast cells Ear tissue was fixed with Carnoy (60% ethanol, 30% chloroform, 10% acetic acid) overnight at 4 C, afterwards
 overnight at 4 C, afterwards") Toluidin-Staining of mast cells Ear tissue was fixed with Carnoy (60% ethanol, 30% chloroform, 10% acetic acid) overnight at 4 C, afterwards incubated in 100 % ethanol overnight at 4 C and embedded in
Toluidin-Staining of mast cells Ear tissue was fixed with Carnoy (60% ethanol, 30% chloroform, 10% acetic acid) overnight at 4 C, afterwards incubated in 100 % ethanol overnight at 4 C and embedded in
BHP 2-7 and Nthy-ori 3-1 cells were grown in RPMI1640 medium (Hyclone) supplemented with 10% fetal bovine serum (Gibco), 2mM L-glutamine, and 100 U/mL
 supplemented with 10% fetal bovine serum (Gibco), 2mM L-glutamine, and 100 U/mL") 1 2 3 4 Materials and Methods Cell culture BHP 2-7 and Nthy-ori 3-1 cells were grown in RPMI1640 medium (Hyclone) 5 supplemented with 10% fetal bovine serum (Gibco), 2mM L-glutamine, and 100 U/mL 6 penicillin-streptomycin.
1 2 3 4 Materials and Methods Cell culture BHP 2-7 and Nthy-ori 3-1 cells were grown in RPMI1640 medium (Hyclone) 5 supplemented with 10% fetal bovine serum (Gibco), 2mM L-glutamine, and 100 U/mL 6 penicillin-streptomycin.
Journal of Cell Science Supplementary information. Arl8b +/- Arl8b -/- Inset B. electron density. genotype
 J. Cell Sci. : doi:.4/jcs.59: Supplementary information E9. A Arl8b /- Arl8b -/- Arl8b Arl8b non-specific band Gapdh Tbp E7.5 HE Inset B D Control al am hf C E Arl8b -/- al am hf E8.5 F low middle high
J. Cell Sci. : doi:.4/jcs.59: Supplementary information E9. A Arl8b /- Arl8b -/- Arl8b Arl8b non-specific band Gapdh Tbp E7.5 HE Inset B D Control al am hf C E Arl8b -/- al am hf E8.5 F low middle high
Supplementary Figure 1
 Supplementary Figure 1 3 3 3 1 1 Bregma -1.6mm 3 : Bregma Ref) Http://www.mbl.org/atlas165/atlas165_start.html Bregma -.18mm Supplementary Figure 1 Schematic representation of the utilized brain slice
Supplementary Figure 1 3 3 3 1 1 Bregma -1.6mm 3 : Bregma Ref) Http://www.mbl.org/atlas165/atlas165_start.html Bregma -.18mm Supplementary Figure 1 Schematic representation of the utilized brain slice
Supplementary Figure 1
 Supplementary Figure 1 Supplementary Figure 1: Cryopreservation alters CD62L expression by CD4 T cells. Freshly isolated (left) or cryopreserved PBMCs (right) were stained with the mix of antibodies described
Supplementary Figure 1 Supplementary Figure 1: Cryopreservation alters CD62L expression by CD4 T cells. Freshly isolated (left) or cryopreserved PBMCs (right) were stained with the mix of antibodies described
Citation for published version (APA): Oosterveer, M. H. (2009). Control of metabolic flux by nutrient sensors Groningen: s.n.
: Oosterveer, M. H. (2009). Control of metabolic flux by nutrient sensors Groningen: s.n.") University of Groningen Control of metabolic flux by nutrient sensors Oosterveer, Maaike IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite from it.
University of Groningen Control of metabolic flux by nutrient sensors Oosterveer, Maaike IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite from it.
Received 9 September 2004/Accepted 10 December 2004
 JOURNAL OF VIROLOGY, Apr. 2005, p. 5035 5046 Vol. 79, No. 8 0022-538X/05/$08.00 0 doi:10.1128/jvi.79.8.5935 5046.2005 Copyright 2005, American Society for Microbiology. All Rights Reserved. Two Gamma Interferon-Activated
JOURNAL OF VIROLOGY, Apr. 2005, p. 5035 5046 Vol. 79, No. 8 0022-538X/05/$08.00 0 doi:10.1128/jvi.79.8.5935 5046.2005 Copyright 2005, American Society for Microbiology. All Rights Reserved. Two Gamma Interferon-Activated
Beta Thalassemia Sami Khuri Department of Computer Science San José State University Spring 2015
 Bioinformatics in Medical Product Development SMPD 287 Three Beta Thalassemia Sami Khuri Department of Computer Science San José State University Hemoglobin Outline Anatomy of a gene Hemoglobinopathies
Bioinformatics in Medical Product Development SMPD 287 Three Beta Thalassemia Sami Khuri Department of Computer Science San José State University Hemoglobin Outline Anatomy of a gene Hemoglobinopathies
Nature Immunology: doi: /ni.3836
 Supplementary Figure 1 Recombinant LIGHT-VTP induces pericyte contractility and endothelial cell activation. (a) Western blot showing purification steps for full length murine LIGHT-VTP (CGKRK) protein:
Supplementary Figure 1 Recombinant LIGHT-VTP induces pericyte contractility and endothelial cell activation. (a) Western blot showing purification steps for full length murine LIGHT-VTP (CGKRK) protein:
SUPPLEMENTARY DATA. Supplementary Table 1. Primer sequences for qrt-pcr
 Supplementary Table 1. Primer sequences for qrt-pcr Gene PRDM16 UCP1 PGC1α Dio2 Elovl3 Cidea Cox8b PPARγ AP2 mttfam CyCs Nampt NRF1 16s-rRNA Hexokinase 2, intron 9 β-actin Primer Sequences 5'-CCA CCA GCG
Supplementary Table 1. Primer sequences for qrt-pcr Gene PRDM16 UCP1 PGC1α Dio2 Elovl3 Cidea Cox8b PPARγ AP2 mttfam CyCs Nampt NRF1 16s-rRNA Hexokinase 2, intron 9 β-actin Primer Sequences 5'-CCA CCA GCG
Nucleotide Sequence of the Australian Bluetongue Virus Serotype 1 RNA Segment 10
 J. gen. Virol. (1988), 69, 945-949. Printed in Great Britain 945 Key words: BTV/genome segment lo/nucleotide sequence Nucleotide Sequence of the Australian Bluetongue Virus Serotype 1 RNA Segment 10 By
J. gen. Virol. (1988), 69, 945-949. Printed in Great Britain 945 Key words: BTV/genome segment lo/nucleotide sequence Nucleotide Sequence of the Australian Bluetongue Virus Serotype 1 RNA Segment 10 By
Supplemental Information. Cancer-Associated Fibroblasts Neutralize. the Anti-tumor Effect of CSF1 Receptor Blockade
 Cancer Cell, Volume 32 Supplemental Information Cancer-Associated Fibroblasts Neutralize the Anti-tumor Effect of CSF1 Receptor Blockade by Inducing PMN-MDSC Infiltration of Tumors Vinit Kumar, Laxminarasimha
Cancer Cell, Volume 32 Supplemental Information Cancer-Associated Fibroblasts Neutralize the Anti-tumor Effect of CSF1 Receptor Blockade by Inducing PMN-MDSC Infiltration of Tumors Vinit Kumar, Laxminarasimha
Analysis of Human Cytomegalovirus orilyt Sequence Requirements in the Context of the Viral Genome
 JOURNAL OF VIROLOGY, Mar. 2005, p. 3615 3626 Vol. 79, No. 6 0022-538X/05/$08.00 0 doi:10.1128/jvi.79.6.3615 3626.2005 Copyright 2005, American Society for Microbiology. All Rights Reserved. Analysis of
JOURNAL OF VIROLOGY, Mar. 2005, p. 3615 3626 Vol. 79, No. 6 0022-538X/05/$08.00 0 doi:10.1128/jvi.79.6.3615 3626.2005 Copyright 2005, American Society for Microbiology. All Rights Reserved. Analysis of
TetR repressor-based bioreporters for the detection of doxycycline using Escherichia
 Supplementary materials TetR repressor-based bioreporters for the detection of doxycycline using Escherichia coli and Acinetobacter oleivorans Hyerim Hong and Woojun Park * Department of Environmental
Supplementary materials TetR repressor-based bioreporters for the detection of doxycycline using Escherichia coli and Acinetobacter oleivorans Hyerim Hong and Woojun Park * Department of Environmental
*To whom correspondence should be addressed. This PDF file includes:
 www.sciencemag.org/cgi/content/full/science.1212182/dc1 Supporting Online Material for Partial Retraction to Detection of an Infectious Retrovirus, XMRV, in Blood Cells of Patients with Chronic Fatigue
www.sciencemag.org/cgi/content/full/science.1212182/dc1 Supporting Online Material for Partial Retraction to Detection of an Infectious Retrovirus, XMRV, in Blood Cells of Patients with Chronic Fatigue
L I F E S C I E N C E S
 1a L I F E S C I E N C E S 5 -UUA AUA UUC GAA AGC UGC AUC GAA AAC UGU GAA UCA-3 5 -TTA ATA TTC GAA AGC TGC ATC GAA AAC TGT GAA TCA-3 3 -AAT TAT AAG CTT TCG ACG TAG CTT TTG ACA CTT AGT-5 OCTOBER 31, 2006
1a L I F E S C I E N C E S 5 -UUA AUA UUC GAA AGC UGC AUC GAA AAC UGU GAA UCA-3 5 -TTA ATA TTC GAA AGC TGC ATC GAA AAC TGT GAA TCA-3 3 -AAT TAT AAG CTT TCG ACG TAG CTT TTG ACA CTT AGT-5 OCTOBER 31, 2006
CIRCRESAHA/2004/098145/R1 - ONLINE 1. Validation by Semi-quantitative Real-Time Reverse Transcription PCR
 CIRCRESAHA/2004/098145/R1 - ONLINE 1 Expanded Materials and Methods Validation by Semi-quantitative Real-Time Reverse Transcription PCR Expression patterns of 13 genes (Online Table 2), selected with respect
CIRCRESAHA/2004/098145/R1 - ONLINE 1 Expanded Materials and Methods Validation by Semi-quantitative Real-Time Reverse Transcription PCR Expression patterns of 13 genes (Online Table 2), selected with respect
Phylogenetic analysis of human and chicken importins. Only five of six importins were studied because
 Supplementary Figure S1 Phylogenetic analysis of human and chicken importins. Only five of six importins were studied because importin-α6 was shown to be testis-specific. Human and chicken importin protein
Supplementary Figure S1 Phylogenetic analysis of human and chicken importins. Only five of six importins were studied because importin-α6 was shown to be testis-specific. Human and chicken importin protein
Supporting Information
 Supporting Information Malapeira et al. 10.1073/pnas.1217022110 SI Materials and Methods Plant Material and Growth Conditions. A. thaliana seedlings were stratified at 4 C in the dark for 3 d on Murashige
Supporting Information Malapeira et al. 10.1073/pnas.1217022110 SI Materials and Methods Plant Material and Growth Conditions. A. thaliana seedlings were stratified at 4 C in the dark for 3 d on Murashige
Cross-talk between mineralocorticoid and angiotensin II signaling for cardiac
 ONLINE SUPPLEMENT TO Crosstalk between mineralocorticoid and angiotensin II signaling for cardiac remodeling An Di ZHANG,,3, Aurelie NGUYEN DINH CAT*,,3, Christelle SOUKASEUM *,,3, Brigitte ESCOUBET, 4,
ONLINE SUPPLEMENT TO Crosstalk between mineralocorticoid and angiotensin II signaling for cardiac remodeling An Di ZHANG,,3, Aurelie NGUYEN DINH CAT*,,3, Christelle SOUKASEUM *,,3, Brigitte ESCOUBET, 4,
Lezione 10. Sommario. Bioinformatica. Lezione 10: Sintesi proteica Synthesis of proteins Central dogma: DNA makes RNA makes proteins Genetic code
 Lezione 10 Bioinformatica Mauro Ceccanti e Alberto Paoluzzi Lezione 10: Sintesi proteica Synthesis of proteins Dip. Informatica e Automazione Università Roma Tre Dip. Medicina Clinica Università La Sapienza
Lezione 10 Bioinformatica Mauro Ceccanti e Alberto Paoluzzi Lezione 10: Sintesi proteica Synthesis of proteins Dip. Informatica e Automazione Università Roma Tre Dip. Medicina Clinica Università La Sapienza
Detection of 549 new HLA alleles in potential stem cell donors from the United States, Poland and Germany
 HLA ISSN 2059-2302 BRIEF COMMUNICATION Detection of 549 new HLA alleles in potential stem cell donors from the United States, Poland and Germany C. J. Hernández-Frederick 1, N. Cereb 2,A.S.Giani 1, J.
HLA ISSN 2059-2302 BRIEF COMMUNICATION Detection of 549 new HLA alleles in potential stem cell donors from the United States, Poland and Germany C. J. Hernández-Frederick 1, N. Cereb 2,A.S.Giani 1, J.
Supplementary Figure 1
 Supplementary Figure 1 Supplementary Figure 1. Lats1/2 deleted ihbs and ihps showed decreased transcripts of hepatocyte related genes (a and b) Western blots (a) and recombination PCR (b) of control and
Supplementary Figure 1 Supplementary Figure 1. Lats1/2 deleted ihbs and ihps showed decreased transcripts of hepatocyte related genes (a and b) Western blots (a) and recombination PCR (b) of control and
Single-Molecule Analysis of Gene Expression Using Two-Color RNA- Labeling in Live Yeast
 Supplemental Figures, Tables and Results Single-Molecule Analysis of Gene Expression Using Two-Color RNA- Labeling in Live Yeast Sami Hocine 1, Pascal Raymond 2, Daniel Zenklusen 2, Jeffrey A. Chao 1 &
Supplemental Figures, Tables and Results Single-Molecule Analysis of Gene Expression Using Two-Color RNA- Labeling in Live Yeast Sami Hocine 1, Pascal Raymond 2, Daniel Zenklusen 2, Jeffrey A. Chao 1 &
Description of Supplementary Files. File Name: Supplementary Information Description: Supplementary Figures and Supplementary Tables
 Description of Supplementary Files File Name: Supplementary Information Description: Supplementary Figures and Supplementary Tables Supplementary Figure 1: (A), HCT116 IDH1-WT and IDH1-R132H cells were
Description of Supplementary Files File Name: Supplementary Information Description: Supplementary Figures and Supplementary Tables Supplementary Figure 1: (A), HCT116 IDH1-WT and IDH1-R132H cells were
A basic helix loop helix transcription factor controls cell growth
 A basic helix loop helix transcription factor controls cell growth and size in root hairs Keke Yi 1,2, Benoît Menand 1,3, Elizabeth Bell 1, Liam Dolan 1,4 Supplementary note Low soil phosphate availability
A basic helix loop helix transcription factor controls cell growth and size in root hairs Keke Yi 1,2, Benoît Menand 1,3, Elizabeth Bell 1, Liam Dolan 1,4 Supplementary note Low soil phosphate availability
CHAPTER 4 RESULTS. showed that all three replicates had similar growth trends (Figure 4.1) (p<0.05; p=0.0000)
 (p<0.05; p=0.0000)") CHAPTER 4 RESULTS 4.1 Growth Characterization of C. vulgaris 4.1.1 Optical Density Growth study of Chlorella vulgaris based on optical density at 620 nm (OD 620 ) showed that all three replicates had similar
CHAPTER 4 RESULTS 4.1 Growth Characterization of C. vulgaris 4.1.1 Optical Density Growth study of Chlorella vulgaris based on optical density at 620 nm (OD 620 ) showed that all three replicates had similar
SUPPLEMENTARY INFORMATION
 doi: 10.1038/nature05883 SUPPLEMENTARY INFORMATION Supplemental Figure 1 Prostaglandin agonists and antagonists alter runx1/cmyb expression. a-e, Embryos were exposed to (b) PGE2 and (c) PGI2 (20μM) and
doi: 10.1038/nature05883 SUPPLEMENTARY INFORMATION Supplemental Figure 1 Prostaglandin agonists and antagonists alter runx1/cmyb expression. a-e, Embryos were exposed to (b) PGE2 and (c) PGI2 (20μM) and
Astaxanthin prevents and reverses diet-induced insulin resistance and. steatohepatitis in mice: A comparison with vitamin E
 Supplementary Information Astaxanthin prevents and reverses diet-induced insulin resistance and steatohepatitis in mice: A comparison with vitamin E Yinhua Ni, 1,2 Mayumi Nagashimada, 1 Fen Zhuge, 1 Lili
Supplementary Information Astaxanthin prevents and reverses diet-induced insulin resistance and steatohepatitis in mice: A comparison with vitamin E Yinhua Ni, 1,2 Mayumi Nagashimada, 1 Fen Zhuge, 1 Lili
Supplementary Figure 1a
 Supplementary Figure 1a Hours: E-cadherin TGF-β On TGF-β Off 0 12 24 36 48 24 48 72 Vimentin βactin Fig. S1a. Treatment of AML12 cells with TGF-β induces EMT. Treatment of AML12 cells with TGF-β results
Supplementary Figure 1a Hours: E-cadherin TGF-β On TGF-β Off 0 12 24 36 48 24 48 72 Vimentin βactin Fig. S1a. Treatment of AML12 cells with TGF-β induces EMT. Treatment of AML12 cells with TGF-β results
Supplementary Materials and Methods
 DD2 suppresses tumorigenicity of ovarian cancer cells by limiting cancer stem cell population Chunhua Han et al. Supplementary Materials and Methods Analysis of publicly available datasets: To analyze
DD2 suppresses tumorigenicity of ovarian cancer cells by limiting cancer stem cell population Chunhua Han et al. Supplementary Materials and Methods Analysis of publicly available datasets: To analyze
Elizabeth A. White, Charles L. Clark, Veronica Sanchez, and Deborah H. Spector*
 JOURNAL OF VIROLOGY, Feb. 2004, p. 1817 1830 Vol. 78, No. 4 0022-538X/04/$08.00 0 DOI: 10.1128/JVI.78.4.1817 1830.2004 Copyright 2004, American Society for Microbiology. All Rights Reserved. Small Internal
JOURNAL OF VIROLOGY, Feb. 2004, p. 1817 1830 Vol. 78, No. 4 0022-538X/04/$08.00 0 DOI: 10.1128/JVI.78.4.1817 1830.2004 Copyright 2004, American Society for Microbiology. All Rights Reserved. Small Internal
Human Cytomegalovirus with IE-2 (UL122) Deleted Fails To Express Early Lytic Genes
 Deleted Fails To Express Early Lytic Genes") JOURNAL OF VIROLOGY, Feb. 2001, p. 1870 1878 Vol. 75, No. 4 0022-538X/01/$04.00 0 DOI: 10.1128/JVI.75.4.1870 1878.2001 Copyright 2001, American Society for Microbiology. All Rights Reserved. Human Cytomegalovirus
JOURNAL OF VIROLOGY, Feb. 2001, p. 1870 1878 Vol. 75, No. 4 0022-538X/01/$04.00 0 DOI: 10.1128/JVI.75.4.1870 1878.2001 Copyright 2001, American Society for Microbiology. All Rights Reserved. Human Cytomegalovirus
Supplemental Information. Th17 Lymphocytes Induce Neuronal. Cell Death in a Human ipsc-based. Model of Parkinson's Disease
 Cell Stem Cell, Volume 23 Supplemental Information Th17 Lymphocytes Induce Neuronal Cell Death in a Human ipsc-based Model of Parkinson's Disease Annika Sommer, Franz Maxreiter, Florian Krach, Tanja Fadler,
Cell Stem Cell, Volume 23 Supplemental Information Th17 Lymphocytes Induce Neuronal Cell Death in a Human ipsc-based Model of Parkinson's Disease Annika Sommer, Franz Maxreiter, Florian Krach, Tanja Fadler,
Supplementary Figure 1
 Metastatic melanoma Primary melanoma Healthy human skin Supplementary Figure 1 CD22 IgG4 Supplementary Figure 1: Immunohisochemical analysis of CD22+ (left) and IgG4 (right), cells (shown in red and indicated
Metastatic melanoma Primary melanoma Healthy human skin Supplementary Figure 1 CD22 IgG4 Supplementary Figure 1: Immunohisochemical analysis of CD22+ (left) and IgG4 (right), cells (shown in red and indicated
ice-cold 70% ethanol with gentle vortexing, incubated at -20 C for 4 hours, and washed with PBS.
 Cell cycle analysis For cell cycle analysis, single cell suspensions of E12.5 fetal liver cells were suspended in 4 ml ice-cold 7% ethanol with gentle vortexing, incubated at -2 C for 4 hours, and washed
Cell cycle analysis For cell cycle analysis, single cell suspensions of E12.5 fetal liver cells were suspended in 4 ml ice-cold 7% ethanol with gentle vortexing, incubated at -2 C for 4 hours, and washed
Baseline clinical characteristics for the 81 CMML patients Routine diagnostic testing and statistical analyses... 3
 Next-Generation Sequencing Technology Reveals a Characteristic Pattern of Molecular Mutations in 72.8% of Chronic Myelomonocytic Leukemia (CMML) by Detecting Frequent Alterations in TET2, CBL, RAS, and
Next-Generation Sequencing Technology Reveals a Characteristic Pattern of Molecular Mutations in 72.8% of Chronic Myelomonocytic Leukemia (CMML) by Detecting Frequent Alterations in TET2, CBL, RAS, and
Eukaryotic transcription (III)
") Eukaryotic transcription (III) 1. Chromosome and chromatin structure Chromatin, chromatid, and chromosome chromatin Genomes exist as chromatins before or after cell division (interphase) but as chromatids
Eukaryotic transcription (III) 1. Chromosome and chromatin structure Chromatin, chromatid, and chromosome chromatin Genomes exist as chromatins before or after cell division (interphase) but as chromatids
Supplemental Figures: Supplemental Figure 1
 Supplemental Figures: Supplemental Figure 1 Suppl. Figure 1. BM-DC infection with H. pylori does not induce cytotoxicity and treatment of BM-DCs with H. pylori sonicate, but not heat-inactivated bacteria,
Supplemental Figures: Supplemental Figure 1 Suppl. Figure 1. BM-DC infection with H. pylori does not induce cytotoxicity and treatment of BM-DCs with H. pylori sonicate, but not heat-inactivated bacteria,
SUPPORTING INFORMATION
 SUPPORTING INFORMATION Biology is different in small volumes: endogenous signals shape phenotype of primary hepatocytes cultured in microfluidic channels Amranul Haque, Pantea Gheibi, Yandong Gao, Elena
SUPPORTING INFORMATION Biology is different in small volumes: endogenous signals shape phenotype of primary hepatocytes cultured in microfluidic channels Amranul Haque, Pantea Gheibi, Yandong Gao, Elena
Supplementary Information
 Supplementary Information Remodeling of heterochromatin structure slows neuropathological progression and prolongs survival in an animal model of Huntington s disease Junghee Lee, Yu Jin Hwang, Yunha Kim,
Supplementary Information Remodeling of heterochromatin structure slows neuropathological progression and prolongs survival in an animal model of Huntington s disease Junghee Lee, Yu Jin Hwang, Yunha Kim,
Expression of Selected Inflammatory Cytokine Genes in Bladder Biopsies
 Borneo Journal of Resource Science and Technology (2013) 3(2): 15-20 Expression of Selected Inflammatory Cytokine Genes in Bladder Biopsies EDMUND UI-HANG SIM *1, NUR DIANA ANUAR 2, TENG-AIK ONG 3, GUAN-
Borneo Journal of Resource Science and Technology (2013) 3(2): 15-20 Expression of Selected Inflammatory Cytokine Genes in Bladder Biopsies EDMUND UI-HANG SIM *1, NUR DIANA ANUAR 2, TENG-AIK ONG 3, GUAN-
L I F E S C I E N C E S
 1a L I F E S C I E N C E S 5 -UUA AUA UUC GAA AGC UGC AUC GAA AAC UGU GAA UCA-3 5 -TTA ATA TTC GAA AGC TGC ATC GAA AAC TGT GAA TCA-3 3 -AAT TAT AAG CTT TCG ACG TAG CTT TTG ACA CTT AGT-5 NOVEMBER 2, 2006
1a L I F E S C I E N C E S 5 -UUA AUA UUC GAA AGC UGC AUC GAA AAC UGU GAA UCA-3 5 -TTA ATA TTC GAA AGC TGC ATC GAA AAC TGT GAA TCA-3 3 -AAT TAT AAG CTT TCG ACG TAG CTT TTG ACA CTT AGT-5 NOVEMBER 2, 2006
Characterizing intra-host influenza virus populations to predict emergence
 Characterizing intra-host influenza virus populations to predict emergence June 12, 2012 Forum on Microbial Threats Washington, DC Elodie Ghedin Center for Vaccine Research Dept. Computational & Systems
Characterizing intra-host influenza virus populations to predict emergence June 12, 2012 Forum on Microbial Threats Washington, DC Elodie Ghedin Center for Vaccine Research Dept. Computational & Systems
The AP-1 binding motifs within the human cytomegalovirus. major immediate-early enhancer are functionally redundant and
 JVI Accepts, published online ahead of print on 24 November 2010 J. Virol. doi:10.1128/jvi.01713-10 Copyright 2010, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights
JVI Accepts, published online ahead of print on 24 November 2010 J. Virol. doi:10.1128/jvi.01713-10 Copyright 2010, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights
HCV Persistence and Immune Evasion in the Absence of Memory T Cell Help.
 SOM Text HCV Persistence and Immune Evasion in the Absence of Memory T Cell Help. Arash Grakoui 1, Naglaa H. Shoukry 2, David J. Woollard 2, Jin-Hwan Han 1, Holly L. Hanson 1, John Ghrayeb 3, Krishna K.
SOM Text HCV Persistence and Immune Evasion in the Absence of Memory T Cell Help. Arash Grakoui 1, Naglaa H. Shoukry 2, David J. Woollard 2, Jin-Hwan Han 1, Holly L. Hanson 1, John Ghrayeb 3, Krishna K.
Cancer Genetics 204 (2011) 45e52
 45e52") Cancer Genetics 204 (2011) 45e52 Exon scanning by reverse transcriptaseepolymerase chain reaction for detection of known and novel EML4eALK fusion variants in nonesmall cell lung cancer Heather R. Sanders
Cancer Genetics 204 (2011) 45e52 Exon scanning by reverse transcriptaseepolymerase chain reaction for detection of known and novel EML4eALK fusion variants in nonesmall cell lung cancer Heather R. Sanders
without LOI phenotype by breeding female wild-type C57BL/6J and male H19 +/.
 Sakatani et al. 1 Supporting Online Material Materials and methods Mice and genotyping: H19 mutant mice with C57BL/6J background carrying a deletion in the structural H19 gene (3 kb) and 10 kb of 5 flanking
Sakatani et al. 1 Supporting Online Material Materials and methods Mice and genotyping: H19 mutant mice with C57BL/6J background carrying a deletion in the structural H19 gene (3 kb) and 10 kb of 5 flanking
Supplementary Information. Bamboo shoot fiber prevents obesity in mice by. modulating the gut microbiota
 Supplementary Information Bamboo shoot fiber prevents obesity in mice by modulating the gut microbiota Xiufen Li 1,2, Juan Guo 1, Kailong Ji 1,2, and Ping Zhang 1,* 1 Key Laboratory of Tropical Plant Resources
Supplementary Information Bamboo shoot fiber prevents obesity in mice by modulating the gut microbiota Xiufen Li 1,2, Juan Guo 1, Kailong Ji 1,2, and Ping Zhang 1,* 1 Key Laboratory of Tropical Plant Resources
The Effect of Epstein-Barr Virus Latent Membrane Protein 2 Expression on the Kinetics of Early B Cell Infection
 The Effect of Epstein-Barr Virus Latent Membrane Protein 2 Expression on the Kinetics of Early B Cell Infection Laura R. Wasil, Monica J. Tomaszewski, Aki Hoji, David T. Rowe* Department of Infectious
The Effect of Epstein-Barr Virus Latent Membrane Protein 2 Expression on the Kinetics of Early B Cell Infection Laura R. Wasil, Monica J. Tomaszewski, Aki Hoji, David T. Rowe* Department of Infectious
PATIENTS AND METHODS. Subjects
 PATIENTS AND METHODS Subjects Twenty-nine morbidly obese subjects involved in a gastric surgery program were enrolled in the study between October 25 and March 21. Bariatric surgery was performed in patients
PATIENTS AND METHODS Subjects Twenty-nine morbidly obese subjects involved in a gastric surgery program were enrolled in the study between October 25 and March 21. Bariatric surgery was performed in patients
Received 13 August 2010/Accepted 10 November 2010
 JOURNAL OF VIROLOGY, Feb. 2011, p. 1732 1746 Vol. 85, No. 4 0022-538X/11/$12.00 doi:10.1128/jvi.01713-10 Copyright 2011, American Society for Microbiology. All Rights Reserved. The Activator Protein 1
JOURNAL OF VIROLOGY, Feb. 2011, p. 1732 1746 Vol. 85, No. 4 0022-538X/11/$12.00 doi:10.1128/jvi.01713-10 Copyright 2011, American Society for Microbiology. All Rights Reserved. The Activator Protein 1
Identification of a Mouse Cytomegalovirus Gene Selectively Targeting CD86 Expression on Antigen-Presenting Cells
 JOURNAL OF VIROLOGY, Dec. 2004, p. 13062 13071 Vol. 78, No. 23 0022-538X/04/$08.00 0 DOI: 10.1128/JVI.78.23.13062 13071.2004 Copyright 2004, American Society for Microbiology. All Rights Reserved. Identification
JOURNAL OF VIROLOGY, Dec. 2004, p. 13062 13071 Vol. 78, No. 23 0022-538X/04/$08.00 0 DOI: 10.1128/JVI.78.23.13062 13071.2004 Copyright 2004, American Society for Microbiology. All Rights Reserved. Identification
Integration Solutions
 Integration Solutions (1) a) With no active glycosyltransferase of either type, an ii individual would not be able to add any sugars to the O form of the lipopolysaccharide. Thus, the only lipopolysaccharide
Integration Solutions (1) a) With no active glycosyltransferase of either type, an ii individual would not be able to add any sugars to the O form of the lipopolysaccharide. Thus, the only lipopolysaccharide
HEK293FT cells were transiently transfected with reporters, N3-ICD construct and
 Supplementary Information Luciferase reporter assay HEK293FT cells were transiently transfected with reporters, N3-ICD construct and increased amounts of wild type or kinase inactive EGFR. Transfections
Supplementary Information Luciferase reporter assay HEK293FT cells were transiently transfected with reporters, N3-ICD construct and increased amounts of wild type or kinase inactive EGFR. Transfections
SUPPLEMENTAL METHODS Cell culture RNA extraction and analysis Immunohistochemical analysis and laser capture microdissection (LCM)
") SUPPLEMENTAL METHODS Cell culture Human peripheral blood mononuclear cells were isolated from healthy donors by Ficoll density gradient centrifugation. Monocyte differentiation to resting macrophages ()
SUPPLEMENTAL METHODS Cell culture Human peripheral blood mononuclear cells were isolated from healthy donors by Ficoll density gradient centrifugation. Monocyte differentiation to resting macrophages ()
Mutation Screening and Association Studies of the Human UCP 3 Gene in Normoglycemic and NIDDM Morbidly Obese Patients
 Mutation Screening and Association Studies of the Human UCP 3 Gene in Normoglycemic and NIDDM Morbidly Obese Patients Shuichi OTABE, Karine CLEMENT, Séverine DUBOIS, Frederic LEPRETRE, Veronique PELLOUX,
Mutation Screening and Association Studies of the Human UCP 3 Gene in Normoglycemic and NIDDM Morbidly Obese Patients Shuichi OTABE, Karine CLEMENT, Séverine DUBOIS, Frederic LEPRETRE, Veronique PELLOUX,
A Rapid and Sensitive Chip-based Assay for Detection of rpob Gene Mutations Conferring Rifampicin Resistance in Mycobacterium tuberculosis (TB).
.") A Rapid and Sensitive Chip-based Assay for Detection of rpob Gene Mutations Conferring Rifampicin Resistance in Mycobacterium tuberculosis (TB). Wanyuan Ao, Steve Aldous, Evelyn Woodruff, Brian Hicke,
A Rapid and Sensitive Chip-based Assay for Detection of rpob Gene Mutations Conferring Rifampicin Resistance in Mycobacterium tuberculosis (TB). Wanyuan Ao, Steve Aldous, Evelyn Woodruff, Brian Hicke,
Loyer, et al. microrna-21 contributes to NASH Suppl 1/15
 Loyer, et al. microrna-21 contributes to NASH Suppl 1/15 SUPPLEMENTARY MATERIAL: Liver MicroRNA-21 is Overexpressed in Non Alcoholic Steatohepatitis and Contributes to the Disease in Experimental Models
Loyer, et al. microrna-21 contributes to NASH Suppl 1/15 SUPPLEMENTARY MATERIAL: Liver MicroRNA-21 is Overexpressed in Non Alcoholic Steatohepatitis and Contributes to the Disease in Experimental Models
The functional investigation of the interaction between TATA-associated factor 3 (TAF3) and p53 protein
 and p53 protein") THESIS BOOK The functional investigation of the interaction between TATA-associated factor 3 (TAF3) and p53 protein Orsolya Buzás-Bereczki Supervisors: Dr. Éva Bálint Dr. Imre Miklós Boros University of
THESIS BOOK The functional investigation of the interaction between TATA-associated factor 3 (TAF3) and p53 protein Orsolya Buzás-Bereczki Supervisors: Dr. Éva Bálint Dr. Imre Miklós Boros University of
SUPPLEMENTARY INFORMATION Glucosylceramide synthase (GlcT-1) in the fat body controls energy metabolism in Drosophila
 in the fat body controls energy metabolism in Drosophila") SUPPLEMENTARY INFORMATION Glucosylceramide synthase (GlcT-1) in the fat body controls energy metabolism in Drosophila Ayako Kohyama-Koganeya, 1,2 Takuji Nabetani, 1 Masayuki Miura, 2,3 Yoshio Hirabayashi
SUPPLEMENTARY INFORMATION Glucosylceramide synthase (GlcT-1) in the fat body controls energy metabolism in Drosophila Ayako Kohyama-Koganeya, 1,2 Takuji Nabetani, 1 Masayuki Miura, 2,3 Yoshio Hirabayashi
Journal of Microbes and Infection,June 2007,Vol 2,No. 2. (HBsAg)2 , (PCR) 1762/ 1764
2 , (PCR) 1762/ 1764") 68 2007 6 2 2 Journal of Microbes and Infection,June 2007,Vol 2,No. 2 2 S 1 1 1 2 2 3 1 (HBsAg)2 ( YIC) S 5 30g 60g YIC ( HBV) DNA > 2 log10 e (HBeAg), 6 DNA, 1 YIC 1, (PCR) (0 ) (44 ) HBV DNA S 2, S a
68 2007 6 2 2 Journal of Microbes and Infection,June 2007,Vol 2,No. 2 2 S 1 1 1 2 2 3 1 (HBsAg)2 ( YIC) S 5 30g 60g YIC ( HBV) DNA > 2 log10 e (HBeAg), 6 DNA, 1 YIC 1, (PCR) (0 ) (44 ) HBV DNA S 2, S a
Influenza viruses are classified as members of the family
 Rewiring the RNAs of influenza virus to prevent reassortment Qinshan Gao a and Peter Palese a,b,1 Departments of a Microbiology and b Medicine, Mount Sinai School of Medicine, New York, NY 10029 Contributed
Rewiring the RNAs of influenza virus to prevent reassortment Qinshan Gao a and Peter Palese a,b,1 Departments of a Microbiology and b Medicine, Mount Sinai School of Medicine, New York, NY 10029 Contributed
Isolation and Genetic Characterization of New Reassortant H3N1 Swine Influenza Virus from Pigs in the Midwestern United States
 JOURNAL OF VIROLOGY, May 2006, p. 5092 5096 Vol. 80, No. 10 0022-538X/06/$08.00 0 doi:10.1128/jvi.80.10.5092 5096.2006 Copyright 2006, American Society for Microbiology. All Rights Reserved. Isolation
JOURNAL OF VIROLOGY, May 2006, p. 5092 5096 Vol. 80, No. 10 0022-538X/06/$08.00 0 doi:10.1128/jvi.80.10.5092 5096.2006 Copyright 2006, American Society for Microbiology. All Rights Reserved. Isolation
Replication and Transcription Activator (RTA) of Murine Gammaherpesvirus 68 Binds to an RTA-Responsive Element and Activates the Expression of ORF18
 of Murine Gammaherpesvirus 68 Binds to an RTA-Responsive Element and Activates the Expression of ORF18") REFERENCES CONTENT ALERTS Replication and Transcription Activator (RTA) of Murine Gammaherpesvirus 68 Binds to an RTA-Responsive Element and Activates the Expression of ORF18 Yun Hong, Jing Qi, Danyang
REFERENCES CONTENT ALERTS Replication and Transcription Activator (RTA) of Murine Gammaherpesvirus 68 Binds to an RTA-Responsive Element and Activates the Expression of ORF18 Yun Hong, Jing Qi, Danyang
Supplementary information
 Supplementary information Human Cytomegalovirus MicroRNA mir-us4-1 Inhibits CD8 + T Cell Response by Targeting ERAP1 Sungchul Kim, Sanghyun Lee, Jinwook Shin, Youngkyun Kim, Irini Evnouchidou, Donghyun
Supplementary information Human Cytomegalovirus MicroRNA mir-us4-1 Inhibits CD8 + T Cell Response by Targeting ERAP1 Sungchul Kim, Sanghyun Lee, Jinwook Shin, Youngkyun Kim, Irini Evnouchidou, Donghyun
What do you think of when you here the word genome?
 What do you think of when you here the word genome? What do you think of when you here the word genome? Personal Genomics Outline Review of pre-lab work Genomics and Medicine Case Overview & Assignment
What do you think of when you here the word genome? What do you think of when you here the word genome? Personal Genomics Outline Review of pre-lab work Genomics and Medicine Case Overview & Assignment
Formylpeptide receptor2 contributes to colon epithelial homeostasis, inflammation, and tumorigenesis
 Supplementary Data Formylpeptide receptor2 contributes to colon epithelial homeostasis, inflammation, and tumorigenesis Keqiang Chen, Mingyong Liu, Ying Liu, Teizo Yoshimura, Wei Shen, Yingying Le, Scott
Supplementary Data Formylpeptide receptor2 contributes to colon epithelial homeostasis, inflammation, and tumorigenesis Keqiang Chen, Mingyong Liu, Ying Liu, Teizo Yoshimura, Wei Shen, Yingying Le, Scott
Bacterial Gene Finding CMSC 423
 Bacterial Gene Finding CMSC 423 Finding Signals in DNA We just have a long string of A, C, G, Ts. How can we find the signals encoded in it? Suppose you encountered a language you didn t know. How would
Bacterial Gene Finding CMSC 423 Finding Signals in DNA We just have a long string of A, C, G, Ts. How can we find the signals encoded in it? Suppose you encountered a language you didn t know. How would
Resistance to Tetracycline Antibiotics by Wangrong Yang, Ian F. Moore, Kalinka P. Koteva, Donald W. Hughes, David C. Bareich and Gerard D. Wright.
 Supplementary Data for TetX is a Flavin-Dependent Monooxygenase Conferring Resistance to Tetracycline Antibiotics by Wangrong Yang, Ian F. Moore, Kalinka P. Koteva, Donald W. Hughes, David C. Bareich and
Supplementary Data for TetX is a Flavin-Dependent Monooxygenase Conferring Resistance to Tetracycline Antibiotics by Wangrong Yang, Ian F. Moore, Kalinka P. Koteva, Donald W. Hughes, David C. Bareich and
Construction of a hepatocellular carcinoma cell line that stably expresses stathmin with a Ser25 phosphorylation site mutation
 Construction of a hepatocellular carcinoma cell line that stably expresses stathmin with a Ser25 phosphorylation site mutation J. Du 1, Z.H. Tao 2, J. Li 2, Y.K. Liu 3 and L. Gan 2 1 Department of Chemistry,
Construction of a hepatocellular carcinoma cell line that stably expresses stathmin with a Ser25 phosphorylation site mutation J. Du 1, Z.H. Tao 2, J. Li 2, Y.K. Liu 3 and L. Gan 2 1 Department of Chemistry,
Development of RT-qPCR-based molecular diagnostic assays for therapeutic target selection of breast cancer patients
 Development of RT-qPCR-based molecular diagnostic assays for therapeutic target selection of breast cancer patients Sangjung Park The Graduate School Yonsei University Department of Biomedical Laboratory
Development of RT-qPCR-based molecular diagnostic assays for therapeutic target selection of breast cancer patients Sangjung Park The Graduate School Yonsei University Department of Biomedical Laboratory
Supporting Information. Mutational analysis of a phenazine biosynthetic gene cluster in
 Supporting Information for Mutational analysis of a phenazine biosynthetic gene cluster in Streptomyces anulatus 9663 Orwah Saleh 1, Katrin Flinspach 1, Lucia Westrich 1, Andreas Kulik 2, Bertolt Gust
Supporting Information for Mutational analysis of a phenazine biosynthetic gene cluster in Streptomyces anulatus 9663 Orwah Saleh 1, Katrin Flinspach 1, Lucia Westrich 1, Andreas Kulik 2, Bertolt Gust
Recommended laboratory tests to identify influenza A/H5 virus in specimens from patients with an influenza-like illness
 World Health Organization Recommended laboratory tests to identify influenza A/H5 virus in specimens from patients with an influenza-like illness General information Highly pathogenic avian influenza (HPAI)
World Health Organization Recommended laboratory tests to identify influenza A/H5 virus in specimens from patients with an influenza-like illness General information Highly pathogenic avian influenza (HPAI)
Supplemental Materials and Methods Plasmids and viruses Quantitative Reverse Transcription PCR Generation of molecular standard for quantitative PCR
 Supplemental Materials and Methods Plasmids and viruses To generate pseudotyped viruses, the previously described recombinant plasmids pnl4-3-δnef-gfp or pnl4-3-δ6-drgfp and a vector expressing HIV-1 X4
Supplemental Materials and Methods Plasmids and viruses To generate pseudotyped viruses, the previously described recombinant plasmids pnl4-3-δnef-gfp or pnl4-3-δ6-drgfp and a vector expressing HIV-1 X4
Human cytomegalovirus sequences expressed in latently infected individuals promote a latent infection in vitro
 IMMUNOBIOLOGY Human cytomegalovirus sequences expressed in latently infected individuals promote a latent infection in vitro Felicia Goodrum, 1 Matthew Reeves, 2 John Sinclair, 2 Kevin High, 3 and Thomas
IMMUNOBIOLOGY Human cytomegalovirus sequences expressed in latently infected individuals promote a latent infection in vitro Felicia Goodrum, 1 Matthew Reeves, 2 John Sinclair, 2 Kevin High, 3 and Thomas