Trafficking and Replication Patterns Reveal Splenic. Macrophages As Major Targets of Dengue Virus in Mice
|
|
|
- Lindsey Stevenson
- 5 years ago
- Views:
Transcription
1 JVI Accepts, published online ahead of print on 29 August 2012 J. Virol. doi: /jvi Copyright 2012, American Society for Microbiology. All Rights Reserved. 1 2 Trafficking and Replication Patterns Reveal Splenic Macrophages As Major Targets of Dengue Virus in Mice Tyler R. Prestwood, Monica M. May, Emily M. Plummer, Malika M. Morar, Lauren E. Yauch, and Sujan Shresta* Division of Vaccine Discovery, La Jolla Institute for Allergy and Immunology, 9420 Athena Circle, La Jolla, CA 92037, USA * Correspondence and requests for materials should be addressed to S.S.: sujan@liai.org Running Title: Dengue virus infects splenic macrophages in mice Word Count (abstract): 223 Word Count (text): 5,500
2 ABSTRACT Human post-mortem studies of natural dengue virus (DENV) infection have reported systemically distributed viral antigen. Although it is widely accepted that DENV infects mononuclear phagocytes, the sequence in which specific tissues and cell types are targeted remains uncharacterized. We have previously reported that mice lacking interferon-α/β and -γ receptors permit high levels of DENV replication and show signs of systemic disease (28). Here, we demonstrate that within 6 hours, DENV traffics to and replicates in both CD169 + and SIGN-R1 + macrophages of the splenic marginal zone or draining lymph node, respectively, following intravenous or intrafootpad inoculation. Subsequently, high levels of replication are detected in F4/80 + splenic red pulp macrophages and in the bone marrow, lymph nodes, and Peyer s patches. Intravenously inoculated mice begin to succumb to dengue disease 72 hours after infection, at which time viral replication occurs systemically, except in lymphoid tissues. In particular, high levels of replication occur in CD68 + macrophages of the kidneys, heart, thymus, and gastrointestinal tract. Over the course of infection, proportionately large quantities of DENV traffic to the liver and spleen. However, late during infection, viral trafficking to the spleen decreases, while trafficking to the liver, thymus and kidneys increases. The present study demonstrates that macrophage populations, initially in the spleen and other lymphoid tissues, and later in non-lymphoid tissues, are major targets of DENV infection in vivo. 2
3 INTRODUCTION The four serotypes of dengue virus (DENV1-4) belong to the Flaviviridae family and cause a mosquito-borne, febrile illness in more than 50 million people per year. In nearly 500,000 cases per year, individuals develop severe hemorrhage and sometimes shock, resulting in more than 25,000 deaths annually (12). The 10.7 kb, positive-stranded RNA genome of DENV encodes a polyprotein that is cleaved into 3 structural proteins, envelope (E), pre-membrane/membrane (prm/m), and capsid (C), and 7 nonstructural (NS) proteins, NS1, NS2A, NS2B, NS3, NS4A, NS4B, and NS5. During replication, all DENV proteins and both positive- and negative-sense DENV RNA are produced intracellularly, while only positive-sense RNA, the structural proteins, and NS1 are known to be secreted at high levels. Thus, based on current knowledge, NS proteins, with the exception of NS1, are predominantly restricted to cells in which DENV replicates. The cellular tropism of DENV has been investigated in humans (2, 5, 6, 8, 13, 15, 16, 25, 29), and mice (3, 9, 20, 38). In human studies, DENV antigen has been reported in the skin, liver, brain, kidney, spleen, lymph nodes, lungs, stomach, and intestine (2, 5, 6, 8, 13, 15, 16, 25, 29). These studies have mainly relied upon detection of secreted DENV antigens, which may actually localize elsewhere, complicating interpretation of results. Recently, two studies have detected DENV NS antigen in the lymph nodes, spleen, liver, and bone marrow cells of mice (2, 9). With the exception of the study by Fink et al. (9), infected cell types have not been identified by co-staining for specific marker expression. Nevertheless, these studies have consistently reported the major targets of DENV to be 3
4 66 67 mononuclear phagocytes. The exact cell populations targeted and sequence of infection at the tissue and cellular level, however, remains unclear Previously, we reported that 129/Sv mice lacking type I and II interferon receptors (AG129 mice) are susceptible to lethal disease when challenged with a mouse-passaged DENV2 strain (28, 32, 38). In this experimental setup, the kinetics of DENV dissemination was followed by qrt-pcr (28, 38). Here, we use fluorescent immunohistochemistry (FIHC) to characterize the sequence in which specific tissues and cell types are targeted by analyzing nearly every major tissue for DENV NS3 at several time points over the course of infection and by determining the precise identity of NS3- expressing cells through co-staining with cell type-specific markers. Additionally, we examine viral trafficking using DENV-labeled with the fluorescent lipophilic dye DiD to quantify DENV extravasation at several time points over the course of infection. The present study characterizes the kinetics of DENV infection and trafficking at the tissue and cellular level in vivo. 4
5 MATERIALS AND METHODS Mice 129/Sv mice deficient in the IFN-α/β receptor (A129) or the IFN-α/β receptor and the IFN-γ receptor (AG129) were housed under specific pathogen-free conditions. All experiments were approved by the Animal Care Committee at the La Jolla Institute for Allergy and Immunology (LIAI). Mice between 5 and 6 weeks of age were inoculated intravenously (i.v.) in the lateral tail vein with 1.5x10 11 GE (3x10 6 PFU) or 5x10 8 GE (1x10 4 PFU) in 200 µl or intrafootpad (i.f.) with 1.5x10 11 GE in 30 µl of the DENV2 strain E124/128-IC diluted in PBS (Invitrogen) containing 10% fetal bovine serum (FBS, Gemini Bio-Products). For DiD-labeled virus experiments, mice were additionally inoculated i.v. 2 hours before harvesting tissues. Cell cultures and viral stocks C6/36 cells were obtained from the American Type Culture Collection and cultured at 28 C in Leibovitz s L-15 medium (Gibco) supplemented with penicillin, streptomycin, HEPES, and 10% FBS. DENV2 strain E124/128-IC (from pe124/128-ic) was generated as previously described (28). Briefly, an infectious cdna clone of PL046 (ppl046-ic) was previously constructed and site-directed mutagenesis was performed to introduce the two mutations identified in D2S10. The plasmid was linearized with XbaI and gel purified using a QIAquick gel extraction kit (Qiagen). Infectious RNA was generated by in vitro transcription with a RiboMax large-scale RNA production system (T7, Promega) and transfected into BHK-21 cells with Lipofectamine (Invitrogen). The entire genome of the viral stocks was confirmed by sequencing. Virus was amplified twice in C6/36 cells 5
6 and purified using a sucrose density gradient, as previously described (28). Genomic equivalents (GE) were quantified by real-time reverse transcription-polymerase chain reaction (RT-PCR) DiD-labeled DENV stocks Solid 1,1'-dioctadecyl-3,3,3',3'-tetramethylindodicarbocyanine, 4-chlorobenzenesulfonate salt (DiD, Invitrogen) was reconstituted at 20 mg/ml in DMSO and then diluted 1:10 in 100% ethanol. Twice-amplified stocks of E124/128-IC were used to grow stocks for DiD labeling. The third amplification was performed in CELLine AD 1000 adherent cell chambers (Wilson Wolf) and virus in the supernatant was concentrated by ultracentrifugation, resuspended, and purified over a sucrose density gradient. After dialysis against PBS, virus was incubated with DiD at a 1:38,000 molar ratio of virus:did for 1 hour at room temperature. After incubation, virus was re-purified over a sucrose density gradient, which yielded a distinct band of labeled virus. Virus was dialyzed against PBS, aliquoted, and stored at -80ºC. Infectivity of DiD-labeled virus was reduced to 44% of the unlabeled virus when normalized to GE. Immunohistochemistry Tissues were embedded in O.C.T. compound (Sakura). Sections (6 μm) were cut and stored at -80 C. Frozen sections were thawed and fixed for 10 min in acetone at 25 C, followed by 8 min on ice in 1% paraformaldehyde in 100 mm dibasic sodium phosphate (ph 7.4) containing 60 mm lysine and 7 mm sodium periodate. 6
7 To stain for DENV replication, tissue sections were blocked with Avidin/Biotin Blocking Kit (Vector), 5% normal goat serum (Caltag Laboratories) and 5% mouse serum in PBS. Sections were stained overnight with affinity purified rabbit polyclonal anti-denv NS3 (a generous gift from the Novartis Institute for Tropical Diseases, Singapore) and purified mouse anti-denv prm/m (clone 2H2) labeled with EZ-Link NHS Biotin (Pierce). After washing, sections were stained with DyLight 649-labeled goat anti-rabbit IgG (Jackson Immunoresearch) and PE-labeled streptavidin (ebioscience). NS3 staining with DiD visualization For DiD and NS3 staining experiments, sections were prepared, fixed, and blocked as stated above and then stained overnight with purified rabbit polyclonal anti-denv NS3. After washing, sections were stained with DyLight 549-labeled goat anti-rabbit IgG (Jackson Immunoresearch). Marginal zone macrophage staining with DiD visualization Sections were prepared, fixed, and blocked as stated above and then stained with rat antimouse CD169 (clone MOMA-1, BMA Biomedicals), biotinylated rat anti-mouse SIGN- R1 (clone ER-TR9, BMA Biomedicals), and rat anti-mouse MARCO (clone ED31, AbD Serotec). After washing, sections were stained with biotinylated goat anti-rat IgG (BD), followed by additional washing and staining of sections with PE-labeled streptavidin (BD). 7
8 Identification of infected splenocytes Sections were prepared, fixed, and blocked as stated above. For marginal zone macrophage markers, sections were stained overnight with rat anti-mouse CD169 (clone MOMA-1), biotinylated rat anti-mouse SIGN-R1 (clone ER-TR9), and purified rabbit polyclonal anti-denv NS3. After washing, sections were stained with PE-conjugated mouse anti-rat IgG2a (BD), FITC-conjugated streptavidin (BD), and DyLight 649- labeled goat anti-rabbit IgG. Finally, after additional washing, sections were stained with AF488-conjugated goat anti-fitc (Invitrogen). For CD11b and CD11c, sections were stained overnight with FITC-labeled Armenian hamster anti-mouse CD11c (clone HL3, BD), PE-labeled rat anti-mouse CD11b (clone M1/70, BD), and purified rabbit polyclonal anti-denv NS3. After washing, sections were stained with PE-conjugated goat anti-rat IgG, AF488-conjugated goat anti-fitc, and DyLight 649-labeled goat anti-rabbit IgG (Jackson ImmunoResearch Laboratories). Lastly, for pan-macrophage and red pulp macrophage markers, sections were stained overnight with FITC-labeled rat anti-f4/80 (ebioscience), biotinylated rat anti-mouse CD68 (clone FA-11, AbD Serotec), and purified rabbit polyclonal anti-denv NS3. After washing, sections were stained with PE-conjugated streptavidin, AF488-conjugated goat anti-fitc, and DyLight 649-labeled goat anti-rabbit IgG. 8
9 Identification of infected cells in lymph nodes Sections were prepared, fixed, and blocked as stated above. For MARCO and SIGN-R1 staining, sections were stained overnight with purified rat anti-marco (clone ED31, AbD Serotec), biotinylated rat anti-mouse SIGN-R1 (clone ER-TR9), and purified rabbit polyclonal anti-denv NS3. After washing, sections were stained with PE-conjugated streptavidin (BD), FITC-conjugated mouse anti-rat IgG1 (BD), and DyLight 649-labeled goat anti-rabbit IgG (Jackson ImmunoResearch Laboratories). For CD68 and CD169 staining, sections were stained overnight with biotinylated rat antimouse CD68 (clone FA-11, AbD Serotec), purified rat anti-mouse CD169 (clone MOMA-1), and purified rabbit polyclonal anti-denv NS3. After washing, sections were stained PE-conjugated mouse anti-rat IgG2a (BD), FITC-conjugated streptavidin (BD), and DyLight 649-labeled goat anti-rabbit IgG (Jackson ImmunoResearch Laboratories). Comparison of infected spleen cells in A129 and AG129 mice Tissues sections were prepared, fixed, and blocked as stated above. For marginal zone macrophage markers, sections were stained overnight with rat anti-mouse CD169 (clone MOMA-1), biotinylated rat anti-mouse SIGN-R1 (clone ER-TR9), and purified rabbit polyclonal anti-denv NS3. After washing, sections were stained with PE-conjugated mouse anti-rat IgG2a and FITC-conjugated goat anti-rabbit IgG. Finally, after additional washing, sections were stained with AF488-conjugated goat anti-fitc and APC-labeled streptavidin (ebioscience). 9
10 For pan-macrophage and red pulp macrophage markers, sections were stained overnight with PE-labeled rat anti-f4/80 (clone BM8, Biolegend), biotinylated rat anti-mouse CD68 (clone FA-11, AbD Serotec), and purified rabbit polyclonal anti-denv NS3. After washing, sections were stained with FITC-conjugated goat anti-rabbit IgG. Finally, after additional washing, sections were stained with AF488-conjugated goat anti-fitc and APC-labeled streptavidin. Identification of infected cells in the kidney, heart and thymus For cell type identification, sections were prepared, fixed, and blocked as stated above. Sections were stained overnight with FITC-labeled rat anti-mouse CD11b (clone M1/70), biotinylated rat anti-mouse CD68 (clone FA-11), and purified rabbit polyclonal anti- DENV NS3. After washing, sections were stained with PE-conjugated streptavidin, AF488-conjugated goat anti-fitc, and DyLight 649-labeled goat anti-rabbit IgG. Image acquisition and preparation Images were recorded using a Marianas deconvolution fluorescence microscope (3i) or DM6000B upright laser scanning confocal microscope (Leica). Final preparation of all images was performed using ImageJ software (NIH), Adobe Photoshop, and Adobe Illustrator (Adobe Systems). Quantification of DiD For quantitative analysis, images were processed with the noise reduction despeckle function in ImageJ. DiD intensity was then quantified using the Batch>Measure function 10
11 for 6-24 non-overlapping low-power images from 2 tissues sections for each of the 5 mice per group per time point. Data were graphed and analyzed with Prism 5.0 (GraphPad). Statistical significance was determined using the unpaired t-test
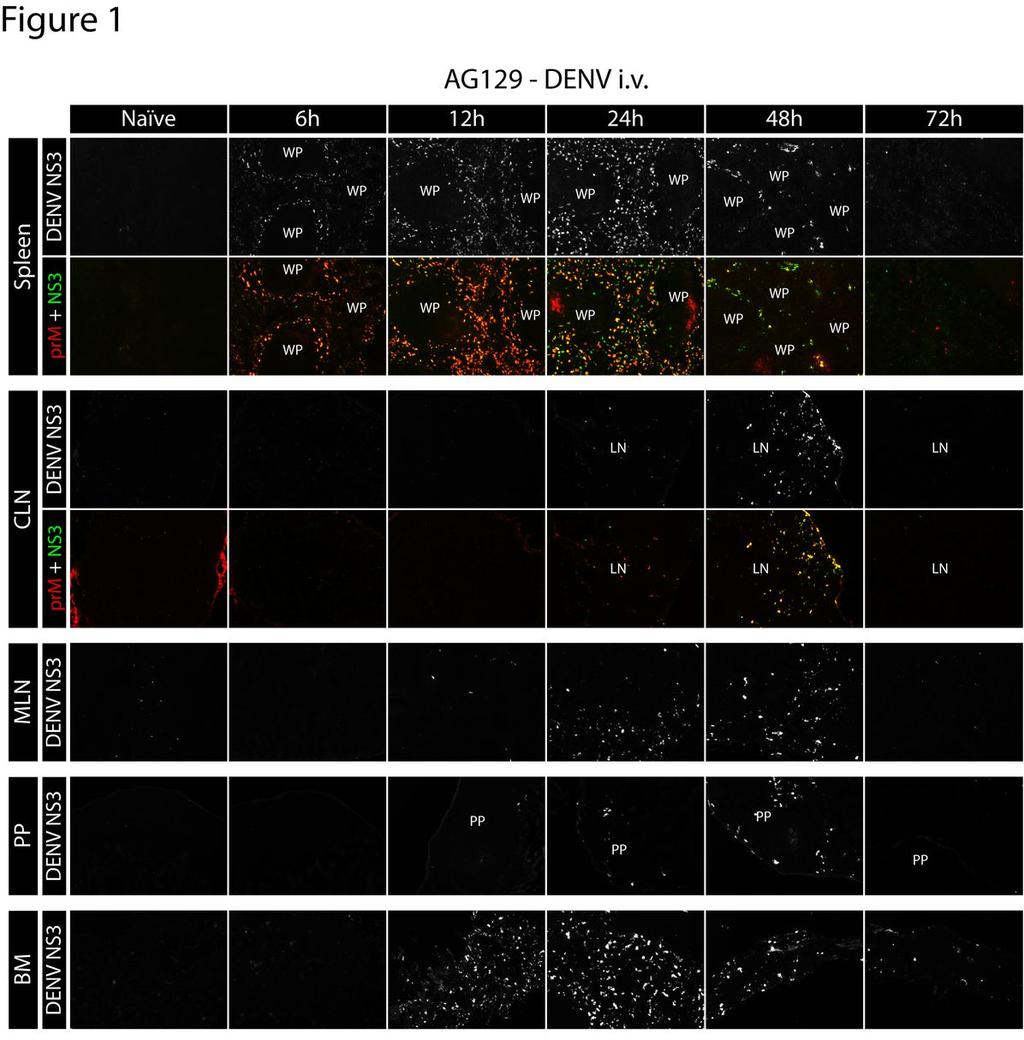
12 RESULTS Early DENV replication occurs in lymphoid tissues We previously reported that serum levels of DENV virions decrease rapidly following intravenous (i.v.) inoculation of AG129 mice (28). However, levels of DENV RNA in the serum begin to increase between 5 and 7 hours after infection, suggesting that initial viral replication in vivo occurs in less than 7 hours. To screen for DENV replication patterns characteristic of lethal infection and to coordinate with our previous RNA kinetics study (28), we utilized a high dose of DENV. AG129 mice were inoculated i.v. with 1.5x10 11 genomic equivalents (GE) of the DENV2 strain E124/128-IC. At 0 (naïve mice), 6, 12, 24, 48, and 72 hours after infection, the following tissues were collected: liver, heart, lung, thymus, spleen, kidney, stomach, small intestine (including Peyer s patches), large intestine, mesenteric lymph nodes (MLN), cutaneous lymph nodes (CLN: brachial, axillary, inguinal LNs were pooled because of equivalent trends), skin, blood, spinal cord, brain, and bone marrow. For analysis of viral replication, 6 µm frozen tissue sections were cut, mounted, and subsequently stained with rabbit polyclonal antibodies raised against purified NS3, in combination with biotinylated mouse anti-denv prm/m monoclonal antibody (clone 2H2). By 6 hours after infection, DENV replication was detected in the spleen (Figure 1), at which time both NS3 and prm/m were present in the marginal zone encircling the white pulp. At 12 hours after infection, NS3 expression was still present in the marginal zone of the spleen, although the circular staining pattern had extended into the red pulp and become more diffuse. At 24 hours after infection, DENV NS3 was distributed 12
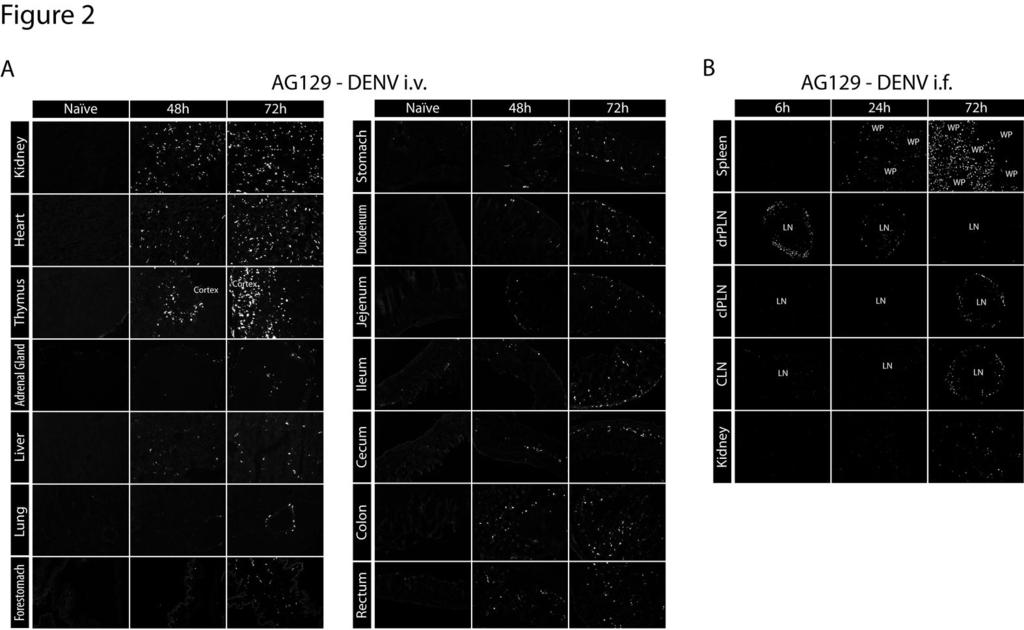
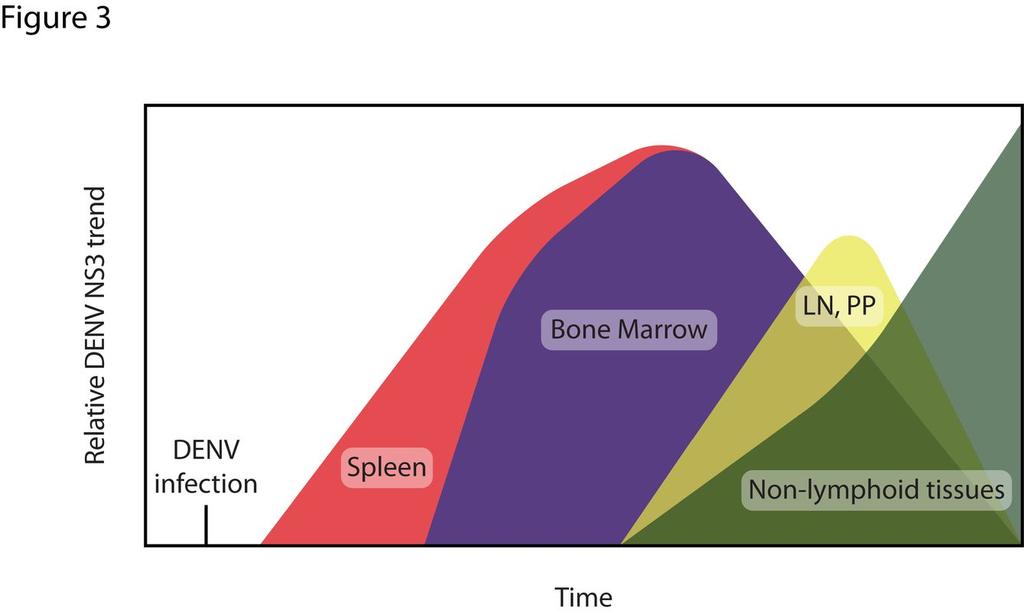
13 throughout the red pulp. By 48 and 72 hours after infection, levels of NS3 in the spleen had decreased substantially, but low levels continue to be detectable in the red pulp at the 72 hour time point. Staining with the anti-denv prm/m antibody 2H2 showed near complete colocalization with anti-ns3 staining, confirming specificity to sites of DENV replication. The kinetics of DENV infection in other lymphoid tissues was similar to that observed in the spleen (Figure 1). In the bone marrow, NS3 expression became detectable at 12 hours and peaked at 24 hours after infection. In the lymph nodes and Peyer s patches, infection was consistently observed 24 hours after inoculation and peaked at 48 hours. As in the spleen, infection in the bone marrow, lymph nodes, and Peyer s patches decreased substantially and became almost undetectable by 72 hours. Collectively, these results demonstrate that systemic lymphoid tissues are targeted by DENV before nonlymphoid tissues in this model. Late DENV replication occurs in non-lymphoid tissues We additionally examined DENV replication in non-lymphoid tissues including the kidney, heart, thymus, adrenal gland, liver, lung, forestomach, stomach, jejunum, duodenum, ileum, cecum, colon, and rectum (Figure 2A). In contrast to replication in lymphoid tissues, replication was undetectable in non-lymphoid tissues early after infection, but became detectable between 24 and 48 hours and continued to increase until 72 hours (Figure 2A). Notably, a high level of DENV NS3 expression was detected in the heart, thymus, and kidneys. Several of the non-lymphoid tissues analyzed, including the 13
14 brain, spinal cord, esophagus, blood, bladder, and skin, showed little or no viral replication, even 72 hours after infection (data not shown). Based on these data, a model of DENV dissemination was developed (Figure 3). These findings are in agreement with our previous kinetic study measuring DENV RNA levels in tissues (28). DENV replication occurs in the draining popliteal lymph node following intrafootpad inoculation To validate our results using a more relevant route of infection than the i.v. route (34), AG129 mice were inoculated intrafootpad with 1.5x10 11 GE of E124/128-IC. The kinetics of systemic DENV dissemination was assessed by monitoring viral RNA levels in the serum at 6, 12, 24, 48, 72 and 96 hours after infection. Levels of DENV RNA were almost undetectable at 6 hours but increased steadily, peaking at 72 hours and decreasing by 96 hours after infection (data not shown). Based on these kinetics, the spleen, draining popliteal lymph node (PLN), contralateral PLN, cutaneous lymph nodes (CLN), and kidneys were harvested 6, 24, and 72 hours after infection. Tissues were stained for the presence of DENV NS3. At 6 hours after infection, NS3 expression was detected only in the draining PLN (Figure 2B). By 24 hours, NS3 expression was observed at a reduced level in the draining PLN, while it became detectable in the marginal zone of the spleen. By 72 hours, NS3 expression was detected in the contralateral PLN as well as distant CLNs, splenic red pulp, and kidneys, but not the draining PLN. These results show that the pattern of DENV replication (as determined by staining for NS3 expression) following intrafootpad inoculation mirrors results obtained with intravenous inoculation: early replication in the draining lymphoid tissue 14
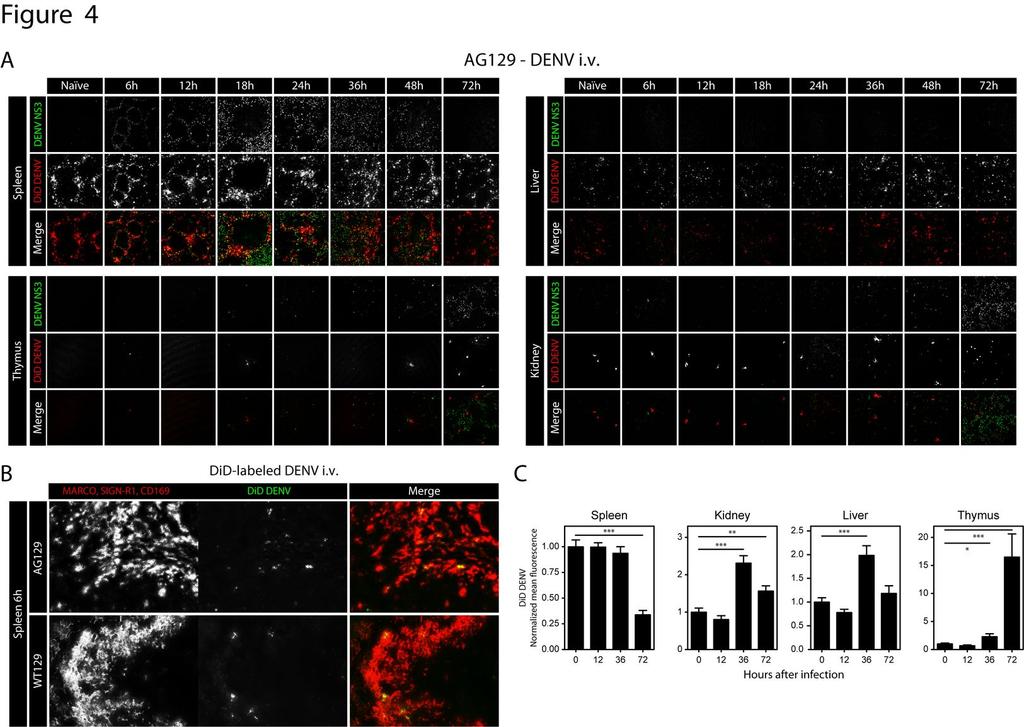
15 followed by systemic dissemination to other lymphoid tissues, and lastly, replication in non-lymphoid tissues Intravenous DENV particles traffic to macrophages in the marginal zone of the spleen Based on the pattern of initial replication in the splenic marginal zone and subsequent dissemination to the red pulp, we hypothesized that the ability of the splenic marginal zone to absorb blood-borne DENV particles diminishes over the course of infection, resulting in leakage of viral particles into the red pulp. To investigate this hypothesis, mice were inoculated i.v. with 1.5x10 11 GE E124/128-IC, as in previous experiments, and administered an additional 5x10 10 GE of E124/128-IC labeled with DiD, a lipophilic farred fluorescent dye, 2 hours before tissue harvest at 0 (naïve mice), 6, 12, 18, 24, 36, 48, and 72 hours after the initial inoculation (Figure 4A). When large amounts of DiD are incorporated into the lipid bilayer of the virion, it undergoes autoquenching and emits only low levels of fluorescence. However, once the lipid bilayer fuses with the host cell membrane, the probe is diluted and renders target cells highly fluorescent. In naïve mice and mice 6 hours after DENV infection, DiD signal in the spleen was restricted to the marginal zone (Figure 4A). DiD distribution became more diffuse in spleens of mice 12 and 18 hours after infection. After 24 hours, dissemination of DENV particles was readily observed throughout the red pulp. However, at all time points after infection, the majority of the DiD accumulated in rings within the marginal zone of the spleen, implying that DENV particles primarily trafficked here throughout infection, even at time points when most of the NS3 signal was located in the red pulp (Figure 4A, hours). 15
16 Because macrophages of the marginal zone often serve to filter blood-borne virus, we speculated that these cells were the initial targets of intravenous DENV. To investigate this, AG129 and WT129 mice were inoculated with 5x10 10 GE of DiD-labeled E124/128- IC, and spleens were harvested 6 hours later. Tissues were sectioned and stained with antibodies against MARCO, CD169 and SIGN-R1, which label the heterogenous pool of macrophages in the marginal zone. Nearly all of the DiD signal in both AG129 and wildtype 129/SV (WT129) mice (Figure 4B) localized to cells expressing marginal zone macrophage markers, implicating these cells as the initial targets of DENV in this model, regardless of host immune status. DENV trafficking to the liver, kidneys and thymus increases over the course of infection, while trafficking to the spleen decreases To determine how viral trafficking and fusion change with time in the major target tissues, the spleen, liver, thymus, and kidneys were analyzed for the presence of DiD and DENV NS3 (Figure 4A). DiD signal intensity was quantified on 6 24 images of the spleen, liver, kidneys, and thymus for 5 mice at 0 (naïve mice), 12, 36, and 72 hours after infection. These results are summarized in Figure 4C. Large amounts of DENV trafficked to the spleen and liver, with the amounts decreasing in the spleen and increasing in the liver over the course of infection. Similar to the liver, the kidneys and thymus had increasing levels of DiD at late time points after infection, albeit less virus trafficked to these tissues than to the spleen and liver. These data show that the bloodborne DENV trafficking pattern changes during infection, favoring early dissemination to lymphoid tissues and late dissemination to non-lymphoid tissues. 16
17 DENV initially infects splenic macrophages in the marginal zone, and later spreads to red pulp macrophages In order to identify the DENV-permissive cells in the spleen of AG129 mice, we sought to distinguish between macrophages, monocytes and dendritic cells (DCs), all of which have been reported to be targeted by DENV in humans. Macrophages in the spleen exist as several heterogeneous, overlapping populations (23). Sialoadhesin, or CD169, is expressed by marginal metallophilic macrophages (MMMs), which reside closest to the white pulp in the innermost part of the marginal zone. SIGN-R1 (a murine homolog of DC-SIGN) and MARCO are expressed by two highly overlapping populations of macrophages in the outer part of the marginal zone, collectively termed marginal zone macrophages (MZMs). We have observed that some of the outer MARCO and SIGN- R1 expressing MZMs also express F4/80. Additionally, we have observed a marked upregulation of MARCO expression on F4/80 + red pulp macrophages between 12 and 24 hours after infection (data not shown). MARCO was therefore not used to identify splenocytes in this study. Lastly, splenic red pulp macrophages (RPMs) can be distinguished based on their high level of F4/80 expression and lack of MZM and MMM markers. Thus, to differentiate the mononuclear phagocytes potentially permissive to DENV infection, we utilized CD11c (dendritic cell marker), CD11b (present at high levels on monocytes and neutrophils), CD169 and SIGN-R1 (present on macrophages of the marginal zone), F4/80 (present on macrophages of the red pulp), and CD68 (expressed at high levels in macrophages). 17
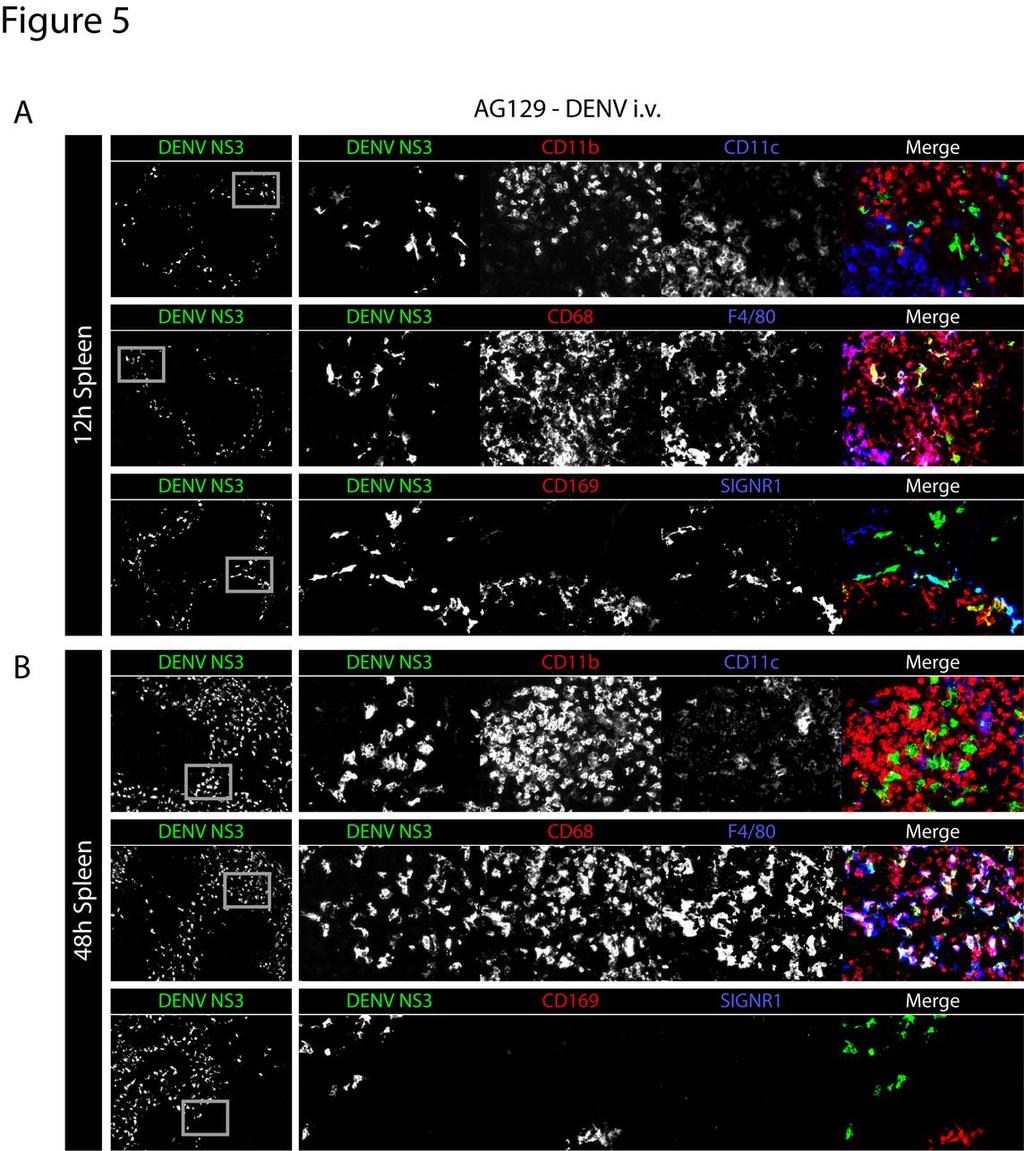
18 Using a sublethal dose of virus (5x10 8 GE of E124/128-IC) to more accurately model a natural human infection, AG129 mice were inoculated intravenously and spleens were harvested 12 and 48 hours after infection. At this dose, replication was observed selectively in the marginal zone at 12 hours. DENV NS3-expressing cells were distinct from cells expressing high levels of CD11b or CD11c (Figure 5A, top row). Instead, NS3 localized to cells expressing CD68, some of which also expressed F4/80 (Figure 5A, middle row) and most of which were also found to express CD169 and/or SIGN-R1, identifying them as MMMs and MZMs, respectively, (Figure 5A, bottom row). At 48 hours, DENV NS3-expressing cells did not express high levels of CD11b, CD11c, CD169 or SIGN-R1 (Figure 5B, top and bottom rows), but instead expressed high levels of CD68 and F4/80 (Figure 5B, middle row). This cell marker expression and cell localization is consistent with RPMs. Thus, in this model of primary DENV infection, virus replicates early in the spleen in the MZMs and MMMs, and later in RPMs. Additionally, we did not observe DENV replication in CD11c high-expressing dendritic cells or CD11b high-expressing monocytes or neutrophils. Interferon-γ signaling restricts DENV replication in macrophages of the red pulp, but not in macrophages of the marginal zone To validate our tropism findings in less immunocompromised mice than AG129, we next sought to determine the pattern of DENV replication in the spleens of A129 mice, which lack the interferon-α/β receptor, but have functional IFN-γ signaling. A129 and AG129 mice were infected i.v. with 5x10 8 GE of E124/128-IC, and spleens were harvested at 12, 24, 48, and 72 hours after infection. Frozen sections of the spleen were stained for 18
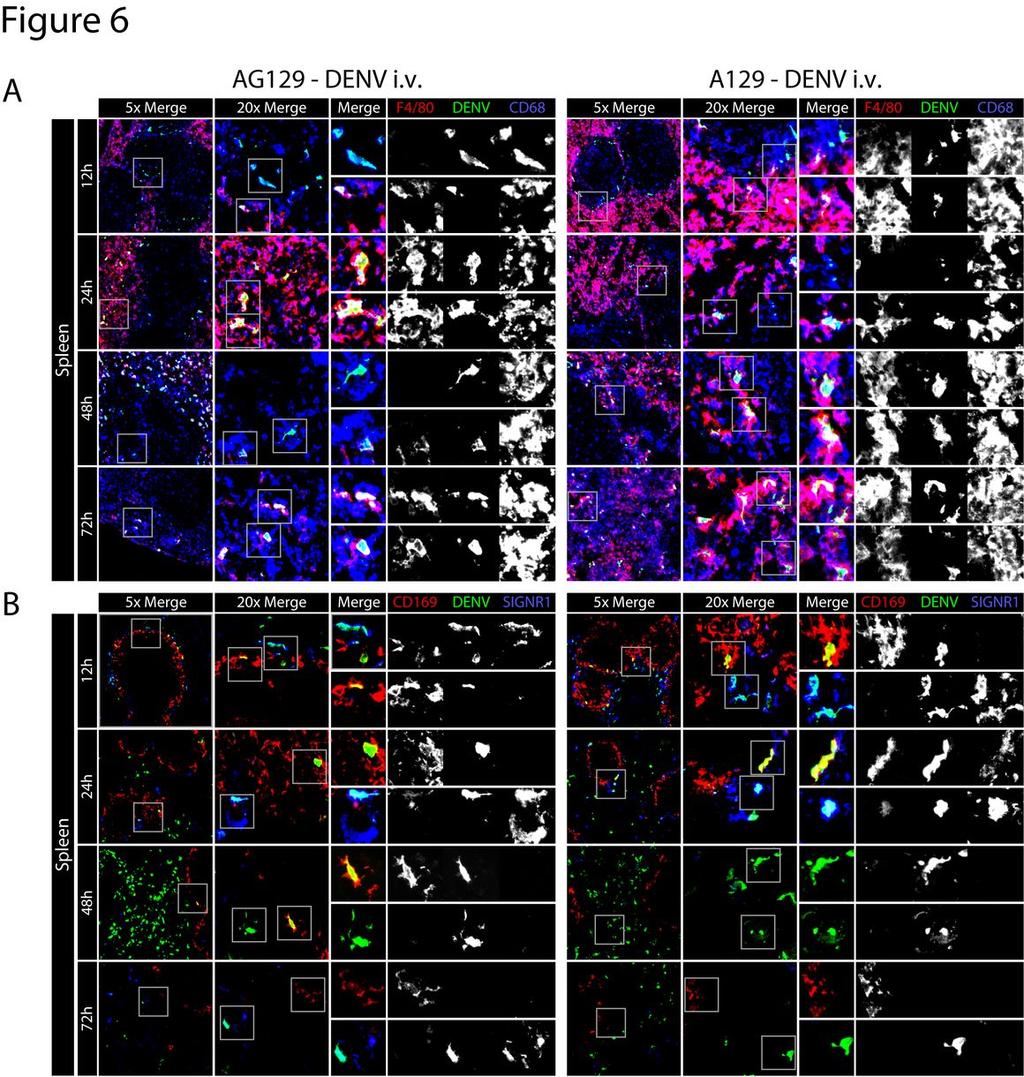
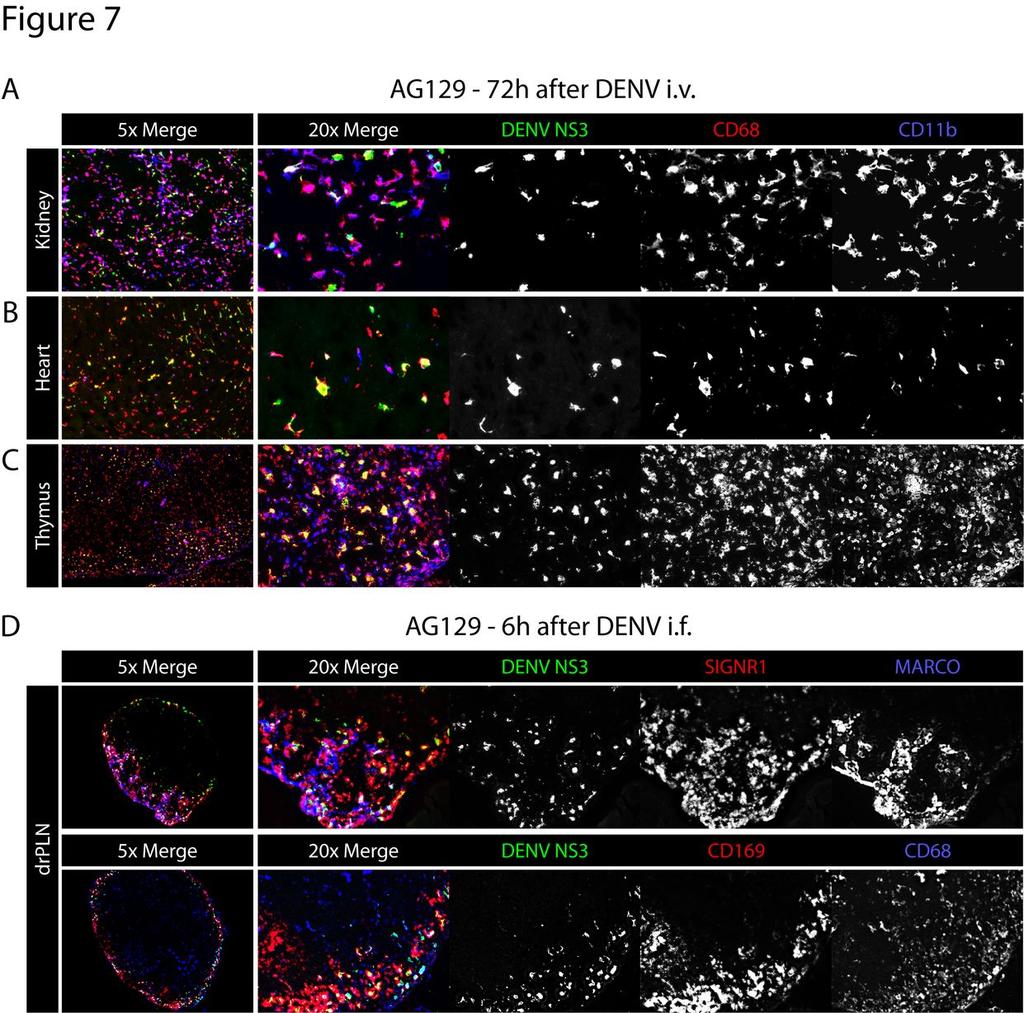
19 DENV NS3 together with either F4/80 and CD68 (Figure 6A), or CD169 and SIGN-R1 (Figure 6B). At 12 and 24 hours post-inoculation, A129 and AG129 mice showed an 386 approximately equivalent infection occurring predominantly in CD169 + MMMs and SIGN-R1 + MZMs. At 48 hours, however, infection in F4/80 + RPMs was strikingly reduced in A129 mice compared to AG129 (Figure 6A). Thus, in this experimental setup, IFN-γ receptor signaling does not affect initial DENV replication in MMMs or MZMs, but is able to reduce replication in RPMs. Additionally, DENV cellular tropism in this model is unaffected by IFN-γ receptor signaling. DENV systemically infects CD68 + macrophages late in infection We have previously reported that in AG129 mice, DENV replicates in CD68 + lamina propria macrophages of the small intestine at late time points after infection (38). To determine whether macrophages are the cells infected at late time points in other tissues, AG129 mice were inoculated i.v. with 1.5x10 11 GE of E124/128-IC, and 72 hours later the kidney, heart and thymus were harvested and stained for CD11b and CD68. In the kidney, heart, and thymus, most of the DENV NS3 signal was present in cells expressing CD68 and, additionally but less often, CD11b (Figure 7A). These results confirm that macrophages in many non-lymphoid tissues are late targets of DENV infection. 19
20 DENV replicates in macrophages of draining lymph nodes following intrafootpad inoculation Initial screening studies for DENV replication following intrafootpad inoculation revealed early replication in the draining PLNs of AG129 mice. To identify the cells in the lymph node targeted by DENV, AG129 mice were inoculated i.f. with 1.5x10 11 GE of E124/128-IC, and PLNs were harvested 6 hours after infection. Frozen sections of the PLNs were stained for DENV NS3 together with either MARCO and SIGN-R1 (Figure 7B, top row) or CD169 and CD68 (Figure 7B, bottom row). As in the spleen, cells expressing these markers were overlapping, and cells expressing each or combinations were found to permit DENV replication. These results show that initial DENV infection occurs in macrophages of draining PLNs within 6 hours following intrafootpad inoculation. Downloaded from on October 2, 2018 by guest 20
21 DISCUSSION In this study, we sought to characterize the tissues and cells targeted by DENV in vivo and the sequence in which they are infected. Following i.v. inoculation, DENV initially traffics to the spleen where it infects macrophages of the marginal zone, including MZMs and MMMs. Following i.f. inoculation, DENV initially targets macrophages of the draining popliteal LN before entering the blood and subsequently infecting the splenic marginal zone. Although blood-borne DENV continues to traffic to the marginal zone throughout the course of infection, replication in the marginal zone decreases and bloodborne DENV later disseminates into the red pulp, where it infects RPMs. The initial wave of replication in the spleen is followed by replication in the bone marrow and then lymph nodes and Peyer s patches. Finally, late during infection, when mice are becoming overtly ill, replication decreases in the lymphoid tissues and increases in macrophages of non-lymphoid tissues, most notably in the kidneys, heart, thymus, and GI tract. In humans, a broad array of tissues and cell types have been reported to harbor DENV: perivascular mononuclear cells of the skin; hepatocytes, endothelial cells, and Kupffer cells of the liver; cortical neurons, astrocytes, microglia, Purkinje s cells, and choroid plexus epithelial cells of the brain; immunoblasts, mononuclear cells, and macrophages of the kidney; immunoblasts, lymphocytes, plasma cells, and macrophages of the spleen, lymph nodes, and kidneys; endothelial cells of the spleen; alveolar macrophages of the lung; mucosal lymphocytes and lamina propria plasma cells of the stomach; and plasma and mononuclear cells of the small and large intestine (2, 5, 6, 8, 13, 15, 16, 25, 29). Importantly, because these studies detected components of mature viral particles and 21
22 identified cells based on cellular morphology alone or with non-specific markers in serial sections, it is unclear whether these studies have accurately identified sites of viral replication. Despite these caveats, the published studies with human tissues have set a framework for understanding DENV behavior in vivo. Due to logistical complications of studies with human tissues, small animal studies can provide critical information to elucidate viral behavior in vivo. For example, the present study investigated the kinetics of DENV dissemination by analyzing several time points and tissues after infection at a consistent viral dose, which is nearly impossible in human studies. However, the use of non-natural mammalian hosts, particularly mice lacking IFN receptors, indicates potential caveats for interpretation of our results. Both type I and type II IFN have been reported to restrict the cellular and tissue tropism of flaviviruses in mice (30, 31), which may result in findings that do not represent what occurs in natural human infections. However, DENV, like other flaviviruses, possesses mechanisms to overcome or circumvent IFN responses in humans and non-human primates (7, 17, 18, 22, 24), and this ability appears to be species-specific for a number of mosquito-borne viruses (1, 4, 24). As we are not currently able to validate our findings with human samples, it remains unclear whether other differences between mice and humans could be confounding our results. For example, there may be cell populations targeted by DENV in humans that are either not present or not infected in mice. Thus, although small animal studies are invaluable for deducing the nature of virus replication and virus-host interactions in vivo, it is crucial that findings are evaluated in focused human studies, even if the same rigor is not possible. 22
23 Identification of the precise cellular targets of DENV in vivo has been hindered by the heterogeneity of mononuclear phagocytes present in different tissues and even within a given tissue. Macrophages of the spleen in particular are highly heterogeneous and, therefore, difficult to distinguish based on cell markers (23). Certain markers, such as CD11b, are expressed at variable levels on multiple cell populations. Moreover, the populations of cells present in a tissue, as well as cell marker expression, can change under inflammatory conditions such as viral infections. This may help explain the inconsistent reports in the literature of individual markers on specific cell populations. For example, RPMs have been variably reported to lack or express CD11b (11, 19, 23, 37). FIHC is thus particularly useful in identifying cell populations, including monocytes/neutrophils (CD11b high ), MZMs (SIGN-R1 + ), MMMs (CD169 + ), and RPMs (F4/80 + ), because it is based on anatomic location, cellular morphology, and expression of cell population-specific markers. Tissue and cellular tropism of DENV has been investigated previously in AG129 mice, both by our group and others (2, 3, 9, 20, 38). It has been reported that in the spleen, DENV infection occurs in CD11c + DCs and in CD11b high-expressing, F4/80 + macrophages (2, 20). In contrast to these publications, we did not find evidence that CD11c + DCs in the spleen are major targets of DENV, nor did we find evidence of notable CD11b expression on DENV-infected F4/80 + red pulp macrophages. Instead, our findings demonstrated that MMMs, MZMs, and RPMs, but not cells expressing high levels of CD11b or CD11c, were highly permissive to DENV infection. In agreement with the present study, Balsitis et al. identified the red pulp and marginal zone as the 23
24 485 dominant compartments for DENV infection in the spleen (2) and Fink et al. reported 486 infection of CD169 + subcapsular sinus macrophages (SCSMs) of the draining LN 487 following subcutaneous administration of DENV (9). In the present study, SCSMs and 488 the equivalent CD169 + cells in the marginal zone of the spleen, respectively, were infected following i.f. and i.v. routes of administration, and all of the lymph nodes and Peyer s patches analyzed 48 hours after infection contained high levels of DENV NS3. These particular macrophage populations of the splenic marginal zone and subcapsular sinus of lymph nodes have been shown to be important for clearance of high molecular mass antigens including viral particles from blood and lymph (21, 26, 33). These cells are believed to permit low levels of viral replication in order to amplify antigen and facilitate generation of high affinity antibodies (14). Additionally, many viruses, including DENV, bind to DC-SIGN (36), which is the human homolog of murine SIGN- R1. This molecule is expressed at high levels on macrophages of the marginal zone and subcapsular sinus and it acts as a pattern recognition receptor for sampling antigens in the surrounding fluid (10). These characteristics all support results that these macrophage populations, which possess molecules thought to be responsible for permitting DENV infection, are initial interaction partners and important initial targets of blood- and lymphborne DENV. To our knowledge, this is the first use of fluorescence-tagged viral particles for tracking the temporal and spatial patterns of DENV trafficking in vivo. The far-red fluorescent, lipophilic dye DiD has previously been used for single DENV particle tracking to 24
25 investigate cell entry in vitro (35). Here, DiD-labeled DENV particles were utilized in vivo to visualize the fate of the injected viral particles. In contrast to tissue section staining for DENV NS3, DiD-labeled DENV particles can be used to probe the outcome of blood-borne DENV in order to visualize trafficking at any time during infection. Our results show that high levels of DENV traffic to cells of the liver and the spleen, initially in the marginal zone and later also in the red pulp. The amount of DiD accumulating in the spleen decreases over time, while it increases in the liver, kidneys and thymus. Although kinetically not identical, this trend mirrors levels of viral replication within these organs over time, suggesting that extravasation of viral particles may dictate the sequence of cells and tissues targeted by DENV in vivo. Our results contribute to the understanding of DENV tropism in vivo over the course of infection. We show that DENV inoculated via a peripheral route initially targets draining lymph nodes. Once DENV enters the blood, it first targets the spleen and bone marrow and later spreads to the lymph nodes and Peyer s patches, and finally to non-lymphoid tissues throughout the mouse. These findings augment our published studies and show that DENV largely infects macrophages in a number of tissues of AG129 mice: the kidneys, heart, thymus, and gastrointestinal tract, marginal zone and red pulp of the spleen, subcapsular sinus of the lymph nodes, as well as sinusoidal endothelial cells of the liver (9, 38). By elucidating the pattern of DENV replication in mice, our study provides an overall framework for DENV behavior in vivo to guide subsequent investigations of DENV tropism in naturally infected humans. 25
26 FIGURE LEGENDS Figure 1 Dengue virus first infects the spleen, followed by bone marrow, and then lymph nodes, and Peyer s patches after intravenous inoculation. Representative immunohistochemical staining of tissue sections from naïve AG129 mice, and at 6, 12, 24, 48, and 72 hours after infection (1.5x10 11 GE E124/128-IC i.v. on day 0). Spleens, cutaneous lymph nodes (CLN: axillary, brachial, and inguinal lymph nodes), mesenteric lymph nodes (MLN), Peyer s patches (PP) and bone marrow (BM) were sectioned and stained for DENV NS3 (white when alone, green when shown with prm/m) and DENV prm/m (antibody clone 2H2, shown in red). Results are representative of 3 mice. WP labels the white pulp. LN labels the center of the lymph nodes. PP labels the center of the Peyer s patches. 26
27 Figure 2 Dengue virus infects systemic non-lymphoid tissues late during infection. Representative immunohistochemical staining of tissue sections from AG129 mice. A) Tissues were harvested from naïve mice and mice 48 and 72 hours after infection (1.5x10 11 GE E124/128-IC i.v. on day 0) and stained for DENV NS3 (white) in the kidney, heart, thymus, adrenal gland, liver, lung, forestomach, stomach, duodenum, jejunum, ileum, cecum, colon, and rectum. Results are representative of 3 mice. B) Tissues were harvested from mice 6, 24, and 72 hours after infection (1.5x10 11 GE E124/128-IC i.f. on day 0) and stained for DENV NS3 (white) in the spleen, draining popliteal lymph node (drpln), contralateral popliteal lymph node (clpln), cutaneous lymph nodes (CLN: axillary, brachial, and inguinal lymph nodes), and kidney. Results are representative of 3 mice. Downloaded from on October 2, 2018 by guest 27
28 Figure 3 A schematic depicting the kinetics of DENV tissue tropism over the course of infection in mice Model of DENV infection. DENV first infects the spleen, followed by the bone marrow, and then the lymph nodes and Peyer s patches (LN, PP). Infection decreases in these tissues while it increases systemically in non-lymphoid tissues at which point mice become moribund. Downloaded from on October 2, 2018 by guest 28
29 Figure 4 Blood-borne DENV particles traffic mainly to the spleen and liver (A) Representative immunohistochemical staining of spleen, liver, thymus, and kidney from naïve AG129 mice, and 6, 12, 18, 24, 36, 48, and 72 hours after infection (1.5x10 11 GE E124/128-IC i.v. on day 0 and 5x10 10 GE DiD-labeled E124/128-IC i.v. 2 hours before harvest). Sections were stained for DENV NS3 (white, top row of each panel) and DiD in the tissue was imaged (white, middle row of each panel). Merged images are shown on the bottom row of each panel (NS3 is shown as green and DiD is shown as red). B) Representative immunohistochemical staining of spleen from AG129 (top) or WT129 (bottom) mice 6 hours after infection (5x10 10 GE DiD-labeled E124/128-IC i.v.). Sections were stained for CD169, SIGN-R1, and MARCO (white, left) and DiD in the tissue was imaged (white, middle). The merged image is shown on the right (DiD is shown as green and CD169, SIGN-R1, and MARCO are shown as red). C) Summary of results from panels (A). DiD intensity was quantified from 6-24 images per mouse with 5 mice per time point. Symbols and error bars represent means ± SEM. P- values from two-tailed unpaired t-test, CI 95%: * P 0.05, ** P 0.01, *** P
30 Figure 5 In the spleen, DENV first infects macrophages of the marginal zone, and later infects macrophages of the red pulp A) Representative immunohistochemical staining of spleen sections from AG129 mice 12 hours after infection (5x10 8 GE E124/128-IC i.v. on day 0). Tissue sections were stained for DENV NS3 (green) and CD11b and CD11c (red and blue, respectively, top row), CD68 and F4/80 (red and blue, respectively, middle row), and CD169 and SIGN-R1 (red and blue, respectively, bottom row). Low magnification (5x) images are shown on the left with DENV NS3 alone, while each individual channel and the merged image are shown on the right. Grey boxes on the DENV NS3 panels depict areas of enlargement shown on the right. Results are representative of 3 mice. B) Representative immunohistochemical staining of spleen sections from AG129 mice 48 hours after infection (5x10 8 GE E124/128-IC i.v. on day 0). Sections are stained as in A. Results are representative of 3 mice. 30
31 Figure 6 Interferon-γ-receptor signaling limits DENV replication in splenic red pulp macrophages A) Representative immunohistochemical staining of spleen sections from AG129 or A129 mice 12, 24, 48, and 72 after infection (5x10 8 GE E124/128-IC i.v. on day 0). Tissue sections were stained for F4/80 (red), DENV NS3 (green), and CD68 (blue). Low magnification (5x) merged images are shown on the left as merged images only, medium magnification (20x) images are shown in the middle as merged images only, and high magnification (33x) images are shown on the right as the merged image followed by each individual channel (F4/80, DENV NS3, and CD68). Grey boxes on the left depict the area of enlargement shown on the immediate right. Two enlarged areas from each middle magnification image are shown. Results are representative of 3 mice. B) Representative immunohistochemical staining of spleen sections from AG129 or A129 mice 12, 24, 48, and 72 after infection (5x10 8 GE E124/128-IC i.v. on day 0), with each time point represented by a row. Images are arranged as in A, except tissue sections were stained for CD169 (red), DENV NS3 (green), and SIGN-R1 (blue). Results are representative of 3 mice. 31
32 Figure 7 DENV infects macrophages of the kidney, heart, thymus and lymph nodes A) Representative immunohistochemical staining of kidney, heart, and thymus sections from AG129 mice 72 hours after infection (1.5x10 11 GE E124/128-IC i.v. on day 0). Tissue sections were stained for DENV NS3 (green), CD68 (red), and CD11b (blue). 5x magnification of all channels is shown on left and 20x magnification images on the right with each channel shown separately in white. Results are representative of 3 mice. B) Representative immunohistochemical staining of draining popliteal lymph node sections from AG129 mice 6 hours after infection (1.5x10 11 GE E124/128-IC i.f.). Tissue sections were stained for DENV NS3 (green), SIGN-R1 (red), and MARCO (blue) in the upper panel and DENV NS3 (green), CD169 (red), and CD68 (blue) in the lower panel. 5x magnification of all channels is shown on left and 20x magnification images on the right with each channel shown separately in white. Results are representative of 3 mice. 32
33 ACKNOWLEDGEMENTS We thank Drs. Chris Benedict, Raphaël M. Zellweger, and Stuart T. Perry at LIAI for experimental advice. We thank Dr. Olga Turovskaya, Sarala Joshi, Steven Lada, and Dr. Satoshi Fukuyama for technical help. This work was supported by NIH grants U01 AI to S.S. and U54 AI from Southeast Regional Center of Excellence for Emerging Infections and Biodefense to P.F. Sparling. The authors declare they have no competing financial interests. Downloaded from on October 2, 2018 by guest 33
34 REFERENCES 1. Ashour, J., J. Morrison, M. Laurent-Rolle, A. Belicha-Villanueva, C. R. Plumlee, D. Bernal-Rubio, K. L. Williams, E. Harris, A. Fernandez-Sesma, C. Schindler, and A. Garcia-Sastre Mouse STAT2 restricts early dengue virus replication. Cell host & microbe 8: Balsitis, S. J., J. Coloma, G. Castro, A. Alava, D. Flores, J. H. McKerrow, P. R. Beatty, and E. Harris Tropism of dengue virus in mice and humans defined by viral nonstructural protein 3-specific immunostaining. Am J Trop Med Hyg 80: Balsitis, S. J., K. L. Williams, R. Lachica, D. Flores, J. L. Kyle, E. Mehlhop, S. Johnson, M. S. Diamond, P. R. Beatty, and E. Harris Lethal antibody enhancement of dengue disease in mice is prevented by Fc modification. PLoS Pathog 6:e Bente, D. A., J. B. Alimonti, W. J. Shieh, G. Camus, U. Stroher, S. Zaki, and S. M. Jones Pathogenesis and immune response of Crimean-Congo hemorrhagic fever virus in a STAT-1 knockout mouse model. Journal of virology 84: Bhoopat, L., N. Bhamarapravati, C. Attasiri, S. Yoksarn, B. Chaiwun, S. Khunamornpong, and V. Sirisanthana Immunohistochemical characterization of a new monoclonal antibody reactive with dengue virusinfected cells in frozen tissue using immunoperoxidase technique. Asian Pac J Allergy Immunol 14: Boonpucknavig, S., V. Boonpucknavig, N. Bhamarapravati, and S. Nimmannitya Immunofluorescence study of skin rash in patients with dengue hemorrhagic fever. Arch Pathol Lab Med 103: Chung, K. M., M. K. Liszewski, G. Nybakken, A. E. Davis, R. R. Townsend, D. H. Fremont, J. P. Atkinson, and M. S. Diamond West Nile virus nonstructural protein NS1 inhibits complement activation by binding the regulatory protein factor H. Proc Natl Acad Sci U S A 103: Couvelard, A., P. Marianneau, C. Bedel, M. T. Drouet, F. Vachon, D. Henin, and V. Deubel Report of a fatal case of dengue infection with hepatitis: demonstration of dengue antigens in hepatocytes and liver apoptosis. Hum Pathol 30: Fink, K., C. Ng, C. Nkenfou, S. G. Vasudevan, N. van Rooijen, and W. Schul Depletion of macrophages in mice results in higher dengue virus titers and highlights the role of macrophages for virus control. Eur J Immunol 39: Geijtenbeek, T. B., P. C. Groot, M. A. Nolte, S. J. van Vliet, S. T. Gangaram- Panday, G. C. van Duijnhoven, G. Kraal, A. J. van Oosterhout, and Y. van Kooyk Marginal zone macrophages express a murine homologue of DC- SIGN that captures blood-borne antigens in vivo. Blood 100: Gorgani, N. N., Y. Ma, and H. F. Clark Gene signatures reflect the marked heterogeneity of tissue-resident macrophages. Immunol Cell Biol 86:
35 Gubler, D. J., and M. Meltzer Impact of dengue/dengue hemorrhagic fever on the developing world. Adv Virus Res 53: Hall, W. C., T. P. Crowell, D. M. Watts, V. L. Barros, H. Kruger, F. Pinheiro, and C. J. Peters Demonstration of yellow fever and dengue antigens in formalin-fixed paraffin-embedded human liver by immunohistochemical analysis. Am J Trop Med Hyg 45: Honke, N., N. Shaabani, G. Cadeddu, U. R. Sorg, D. E. Zhang, M. Trilling, K. Klingel, M. Sauter, R. Kandolf, N. Gailus, N. van Rooijen, C. Burkart, S. E. Baldus, M. Grusdat, M. Lohning, H. Hengel, K. Pfeffer, M. Tanaka, D. Haussinger, M. Recher, P. A. Lang, and K. S. Lang Enforced viral replication activates adaptive immunity and is essential for the control of a cytopathic virus. Nat Immunol 13: Huerre, M. R., N. T. Lan, P. Marianneau, N. B. Hue, H. Khun, N. T. Hung, N. T. Khen, M. T. Drouet, V. T. Huong, D. Q. Ha, Y. Buisson, and V. Deubel Liver histopathology and biological correlates in five cases of fatal dengue fever in Vietnamese children. Virchows Arch 438: Jessie, K., M. Y. Fong, S. Devi, S. K. Lam, and K. T. Wong Localization of dengue virus in naturally infected human tissues, by immunohistochemistry and in situ hybridization. J Infect Dis 189: Jones, M., A. Davidson, L. Hibbert, P. Gruenwald, J. Schlaak, S. Ball, G. R. Foster, and M. Jacobs Dengue virus inhibits alpha interferon signaling by reducing STAT2 expression. J Virol 79: Keller, B. C., B. L. Fredericksen, M. A. Samuel, R. E. Mock, P. W. Mason, M. S. Diamond, and M. Gale, Jr Resistance to alpha/beta interferon is a determinant of West Nile virus replication fitness and virulence. Journal of virology 80: Kirby, A. C., L. Beattie, A. Maroof, N. van Rooijen, and P. M. Kaye SIGNR1-negative red pulp macrophages protect against acute streptococcal sepsis after Leishmania donovani-induced loss of marginal zone macrophages. Am J Pathol 175: Kyle, J. L., P. R. Beatty, and E. Harris Dengue virus infects macrophages and dendritic cells in a mouse model of infection. J Infect Dis 195: Lang, P. A., M. Recher, N. Honke, S. Scheu, S. Borkens, N. Gailus, C. Krings, A. Meryk, A. Kulawik, L. Cervantes-Barragan, N. Van Rooijen, U. Kalinke, B. Ludewig, H. Hengartner, N. Harris, D. Haussinger, P. S. Ohashi, R. M. Zinkernagel, and K. S. Lang Tissue macrophages suppress viral replication and prevent severe immunopathology in an interferon-i-dependent manner in mice. Hepatology 52: Lin, R. J., C. L. Liao, E. Lin, and Y. L. Lin Blocking of the alpha interferon-induced Jak-Stat signaling pathway by Japanese encephalitis virus infection. J Virol 78: Lloyd, C. M., A. R. Phillips, G. J. Cooper, and P. R. Dunbar Threecolour fluorescence immunohistochemistry reveals the diversity of cells staining for macrophage markers in murine spleen and liver. J Immunol Methods 334:
36 Meier, K. C., C. L. Gardner, M. V. Khoretonenko, W. B. Klimstra, and K. D. Ryman A mouse model for studying viscerotropic disease caused by yellow fever virus infection. PLoS pathogens 5:e Miagostovich, M. P., R. G. Ramos, A. F. Nicol, R. M. Nogueira, T. Cuzzi- Maya, A. V. Oliveira, R. S. Marchevsky, R. P. Mesquita, and H. G. Schatzmayr Retrospective study on dengue fatal cases. Clin Neuropathol 16: Oehen, S., B. Odermatt, U. Karrer, H. Hengartner, R. Zinkernagel, and C. Lopez-Macias Marginal zone macrophages and immune responses against viruses. J Immunol 169: Perry, S. T., T. R. Prestwood, S. M. Lada, C. A. Benedict, and S. Shresta Cardif-mediated signaling controls the initial innate response to dengue virus in vivo. J Virol 83: Prestwood, T. R., D. M. Prigozhin, K. L. Sharar, R. M. Zellweger, and S. Shresta A mouse-passaged dengue virus strain with reduced affinity for heparan sulfate causes severe disease in mice by establishing increased systemic viral loads. J Virol 82: Ramos, C., G. Sanchez, R. H. Pando, J. Baquera, D. Hernandez, J. Mota, J. Ramos, A. Flores, and E. Llausas Dengue virus in the brain of a fatal case of hemorrhagic dengue fever. J Neurovirol 4: Ryman, K. D., W. B. Klimstra, K. B. Nguyen, C. A. Biron, and R. E. Johnston Alpha/beta interferon protects adult mice from fatal Sindbis virus infection and is an important determinant of cell and tissue tropism. J Virol 74: Samuel, M. A., and M. S. Diamond Alpha/beta interferon protects against lethal West Nile virus infection by restricting cellular tropism and enhancing neuronal survival. J Virol 79: Shresta, S., K. L. Sharar, D. M. Prigozhin, P. R. Beatty, and E. Harris Murine model for dengue virus-induced lethal disease with increased vascular permeability. J Virol 80: Steiniger, B., and P. Barth Microanatomy and function of the spleen. Adv Anat Embryol Cell Biol 151:III-IX, Styer, L. M., K. A. Kent, R. G. Albright, C. J. Bennett, L. D. Kramer, and K. A. Bernard Mosquitoes inoculate high doses of West Nile virus as they probe and feed on live hosts. PLoS pathogens 3: van der Schaar, H. M., M. J. Rust, C. Chen, H. van der Ende-Metselaar, J. Wilschut, X. Zhuang, and J. M. Smit Dissecting the cell entry pathway of dengue virus by single-particle tracking in living cells. PLoS Pathog 4:e van Kooyk, Y., and T. B. Geijtenbeek DC-SIGN: escape mechanism for pathogens. Nat Rev Immunol 3: Witmer-Pack, M. D., D. A. Hughes, G. Schuler, L. Lawson, A. McWilliam, K. Inaba, R. M. Steinman, and S. Gordon Identification of macrophages and dendritic cells in the osteopetrotic (op/op) mouse. J Cell Sci 104 ( Pt 4):
37 Zellweger, R. M., T. R. Prestwood, and S. Shresta Enhanced infection of liver sinusoidal endothelial cells in a mouse model of antibody-induced severe dengue disease. Cell Host Microbe 7:
38
39
40
41
42
43
44
SUPPLEMENTARY INFORMATION
 doi:1.138/nature1554 a TNF-α + in CD4 + cells [%] 1 GF SPF 6 b IL-1 + in CD4 + cells [%] 5 4 3 2 1 Supplementary Figure 1. Effect of microbiota on cytokine profiles of T cells in GALT. Frequencies of TNF-α
doi:1.138/nature1554 a TNF-α + in CD4 + cells [%] 1 GF SPF 6 b IL-1 + in CD4 + cells [%] 5 4 3 2 1 Supplementary Figure 1. Effect of microbiota on cytokine profiles of T cells in GALT. Frequencies of TNF-α
Supporting Information
 Supporting Information Idoyaga et al. 10.1073/pnas.0812247106 SSC a) Single cell suspension 99 Aqua b) Live cells 96 -W c) Singlets 92 -A CD19+ER119 d) CD19 ER119 cells 97 CD3 e) CD3 cells 27 f) DX5 cells
Supporting Information Idoyaga et al. 10.1073/pnas.0812247106 SSC a) Single cell suspension 99 Aqua b) Live cells 96 -W c) Singlets 92 -A CD19+ER119 d) CD19 ER119 cells 97 CD3 e) CD3 cells 27 f) DX5 cells
Hepatitis virus immunity. Mar 9, 2005 Rehermann and Nascimbeni review Crispe review
 Hepatitis virus immunity Mar 9, 2005 Rehermann and Nascimbeni review Crispe review HBV & HCV infection outcomes Both viruses cause immune-mediated active and chronic hepatitis HBV Vertical transmission
Hepatitis virus immunity Mar 9, 2005 Rehermann and Nascimbeni review Crispe review HBV & HCV infection outcomes Both viruses cause immune-mediated active and chronic hepatitis HBV Vertical transmission
Supplementary Figures
 Inhibition of Pulmonary Anti Bacterial Defense by IFN γ During Recovery from Influenza Infection By Keer Sun and Dennis W. Metzger Supplementary Figures d a Ly6G Percentage survival f 1 75 5 1 25 1 5 1
Inhibition of Pulmonary Anti Bacterial Defense by IFN γ During Recovery from Influenza Infection By Keer Sun and Dennis W. Metzger Supplementary Figures d a Ly6G Percentage survival f 1 75 5 1 25 1 5 1
Supplementary Figure 1. Characterization of basophils after reconstitution of SCID mice
 Supplementary figure legends Supplementary Figure 1. Characterization of after reconstitution of SCID mice with CD4 + CD62L + T cells. (A-C) SCID mice (n = 6 / group) were reconstituted with 2 x 1 6 CD4
Supplementary figure legends Supplementary Figure 1. Characterization of after reconstitution of SCID mice with CD4 + CD62L + T cells. (A-C) SCID mice (n = 6 / group) were reconstituted with 2 x 1 6 CD4
PBS Class #2 Introduction to the Immune System part II Suggested reading: Abbas, pgs , 27-30
 PBS 803 - Class #2 Introduction to the Immune System part II Suggested reading: Abbas, pgs. 15-25, 27-30 Learning Objectives Compare and contrast the maturation of B and T lymphocytes Compare and contrast
PBS 803 - Class #2 Introduction to the Immune System part II Suggested reading: Abbas, pgs. 15-25, 27-30 Learning Objectives Compare and contrast the maturation of B and T lymphocytes Compare and contrast
Cell isolation. Spleen and lymph nodes (axillary, inguinal) were removed from mice
 were removed from mice") Supplementary Methods: Cell isolation. Spleen and lymph nodes (axillary, inguinal) were removed from mice and gently meshed in DMEM containing 10% FBS to prepare for single cell suspensions. CD4 + CD25
Supplementary Methods: Cell isolation. Spleen and lymph nodes (axillary, inguinal) were removed from mice and gently meshed in DMEM containing 10% FBS to prepare for single cell suspensions. CD4 + CD25
IL-34 is a tissue-restricted ligand of CSF1R required for the development of Langerhans cells and microglia
 Supplementary Figures IL-34 is a tissue-restricted ligand of CSF1R required for the development of Langerhans cells and microglia Yaming Wang, Kristy J. Szretter, William Vermi, Susan Gilfillan, Cristina
Supplementary Figures IL-34 is a tissue-restricted ligand of CSF1R required for the development of Langerhans cells and microglia Yaming Wang, Kristy J. Szretter, William Vermi, Susan Gilfillan, Cristina
Supplementary Figure 1. Generation of knockin mice expressing L-selectinN138G. (a) Schematics of the Sellg allele (top), the targeting vector, the
 Schematics of the Sellg allele (top), the targeting vector, the") Supplementary Figure 1. Generation of knockin mice expressing L-selectinN138G. (a) Schematics of the Sellg allele (top), the targeting vector, the targeted allele in ES cells, and the mutant allele in
Supplementary Figure 1. Generation of knockin mice expressing L-selectinN138G. (a) Schematics of the Sellg allele (top), the targeting vector, the targeted allele in ES cells, and the mutant allele in
Chapter 24 The Immune System
 Chapter 24 The Immune System The Immune System Layered defense system The skin and chemical barriers The innate and adaptive immune systems Immunity The body s ability to recognize and destroy specific
Chapter 24 The Immune System The Immune System Layered defense system The skin and chemical barriers The innate and adaptive immune systems Immunity The body s ability to recognize and destroy specific
Figure S1. Generation of inducible PTEN deficient mice and the BMMCs (A) B6.129 Pten loxp/loxp mice were mated with B6.
 B6.129 Pten loxp/loxp mice were mated with B6.") Figure S1. Generation of inducible PTEN deficient mice and the BMMCs (A) B6.129 Pten loxp/loxp mice were mated with B6.129-Gt(ROSA)26Sor tm1(cre/ert2)tyj /J mice. To induce deletion of the Pten locus,
Figure S1. Generation of inducible PTEN deficient mice and the BMMCs (A) B6.129 Pten loxp/loxp mice were mated with B6.129-Gt(ROSA)26Sor tm1(cre/ert2)tyj /J mice. To induce deletion of the Pten locus,
Blocking antibodies and peptides. Rat anti-mouse PD-1 (29F.1A12, rat IgG2a, k), PD-
, PD-") Supplementary Methods Blocking antibodies and peptides. Rat anti-mouse PD-1 (29F.1A12, rat IgG2a, k), PD- L1 (10F.9G2, rat IgG2b, k), and PD-L2 (3.2, mouse IgG1) have been described (24). Anti-CTLA-4 (clone
Supplementary Methods Blocking antibodies and peptides. Rat anti-mouse PD-1 (29F.1A12, rat IgG2a, k), PD- L1 (10F.9G2, rat IgG2b, k), and PD-L2 (3.2, mouse IgG1) have been described (24). Anti-CTLA-4 (clone
(a) Significant biological processes (upper panel) and disease biomarkers (lower panel)
 Significant biological processes (upper panel) and disease biomarkers (lower panel)") Supplementary Figure 1. Functional enrichment analyses of secretomic proteins. (a) Significant biological processes (upper panel) and disease biomarkers (lower panel) 2 involved by hrab37-mediated secretory
Supplementary Figure 1. Functional enrichment analyses of secretomic proteins. (a) Significant biological processes (upper panel) and disease biomarkers (lower panel) 2 involved by hrab37-mediated secretory
SUPPORTING INFORMATIONS
 SUPPORTING INFORMATIONS Mice MT/ret RetCD3ε KO α-cd25 treated MT/ret Age 1 month 3 mnths 6 months 1 month 3 months 6 months 1 month 3 months 6 months 2/87 Survival 87/87 incidence of 17/87 1 ary tumor
SUPPORTING INFORMATIONS Mice MT/ret RetCD3ε KO α-cd25 treated MT/ret Age 1 month 3 mnths 6 months 1 month 3 months 6 months 1 month 3 months 6 months 2/87 Survival 87/87 incidence of 17/87 1 ary tumor
MACROPHAGE "MONOCYTES" SURFACE RECEPTORS
 LECTURE: 13 Title: MACROPHAGE "MONOCYTES" SURFACE RECEPTORS LEARNING OBJECTIVES: The student should be able to: Describe the blood monocytes (size, and shape of nucleus). Enumerate some of the monocytes
LECTURE: 13 Title: MACROPHAGE "MONOCYTES" SURFACE RECEPTORS LEARNING OBJECTIVES: The student should be able to: Describe the blood monocytes (size, and shape of nucleus). Enumerate some of the monocytes
NS1 Protein Expression in the JaOArS982 Strain of Japanese Encephalitis Virus Does Not Enhance Virulence in Mice
 Tropical Medicine and Health Vol. 43 No.4, 2015, 233 237 doi: 10.2149/tmh.2015-27 Copyright 2015 by The Japanese Society of Tropical Medicine 233 Short Communications NS1 Protein Expression in the JaOArS982
Tropical Medicine and Health Vol. 43 No.4, 2015, 233 237 doi: 10.2149/tmh.2015-27 Copyright 2015 by The Japanese Society of Tropical Medicine 233 Short Communications NS1 Protein Expression in the JaOArS982
Medical Virology Immunology. Dr. Sameer Naji, MB, BCh, PhD (UK) Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University
 Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University") Medical Virology Immunology Dr. Sameer Naji, MB, BCh, PhD (UK) Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University Human blood cells Phases of immune responses Microbe Naïve
Medical Virology Immunology Dr. Sameer Naji, MB, BCh, PhD (UK) Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University Human blood cells Phases of immune responses Microbe Naïve
FOR OPTIMAL GUT HEALTH KEMIN.COM/GUTHEALTH
 FOR OPTIMAL GUT HEALTH KEMIN.COM/GUTHEALTH ALETA A SOURCE OF 1,3-BETA GLUCANS Aleta is highly bioavailable, offering a concentration greater than 5% of 1,3-beta glucans. Aleta provides a consistent response
FOR OPTIMAL GUT HEALTH KEMIN.COM/GUTHEALTH ALETA A SOURCE OF 1,3-BETA GLUCANS Aleta is highly bioavailable, offering a concentration greater than 5% of 1,3-beta glucans. Aleta provides a consistent response
SUPPLEMENTARY INFORMATION
 Complete but curtailed T-cell response to very-low-affinity antigen Dietmar Zehn, Sarah Y. Lee & Michael J. Bevan Supp. Fig. 1: TCR chain usage among endogenous K b /Ova reactive T cells. C57BL/6 mice
Complete but curtailed T-cell response to very-low-affinity antigen Dietmar Zehn, Sarah Y. Lee & Michael J. Bevan Supp. Fig. 1: TCR chain usage among endogenous K b /Ova reactive T cells. C57BL/6 mice
Chapter10 Immune system
 Chapter10 Immune system Lyu Zhengmei Department of Histology and Embryology, Anhui Medical University Ⅰ.General Introduction Function ------ Defense The human body immune system has the ability to distinguish
Chapter10 Immune system Lyu Zhengmei Department of Histology and Embryology, Anhui Medical University Ⅰ.General Introduction Function ------ Defense The human body immune system has the ability to distinguish
VEGFR2-Mediated Vascular Dilation as a Mechanism of VEGF-Induced Anemia and Bone Marrow Cell Mobilization
 Cell Reports, Volume 9 Supplemental Information VEGFR2-Mediated Vascular Dilation as a Mechanism of VEGF-Induced Anemia and Bone Marrow Cell Mobilization Sharon Lim, Yin Zhang, Danfang Zhang, Fang Chen,
Cell Reports, Volume 9 Supplemental Information VEGFR2-Mediated Vascular Dilation as a Mechanism of VEGF-Induced Anemia and Bone Marrow Cell Mobilization Sharon Lim, Yin Zhang, Danfang Zhang, Fang Chen,
Supplementary Figure 1 Lymphocytes can be tracked for at least 4 weeks after
 Supplementary Figure 1 Lymphocytes can be tracked for at least 4 weeks after photoconversion by using H2B-Dendra2. 4-5 PPs of H2B-Dendra2 BM chimeras were photoconverted and analyzed 7 days (upper panel)
Supplementary Figure 1 Lymphocytes can be tracked for at least 4 weeks after photoconversion by using H2B-Dendra2. 4-5 PPs of H2B-Dendra2 BM chimeras were photoconverted and analyzed 7 days (upper panel)
Washington University School of Medicine, St Louis. MO Division of Infectious
 JVI Accepts, published online ahead of print on 27 June 2012 J. Virol. doi:10.1128/jvi.00673-12 Copyright 2012, American Society for Microbiology. All Rights Reserved. 1 2 CD8 + T cells use TRAIL to restrict
JVI Accepts, published online ahead of print on 27 June 2012 J. Virol. doi:10.1128/jvi.00673-12 Copyright 2012, American Society for Microbiology. All Rights Reserved. 1 2 CD8 + T cells use TRAIL to restrict
Islet viability assay and Glucose Stimulated Insulin Secretion assay RT-PCR and Western Blot
 Islet viability assay and Glucose Stimulated Insulin Secretion assay Islet cell viability was determined by colorimetric (3-(4,5-dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide assay using CellTiter
Islet viability assay and Glucose Stimulated Insulin Secretion assay Islet cell viability was determined by colorimetric (3-(4,5-dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide assay using CellTiter
Pre-clinical Development of a Dengue Vaccine. Jeremy Brett Sanofi Pasteur, Singapore
 Pre-clinical Development of a Dengue Vaccine Jeremy Brett Sanofi Pasteur, Singapore Dengue Vaccine B Guy 1 Talk flow Introduction: What are the challenges of dengue vaccine development? The Virus The host
Pre-clinical Development of a Dengue Vaccine Jeremy Brett Sanofi Pasteur, Singapore Dengue Vaccine B Guy 1 Talk flow Introduction: What are the challenges of dengue vaccine development? The Virus The host
Supporting Online Material for
 www.sciencemag.org/cgi/content/full/1175194/dc1 Supporting Online Material for A Vital Role for Interleukin-21 in the Control of a Chronic Viral Infection John S. Yi, Ming Du, Allan J. Zajac* *To whom
www.sciencemag.org/cgi/content/full/1175194/dc1 Supporting Online Material for A Vital Role for Interleukin-21 in the Control of a Chronic Viral Infection John S. Yi, Ming Du, Allan J. Zajac* *To whom
CD14 + S100A9 + Monocytic Myeloid-Derived Suppressor Cells and Their Clinical Relevance in Non-Small Cell Lung Cancer
 CD14 + S1A9 + Monocytic Myeloid-Derived Suppressor Cells and Their Clinical Relevance in Non-Small Cell Lung Cancer Po-Hao, Feng M.D., Kang-Yun, Lee, M.D. Ph.D., Ya-Ling Chang, Yao-Fei Chan, Lu- Wei, Kuo,Ting-Yu
CD14 + S1A9 + Monocytic Myeloid-Derived Suppressor Cells and Their Clinical Relevance in Non-Small Cell Lung Cancer Po-Hao, Feng M.D., Kang-Yun, Lee, M.D. Ph.D., Ya-Ling Chang, Yao-Fei Chan, Lu- Wei, Kuo,Ting-Yu
The peripheral (secondary) lymphoid tissues
 lymphoid tissues") The peripheral (secondary) lymphoid tissues The peripheral (secondary) lymphoid tissues : are the lymph nodes, spleen, Mucosal associated lymphoid tissue (MALT). All secondary lymphoid organs have one
The peripheral (secondary) lymphoid tissues The peripheral (secondary) lymphoid tissues : are the lymph nodes, spleen, Mucosal associated lymphoid tissue (MALT). All secondary lymphoid organs have one
Trim29 gene-targeting strategy. (a) Genotyping of wildtype mice (+/+), Trim29 heterozygous mice (+/ ) and homozygous mice ( / ).
 Genotyping of wildtype mice (+/+), Trim29 heterozygous mice (+/ ) and homozygous mice ( / ).") Supplementary Figure 1 Trim29 gene-targeting strategy. (a) Genotyping of wildtype mice (+/+), Trim29 heterozygous mice (+/ ) and homozygous mice ( / ). (b) Immunoblot analysis of TRIM29 in lung primary
Supplementary Figure 1 Trim29 gene-targeting strategy. (a) Genotyping of wildtype mice (+/+), Trim29 heterozygous mice (+/ ) and homozygous mice ( / ). (b) Immunoblot analysis of TRIM29 in lung primary
Application of μmacs Streptavidin MicroBeads for the analysis of HIV-1 directly from patient plasma
 Excerpt from MACS&more Vol 8 1/2004 Application of μmacs Streptavidin MicroBeads for the analysis of HIV-1 directly from patient plasma L. Davis Lupo and Salvatore T. Butera HIV and Retrovirology Branch,
Excerpt from MACS&more Vol 8 1/2004 Application of μmacs Streptavidin MicroBeads for the analysis of HIV-1 directly from patient plasma L. Davis Lupo and Salvatore T. Butera HIV and Retrovirology Branch,
Interferon γ regulates idiopathic pneumonia syndrome, a. Th17 + CD4 + T-cell-mediated GvH disease
 Interferon γ regulates idiopathic pneumonia syndrome, a Th17 + CD4 + T-cell-mediated GvH disease Nora Mauermann, Julia Burian, Christophe von Garnier, Stefan Dirnhofer, Davide Germano, Christine Schuett,
Interferon γ regulates idiopathic pneumonia syndrome, a Th17 + CD4 + T-cell-mediated GvH disease Nora Mauermann, Julia Burian, Christophe von Garnier, Stefan Dirnhofer, Davide Germano, Christine Schuett,
Chapter 13 Lymphatic and Immune Systems
 The Chapter 13 Lymphatic and Immune Systems 1 The Lymphatic Vessels Lymphoid Organs Three functions contribute to homeostasis 1. Return excess tissue fluid to the bloodstream 2. Help defend the body against
The Chapter 13 Lymphatic and Immune Systems 1 The Lymphatic Vessels Lymphoid Organs Three functions contribute to homeostasis 1. Return excess tissue fluid to the bloodstream 2. Help defend the body against
Supporting Information
 Supporting Information Valkenburg et al. 10.1073/pnas.1403684111 SI Materials and Methods ELISA and Microneutralization. Sera were treated with Receptor Destroying Enzyme II (RDE II, Accurate) before ELISA
Supporting Information Valkenburg et al. 10.1073/pnas.1403684111 SI Materials and Methods ELISA and Microneutralization. Sera were treated with Receptor Destroying Enzyme II (RDE II, Accurate) before ELISA
NON HUMAN PRIMATE BIOMEDICAL RESEARCH FOR TACKLING EMERGING INFECTIOUS DISEASES (II): ZIKA VIRUS. Special series on Laboratory Animal Science
: ZIKA VIRUS. Special series on Laboratory Animal Science") NON HUMAN PRIMATE BIOMEDICAL RESEARCH FOR TACKLING EMERGING INFECTIOUS DISEASES (II): ZIKA VIRUS Special series on Laboratory Animal Science SPECIAL SERIES ON LABORATORY ANIMAL SCIENCE Signature by Adriano
NON HUMAN PRIMATE BIOMEDICAL RESEARCH FOR TACKLING EMERGING INFECTIOUS DISEASES (II): ZIKA VIRUS Special series on Laboratory Animal Science SPECIAL SERIES ON LABORATORY ANIMAL SCIENCE Signature by Adriano
Effector mechanisms of cell-mediated immunity: Properties of effector, memory and regulatory T cells
 ICI Basic Immunology course Effector mechanisms of cell-mediated immunity: Properties of effector, memory and regulatory T cells Abul K. Abbas, MD UCSF Stages in the development of T cell responses: induction
ICI Basic Immunology course Effector mechanisms of cell-mediated immunity: Properties of effector, memory and regulatory T cells Abul K. Abbas, MD UCSF Stages in the development of T cell responses: induction
NOTES. The Naturally Attenuated Kunjin Strain of West Nile Virus Shows Enhanced Sensitivity to the Host Type I Interferon Response
 JOURNAL OF VIROLOGY, June 2011, p. 5664 5668 Vol. 85, No. 11 0022-538X/11/$12.00 doi:10.1128/jvi.00232-11 Copyright 2011, American Society for Microbiology. All Rights Reserved. NOTES The Naturally Attenuated
JOURNAL OF VIROLOGY, June 2011, p. 5664 5668 Vol. 85, No. 11 0022-538X/11/$12.00 doi:10.1128/jvi.00232-11 Copyright 2011, American Society for Microbiology. All Rights Reserved. NOTES The Naturally Attenuated
Introduction to Immunology Part 2 September 30, Dan Stetson
 Introduction to Immunology Part 2 September 30, 2016 Dan Stetson stetson@uw.edu 441 Lecture #2 Slide 1 of 26 CLASS ANNOUNCEMENT PLEASE NO TREE NUTS IN CLASS!!! (Peanuts, walnuts, almonds, cashews, etc)
Introduction to Immunology Part 2 September 30, 2016 Dan Stetson stetson@uw.edu 441 Lecture #2 Slide 1 of 26 CLASS ANNOUNCEMENT PLEASE NO TREE NUTS IN CLASS!!! (Peanuts, walnuts, almonds, cashews, etc)
Integrin v 3 targeted therapy for Kaposi s sarcoma with an in vitro evolved antibody 1
 Integrin v 3 targeted therapy for Kaposi s sarcoma with an in vitro evolved antibody 1 CHRISTOPH RADER, 2 MIKHAIL POPKOV, JOHN A. NEVES, AND CARLOS F. BARBAS III 2 Department of Molecular Biology and The
Integrin v 3 targeted therapy for Kaposi s sarcoma with an in vitro evolved antibody 1 CHRISTOPH RADER, 2 MIKHAIL POPKOV, JOHN A. NEVES, AND CARLOS F. BARBAS III 2 Department of Molecular Biology and The
PBMC from each patient were suspended in AIM V medium (Invitrogen) with 5% human
 with 5% human") Anti-CD19-CAR transduced T-cell preparation PBMC from each patient were suspended in AIM V medium (Invitrogen) with 5% human AB serum (Gemini) and 300 international units/ml IL-2 (Novartis). T cell proliferation
Anti-CD19-CAR transduced T-cell preparation PBMC from each patient were suspended in AIM V medium (Invitrogen) with 5% human AB serum (Gemini) and 300 international units/ml IL-2 (Novartis). T cell proliferation
effect on the upregulation of these cell surface markers. The mean peak fluorescence intensity
 SUPPLEMENTARY FIGURE 1 Supplementary Figure 1 ASIC1 disruption or blockade does not effect in vitro and in vivo antigen-presenting cell activation. (a) Flow cytometric analysis of cell surface molecules
SUPPLEMENTARY FIGURE 1 Supplementary Figure 1 ASIC1 disruption or blockade does not effect in vitro and in vivo antigen-presenting cell activation. (a) Flow cytometric analysis of cell surface molecules
Supplemental Table I.
 Supplemental Table I Male / Mean ± SEM n Mean ± SEM n Body weight, g 29.2±0.4 17 29.7±0.5 17 Total cholesterol, mg/dl 534.0±30.8 17 561.6±26.1 17 HDL-cholesterol, mg/dl 9.6±0.8 17 10.1±0.7 17 Triglycerides,
Supplemental Table I Male / Mean ± SEM n Mean ± SEM n Body weight, g 29.2±0.4 17 29.7±0.5 17 Total cholesterol, mg/dl 534.0±30.8 17 561.6±26.1 17 HDL-cholesterol, mg/dl 9.6±0.8 17 10.1±0.7 17 Triglycerides,
Question 1. Kupffer cells, microglial cells and osteoclasts are all examples of what type of immune system cell?
 Abbas Chapter 2: Sarah Spriet February 8, 2015 Question 1. Kupffer cells, microglial cells and osteoclasts are all examples of what type of immune system cell? a. Dendritic cells b. Macrophages c. Monocytes
Abbas Chapter 2: Sarah Spriet February 8, 2015 Question 1. Kupffer cells, microglial cells and osteoclasts are all examples of what type of immune system cell? a. Dendritic cells b. Macrophages c. Monocytes
Quantifying Lipid Contents in Enveloped Virus Particles with Plasmonic Nanoparticles
 Quantifying Lipid Contents in Enveloped Virus Particles with Plasmonic Nanoparticles Amin Feizpour Reinhard Lab Department of Chemistry and the Photonics Center, Boston University, Boston, MA May 2014
Quantifying Lipid Contents in Enveloped Virus Particles with Plasmonic Nanoparticles Amin Feizpour Reinhard Lab Department of Chemistry and the Photonics Center, Boston University, Boston, MA May 2014
Putting it Together. Stephen Canfield Secondary Lymphoid System. Tonsil Anterior Cervical LN s
 Putting it Together Stephen Canfield smc12@columbia.edu Secondary Lymphoid System Tonsil Anterior Cervical LN s Axillary LN s Mediastinal/Retroperitoneal LN s Thoracic Duct Appendix Spleen Inguinal LN
Putting it Together Stephen Canfield smc12@columbia.edu Secondary Lymphoid System Tonsil Anterior Cervical LN s Axillary LN s Mediastinal/Retroperitoneal LN s Thoracic Duct Appendix Spleen Inguinal LN
Supplementary Materials for
 immunology.sciencemag.org/cgi/content/full/2/16/eaan6049/dc1 Supplementary Materials for Enzymatic synthesis of core 2 O-glycans governs the tissue-trafficking potential of memory CD8 + T cells Jossef
immunology.sciencemag.org/cgi/content/full/2/16/eaan6049/dc1 Supplementary Materials for Enzymatic synthesis of core 2 O-glycans governs the tissue-trafficking potential of memory CD8 + T cells Jossef
The four serotypes of dengue virus (DENV1 to -4) belong to the
 belong to the") Gamma Interferon (IFN- ) Receptor Restricts Systemic Dengue Virus Replication and Prevents Paralysis in IFN- / Receptor-Deficient Mice Tyler R. Prestwood, Malika M. Morar, Raphaël M. Zellweger, Robyn Miller,
Gamma Interferon (IFN- ) Receptor Restricts Systemic Dengue Virus Replication and Prevents Paralysis in IFN- / Receptor-Deficient Mice Tyler R. Prestwood, Malika M. Morar, Raphaël M. Zellweger, Robyn Miller,
MATERIALS AND METHODS. Neutralizing antibodies specific to mouse Dll1, Dll4, J1 and J2 were prepared as described. 1,2 All
 MATERIALS AND METHODS Antibodies (Abs), flow cytometry analysis and cell lines Neutralizing antibodies specific to mouse Dll1, Dll4, J1 and J2 were prepared as described. 1,2 All other antibodies used
MATERIALS AND METHODS Antibodies (Abs), flow cytometry analysis and cell lines Neutralizing antibodies specific to mouse Dll1, Dll4, J1 and J2 were prepared as described. 1,2 All other antibodies used
Lymphatic System and Immune System. Blood capillaries. Lymphatic vessels/ lymph nodes. Then, identify by labeling these specific structures in part B.
 Name: Date: Period: Lymphatic System and Immune System 1. Figure 21.1 provides an overview of the lymphatic vessels. In part A the relationship between lymphatic vessels and the blood vessels of the cardiovascular
Name: Date: Period: Lymphatic System and Immune System 1. Figure 21.1 provides an overview of the lymphatic vessels. In part A the relationship between lymphatic vessels and the blood vessels of the cardiovascular
Determination of the temporal pattern and importance of BALF1 expression in Epstein-Barr viral infection
 Determination of the temporal pattern and importance of BALF1 expression in Epstein-Barr viral infection Melissa Mihelidakis May 6, 2004 7.340 Research Proposal Introduction Apoptosis, or programmed cell
Determination of the temporal pattern and importance of BALF1 expression in Epstein-Barr viral infection Melissa Mihelidakis May 6, 2004 7.340 Research Proposal Introduction Apoptosis, or programmed cell
Chapter 2 (pages 22 33): Cells and Tissues of the Immune System. Prepared by Kristen Dazy, MD, Scripps Clinic Medical Group
: Cells and Tissues of the Immune System. Prepared by Kristen Dazy, MD, Scripps Clinic Medical Group") Allergy and Immunology Review Corner: Cellular and Molecular Immunology, 8th Edition By Abul K. Abbas, MBBS; Andrew H. H. Lichtman, MD, PhD; and Shiv Pillai, MBBS, PhD. Chapter 2 (pages 22 33): Cells and
Allergy and Immunology Review Corner: Cellular and Molecular Immunology, 8th Edition By Abul K. Abbas, MBBS; Andrew H. H. Lichtman, MD, PhD; and Shiv Pillai, MBBS, PhD. Chapter 2 (pages 22 33): Cells and
SUPPLEMENTARY INFORMATION
 Supplemental Figure 1. Furin is efficiently deleted in CD4 + and CD8 + T cells. a, Western blot for furin and actin proteins in CD4cre-fur f/f and fur f/f Th1 cells. Wild-type and furin-deficient CD4 +
Supplemental Figure 1. Furin is efficiently deleted in CD4 + and CD8 + T cells. a, Western blot for furin and actin proteins in CD4cre-fur f/f and fur f/f Th1 cells. Wild-type and furin-deficient CD4 +
LIST OF ORGANS FOR HISTOPATHOLOGICAL ANALYSIS:!! Neural!!!!!!Respiratory:! Brain : Cerebrum,!!! Lungs and trachea! Olfactory, Cerebellum!!!!Other:!
 LIST OF ORGANS FOR HISTOPATHOLOGICAL ANALYSIS:!! Neural!!!!!!Respiratory:! Brain : Cerebrum,!!! Lungs and trachea! Olfactory, Cerebellum!!!!Other:! Spinal cord and peripheral nerves! Eyes, Inner ear, nasal
LIST OF ORGANS FOR HISTOPATHOLOGICAL ANALYSIS:!! Neural!!!!!!Respiratory:! Brain : Cerebrum,!!! Lungs and trachea! Olfactory, Cerebellum!!!!Other:! Spinal cord and peripheral nerves! Eyes, Inner ear, nasal
B6/COLODR/SPL/11C/83/LAP/#2.006 B6/COLODR/SPL/11C/86/LAP/#2.016 CD11C B6/COLODR/SPL/11C/80/LAP/#2.011 CD11C
 CD3-specific antibody-induced immune tolerance and suppression of autoimmune encephalomyelitis involves TGF-β production through phagocytes digesting apoptotic T cells Sylvain Perruche 1,3, Pin Zhang 1,
CD3-specific antibody-induced immune tolerance and suppression of autoimmune encephalomyelitis involves TGF-β production through phagocytes digesting apoptotic T cells Sylvain Perruche 1,3, Pin Zhang 1,
Product Datasheet. HLA ABC Antibody (W6/32) NB Unit Size: 0.25 mg. Store at -20C. Avoid freeze-thaw cycles. Reviews: 1 Publications: 22
 NB Unit Size: 0.25 mg. Store at -20C. Avoid freeze-thaw cycles. Reviews: 1 Publications: 22") Product Datasheet HLA ABC Antibody (W6/32) NB100-64775 Unit Size: 0.25 mg Store at -20C. Avoid freeze-thaw cycles. Reviews: 1 Publications: 22 Protocols, Publications, Related Products, Reviews, Research
Product Datasheet HLA ABC Antibody (W6/32) NB100-64775 Unit Size: 0.25 mg Store at -20C. Avoid freeze-thaw cycles. Reviews: 1 Publications: 22 Protocols, Publications, Related Products, Reviews, Research
The Immune System. These are classified as the Innate and Adaptive Immune Responses. Innate Immunity
 The Immune System Biological mechanisms that defend an organism must be 1. triggered by a stimulus upon injury or pathogen attack 2. able to counteract the injury or invasion 3. able to recognise foreign
The Immune System Biological mechanisms that defend an organism must be 1. triggered by a stimulus upon injury or pathogen attack 2. able to counteract the injury or invasion 3. able to recognise foreign
Chapter 35 Active Reading Guide The Immune System
 Name: AP Biology Mr. Croft Chapter 35 Active Reading Guide The Immune System Section 1 Phagocytosis plays an important role in the immune systems of both invertebrates and vertebrates. Review the process
Name: AP Biology Mr. Croft Chapter 35 Active Reading Guide The Immune System Section 1 Phagocytosis plays an important role in the immune systems of both invertebrates and vertebrates. Review the process
Lymphoid Organs and Lymphocyte Trafficking. Dr. Issa Abu-Dayyeh
 Lymphoid Organs and Lymphocyte Trafficking Dr. Issa Abu-Dayyeh Invader recognition Where does invader recognition take place?? Secondary lymphoid organs: Lymph nodes Spleen Mucosal-associated lymphoid
Lymphoid Organs and Lymphocyte Trafficking Dr. Issa Abu-Dayyeh Invader recognition Where does invader recognition take place?? Secondary lymphoid organs: Lymph nodes Spleen Mucosal-associated lymphoid
Supplementary Data 1. Alanine substitutions and position variants of APNCYGNIPL. Applied in
 Supplementary Data 1. Alanine substitutions and position variants of APNCYGNIPL. Applied in Supplementary Fig. 2 Substitution Sequence Position variant Sequence original APNCYGNIPL original APNCYGNIPL
Supplementary Data 1. Alanine substitutions and position variants of APNCYGNIPL. Applied in Supplementary Fig. 2 Substitution Sequence Position variant Sequence original APNCYGNIPL original APNCYGNIPL
OBJECTIVES. The Amazing Immune System
 The Amazing Immune System Graphic source: (l) Jeanne Kelly, NIAID; (r) Wikimedia Commons OBJECTIVES Describe at least three components of the immune system Describe the role in our immune response of at
The Amazing Immune System Graphic source: (l) Jeanne Kelly, NIAID; (r) Wikimedia Commons OBJECTIVES Describe at least three components of the immune system Describe the role in our immune response of at
Nature Immunology: doi: /ni Supplementary Figure 1. Examples of staining for each antibody used for the mass cytometry analysis.
 Supplementary Figure 1 Examples of staining for each antibody used for the mass cytometry analysis. To illustrate the functionality of each antibody probe, representative plots illustrating the expected
Supplementary Figure 1 Examples of staining for each antibody used for the mass cytometry analysis. To illustrate the functionality of each antibody probe, representative plots illustrating the expected
Antigen Presentation and T Lymphocyte Activation. Abul K. Abbas UCSF. FOCiS
 1 Antigen Presentation and T Lymphocyte Activation Abul K. Abbas UCSF FOCiS 2 Lecture outline Dendritic cells and antigen presentation The role of the MHC T cell activation Costimulation, the B7:CD28 family
1 Antigen Presentation and T Lymphocyte Activation Abul K. Abbas UCSF FOCiS 2 Lecture outline Dendritic cells and antigen presentation The role of the MHC T cell activation Costimulation, the B7:CD28 family
NK cell flow cytometric assay In vivo DC viability and migration assay
 NK cell flow cytometric assay 6 NK cells were purified, by negative selection with the NK Cell Isolation Kit (Miltenyi iotec), from spleen and lymph nodes of 6 RAG1KO mice, injected the day before with
NK cell flow cytometric assay 6 NK cells were purified, by negative selection with the NK Cell Isolation Kit (Miltenyi iotec), from spleen and lymph nodes of 6 RAG1KO mice, injected the day before with
Supporting Information
 Supporting Information Aldridge et al. 10.1073/pnas.0900655106 Fig. S1. Flow diagram of sublethal (a) and lethal (b) influenza virus infections. (a) Infection of lung epithelial cells by influenza virus
Supporting Information Aldridge et al. 10.1073/pnas.0900655106 Fig. S1. Flow diagram of sublethal (a) and lethal (b) influenza virus infections. (a) Infection of lung epithelial cells by influenza virus
Chapter 12: The Lymphatic System
 Chapter 12: The Lymphatic System Immune System Composed of many nonspecific and specific defenses Lymphatic System also plays an important role in establishing immunity Lymphatic System Major components
Chapter 12: The Lymphatic System Immune System Composed of many nonspecific and specific defenses Lymphatic System also plays an important role in establishing immunity Lymphatic System Major components
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature10134 Supplementary Figure 1. Anti-inflammatory activity of sfc. a, Autoantibody immune complexes crosslink activating Fc receptors, promoting activation of macrophages, and WWW.NATURE.COM/NATURE
doi:10.1038/nature10134 Supplementary Figure 1. Anti-inflammatory activity of sfc. a, Autoantibody immune complexes crosslink activating Fc receptors, promoting activation of macrophages, and WWW.NATURE.COM/NATURE
Supplementary Figure 1.
 Supplementary Figure 1. Female Pro-ins2 -/- mice at 5-6 weeks of age were either inoculated i.p. with a single dose of CVB4 (1x10 5 PFU/mouse) or PBS and treated with αgalcer or control vehicle. On day
Supplementary Figure 1. Female Pro-ins2 -/- mice at 5-6 weeks of age were either inoculated i.p. with a single dose of CVB4 (1x10 5 PFU/mouse) or PBS and treated with αgalcer or control vehicle. On day
Chapter 17. The Lymphatic System and Immunity. Copyright 2010, John Wiley & Sons, Inc.
 Chapter 17 The Lymphatic System and Immunity Immunity Innate Immunity Fast, non-specific and no memory Barriers, ph extremes, Phagocytes & NK cells, fever, inflammation, complement, interferon Adaptive
Chapter 17 The Lymphatic System and Immunity Immunity Innate Immunity Fast, non-specific and no memory Barriers, ph extremes, Phagocytes & NK cells, fever, inflammation, complement, interferon Adaptive
Principles of Adaptive Immunity
 Principles of Adaptive Immunity Chapter 3 Parham Hans de Haard 17 th of May 2010 Agenda Recognition molecules of adaptive immune system Features adaptive immune system Immunoglobulins and T-cell receptors
Principles of Adaptive Immunity Chapter 3 Parham Hans de Haard 17 th of May 2010 Agenda Recognition molecules of adaptive immune system Features adaptive immune system Immunoglobulins and T-cell receptors
Study Guide 23, 24 & 47
 Study Guide 23, 24 & 47 STUDY GUIDE SECTION 23-3 Bacteria and Humans Name Period Date 1. One bacterial disease that is transmitted by contaminated drinking water is a. Lyme disease b. gonorrhea c. tuberculosis
Study Guide 23, 24 & 47 STUDY GUIDE SECTION 23-3 Bacteria and Humans Name Period Date 1. One bacterial disease that is transmitted by contaminated drinking water is a. Lyme disease b. gonorrhea c. tuberculosis
Unit 5 The Human Immune Response to Infection
 Unit 5 The Human Immune Response to Infection Unit 5-page 1 FOM Chapter 21 Resistance and the Immune System: Innate Immunity Preview: In Chapter 21, we will learn about the branch of the immune system
Unit 5 The Human Immune Response to Infection Unit 5-page 1 FOM Chapter 21 Resistance and the Immune System: Innate Immunity Preview: In Chapter 21, we will learn about the branch of the immune system
SUPPLEMENTARY METHODS
 SUPPLEMENTARY METHODS Histological analysis. Colonic tissues were collected from 5 parts of the middle colon on day 7 after the start of DSS treatment, and then were cut into segments, fixed with 4% paraformaldehyde,
SUPPLEMENTARY METHODS Histological analysis. Colonic tissues were collected from 5 parts of the middle colon on day 7 after the start of DSS treatment, and then were cut into segments, fixed with 4% paraformaldehyde,
Nature Immunology: doi: /ni Supplementary Figure 1. Huwe1 has high expression in HSCs and is necessary for quiescence.
 Supplementary Figure 1 Huwe1 has high expression in HSCs and is necessary for quiescence. (a) Heat map visualizing expression of genes with a known function in ubiquitin-mediated proteolysis (KEGG: Ubiquitin
Supplementary Figure 1 Huwe1 has high expression in HSCs and is necessary for quiescence. (a) Heat map visualizing expression of genes with a known function in ubiquitin-mediated proteolysis (KEGG: Ubiquitin
BMDCs were generated in vitro from bone marrow cells cultured in 10 % RPMI supplemented
 Supplemental Materials Figure S1. Cultured BMDCs express CD11c BMDCs were generated in vitro from bone marrow cells cultured in 10 % RPMI supplemented with 15 ng/ml GM-CSF. Media was changed and fresh
Supplemental Materials Figure S1. Cultured BMDCs express CD11c BMDCs were generated in vitro from bone marrow cells cultured in 10 % RPMI supplemented with 15 ng/ml GM-CSF. Media was changed and fresh
What is the role of animal models in studying protective titres and the need for establishing surrogates/correlates of protection?
 What is the role of animal models in studying protective titres and the need for establishing surrogates/correlates of protection? Alan D.T. Barrett Department of Pathology and Sealy Center for Vaccine
What is the role of animal models in studying protective titres and the need for establishing surrogates/correlates of protection? Alan D.T. Barrett Department of Pathology and Sealy Center for Vaccine
Solid-in-oil peptide nanocarriers for transcutaneous cancer vaccine delivery. against melanoma
 Supplementary Information for Solid-in-oil peptide nanocarriers for transcutaneous cancer vaccine delivery against melanoma Rie Wakabayashi,,a,b Masato Sakuragi,,a Shuto Kozaka, a Yoshiro Tahara, a Noriho
Supplementary Information for Solid-in-oil peptide nanocarriers for transcutaneous cancer vaccine delivery against melanoma Rie Wakabayashi,,a,b Masato Sakuragi,,a Shuto Kozaka, a Yoshiro Tahara, a Noriho
and follicular helper T cells is Egr2-dependent. (a) Diagrammatic representation of the
 Diagrammatic representation of the") Supplementary Figure 1. LAG3 + Treg-mediated regulation of germinal center B cells and follicular helper T cells is Egr2-dependent. (a) Diagrammatic representation of the experimental protocol for the
Supplementary Figure 1. LAG3 + Treg-mediated regulation of germinal center B cells and follicular helper T cells is Egr2-dependent. (a) Diagrammatic representation of the experimental protocol for the
Figure S1. PMVs from THP-1 cells expose phosphatidylserine and carry actin. A) Flow
 Flow") SUPPLEMENTARY DATA Supplementary Figure Legends Figure S1. PMVs from THP-1 cells expose phosphatidylserine and carry actin. A) Flow cytometry analysis of PMVs labelled with annexin-v-pe (Guava technologies)
SUPPLEMENTARY DATA Supplementary Figure Legends Figure S1. PMVs from THP-1 cells expose phosphatidylserine and carry actin. A) Flow cytometry analysis of PMVs labelled with annexin-v-pe (Guava technologies)
Chapter 3, Part A (Pages 37-45): Leukocyte Migration into Tissues
: Leukocyte Migration into Tissues") Allergy and Immunology Review Corner: Chapter 3, Part A (pages 37-45) of Cellular and Molecular Immunology (Seventh Edition), by Abul K. Abbas, Andrew H. Lichtman and Shiv Pillai. Chapter 3, Part A (Pages
Allergy and Immunology Review Corner: Chapter 3, Part A (pages 37-45) of Cellular and Molecular Immunology (Seventh Edition), by Abul K. Abbas, Andrew H. Lichtman and Shiv Pillai. Chapter 3, Part A (Pages
Gastroenteritis and viral infections
 Gastroenteritis and viral infections A Large number of viruses are found in the human gut; these include some that are associated with gastroenteritis Rotaviruses Adenoviruses 40/41 Caliciviruses Norwalk-like
Gastroenteritis and viral infections A Large number of viruses are found in the human gut; these include some that are associated with gastroenteritis Rotaviruses Adenoviruses 40/41 Caliciviruses Norwalk-like
Dengue Virus Infects Macrophages and Dendritic Cells in a Mouse Model of Infection
 MAJOR ARTICLE Dengue Virus Infects Macrophages and Dendritic Cells in a Mouse Model of Infection Jennifer L. Kyle, 1,2 P. Robert Beatty, 3 and Eva Harris 1,2 1 Division of Infectious Diseases, School of
MAJOR ARTICLE Dengue Virus Infects Macrophages and Dendritic Cells in a Mouse Model of Infection Jennifer L. Kyle, 1,2 P. Robert Beatty, 3 and Eva Harris 1,2 1 Division of Infectious Diseases, School of
Mucosal Immunology Sophomore Dental and Optometry Microbiology Section I: Immunology. Robin Lorenz
 Mucosal Immunology Sophomore Dental and Optometry Microbiology Section I: Immunology Robin Lorenz rlorenz@uab.edu Why do we Need to Understand How the Mucosal Immune System Works? The mucosa is the major
Mucosal Immunology Sophomore Dental and Optometry Microbiology Section I: Immunology Robin Lorenz rlorenz@uab.edu Why do we Need to Understand How the Mucosal Immune System Works? The mucosa is the major
Superior Fluorescent Labeling Dyes Spanning the Full Visible Spectrum...1. Trademarks: HiLyte Fluor (AnaSpec, Inc.)
") Table of Contents Fluor TM Labeling Dyes Superior Fluorescent Labeling Dyes Spanning the Full Visible Spectrum....1 Fluor TM 405 Dye, an Excellent Alternative to Alexa Fluor 405 & DyLight 405....2 Fluor
Table of Contents Fluor TM Labeling Dyes Superior Fluorescent Labeling Dyes Spanning the Full Visible Spectrum....1 Fluor TM 405 Dye, an Excellent Alternative to Alexa Fluor 405 & DyLight 405....2 Fluor
Title of file for HTML: Supplementary Information Description: Supplementary Figures and Supplementary Table
 Title of file for HTML: Supplementary Information Description: Supplementary Figures and Supplementary Table Title of file for HTML: Peer Review File Description: Innate Scavenger Receptor-A regulates
Title of file for HTML: Supplementary Information Description: Supplementary Figures and Supplementary Table Title of file for HTML: Peer Review File Description: Innate Scavenger Receptor-A regulates
Supporting Information Table of Contents
 Supporting Information Table of Contents Supporting Information Figure 1 Page 2 Supporting Information Figure 2 Page 4 Supporting Information Figure 3 Page 5 Supporting Information Figure 4 Page 6 Supporting
Supporting Information Table of Contents Supporting Information Figure 1 Page 2 Supporting Information Figure 2 Page 4 Supporting Information Figure 3 Page 5 Supporting Information Figure 4 Page 6 Supporting
sequences of a styx mutant reveals a T to A transversion in the donor splice site of intron 5
 sfigure 1 Styx mutant mice recapitulate the phenotype of SHIP -/- mice. (A) Analysis of the genomic sequences of a styx mutant reveals a T to A transversion in the donor splice site of intron 5 (GTAAC
sfigure 1 Styx mutant mice recapitulate the phenotype of SHIP -/- mice. (A) Analysis of the genomic sequences of a styx mutant reveals a T to A transversion in the donor splice site of intron 5 (GTAAC
Coronaviruses cause acute, mild upper respiratory infection (common cold).
.") Coronaviruses David A. J. Tyrrell Steven H. Myint GENERAL CONCEPTS Clinical Presentation Coronaviruses cause acute, mild upper respiratory infection (common cold). Structure Spherical or pleomorphic enveloped
Coronaviruses David A. J. Tyrrell Steven H. Myint GENERAL CONCEPTS Clinical Presentation Coronaviruses cause acute, mild upper respiratory infection (common cold). Structure Spherical or pleomorphic enveloped
4. TEXTBOOK: ABUL K. ABBAS. ANDREW H. LICHTMAN. CELLULAR AND MOLECULAR IMMUNOLOGY. 5 TH EDITION. Chapter 2. pg
 LECTURE: 03 Title: CELLS INVOLVED IN THE IMMUNE RESPONSE LEARNING OBJECTIVES: The student should be able to: Identify the organs where the process of the blood formation occurs. Identify the main cell
LECTURE: 03 Title: CELLS INVOLVED IN THE IMMUNE RESPONSE LEARNING OBJECTIVES: The student should be able to: Identify the organs where the process of the blood formation occurs. Identify the main cell
SUPPLEMENTARY INFORMATION. Supplementary Figures S1-S9. Supplementary Methods
 SUPPLEMENTARY INFORMATION SUMO1 modification of PTEN regulates tumorigenesis by controlling its association with the plasma membrane Jian Huang 1,2#, Jie Yan 1,2#, Jian Zhang 3#, Shiguo Zhu 1, Yanli Wang
SUPPLEMENTARY INFORMATION SUMO1 modification of PTEN regulates tumorigenesis by controlling its association with the plasma membrane Jian Huang 1,2#, Jie Yan 1,2#, Jian Zhang 3#, Shiguo Zhu 1, Yanli Wang
2. The normal of the gut, and vagina keep the growth of pathogens in check. 3. in the respiratory tract sweep out bacteria and particles.
 Chapter 39 Immunity I. Three Lines of Defense A. Surface Barriers to Invasion 1. is an important barrier. 2. The normal of the gut, and vagina keep the growth of pathogens in check. 3. in the respiratory
Chapter 39 Immunity I. Three Lines of Defense A. Surface Barriers to Invasion 1. is an important barrier. 2. The normal of the gut, and vagina keep the growth of pathogens in check. 3. in the respiratory
L-selectin Is Essential for Delivery of Activated CD8 + T Cells to Virus-Infected Organs for Protective Immunity
 Cell Reports Supplemental Information L-selectin Is Essential for Delivery of Activated CD8 + T Cells to Virus-Infected Organs for Protective Immunity Rebar N. Mohammed, H. Angharad Watson, Miriam Vigar,
Cell Reports Supplemental Information L-selectin Is Essential for Delivery of Activated CD8 + T Cells to Virus-Infected Organs for Protective Immunity Rebar N. Mohammed, H. Angharad Watson, Miriam Vigar,
SUPPLEMENTARY INFORMATION
 1. Supplementary Figures and Legends Supplementary Fig. 1. S1P-mediated transcriptional regulation of integrins expressed in OP/monocytoid cells. Real-time quantitative PCR analyses of mrna for two integrins,
1. Supplementary Figures and Legends Supplementary Fig. 1. S1P-mediated transcriptional regulation of integrins expressed in OP/monocytoid cells. Real-time quantitative PCR analyses of mrna for two integrins,
Fine Mapping of a cis-acting Sequence Element in Yellow Fever Virus RNA That Is Required for RNA Replication and Cyclization
 JOURNAL OF VIROLOGY, Feb. 2003, p. 2265 2270 Vol. 77, No. 3 0022-538X/03/$08.00 0 DOI: 10.1128/JVI.77.3.2265 2270.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved. Fine Mapping
JOURNAL OF VIROLOGY, Feb. 2003, p. 2265 2270 Vol. 77, No. 3 0022-538X/03/$08.00 0 DOI: 10.1128/JVI.77.3.2265 2270.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved. Fine Mapping
Suppl Video: Tumor cells (green) and monocytes (white) are seeded on a confluent endothelial
 and monocytes (white) are seeded on a confluent endothelial") Supplementary Information Häuselmann et al. Monocyte induction of E-selectin-mediated endothelial activation releases VE-cadherin junctions to promote tumor cell extravasation in the metastasis cascade
Supplementary Information Häuselmann et al. Monocyte induction of E-selectin-mediated endothelial activation releases VE-cadherin junctions to promote tumor cell extravasation in the metastasis cascade
Molecular and Cellular Basis of Immune Protection of Mucosal Surfaces
 Molecular and Cellular Basis of Immune Protection of Mucosal Surfaces Department of Biologic & Materials Sciences School of Dentistry University of Michigan Ann Arbor, Michigan 48109-1078 1 Image quality
Molecular and Cellular Basis of Immune Protection of Mucosal Surfaces Department of Biologic & Materials Sciences School of Dentistry University of Michigan Ann Arbor, Michigan 48109-1078 1 Image quality
Supplementary Information:
 Supplementary Information: Follicular regulatory T cells with Bcl6 expression suppress germinal center reactions by Yeonseok Chung, Shinya Tanaka, Fuliang Chu, Roza Nurieva, Gustavo J. Martinez, Seema
Supplementary Information: Follicular regulatory T cells with Bcl6 expression suppress germinal center reactions by Yeonseok Chung, Shinya Tanaka, Fuliang Chu, Roza Nurieva, Gustavo J. Martinez, Seema
General information. Cell mediated immunity. 455 LSA, Tuesday 11 to noon. Anytime after class.
 General information Cell mediated immunity 455 LSA, Tuesday 11 to noon Anytime after class T-cell precursors Thymus Naive T-cells (CD8 or CD4) email: lcoscoy@berkeley.edu edu Use MCB150 as subject line
General information Cell mediated immunity 455 LSA, Tuesday 11 to noon Anytime after class T-cell precursors Thymus Naive T-cells (CD8 or CD4) email: lcoscoy@berkeley.edu edu Use MCB150 as subject line
Supplementary Figure 1. H-PGDS deficiency does not affect GI tract functions and anaphylactic reaction. (a) Representative pictures of H&E-stained
 Representative pictures of H&E-stained") 1 2 3 4 5 6 7 8 9 10 11 Supplementary Figure 1. H-PGDS deficiency does not affect GI tract functions and anaphylactic reaction. (a) Representative pictures of H&E-stained jejunum sections ( 200 magnification;
1 2 3 4 5 6 7 8 9 10 11 Supplementary Figure 1. H-PGDS deficiency does not affect GI tract functions and anaphylactic reaction. (a) Representative pictures of H&E-stained jejunum sections ( 200 magnification;
T-cell activation T cells migrate to secondary lymphoid tissues where they interact with antigen, antigen-presenting cells, and other lymphocytes:
 Interactions between innate immunity & adaptive immunity What happens to T cells after they leave the thymus? Naïve T cells exit the thymus and enter the bloodstream. If they remain in the bloodstream,
Interactions between innate immunity & adaptive immunity What happens to T cells after they leave the thymus? Naïve T cells exit the thymus and enter the bloodstream. If they remain in the bloodstream,
T-cell activation T cells migrate to secondary lymphoid tissues where they interact with antigen, antigen-presenting cells, and other lymphocytes:
 Interactions between innate immunity & adaptive immunity What happens to T cells after they leave the thymus? Naïve T cells exit the thymus and enter the bloodstream. If they remain in the bloodstream,
Interactions between innate immunity & adaptive immunity What happens to T cells after they leave the thymus? Naïve T cells exit the thymus and enter the bloodstream. If they remain in the bloodstream,
Supplementary Fig. 1. Delivery of mirnas via Red Fluorescent Protein.
 prfp-vector RFP Exon1 Intron RFP Exon2 prfp-mir-124 mir-93/124 RFP Exon1 Intron RFP Exon2 Untransfected prfp-vector prfp-mir-93 prfp-mir-124 Supplementary Fig. 1. Delivery of mirnas via Red Fluorescent
prfp-vector RFP Exon1 Intron RFP Exon2 prfp-mir-124 mir-93/124 RFP Exon1 Intron RFP Exon2 Untransfected prfp-vector prfp-mir-93 prfp-mir-124 Supplementary Fig. 1. Delivery of mirnas via Red Fluorescent