Transcriptional co-activator LEDGF/p75 modulates HIV-1 integrase ACCEPTED. mediated concerted integration
|
|
|
- Cory Richardson
- 5 years ago
- Views:
Transcription
1 JVI Accepts, published online ahead of print on 31 January 2007 J. Virol. doi: /jvi Copyright 2007, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved Transcriptional co-activator LEDGF/p75 modulates HIV-1 integrase mediated concerted integration Krishan K. Pandey*, Sapna Sinha, and Duane P. Grandgenett* Institute for Molecular Virology, Saint Louis University Health Sciences Center, St. Louis, MO 63110, USA Running Title: LEDGF Interactions with HIV-1 Synaptic Complexes Word Count in Abstract: 212 Word Count for Text: 6684 *Corresponding authors: Krishan K Pandey, Institute for Molecular Virology, 3681 Park Avenue, Saint Louis University Health Sciences Center, St. Louis, MO 63110, USA. Tel: (314) Fax: (314) pandeykk@slu.edu Duane P. Grandgenett, Institute for Molecular Virology, 3681 Park Avenue, Saint Louis University Health Sciences Center, St. Louis, MO 63110, USA. Tel: (314) Fax: (314) grandgdp@slu.edu 1
2 21 ABSTRACT Human transcriptional co-activator p75/lense epithelium-derived growth factor (LEDGF) binds HIV-1 integrase (IN). We studied the effects of LEDGF on the assembly of and activity of HIV-1 synaptic complexes, which upon association with target, mediate concerted integration of viral DNA substrates in vitro. We found that while augmenting single-ended viral DNA integration into target DNA, the host factor in a concentration dependent manner was able to either stimulate or abrogate concerted integration. LEDGF modestly stimulated (two- to threefold) concerted integration at low molar ratios to IN (<1). The modest stimulation was independent of solution conditions and several different viral DNA substrates. In solution, concerted integration was inhibited if the molar ratios of LEDGF to IN were >1, apparently due to the disruption of IN-IN interactions essential for formation of active synaptic complexes, prior to their association with circular target. Isolated IN binding domain (IBD) of LEDGF was sufficient to stimulate and inhibit concerted integration as observed with full-length protein, albeit at lower efficiencies. Our data shows that LEDGF differentially affects IN-DNA complexes mediating single-ended viral DNA integration and synaptic complexes mediating concerted integration. Synaptic complexes associated with target, termed strand transfer complexes (STC), are resistant to disruption by high concentrations of LEDGF. The results suggest that LEDGF may influence HIV-1 integration in vivo. 2
3 39 INTRODUCTION Retroviruses are unique in that they insert DNA replicas of their genomes into host cell chromosomal DNA. Following reverse transcription of the viral RNA genome into linear DNA, preintegration complexes (PIC) are formed that contain the viral integrase (IN) and several other viral and cellular proteins (2). The PIC traverses the nuclear envelope and associates with chromosomes, where IN inserts the viral genome. Isolated cytoplasmic PIC can insert the viral DNA ends in a concerted fashion into exogenously supplied target DNA substrates in vitro (3, 5, 6, 22, 33, 50). Major advances have occurred in identifying target sites and understanding the basis for target site selectivity exhibited by human immunodeficiency virus (HIV-1), murine leukemia virus (MLV), and avian sarcoma-leukemia virus (ASLV) in human chromosomes (25, 34, 36, 37, 51). HIV-1 predominantly integrates within genes, having a preference for those that are actively transcribing. MLV prefers integration into or near transcription start sites, while ASLV exhibits a slight preference for genes and no preference for transcription start sites. These results suggest that HIV-1 and MLV may interact with chromatin-associated factors and/or transcriptional co- factors that direct the PIC to transcription units in the case of HIV-1, and to or near transcriptional start sites in the case of MLV (16). Simian immunodeficiency virus (SIV) selectivity for integration into human chromosomes is similar to HIV-1 (14). Retrovirus site selection has implications for human gene therapy (46). Several cellular proteins were observed to co-fractionate with the PIC; their proposed roles range from trafficking and transport of the PIC into the nucleus, structural maintenance and selection of host target sites (40, 42). A human transcriptional co-activator, LEDGF/p75 (17), 3
4 was identified as a high-affinity interactor to ectopically expressed HIV-1 IN in human cells (9). LEDGF was later shown to associate with divergent feline immunodeficiency virus (FIV) but not MLV IN (30). Functional HIV-1 and FIV PICs could be immunoprecipitated with anti-ledgf antibody (30). LEDGF specifically interacts with the catalytic core domain (CCD) of HIV-1 IN in a stoichiometric ratio of 1 (7, 10, 31). The HIV-1 IN binding domain (IBD) in LEDGF maps to a highly conserved region within the C-terminal region of the protein (7, 8, 10). Depletion of LEDGF by RNAi results in a loss of stable association between IN and chromatin, suggesting that LEDGF tethers IN to cellular DNA (15, 30, 32, 45). Furthermore, HIV-1 IN mutant (Q168A) deficient for binding to LEDGF failed to associate with chromosomes (15). RNAi-mediated depletion of LEDGF appears to have little effect on HIV-1 replication (30, 43) although other LEDGF depletion studies suggested a two- to four fold reduction in HIV-1 replication with the block occurring at the integration step (44). In another LEDGF depletion study, an average 53% reduced infection and a 74% decrease in integration was observed in HIV-1 infected macrophages suggesting a modest involvement of LEDGF in integration (52). Further intensified RNAi and dominant negative protein approaches suggested that LEDGF is an essential HIV-1 integration cofactor (29). Lastly, genomic analyses of HIV-1 infected cells depleted of LEDGF as compared to non-depleted cells indicate that the protein is involved in the selection of chromosomal integration sites (13). These studies suggest that LEDGF is important for HIV-1 integration although its precise functions remain to be established. The in vitro biochemical dissection of retroviral integration provides important mechanistic insights into the integration process in vivo. Synaptic complexes consisting of purified IN and viral DNA substrates can efficiently insert two viral DNA ends in a concerted fashion into circular target DNA (12, 47-49), in a process here termed full-site integration (Fig. 1). The 4
5 insertion of one DNA substrate end into target DNA produces a circular half-site (CHS) integration product. IN in detergent-lysed HIV-1 virions (19) as well as recombinant HIV-1 IN catalyze these reactions (20, 23, 26, 38, 39). Although LEDGF has been demonstrated to stimulate gross strand transfer activity of HIV-1 IN in vitro (8-10, 41), the effect of the co-factor on full-site integration has not been addressed. In this report, we show that the concentration of LEDGF modulates synaptic complexes but not strand transfer complexes (STC)(Fig. 1) that produce full-site products. Modest stimulation of full-site integration (two- to threefold) is observed at molar ratios of LEDGF to IN (<1) while at ratios >1, LEDGF disrupts IN-IN interactions necessary for assembly of synaptic complexes in solution. MATERIALS AND METHODS Production and labeling of DNA substrates. The linear 4.1 kbp and 0.48 kbp DNA substrates containing HIV-1 wt U3 and U5 long terminal repeats (LTR) attachment ends were previously described (38, 39). NdeI digestion of the plasmids produced linear substrates with 3 OH recessed ends containing a 5 TA overhang (Fig. 1). To obtain a 3.4 kbp single-ended 3 OH recessed U5 substrate, the NdeI linearized and dephosphorylated 4.1 kbp plasmid was digested with SacI. The DNA donor substrates were 5 end-labeled on the non-catalytic strand with [γ- 32 ] ATP and T4 polynucleotide kinase. The specific activities were ~1,500-2,000 cpm (Cerenkov) per ng of double end-labeled donor substrates and ~1,000 cpm for single-end labeled substrates. The fragments were isolated from agarose gels and extracted using the Qiaquick Gel Extraction Kit (Qiagen, Valencia, CA) or electroelution. Supercoiled pbsk2 -Zeo (2.7 kbp), kindly provided by Karen Moreau (35), was used as target DNA. 5
6 A HIV-1 linear 4.7 kbp DNA containing wt U3 and U5 LTR blunt-ends (11) was also used for strand transfer. ScaI digestion of the plasmid produced a substrate containing HIV-1 natural DNA blunt-ends (11). To obtain single-ended U5 and U3 blunt-end substrates, the ScaI linearized and dephosphorylated plasmid was further digested with NcoI to yield a 1.6 kbp U5 substrate (Fig. 1) and a 2.4 kbp U3 substrate. Preprocessed natural DNA fragments containing 5 AC overhangs were constructed by ligation of 93 bp ds oligonucleotides containing HIV-1 U5 LTR sequences to a puc19 derived fragment. The ligated DNA fragments were purified by agarose gel electrophoresis and end-labeled. The synthetic 5 AC fragment was 1.5 kbp in length and the nonspecific end was blunt-ended (Fig. 1). Integration assay. The HIV-1 integration assays with all of the DNA substrates (Fig. 1) were performed as described (38). Briefly, HIV-1 IN was assembled with the donor substrate in presence of 20 mm HEPES (ph 7.0), 5 mm dithiothreitol, 10 mm MgCl 2, 25 µm ZnCl 2, 50 mm NaCl and 10% polyethylene glycol (6000 daltons) at 14ºC for 15 min. The NaCl concentration was varied between 50 mm and 125 mm as indicated. The IN and donor concentrations are described in each experiment. The standard reaction volumes were 200 µl. The reactions were initiated by addition of 500 ng of supercoiled target DNA and incubated for 2 hrs at 37ºC. The above reactions were stopped by addition of EDTA, SDS, and proteinase K to the final concentration of 25 mm, 0.5%, and 1 mg/ml, respectively. Equivalent amounts of labeled integration products (~10,000 to 20,000 cpm) were subjected to 0.8% or 1.5% agarose gel electrophoresis in TBE buffer for 10 to 16 h at 100 volts, depending upon the size of the donor. Please note that the full-site and CHS products migrate at different positions on the two different agarose gels. The gels were dried and exposed to a PhosphorImager screen. Full-site and CHS products were quantified using a Storm 860 system (Amersham Biosciences). To evaluate the 6
7 differential effects of LEDGF upon the assembly and catalytic properties of synaptic complexes and STC, LEDGF was added either to assembled synaptic complexes at 14ºC or was bound with HIV-1 IN at 14ºC prior to the addition of donor (Fig. 1). In other sets of experiments, target was mixed with assembled synaptic complexes prior to adding LEDGF. Lastly, preformed STC were allowed to form for 2 h at 37ºC and then challenged with LEDGF prior to the analysis on native 0.7% agarose gels. Identification and evaluation of STC by native agarose gels. The synaptic complexes were assembled as described in the previous section. After the addition of target, STCs were allowed to form at 37 C for 2h. The reactions were stopped by adding 20 mm EDTA and the products were subjected to electrophoresis on a 0.7% native agarose gel in TBE buffer without 1 M urea (27). Two-dimensional gel electrophoresis of complexes separated on agarose gels was performed as described (27). Purification of recombinant proteins. IN from the pny clone was expressed in E. coli BL21 (DE3) cells and purified to near homogeneity as described (38). Concentrations of HIV-1 IN were determined using RSV IN as a standard (12). LEDGF was purified as previously described (9, 10) and its concentration was determined using the molar extinction coefficient value of 16,500 M -1 cm -1 determined from its amino acid sequence. Isolated IBD of LEDGF (residues ) and full-length mutant LEDGF (D336N) were prepared as described (10). Analysis of integration products. The integration products obtained with HIV-1 IN and the 4.1 kb donor substrate were previously analyzed by EcoRI digestion and DNA sequencing of the viral DNA-host junctions (38). The full-site integration (9.5 kbp) products obtained with the single-ended U5 donor (3.4 kb) were also digested with EcoRI and yielded the correct 2.9 kb U5- target-u5 product (38). The 1.6 kbp DNA substrate containing blunt-ended U5 produced the 7
8 predicted 5.9 kbp full-site product that upon digestion by either HpaI, MluI, or Bsu36I produced the predicted full-site DNA fragments (data not shown). The viral DNA-host junctions of the full-site integration products derived from coupled U5-U5 and U5-U3 substrates were sequenced. The U5-U5 full-site products were isolated from a 0.8% agarose gel and purified by electroelution. The integration products were amplified by PCR using U5 LTR sequence as primers (forward as well as reverse 5 GTCAGTGTGGAAAATCTCTAGC 3 ) using PfuUltra Hotstart PCR Master Mix (Stratagene). PCR cycling parameters included initial denaturation at 95 C for 2 min, followed by 30 cycles at 95 C for 1 min, 56 C for 1 min and 72 C for 3.5 min. Final extension was for 10 min at 72 C. PCR products (~2.7 kb) were purified from agarose gels by Qiaquick Gel extraction kit and cloned in Zero Blunt PCR vector (Invitrogen). Recombinant clones were sequenced using custom primers (KKPBlunt244 5 GGTGACGCGTTAGAATACTCAAGC 3 and ccd-r 5 GCCCCGGCGTGTCAATAATATC 3 ) to analyze the LTR-target junction and host site duplications. The U5-U3 full site products were isolated from the gel, digested with BglII and BamHI. The 3.1 kb products were gel purified; self ligated and used to transform E. coli DH5α cells. Recombinants were sequenced using custom primers 6176U5BglII 5 CTGACCTGCCAGATCTGCTAACTAGGGAACCCAC 3 and 6176U3BamHI (5 CGCCCGCCACTAGTGGATCCAAAGAATTCTATC 3 ). RESULTS LEDGF, in a concentration dependent manner, differentially interacts with synaptic complexes and other IN-DNA complexes mediating CHS integration. Full-site integration by HIV-1 synaptic complexes in vitro allows functional analysis of putative cellular cofactors (Fig. 1). The production of full-site integration products by HIV-1 synaptic complexes is dependent 8
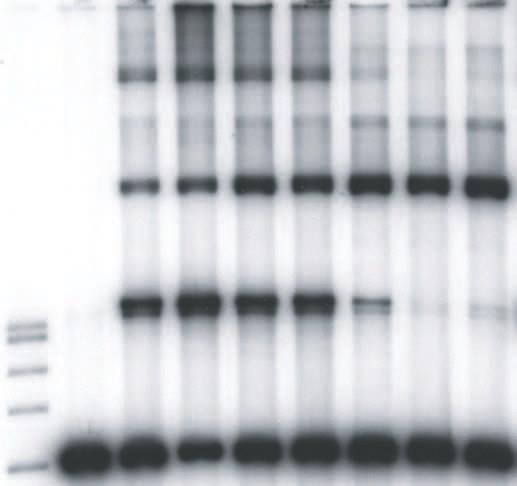
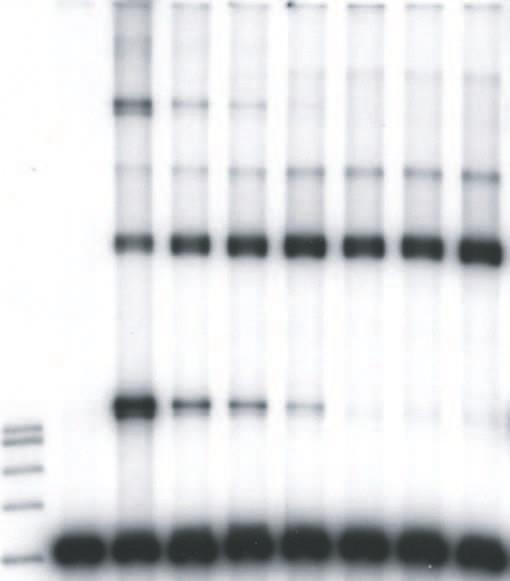
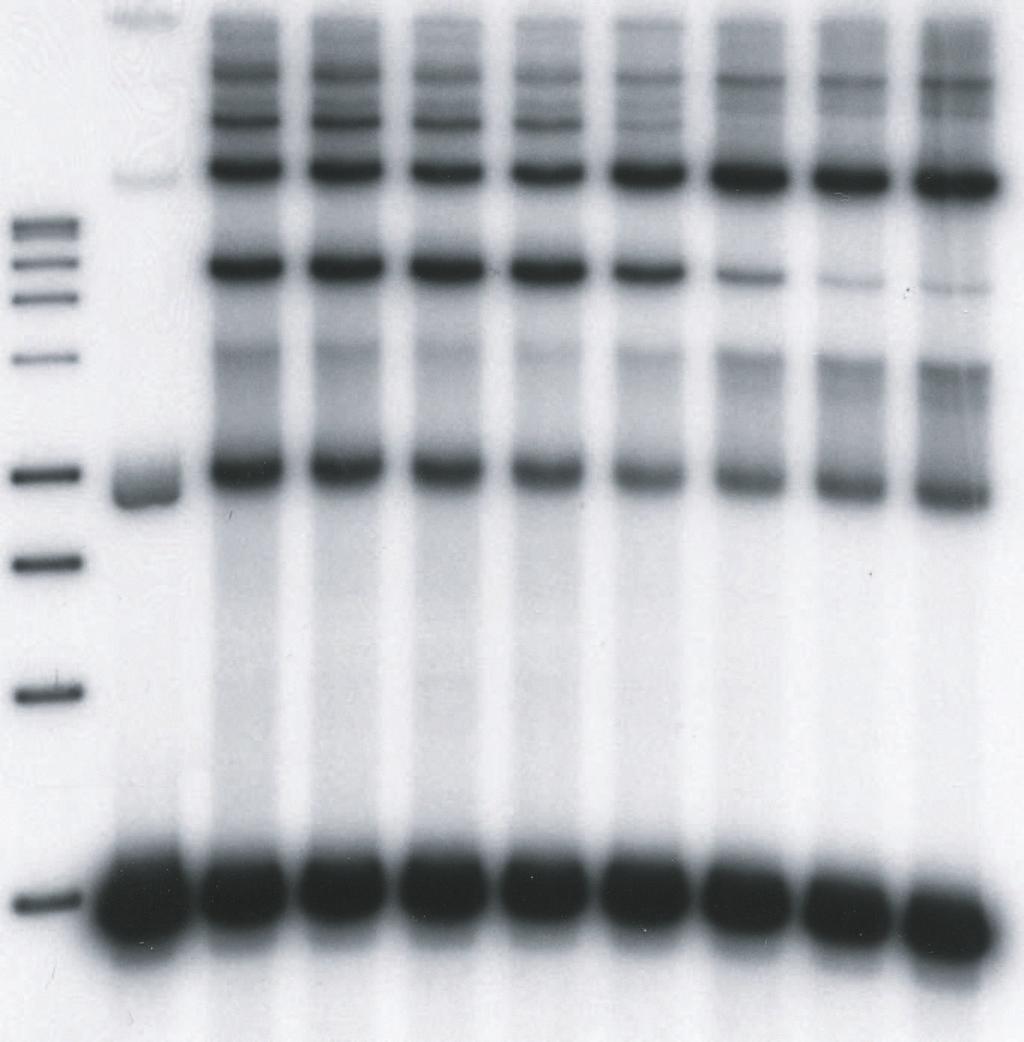
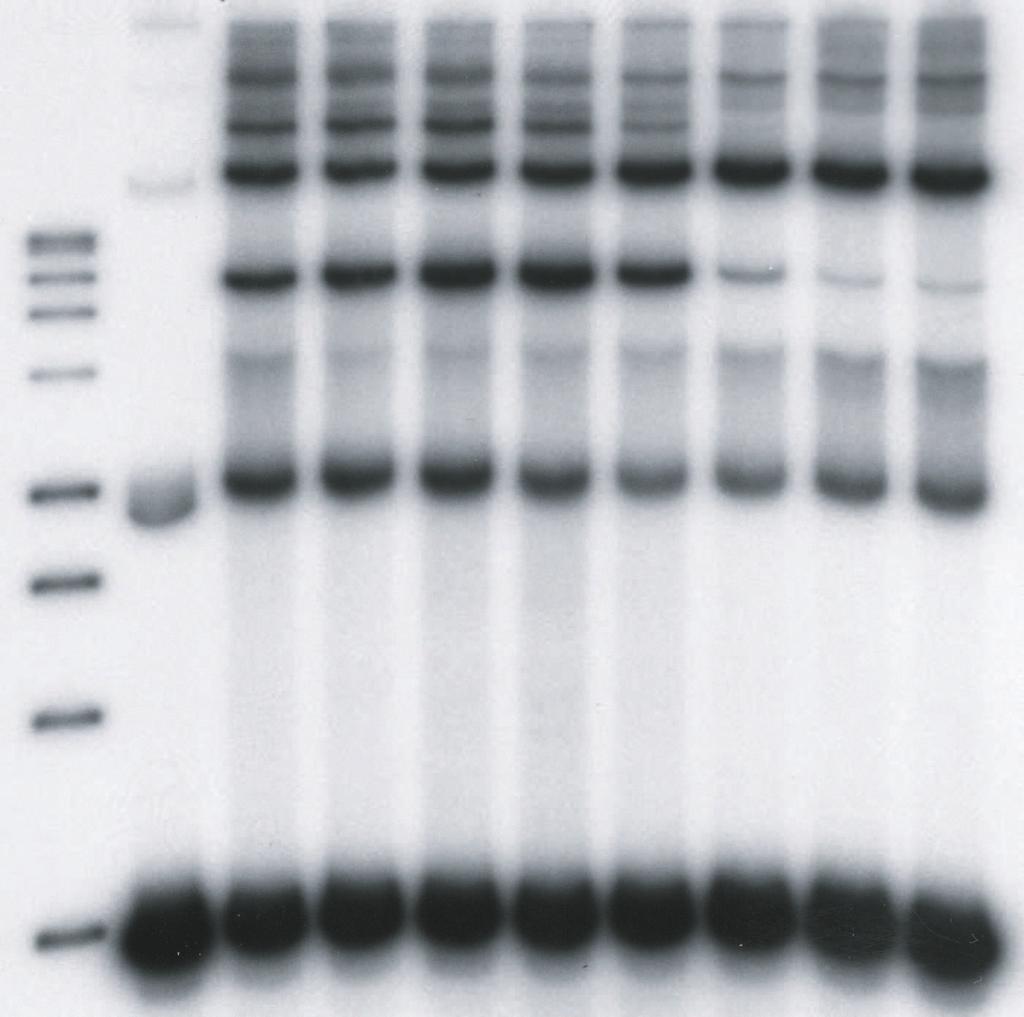
9 upon the concentrations of HIV-1 IN in the reaction mixtures, the length and concentrations of the DNA substrates, the use of preprocessed or blunt-ended viral substrates, and the solution conditions (19, 20, 26, 27, 38, 39). We explored these parameters to investigate the effects of LEDGF on HIV-1 integration in vitro. We first used a 4.1 kbp DNA substrate containing preprocessed 3 OH recessed U3 and U5 ends with 5 TA overhangs to examine the effects of LEDGF on the assembly and catalytic activity of HIV-1 synaptic complexes (Fig. 1). HIV-1 IN (5 nm) was assembled onto the 4.1 kbp donor (0.2 nm), prior to the addition of increasing concentrations of LEDGF in the assembly mixture, followed by analysis of strand transfer products (Fig. 2A). LEDGF at 1.5 and 3.7 nm stimulated full-site integration activity approximately 2.0 and 1.3-fold, respectively, (Fig. 2A, lanes 4 and 5) in comparison to IN without LEDGF (lane 3). A minor donor-donor product that migrates slightly faster than the full-site product (38), is also affected by LEDGF (Fig. 2A). Higher concentrations of LEDGF (>7.5 nm) in the assembly mixture eliminated full-site integration activity presumably by preventing assembly of the synaptic complex (Fig. 2A, lanes 7 to 9). In contrast, the production of CHS products was increased at all concentrations of LEDGF using the preprocessed 3 OH substrate (Fig. 2A, lanes 3 to 9). Under conditions of this assay, the CHS products were enhanced ~5-fold (Fig. 2A, lane 9). Several other integration products migrating above the CHS products were inhibited, stimulated or unaffected at various LEDGF concentrations. The data suggests that LEDGF, in a concentration dependent manner, differentially interacts with synaptic complexes capable of full-site activity, and with nucleoprotein complexes that possess CHS activity. Order-of-addition affects the assembly of the HIV-1 synaptic complex. Binding of LEDGF to IN prior to addition of the donor with 5 TA overhangs at 14 C essentially prevented 9
10 the accumulation of the full-site product (Fig. 2B, lanes 4 to 9) in comparison to assembly in the absence of LEDGF (Fig. 2B, lane 3). These results suggest that binding of LEDGF to IN prevents assembly of synaptic complexes, presumably by steric hindrance or conformational alterations in IN. In contrast, CHS products were enhanced at all LEDGF concentrations. The CHS activity was enhanced ~4-fold at 37 nm LEDGF (Fig. 2B, lane 9). Effects of DNA substrate and solution conditions on the properties of LEDGF. A 3.4 kbp single-ended U5 donor (0.3 nm) containing preprocessed 5 TA ends (Fig. 1) was assembled with HIV-1 IN (5 nm). Addition of LEDGF to assembled synaptic complexes slightly stimulated the production of full-site products at low LEDGF concentrations while at higher LEDGF concentrations (10 nm) full-site integration was again disrupted (data not shown). On the other hand, the CHS products were reproducibly enhanced by LEDGF. The interactions between LEDGF and IN within synaptic complexes are not overly sensitive to concentrations of components and solution conditions. A 0.48 kbp donor (2.5 nm) containing preprocessed HIV-1 U3 and U5 att sites with 5 TA overhangs was assembled with HIV-1 IN (16 nm) on ice in the presence of 12% dimethyl sulfoxide and 100 mm NaCl (39). After assembly, varying concentrations of LEDGF were added, followed by target DNA and incubation at 37 C (data not shown). The results suggest that even under significantly altered assay conditions, LEDGF interactions with assembled synaptic complexes were similar to those previously observed (Fig. 2A). In particular, the disruption of full-site integration only occurred at stoichiometric ratios of LEDGF to IN >1. Interactions of LEDGF with synaptic complexes containing DNA with natural blunt or 3 OH recessed termini. Processing of the natural HIV-1 DNA blunt ends by IN channels the reaction to concerted or full-site integration (26). Synaptic complexes were assembled using a 10
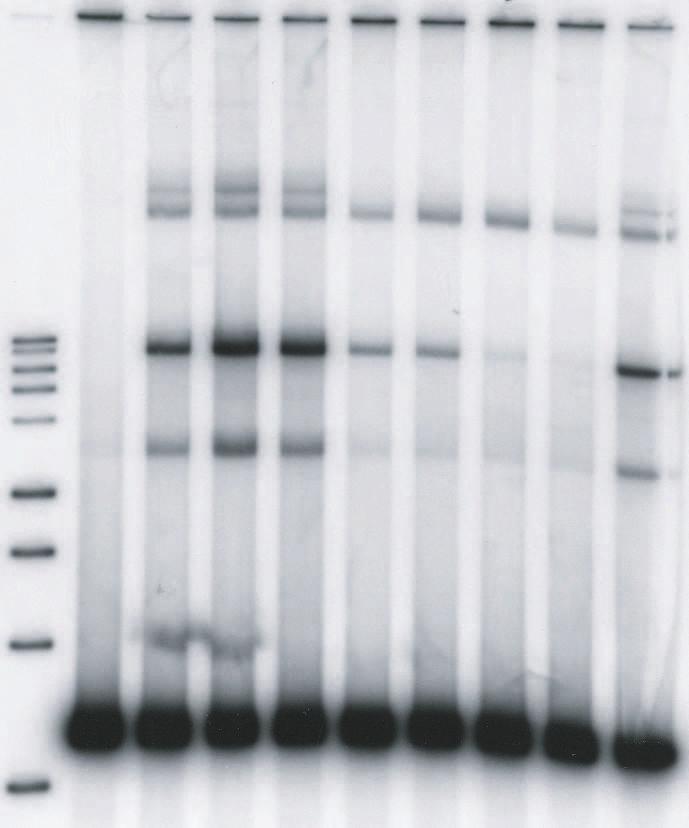
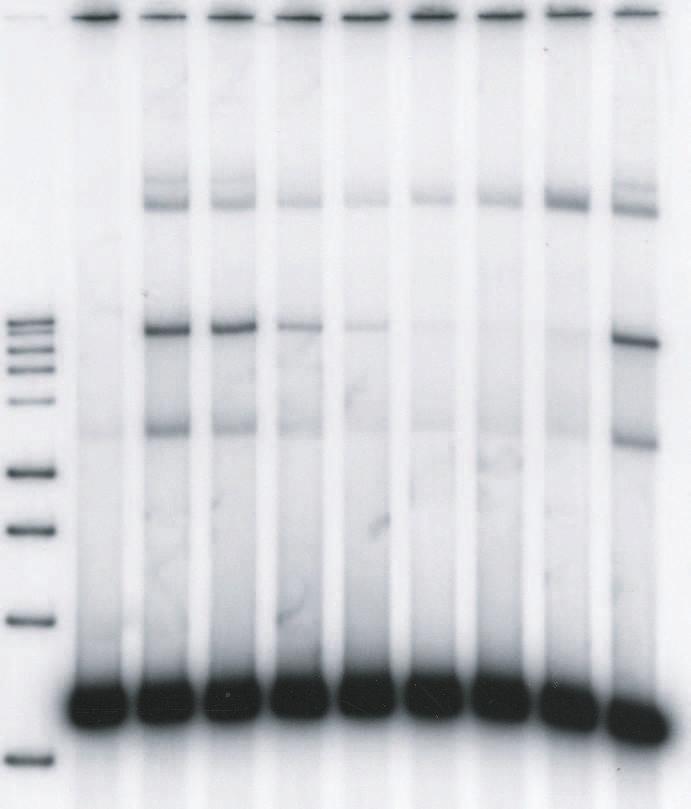
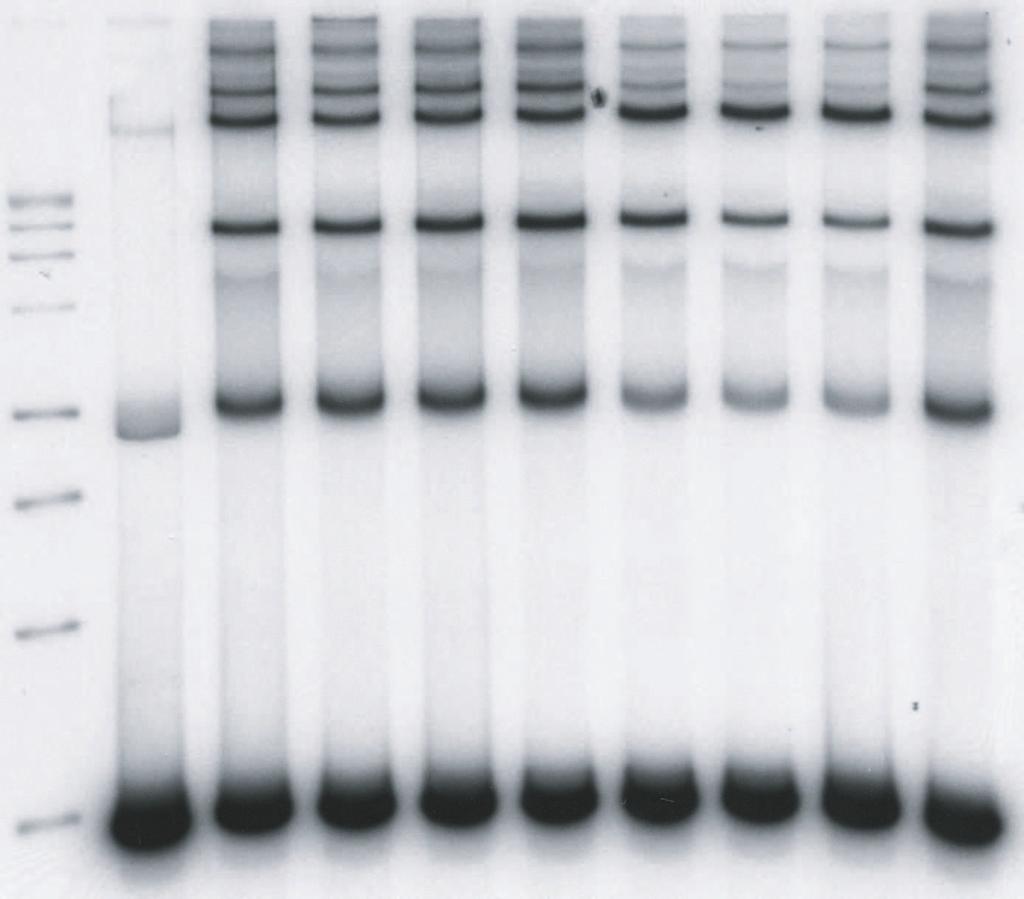
11 kbp donor (0.52 nm) containing a single U5 blunt end and IN (10 nm) at 14 C in 125 mm NaCl (Fig. 3). LEDGF at 2.5 nm and 5 nm stimulated production of the full-site integration products ~2 fold (Fig. 3A, lanes 4 and 5) in comparison to its absence (Fig. 3A, lanes 3 and 10). Titration experiments established that the optimum stimulation (~2-fold) of full-site integration required ~3 nm LEDGF at 100 mm NaCl in the assembly mixtures (Table 1)(data not shown). Restriction analysis of the full-site products using the 1.6 kbp blunt-ended donor and DNA sequence analysis of the viral-host junctions verified that the full-site integration reaction had occurred (26). Sequence analysis of the donor-target junctions verified that the HIV-1 5 bp host site duplications were present with a fidelity of 76% for U5-U5 products (47 plasmid clones) and 88% for U5-U3 products (9 plasmid clones). The other recombinants possessed small size deletions at the target site. Higher concentrations of LEDGF prevented assembly of the synaptic complexes in the assembly buffer (Fig. 3A, lanes 6 to 9) with little observable effects on CHS production. Prior binding of LEDGF to IN before the addition of the 1.6 kbp blunt ended donor almost completely prevented assembly of synaptic complexes even at 5 nm LEDGF (Fig. 3B). Similar results were obtained using both order of addition schemes with LEDGF, if a blunt-ended donor containing U3 LTR sequences was used (data not shown). The results suggest that LEDGF at low nm concentrations enhances assembly of synaptic complexes with blunt ends and subsequent fullsite integration activity. However, at higher concentrations, LEDGF efficiently prevented further assembly of synaptic complexes with the natural blunt-ended substrate. The processing of DNA ends by IN in HIV-1 PIC occurs soon after reverse transcription and prior to transport of the PIC across the nuclear envelope (33). We tested HIV-1 synaptic complexes containing a 1.5 kbp donor possessing 3 OH recessed ends with a natural 5 AC 11
12 overhang for interactions with LEDGF. The donor (0.42 nm) was assembled with IN (10 nm) at 14 C in 125 mm NaCl (Fig. 4). Titration of LEDGF demonstrated that there was slight stimulation (1.3-fold) of full-site integration activity (Fig. 4A, lanes 4 to 6) while full-site activity started to decrease at greater than 10 nm LEDGF in the assembly buffer (Fig. 4A, lanes 7 to 10)(Fig. 4B). Higher concentrations of LEDGF are required to inhibit full-site activity with 3 OH recessed ended (Fig. 4B) than with blunt-end donors (Fig. 3). The results indicate that low concentrations of LEDGF modestly stimulated full-site integration, while higher concentrations of LEDGF prevents assembly of the synaptic complexes (see Fig. 10 for model). To further investigate the effect of LEDGF on full-site integration activity using the 1.6 kbp U5 blunt-ended donor, we executed another order-of-addition experiment where LEDGF (50 nm)(fig. 3) was or was not added at different times after initiation of strand transfer at 37 C. Addition of LEDGF at 30, 45 and 60 min time points after initiation of strand transfer had a minimum effect on the total amount of full-site product produced in 2 h at 37 C in comparison, to the appropriate controls without LEDGF (data not shown). Addition of LEDGF at 0 and 15 min had a significant effect similar to preincubation with LEDGF (Fig. 3). The results suggest that although LEDGF prevents assembly of the synaptic complex, the STC (Fig. 1) formed later in the reaction is apparently resistant to disruption by LEDGF (see Fig. 8). A subset of the donor-donor integration products responds to LEDGF in the same way as full-site products resulting from synaptic complexes. Presumably, juxtaposition of two donor molecules is required prior to presentation of target to produce the full-site product (Fig. 1). Assembly of RSV and AMV IN synaptic complexes occurs with two DNA donors containing 3 OH recessed ends at 14 C without target present (1). Intriguingly, the production of HIV kbp donor-donor products from two 1.6 kbp donors containing blunt ends (Fig. 3A, lanes 3 to 5; 12
13 Fig. 5) and of 3 kbp donor-donor products from two 1.5 kbp donors with 5 AC overhangs (Fig. 4 and Fig. 6) paralleled the production of full-site products because each were affected by LEDGF. Approximately 60% of the complexes responsible for producing the donor-donor products with the 1.5 kbp 5 AC donor are eliminated by LEDGF (Fig. 4B and Fig. 6) and completely by it with the 1.6 kbp DNA donor with blunt ends (Fig. 3 and Fig. 5), prior to strand transfer. Restriction and DNA sequence analysis of a 9.4 kbp donor-donor product from two 4.7 kbp substrates with blunt-ends suggest that end joining occurs preferentially near the LTR ends (11). Similar results were obtained with restriction analysis of the 3.2 kbp donor-donor products obtained with the 1.6-kbp donor (data not shown). In contrast, the Y-type donor-donor products (Fig. 1) (11, 21, 39) migrating between 3 kbp and 4 kbp were not attenuated by LEDGF (Fig. 4 to Fig. 6). The modest stimulation and inhibition of the specific size donor-donor integration products parallels that of the full-site products suggesting that LEDGF modulates interactions between IN subunits bound at two LTR ends but not with CHS and Y-type products containing one LTR end. Interactions of isolated LEDGF-IBD with synaptic complexes are similar to those of full-length LEDGF. LEDGF is a modular protein that interacts with HIV-1 IN via a small alpha-helical domain, IBD (residues ), located within its C-terminal region (8, 10, 45). In addition to the IBD, the full-length protein contains other structural and functional elements, such as N-terminal PWWP domain, a nuclear localization signal, and a pair of putative AThooks (8, 31, 41, 45). The IBD of LEDGF specifically interacts with the CCD and likely the N- terminal domain of HIV-1 IN (4, 7, 10, 31). Unlike full-length protein, IBD does not bind DNA in vitro (41). The IBD was essential but not sufficient to stimulate HIV-1 IN activity in uncoupled stand transfer assays (8). 13
14 Synaptic complexes were assembled with a 1.6 kbp DNA (0.42 nm) containing a single U5 blunt end and IN (10 nm) at 50 mm NaCl prior to addition of LEDGF fragment containing the IBD (residues ) and further incubation at 14 C (Fig. 5). The NaCl concentration was lowered to 50 mm NaCl instead of 125 mm NaCl (Fig. 3 and Fig. 4) to help mediate more efficient 3 OH processing of the blunt ended substrate (26). Slight stimulation was observed for full-site integration and the specific 3.2 kbp donor-donor products up to 10 nm IBD. Thereafter, the full-site and the 3.2 kbp donor-donor integration reactions were essentially inhibited while the CHS integration was unaffected. This result indicates that local allosteric and/or steric effects of the IN-LEDGF-IBD interactions are sufficient to prevent assembly of the synaptic complex. Synaptic complexes containing a 5 AC overhang display increased resistance to the inhibitory effects of LEDGF and LEDGF-IBD. We wanted to determine if LEDGF and IBD binding to IN prior to assembly of the synaptic complex with the 5 AC overhang donor was as effective as LEDGF binding to IN using the 4.1 kbp donor with a 5 TA overhang (Fig. 2B) or 1.6 kbp blunt ended donor (Fig. 3B). Interestingly, there was no significant inhibition of any strand transfer products by IBD at low concentrations when it was bound to IN prior to the addition of preprocessed 1.5 kbp 5 AC overhang donor (Fig. 6A, lanes 4 to 6). Approximately 50% inhibition of full-site products was observed only at higher IBD concentrations (Fig. 6A, lanes 8 and 9). Likewise, pre-incubation of LEDGF with IN prior to addition of the 5 AC donor resulted in no inhibition and only at higher concentrations of LEDGF was inhibition observed (Fig. 6B, lanes 8 to 10). The results suggest that, i) catalytic activities of HIV-1 IN are not affected by engagement of the LEDGF-binding site on the IN CCD per se, ii) the association of IN with the 5 AC donor end in synaptic complexes is higher than that observed with either the 5 TA donor end (Fig. 2) or the natural blunt-ended DNA (Fig. 3) thus resulting in more efficient 14
15 assembly and/or increased stability of the synaptic complexes, and iii) LEDGF is more effective than IBD in preventing assembly of the synaptic complexes at higher nm concentrations (Fig. 6). LEDGF does not disrupt the STC. The time-dependent formation of the STC (Fig. 1) occurs through different stages that results in a stable IN-DNA complex containing the full-site integration product (27). The STC is stable upon electrophoresis in 0.8% agarose gel with 1 M urea and is resistant to treatment with either 0.6 M NaCl, 0.1% NP40, or 10 µg/ml heparin (27). We determined that the STC could be identified on 0.7% native agarose gels (Fig. 7A) and is resistant to disruption by LEDGF (Fig. 8). To produce the STC, IN (15 nm) was assembled with 1.6 kbp U5 blunt-ended donor at 14 C for 15 min, followed by the addition of target and further incubation for 2 h at 37 C. Aliquots of the sample were subjected to 0.7% native agarose gel electrophoresis at 4 C (Fig. 7A, lane 3). Following separation of the complexes on the native gel (Fig. 7A, lane 3), the native gel was treated with SDS and was subjected to two-dimensional gel electrophoresis (Fig. 7A, right panel). The STC and CHS on the native gel gave rise to the predicted strand transfer products as previously demonstrated by two-dimensional gel analysis (27). From PhosphorImager analysis, the STC (Fig. 7A, lane 3) contained only full-site product with minor amounts of CHS and free donor (Fig. 7A, right panel). The two slower migrating bands (marked with dots)(fig. 7A, lane 3) each contained ~80% full-site product and other minor products all of which, are apparently looped together by IN in a nonspecific fashion (11, 21). The smeared products above the STC (Fig. 7A, lane 3) contained mostly full-site and 3.2 kbp donordonor/y-type products (Fig. 7A, right panel). The unknown higher molecular weight product probably represents integration of another donor into the full-site product or other DNA structures. A sample was also deproteinized and analyzed on 0.7% agarose to quantify the integration products (Fig. 7A, gel positioned below two-dimensional gel). The % donor 15
16 incorporated into full-site, CHS, and 3.2 kbp donor-donor/y-type products were 19%, 6% and 9%, respectively. Lastly, nearly equivalent quantities of STCs containing full-site product were evident on a native 0.7% agarose gel in comparison to the same sample on a gel containing 1 M urea, as determined by PhosphorImager analysis (Fig. 7B). The data demonstrates that STC can be identified on native agarose gels even in the presence of nonspecific DNA binding by IN. From our earlier results, we conclude that LEDGF can stimulate the assembly process at low nm concentrations while at high concentrations it prevents assembly of the synaptic complex. In preliminary studies, addition of 50 nm LEDGF to the assembly mixture (Fig. 3, lane 9) was found to essentially inhibit all full-site integration activity with the U5 blunt-ended donor as well as detection of the STC in 0.7% native agarose gels (data not shown). Next, we determined that preformed STCs are resistant to disruption by LEDGF. STC were formed with IN (15 nm) for 2h at 37 C followed by addition of EDTA to prevent further catalysis. These samples were then subjected to challenge by LEDGF. In one set of samples, 50 nm LEDGF was added and incubated for different time intervals at 14 C (Fig. 8A, lanes 5, 7, and 9). In the parallel set, samples were maintained at 14 C in the absence of LEDGF (Fig. 8A, lanes 4, 6, and 8). The samples were run on a native 0.7% agarose gel, without (Fig. 8A) or with deproteinization (Fig. 8B). From PhosphorImager data, the STC and the two slower migrating bands (Fig. 8A, lanes 5, 7, and 9) that contained full-site products (see Fig. 7, right panel) were essentially stable (~85%) on native gels upon LEDGF challenge in comparison to their control lanes without LEDGF (Fig. 8A, lanes 4, 6, and 8). LEDGF also bound to the DNA in the STC, CHS, 3.2 kbp donor-donor and free donor because of all these bands on the native agarose gel are shifted slightly upwards (Fig. 8, lanes 5, 7, and 9). In all of the samples, the % donor incorporated into full-site and the 3.2 kbp D-D products were essentially equivalent (30% and 15%, respectively)(fig. 8B, lanes 3 16
17 to 9). Two-dimensional studies of the 15 min samples not treated (Fig. 8C) or treated (Fig. 8D) with LEDGF on the native gel (Fig. 8A, lanes 4 and 5, respectively) demonstrated that LEDGF did not modify the amounts of full-site, D-D and other products in comparison to the untreated control, as defined by PhosphorImager analysis. Two-dimensional analysis of the 30 min samples produced similar results (data not shown). The results suggest that LEDGF does not significantly disrupt the STC. LEDGF does not modulate RSV IN mediated full-site integration. LEDGF binding appears to be specific to lentiviral IN (4, 30). Concordantly, a titration of LEDGF did not specifically affect recombinant RSV IN activities in vitro. There was no stimulation of CHS or full-site integration (Fig. 9). These results indicate that a direct interaction between LEDGF and HIV-1 IN is essential for the observed modest stimulation and disruption of IN-IN interactions necessary for full-site integration. DISCUSSION LEDGF differentially modulates synaptic complexes capable of full-site integration and IN-DNA complexes producing CHS products in vitro. LEDGF binds tightly and specifically to HIV-1 IN, primarily recognizing the catalytic core domain of the viral enzyme in the absence of viral DNA (7, 10, 32). Reconstituted synaptic complexes (26, 38, 39, 47, 48) proceed through a series of nucleoprotein complexes throughout the pathway to full-site integration (27). Through the monitoring of specific integration products, we found that the strand transfer activity of synaptic complexes is modulated, depending upon the structure of and the concentration of LEDGF. In one order of addition scheme, IN was bound to viral DNA at 14 C prior to the addition of LEDGF. At low molar ratios of LEDGF to IN (<1), there was a 17
18 modest stimulation of full-site product formation, presumably due to a facilitation of the assembly process which results in a ~two- to threefold increase in the number of active synaptic complexes formed. At higher molar ratios (>1), LEDGF effected a concentration dependent attenuation of full-site integration. When present at high concentrations in solution, LEDGF disrupts IN-IN interactions necessary for assembly of synaptic complexes, resulting in a quantitative conversion to CHS products. LEDGF challenge experiments showed that the preformed STC is resistant to disruption (Fig. 10). Correlations to integration studies in vivo. In vivo knockdown studies have brought forth conflicting results regarding the role of LEDGF in HIV-1 replication. The depletion of LEDGF by RNAi appears to have little effect on virus replication, possibly because an estimated ~2.5 x 10 4 copies of LEDGF persist per cell in these depletion experiments (28, 43). Estimation of the number of LEDGF copies in control HeLa-P4 cells is ~4 x In contrast, another LEDGF depletion study suggests a two- to fourfold reduction of HIV-1 replication with the defect occurring at the integration step (44). Inhibition of HIV-1 integration (74%) in macrophages depleted of LEDGF (70 to 80%) suggests a modest involvement of LEDGF in integration (52). Fractionally minute levels of chromatin-associated LEDGF appear sufficient for HIV-1 integration (29). Recent knockout studies of LEDGF in mouse embryo fibroblasts (LEDGF, -/-) infected by HIV-1 resulted in an approximate five- to tenfold reductions in virus titer, which, in a preliminary experiment, was found to directly correlated with a fivefold reduction in PIC activity in vitro as compared to results obtained with virus-infected control cells (personal communication, Vandegraaff, et al.). The results suggest that the concentration of and location of LEDGF in cells, in an undefined manner, modulates HIV-1 integration in vivo. 18
19 Our control reactions without LEDGF may represent PIC in cells devoid of LEDGF or saltstripped PIC (43). Addition of LEDGF to synaptic complexes at low molar ratios of LEDGF to IN (<1) stimulated (two- to threefold) the assembly of synaptic complexes (Fig. 2 to 6) that mediated full-site integration (Table 1). The effects observed with LEDGF at low nm concentrations in our studies corroborate the above knockout and knockdown studies of LEDGF in vivo. Possibly, this modest stimulation of full-site integration by LEDGF is due to its ability to promote tethering of synaptic complexes to target DNA (15, 29, 30, 32, 45) and/or increase the binding affinity of HIV-1 IN to DNA (4). Importantly, the assembly of synaptic complexes (Fig. 2 to 6) proceeds normally in the presence of low concentrations of LEDGF suggesting that, as in HIV-1 PIC which contain LEDGF (30), the juxtaposition of the viral ends is not hindered by this protein. The actual number of molecules of LEDGF in wt and depleted PIC in vivo are unknown. Integration activity in salt-stripped PICs is stimulated ~40%, ~6%, and ~32% by the addition of LEDGF, IBD, and LEDGF mutant (D366N), respectively, relative to the starting PIC activity (43). The stimulation of full-site integration observed at low concentrations of LEDGF correlates with these PIC reconstitution studies, implicating a functional role for LEDGF in retrovirus integration. Additional compelling evidence for the involvement of LEDGF in HIV-1 integration comes from knockdown studies of LEDGF that demonstrate a redistribution of integration target sites upon depletion of LEDGF in the infected cells (13). Although statistically significant, the redistribution of integration sites was not absolute, leading to ~5-20% reduction of integration in transcription units depending on the cell type and ~25% reduction of integration events into genes responsive to LEDGF (13). The partial redistribution of integration sites from transcription units to CG regions may suggest that overall integration is not always tightly controlled by 19
20 LEDGF or by other cellular factors (13). These data suggest that IN is possibly involved in target site selection in cells that may proceed, in some cases, independently of cellular proteins (13, 34, 37). Synaptic complexes are fully functional in the absence of LEGDF (Fig. 2 to Fig. 8)(27, 38, 39) possibly, supporting the observation that LEDGF has a modest involvement in HIV-1 integration in macrophages (52) and the partial redistribution of integration sites (13). Because LEDGF, at low concentrations promotes legitimate full-site integration, it would be of interest to determine under appropriate assay conditions, if LEDGF affects target site selection in vitro, thus extending the above in vivo studies. It will be interesting also to determine if the fidelity of the HIV-1 5 bp host site duplications observed in vitro, which occurs at ~75% (18, 26, 38, 39), is significantly improved at low nm concentrations of LEDGF with several of the donor substrates used in this study. This will further show that low concentrations of LEDGF impart stability to the synaptic complex and in analogy, to the PIC. What are the concentration and structure requirements for LEDGF mediated disruption of IN-IN interactions necessary for assembly of synaptic complexes and maintenance of STCs? The PIC transverses the cytoplasm and is transported into the nucleus, suggesting that it possesses structural stability. Synaptic complexes formed in vitro with avian IN are stable (1, 48) and particularly when the HIV-1 synaptic complex is associated with target, forming the STC (27). The assembly and stability of these structures is due to yet unidentified IN-IN interactions that are required for these two macromolecular structures. We performed order of addition experiments with LEDGF (Fig. 1) to investigate how it affects the assembly of the synaptic complex. Pre-incubation of LEDGF with HIV-1 IN prior to the addition of a donor possessing a 5 TA overhang completely prevents the formation of synaptic complexes, while IN-DNA complexes producing CHS products are significantly 20
21 stimulated with increasing concentrations of LEDGF (Fig. 2B). This steady increase in CHS products occurs in the near complete absence of any synaptic complexes (Fig. 2B, lanes 4 to 9) suggesting LEDGF assists the binding of IN to single DNA ends (4). These results stand in contrast to the analogous situation in which the donor possesses a natural 5 AC overhang (Fig. 6B). The synaptic complexes with the 5 AC overhang donor induces stronger IN-IN interactions required for assembly as compared to the synaptic complexes with 5 TA overhang (Fig. 2B). A recent report shows that the 5 -C is involved in a hydrogen bond with the Gln-148 of IN stabilizing the conformation of the flexible loop adjacent to the active site of IN (24). Our results suggest that the configuration of IN associated with the natural 3 OH recessed ends may be optimized for interacting with LEDGF. In a different order of addition scheme, IN was assembled with viral DNA at 14 C prior to the addition of LEDGF. The tendency of LEDGF to more readily prevent the assembly of synaptic complexes using the blunt-ended 5 AC donor substrate (Fig. 3) in comparison to the 5 AC overhang substrate (Fig. 4) further suggests that the IN-IN interactions associated with the latter substrate contribute to the stable structure of the IN-DNA complex found in the cytoplasmic PIC containing 3 OH recessed ends. Titration experiments showed that LEDGF up to 100 nm, does not inhibit 3 OH processing of native blunt-ended donor by IN at 10 nm (data not shown). Upon assembly of IN with the 5 AC overhang substrate prior to addition of LEDGF, facilitation of assembly of synaptic complexes proceeds at molar ratios of LEDGF to IN <1 (Fig. 4). In contrast with our experimental conditions where synaptic complexes are in equilibrium with high concentrations of LEDGF in solution (LEDGF to IN >1), PICs are exposed to the restrictions imposed by the complex cellular environment where, only minute amounts of LEDGF located on chromatin may interact with the PIC (29). 21
22 What are the structural requirements of LEDGF for both the modest stimulation of full-site products and assembly of the synaptic complex? The IBD of LEDGF specifically interacts with the CCD of IN (7, 10) but this specific interface is likely to signify only a portion of the total contacts between LEDGF and IN (32). Slight stimulation of full-site activity was observed with the IBD while disruption of this activity was similar to LEDGF but less effective (Fig. 5 and Fig. 6). Even though the IBD fragment and LEDGF containing the D366N mutation does not interact with IN in pull-down experiments (10), high concentration of full-length LEDGF containing this mutation appears to behave similar to wt LEDGF with the native blunt-ended and 5 AC overhang substrates (data not shown). However, unlike LEDGF, it does not modestly stimulate full-site integration as reported earlier for PIC (43). Disruption of full-site integration by isolated IBD, full length wt LEDGF as well as its mutant D366N occurred essentially at the same molar ratio of each respective protein to IN. These results suggest that the interactions of LEDGF with IN in the context of synaptic complexes is extensive in disrupting subunit interactions necessary for assembly. However, preformed STC were essentially resistant to LEDGF at 50 nm (Fig. 8) which at this concentration, completely prevented full-site integration when added before assembly of the synaptic complex (Fig. 2 to 6). It is unknown whether the resistance of STC to challenge by LEDGF is due to a lack of interaction with the CCD or other sites on IN subunits within the STC. ACKNOWLEDGEMENTS This work was supported by National Institutes of Allergy and Infectious Diseases Grant AI31334 to D.P.G. We thank Peter Cherepanov for his generous supply of wt and mutant LEDGF proteins, plasmids and for his discussions and comments on the manuscript. We thank 22
23 both Jacob Zahm for his comments and several experiments and Sibes Bera for producing the U5 and U3 LTR donor substrates that contained the 5 AC overhang. We also like to thank Nickolas Vandergraaff and Alan Engelman for sharing results prior to publication. REFERENCES 1. Bera, S., A. C. Vora, R. Chiu, T. Heyduk, and D. P. Grandgenett Synaptic complex formation of two retrovirus DNA attachment sites by integrase: a fluorescence energy transfer study. Biochemistry 44: Brown, P. O Integration, p In J. M. Coffin, Hughes, S. Varmus, H. eds (ed.), Retroviruses. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY. 3. Brown, P. O., B. Bowerman, H. E. Varmus, and J. M. Bishop Correct integration of retroviral DNA in vitro. Cell 49: Busschots, K., J. Vercammen, S. Emiliani, R. Benarous, Y. Engelborghs, F. Christ, and Z. Debyser The interaction of LEDGF/p75 with integrase is lentivirus-specific and promotes DNA binding. J. Biol. Chem. 280: Chen, H., and A. Engelman Asymmetric processing of human immunodeficiency virus type 1 cdna in vivo: implications for functional end coupling during the chemical steps of DNA transposition. Mol. Cell Biol. 21:
24 Chen, H., S. Q. Wei, and A. Engelman Multiple integrase functions are required to form the native structure of the human immunodeficiency virus type I intasome. J. Biol. Chem. 274: Cherepanov, P., A. L. Ambrosio, S. Rahman, T. Ellenberger, and A. Engelman Structural basis for the recognition between HIV-1 integrase and transcriptional coactivator p75. Proc. Natl. Acad. Sci. U S A 102: Cherepanov, P., E. Devroe, P. A. Silver, and A. Engelman Identification of an evolutionarily conserved domain in human lens epithelium-derived growth factor/transcriptional co-activator p75 (LEDGF/p75) that binds HIV-1 integrase. J. Biol. Chem. 279: Cherepanov, P., G. Maertens, P. Proost, B. Devreese, J. Van Beeumen, Y. Engelborghs, E. De Clercq, and Z. Debyser HIV-1 integrase forms stable tetramers and associates with LEDGF/p75 protein in human cells. J. Biol. Chem. 278: Cherepanov, P., Z. Y. Sun, S. Rahman, G. Maertens, G. Wagner, and A. Engelman Solution structure of the HIV-1 integrase-binding domain in LEDGF/p75. Nat. Struct. Mol. Biol. 12: Cherepanov, P., D. Surratt, J. Toelen, W. Pluymers, J. Griffith, E. De Clercq, and Z. Debyser Activity of recombinant HIV-1 integrase on mini-hiv DNA. Nucleic Acids Res. 27:
25 Chiu, R., and D. P. Grandgenett Molecular and genetic determinants of Rous sarcoma virus integrase for concerted DNA integration. J. Virol. 77: Ciuffi, A., M. Llano, E. Poeschla, C. Hoffmann, J. Leipzig, P. Shinn, J. R. Ecker, and F. Bushman A role for LEDGF/p75 in targeting HIV DNA integration. Nat. Med. 11: Crise, B., Y. Li, C. Yuan, D. R. Morcock, D. Whitby, D. J. Munroe, L. O. Arthur, and X. Wu Simian immunodeficiency virus integration preference is similar to that of human immunodeficiency virus type 1. J. Virol. 79: Emiliani, S., A. Mousnier, K. Busschots, M. Maroun, B. Van Maele, D. Tempe, L. Vandekerckhove, F. Moisant, L. Ben-Slama, M. Witvrouw, F. Christ, J. C. Rain, C. Dargemont, Z. Debyser, and R. Benarous Integrase Mutants Defective for Interaction with LEDGF/p75 Are Impaired in Chromosome Tethering and HIV-1 Replication. J. Biol. Chem. 280: Engelman, A The ups and downs of gene expression and retroviral DNA integration. Proc. Natl. Acad. Sci. U S A 102: Ge, H., Y. Si, and R. G. Roeder Isolation of cdnas encoding novel transcription coactivators p52 and p75 reveals an alternate regulatory mechanism of transcriptional activation. Embo J. 17:
26 Goodarzi, G., R. Chiu, K. Brackmann, K. Kohn, Y. Pommier, and D. P. Grandgenett Host site selection for concerted integration by human immunodeficiency virus type-1 virions in vitro. Virology 231: Goodarzi, G., G. J. Im, K. Brackmann, and D. Grandgenett Concerted integration of retrovirus-like DNA by human immunodeficiency virus type 1 integrase. J. Virol. 69: Goodarzi, G., M. Pursley, P. Felock, M. Witmer, D. Hazuda, K. Brackmann, and D. Grandgenett Efficiency and fidelity of full-site integration reactions using recombinant simian immunodeficiency virus integrase. J. Virol. 73: Grandgenett, D. P., R. B. Inman, A. C. Vora, and M. L. Fitzgerald Comparison of DNA binding and integration half-site selection by avian myeloblastosis virus integrase. J. Virol. 67: Hazuda, D. J., P. Felock, M. Witmer, A. Wolfe, K. Stillmock, J. A. Grobler, A. Espeseth, L. Gabryelski, W. Schleif, C. Blau, and M. D. Miller Inhibitors of strand transfer that prevent integration and inhibit HIV-1 replication in cells. Science 287: Hindmarsh, P., T. Ridky, R. Reeves, M. Andrake, A. M. Skalka, and J. Leis HMG protein family members stimulate human immunodeficiency virus type 1 and avian sarcoma virus concerted DNA integration in vitro. J. Virol. 73:
27 Johnson, A. A., W. Santos, G. C. Pais, C. Marchand, R. Amin, T. R. Burke, Jr., G. Verdine, and Y. Pommier HIV-1 integration requires the viral DNA end (5'-C) interaction with glutamine 148 of the integrase flexible loop. J. Biol. Chem. 281: Lewinski, M. K., and F. D. Bushman Retroviral DNA integration--mechanism and consequences. Adv. Genet. 55: Li, M., and R. Craigie Processing of viral DNA ends channels the HIV-1 integration reaction to concerted integration. J. Biol. Chem. 280: Li, M., M. Mizuuchi, T. R. Burke, Jr., and R. Craigie Retroviral DNA integration: reaction pathway and critical intermediates. EMBO J. 25: Llano, M., S. Delgado, M. Vanegas, and E. M. Poeschla Lens epithelium-derived growth factor/p75 prevents proteasomal degradation of HIV-1 integrase. J. Biol. Chem. 279: Llano, M., D. T. Saenz, A. Meehan, P. Wongthida, M. Peretz, W. H. Walker, W. Teo, and E. M. Poeschla An Essential Role for LEDGF/p75 in HIV Integration. Science 314: Llano, M., M. Vanegas, O. Fregoso, D. Saenz, S. Chung, M. Peretz, and E. M. Poeschla LEDGF/p75 determines cellular trafficking of diverse lentiviral but not murine oncoretroviral integrase proteins and is a component of functional lentiviral preintegration complexes. J. Virol. 78:
28 Maertens, G., P. Cherepanov, Z. Debyser, Y. Engelborghs, and A. Engelman Identification and characterization of a functional nuclear localization signal in the HIV-1 integrase interactor LEDGF/p75. J. Biol. Chem. 279: Maertens, G., P. Cherepanov, W. Pluymers, K. Busschots, E. De Clercq, Z. Debyser, and Y. Engelborghs LEDGF/p75 is essential for nuclear and chromosomal targeting of HIV-1 integrase in human cells. J. Biol. Chem. 278: Miller, M. D., C. M. Farnet, and F. D. Bushman Human immunodeficiency virus type 1 preintegration complexes: studies of organization and composition. J. Virol. 71: Mitchell, R. S., B. F. Beitzel, A. R. Schroder, P. Shinn, H. Chen, C. C. Berry, J. R. Ecker, and F. D. Bushman Retroviral DNA integration: ASLV, HIV, and MLV show distinct target site preferences. PLoS Biol. 2:E Moreau, K., C. Faure, G. Verdier, and C. Ronfort Analysis of conserved and non- conserved amino acids critical for ALSV (Avian leukemia and sarcoma viruses) integrase functions in vitro. Arch. Virol. 147: Narezkina, A., K. D. Taganov, S. Litwin, R. Stoyanova, J. Hayashi, C. Seeger, A. M. Skalka, and R. A. Katz Genome-wide analyses of avian sarcoma virus integration sites. J. Virol. 78: Schroder, A. R., P. Shinn, H. Chen, C. Berry, J. R. Ecker, and F. Bushman HIV- 1 integration in the human genome favors active genes and local hotspots. Cell 110:
29 Sinha, S., and D. Grandgenett Recombinant HIV-1 integrase exhibits a capacity for full-site integration in vitro that is comparable to that of purified preintegraton complexes from virus-infected cells. J. Virol. 79: Sinha, S., M. H. Pursley, and D. P. Grandgenett Efficient concerted integration by recombinant human immunodeficiency virus type 1 integrase without cellular or viral cofactors. J. Virol. 76: Turlure, F., E. Devroe, P. A. Silver, and A. Engelman Human cell proteins and human immunodeficiency virus DNA integration. Front. Biosci. 9: Turlure, F., G. Maertens, S. Rahman, P. Cherepanov, and A. Engelman A tripartite DNA-binding element, comprised of the nuclear localization signal and two AT- hook motifs, mediates the association of LEDGF/p75 with chromatin in vivo. Nucleic Acids Res. 34: Van Maele, B., and Z. Debyser HIV-1 integration: an interplay between HIV-1 integrase, cellular and viral proteins. AIDS Rev. 7: Vandegraaff, N., E. Devroe, F. Turlure, P. A. Silver, and A. Engelman Biochemical and genetic analyses of integrase-interacting proteins lens epithelium-derived growth factor (LEDGF)/p75 and hepatoma-derived growth factor related protein 2 (HRP2) in preintegration complex function and HIV-1 replication. Virology 80: Vandekerckhove, L., F. Christ, B. Van Maele, J. De Rijck, R. Gijsbers, C. Van den Haute, M. Witvrouw, and Z. Debyser Transient and stable knockdown of the 29
30 integrase cofactor LEDGF/p75 reveals its role in the replication cycle of human immunodeficiency virus. J. Virol. 80: Vanegas, M., M. Llano, S. Delgado, D. Thompson, M. Peretz, and E. Poeschla Identification of the LEDGF/p75 HIV-1 integrase-interaction domain and NLS reveals NLS- independent chromatin tethering. J. Cell Sci. 118: Verma, I. M., and M. D. Weitzman Gene Therapy: Twenty-First Century Medicine. Annu. Rev. Biochem. 74: Vora, A., S. Bera, and D. Grandgenett Structural organization of avian retrovirus integrase in assembled intasomes mediating full-site integration. J. Biol. Chem. 279: Vora, A., and D. P. Grandgenett DNase protection analysis of retrovirus integrase at the viral DNA ends for full-site integration in vitro. J. Virol. 75: Vora, A. C., and D. P. Grandgenett Assembly and catalytic properties of retrovirus integrase-dna complexes capable of efficiently performing concerted integration. J. Virol. 69: Wei, S. Q., K. Mizuuchi, and R. Craigie A large nucleoprotein assembly at the ends of the viral DNA mediates retroviral DNA integration. Embo J. 16: Wu, X., Y. Li, B. Crise, and S. M. Burgess Transcription start regions in the human genome are favored targets for MLV integration. Science 300:
31 Zielske, S. P., and M. Stevenson Modest but Reproducible Inhibition of Human Immunodeficiency Virus Type 1 Infection in Macrophages following LEDGFp75 Silencing. J. Virol. 80: FIGURE LEGENDS FIG. 1. Assembly of HIV-1 synaptic complexes and the production of DNA integration products. A linear donor containing U3 and U5 ends is assembled with IN at 14 C to form a synaptic complex (top). Circular target DNA is added to allow strand transfer activity at 37 C. The strand transfer complex (STC) produces the full-site integration product. Other products produced by different IN-DNA complexes include circular half-site (CHS) and donor-donor products equal to twice the length of the original donor and other Y-type structures. The arrows indicate the different order of addition of LEDGF. At the bottom, different substrates having U5- U3 LTR ends or only U5 LTR ends used in this study are shown. FIG. 2. LEDGF differentially affects full-site and CHS integration. (A) Synaptic complexes were assembled with IN (5 nm) and the 4.1-kbp donor (0.2 nm) possessing U3 and U5 ends with 5 TA overhangs. LEDGF was added at the indicated final concentrations and incubated for an additional 15 min at 14 C. Strand transfer was initiated by the addition of target and incubation for 2 h at 37 C. All lanes of Fig. 2A are identical to 2B except for the order of addition shown at the top. P is for products. The DNA products and input donor are indicated on the left. Lanes 1 and 2 contain a 1 kbp DNA ladder (Promega) and donor only, respectively. Lane 3 contains IN only; it incorporated ~13% and 4% of the donor into full-site (FS) and circular half-site (CHS) 31
32 products, respectively. The full-site products in lanes 4 to 6 in Fig. 2A were ~30%, 16%, and 12%, respectively. The CHS products in lane 9 were ~22%. (B) Identical conditions were employed as in A except IN and LEDGF were preincubated together prior to the addition of donor (top). The CHS products in lane 9 were 15%. The experiments in A and B were performed in a simultaneous manner. FIG. 3. LEDGF stimulates HIV-1 synaptic complexes containing a donor with a natural U5 blunt-end. (A) Synaptic complexes were assembled with a 1.6 kbp substrate containing a single U5 blunt-end (0.52 nm) and IN (10 nm) in 125 mm NaCl at 14 C. LEDGF was added at the indicated concentrations (top) and incubated for an additional 15 min. Strand transfer was initiated by the addition of target and incubation for 2 h at 37 C. The DNA products are indicated on the left. Lane 1 contains Promega 1 kbp markers and lane 2 contains donor only. Two controls with IN only (lanes 3 and 10) incorporated 5% and 1% of the donor into full-site and CHS products, respectively. The % donor incorporated into full-site products at 2.5 and 5 nm LEDGF (lanes 4 and 5) were 11% and 8%, respectively. Three similar independent experiments averaged an ~2.3-fold increase of full-site products at these LEDGF concentrations. A 3.2 kbp donor-donor product representing twice the size of the input donor was also produced (Fig. 1). The donor-donor products in lanes 4 and 5 were 6% and 2%, respectively. (B) Identical conditions were employed as in A except IN and LEDGF were preincubated together prior to the addition of donor. The experiments in A and B were performed in a simultaneous manner FIG. 4. High concentrations of LEDGF disrupt synaptic complex formation in a quantitative manner into IN-DNA complexes mediating CHS integration. (A) Synaptic complexes were assembled with a 1.5 kbp DNA substrate (0.52 nm) containing a single U5 end with 5 -AC overhang and IN (10 nm) in 125 mm NaCl at 14 C. Lane 1 contains Promega 1 kbp markers and 32
33 lane 2 contains donor only. The amount of the DNA fragment in lane 2 migrating below the 3 kbp marker in lane 1 is variable between experiments (Fig. 3 and Fig. 6) and is apparently due to non-specific interactions of the donor overhangs upon electrophoresis. LEDGF was added at the indicated final concentrations (top) and incubated for an additional 15 min. Strand transfer was initiated by adding target and incubation for 2 h at 37 C. Products are indicated on the left. (B) Graphic representation of the % donor incorporated into full-site, CHS, and 3.0 kbp donor-donor products. FIG. 5. The LEDGF-IBD affects full-site integration similarly as full-length LEDGF. Synaptic complexes were assembled with a 1.6 kbp substrate containing a single U5 blunt-end donor (0.52 nm) and IN (10 nm) in 50 mm NaCl at 14 C. IBD at the indicated final concentrations (top) were added and incubated for an additional 15 min. Strand transfer was initiated by adding target and incubation for 2 h at 37 C. Lanes 1 and 2 contain Promega 1 kbp markers and donor, respectively. Two controls with IN only (lanes 3 and 10) incorporated 16%, 7% and 7% of the donor into full-site, CHS and 3.2 kb donor-donor products, respectively. At 10 nm IBD (lane 5), IN incorporated 21%, 9%, and 9% into these same products, respectively. FIG. 6. LEDGF is more effective than LEDGF-IBD to disrupt synaptic complexes. (A) IN (10 nm) and IBD at the indicated concentrations were incubated together for 15 min in 125 mm NaCl at 14 C. Synaptic complexes were formed with the addition of the 1.5 kbp 5 AC donor (0.52 nm). Strand transfer was initiated by adding target and incubation for 2 h at 37 C. Lane 1 contains Promega 1 kbp markers and lane 2 contains donor only. Two controls with IN only (lanes 3 and 10) incorporated 8%, 8%, and 8% of the donor into full-site, CHS and 3.0 kbp donor-donor products, respectively. At 75 nm IBD (lane 9), the same products were 4%, 10% and 3%, respectively. Products are indicated on the left. (B) The assembly and assay conditions 33
34 are the same as described in A except LEDGF was used. IN incorporated 8%, 11%, and 8% of the donor into full-site, CHS, and 3.0 kbp donor-donor products, respectively (lane 3) FIG. 7. electrophoresis. Formation of STC and their analysis by native agarose and two-dimensional gel STCs were formed for 2 h at 37 C, treated with 20 mm EDTA, and the samples were subjected to electrophoresis on a native 0.7% agarose gels in TBE buffer at 4 C for 16 ½ h. STC were formed with IN at 15 nm (lane 3). The labeling for STC and CHS is indicated on the left. A 1 kbp ladder and control DNA without IN are in lanes 1 and 2, respectively. The dots ( ) on the native gel indicate that the slower migrating bands contained full-site and other products as well as free donor. The arrow indicates the direction for electrophoresis. Right Panel-An aliquot of the sample shown in lane 3 was subjected to electrophoresis on another native 0.7% agarose gel. After SDS treatment, the gel was run in the second dimension as indicated at the top with an arrow. The 0.7% agarose gels for the native and deproteinized samples are positioned on the left and bottom of the two-dimensional gel, respectively. The full-site (FS), CHS, the 3.2 kbp donor-donor/y-type structures and free donor are indicated on the deproteinized gel located at the bottom. The unknown larger size product probably represents another integration event into full-site and/or CHS products. B. STC were formed and subjected to 0.7% agarose gel electrophoresis without (-) and with (+) 1 M urea in the gel FIG. 8. STCs are resistant to challenge by LEDGF. (A) Synaptic complexes were assembled with a 1.6 kbp substrate containing a single U5 blunt-end (0.52 nm) and IN (15 nm) at 14 C. Samples were incubated at 37 C for 2h after adding target to enable the formation of STC. The samples were treated with 30 mm EDTA. In one set of samples, 50 nm LEDGF was added (+) 34
35 and samples were further incubated at 14 C at indicated times. As controls in another set, there was no LEDGF added (-). The samples were subjected to electrophoresis on a native 0.7% agarose gel for 14 h. The STC and CHS were marked on the left. (B) A portion of the above samples were deproteinized and were subjected to electrophoresis on 0.7% agarose. The FS, CHS, and donor-donor products are marked on the right. (C) An aliquot from untreated STC (lane 4) was subjected to two-dimensional gel analysis as described in Fig. 7A. (D) An aliquot from LEDGF-treated (lane 5) was also subjected to two-dimensional gel analysis. FIG. 9. Titration of LEDGF with RSV IN. Recombinant RSV IN (5 nm) was assembled with a 3.4 kbp donor (0.21 nm) that contained a single wt U3 LTR end for 30 min on ice. LEDGF was added at the final concentration (top) for an additional 10 min. Supercoiled target was added and the samples were incubated at 37 C for 20 min. The assay conditions were previously described (48) with the final NaCl concentration at 0.2 M. Lanes 1 and 10 were without LEDGF. The % donor incorporated into FS and CHS products in these two lanes averaged 31 and 3, respectively. With increasing concentrations of LEDGF (lanes 2 to 9), the % donor incorporated into FS and CHS products averaged 33% and 2.6%, respectively. Part of the sample was lost in lane 9. FIG. 10. A model for LEDGF interactions with the HIV-1 synaptic complex and STC. From left to right, HIV-1 IN is assembled with a viral DNA donor to form a synaptic complex. For simplicity, the two DNA donors have only one LTR end displayed. IN is represented as a dimer while LEDGF as a monomer. At a low molar ratio of LEDGF to an IN tetramer in a synaptic complex, LEDGF modestly enhances the production of synaptic complexes that produce full-site integration products. For host site selection in vivo, a specific low stoichiometry between LEDGF and IN is implied (29). At a high molar ratio of LEDGF to IN in solution, LEDGF prevents assembly of synaptic complexes. The STC is stable upon challenge by high LEDGF 35
36 concentration. The association and dissociation constants for LEDGF binding to IN within the synaptic complex in vitro are unknown but appear to be in the low nm range. TABLE 1. Summary of LEDGF stimulation of HIV-1 IN mediated full-site integration. Donor type Fold Stimulation a P value b Molar Ratio c (LEDGF:IN) Preprocessed donors 4.1 kbp 5 -TA (U5-U3) 1.33 ± 0.15 < kbp 5 -AC (U5) 1.26 ± 0.07 < Blunt ended Donor 1.6 kbp DNA 5 -AC (U5) 2.35 ± 0.32 < a LEDGF stimulated HIV-1 full-site integration with different preprocessed 3 OH recessed and blunt-ended substrates. b P values for fold stimulation by LEDGF were determined using ANOVA one factor test. c LEDGF concentration was determined as a monomer while IN was a tetramer. 36
37
38
39
40

41
42
HIV-1 Integrase Strand Transfer Inhibitors: Novel Insights into their Mechanism of Action
 CMMETARY SPECIAL ISSUE HIV-1 Integrase Strand Transfer Inhibitors: ovel Insights into their Mechanism of Action Krishan K. Pandey and Duane P. Grandgenett Institute for Molecular Virology, Saint Louis
CMMETARY SPECIAL ISSUE HIV-1 Integrase Strand Transfer Inhibitors: ovel Insights into their Mechanism of Action Krishan K. Pandey and Duane P. Grandgenett Institute for Molecular Virology, Saint Louis
Recombinant Protein Expression Retroviral system
 Recombinant Protein Expression Retroviral system Viruses Contains genome DNA or RNA Genome encased in a protein coat or capsid. Some viruses have membrane covering protein coat enveloped virus Ø Essential
Recombinant Protein Expression Retroviral system Viruses Contains genome DNA or RNA Genome encased in a protein coat or capsid. Some viruses have membrane covering protein coat enveloped virus Ø Essential
The HIV life cycle. integration. virus production. entry. transcription. reverse transcription. nuclear import
 The HIV life cycle entry reverse transcription transcription integration virus production nuclear import Hazuda 2012 Integration Insertion of the viral DNA into host chromosomal DNA, essential step in
The HIV life cycle entry reverse transcription transcription integration virus production nuclear import Hazuda 2012 Integration Insertion of the viral DNA into host chromosomal DNA, essential step in
Outer domains of integrase within retroviral intasomes are dispensible for catalysis of DNA integration
 Outer domains of integrase within retroviral intasomes are dispensible for catalysis of DNA integration Min Li, Shiqiang Lin, and Robert Craigie* Laboratory of Molecular Biology, National Institute of
Outer domains of integrase within retroviral intasomes are dispensible for catalysis of DNA integration Min Li, Shiqiang Lin, and Robert Craigie* Laboratory of Molecular Biology, National Institute of
Intasome architecture and chromatin density modulate retroviral integration into nucleosome
 Benleulmi et al. Retrovirology (2015) 12:13 DOI 10.1186/s12977-015-0145-9 RESEARCH Open Access Intasome architecture and chromatin density modulate retroviral integration into nucleosome Mohamed Salah
Benleulmi et al. Retrovirology (2015) 12:13 DOI 10.1186/s12977-015-0145-9 RESEARCH Open Access Intasome architecture and chromatin density modulate retroviral integration into nucleosome Mohamed Salah
The Lentiviral Integrase Binding Protein LEDGF/p75 and HIV-1 Replication
 Review The Lentiviral Integrase Binding Protein LEDGF/p75 and HIV-1 Replication Alan Engelman 1 *, Peter Cherepanov 2 * 1 Department of Cancer Immunology and AIDS, Dana-Farber Cancer Institute, Division
Review The Lentiviral Integrase Binding Protein LEDGF/p75 and HIV-1 Replication Alan Engelman 1 *, Peter Cherepanov 2 * 1 Department of Cancer Immunology and AIDS, Dana-Farber Cancer Institute, Division
Modeling the HIV-1 Intasome: A Prototype View of the Target of Integrase Inhibitors
 Viruses 2010, 2, 2777-2781; doi:10.3390/v2122777 OPEN ACCESS viruses ISSN 1999-4915 www.mdpi.com/journal/viruses Commentary Modeling the HIV-1 Intasome: A Prototype View of the Target of Integrase Inhibitors
Viruses 2010, 2, 2777-2781; doi:10.3390/v2122777 OPEN ACCESS viruses ISSN 1999-4915 www.mdpi.com/journal/viruses Commentary Modeling the HIV-1 Intasome: A Prototype View of the Target of Integrase Inhibitors
VIROLOGY. Engineering Viral Genomes: Retrovirus Vectors
 VIROLOGY Engineering Viral Genomes: Retrovirus Vectors Viral vectors Retrovirus replicative cycle Most mammalian retroviruses use trna PRO, trna Lys3, trna Lys1,2 The partially unfolded trna is annealed
VIROLOGY Engineering Viral Genomes: Retrovirus Vectors Viral vectors Retrovirus replicative cycle Most mammalian retroviruses use trna PRO, trna Lys3, trna Lys1,2 The partially unfolded trna is annealed
The Interaction of LEDGF/p75 with Integrase Is Lentivirus-specific and Promotes DNA Binding*
 THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 280, No. 18, Issue of May 6, pp. 17841 17847, 2005 2005 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. The Interaction of
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 280, No. 18, Issue of May 6, pp. 17841 17847, 2005 2005 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. The Interaction of
7.012 Quiz 3 Answers
 MIT Biology Department 7.012: Introductory Biology - Fall 2004 Instructors: Professor Eric Lander, Professor Robert A. Weinberg, Dr. Claudette Gardel Friday 11/12/04 7.012 Quiz 3 Answers A > 85 B 72-84
MIT Biology Department 7.012: Introductory Biology - Fall 2004 Instructors: Professor Eric Lander, Professor Robert A. Weinberg, Dr. Claudette Gardel Friday 11/12/04 7.012 Quiz 3 Answers A > 85 B 72-84
Introduction retroposon
 17.1 - Introduction A retrovirus is an RNA virus able to convert its sequence into DNA by reverse transcription A retroposon (retrotransposon) is a transposon that mobilizes via an RNA form; the DNA element
17.1 - Introduction A retrovirus is an RNA virus able to convert its sequence into DNA by reverse transcription A retroposon (retrotransposon) is a transposon that mobilizes via an RNA form; the DNA element
Use of double- stranded DNA mini- circles to characterize the covalent topoisomerase- DNA complex
 SUPPLEMENTARY DATA Use of double- stranded DNA mini- circles to characterize the covalent topoisomerase- DNA complex Armêl Millet 1, François Strauss 1 and Emmanuelle Delagoutte 1 1 Structure et Instabilité
SUPPLEMENTARY DATA Use of double- stranded DNA mini- circles to characterize the covalent topoisomerase- DNA complex Armêl Millet 1, François Strauss 1 and Emmanuelle Delagoutte 1 1 Structure et Instabilité
Structural vs. nonstructural proteins
 Why would you want to study proteins associated with viruses or virus infection? Receptors Mechanism of uncoating How is gene expression carried out, exclusively by viral enzymes? Gene expression phases?
Why would you want to study proteins associated with viruses or virus infection? Receptors Mechanism of uncoating How is gene expression carried out, exclusively by viral enzymes? Gene expression phases?
Retroviruses. ---The name retrovirus comes from the enzyme, reverse transcriptase.
 Retroviruses ---The name retrovirus comes from the enzyme, reverse transcriptase. ---Reverse transcriptase (RT) converts the RNA genome present in the virus particle into DNA. ---RT discovered in 1970.
Retroviruses ---The name retrovirus comes from the enzyme, reverse transcriptase. ---Reverse transcriptase (RT) converts the RNA genome present in the virus particle into DNA. ---RT discovered in 1970.
Supplemental Materials and Methods Plasmids and viruses Quantitative Reverse Transcription PCR Generation of molecular standard for quantitative PCR
 Supplemental Materials and Methods Plasmids and viruses To generate pseudotyped viruses, the previously described recombinant plasmids pnl4-3-δnef-gfp or pnl4-3-δ6-drgfp and a vector expressing HIV-1 X4
Supplemental Materials and Methods Plasmids and viruses To generate pseudotyped viruses, the previously described recombinant plasmids pnl4-3-δnef-gfp or pnl4-3-δ6-drgfp and a vector expressing HIV-1 X4
For the 5 GATC-overhang two-oligo adaptors set up the following reactions in 96-well plate format:
 Supplementary Protocol 1. Adaptor preparation: For the 5 GATC-overhang two-oligo adaptors set up the following reactions in 96-well plate format: Per reaction X96 10X NEBuffer 2 10 µl 10 µl x 96 5 -GATC
Supplementary Protocol 1. Adaptor preparation: For the 5 GATC-overhang two-oligo adaptors set up the following reactions in 96-well plate format: Per reaction X96 10X NEBuffer 2 10 µl 10 µl x 96 5 -GATC
Sequences in the 5 and 3 R Elements of Human Immunodeficiency Virus Type 1 Critical for Efficient Reverse Transcription
 JOURNAL OF VIROLOGY, Sept. 2000, p. 8324 8334 Vol. 74, No. 18 0022-538X/00/$04.00 0 Copyright 2000, American Society for Microbiology. All Rights Reserved. Sequences in the 5 and 3 R Elements of Human
JOURNAL OF VIROLOGY, Sept. 2000, p. 8324 8334 Vol. 74, No. 18 0022-538X/00/$04.00 0 Copyright 2000, American Society for Microbiology. All Rights Reserved. Sequences in the 5 and 3 R Elements of Human
Supplementary Material
 Supplementary Material Nuclear import of purified HIV-1 Integrase. Integrase remains associated to the RTC throughout the infection process until provirus integration occurs and is therefore one likely
Supplementary Material Nuclear import of purified HIV-1 Integrase. Integrase remains associated to the RTC throughout the infection process until provirus integration occurs and is therefore one likely
Certificate of Analysis
 Certificate of Analysis Catalog No. Amount Lot Number 631987 10 μg Specified on product label. Product Information plvx-ef1α-ires-mcherry is a bicistronic lentiviral expression vector that can be used
Certificate of Analysis Catalog No. Amount Lot Number 631987 10 μg Specified on product label. Product Information plvx-ef1α-ires-mcherry is a bicistronic lentiviral expression vector that can be used
Viral Vectors In The Research Laboratory: Just How Safe Are They? Dawn P. Wooley, Ph.D., SM(NRM), RBP, CBSP
, RBP, CBSP") Viral Vectors In The Research Laboratory: Just How Safe Are They? Dawn P. Wooley, Ph.D., SM(NRM), RBP, CBSP 1 Learning Objectives Recognize hazards associated with viral vectors in research and animal
Viral Vectors In The Research Laboratory: Just How Safe Are They? Dawn P. Wooley, Ph.D., SM(NRM), RBP, CBSP 1 Learning Objectives Recognize hazards associated with viral vectors in research and animal
Jumpstart your research with ViraPower Lentiviral Expression Systems
 ViraPower Lentiviral Expression Systems Jumpstart your research with ViraPower Lentiviral Expression Systems With ViraPower Lentiviral Systems you can: Efficiently transduce both dividing and non-dividing
ViraPower Lentiviral Expression Systems Jumpstart your research with ViraPower Lentiviral Expression Systems With ViraPower Lentiviral Systems you can: Efficiently transduce both dividing and non-dividing
The Barrier-to-Autointegration Factor Is a Component of Functional Human Immunodeficiency Virus Type 1 Preintegration Complexes
 JOURNAL OF VIROLOGY, Apr. 2003, p. 5030 5036 Vol. 77, No. 8 0022-538X/03/$08.00 0 DOI: 10.1128/JVI.77.8.5030 5036.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved. The Barrier-to-Autointegration
JOURNAL OF VIROLOGY, Apr. 2003, p. 5030 5036 Vol. 77, No. 8 0022-538X/03/$08.00 0 DOI: 10.1128/JVI.77.8.5030 5036.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved. The Barrier-to-Autointegration
Engineered Hyperactive Integrase for Concerted HIV-1 DNA Integration
 Engineered Hyperactive Integrase for Concerted HIV-1 DNA Integration The Harvard community has made this article openly available. Please share how this access benefits you. Your story matters. Citation
Engineered Hyperactive Integrase for Concerted HIV-1 DNA Integration The Harvard community has made this article openly available. Please share how this access benefits you. Your story matters. Citation
Fayth K. Yoshimura, Ph.D. September 7, of 7 HIV - BASIC PROPERTIES
 1 of 7 I. Viral Origin. A. Retrovirus - animal lentiviruses. HIV - BASIC PROPERTIES 1. HIV is a member of the Retrovirus family and more specifically it is a member of the Lentivirus genus of this family.
1 of 7 I. Viral Origin. A. Retrovirus - animal lentiviruses. HIV - BASIC PROPERTIES 1. HIV is a member of the Retrovirus family and more specifically it is a member of the Lentivirus genus of this family.
Lecture 2: Virology. I. Background
 Lecture 2: Virology I. Background A. Properties 1. Simple biological systems a. Aggregates of nucleic acids and protein 2. Non-living a. Cannot reproduce or carry out metabolic activities outside of a
Lecture 2: Virology I. Background A. Properties 1. Simple biological systems a. Aggregates of nucleic acids and protein 2. Non-living a. Cannot reproduce or carry out metabolic activities outside of a
Characterization of HIV-1 integrase N-terminal mutant viruses
 Virology 360 (2007) 129 135 www.elsevier.com/locate/yviro Characterization of HIV-1 integrase N-terminal mutant viruses Aliza G. Lloyd, Yen Shing Ng, Mark A. Muesing, Viviana Simon 1, Lubbertus C.F. Mulder,1
Virology 360 (2007) 129 135 www.elsevier.com/locate/yviro Characterization of HIV-1 integrase N-terminal mutant viruses Aliza G. Lloyd, Yen Shing Ng, Mark A. Muesing, Viviana Simon 1, Lubbertus C.F. Mulder,1
Supplementary Information. Supplementary Figure 1
 Supplementary Information Supplementary Figure 1 1 Supplementary Figure 1. Functional assay of the hcas9-2a-mcherry construct (a) Gene correction of a mutant EGFP reporter cell line mediated by hcas9 or
Supplementary Information Supplementary Figure 1 1 Supplementary Figure 1. Functional assay of the hcas9-2a-mcherry construct (a) Gene correction of a mutant EGFP reporter cell line mediated by hcas9 or
Functional coupling between HIV-1 integrase and the SWI/SNF chromatin remodeling complex for efficient in vitro integration into stable nucleosomes.
 Functional coupling between HIV-1 integrase and the SWI/SNF chromatin remodeling complex for efficient in vitro integration into stable nucleosomes. Paul Lesbats, Yair Botbol, Guillaume Chevereau, Cédric
Functional coupling between HIV-1 integrase and the SWI/SNF chromatin remodeling complex for efficient in vitro integration into stable nucleosomes. Paul Lesbats, Yair Botbol, Guillaume Chevereau, Cédric
STRUCTURAL STUDIES OF THE HIV-1 PRE-INTEGRATION COMPLEX IN/LEDGF INTERACTION INHIBITORS
 STRUCTURAL STUDIES OF THE HIV-1 PRE-INTEGRATION COMPLEX IN/LEDGF INTERACTION INHIBITORS Marc Ruff Laboratory of Integrated structural biology IGBMC, Illkirch, France ruff@igbmc.fr Schematic diagram of
STRUCTURAL STUDIES OF THE HIV-1 PRE-INTEGRATION COMPLEX IN/LEDGF INTERACTION INHIBITORS Marc Ruff Laboratory of Integrated structural biology IGBMC, Illkirch, France ruff@igbmc.fr Schematic diagram of
October 26, Lecture Readings. Vesicular Trafficking, Secretory Pathway, HIV Assembly and Exit from Cell
 October 26, 2006 Vesicular Trafficking, Secretory Pathway, HIV Assembly and Exit from Cell 1. Secretory pathway a. Formation of coated vesicles b. SNAREs and vesicle targeting 2. Membrane fusion a. SNAREs
October 26, 2006 Vesicular Trafficking, Secretory Pathway, HIV Assembly and Exit from Cell 1. Secretory pathway a. Formation of coated vesicles b. SNAREs and vesicle targeting 2. Membrane fusion a. SNAREs
Mechanisms of alternative splicing regulation
 Mechanisms of alternative splicing regulation The number of mechanisms that are known to be involved in splicing regulation approximates the number of splicing decisions that have been analyzed in detail.
Mechanisms of alternative splicing regulation The number of mechanisms that are known to be involved in splicing regulation approximates the number of splicing decisions that have been analyzed in detail.
VIRUSES AND CANCER Michael Lea
 VIRUSES AND CANCER 2010 Michael Lea VIRAL ONCOLOGY - LECTURE OUTLINE 1. Historical Review 2. Viruses Associated with Cancer 3. RNA Tumor Viruses 4. DNA Tumor Viruses HISTORICAL REVIEW Historical Review
VIRUSES AND CANCER 2010 Michael Lea VIRAL ONCOLOGY - LECTURE OUTLINE 1. Historical Review 2. Viruses Associated with Cancer 3. RNA Tumor Viruses 4. DNA Tumor Viruses HISTORICAL REVIEW Historical Review
Retro-X qrt-pcr Titration Kit User Manual
 Takara Bio USA Retro-X qrt-pcr Titration Kit User Manual Cat. No. 631453 PT3952-1 (030218) 1290 Terra Bella Avenue, Mountain View, CA 94043, USA U.S. Technical Support: techus@takarabio.com United States/Canada
Takara Bio USA Retro-X qrt-pcr Titration Kit User Manual Cat. No. 631453 PT3952-1 (030218) 1290 Terra Bella Avenue, Mountain View, CA 94043, USA U.S. Technical Support: techus@takarabio.com United States/Canada
Substrate variations that affect the nucleic acid clamp activity of reverse transcriptases
 Substrate variations that affect the nucleic acid clamp activity of reverse transcriptases Iris Oz-Gleenberg, Eytan Herzig, Nickolay Voronin and Amnon Hizi* Department of Cell and Developmental Biology,
Substrate variations that affect the nucleic acid clamp activity of reverse transcriptases Iris Oz-Gleenberg, Eytan Herzig, Nickolay Voronin and Amnon Hizi* Department of Cell and Developmental Biology,
Section 6. Junaid Malek, M.D.
 Section 6 Junaid Malek, M.D. The Golgi and gp160 gp160 transported from ER to the Golgi in coated vesicles These coated vesicles fuse to the cis portion of the Golgi and deposit their cargo in the cisternae
Section 6 Junaid Malek, M.D. The Golgi and gp160 gp160 transported from ER to the Golgi in coated vesicles These coated vesicles fuse to the cis portion of the Golgi and deposit their cargo in the cisternae
Certificate of Analysis
 Certificate of Analysis plvx-ef1α-ires-puro Vector Table of Contents Product Information... 1 Description... 2 Location of Features... 3 Additional Information... 3 Quality Control Data... 4 Catalog No.
Certificate of Analysis plvx-ef1α-ires-puro Vector Table of Contents Product Information... 1 Description... 2 Location of Features... 3 Additional Information... 3 Quality Control Data... 4 Catalog No.
Viruses. Picture from:
 Viruses Understand the structure of bacteriophages & human immunodeficiency virus (HIV) Appreciate that viruses replicate in host cells (thereby destroying them) Picture from: http://eands.caltech.edu/articles/lxvii1/viruses.html
Viruses Understand the structure of bacteriophages & human immunodeficiency virus (HIV) Appreciate that viruses replicate in host cells (thereby destroying them) Picture from: http://eands.caltech.edu/articles/lxvii1/viruses.html
Polyomaviridae. Spring
 Polyomaviridae Spring 2002 331 Antibody Prevalence for BK & JC Viruses Spring 2002 332 Polyoma Viruses General characteristics Papovaviridae: PA - papilloma; PO - polyoma; VA - vacuolating agent a. 45nm
Polyomaviridae Spring 2002 331 Antibody Prevalence for BK & JC Viruses Spring 2002 332 Polyoma Viruses General characteristics Papovaviridae: PA - papilloma; PO - polyoma; VA - vacuolating agent a. 45nm
Hepatitis B Antiviral Drug Development Multi-Marker Screening Assay
 Hepatitis B Antiviral Drug Development Multi-Marker Screening Assay Background ImQuest BioSciences has developed and qualified a single-plate method to expedite the screening of antiviral agents against
Hepatitis B Antiviral Drug Development Multi-Marker Screening Assay Background ImQuest BioSciences has developed and qualified a single-plate method to expedite the screening of antiviral agents against
JANUARY 27, 2006 VOLUME 281 NUMBER 4 JOURNAL OF BIOLOGICAL CHEMISTRY 1943
 THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 281, NO. 4, pp. 1943 1955, January 27, 2006 2006 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A. Sequence, Distance,
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 281, NO. 4, pp. 1943 1955, January 27, 2006 2006 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A. Sequence, Distance,
LESSON 4.4 WORKBOOK. How viruses make us sick: Viral Replication
 DEFINITIONS OF TERMS Eukaryotic: Non-bacterial cell type (bacteria are prokaryotes).. LESSON 4.4 WORKBOOK How viruses make us sick: Viral Replication This lesson extends the principles we learned in Unit
DEFINITIONS OF TERMS Eukaryotic: Non-bacterial cell type (bacteria are prokaryotes).. LESSON 4.4 WORKBOOK How viruses make us sick: Viral Replication This lesson extends the principles we learned in Unit
Chapter 19: Viruses. 1. Viral Structure & Reproduction. 2. Bacteriophages. 3. Animal Viruses. 4. Viroids & Prions
 Chapter 19: Viruses 1. Viral Structure & Reproduction 2. Bacteriophages 3. Animal Viruses 4. Viroids & Prions 1. Viral Structure & Reproduction Chapter Reading pp. 393-396 What exactly is a Virus? Viruses
Chapter 19: Viruses 1. Viral Structure & Reproduction 2. Bacteriophages 3. Animal Viruses 4. Viroids & Prions 1. Viral Structure & Reproduction Chapter Reading pp. 393-396 What exactly is a Virus? Viruses
Name Section Problem Set 6
 Name Section 7.012 Problem Set 6 Question 1 The viral family Orthomyxoviridae contains the influenza A, B and C viruses. These viruses have a (-)ss RNA genome surrounded by a capsid composed of lipids
Name Section 7.012 Problem Set 6 Question 1 The viral family Orthomyxoviridae contains the influenza A, B and C viruses. These viruses have a (-)ss RNA genome surrounded by a capsid composed of lipids
Life Sciences 1A Midterm Exam 2. November 13, 2006
 Name: TF: Section Time Life Sciences 1A Midterm Exam 2 November 13, 2006 Please write legibly in the space provided below each question. You may not use calculators on this exam. We prefer that you use
Name: TF: Section Time Life Sciences 1A Midterm Exam 2 November 13, 2006 Please write legibly in the space provided below each question. You may not use calculators on this exam. We prefer that you use
Watson-Crick Model of B-DNA
 Reading: Ch8; 285-290 Ch24; 963-978 Problems: Ch8 (text); 9 Ch8 (study-guide: facts); 3 Ch24 (text); 5,7,9,10,14,16 Ch24 (study-guide: applying); 1 Ch24 (study-guide: facts); 1,2,4 NEXT Reading: Ch1; 29-34
Reading: Ch8; 285-290 Ch24; 963-978 Problems: Ch8 (text); 9 Ch8 (study-guide: facts); 3 Ch24 (text); 5,7,9,10,14,16 Ch24 (study-guide: applying); 1 Ch24 (study-guide: facts); 1,2,4 NEXT Reading: Ch1; 29-34
Molecular Mechanisms by Which Human Immunodeficiency Virus Type 1 Integrase Stimulates the Early Steps of Reverse Transcription
 JOURNAL OF VIROLOGY, Sept. 2007, p. 10037 10046 Vol. 81, No. 18 0022-538X/07/$08.00 0 doi:10.1128/jvi.00519-07 Copyright 2007, American Society for Microbiology. All Rights Reserved. Molecular Mechanisms
JOURNAL OF VIROLOGY, Sept. 2007, p. 10037 10046 Vol. 81, No. 18 0022-538X/07/$08.00 0 doi:10.1128/jvi.00519-07 Copyright 2007, American Society for Microbiology. All Rights Reserved. Molecular Mechanisms
Chapter 19: Viruses. 1. Viral Structure & Reproduction. What exactly is a Virus? 11/7/ Viral Structure & Reproduction. 2.
 Chapter 19: Viruses 1. Viral Structure & Reproduction 2. Bacteriophages 3. Animal Viruses 4. Viroids & Prions 1. Viral Structure & Reproduction Chapter Reading pp. 393-396 What exactly is a Virus? Viruses
Chapter 19: Viruses 1. Viral Structure & Reproduction 2. Bacteriophages 3. Animal Viruses 4. Viroids & Prions 1. Viral Structure & Reproduction Chapter Reading pp. 393-396 What exactly is a Virus? Viruses
In vitro DNase I foot printing. In vitro DNase I footprinting was performed as described
 Supplemental Methods In vitro DNase I foot printing. In vitro DNase I footprinting was performed as described previously 1 2 using 32P-labeled 211 bp fragment from 3 HS1. Footprinting reaction mixes contained
Supplemental Methods In vitro DNase I foot printing. In vitro DNase I footprinting was performed as described previously 1 2 using 32P-labeled 211 bp fragment from 3 HS1. Footprinting reaction mixes contained
1. Identify and characterize interesting phenomena! 2. Characterization should stimulate some questions/models! 3. Combine biochemistry and genetics
 1. Identify and characterize interesting phenomena! 2. Characterization should stimulate some questions/models! 3. Combine biochemistry and genetics to gain mechanistic insight! 4. Return to step 2, as
1. Identify and characterize interesting phenomena! 2. Characterization should stimulate some questions/models! 3. Combine biochemistry and genetics to gain mechanistic insight! 4. Return to step 2, as
HIV Nuclear Entry: Clearing the Fog
 Viruses 2010, 2, 1190-1194; doi:10.3390/v2051190 OPEN ACCESS viruses ISSN 1999-4915 www.mdpi.com/journal/viruses Commentary HIV Nuclear Entry: Clearing the Fog Vaibhav B. Shah and Christopher Aiken * Department
Viruses 2010, 2, 1190-1194; doi:10.3390/v2051190 OPEN ACCESS viruses ISSN 1999-4915 www.mdpi.com/journal/viruses Commentary HIV Nuclear Entry: Clearing the Fog Vaibhav B. Shah and Christopher Aiken * Department
Hepadnaviruses: Variations on the Retrovirus Theme
 WBV21 6/27/03 11:34 PM Page 377 Hepadnaviruses: Variations on the Retrovirus Theme 21 CHAPTER The virion and the viral genome The viral replication cycle The pathogenesis of hepatitis B virus A plant hepadnavirus
WBV21 6/27/03 11:34 PM Page 377 Hepadnaviruses: Variations on the Retrovirus Theme 21 CHAPTER The virion and the viral genome The viral replication cycle The pathogenesis of hepatitis B virus A plant hepadnavirus
Supplementary Figure 1
 Supplementary Figure 1 Asymmetrical function of 5p and 3p arms of mir-181 and mir-30 families and mir-142 and mir-154. (a) Control experiments using mirna sensor vector and empty pri-mirna overexpression
Supplementary Figure 1 Asymmetrical function of 5p and 3p arms of mir-181 and mir-30 families and mir-142 and mir-154. (a) Control experiments using mirna sensor vector and empty pri-mirna overexpression
Statin inhibition of HMG-CoA reductase: a 3-dimensional view
 Atherosclerosis Supplements 4 (2003) 3/8 www.elsevier.com/locate/atherosclerosis Statin inhibition of HMG-CoA reductase: a 3-dimensional view Eva Istvan * Department of Molecular Microbiology, Howard Hughes
Atherosclerosis Supplements 4 (2003) 3/8 www.elsevier.com/locate/atherosclerosis Statin inhibition of HMG-CoA reductase: a 3-dimensional view Eva Istvan * Department of Molecular Microbiology, Howard Hughes
Nature Methods: doi: /nmeth Supplementary Figure 1
 Supplementary Figure 1 Subtiligase-catalyzed ligations with ubiquitin thioesters and 10-mer biotinylated peptides. (a) General scheme for ligations between ubiquitin thioesters and 10-mer, biotinylated
Supplementary Figure 1 Subtiligase-catalyzed ligations with ubiquitin thioesters and 10-mer biotinylated peptides. (a) General scheme for ligations between ubiquitin thioesters and 10-mer, biotinylated
Identification of Mutation(s) in. Associated with Neutralization Resistance. Miah Blomquist
 in. Associated with Neutralization Resistance. Miah Blomquist") Identification of Mutation(s) in the HIV 1 gp41 Subunit Associated with Neutralization Resistance Miah Blomquist What is HIV 1? HIV-1 is an epidemic that affects over 34 million people worldwide. HIV-1
Identification of Mutation(s) in the HIV 1 gp41 Subunit Associated with Neutralization Resistance Miah Blomquist What is HIV 1? HIV-1 is an epidemic that affects over 34 million people worldwide. HIV-1
Frequent Segregation of More-Defective Variants from a Rous Sarcoma Virus Packaging Mutant, TK15
 JOURNAL OF VIROLOGY, Oct. 1987, p. 3208-3213 0022-538X/87/103208-06$02.00/0 Copyright 1987, American Society for Microbiology Vol. 61, No. 10 Frequent Segregation of More-Defective Variants from a Rous
JOURNAL OF VIROLOGY, Oct. 1987, p. 3208-3213 0022-538X/87/103208-06$02.00/0 Copyright 1987, American Society for Microbiology Vol. 61, No. 10 Frequent Segregation of More-Defective Variants from a Rous
 CDC site UNAIDS Aids Knowledge Base http://www.cdc.gov/hiv/dhap.htm http://hivinsite.ucsf.edu/insite.jsp?page=kb National Institute of Allergy and Infectious Diseases http://www.niaid.nih.gov/default.htm
CDC site UNAIDS Aids Knowledge Base http://www.cdc.gov/hiv/dhap.htm http://hivinsite.ucsf.edu/insite.jsp?page=kb National Institute of Allergy and Infectious Diseases http://www.niaid.nih.gov/default.htm
7.012 Problem Set 6 Solutions
 Name Section 7.012 Problem Set 6 Solutions Question 1 The viral family Orthomyxoviridae contains the influenza A, B and C viruses. These viruses have a (-)ss RNA genome surrounded by a capsid composed
Name Section 7.012 Problem Set 6 Solutions Question 1 The viral family Orthomyxoviridae contains the influenza A, B and C viruses. These viruses have a (-)ss RNA genome surrounded by a capsid composed
Choosing Between Lentivirus and Adeno-associated Virus For DNA Delivery
 Choosing Between Lentivirus and Adeno-associated Virus For DNA Delivery Presenter: April 12, 2017 Ed Davis, Ph.D. Senior Application Scientist GeneCopoeia, Inc. Outline Introduction to GeneCopoeia Lentiviral
Choosing Between Lentivirus and Adeno-associated Virus For DNA Delivery Presenter: April 12, 2017 Ed Davis, Ph.D. Senior Application Scientist GeneCopoeia, Inc. Outline Introduction to GeneCopoeia Lentiviral
Human Immunodeficiency Virus
 Human Immunodeficiency Virus Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Viruses and hosts Lentivirus from Latin lentis (slow), for slow progression of disease
Human Immunodeficiency Virus Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Viruses and hosts Lentivirus from Latin lentis (slow), for slow progression of disease
Julianne Edwards. Retroviruses. Spring 2010
 Retroviruses Spring 2010 A retrovirus can simply be referred to as an infectious particle which replicates backwards even though there are many different types of retroviruses. More specifically, a retrovirus
Retroviruses Spring 2010 A retrovirus can simply be referred to as an infectious particle which replicates backwards even though there are many different types of retroviruses. More specifically, a retrovirus
Materials and Methods , The two-hybrid principle.
 The enzymatic activity of an unknown protein which cleaves the phosphodiester bond between the tyrosine residue of a viral protein and the 5 terminus of the picornavirus RNA Introduction Every day there
The enzymatic activity of an unknown protein which cleaves the phosphodiester bond between the tyrosine residue of a viral protein and the 5 terminus of the picornavirus RNA Introduction Every day there
TNPO3 is Required for HIV-1 Replication After Nuclear Import but Prior to. Integration and Binds the HIV-1 Core. and Felipe Diaz-Griffero *1
 JVI Accepts, published online ahead of print on 7 March 2012 J. Virol. doi:10.1128/jvi.00451-12 Copyright 2012, American Society for Microbiology. All Rights Reserved. 1 2 3 4 TNPO3 is Required for HIV-1
JVI Accepts, published online ahead of print on 7 March 2012 J. Virol. doi:10.1128/jvi.00451-12 Copyright 2012, American Society for Microbiology. All Rights Reserved. 1 2 3 4 TNPO3 is Required for HIV-1
Product Manual. Omni-Array Sense Strand mrna Amplification Kit, 2 ng to 100 ng Version Catalog No.: Reactions
 Genetic Tools and Reagents Universal mrna amplification, sense strand amplification, antisense amplification, cdna synthesis, micro arrays, gene expression, human, mouse, rat, guinea pig, cloning Omni-Array
Genetic Tools and Reagents Universal mrna amplification, sense strand amplification, antisense amplification, cdna synthesis, micro arrays, gene expression, human, mouse, rat, guinea pig, cloning Omni-Array
hemagglutinin and the neuraminidase genes (RNA/recombinant viruses/polyacrylamide gel electrophoresis/genetics)
") Proc. Natl. Acad. Sci. USA Vol. 73, No. 6, pp. 242-246, June 976 Microbiology Mapping of the influenza virus genome: Identification of the hemagglutinin and the neuraminidase genes (RNA/recombinant viruses/polyacrylamide
Proc. Natl. Acad. Sci. USA Vol. 73, No. 6, pp. 242-246, June 976 Microbiology Mapping of the influenza virus genome: Identification of the hemagglutinin and the neuraminidase genes (RNA/recombinant viruses/polyacrylamide
Template Dimerization Promotes an Acceptor Invasion-Induced Transfer Mechanism during Human Immunodeficiency Virus Type 1 Minus-Strand Synthesis
 JOURNAL OF VIROLOGY, Apr. 2003, p. 4710 4721 Vol. 77, No. 8 0022-538X/03/$08.00 0 DOI: 10.1128/JVI.77.8.4710 4721.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved. Template Dimerization
JOURNAL OF VIROLOGY, Apr. 2003, p. 4710 4721 Vol. 77, No. 8 0022-538X/03/$08.00 0 DOI: 10.1128/JVI.77.8.4710 4721.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved. Template Dimerization
BIL 256 Cell and Molecular Biology Lab Spring, Tissue-Specific Isoenzymes
 BIL 256 Cell and Molecular Biology Lab Spring, 2007 Background Information Tissue-Specific Isoenzymes A. BIOCHEMISTRY The basic pattern of glucose oxidation is outlined in Figure 3-1. Glucose is split
BIL 256 Cell and Molecular Biology Lab Spring, 2007 Background Information Tissue-Specific Isoenzymes A. BIOCHEMISTRY The basic pattern of glucose oxidation is outlined in Figure 3-1. Glucose is split
Under the Radar Screen: How Bugs Trick Our Immune Defenses
 Under the Radar Screen: How Bugs Trick Our Immune Defenses Session 7: Cytokines Marie-Eve Paquet and Gijsbert Grotenbreg Whitehead Institute for Biomedical Research HHV-8 Discovered in the 1980 s at the
Under the Radar Screen: How Bugs Trick Our Immune Defenses Session 7: Cytokines Marie-Eve Paquet and Gijsbert Grotenbreg Whitehead Institute for Biomedical Research HHV-8 Discovered in the 1980 s at the
Received 13 October 2003/Accepted 27 January 2004
 JOURNAL OF VIROLOGY, June 2004, p. 5835 5847 Vol. 78, No. 11 0022-538X/04/$08.00 0 DOI: 10.1128/JVI.78.11.5835 5847.2004 Copyright 2004, American Society for Microbiology. All Rights Reserved. Human Immunodeficiency
JOURNAL OF VIROLOGY, June 2004, p. 5835 5847 Vol. 78, No. 11 0022-538X/04/$08.00 0 DOI: 10.1128/JVI.78.11.5835 5847.2004 Copyright 2004, American Society for Microbiology. All Rights Reserved. Human Immunodeficiency
Fayth K. Yoshimura, Ph.D. September 7, of 7 RETROVIRUSES. 2. HTLV-II causes hairy T-cell leukemia
 1 of 7 I. Diseases Caused by Retroviruses RETROVIRUSES A. Human retroviruses that cause cancers 1. HTLV-I causes adult T-cell leukemia and tropical spastic paraparesis 2. HTLV-II causes hairy T-cell leukemia
1 of 7 I. Diseases Caused by Retroviruses RETROVIRUSES A. Human retroviruses that cause cancers 1. HTLV-I causes adult T-cell leukemia and tropical spastic paraparesis 2. HTLV-II causes hairy T-cell leukemia
L I F E S C I E N C E S
 1a L I F E S C I E N C E S 5 -UUA AUA UUC GAA AGC UGC AUC GAA AAC UGU GAA UCA-3 5 -TTA ATA TTC GAA AGC TGC ATC GAA AAC TGT GAA TCA-3 3 -AAT TAT AAG CTT TCG ACG TAG CTT TTG ACA CTT AGT-5 OCTOBER 31, 2006
1a L I F E S C I E N C E S 5 -UUA AUA UUC GAA AGC UGC AUC GAA AAC UGU GAA UCA-3 5 -TTA ATA TTC GAA AGC TGC ATC GAA AAC TGT GAA TCA-3 3 -AAT TAT AAG CTT TCG ACG TAG CTT TTG ACA CTT AGT-5 OCTOBER 31, 2006
What causes cancer? Physical factors (radiation, ionization) Chemical factors (carcinogens) Biological factors (virus, bacteria, parasite)
 Chemical factors (carcinogens) Biological factors (virus, bacteria, parasite)") Oncogenes What causes cancer? Chemical factors (carcinogens) Physical factors (radiation, ionization) Biological factors (virus, bacteria, parasite) DNA Mutation or damage Oncogenes Tumor suppressor genes
Oncogenes What causes cancer? Chemical factors (carcinogens) Physical factors (radiation, ionization) Biological factors (virus, bacteria, parasite) DNA Mutation or damage Oncogenes Tumor suppressor genes
Reverse Genetics of RNA Viruses
 Reverse Genetics of RNA Viruses Reverse Genetics (RG) he creation of a virus with a fulllength copy of the viral genome he most powerful tool in modern virology RG of RNA viruses Generation or recovery
Reverse Genetics of RNA Viruses Reverse Genetics (RG) he creation of a virus with a fulllength copy of the viral genome he most powerful tool in modern virology RG of RNA viruses Generation or recovery
AP Biology Summer Assignment Chapter 3 Quiz
 AP Biology Summer Assignment Chapter 3 Quiz 2016-17 Multiple Choice Identify the choice that best completes the statement or answers the question. 1. All of the following are found in a DNA nucleotide
AP Biology Summer Assignment Chapter 3 Quiz 2016-17 Multiple Choice Identify the choice that best completes the statement or answers the question. 1. All of the following are found in a DNA nucleotide
TRANSPORT OF AMINO ACIDS IN INTACT 3T3 AND SV3T3 CELLS. Binding Activity for Leucine in Membrane Preparations of Ehrlich Ascites Tumor Cells
 Journal of Supramolecular Structure 4:441 (401)-447 (407) (1976) TRANSPORT OF AMINO ACIDS IN INTACT 3T3 AND SV3T3 CELLS. Binding Activity for Leucine in Membrane Preparations of Ehrlich Ascites Tumor Cells
Journal of Supramolecular Structure 4:441 (401)-447 (407) (1976) TRANSPORT OF AMINO ACIDS IN INTACT 3T3 AND SV3T3 CELLS. Binding Activity for Leucine in Membrane Preparations of Ehrlich Ascites Tumor Cells
Overview: Chapter 19 Viruses: A Borrowed Life
 Overview: Chapter 19 Viruses: A Borrowed Life Viruses called bacteriophages can infect and set in motion a genetic takeover of bacteria, such as Escherichia coli Viruses lead a kind of borrowed life between
Overview: Chapter 19 Viruses: A Borrowed Life Viruses called bacteriophages can infect and set in motion a genetic takeover of bacteria, such as Escherichia coli Viruses lead a kind of borrowed life between
Overview of the Expressway Cell-Free Expression Systems. Expressway Mini Cell-Free Expression System
 Overview of the Expressway Cell-Free Expression Systems The Expressway Cell-Free Expression Systems use an efficient coupled transcription and translation reaction to produce up to milligram quantities
Overview of the Expressway Cell-Free Expression Systems The Expressway Cell-Free Expression Systems use an efficient coupled transcription and translation reaction to produce up to milligram quantities
Virology Introduction. Definitions. Introduction. Structure of virus. Virus transmission. Classification of virus. DNA Virus. RNA Virus. Treatment.
 DEVH Virology Introduction Definitions. Introduction. Structure of virus. Virus transmission. Classification of virus. DNA Virus. RNA Virus. Treatment. Definitions Virology: The science which study the
DEVH Virology Introduction Definitions. Introduction. Structure of virus. Virus transmission. Classification of virus. DNA Virus. RNA Virus. Treatment. Definitions Virology: The science which study the
Translation. Host Cell Shutoff 1) Initiation of eukaryotic translation involves many initiation factors
 Initiation of eukaryotic translation involves many initiation factors") Translation Questions? 1) How does poliovirus shutoff eukaryotic translation? 2) If eukaryotic messages are not translated how can poliovirus get its message translated? Host Cell Shutoff 1) Initiation
Translation Questions? 1) How does poliovirus shutoff eukaryotic translation? 2) If eukaryotic messages are not translated how can poliovirus get its message translated? Host Cell Shutoff 1) Initiation
Conditional and reversible disruption of essential herpesvirus protein functions
 nature methods Conditional and reversible disruption of essential herpesvirus protein functions Mandy Glaß, Andreas Busche, Karen Wagner, Martin Messerle & Eva Maria Borst Supplementary figures and text:
nature methods Conditional and reversible disruption of essential herpesvirus protein functions Mandy Glaß, Andreas Busche, Karen Wagner, Martin Messerle & Eva Maria Borst Supplementary figures and text:
The U1 snrnp Base Pairs with the 5 Splice Site within a Penta-snRNP Complex
 MOLECULAR AND CELLULAR BIOLOGY, May 2003, p. 3442 3455 Vol. 23, No. 10 0270-7306/03/$08.00 0 DOI: 10.1128/MCB.23.10.3442 3455.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved.
MOLECULAR AND CELLULAR BIOLOGY, May 2003, p. 3442 3455 Vol. 23, No. 10 0270-7306/03/$08.00 0 DOI: 10.1128/MCB.23.10.3442 3455.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved.
Tel: ; Fax: ;
 Tel.: +98 216 696 9291; Fax: +98 216 696 9291; E-mail: mrasadeghi@pasteur.ac.ir Tel: +98 916 113 7679; Fax: +98 613 333 6380; E-mail: abakhshi_e@ajums.ac.ir A Soluble Chromatin-bound MOI 0 1 5 0 1 5 HDAC2
Tel.: +98 216 696 9291; Fax: +98 216 696 9291; E-mail: mrasadeghi@pasteur.ac.ir Tel: +98 916 113 7679; Fax: +98 613 333 6380; E-mail: abakhshi_e@ajums.ac.ir A Soluble Chromatin-bound MOI 0 1 5 0 1 5 HDAC2
Review Article The Role of TNPO3 in HIV-1 Replication
 Molecular Biology International Volume 2012, Article ID 868597, 6 pages doi:10.1155/2012/868597 Review Article The Role of TNPO3 in HIV-1 Replication Felipe Diaz-Griffero Department of Microbiology and
Molecular Biology International Volume 2012, Article ID 868597, 6 pages doi:10.1155/2012/868597 Review Article The Role of TNPO3 in HIV-1 Replication Felipe Diaz-Griffero Department of Microbiology and
Department of Microbiology, School of Medicine, Box , University of Washington, Seattle, WA 98195, USA
 Virology 366 (2007) 361 376 www.elsevier.com/locate/yviro Substitution of alanine for tyrosine-64 in the fingers subdomain of M-MuLV reverse transcriptase impairs strand displacement synthesis and blocks
Virology 366 (2007) 361 376 www.elsevier.com/locate/yviro Substitution of alanine for tyrosine-64 in the fingers subdomain of M-MuLV reverse transcriptase impairs strand displacement synthesis and blocks
Your Gene ATG GGT. pd1118 EF1a-ORF, Lenti-ElecD 7803 bp
 Mammalian Expression Vectors has mammalian expression vectors suitable for transient or stable expression. These vectors are available with features including various promoters, markers, and fusions. Lentiviral
Mammalian Expression Vectors has mammalian expression vectors suitable for transient or stable expression. These vectors are available with features including various promoters, markers, and fusions. Lentiviral
Chemistry 107 Exam 4 Study Guide
 Chemistry 107 Exam 4 Study Guide Chapter 10 10.1 Recognize that enzyme catalyze reactions by lowering activation energies. Know the definition of a catalyst. Differentiate between absolute, relative and
Chemistry 107 Exam 4 Study Guide Chapter 10 10.1 Recognize that enzyme catalyze reactions by lowering activation energies. Know the definition of a catalyst. Differentiate between absolute, relative and
Use of Patient-Derived Human Immunodeficiency Virus Type 1 Integrases To Identify a Protein Residue That Affects Target Site Selection
 JOURNAL OF VIROLOGY, Aug. 2001, p. 7756 7762 Vol. 75, No. 16 0022-538X/01/$04.00 0 DOI: 10.1128/JVI.75.16.7756 7762.2001 Copyright 2001, American Society for Microbiology. All Rights Reserved. Use of Patient-Derived
JOURNAL OF VIROLOGY, Aug. 2001, p. 7756 7762 Vol. 75, No. 16 0022-538X/01/$04.00 0 DOI: 10.1128/JVI.75.16.7756 7762.2001 Copyright 2001, American Society for Microbiology. All Rights Reserved. Use of Patient-Derived
DATA SHEET. Provided: 500 µl of 5.6 mm Tris HCl, 4.4 mm Tris base, 0.05% sodium azide 0.1 mm EDTA, 5 mg/liter calf thymus DNA.
 Viral Load DNA >> Standard PCR standard 0 Copies Catalog Number: 1122 Lot Number: 150298 Release Category: A Provided: 500 µl of 5.6 mm Tris HCl, 4.4 mm Tris base, 0.05% sodium azide 0.1 mm EDTA, 5 mg/liter
Viral Load DNA >> Standard PCR standard 0 Copies Catalog Number: 1122 Lot Number: 150298 Release Category: A Provided: 500 µl of 5.6 mm Tris HCl, 4.4 mm Tris base, 0.05% sodium azide 0.1 mm EDTA, 5 mg/liter
The rabbit femoral artery was prepared and each arterial ring was permeabilized
 Online Supplement Nakmura et al. cgmp-dependent relaxation of smooth muscle Materials and Methods Measurement of tension The rabbit femoral artery was prepared and each arterial ring was permeabilized
Online Supplement Nakmura et al. cgmp-dependent relaxation of smooth muscle Materials and Methods Measurement of tension The rabbit femoral artery was prepared and each arterial ring was permeabilized
2) What is the difference between a non-enveloped virion and an enveloped virion? (4 pts)
 What is the difference between a non-enveloped virion and an enveloped virion? (4 pts)") Micro 260 SFCC Spring 2010 Name: All diagrams and drawings shall be hand drawn (do not photo-copied from a publication then cut and pasted into work sheet). Do not copy other student s answers. Para phase
Micro 260 SFCC Spring 2010 Name: All diagrams and drawings shall be hand drawn (do not photo-copied from a publication then cut and pasted into work sheet). Do not copy other student s answers. Para phase
Norgen s HIV proviral DNA PCR Kit was developed and validated to be used with the following PCR instruments: Qiagen Rotor-Gene Q BioRad icycler
 3430 Schmon Parkway Thorold, ON, Canada L2V 4Y6 Phone: (905) 227-8848 Fax: (905) 227-1061 Email: techsupport@norgenbiotek.com HIV Proviral DNA PCR Kit Product # 33840 Product Insert Background Information
3430 Schmon Parkway Thorold, ON, Canada L2V 4Y6 Phone: (905) 227-8848 Fax: (905) 227-1061 Email: techsupport@norgenbiotek.com HIV Proviral DNA PCR Kit Product # 33840 Product Insert Background Information
1) DNA unzips - hydrogen bonds between base pairs are broken by special enzymes.
 DNA unzips - hydrogen bonds between base pairs are broken by special enzymes.") Biology 12 Cell Cycle To divide, a cell must complete several important tasks: it must grow, during which it performs protein synthesis (G1 phase) replicate its genetic material /DNA (S phase), and physically
Biology 12 Cell Cycle To divide, a cell must complete several important tasks: it must grow, during which it performs protein synthesis (G1 phase) replicate its genetic material /DNA (S phase), and physically
Viruses. Rotavirus (causes stomach flu) HIV virus
 HIV virus") Viruses Rotavirus (causes stomach flu) HIV virus What is a virus? A virus is a microscopic, infectious agent that may infect any type of living cell. Viruses must infect living cells in order to make more
Viruses Rotavirus (causes stomach flu) HIV virus What is a virus? A virus is a microscopic, infectious agent that may infect any type of living cell. Viruses must infect living cells in order to make more
Norgen s HIV Proviral DNA PCR Kit was developed and validated to be used with the following PCR instruments: Qiagen Rotor-Gene Q BioRad T1000 Cycler
 3430 Schmon Parkway Thorold, ON, Canada L2V 4Y6 Phone: 866-667-4362 (905) 227-8848 Fax: (905) 227-1061 Email: techsupport@norgenbiotek.com HIV Proviral DNA PCR Kit Product# 33840 Product Insert Intended
3430 Schmon Parkway Thorold, ON, Canada L2V 4Y6 Phone: 866-667-4362 (905) 227-8848 Fax: (905) 227-1061 Email: techsupport@norgenbiotek.com HIV Proviral DNA PCR Kit Product# 33840 Product Insert Intended
Virus and Prokaryotic Gene Regulation - 1
 Virus and Prokaryotic Gene Regulation - 1 We have discussed the molecular structure of DNA and its function in DNA duplication and in transcription and protein synthesis. We now turn to how cells regulate
Virus and Prokaryotic Gene Regulation - 1 We have discussed the molecular structure of DNA and its function in DNA duplication and in transcription and protein synthesis. We now turn to how cells regulate
A complete next-generation sequencing workfl ow for circulating cell-free DNA isolation and analysis
 APPLICATION NOTE Cell-Free DNA Isolation Kit A complete next-generation sequencing workfl ow for circulating cell-free DNA isolation and analysis Abstract Circulating cell-free DNA (cfdna) has been shown
APPLICATION NOTE Cell-Free DNA Isolation Kit A complete next-generation sequencing workfl ow for circulating cell-free DNA isolation and analysis Abstract Circulating cell-free DNA (cfdna) has been shown
igem2013 Microbiology BMB SDU
 igem2013 Microbiology BMB SDU Project type: Creation of Biobrick with Prenyltransferase Project title: Biobrick_Prenyltransferase Sub project: Creation date: 13.08.09 Written by: SIS Performed by: PRA,
igem2013 Microbiology BMB SDU Project type: Creation of Biobrick with Prenyltransferase Project title: Biobrick_Prenyltransferase Sub project: Creation date: 13.08.09 Written by: SIS Performed by: PRA,
STRUCTURE, GENERAL CHARACTERISTICS AND REPRODUCTION OF VIRUSES
 STRUCTURE, GENERAL CHARACTERISTICS AND REPRODUCTION OF VIRUSES Introduction Viruses are noncellular genetic elements that use a living cell for their replication and have an extracellular state. Viruses
STRUCTURE, GENERAL CHARACTERISTICS AND REPRODUCTION OF VIRUSES Introduction Viruses are noncellular genetic elements that use a living cell for their replication and have an extracellular state. Viruses
~Lentivirus production~
 ~Lentivirus production~ May 30, 2008 RNAi core R&D group member Lentivirus Production Session Lentivirus!!! Is it health threatening to lab technician? What s so good about this RNAi library? How to produce
~Lentivirus production~ May 30, 2008 RNAi core R&D group member Lentivirus Production Session Lentivirus!!! Is it health threatening to lab technician? What s so good about this RNAi library? How to produce