Ultrastructure of the ascus apical apparatus and ascospore wall in Ombrophila hemiamyloidea (Helotiales, Ascomycota)
|
|
|
- Ruby Palmer
- 5 years ago
- Views:
Transcription
1 Nova Hedwigia Stuttgart, November 2003 Ultrastructure of the ascus apical apparatus and ascospore wall in Ombrophila hemiamyloidea (Helotiales, Ascomycota) by Gerard J.M. Verkley Centraalbureau voor Schimmelcultures, Fungal Biodiversity Center P.O. Box 85167, NL-3508 AD Utrecht, The Netherlands verkley@cbs.knaw.nl With 11 figures Verkley, G.J.M. (2003): Ultrastructure of the ascus apical apparatus and ascospore wall in Ombrophila hemiamyloidea (Helotiales, Ascomycota). Nova Hedwigia 77: Abstract: The ultrastructure of the ascus apical apparatus, lateral ascus wall and ascospore wall of Ombrophila hemiamyloidea was studied with TEM. The apical apparatus is composed of a well-developed apical thickening that surrounds an apical chamber and is fully occupied by a broad annulus. This annulus initially consists of a homogeneous matrix, but, due to changes in ultrastructure and PA-TCH-SP reactivity, two annular subregions become distinguishable as the ascus matures. These features do not correlate with the hemiamyloid properties of the annulus in Lugol s iodine solution, which imparts a distinct and homogeneous red colour to both of these subregions in mature asci. After dehiscence, the annulus is everted over an angle of about 90. The mature ascospore wall is composed of an endospore covered by an epispore. The endospore is lined by an inner wall layer which is continuous with the septal walls. Comparison with ultrastructure of the ascus and ascospores of Ombrophila violacea, the type species of the genus Ombrophila, indicates that O. hemiamyloidea does not belong in this genus. The general morphology of the apical apparatus and the structure and reactivity of the annulus agree with the Pezicula ascus type. The results indicate that O. hemiamyloidea is more closely related to Pezicula and related genera of the family Dermateaceae than to the Vibrisseaceae or the Mollisiaceae as previously been suggested. Key words: Dermateaceae, Helotiaceae, Mollisiaceae, Pezicula, Vibrisseaceae, wall structure. Introduction In fall of 1991 I collected a discomycete with relatively large, white, sessile apothecia on a piece of bare wood submerged in a forest stream. Although it showed a number of distinctive microscopic characters, I was unable to identify it at that time. The DOI: / / 2003/ / 03/ $ J. Cramer in der Gebrüder Borntraeger Verlagsbuchha ndlung, D Berlin D Stuttgart 271
2 inoperculate asci were profoundly hemiamyloid, i.e., the apical apparatus became intensely red-brown when Lugol s iodine solution (IKI) was added to water mounts (reaction type rr in the scheme of Baral 1987a). No reaction was observed in asci mounted in Melzer s reagent (Mlz), but if the asci were first treated with KOH, they blued in both IKI and Mlz. In IKI, a purplish-red to pink reaction was observed on the outer surface of the mature ascospore walls, and occasionally also in the material covering the paraphyses. Rr-type hemiamyloid asci are known in relatively few Helotiales (Baral 1987a). Of the genera known to have hemiamyloid asci, only Pezicula Tul. & C. Tul. has been studied at the ultrastructural level (Bellemère 1977). To determine if the asci of the present fungus were similar to those of Pezicula spp., I fixed some of the fresh material and studied it with transmission electron microscopy (TEM). Its identity remained unclear until recently, when the new species Ombrophila hemiamyloidea Baral & A. Gminder was published (Baral 1999). After examination of the holotyp e of O. hemiamyloidea, the identification could be confirmed. Baral (1999) pointed out that O. hemiamyloidea exhibited an exceptional combination of characters. He stated that this taxon formed a link between three families, viz. Mollisiaceae (Dermateaceae Fr., p.p.), Vibrisseaceae Korf, and Helotiaceae Rehm. He placed it in the genus Ombrophila Fr. of the family Helotiaceae mainly because of its medullary excipulum of textura intricata embedded in a gelatinous matrix with numerous crystals, as well as its hyaline ectal excipulum. Other characters of O. hemiamyloidea indicated possible affinities to members of the Mollisia-Pyrenopeziza complex (Dermateaceae), viz., the almost perfectly spherical cells in the nongelatinised ectal excipulum, and the yellow KOH reaction of the refractive vacuolar bodies in the paraphyses. Paraphyses of similar shape, and spores with similar IKI reactions in their sheaths are known in members of the mainly aquatic Vibrisseaceae (Baral 1999). Many genera in the Dermateaceae and Helotiaceae are poorly delimited due to the paucity of synapomorphic characters. Few have been revised or studied in culture, and molecular phylogenetic data for certain groups have only recently become available (Collado et al. 2002, Gernandt et al. 2001, Verkley 1999, Abeln et al. 2000, Crous et al. 2003). The taxonomy of the inoperculate discomycetes is far from settled, and mycologists have used light microscopy to study ascal morphology with refined methods as a possible source of useful characters (Chadefaud 1964, 1973, Baral 1987b, Triebel & Baral 1996) and with TEM (Bellemère 1975, 1977, Bellemère et al. 1987, Verkley 1992, 1993a and b, 1994, 1995a and b, 1996, Lenurm & Raitviir 2000, 2001). These studies have revealed a large diversity in structure and function of the ascus apical apparatus in the Helotiales. TEM analysis of the asci of O. hemiamyloidea could contribute to a better understanding of the structure of hemiamyloid asci, and also throw more light on the relationships of this fungus with other Helotiales. The results of the present study are compared with ultrastructural data available for other helotialean fungi, and the implications for the taxonomic position of O. hemiamyloidea are discussed. 272
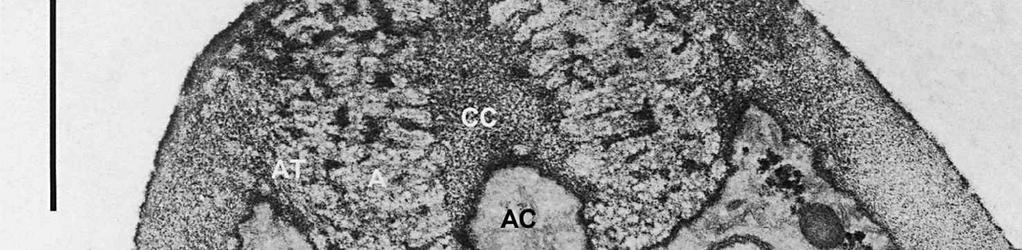
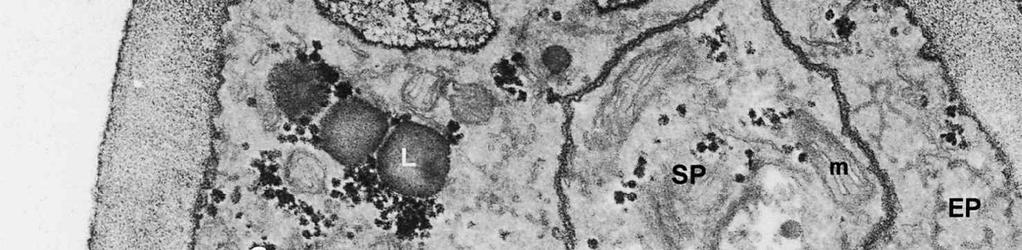
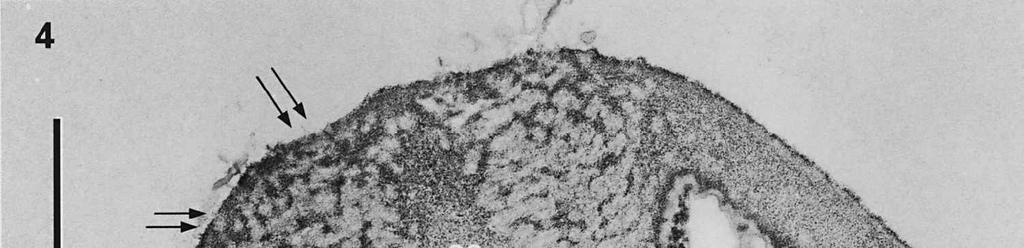
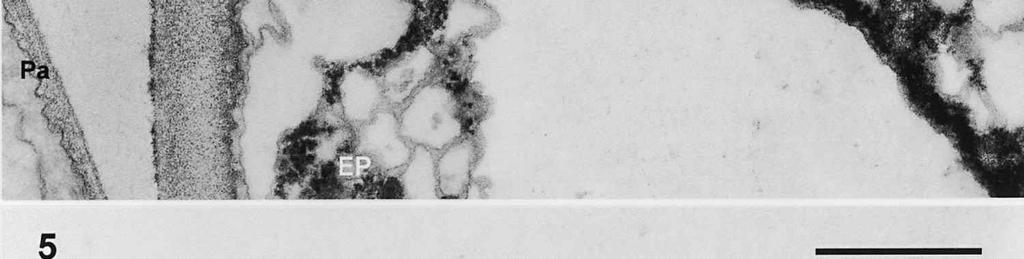
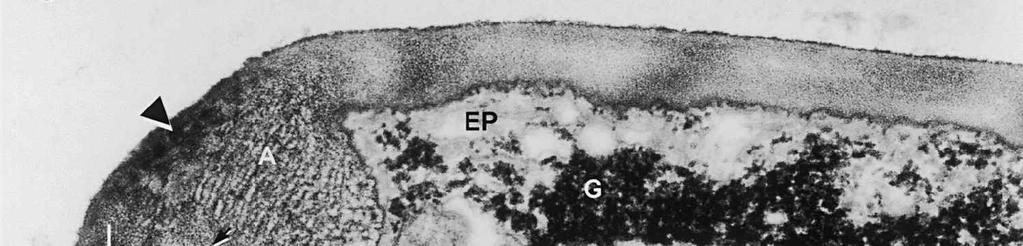
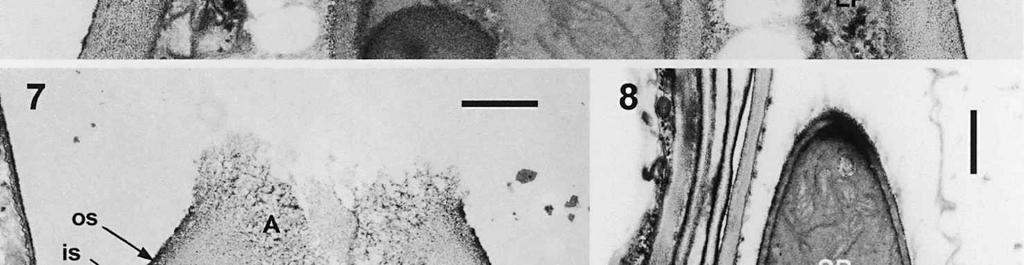
3 Material and methods MATERIAL EXAMINED. The Netherlands, prov. Gelderland, Ubbergen, Filosofendal, on bare wood of Alnus glutinosa, fully submerged in a rivulet, 28.X.1991, G. Verkley (used for TEM, vaucher in L ); Germany: Baden-Württemberg, Heidelberg, Ziegelhausen, Bärenbachtal, on Carpinus betulus, 22.XI.1997, J. Haedeke, H.B (Holotypus of O. hemiamyloidea, M). SPECIMEN PREPARATION FOR TEM. Part of the collection from the Netherlands cited above was prepared for TEM in the fresh state. The identity of the dried voucher was later confirmed by lightmicroscopic comparison with the holotype material of O. hemiamyloidea. For electron microscopy, parts of fruit-bodies were fixed in 1% glutaraldehyde in 0.1 M sodium cacodylate buffer (ph 7.2) at 4 C, washed in buffer and postfixed in similarly buffered 1% osmium tetroxide at room temperature. Material was then dehydrated in a graded series of ethanol, stained in 1% uranyl acetate for 10 min, and embedded in Epon 3/7. Ultrathin sections were cut using a Diatome diamond knife on a Reichert Jung Ultracut E ultratome. Sections were picked up on 200 mesh gold grids and treated with periodic acid (PA)-thiocarbohydrazide (TCH)-silver proteinate (SP; collectively PA-TCH-SP) as described by Verkley (1992), modified from Thiéry (1967), in short: 1% aqueous PA, 30 min; water rinse; 0.2% TCH in 20% acetic acid, 60 min; rinses in 10, 5 and 2.5% acetic acid followed by water; 1% aqueous SP (Prolabo, Paris, France), 25 min in the dark; water rinse. Shortly after this treatment material was examined with a Philips EM 300 at 60 kv. TERMINOLOGY. The terminology used for ascus structures is fully explained in Verkley (1992, 1994). Results ASCUS APICAL APPARATUS. The ascus apex is conical to conical-rounded. In the earliest stage of development observed, when the ascus is still elongating, two layers can already be distinguished in the lateral ascus wall (Fig. 1). The outer layer, nm thick, consists of two strata: a strongly PA-TCH-SP reactive outer stratum, and a less but homogeneously reactive inner stratum (Fig. 2). The inner layer is thinner ( nm) than the outer layer, and shows a weak and granular reactivity. It has no internal stratification (Fig. 1). The apical ascoplasm is rich in mitochondria, endoplasmatic reticulum, and aggregations of glycogen particles. Vesicles containing reactive material are found in close proximity to the inner wall layer in the tip of the ascus, which still lacks a differentiated apical apparatus at this stage (Fig. 1). Further towards the base, the ascoplasm contains numerous lipid bodies. During formation of the apical apparatus, vesicles are again observed close to the inner apical wall (Fig. 2). The apparatus is completed before ascospore delimitation. At this time, the inner layer of the lateral ascus wall has reached its mature thickness of nm. The inner layer increases abruptly in thickness in the apical apparatus (single short arrow, Fig. 2), where it forms a well-developed apical thickening. Here and in the lateral ascus wall, the outer layer is about equal in thickness. The apical thickening is fully occupied by the annulus and encloses a conspicuous apical chamber (Figs 3, 4). There is no annular protrusion. In the young and immature ascus, the annulus is characterized by densely packed, more or less flattened discontinuous layers of weakly PA-TCH-SP reactive, fibrillar to amorphous material. The layered material is oriented more or less parallel to the inner surface of the wall and embedded within a moderately reactive matrix (Figs 3, 4). Irregular patches of strongly reactive material are found throughout the annulus, but in some asci concentrations of such patches are also found in the part closest to the outer wall surface. The central 273
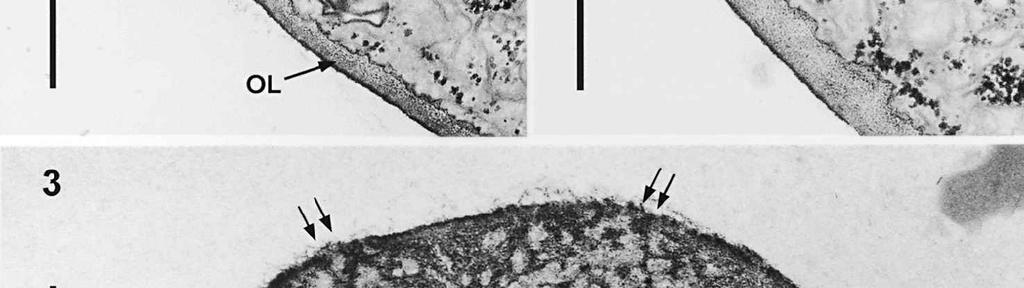
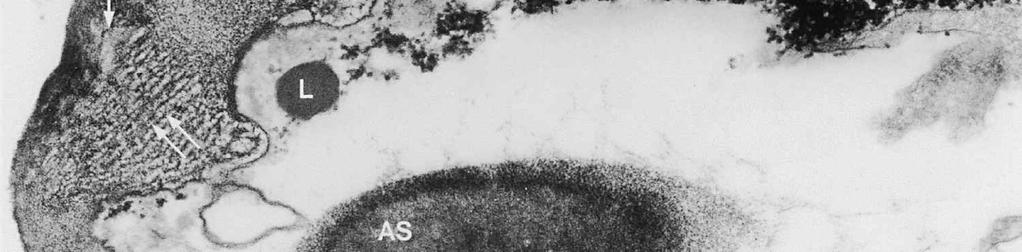
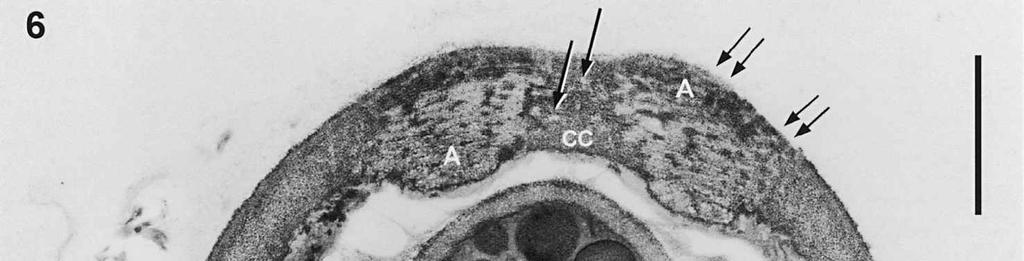
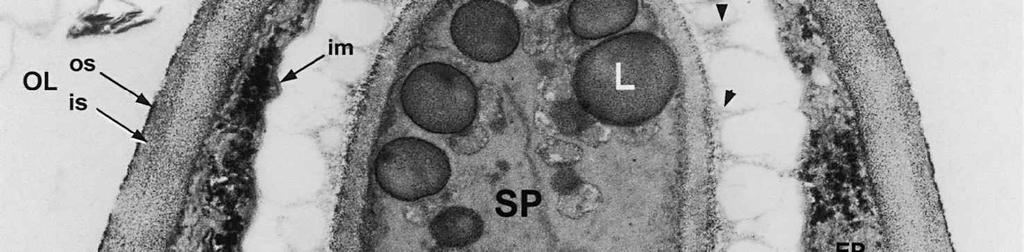
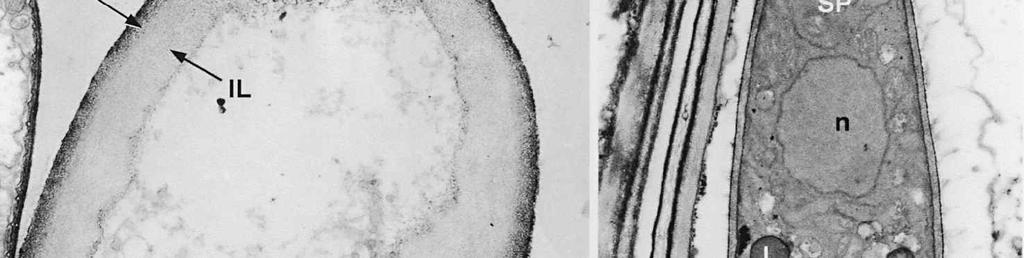
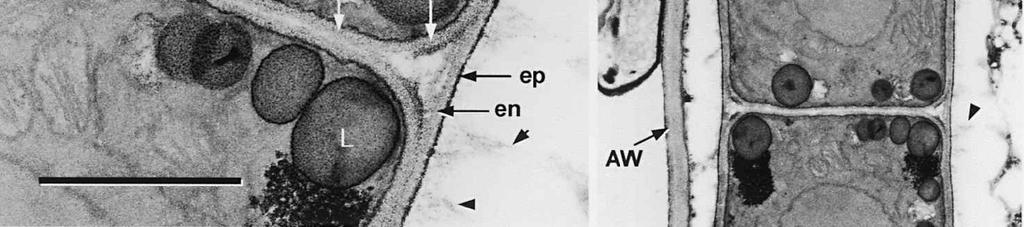
4 cylinder is composed of a homogeneous, moderately reactive, granular matrix. The strongest PA-TCH-SP reactivity within the central cylinder is often seen close to the inner and outer surfaces of the wall. Patches of less reactive material are also observed in the inner layer of the lateral wall immediately adjacent to the apical thickening. At this stage, the outer stratum of the outer layer can already be partly eroded over the apical thickening (double arrows, Figs 3, 4). As the ascospore walls are being formed in the immature ascus, the apical apparatus becomes more compressed, and changes are observed in structure and reactivity of the annulus (Fig. 5). In the lower two-thirds of the annulus, fine layers of strongly and weakly reactive material appear to alternate (double white arrows, Fig. 5), while in the upper part of the annulus overall reactivity increases (large arrowhead, Fig. 5), except for a limited area directly surrounding the central cylinder (single white arrow, Fig. 5). In the middle of the central cylinder the reactivity also seemed to decrease in some asci (small arrow, Fig. 5). In the mature ascus with fully developed ascospores, the apical apparatus is strongly compressed (Fig. 6). The upper and lower parts of the annulus differ even more profoundly in reactivity, and an upper and a lower subregion can be distinguished shortly before dehiscence. In the central cylinder patches of material with decreased reactivity are also observed (single arrows, Fig. 6), and the outer stratum of the outer layer is now almost completely eroded away from the apical apparatus in most asci (double arrows, Fig. 6). After dehiscence, the annulus is everted over an angle of about 90 (Fig. 7). Few remnants of the central cylinder can be found. The remaining material of the apical thickening swells and disintegrates rapidly, as does the inner layer of the lateral wall. ASCOSPORE WALL. After ascospore delimitation is complete, the investing membrane and spore plasma membrane initially lie closely together (Fig. 3). They then become separated as material is deposited between these membranes to build the ascospore wall (Figs 6, 8, 9). Intermediate stages of spore wall development were insufficiently preserved in the material studied to allow description of this aspect of morphogenesis. Explanation to legends Abbreviations used in Figs A, annulus; AC, apical chamber; AS, ascospore; AT, apical thickening; AW, lateral ascus wall; CC, central cylinder; EP, epiplasm; en, endospore; ep, epispore; ER, endoplasmatic reticulum; G, glycogen; IL, inner layer of ascus wall; im, investing membrane; is, inner stratum; L, lipid body; m, mitochondrion; n, nucleus; OL, outer layer of ascus wall; os, outer stratum; Pa, paraphysis; SP, sporoplasm; spm, spore plasma membrane; Va, vacuole. Figs 1-3. Longitudinal median sections of asci of Ombrophila hemiamyloidea. 1. Young ascus, still elongating, prior to the formation of the apical apparatus. 2. Young ascus, during formation of the apical apparatus. Single short arrow indicates where inner layer increases in thickness to form the apical thickening. 3. Immature ascus, just after completion of ascospore delimitation. Double arrows indicate where the outer stratum of the outer layer is eroded above the apical apparatus. Scale bars = 1 µm. 274
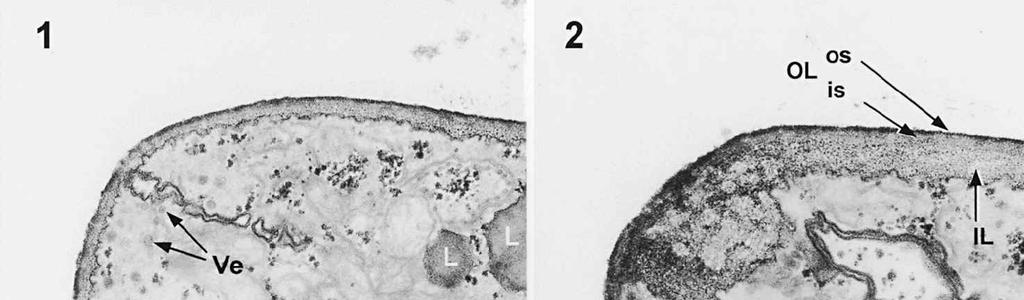
5 275
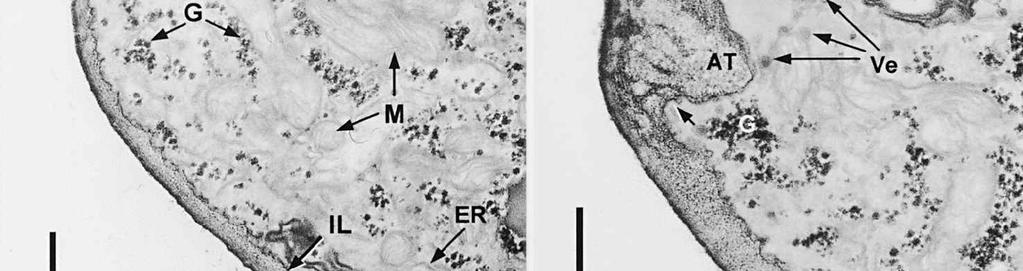
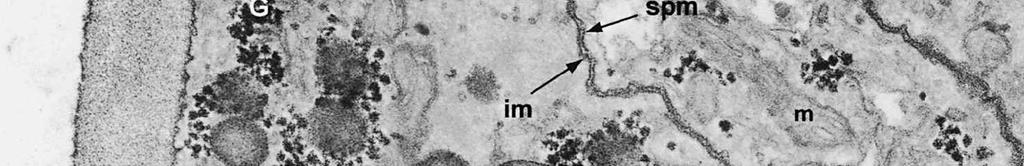
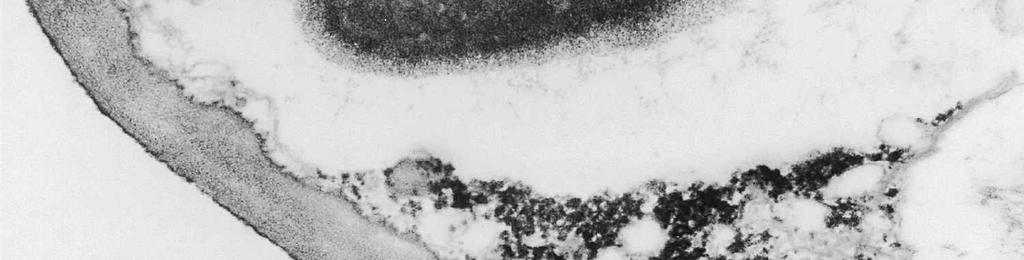
6 The mature ascospore wall is characterized by a moderately reactive, nm thick endospore, which is lined by a slightly less or equally reactive inner wall layer nm thick (Fig. 9). This inner wall layer curves around a 90 bend at the transverse septum, and from two directions forms one of the two layers that make up the spore septum. These septa are formed prior to ascus dehiscence (Figs 8, 9). The endospore is covered by a strongly reactive epispore, which is about 20 nm thick. Between the wall and the investing membrane, a reactive matrix substance is observed which in places is in contact with the epispore (arrowheads, Figs 6, 8, 9). Discussion The structure of the ascus wall and apical apparatus The ultrastructural data on the general morphology of the apical apparatus and its reactivity in O. hemiamyloidea are summarized diagrammatically in Figs 10 and 11. The lateral ascus wall is two-layered as in other helotialean taxa studied (Verkley 1995b). The inner and outer stratum of the outer layer agree with those observed in other Helotiales. These strata generally correspond with the a- and b-layer of the ascus wall reported in the studies by Bellemère (1975, 1977), while the inner layer corresponds to the c- and d-layer. In O. hemiamyloidea, however, no such stratification was observed within the inner layer of the lateral ascus wall. As in most Helotialean species investigated, only the inner layer increases in thickness in the apical apparatus of O. hemiamyloidea. This species showed a distinct apex maturation pattern (Verkley 1992), i.e., ultrastructure and PA-TCH-SP reactivity of the apical apparatus changed several times between its completion shortly before ascospore delimitation and the time just prior to dehiscence. A pronounced differentiation of subregions within the annulus, as observed in mature asci of O. hemiamyloidea, is rare in the Helotiales. It has so far only been observed in the ascus of Hymenoscyphus consobrinus (Boud.) Hengstmengel (Verkley 1993b). Based on his TEM studies of several members of the Helotiales, Bellemère (1977) suggested that the ascus apical apparatus of these fungi contained two rings, an outer ring ( anneau supérieur ) composed of c-layer material of the ascus wall, and an inner ring ( anneau inférieur ) composed of d- layer material. Bellemère et al. (1987), however, no longer insisted on the linkage of these layers and rings. According to Baral (1987b), the annulus of Helotiales consists of three separate rings, which correspond with three layers of the ascus wall. His observations, however, were all made with the light microscope. I regard the annulus of O. hemiamyloidea as a single functional and structural unit, a single ring, which Figs 4, 5. Longitudinal median sections of asci of Ombrophila hemiamyloidea. 4. Immature ascus, early stage. Double arrows indicate where the outer stratum of the outer layer is eroded above the apical apparatus. 5. Immature ascus, advanced stage with compressed apical apparatus. Double white arrows point to alternating weakly and strongly reactive layers within the annulus. In the upper part of the annulus, overall reactivity increases (large arrowhead), except for a limited area directly surrounding the central cylinder (single white arrow). The middle of the central cylinder may also show a decrease in reactivity (small arrow). Scale bars = 1 µm. 276
7 277
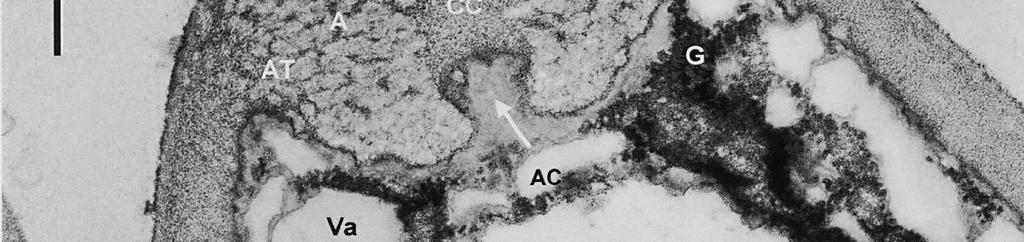
8 is developed during apex formation as a homogeneous and specialized region of the inner layer of the ascus wall. No stratification of the inner layer corresponding with the annular subregions was observed in the ascus wall of O. hemiamyloidea. In Helotiales with euamyloid asci (always blueing in IKI and Mlz; reaction type bb), the parts in the apical apparatus blueing in iodine under the light microscope largely correspond with structurally differentiated parts observed with TEM, which usually also react strongly with PA-TCH-SP. A marked exception is Trichopeziza mollissima (Lasch) Fuckel. The annulus of that species blues intensely in iodine, yet is entirely non-reactive in PA-TCH-SP. In the wall material of the apical thickening surrounding the annulus of T. mollisima the situation is reversed (Verkley 1996). Also in species of Sclerotiniaceae (Verkley 1993a), the euamyloid annulus was often less PA-TCH- SP reactive than the surrounding, inamyloid apical thickening. I therefore conclude that iodine reactions observed in fresh material need not strictly correlate with PA- TCH-SP reactivity in the same material after fixation. Interestingly, the annular material of Calycina herbarum (Pers. : Fr.) S. F. Gray (syn. Hymenoscyphus herbarum (Pers. : Fr.) Dennis) loses much of its reactivity as the ascus reaches full maturity. According to Baral (1987a), the asci of this species are hemiamyloid of type rb, staining red above and blue below an iodine concentration of 0.08% in IKI. But in the same species I have only seen amyloid asci that also blue at higher iodine concentrations. Lizon (1992) also reported that the ascus in this species blues distinctly in Melzer s reagent. In mature asci of O. hemiamyloidea, the annular subregions react with different intensity to PA-TCH-SP, but under the light microscope the red staining in IKI is consistently intense. Hence, the PA-TCH-SP reactivity and hemiamyloidity (reaction type rr) are not correlated either. The taxonomic position of Ombrophila hemiamyloidea As pointed out by Baral (1999), the taxonomic position of O. hemiamyloidea is still uncertain. It could have affinities to the families Dermateaceae and Vibrisseaceae, as well as to the Helotiaceae in which it is currently placed. The ultrastructural data of the ascus apical apparatus and mature ascospore wall conflict with the placement of O. hemiamyloidea in the genus Ombrophila. In O. violacea Fr., the type species of Ombrophila, the annulus is a narrow, discontinuous structure embedded in a welldeveloped apical thickening, which shows a relatively small, persistent, and highly reactive annular protrusion (Ombrophila type of apical apparatus, fide Verkley 1992). In O. hemiamyloidea, the annulus fully occupies the apical thickening, which does Figs 6-9. Longitudinal median sections of asci and ascospores of Ombrophila hemiamyloidea. 6. Mature ascus. Single arrows point to patches of material with decreased reactivity within the central cylinder. Note the erosion of the outer stratum of the outer layer over most of the apical apparatus (double arrows). Arrowheads point to the reactive matrix substance in contact with the epispore of the ascospore wall. 7. Dehisced ascus. 8. Mature, 3-septate ascospore within a mature ascus. Arrowheads indicate the reactive matrix substance in contact with the epispore. 9. Detail of ascospore wall in a mature ascospore near a transverse spore septum. Arrowheads indicate the reactive matrix substance in contact with the epispore. Scale bars = 1 µm. 278
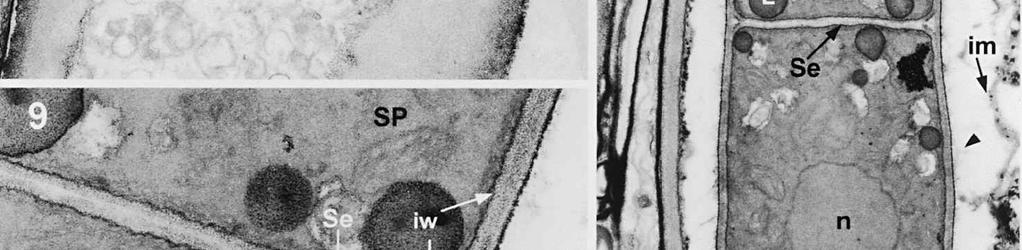
9 279
10 Figs 10, 11. Diagrammatic schemes of the ascus apical apparatus and lateral ascus wall of Ombrophila hemiamyloidea, showing the relative PA-TCH-SP reactivity on the left, and layers, strata and annular region on the right of each scheme. 10. Early immature ascus. 11. Fully mature ascus. 280
11 not contain an annular protrusion. Moreover, the asci of O. violacea are amyloid, and hemiamyloid asci have thus far not been reported in other species of Ombrophila. The apical apparatus of Neobulgaria pura (Fr. : Fr.) Petr. was found to be similar to that of O. violacea, and therefore it is of the Ombrophila type (Verkley 1995b). The walls of mature ascospores of O. hemiamyloidea show a compact and homogeneous epispore of strongly PA-TCH-SP reactive amorphous material. In contrast, mature ascospores of O. violacea show a layer of strongly reactive bands which are oriented perpendicularly to the wall surface, covered by a diffuse matrix that forms a gelatinous sheath. The spore walls of N. pura are similar to those of O. violacea, but the outermost layer consists of bristles instead of bands. In O. hemiamyloidea, the epispore, and perhaps also the loose material surrounding it, are probably responsible for the reaction in IKI of the ascospore walls. Such reactions are unknown in other Ombrophila species. The general morphology of the apical apparatus and the structure and reactivity of the annulus of O. hemiamyloidea agree well with those observed in two Pezicula species in the family Dermateaceae (Bellemère 1977). Bellemère depicted a relatively narrow annulus in his diagrammatic scheme of the Pezicula-type ascus and, as a consequence, called the most protruding part of the apical thickening pendentif (annular protrusion). However, the electron micrograph of his Plate 1b clearly shows that the annulus is in fact much broader than described, occupying the entire apical thickening. Bellemère (1977) described a differentiation of the upper and lower regions of the annulus of Pezicula in terms of PA-TCH-SP reactivity. A very similar differentiation is also observed in O. hemiamyloidea. Bellemère (1977) did not note changes during apex maturation in Pezicula. No image or description of a dehisced ascus was provided but, based on my light-microscopic observations, the mode of dehiscence appears similar to that in O. hemiamyloidea. The a- and b-layers in Pezicula agree well with the outer and inner stratum of the outer layer in O. hemiamyloidea. However, no thin, strongly reactive zone ( un très mince liseré riche en polysaccharides ), which separates the c- and d-layers in Pezicula, was observed in O. hemiamyloidea, nor was any stratification seen within the inner layer of the ascus wall. But these are only minor differences, and I conclude that the ascus apical apparatus of O. hemiamyloidea is of the Pezicula type. Species of Pezicula typically also show hemiamyloid reactions in fresh or recently dried material. In addition, shape and septation of the ascospores are the same in Pezicula. However, under normal conditions, Pezicula species form septa after discharge, whereas in O. hemiamyloidea septation occurs prior to discharge (personal observations; Baral 1999). According to Bellemère (1977), the genera Geoglossum Pers. and Microglossum Gillet also have Pezicula-type asci but, in earlier investigations of G. nigritum Cooke, G. cookeianum Nannf., and Microglossum viride (Pers.) Gillet, I found significant morpholo gical differences in the apparatus, and proposed the distinction of Geoglossum and Microglossum types (Verkley 1994, 1995b). Bellemère (1977) also regarded the asci of Polydesmia Boud. and Melittosporium Sacc. as probably being of Pezicula type, of which he considered the ascus of Dermea cerasi (Pers. : Fr.) Fr. to be a mere variant. 281
12 The family Dermateaceae as traditionally defined is highly unnatural (Nauta & Spooner 1999). It encompasses at least two distantly related entities: one containing Dermea Fr., the type genus of the family, as well as related genera such as Pezicula and Neofabraea H. S. Jacks. (Abeln et al. 2000, De Jong et al. 2001), and a second group that contains the mollisioid fungi and is sometimes informally referred to as the Mollisiaceae (Baral 1999). Little is known about the ultrastructure of the apical apparatus in these fungi. Bellemère (1977) mentioned that the ascus apical apparatus of Tapesia (= Mollisia) strobilicola (Rehm) Sacc. (as Tapesia strobilina), T. cf. cinerea, and Pyrenopeziza rubi (Fr. : Fr.) Rehm shared certain features with that of Bulgaria inquinans Fr. : Fr., but he did not present micrographs for these species. The ultrastructure of the relatively small, amyloid apical apparatus of T. strobilicola (Bellemère et al. 1987) differs considerably from that of O. hemiamyloidea, and also from Bulgaria inquinans (Bellemère 1977, Verkley 1992). Baral (1999) suggested that O. hemiamyloidea might be closely related to Belonopsis (Sacc.) Rehm, on account of the abundant crystals in the medullary excipulum. He also suggested a possible affinity with Niptera Fr. (including Nimbomollisia Nannf.), on account of similar spore morphology and septation, as well as the presence of an IKI-red gel around the ascospores. Based on morphological criteria and results of sequence analysis of the complete small subunit of the ribosomal RNA gene, Döring & Triebel (1998) argued that Bulgaria Fr. is best placed in a separate family within the order Helotiales. Kirk et al. (2001) subsequently accepted the family Bulgariaceae Fr. in the ninth edition of the Dictionary of the Fungi. I recognized the apical apparatus of Bulgaria inquinans as a distinct type, Bulgaria, differing in general morphology as well as mode of dehiscence from those of Ombrophila violacea and Neobulgaria pura (Ombrophila type), Ascocoryne (Hymenoscyphus type), and Claussenomyces (Leotia type) (Bellemère 1969, 1977 Verkley 1992, 1995b). The annulus and apical thickening of Bulgaria inquinans and O. hemiamyloidea are similar in relative proportions, but there are also important differences between the species. For example, the apical apparatus of Bulgaria inquinans is not everted after dehiscence, while that of O. hemiamyloidea everts to an angle of about 90. Baral (1999) summarized the characters that O. hemiamyloidea has in common with certain Vibrisseaceae. For example, Leucovibrissea obconica (Kanouse) Korf, the type species of the unispecific genus Leucovibrissea (A. Sánchez) Korf, forms numerous rhomboid crystals and druses in the medullary excipulum as O. hemiamyloidea does but, contrasting with O. hemiamyloidea, it also forms them on the hyaline ectal excipulum (Baral 1999). Nevertheless, the ascus apical apparatus of O. hemiamyloidea differs greatly from that of members of the Vibrisseaceae studied so far. The amyloid ascus apex of Vibrissea decolorans (Saut.) Sánchez & Korf was recognized as a distinct type (Bellemère 1977). The Vibrissea-type ascus is characterized mainly by the presence of a relatively small but well-developed apical thickening containing a reactive annulus which is associated with a distinct annular protrusion. This complex structure is surrounded by a relatively large, well-developed subapical thickening ( bourrelet sous-apical ), which is composed of material considered to belong to the d2-layer of the ascus wall. This thickening encloses a 282
13 part of the epiplasm, with on top a mass of amorphous material ( calotte épiplasmique, Bellemère 1977). The ascus apical apparatus of Apostemidium fiscellum (P. Karst.) P. Karst. shows a very similar ultrastructure (Verkley unpubl. data), and is also of the Vibrissea type. Sánchez & Korf (1966) upheld a broad concept of Vibrissea Fr., including in it species with stipitate and sessile apothecia classified in Apostemidium P. Karst. by others. When Korf (1990) erected the family Vibrisseaceae, he included only three genera that occurred in aquatic habitats and produced filiform ascospores, viz. Vibrissea, Chlorovibrissea L.M. Kohn, and Leucovibrissea. He considered these genera to be distant from all other inoperculates. The asci of Chlorovibrissea and Leucovibrissea seem very similar to those of Vibrissea in light microscopy, but no ultrastructural data are available. Iodine reactions of asci vary among these fungi from amyloid to inamyloid, but hemiamyloid asci are unknown in the Vibrisseaceae (Baral 1999). Conclusions - The data on the ultrastructure of the ascus apical apparatus and mature ascospore wall do not support the placement of O. hemiamyloidea in the genus Ombrophila. These data give no indication of a close relationship with other Leotiaceae or Vibrisseaceae which have so far been studied with TEM. - The ascus apical apparatus of O. hemiamyloidea is considered to belong to the Pezicula type, and both the ultrastructure and hemiamyloid reaction of the ascus indicate possible relationships with the Dermateaceae in a restricted sense, viz. Dermea and related genera including Pezicula, but excluding the Mollisiaceae. - Molecular studies of selected members of the family Dermateaceae will be necessary to corroborate these findings, and decide about the generic classification of O. hemiamyloidea. Acknowledgements Dr Dagmar Triebel is kindly thanked for loan of the holotype material of O. hemiamyloidea from the Botanische Staatssammlung München. Drs R. C. Summerbell, J. van Brummelen, and P. W. Crous are kindly thanked for critical reading of the manuscript. References ABELN, E.C., M.A. DE PAGTER & G.J.M. VERKLEY (2000): Phylogeny of Pezicula, Dermea and Neofabraea inferred from partial sequences of the nuclear ribosomal RNA gene cluster. Mycologia 92: BARAL, H.O. (1987a): Lugol s solution/iki versus Melzer s reagent: hemiamyloidity a universal feature of the ascus wall. - Mycotaxon 29: BARAL, H.O. (1987b): Der Apikalapparat der Helotiales. Eine lichtmikroskopische Studie über Arten mit Amyloidring. - Z. Mykol. 53: BARAL, H.O. (1999): Ombrophila hemiamyloidea (Leotiales), a new aquatic discomycete. - Mycol. Bavar. 3:
14 BELLEMÈRE, A. (1969): Quelques observations relatives à l infrastructure de l appareil apical des asques du Bulgaria inquinans Fr. (Discomycète inoperculé). - C. R. Hebd. Séanc. Acad. Sci., Paris. Sér. D, 268 : BELLEMÈRE, A. (1975): Étude ultrastructuraledes asques: la paroi de l appareil apical, la paroi des ascospores chez des Discomycètes inoperculés et des Hystériales. - Physiol. Végét. 13: BELLEMÈRE, A. (1977): L appareil apical de l asque chez quelques discomycètes: Étude ultrastructurale comparative. - Rev. Mycol. 41: BELLEMÈRE, A., M.C. JANEX-FAVRE & A. PARGUEY-LEDUC (1987): Marius Chadefaud et les asques: données inédites. Études ultrastructurales complémentaires. - Bull. Soc. Bot. Fr. 134, Lettres bot. 3: CHADEFAUD, M. (1964): Sur l origine et la structure des asques du type annellascé. - C. R. Hebd. Séanc. Acad. Sci., Paris, Sér. D, 244: CHADEFAUD, M. (1973): Les asques et la systématique des Ascomycètes (1). - Bull. Trimestriel Soc. Mycol. Fr. 89: COLLADO, J., V. RUBIO, R. GALÁN, G. PLATAS, F. ARENAL, V. GONZÁLEZ, J. SÁNCHEZBALLESTEROS, M. VILLARREAL, H.O. BARAL & F. PELÁEZ (2002). Molecular phylogeny of ascomycetes of the Helotiales.- Abstr. Seventh Int. Mycol. Congr., Oslo. No CROUS, P., J.Z. GROENEWALD & W. GAMS (2003): Eyespot of cereals revisited: ITS phylogeny reveals new species relationships. - Eur. J. Pl. Pathol. In press. DE JONG, S.N., C.A. LEVESQUE, G.J.M. VERKLEY, E.C. ABELN, J.E. RAHE & P.G. BROWN (2001): Phylogenetic relationships among Neofabraea species causing tree cankers and bull s-eye rot of apple based on DNA sequencing of ITS nuclear rdna, mitochondrial rdna, and the ßtubulin gene. - Mycol. Res. 105: DÖRING, H. & D. TRIEBEL (1998): Phylogenetic relationhips of Bulgaria inferred by 18S rdna sequence analysis. - Cryptog., Bryol. Lichénol., 19: GERNANDT, D.S., J.L. PLATT, J.K. STONE, J.S. SPATAFORA, A. HOLST-JENSEN, R.C. HAMELIN & L.M. KOHN (2001): Phylogenetics of Helotiales and Rhytismatales based on partial small subunit nuclear ribosomal DNA sequences. - Mycologia 93: KIRK, P.M., P.F. CANNON, J.C. DAVID & J.A. STALPERS, eds. (2001): Ainsworth & Bisby s Dictionary of the Fungi, 9th ed. - CABI, Wallingford. KORF, R.P. (1990): Discomycete systematics today: a look at some unanswered questions in a group of unitunicate ascomycetes. - Mycosystema 3: LENURM, K. & A. RAITVIIR (2000): The ultrastructure of Belonidium aeruginosum Mont. & Durieu (Hyaloscyphaceae, Helotiales). - Folia Cryptog. Eston. 36: LENURM, K. & A. RAITVIIR (2001): The ultrastructure of Lachnellula willkommii (Hyaloscyphaceae, Helotiales, Ascomycetes). - Folia Cryptog. Eston. 36: LIZON, P. (1992): The genus Hymenoscyphus (Helotiales) in Slovakia, Czechoslovakia. - Mycotaxon 45: NAUTA, M.M. & B. SPOONER (1999). British Dermateaceae: 1. Introduction. - Mycologist 13: 3-6. SÁNCHEZ, A. & R.P. KORF (1966). The genus Vibrissea, and the generic names Leptosporium, Apostemidium, Gorgoniceps and Ophiogloea. - Mycologia 58: THIÉRY, J.P. (1967): Mise en évidence des polysaccharides sur coupes fines en microscopie électronique. - J. Microsc. 6: TRIEBEL, D & H.O. BARAL (1996): Notes on the ascus types in Crocicreas (Leotiales, Ascomycetes) with a characterization of selected taxa. - Sendtnera 3:
15 VERKLEY, G.J.M. (1992): Ultrastructure of the apical apparatus of asci in Ombrophila violacea, Neobulgaria pura and Bulgaria inquinans (Leotiales). - Persoonia 15: VERKLEY, G.J.M. (1993a): Ultrastructure of the ascus apical apparatus in ten species of Sclerotiniaceae. - Mycol. Res. 97: VERKLEY, G.J.M. (1993b): Ultrastructure of the ascus apical apparatus in Hymenoscyphus and other genera of the Hymenoscyphoideae (Leotiales, Ascomycotina). - Persoonia 15: VERKLEY, G.J.M. (1994): Ultrastructure of the ascus apical apparatus in Leotia lubrica and some Geoglossaceae. - Persoonia 15: VERKLEY, G.J.M. (1995a): Ultrastructure of the ascus apical apparatus in species of Cenangium, Encoelia, Claussenomyces and Ascocoryne. - Mycol. Res. 99: VERKLEY, G.J.M. (1995b): The ascus apical apparatus in Leotiales: an evaluation of ultrastructural characters as phylogenetic markers in the families Sclerotiniaceae, Leotiaceae, and Geoglossaceae. Ph. D. Thesis, Rijksuniversiteit Leiden. VERKLEY, G.J.M. (1996): Ultrastructure of the ascus in the genera Lachnum and Trichopeziza (Hyaloscyphaceae, Ascomycotina). - Nova Hedwigia 63: VERKLEY, G.J.M. (1999): A monograph of the genus Pezicula and its anamorphs. - Stud. Mycol. 44: Received 28 April 2003, accepted in revised form 20 May
(From The Rockefeller Institute) Materials and Methods. Observations with the Electron Microscope
 Materials and Methods. Observations with the Electron Microscope") ELECTRON MICROSCOPE STUDY OF THE DEVELOPMENT OF THE PAPILLOMA VIRUS IN THE SKIN OF THE RABBIT* BY ROBERT S. STONE,~ M.D., RICHARD E. SHOPE, M.D., DAN H. MOORE, P,~.D. (From The Rockefeller Institute) PLATES
ELECTRON MICROSCOPE STUDY OF THE DEVELOPMENT OF THE PAPILLOMA VIRUS IN THE SKIN OF THE RABBIT* BY ROBERT S. STONE,~ M.D., RICHARD E. SHOPE, M.D., DAN H. MOORE, P,~.D. (From The Rockefeller Institute) PLATES
The ultrastructure of the egg and
 The ultrastructure of the egg and central cell of Petunia J.L. van Went Botanisch Laboratorium, Universiteit, Nijmegen SUMMARY The egg and central cell of Petunia hybrida undergo a number of changes and
The ultrastructure of the egg and central cell of Petunia J.L. van Went Botanisch Laboratorium, Universiteit, Nijmegen SUMMARY The egg and central cell of Petunia hybrida undergo a number of changes and
Intercellular Matrix in Colonies of Candida
 JouRNAL OF BAcTEROLOGY, Sept. 1975, p. 1139-1143 Vol. 123, No. 3 Copyright 0 1975 American Society for Microbiology Printed in U.S.A. ntercellular Matrix in Colonies of Candida K. R. JOSH, J. B. GAVN,*
JouRNAL OF BAcTEROLOGY, Sept. 1975, p. 1139-1143 Vol. 123, No. 3 Copyright 0 1975 American Society for Microbiology Printed in U.S.A. ntercellular Matrix in Colonies of Candida K. R. JOSH, J. B. GAVN,*
The Fine Structure of the Epithelial Cells of the Mouse Prostate* II. Ventral Lobe Epithelium
 Published Online: 1 June, 1960 Supp Info: http://doi.org/10.1083/jcb.7.3.511 Downloaded from jcb.rupress.org on September 28, 2018 The Fine Structure of the Epithelial Cells of the Mouse Prostate* II.
Published Online: 1 June, 1960 Supp Info: http://doi.org/10.1083/jcb.7.3.511 Downloaded from jcb.rupress.org on September 28, 2018 The Fine Structure of the Epithelial Cells of the Mouse Prostate* II.
An Electron-Microscope Study of Germination of Conidia of Botrytis cinerea
 J. gen. Microbiol. (1963), 33, 43-46 With 2 plates Printed in Great Britain 43 An Electron-Microscope Study of Germination of Conidia of Botrytis cinerea BY LILIAN E. HAWKER AND R. J. HENDY Department
J. gen. Microbiol. (1963), 33, 43-46 With 2 plates Printed in Great Britain 43 An Electron-Microscope Study of Germination of Conidia of Botrytis cinerea BY LILIAN E. HAWKER AND R. J. HENDY Department
COMPARATIVE DISTRIBUTION OF CARBOHYDRATES AND LIPID DROPLETS IN THE GOLGI APPARATUS OF INTESTINAL ABSORPTIVE CELLS
 COMPARATIVE DISTRIBUTION OF CARBOHYDRATES AND LIPID DROPLETS IN THE GOLGI APPARATUS OF INTESTINAL ABSORPTIVE CELLS JEAN A. SAGE and RALPH A. JERSILD, JR. Medical Center, Indianapolis, Indiana 46202 From
COMPARATIVE DISTRIBUTION OF CARBOHYDRATES AND LIPID DROPLETS IN THE GOLGI APPARATUS OF INTESTINAL ABSORPTIVE CELLS JEAN A. SAGE and RALPH A. JERSILD, JR. Medical Center, Indianapolis, Indiana 46202 From
PERSOONIA. Part 2, pp (1962) Studies on discomycetez I. of Ascobolus and Saccobolus. herbarium, Rijksherbarium, Leiden. inquirenda. on loan.
 Studies on discomycetez I. of Ascobolus and Saccobolus. herbarium, Rijksherbarium, Leiden. inquirenda. on loan.") FIG. PERSOONIA Published by the Rijksherbarium, Leiden Volume 2, Part 2, pp. 195-199 (1962) Studies on discomycetezi. s of species of Ascobolus and Saccobolus in Spegazzini s herbarium J. van Brummelen
FIG. PERSOONIA Published by the Rijksherbarium, Leiden Volume 2, Part 2, pp. 195-199 (1962) Studies on discomycetezi. s of species of Ascobolus and Saccobolus in Spegazzini s herbarium J. van Brummelen
Cell Overview. Hanan Jafar BDS.MSc.PhD
 Cell Overview Hanan Jafar BDS.MSc.PhD THE CELL is made of: 1- Nucleus 2- Cell Membrane 3- Cytoplasm THE CELL Formed of: 1. Nuclear envelope 2. Chromatin 3. Nucleolus 4. Nucleoplasm (nuclear matrix) NUCLEUS
Cell Overview Hanan Jafar BDS.MSc.PhD THE CELL is made of: 1- Nucleus 2- Cell Membrane 3- Cytoplasm THE CELL Formed of: 1. Nuclear envelope 2. Chromatin 3. Nucleolus 4. Nucleoplasm (nuclear matrix) NUCLEUS
7'cA 5P 'Z /' IA.N 5 '7S abra'ry TECHNiCAL F:-'.. THE INSTITUTE OF PAPER CHEMISTRY, APPLETON, WISCONSIN IPC TECHNICAL PAPER SERIES NUMBER 16
 7'cA 5P 'Z /' IA.N 5 '7S abra'ry TECHNiCAL F:-'.. THE INSTITUTE OF PAPER CHEMISTRY, APPLETON, WISCONSIN IPC TECHNICAL PAPER SERIES NUMBER 16 DIFFERENTIATION OF TANNIN, LIPID, AND STARCH IN CULTURED PLANT
7'cA 5P 'Z /' IA.N 5 '7S abra'ry TECHNiCAL F:-'.. THE INSTITUTE OF PAPER CHEMISTRY, APPLETON, WISCONSIN IPC TECHNICAL PAPER SERIES NUMBER 16 DIFFERENTIATION OF TANNIN, LIPID, AND STARCH IN CULTURED PLANT
Ultrastructure of Connective Tissue Cells of Giant African Snails Achatina fulica (Bowdich)
") Kasetsart J. (Nat. Sci.) 36 : 285-290 (2002) Ultrastructure of Connective Tissue Cells of Giant African Snails Achatina fulica (Bowdich) Viyada Seehabutr ABSTRACT The connective tissue sheath of cerebral
Kasetsart J. (Nat. Sci.) 36 : 285-290 (2002) Ultrastructure of Connective Tissue Cells of Giant African Snails Achatina fulica (Bowdich) Viyada Seehabutr ABSTRACT The connective tissue sheath of cerebral
Ascospores of Metschnikowia Kamienski
 JOURNAL OF BACTERIOLOGY, July 1973, p. 316-322 Copyright 0 1973 American Society for Microbiology Vol. 115, No. 1 Printed in U.SA. Electron Micrograph Study of the Asci and Ascospores of Metschnikowia
JOURNAL OF BACTERIOLOGY, July 1973, p. 316-322 Copyright 0 1973 American Society for Microbiology Vol. 115, No. 1 Printed in U.SA. Electron Micrograph Study of the Asci and Ascospores of Metschnikowia
the structure of their ducts has been
 Tza JOURNAL 0? INVEa'riGATrVN DEBMATOLOOT Copyright t 1966 by The Williams & Wilkins Co. Vol. 46, No. I Printed in U.S.A. AN ELECTRON MICROSCOPIC STUDY OF THE ADULT HUMAN APOCRINE DUCT* KEN HASHIMOTO,
Tza JOURNAL 0? INVEa'riGATrVN DEBMATOLOOT Copyright t 1966 by The Williams & Wilkins Co. Vol. 46, No. I Printed in U.S.A. AN ELECTRON MICROSCOPIC STUDY OF THE ADULT HUMAN APOCRINE DUCT* KEN HASHIMOTO,
Zhuang, W.Y. (1999). Discomycetes of tropical China. VI. Additional species from Guangxi. Fungal Diversity 3:
. Discomycetes of tropical China. VI. Additional species from Guangxi. Fungal Diversity 3:") Fungal Diversity Discomycetes Guangxi of tropical China. VI. Additional species from Wen-Ying Zhuang Systematic Mycology and Lichenology Laboratory, Institute of Microbiology, Chinese Academy of Sciences,
Fungal Diversity Discomycetes Guangxi of tropical China. VI. Additional species from Wen-Ying Zhuang Systematic Mycology and Lichenology Laboratory, Institute of Microbiology, Chinese Academy of Sciences,
Biology. Dr. Khalida Ibrahim
 Biology Dr. Khalida Ibrahim BONE TISSUE Bone tissue is a specialized form of connective tissue and is the main element of the skeletal tissues. It is composed of cells and an extracellular matrix in which
Biology Dr. Khalida Ibrahim BONE TISSUE Bone tissue is a specialized form of connective tissue and is the main element of the skeletal tissues. It is composed of cells and an extracellular matrix in which
AN ELECTRON-MICROSCOPIC STUDY OF THE STARCH-CONTAINING PLASTIDS IN THE FERN TODEA BARBARA
 J. Cell Sci. 4, 211-221 (1969) 211 Printed in Great Britain AN ELECTRON-MICROSCOPIC STUDY OF THE STARCH-CONTAINING PLASTIDS IN THE FERN TODEA BARBARA H. M. SMITH* AND D. S. SMITHf Department of Biology,
J. Cell Sci. 4, 211-221 (1969) 211 Printed in Great Britain AN ELECTRON-MICROSCOPIC STUDY OF THE STARCH-CONTAINING PLASTIDS IN THE FERN TODEA BARBARA H. M. SMITH* AND D. S. SMITHf Department of Biology,
IT has been shown (Chou, 1957 a, b) that there are three kinds of lipid
 that there are three kinds of lipid") 279 The Ultra-fine Structure of Lipid Globules in the Neurones of Helix aspersa By J. T. Y. CHOU and G. A. MEEK (From the Cytological Laboratory, Department of Zoology; and Department of Human Anatomy,
279 The Ultra-fine Structure of Lipid Globules in the Neurones of Helix aspersa By J. T. Y. CHOU and G. A. MEEK (From the Cytological Laboratory, Department of Zoology; and Department of Human Anatomy,
Luciotrichus lasioboloides, a new genus and a new species of the Pezizales
 CZECH MYCOL. 47 (4), 1994 Luciotrichus lasioboloides, a new genus and a new species of the Pezizales RICARDO GAL..\N and AIN RAITVIIR 1 ) Department of Plant Biology, University of Alcalli 288 71 Alcalli
CZECH MYCOL. 47 (4), 1994 Luciotrichus lasioboloides, a new genus and a new species of the Pezizales RICARDO GAL..\N and AIN RAITVIIR 1 ) Department of Plant Biology, University of Alcalli 288 71 Alcalli
A Compact and a Dispersed Form of the Golgi Apparatus
 A Compact and a Dispersed Form of the Golgi Apparatus of Fish Liver 1 D. James Morre and Carole A. Lembi Department of Botany and Plant Pathology Purdue University, Lafayette, Indiana 47907, and H. H.
A Compact and a Dispersed Form of the Golgi Apparatus of Fish Liver 1 D. James Morre and Carole A. Lembi Department of Botany and Plant Pathology Purdue University, Lafayette, Indiana 47907, and H. H.
Basophilic. Basophilic structures are stained by basic dyes: Mnemonic: Basophilic = Blue
 Cell Overview Basophilic Basophilic structures are stained by basic dyes: Basic dyes are positive Basophilic structures are negative (ex. DNA, RNA, ribosomes, RER) Mnemonic: Basophilic = Blue Acidophilic
Cell Overview Basophilic Basophilic structures are stained by basic dyes: Basic dyes are positive Basophilic structures are negative (ex. DNA, RNA, ribosomes, RER) Mnemonic: Basophilic = Blue Acidophilic
Electron Microscopy of Small Cells: Mycoplasma hominis
 JOURNAL of BAcTRiowOY, Dc. 1969, p. 1402-1408 Copyright 0 1969 American Society for Microbiology Vol. 100, No. 3 Printed In U.S.A. NOTES Electron Microscopy of Small Cells: Mycoplasma hominis JACK MANILOFF
JOURNAL of BAcTRiowOY, Dc. 1969, p. 1402-1408 Copyright 0 1969 American Society for Microbiology Vol. 100, No. 3 Printed In U.S.A. NOTES Electron Microscopy of Small Cells: Mycoplasma hominis JACK MANILOFF
New aspect of hepatic nuclear glycogenosis
 J. clin. Path. (1968), 21, 19 New aspect of hepatic nuclear glycogenosis in diabetes1 F. CARAMIA, F. G. GHERGO, C. BRANCIARI, AND G. MENGHINI From the Institute of General Pathology, University of Rome,
J. clin. Path. (1968), 21, 19 New aspect of hepatic nuclear glycogenosis in diabetes1 F. CARAMIA, F. G. GHERGO, C. BRANCIARI, AND G. MENGHINI From the Institute of General Pathology, University of Rome,
A Tour of the Cell. Chapter 6. Biology Eighth Edition Neil Campbell and Jane Reece. PowerPoint Lecture Presentations for
 Chapter 6 A Tour of the Cell PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions from Joan Sharp
Chapter 6 A Tour of the Cell PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions from Joan Sharp
DEVELOPMENT AND DISPERSAL OF P-PROTEIN IN THE PHLOEM OF COLEUS BLUMEI BENTH.
 J. Cell Sci. 4, 155-169 (1969) 155 Printed in Great Britain DEVELOPMENT AND DISPERSAL OF P-PROTEIN IN THE PHLOEM OF COLEUS BLUMEI BENTH. M. W. STEER AND E. H. NEWCOMB Department of Botany, University of
J. Cell Sci. 4, 155-169 (1969) 155 Printed in Great Britain DEVELOPMENT AND DISPERSAL OF P-PROTEIN IN THE PHLOEM OF COLEUS BLUMEI BENTH. M. W. STEER AND E. H. NEWCOMB Department of Botany, University of
ELECTRON MICROSCOPIC STUDIES ON EQUINE ENCEPHALOSIS VIRUS
 Onderstepoort]. vet. Res. 40 (2), 53-58 (1973) ELECTRON MICROSCOPIC STUDIES ON EQUINE ENCEPHALOSIS VIRUS G. LECATSAS, B. J. ERASMUS and H. J. ELS, Veterinary Research Institute, Onderstepoort ABSTRACT
Onderstepoort]. vet. Res. 40 (2), 53-58 (1973) ELECTRON MICROSCOPIC STUDIES ON EQUINE ENCEPHALOSIS VIRUS G. LECATSAS, B. J. ERASMUS and H. J. ELS, Veterinary Research Institute, Onderstepoort ABSTRACT
Initially, the patients did not receive extra vitamin E except for a very
 EFFECT OF VITAMIN E ON MEMBRANES OF THE INTESTINAL CELL BY I. MOLENAAR, F. A. HOMMES, W. G. BRAAMS, AND H. A. POLMAN CENTER FOR MEDICAL ELECTRON MICROSCOPY AND DEPARTMENT OF PEDIATRICS, UNIVERSITY OF GRONINGEN,
EFFECT OF VITAMIN E ON MEMBRANES OF THE INTESTINAL CELL BY I. MOLENAAR, F. A. HOMMES, W. G. BRAAMS, AND H. A. POLMAN CENTER FOR MEDICAL ELECTRON MICROSCOPY AND DEPARTMENT OF PEDIATRICS, UNIVERSITY OF GRONINGEN,
North American Pezizales: Greletia and Marcellina
 V Sydowia, Annales Mycologici Ser. II. Vol. 38: 235-240 (1985) Verlag Ferdinand Berger & Söhne Gesellschaft m.b.h., 3580 Horn, Austria North American Pezizales: Greletia and Marcellina Donald H. PFISTER
V Sydowia, Annales Mycologici Ser. II. Vol. 38: 235-240 (1985) Verlag Ferdinand Berger & Söhne Gesellschaft m.b.h., 3580 Horn, Austria North American Pezizales: Greletia and Marcellina Donald H. PFISTER
A Tour of the Cell. reference: Chapter 6. Reference: Chapter 2
 A Tour of the Cell reference: Chapter 6 Reference: Chapter 2 Monkey Fibroblast Cells stained with fluorescent dyes to show the nucleus (blue) and cytoskeleton (yellow and red fibers), image courtesy of
A Tour of the Cell reference: Chapter 6 Reference: Chapter 2 Monkey Fibroblast Cells stained with fluorescent dyes to show the nucleus (blue) and cytoskeleton (yellow and red fibers), image courtesy of
Growth and repair: Cartilage is a vascular tissues that receives nutrients by diffusion through its matrix, cartilage grow by 2 mechanisms:
 Skeletal connective tissues: (cartilage and bone): Cartilage and bone are specialized connective tissues both adapted to serve as skeletal framework in most vertebrates the presence of solid inter cellular
Skeletal connective tissues: (cartilage and bone): Cartilage and bone are specialized connective tissues both adapted to serve as skeletal framework in most vertebrates the presence of solid inter cellular
Dentin Formation(Dentinogenesis)
") Lecture four Dr. Wajnaa Oral Histology Dentin Formation(Dentinogenesis) Dentinogenesis begins at the cusp tips after the odontoblasts have differentiated and begin collagen production. Dentinogenesis growth
Lecture four Dr. Wajnaa Oral Histology Dentin Formation(Dentinogenesis) Dentinogenesis begins at the cusp tips after the odontoblasts have differentiated and begin collagen production. Dentinogenesis growth
BIOSC 041. v Today s lecture. v Today s lab. v Note- Monday is a holiday good time to do some reading!
 BIOSC 041 v Today s lecture Review questions Chapter 6, Cells More review questions v Today s lab Quick review of lab safety The Scientific Method start thinking about which environments you might want
BIOSC 041 v Today s lecture Review questions Chapter 6, Cells More review questions v Today s lab Quick review of lab safety The Scientific Method start thinking about which environments you might want
Fine Structure of the Normal Trigeminal Ganglion in the Cat and Monkey*
 Fine Structure of the Normal Trigeminal Ganglion in the Cat and Monkey* DAVID S. MAXWELL, PH.D. Principal Contributor and Leader of Discussion HE inclusion of animal material m a y be justified as a means
Fine Structure of the Normal Trigeminal Ganglion in the Cat and Monkey* DAVID S. MAXWELL, PH.D. Principal Contributor and Leader of Discussion HE inclusion of animal material m a y be justified as a means
POLLEN-WALL PROTEINS: ELECTRON- MICROSCOPIC LOCALIZATION OF ACID PHOSPHATASE IN THE INTINE OF CROCUS VERNUS
 J. Cell Sci. 8, 727-733 (197O 727 Printed in Great Britain POLLEN-WALL PROTEINS: ELECTRON- MICROSCOPIC LOCALIZATION OF ACID PHOSPHATASE IN THE INTINE OF CROCUS VERNUS R.B. KNOX* AND J. HESLOP-HARRISONf
J. Cell Sci. 8, 727-733 (197O 727 Printed in Great Britain POLLEN-WALL PROTEINS: ELECTRON- MICROSCOPIC LOCALIZATION OF ACID PHOSPHATASE IN THE INTINE OF CROCUS VERNUS R.B. KNOX* AND J. HESLOP-HARRISONf
Cell Structure & Function. Source:
 Cell Structure & Function Source: http://koning.ecsu.ctstateu.edu/cell/cell.html Definition of Cell A cell is the smallest unit that is capable of performing life functions. http://web.jjay.cuny.edu/~acarpi/nsc/images/cell.gif
Cell Structure & Function Source: http://koning.ecsu.ctstateu.edu/cell/cell.html Definition of Cell A cell is the smallest unit that is capable of performing life functions. http://web.jjay.cuny.edu/~acarpi/nsc/images/cell.gif
Chapter 7. (7-1 and 7-2) A Tour of the Cell
 A Tour of the Cell") Chapter 7 (7-1 and 7-2) A Tour of the Cell Microscopes as Windows to the World of Cells Cells were first described in 1665 by Robert Hooke. By the mid-1800s, the accumulation of scientific evidence led
Chapter 7 (7-1 and 7-2) A Tour of the Cell Microscopes as Windows to the World of Cells Cells were first described in 1665 by Robert Hooke. By the mid-1800s, the accumulation of scientific evidence led
An exceptional find on rabbit dung from Italy : third record worldwide of Ascobolus perforatus
 An exceptional find on rabbit dung from Italy : third record worldwide of Ascobolus perforatus Doveri F* Via dei Funaioli 22, I 57126 Livorno. Doveri F. 2012 An exceptional find on rabbit dung from Italy
An exceptional find on rabbit dung from Italy : third record worldwide of Ascobolus perforatus Doveri F* Via dei Funaioli 22, I 57126 Livorno. Doveri F. 2012 An exceptional find on rabbit dung from Italy
psittaci by Silver-Methenamine Staining and
 JOURNAL OF BACTERIOLOGY, July 1972, p. 267-271 Copyright 1972 American Society for Microbiology Vol. 111, No. 1 Printed in U.S.A. Location of Polysaccharide on Chlamydia psittaci by Silver-Methenamine
JOURNAL OF BACTERIOLOGY, July 1972, p. 267-271 Copyright 1972 American Society for Microbiology Vol. 111, No. 1 Printed in U.S.A. Location of Polysaccharide on Chlamydia psittaci by Silver-Methenamine
Eukaryotic cells are essentially two envelope systems. Nuclear materials are separated from cytoplasm by nuclear membrane. Complex structure Also
 Dr. Gugale Pritesh Ramanlal M.Sc., Ph.D., B.Ed., D.M.L.T. Email id - pritesh.gugale09@gmail.com Contact numbernumber- 8446475310 Eukaryotic cells are essentially two envelope systems. Nuclear materials
Dr. Gugale Pritesh Ramanlal M.Sc., Ph.D., B.Ed., D.M.L.T. Email id - pritesh.gugale09@gmail.com Contact numbernumber- 8446475310 Eukaryotic cells are essentially two envelope systems. Nuclear materials
Published Online: 25 November, 1956 Supp Info: on November 16, 2018 jcb.rupress.org Downloaded from
 Published Online: 25 November, 1956 Supp Info: http://doi.org/10.1083/jcb.2.6.799 Downloaded from jcb.rupress.org on November 16, 2018 B~IEF NOrmS 799 Permanganate--A New Fixative for Electron Microscopy.*
Published Online: 25 November, 1956 Supp Info: http://doi.org/10.1083/jcb.2.6.799 Downloaded from jcb.rupress.org on November 16, 2018 B~IEF NOrmS 799 Permanganate--A New Fixative for Electron Microscopy.*
Comparison of infrastructure of Germinating Pea Leaves Prepared by High-Pressure Freezing-Freeze Substitution and Conventional Chemical Fixation
 / Electron Mlcrosc 44: 104-109 (1995) Comparison of infrastructure of Germinating Pea Leaves Prepared by High-Pressure Freezing-Freeze Substitution and Conventional Chemical Fixation Yasuko Kaneko and
/ Electron Mlcrosc 44: 104-109 (1995) Comparison of infrastructure of Germinating Pea Leaves Prepared by High-Pressure Freezing-Freeze Substitution and Conventional Chemical Fixation Yasuko Kaneko and
THE APPEARANCE OF CARBOHYDRATE- RICH MATERIAL IN THE DEVELOPING GOLGI APPARATUS OF AMOEBAE
 J. CellSci. 47, 55-63(1981) 55 Printed in Great Britain Company of Biologists Limited 1981 THE APPEARANCE OF CARBOHYDRATE- RICH MATERIAL IN THE DEVELOPING GOLGI APPARATUS OF AMOEBAE CHARLES J. FLICKINGER
J. CellSci. 47, 55-63(1981) 55 Printed in Great Britain Company of Biologists Limited 1981 THE APPEARANCE OF CARBOHYDRATE- RICH MATERIAL IN THE DEVELOPING GOLGI APPARATUS OF AMOEBAE CHARLES J. FLICKINGER
The Microscopic World of Cells. The Microscopic World of Cells. The Microscopic World of Cells 9/21/2012
 Organisms are either: Single-celled, such as most prokaryotes and protists or Multicelled, such as plants, animals, and most fungi How do we study cells? Light microscopes can be used to explore the structures
Organisms are either: Single-celled, such as most prokaryotes and protists or Multicelled, such as plants, animals, and most fungi How do we study cells? Light microscopes can be used to explore the structures
Skeletal muscle. General features :
 Muscular tissues In the first embryonic life the muscular tissues arise from mesoderm, The function of movement in multicellular organisms is usually assumed by specialized cells called muscle fibers which
Muscular tissues In the first embryonic life the muscular tissues arise from mesoderm, The function of movement in multicellular organisms is usually assumed by specialized cells called muscle fibers which
Some Observations on the Fine Structure of the Goblet Cells. Special Reference to the Well-Developed Agranular Endoplasmic Reticulum
 Okajimas Folia Anat. Jpn., 58(4-6) : 583-594, March 1982 Some Observations on the Fine Structure of the Goblet Cells in the Nasal Respiratory Epithelium of the Rat, with Special Reference to the Well-Developed
Okajimas Folia Anat. Jpn., 58(4-6) : 583-594, March 1982 Some Observations on the Fine Structure of the Goblet Cells in the Nasal Respiratory Epithelium of the Rat, with Special Reference to the Well-Developed
SBI3U7 Cell Structure & Organelles. 2.2 Prokaryotic Cells 2.3 Eukaryotic Cells
 SBI3U7 Cell Structure & Organelles 2.2 Prokaryotic Cells 2.3 Eukaryotic Cells No nucleus Prokaryotic Cells No membrane bound organelles Has a nucleus Eukaryotic Cells Membrane bound organelles Unicellular
SBI3U7 Cell Structure & Organelles 2.2 Prokaryotic Cells 2.3 Eukaryotic Cells No nucleus Prokaryotic Cells No membrane bound organelles Has a nucleus Eukaryotic Cells Membrane bound organelles Unicellular
Fig. LPS in Gram negative bacteria
 Structure of bacterial cell Dentistry college - first class Medical biology- Lec.3 Lecturer D. Hanan S A- Cell wall ***Chemical composition of the cell wall Bacteria are divided into two separated groups
Structure of bacterial cell Dentistry college - first class Medical biology- Lec.3 Lecturer D. Hanan S A- Cell wall ***Chemical composition of the cell wall Bacteria are divided into two separated groups
STUDIES OF THE HUMAN UNFERTILIZED TUBAL OVUM*t
 FERTILITY AND STERILITY Copyright @ 1973 by The Williams & Wilkins Co. Vol. 24, No.8, August 1973 Printed in U.S.A. STUDIES OF THE HUMAN UNFERTILIZED TUBAL OVUM*t C. NORIEGA, M.D., AND C. OBERTI, M.D.
FERTILITY AND STERILITY Copyright @ 1973 by The Williams & Wilkins Co. Vol. 24, No.8, August 1973 Printed in U.S.A. STUDIES OF THE HUMAN UNFERTILIZED TUBAL OVUM*t C. NORIEGA, M.D., AND C. OBERTI, M.D.
Cytosol the fluid Cytoplasm cell interior, everything outside the nucleus but within the cell membrane, includes the organelles, cytosol, and
 Cell Organelles Plasma Membrane comprised of a phospholipid bilayer and embedded proteins Outer surface has oligosaccharides separates the cells s contents from its surroundings Cytosol the fluid Cytoplasm
Cell Organelles Plasma Membrane comprised of a phospholipid bilayer and embedded proteins Outer surface has oligosaccharides separates the cells s contents from its surroundings Cytosol the fluid Cytoplasm
BALLISTICS IN CERTAIN ASCOMYCETES
 BALLISTICS IN CERTAIN ASCOMYCETES BY C. T. INGOLD Birkbeck College, University of London (Received 5 January 1961) (With 3 figures in the te.xt) SUMMARY Horizontal spore discharge has been studied in the
BALLISTICS IN CERTAIN ASCOMYCETES BY C. T. INGOLD Birkbeck College, University of London (Received 5 January 1961) (With 3 figures in the te.xt) SUMMARY Horizontal spore discharge has been studied in the
EDUCATIONAL COMMENTARY MORPHOLOGIC ABNORMALITIES IN LEUKOCYTES
 EDUCATIONAL COMMENTARY MORPHOLOGIC ABNORMALITIES IN LEUKOCYTES Educational commentary is provided through our affiliation with the American Society for Clinical Pathology (ASCP). To obtain FREE CME/CMLE
EDUCATIONAL COMMENTARY MORPHOLOGIC ABNORMALITIES IN LEUKOCYTES Educational commentary is provided through our affiliation with the American Society for Clinical Pathology (ASCP). To obtain FREE CME/CMLE
Supplemental Data. Wang et al. (2013). Plant Cell /tpc
. Plant Cell /tpc") Supplemental Data. Wang et al. (2013). Plant Cell 10.1105/tpc.112.108993 Supplemental Figure 1. 3-MA Treatment Reduces the Growth of Seedlings. Two-week-old Nicotiana benthamiana seedlings germinated on
Supplemental Data. Wang et al. (2013). Plant Cell 10.1105/tpc.112.108993 Supplemental Figure 1. 3-MA Treatment Reduces the Growth of Seedlings. Two-week-old Nicotiana benthamiana seedlings germinated on
Glycogen Aggregates in Cardiac Muscle Cell: A Cytopathological Study on Endomyocardial Biopsies
 Arch. histol. jap., Vol. 45, No. 4 (1982) p. 347-354 Glycogen Aggregates in Cardiac Muscle Cell: A Cytopathological Study on Endomyocardial Biopsies Kazumasa MIURA, Tohru IZUMI, Junichi FUKUDA, Masaru
Arch. histol. jap., Vol. 45, No. 4 (1982) p. 347-354 Glycogen Aggregates in Cardiac Muscle Cell: A Cytopathological Study on Endomyocardial Biopsies Kazumasa MIURA, Tohru IZUMI, Junichi FUKUDA, Masaru
Ultrastructural Study of Human Natural Killer CNK) Cell*)
 Cell*)") Hiroshima Journal of Medical Sciences Vol. 31, No. 1, March, 1982 HJIM 31-6 31 Ultrastructural Study of Human Natural Killer CNK) Cell*) Yoshinori KAWAGUCHI, Eishi KITTAKA, Yoshito TANAKA, Takeo TANAKA
Hiroshima Journal of Medical Sciences Vol. 31, No. 1, March, 1982 HJIM 31-6 31 Ultrastructural Study of Human Natural Killer CNK) Cell*) Yoshinori KAWAGUCHI, Eishi KITTAKA, Yoshito TANAKA, Takeo TANAKA
Microbiology: A Systems Approach
 Microbiology: A Systems Approach First Edition Cowan & Talaro Chapter 4 Prokaryotic Profiles: the Bacteria and the Archaea Chapter 4 Fig. 4.1 3 3 parts flagella filament long, thin, helical structure composed
Microbiology: A Systems Approach First Edition Cowan & Talaro Chapter 4 Prokaryotic Profiles: the Bacteria and the Archaea Chapter 4 Fig. 4.1 3 3 parts flagella filament long, thin, helical structure composed
FIXATION BY MEANS OF GLUTARALDEHYDE-HYDROGEN PEROXIDE REACTION PRODUCTS
 FIXATION BY MEANS OF GLUTARALDEHYDE-HYDROGEN PEROXIDE REACTION PRODUCTS CAMILLO PERACCHIA and BRANT S. MITTLER. From the Department of Anatomy, Duke University Medical Center, Durham, North Carolina 27706,
FIXATION BY MEANS OF GLUTARALDEHYDE-HYDROGEN PEROXIDE REACTION PRODUCTS CAMILLO PERACCHIA and BRANT S. MITTLER. From the Department of Anatomy, Duke University Medical Center, Durham, North Carolina 27706,
Topic 03 Prokaryotes (3.3)
") Topic 03 Prokaryotes (3.3) Topics Characteristics (comparison) External Structures Cell Envelope Internal Structures Cell Shapes, Arrangement, and Sizes Classification 1 Relative size of bacterial cell
Topic 03 Prokaryotes (3.3) Topics Characteristics (comparison) External Structures Cell Envelope Internal Structures Cell Shapes, Arrangement, and Sizes Classification 1 Relative size of bacterial cell
Two new intertidallignicolous Swampomyces species from Red Sea mangroves in Egypt
 Fungal Diversity Two new intertidallignicolous Swampomyces species from Red Sea mangroves in Egypt M.A. Abdel-Wahab1, H. El-Sharouneyl and E.B.G. Jones2 IDepat1ment of Botany, South Valley University,
Fungal Diversity Two new intertidallignicolous Swampomyces species from Red Sea mangroves in Egypt M.A. Abdel-Wahab1, H. El-Sharouneyl and E.B.G. Jones2 IDepat1ment of Botany, South Valley University,
Yara shwabkeh. Osama Alkhader. Heba Kalbouneh
 2 Yara shwabkeh Osama Alkhader Heba Kalbouneh CELL OVERVIEW -Note ; the important thing is to know how the organelles appear under the microscope - the stains we usually use in Histology are composed of
2 Yara shwabkeh Osama Alkhader Heba Kalbouneh CELL OVERVIEW -Note ; the important thing is to know how the organelles appear under the microscope - the stains we usually use in Histology are composed of
A Tour of the Cell. reference: Chapter 6. Reference: Chapter 2
 A Tour of the Cell reference: Chapter 6 Reference: Chapter 2 Monkey Fibroblast Cells stained with fluorescent dyes to show the nucleus (blue) and cytoskeleton (yellow and red fibers), image courtesy of
A Tour of the Cell reference: Chapter 6 Reference: Chapter 2 Monkey Fibroblast Cells stained with fluorescent dyes to show the nucleus (blue) and cytoskeleton (yellow and red fibers), image courtesy of
Chapter 2 Cell. Zhou Li Prof. Dept. of Histology and Embryology
 Chapter 2 Cell Zhou Li Prof. Dept. of Histology and Embryology The inner life of the cell Ⅰ. Plasma membrane (Plasmalemma) 1.1 The structure Unit membrane: inner layer 3-layered structure outer layer mediat
Chapter 2 Cell Zhou Li Prof. Dept. of Histology and Embryology The inner life of the cell Ⅰ. Plasma membrane (Plasmalemma) 1.1 The structure Unit membrane: inner layer 3-layered structure outer layer mediat
10/13/11. Cell Theory. Cell Structure
 Cell Structure Grade 12 Biology Cell Theory All organisms are composed of one or more cells. Cells are the smallest living units of all living organisms. Cells arise only by division of a previously existing
Cell Structure Grade 12 Biology Cell Theory All organisms are composed of one or more cells. Cells are the smallest living units of all living organisms. Cells arise only by division of a previously existing
Further Observations on the Structure of Influenza Viruses A and C
 J. gen. ViroL (I969), 4, 365-370 With 2 plates Printed in Great Britain 365 Further Observations on the Structure of Influenza Viruses A and C By K. APOSTOLOV The Wellcome Research Laboratories, Beckenham,
J. gen. ViroL (I969), 4, 365-370 With 2 plates Printed in Great Britain 365 Further Observations on the Structure of Influenza Viruses A and C By K. APOSTOLOV The Wellcome Research Laboratories, Beckenham,
Ch 4. Functional Anatomy of Prokaryotic and Eukaryotic Cells
 Ch 4 Functional Anatomy of Prokaryotic and Eukaryotic Cells Objectives Compare and contrast the overall cell structure of prokaryotes and eukaryotes. Identify the three basic shapes of bacteria. Describe
Ch 4 Functional Anatomy of Prokaryotic and Eukaryotic Cells Objectives Compare and contrast the overall cell structure of prokaryotes and eukaryotes. Identify the three basic shapes of bacteria. Describe
Preliminary and Short Report
 Tuz JOURNAL OF INvRSrIOATIVE DERMATOLOOT Copyright 1965 by The Williams & Wilkins Co. Vol. 45, No. 3 Printed in U.S.A. Preliminary and Short Report ULTRASTRUCTTJRE OF BLASTOSPORES OF CANDIDA ALBICANS AFTER
Tuz JOURNAL OF INvRSrIOATIVE DERMATOLOOT Copyright 1965 by The Williams & Wilkins Co. Vol. 45, No. 3 Printed in U.S.A. Preliminary and Short Report ULTRASTRUCTTJRE OF BLASTOSPORES OF CANDIDA ALBICANS AFTER
ELECTRON MICROSCOPIC STUDY OF THE FORMATION OF BLUETONGUE VIRUS*
 Onderstepoort J. vet. Res. (1968), 35 (1), 139-150 Printed in the Repub. of S. Afr. by The Government Printer, Pretoria ELECTRON MICROSCOPIC STUDY OF THE FORMATION OF BLUETONGUE VIRUS* G. LECATSAS, Veterinary
Onderstepoort J. vet. Res. (1968), 35 (1), 139-150 Printed in the Repub. of S. Afr. by The Government Printer, Pretoria ELECTRON MICROSCOPIC STUDY OF THE FORMATION OF BLUETONGUE VIRUS* G. LECATSAS, Veterinary
TRANSFER OF PREMELANOSOMES INTO THE KERATINIZING CELLS OF ALBINO HAIR FOLLICLE
 TRANSFER OF PREMELANOSOMES INTO THE KERATINIZING CELLS OF ALBINO HAIR FOLLICLE PAUL F. PARAKKAL. From the Department of Dermatology, Boston University School of Medicine, Boston, Massachusetts 02118 INTRODUCTION
TRANSFER OF PREMELANOSOMES INTO THE KERATINIZING CELLS OF ALBINO HAIR FOLLICLE PAUL F. PARAKKAL. From the Department of Dermatology, Boston University School of Medicine, Boston, Massachusetts 02118 INTRODUCTION
BONE LABORATORY DEMONSTRATIONS. These demonstrations are found on the bulletin boards outside the MCO Bookstore.
 BONE LABORATORY DEMONSTRATIONS These demonstrations are found on the bulletin boards outside the MCO Bookstore. COMPACT & TRABECULAR BONE - LM When viewed under the polarizing light microscope, the layering
BONE LABORATORY DEMONSTRATIONS These demonstrations are found on the bulletin boards outside the MCO Bookstore. COMPACT & TRABECULAR BONE - LM When viewed under the polarizing light microscope, the layering
The Cytoplasm Li Shulei Department of Histology & Embryology
 The Cytoplasm Li Shulei lishulei@tom.com Department of Histology & Embryology Cell components Cytoplasm Plasma membrane Organelles Cytoplasmic deposits Cytoskeleton Cytosol ( Matrix ) Nucleus Plasma membrane
The Cytoplasm Li Shulei lishulei@tom.com Department of Histology & Embryology Cell components Cytoplasm Plasma membrane Organelles Cytoplasmic deposits Cytoskeleton Cytosol ( Matrix ) Nucleus Plasma membrane
Yara Saddam. Amr Alkhatib. Ihsan
 1 Yara Saddam Amr Alkhatib Ihsan NOTE: Yellow highlighting=correction/addition to the previous version of the sheet. Histology (micro anatomy) :- the study of tissues and how they are arranged into organs.
1 Yara Saddam Amr Alkhatib Ihsan NOTE: Yellow highlighting=correction/addition to the previous version of the sheet. Histology (micro anatomy) :- the study of tissues and how they are arranged into organs.
Electron Microscope Studies of HeLa Cells Infected with Herpes Virus
 244 STOKER, M. G. P., SMITH, K. M. & Ross, R. W. (1958). J. gen. Microbiol. 19,244-249 Electron Microscope Studies of HeLa Cells Infected with Herpes Virus BY M: G. P. STOKER, K. M. SMITH AND R. W. ROSS
244 STOKER, M. G. P., SMITH, K. M. & Ross, R. W. (1958). J. gen. Microbiol. 19,244-249 Electron Microscope Studies of HeLa Cells Infected with Herpes Virus BY M: G. P. STOKER, K. M. SMITH AND R. W. ROSS
Cell Structure. Present in animal cell. Present in plant cell. Organelle. Function. strength, resist pressure created when water enters
 Cell Structure Though eukaryotic cells contain many organelles, it is important to know which are in plant cells, which are in animal cells and what their functions are. Organelle Present in plant cell
Cell Structure Though eukaryotic cells contain many organelles, it is important to know which are in plant cells, which are in animal cells and what their functions are. Organelle Present in plant cell
Starch grains - excess sugars
 (a) Membrane system - site of light reactions (photosynthesis) - chlorpophyll pigments - enzymes - electron carriers - flattened, fluid-filled sacs (called thylakoids which are stacked to form grana) -
(a) Membrane system - site of light reactions (photosynthesis) - chlorpophyll pigments - enzymes - electron carriers - flattened, fluid-filled sacs (called thylakoids which are stacked to form grana) -
INVESTIGATIVE OPHTHALMOLOGY. Corneal and conjunctival changes in dysproteinemia
 August 1969 Volume 8, Number 4 INVESTIGATIVE OPHTHALMOLOGY Corneal and conjunctival changes in dysproteinemia 7?. M. H. Pinkerton and David M. Robertson A case of dysproteinemia with corneal and conjunctival
August 1969 Volume 8, Number 4 INVESTIGATIVE OPHTHALMOLOGY Corneal and conjunctival changes in dysproteinemia 7?. M. H. Pinkerton and David M. Robertson A case of dysproteinemia with corneal and conjunctival
Chapter 4. Cartilage and Bone. Li Shu-Lei instructor. Dept. Histology and Embryology, School of Basic Medical Sciences, Jilin University
 Chapter 4 Cartilage and Bone Li Shu-Lei instructor Dept. Histology and Embryology, School of Basic Medical Sciences, Jilin University I Cartilage a specialized connective tissue Characterizers: Cartilage
Chapter 4 Cartilage and Bone Li Shu-Lei instructor Dept. Histology and Embryology, School of Basic Medical Sciences, Jilin University I Cartilage a specialized connective tissue Characterizers: Cartilage
1. General characteristics of muscle tissues: 2. A. Skeletal muscle tissue ("striated muscle tissue")
") 1. General characteristics of muscle tissues: Muscle fibers, AKA, muscle cells Vascularized. Other tissues dense and loose C.T. nerves and nerve fibers Muscle fibers (muscle cells) close together. From
1. General characteristics of muscle tissues: Muscle fibers, AKA, muscle cells Vascularized. Other tissues dense and loose C.T. nerves and nerve fibers Muscle fibers (muscle cells) close together. From
A Tour of the Cell. Chapter 4. Most cells are microscopic. Cells vary in size and shape
 Chapter 4 A Tour of the Cell Most cells are microscopic Cells vary in size and shape 10 m Human height 1 m Length of some nerve and muscle cells 100 mm (10 cm) 10 mm (1 cm) Chicken egg Unaided eye 1 mm
Chapter 4 A Tour of the Cell Most cells are microscopic Cells vary in size and shape 10 m Human height 1 m Length of some nerve and muscle cells 100 mm (10 cm) 10 mm (1 cm) Chicken egg Unaided eye 1 mm
Ch. 6 A Tour of the Cell BIOL 222
 Ch. 6 A Tour of the Cell BIOL 222 Overview: The Fundamental Units of Life All organisms are made of cells The cell is the simplest collec=on of ma>er that can live Cell structure is correlated to cellular
Ch. 6 A Tour of the Cell BIOL 222 Overview: The Fundamental Units of Life All organisms are made of cells The cell is the simplest collec=on of ma>er that can live Cell structure is correlated to cellular
Pathology and Morphogenesis of a Granulosis Virus of the Diamondback Moth
 20 Pathology and Morphogenesis of a Granulosis Virus of the Diamondback Moth Tetsu Asayama Plant Protection Laboratory, Aichi-Ken Agricultural Research Center, Nagakute, Aichi 480-1 1, Japan Abstract In
20 Pathology and Morphogenesis of a Granulosis Virus of the Diamondback Moth Tetsu Asayama Plant Protection Laboratory, Aichi-Ken Agricultural Research Center, Nagakute, Aichi 480-1 1, Japan Abstract In
This work is licensed under a Creative Commons Attribution- NonCommercial-NoDerivatives 4.0 International License.
 Title A new freshwater species of Saccardoella from Hong Kong and South Africa Author(s) Tsui, KM; Hyde, KD; Hodgkiss, IJ; Goh, TK Citation Mycologia, 1998, v. 90 n. 4, p. 701-704 Issued Date 1998 URL
Title A new freshwater species of Saccardoella from Hong Kong and South Africa Author(s) Tsui, KM; Hyde, KD; Hodgkiss, IJ; Goh, TK Citation Mycologia, 1998, v. 90 n. 4, p. 701-704 Issued Date 1998 URL
LOW-ANGLE X-RAY DIFFRACTION AND ELECTRON-MICROSCOPE STUDIES OF ISOLATED CELL MEMBRANES
 J. Cell Sci. I, 287-296 (1966) 287 Printed in Great Britain LOW-ANGLE X-RAY DIFFRACTION AND ELECTRON-MICROSCOPE STUDIES OF ISOLATED CELL MEMBRANES J. B. FINEAN, R. COLEMAN, W. G. GREEN* AND A. R. LIMBRICK
J. Cell Sci. I, 287-296 (1966) 287 Printed in Great Britain LOW-ANGLE X-RAY DIFFRACTION AND ELECTRON-MICROSCOPE STUDIES OF ISOLATED CELL MEMBRANES J. B. FINEAN, R. COLEMAN, W. G. GREEN* AND A. R. LIMBRICK
A Tour of the Cell. Chapter 6. Biology. Edited by Shawn Lester. Inner Life of Cell. Eighth Edition Neil Campbell and Jane Reece
 Chapter 6 A Tour of the Cell Inner Life of Cell Edited by Shawn Lester PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin
Chapter 6 A Tour of the Cell Inner Life of Cell Edited by Shawn Lester PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin
Distribution of the Two Kinds of Myofilaments in Insect Muscles
 AM. ZOOLOCIST, 7:451-456 (1967). Distribution of the Two Kinds of Myofilaments in Insect Muscles JACQUES AUBER Laboratoire de Cytologie, Faculte des Sciences, Paris, et Laboratoire de Microscopie Electronique
AM. ZOOLOCIST, 7:451-456 (1967). Distribution of the Two Kinds of Myofilaments in Insect Muscles JACQUES AUBER Laboratoire de Cytologie, Faculte des Sciences, Paris, et Laboratoire de Microscopie Electronique
CELLS CONTAINING LANGERHANS GRANULES IN HUMAN LYMPH NODES OF DERMATOPATHIC LYMPHADENOPATHY*
 THS JOURNAL OF INVEBTIOATIVR DERMATOLOGY Copyright 1969 by The Williams & Wilkins Co. Vol. 93, No. 4 Printed in U.S.A. CELLS CONTAINING LANGERHANS GRANULES IN HUMAN LYMPH NODES OF DERMATOPATHIC LYMPHADENOPATHY*
THS JOURNAL OF INVEBTIOATIVR DERMATOLOGY Copyright 1969 by The Williams & Wilkins Co. Vol. 93, No. 4 Printed in U.S.A. CELLS CONTAINING LANGERHANS GRANULES IN HUMAN LYMPH NODES OF DERMATOPATHIC LYMPHADENOPATHY*
*Need a summary just not covered yet!
 *Need a summary just not covered yet! Organelle Pupil Organelle Pupil Nucleus KO N Microtubules + centrioles LF Endoplasmic reticulum ED Plant cell wall OC Ribosomes KL Chloroplasts RK Golgi apparatus
*Need a summary just not covered yet! Organelle Pupil Organelle Pupil Nucleus KO N Microtubules + centrioles LF Endoplasmic reticulum ED Plant cell wall OC Ribosomes KL Chloroplasts RK Golgi apparatus
Supplementary Figure 1.
 Supplementary Figure 1. Visualization of endoplasmic reticulum-mitochondria interaction by in situ proximity ligation assay. A) Illustration of targeted proteins in mitochondria (M), endoplasmic reticulum
Supplementary Figure 1. Visualization of endoplasmic reticulum-mitochondria interaction by in situ proximity ligation assay. A) Illustration of targeted proteins in mitochondria (M), endoplasmic reticulum
AET-treated normal red cells (PNH-like cells)
") J. clin. Path., 1971, 24, 677-684 Electron microscope study of PNH red cells and AET-treated normal red cells (PNH-like cells) S. M. LEWIS, G. LAMBERTENGHI, S. FERRONE, AND G. SIRCHIA From the Department
J. clin. Path., 1971, 24, 677-684 Electron microscope study of PNH red cells and AET-treated normal red cells (PNH-like cells) S. M. LEWIS, G. LAMBERTENGHI, S. FERRONE, AND G. SIRCHIA From the Department
Thursday, October 16 th
 Thursday, October 16 th Good morning. Those of you needing to take the Enzymes and Energy Quiz will start very soon. Students who took the quiz Wednesday: Please QUIETLY work on the chapter 6 reading guide.
Thursday, October 16 th Good morning. Those of you needing to take the Enzymes and Energy Quiz will start very soon. Students who took the quiz Wednesday: Please QUIETLY work on the chapter 6 reading guide.
Plasma Membrane. comprised of a phospholipid bilayer and embedded proteins separates the cells s contents from its surroundings
 Cell Organelles Plasma Membrane comprised of a phospholipid bilayer and embedded proteins separates the cells s contents from its surroundings Cytosol the fluid Cytoplasm cell interior, everything outside
Cell Organelles Plasma Membrane comprised of a phospholipid bilayer and embedded proteins separates the cells s contents from its surroundings Cytosol the fluid Cytoplasm cell interior, everything outside
Cell Structure and Function. Biology 12 Unit 1 Cell Structure and Function Inquiry into Life pages and 68-69
 Cell Structure and Function Biology 12 Unit 1 Cell Structure and Function Inquiry into Life pages 45 59 and 68-69 Assignments for this Unit Pick up the notes/worksheet for this unit and the project There
Cell Structure and Function Biology 12 Unit 1 Cell Structure and Function Inquiry into Life pages 45 59 and 68-69 Assignments for this Unit Pick up the notes/worksheet for this unit and the project There
Ultrastructure of Azotobacter vinelandii
 JOURNAL OF BACTERIOLoGY, Nov. 1970, p. 933-939 Vol. 104, No. 2 Copyright a 1970 American Society for Microbiology Printed in U.S.A. Ultrastructure of Azotobacter vinelandii G. R. VELA, G. D. CAGLE, AND
JOURNAL OF BACTERIOLoGY, Nov. 1970, p. 933-939 Vol. 104, No. 2 Copyright a 1970 American Society for Microbiology Printed in U.S.A. Ultrastructure of Azotobacter vinelandii G. R. VELA, G. D. CAGLE, AND
Notes on the Effects of Pretreatment of Spores on the Iodinophile Vacuole of Henneguya exilisl
 Proceedings of the Iowa Academy of Science Volume 76 Annual Issue Article 57 1969 Notes on the Effects of Pretreatment of Spores on the Iodinophile Vacuole of Henneguya exilisl Dennis Galinsky Drake University
Proceedings of the Iowa Academy of Science Volume 76 Annual Issue Article 57 1969 Notes on the Effects of Pretreatment of Spores on the Iodinophile Vacuole of Henneguya exilisl Dennis Galinsky Drake University
:1c.c :& Preliminary and Short Report GRANULE FORMATION IN THE LANGERHANS CELL* structure with rounded ends and a striated lamella
 THE JOURNAL OF INVESTIGATIVE DERMATOLOGY Copyright 1566 by The Williams & Wilkins Co. Vol. 7, No. 5 Printed in U.S.A. Preliminary and Short Report GRANULE FORMATION IN THE LANGERHANS CELL* ALVIN S. ZELICKSON,
THE JOURNAL OF INVESTIGATIVE DERMATOLOGY Copyright 1566 by The Williams & Wilkins Co. Vol. 7, No. 5 Printed in U.S.A. Preliminary and Short Report GRANULE FORMATION IN THE LANGERHANS CELL* ALVIN S. ZELICKSON,
Early scientists who observed cells made detailed sketches of what they saw.
 Early scientists who observed cells made detailed sketches of what they saw. Early scientists who observed cells made detailed sketches of what they saw. CORK Early scientists who observed cells made detailed
Early scientists who observed cells made detailed sketches of what they saw. Early scientists who observed cells made detailed sketches of what they saw. CORK Early scientists who observed cells made detailed
An electron microscope study of the tegument and associated structures of Dipylidium Caninum. By L. T. THREADGOLD
 An electron microscope study of the tegument and associated structures of Dipylidium Caninum By L. T. THREADGOLD (From the Department of Anatomy, Dalhousie University, Halifax, Nova Scotia, Canada. Present
An electron microscope study of the tegument and associated structures of Dipylidium Caninum By L. T. THREADGOLD (From the Department of Anatomy, Dalhousie University, Halifax, Nova Scotia, Canada. Present
Human height. Length of some nerve and muscle cells. Chicken egg. Frog egg. Most plant and animal cells Nucleus Most bacteria Mitochondrion
 10 m 1 m 0.1 m 1 cm Human height Length of some nerve and muscle cells Chicken egg Unaided eye 1 mm Frog egg 100 µm 10 µm 1 µm 100 nm 10 nm Most plant and animal cells Nucleus Most bacteria Mitochondrion
10 m 1 m 0.1 m 1 cm Human height Length of some nerve and muscle cells Chicken egg Unaided eye 1 mm Frog egg 100 µm 10 µm 1 µm 100 nm 10 nm Most plant and animal cells Nucleus Most bacteria Mitochondrion
Laboratory diagnosis of parasitic diseases. (Amoebiasis)
") Laboratory diagnosis of parasitic diseases (Amoebiasis) Sarah Alharbi Clinical Laboratory department Collage of Applied Medical Sciences King Saud University This document contains materials modified or
Laboratory diagnosis of parasitic diseases (Amoebiasis) Sarah Alharbi Clinical Laboratory department Collage of Applied Medical Sciences King Saud University This document contains materials modified or
Don t Freak Out. Test on cell organelle on Friday!
 Cell Structure 1 Don t Freak Out Test on cell organelle on Friday! This test should be a buffer test and help raise your overall test score. All information will come from this week! 2 Cells Provide Compartments
Cell Structure 1 Don t Freak Out Test on cell organelle on Friday! This test should be a buffer test and help raise your overall test score. All information will come from this week! 2 Cells Provide Compartments
Microscopic Anatomy of Inferior Medullary Velum Of Cerebellum
 32 J Anat. Soc. India 51(1) 32-34 (2002) Microscopic Anatomy of Of Cerebellum Arora, N.K. Department of Anatomy, Government Medical College, Chandigarh INDIA. Abstract. A study of the inferior medullary
32 J Anat. Soc. India 51(1) 32-34 (2002) Microscopic Anatomy of Of Cerebellum Arora, N.K. Department of Anatomy, Government Medical College, Chandigarh INDIA. Abstract. A study of the inferior medullary
Acta Medica Okayama. Electron microscopic demonstration of a new virus isolated from a patient with SMON. Zensuke Ota DECEMBER 1970
 Acta Medica Okayama Volume 24, Issue 6 1970 Article 3 DECEMBER 1970 Electron microscopic demonstration of a new virus isolated from a patient with SMON Zensuke Ota Okayama University, Copyright c 1999
Acta Medica Okayama Volume 24, Issue 6 1970 Article 3 DECEMBER 1970 Electron microscopic demonstration of a new virus isolated from a patient with SMON Zensuke Ota Okayama University, Copyright c 1999
Observations on Erysiphe platani in Germany
 Observations on Erysiphe platani in Germany Kirschner R* Department of Life Sciences, National Central University, Jhongli City, 320 Taoyuan County, Taiwan R.O.C. Kirschner R. 2011 Observations on Erysiphe
Observations on Erysiphe platani in Germany Kirschner R* Department of Life Sciences, National Central University, Jhongli City, 320 Taoyuan County, Taiwan R.O.C. Kirschner R. 2011 Observations on Erysiphe
The fine structure of the cortical layers in Paramecium aurelia
 129 The fine structure of the cortical layers in Paramecium aurelia By JOHN M. STEWART and ALAN R. MUIR (From the Department of Natural History, University of Aberdeen, and the Department of Anatomy, University
129 The fine structure of the cortical layers in Paramecium aurelia By JOHN M. STEWART and ALAN R. MUIR (From the Department of Natural History, University of Aberdeen, and the Department of Anatomy, University