SUPPLEMENTARY INFORMATION
|
|
|
- Bertram Murphy
- 5 years ago
- Views:
Transcription

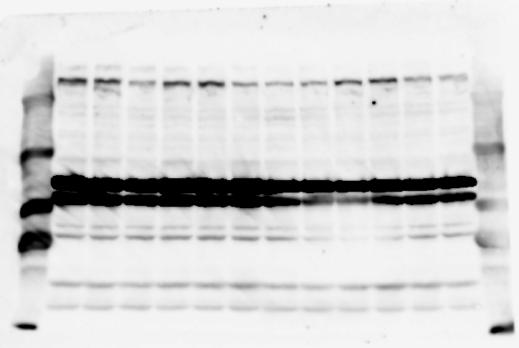

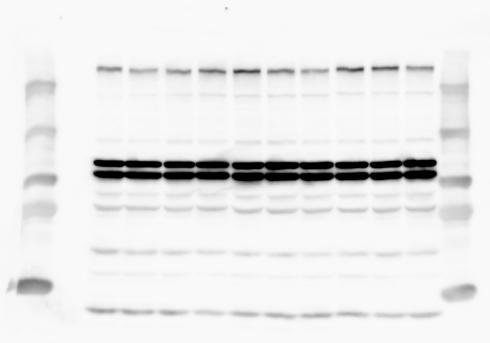

1 SUPPLEMENTARY INFORMATION doi: /nature19357 Figure 1a Chd8 +/+ Chd8 +/ΔSL Chd8 +/+ Chd8 +/ΔL E10.5_Whole brain E10.5_Whole brain E10.5_Whole brain E14.5_Whole brain E14.5_Whole brain E14.5_Whole brain E18.5_Whole brain E18.5_Whole brain E18.5_Whole brain E18.5_Whole brain 1
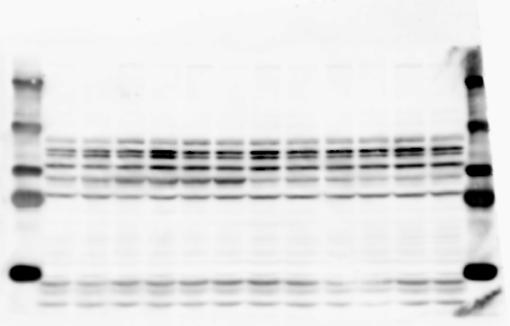
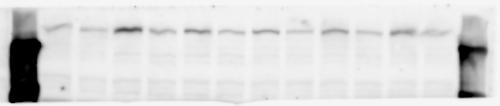
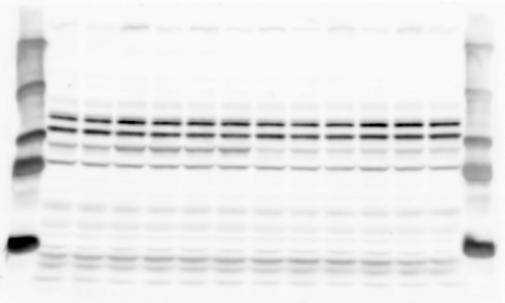
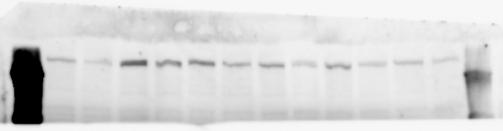
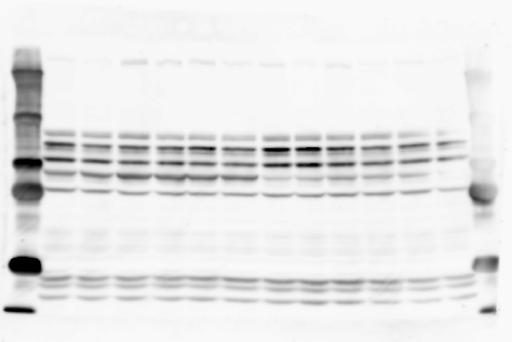
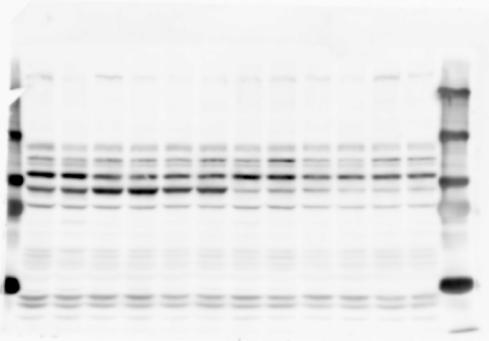

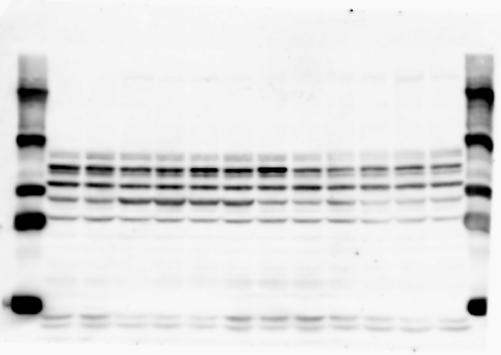
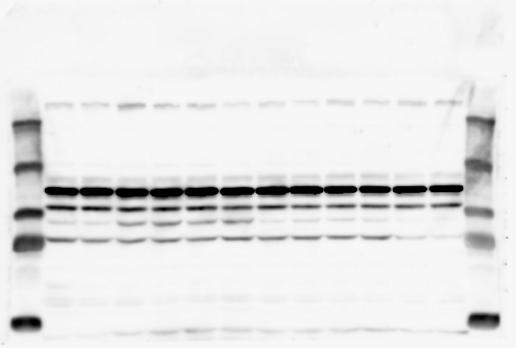
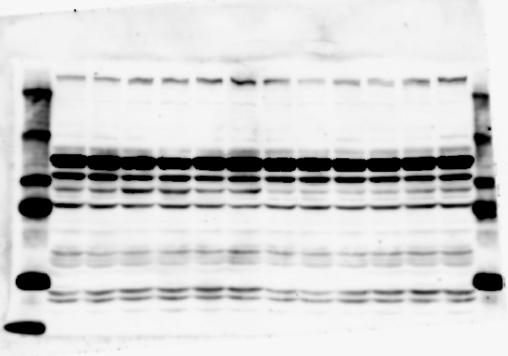
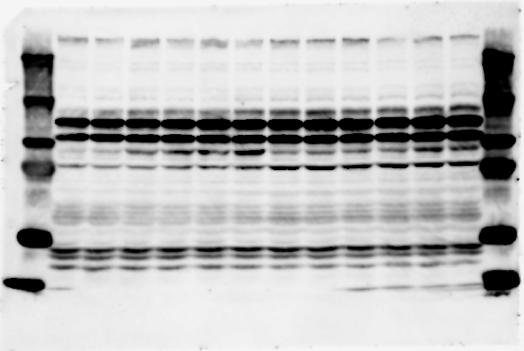

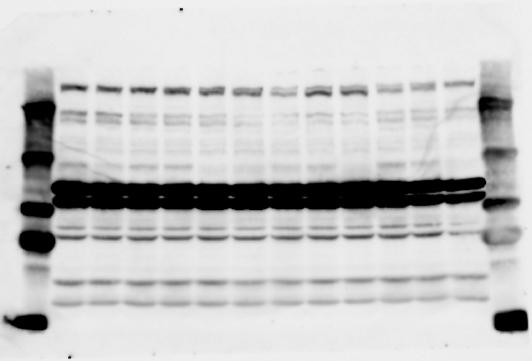
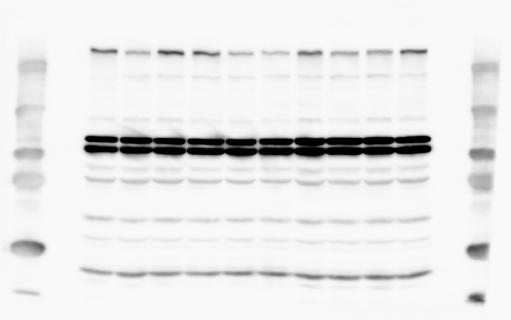
2 RESEARCH SUPPLEMENTARY INFORMATION Figure 1a Chd8 +/+ Chd8 +/ΔSL Chd8 +/+ Chd8 +/ΔL OB CTX HIP STR TH HY MB CB P 2
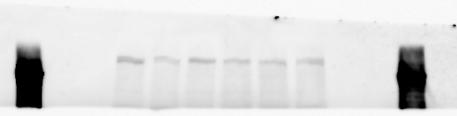
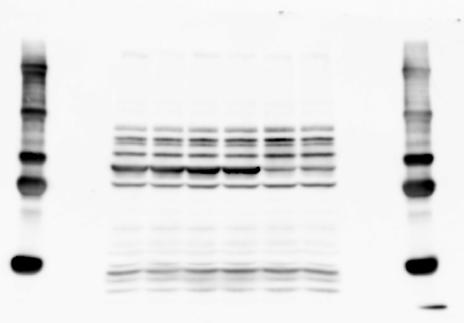
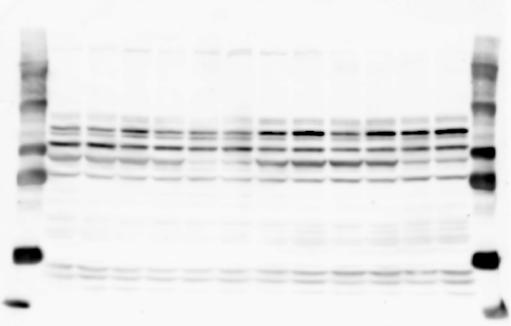
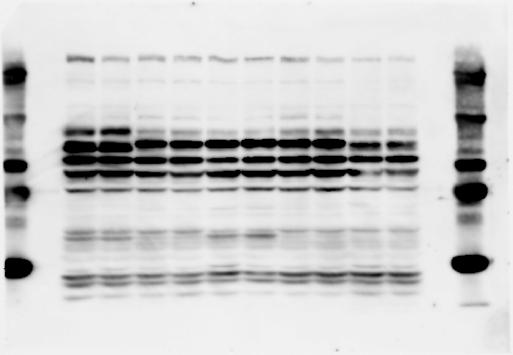
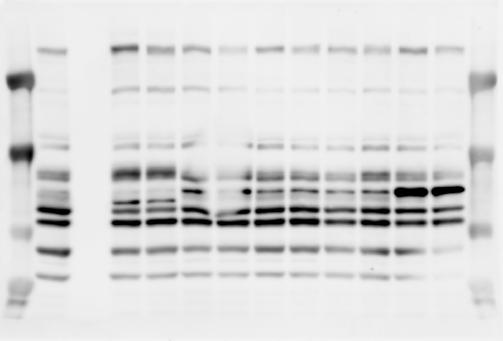

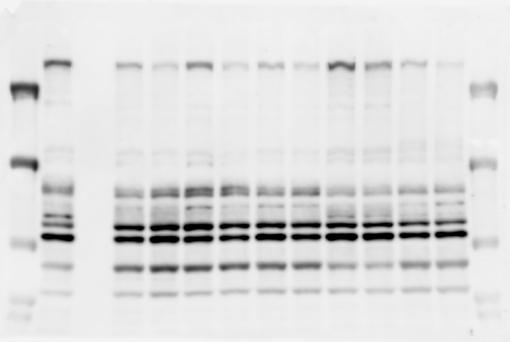
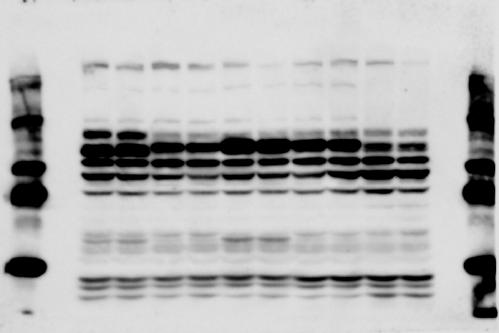
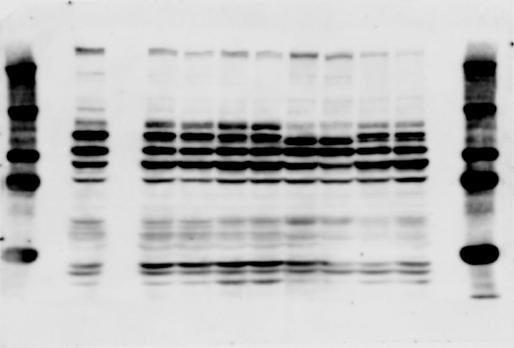
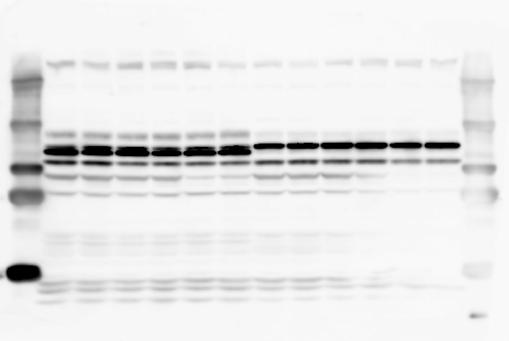
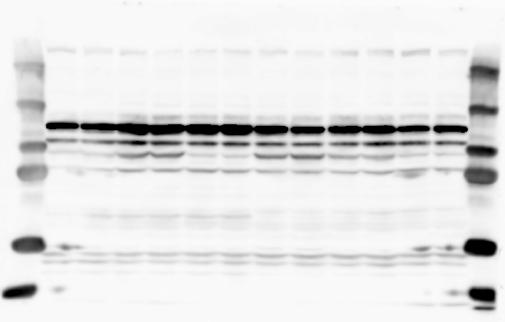
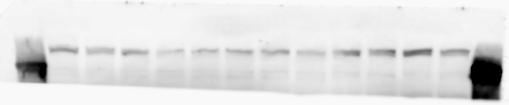
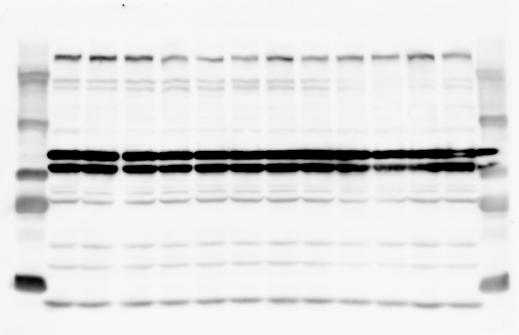
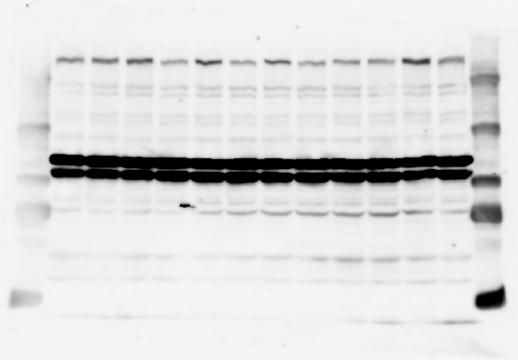
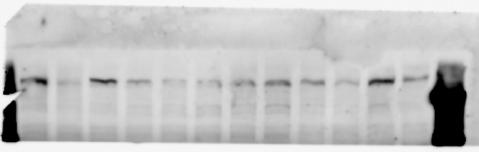
3 SUPPLEMENTARY INFORMATION RESEARCH Katayama et al. Supplementary Figure 1 +/+ Chd8 Figure 1a OB CTX HIP STR +/ΔSL Chd8 OB CTX HIP +/+ Chd8 STR +/ΔL Chd8 TH OB HIP STR TH CTX TH HY MB CB P HY MB CB P OB HIP CTX CB HY MB TH STR P CB MB HY P W W W. N A T U R E. C O M / N A T U R E 3

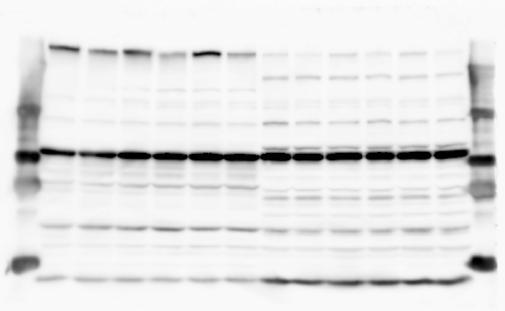
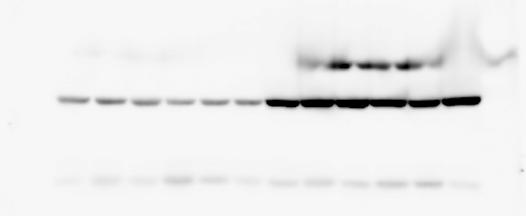
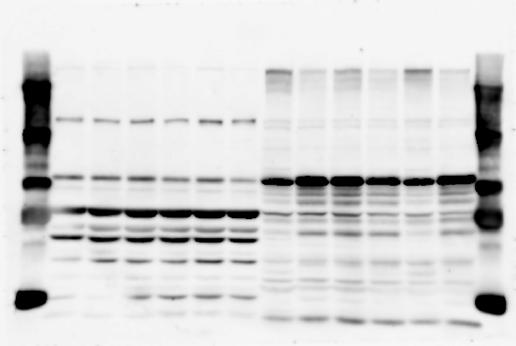

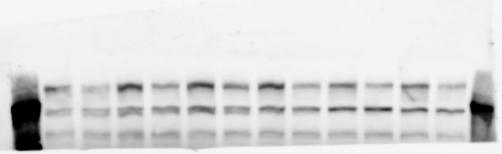
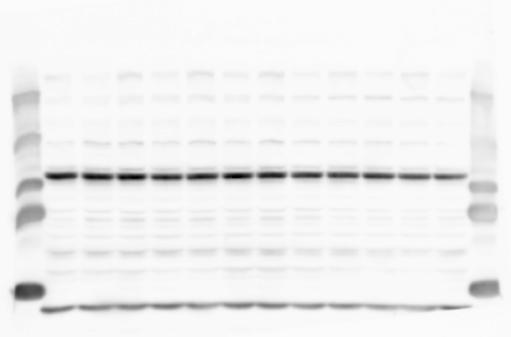
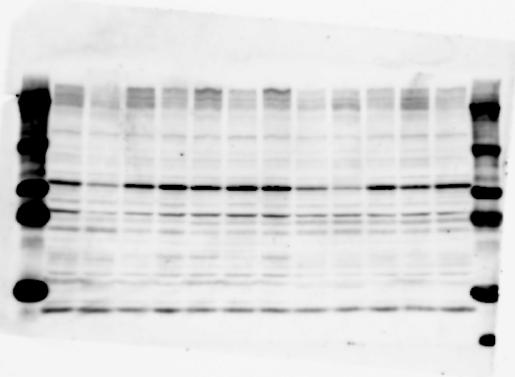
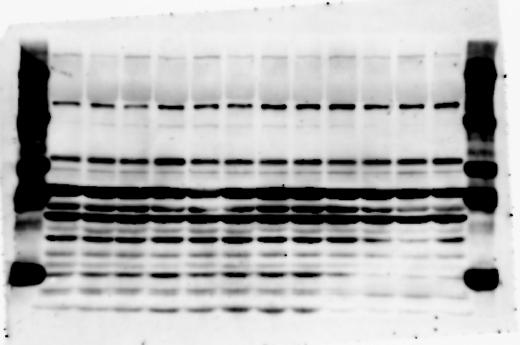
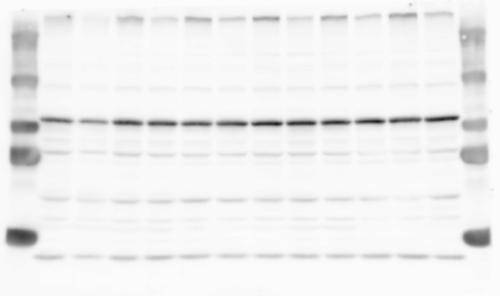
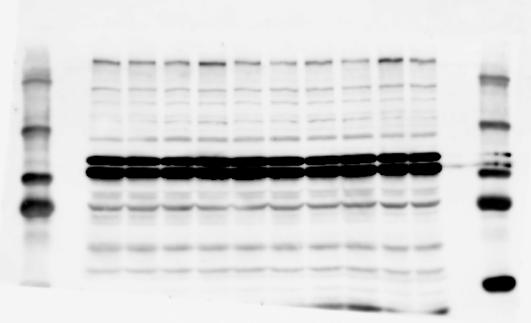
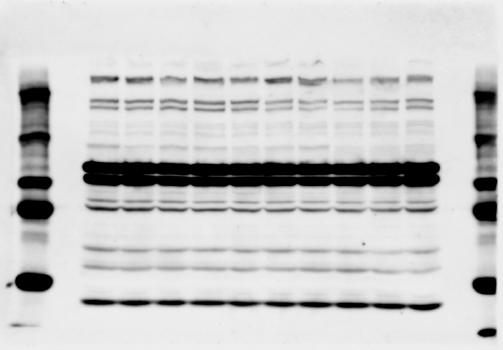
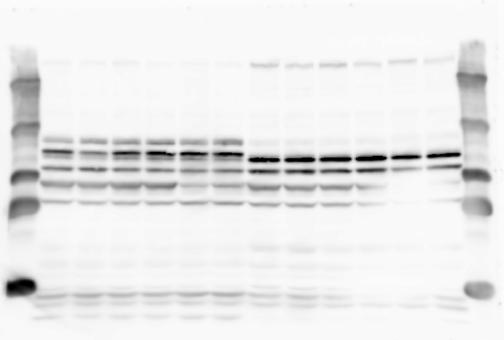
4 RESEARCH SUPPLEMENTARY INFORMATION Extended Data Figure 2e, f Chd8 +/+ Chd8 +/ΔSL Chd8 +/+ Chd8 +/ΔL Heart Thymus Spleen Testis Heart Spleen Thymus Testis 4












: 0 0.")
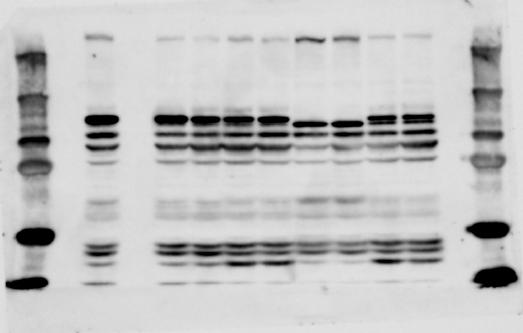
5 SUPPLEMENTARY INFORMATION RESEARCH Figure 4e Extended Data Figure 1f IP IgG Anti- Input Chd8+/+ Chd8+/ΔL Chd8+/+ Chd8+/ΔL Chd8+/+ Chd8+/ΔL E10.5 Whole brain E14.5 Whole brain E18.5 Whole brain Adult Olfactory bulb Chd8+/+ Chd8+/ΔL REST Extended Data Figure 6a CHD8 antibodies : A A Chd8 genotype : Chd8+/+ Chd8L F/F A A Chd8+/+ Chd8L F/F Anti- mab Chd8+/+ Chd8L F/F Anti-CHD8 pab Chd8+/+ Chd8L F/F Extended Data Figure 1c 4-OHT (1 μm) : kb Chd8+/+ Chd8+/ΔL Wild-type Deletion 3 Extended Data Figure 10f Extended Data Figure 10h Input IP: FLAG Chd8 +/+ Chd8 +/ΔL Chd8 ΔL/ΔL Extended Data Figure 1d Chd8+/+ CAG-CreER T2 /Chd8+/+ Chd8L F/F CAG-CreER T2 /Chd8L F/F FLAG : - FOXP1 - FOXP1 REST CHX (h): REST FOXP
6 RESEARCH SUPPLEMENTARY INFORMATION Supplementary Discussion Antibody specificity We used rabbit polyclonal antibodies to pan-chd8 (anti-chd8 pab) for immunoblot analysis and a rat monoclonal antibody to CHD8 L (anti-chd8 L mab) for ChIP and immunoprecipitation in the present study. We examined the specificity of these antibodies generated in-house by immunoblot and ChIP analyses with lysates prepared from WT or CHD8 L -null MEFs (the latter of which comprised CAG-CreER T2 /Chd8 F/F L MEFs treated with 4-hydroxytamoxifen [4-OHT]). The signal corresponding to CHD8 L was not detected in the CHD8 L -null MEFs by immunoblot analysis with either anti-chd8 pab or anti-chd8 L mab (Extended Data Fig. 6a). ChIP with anti-chd8 L mab also revealed that CHD8 L binding to target genes identified in previous studies, including Pten, Nras, and Rac1, was greatly diminished in CHD8 L -null MEFs (Extended Data Fig. 6b). These results together show that our anti-chd8 L mab is highly specific for CHD8 L, and they suggest that the data obtained with this antibody in the present study are reliable. Unexpectedly, ChIP signals with one of the antibody preparations (A A, Bethyl Laboratories) used in previous studies 20,46 were not diminished in CHD8 L -null cells in our experiments (Extended Data Fig. 6b), suggesting that the results of ChIP analysis with this antibody preparation may include nonspecific artifacts. CHD8 L and CHD8 S expression We examined the abundance of CHD8 L and CHD8 S isoforms at both mrna and protein levels in tissues of E10.5, E14.5, E18.5, and adult Chd8 +/+, Chd8 +/ SL, and Chd8 +/ L mice including various brain regions as well as tissues such as heart, thymus, spleen, and testis in which CHD8 is relatively abundant in adult mice 12 (Fig. 1a and Extended Data Fig. 2). In most cases, CHD8 L protein levels in the whole brain or brain regions of Chd8 +/ SL and Chd8 +/ L mice were ~70% of those in WT mice (Fig. 1a), whereas this value was ~% for the other tissues examined (Extended Data Fig. 2e). The amounts of Chd8 L and Chd8 S 6
7 SUPPLEMENTARY INFORMATION RESEARCH mrnas were reduced by ~% in all tissues and at all developmental stages in the corresponding mutant mice (Extended Data Fig. 2a d), suggesting that the dosage compensation for CHD8 L haploinsufficiency in the brain occurs at a posttranscriptional level. We did not detect shorter proteins that were present only in the Chd8 +/ L brain (Extended Data Fig. 1f) or in CHD8 L -null MEFs (Extended Data Fig. 6a), suggesting that any such truncated forms of CHD8 produced from the mutant allele are degraded immediately after translation or otherwise unstable. Unfortunately, our in-house rabbit polyclonal antibodies to pan-chd8 (anti-chd8 pab) detected a nonspecific band with an electrophoretic mobility similar to that of CHD8 S in mouse brain (unpublished results), making it technically difficult to quantify CHD8 S protein and therefore to determine the stoichiometry of CHD8 L and CHD8 S isoforms in the brain. Of note, the abundance of Chd8 S mrna was up-regulated in the brain at certain developmental stages and in most regions of the adult brain in Chd8 +/ L mice (Extended Data Fig. 2b), suggesting that CHD8 might suppress its own gene promoter in a negative feedback loop. This moderate increase in the abundance of Chd8 S mrna in the brain might result in a corresponding increase in the amount of CHD8 S protein in Chd8 +/ L mice. Furthermore, it is difficult to experimentally exclude the possibility that such a compensatory up-regulation of CHD8 S protein might affect the ASD-like phenotypes of Chd8 +/ L mice. However, given that the results of the behavioural tests were similar for Chd8 +/ SL and Chd8 +/ L mice (Fig. 1c g, Fig. 2, and Extended Data Fig. 4) and that many mutations in the human CHD8 gene are localized in the region specific to the CHD8 L isoform (Extended Data Fig. 1a), it is likely that the abundance of CHD8 L protein, rather than that of CHD8 S, is an important determinant of ASD phenotypes. The presence of the CHD8 S isoform thus does not appear to substantially affect ASD phenotypes in human or mouse. Presence of the CHD8 S isoform in humans In our previous study 12, immunoblot analysis with our in-house anti-chd8 pab revealed the presence of the CHD8 S isoform in HEK293T, U2OS, SaOS2, HeLa, and HCT116 cell lines, all of which are derived from human. Indeed, we recently isolated a cdna encoding CHD8 S 7
8 RESEARCH SUPPLEMENTARY INFORMATION from human-derived cells (unpublished results). We have no evidence for an interaction between the long and short protein products. Although we cannot exclude the possibility that CHD8 S might function in a dominant negative manner, this appears unlikely given the absence of substantial phenotypic differences between Chd8 +/ SL and Chd8 +/ L mice. Genes whose expression is altered in Chd8 mutant mice We found only a few genes whose expression level was changed by a factor of >2 or <0.5 and with a P value of <0.05 in the brain of Chd8 mutant mice, and even these genes only just fulfilled these criteria (Fig. 3c and Extended Data Fig. 9b f). Such genes include those for noncoding RNAs and a pseudogene and have no known connection to neural function (Extended Data Fig. 9g). It is therefore unlikely that the changes in the expression of such genes contribute to the ASD-like phenotypes of Chd8 mutant mice. CHD8 haploinsufficiency thus does not appear to result in prominent changes in the expression of a few specific genes but rather gives rise to small but global changes in gene expression in mouse brain, reminiscent of the brain of human ASD patients. CHD8 binding pattern in Chd8 mutant mice Comparison of the ChIP-seq results between WT and Chd8 +/ L mice revealed that the pattern of CHD8 binding to transcription start sites (TSSs) in adult brain did not differ significantly between the two genotypes (Extended Data Fig. 9h) regardless of changes in gene expression (Extended Data Fig. 9i). CHD8 binding thus appears to decrease uniformly in the brain of Chd8 +/ L mice with no change in the binding pattern. The specific changes in gene expression apparent in the brain of Chd8 +/ L mice are thus not likely attributable to changes in the pattern of CHD8 binding. Instead, the sensitivity to CHD8 haploinsufficiency may differ among genes. Our results suggest that many REST target genes are included in such genes whose expression is sensitive to CHD8 haploinsufficiency. Of course, REST target genes that do not associate with CHD8 are not influenced by CHD8 haploinsufficiency (Fig. 4f). 8
9 SUPPLEMENTARY INFORMATION RESEARCH Comparison of our results with those of previous studies We compared our ChIP-seq results with those of previous studies 20,21,46 with regard to the degree of overlap in CHD8 binding peaks (Extended Data Fig. 8a) and in CHD8 target genes (Extended Data Fig. 8b e). Our data were found to overlap with those of the previous studies to some extent. Given that the samples and antibodies used differ among these various studies, some level of inconsistency in results is to be expected. As mentioned above with regard to antibody specificity, we found that ChIP signals obtained with one of the anti-chd8 preparations used in two of the previous studies 20,46 were not diminished in CHD8-null cells, suggesting that the results of ChIP experiments with these antibodies may include nonspecific artifacts. We also compared our RNA-seq results with those of the previous studies 20,21. We found that the gene set for REST was markedly affected in the brain of Chd8 mutant mice and of human ASD patients (Fig. 4a, b, d), whereas such a decrease in REST target gene expression was not detected by GSEA with the results of these two previous studies 20,21 (Extended Data Fig. 10b, c). This difference in the detection of these genes between our study and previous studies might be attributable to the difference in the samples analyzed: We examined Chd8 mutant mouse brain (30% to % reduction in CHD8 protein abundance), whereas the other groups used neural stem cells differentiated from human induced pluripotent stem cells (ipscs) and subjected to RNA interference mediated knockdown of CHD8 (70% to 90% reduction in CHD8 protein abundance). The relation between CHD8 and REST is thus revealed only in our study. REST expression in hetmt mice GSEA with Hallmark gene sets revealed that the Wnt/ -catenin pathway appeared to be slightly activated in the brain of Chd8 +/ L mice only at E14.5 (P = 0.043) (Extended Data Fig. 9k). REST was previously shown to be activated by the Wnt signaling pathway 47. Indeed, we found that the amount of Rest mrna tended to be increased in the brain of hetmt mice at E14.5 (P = for Chd8 +/ SL mice and P = for Chd8 +/ L mice) (Extended Data Fig. 9
10 RESEARCH SUPPLEMENTARY INFORMATION 10d), consistent with the Wnt activation apparent at this stage. Given that REST was shown to increase its own expression through the action of the microrna mir-9 in a positive feedback loop 48,49, this small increase in the amount of Rest mrna might reflect a secondary effect of REST activation by CHD8 haploinsufficiency. Mechanistic insight It will be important to explore the mechanistic connection between changes in gene expression profiles and the behavioural abnormalities in our Chd8 mutant mice. We found that CHD8 physically binds to REST and suppresses its function. The importance of the CHD8-REST axis in ASD etiology is supported by two additional observations. First, GSEA revealed that expression of REST target genes was down-regulated in the brain of human ASD patients (Fig. 4d), as seen in our mutant mice (Fig. 4b). Second, expression of REST target genes that bind CHD8 was down-regulated in the brain of Chd8 +/ L mice compared with WT mice, whereas that of such genes that do not bind CHD8 did not show such enrichment (Fig. 4f). These results thus suggest that the CHD8-REST axis plays a key role in ASD pathogenesis. There are four possible mechanisms by which CHD8 might interfere with REST: (1) It inhibits REST activity; (2) it induces dissociation of REST from target genes; (3) it attenuates the expression of Rest; and (4) it promotes REST degradation. With regard to possibility (2), genome-wide ChIP-seq analysis for REST with the brain of WT and CHD8-haploinsufficient mouse embryos at E14.5 revealed no significant difference in REST binding to the genes with or without bound CHD8 between the two genotypes (Extended Data Fig. 10g). These results thus indicate that CHD8 does not prevent or reverse the binding of REST to its target genes. With regard to possibility (3), the amount of Rest mrna was slightly increased in the brain of the heterozygous mutant mice at E14.5 as mentioned above. Regardless of this finding, however, ChIP-seq data showing that the amount of REST bound to its target genes was unchanged in Chd8 heterozygous mutant mice (Extended Data Fig. 10g) indicate that the slight increase in Rest mrna abundance is not likely responsible for 10
11 SUPPLEMENTARY INFORMATION RESEARCH the ASD-like phenotypes. With regard to possibility (4), cycloheximide chase analysis revealed that the stability of REST protein did not differ among WT, Chd8 +/ L, and Chd8 L/ L MEFs (Extended Data Fig. 10h), excluding the possibility that CHD8 inhibits REST function by promoting REST degradation. We therefore conclude that CHD8 inhibits REST activity while REST remains bound to its target genes. CHD8 binding might interfere with the formation of a protein complex that usually associates with REST and mediates its suppressor function such as CoREST, Sin3, LSD1, or histone deacetylase 27,28, or CHD8 binding might antagonize the suppressor function of REST either by itself or by recruiting other transcriptional activators such as the MLL histone methyltransferase complex 46,. In general, however, it is technically difficult at present to explore the molecular mechanisms by which a global modulator of gene expression affects a particular phenotype. For example, mutation of MeCP2, one of the most well-studied molecules in the field of ASD-related disease, results in Rett syndrome with an autistic phenotype 51. It remains to be fully understood how MeCP2 mutation gives rise to ASD-like phenotypes, however, with studies to shed light on the mechanistic connection still being in progress 16, Another example is provided by Brg1, one of the most well-characterized chromatin-remodeling factors. Brg1 mutation results in a variety of phenotypes, and altered Brg1 function has been suggested to underlie many human diseases The precise causal relation between Brg1 mutation and associated phenotypes also remains unclear. These examples suggest that chromatin remodelers or epigenetic modifiers with broad specificity might target thousands of genes, and abnormalities resulting from their mutation might be attributable to combined effects on the expression of many of these genes. Such appears to be the case for CHD8. Although we believe that REST activation may be a key causal factor, we do not conclude that REST alone is responsible for ASD etiology. Rather, it is likely that the combination of altered expression of many genes including REST target genes induced by CHD8 haploinsufficiency shapes ASD phenotypes. The discovery of the physical and functional interaction between CHD8 and REST may prove to be important in itself, however, given that REST is a master regulator of neurogenesis. Indeed, GSEA revealed that 11
12 RESEARCH SUPPLEMENTARY INFORMATION expression of REST target genes is down-regulated in the brain of human ASD patients (Fig. 4d), as it is in our mutant mice (Fig. 4b). Deregulated activation of REST resulting from CHD8 haploinsufficiency might thus give rise to neurodevelopmental delay and the consequent development of ASD phenotypes. Identification of the CHD8-REST axis may therefore provide important insight into the pathogenesis of ASD as well as serve as the basis for development of new treatments for this substantial medical and social problem. 12
13 SUPPLEMENTARY INFORMATION RESEARCH Supplementary references 47 Lu, T. et al. REST and stress resistance in ageing and Alzheimer's disease. Nature 7, (2014). 48 Packer, A. N., Xing, Y., Harper, S. Q., Jones, L. & Davidson, B. L. The bifunctional microrna mir-9/mir-9* regulates REST and CoREST and is downregulated in Huntington's disease. J. Neurosci. 28, (2008). 49 Laneve, P. et al. A minicircuitry involving REST and CREB controls mir-9-2 expression during human neuronal differentiation. Nucleic Acids Res. 38, (2010). Dou, Y. et al. Physical association and coordinate function of the H3 K4 methyltransferase MLL1 and the H4 K16 acetyltransferase MOF. Cell 121, (2005). 51 Chahrour, M. & Zoghbi, H. Y. The story of Rett syndrome: from clinic to neurobiology. Neuron 56, (2007). 52 Shahbazian, M. et al. Mice with truncated MeCP2 recapitulate many Rett syndrome features and display hyperacetylation of histone H3. Neuron 35, (2002). 53 Guy, J., Gan, J., Selfridge, J., Cobb, S. & Bird, A. Reversal of neurological defects in a mouse model of Rett syndrome. Science 315, (2007). 54 Gabel, H. W. et al. Disruption of DNA-methylation-dependent long gene repression in Rett syndrome. Nature 522, (2015). 55 Sztainberg, Y. et al. Reversal of phenotypes in MECP2 duplication mice using genetic rescue or antisense oligonucleotides. Nature 528, (2015). 56 Wilson, B. G. & Roberts, C. W. SWI/SNF nucleosome remodellers and cancer. Nat. Rev. Cancer 11, (2011). 57 Hang, C. T. et al. Chromatin regulation by Brg1 underlies heart muscle development and disease. Nature 466, (2010). 58 Zhang, Z. et al. Autism-associated chromatin regulator Brg1/SmarcA4 is required for synapse development and myocyte rnhancer factor 2-mediated dynapse remodeling. Mol. Cell. Biol. 36, (2015). 59 Ho, L. et al. An embryonic stem cell chromatin remodeling complex, esbaf, is an essential component of the core pluripotency transcriptional network. Proc. Natl. Acad. Sci. U. S. A. 106, (2009). 13
Epigenetics: The Future of Psychology & Neuroscience. Richard E. Brown Psychology Department Dalhousie University Halifax, NS, B3H 4J1
 Epigenetics: The Future of Psychology & Neuroscience Richard E. Brown Psychology Department Dalhousie University Halifax, NS, B3H 4J1 Nature versus Nurture Despite the belief that the Nature vs. Nurture
Epigenetics: The Future of Psychology & Neuroscience Richard E. Brown Psychology Department Dalhousie University Halifax, NS, B3H 4J1 Nature versus Nurture Despite the belief that the Nature vs. Nurture
SUPPLEMENTARY INFORMATION
 Supplementary Discussion The cell cycle machinery and the DNA damage response network are highly interconnected and co-regulated in assuring faithful duplication and partition of genetic materials into
Supplementary Discussion The cell cycle machinery and the DNA damage response network are highly interconnected and co-regulated in assuring faithful duplication and partition of genetic materials into
Is Intrinsic Hyperexcitability in CA3 the Culprit for Seizures in Rett Syndrome?
 Current Literature In Basic Science Is Intrinsic Hyperexcitability in CA3 the Culprit for Seizures in Rett Syndrome? Network Hyperexcitability in Hippocampal Slices From Mecp2 Mutant Mice Revealed by Voltage-Sensitive
Current Literature In Basic Science Is Intrinsic Hyperexcitability in CA3 the Culprit for Seizures in Rett Syndrome? Network Hyperexcitability in Hippocampal Slices From Mecp2 Mutant Mice Revealed by Voltage-Sensitive
Gene Regulation Part 2
 Michael Cummings Chapter 9 Gene Regulation Part 2 David Reisman University of South Carolina Other topics in Chp 9 Part 2 Protein folding diseases Most diseases are caused by mutations in the DNA that
Michael Cummings Chapter 9 Gene Regulation Part 2 David Reisman University of South Carolina Other topics in Chp 9 Part 2 Protein folding diseases Most diseases are caused by mutations in the DNA that
Eukaryotic transcription (III)
") Eukaryotic transcription (III) 1. Chromosome and chromatin structure Chromatin, chromatid, and chromosome chromatin Genomes exist as chromatins before or after cell division (interphase) but as chromatids
Eukaryotic transcription (III) 1. Chromosome and chromatin structure Chromatin, chromatid, and chromosome chromatin Genomes exist as chromatins before or after cell division (interphase) but as chromatids
Supplementary Figure 1. Efficiency of Mll4 deletion and its effect on T cell populations in the periphery. Nature Immunology: doi: /ni.
 Supplementary Figure 1 Efficiency of Mll4 deletion and its effect on T cell populations in the periphery. Expression of Mll4 floxed alleles (16-19) in naive CD4 + T cells isolated from lymph nodes and
Supplementary Figure 1 Efficiency of Mll4 deletion and its effect on T cell populations in the periphery. Expression of Mll4 floxed alleles (16-19) in naive CD4 + T cells isolated from lymph nodes and
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature12652 Supplementary Figure 1. PRDM16 interacts with endogenous EHMT1 in brown adipocytes. Immunoprecipitation of PRDM16 complex by flag antibody (M2) followed by Western blot analysis
doi:10.1038/nature12652 Supplementary Figure 1. PRDM16 interacts with endogenous EHMT1 in brown adipocytes. Immunoprecipitation of PRDM16 complex by flag antibody (M2) followed by Western blot analysis
Comparison of open chromatin regions between dentate granule cells and other tissues and neural cell types.
 Supplementary Figure 1 Comparison of open chromatin regions between dentate granule cells and other tissues and neural cell types. (a) Pearson correlation heatmap among open chromatin profiles of different
Supplementary Figure 1 Comparison of open chromatin regions between dentate granule cells and other tissues and neural cell types. (a) Pearson correlation heatmap among open chromatin profiles of different
The Biology and Genetics of Cells and Organisms The Biology of Cancer
 The Biology and Genetics of Cells and Organisms The Biology of Cancer Mendel and Genetics How many distinct genes are present in the genomes of mammals? - 21,000 for human. - Genetic information is carried
The Biology and Genetics of Cells and Organisms The Biology of Cancer Mendel and Genetics How many distinct genes are present in the genomes of mammals? - 21,000 for human. - Genetic information is carried
Not IN Our Genes - A Different Kind of Inheritance.! Christopher Phiel, Ph.D. University of Colorado Denver Mini-STEM School February 4, 2014
 Not IN Our Genes - A Different Kind of Inheritance! Christopher Phiel, Ph.D. University of Colorado Denver Mini-STEM School February 4, 2014 Epigenetics in Mainstream Media Epigenetics *Current definition:
Not IN Our Genes - A Different Kind of Inheritance! Christopher Phiel, Ph.D. University of Colorado Denver Mini-STEM School February 4, 2014 Epigenetics in Mainstream Media Epigenetics *Current definition:
SUPPLEMENTARY INFORMATION
 DOI: 10.1038/ncb2566 Figure S1 CDKL5 protein expression pattern and localization in mouse brain. (a) Multiple-tissue western blot from a postnatal day (P) 21 mouse probed with an antibody against CDKL5.
DOI: 10.1038/ncb2566 Figure S1 CDKL5 protein expression pattern and localization in mouse brain. (a) Multiple-tissue western blot from a postnatal day (P) 21 mouse probed with an antibody against CDKL5.
The Role of Smoking in Cocaine. Addiction
 The Role of Smoking in Cocaine Addiction Luca Colnaghi Eric Kandel Laboratory Columbia University in the City of New York Department of Neuroscience Index 1- The Brain, memory, metaplasticity 2- Cocaine
The Role of Smoking in Cocaine Addiction Luca Colnaghi Eric Kandel Laboratory Columbia University in the City of New York Department of Neuroscience Index 1- The Brain, memory, metaplasticity 2- Cocaine
Marta Puerto Plasencia. microrna sponges
 Marta Puerto Plasencia microrna sponges Introduction microrna CircularRNA Publications Conclusions The most well-studied regions in the human genome belong to proteincoding genes. Coding exons are 1.5%
Marta Puerto Plasencia microrna sponges Introduction microrna CircularRNA Publications Conclusions The most well-studied regions in the human genome belong to proteincoding genes. Coding exons are 1.5%
Supplementary Figure 1 (Related with Figure 4). Molecular consequences of Eed deletion. (a) ChIP analysis identifies 3925 genes that are associated
. Molecular consequences of Eed deletion. (a) ChIP analysis identifies 3925 genes that are associated") Supplementary Figure 1 (Related with Figure 4). Molecular consequences of Eed deletion. (a) ChIP analysis identifies 3925 genes that are associated with the H3K27me3 mark in chondrocytes (see Table S1,
Supplementary Figure 1 (Related with Figure 4). Molecular consequences of Eed deletion. (a) ChIP analysis identifies 3925 genes that are associated with the H3K27me3 mark in chondrocytes (see Table S1,
Analysis of Massively Parallel Sequencing Data Application of Illumina Sequencing to the Genetics of Human Cancers
 Analysis of Massively Parallel Sequencing Data Application of Illumina Sequencing to the Genetics of Human Cancers Gordon Blackshields Senior Bioinformatician Source BioScience 1 To Cancer Genetics Studies
Analysis of Massively Parallel Sequencing Data Application of Illumina Sequencing to the Genetics of Human Cancers Gordon Blackshields Senior Bioinformatician Source BioScience 1 To Cancer Genetics Studies
BIO360 Fall 2013 Quiz 1
 BIO360 Fall 2013 Quiz 1 1. Examine the diagram below. There are two homologous copies of chromosome one and the allele of YFG carried on the light gray chromosome has undergone a loss-of-function mutation.
BIO360 Fall 2013 Quiz 1 1. Examine the diagram below. There are two homologous copies of chromosome one and the allele of YFG carried on the light gray chromosome has undergone a loss-of-function mutation.
Supplementary Figure S1: Defective heterochromatin repair in HGPS progeroid cells
 Supplementary Figure S1: Defective heterochromatin repair in HGPS progeroid cells Immunofluorescence staining of H3K9me3 and 53BP1 in PH and HGADFN003 (HG003) cells at 24 h after γ-irradiation. Scale bar,
Supplementary Figure S1: Defective heterochromatin repair in HGPS progeroid cells Immunofluorescence staining of H3K9me3 and 53BP1 in PH and HGADFN003 (HG003) cells at 24 h after γ-irradiation. Scale bar,
SUPPLEMENTARY FIGURES
 SUPPLEMENTARY FIGURES 1 Supplementary Figure 1, Adult hippocampal QNPs and TAPs uniformly express REST a-b) Confocal images of adult hippocampal mouse sections showing GFAP (green), Sox2 (red), and REST
SUPPLEMENTARY FIGURES 1 Supplementary Figure 1, Adult hippocampal QNPs and TAPs uniformly express REST a-b) Confocal images of adult hippocampal mouse sections showing GFAP (green), Sox2 (red), and REST
Prokaryotes and eukaryotes alter gene expression in response to their changing environment
 Chapter 18 Prokaryotes and eukaryotes alter gene expression in response to their changing environment In multicellular eukaryotes, gene expression regulates development and is responsible for differences
Chapter 18 Prokaryotes and eukaryotes alter gene expression in response to their changing environment In multicellular eukaryotes, gene expression regulates development and is responsible for differences
Raymond Auerbach PhD Candidate, Yale University Gerstein and Snyder Labs August 30, 2012
 Elucidating Transcriptional Regulation at Multiple Scales Using High-Throughput Sequencing, Data Integration, and Computational Methods Raymond Auerbach PhD Candidate, Yale University Gerstein and Snyder
Elucidating Transcriptional Regulation at Multiple Scales Using High-Throughput Sequencing, Data Integration, and Computational Methods Raymond Auerbach PhD Candidate, Yale University Gerstein and Snyder
Histones modifications and variants
 Histones modifications and variants Dr. Institute of Molecular Biology, Johannes Gutenberg University, Mainz www.imb.de Lecture Objectives 1. Chromatin structure and function Chromatin and cell state Nucleosome
Histones modifications and variants Dr. Institute of Molecular Biology, Johannes Gutenberg University, Mainz www.imb.de Lecture Objectives 1. Chromatin structure and function Chromatin and cell state Nucleosome
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature12215 Supplementary Figure 1. The effects of full and dissociated GR agonists in supporting BFU-E self-renewal divisions. BFU-Es were cultured in self-renewal medium with indicated GR
doi:10.1038/nature12215 Supplementary Figure 1. The effects of full and dissociated GR agonists in supporting BFU-E self-renewal divisions. BFU-Es were cultured in self-renewal medium with indicated GR
(A) SW480, DLD1, RKO and HCT116 cells were treated with DMSO or XAV939 (5 µm)
 SW480, DLD1, RKO and HCT116 cells were treated with DMSO or XAV939 (5 µm)") Supplementary Figure Legends Figure S1. Tankyrase inhibition suppresses cell proliferation in an axin/β-catenin independent manner. (A) SW480, DLD1, RKO and HCT116 cells were treated with DMSO or XAV939
Supplementary Figure Legends Figure S1. Tankyrase inhibition suppresses cell proliferation in an axin/β-catenin independent manner. (A) SW480, DLD1, RKO and HCT116 cells were treated with DMSO or XAV939
Systems Analysis Of Chromatin-Related Protein Complexes In Cancer READ ONLINE
 Systems Analysis Of Chromatin-Related Protein Complexes In Cancer READ ONLINE If looking for the book Systems Analysis of Chromatin-Related Protein Complexes in Cancer in pdf format, then you have come
Systems Analysis Of Chromatin-Related Protein Complexes In Cancer READ ONLINE If looking for the book Systems Analysis of Chromatin-Related Protein Complexes in Cancer in pdf format, then you have come
Epigenetics. Lyle Armstrong. UJ Taylor & Francis Group. f'ci Garland Science NEW YORK AND LONDON
 ... Epigenetics Lyle Armstrong f'ci Garland Science UJ Taylor & Francis Group NEW YORK AND LONDON Contents CHAPTER 1 INTRODUCTION TO 3.2 CHROMATIN ARCHITECTURE 21 THE STUDY OF EPIGENETICS 1.1 THE CORE
... Epigenetics Lyle Armstrong f'ci Garland Science UJ Taylor & Francis Group NEW YORK AND LONDON Contents CHAPTER 1 INTRODUCTION TO 3.2 CHROMATIN ARCHITECTURE 21 THE STUDY OF EPIGENETICS 1.1 THE CORE
Transcriptional control in Eukaryotes: (chapter 13 pp276) Chromatin structure affects gene expression. Chromatin Array of nuc
 Chromatin structure affects gene expression. Chromatin Array of nuc") Transcriptional control in Eukaryotes: (chapter 13 pp276) Chromatin structure affects gene expression Chromatin Array of nuc 1 Transcriptional control in Eukaryotes: Chromatin undergoes structural changes
Transcriptional control in Eukaryotes: (chapter 13 pp276) Chromatin structure affects gene expression Chromatin Array of nuc 1 Transcriptional control in Eukaryotes: Chromatin undergoes structural changes
Supplementary information
 Supplementary information Human Cytomegalovirus MicroRNA mir-us4-1 Inhibits CD8 + T Cell Response by Targeting ERAP1 Sungchul Kim, Sanghyun Lee, Jinwook Shin, Youngkyun Kim, Irini Evnouchidou, Donghyun
Supplementary information Human Cytomegalovirus MicroRNA mir-us4-1 Inhibits CD8 + T Cell Response by Targeting ERAP1 Sungchul Kim, Sanghyun Lee, Jinwook Shin, Youngkyun Kim, Irini Evnouchidou, Donghyun
Epigenetic Principles and Mechanisms Underlying Nervous System Function in Health and Disease Mark F. Mehler MD, FAAN
 Epigenetic Principles and Mechanisms Underlying Nervous System Function in Health and Disease Mark F. Mehler MD, FAAN Institute for Brain Disorders and Neural Regeneration F.M. Kirby Program in Neural
Epigenetic Principles and Mechanisms Underlying Nervous System Function in Health and Disease Mark F. Mehler MD, FAAN Institute for Brain Disorders and Neural Regeneration F.M. Kirby Program in Neural
Profiles of gene expression & diagnosis/prognosis of cancer. MCs in Advanced Genetics Ainoa Planas Riverola
 Profiles of gene expression & diagnosis/prognosis of cancer MCs in Advanced Genetics Ainoa Planas Riverola Gene expression profiles Gene expression profiling Used in molecular biology, it measures the
Profiles of gene expression & diagnosis/prognosis of cancer MCs in Advanced Genetics Ainoa Planas Riverola Gene expression profiles Gene expression profiling Used in molecular biology, it measures the
Nature Immunology: doi: /ni Supplementary Figure 1. Characteristics of SEs in T reg and T conv cells.
 Supplementary Figure 1 Characteristics of SEs in T reg and T conv cells. (a) Patterns of indicated transcription factor-binding at SEs and surrounding regions in T reg and T conv cells. Average normalized
Supplementary Figure 1 Characteristics of SEs in T reg and T conv cells. (a) Patterns of indicated transcription factor-binding at SEs and surrounding regions in T reg and T conv cells. Average normalized
BIOL2005 WORKSHEET 2008
 BIOL2005 WORKSHEET 2008 Answer all 6 questions in the space provided using additional sheets where necessary. Hand your completed answers in to the Biology office by 3 p.m. Friday 8th February. 1. Your
BIOL2005 WORKSHEET 2008 Answer all 6 questions in the space provided using additional sheets where necessary. Hand your completed answers in to the Biology office by 3 p.m. Friday 8th February. 1. Your
Accessing and Using ENCODE Data Dr. Peggy J. Farnham
 1 William M Keck Professor of Biochemistry Keck School of Medicine University of Southern California How many human genes are encoded in our 3x10 9 bp? C. elegans (worm) 959 cells and 1x10 8 bp 20,000
1 William M Keck Professor of Biochemistry Keck School of Medicine University of Southern California How many human genes are encoded in our 3x10 9 bp? C. elegans (worm) 959 cells and 1x10 8 bp 20,000
p53 cooperates with DNA methylation and a suicidal interferon response to maintain epigenetic silencing of repeats and noncoding RNAs
 p53 cooperates with DNA methylation and a suicidal interferon response to maintain epigenetic silencing of repeats and noncoding RNAs 2013, Katerina I. Leonova et al. Kolmogorov Mikhail Noncoding DNA Mammalian
p53 cooperates with DNA methylation and a suicidal interferon response to maintain epigenetic silencing of repeats and noncoding RNAs 2013, Katerina I. Leonova et al. Kolmogorov Mikhail Noncoding DNA Mammalian
Supplementary Figure 1.TRIM33 binds β-catenin in the nucleus. a & b, Co-IP of endogenous TRIM33 with β-catenin in HT-29 cells (a) and HEK 293T cells
 and HEK 293T cells") Supplementary Figure 1.TRIM33 binds β-catenin in the nucleus. a & b, Co-IP of endogenous TRIM33 with β-catenin in HT-29 cells (a) and HEK 293T cells (b). TRIM33 was immunoprecipitated, and the amount of
Supplementary Figure 1.TRIM33 binds β-catenin in the nucleus. a & b, Co-IP of endogenous TRIM33 with β-catenin in HT-29 cells (a) and HEK 293T cells (b). TRIM33 was immunoprecipitated, and the amount of
Lysine methyltransferase SMYD2 promotes cyst growth in autosomal dominant polycystic kidney disease
 Lysine methyltransferase SMYD2 promotes cyst growth in autosomal dominant polycystic kidney disease Linda Xiaoyan Li,, Julien Sage, Xiaogang Li J Clin Invest. 2017;127(7):2751-2764. https://doi.org/10.1172/jci90921.
Lysine methyltransferase SMYD2 promotes cyst growth in autosomal dominant polycystic kidney disease Linda Xiaoyan Li,, Julien Sage, Xiaogang Li J Clin Invest. 2017;127(7):2751-2764. https://doi.org/10.1172/jci90921.
SUPPLEMENTAL FIGURE LEGENDS
 SUPPLEMENTAL FIGURE LEGENDS Supplemental Figure S1: Endogenous interaction between RNF2 and H2AX: Whole cell extracts from 293T were subjected to immunoprecipitation with anti-rnf2 or anti-γ-h2ax antibodies
SUPPLEMENTAL FIGURE LEGENDS Supplemental Figure S1: Endogenous interaction between RNF2 and H2AX: Whole cell extracts from 293T were subjected to immunoprecipitation with anti-rnf2 or anti-γ-h2ax antibodies
MicroRNA-mediated incoherent feedforward motifs are robust
 The Second International Symposium on Optimization and Systems Biology (OSB 8) Lijiang, China, October 31 November 3, 8 Copyright 8 ORSC & APORC, pp. 62 67 MicroRNA-mediated incoherent feedforward motifs
The Second International Symposium on Optimization and Systems Biology (OSB 8) Lijiang, China, October 31 November 3, 8 Copyright 8 ORSC & APORC, pp. 62 67 MicroRNA-mediated incoherent feedforward motifs
Computational Analysis of UHT Sequences Histone modifications, CAGE, RNA-Seq
 Computational Analysis of UHT Sequences Histone modifications, CAGE, RNA-Seq Philipp Bucher Wednesday January 21, 2009 SIB graduate school course EPFL, Lausanne ChIP-seq against histone variants: Biological
Computational Analysis of UHT Sequences Histone modifications, CAGE, RNA-Seq Philipp Bucher Wednesday January 21, 2009 SIB graduate school course EPFL, Lausanne ChIP-seq against histone variants: Biological
Soft Agar Assay. For each cell pool, 100,000 cells were resuspended in 0.35% (w/v)
") SUPPLEMENTARY MATERIAL AND METHODS Soft Agar Assay. For each cell pool, 100,000 cells were resuspended in 0.35% (w/v) top agar (LONZA, SeaKem LE Agarose cat.5004) and plated onto 0.5% (w/v) basal agar.
SUPPLEMENTARY MATERIAL AND METHODS Soft Agar Assay. For each cell pool, 100,000 cells were resuspended in 0.35% (w/v) top agar (LONZA, SeaKem LE Agarose cat.5004) and plated onto 0.5% (w/v) basal agar.
Supporting Online Material for
 www.sciencemag.org/cgi/content/full/1171320/dc1 Supporting Online Material for A Frazzled/DCC-Dependent Transcriptional Switch Regulates Midline Axon Guidance Long Yang, David S. Garbe, Greg J. Bashaw*
www.sciencemag.org/cgi/content/full/1171320/dc1 Supporting Online Material for A Frazzled/DCC-Dependent Transcriptional Switch Regulates Midline Axon Guidance Long Yang, David S. Garbe, Greg J. Bashaw*
7SK ChIRP-seq is specifically RNA dependent and conserved between mice and humans.
 Supplementary Figure 1 7SK ChIRP-seq is specifically RNA dependent and conserved between mice and humans. Regions targeted by the Even and Odd ChIRP probes mapped to a secondary structure model 56 of the
Supplementary Figure 1 7SK ChIRP-seq is specifically RNA dependent and conserved between mice and humans. Regions targeted by the Even and Odd ChIRP probes mapped to a secondary structure model 56 of the
Transcriptional and Epigenetic Mechanisms of Addiction
 Transcriptional and Epigenetic Mechanisms of Addiction Eric J. Nestler Mount Sinai School of Medicine New York, NY Dr. Ray Fuller There is every reason to be optimistic that in the future we will find
Transcriptional and Epigenetic Mechanisms of Addiction Eric J. Nestler Mount Sinai School of Medicine New York, NY Dr. Ray Fuller There is every reason to be optimistic that in the future we will find
supplementary information
 DOI: 10.1038/ncb1875 Figure S1 (a) The 79 surgical specimens from NSCLC patients were analysed by immunohistochemistry with an anti-p53 antibody and control serum (data not shown). The normal bronchi served
DOI: 10.1038/ncb1875 Figure S1 (a) The 79 surgical specimens from NSCLC patients were analysed by immunohistochemistry with an anti-p53 antibody and control serum (data not shown). The normal bronchi served
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature10643 Supplementary Table 1. Identification of hecw-1 coding polymorphisms at amino acid positions 322 and 325 in 162 strains of C. elegans. WWW.NATURE.COM/NATURE 1 Supplementary Figure
doi:10.1038/nature10643 Supplementary Table 1. Identification of hecw-1 coding polymorphisms at amino acid positions 322 and 325 in 162 strains of C. elegans. WWW.NATURE.COM/NATURE 1 Supplementary Figure
Overview: Conducting the Genetic Orchestra Prokaryotes and eukaryotes alter gene expression in response to their changing environment
 Overview: Conducting the Genetic Orchestra Prokaryotes and eukaryotes alter gene expression in response to their changing environment In multicellular eukaryotes, gene expression regulates development
Overview: Conducting the Genetic Orchestra Prokaryotes and eukaryotes alter gene expression in response to their changing environment In multicellular eukaryotes, gene expression regulates development
Fragile X Syndrome. Genetics, Epigenetics & the Role of Unprogrammed Events in the expression of a Phenotype
 Fragile X Syndrome Genetics, Epigenetics & the Role of Unprogrammed Events in the expression of a Phenotype A loss of function of the FMR-1 gene results in severe learning problems, intellectual disability
Fragile X Syndrome Genetics, Epigenetics & the Role of Unprogrammed Events in the expression of a Phenotype A loss of function of the FMR-1 gene results in severe learning problems, intellectual disability
Supplementary Information. Preferential associations between co-regulated genes reveal a. transcriptional interactome in erythroid cells
 Supplementary Information Preferential associations between co-regulated genes reveal a transcriptional interactome in erythroid cells Stefan Schoenfelder, * Tom Sexton, * Lyubomira Chakalova, * Nathan
Supplementary Information Preferential associations between co-regulated genes reveal a transcriptional interactome in erythroid cells Stefan Schoenfelder, * Tom Sexton, * Lyubomira Chakalova, * Nathan
Are you the way you are because of the
 EPIGENETICS Are you the way you are because of the It s my fault!! Nurture Genes you inherited from your parents? Nature Experiences during your life? Similar DNA Asthma, Autism, TWINS Bipolar Disorders
EPIGENETICS Are you the way you are because of the It s my fault!! Nurture Genes you inherited from your parents? Nature Experiences during your life? Similar DNA Asthma, Autism, TWINS Bipolar Disorders
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature10495 WWW.NATURE.COM/NATURE 1 2 WWW.NATURE.COM/NATURE WWW.NATURE.COM/NATURE 3 4 WWW.NATURE.COM/NATURE WWW.NATURE.COM/NATURE 5 6 WWW.NATURE.COM/NATURE WWW.NATURE.COM/NATURE 7 8 WWW.NATURE.COM/NATURE
doi:10.1038/nature10495 WWW.NATURE.COM/NATURE 1 2 WWW.NATURE.COM/NATURE WWW.NATURE.COM/NATURE 3 4 WWW.NATURE.COM/NATURE WWW.NATURE.COM/NATURE 5 6 WWW.NATURE.COM/NATURE WWW.NATURE.COM/NATURE 7 8 WWW.NATURE.COM/NATURE
Stem Cell Epigenetics
 Stem Cell Epigenetics Philippe Collas University of Oslo Institute of Basic Medical Sciences Norwegian Center for Stem Cell Research www.collaslab.com Source of stem cells in the body Somatic ( adult )
Stem Cell Epigenetics Philippe Collas University of Oslo Institute of Basic Medical Sciences Norwegian Center for Stem Cell Research www.collaslab.com Source of stem cells in the body Somatic ( adult )
Discovery of Schizophrenia Drug Targets from DISC1 Mechanisms Atsushi Kamiya M.D., Ph.D.
 Discovery of Schizophrenia Drug Targets 1 Assistant Professor Department of Psychiatry and Behavioral Sciences Johns Hopkins University School of Medicine akamiya1@jhmi.edu How does neurobiology offer
Discovery of Schizophrenia Drug Targets 1 Assistant Professor Department of Psychiatry and Behavioral Sciences Johns Hopkins University School of Medicine akamiya1@jhmi.edu How does neurobiology offer
Big brains may hold clues to origins of autism
 VIEWPOINT Big brains may hold clues to origins of autism BY KONSTANTINOS ZARBALIS 23 FEBRUARY 2016 A persistent challenge to improving our understanding of autism is the fact that no single neurological
VIEWPOINT Big brains may hold clues to origins of autism BY KONSTANTINOS ZARBALIS 23 FEBRUARY 2016 A persistent challenge to improving our understanding of autism is the fact that no single neurological
RESEARCHER S NAME: Làszlò Tora RESEARCHER S ORGANISATION: Institut de Génétique et de Biologie Moléculaire et Cellulaire (IGBMC)
") Thursday 5 November EU-India PARTNERING EVENT Theme: Health RESEARCHER S NAME: Làszlò Tora RESEARCHER S ORGANISATION: Institut de Génétique et de Biologie Moléculaire et Cellulaire (IGBMC) CNRS, INSERM,
Thursday 5 November EU-India PARTNERING EVENT Theme: Health RESEARCHER S NAME: Làszlò Tora RESEARCHER S ORGANISATION: Institut de Génétique et de Biologie Moléculaire et Cellulaire (IGBMC) CNRS, INSERM,
Eukaryotic Gene Regulation
 Eukaryotic Gene Regulation Chapter 19: Control of Eukaryotic Genome The BIG Questions How are genes turned on & off in eukaryotes? How do cells with the same genes differentiate to perform completely different,
Eukaryotic Gene Regulation Chapter 19: Control of Eukaryotic Genome The BIG Questions How are genes turned on & off in eukaryotes? How do cells with the same genes differentiate to perform completely different,
Allelic reprogramming of the histone modification H3K4me3 in early mammalian development
 Allelic reprogramming of the histone modification H3K4me3 in early mammalian development 张戈 Method and material STAR ChIP seq (small-scale TELP-assisted rapid ChIP seq) 200 mouse embryonic stem cells PWK/PhJ
Allelic reprogramming of the histone modification H3K4me3 in early mammalian development 张戈 Method and material STAR ChIP seq (small-scale TELP-assisted rapid ChIP seq) 200 mouse embryonic stem cells PWK/PhJ
Effects of UBL5 knockdown on cell cycle distribution and sister chromatid cohesion
 Supplementary Figure S1. Effects of UBL5 knockdown on cell cycle distribution and sister chromatid cohesion A. Representative examples of flow cytometry profiles of HeLa cells transfected with indicated
Supplementary Figure S1. Effects of UBL5 knockdown on cell cycle distribution and sister chromatid cohesion A. Representative examples of flow cytometry profiles of HeLa cells transfected with indicated
A Genetic Program for Embryonic Development
 Concept 18.4: A program of differential gene expression leads to the different cell types in a multicellular organism During embryonic development, a fertilized egg gives rise to many different cell types
Concept 18.4: A program of differential gene expression leads to the different cell types in a multicellular organism During embryonic development, a fertilized egg gives rise to many different cell types
Sirt1 Hmg20b Gm (0.17) 24 (17.3) 877 (857)
 24 (17.3) 877 (857)") 3 (0.17) 24 (17.3) Sirt1 Hmg20 Gm4763 877 (857) c d Suppl. Figure 1. Screen validation for top candidate antagonists of Dot1L (a) Numer of genes with one (gray), two (cyan) or three (red) shrna scored
3 (0.17) 24 (17.3) Sirt1 Hmg20 Gm4763 877 (857) c d Suppl. Figure 1. Screen validation for top candidate antagonists of Dot1L (a) Numer of genes with one (gray), two (cyan) or three (red) shrna scored
Today. Genomic Imprinting & X-Inactivation
 Today 1. Quiz (~12 min) 2. Genomic imprinting in mammals 3. X-chromosome inactivation in mammals Note that readings on Dosage Compensation and Genomic Imprinting in Mammals are on our web site. Genomic
Today 1. Quiz (~12 min) 2. Genomic imprinting in mammals 3. X-chromosome inactivation in mammals Note that readings on Dosage Compensation and Genomic Imprinting in Mammals are on our web site. Genomic
Genetics and Genomics in Medicine Chapter 6 Questions
 Genetics and Genomics in Medicine Chapter 6 Questions Multiple Choice Questions Question 6.1 With respect to the interconversion between open and condensed chromatin shown below: Which of the directions
Genetics and Genomics in Medicine Chapter 6 Questions Multiple Choice Questions Question 6.1 With respect to the interconversion between open and condensed chromatin shown below: Which of the directions
Epigenetics q&more 01.11
 Laurie. Knight, istockphoto.com Epigenetics 6 Bookmarks About the reading of genes in the Book of Life Prof. Dr. Manfred Jung, Julia M. Wagner, Institute for Pharmaceutical Sciences, Albert-Ludwig-University
Laurie. Knight, istockphoto.com Epigenetics 6 Bookmarks About the reading of genes in the Book of Life Prof. Dr. Manfred Jung, Julia M. Wagner, Institute for Pharmaceutical Sciences, Albert-Ludwig-University
EPIGENOMICS PROFILING SERVICES
 EPIGENOMICS PROFILING SERVICES Chromatin analysis DNA methylation analysis RNA-seq analysis Diagenode helps you uncover the mysteries of epigenetics PAGE 3 Integrative epigenomics analysis DNA methylation
EPIGENOMICS PROFILING SERVICES Chromatin analysis DNA methylation analysis RNA-seq analysis Diagenode helps you uncover the mysteries of epigenetics PAGE 3 Integrative epigenomics analysis DNA methylation
mirna Dr. S Hosseini-Asl
 mirna Dr. S Hosseini-Asl 1 2 MicroRNAs (mirnas) are small noncoding RNAs which enhance the cleavage or translational repression of specific mrna with recognition site(s) in the 3 - untranslated region
mirna Dr. S Hosseini-Asl 1 2 MicroRNAs (mirnas) are small noncoding RNAs which enhance the cleavage or translational repression of specific mrna with recognition site(s) in the 3 - untranslated region
Nature Immunology: doi: /ni Supplementary Figure 1 33,312. Aire rep 1. Aire rep 2 # 44,325 # 44,055. Aire rep 1. Aire rep 2.
 a 33,312 b rep 1 rep 1 # 44,325 rep 2 # 44,055 [0-84] rep 2 [0-84] 1810043G02Rik Pfkl Dnmt3l Icosl rep 1 [0-165] rep 2 [0-165] Rps14 Cd74 Mir5107 Tcof1 rep 1 [0-69] rep 2 [0-68] Id3 E2f2 Asap3 rep 1 [0-141]
a 33,312 b rep 1 rep 1 # 44,325 rep 2 # 44,055 [0-84] rep 2 [0-84] 1810043G02Rik Pfkl Dnmt3l Icosl rep 1 [0-165] rep 2 [0-165] Rps14 Cd74 Mir5107 Tcof1 rep 1 [0-69] rep 2 [0-68] Id3 E2f2 Asap3 rep 1 [0-141]
(a) Schematic diagram of the FS mutation of UVRAG in exon 8 containing the highly instable
 Schematic diagram of the FS mutation of UVRAG in exon 8 containing the highly instable") Supplementary Figure 1. Frameshift (FS) mutation in UVRAG. (a) Schematic diagram of the FS mutation of UVRAG in exon 8 containing the highly instable A 10 DNA repeat, generating a premature stop codon
Supplementary Figure 1. Frameshift (FS) mutation in UVRAG. (a) Schematic diagram of the FS mutation of UVRAG in exon 8 containing the highly instable A 10 DNA repeat, generating a premature stop codon
Selective filtering defect at the axon initial segment in Alzheimer s disease mouse models. Yu Wu
 Selective filtering defect at the axon initial segment in Alzheimer s disease mouse models Yu Wu Alzheimer s Disease (AD) Mouse models: APP/PS1, PS1δE9, APPswe, hps1 Wirths, O. et al, Acta neuropathologica
Selective filtering defect at the axon initial segment in Alzheimer s disease mouse models Yu Wu Alzheimer s Disease (AD) Mouse models: APP/PS1, PS1δE9, APPswe, hps1 Wirths, O. et al, Acta neuropathologica
Nature Structural & Molecular Biology: doi: /nsmb Supplementary Figure 1. Differential expression of mirnas from the pri-mir-17-92a locus.
 Supplementary Figure 1 Differential expression of mirnas from the pri-mir-17-92a locus. (a) The mir-17-92a expression unit in the third intron of the host mir-17hg transcript. (b,c) Impact of knockdown
Supplementary Figure 1 Differential expression of mirnas from the pri-mir-17-92a locus. (a) The mir-17-92a expression unit in the third intron of the host mir-17hg transcript. (b,c) Impact of knockdown
The Amazing Brain Webinar Series: Select Topics in Neuroscience and Child Development for the Clinician
 The Amazing Brain Webinar Series: Select Topics in Neuroscience and Child Development for the Clinician Part VII Recent Advances in the Genetics of Autism Spectrum Disorders Abha R. Gupta, MD, PhD Jointly
The Amazing Brain Webinar Series: Select Topics in Neuroscience and Child Development for the Clinician Part VII Recent Advances in the Genetics of Autism Spectrum Disorders Abha R. Gupta, MD, PhD Jointly
RUNX1 and FPD/AML Translational Research. The Leukemia and Lymphoma Society / Babich Family Foundation Partnership. September 2016
 www.lls.org www.runx1.com RUNX1 and FPD/AML Translational Research The Leukemia and Lymphoma Society / Babich Family Foundation Partnership September 2016 Prepared by L. Greenberger, PhD Chief Scientific
www.lls.org www.runx1.com RUNX1 and FPD/AML Translational Research The Leukemia and Lymphoma Society / Babich Family Foundation Partnership September 2016 Prepared by L. Greenberger, PhD Chief Scientific
Nature Medicine: doi: /nm.4322
 1 2 3 4 5 6 7 8 9 10 11 Supplementary Figure 1. Predicted RNA structure of 3 UTR and sequence alignment of deleted nucleotides. (a) Predicted RNA secondary structure of ZIKV 3 UTR. The stem-loop structure
1 2 3 4 5 6 7 8 9 10 11 Supplementary Figure 1. Predicted RNA structure of 3 UTR and sequence alignment of deleted nucleotides. (a) Predicted RNA secondary structure of ZIKV 3 UTR. The stem-loop structure
Supplementary Figure 1. MAT IIα is Acetylated at Lysine 81.
 IP: Flag a Mascot PTM Modified Mass Error Position Gene Names Score Score Sequence m/z [ppm] 81 MAT2A;AMS2;MATA2 35.6 137.28 _AAVDYQK(ac)VVR_ 595.83-2.28 b Pre-immu After-immu Flag- WT K81R WT K81R / Flag
IP: Flag a Mascot PTM Modified Mass Error Position Gene Names Score Score Sequence m/z [ppm] 81 MAT2A;AMS2;MATA2 35.6 137.28 _AAVDYQK(ac)VVR_ 595.83-2.28 b Pre-immu After-immu Flag- WT K81R WT K81R / Flag
Supplemental Data Macrophage Migration Inhibitory Factor MIF Interferes with the Rb-E2F Pathway
 Supplemental Data Macrophage Migration Inhibitory Factor MIF Interferes with the Rb-E2F Pathway S1 Oleksi Petrenko and Ute M. Moll Figure S1. MIF-Deficient Cells Have Reduced Transforming Ability (A) Soft
Supplemental Data Macrophage Migration Inhibitory Factor MIF Interferes with the Rb-E2F Pathway S1 Oleksi Petrenko and Ute M. Moll Figure S1. MIF-Deficient Cells Have Reduced Transforming Ability (A) Soft
Breast cancer. Risk factors you cannot change include: Treatment Plan Selection. Inferring Transcriptional Module from Breast Cancer Profile Data
 Breast cancer Inferring Transcriptional Module from Breast Cancer Profile Data Breast Cancer and Targeted Therapy Microarray Profile Data Inferring Transcriptional Module Methods CSC 177 Data Warehousing
Breast cancer Inferring Transcriptional Module from Breast Cancer Profile Data Breast Cancer and Targeted Therapy Microarray Profile Data Inferring Transcriptional Module Methods CSC 177 Data Warehousing
Mechanisms of alternative splicing regulation
 Mechanisms of alternative splicing regulation The number of mechanisms that are known to be involved in splicing regulation approximates the number of splicing decisions that have been analyzed in detail.
Mechanisms of alternative splicing regulation The number of mechanisms that are known to be involved in splicing regulation approximates the number of splicing decisions that have been analyzed in detail.
3) The sheer number of and inconsistency between different animal models used make the paper difficult to follow and may impact data interpretation:
 The sheer number of and inconsistency between different animal models used make the paper difficult to follow and may impact data interpretation:") Reviewers' comments: Reviewer #1 (Remarks to the Author): In this manuscript, Pasricha et al. show that iron deficiency (ID) and stimulated erythropoiesis suppress hepcidin via distinct processes. They
Reviewers' comments: Reviewer #1 (Remarks to the Author): In this manuscript, Pasricha et al. show that iron deficiency (ID) and stimulated erythropoiesis suppress hepcidin via distinct processes. They
Ch. 18 Regulation of Gene Expression
 Ch. 18 Regulation of Gene Expression 1 Human genome has around 23,688 genes (Scientific American 2/2006) Essential Questions: How is transcription regulated? How are genes expressed? 2 Bacteria regulate
Ch. 18 Regulation of Gene Expression 1 Human genome has around 23,688 genes (Scientific American 2/2006) Essential Questions: How is transcription regulated? How are genes expressed? 2 Bacteria regulate
DNA methylation & demethylation
 DNA methylation & demethylation Lars Schomacher (Group Christof Niehrs) What is Epigenetics? Epigenetics is the study of heritable changes in gene expression (active versus inactive genes) that do not
DNA methylation & demethylation Lars Schomacher (Group Christof Niehrs) What is Epigenetics? Epigenetics is the study of heritable changes in gene expression (active versus inactive genes) that do not
a) List of KMTs targeted in the shrna screen. The official symbol, KMT designation,
 List of KMTs targeted in the shrna screen. The official symbol, KMT designation,") Supplementary Information Supplementary Figures Supplementary Figure 1. a) List of KMTs targeted in the shrna screen. The official symbol, KMT designation, gene ID and specifities are provided. Those highlighted
Supplementary Information Supplementary Figures Supplementary Figure 1. a) List of KMTs targeted in the shrna screen. The official symbol, KMT designation, gene ID and specifities are provided. Those highlighted
The functional investigation of the interaction between TATA-associated factor 3 (TAF3) and p53 protein
 and p53 protein") THESIS BOOK The functional investigation of the interaction between TATA-associated factor 3 (TAF3) and p53 protein Orsolya Buzás-Bereczki Supervisors: Dr. Éva Bálint Dr. Imre Miklós Boros University of
THESIS BOOK The functional investigation of the interaction between TATA-associated factor 3 (TAF3) and p53 protein Orsolya Buzás-Bereczki Supervisors: Dr. Éva Bálint Dr. Imre Miklós Boros University of
Regulation of Gene Expression in Eukaryotes
 Ch. 19 Regulation of Gene Expression in Eukaryotes BIOL 222 Differential Gene Expression in Eukaryotes Signal Cells in a multicellular eukaryotic organism genetically identical differential gene expression
Ch. 19 Regulation of Gene Expression in Eukaryotes BIOL 222 Differential Gene Expression in Eukaryotes Signal Cells in a multicellular eukaryotic organism genetically identical differential gene expression
Alpha thalassemia mental retardation X-linked. Acquired alpha-thalassemia myelodysplastic syndrome
 Alpha thalassemia mental retardation X-linked Acquired alpha-thalassemia myelodysplastic syndrome (Alpha thalassemia mental retardation X-linked) Acquired alpha-thalassemia myelodysplastic syndrome Schematic
Alpha thalassemia mental retardation X-linked Acquired alpha-thalassemia myelodysplastic syndrome (Alpha thalassemia mental retardation X-linked) Acquired alpha-thalassemia myelodysplastic syndrome Schematic
A. List of selected proteins with high SILAC (H/L) ratios identified in mass
 ratios identified in mass") Supplementary material Figure S1. Interaction between UBL5 and FANCI A. List of selected proteins with high SILAC (H/L) ratios identified in mass spectrometry (MS)-based analysis of UBL5-interacting proteins,
Supplementary material Figure S1. Interaction between UBL5 and FANCI A. List of selected proteins with high SILAC (H/L) ratios identified in mass spectrometry (MS)-based analysis of UBL5-interacting proteins,
Supplementary Information for
 Supplementary Information for Involvement of urinary bladder Connexin43 and the circadian clock in the coordination of diurnal micturition rhythm Hiromitsu Negoro, 1,2 Akihiro Kanematsu, 1,3 Masao Doi,
Supplementary Information for Involvement of urinary bladder Connexin43 and the circadian clock in the coordination of diurnal micturition rhythm Hiromitsu Negoro, 1,2 Akihiro Kanematsu, 1,3 Masao Doi,
Session 6: Integration of epigenetic data. Peter J Park Department of Biomedical Informatics Harvard Medical School July 18-19, 2016
 Session 6: Integration of epigenetic data Peter J Park Department of Biomedical Informatics Harvard Medical School July 18-19, 2016 Utilizing complimentary datasets Frequent mutations in chromatin regulators
Session 6: Integration of epigenetic data Peter J Park Department of Biomedical Informatics Harvard Medical School July 18-19, 2016 Utilizing complimentary datasets Frequent mutations in chromatin regulators
Kabuki syndrome: A potentially treatable cause of intellectual disability.
 Qydezia Barrett Kabuki syndrome: A potentially treatable cause of intellectual disability. Hans Tómas Björnsson MD PhD East Coast Kabuki Conference June 2015 What is DNA? DNA is the language of life (how
Qydezia Barrett Kabuki syndrome: A potentially treatable cause of intellectual disability. Hans Tómas Björnsson MD PhD East Coast Kabuki Conference June 2015 What is DNA? DNA is the language of life (how
Figure S1, Beyer et al.
 Figure S1, eyer et al. Pax7 Myogenin si sitrl Hoechst T = 72h 14 1.8.6.4.2 12 1 8 6 4 2 24h 48h 96h diff. sitrl siset1 212 72h diff. b1 td r t Se km MyH Vinculin Myogenin β-ctin Vinculin MW b1 ka td r
Figure S1, eyer et al. Pax7 Myogenin si sitrl Hoechst T = 72h 14 1.8.6.4.2 12 1 8 6 4 2 24h 48h 96h diff. sitrl siset1 212 72h diff. b1 td r t Se km MyH Vinculin Myogenin β-ctin Vinculin MW b1 ka td r
Processing, integrating and analysing chromatin immunoprecipitation followed by sequencing (ChIP-seq) data
 data") Processing, integrating and analysing chromatin immunoprecipitation followed by sequencing (ChIP-seq) data Bioinformatics methods, models and applications to disease Alex Essebier ChIP-seq experiment To
Processing, integrating and analysing chromatin immunoprecipitation followed by sequencing (ChIP-seq) data Bioinformatics methods, models and applications to disease Alex Essebier ChIP-seq experiment To
Supplementary Figures
 J. Cell Sci. 128: doi:10.1242/jcs.173807: Supplementary Material Supplementary Figures Fig. S1 Fig. S1. Description and/or validation of reagents used. All panels show Drosophila tissues oriented with
J. Cell Sci. 128: doi:10.1242/jcs.173807: Supplementary Material Supplementary Figures Fig. S1 Fig. S1. Description and/or validation of reagents used. All panels show Drosophila tissues oriented with
Myelin suppresses axon regeneration by PIR-B/SHPmediated inhibition of Trk activity
 Manuscript EMBO-2010-76298 Myelin suppresses axon regeneration by PIR-B/SHPmediated inhibition of Trk activity Yuki Fujita, Shota Endo, Toshiyuki Takai and Toshihide Yamashita Corresponding author: Toshihide
Manuscript EMBO-2010-76298 Myelin suppresses axon regeneration by PIR-B/SHPmediated inhibition of Trk activity Yuki Fujita, Shota Endo, Toshiyuki Takai and Toshihide Yamashita Corresponding author: Toshihide
SUPPLEMENTARY INFORMATION
 DOI: 10.1038/ncb2988 Supplementary Figure 1 Kif7 L130P encodes a stable protein that does not localize to cilia tips. (a) Immunoblot with KIF7 antibody in cell lysates of wild-type, Kif7 L130P and Kif7
DOI: 10.1038/ncb2988 Supplementary Figure 1 Kif7 L130P encodes a stable protein that does not localize to cilia tips. (a) Immunoblot with KIF7 antibody in cell lysates of wild-type, Kif7 L130P and Kif7
NFIL3/E4BP4 controls Type 2 T helper cell cytokine expression
 Manuscript EMBO-2010-75657 NFIL3/E4BP4 controls Type 2 T helper cell cytokine expression Masaki Kashiwada, Suzanne L. Cassel, John D. Colgan and Paul B. Rothman Corresponding author: Paul Rothman, University
Manuscript EMBO-2010-75657 NFIL3/E4BP4 controls Type 2 T helper cell cytokine expression Masaki Kashiwada, Suzanne L. Cassel, John D. Colgan and Paul B. Rothman Corresponding author: Paul Rothman, University
Early Embryonic Development
 Early Embryonic Development Maternal effect gene products set the stage by controlling the expression of the first embryonic genes. 1. Transcription factors 2. Receptors 3. Regulatory proteins Maternal
Early Embryonic Development Maternal effect gene products set the stage by controlling the expression of the first embryonic genes. 1. Transcription factors 2. Receptors 3. Regulatory proteins Maternal
Supplementary Information Supplementary Fig. 1. Elevated Usp9x in melanoma and NRAS mutant melanoma cells are dependent on NRAS for 3D growth.
 Supplementary Information Supplementary Fig. 1. Elevated Usp9x in melanoma and NRAS mutant melanoma cells are dependent on NRAS for 3D growth. a. Immunoblot for Usp9x protein in NRAS mutant melanoma cells
Supplementary Information Supplementary Fig. 1. Elevated Usp9x in melanoma and NRAS mutant melanoma cells are dependent on NRAS for 3D growth. a. Immunoblot for Usp9x protein in NRAS mutant melanoma cells
Yue Wei 1, Rui Chen 2, Carlos E. Bueso-Ramos 3, Hui Yang 1, and Guillermo Garcia-Manero 1
 Genome-wide CHIP-Seq Analysis of Histone Methylation Reveals Modulators of NF- B Signaling And the Histone Demethylase JMJD3 Implicated in Myelodysplastic Syndrome Yue Wei 1, Rui Chen 2, Carlos E. Bueso-Ramos
Genome-wide CHIP-Seq Analysis of Histone Methylation Reveals Modulators of NF- B Signaling And the Histone Demethylase JMJD3 Implicated in Myelodysplastic Syndrome Yue Wei 1, Rui Chen 2, Carlos E. Bueso-Ramos
Supplement Figure S1. Real Time PCR analysis of mrna levels of C/EBPα and PU.1 in wild type (WT) and NQO1-null (NQO1-/-) mice.
 and NQO1-null (NQO1-/-) mice.") competes with 20S proteasome for binding with C/EBP leading to its stabilization and Relative mrna levels Supplement Figure S1. Real Time PCR analysis of mrna levels of C/EBPα and PU.1 in wild type (WT)
competes with 20S proteasome for binding with C/EBP leading to its stabilization and Relative mrna levels Supplement Figure S1. Real Time PCR analysis of mrna levels of C/EBPα and PU.1 in wild type (WT)
MeCP2 and psychostimulantinduced behavioral adaptations. Anne E. West, M.D., Ph.D. Department of Neurobiology Duke University Medical Center
 MeCP2 and psychostimulantinduced behavioral adaptations Anne E. West, M.D., Ph.D. Department of Neurobiology Duke University Medical Center Psychostimulants and antidepressants slowly change behavior Psychostimulants
MeCP2 and psychostimulantinduced behavioral adaptations Anne E. West, M.D., Ph.D. Department of Neurobiology Duke University Medical Center Psychostimulants and antidepressants slowly change behavior Psychostimulants
Biochemical Determinants Governing Redox Regulated Changes in Gene Expression and Chromatin Structure
 Biochemical Determinants Governing Redox Regulated Changes in Gene Expression and Chromatin Structure Frederick E. Domann, Ph.D. Associate Professor of Radiation Oncology The University of Iowa Iowa City,
Biochemical Determinants Governing Redox Regulated Changes in Gene Expression and Chromatin Structure Frederick E. Domann, Ph.D. Associate Professor of Radiation Oncology The University of Iowa Iowa City,
Jayanti Tokas 1, Puneet Tokas 2, Shailini Jain 3 and Hariom Yadav 3
 Jayanti Tokas 1, Puneet Tokas 2, Shailini Jain 3 and Hariom Yadav 3 1 Department of Biotechnology, JMIT, Radaur, Haryana, India 2 KITM, Kurukshetra, Haryana, India 3 NIDDK, National Institute of Health,
Jayanti Tokas 1, Puneet Tokas 2, Shailini Jain 3 and Hariom Yadav 3 1 Department of Biotechnology, JMIT, Radaur, Haryana, India 2 KITM, Kurukshetra, Haryana, India 3 NIDDK, National Institute of Health,
Muscular Dystrophy. Biol 405 Molecular Medicine
 Muscular Dystrophy Biol 405 Molecular Medicine Duchenne muscular dystrophy Duchenne muscular dystrophy is a neuromuscular disease that occurs in ~ 1/3,500 male births. The disease causes developmental
Muscular Dystrophy Biol 405 Molecular Medicine Duchenne muscular dystrophy Duchenne muscular dystrophy is a neuromuscular disease that occurs in ~ 1/3,500 male births. The disease causes developmental
Epigenetics Armstrong_Prelims.indd 1 04/11/2013 3:28 pm
 Epigenetics Epigenetics Lyle Armstrong vi Online resources Accessible from www.garlandscience.com, the Student and Instructor Resource Websites provide learning and teaching tools created for Epigenetics.
Epigenetics Epigenetics Lyle Armstrong vi Online resources Accessible from www.garlandscience.com, the Student and Instructor Resource Websites provide learning and teaching tools created for Epigenetics.