Characterization of Jak, STAT, and Src interactions in Head and Neck Squamous Cell Carcinoma
|
|
|
- Raymond Fitzgerald
- 6 years ago
- Views:
Transcription

. UT GSBS Dissertations and Theses (Open Access). 387.")
1 Texas Medical Center Library UT GSBS Dissertations and Theses (Open Access) Graduate School of Biomedical Sciences Characterization of Jak, STAT, and Src interactions in Head and Neck Squamous Cell Carcinoma Reshma Jaseja Reshma Jaseja Follow this and additional works at: Part of the Biology Commons, Cancer Biology Commons, Cell Biology Commons, Laboratory and Basic Science Research Commons, and the Molecular Biology Commons Recommended Citation Jaseja, Reshma and Jaseja, Reshma, "Characterization of Jak, STAT, and Src interactions in Head and Neck Squamous Cell Carcinoma" (2013). UT GSBS Dissertations and Theses (Open Access) This Thesis (MS) is brought to you for free and open access by the Graduate School of Biomedical Sciences at It has been accepted for inclusion in UT GSBS Dissertations and Theses (Open Access) by an authorized administrator of For more information, please contact
2 Characterization of Jak, STAT, and Src interactions in Head and Neck Squamous Cell Carcinoma APPROVED: by Reshma Jaseja, B.S Faye M. Johnson, MD PhD Supervisory Professor Gary Gallick, PhD Pierre McCrea, PhD Michael Davies, MD PhD Don Gibbons, MD PhD APPROVED: Dean, The University of Texas Graduate School of Biomedical Sciences at Houston
3 Characterization of Jak, STAT, and Src interactions in Head and Neck Squamous Cell Carcinoma A THESIS Presented to the Faculty of The University of Texas Health Science Center at Houston and The University of Texas MD Anderson Cancer Center Graduate School of Biomedical Sciences In Partial Fulfillment of the Requirements for the Degree of MASTER OF SCIENCE by Reshma Jaseja, B.S Houston, Texas August, 2013
4 Dedicated to my family iii
5 Acknowledgements: I would like to express my gratitude to my mentor Dr. Faye Johnson for her kindness, patience, and support while she gave me guidance for the past two years. I am also grateful for the help I received from the members of the lab: Dr. Banibrata Sen, Dr. Courtney Nicholas, Dr. Tuhina Mazumdar, Dr. Shaohua Peng, and Dr. Renata Ferrarotto. I would also like to thank my committee members Dr. Gary Gallick, Dr. Pierre McCrea, Dr. Michael Davies, and Dr. Don Gibbons for their advice given during committee meetings. And lastly, I would like to thank my wonderful parents Mahesh Jaseja and Premila Jaseja, as well as my sister Vanita Jaseja, for their constant support and encouragement. iv
6 Characterization of Jak, STAT and Src Interactions in Head and Neck Squamous Cell Carcinoma Reshma Jaseja, B.S. Supervisory Professor: Faye M. Johnson, M.D., Ph.D. Recurrence of Head and Neck Squamous Cell Carcinoma (HNSCC) is common; thus, it is essential to improve the effectiveness and reduce toxicity of current treatments. Proteins in the Src/Jak/STAT pathway represent potential therapeutic targets, as this pathway is hyperactive in HNSCC and it has roles in cell migration, metastasis, proliferation, survival, and angiogenesis. During short-term Src inhibition, Janus kinase (Jak) 2, and signal transducer and activator of transcription (STAT) 3 and STAT5 are dephosphorylated and inactivated. Following sustained Src inhibition, STAT5 remains inactive, but Jak2 and STAT3 are reactivated following their early inhibition. To further characterize the mechanism of this novel feedback pathway we performed several experiments to look at the interactions between Src, Jak2, STAT5 and STAT3. We attempted to develop a non-radioactive kinase assay using purified recombinant Jak2 and Src proteins, but found that phospho-tyrosine antibodies were non-specifically binding to purified recombinant proteins. We then performed in vitro kinase assays (IVKAs) using purified recombinant Jak2, Src, STAT3, and STAT5 proteins with and without Src and Jak2 pharmacologic inhibitors. We also examined the interactions of these proteins in intact HNSCC cells. We found that recombinant Jak2, STAT3, and STAT5 are direct substrates of Src and that recombinant Src, STAT3, and STAT5 are direct substrates of Jak2 in the IVKA. To our knowledge, the finding that Src is a Jak substrate is novel and has not been v
7 shown before. In intact HNSCC cells we find that STAT3 can be reactivated despite continuous Src inhibition and that STAT5 continues to be inhibited despite Jak2 reactivation. Also, Jak2 inhibition did not affect Src or STAT5 activity but it did cause STAT3 inhibition. We hypothesized that the differences between the intact cells and the IVKA assays were due to a potential need for binding partners in intact HNSCC cells. One potential binding partner that we examined is the epidermal growth factor receptor (EGFR). We found that EGFR activation caused increased activation of Src and STAT5 but not Jak2. Our results demonstrate that although STAT3 and STAT5 are capable of being Src and Jak2 substrates, in intact HNSCC cells Src predominantly regulates STAT5 and Jak2 regulates STAT3. Regulation of STAT5 by Src may involve interactions between Src and EGFR. This knowledge along with future studies will better define the mechanisms of STAT regulation in HNSCC cells and ultimately result in an ideal combination of therapeutic agents for HNSCC. vi
8 TABLE OF CONTENTS: Approval Sheet...i Title Page...ii Dedication...iii Acknowledgements...iv Abstract...v Table of Contents...vii List of Figures...ix Chapter 1. Introduction...1 Head and Neck Squamous Cell Carcinoma Background...2 Commonly altered signaling pathways in HNSCC...4 Src Family Kinases in HNSCC...7 Integrins...9 The Jak-STAT pathway...10 STAT3 Reactivation Following Sustained Src Inhibition in HNSCC...14 Hypothesis...17 Chapter 2. Materials and Methods...18 Cells and Reagents...19 In Vitro Kinase Assays...20 Cell Culture...21 Western Blot Analysis...22 Immunoprecipitation...22 Chapter 3. Results...23 vii
9 Commercially available anti-phosphotyrosine antibodies bind nonspecifically to recombinant JAK2 and Src...24 Src and Jak2 are direct substrates of each other in a cell-free in vitro kinase assay...29 Src regulates STAT5 but not STAT3 activity in intact HNSCC cell lines...33 Jak regulates STAT3 but not STAT5 in intact HNSCC cell lines...33 STAT5 is a direct substrates of both Jak2 and Src in a cell-free in vitro kinase assay...35 STAT3 is a direct substrate of both Jak2 and Src in a cell-free in vitro kinase assay...35 EGFR activation increases Src and STAT5 activity in HNSCC cells...39 Focal Adhesion Kinase may regulate Src...41 Chapter 4. Discussion...42 Jak and Src Interactions...43 Regulation of STAT3 and STAT5 Activation...44 Src Activation...47 Future Directions...48 Chapter 5. References...51 Vita...67 viii
10 LIST OF FIGURES: Figure 1 Anatomy of the Head and Neck...2 Figure 2 The PI3K/Akt pathway and common mutations...6 Figure 3 Src kinase structure and domains...7 Figure 4 Active and Inactive Conformations of Src...8 Figure 5 Structure of Jak kinase and domains...11 Figure 6 The Jak/STAT pathway...12 Figure 7 STAT structure, domains, and functions...13 Figure 8 Schematic of basal, acute, and chronic Src inhibition and its effects on Jak2 and STATs...16 Figure 9 Anti-phosphotyrosine antibodies bind non-specifically to recombinant Jak2 and Src...26 Figure 10 Recombinant Jak2 is a direct Src substrate and Src is a Jak2 substrate...31 Figure 11 Src predominately regulates STAT5 and Jak predominately regulates STAT3 in intact HNSCC cells...34 Figure 12 STAT5A is a direct substrate of both Src and Jak Figure 13 STAT3 is a direct substrate of both Jak2 and Src...38 Figure 14 EGFR co-immunoprecipitates with Src in TU167 cells...40 Figure 15 FAK may play a role in Src regulation, but is not a major regulator...41 ix
11 CHAPTER 1: INTRODUCTION 1
 with cancer suffer from HNSCC.")
12 Head and Neck Squamous Cell Carcinoma Background Head and Neck Squamous Cell Carcinoma (HNSCC) is the sixth most common type of cancer worldwide (1). Approximately 3 to 5% of patients in the United States (US) with cancer suffer from HNSCC. It is estimated that in the US about 53,640 people will acquire head and neck cancer in 2013 and of these, 11,520 people will die. Generally, twice as many men suffer from HNSCC as women and a majority of people are diagnosed after the age of 50. This type of cancer begins in the squamous cells that line the head and neck mucosal surfaces (2). These surfaces include those of the pharynx, larynx, oral cavity, paranasal sinuses, and nasal cavity (Figure 1). Figure 1. Anatomy of the Head and Neck. Permission to use this image was given by Ms. Terese Winslow LLC. 2
13 Many risk factors can contribute to HNSCC. About 85% of head and neck cancers are caused by tobacco use. Human papilloma virus (HPV), particularly subtypes 16 and 18, is a major risk factor for oropharyngeal cancer; the incidence of HPV-related oropharyngeal cancer is increasing in the US (3). Other risk factors include alcohol consumption, exposure to the sun (lip), Epstein-Barr virus (nasopharynx), male gender, advanced age, Asian race, poor oral hygiene, and others (4, 5). The choice of treatment depends principally on the location of the primary tumor and the stage of the cancer. Treatments include surgery, targeted therapy, radiotherapy, chemotherapy, or a combination of these therapies. All of these therapies can result in uncomfortable and life-altering side effects that may impact speech, eating, hearing, vision, and appearance. For example, surgery may result in the removal of the larynx (voice box); radiation therapy can cause hypothyroidism and severe xerostomia by destruction of the normal thyroid and salivary tissue respectively; chemotherapy can enhance the effects of radiation and can also cause systemic side effects such as infection, hair loss, fatigue, neuropathy, nausea and vomiting (2). Targeted therapy inhibits specific proteins that allow the cancer cells to grow and survive; there is a continuous need for research to identify specific targets so that inhibitors or other treatments can be used against them. Cetuximab, a monoclonal antibody against epidermal growth factor receptor (EGFR) is the only currently approved targeted therapy against HNSCC (6). Cetuximab is used in 3
14 combination with chemotherapy or as monotherapy for metastatic disease or in combination with radiotherapy for locally advanced HNSCC. Commonly altered signaling pathways in HNSCC Targeted therapy is an important and effective way in which cancer patients are treated. For targeted therapy to be effective, it is essential to know which signaling pathways are activated and drive cancer survival and progression in a specific type of cancer. Previous research has shown that several signaling pathways may contribute to the progression of HNSCC. EGFR has been targeted for the treatment of HNSCC and other cancer types. In non-small cell lung cancer (NSCLC) EGFR may be activated through mutation. Although activating mutations in EGFR are rare in HNSCC, EGFR is overexpressed, often by gene amplification, in this type of cancer in 80-90% of all cases (7). EGFR is a transmembrane glycoprotein that is part of the ErbB/Her family of tyrosine kinase receptors. When EGFR binds its ligand such as Epidermal Growth Factor (EGF) or transforming growth factor (TGF ), the receptor experiences a conformational change that exposes two previously occluded sites within EGFR and allows dimerization with another EGFR or ErbB/Her family member. Receptor dimerization allows EGFR autophosphorylation and the activation of its kinase activity. EGFR has several substrates which subsequently cause activation of downstream pathways that are involved in proliferation, survival, and migration of the cancer cell. One of the best-studied pathways downstream of EGFR and other cell surface receptors is the RAS/RAF/MAPK pathway. Previous 4
15 studies have shown that EGFR overexpression in HNSCC correlates with poor clinical outcome (8); EGFR tyrosine kinase inhibitors have been approved for clinical use and include gefitinib, erlotinib, and lapatinib (1, 9). Another important signaling pathway in HNSCC is the PI3K/Akt/mTOR pathway. It has been discovered through whole-exome sequencing that about 20-30% of HNSCC tumors that were analyzed had activating mutations in phophoinositide 3-kinase (PI3K) (10). Class IA PI3Ks are a family of kinases that phosphorylate phosphatidylinositols such as phosphatidylinositol-4,5,bisphosphate (PI(4,5)P 2 ) to produce phosphatidylinositol-3,4,5,trisphosphate (PI(3,4,5)P 3 ), which is a second messenger. This second messenger then binds to 3 -phosphoinositidedependent kinase 1 (PDK1) at the Plekstrin Homology (PH) domain and Akt. This binding causes their translocation to the cell membrane and allows PDK1 to phosphorylate and activate Akt. Activated Akt has multiple substrates, including mtorc1, that are necessary for many cellular processes such as cell survival (Figure 2). This pathway becomes constitutively active when there is an activating mutation in the PIK3CA gene. Mutations can also occur in other parts of this pathway including Akt, other PI3K family members, and PTEN, a phosphatase which acts to inhibit Akt by dephosphorylating (PI(3,4,5)P 3 ). Components of this pathway can be altered by mutation, deletion, amplifications, and loss (11). 5

16 Figure 2. The PI3K/Akt pathway and common mutations. Reprinted by permission from Macmillan Publishers Ltd: Nature Reviews: Drug Discovery. Faivre, S., G. Kroemer, and E. Raymond Current development of mtor inhibitors as anticancer agents. Nature reviews. Drug discovery 5: Other commonly activated pathways in HNSCC are Src and the JAK/STAT pathways described in more detail below. Common somatic mutations include those in FAT1, HRAS, MLL2, CASP8, FBXW7, and CDKN2A (12). Several tumor suppressors are also commonly mutated in HNSCC and these include p53 and NOTCH1(12). 6
.")
17 Src Family Kinases in HNSCC Src belongs to a family of non-receptor tyrosine kinases that includes Yes, Fyn, Lyn, Lck, Blk, Fgr, and Hck in mammals. Src was the first proto-oncogene to be discovered and is an important protein that has roles in cancer cell migration, proliferation, survival, differentiation, invasion and angiogenesis (13). Figure 3. Src kinase structure showing all domains. Reprinted by permission from Macmillan Publishers Ltd: Oncogene, The Src protein has several domains that contribute to and regulate its function as a kinase. Src contains a catalytic domain (SH1 domain), SH2 domain, SH3 domain, SH4 domain, and a unique domain (Figure 3). The catalytic domain or kinase domain (SH1) contains an activation loop where the tyrosine 419 (humans) is located. Src kinases also have a small C-terminal cytoplasmic tail, which contains a phosphorylation site at tyrosine 527. This is an inhibitory phosphorylation site. The SH2 and SH3 domains are non-catalytic domains that 7
.")
18 together act as a regulatory unit. These two domains allow Src to be in an inactive confirmation when there is intramolecular binding (13). (Binding of the Src SH2 domain to phosphopeptides is determined by residues in both the SH2 domain and the phosphopeptides). The SH2 and SH3 domains allow for the recognition of other proteins to bind to Src via phosphotyrosine motifs and proline-rich motifs and once this binding occurs, the protein unfolds and becomes activated because the kinase phosphorylation site is accessible (Figure 4) (14). Figure 4. Active and Inactive conformations of Src. Reprinted from Molecular Cell, Vol 3, Weqing Xu, Amish Doshi, Ming Lei, Michael J Eck, Stephen C Harrison, , 1999, with permission from Elsevier. 8
19 There is increased Src expression in HNSCC compared to the normal mucosal surfaces (15). Previous reports have also shown that Platelet Derived Growth Factor Receptor (PDGFR), EGFR, and Focal Adhesion Kinase (FAK), among many other molecules, can activate Src (16-20). There has also been evidence to suggest that the cytoplasmic domain of beta integrin may have a role in Src activation (21). The integrins are described in more detail below. Src directly phosphorylates many proteins. Its substrates include STATs, cell adhesion receptors, G-protein coupled receptors, Hepatocyte growth factor receptor (Met), and components of the FAK, PI3K-Akt, MAPK, and Jak/STAT pathways (22-25). It is known that Src activates STATs, particularly STAT3 and STAT5, independently of Jak activation (26). Previous reports from this laboratory also show that with Src inhibition, both STAT5 and STAT3 are initially inhibited (27). Integrins As mentioned previously, integrins act upstream of Src. Integrins are needed for cell adhesion to the extracellular matrix, which is important for cell survival, differentiation, growth, migration, etc. Integrin signaling occurs through the interaction of protein partners such as FAK, adaptor proteins, and integrin-like kinase (ILK). Integrins are made up of an α chain and a β chain. There are 18 α chains and 8 β chains. One of the most common chains found in integrin heterodimers (in epithelial cells) is the β1 integrin. Cytoplasmic proteins and adaptor proteins bind to the cytoplasmic tails of integrins, which leads to signaling (28). 9
20 The Jak-STAT pathway The main components of the Jak/STAT pathway include a cytokine or growth factor receptor, Janus Kinases (Jaks) and Signal Transducers and Activators of Transcription (STATs). Janus Kinases (Jaks) are a family of non-receptor tyrosine kinases; this family includes Jak1, Jak2, Jak3, and Tyk2. A diagram of the Jak structure is shown below (Figure 5). Jaks contain 7 Jak Homology (JH) domains. The JH1 domain, which is at the C-terminal end, is also the catalytic domain. This domain contains phosphotyrosines that are necessary for the activation of Jaks (tyrosine 1007/1008 for Jak2). The JH2 domain is the pseudo-kinase domain. This domain acts to negatively regulate the protein; when tyrosines on this domain are phosphorylated, the activity of the JH1 domain is inhibited. JH3 through JH7 domains are also regulatory domains and are needed for receptor interactions and includes the FERM (F -4.1 protein, E -ezrin, R - radixin and M moesin) domain (29). 10

21 Figure 5. Structure of Jak Kinase. (a) shows the main Jak domains and (b) shows the Jak homology domains. Reprinted from Trends in Molecular Medicine, Vol 11, Chloe James, Valerie Ugo, Nicole Casadevall, Stefan N. Constantinescu, William Vainchenker, A JAK2 mutation in myeloproliferative disorders: pathogenesis and therapeutic and scientific prospects, , 2005, with permission from Elsevier. The Jak/STAT pathway is primarily activated by cytokine and growth factors. When the receptor s ligand binds, receptor dimerization occurs to form homodimers or heterodimers allowing the associated Jaks to trans-phosphorylate each other to become activated, which then allows them to phosphorylate the interacting receptors and STATs. Phosphorylation of STATs causes them to dimerize and translocate to the nucleus, where they can bind to specific DNA sequences and cause transcription of their target genes (Figure 6) (30). 11

22 Figure 6. The Jak-STAT pathway. Reprinted by permission from Macmillan Publishers Ltd: Nature Reviews Immunology. Shuai, K., and B. Liu Regulation of JAK-STAT signalling in the immune system. Nature reviews. Immunology 3: The STAT proteins are transcription factors that regulate cell survival and differentiation. There are 7 STAT family members which are STAT1, STAT2, STAT3, STAT4, STAT5A, STAT5B, and STAT6. Unphosphorylated STATs are located in the cytoplasm. After phosphorylation they dimerize through the interactions of their SH2 domains and move to the nucleus where they bind to specific sequences on the DNA called Gamma-activated sites (GAS) and stimulate 12

23 transcription (31) (Figure 7). The two STAT family members that are implicated in HNSCC progression are STAT3 and STAT5 (32, 33). STAT3 is activated principally through phosphorylation of tyrosine 705 and this activation results in its mediation of important cell processes such as cell proliferation and apoptosis. STAT5 has two isoforms, STAT5A and STAT5B and they are activated by phosphorylation of tyrosine 694. Figure 7. STAT structure showing different domains and their functions. Reprinted from The Jounal of allergy and clinical immunology, Vol 119, Weiguo Chen,Gurjit K. Khurana Hershey, Signal transducer and activator of transcription signals in allergic disease, 2007, with permission from Elsevier. This pathway can be regulated at several levels. For the negative regulation of this pathway, there are three types of negative regulators. These include the Suppressor of Cytokine Signaling (SOCS) proteins, protein inhibitors of activated STATs (PIAs), and Protein Tyrosine Phosphatases (PTPs). SOCS inhibit the kinase activity of Jaks, aid with ubiquitin-mediated degradation, and prevent STATs from 13
24 binding to cytokine receptors. The PIAs negatively regulate STAT transcription and PTPs dephosphorylate Jak kinases (27). Although this pathway has been researched extensively, there are still many details of the pathway that are unknown. STAT3 Reactivation Following Sustained Src Inhibition in HNSCC Previous studies have shown that Src inhibition using an ATP-competitive Src kinase inhibitor such as dasatinib results in rapid (30 min) Jak2, STAT3, and STAT5 dephosphorylation and inactivation (27). During sustained Src inhibition, decreased STAT3 activation is only temporary but STAT5 inhibition was sustained. By 4-7 hours STAT3 becomes reactivated. This is also similar to Jak2 in that sustained Src inhibition results in initial Jak2 inhibition and Jak2 kinase activity recovers by 7 hours. In a previous study, it was shown that STAT3 reactivation occurs via Jak2 kinase activity and Jak-STAT3 binding. The investigators showed that when they depleted Jak2, they also saw an inhibition of STAT3 (Figure 8) (27). Because Jak2 regulates STAT3, when Jak2 is chronically inhibited so is STAT3 (Figure 8) (27). The reactivation of STAT3 is dependent upon Jak2 and Tyk2. This was shown by depleting both Jak2 or Tyk2 using sirna. Depletion of Tyk2 or JAK2 caused a partial decrease in STAT3 reactivation after Src inhibition. When both Jak2 and Tyk2 were knockdown at the same time, there was complete inhibition of STAT3 reactivation (27). The reactivation of STAT3 and Jak2 is biologically important. When STAT3 14
25 was completely abrogated along with Src, there was significant cell cycle arrest. There was also an increase in apoptosis when cells were treated with both a Src inhibitor and STAT3 sirna compared to when cells were just treated with the Src inhibitor. This suggests that STAT3 and Src work together to maintain cell survival and proliferation and that STAT3 reactivation lessens the effects of Src inhibition on cancer cell survival and proliferation. The reactivation of Jak2, Tyk2, and STAT3 is due to the loss of SOCS2 expression. STAT5 regulates SOCS2 transcription (34). The loss of STAT5 activity following Src inhibition leads to the decrease of SOCS protein levels. This prevents SOCS2 from inhibiting Jak2, interfering with Jak2-STAT3 binding and Jak2 kinase activity. 15

26 Figure 8. Schematic of basal, acute, and chronic Src inhibition and its effect on Jak2 and STAT activation. The basal state shows activation of Src, Jak2, STAT3, and STAT5. Acute Src inhibition results in inhibition of Jak2, STAT3, and STAT5 activity. Chronic Src inhibition results in STAT3 and Jak2 reactivation and durable STAT5 inhibition. In a negative feedback pathway, SOCS binds to and inhibits Jak2 from binding to STAT3. Taken from Courtney Nicholas with permission, PhD. 16
27 Hypothesis The model described above provides a framework for a feedback loop in which the inhibition of Src leads to Jak, STAT3, and STAT5 inhibition. sustained inhibition of STAT5 leads to the loss of SOCS2 transcription. The The decrease of SOCS2 protein levels allows for Jak2 and STAT3 reactivation by removing an inhibitory effect on Jak2-STAT3 binding and Jak2 kinase activity. However, this model has several unanswered questions. Although we know that Src inhibition leads to Jak2, STAT3, and STAT5 inhibition, we do not know if these molecules are direct substrates of Src. We are also uncertain about how STAT5 and STAT3 are regulated within the cells. We hypothesize that they are regulated independently by Src and Jak2 respectively. We also do not know the primary regulator of Src in HNSCC cells. It is possible that EGFR, FAK, integrins, a combination of these, or other upstream pathways may regulate Src activation in HNSCC. This thesis examines these questions by manipulating the pathway by use of inhibitors and stimulation using recombinant cytokines and growth factors. In vitro kinase assays were also performed to look at the interaction of Jak2, Src, STAT3, and STAT5 in a cell free system to compare these interactions with the interactions that occur in intact HNSCC cells. Better understanding of the Jak/STAT pathway is essential so that ultimately better treatments can be developed in order to improve patient survival and lifestyle. 17
28 CHAPTER 2: MATERIALS AND METHODS 18
29 Cells and reagents Human HNSCC cell lines OSC19 and TU167 were obtained from Dr. Jeffrey Myers, MD Anderson (35). Cell lines were authenticated by DNA fingerprinting and tested for mycoplasma and other contaminants by mouse antibody production (MAP) testing. Dasatinib, pyridone 6, saracatinib, PF , and imatinib were purchased from Selleck Chemicals (Houston, TX) and prepared as stock solutions of 10 or 100 mmol/l in DMSO. Cytokines and Growth factors (PDGF, IGF, EGF, IL- 6, and TGF-α) were purchased from Peprotech (Rocky Hill, New Jersey). Full length human, recombinant His-tagged Src expressed in insect cells (66 kda) was purchased from Sigma-Aldrich. Human, recombinant Jak2 containing the JH1 and JH2 domains (amino acids , 99kDa) expressed in Baculovirus infected Sf9 cells was purchased from Sigma-Aldrich and Invitrogen. Human STAT5A was purchased from Novus Biologicals. This recombinant protein contains full length STAT5A and a GST tag resulting in a protein of about 115 kda that was expressed in an in vitro wheat germ system. Human STAT3 was purchased from Abcam (Cambridge, MA). This protein is a full-length recombinant protein that was expressed in Sf9 insect cells and contains an N-terminal GST tag resulting in a protein of about120 kda. Antibodies psrc (Y416) CS2101, total Src CS2110, pjak2(y1007/1008) CS3771, pstat3(y705) CS9131, Total Stat3 CS9139, pstat5(y694) CS9351, Total STAT5 CS9358, pegfr (Y1068) CS3777, total EGFR CS4267, pfak(y576) CS3281, pfak(y925) CS3284, pfak(y861) CS3283, total Fak CS3285, total PDGFRβ CS3169, and β1 Integrin CS9699 were all purchased from Cell Signaling. 19
30 ppdgfr(y572/574) # G and pfak(y397) #44624G were purchased from Invitrogen. Total Jak2 (sc-294) was purchased from Santa Cruz. Phospho-Paxillin was purchased from Abcam For immunoprecipitations, the Jak2 antibody (AHO1352) was purchased from Invitrogen and the Src antibody (OP07A) was purchased from Calbiochem. In Vitro kinase assays This assay was done as previously described with the following modifications (27). Recombinant proteins [Src (435 nmol/l), Jak2 (870 nmol/l), STAT5 (870 nmol/l), and/or STAT3 (870 nmol/l)] were incubated with DMSO control, 2 μm saracatinib, 100 nm dasatinib, or 20 mm pyridone 6 for 30 minutes after which HBV buffer [50 mm/l HEPES, 100 umol/l sodium orthovanadate (ph 7.4), 1 M/L MgCl 2, 1 M/L MnCl 2 and 10 μci (y- 32 P)ATP (3000 Ci/mmol; 1 Ci= 37 GBq)] was added and the samples incubated for another 15 min at room temperature. The reaction was stopped with 3X sample buffer (2% SDS, 5% beta-mercaptoethanol, mol/l Tris (ph 6.8), 1 mmol/l EDTA, 0.02% bromphenol blue). The samples were then boiled for 5 min and separated on 7.5% SDS-PAGE. The gel was fixed with fixing solution [45% Methanol, 45% distilled water, 10% Acetic Acid] for 20 min two times and boiled with 1 mol/l KOH for 10 minutes. The gel was then placed back in fixing solution for another 20 min and then dried using a gel dryer for 3 hr. The radiolabeled proteins were detected by autoradiography. 20
31 Cell culture HNSCC cells were grown in DMEM containing 10% Bovine Serum in a humidified CO 2 incubator at 37 C. They were grown on 100 mm tissue-culture dishes in a monolayer. Western blot analysis: Subconfluent HNSCC cells were washed with phosphate-buffered saline without calcium and magnesium. The cells were then incubated for 5 minutes with trypsin (Sigma-Aldrich) and spun down for 5 min at rpm at 4 C. The cell pellet was washed with PBS and lysed using lysis buffer containing 50 mmol/l Trizma base (ph8; Sigma), 1 % Triton X-100, 150 mmol/l NaCL, 20 ug/ml leupeptin, 10 ug/ml aprotinin, 1 mmol/l pehnylmethylsulfonyl fluoride, 1 mmol/l sodium vanadate and Protease and phosphatase inhibitors and were incubated for 20 mins at 4 degrees on a rotator. The lysates were centrifuged for 10 minutes at 14,000 rpm and the supernatant was collected. The protein concentration of each sample was determined using BCA reagents as per manufacturer s instructions (Thermo Scientific). Equal amounts of each sample was taken and mixed with the same volume of 2X sample buffer and resolved by SDS-PAGE and transferred to nitrocellulose membranes. The membranes were blocked in 5% milk in TBS-T for 1 hr at room temperature and then probed with primary antibody overnight. The primary antibody was incubated overnight at 4 C and then the membrane was washed with TBS-T 3 times for 5 min. The secondary antibody conjugated with horseradish peroxidase in 5% milk in TBS-T was then added for 1-2 hr at room 21
32 temperature and the membrane was washed again with TBS-T 3 times for 5 min. Protein bands were detected using Enhanced chemiluminescence reagent (Pierce). Immunoprecipitation: Tu167 and OSC19 cells were collected and lysed using immunoprecipitation (IP) lysis buffer (20 m M Tris-Cl ph 7.5, 150 mm NaCl, 1 mm EDTA, 1 mm EGTA, 1% NP-40, 5% glycerol). Lysates containing 800 μg of protein were pre-cleared using protein A and G sepharose beads for 1 hour. The cleared lysate was incubated with the indicated antibody or IgG control antibody and agarose beads for an hour. The beads were then washed 3 times using IP lysis buffer. Sample buffer was added to each sample and then each sample was boiled for 5 min and loaded onto SDS-polyacrylamide gels, transferred to nitrocellulose membranes, and probed with primary antibodies as indicated. The secondary antibody conjugated with horseradish peroxidase was then added and detected using enhanced chemiluminescence reagent. 22
33 CHAPTER 3: RESULTS 23
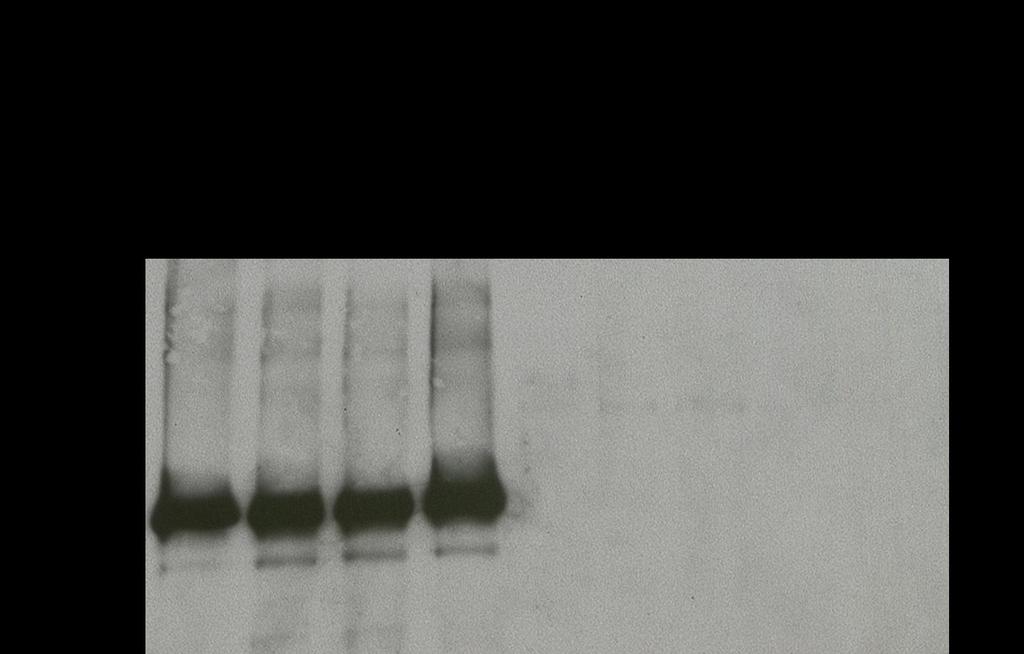
34 Commercially available anti-phosphotyrosine antibodies bind non-specifically to recombinant Jak2 and Src To facilitate our research examining Src, Jak, and STAT interactions and to enhance lab safety, we attempted to develop an in vitro kinase assay (IVKA) using recombinant purified proteins, Src and Jak2, and non-radiolabeled ATP. Recombinant Jak2 and Src were incubated with the Src inhibitor, dasatinib, the Jak inhibitor, pyridone 6, or vehicle control for 30 min in the presence of 80 mm ATP and the exogenous substrate enolase. Following incubation, the samples were run on SDS-PAGE and the membrane was probed with one of two phospho-tyrosine antibodies (4G10 or py100). We did not observe a significant band corresponding to enolase. When Src was incubated with dasatinib, there was no reduction in the staining of the 60 kda band corresponding to Src. Likewise, when Jak2 was incubated with pyridone 6; there was no reduction of the 130 kda Jak2 band (Figure 9A, B). To investigate the possibility of the recombinant proteins already being phosphorylated prior to performing the assay, a phosphatase, protein-tyrosine phosphatase 1B (PTP1B), was added to the IVKA reaction mixture without the addition of ATP. The results of this assay were similar to the previous one, showing no reduction in the staining of both the Src and Jak2 bands (Figure 9C). To examine the efficacy of the phosphatase, we immunoprecipitated epidermal growth factor receptor (EGFR) from TU167 cells with and without the addition of exogenous EGF to stimulate phosphorylation of EGFR. After the immunoprecipitation was performed, PTP1B was added to one sample to dephosphorylate the receptor. The lysates were run on SDS-PAGE. The 24
35 membrane was then probed with pegfr (Y1068). As expected, EGF stimulation led to a significant increase in pegfr. There was no pegfr band in the sample containing EGF and the phosphatase, indicating that PTP1B dephosphorylated EGFR (Figure 9D). We concluded that the phospho-tyrosine antibodies were nonspecifically binding to the recombinant proteins, making this non-radioactive IVKA unusable with currently available reagents. 25
36 26
37 27
 and Jak (5µM pyridone 6) inhibitors did not reduce the staining of Src and Jak2 with")
,")
38 Figure 9. Anti-phosphotyrosine antibodies bind non-specifically to recombinant Jak2 and Src. Potent Src (100 nm dasatinib) and Jak (5µM pyridone 6) inhibitors did not reduce the staining of Src and Jak2 with the antiphospho tyrosine antibodies 4G10 (A) or py100 (B). PTP1B phosphatase did not reduce the staining by 4G10 of the bands corresponding to the recombinant proteins (C), but did effectively dephosphorylate EGFR immunoprecipitated from TU167 cells (D). 28
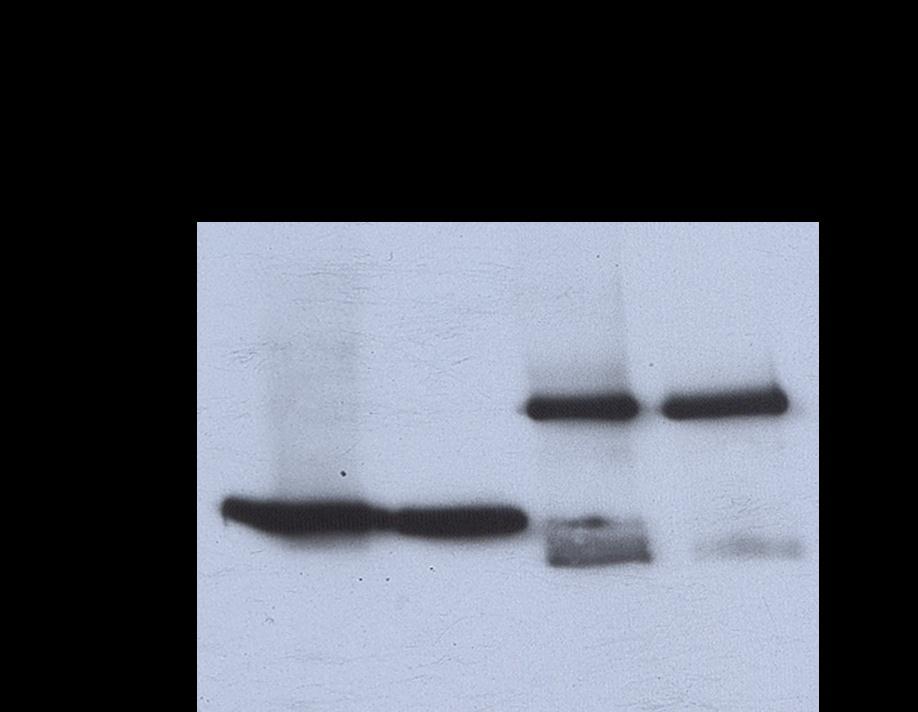
39 Src and Jak2 are direct substrates of each other in a cell-free, in vitro kinase assay The non-radioactive IVKA was unusable, so the established IVKA method was used to explore Src, Jak2, and STAT interactions for the remainder of the study. In this method, recombinant proteins are incubated with the Src inhibitor dasatinib, the Jak inhibitor pyridone 6, or vehicle control for 30 min in the presence of 10 mci ( - 32 P)ATP. As expected, Src acted as its own substrate and Src kinase activity was inhibited by dasatinib (Figure 10A, lanes 2-3). Likewise, Jak2 was autophosphorylated and its kinase activity was inhibited with pyridone 6 (Figure 10A, lanes 5 and 7). Surprisingly, dasatinib also partially inhibited Jak2 in this assay (Figure 10A, lane 8) an effect we had not observed in Jak2 that was immunoprecipitated from intact cells incubated with dasatinib (27). Since dasatinib inhibited recombinant Jak2 we used saracatinib, a more specific Src inhibitor, in subsequent experiments (Figure 10B-D). When Jak2 was incubated with saracatinib at 200 nm, there was no reduction in Jak2 phosphorylation (Figure 10B, lanes 5 and 8), indicating that Jak2 kinase activity was not being affected by this inhibitor. However, 200 nm saracatinib only partially inhibited Src (Figure 10B, lanes 2-3). Increasing the saracatinib concentration to 2 M led to full Src inhibition (Figure 10C, lanes 1 and 3) without Jak2 inhibtion (Figure 10C, lanes 2 and 4) To determine if Src could phosphorylate Jak2, we incubated the proteins together in the presence of pyridone 6. When Src and Jak2 were incubated together, we saw that both proteins were phosphorylated (Figure 10A-C, lane 6). 29
40 When pyridone 6 was added to the combination, inhibiting Jak2 kinase activity, Jak2 was still phosphorylated (Figure 10A-B, lane 10). This demonstrates that recombinant Jak2 is a direct Src substrate. When 2 M saracatinib was added to the combination of Src and Jak2 proteins, Src phosphorylation was observed (Figure 10C, lane 9). The degree of Src phosphorylation was similar to that of the Jak2 and Src combination demonstrating that Jak2 can phosphorylate recombinant Src. 30
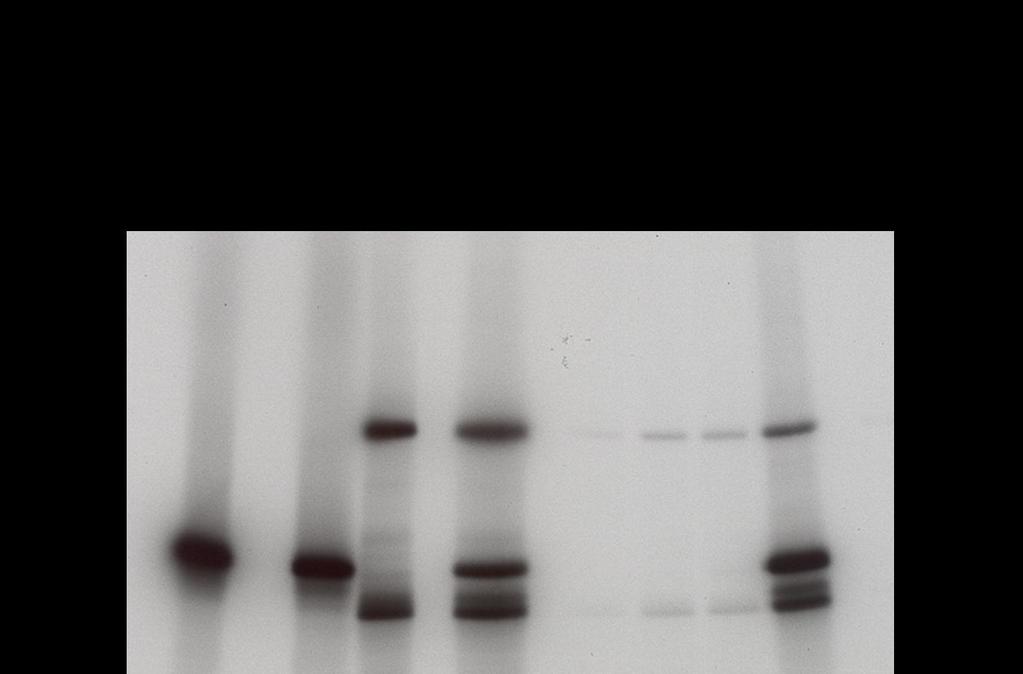
41 31

, 2 µm saracatinib (C), and/or 20 µm pyridone 6 as")
42 Figure 10. Recombinant Jak2 is a direct Src substrate and Src is a Jak2 substrate. Recombinant Src and Jak2 were incubated with 100 nm dasatinib (A), 200 nm saracatinib (B), 2 µm saracatinib (C), and/or 20 µm pyridone 6 as indicated in an IVKA and phospho proteins detected by autoradiography. 32
43 Src regulates STAT5 but not STAT3 activity in intact HNSCC cell lines Previously published data demonstrated that Src inhibition leads to marked and sustained decreased STAT5 phosphorylation despite recovery of Jak2 activity after 4-7 h (27, 34). This suggests that Src, and not Jak2, regulates STAT5 activity in HNSCC cells. To examine potential activators of STAT5 further, we treated OSC19 and TU167 cells with interleukin-6 (IL-6), epidermal growth factor (EGF), dasatinib, pyridone 6, or saracatinib and examined Src and STAT activation using Western blotting (Figure 11). As expected, IL-6 had no significant effect upon Src phosphorylation; dasatinib and 1 µm saracatinib inhibited Src completely; and pyridone 6 did not change Src phosphorylation levels. The lower concentration of saracatinib (75 nm) did not completely inhibit Src. STAT5 phosphorylation paralleled that of Src but STAT3 activation was unaffected by Src inhibition demonstrating that Src regulates STAT5 but not STAT3 activity. Jak regulates STAT3 but not STAT5 in intact HNSCC cell lines We found a significant increase in the phosphorylation of STAT3 in IL-6 stimulated samples and complete inhibition with pyridone 6. IL6 and pyridone 6 did not affect STAT5 activation (Figure 11). Since pyridone 6 is a pan-jak inhibitor and IL6 stimulates Jak2, we concluded that Jak was regulating STAT3 but not STAT5 in HNSCC cells. 33


44 Figure 11. Src predominately regulates STAT5 and Jak predominately regulates STAT3 in intact HNSCC cells. TU167 and OSC19 cells were treated with 100 ng/ml IL-6 or 10 ng/ml EGF for 5 min; 100 nm dasatinib for 1 h; 5 M pyridone 6 for 1 h; or 75 nm or 1 M saracatinib for 1 or 7 h as indicated. Cell lysates were subjected to Western blotting and expression of various proteins was measured using the indicated antibodies. 34
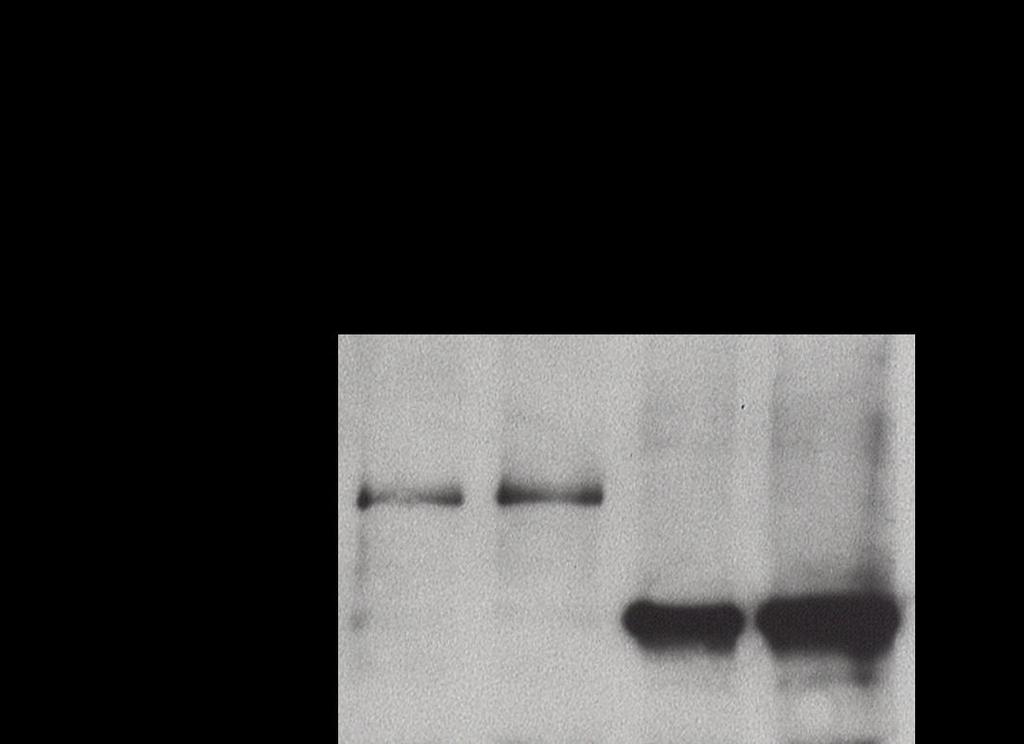
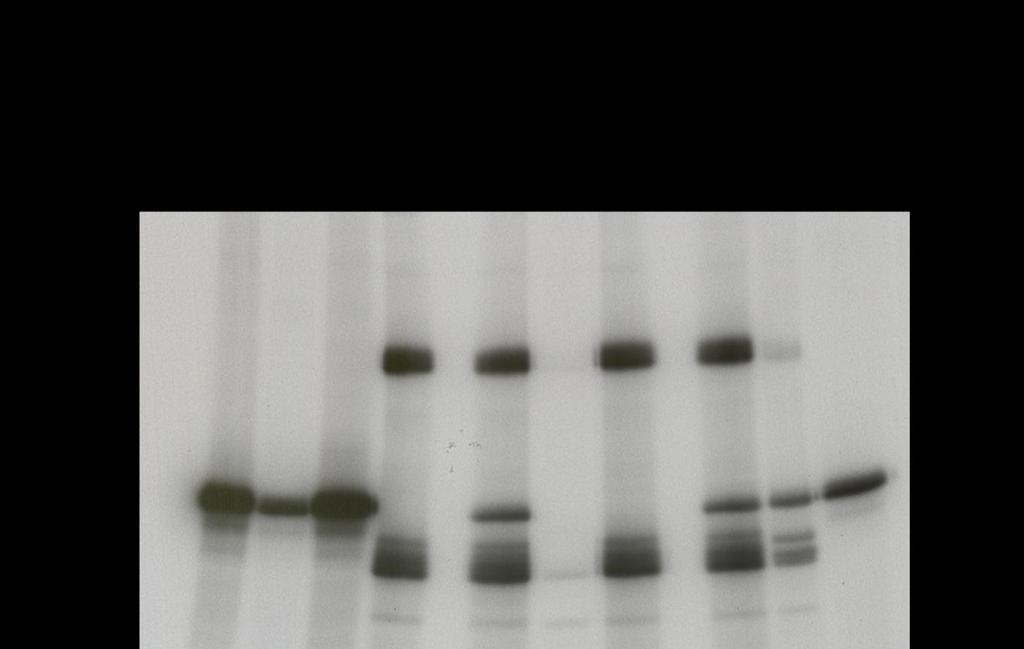
45 STAT5 is a direct substrate of both Jak2 and Src in a cell-free, in vitro kinase assay In intact HNSCC cells the inhibition of Src reduced STAT5 activation, but Jak inhibition did not. In contrast, others have demonstrated that STAT5 can be regulated by Jak family members (36, 37). We performed an IVKA to determine if STAT5 is a direct substrate of Src or Jak2. Recombinant Jak2, Src, and STAT5A proteins were incubated with 32 P-labeled ATP both with and without dasatinib or pyridone 6. Upon incubation of Src and STAT5A, STAT5A was phosphorylated indicating that STAT5A can be a direct substrate of Src (Figure 12, lane 2). When Src and STAT5A were incubated with dasatinib, the dasatinib inhibited Src kinase activity and no STAT5A phosphorylation occurred (Figure 12, lane 3). When Jak2 and STAT5A were incubated together, STAT5A was phosphorylated, indicating that STAT5A can also be a direct substrate of Jak2 in a cell free system (Figure 12, lane 4). When Jak2 and STAT5A were incubated with pyridone 6, this inhibitor inhibited Jak2 kinase activity so no STAT5A phosphorylation occurred (Figure 12, lane 5). STAT3 is a direct substrate of both Jak2 and Src in a cell free, in vitro kinase assay Analogous to the experiments above, we performed an IVKA to determine if STAT3 is a direct substrate of Src or Jak2. Upon incubation of Src and STAT3, STAT3 was phosphorylated indicating that STAT3 can be a direct substrate of Src (Figure 13, lane 4). When saracatinib was added, STAT3 phosphorylation was inhibited (Figure 13, lane 6). When Jak2 and STAT3 were incubated together, 35
46 STAT3 was phosphorylated. Likewise, Jak2 phosphorylated STAT3 which was inhibited by pyridone 6 (Figure 13, lanes 5 and 7). Although equimolar protein amounts were used, the degree of STAT5A and STAT3 phosphorylation by Jak2 kinase activity was much lower than that caused by Src. 36

47 Figure 12. STAT5A is a direct substrate of both Src and Jak2. Recombinant Jak2, Src, and STAT5A were incubated with dasatinib, pyridone 6, or vehicle control as indicated. The proteins were subjected to an IVKA with 32 P-labeled ATP and phosphorylated proteins detected by autoradiography. 37

48 Figure 13. STAT3 is a direct substrate of both Jak2 and Src. Recombinant Jak2, Src, and STAT3 were incubated with dasatinib or pyridone 6 or vehicle control as indicated. The proteins were subjected to an IVKA using with 32 P-labeled ATP and phosphorylated proteins detected by autoradiography. 38
49 EGFR activation increases Src and STAT5 activity in HNSCC cells Our data suggest that Src predominantly regulates STAT5 in HNSCC cells but Jak2 can also phosphorylate recombinant STAT5A, suggesting that substrate specificity in intact cells may be regulated by proximity. Furthermore, since STAT5, STAT3, Src and Jak2 are all located primarily in the cytoplasm (C. Nicholas, unpublished data) substrate specificity is likely regulated by differential protein binding partners. To determine the upstream activators of Src and STAT5 in HNSCC we first examined the effect of EGFR stimulation on their activation because it has previously been reported to stimulate Src in HNSCC (38). As expected, we did detect increased EGFR activation in the EGF and TGF-α stimulated cells. Both EGFR ligands led to activation of Src and STAT5 in TU167 and Osc19 cells although the stimulation was more marked in Osc19 cells (Figure 11 and 14). Basal expression of pegfr (Y1086) was undetectable and not affected by stimulation with IL-6 or incubation with dasatinib, pyridone 6 or saracatinib (Figure 11). Tyrosine 1086 was examined because it is not a Srcdependent phosphorylation site (Jak and Grb2-dependent) (39). To test for Src and EGFR interactions, we performed a Src immunoprecipitation (IP) from HNSCC cells incubated with EGF, TGF-α, or saracatinib (Figure 14A-B). In all conditions, Src and EGFR co-immunoprecipitated in TU167 cells but this interaction was not detectable in Osc-19 cells. Interestingly, despite robust Src activation detected on the Western blots following EGF and TGF- stimulation, this activation was not detected on the IP samples suggesting that the 39
 (Y1068) (Y1068) Figure 14.")

50 anti-total Src antibody used for the IP was saturated and preferentially bound to phosphorylated Src. (Y1068) (Y1068) (Y1068) Figure 14. EGFR co-immunoprecipitates with Src in TU167 cells. Src immunoprecipitation of cells treated with EGF, TGF-α, saracatinib for 1 and 7 h in (A) TU167 cells and (B) OSC19 cells. 40
.")
51 Band density Focal Adhesion Kinase may regulate Src Activity in HNSCC To further investigate the upstream activators of Src, we examined the effects of FAK activity on Src activation. We treated HNSCC cells with an ATPcompetitive FAK inhibitor, PF , for 4 hours at different concentrations (100 nm, 500 nm, 1 µm, and 3 µm). We saw that with increasing doses, there was a gradual decrease in Src activity, but this decrease was very minimal (Figure 15). Likewise there was also a decrease in ppaxillin, a known substrate of FAK. B FAK inhibitor (PF ), ng/ml Figure 8. FAK may play a role in Src regulation. TU167 cells were treated with increasing concentrations of PF and probed with pfak(y397), psrc, and ppaxillin (Y118) (A).(B) shows densitometry. 41
52 CHAPTER 4: DISCUSSION 42
53 In this study, we examined the interactions of Jak2, STAT3, STAT5 and Src in HNSCC cells. Previous work from our laboratory defined a feedback pathway in which sustained Src inhibition led to the loss of STAT5A-driven SOCS2 expression which allowed Jak2 kinase and STAT3 activities to recover. In this work, we demonstrated that Jak2, STAT3, and STAT5A are direct substrates of Src in an IVKA with purified proteins. Likewise Src, STAT3, and STAT5A are all substrates of Jak2 in an IVKA. In contrast, in intact HNSCC cells, Src inhibition leads primarily to STAT5 inhibition with transient, minimal effects on Jak2 and STAT3. Jak inhibition in intact HNSCC cells does not affect STAT5 or Src activity but does result in STAT3 inhibition. The contrast between the IVKA assays with recombinant proteins suggests that substrate specificity in intact cells may be regulated by differential protein binding partners. One potential protein that may mediate the interactions between these proteins is EGFR and we demonstrated that EGFR activation leads to Src and STAT5 activation. Jak and Src Interactions Previous studies have shown that in intact breast cancer cells, upon stimulation, Jak2 and Src bind to each other (40). However, it has not yet been shown whether they directly phosphorylate each other. This is the first report in HNSCC that investigates a novel relationship between Jak2 and Src. Our in vitro kinase assays show that Src is a direct substrate of Jak2 and Jak2 is a direct substrate of Src. These results are significant because within intact cells, when Src is inhibited, there is initial inhibition of Jak2 activity as well, but Jak2 is recovered after some time. However when we inhibit Jak2 in intact cells, we do not find Src 43
54 inhibition. This leads to the possibility that Jak and Src do not physically interact because they bind to distinct scaffolding proteins. These results demonstrate that Jak2 is a direct Src substrate in intact cells but that Src is not a Jak2 substrate. As discussed below, possible reasons for the discrepancies between the IVKAs and the intact cells are that the Jak2 protein used in the IVKA is a truncated protein or that Src is more easily accessible for phosphorylation by Jak2 in the cell free system. Regulation of STAT3 and STAT5 Activation The STAT family of transcription factors, especially STAT3 and STAT5, regulates oncogenic signaling in many different tumor types (41). STATs are Src substrates and can mediate Src s biological effects including proliferation, survival, and angiogenesis (42). In HNSCC cells, c-src interacts with STAT3 and STAT5 (co-immunoprecipitation) and Src inhibition results in reduced STAT3 and STAT5 activation and reduced cell proliferation (43). Inhibition of STAT3 in HNSCC leads to increased apoptosis, decreased proliferation, and decreased tumor size (44, 45). STAT3 activation leads to the increased expression of downstream target genes (e.g., Bcl-XL, cyclin D1, survivin) and increased cell proliferation and tumor growth in vivo (46). STAT3 is essential for Src-mediated angiogenesis; following activation by Src, STAT3 binds to the promoter of vascular endothelial growth factor (VEGF), leading to increased VEGF production (47, 48). Activation of STAT3 is required for v-src-mediated transformation (49) suggesting that STAT3 is a key mediator of the oncogenic effects of c-src. 44
55 In addition to regulation by Src, STAT3 can be activated by Jaks. Jaks are essential for cytokine signaling. Jaks constitutively bind to cytokine receptors which do not have intrinsic kinase activity. Upon ligand binding, the cytokine receptor undergoes oligomerization resulting in Jak activation by trans-phosphorylation. The activated Jak molecules then phosphorylate the cytokine receptor allowing for the binding of the monomeric, inactive STATs that are present in the cytoplasm. Once bound to the cytokine receptors, STATs become Jak substrates. Phosphorylated STATs undergo dimerization and nuclear translocation (42). The molecular mechanisms leading to Jak activation are not fully understood and this simplified model understates the complexity of these pathway interactions. Two key examples are that: activation of Jak2 can occur, albeit at a lower level, in the absence of Y1007/1008 phosphorylation, theoretically obviating the need for activation by an upstream kinase (50); and Jaks can be activated by oncogenic tyrosine kinases, independent of cytokine receptors (51-53). There has been evidence to suggest that Jak does not activate STAT5. After sustained Src inhibition as already mentioned, Jak2 is reactivated. With Jak2 reactivation STAT3 reactivation was also seen, but STAT5 remained inhibited (27). As part of this study we also looked at the regulation of STAT3 and STAT5 by Src and Jak2. Our results demonstrate that STAT5A can be directly phosphorylated by both Src and Jak2 in an IVKA but that in intact HNSCC cells Src, but not Jak2, regulates STAT5 activity. Similarly, we also found that STAT3 can be phosphorylated by both Jak2 and Src in an IVKA but in intact cells Jak2 regulates STAT3 activity. Another discrepancy between the IVKA and the intact cell data was 45
56 that the degree of phosphorylation of the STAT proteins by Jak2 was much lower than that of Src. There are several possible reasons to explain the discrepancies between the two systems. First a receptor and/or adaptor proteins may be needed to complex with Jak2 and STAT3 for STAT3 phosphorylation to occur. This is consistent with what we know about how Jak functions in cells. This complex may be needed to affect conformational changes in the involved proteins or simply to bring them into proximity for enzymatic interactions. Another reason for the lower Jak2 kinase activity toward STAT3 in the IVKA may be due to the recombinant Jak2 being a truncated protein. While the domains that we think are the most important for kinase activity and regulation, the JH1 and JH2 domains, are still present, the recombinant Jak2 lacks the 20 kda region on the N-terminal end. This loss may affect Jak2 interactions with its substrates. It is also possible that Jak2 may have a slower enzyme kinetics or that the accessibility of both Jak2 and Src to the STAT proteins may be better In Vitro compared to intact cells. When we investigated the regulation of STAT3 and STAT5 in intact HNSCC cells, we saw that Jak2 was the primary regulator of STAT3. When Src was inhibited with dasatinib and saracatinib, there was no sustained decrease in STAT3 activity, but when Jak2 was inhibited with pyridone 6, we saw complete inhibition of STAT3. When we inhibited Jak2 and looked at STAT5 activation, there was no change; whereas when Src was inhibited, there was complete inhibition of STAT5. These results suggest that Src primarily regulates STAT5. 46
57 Src Activation Src participates in several cellular functions in cancer cells including cell cycle progression, immune recognition, adhesion, spreading, migration, apoptosis regulation, and differentiation (54, 55). Although v-src, which lacks the C-terminal regulatory domain and is constitutively active, transforms fibroblasts (56), c-src is only weakly transforming. HNSCC cells transfected with a dominant active c-src showed enhanced tumor growth and invasion (57). Inhibition of c-src activity using both molecular approaches and pharmacologic inhibitors leads to reduced anchorage-independent growth (58, 59), proliferation (60, 61), tumor growth in vivo (62, 63), invasion, migration (64-71), metastasis (72-76), vascularity (77-79), and survival (59). In HNSCC specifically, inhibition of c-src leads to decreased invasion, migration and proliferation (38, 43, 57, 64, 80). Aberrant c-src expression has been demonstrated in multiple epithelial cancers (81) where its expression and activation correlate with malignant progression (82-88), advanced clinical stage, and poor survival (89). c-src is expressed in areas of hyperproliferation in HNSCC, dysplastic oral epithelium, and benign oral mucosal lesions (90). Several Src members are expressed and activated in multiple HNSCC cell lines (38, 43, 91). Expression of activated c-src is higher in human HNSCC tumor tissue than in normal mucosa and correlates with an invasive, poorly differentiated phenotype and advanced nodal stage (15). Src can be activated by Platelet Derived Growth Factor Receptor (PDGFR), EGFR Focal Adhesion Kinase (FAK), G-coupled protein receptors (GCPRs), and integrins (16-21). We examined the effect of EGFR on Src activation because 47
G-Protein Signaling. Introduction to intracellular signaling. Dr. SARRAY Sameh, Ph.D
 G-Protein Signaling Introduction to intracellular signaling Dr. SARRAY Sameh, Ph.D Cell signaling Cells communicate via extracellular signaling molecules (Hormones, growth factors and neurotransmitters
G-Protein Signaling Introduction to intracellular signaling Dr. SARRAY Sameh, Ph.D Cell signaling Cells communicate via extracellular signaling molecules (Hormones, growth factors and neurotransmitters
Signaling. Dr. Sujata Persad Katz Group Centre for Pharmacy & Health research
 Signaling Dr. Sujata Persad 3-020 Katz Group Centre for Pharmacy & Health research E-mail:sujata.persad@ualberta.ca 1 Growth Factor Receptors and Other Signaling Pathways What we will cover today: How
Signaling Dr. Sujata Persad 3-020 Katz Group Centre for Pharmacy & Health research E-mail:sujata.persad@ualberta.ca 1 Growth Factor Receptors and Other Signaling Pathways What we will cover today: How
Enzyme-coupled Receptors. Cell-surface receptors 1. Ion-channel-coupled receptors 2. G-protein-coupled receptors 3. Enzyme-coupled receptors
 Enzyme-coupled Receptors Cell-surface receptors 1. Ion-channel-coupled receptors 2. G-protein-coupled receptors 3. Enzyme-coupled receptors Cell-surface receptors allow a flow of ions across the plasma
Enzyme-coupled Receptors Cell-surface receptors 1. Ion-channel-coupled receptors 2. G-protein-coupled receptors 3. Enzyme-coupled receptors Cell-surface receptors allow a flow of ions across the plasma
Phospho-AKT Sampler Kit
 Phospho-AKT Sampler Kit E 0 5 1 0 0 3 Kits Includes Cat. Quantity Application Reactivity Source Akt (Ab-473) Antibody E021054-1 50μg/50μl IHC, WB Human, Mouse, Rat Rabbit Akt (Phospho-Ser473) Antibody
Phospho-AKT Sampler Kit E 0 5 1 0 0 3 Kits Includes Cat. Quantity Application Reactivity Source Akt (Ab-473) Antibody E021054-1 50μg/50μl IHC, WB Human, Mouse, Rat Rabbit Akt (Phospho-Ser473) Antibody
Receptor mediated Signal Transduction
 Receptor mediated Signal Transduction G-protein-linked receptors adenylyl cyclase camp PKA Organization of receptor protein-tyrosine kinases From G.M. Cooper, The Cell. A molecular approach, 2004, third
Receptor mediated Signal Transduction G-protein-linked receptors adenylyl cyclase camp PKA Organization of receptor protein-tyrosine kinases From G.M. Cooper, The Cell. A molecular approach, 2004, third
Discovery and Optimization of Inhibitors of STAT3 Activation for the Treatment of Squamous Cell Carcinoma of the Head and Neck
 Discovery and ptimization of Inhibitors of STAT3 Activation for the Treatment of Squamous Cell Carcinoma of the Head and Neck Feng Zhang Wipf Group Research Topic Seminar 02-09-2013 1 Feng Zhang @ Wipf
Discovery and ptimization of Inhibitors of STAT3 Activation for the Treatment of Squamous Cell Carcinoma of the Head and Neck Feng Zhang Wipf Group Research Topic Seminar 02-09-2013 1 Feng Zhang @ Wipf
Cell Signaling part 2
 15 Cell Signaling part 2 Functions of Cell Surface Receptors Other cell surface receptors are directly linked to intracellular enzymes. The largest family of these is the receptor protein tyrosine kinases,
15 Cell Signaling part 2 Functions of Cell Surface Receptors Other cell surface receptors are directly linked to intracellular enzymes. The largest family of these is the receptor protein tyrosine kinases,
MCB*4010 Midterm Exam / Winter 2008
 MCB*4010 Midterm Exam / Winter 2008 Name: ID: Instructions: Answer all 4 questions. The number of marks for each question indicates how many points you need to provide. Write your answers in point form,
MCB*4010 Midterm Exam / Winter 2008 Name: ID: Instructions: Answer all 4 questions. The number of marks for each question indicates how many points you need to provide. Write your answers in point form,
Protocol for Gene Transfection & Western Blotting
 The schedule and the manual of basic techniques for cell culture Advanced Protocol for Gene Transfection & Western Blotting Schedule Day 1 26/07/2008 Transfection Day 3 28/07/2008 Cell lysis Immunoprecipitation
The schedule and the manual of basic techniques for cell culture Advanced Protocol for Gene Transfection & Western Blotting Schedule Day 1 26/07/2008 Transfection Day 3 28/07/2008 Cell lysis Immunoprecipitation
THE HALLMARKS OF CANCER
 THE HALLMARKS OF CANCER ONCOGENES - Most of the oncogenes were first identified in retroviruses: EGFR (ErbB), Src, Ras, Myc, PI3K and others (slightly more than 30) - Mutated cellular genes incorporated
THE HALLMARKS OF CANCER ONCOGENES - Most of the oncogenes were first identified in retroviruses: EGFR (ErbB), Src, Ras, Myc, PI3K and others (slightly more than 30) - Mutated cellular genes incorporated
The Schedule and the Manual of Basic Techniques for Cell Culture
 The Schedule and the Manual of Basic Techniques for Cell Culture 1 Materials Calcium Phosphate Transfection Kit: Invitrogen Cat.No.K2780-01 Falcon tube (Cat No.35-2054:12 x 75 mm, 5 ml tube) Cell: 293
The Schedule and the Manual of Basic Techniques for Cell Culture 1 Materials Calcium Phosphate Transfection Kit: Invitrogen Cat.No.K2780-01 Falcon tube (Cat No.35-2054:12 x 75 mm, 5 ml tube) Cell: 293
SUPPLEMENTARY INFORMATION
 Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
KEY CONCEPT QUESTIONS IN SIGNAL TRANSDUCTION
 Signal Transduction - Part 2 Key Concepts - Receptor tyrosine kinases control cell metabolism and proliferation Growth factor signaling through Ras Mutated cell signaling genes in cancer cells are called
Signal Transduction - Part 2 Key Concepts - Receptor tyrosine kinases control cell metabolism and proliferation Growth factor signaling through Ras Mutated cell signaling genes in cancer cells are called
RayBio KinaseSTAR TM Akt Activity Assay Kit
 Activity Assay Kit User Manual Version 1.0 March 13, 2015 RayBio KinaseSTAR TM Akt Activity Kit Protocol (Cat#: 68AT-Akt-S40) RayBiotech, Inc. We Provide You With Excellent Support And Service Tel:(Toll
Activity Assay Kit User Manual Version 1.0 March 13, 2015 RayBio KinaseSTAR TM Akt Activity Kit Protocol (Cat#: 68AT-Akt-S40) RayBiotech, Inc. We Provide You With Excellent Support And Service Tel:(Toll
Signal Transduction Pathway Smorgasbord
 Molecular Cell Biology Lecture. Oct 28, 2014 Signal Transduction Pathway Smorgasbord Ron Bose, MD PhD Biochemistry and Molecular Cell Biology Programs Washington University School of Medicine Outline 1.
Molecular Cell Biology Lecture. Oct 28, 2014 Signal Transduction Pathway Smorgasbord Ron Bose, MD PhD Biochemistry and Molecular Cell Biology Programs Washington University School of Medicine Outline 1.
Biol403 MAP kinase signalling
 Biol403 MAP kinase signalling The mitogen activated protein kinase (MAPK) pathway is a signalling cascade activated by a diverse range of effectors. The cascade regulates many cellular activities including
Biol403 MAP kinase signalling The mitogen activated protein kinase (MAPK) pathway is a signalling cascade activated by a diverse range of effectors. The cascade regulates many cellular activities including
Principles of Genetics and Molecular Biology
 Cell signaling Dr. Diala Abu-Hassan, DDS, PhD School of Medicine Dr.abuhassand@gmail.com Principles of Genetics and Molecular Biology www.cs.montana.edu Modes of cell signaling Direct interaction of a
Cell signaling Dr. Diala Abu-Hassan, DDS, PhD School of Medicine Dr.abuhassand@gmail.com Principles of Genetics and Molecular Biology www.cs.montana.edu Modes of cell signaling Direct interaction of a
Effects of Second Messengers
 Effects of Second Messengers Inositol trisphosphate Diacylglycerol Opens Calcium Channels Binding to IP 3 -gated Channel Cooperative binding Activates Protein Kinase C is required Phosphorylation of many
Effects of Second Messengers Inositol trisphosphate Diacylglycerol Opens Calcium Channels Binding to IP 3 -gated Channel Cooperative binding Activates Protein Kinase C is required Phosphorylation of many
CYTOKINE RECEPTORS AND SIGNAL TRANSDUCTION
 CYTOKINE RECEPTORS AND SIGNAL TRANSDUCTION What is Cytokine? Secreted popypeptide (protein) involved in cell-to-cell signaling. Acts in paracrine or autocrine fashion through specific cellular receptors.
CYTOKINE RECEPTORS AND SIGNAL TRANSDUCTION What is Cytokine? Secreted popypeptide (protein) involved in cell-to-cell signaling. Acts in paracrine or autocrine fashion through specific cellular receptors.
UNIVERSITY OF MEDICINE AND PHARMACY CRAIOVA PhD SCHOOL. PhD THESIS
 UNIVERSITY OF MEDICINE AND PHARMACY CRAIOVA PhD SCHOOL PhD THESIS THE IMPORTANCE OF TUMOR ANGIOGENESIS IN CEREBRAL TUMOR DIAGNOSIS AND THERAPY ABSTRACT PhD COORDINATOR: Prof. univ. dr. DRICU Anica PhD
UNIVERSITY OF MEDICINE AND PHARMACY CRAIOVA PhD SCHOOL PhD THESIS THE IMPORTANCE OF TUMOR ANGIOGENESIS IN CEREBRAL TUMOR DIAGNOSIS AND THERAPY ABSTRACT PhD COORDINATOR: Prof. univ. dr. DRICU Anica PhD
Supplementary data Supplementary Figure 1 Supplementary Figure 2
 Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
(PDGF), 9 ( -2 (FGF-2), SMO
, 9 ( -2 (FGF-2), SMO") Abstract An ethanol extract from shark muscle has been shown to have potent angiogenic activity when mixed together with olive oil in a ratio of 1part extract to 9 parts olive oil. This mixture has been
Abstract An ethanol extract from shark muscle has been shown to have potent angiogenic activity when mixed together with olive oil in a ratio of 1part extract to 9 parts olive oil. This mixture has been
Signaling Through Immune System Receptors (Ch. 7)
") Signaling Through Immune System Receptors (Ch. 7) 1. General principles of signal transduction and propagation. 2. Antigen receptor signaling and lymphocyte activation. 3. Other receptors and signaling
Signaling Through Immune System Receptors (Ch. 7) 1. General principles of signal transduction and propagation. 2. Antigen receptor signaling and lymphocyte activation. 3. Other receptors and signaling
Molecular Cell Biology - Problem Drill 19: Cell Signaling Pathways and Gene Expression
 Molecular Cell Biology - Problem Drill 19: Cell Signaling Pathways and Gene Expression Question No. 1 of 10 1. Which statement about cell signaling is correct? Question #1 (A) Cell signaling involves receiving
Molecular Cell Biology - Problem Drill 19: Cell Signaling Pathways and Gene Expression Question No. 1 of 10 1. Which statement about cell signaling is correct? Question #1 (A) Cell signaling involves receiving
A Hepatocyte Growth Factor Receptor (Met) Insulin Receptor hybrid governs hepatic glucose metabolism SUPPLEMENTARY FIGURES, LEGENDS AND METHODS
 Insulin Receptor hybrid governs hepatic glucose metabolism SUPPLEMENTARY FIGURES, LEGENDS AND METHODS") A Hepatocyte Growth Factor Receptor (Met) Insulin Receptor hybrid governs hepatic glucose metabolism Arlee Fafalios, Jihong Ma, Xinping Tan, John Stoops, Jianhua Luo, Marie C. DeFrances and Reza Zarnegar
A Hepatocyte Growth Factor Receptor (Met) Insulin Receptor hybrid governs hepatic glucose metabolism Arlee Fafalios, Jihong Ma, Xinping Tan, John Stoops, Jianhua Luo, Marie C. DeFrances and Reza Zarnegar
Lecture: CHAPTER 13 Signal Transduction Pathways
 Lecture: 10 17 2016 CHAPTER 13 Signal Transduction Pathways Chapter 13 Outline Signal transduction cascades have many components in common: 1. Release of a primary message as a response to a physiological
Lecture: 10 17 2016 CHAPTER 13 Signal Transduction Pathways Chapter 13 Outline Signal transduction cascades have many components in common: 1. Release of a primary message as a response to a physiological
Growth and Differentiation Phosphorylation Sampler Kit
 Growth and Differentiation Phosphorylation Sampler Kit E 0 5 1 0 1 4 Kits Includes Cat. Quantity Application Reactivity Source Akt (Phospho-Ser473) E011054-1 50μg/50μl IHC, WB Human, Mouse, Rat Rabbit
Growth and Differentiation Phosphorylation Sampler Kit E 0 5 1 0 1 4 Kits Includes Cat. Quantity Application Reactivity Source Akt (Phospho-Ser473) E011054-1 50μg/50μl IHC, WB Human, Mouse, Rat Rabbit
EGFR: fundamenteel en klinisch
 EGFR: fundamenteel en klinisch Guido Lammering MAASTRO Clinic Maastricht, NL What is EGFR?? The EGFR some facts 1186 amino acids 170 kda Expressed by all cells of epithelial origin Increased activation
EGFR: fundamenteel en klinisch Guido Lammering MAASTRO Clinic Maastricht, NL What is EGFR?? The EGFR some facts 1186 amino acids 170 kda Expressed by all cells of epithelial origin Increased activation
VIII Curso Internacional del PIRRECV. Some molecular mechanisms of cancer
 VIII Curso Internacional del PIRRECV Some molecular mechanisms of cancer Laboratorio de Comunicaciones Celulares, Centro FONDAP Estudios Moleculares de la Celula (CEMC), ICBM, Facultad de Medicina, Universidad
VIII Curso Internacional del PIRRECV Some molecular mechanisms of cancer Laboratorio de Comunicaciones Celulares, Centro FONDAP Estudios Moleculares de la Celula (CEMC), ICBM, Facultad de Medicina, Universidad
Antibodies. Product Name Cat. No. Size. GH (Growth Hormone) Antibody µg, 100 µg GM-CSF Antibody µg, 100 µg
 Antibody µg, 100 µg GM-CSF Antibody µg, 100 µg") JAK/STAT Signaling The JAK/STAT signaling pathway transmits a signal from outside the cell eventually resulting in DNA transcription and activity in the cell. Leptin (a product of the obese gene) can induce
JAK/STAT Signaling The JAK/STAT signaling pathway transmits a signal from outside the cell eventually resulting in DNA transcription and activity in the cell. Leptin (a product of the obese gene) can induce
Concise Reference. HER2 Testing in Breast Cancer. Mary Falzon, Angelica Fasolo, Michael Gandy, Luca Gianni & Stefania Zambelli
 Concise Reference Testing in Breast Cancer Mary Falzon, Angelica Fasolo, Michael Gandy, Luca Gianni & Stefania Zambelli Extracted from Handbook of -Targeted Agents in Breast Cancer ublished by Springer
Concise Reference Testing in Breast Cancer Mary Falzon, Angelica Fasolo, Michael Gandy, Luca Gianni & Stefania Zambelli Extracted from Handbook of -Targeted Agents in Breast Cancer ublished by Springer
MEK1 Assay Kit 1 Catalog # Lot # 16875
 MEK1 Assay Kit 1 Kit Components Assay Dilution Buffer (ADB), Catalog # 20-108. Three vials, each containing 1.0ml of assay dilution buffer (20mM MOPS, ph 7.2, 25mM ß-glycerol phosphate, 5mM EGTA, 1mM sodium
MEK1 Assay Kit 1 Kit Components Assay Dilution Buffer (ADB), Catalog # 20-108. Three vials, each containing 1.0ml of assay dilution buffer (20mM MOPS, ph 7.2, 25mM ß-glycerol phosphate, 5mM EGTA, 1mM sodium
7/6/2015. Cancer Related Deaths: United States. Management of NSCLC TODAY. Emerging mutations as predictive biomarkers in lung cancer: Overview
 Emerging mutations as predictive biomarkers in lung cancer: Overview Kirtee Raparia, MD Assistant Professor of Pathology Cancer Related Deaths: United States Men Lung and bronchus 28% Prostate 10% Colon
Emerging mutations as predictive biomarkers in lung cancer: Overview Kirtee Raparia, MD Assistant Professor of Pathology Cancer Related Deaths: United States Men Lung and bronchus 28% Prostate 10% Colon
Cell cycle, signaling to cell cycle, and molecular basis of oncogenesis
 Cell cycle, signaling to cell cycle, and molecular basis of oncogenesis MUDr. Jiří Vachtenheim, CSc. CELL CYCLE - SUMMARY Basic terminology: Cyclins conserved proteins with homologous regions; their cellular
Cell cycle, signaling to cell cycle, and molecular basis of oncogenesis MUDr. Jiří Vachtenheim, CSc. CELL CYCLE - SUMMARY Basic terminology: Cyclins conserved proteins with homologous regions; their cellular
Signal Transduction: G-Protein Coupled Receptors
 Signal Transduction: G-Protein Coupled Receptors Federle, M. (2017). Lectures 4-5: Signal Transduction parts 1&2: nuclear receptors and GPCRs. Lecture presented at PHAR 423 Lecture in UIC College of Pharmacy,
Signal Transduction: G-Protein Coupled Receptors Federle, M. (2017). Lectures 4-5: Signal Transduction parts 1&2: nuclear receptors and GPCRs. Lecture presented at PHAR 423 Lecture in UIC College of Pharmacy,
Chapter 15: Signal transduction
 Chapter 15: Signal transduction Know the terminology: Enzyme-linked receptor, G-protein linked receptor, nuclear hormone receptor, G-protein, adaptor protein, scaffolding protein, SH2 domain, MAPK, Ras,
Chapter 15: Signal transduction Know the terminology: Enzyme-linked receptor, G-protein linked receptor, nuclear hormone receptor, G-protein, adaptor protein, scaffolding protein, SH2 domain, MAPK, Ras,
Propagation of the Signal
 OpenStax-CNX module: m44452 1 Propagation of the Signal OpenStax College This work is produced by OpenStax-CNX and licensed under the Creative Commons Attribution License 3.0 By the end of this section,
OpenStax-CNX module: m44452 1 Propagation of the Signal OpenStax College This work is produced by OpenStax-CNX and licensed under the Creative Commons Attribution License 3.0 By the end of this section,
RAS Genes. The ras superfamily of genes encodes small GTP binding proteins that are responsible for the regulation of many cellular processes.
 ۱ RAS Genes The ras superfamily of genes encodes small GTP binding proteins that are responsible for the regulation of many cellular processes. Oncogenic ras genes in human cells include H ras, N ras,
۱ RAS Genes The ras superfamily of genes encodes small GTP binding proteins that are responsible for the regulation of many cellular processes. Oncogenic ras genes in human cells include H ras, N ras,
TFEB-mediated increase in peripheral lysosomes regulates. Store Operated Calcium Entry
 TFEB-mediated increase in peripheral lysosomes regulates Store Operated Calcium Entry Luigi Sbano, Massimo Bonora, Saverio Marchi, Federica Baldassari, Diego L. Medina, Andrea Ballabio, Carlotta Giorgi
TFEB-mediated increase in peripheral lysosomes regulates Store Operated Calcium Entry Luigi Sbano, Massimo Bonora, Saverio Marchi, Federica Baldassari, Diego L. Medina, Andrea Ballabio, Carlotta Giorgi
Cancer. The fundamental defect is. unregulated cell division. Properties of Cancerous Cells. Causes of Cancer. Altered growth and proliferation
 Cancer The fundamental defect is unregulated cell division. Properties of Cancerous Cells Altered growth and proliferation Loss of growth factor dependence Loss of contact inhibition Immortalization Alterated
Cancer The fundamental defect is unregulated cell division. Properties of Cancerous Cells Altered growth and proliferation Loss of growth factor dependence Loss of contact inhibition Immortalization Alterated
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION doi:1.138/nature9814 a A SHARPIN FL B SHARPIN ΔNZF C SHARPIN T38L, F39V b His-SHARPIN FL -1xUb -2xUb -4xUb α-his c Linear 4xUb -SHARPIN FL -SHARPIN TF_LV -SHARPINΔNZF -SHARPIN
SUPPLEMENTARY INFORMATION doi:1.138/nature9814 a A SHARPIN FL B SHARPIN ΔNZF C SHARPIN T38L, F39V b His-SHARPIN FL -1xUb -2xUb -4xUb α-his c Linear 4xUb -SHARPIN FL -SHARPIN TF_LV -SHARPINΔNZF -SHARPIN
Genome of Hepatitis B Virus. VIRAL ONCOGENE Dr. Yahwardiah Siregar, PhD Dr. Sry Suryani Widjaja, Mkes Biochemistry Department
 Genome of Hepatitis B Virus VIRAL ONCOGENE Dr. Yahwardiah Siregar, PhD Dr. Sry Suryani Widjaja, Mkes Biochemistry Department Proto Oncogen and Oncogen Oncogen Proteins that possess the ability to cause
Genome of Hepatitis B Virus VIRAL ONCOGENE Dr. Yahwardiah Siregar, PhD Dr. Sry Suryani Widjaja, Mkes Biochemistry Department Proto Oncogen and Oncogen Oncogen Proteins that possess the ability to cause
CHAPTER 3: EGFR ACTIVATION IMPACTS ON FAK PROTEIN EXPRESSION AND PHOSPHORYLATION STATUS IN HOSCC CELL LINES
 CHAPTER 3: EGFR ACTIVATION IMPACTS ON FAK PROTEIN EXPRESSION AND PHOSPHORYLATION STATUS IN HOSCC CELL LINES 3.1 Introduction Developmental processes such as cell migration depend on signals from both the
CHAPTER 3: EGFR ACTIVATION IMPACTS ON FAK PROTEIN EXPRESSION AND PHOSPHORYLATION STATUS IN HOSCC CELL LINES 3.1 Introduction Developmental processes such as cell migration depend on signals from both the
The PI3K/AKT axis. Dr. Lucio Crinò Medical Oncology Division Azienda Ospedaliera-Perugia. Introduction
 The PI3K/AKT axis Dr. Lucio Crinò Medical Oncology Division Azienda Ospedaliera-Perugia Introduction Phosphoinositide 3-kinase (PI3K) pathway are a family of lipid kinases discovered in 1980s. They have
The PI3K/AKT axis Dr. Lucio Crinò Medical Oncology Division Azienda Ospedaliera-Perugia Introduction Phosphoinositide 3-kinase (PI3K) pathway are a family of lipid kinases discovered in 1980s. They have
2013 W. H. Freeman and Company. 12 Signal Transduction
 2013 W. H. Freeman and Company 12 Signal Transduction CHAPTER 12 Signal Transduction Key topics: General features of signal transduction Structure and function of G protein coupled receptors Structure
2013 W. H. Freeman and Company 12 Signal Transduction CHAPTER 12 Signal Transduction Key topics: General features of signal transduction Structure and function of G protein coupled receptors Structure
C) The graph should look exactly like the graph on the left (Mut1 cells + Mating Pheromone for 3 hours at 25 degrees). The cells arrest in G1.
 The graph should look exactly like the graph on the left (Mut1 cells + Mating Pheromone for 3 hours at 25 degrees). The cells arrest in G1.") 706-2000-Exam 4 Answer Key 1) The question asks you to explain peaks A and B in the top graph. The other two graphs were there to give you hints. The question did not ask for these other two graphs to
706-2000-Exam 4 Answer Key 1) The question asks you to explain peaks A and B in the top graph. The other two graphs were there to give you hints. The question did not ask for these other two graphs to
Data Sheet TIGIT / NFAT Reporter - Jurkat Cell Line Catalog #60538
 Data Sheet TIGIT / NFAT Reporter - Jurkat Cell Line Catalog #60538 Background: TIGIT is a co-inhibitory receptor that is highly expressed in Natural Killer (NK) cells, activated CD4+, CD8+ and regulatory
Data Sheet TIGIT / NFAT Reporter - Jurkat Cell Line Catalog #60538 Background: TIGIT is a co-inhibitory receptor that is highly expressed in Natural Killer (NK) cells, activated CD4+, CD8+ and regulatory
Supplementary Materials for
 www.sciencesignaling.org/cgi/content/full/8/364/ra18/dc1 Supplementary Materials for The tyrosine phosphatase (Pez) inhibits metastasis by altering protein trafficking Leila Belle, Naveid Ali, Ana Lonic,
www.sciencesignaling.org/cgi/content/full/8/364/ra18/dc1 Supplementary Materials for The tyrosine phosphatase (Pez) inhibits metastasis by altering protein trafficking Leila Belle, Naveid Ali, Ana Lonic,
Signal-Transduction Cascades - 2. The Phosphoinositide Cascade
 Signal-Transduction Cascades - 2 The Phosphoinositide Cascade Calcium ion as a second messenger Tyrosine kinase and receptor dimerization scribd.com Faisal Khatib JU The Phosphoinositide Cascade Used by
Signal-Transduction Cascades - 2 The Phosphoinositide Cascade Calcium ion as a second messenger Tyrosine kinase and receptor dimerization scribd.com Faisal Khatib JU The Phosphoinositide Cascade Used by
Cancer. The fundamental defect is. unregulated cell division. Properties of Cancerous Cells. Causes of Cancer. Altered growth and proliferation
 Cancer The fundamental defect is unregulated cell division. Properties of Cancerous Cells Altered growth and proliferation Loss of growth factor dependence Loss of contact inhibition Immortalization Alterated
Cancer The fundamental defect is unregulated cell division. Properties of Cancerous Cells Altered growth and proliferation Loss of growth factor dependence Loss of contact inhibition Immortalization Alterated
Supporting Online Material Material and Methods References Supplemental Figures S1, S2, and S3
 Supporting Online Material Material and Methods References Supplemental Figures S1, S2, and S3 Sarbassov et al. 1 Material and Methods Materials Reagents were obtained from the following sources: protein
Supporting Online Material Material and Methods References Supplemental Figures S1, S2, and S3 Sarbassov et al. 1 Material and Methods Materials Reagents were obtained from the following sources: protein
Signal Transduction Pathways. Part 2
 Signal Transduction Pathways Part 2 GPCRs G-protein coupled receptors > 700 GPCRs in humans Mediate responses to senses taste, smell, sight ~ 1000 GPCRs mediate sense of smell in mouse Half of all known
Signal Transduction Pathways Part 2 GPCRs G-protein coupled receptors > 700 GPCRs in humans Mediate responses to senses taste, smell, sight ~ 1000 GPCRs mediate sense of smell in mouse Half of all known
Lecture 7: Signaling Through Lymphocyte Receptors
 Lecture 7: Signaling Through Lymphocyte Receptors Questions to Consider After recognition of its cognate MHC:peptide, how does the T cell receptor activate immune response genes? What are the structural
Lecture 7: Signaling Through Lymphocyte Receptors Questions to Consider After recognition of its cognate MHC:peptide, how does the T cell receptor activate immune response genes? What are the structural
TSH Receptor Monoclonal Antibody (49) Catalog Number MA3-218 Product data sheet
 Catalog Number MA3-218 Product data sheet") Website: thermofisher.com Customer Service (US): 1 800 955 6288 ext. 1 Technical Support (US): 1 800 955 6288 ext. 441 TSH Receptor Monoclonal Antibody (49) Catalog Number MA3-218 Product data sheet Details
Website: thermofisher.com Customer Service (US): 1 800 955 6288 ext. 1 Technical Support (US): 1 800 955 6288 ext. 441 TSH Receptor Monoclonal Antibody (49) Catalog Number MA3-218 Product data sheet Details
1. Activated receptor tyrosine kinases (RTKs) phosphorylates themselves
 phosphorylates themselves") Enzyme-coupled receptors Transmembrane proteins Ligand-binding domain on the outer surface Cytoplasmic domain acts as an enzyme itself or forms a complex with enzyme 1. Activated receptor tyrosine kinases
Enzyme-coupled receptors Transmembrane proteins Ligand-binding domain on the outer surface Cytoplasmic domain acts as an enzyme itself or forms a complex with enzyme 1. Activated receptor tyrosine kinases
Src-INACTIVE / Src-INACTIVE
 Biology 169 -- Exam 1 February 2003 Answer each question, noting carefully the instructions for each. Repeat- Read the instructions for each question before answering!!! Be as specific as possible in each
Biology 169 -- Exam 1 February 2003 Answer each question, noting carefully the instructions for each. Repeat- Read the instructions for each question before answering!!! Be as specific as possible in each
Cellular Signaling Pathways. Signaling Overview
 Cellular Signaling Pathways Signaling Overview Signaling steps Synthesis and release of signaling molecules (ligands) by the signaling cell. Transport of the signal to the target cell Detection of the
Cellular Signaling Pathways Signaling Overview Signaling steps Synthesis and release of signaling molecules (ligands) by the signaling cell. Transport of the signal to the target cell Detection of the
Imaging of glycolytic metabolism in primary glioblastoma cells with
 63 Chapter 5 Imaging of glycolytic metabolism in primary glioblastoma cells with RIMChip 5.1. Introduction Glioblastoma(GBM) is one of the most common brain tumors 1. It is composed of heterogeneous subpopulations
63 Chapter 5 Imaging of glycolytic metabolism in primary glioblastoma cells with RIMChip 5.1. Introduction Glioblastoma(GBM) is one of the most common brain tumors 1. It is composed of heterogeneous subpopulations
SUPPLEMENTARY INFORMATION
 In the format provided by the authors and unedited. 2 3 4 DOI: 10.1038/NMAT4893 EGFR and HER2 activate rigidity sensing only on rigid matrices Mayur Saxena 1,*, Shuaimin Liu 2,*, Bo Yang 3, Cynthia Hajal
In the format provided by the authors and unedited. 2 3 4 DOI: 10.1038/NMAT4893 EGFR and HER2 activate rigidity sensing only on rigid matrices Mayur Saxena 1,*, Shuaimin Liu 2,*, Bo Yang 3, Cynthia Hajal
Tyrosine kinases. Cell surface receptors ligand binding. Producer cell RNA. Target cell
 Tyrosine kinases http://msbl.helsinki.fi/tkseminar roducer cell Signaling molecules Receptor binding Signal transduction Target cell Activation of Gene expression RNA Biological responses proliferation,
Tyrosine kinases http://msbl.helsinki.fi/tkseminar roducer cell Signaling molecules Receptor binding Signal transduction Target cell Activation of Gene expression RNA Biological responses proliferation,
Receptors Functions and Signal Transduction- L4- L5
 Receptors Functions and Signal Transduction- L4- L5 Faisal I. Mohammed, MD, PhD University of Jordan 1 PKC Phosphorylates many substrates, can activate kinase pathway, gene regulation PLC- signaling pathway
Receptors Functions and Signal Transduction- L4- L5 Faisal I. Mohammed, MD, PhD University of Jordan 1 PKC Phosphorylates many substrates, can activate kinase pathway, gene regulation PLC- signaling pathway
Supplementary Figure 1. Normal T lymphocyte populations in Dapk -/- mice. (a) Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk
 Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk") Supplementary Figure 1. Normal T lymphocyte populations in Dapk -/- mice. (a) Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk -/- mice were stained for expression of CD4 and CD8.
Supplementary Figure 1. Normal T lymphocyte populations in Dapk -/- mice. (a) Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk -/- mice were stained for expression of CD4 and CD8.
TRAF6 ubiquitinates TGFβ type I receptor to promote its cleavage and nuclear translocation in cancer
 Supplementary Information TRAF6 ubiquitinates TGFβ type I receptor to promote its cleavage and nuclear translocation in cancer Yabing Mu, Reshma Sundar, Noopur Thakur, Maria Ekman, Shyam Kumar Gudey, Mariya
Supplementary Information TRAF6 ubiquitinates TGFβ type I receptor to promote its cleavage and nuclear translocation in cancer Yabing Mu, Reshma Sundar, Noopur Thakur, Maria Ekman, Shyam Kumar Gudey, Mariya
Negative Regulation of c-myc Oncogenic Activity Through the Tumor Suppressor PP2A-B56α
 Negative Regulation of c-myc Oncogenic Activity Through the Tumor Suppressor PP2A-B56α Mahnaz Janghorban, PhD Dr. Rosalie Sears lab 2/8/2015 Zanjan University Content 1. Background (keywords: c-myc, PP2A,
Negative Regulation of c-myc Oncogenic Activity Through the Tumor Suppressor PP2A-B56α Mahnaz Janghorban, PhD Dr. Rosalie Sears lab 2/8/2015 Zanjan University Content 1. Background (keywords: c-myc, PP2A,
Phospholipase C γ Prof. Graham Carpenter
 Graham Carpenter, h.d. rofessor of Biochemistry Cornelia Crooke Department of Biochemistry Vanderbilt University School of Medicine, Nashville, TN 1 Receptor Tyrosine Kinases GF Extracellular M Intracellular
Graham Carpenter, h.d. rofessor of Biochemistry Cornelia Crooke Department of Biochemistry Vanderbilt University School of Medicine, Nashville, TN 1 Receptor Tyrosine Kinases GF Extracellular M Intracellular
Nature Methods: doi: /nmeth Supplementary Figure 1
 Supplementary Figure 1 Subtiligase-catalyzed ligations with ubiquitin thioesters and 10-mer biotinylated peptides. (a) General scheme for ligations between ubiquitin thioesters and 10-mer, biotinylated
Supplementary Figure 1 Subtiligase-catalyzed ligations with ubiquitin thioesters and 10-mer biotinylated peptides. (a) General scheme for ligations between ubiquitin thioesters and 10-mer, biotinylated
Chromatin IP (Isw2) Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles.
 Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles.") Chromatin IP (Isw2) 7/01 Toshi last update: 06/15 Reagents Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles. 2.5 M glycine. TBS:
Chromatin IP (Isw2) 7/01 Toshi last update: 06/15 Reagents Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles. 2.5 M glycine. TBS:
Evaluation of the Small Molecular Weight G-Protein Ras in the Interaction of Growth Hormone and Epidermal Growth Factor Signaling
 Evaluation of the Small Molecular Weight G-Protein Ras in the Interaction of Growth Hormone and Epidermal Growth Factor Signaling Danielle Mollet TVI Community College & University of New Mexico Dr. Paul
Evaluation of the Small Molecular Weight G-Protein Ras in the Interaction of Growth Hormone and Epidermal Growth Factor Signaling Danielle Mollet TVI Community College & University of New Mexico Dr. Paul
Cancer. Questions about cancer. What is cancer? What causes unregulated cell growth? What regulates cell growth? What causes DNA damage?
 Questions about cancer What is cancer? Cancer Gil McVean, Department of Statistics, Oxford What causes unregulated cell growth? What regulates cell growth? What causes DNA damage? What are the steps in
Questions about cancer What is cancer? Cancer Gil McVean, Department of Statistics, Oxford What causes unregulated cell growth? What regulates cell growth? What causes DNA damage? What are the steps in
hexahistidine tagged GRP78 devoid of the KDEL motif (GRP78-His) on SDS-PAGE. This
 on SDS-PAGE. This") SUPPLEMENTAL FIGURE LEGEND Fig. S1. Generation and characterization of. (A) Coomassie staining of soluble hexahistidine tagged GRP78 devoid of the KDEL motif (GRP78-His) on SDS-PAGE. This protein was expressed
SUPPLEMENTAL FIGURE LEGEND Fig. S1. Generation and characterization of. (A) Coomassie staining of soluble hexahistidine tagged GRP78 devoid of the KDEL motif (GRP78-His) on SDS-PAGE. This protein was expressed
Signal Transduction: Information Metabolism. Chem 454: Regulatory Mechanisms in Biochemistry University of Wisconsin-Eau Claire
 Signal Transduction: Information Metabolism Chem 454: Regulatory Mechanisms in Biochemistry University of Wisconsin-Eau Claire Introduction Information Metabolism How cells receive, process and respond
Signal Transduction: Information Metabolism Chem 454: Regulatory Mechanisms in Biochemistry University of Wisconsin-Eau Claire Introduction Information Metabolism How cells receive, process and respond
Chapter 20. Cell - Cell Signaling: Hormones and Receptors. Three general types of extracellular signaling. endocrine signaling. paracrine signaling
 Chapter 20 Cell - Cell Signaling: Hormones and Receptors Three general types of extracellular signaling endocrine signaling paracrine signaling autocrine signaling Endocrine Signaling - signaling molecules
Chapter 20 Cell - Cell Signaling: Hormones and Receptors Three general types of extracellular signaling endocrine signaling paracrine signaling autocrine signaling Endocrine Signaling - signaling molecules
supplementary information
 DOI: 10.1038/ncb1875 Figure S1 (a) The 79 surgical specimens from NSCLC patients were analysed by immunohistochemistry with an anti-p53 antibody and control serum (data not shown). The normal bronchi served
DOI: 10.1038/ncb1875 Figure S1 (a) The 79 surgical specimens from NSCLC patients were analysed by immunohistochemistry with an anti-p53 antibody and control serum (data not shown). The normal bronchi served
number Done by Corrected by Doctor Maha Shomaf
 number 19 Done by Waseem Abo-Obeida Corrected by Abdullah Zreiqat Doctor Maha Shomaf Carcinogenesis: the molecular basis of cancer. Non-lethal genetic damage lies at the heart of carcinogenesis and leads
number 19 Done by Waseem Abo-Obeida Corrected by Abdullah Zreiqat Doctor Maha Shomaf Carcinogenesis: the molecular basis of cancer. Non-lethal genetic damage lies at the heart of carcinogenesis and leads
The Tissue Engineer s Toolkit
 The Tissue Engineer s Toolkit Stimuli Detection and Response Ken Webb, Ph. D. Assistant Professor Dept. of Bioengineering Clemson University Environmental Stimulus-Cellular Response Environmental Stimuli
The Tissue Engineer s Toolkit Stimuli Detection and Response Ken Webb, Ph. D. Assistant Professor Dept. of Bioengineering Clemson University Environmental Stimulus-Cellular Response Environmental Stimuli
Signal Transduction I
 Signal Transduction I Prof. Tianhua Zhou Department of Cell Biology Zhejiang University School of Medicine Office hours by appointment tzhou@zju.edu.cn Signal transduction: Key contents for signal transduction:
Signal Transduction I Prof. Tianhua Zhou Department of Cell Biology Zhejiang University School of Medicine Office hours by appointment tzhou@zju.edu.cn Signal transduction: Key contents for signal transduction:
Introduction. Cancer Biology. Tumor-suppressor genes. Proto-oncogenes. DNA stability genes. Mechanisms of carcinogenesis.
 Cancer Biology Chapter 18 Eric J. Hall., Amato Giaccia, Radiobiology for the Radiologist Introduction Tissue homeostasis depends on the regulated cell division and self-elimination (programmed cell death)
Cancer Biology Chapter 18 Eric J. Hall., Amato Giaccia, Radiobiology for the Radiologist Introduction Tissue homeostasis depends on the regulated cell division and self-elimination (programmed cell death)
609G: Concepts of Cancer Genetics and Treatments (3 credits)
") Master of Chemical and Life Sciences Program College of Computer, Mathematical, and Natural Sciences 609G: Concepts of Cancer Genetics and Treatments (3 credits) Text books: Principles of Cancer Genetics,
Master of Chemical and Life Sciences Program College of Computer, Mathematical, and Natural Sciences 609G: Concepts of Cancer Genetics and Treatments (3 credits) Text books: Principles of Cancer Genetics,
Cell Cell Communication
 IBS 8102 Cell, Molecular, and Developmental Biology Cell Cell Communication January 29, 2008 Communicate What? Why do cells communicate? To govern or modify each other for the benefit of the organism differentiate
IBS 8102 Cell, Molecular, and Developmental Biology Cell Cell Communication January 29, 2008 Communicate What? Why do cells communicate? To govern or modify each other for the benefit of the organism differentiate
Supplementary Information POLO-LIKE KINASE 1 FACILITATES LOSS OF PTEN-INDUCED PROSTATE CANCER FORMATION
 Supplementary Information POLO-LIKE KINASE 1 FACILITATES LOSS OF PTEN-INDUCED PROSTATE CANCER FORMATION X. Shawn Liu 1, 3, Bing Song 2, 3, Bennett D. Elzey 3, 4, Timothy L. Ratliff 3, 4, Stephen F. Konieczny
Supplementary Information POLO-LIKE KINASE 1 FACILITATES LOSS OF PTEN-INDUCED PROSTATE CANCER FORMATION X. Shawn Liu 1, 3, Bing Song 2, 3, Bennett D. Elzey 3, 4, Timothy L. Ratliff 3, 4, Stephen F. Konieczny
Computational Systems Biology: Biology X
 Bud Mishra Room 1002, 715 Broadway, Courant Institute, NYU, New York, USA L#10:(November-22-2010) Cancer and Signals 1 1 Micro-Environment Story How does the rest of our body influences the cancer cell?
Bud Mishra Room 1002, 715 Broadway, Courant Institute, NYU, New York, USA L#10:(November-22-2010) Cancer and Signals 1 1 Micro-Environment Story How does the rest of our body influences the cancer cell?
Genetics and Cancer Ch 20
 Genetics and Cancer Ch 20 Cancer is genetic Hereditary cancers Predisposition genes Ex. some forms of colon cancer Sporadic cancers ~90% of cancers Descendants of cancerous cells all cancerous (clonal)
Genetics and Cancer Ch 20 Cancer is genetic Hereditary cancers Predisposition genes Ex. some forms of colon cancer Sporadic cancers ~90% of cancers Descendants of cancerous cells all cancerous (clonal)
A RENAISSANCE FOR SRC
 A RENAISSANCE FOR SRC Timothy J. Yeatman The c-src non-receptor tyrosine kinase is overexpressed and activated in a large number of human malignancies and has been linked to the development of cancer and
A RENAISSANCE FOR SRC Timothy J. Yeatman The c-src non-receptor tyrosine kinase is overexpressed and activated in a large number of human malignancies and has been linked to the development of cancer and
Interleukin-6; pathogenesis and treatment of autoimmune inflammatory diseases
 54 Review Article Interleukin-6; pathogenesis and treatment of autoimmune inflammatory diseases Toshio Tanaka 1, 2), Masashi Narazaki 3), Kazuya Masuda 4) and Tadamitsu Kishimoto 4, ) 1) Department of
54 Review Article Interleukin-6; pathogenesis and treatment of autoimmune inflammatory diseases Toshio Tanaka 1, 2), Masashi Narazaki 3), Kazuya Masuda 4) and Tadamitsu Kishimoto 4, ) 1) Department of
Western Immunoblotting Preparation of Samples:
 Western Immunoblotting Preparation of Samples: Total Protein Extraction from Culture Cells: Take off the medium Wash culture with 1 x PBS 1 ml hot Cell-lysis Solution into T75 flask Scrap out the cells
Western Immunoblotting Preparation of Samples: Total Protein Extraction from Culture Cells: Take off the medium Wash culture with 1 x PBS 1 ml hot Cell-lysis Solution into T75 flask Scrap out the cells
PI3K Background. The SignalRx R & D pipeline is shown below followed by a brief description of each program:
 PI3K Background The phosphatidylinositol 3-kinase (PI3K) pathway is a key cell signaling node whose dysregulation commonly results in the transformation of normal cells into cancer cells. The role of PI3K
PI3K Background The phosphatidylinositol 3-kinase (PI3K) pathway is a key cell signaling node whose dysregulation commonly results in the transformation of normal cells into cancer cells. The role of PI3K
Protein tyrosine kinase signaling
 rotein tyrosine kinase signaling Serge ROCHE CRBM CNRS/Montpellier University serge.roche@crbm.cnrs.fr rotein phosphorylation on Tyr A central mechanism to control cell communication in a multicellular
rotein tyrosine kinase signaling Serge ROCHE CRBM CNRS/Montpellier University serge.roche@crbm.cnrs.fr rotein phosphorylation on Tyr A central mechanism to control cell communication in a multicellular
A particular set of insults induces apoptosis (part 1), which, if inhibited, can switch to autophagy. At least in some cellular settings, autophagy se
, which, if inhibited, can switch to autophagy. At least in some cellular settings, autophagy se") A particular set of insults induces apoptosis (part 1), which, if inhibited, can switch to autophagy. At least in some cellular settings, autophagy serves as a defence mechanism that prevents or retards
A particular set of insults induces apoptosis (part 1), which, if inhibited, can switch to autophagy. At least in some cellular settings, autophagy serves as a defence mechanism that prevents or retards
SUPPLEMENTARY INFORMATION
 Supplementary Table 1. Cell sphingolipids and S1P bound to endogenous TRAF2. Sphingolipid Cell pmol/mg TRAF2 immunoprecipitate pmol/mg Sphingomyelin 4200 ± 250 Not detected Monohexosylceramide 311 ± 18
Supplementary Table 1. Cell sphingolipids and S1P bound to endogenous TRAF2. Sphingolipid Cell pmol/mg TRAF2 immunoprecipitate pmol/mg Sphingomyelin 4200 ± 250 Not detected Monohexosylceramide 311 ± 18
Functional Limitations
 Regulation of the Cell Cycle Chapter 12 Pg. 228 245 Functional Limitations Various factors determine whether and when a cell divides. Two functional limitations for cell size limit growth or influence
Regulation of the Cell Cycle Chapter 12 Pg. 228 245 Functional Limitations Various factors determine whether and when a cell divides. Two functional limitations for cell size limit growth or influence
RayBio Human PPAR-alpha Transcription Factor Activity Assay Kit
 RayBio Human PPAR-alpha Transcription Factor Activity Assay Kit Catalog #: TFEH-PPARa User Manual Jan 5, 2018 3607 Parkway Lane, Suite 200 Norcross, GA 30092 Tel: 1-888-494-8555 (Toll Free) or 770-729-2992,
RayBio Human PPAR-alpha Transcription Factor Activity Assay Kit Catalog #: TFEH-PPARa User Manual Jan 5, 2018 3607 Parkway Lane, Suite 200 Norcross, GA 30092 Tel: 1-888-494-8555 (Toll Free) or 770-729-2992,
Supplementary Information Supplementary Fig. 1. Elevated Usp9x in melanoma and NRAS mutant melanoma cells are dependent on NRAS for 3D growth.
 Supplementary Information Supplementary Fig. 1. Elevated Usp9x in melanoma and NRAS mutant melanoma cells are dependent on NRAS for 3D growth. a. Immunoblot for Usp9x protein in NRAS mutant melanoma cells
Supplementary Information Supplementary Fig. 1. Elevated Usp9x in melanoma and NRAS mutant melanoma cells are dependent on NRAS for 3D growth. a. Immunoblot for Usp9x protein in NRAS mutant melanoma cells
Problem Set 8 Key 1 of 8
 7.06 2003 Problem Set 8 Key 1 of 8 7.06 2003 Problem Set 8 Key 1. As a bright MD/PhD, you are interested in questions about the control of cell number in the body. Recently, you've seen three patients
7.06 2003 Problem Set 8 Key 1 of 8 7.06 2003 Problem Set 8 Key 1. As a bright MD/PhD, you are interested in questions about the control of cell number in the body. Recently, you've seen three patients
Cellular Physiology (PHSI3009) Contents:
 Contents:") Cellular Physiology (PHSI3009) Contents: Cell membranes and communication 2 nd messenger systems G-coupled protein signalling Calcium signalling Small G-protein signalling o RAS o MAPK o PI3K RHO GTPases
Cellular Physiology (PHSI3009) Contents: Cell membranes and communication 2 nd messenger systems G-coupled protein signalling Calcium signalling Small G-protein signalling o RAS o MAPK o PI3K RHO GTPases
SUPPLEMENTAL MATERIALS AND METHODS. Puromycin-synchronized metabolic labelling - Transfected HepG2 cells were depleted of
 SUPPLEMENTAL MATERIALS AND METHODS Puromycin-synchronized metabolic labelling - Transfected HepG2 cells were depleted of cysteine and methionine and then treated with 10 μm puromycin in depletion medium
SUPPLEMENTAL MATERIALS AND METHODS Puromycin-synchronized metabolic labelling - Transfected HepG2 cells were depleted of cysteine and methionine and then treated with 10 μm puromycin in depletion medium
Human Lung Cancer Pathology and Cellular Biology Mouse Lung Tumor Workshop
 Human Lung Cancer Pathology and Cellular Biology Mouse Lung Tumor Workshop Jan 7 th and 8 th, 2014 Brigitte Gomperts, MD University of California, Los Angeles Lung Structure and Function Airway Epithelial
Human Lung Cancer Pathology and Cellular Biology Mouse Lung Tumor Workshop Jan 7 th and 8 th, 2014 Brigitte Gomperts, MD University of California, Los Angeles Lung Structure and Function Airway Epithelial
Deregulation of signal transduction and cell cycle in Cancer
 Deregulation of signal transduction and cell cycle in Cancer Tuangporn Suthiphongchai, Ph.D. Department of Biochemistry Faculty of Science, Mahidol University Email: tuangporn.sut@mahidol.ac.th Room Pr324
Deregulation of signal transduction and cell cycle in Cancer Tuangporn Suthiphongchai, Ph.D. Department of Biochemistry Faculty of Science, Mahidol University Email: tuangporn.sut@mahidol.ac.th Room Pr324
The Immunoassay Guide to Successful Mass Spectrometry. Orr Sharpe Robinson Lab SUMS User Meeting October 29, 2013
 The Immunoassay Guide to Successful Mass Spectrometry Orr Sharpe Robinson Lab SUMS User Meeting October 29, 2013 What is it? Hey! Look at that! Something is reacting in here! I just wish I knew what it
The Immunoassay Guide to Successful Mass Spectrometry Orr Sharpe Robinson Lab SUMS User Meeting October 29, 2013 What is it? Hey! Look at that! Something is reacting in here! I just wish I knew what it
Genetics of Cancer Lecture 32 Cancer II. Prof. Bevin Engelward, MIT Biological Engineering Department
 Genetics of Cancer Lecture 32 Cancer II rof. Bevin Engelward, MIT Biological Engineering Department Why Cancer Matters New Cancer Cases in 1997 Cancer Deaths in 1997 Genetics of Cancer: Today: What types
Genetics of Cancer Lecture 32 Cancer II rof. Bevin Engelward, MIT Biological Engineering Department Why Cancer Matters New Cancer Cases in 1997 Cancer Deaths in 1997 Genetics of Cancer: Today: What types
Targeting the ERBB family in cancer: couples therapy
 OPINION Targeting the ERBB family in cancer: couples therapy Niall Tebbutt, Mikkel W. Pedersen and Terrance G. Johns Abstract The ERBB family of receptor tyrosine kinases has a central role in the tumorigenesis
OPINION Targeting the ERBB family in cancer: couples therapy Niall Tebbutt, Mikkel W. Pedersen and Terrance G. Johns Abstract The ERBB family of receptor tyrosine kinases has a central role in the tumorigenesis