Wait Anaphase Signals Are Not Confined to the Mitotic Spindle
|
|
|
- Kelley Manning
- 6 years ago
- Views:
Transcription
1 Wait Anaphase Signals Are Not Confined to the Mitotic Spindle Lydia Reńe Heasley 1*, Steven M. Markus 1*, and Jennifer G. DeLuca 1* 1 Colorado State University, Fort Collins, CO *Corresponding authors 28,728 characters Abstract The Spindle Assembly Checkpoint ensures the faithful inheritance of chromosomes by arresting mitotic progression in the presence of kinetochores that are not attached to spindle microtubules. This is achieved through inhibition of the Anaphase Promoting Complex/Cyclosome by a kinetochore-derived wait anaphase signal known as the Mitotic Checkpoint Complex. It remains unclear whether the localization and activity of these inhibitory complexes are restricted to the mitotic spindle compartment or are diffusible throughout the cytoplasm. Here we report that wait anaphase signals are indeed able to diffuse outside the confines of the mitotic spindle compartment. Using a cell fusion approach to generate multinucleate cells, we investigate the effects of checkpoint signals derived from one spindle compartment on a neighboring spindle compartment. We find that spindle compartments in close proximity wait for one another to align all chromosomes before entering anaphase synchronously. Synchrony is disrupted in cells with increased inter-spindle distances and cellular constrictions between spindle compartments. In addition, when mitotic cells are fused with interphase cells, wait anaphase signals are diluted, resulting in premature mitotic exit. Overall, our studies reveal that anaphase inhibitors are diffusible and active outside the confines of the mitotic spindle from which they are derived.
2 Introduction Accurate chromosome inheritance during cell division is necessary for the development and maintenance of all organisms. Failure to properly segregate genetic material results in the generation of aneuploid cells (cells with too few or too many chromosomes), an event associated with disease states such as infertility and cancer 1. Thus, cells have developed an elegant surveillance system called the Spindle Assembly Checkpoint (SAC), which suspends the initiation of anaphase the separation of chromatids toward opposite cell poles until all the chromosomes are positioned to be equally inherited 2. The SAC monitors attachment of spindle microtubules (MTs) to large protein complexes called kinetochores (KTs), which reside at the centromere of each chromosome 3. Unbound KTs generate a molecular signal that ultimately manifests in the cell-wide inhibition of anaphase onset 4. This wait anaphase signal, which is comprised of a multimeric protein complex called the mitotic checkpoint (MCC), is generated when Mad1 induces a conformational change in Mad2 from an open to a closed state 5-8. It is upon the scaffold of a closed Mad2 molecule that the MCC which consists of Mad2, Bub3, BubR1, and Cdc20 is assembled 9,10. The MCC delays anaphase onset by inhibiting the activity of the anaphase promoting complex/cyclosome (APC/C) the aptly named E3-ubiquitin ligase whose action is required for progression into anaphase through sequestration of the APC/C activating subunit Cdc Inhibition of the APC/C prevents the targeting and subsequent degradation of key mitosis-promoting factors including Cyclin B and Securin 2. This checkpoint is exceptionally robust. Studies have shown that the inhibitory wait anaphase signals generated by even a single unbound KT can arrest a cell in mitosis 4,12. Nearly three decades of intensive research have led to the characterization of the molecular mechanisms by which the SAC senses and responds to chromosome attachment defects. However, we still do not fully understand how inhibitory MCC complexes generated from a single KT can influence the activities of the entire cell. This mechanism no doubt relies on the diffusible properties of KT-derived inhibitory signals.
3 In 1997, Rieder et al. observed mitosis in fused mammalian cells possessing two spindle compartments, defined as an assembled spindle and the associated molecular components that originate from a single nucleus 13. They made two key observations: 1) the unattached KTs in one spindle compartment did not cause a mitotic checkpoint arrest in the neighboring spindle compartment, and 2) when one spindle compartment initiated anaphase, the neighboring spindle compartment also initiated anaphase regardless of the alignment status of its own chromosomes. These observations prompted the authors to conclude that the activity and diffusibility of wait anaphase signals (i.e., active MCC complexes) were restricted to the spindle from which they were generated, but go anaphase signals were global and dominant. The notion that MCC molecules are spindle restricted has remained a dominant model in the field. Evidence supporting the MCC restriction model includes the discovery of the spindle matrix: a proteinaceous fusiform structure that embodies the mitotic spindle Work using Drosophila and human cells has shown that Mad1 and Mad2 remain enriched within this structure, suggesting that indeed, components of the SAC and MCC may be restricted in their diffusion away from the spindle compartment Recent characterization of the molecular mechanisms underlying SAC signaling has raised the possibility that MCC complexes may not be confined to the spindle compartment, and may in fact diffuse throughout the cytoplasm of a mitotic cell 7,8, For example, an important prediction of the spindle restriction model is that the target of the MCC the APC/C should also be restricted to the spindle compartment such that it can be sufficiently inhibited by the MCC to prevent anaphase onset. If the APC/C CDC20 pool was not spindle restricted but MCC complexes were, then the non-spindle restricted pool of APC/C CDC20 would be active, and would thus promote anaphase onset irrespective of KT-MT attachment status. However, in mitotic cells, subcellular pools of the APC/C are found both within the spindle compartment (e.g., spindle poles, kinetochores, chromosomes), and throughout the cytoplasm, suggesting that there may be a cytoplasmic inhibitor of the APC/C
4 In this article, we present findings that demonstrate that wait anaphase signals are not restricted to the mitotic spindle compartment from which they originate, and are in fact diffusible. Similar to the cell fusion experiments conducted by Rieder et. al., we have fused mitotic cells and examined the behavior of spindle compartments that share a common cytoplasm. We set out to test three predictions of how wait anaphase signals should behave if they are restricted to the confines of the mitotic spindle. First, spindle compartment-restricted wait anaphase signals should be unable to influence the behavior and mitotic progression of other spindles within a shared cytoplasm. Second, spindle compartment-restricted wait anaphase signals should be insensitive to cellular diffusion barriers. Finally, inhibitory activities of spindle compartment-restricted wait anaphase signals should not be affected by cytoplasmic dilution. In contrast to earlier results, we find that mitotic spindles within close proximity wait for one another to align their chromosomes before initiating anaphase in synchrony. In contrast, spindles that remain far apart or are separated by a cellular constriction do not undergo synchronous anaphase. We also find that when mitotic cells are fused with interphase cells, pre-existing mitotic spindle compartments exit mitosis prematurely, suggesting that the wait anaphase signals become diluted by non-mitotic cytoplasm. These observations support a model in which KT-derived wait anaphase signals can diffuse away from the source spindle compartment and into the cytoplasm to levels that are sufficient to prevent anaphase onset. Our findings provide new insight into the molecular mechanisms governing the activity of the Spindle Assembly Checkpoint. Results Synchronized and Fused PtK1 Cells Exhibit Normal Mitotic Timing To enrich for mitotic PtK1 cells for use in our fusion experiments, we treated cells with the CDK1 inhibitor RO3306 to arrest cells at the G2/M boundary 30. We found that ~30% of cells entered mitosis within one hour following washout from a 20-hour treatment with RO3306 (Figure 1A). We compared the timing of mitotic progression in asynchronous cells, synchronized cells, and synchronized cells exposed to fusion media (but those that remained unfused) and found that treatment with RO3306 or
5 fusion media did not significantly impact the duration of prometaphase (defined as the time from nuclear envelope breakdown NEBD to chromosome alignment) or metaphase (defined as the time from chromosome alignment to chromatid separation; Figure 1B and 1C). Using partially synchronized populations of PtK1 cells, we generated fused cells possessing multiple spindle compartments within a single shared cytoplasm (see Materials and Methods). Spindles Wait for One Another to Initiate Anaphase From their study of fused mitotic cells, Reider et al. concluded that wait anaphase signals were restricted to the spindle compartment from which they were generated 13. They based this conclusion on the observation that in bi-spindled cells, unbound kinetochores in one spindle compartment did not delay anaphase onset in the neighboring compartment. However, the observed behaviors could also be explained if: a) the inhibitory MCC complexes were diffusible, but b) the spindle compartments were too far away from one another for the MCC complexes generated from one compartment to impose a mitotic arrest on the neighboring compartment. If this were the case, we would predict that spindle compartments in sufficiently close proximity would exchange/share wait anaphase signals, and only when the SAC of both compartments were satisfied would synchronous anaphase ensue (Figure 2A, left). To determine whether spindle compartments share wait anaphase signals, we imaged fused mitotic cells over time and scored the frequency of synchronous anaphase onset. Strikingly, time-lapse imaging revealed numerous fused bi-spindled cells in which the first spindle to reach metaphase (the early spindle) appeared to delay anaphase onset until the other spindle (the late spindle) aligned all of its chromosomes. Shortly after this occurred, both spindle compartments synchronously entered anaphase (Figure 2B). In these cases, the early spindle delayed anaphase onset for up to three times longer (maximum = 30 min, average = 21.2 min) than the mean duration of metaphase in non-fused cells (average = 9.4 min; Fig. 1B and 2C, Single Spindle ). Interestingly, once the late spindle completed aligning its chromosomes, it progressed into anaphase more quickly than control spindles (5.7 min versus
6 9.4 min; p < 0.001; Figure 2C). This supports the notion that cells become primed to exit mitosis as they accumulate KT attachments to spindle microtubules. Previous studies indicate that this priming is due to a combination of: a) decreased formation of new inhibitory MCC complexes, and b) active disassembly of existing MCC complexes 12,19. It should be noted that synchronous anaphase between two spindles was observed in the seminal study using fused cells to study checkpoint activity conducted by Rieder and colleagues 13. However, the authors did not include these cells in their analysis due to the high variability of metaphase duration that they had calculated previously for PtK1 cells. Specifically, they reported that the time spent in metaphase varied from 9-49 min 13. Because of this high variability in metaphase duration, it could not be determined if one spindle was delayed in anaphase onset due to its neighbor spindle, or alternatively, if the mitotic timing of the delayed spindle fell into the reported normal range. In our own studies, we found that the duration of metaphase in PtK1 cells was shorter (9.4 min) and that the range was narrower (from 3-18 min) than reported in previous studies. Thus, we are confident that the behavior resulting in synchronous anaphase is not due to variability of mitotic timing, but rather cell-wide inhibition of anaphase onset imparted by the spindle containing unbound KTs. In addition, the authors of the original paper included fused cells possessing monopolar or multipolar spindles in their analysis 13. Due to the complex signaling and mitotic timing that occurs in such abnormal spindles, we excluded cells containing multipolar or monopolar spindles from our analysis. Asynchronous Anaphase Occurs When Wait Anaphase Signals Are Subject To Diffusion Barriers To obtain a larger sample size than was feasible with our live-cell experiments, we implemented a fixedcell protocol to analyze mitotic cell fusion events. For these experiments, we first fused synchronized PtK1 cells cultured on coverslips as described above. Following exposure to fusion media, the cells were returned to normal growth conditions for one hour to allow for progression into mitosis, and then subjected to fixation and immunostaining. We determined the stage of mitotic progression in each spindle
7 as follows: 1) using DIC microscopy, we assessed whether sister chromatids were attached or separated, as anaphase chromatids become separated from one another and move towards opposite spindle poles; 2) we used immunofluorescence to visualize the mitotic spindle, as anaphase spindles contain short KT fibers near the poles and long interpolar MTs 31 ; 3) finally, we used immunofluorescence to detect the SAC protein Mad1, which localizes to unattached, but not attached KTs (see Figure 3B, merged insets) 32. As above, we predicted that if wait anaphase signals were diffusible, then one spindle with an unattached kinetochore should generate sufficient signal to keep cytoplasmic (i.e., diffusive) MCC levels above the threshold needed to keep both spindles in mitosis (Figure 3Ai, top). Alternatively, if wait anaphase signals were spindle-restricted, then a shared cytoplasmic volume would have no effect on anaphase onset of each spindle (Figure 3Ai, bottom). Using our fixed cell approach, we observed many cells (44/55) in which two mitotic spindles appeared to be undergoing synchronous anaphase (Figure 3B, left). Interestingly, and in stark contrast, we also observed cells in which only one of the two spindles was undergoing anaphase, indicating asynchrony of anaphase onset (11/55; Figure 3B, right). We noticed that the spindles within these asynchronous anaphase cells were at greater distances from one another than the spindles in cells that underwent synchronous anaphase (Figure 3B; compare Synchronous to Asynchronous ). Additionally, cells that underwent asynchronous anaphase typically formed a cytoplasmic constriction point between the two spindles (Figure 3B, right, and 3D). If wait anaphase signals are confined to the spindle compartment, these signals should be insensitive to diffusion barriers between two spindle compartments within a shared cytoplasm (Figure 3Aii and iii, bottom). However, if these signals are indeed diffusible in the cytoplasm, then both inter-spindle distance and degree of cytoplasmic constriction would be expected to alter the extent over which the inhibitory complexes can act (Figure 3Aii and iii, top). We measured the inter-spindle distance in fused cells and correlated this distance to the anaphase behavior in each cell (synchronous vs. asynchronous). We found that synchronous anaphase spindles were typically in close proximity to one other within the fused cell. Specifically, we found that spindle compartments <~30 µm apart underwent synchronous anaphase,
8 whereas spindles >~40 µm apart underwent asynchronous anaphase (Figure 3C). Interestingly, we found that asynchronous cells possessed narrower cytoplasmic constrictions than synchronous cells (Figure 3B and E). We quantified the extent of this constriction for each cell by calculating their constriction coefficient, which was defined as the ratio between the narrowest and widest points for a given fused cell (see Figure 3A; w, widest point; c, constriction). Our live-cell studies revealed that these constrictions develop as mitosis progresses (see Figure 3D). In the first frame of Figure 3D (time = 0 ), there is a modest degree of constriction (constriction coefficient at t 0 = 0.36), and both nuclei are in prophase. As mitosis progresses, the constriction becomes more pronounced (constriction coefficient at t 50 = 0.15) and the spindle compartments undergo asynchronous anaphase (left spindle at 50 ; right spindle at 70 ; see A ). We interpret the asynchronous anaphase onset in such cells to be a consequence of the presence of a barrier to diffusion of wait anaphase signals between the two spindles. As further evidence that wait anaphase signals are subject to diffusion barriers, we observed both synchrony and asynchrony of anaphase in a single fused trinucleate cell (Figure 4). Two of the nuclei remained within 13 µm of one another while the third nucleus was situated approximately 68 µm away from the other two. All three nuclei entered mitosis at the same time, as evidenced by the synchrony of nuclear envelope breakdown (NEBD; at 20 ). This was expected because it is known that Cyclin B/CDK1 is a powerful inducer of mitosis, even in large syncytia containing multiple nuclei Following NEBD, all three nuclei formed distinct mitotic spindles. As was commonly seen throughout these experiments, a cellular constriction formed between the two closely apposed spindles and the third distant spindle. Sixtyfive minutes following NEBD (at 85 ), the closely apposed spindles underwent synchronous anaphase. However, the distant spindle, now separated by a cytoplasmic constriction did not undergo anaphase until 140, approximately 120 min after NEBD, and 55 after the other spindles. These observations are consistent with our observations in bi-spindled cells, and lend additional support to the notion that wait anaphase signals are indeed diffusible. Thus, factors that can influence the extent of SAC protein
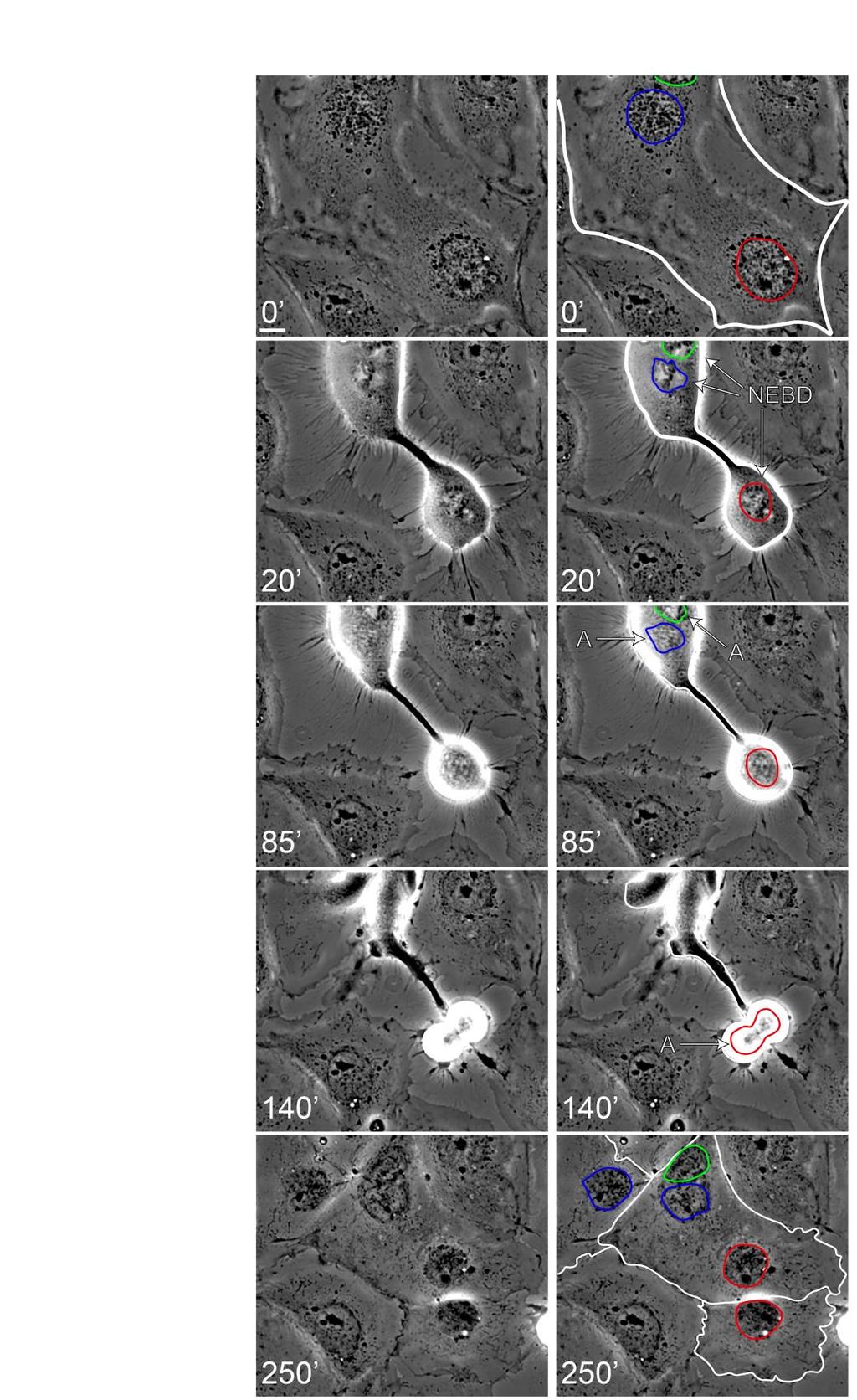
9 diffusion, including inter-spindle distance, a diffusion barrier such as a cytoplasmic constriction, or a combination of both, can provide spindles with autonomy with respect to mitotic progression. Dilution of Wait Anaphase Complexes Affects the Strength of SAC Signaling Whereas spindle compartment-restricted wait anaphase complexes should be unaffected by changes in cytoplasmic volume, diffusible wait anaphase signals should be sensitive to cytoplasmic dilution. We predicted that if cytoplasmic wait anaphase complexes were diluted by additional non-mitotic cytoplasm, the SAC might become compromised, causing spindle compartments to prematurely exit mitosis 36,37 (Figure 5A). To test this prediction, we exposed an asynchronous population of PtK1 cells to fusion media in order to observe fusion events between mitotic and interphase cells. Indeed, the majority of these fusion events resulted in premature mitotic exit (Figure 5B and C, and data not shown). We observed two types of aberrant mitotic exit resulting from fusion of a mitotic cell with a non-mitotic cell. The first and most common type of premature mitotic exit consisted of sister chromatid separation and chromosome decondensation without chromosome alignment (Figure 5B). Figure 5B shows an example of this type of mitotic exit occurring in a fused cell containing three nuclei, each in different phases of the cell cycle: the top spindle compartment is in prometaphase ( PM ), the bottom left spindle compartment is in late anaphase ( A ), and the bottom right nucleus is in interphase ( I ). Shortly following the fusion event, the prometaphase spindle compartment can be observed exiting mitosis without aligning its chromosomes or initiating anaphase (see Exit at 30 ). The other, less commonly observed example of premature mitotic exit consisted of anaphase onset in the presence of unaligned chromosomes (Figure 5C). In this example, which shows a fused cell containing a prometaphase nucleus ( PM, top) and two interphase nuclei ( I, bottom), the prophase nucleus proceeds to form a bipolar spindle ( BPS at 65 ; unlike the example shown in Fig. 5B). However, anaphase onset ( A at 70 ) occurs in spite of the presence of unaligned chromosomes (see arrowheads at 70 ). Interestingly, but not surprisingly, the two interphase nuclei undergo premature chromosome condensation ( PCC at 65 ; a sign of mitotic entry), similar to our observations described above (see Figure 4, and associated text). We interpret the
10 premature mitotic exit observed in these fused cells to be indicative of perturbed SAC activity, and attribute this dysfunction to the dilution of cytoplasmic wait anaphase signals. Although our data suggest a model whereby mitotic exit results from dilution of active wait anaphase complexes, we cannot formally rule out the possibility that cytoplasmic factors exist in the interphase cells that could potentially promote the disassembly of MCC complexes, and thereby promote premature mitotic exit. Notably, when we performed the same experiment with RO3306 synchronized cells (i.e., arrested at G2/M), mitotic spindle compartments waited at metaphase for the interphase nuclei to enter mitosis and align their chromosomes prior to a synchronous anaphase onset (Figure 5D). This is not unexpected, since RO3306 leads to the accumulation of cells in late G2, a point at which cells are competent to enter mitosis and generate active MCC complexes 10, Discussion In this study we employed cell fusion experiments to test the hypothesis that wait anaphase signals are restricted to the spindle compartment within which they are generated. This hypothesis, which was originally proposed by Reider et al. in 1997, has remained a prominent theory in the field of SAC signaling and mitosis. However, as we have gained a more detailed understanding of SAC signaling and the molecules involved, it is difficult to conceive of a mechanism by which SAC components would be restricted to the spindle region, and thus excluded from the cytoplasm outside of the spindle. We therefore directly tested the hypothesis using predictions of how fused, multi-spindled cells would behave during mitosis if the wait anaphase signals were spindle restricted, or diffusible. If such signals are indeed spindle-restricted, we predicted that: a) spindle compartments in a shared cytoplasm would progress through mitosis autonomously, b) spindle compartments in a shared cytoplasm would be unaffected by diffusion barriers, and c) mitotic progression would be unaffected by the introduction of additional cytoplasmic volume. For each of these predictions, we observed cellular behaviors that instead indicate that wait anaphase signals are in fact diffusible outside the confines of the spindle compartment.
11 Specifically, we found that: a) spindle compartments within a shared cytoplasm exhibit synchronized mitotic progression (i.e., undergo simultaneous anaphase onset), b) anaphase synchrony is disrupted in the presence of a diffusion barrier such as a cytoplasmic constriction, and c) spindle compartments prematurely exit mitosis when exposed to non-mitotic cytoplasm via cell fusion with an interphase cell. These findings provide new insight into the molecular biology of the SAC with specific attention to where in the cell wait anaphase signals are active. These results have important implications for our understanding of mitotic checkpoint dysfunction during embryonic cell division. It has been found that the embryonic cells of most animal species lack a robust SAC until embryos reach the mid-blastula transition 37,41,42. As a result, chromosome segregation errors occur at a higher frequency early in embryogenesis due to premature mitotic exit 43. The predominant hypothesis to explain deficient checkpoint function in these cells is that the large cytoplasmic volume of embryonic cells might dilute the inhibitory wait anaphase signal, thus reducing the strength of the SAC. Thus, a defined kinetochore:cytoplasm ratio is required to ensure appropriately-timed and error-free mitoses. This model has gained support from a recent study by Galli et al., in which the authors measured checkpoint strength in the embryos of the nematode C. elegans 37. As is the case in many animals, the early embryonic cells of C. elegans divide such that the cellular volume is reduced at each round of division (i.e., they undergo reductive divisions). The resulting cells are of equal ploidy (and thus contain equal numbers of kinetochores) but are smaller in volume. Galli et al. found that as cell volume decreased, the ability of cells to maintain a nocodazole-induced mitotic arrest improved 37. A critical assumption of this kinetochore:cytoplasm hypothesis is that inhibitory wait anaphase signals can diffuse outside of the confines of the spindle and into the cytoplasm. It is unclear how this signal could become diluted if it was restricted to the spindle compartment. Our studies offer clarification as to how dilution of wait anaphase signals could occur.
12 Within the human body, there are very few cell types that maintain a multinucleate state (e.g., skeletal and placental tissue). However, our findings that wait anaphase signals can be communicated through the cytoplasm between two spindle compartments have direct application to the behavior of the SAC in mononucleate cells. For example, during mitosis, it is not uncommon for chromosomes to become isolated outside the confines of the spindle compartment. These chromosomes, whether they are stuck behind the spindle poles or simply distal from the spindle compartment are known to be competent to: a) recruit SAC components such as Mad1, and b) cause a delay in mitotic progression 12. Our results here reveal the physical basis for this delay, which we propose is due to the diffusion of active MCC complexes away from these unattached chromosomes throughout the cell, which consequently globally inhibit the APC/C. Materials and Methods Cell Culture, Synchronization and Stable Cell Line Generation PtK1 cells were cultured in 5% CO 2 in Ham s F-12 medium supplemented with 15% FBS and 1% penicillin/streptomycin. PtK1 cells were partially synchronized using the CDK1 inhibitor RO3306 (Sigma). Cells were treated with 9μM RO3306 (in DMSO) for 20 hrs, and subsequently released by washing with PBS 4-5 times and then replaced into drug-free media. To generate stable H2B-GFP expressing PtK1 cells, the following method was used. HEK-293T (the viral host) cells were transfected with ppax2 and pmd2.g viral packaging plasmids along with pgk:h2b-egfp vector (Addgene). PtK1 cells were then infected with resulting virus and cultured for 24 hours. Subsequently, fresh virus was added to these cells, which were then cultured for an additional 24 hours. 48 hours after the initial infection, cells were grown in selection media (Ham s F12 supplemented with Hygromycin B). Single colonies were then picked, expanded in 24 well plates in selection media, and then visually screened via fluorescence microscopy for H2B-GFP expression.
13 Cell Fusion PtK1 cells were fused as previously described 44. Fusion media was made by melting 8 g of poly-ethylene glycol 3500 (PEG) in an autoclave, cooling the molten PEG to 45 C, and diluting with 2 ml of DMSO and 10 ml of pre-warmed (37 C) Dulbecco s Modified Eagle Medium (DMEM) without FBS. To fuse cells, normal growth media was aspirated and fusion media was added to the cells for seconds. Cells were then repeatedly washed with PBS (4-5 times) until residual fusion media was removed, and replaced into Ham s F-12 medium supplemented with 15% FBS and 1% penicillin/streptomycin. Immunofluorescence Coverslip cultures of fused PtK1 cells were rinsed in PHEM Buffer (60 mm PIPES, 25 mm HEPES, 10 mm EGTA, 8 mm MgSO 4, ph 7.0), lysed in PHEM buffer supplemented with 1% Triton X-100 for 1 minute, and then fixed in 4% paraformaldehyde for 20 min. Cells were rinsed 3 x 5 minutes in PHEM buffer supplemented with 0.5% Triton X-100, and then blocked for 1 hour at room temperature with 10% boiled donkey serum (BDS) in PHEM. Primary antibodies were diluted in 5% BDS in PHEM as follows: mouse anti-α-tubulin, 1:300 (Sigma, T6199), and rabbit anti-mad1, 1:500 (Gene Tex , lot 40030). Cells were incubated with primary antibodies overnight at 4 C, and then rinsed 3 5 min in PHEM supplemented with 0.05% Triton X-100. Cells were incubated with appropriate secondary antibodies (donkey anti-mouse, or donkey anti-rabbit) conjugated to Alexa Fluor 647 or 555 (Jackson ImmunoResearch Laboratories) diluted 1:300 in 5% BDS in PHEM for 45 minutes at room temperature. Cells were then rinsed with PHEM supplemented with 0.05% Triton X min, rinsed once with PHEM, and mounted onto slides using mounting media (20 mm Tris, ph 8.0, 0.5% N-propyl gallate, and 90% glycerol). Coverslips were sealed to the slides using fingernail polish. Microscopy and Analysis Cells were imaged with a DeltaVision Personal DV (Applied Precision) imaging system equipped with a 40X/0.75NA Ph UPlanFL objective (Olympus) and a CoolSNAP HQ2 (Photometrics/Roper Scientific)
14 camera. The system was controlled with SoftWorx acquisition software (Applied Precision). Images for fixed-cell experiments (Figure 3B, C and E) were acquired as z-stacks with 200 nm step sizes. For live cell experiments, (Figures 1, 2, 3D, 4, and 5) cells were maintained at 37 C using an environmental chamber (Precision Control, Seattle, WA), and a single z-plane was acquired every 3 or 5 minutes depending on the experiment. Measuring Inter-spindle Distance and Constriction Coefficients Inter-spindle distance was measured in Fiji (NIH ImageJ) by determining the distance (i.e., the most direct intracellular path) between the centroid of each spindle (centroids were approximated by determining the midpoint between spindle poles). Because PtK1 cells remain flat during mitosis, distances in the z dimension are negligible, and were not used (not shown). We noted that fused asynchronous cells frequently displayed a severe constriction between the two spindles. When the constriction was very narrow (i.e., a constriction coefficient 0.1; see Figure 3Aii), it became difficult to determine whether cells were fused. Thus, we only included cells in which the constriction width was 10% the maximum width of the cells (i.e., constriction coefficient > 0.1). Constriction coefficient values were calculated by dividing the width of the narrowest point of the constriction ( c ) by the width of the widest point ( w ) in the cells (see Figure 3Aii). Acknowledgments This work was supported by the NIH (R01 GM to J.G.D and R01 GM to S.M.M.) and the NSF ( to J.G.D. and S.M.M.). We thank Keith DeLuca for technical assistance and stable cell generation.
15 Literature Cited 1. Santaguida S, Amon A. Short- and long-term effects of chromosome mis-segregation and aneuploidy. Nature Reviews Molecular Cell Biology. 2015;16(8): doi: /nrm Musacchio A. The Molecular Biology of Spindle Assembly Checkpoint Signaling Dynamics. Current biology: CB. 2015;25(20):R1002-R1018. doi: /j.cub Kops GJPL, Shah JV. Connecting up and clearing out: how kinetochore attachment silences the spindle assembly checkpoint. Chromosoma. 2012;121(5): doi: /s Rieder CL, Cole RW, Khodjakov A, Sluder G. The checkpoint delaying anaphase in response to chromosome monoorientation is mediated by an inhibitory signal produced by unattached kinetochores. The Journal of Cell Biology. 1995;130(4): Mapelli M, Musacchio A. MAD contortions: conformational dimerization boosts spindle checkpoint signaling. Curr Opin Struct Biol. 2007;17(6): doi: /j.sbi Luo X, Tang Z, Xia G, et al. The Mad2 spindle checkpoint protein has two distinct natively folded states. Nat Struct Mol Biol. 2004;11(4): Ji Z, Gao H, Jia L, Li B, Yu H. A sequential multi-target Mps1 phosphorylation cascade promotes spindle checkpoint signaling. elife. 2017;6. doi: /elife Faesen AC, Thanasoula M, Maffini S, et al. Basis of catalytic assembly of the mitotic checkpoint complex. Nature. January doi: /nature Han JS, Holland AJ, Fachinetti D, Kulukian A, Cetin B, Cleveland DW. Catalytic assembly of the mitotic checkpoint inhibitor BubR1-Cdc20 by a Mad2-induced functional switch in Cdc20. Molecular Cell. 2013;51(1): doi: /j.molcel Sudakin V, Chan GK, Yen TJ. Checkpoint inhibition of the APC/C in HeLa cells is mediated by a complex of BUBR1, BUB3, CDC20, and MAD2. The Journal of Cell Biology. 2001;154(5): doi: /jcb Yamaguchi M, VanderLinden R, Weissmann F, et al. Cryo-EM of Mitotic Checkpoint Complex- Bound APC/C Reveals Reciprocal and Conformational Regulation of Ubiquitin Ligation. Molecular Cell. 2016;63(4): doi: /j.molcel Collin P, Nashchekina O, Walker R, Pines J. The spindle assembly checkpoint works like a rheostat rather than a toggle switch. Nature Cell Biology. 2013;15(11): doi: /ncb Rieder CL, Khodjakov A, Paliulis LV, Fortier TM, Cole RW, Sluder G. Mitosis in vertebrate somatic cells with two spindles: Implications for the metaphase/anaphase transition checkpoint and cleavage. Proceedings of the National Academy of Sciences of the United States of America. 1997;94(10):
16 14. Lince-Faria M, Maffini S, Orr B, et al. Spatiotemporal control of mitosis by the conserved spindle matrix protein Megator. The Journal of Cell Biology. 2009;184(5): Yao C, Rath U, Maiato H, et al. A nuclear-derived proteinaceous matrix embeds the microtubule spindle apparatus during mitosis. Molecular Biology of the Cell. 2012;23(18): doi: /mbc.e Schweizer N, Pawar N, Weiss M, Maiato H. An organelle-exclusion envelope assists mitosis and underlies distinct molecular crowding in the spindle region. The Journal of Cell Biology. 2015;210(5): doi: /jcb De Souza CP, Hashmi SB, Nayak T, Oakley B, Osmani SA. Mlp1 acts as a mitotic scaffold to spatially regulate spindle assembly checkpoint proteins in Aspergillus nidulans. Molecular Biology of the Cell. 2009;20(8): doi: /mbc.e Schindelin J, Arganda-Carreras I, Frise E, et al. Fiji: an open-source platform for biological-image analysis. Nature Methods. 2012;9(7): doi: /nmeth Ciliberto A, Shah JV. A quantitative systems view of the spindle assembly checkpoint. The EMBO journal. 2009;28(15): Vanoosthuyse V, Hardwick KG. Overcoming inhibition in the spindle checkpoint. Genes \& Development. 2009;23(24): Cleveland DW, Mao Y, Sullivan KF. Centromeres and kinetochores: from epigenetics to mitotic checkpoint signaling. Cell. 2003;112(4): Yu H. Regulation of APC-Cdc20 by the spindle checkpoint. Current Opinion in Cell Biology. 2002;14(6): Lara-Gonzalez P, Westhorpe FG, Taylor SS. The spindle assembly checkpoint. Current biology: CB. 2012;22(22):R966-R980. doi: /j.cub Hardwick KG. Checkpoint signalling: Mad2 conformers and signal propagation. Current biology: CB. 2005;15(4):R122-R124. doi: /j.cub Acquaviva C, Herzog F, Kraft C, Pines J. The anaphase promoting complex/cyclosome is recruited to centromeres by the spindle assembly checkpoint. Nature Cell Biology. 2004;6(9): Sivakumar S, Gorbsky GJ. Spatiotemporal regulation of the anaphase-promoting complex in mitosis. Nature Reviews Molecular Cell Biology. 2015;16(2): doi: /nrm Topper LM, Campbell MS, Tugendreich S, et al. The dephosphorylated form of the anaphasepromoting complex protein Cdc27/Apc3 concentrates on kinetochores and chromosome arms in mitosis. Cell Cycle (Georgetown, Tex). 2002;1(4): Torres JZ, Ban KH, Jackson PK. A specific form of phospho protein phosphatase 2 regulates anaphase-promoting complex/cyclosome association with spindle poles. Molecular Biology of the Cell. 2010;21(6): doi: /mbc.e
17 29. Sivakumar S, Daum JR, Tipton AR, Rankin S, Gorbsky GJ. The spindle and kinetochoreassociated (Ska) complex enhances binding of the anaphase-promoting complex/cyclosome (APC/C) to chromosomes and promotes mitotic exit. Molecular Biology of the Cell. 2014;25(5): doi: /mbc.e Vassilev LT, Tovar C, Chen S, et al. Selective small-molecule inhibitor reveals critical mitotic functions of human CDK1. Proceedings of the National Academy of Sciences of the United States of America. 2006;103(28): McIntosh JR. Mitosis. Cold Spring Harb Perspect Biol. 2016;8(9). doi: /cshperspect.a Howell BJ, Moree B, Farrar EM, Stewart S, Fang G, Salmon ED. Spindle checkpoint protein dynamics at kinetochores in living cells. Current biology: CB. 2004;14(11): doi: /j.cub Johnson RT, Rao PN, Hughes HD. Mammalian cell fusion. 3. A HeLa cell inducer of premature chromosome condensation active in cells from a variety of animal species. J Cell Physiol. 1970;76(2): doi: /jcp Johnson RT, Rao PN. Mammalian cell fusion: induction of premature chromosome condensation in interphase nuclei. Nature. 1970;226(5247): Rao PN, Johnson RT. Mammalian cell fusion: studies on the regulation of DNA synthesis and mitosis. Nature. 1970;225(5228): Howell BJ, Hoffman DB, Fang G, Murray AW, Salmon ED. Visualization of Mad2 dynamics at kinetochores, along spindle fibers, and at spindle poles in living cells. The Journal of Cell Biology. 2000;150(6): Galli M, Morgan DO. Cell Size Determines the Strength of the Spindle Assembly Checkpoint during Embryonic Development. Developmental Cell. 2016;36(3): doi: /j.devcel Fraschini R, Beretta A, Sironi L, Musacchio A, Lucchini G, Piatti S. Bub3 interaction with Mad2, Mad3 and Cdc20 is mediated by WD40 repeats and does not require intact kinetochores. The EMBO journal. 2001;20(23): doi: /emboj/ Maciejowski J, George KA, Terret M-E, Zhang C, Shokat KM, Jallepalli PV. Mps1 directs the assembly of Cdc20 inhibitory complexes during interphase and mitosis to control M phase timing and spindle checkpoint signaling. The Journal of Cell Biology. 2010;190(1): doi: /jcb Rodriguez-Bravo V, Maciejowski J, Corona J, et al. Nuclear pores protect genome integrity by assembling a premitotic and Mad1-dependent anaphase inhibitor. Cell. 2014;156(5): doi: /j.cell Hara K, Tydeman P, Kirschner M. A cytoplasmic clock with the same period as the division cycle in Xenopus eggs. Proceedings of the National Academy of Sciences of the United States of America. 1980;77(1):
:129-137.")
18 42. Zhang M, Kothari P, Lampson MA. Spindle Assembly Checkpoint Acquisition at the Mid- Blastula Transition. Foltz D, ed. PloS One. 2015;10(3):e Daphnis DD, Delhanty JDA, Jerkovic S, Geyer J, Craft I, Harper JC. Detailed FISH analysis of day 5 human embryos reveals the mechanisms leading to mosaic aneuploidy. Hum Reprod. 2005;20(1): doi: /humrep/deh Peterson SP, Berns MW. Mitosis in flat PTK2-human hybrid cells. Experimental Cell Research. 1979;120(2):
19 Figure 1. The CDK1 inhibitor RO3306 partially synchronizes PtK1 cells in mitosis without affecting mitotic timing. A) Plot depicting fraction of cells in the indicated phase of the cell cycle in the absence or presence of RO3306, followed by drug washout, as indicated. A 20-hour RO3306 treatment followed by drug washout leads to a partial enrichment of cells in mitosis. B) Duration of prometaphase (nuclear envelope breakdown to metaphase, or NEBD-M) and metaphase (metaphase to anaphase, or M-A) in asynchronous cells, RO3306 synchronized cells, or RO3306 synchronized cells exposed to fusion media (specifically those that remain unfused; see Materials and Methods). Significance was determined using 1-way analysis of variance tests (n.s., not significant). C) Example time frames depicting mitotic progression (starting at late prometaphase) of a PtK1 cell stably expressing H2B-GFP treated with RO3306 for 20 hours and then released into non-ro3306 containing media. Scale bar is 10µm.
 Models describing our predictions that diffusible wait anaphase signals should synchronize anaphase onset of multiple mitotic spindle compartments (left), but spindle-restricted signals should not")
20 Figure 2. Spindles wait to initiate anaphase together. A) Models describing our predictions that diffusible wait anaphase signals should synchronize anaphase onset of multiple mitotic spindle compartments (left), but spindle-restricted signals should not (right). B) Representative time-course of two mitotic cells stably expressing H2B-GFP fusing (at 15 ) and undergoing synchronous anaphase ( M, metaphase spindle; A, anaphase spindle). C) Quantification of metaphase duration in early and late spindles in synchronous anaphase cells (see text). Data set of RO3306 synchronized, fusion media exposed cells from
21 Figure 1B is included for comparative purposes ( Single Spindle ). Statistical significance was determined using unpaired T-tests (**, p 0.001; ***, p ).
22
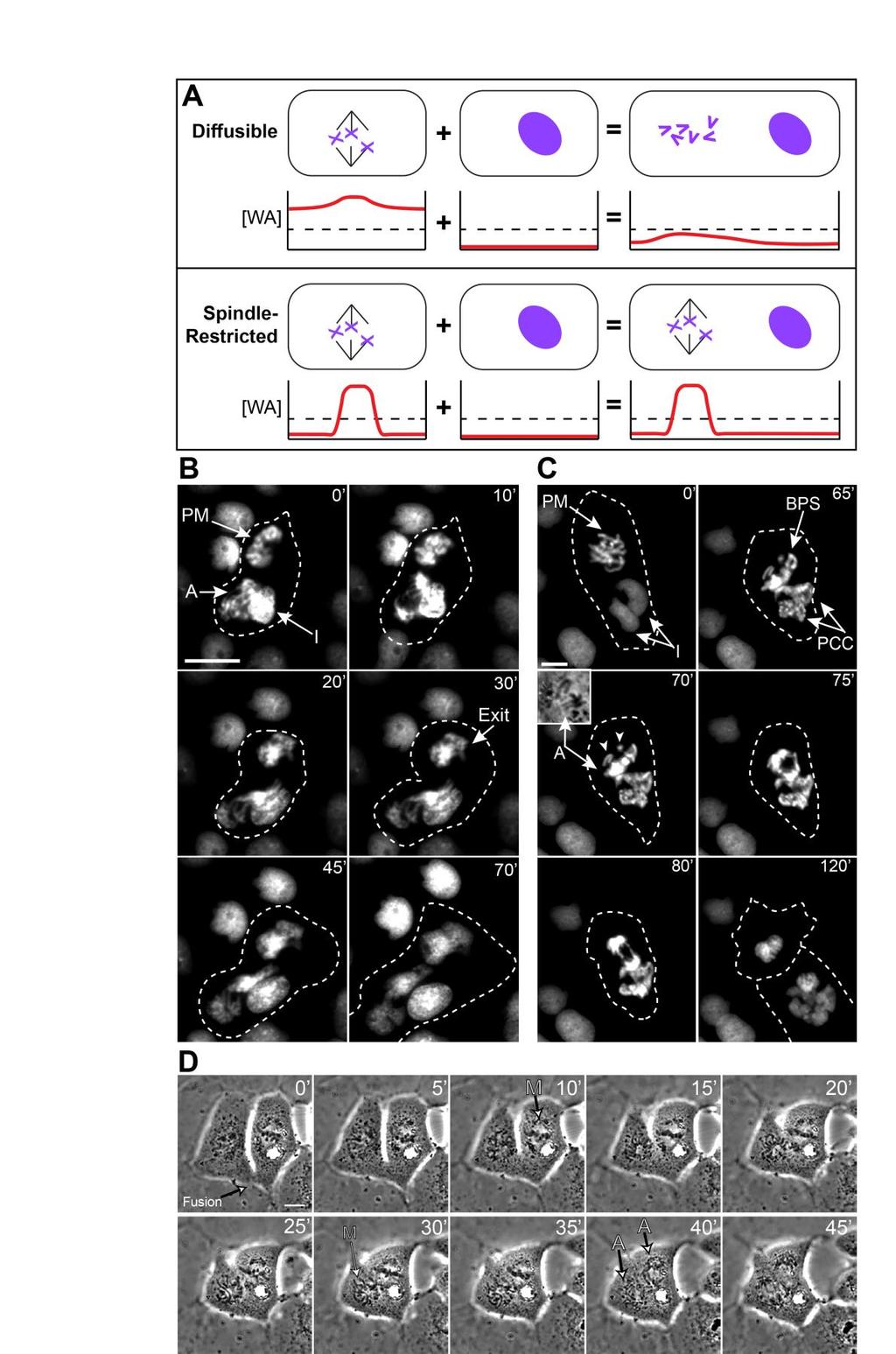
23 Figure 3. Synchrony of anaphase onset correlates with reduced inter-spindle distance and cytoplasmic constriction in bi-spindled cells. A) A model describing our predictions for how diffusible and spindlerestricted wait anaphase complexes would respond to three different cellular scenarios: (i) one in which two spindles are in relative close proximity and have no cellular constriction; (ii) one in which the spindles are relatively distal, and have only a moderate cellular constriction; and (iii) one in which the spindles are relatively distal, and have a severe cellular constriction. Cartoon graphs represent the concentration of wait anaphase complexes ([WA]) across the length of a bi-spindled cell (red line). The black dashed line denotes the threshold of wait anaphase signals needed to prevent the spindle compartments from initiating anaphase. For diffusible complexes, less constriction (i and ii) will allow the levels of active MCC complexes to remain above the threshold necessary to prevent mitotic exit across the entirety of the cell. However, when the cell is constricted to the point of restricting diffusion from one side of the cell to the other (iii), active MCC complexes are restricted in the extent of their reach, and thus cannot influence the behavior of multiple spindle compartments. In iii, top, a spindle compartment in metaphase (right) was not prevented from entering anaphase because the constriction impeded the diffusion of active MCC molecules from the spindle compartment in prometaphase (left). If MCC molecules are restricted to the spindle compartment, spindle behavior would be autonomous to each spindle regardless of any barriers to diffusion between the two compartments ( w, widest dimension for a given cell; c, constriction, or narrowest point for a given cell). B) Micrographs of fused, fixed, and stained (as indicated) mitotic cells with two spindles undergoing either synchronous anaphase (left) or asynchronous anaphase (right). C) Plot depicting anaphase behavior in relation to inter-spindle distance, as measured from fixed cells. Purple and green color gradients indicate approximate distance thresholds in which spindles have a high probability of being synchronous (<~30 µm) or asynchronous (>~40 µm), respectively. D) Representative time-lapse images of a fused cell (with one nucleus stably expressing H2B-GFP) developing a cytoplasmic constriction. Insets at 60 and 70 depict H2B-GFP fluorescence. E) Plot depicting anaphase behavior in relation to the constriction coefficient, as measured from fixed cells.
24 Statistical significance was determined using unpaired T-tests. Scale bars are 10µm. For panels C and E, ***, p
25
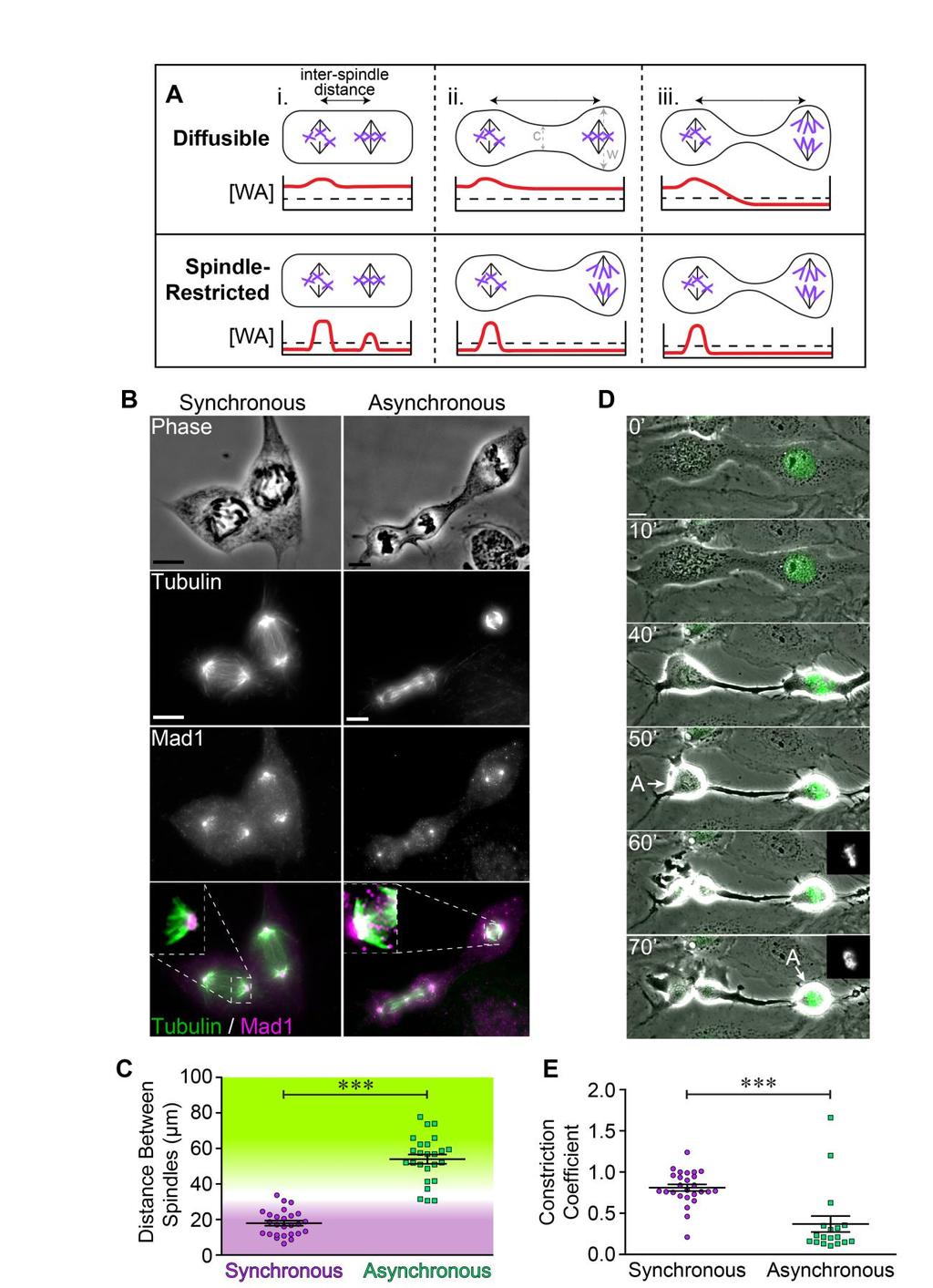
26 Figure 4. Anaphase synchrony is distance dependent. In this example of a trinucleate cell (unannotated on the left, annotated on the right), all nuclei are in prophase at 0. Two nuclei are clustered at the top of the cell (bounded by blue and green outlines in annotated images on the right). The green nucleus is just out of the field of view at 0, but moves into view in subsequent frames. The third nucleus is at the bottom of the cell (bounded by a red outline). By 20, all three nuclei have initiated NEBD, a cellular constriction has formed between the closely apposed nuclei (blue and green) and the distal nucleus (red), and the blue and green nuclei begin to align their chromosomes. At 85, the blue and green spindle compartments initiate anaphase synchronously while the red spindle compartment remains in prometaphase. At 140, the red spindle compartment initiates anaphase, 55 after the blue and green spindle compartments initiated anaphase. At 250, all cell products from the three mitotic spindles re-enter interphase. The nuclei produced from each mitotic spindle at 250 are outlined in their respective colors. One of the green daughter cells is out of frame. Scale bars are 10 µm.
27
28 Figure 5. Dilution of wait anaphase signals affects SAC activity. A) A model describing how diffusible and spindle restricted wait anaphase complexes should respond to cytoplasmic dilution. As in Figure 3A, cartoon graphs depict the concentration of wait anaphase complexes across the cell (red line). The threshold of wait anaphase signal concentration required to prevent anaphase onset is denoted with a black dashed line. If wait anaphase signals are diffusible, the addition of non-mitotic cytoplasm via fusion with a non-mitotic cell (cells on the right in each example) would dilute the overall concentration of wait anaphase complexes. This would result in premature mitotic exit (i.e., spindle disassembly, as depicted in the fused cell cartoon, top) since the concentration of wait anaphase signaling molecules (i.e., active MCC complexes) drops below the threshold required for SAC-mediated arrest. Conversely, if active MCC complexes are restricted to the spindle compartment, addition of cytoplasm would not perturb SAC activity, and each spindle compartment would progress through mitosis with normal timing. B-C) Representative time-lapse images of fused H2B-GFP expressing cells in which mitotic spindle compartments prematurely exit mitosis following fusion with non-mitotic cells (see text). Inset in panel C (at 70 ) shows separation of sister chromatids as an indicator of anaphase onset. D) Representative timelapse images of fused RO3306 synchronized cells demonstrating that cells already in mitosis and G2 prior to cell fusion undergo synchronous anaphase. In this binucleate cell, the spindle compartment in metaphase ( M, at 10 ) delays anaphase onset until the interphase nucleus (left; presumably in G2 phase at point of fusion) enters mitosis (30 ) and aligns its chromosomes, at which point both spindle compartments initiate anaphase (40 ). Scale bars are 10µm ( M, metaphase; A, anaphase; PM, prometaphase; I, interphase; Exit, mitotic exit; BPS, bipolar spindle; PCC, premature chromosome condensation; arrowheads, unaligned chromosomes).
基醫所. The Cell Cycle. Chi-Wu Chiang, Ph.D. IMM, NCKU
 基醫所 The Cell Cycle Chi-Wu Chiang, Ph.D. IMM, NCKU 1 1 Introduction to cell cycle and cell cycle checkpoints 2 2 Cell cycle A cell reproduces by performing an orderly sequence of events in which it duplicates
基醫所 The Cell Cycle Chi-Wu Chiang, Ph.D. IMM, NCKU 1 1 Introduction to cell cycle and cell cycle checkpoints 2 2 Cell cycle A cell reproduces by performing an orderly sequence of events in which it duplicates
Regulators of Cell Cycle Progression
 Regulators of Cell Cycle Progression Studies of Cdk s and cyclins in genetically modified mice reveal a high level of plasticity, allowing different cyclins and Cdk s to compensate for the loss of one
Regulators of Cell Cycle Progression Studies of Cdk s and cyclins in genetically modified mice reveal a high level of plasticity, allowing different cyclins and Cdk s to compensate for the loss of one
Chromosomes Can Congress To The Metaphase Plate Prior. To Bi-Orientation
 Chromosomes Can Congress To The Metaphase Plate Prior To Bi-Orientation Tarun M. Kapoor 1,2, Michael Lampson 1, Polla Hergert 3, Lisa Cameron 2,4, Daniela Cimini 4, E.D. Salmon 2,4, Bruce F. McEwen 3,
Chromosomes Can Congress To The Metaphase Plate Prior To Bi-Orientation Tarun M. Kapoor 1,2, Michael Lampson 1, Polla Hergert 3, Lisa Cameron 2,4, Daniela Cimini 4, E.D. Salmon 2,4, Bruce F. McEwen 3,
How Cells Divide. Chapter 10
 How Cells Divide Chapter 10 Bacterial Cell Division Bacteria divide by binary fission. -the single, circular bacterial chromosome is replicated -replication begins at the origin of replication and proceeds
How Cells Divide Chapter 10 Bacterial Cell Division Bacteria divide by binary fission. -the single, circular bacterial chromosome is replicated -replication begins at the origin of replication and proceeds
Molecular Cell Biology - Problem Drill 22: The Mechanics of Cell Division
 Molecular Cell Biology - Problem Drill 22: The Mechanics of Cell Division Question No. 1 of 10 1. Which of the following statements about mitosis is correct? Question #1 (A) Mitosis involves the dividing
Molecular Cell Biology - Problem Drill 22: The Mechanics of Cell Division Question No. 1 of 10 1. Which of the following statements about mitosis is correct? Question #1 (A) Mitosis involves the dividing
2014 Pearson Education, Inc.
 2 The Cell Cycle CAMPBELL BIOLOGY TENTH EDITION Reece Urry Cain Wasserman Minorsky Jackson The Key Roles of Cell Division The ability of organisms to produce more of their own kind best distinguishes living
2 The Cell Cycle CAMPBELL BIOLOGY TENTH EDITION Reece Urry Cain Wasserman Minorsky Jackson The Key Roles of Cell Division The ability of organisms to produce more of their own kind best distinguishes living
The Cell Cycle. Packet #9. Thursday, August 20, 2015
 1 The Cell Cycle Packet #9 2 Introduction Cell Cycle An ordered sequence of events in the life of a dividing eukaryotic cell and is a cellular asexual reproduction. The contents of the parent s cell nucleus
1 The Cell Cycle Packet #9 2 Introduction Cell Cycle An ordered sequence of events in the life of a dividing eukaryotic cell and is a cellular asexual reproduction. The contents of the parent s cell nucleus
LECTURE PRESENTATIONS
 LECTURE PRESENTATIONS For CAMPBELL BIOLOGY, NINTH EDITION Jane B. Reece, Lisa A. Urry, Michael L. Cain, Steven A. Wasserman, Peter V. Minorsky, Robert B. Jackson Chapter 12 The Cell Cycle Lectures by Erin
LECTURE PRESENTATIONS For CAMPBELL BIOLOGY, NINTH EDITION Jane B. Reece, Lisa A. Urry, Michael L. Cain, Steven A. Wasserman, Peter V. Minorsky, Robert B. Jackson Chapter 12 The Cell Cycle Lectures by Erin
BIOLOGY. The Cell Cycle CAMPBELL. Reece Urry Cain Wasserman Minorsky Jackson. Lecture Presentation by Nicole Tunbridge and Kathleen Fitzpatrick
 CAMPBELL BIOLOGY TENTH EDITION Reece Urry Cain Wasserman Minorsky Jackson 12 The Cell Cycle Lecture Presentation by Nicole Tunbridge and Kathleen Fitzpatrick The Key Roles of Cell Division The ability
CAMPBELL BIOLOGY TENTH EDITION Reece Urry Cain Wasserman Minorsky Jackson 12 The Cell Cycle Lecture Presentation by Nicole Tunbridge and Kathleen Fitzpatrick The Key Roles of Cell Division The ability
Mitosis THE CELL CYCLE. In unicellular organisms, division of one cell reproduces the entire organism Multicellular organisms use cell division for..
 Mitosis THE CELL CYCLE In unicellular organisms, division of one cell reproduces the entire organism Multicellular organisms use cell division for.. Development from a fertilized cell Growth Repair Cell
Mitosis THE CELL CYCLE In unicellular organisms, division of one cell reproduces the entire organism Multicellular organisms use cell division for.. Development from a fertilized cell Growth Repair Cell
LECTURE PRESENTATIONS
 LECTURE PRESENTATIONS For CAMPBELL BIOLOGY, NINTH EDITION Jane B. Reece, Lisa A. Urry, Michael L. Cain, Steven A. Wasserman, Peter V. Minorsky, Robert B. Jackson Chapter 12 The Cell Cycle Lectures by Erin
LECTURE PRESENTATIONS For CAMPBELL BIOLOGY, NINTH EDITION Jane B. Reece, Lisa A. Urry, Michael L. Cain, Steven A. Wasserman, Peter V. Minorsky, Robert B. Jackson Chapter 12 The Cell Cycle Lectures by Erin
Lecture 10. G1/S Regulation and Cell Cycle Checkpoints. G1/S regulation and growth control G2 repair checkpoint Spindle assembly or mitotic checkpoint
 Lecture 10 G1/S Regulation and Cell Cycle Checkpoints Outline: G1/S regulation and growth control G2 repair checkpoint Spindle assembly or mitotic checkpoint Paper: The roles of Fzy/Cdc20 and Fzr/Cdh1
Lecture 10 G1/S Regulation and Cell Cycle Checkpoints Outline: G1/S regulation and growth control G2 repair checkpoint Spindle assembly or mitotic checkpoint Paper: The roles of Fzy/Cdc20 and Fzr/Cdh1
Outline Interphase Mitotic Stage Cell Cycle Control Apoptosis Mitosis Mitosis in Animal Cells Cytokinesis Cancer Prokaryotic Cell Division
 The Cell Cycle and Cellular Reproduction Chapter 9 Outline Interphase Mitotic Stage Cell Cycle Control Apoptosis Mitosis Mitosis in Animal Cells Cytokinesis Cancer Prokaryotic Cell Division 1 2 Interphase
The Cell Cycle and Cellular Reproduction Chapter 9 Outline Interphase Mitotic Stage Cell Cycle Control Apoptosis Mitosis Mitosis in Animal Cells Cytokinesis Cancer Prokaryotic Cell Division 1 2 Interphase
Mitosis. AND Cell DiVISION
 Mitosis AND Cell DiVISION Cell Division Characteristic of living things: ability to reproduce their own kind. Cell division purpose: When unicellular organisms such as amoeba divide to form offspring reproduction
Mitosis AND Cell DiVISION Cell Division Characteristic of living things: ability to reproduce their own kind. Cell division purpose: When unicellular organisms such as amoeba divide to form offspring reproduction
CELL CYCLE INTRODUCTION PART I ANIMAL CELL CYCLE INTERPHASE
 CELL CYCLE INTRODUCTION The nuclei in cells of eukaryotic organisms contain chromosomes with clusters of genes, discrete units of hereditary information consisting of double-stranded DNA. Structural proteins
CELL CYCLE INTRODUCTION The nuclei in cells of eukaryotic organisms contain chromosomes with clusters of genes, discrete units of hereditary information consisting of double-stranded DNA. Structural proteins
Mad2-independent spindle assembly checkpoint activation and controlled. metaphase-anaphase transition in Drosophila S2 cells
 Mad2-independent spindle assembly checkpoint activation and controlled metaphase-anaphase transition in Drosophila S2 cells Bernardo Orr 1, Hassan Bousbaa 2 and Claudio E. Sunkel 1,3,4 1) Instituto de
Mad2-independent spindle assembly checkpoint activation and controlled metaphase-anaphase transition in Drosophila S2 cells Bernardo Orr 1, Hassan Bousbaa 2 and Claudio E. Sunkel 1,3,4 1) Instituto de
Cell Cycle. Trends in Cell Biology
 Cell Cycle Trends in Cell Biology Cell Cycle The orderly sequence of events by which a cell duplicates its contents and divides into two Daughter Cells Activities of a cell from one cell division to the
Cell Cycle Trends in Cell Biology Cell Cycle The orderly sequence of events by which a cell duplicates its contents and divides into two Daughter Cells Activities of a cell from one cell division to the
T H E J O U R N A L O F C E L L B I O L O G Y
 T H E J O U R N A L O F C E L L B I O L O G Y Supplemental material Lu et al., http://www.jcb.org/cgi/content/full/jcb.201012063/dc1 Figure S1. Kinetics of nuclear envelope assembly, recruitment of Nup133
T H E J O U R N A L O F C E L L B I O L O G Y Supplemental material Lu et al., http://www.jcb.org/cgi/content/full/jcb.201012063/dc1 Figure S1. Kinetics of nuclear envelope assembly, recruitment of Nup133
The Cell Cycle 4/10/12. Chapter 12. Overview: The Key Roles of Cell Division
 LECTURE PRESENTATIONS For CAMPBELL BIOLOGY, NINTH EDITION Jane B. Reece, Lisa A. Urry, Michael L. Cain, Steven A. Wasserman, Peter V. Minorsky, Robert B. Jackson Chapter 12 The Cell Cycle Lectures by Erin
LECTURE PRESENTATIONS For CAMPBELL BIOLOGY, NINTH EDITION Jane B. Reece, Lisa A. Urry, Michael L. Cain, Steven A. Wasserman, Peter V. Minorsky, Robert B. Jackson Chapter 12 The Cell Cycle Lectures by Erin
The Cell Cycle. Dr. SARRAY Sameh, Ph.D
 The Cell Cycle Dr. SARRAY Sameh, Ph.D Overview When an organism requires additional cells (either for growth or replacement of lost cells), new cells are produced by cell division (mitosis) Somatic cells
The Cell Cycle Dr. SARRAY Sameh, Ph.D Overview When an organism requires additional cells (either for growth or replacement of lost cells), new cells are produced by cell division (mitosis) Somatic cells
CELL CYCLE INTRODUCTION PART I ANIMAL CELL CYCLE INTERPHASE EVOLUTION/HEREDITY UNIT. Activity #3
 AP BIOLOGY EVOLUTION/HEREDITY UNIT Unit 1 Part 3 Chapter 12 Activity #3 INTRODUCTION CELL CYCLE NAME DATE PERIOD The nuclei in cells of eukaryotic organisms contain chromosomes with clusters of genes,
AP BIOLOGY EVOLUTION/HEREDITY UNIT Unit 1 Part 3 Chapter 12 Activity #3 INTRODUCTION CELL CYCLE NAME DATE PERIOD The nuclei in cells of eukaryotic organisms contain chromosomes with clusters of genes,
The Cell Cycle. Chapter 12. PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece
 Chapter 12 The Cell Cycle PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions from Joan Sharp Overview:
Chapter 12 The Cell Cycle PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions from Joan Sharp Overview:
The Cell Cycle CAMPBELL BIOLOGY IN FOCUS SECOND EDITION URRY CAIN WASSERMAN MINORSKY REECE
 CAMPBELL BIOLOGY IN FOCUS URRY CAIN WASSERMAN MINORSKY REECE 9 The Cell Cycle Lecture Presentations by Kathleen Fitzpatrick and Nicole Tunbridge, Simon Fraser University SECOND EDITION Overview: The Key
CAMPBELL BIOLOGY IN FOCUS URRY CAIN WASSERMAN MINORSKY REECE 9 The Cell Cycle Lecture Presentations by Kathleen Fitzpatrick and Nicole Tunbridge, Simon Fraser University SECOND EDITION Overview: The Key
Cell Cycle, Mitosis, and Microtubules. LS1A Final Exam Review Friday 1/12/07. Processes occurring during cell cycle
 Cell Cycle, Mitosis, and Microtubules LS1A Final Exam Review Friday 1/12/07 Processes occurring during cell cycle Replicate chromosomes Segregate chromosomes Cell divides Cell grows Cell Growth 1 The standard
Cell Cycle, Mitosis, and Microtubules LS1A Final Exam Review Friday 1/12/07 Processes occurring during cell cycle Replicate chromosomes Segregate chromosomes Cell divides Cell grows Cell Growth 1 The standard
University of Bristol - Explore Bristol Research
 Shandilya, J., Medler, K., & Roberts, S. G. E. (2016). Regulation of AURORA B function by mitotic checkpoint protein MAD2. Cell Cycle, 15(16), 2196-2201. https://doi.org/10.1080/15384101.2016.1200773 Peer
Shandilya, J., Medler, K., & Roberts, S. G. E. (2016). Regulation of AURORA B function by mitotic checkpoint protein MAD2. Cell Cycle, 15(16), 2196-2201. https://doi.org/10.1080/15384101.2016.1200773 Peer
General Biology. Overview: The Key Roles of Cell Division The continuity of life is based upon the reproduction of cells, or cell division
 General Biology Course No: BNG2003" Credits: 3.00 " " " 8. The Cell Cycle Prof. Dr. Klaus Heese Overview: The Key Roles of Cell Division The continuity of life is based upon the reproduction of cells,
General Biology Course No: BNG2003" Credits: 3.00 " " " 8. The Cell Cycle Prof. Dr. Klaus Heese Overview: The Key Roles of Cell Division The continuity of life is based upon the reproduction of cells,
-The cell s hereditary endowment of DNA -Usually packaged into chromosomes for manageability
 Binary Fission-Bacterial Cell Division -Asexual reproduction of prokaryotes -No mitosis -Circular DNA and organelles replicate, the copies migrate to opposite sides of the elongating cell, and the cell
Binary Fission-Bacterial Cell Division -Asexual reproduction of prokaryotes -No mitosis -Circular DNA and organelles replicate, the copies migrate to opposite sides of the elongating cell, and the cell
The Cell Cycle and How Cells Divide
 The Cell Cycle and How Cells Divide 1 Phases of the Cell Cycle The cell cycle consists of Interphase normal cell activity The mitotic phase cell divsion INTERPHASE Growth G 1 (DNA synthesis) Growth G 2
The Cell Cycle and How Cells Divide 1 Phases of the Cell Cycle The cell cycle consists of Interphase normal cell activity The mitotic phase cell divsion INTERPHASE Growth G 1 (DNA synthesis) Growth G 2
The Cell Cycle. Chapter 12. Biology Eighth Edition Neil Campbell and Jane Reece. PowerPoint Lecture Presentations for
 Chapter 12 The Cell Cycle PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions from Joan Sharp Copyright
Chapter 12 The Cell Cycle PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions from Joan Sharp Copyright
Bacterial cell. Origin of replication. Septum
 Bacterial cell Bacterial chromosome: Double-stranded DNA Origin of replication Septum 1 2 3 Chromosome Rosettes of Chromatin Loops Scaffold protein Chromatin Loop Solenoid Scaffold protein Chromatin loop
Bacterial cell Bacterial chromosome: Double-stranded DNA Origin of replication Septum 1 2 3 Chromosome Rosettes of Chromatin Loops Scaffold protein Chromatin Loop Solenoid Scaffold protein Chromatin loop
Name. A.P. Biology Chapter 12 The Cell Cycle
 A.P. Biology Chapter 12 The Cell Cycle Name Living species MUST possess the ability to r if they are to flourish. The Cell Cycle follows the life of a cell from its o until its d. The Key Roles Of Cell
A.P. Biology Chapter 12 The Cell Cycle Name Living species MUST possess the ability to r if they are to flourish. The Cell Cycle follows the life of a cell from its o until its d. The Key Roles Of Cell
Cell Division and Mitosis
 Chromatin-Uncoiled DNA during interphase Cell Division and Mitosis Chromosomes-Tightly coiled DNA Chromatid-One half of a duplicated chromosome. Each is identical and called sister chromatids Centromere-The
Chromatin-Uncoiled DNA during interphase Cell Division and Mitosis Chromosomes-Tightly coiled DNA Chromatid-One half of a duplicated chromosome. Each is identical and called sister chromatids Centromere-The
(a) Reproduction. (b) Growth and development. (c) Tissue renewal
 Reproduction. (b) Growth and development. (c) Tissue renewal") 100 µm 200 µm 20 µm (a) Reproduction (b) Growth and development (c) Tissue renewal 1 20 µm 2 0.5 µm Chromosomes DNA molecules Chromosome arm Centromere Chromosome duplication (including DNA synthesis)
100 µm 200 µm 20 µm (a) Reproduction (b) Growth and development (c) Tissue renewal 1 20 µm 2 0.5 µm Chromosomes DNA molecules Chromosome arm Centromere Chromosome duplication (including DNA synthesis)
General Biology. Overview: The Key Roles of Cell Division. Unicellular organisms
 General Biology Course No: BNG2003 Credits: 3.00 8. The Cell Cycle Prof. Dr. Klaus Heese Overview: The Key Roles of Cell Division The continuity of life is based upon the reproduction of cells, or cell
General Biology Course No: BNG2003 Credits: 3.00 8. The Cell Cycle Prof. Dr. Klaus Heese Overview: The Key Roles of Cell Division The continuity of life is based upon the reproduction of cells, or cell
BIOLOGY - CLUTCH CH.12 - CELL DIVISION.
 !! www.clutchprep.com CONCEPT: CELL DIVISION Cell division is the process by which one cell splits into two or more daughter cells. Cell division generally requires that cells produce enough materials,
!! www.clutchprep.com CONCEPT: CELL DIVISION Cell division is the process by which one cell splits into two or more daughter cells. Cell division generally requires that cells produce enough materials,
Biology is the only subject in which multiplication is the same thing as division
 Biology is the only subject in which multiplication is the same thing as division 2007-2008 The Cell Cycle: Cell Growth, Cell Division Ch. 10 Where it all began You started as a cell smaller than a period
Biology is the only subject in which multiplication is the same thing as division 2007-2008 The Cell Cycle: Cell Growth, Cell Division Ch. 10 Where it all began You started as a cell smaller than a period
LECTURE PRESENTATIONS
 LECTURE PRESENTATIONS For CAMPBELL BIOLOGY, NINTH EDITION Jane B. Reece, Lisa A. Urry, Michael L. Cain, Steven A. Wasserman, Peter V. Minorsky, Robert B. Jackson Chapter 12 The Cell Cycle Lectures by Erin
LECTURE PRESENTATIONS For CAMPBELL BIOLOGY, NINTH EDITION Jane B. Reece, Lisa A. Urry, Michael L. Cain, Steven A. Wasserman, Peter V. Minorsky, Robert B. Jackson Chapter 12 The Cell Cycle Lectures by Erin
Chapter 8: Cellular Reproduction
 Chapter 8: Cellular Reproduction 1. The Cell Cycle 2. Mitosis 3. Meiosis 2 Types of Cell Division 2n 1n Mitosis: occurs in somatic cells (almost all cells of the body) generates cells identical to original
Chapter 8: Cellular Reproduction 1. The Cell Cycle 2. Mitosis 3. Meiosis 2 Types of Cell Division 2n 1n Mitosis: occurs in somatic cells (almost all cells of the body) generates cells identical to original
Ploidy and Human Cell Types. Cell Cycle and Mitosis. DNA and Chromosomes. Where It All Began 11/19/2014. Chapter 12 Pg
 Ploidy and Human Cell Types Cell Cycle and Mitosis Chapter 12 Pg. 228 245 Cell Types Somatic cells (body cells) have 46 chromosomes, which is the diploid chromosome number. A diploid cell is a cell with
Ploidy and Human Cell Types Cell Cycle and Mitosis Chapter 12 Pg. 228 245 Cell Types Somatic cells (body cells) have 46 chromosomes, which is the diploid chromosome number. A diploid cell is a cell with
Cell Cycle - Introduction
 Cell Cycle - Introduction Key Concepts Cell division results in two identical cells During cell division the ability to organize DNA in time and space (location in the cell) is critical! The mitotic phase
Cell Cycle - Introduction Key Concepts Cell division results in two identical cells During cell division the ability to organize DNA in time and space (location in the cell) is critical! The mitotic phase
Campbell Biology in Focus (Urry) Chapter 9 The Cell Cycle. 9.1 Multiple-Choice Questions
 Chapter 9 The Cell Cycle. 9.1 Multiple-Choice Questions") Campbell Biology in Focus (Urry) Chapter 9 The Cell Cycle 9.1 Multiple-Choice Questions 1) Starting with a fertilized egg (zygote), a series of five cell divisions would produce an early embryo with how
Campbell Biology in Focus (Urry) Chapter 9 The Cell Cycle 9.1 Multiple-Choice Questions 1) Starting with a fertilized egg (zygote), a series of five cell divisions would produce an early embryo with how
Chapter 8 The Cell Cycle
 What molecule stores your genetic information or determines everything about you? DNA a nucleic acid How are DNA molecules arranged in the nucleus? As you can see DNA is: Chapter 8 The Cell Cycle 1. Arranged
What molecule stores your genetic information or determines everything about you? DNA a nucleic acid How are DNA molecules arranged in the nucleus? As you can see DNA is: Chapter 8 The Cell Cycle 1. Arranged
Reproduction is a fundamental property of life. Cells are the fundamental unit of life. Reproduction occurs at the cellular level with one mother
 Cell Division ision Reproduction is a fundamental property of life. Cells are the fundamental unit of life. Reproduction occurs at the cellular level with one mother cell giving rise to two daughter cells.
Cell Division ision Reproduction is a fundamental property of life. Cells are the fundamental unit of life. Reproduction occurs at the cellular level with one mother cell giving rise to two daughter cells.
The Cell Cycle CHAPTER 12
 The Cell Cycle CHAPTER 12 The Key Roles of Cell Division cell division = reproduction of cells All cells come from pre-exisiting cells Omnis cellula e cellula Unicellular organisms division of 1 cell reproduces
The Cell Cycle CHAPTER 12 The Key Roles of Cell Division cell division = reproduction of cells All cells come from pre-exisiting cells Omnis cellula e cellula Unicellular organisms division of 1 cell reproduces
Origin of replication. Septum
 Bacterial cell Bacterial chromosome: Double-stranded DNA Origin of replication Septum 1 2 3 Chromosome Rosettes of Chromatin Loops Chromatin Loop Solenoid Scaffold protein Scaffold protein Chromatin loop
Bacterial cell Bacterial chromosome: Double-stranded DNA Origin of replication Septum 1 2 3 Chromosome Rosettes of Chromatin Loops Chromatin Loop Solenoid Scaffold protein Scaffold protein Chromatin loop
The Cell Cycle. Chapter 12. Key Concepts in Chapter 12. Overview: The Key Roles of Cell Division. Video: Sea Urchin Embryonic Development (time-lapse)
") Chapter 12 The Cell Cycle Dr. Wendy era Houston Community College Biology 1406 Key Concepts in Chapter 12 1. Most cell division results in genetically identical daughter cells. 2. The mitotic phase alternates
Chapter 12 The Cell Cycle Dr. Wendy era Houston Community College Biology 1406 Key Concepts in Chapter 12 1. Most cell division results in genetically identical daughter cells. 2. The mitotic phase alternates
BIOLOGY 4/6/2015. Cell Cycle - Mitosis. Outline. Overview: The Key Roles of Cell Division. identical daughter cells. I. Overview II.
 2 Cell Cycle - Mitosis CAMPBELL BIOLOGY TENTH EDITION Reece Urry Cain Wasserman Minorsky Jackson Outline I. Overview II. Mitotic Phase I. Prophase II. III. Telophase IV. Cytokinesis III. Binary fission
2 Cell Cycle - Mitosis CAMPBELL BIOLOGY TENTH EDITION Reece Urry Cain Wasserman Minorsky Jackson Outline I. Overview II. Mitotic Phase I. Prophase II. III. Telophase IV. Cytokinesis III. Binary fission
MBoC ARTICLE. Sushama Sivakumar a,b, John R. Daum a, Aaron R. Tipton a, Susannah Rankin a,b, and Gary J. Gorbsky a,b a
 MBoC ARTICLE The spindle and kinetochore associated (Ska) complex enhances binding of the anaphasepromoting complex/cyclosome (APC/C) to chromosomes and promotes mitotic exit Sushama Sivakumar a,b, John
MBoC ARTICLE The spindle and kinetochore associated (Ska) complex enhances binding of the anaphasepromoting complex/cyclosome (APC/C) to chromosomes and promotes mitotic exit Sushama Sivakumar a,b, John
The Cell Cycle 4/10/12. Chapter 12. Overview: The Key Roles of Cell Division
 LECTURE PREENTATION For CAMPBELL BIOLOGY, NINTH EDITION Jane B. Reece, Lisa A. Urry, Michael L. Cain, teven A. Wasserman, Peter V. Minorsky, Robert B. Jackson Chapter 12 The Cell Cycle Overview: The Key
LECTURE PREENTATION For CAMPBELL BIOLOGY, NINTH EDITION Jane B. Reece, Lisa A. Urry, Michael L. Cain, teven A. Wasserman, Peter V. Minorsky, Robert B. Jackson Chapter 12 The Cell Cycle Overview: The Key
Mitosis and the Cell Cycle
 Mitosis and the Cell Cycle Chapter 12 The Cell Cycle: Cell Growth & Cell Division Where it all began You started as a cell smaller than a period at the end of a sentence Getting from there to here Cell
Mitosis and the Cell Cycle Chapter 12 The Cell Cycle: Cell Growth & Cell Division Where it all began You started as a cell smaller than a period at the end of a sentence Getting from there to here Cell
Supplementary Information for. Shi and King, Chromosome Nondisjunction Yields Tetraploid Rather than Aneuploid Cells in Human Cell Lines.
 Supplementary Information for Shi and King, Chromosome Nondisjunction Yields Tetraploid Rather than Aneuploid Cells in Human Cell Lines Contains Supplementary Methods Supplementary Figures 1-7 Supplementary
Supplementary Information for Shi and King, Chromosome Nondisjunction Yields Tetraploid Rather than Aneuploid Cells in Human Cell Lines Contains Supplementary Methods Supplementary Figures 1-7 Supplementary
The Cell Cycle. Chapter 12. Biology Eighth Edition Neil Campbell and Jane Reece. PowerPoint Lecture Presentations for
 Chapter 12 The Cell Cycle PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions from Joan Sharp 1
Chapter 12 The Cell Cycle PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions from Joan Sharp 1
Cellular Reproduction, Part 2: Meiosis Lecture 10 Fall 2008
 Mitosis & 1 Cellular Reproduction, Part 2: Lecture 10 Fall 2008 Mitosis Form of cell division that leads to identical daughter cells with the full complement of DNA Occurs in somatic cells Cells of body
Mitosis & 1 Cellular Reproduction, Part 2: Lecture 10 Fall 2008 Mitosis Form of cell division that leads to identical daughter cells with the full complement of DNA Occurs in somatic cells Cells of body
Chapter 12. The Cell Cycle
 Chapter 12 The Cell Cycle The Key Roles of Cell Division The ability of organisms to produce more of their own kind is the one characteristic that best distinguishes living things from nonliving things.
Chapter 12 The Cell Cycle The Key Roles of Cell Division The ability of organisms to produce more of their own kind is the one characteristic that best distinguishes living things from nonliving things.
Mad2-independent Spindle Assembly Checkpoint Activation and Controlled Metaphase Anaphase Transition D V
 Molecular Biology of the Cell Vol. 18, 850 863, March 2007 Mad2-independent Spindle Assembly Checkpoint Activation and Controlled Metaphase Anaphase Transition D V in Drosophila S2 Cells Bernardo Orr,*
Molecular Biology of the Cell Vol. 18, 850 863, March 2007 Mad2-independent Spindle Assembly Checkpoint Activation and Controlled Metaphase Anaphase Transition D V in Drosophila S2 Cells Bernardo Orr,*
CH 9: The Cell Cycle Overview. Cellular Organization of the Genetic Material. Distribution of Chromosomes During Eukaryotic Cell Division
 CH 9: The Cell Cycle Overview The ability of organisms to produce more of their own kind best distinguishes living things from nonliving matter The continuity of life is based on the reproduction of cells,
CH 9: The Cell Cycle Overview The ability of organisms to produce more of their own kind best distinguishes living things from nonliving matter The continuity of life is based on the reproduction of cells,
Chapter 2. Mitosis and Meiosis
 Chapter 2. Mitosis and Meiosis Chromosome Theory of Heredity What structures within cells correspond to genes? The development of genetics took a major step forward by accepting the notion that the genes
Chapter 2. Mitosis and Meiosis Chromosome Theory of Heredity What structures within cells correspond to genes? The development of genetics took a major step forward by accepting the notion that the genes
BIOLOGY. Cell Cycle - Mitosis. Outline. Overview: The Key Roles of Cell Division. identical daughter cells. I. Overview II.
 2 Cell Cycle - Mitosis CAMPBELL BIOLOGY TENTH EDITION Reece Urry Cain Wasserman Minorsky Jackson Outline I. Overview II. Mitotic Phase I. Prophase II. III. Telophase IV. Cytokinesis III. Binary fission
2 Cell Cycle - Mitosis CAMPBELL BIOLOGY TENTH EDITION Reece Urry Cain Wasserman Minorsky Jackson Outline I. Overview II. Mitotic Phase I. Prophase II. III. Telophase IV. Cytokinesis III. Binary fission
Biology is the only subject in which multiplication is the same thing as division
 The Cell Cycle Biology is the only subject in which multiplication is the same thing as division Why do cells divide? For reproduction asexual reproduction For growth one-celled organisms from fertilized
The Cell Cycle Biology is the only subject in which multiplication is the same thing as division Why do cells divide? For reproduction asexual reproduction For growth one-celled organisms from fertilized
SUPPLEMENTARY INFORMATION
 DOI: 0.038/ncb33 a b c 0 min 6 min 7 min (fixed) DIC -GFP, CenpF 3 µm Nocodazole Single optical plane -GFP, CenpF Max. intensity projection d µm -GFP, CenpF, -GFP CenpF 3-D rendering e f 0 min 4 min 0
DOI: 0.038/ncb33 a b c 0 min 6 min 7 min (fixed) DIC -GFP, CenpF 3 µm Nocodazole Single optical plane -GFP, CenpF Max. intensity projection d µm -GFP, CenpF, -GFP CenpF 3-D rendering e f 0 min 4 min 0
CELL CYCLE REGULATION AND CANCER. Cellular Reproduction II
 CELL CYCLE REGULATION AND CANCER Cellular Reproduction II THE CELL CYCLE Interphase G1- gap phase 1- cell grows and develops S- DNA synthesis phase- cell replicates each chromosome G2- gap phase 2- cell
CELL CYCLE REGULATION AND CANCER Cellular Reproduction II THE CELL CYCLE Interphase G1- gap phase 1- cell grows and develops S- DNA synthesis phase- cell replicates each chromosome G2- gap phase 2- cell
klp-18 (RNAi) Control. supplementary information. starting strain: AV335 [emb-27(g48); GFP::histone; GFP::tubulin] bleach
![klp-18 (RNAi) Control. supplementary information. starting strain: AV335 [emb-27(g48); GFP::histone; GFP::tubulin] bleach](/thumbs/91/104639484.jpg "klp-18 (RNAi) Control. supplementary information. starting strain: AV335 [emb-27(g48); GFP::histone; GFP::tubulin] bleach") DOI: 10.1038/ncb1891 A. starting strain: AV335 [emb-27(g48); GFP::histone; GFP::tubulin] bleach embryos let hatch overnight transfer to RNAi plates; incubate 5 days at 15 C RNAi food L1 worms adult worms
DOI: 10.1038/ncb1891 A. starting strain: AV335 [emb-27(g48); GFP::histone; GFP::tubulin] bleach embryos let hatch overnight transfer to RNAi plates; incubate 5 days at 15 C RNAi food L1 worms adult worms
Mitosis Flap Book Excludes Prometaphase
 Mitosis Flap Book Excludes Prometaphase TEACHER S INSTRUCTIONS 1) Choose one of the foldables from the choices below. Three Color Choices Black & White Cells without Chromosomes Choose this option if you
Mitosis Flap Book Excludes Prometaphase TEACHER S INSTRUCTIONS 1) Choose one of the foldables from the choices below. Three Color Choices Black & White Cells without Chromosomes Choose this option if you
Cell division functions in 1. reproduction, 2. growth, and 3. repair
 Cell division functions in 1. reproduction, 2. growth, and 3. repair What do you think you are looking at here??? Can something like you or I do this??? Fig. 12.1 How did you start out? How did you grow?
Cell division functions in 1. reproduction, 2. growth, and 3. repair What do you think you are looking at here??? Can something like you or I do this??? Fig. 12.1 How did you start out? How did you grow?
Biology is the only subject in which multiplication is the same thing as division
 Biology is the only subject in which multiplication is the same thing as division The Cell Cycle: Cell Growth, Cell Division 2007-2008 2007-2008 Getting from there to here Going from egg to baby. the original
Biology is the only subject in which multiplication is the same thing as division The Cell Cycle: Cell Growth, Cell Division 2007-2008 2007-2008 Getting from there to here Going from egg to baby. the original
Cell cycle and Apoptosis. Chalermchai Mitrpant
 Cell cycle and Apoptosis 2556 Chalermchai Mitrpant Overview of the cell cycle Outline Regulatory mechanisms controlling cell cycle Progression of the cell cycle Checkpoint of the cell cycle Phases of the
Cell cycle and Apoptosis 2556 Chalermchai Mitrpant Overview of the cell cycle Outline Regulatory mechanisms controlling cell cycle Progression of the cell cycle Checkpoint of the cell cycle Phases of the
MITOSIS AND THE CELL CYCLE PowerPoint Notes
 1 Name: Date: MITOSIS AND THE CELL CYCLE PowerPoint Notes THE FUNCTIONS OF CELL DIVISION 1. Cell division is vital for all. living organisms This is the only process that can create. new cells 2. Cell
1 Name: Date: MITOSIS AND THE CELL CYCLE PowerPoint Notes THE FUNCTIONS OF CELL DIVISION 1. Cell division is vital for all. living organisms This is the only process that can create. new cells 2. Cell
10.2 The Cell Cycle *
 OpenStax-CNX module: m52672 1 10.2 The Cell Cycle * Shannon McDermott Based on The Cell Cycle by OpenStax This work is produced by OpenStax-CNX and licensed under the Creative Commons Attribution License
OpenStax-CNX module: m52672 1 10.2 The Cell Cycle * Shannon McDermott Based on The Cell Cycle by OpenStax This work is produced by OpenStax-CNX and licensed under the Creative Commons Attribution License
APGRU4L1 Chap 12 Extra Reading Cell Cycle and Mitosis
 APGRU4L1 Chap 12 Extra Reading Cell Cycle and Mitosis Dr. Ramesh Biology is the only subject in which multiplication is the same thing as division 2007-2008 The Cell Cycle: Cell Growth, Cell Division 2007-2008
APGRU4L1 Chap 12 Extra Reading Cell Cycle and Mitosis Dr. Ramesh Biology is the only subject in which multiplication is the same thing as division 2007-2008 The Cell Cycle: Cell Growth, Cell Division 2007-2008
Chapter 10 Cell Growth and Division
 Chapter 10 Cell Growth and Division 10 1 Cell Growth 2 Limits to Cell Growth The larger a cell becomes, the more demands the cell places on its DNA. In addition, the cell has more trouble moving enough
Chapter 10 Cell Growth and Division 10 1 Cell Growth 2 Limits to Cell Growth The larger a cell becomes, the more demands the cell places on its DNA. In addition, the cell has more trouble moving enough
Unduplicated. Chromosomes. Telophase
 10-2 Cell Division The Cell Cycle Interphase Mitosis Prophase Cytokinesis G 1 S G 2 Chromatin in Parent Nucleus & Daughter Cells Chromatin Daughter Nuclei Telophase Mitotic Anaphase Metaphase Use what
10-2 Cell Division The Cell Cycle Interphase Mitosis Prophase Cytokinesis G 1 S G 2 Chromatin in Parent Nucleus & Daughter Cells Chromatin Daughter Nuclei Telophase Mitotic Anaphase Metaphase Use what
Chapter 10. Cell Growth and Division
 Chapter 10 Cell Growth and Division Cell Growth A. Limits to Cell Growth 1. Two main reasons why cells divide: a. Demands on DNA as the cell get too large Cell Growth b. Moving nutrients and waste across
Chapter 10 Cell Growth and Division Cell Growth A. Limits to Cell Growth 1. Two main reasons why cells divide: a. Demands on DNA as the cell get too large Cell Growth b. Moving nutrients and waste across
Chapter 12 The Cell Cycle: Cell Growth, Cell Division
 Chapter 12 The Cell Cycle: Cell Growth, Cell Division 2007-2008 Where it all began You started as a cell smaller than a period at the end of a sentence And now look at you How did you get from there to
Chapter 12 The Cell Cycle: Cell Growth, Cell Division 2007-2008 Where it all began You started as a cell smaller than a period at the end of a sentence And now look at you How did you get from there to
Monday, October 6 Put these items into the appropriate category:
 Monday, October 6 Put these items into the appropriate category: Active Transport Facilitated Diffusion Osmosis Simple Diffusion The smell of rotten eggs spreading through the room Requires ATP expenditure
Monday, October 6 Put these items into the appropriate category: Active Transport Facilitated Diffusion Osmosis Simple Diffusion The smell of rotten eggs spreading through the room Requires ATP expenditure
The Cell Cycle. Chapter 12. Biology. Edited by Shawn Lester. Eighth Edition Neil Campbell and Jane Reece. PowerPoint Lecture Presentations for
 Chapter 12 The Cell Cycle Edited by Shawn Lester PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions
Chapter 12 The Cell Cycle Edited by Shawn Lester PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions
10-2 Cell Division. Chromosomes
 Cell Division In eukaryotes, cell division occurs in two major stages. The first stage, division of the cell nucleus, is called mitosis. The second stage, division of the cell cytoplasm, is called cytokinesis.
Cell Division In eukaryotes, cell division occurs in two major stages. The first stage, division of the cell nucleus, is called mitosis. The second stage, division of the cell cytoplasm, is called cytokinesis.
The Cell Cycle. Materials 2 pipe cleaners of one color and 2 of another color Drawing paper
 The Cell Cycle Introduction When the cell has reached its growth potential it will begin to divide. Additionally, if a cell has become damaged or worn out it can be replaced by surrounding cells through
The Cell Cycle Introduction When the cell has reached its growth potential it will begin to divide. Additionally, if a cell has become damaged or worn out it can be replaced by surrounding cells through
Stages of Mitosis. Introduction
 Name: Due: Stages of Mitosis Introduction Mitosis, also called karyokinesis, is division of the nucleus and its chromosomes. It is followed by division of the cytoplasm known as cytokinesis. Both mitosis
Name: Due: Stages of Mitosis Introduction Mitosis, also called karyokinesis, is division of the nucleus and its chromosomes. It is followed by division of the cytoplasm known as cytokinesis. Both mitosis
Cell Cycle/Mitosis -Notes-
 Cell Cycle/Mitosis -Notes- LIMITS TO CELL GROWTH The a cell becomes, the more demands the cell places on DNA. Additionally, the cell has more trouble moving enough and wastes across the cell membrane.
Cell Cycle/Mitosis -Notes- LIMITS TO CELL GROWTH The a cell becomes, the more demands the cell places on DNA. Additionally, the cell has more trouble moving enough and wastes across the cell membrane.
Biology is the only subject in which multiplication is the same thing as division
 Biology is the only subject in which multiplication is the same thing as division 2007-2008 The Cell Cycle: Cell Growth, Cell Division 2007-2008 Where it all began You started as a cell smaller than a
Biology is the only subject in which multiplication is the same thing as division 2007-2008 The Cell Cycle: Cell Growth, Cell Division 2007-2008 Where it all began You started as a cell smaller than a
Biology is the only subject in which multiplication is the same thing as division
 Biology is the only subject in which multiplication is the same thing as division 2007-2008 The Cell Cycle: Cell Growth, Cell Division 2007-2008 Getting from there to here Going from egg to baby. the original
Biology is the only subject in which multiplication is the same thing as division 2007-2008 The Cell Cycle: Cell Growth, Cell Division 2007-2008 Getting from there to here Going from egg to baby. the original
Chapter 12. living /non-living? growth repair renew. Reproduction. Reproduction. living /non-living. fertilized egg (zygote) next chapter
 next chapter") Chapter 12 How cells divide Reproduction living /non-living? growth repair renew based on cell division first mitosis - distributes identical sets of chromosomes cell cycle (life) Cell Division in Bacteria
Chapter 12 How cells divide Reproduction living /non-living? growth repair renew based on cell division first mitosis - distributes identical sets of chromosomes cell cycle (life) Cell Division in Bacteria
UNC-Duke Biology Course for Residents Fall Cell Cycle Effects of Radiation
 UNC-Duke Biology Course for Residents Fall 2018 1 Cell Cycle: Sequence of changes in a cell starting with the moment the cell is created by cell division, continuing through the doubling of the DNA and
UNC-Duke Biology Course for Residents Fall 2018 1 Cell Cycle: Sequence of changes in a cell starting with the moment the cell is created by cell division, continuing through the doubling of the DNA and
Mitosis in vertebrate somatic cells with two spindles: Implications for the metaphase anaphase transition checkpoint and cleavage
 Proc. Natl. Acad. Sci. USA Vol. 94, pp. 5107 5112, May 1997 Cell Biology Mitosis in vertebrate somatic cells with two spindles: Implications for the metaphase anaphase transition checkpoint and cleavage
Proc. Natl. Acad. Sci. USA Vol. 94, pp. 5107 5112, May 1997 Cell Biology Mitosis in vertebrate somatic cells with two spindles: Implications for the metaphase anaphase transition checkpoint and cleavage
Biology is the only subject in which multiplication is the same thing as division
 Biology is the only subject in which multiplication is the same thing as division 2007-2008 The Cell Cycle: Cell Growth, Cell Division 2007-2008 Where it all began You started as a cell smaller than a
Biology is the only subject in which multiplication is the same thing as division 2007-2008 The Cell Cycle: Cell Growth, Cell Division 2007-2008 Where it all began You started as a cell smaller than a
Figure S1. HP1α localizes to centromeres in mitosis and interacts with INCENP. (A&B) HeLa
 HeLa") SUPPLEMENTARY FIGURES Figure S1. HP1α localizes to centromeres in mitosis and interacts with INCENP. (A&B) HeLa tet-on cells that stably express HP1α-CFP, HP1β-CFP, or HP1γ-CFP were monitored with livecell
SUPPLEMENTARY FIGURES Figure S1. HP1α localizes to centromeres in mitosis and interacts with INCENP. (A&B) HeLa tet-on cells that stably express HP1α-CFP, HP1β-CFP, or HP1γ-CFP were monitored with livecell
Cell Division. During interphase, a cell s DNA is in a loose form called. It condenses into tightly coiled structures called chromosomes during.
 Cell Division The is a cell s total DNA. Prokaryotes DNA is found mostly in a single called the and also in small circles called. Eukaryotes have several DNA double helices packaged into. During interphase,
Cell Division The is a cell s total DNA. Prokaryotes DNA is found mostly in a single called the and also in small circles called. Eukaryotes have several DNA double helices packaged into. During interphase,
Cell Division. Learning Objectives: Introduction. Revised Fall 2018
 Revised Fall 2018 Cell Division Learning Objectives: 1. Define cell cycle and the ordered sequence of events in the cell cycle (Interphase and The divisional phase or M phase) 2. Explain the stages in
Revised Fall 2018 Cell Division Learning Objectives: 1. Define cell cycle and the ordered sequence of events in the cell cycle (Interphase and The divisional phase or M phase) 2. Explain the stages in
Cell Cycle and Mitosis
 Cell Cycle and Mitosis Name Period A# THE CELL CYCLE The cell cycle, or cell-division cycle, is the series of events that take place in a eukaryotic cell between its formation and the moment it replicates
Cell Cycle and Mitosis Name Period A# THE CELL CYCLE The cell cycle, or cell-division cycle, is the series of events that take place in a eukaryotic cell between its formation and the moment it replicates
T H E J O U R N A L O F C E L L B I O L O G Y
 T H E J O U R N A L O F C E L L B I O L O G Y Supplemental material Posch et al., http://www.jcb.org/cgi/content/full/jcb.200912046/dc1 Figure S1. Biochemical characterization of the interaction between
T H E J O U R N A L O F C E L L B I O L O G Y Supplemental material Posch et al., http://www.jcb.org/cgi/content/full/jcb.200912046/dc1 Figure S1. Biochemical characterization of the interaction between
U3.2.3: Eukaryotic chromosomes are linear DNA molecules associated with histone proteins. (Oxford Biology Course Companion page 151).
.") Cell Division Study Guide U3.2.3: Eukaryotic chromosomes are linear DNA molecules associated with histone proteins. (Oxford Biology Course Companion page 151). 1. Describe the structure of eukaryotic DNA
Cell Division Study Guide U3.2.3: Eukaryotic chromosomes are linear DNA molecules associated with histone proteins. (Oxford Biology Course Companion page 151). 1. Describe the structure of eukaryotic DNA
10-2 Cell Division mitosis. cytokinesis. Chromosomes chromosomes Slide 1 of 38
 In eukaryotes, cell division occurs in two major stages. The first stage, division of the cell nucleus, is called mitosis. The second stage, division of the cell cytoplasm, is called cytokinesis. Chromosomes
In eukaryotes, cell division occurs in two major stages. The first stage, division of the cell nucleus, is called mitosis. The second stage, division of the cell cytoplasm, is called cytokinesis. Chromosomes
Supplementary Figure 1. Mother centrioles can reduplicate while in the close association
 C1-GFP distance (nm) C1-GFP distance (nm) a arrested HeLa cell expressing C1-GFP and Plk1TD-RFP -3 s 1 2 3 4 5 6 7 8 9 11 12 13 14 16 17 18 19 2 21 22 23 24 26 27 28 29 3 b 9 8 7 6 5 4 3 2 arrested HeLa
C1-GFP distance (nm) C1-GFP distance (nm) a arrested HeLa cell expressing C1-GFP and Plk1TD-RFP -3 s 1 2 3 4 5 6 7 8 9 11 12 13 14 16 17 18 19 2 21 22 23 24 26 27 28 29 3 b 9 8 7 6 5 4 3 2 arrested HeLa
Name: Date: Block: 10-2 Cell Division Worksheet
 10-2 Cell Division Worksheet W hat do you think would happen if a cell were simple to split into two, without any advance preparation? Would each daughter cell have everything it needed to survive? Because
10-2 Cell Division Worksheet W hat do you think would happen if a cell were simple to split into two, without any advance preparation? Would each daughter cell have everything it needed to survive? Because
Cellular Reproduction
 Section 1: Cellular Growth Section 2: Mitosis and Cytokinesis Section 3: Cell Cycle Regulation Click on a lesson name to select. Section 1 Cellular Growth Ratio of Surface Area to Volume Section 1 Cellular
Section 1: Cellular Growth Section 2: Mitosis and Cytokinesis Section 3: Cell Cycle Regulation Click on a lesson name to select. Section 1 Cellular Growth Ratio of Surface Area to Volume Section 1 Cellular
Essential Questions. Why are cells relatively small? What are the primary stages of the cell cycle? What are the stages of interphase?
 Essential Questions Why are cells relatively small? What are the primary stages of the cell cycle? What are the stages of interphase? Cellular Growth Vocabulary Review selective permeability New cell cycle
Essential Questions Why are cells relatively small? What are the primary stages of the cell cycle? What are the stages of interphase? Cellular Growth Vocabulary Review selective permeability New cell cycle
Creating Identical Body Cells
 Creating Identical Body Cells 5.A Students will describe the stages of the cell cycle, including DNA replication and mitosis, and the importance of the cell cycle to the growth of organisms 5.D Students
Creating Identical Body Cells 5.A Students will describe the stages of the cell cycle, including DNA replication and mitosis, and the importance of the cell cycle to the growth of organisms 5.D Students
10-2 Cell Division. Slide 1 of 38. End Show. Copyright Pearson Prentice Hall
 1 of 38 Cell Division In eukaryotes, cell division occurs in two major stages. The first stage, division of the cell nucleus, is called mitosis. The second stage, division of the cell cytoplasm, is called
1 of 38 Cell Division In eukaryotes, cell division occurs in two major stages. The first stage, division of the cell nucleus, is called mitosis. The second stage, division of the cell cytoplasm, is called
meiosis asexual reproduction CHAPTER 9 & 10 The Cell Cycle, Meiosis & Sexual Life Cycles Sexual reproduction mitosis
 meiosis asexual reproduction CHAPTER 9 & 10 The Cell Cycle, Meiosis & Sexual Sexual reproduction Life Cycles mitosis Chromosomes Consists of a long DNA molecule (represents thousands of genes) Also consists
meiosis asexual reproduction CHAPTER 9 & 10 The Cell Cycle, Meiosis & Sexual Sexual reproduction Life Cycles mitosis Chromosomes Consists of a long DNA molecule (represents thousands of genes) Also consists