Identification and characterization of retinoblastoma gene mutations disturbing apoptosis in human breast cancers
|
|
|
- Marjory Dorsey
- 6 years ago
- Views:
Transcription
1 Molecular Cancer This Provisional PDF corresponds to the article as it appeared upon acceptance. Fully formatted PDF and full text (HTML) versions will be made available soon. Identification and characterization of retinoblastoma gene mutations disturbing apoptosis in human breast cancers Molecular Cancer 2010, 9:173 doi: / Elisabet Ognedal Berge Stian Knappskog Stephanie Geisler Vidar Staalsen Marec Pacal Anne-Lise Borresen-Dale Pal Puntervoll Johan Richard Lillehaug Per Eystein Lonning ISSN Article type Research Submission date 12 January 2010 Acceptance date 1 July 2010 Publication date 1 July 2010 Article URL This peer-reviewed article was published immediately upon acceptance. It can be downloaded, printed and distributed freely for any purposes (see copyright notice below). Articles in Molecular Cancer are listed in PubMed and archived at PubMed Central. For information about publishing your research in Molecular Cancer or any BioMed Central journal, go to For information about other BioMed Central publications go to Ognedal Berge et al., licensee BioMed Central Ltd. This is an open access article distributed under the terms of the Creative Commons Attribution License ( which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
2 Identification and characterization of retinoblastoma gene mutations disturbing apoptosis in human breast cancers Elisabet Ognedal Berge 1,2,3, Stian Knappskog 1,2,3, Stephanie Geisler *1, Vidar Staalesen 1,2,3, Marec Pacal 4,5, Anne-Lise Børresen-Dale 6,7, Pål Puntervoll 8, Johan Richard Lillehaug 2, Per Eystein Lønning # 1,3 Address: 1 Section of Oncology, Institute of Medicine, University of Bergen, Norway; 2 Department of Molecular Biology, University of Bergen, Norway; 3 Department of Oncology, Haukeland University Hospital, Bergen, Norway; 4 Toronto Western Research Institute, University Health Network; 5 Department of Laboratory Medicine and Pathobiology, University of Toronto, Ontario, Canada; 6 Department of Genetics, Institute for Cancer Research, Oslo University Hospital Radiumhospitalet, Oslo, Norway; 7 Institute of Clinical Medicine, Faculty of Medicine, University of Oslo, Norway; 8 Computational Biology Unit, Bergen Center for Computational Science, University of Bergen, Norway Elisabet Ognedal Berge - Elisabet.Ognedal@med.uib.no; Stian Knappskog - Stian.Knappskog@med.uib.no; Stephanie Geisler - Stephanie.Geisler@vestreviken.no; Vidar.Staalsen - Vidar.Staalesen@med.uib.no; Pål Puntervoll - Pal.Puntervoll@bccs.uib.no; Marec Pacal - igg.pacal@utoronto.ca; Anne-Lise Børresen-Dale - a.l.borresendale@medisin.uio.no; Johan Richard Lillehaug - Johan.Lillehaug@mbi.uib.no; Per Eystein Lønning # - per.lonning@helse-bergen.no # Corresponding author * Current permanent address: Department of Medicine, Section for Oncology, Asker and Bærum Hospital, 1309 Rud, Norway
3 Abstract Background: The tumor suppressor prb plays a key role regulating cell cycle arrest, and disturbances in the RB1 gene have been reported in different cancer forms. However, the literature reports contradictory findings with respect to a pro - versus anti - apoptotic role of prb, and the consequence of alterations in RB1 to chemotherapy sensitivity remains unclear. This study is part of a project investigating alterations in pivotal genes as predictive factors to chemotherapy sensitivity in breast cancer. Results: Analyzing 73 locally advanced (stage III) breast cancers, we identified two somatic and one germline single nucleotide changes, each leading to amino acid substitution in the prb protein (Leu607Ile, Arg698Trp, and Arg621Cys, respectively). This is the first study reporting point mutations affecting RB1 in breast cancer tissue. In addition, MLPA analysis revealed two large multiexon deletions (exons 13 to 27 and exons 21 to 23) with the exons deletion occurring in the tumor also harboring the Leu607Ile mutation. Interestingly, Leu607Ile and Arg621Cys point mutations both localize to the spacer region of the prb protein, a region previously shown to harbor somatic and germline mutations. Multiple sequence alignment across species indicates the spacer to be evolutionary conserved. All three RB1 point mutations encoded nuclear proteins with impaired ability to induce apoptosis compared to wild-type prb in vitro. Notably, three out of four tumors harboring RB1 mutations displayed primary resistance to treatment with either 5-FU/mitomycin or doxorubicin while only 14 out of 64 tumors without mutations were resistant (p = 0.046).
4 Conclusions: Although rare, our findings suggest RB1 mutations to be of pathological importance potentially affecting sensitivity to mitomycin/anthracycline treatment in breast cancer. Background The retinoblastoma gene (RB1) is a tumor suppressor gene. prb, the protein coded for by the RB1 gene, plays a pivotal role in cell cycle regulation, promoting G1/S arrest and growth restriction through inhibition of the E2F transcription factors [1]. Germline mutations affecting the RB1 gene are strongly associated with retinoblastoma development in children, and recent evidence has revealed an increased risk of different malignancies, including breast cancers, among patients cured from hereditary retinoblastoma [2]. Somatic alterations of the RB1 gene have been detected in different malignancies [3-5]. Previous studies have reported allelic imbalance (AI), loss of prb protein expression [3], hypermethylation of the RB1 promoter [6] and, in some rare cases, large intragenic deletions [7] in the RB1 gene in primary breast cancer. However, point mutations (1163T>C and 1544C>T) have, so far, only been detected in a single breast cancer cell line (BT20) [8]. To the best of our knowledge, no point mutations have previously been reported in biopsies from breast carcinomas. While the cellular functions of prb are well characterized, the effect of disturbances in the RB1 gene on tumor growth and response to systemic therapy in breast cancer is incompletely understood. Lack of prb protein and loss of heterozygosity (LOH) at the RB1 locus have been related to triple negative (TNBC) or basal cell-like breast cancer [9,10]. Absence of prb expression has been linked to poor prognosis in breast cancer patients receiving adjuvant
5 endocrine therapy [11,12]. In contrast, loss of expression has been associated with good prognosis in patients receiving chemotherapy [10,12]. However, these findings may not be interpreted as direct evidence that alterations in RB1 predict chemosensitivity [13]. Breast cancer patients are selected for systemic treatment options based on tumor characteristics like histological grading, estrogen receptor expression, and Her-2 status, thus, the patient cohorts referred to above may differ with respect to key biological parameters. Experimental studies have provided contradictory results, revealing loss of prb function to enhance [11,14-17] as well as to reduce [18,19] cell death and sensitivity to chemotherapeutic agents. In the current study, we analyzed 73 breast cancers undergoing pre-surgical treatment with doxorubicin or mitomycin with 5-FU for genetic and epigenetic changes in the RB1 gene. We report for the first time point mutations affecting RB1 in breast cancer tissue. Each mutation lead to amino acid substitution (Leu607Ile, Arg698Trp, and Arg621Cys) in prb. The mutated prb variants were all located to the nuclear compartment and expressed reduced apoptotic capacity compared to wild-type prb. Furthermore, MLPA unveiled two large multiexon deletions (exons 13 to 27 and exons 21 to 23). Most interesting, three out of four tumors harboring RB1 mutations expressed resistance to chemotherapy. Our data provide the first indication that RB1 might be a candidate gene involved in drug resistance. Results Sequencing the RB1 coding exons cdna generated from 73 locally advanced breast cancer samples obtained prior to chemotherapy was analyzed by PCR and DNA sequencing for RB1 mutations. Three tumors were found to harbor a single nucleotide change each, all resulting in amino acid substitutions (Table 1). Each mutation was located within the pocket domain of prb (Figure 1). Two of the
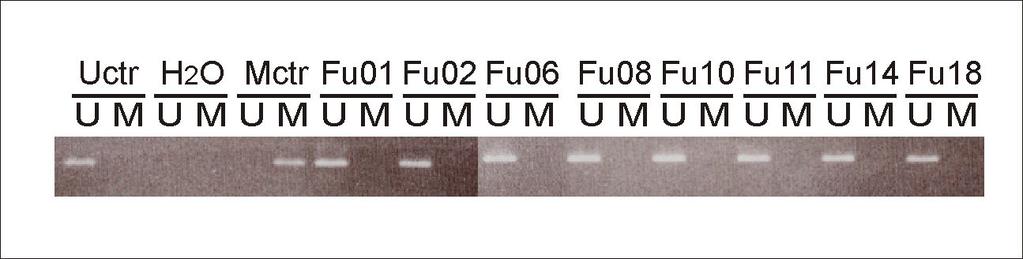
6 mutations were located in exon 19: C1819A (Leu607Ile) and C1861T (Arg621Cys), while the third was located in exon 20: A2092T (Arg698Trp) (nucleotide numbering according to GeneBank sequence L11910 with the A of ATG = number one). Each mutation was verified by amplification and sequencing of genomic DNA from the corresponding tumor. To evaluate whether the mutations were somatic or germline, DNA from corresponding lymphocytes were analyzed. One of these nucleotide changes, C1861T (Arg621Cys), was detected in white blood cells from the affected patient. This patient revealed no family history suggesting hereditary retinoblastoma. Thus, to evaluate whether the Arg621Cys alteration was a common polymorphism, we sequenced DNA from 231 healthy individuals. None of these samples harbored the C1861T nucleotide substitution, arguing against the hypothesis that C1861T could be a polymorphism occurring among >1% of the population (p< 0.10). Promoter analysis In order to explore the occurrence of promoter aberrations as a possible cause for RB1 inactivation, 71 tumors were analyzed for RB1 promoter hypermethylation using methylation specific PCR, and 45 tumors were examined for mutations of the RB1 promoter by sequencing. No hypermethylation (Figure 2) or mutations of the RB1 promoter were detected. Analysing for large exon deletions with use of MLPA Multiplex ligation-dependent probe amplification (MLPA) analysis was performed on tumor DNA from 71 of the samples including the tumors harboring point mutations. For the remaining two samples, genomic DNA was not available. Two individual samples harbored
7 one large multiexon deletion each (exons 13 to 27 and exons 21 to 23), the exons deletion occurring in the tumor also harboring the Leu607Ile point mutation (Table 1). Allelic imbalance MLPA further revealed the RB1 gene to be duplicated in three of the samples (4 %), and 18 of the tumors (25 %) harbored a reduced copy number at the RB1 locus (Table 1). In order to confirm the findings obtained by MLPA, traditional LOH analysis with VNTR/microsatellites was performed for all patients from whom white blood cell DNA was available (n = 43). Allelic imbalance at 13q14 was examined using three markers: D13S263 (located centromeric to RB1), D13S153 (located within intron 2 of RB1), and RB1 (located within intron 20 of RB1). For the informative samples, the findings detected by MLPA were confirmed in all cases (data not shown). RB1 mutations and response to chemotherapy in vivo In this study, all breast cancer tissue samples analyzed (Additional file 1) were obtained from locally advanced primary breast cancers treated in two prospective translational phase III studies [20,21], aiming at identifying markers predicting therapy resistance [22]. All patients included from both protocols are listed in Additional file 1 together with their response to therapy. The total material of 73 tumors included 37 tumors from patients treated with 5-FU and mitomycin in concert (FUMI); eight out of these tumors expressed primary therapy resistance [21]. The remaining 36 patients were selected from a second study exploring resistance to weekly doxorubicin (Dox) among a total of 90 patients [20]. The sub-cohort analyzed here (n=36) contained 9 tumors expressing primary therapy resistance towards
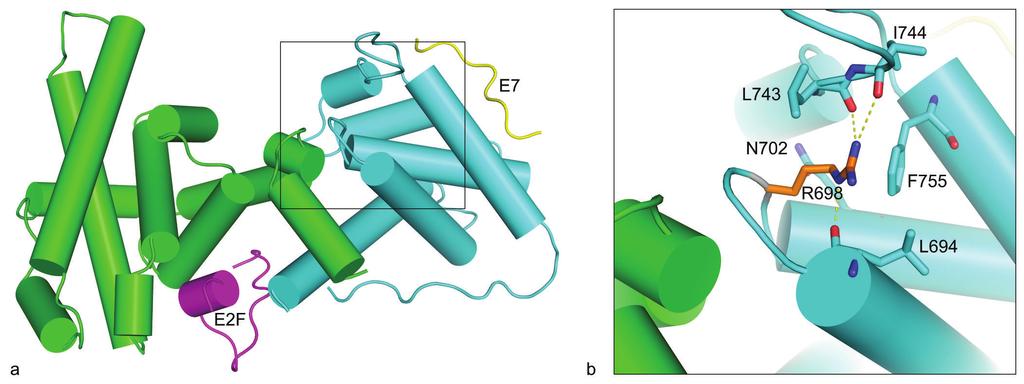
8 doxorubicin together with a random set of 27 patients having an objective response to or stabilization of disease during doxorubicin treatment. Notably, three out of four tumors harboring RB1 mutations (all tumors except the one harboring the Arg698Trp mutation) expressed primary resistance to therapy (Table 1 and Additional file 1). Thus, among 68 tumors analyzed by MLPA and cdna sequencing for which clinical data on response was available, three out of a total of 17 tumors resistant to therapy (PD) harbored RB1 mutations, contrasting only one out of 51 tumors with stable disease or an objective response (p = 0.046). In contrast, no correlation between RB1 allelic imbalance and treatment response was found, and neither mutational status nor AI correlated to overall survival. Multiple sequence alignment of the prb spacer The two point mutations Leu607Ile and Arg621Cys are both located in the spacer region, previously assumed to be non-essential to prb protein function [23]. Employing ClustalX using default parameters [24], a multiple sequence alignment of the prb spacer region including sequences from eight different species was constructed. As shown in Figure 3, the spacer region is fairly well conserved. In fact, the human and mouse RB1-spacer sequences have a higher level of identity than the average human-mouse sequence identity (82 % versus 70 %). This finding indicates that the spacer region is of important for prb function. In silico structural modeling analysis The Arg698Trp mutation is located in the B box of the prb pocket (Figure 4a). In silico structural analysis of the prb pocket [23] revealed the Arg698 residue to form a hydrogen-
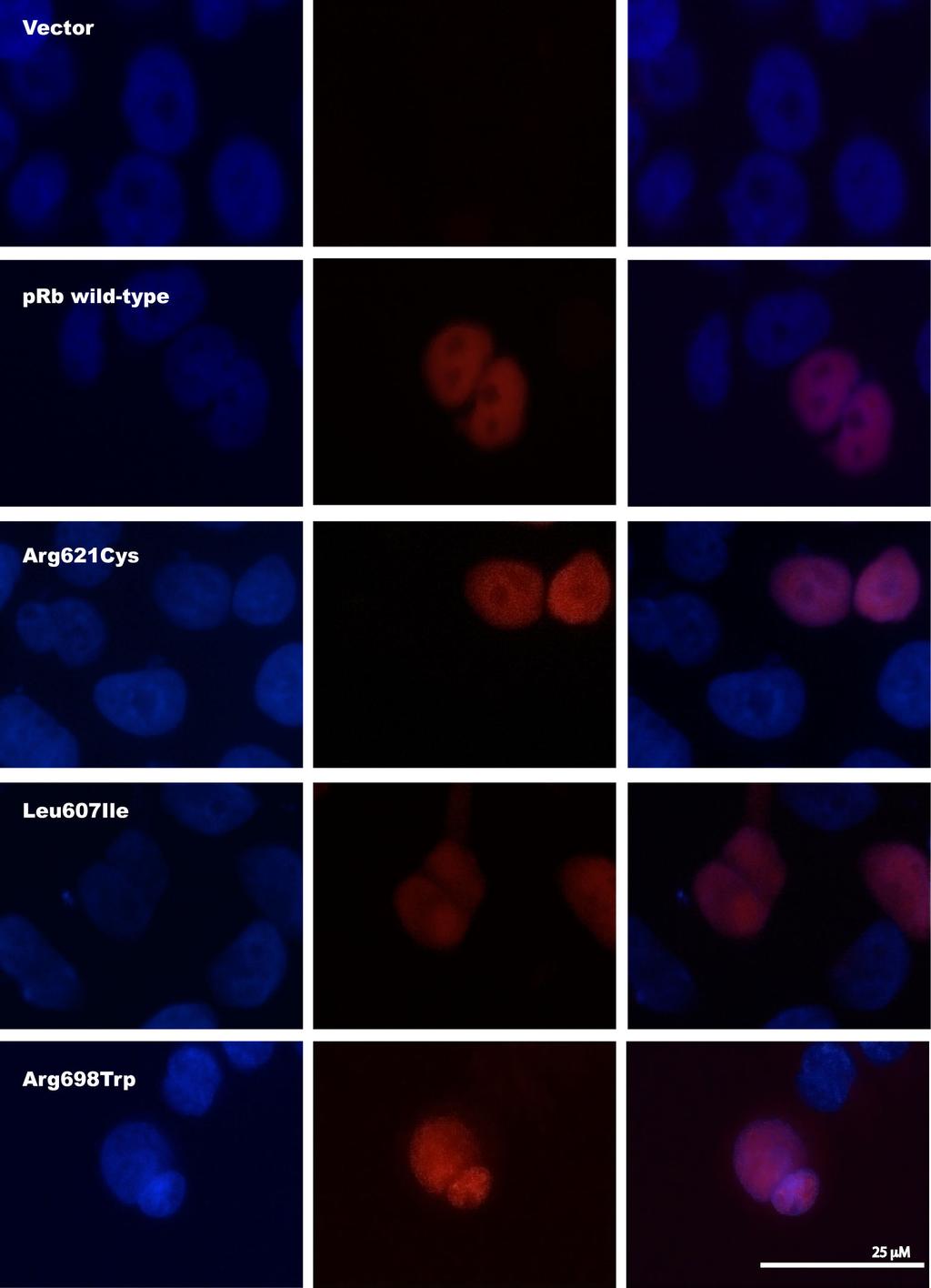
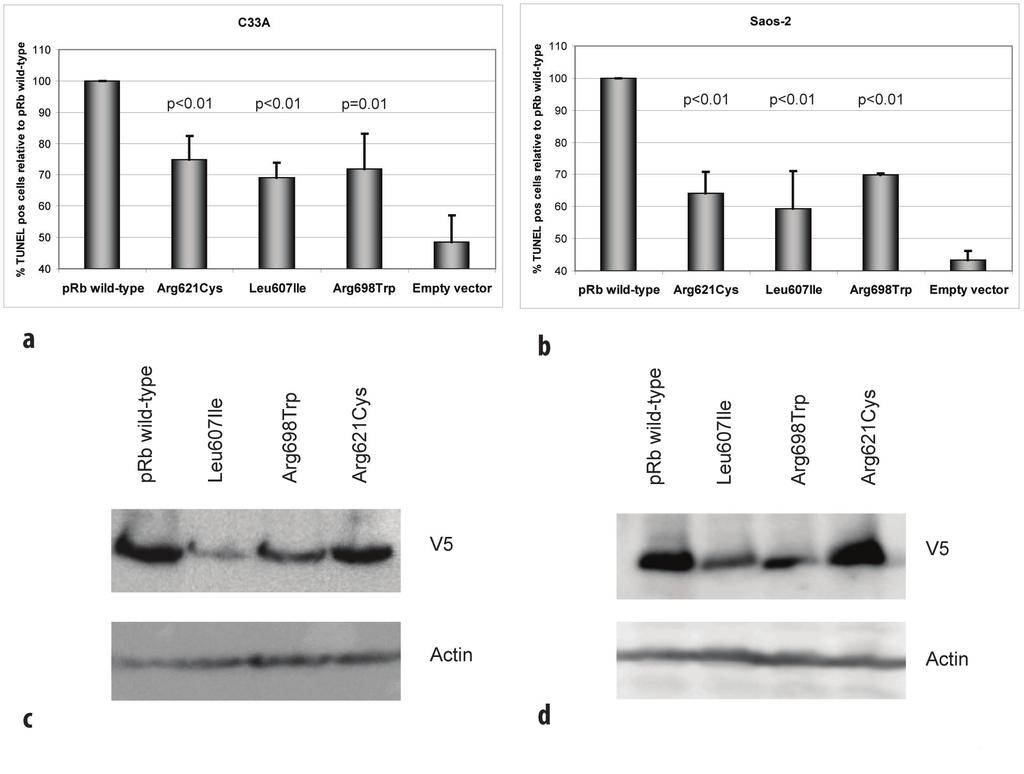
9 bond network (Figure 4b) and predicted Arg698Trp to disrupt this intramolecular hydrogen bond network with a possible structural and functional consequence on the prb protein. Subcellular localization Exploring expression of the mutant proteins in transfected RB1-deficient C-33 A cells, immunostaining revealed positive nuclear staining for all the three prb point mutants (Leu607Ile, Arg698Trp, and Arg621Cys) similar as for prb wild-type (Figure 5). Each control was negative with respect to unspecific fluorescence staining. This indicates that none of the mutants express altered activity due to improper subcellular localization. Apoptotic function and stability of the prb mutant proteins In addition to inhibit cell cycle progression, previous studies of the wild-type prb protein have revealed pro-[19,25,26] as well as anti-[14-17] apoptotic functions in response to among others genotoxic stress like treatment with cytotoxic compounds in vitro. As two of the three point mutations were observed in patients not responding to DNA damaging chemotherapy, we aimed at exploring the ability of the wild-type and mutant prb proteins to mediate apoptosis in RB1-deficient C-33 A cells following treatment with doxorubicin. Analysis by TUNEL assay revealed transfection of wild-type prb to restore apoptosis in response to doxorubicin treatment in C-33 A cells (Figure 6a). While each of the point mutated prb variants expressed some pro-apoptotic function, this was significantly reduced as compared to wild-type protein. Notably, these observations were confirmed in a second RB1-deficient cell line (Saos-2; Figure 6b) revealing the reduced pro-apoptotic effect to occur independent of cell line used (p< 0.01 for all comparisons).
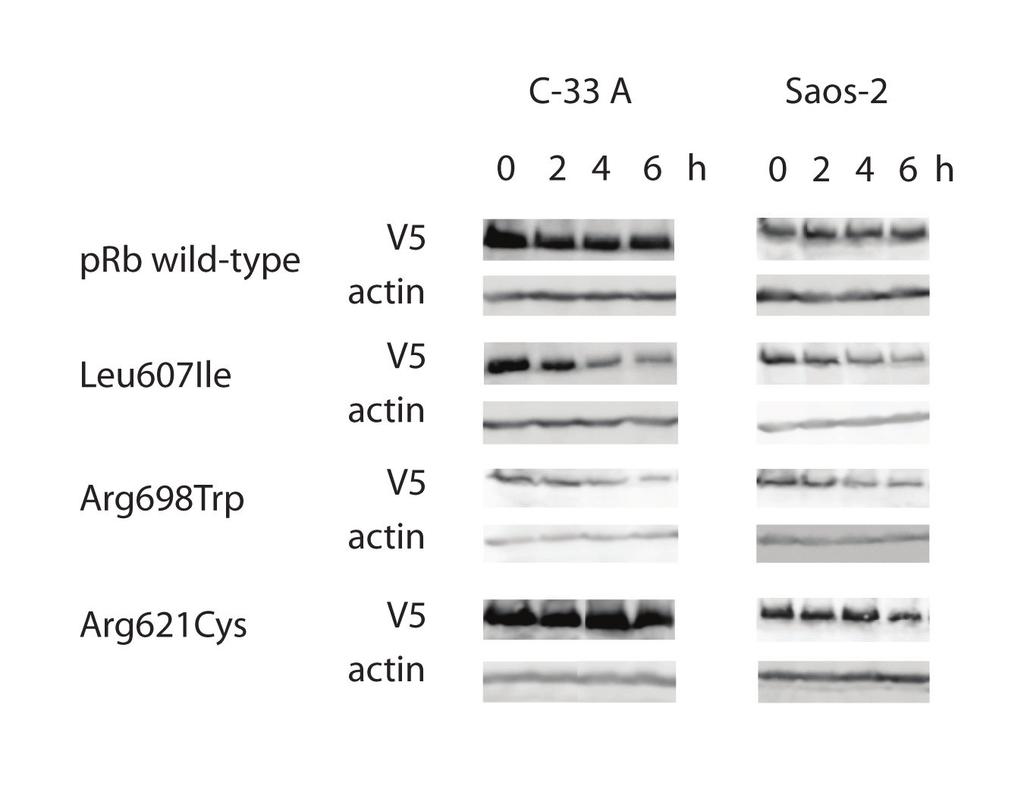
10 Following plasmid transfection with identical amounts of DNA, western blot analyses on samples run in parallel with the TUNEL assays revealed the Leu607Ile and Arg698Trp mutants to reach lower protein amounts as compared to wild-type protein and the Arg621Cys mutant (Figure 6c and 6d). Protein measurement following cycloheximide treatment (Figure 7) revealed both Leu607Ile and Arg698Trp to display reduced stability, thus explaining the mechanism for reduced protein levels. Consistent with the observed apoptotic responses, cell transformation experiments using NIH 3T3 cells revealed a slight, but not significant, increase in foci formation in cells transfected with plasmids carrying the mutant RB1 genes compared to wild-type RB1 (data not shown). Discussion The present work is the first study reporting RB1 point mutations in primary breast carcinomas. Based on our findings that all three point mutated prb proteins expressed reduced pro-apoptotic effect in vitro, and three out of four tumors harboring RB1 mutations were resistant to chemotherapy, our data provide the first indication that RB1 alterations could influence breast cancer chemosensitivity in vivo. Somatic point mutations in RB1 have been detected in different malignancies including bladder [4] and prostate [5] cancer. While previous studies have examined breast cancer samples with respect to RB1 LOH, loss of prb immunostaining [3] and chromosomal rearrangements [7], except for one study reporting no mutations when sequencing the exon 21 only [27] and the finding of point mutations in a single breast cancer cell line [8], we are not
11 aware of any study reporting RB1 point mutations in breast cancer tissue ( In this study, we evaluated potential point mutations through gene sequencing (N=73) and intragenetic deletions by use of MLPA (N=71) across the whole coding region of the RB1 gene in stage III breast cancers. We identified three novel single nucleotide mutations in RB1, each leading to amino acid substitutions. While one of the point mutations was germline, the patient harboring this alteration revealed no family history of retinoblastoma or family clustering of either breast cancer or any other malignancy. Each of the detected point mutations was located in the functionally important pocket domain. The pocket is essential for prb`s interaction with other proteins [28]. It consists of the socalled A and B boxes [23], forming a tight hydrophobic interface, with the two parts being covalently linked by the spacer region (Figure 1). Thus, mutations affecting the A [29] and B [30] boxes of pocket domain are known to give rise to prb proteins with disturbed function. Alterations located in both of these parts have been detected in human sporadic cancers [4,5], but also as germline mutations in patients with retinoblastoma [31,32]. One of the point mutations identified here (Arg698Trp) is located in the B box, and in silico structural analysis suggested this mutation to disrupt a hydrogen-bond network. Furthermore, our findings indicate this mutant to be less stable than the wild-type prb protein (Figure 7). Taken together, these findings suggest that mutations in this location might significantly affect prb function and stability. In contrast, mutations Leu607Ile and Arg621Cys locate to the spacer region. While germline [31-33] as well as somatic [5] mutations within the spacer region have been identified, many authors have considered this domain non-essential to peptide-binding activity of the prb pocket [23]. Our alignment analysis confirmed the spacer domain to be
12 well conserved across different species (Figure 3), and the Leu607Ile mutant protein was considerably less stable compared to prb wild-type These findings, in addition to our results revealing both of these mutations, similar to Arg698Trp, to express a reduced apoptotic function, further indicates a functional role for this region of the prb pocket. Considering the intragenic deletions (exon and exon 13-27), these deletions result in truncated proteins missing half of the B box and both A and B boxes respectively, most likely abolishing the function of the prb pocket in both cases. The retinoblastoma gene encodes a nuclear phosphoprotein which in its unphosphorylated state binds to and inactivates E2F1, causing G1-S phase arrest [1]. Potential additional roles, including pro-apoptotic functions of prb have been suggested [26]. While most studies have indicated an anti-apoptotic role of prb [34], some studies have shown prb to enhance apoptosis following γ-irradation [25] as well as doxorubicin-induced DNA damage [19] which is more in accordance with its role as a tumor suppressor. Here we found the apoptotic response to doxorubicin treatment to be restored by transfecting the RB1-deficient C-33 A and Saos-2 cell lines with wild-type prb, but only to a minor degree when transfecting the prb point mutants, supporting a pro-apoptotic function of prb in response to anthracycline therapy (Figure 6a and 6b). However, the corresponding protein level controls and subsequent stability assays showed that the Leu607Ile and Arg698Trp prb mutants were less stable than the mutant Arg621Cys prb and the prb wild-type proteins which harbored similar protein levels and stabilities. Thus, we believe the relatively low percentage of apoptotic cells observed for all the 3 mutants is due to reduced protein stability in the case of Leu607Ile and Arg698Trp, and due to impaired ability to induce apoptosis in the case of the more stable prb mutant Arg621Cys.
13 Further, in support of a pro-apoptotic role of the prb protein in breast cancer, three out of four tumors harboring RB1 mutations expressed resistance to doxorubicin or mitomycin treatment in vivo. The two drugs have similarities with respect to their mechanisms of antitumor actions. Thus, the in vitro findings on apoptotic response are consistent with our observations on patient drug resistance in vivo. The fact that one of the tumors (Dox 111 harboring Arg698Trp) responded to chemotherapy do not refute such a hypothesis; while we previously reported mutations affecting TP53 [20,21] to be associated with chemoresistance in vivo, the predictive power was not 100%; indicating alternative mechanisms may act in concert [35]. Conclusions In summary, we demonstrate for the first time point mutations in RB1 among breast cancer tumors. Most interestingly, these point mutated genes encode proteins expressing reduced pro-apoptotic effect in vitro, and three out of four tumors harboring RB1 mutations were resistant to chemotherapy. Chemoresistance remains the main obstacle to cure breast cancer as well as most other solid malignancies. Although RB1 mutations were detected in a minority of tumors revealing chemoresistance in vivo (3/17), our present findings with respect to specific RB1 mutations affecting apoptotic response to doxorubicin treatment may point to functional network(s) potentially important for drug resistance in vivo. These findings merit further investigations of factors involved in the RB1 pathway and their role in chemotherapeutic response in breast cancer.
14 Methods Patients This study included patients from two prospective studies addressing the potential role of mutations in TP53 and other genes regarding resistance to treatment with doxorubicin [20] or mitomycin and 5-fluorouracil [21] in locally advanced breast cancer. Both studies were approved by the Regional Ethical Committee, and each patient gave written informed consent to the procedure. Because these studies were designed to explore causes of chemoresistance, we focused on comparing tumors that showed primary drug resistance (progressive disease within 12 weeks) with the combined group of tumors showing stable disease or objective response [22,36]. Thus, for the patients treated with doxorubicin, we analyzed RB1 mutational status in all nine tumors that were resistant to therapy together with a randomly selected subgroup of 27 responding tumors. Regarding the group of patients treated with mitomycin and 5-fluorouracil, we analyzed 37 tumors for the presence of RB1 mutations, including eight patients resistant to therapy [21] and four none evaluable samples. Included were three patients with locally advanced breast cancer, treated with mitomycin and 5-FU, not participating in these studies. Thus, a total of 73 patients were included (Additional file 1). Control subjects Due to the fact that we discovered a novel germline base substitution C1861T (Arg621Cys) in one breast cancer patient, we sought to evaluate its frequency in the general Norwegian population. Thus, we examined blood DNA from 231 healthy women recruited from the national mammographic program into other studies described elsewhere [37]. RNA Purification
15 RNA was purified by Trizol (life Technologies, Inc.) extraction from snap-frozen tissue samples according to the manufacturer s instructions. After extraction, the RNA was dissolved in diethyl pyrocarbonate-treated double-distilled, deionized H 2 O. cdna was synthesized by reverse transcription using Superscript II reverse transcriptase (Invitrogen). DNA Purification Genomic DNA was purified from tumor tissue or lymphocytes using QIAamp DNA Mini Kit (Qiagen) according to the manufacturer s instructions. Multiplex ligand probe amplification (MLPA) MLPA analysis of genomic DNA from 71 patients was performed using the SALSA MLPA RB1 kit (MRC-Holland, Amsterdam, The Netherlands) according to the manufacturer`s instructions. In the patient samples, the peak areas of all MLPA products resulting from RB1 specific probes were first normalized by the average of peak areas resulting from control probes specific for locations other than on chromosomes 13. A ratio was then calculated where this normalized value was divided by the corresponding value from a sample consisting of pooled DNA from 10 healthy individuals. A sample was scored as having a reduced copy number at a specific location if this ratio was below 0.75, and as having an increased copy number if the ratio was above LOH analysis Samples shown by MLPA analysis to harbor AI at the RB1 locus were subsequently analyzed by LOH. Three markers were applied spanning 13q14.1-3: D13S263, centromeric to RB1,
16 D13S153, and RB1, the two latter located within intron 2 and intron 20 of the RB1 gene, respectively. Both D13S263 and D13S153 are microsattelite markers; while RB1 is a variable number tandem repeat (VNTR). All three markers were amplified by PCR using primers as specified in Table 2 and the PCR products were analyzed on an automated DNA sequencer (ABI 3700). Data were analyzed by comparing normal and tumor tissue allele peak-height ratios. A sample was scored as having AI when the ratio between height of tumor and normal sample was less than 0.84 [38]. PCR Amplification of RB1 from cdna Six fragments encompassing the RB1 reading frame starting from nucleotide 89 to 2749 (TGA) were amplified by nested PCR using the primers listed in Table 2. The nucleotides 1 (starting from ATG) to 88 were not covered by our analysis. The frequency of published mutations in this region is approximately 2 % for patients with retinoblastoma ( lsdb.d-lohmann.de/). It was therefore concluded that the risk of missing somatic alterations was acceptably low. Observed mutations were verified by PCR amplification of the affected exons from genomic DNA. In some tumors, no cdna-based products were detected for parts of the RB1 gene. To verify the integrity of the RB1 gene in these patients, we amplified the exons of interest using genomic DNA. PCR was carried out with Dynazyme EXT DNA polymerase (Dynazyme) in a 50 µl solution containing 1X PCR buffer, 1.5 mm MgCl 2, 0.5 mm of each deoxynucleotide tri-phosphate, 5 % DMSO, 0.2 µm of each primer, and 0.5 µl of cdna, 1 µl of first-round PCR-product or ~50 ng genomic DNA was used as template. The RB1 PCR conditions for the first round of the nested PCR for fragments one and two were: An initial 5 min denaturation at 94 o C, 40 cycles of 30 sec at 94 o C, 30 sec at 52.5 o C, and 120 sec at 72 o C, and a final 7 min extension at
17 72 o C. The second round of the nested PCR for fragments one and two was conducted as follows: An initial 5 min denaturation at 94 o C, 40 cycles of 30 sec at 94 o C, 30 sec at 55.6 o C, and 60 sec at 72 o C, and a final 10 min extension at 72 o C. The RB1 PCR conditions for both rounds of the nested PCR for fragments three to six were identical and consisted of: An initial 5 min denaturation at 94 o C, 40 cycles of 30 sec at 94 o C, 30 sec at 45 o C, and 120 sec at 72 o C, and a final 7 min extension at 72 o C. PCR on genomic DNA was done with an initial 5 min denaturation at 94 o C, 40 cycles of 60 sec at 94 o C, 30 sec at an annealing temperature optimized for each exon (Table 2), and 45 sec at 72 o C, and a final 7 min extension at 72 o C. Finally, the RB1 promoter PCR was done with an initial 5 min denaturation at 94 o C, 35 cycles of 30 sec at 94 o C, 30 sec at 53 o C, and 45 sec at 72 o C, and a final 5 min extension at 72 o C. DNA Sequencing Sequencing was performed as described previously [39] using primers specific for the different RB1 fragments (Table 2). The sequences generated were compared with wild-type RB1 (GenBank accession number L11910) for sequence alterations. Promoter Methylation Analysis RB1 promoter methylation status was analyzed by methylation-specific PCR in 71 patients. Genomic DNA from patients was modified using the CpGenome DNA Modification Kit (Intergen), and primers designed specific for methylated (RB1M-S and RB1M-AS, Table 2) and unmethylated (RB1U-S and RB1U-AS, Table 2) DNA [40]. Methylation- and unmethylation-specific PCRs (MSP and USP) were done with AmpliTaq Gold DNA
18 Polymerase (Applied Biosystems) in a 50 µl solution containing 1X PCR buffer, 1.5 mm MgCl 2, 0.5 mm of each deoxynucleotide triphosphate, 0.2 µm of each primer and 25 ng of modified genomic DNA. The methylation-specific PCR was carried out for 35 cycles of 60 sec at 95 o C, 45 sec at 65 o C, and 1 min at 72 o C. The unmethylation-specific PCR was carried out for 35 cycles of 60 sec at 95 o C, 45 sec at 61 o C, and 1 min at 72 o C. Both PCRs were performed with an initial 5 min of denaturation at 95 o C and concluded with 2 min at 72 o C. After amplification, the PCR products were visualized on a 3 % agarose gel. Included in each run was a positive control (CpGenome Universal Methylated DNA, Millipore) and two negative controls (modified DNA from healthy donors and water). Plasmid constructs RB1 wild-type was amplified from cdna by nested PCR using the primers RB1-plasmid-S1 and RB1-plasmid-AS1 in the first, and RB1-plasmid-S and RB-AS-STOP in the second PCR (Table 2). The first PCR was carried out in a 50 µl reaction mixture of 2.5 U KOD XL DNA polymerase (Novagen), 1X PCR buffer, 0.2 mm of each deoxynucleotide triphosphate, 0.2 µm of each primer and 10 µl of cdna. The thermal conditions for the first PCR were as follows: 94 o C for 5 min, 30 cycles of 30 sec at 94 o C, 2 sec at 45 o C and 180 sec at 70 o C and a final extension at 74 o C for 10 min. For the second PCR, a 50 µl reaction solution was made consisting of 0.5 U Dynazyme EXT DNA polymerase, 1X PCR buffer, 5 % DMSO, 0.2 mm of each deoxynucleotide triphosphate, 0.2 µm of each primer and 1 µl of first-round PCR product. The amplification conditions were: 5 min denaturation at 94 o C followed by 40 cycles of 30 sec at 94 o C, 30 sec at 45 o C and 180 sec at 72 o C before a final step at 72 o C for 7 min. The final PCR product was TA-cloned into the expression vector pcdna3.1/v5-his TOPO (Invitrogen) according to the manufacturer s instructions. Using the resulting construct
19 RB1wild-type-V5, corresponding primers (Table 2), and QuickChange Multi Site-Directed Mutagenesis Kit (Stratagene), the following constructs were made; RB1Arg621Cys-V5, RB1Leu607Ile-V5, and RB1Arg698Trp-V5. Cell culture and transfection C-33 A cells and Saos-2 cells were cultured in EMEM and McCoys 5A, respectively, supplemented with 10 % FBS. Both cell lines were purchased from ATCC (American Type Culture Collection). Transfection was performed using Lipofectamine 2000 (Invitrogen) according to the manufacturer`s instruction. Western blot analysis Protein samples were separated on 8% SDS-polyacrylamide gels and blotted onto nitrocellulose membranes (Whatman). The membranes were probed with the following antibodies: anti-v5 (Invitrogen) and anti-actin (Santa Cruz Biotechnology), and the HRPconjugated 2. antibodies sheep anti-mouse (GE Healthcare) and mouse anti-goat (Santa Cruz Biotechnology). Signals were detected using ECL Western blotting Detection Reagents (GE Healthcare). Protein stability Analyses of protein stability using cycloheximide was performed as previously described [41]. After 0, 2, 4, and 6 h, the cells were harvested and lysed in 1 X SDS lysis buffer (0.075 M Tris ph 6.8, 2 % SDS, 20 % glycerol, 0.1 M β-mercaptoethanol, 0.01 % bromphenol blue). The samples were analyzed by SDS-PAGE and western blot using anti-v5 and anti-actin (loading control).
20 Immunofluorescence Cells grown on cover slips were fixed in 3.7% formaldehyde for 20 minutes and washed with 1x PBS. The cells were permeabilized for 12 minutes using 0.1% triton X100 in 1x PBS before blocking with 1% BSA in 1x PBS. Detection of the corresponding proteins was performed using monoclonal mouse anti-v5 and TXR-conjugated goat anti-mouse Ig secondary antibody. The slides were mounted with Vectashield (Vector Laboratories, USA) and examined using a confocal laser scanning microscope (Leica TCS Confocal System attached to a Leica DM RXA microscope). Statistical Analysis Alterations in the RB1 gene were correlated to response to chemotherapy by use of Fisher exact test (p-value given as cumulative, two-sided). Differences between prb wild-type and mutants with respect to induction of apoptosis were evaluated by analyses of variance (ANOVA). Subject to statistical significance, the efficacy of each mutant was compared to prb wild-type with use of the Student test for 2 samples. Statistical calculations were performed using the SPSS 15.0 software and TUNEL assay RB1-deficient C-33 A cells [26] were transfected with pcdna3.1/v5-his-topo vector or one of the four constructs RB1wild-type-V5, RB1Arg621Cys-V5, RB1Leu607Ile-V5, or RB1Arg698Trp-V5. After 24 hours, the cells were treated with 5 µm doxorubicin (adriamycin) for 6 hours, and washed with 1x PBS before cytospins were prepared. The cells were fixed with 3.7 % paraformaldehyde o.n at room temperature and permabilized with 0.1
21 % Triton X-100 in sodium citrate for 2 min on ice. TUNEL reaction mixture (In situ Cell Death Detection Kit, TMR red, Roche) was applied for 2.5 h in dark at 37 o C. Controls were either treated with DNaseI (positive control) or with a TUNEL reaction mixture not containing enzyme. Hoechst was used to visualize the nucleus and the cells were analyzed using Leica DMI6000B epifluorescence microscope (Leica Microsystems). A minimum of 1000 cells for all samples were counted in each of three independent experiments. The same experiment was repeated in Saos-2 cells. Here, the optimal conditions for detection of changes in pro-apoptotic function differed from C-33 A cells. Saos-2 cells were treated with 0.5 µm doxorubicin (adriamycin) for 12 hours before harvest. Abbreviations (AI): Allelic imbalance; (Dox): Doxorubicin; (FUMI): 5-FU/Mitomycin; (LOH): Loss of heterozygosity; (MLPA): Multiplex ligation-dependent probe amplification; (MSP): Methylation-specific PCR; (prb): Retinoblastoma gene product; (RB1): Retinoblastoma tumor suppressor gene; (USP): Unmethylation-specific PCR. Competing interests The authors declare that they have no competing interests. Authors` contributions EOB prepared the manuscript and was responsible for the immunofluoresence experiment. EOB and SK carried out the LOH, MLPA and TUNEL assays and the statistical calculations. SG performed the nucleic acids extraction, sequencing, and methylation experiments
22 supported by VS and EOB. PP was responsible for the multiple sequence alignment and the in silico structural modeling analysis. MP contributed to the design of plasmid constructs, and ALBD was involved in the LOH analysis and helped drafting the manuscript. JRL supervised the laboratory analyses and participated in writing the paper. PEL directed the clinical study and provided the tumor samples. In addition PEL conceived and directed the project, and contributed to the writing of the manuscript. All authors have read and approved the manuscript. Acknowledgements We greatly appreciate the excellent technical assistance of Hilde Johnsen, Beryl Leirvaag, Phoung Vu, Linda Ramsevik, Johanna Huun, Gjertrud T. Iversen, and Christine Eriksen. This work was supported by grants from the Norwegian Cancer Society, Haukeland University Hospital Gene Therapy Program and West Norwegian Health Authorities. Parts of this work were performed at Mohn Cancer Research Laboratory, Haukeland University Hospital. References 1. Weinberg RA: The retinoblastoma protein and cell cycle control. Cell 1995, 81: Marees T, Moll AC, Imhof SM, de Boer MR, Ringens PJ, van Leeuwen FE: Risk of second malignancies in survivors of retinoblastoma: more than 40 years of follow-up. J Natl Cancer Inst 2008, 100: Borg A, Zhang QX, Alm P, Olsson H, Sellberg G: The retinoblastoma gene in breast cancer: allele loss is not correlated with loss of gene protein expression. Cancer Res 1992, 52: Miyamoto H, Shuin T, Torigoe S, Iwasaki Y, Kubota Y: Retinoblastoma gene mutations in primary human bladder cancer. Br J Cancer 1995, 71: Kubota Y, Fujinami K, Uemura H, Dobashi Y, Miyamoto H, Iwasaki Y, Kitamura H, Shuin T: Retinoblastoma gene mutations in primary human prostate cancer. Prostate 1995, 27: Zemliakova VV, Zhevlova AI, Strel'nikov VV, Liubchenko LN, Vishnevskaia Ia V, Tret'iakova VA, Zaletaev DV, Nemtsova MV: [Abnormal methylation of several
23 tumor suppressor genes in sporadic breast cancer]. Mol Biol (Mosk) 2003, 37: T'Ang A, Varley JM, Chakraborty S, Murphree AL, Fung YK: Structural rearrangement of the retinoblastoma gene in human breast carcinoma. Science 1988, 242: Hollestelle A, Nagel JH, Smid M, Lam S, Elstrodt F, Wasielewski M, Ng SS, French PJ, Peeters JK, Rozendaal MJ, Riaz M, Koopman DG, Ten Hagen TL, de Leeuw BH, Zwarthoff EC, Teunisse A, van der Spek PJ, Klijn JG, Dinjens WN, Ethier SP, Clevers H, Jochemsen AG, den Bakker MA, Foekens JA, Martens JW, Schutte M: Distinct gene mutation profiles among luminal-type and basal-type breast cancer cell lines. Breast Cancer Res Treat 2009, DOI /s Herschkowitz JI, He X, Fan C, Perou CM: The functional loss of the retinoblastoma tumour suppressor is a common event in basal-like and luminal B breast carcinomas. Breast Cancer Res 2008, 10:R Trere D, Brighenti E, Donati G, Ceccarelli C, Santini D, Taffurelli M, Montanaro L, Derenzini M: High prevalence of retinoblastoma protein loss in triple-negative breast cancers and its association with a good prognosis in patients treated with adjuvant chemotherapy. Ann Oncol 2009, 20: Bosco EE, Wang Y, Xu H, Zilfou JT, Knudsen KE, Aronow BJ, Lowe SW, Knudsen ES: The retinoblastoma tumor suppressor modifies the therapeutic response of breast cancer. J Clin Invest 2007, 117: Derenzini M, Donati G, Mazzini G, Montanaro L, Vici M, Ceccarelli C, Santini D, Taffurelli M, Trere D: Loss of retinoblastoma tumor suppressor protein makes human breast cancer cells more sensitive to antimetabolite exposure. Clin Cancer Res 2008, 14: Lonning PE, Knappskog S, Staalesen V, Chrisanthar R, Lillehaug JR: Breast cancer prognostication and prediction in the postgenomic era. Ann Oncol 2007, 18: Almasan A, Yin Y, Kelly RE, Lee EY, Bradley A, Li W, Bertino JR, Wahl GM: Deficiency of retinoblastoma protein leads to inappropriate S-phase entry, activation of E2F-responsive genes, and apoptosis. Proc Natl Acad Sci U S A 1995, 92: Knudsen KE, Booth D, Naderi S, Sever-Chroneos Z, Fribourg AF, Hunton IC, Feramisco JR, Wang JY, Knudsen ES: RB-dependent S-phase response to DNA damage. Mol Cell Biol 2000, 20: Stengel KR, Dean JL, Seeley SL, Mayhew CN, Knudsen ES: RB status governs differential sensitivity to cytotoxic and molecularly-targeted therapeutic agents. Cell Cycle 2008, 7: Zagorski WA, Knudsen ES, Reed MF: Retinoblastoma deficiency increases chemosensitivity in lung cancer. Cancer Res 2007, 67: Sharma A, Comstock CE, Knudsen ES, Cao KH, Hess-Wilson JK, Morey LM, Barrera J, Knudsen KE: Retinoblastoma tumor suppressor status is a critical determinant of therapeutic response in prostate cancer cells. Cancer Res 2007, 67: Ianari A, Natale T, Calo E, Ferretti E, Alesse E, Screpanti I, Haigis K, Gulino A, Lees JA: Proapoptotic function of the retinoblastoma tumor suppressor protein. Cancer Cell 2009, 15: Geisler S, Lonning PE, Aas T, Johnsen H, Fluge O, Haugen DF, Lillehaug JR, Akslen LA, Borresen-Dale AL: Influence of TP53 gene alterations and c-erbb-2
24 expression on the response to treatment with doxorubicin in locally advanced breast cancer. Cancer Res 2001, 61: Geisler S, Borresen-Dale AL, Johnsen H, Aas T, Geisler J, Akslen LA, Anker G, Lonning PE: TP53 gene mutations predict the response to neoadjuvant treatment with 5-fluorouracil and mitomycin in locally advanced breast cancer. Clin Cancer Res 2003, 9: Lonning PE: Study of suboptimum treatment response: lessons from breast cancer. Lancet Oncol 2003, 4: Lee JO, Russo AA, Pavletich NP: Structure of the retinoblastoma tumoursuppressor pocket domain bound to a peptide from HPV E7. Nature 1998, 391: Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG: The CLUSTAL_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res 1997, 25: Bowen C, Spiegel S, Gelmann EP: Radiation-induced apoptosis mediated by retinoblastoma protein. Cancer Res 1998, 58: Knudsen KE, Weber E, Arden KC, Cavenee WK, Feramisco JR, Knudsen ES: The retinoblastoma tumor suppressor inhibits cellular proliferation through two distinct mechanisms: inhibition of cell cycle progression and induction of cell death. Oncogene 1999, 18: Hovig E, Smith-Sorensen B, Gebhardt MC, Ryberg D, Lothe R, Borresen AL: No alterations in exon 21 of the RB1 gene in sarcomas and carcinomas of the breast, colon, and lung. Genes Chromosomes Cancer 1992, 5: Morris EJ, Dyson NJ: Retinoblastoma protein partners. Adv Cancer Res 2001, 82: Sellers WR, Novitch BG, Miyake S, Heith A, Otterson GA, Kaye FJ, Lassar AB, Kaelin WG, Jr.: Stable binding to E2F is not required for the retinoblastoma protein to activate transcription, promote differentiation, and suppress tumor cell growth. Genes Dev 1998, 12: Kaye FJ, Kratzke RA, Gerster JL, Horowitz JM: A single amino acid substitution results in a retinoblastoma protein defective in phosphorylation and oncoprotein binding. Proc Natl Acad Sci U S A 1990, 87: Valverde JR, Alonso J, Palacios I, Pestana A: RB1 gene mutation up-date, a metaanalysis based on 932 reported mutations available in a searchable database. BMC Genet 2005, 6: Yu YS, Kim IJ, Ku JL, Park JG: Identification of four novel RB1 germline mutations in Korean retinoblastoma patients. Hum Mutat 2001, 18: Blanquet V, Turleau C, Grossmorand MS, Beaufort CS, Doz F, Besmond C: Spectrum of Germline Mutations in the Rb1 Gene - a Study of 232 Patients with Hereditary and Non Hereditary Retinoblastoma. Human Molecular Genetics 1995, 4: Godefroy N, Lemaire C, Mignotte B, Vayssiere JL: p53 and Retinoblastoma protein (prb): a complex network of interactions. Apoptosis 2006, 11: Lonning PE: Genes causing inherited cancer as beacons to identify the mechanisms of chemoresistance. Trends Mol Med 2004, 10: Aas T, Borresen AL, Geisler S, Smith-Sorensen B, Johnsen H, Varhaug JE, Akslen LA, Lonning PE: Specific P53 mutations are associated with de novo resistance to doxorubicin in breast cancer patients. Nat Med 1996, 2: Gram IT, Bremnes Y, Ursin G, Maskarinec G, Bjurstam N, Lund E: Percentage density, Wolfe's and Tabar's mammographic patterns: agreement and
25 association with risk factors for breast cancer. Breast Cancer Res 2005, 7:R Skotheim RI, Diep CB, Kraggerud SM, Jakobsen KS, Lothe RA: Evaluation of loss of heterozygosity/allelic imbalance scoring in tumor DNA. Cancer Genet Cytogenet 2001, 127: Knappskog S, Chrisanthar R, Staalesen V, Borresen-Dale AL, Gram IT, Lillehaug JR, Lonning PE: Mutations and polymorphisms of the p21b transcript in breast cancer. Int J Cancer 2007, 121: Simpson DJ, Hibberts NA, McNicol AM, Clayton RN, Farrell WE: Loss of prb expression in pituitary adenomas is associated with methylation of the RB1 CpG island. Cancer Res 2000, 60: Berge EO, Staalesen V, Straume AH, Lillehaug JR, Lonning PE: Chk2 splice variants express a dominant-negative effect on the wild-type Chk2 kinase activity. Biochim Biophys Acta 2010, 1803: Figure legends Figure 1 Observed RB1 mutations. (a) Schematic representation of the prb protein. (b) We observed three novel point mutations in RB1: The C1819A mutation (Leu607Ile), the C1861T mutation (Arg621Cys), and the A2092T mutation (Arg698Trp), all leading to amino acid substitutions inside the prb pocket domain. Figure 2 Methylation of the RB1 promoter. RB1 promoter methylation status was analyzed by methylation-specific PCR in 71 patients. Genomic DNA from patients was bisulfite converted. Figure shows products from MSP and USP of a representative selection of patients. Uctr: DNA from healthy donor; Mctr: Universally Methylated Control DNA. Figure 3 Multiple sequence alignment of the prb spacer region. A multiple sequence alignment of the spacer region between the A and B boxes of the prb pocket was generated with ClustalX
26 using default parameters [24]. The locations of two of the mutations reported here are marked with black arrows, and two previously reported mutations are shown in grey [32,33]. Sequences from eight different species were included in the alignment: Human [UNIPROT: P06400 RB_HUMAN], cow [UNIPROT: Q08E68_BOVIN], mouse [UNIPROT: P13405 RB_MOUSE], chicken [UNIPROT: Q90600 RB_CHICK], newt [UNIPROT: Q98966_NOTVI], salmon [UNIPROT: C0H9R0_SALSA], killifish [UNIPROT: Q5J3Q9_FUNHE], and zebrafish [UNIPROT: A0JMQ4_DANRE]. Figure 4 Structural model of the prb pocket domain. (a) The cartoon shows the prb pocket in complex with peptides from the transcription factor E2F (magenta) and the human papilloma virus protein E7 (yellow). The A and B boxes are colored green and cyan, respectively. The model was made from two structures: PDB:1GUX (Rb pocket and E7 peptide) and PDB:1O9K (E2F peptide). (b) Close-up view of the arginine 698 (R698) and amino acids that have at least one atom within a 4Å distance to the side chain atoms of R698. The R698 that is mutated to tryptophan in one of the tumors is located in the B box of the prb pocket, and forms a hydrogen bond network with three backbone carbonyls. Hydrogen bonds to backbone carbonyls of residues L694, L743 and I744 are shown by yellow dotted lines. The structures were visualized using PyMOL ( Figure 5 Subcellular localization of prb mutants. Immunofluorescence staining of RB1-deficient C- 33 A cells transfected with the pcdna3.1/v5-his-topo vector or one of the four plasmids RB1wild-type-V5, RB1Arg621Cys-V5, RB1Leu607Ile-V5, and RB1Arg698Trp-V5. All
27 mutants display nuclear localization. Left: DAPI, Centre: Anti-V5 (Texas Red), Right: Overlay. Figure 6 Mutant prb proteins display reduced apoptotic function. (a) Diagram shows percentage of apoptotic (TUNEL positive) cells following doxorubicin treatment of RB1-deficient C-33 A and (b) Saos-2 cells, transfected with vectors expressing the different prb mutants, relative to prb wild-type (100%). For each mutant, a minimum of 1000 cells were counted in each of three independent experiments. Error bars indicate standard deviations between the three experiments. p-values given are based on analyses of variance (ANOVA) between the individual mutants and prb wild-type. All over, each mutants revealed reduced apoptotic function in both cell lines (p<0.01). (c) Western blot controls for TUNEL assay. Panels displaying expression of prb wild-type and mutants with corresponding actin expression (loading control) from transfections performed in parallel to those used for TUNEL assays in C-33 A and (d) Saos-2. Figure 7 Stability of prb wild-type and mutant proteins. Panels show the protein levels of prb wild-type and mutants including corresponding actin levels as loading control. The samples were harvested 0-6 h after addition of cycloheximide (50 ug/ml) to transfected C-33 A and Saos-2 cells and examined by SDS-PAGE and western blot analyses.
28 Tables Table 1: RB1 alterations observed among patients included in the study Patient 1 Alterations in RB1 Response 4 Large rearrangements 2 Point Mutations 3 Dox19 AI WT PD FUMI 12 AI WT PD Dox 95 Del exon WT PD FUMI 07 Del exon Leu607Ile PD Dox 48 Duplication WT PD Dox 65 WT Arg621Cys (G.l) PD FUMI 39 Duplication WT SD Dox 04 AI WT SD Dox 32 AI WT SD Dox 74 AI WT SD Dox 83 AI WT SD FUMI 25 AI WT SD FUMI 26 AI WT SD FUMI 27 AI WT SD FUMI 44 AI WT SD Dox 15 AI WT PR Dox 109 AI WT PR FUMI 15 AI WT PR FUMI 23 AI WT PR FUMI 29 AI WT PR FUMI 37 AI WT PR Dox 111 AI Arg698Trp PR Dox 39 Duplication WT PR FUMI 30 AI WT NE 1 Dox X, patients treated with doxorubicin; FUMI X, patients treated with FUMI. 2 AI, allelic imbalance; Del, deletion; WT, wild-type. 3 G.l, germline. 4 PD, progressive disease; SD, stable disease; PR, partial response; NE, not evaluable.
29 Table 2: PCR and DNA sequencing primers. Primer name Sense primer Anti sense primer Length (bp) Annealing Temp. ( o C) RB1M 5 -GGG AGT TTC GCG GAC GTG AC-3 5 -ACG TCG AAA CAC GCC CCG RB1U 5 -GGG AGT TTT GTG GAT GTG AT-3 5 -ACA TCA AAA CAC ACC CCA RB1 promoter 5 -CGC CCC AGT TCC CCA CAG A-3 5 -GGC AAC TGA GCG CCG CGT RB1 exon 1 5 -AAC GGG AGT CGG GAG AG-3 5 -AAT CCT GTC ACC ATT CTG C RB1 exon 2 5 -GAT TAT TTT CAT TTG GTA GGC-3 5 -AAA GTG GTA GGA TTA CAG GC RB1 exon 3 5 -TTT TAA CAT AGT ATC CAG TGT GTG-3 5 -TAC ACT TTC ATA ACG GCT CC RB1 exon 4 5 -GAC CCT TCG TTT TCT TAT ATT CTC-3 5 -ATC AGA GTG TAA CCC TAA TAA AAT G RB1 exon 5 5 -ATT GGG AAA ATC TAC TTG AAC-3 5 -TCA AAC TAA CCC TAA CTA TCA AG RB1 exon 6 5 -CAT TCT ATT ATG CAT TTA ACT AAG G-3 5 -CTA ACA GTT AAT AAG CCA AGC AG RB1 exon 7 5 -ATG GAT ATA CTC TAC CCT GCG-3 5 -ATC CTG TCA GCC TTA GAA CC RB1 exon 8 5 -TAA AAG TAG TAG AAT GTT ACC AAG-3 5 -CAG TGA TTC CAG AGT GAG G RB1 exon 9 5 -TTG ACA CCT CTA ACT TAC CCT G-3 5 -TTG GCT AGA TTC TTC TTG GG RB1 exon GAA ATC TGT GCC TCT GTG TG-3 5 -AAA GGT AAC TGT TAT AGG ACA CAC RB1 exon GTT ATC AAT ACC ACC AGG GAG-3 5 -CAA ATC TGA AAC ACT ATA AAG CC RB1 exon AGA CAA GAT GTA TCT GGG TGT AC-3 5 -CAT GAT TTG AAC CCA GTC AG RB1 exon CTT ATT CCC ACA GTG TAT GCC-3 5 -AGC CTG GGT AAC AGA GTG AG RB1 exon ATT CTG ACT ACT TTT ACA TC-3 5 -TTA TGT TAT GGA TAT GGA T RB1 exon ATA TGT GCT TCT TAC CAG T-3 5 -CAC GTT TGA ATG TCT GAG GA RB1 exon CCT CAG ACA TTC AAA CGT GT-3 5 -TTG GTG GAC CCA TTA CAT TA RB1 exon TAA TGT AAT GGG TCC ACC AA-3 5 -TCA AAA TAA TCC CCC TCT CA RB1 exon GAA TGA TGT ATT TAT GCT CA-3 5 -TTC TTT TAT ACT TAC AAT GC RB1 exon CTT TGC CTG ATT TTT GAC AC-3 5 -CAG TGC TGA GAC TCT GGA TTC RB1 exon CAT TTA TGT TTT AGA TGG TTA G-3 5 -GTT TAT TTC GTT TAC ACA AG RB1 exon CAG CCA CTT GCC AAC TTA C-3 5 -CAT AAA CAG AAC CTG GGA AAG RB1-1.r-S2 / AS4 5 -AAC GGG AGT CGG GAG AG-3 5 -GAA TTA CAT TCA CCT CTT CAT CAA G RB1-1.r-S3 / AS2 5 -ATG ATA AAA CTC TTC AGA CTG ATT C-3 5 -TGT CCA CCA AGG TCC TGA G RB1-2.r-frag1-S / AS 5 -AGG AGG ACC CAG AGC AGG AC-3 5 -CCA AGA AAC TTT TAG CAC CAA TG RB1-2.r-frag2-S / AS 5 -CTA CTG AAA TAA ATT CTG CAT TGG T-3 5 -CTC TTC ATC AAG GTT ACT TTT TCG T RB1-2.r-frag3-S / AS 5 -GAA ACA CAG AGA ACA CCA C-3 5 -ATT CTG AGA TGT ACT TCT GCT A RB1-2.r-frag4-S / AS 5 -AGC AAA CTT CTG AAT GAC AAC-3 5 -GAG AGG TAG ATT TCA ATG G RB1-2.r-frag5-S / AS 5 -CTC CAA AGA AAA AAG GTT CAA-3 5 -GGT ATT GGT GAC AAG GTA GG RB1-2.r-frag6-S / AS 5 -GTA TTC TAT AAC TCG GTC TTC A-3 5 -CAT TTC TCT TCC TTG TTT GA RB1-plasmid-S1/ AS1 5 -GGT TTT TCT CAG GGG ACG-3 5 -GTG AGA GAC AAT GAA TCC AGA G RB1-plasmid-S / AS-STOP 5 -CAC AGC TCG CTG GCT CCC-3 5 -TTT CTC TTC CTT GTT TGA G RB1 (Leu607Ile) 5 -GCA GCA GAT ATG TAT ATT TCT CCT GTA AGA TCT CC-3 RB1 (Arg621Cys) 5 -AAA GGT TCA ACT ACG TGT GTA AAT RB1 (Arg698Trp) TCT ACT GC-3 5 -GAA CTC ATG AGA GAC TGG CAT TTG GAC CAA ATT ATG-3 D13S153 F / R 5 -TTG CAC TGT GGA GAT AAA CAC ATA G-3 5 -TCA CAT TGT CTT TTA AGG CAG GAG-3 D13S263 F / R 5 -CCT GGC CTG TTA GTT TTT ATT GTT A-3 5 -CCC AGT CTT GGG TAT GTT TTT A-3 RB1 5 -TGT ATC GGC TAG CCT ATC TC-3 5 -AAT TAA CAA GGT GTG GTG G-3 Additional files Additional file 1 Title: Characteristics of all patients included in the study. Description: Treatment, age, grade, stage of disease, clinical response and outcome of patients included in the study.
30
31 Figure 2

32 Figure 3
33 Figure 4
34 Figure 5
35 Figure 6
36 Figure 7
Identification and characterization of retinoblastoma gene mutations disturbing apoptosis in human breast cancers
 RESEARCH Open Access Identification and characterization of retinoblastoma gene mutations disturbing apoptosis in human breast cancers Elisabet Ognedal Berge 1,2,3, Stian Knappskog 1,2,3, Stephanie Geisler
RESEARCH Open Access Identification and characterization of retinoblastoma gene mutations disturbing apoptosis in human breast cancers Elisabet Ognedal Berge 1,2,3, Stian Knappskog 1,2,3, Stephanie Geisler
Supplementary Appendix
 Supplementary Appendix This appendix has been provided by the authors to give readers additional information about their work. Supplement to: Sherman SI, Wirth LJ, Droz J-P, et al. Motesanib diphosphate
Supplementary Appendix This appendix has been provided by the authors to give readers additional information about their work. Supplement to: Sherman SI, Wirth LJ, Droz J-P, et al. Motesanib diphosphate
a) Primary cultures derived from the pancreas of an 11-week-old Pdx1-Cre; K-MADM-p53
 Primary cultures derived from the pancreas of an 11-week-old Pdx1-Cre; K-MADM-p53") 1 2 3 4 5 6 7 8 9 10 Supplementary Figure 1. Induction of p53 LOH by MADM. a) Primary cultures derived from the pancreas of an 11-week-old Pdx1-Cre; K-MADM-p53 mouse revealed increased p53 KO/KO (green,
1 2 3 4 5 6 7 8 9 10 Supplementary Figure 1. Induction of p53 LOH by MADM. a) Primary cultures derived from the pancreas of an 11-week-old Pdx1-Cre; K-MADM-p53 mouse revealed increased p53 KO/KO (green,
Supplementary Table 3. 3 UTR primer sequences. Primer sequences used to amplify and clone the 3 UTR of each indicated gene are listed.
 Supplemental Figure 1. DLKI-DIO3 mirna/mrna complementarity. Complementarity between the indicated DLK1-DIO3 cluster mirnas and the UTR of SOX2, SOX9, HIF1A, ZEB1, ZEB2, STAT3 and CDH1with mirsvr and PhastCons
Supplemental Figure 1. DLKI-DIO3 mirna/mrna complementarity. Complementarity between the indicated DLK1-DIO3 cluster mirnas and the UTR of SOX2, SOX9, HIF1A, ZEB1, ZEB2, STAT3 and CDH1with mirsvr and PhastCons
c Tuj1(-) apoptotic live 1 DIV 2 DIV 1 DIV 2 DIV Tuj1(+) Tuj1/GFP/DAPI Tuj1 DAPI GFP
 apoptotic live 1 DIV 2 DIV 1 DIV 2 DIV Tuj1(+) Tuj1/GFP/DAPI Tuj1 DAPI GFP") Supplementary Figure 1 Establishment of the gain- and loss-of-function experiments and cell survival assays. a Relative expression of mature mir-484 30 20 10 0 **** **** NCP mir- 484P NCP mir- 484P b Relative
Supplementary Figure 1 Establishment of the gain- and loss-of-function experiments and cell survival assays. a Relative expression of mature mir-484 30 20 10 0 **** **** NCP mir- 484P NCP mir- 484P b Relative
Supplementary Document
 Supplementary Document 1. Supplementary Table legends 2. Supplementary Figure legends 3. Supplementary Tables 4. Supplementary Figures 5. Supplementary References 1. Supplementary Table legends Suppl.
Supplementary Document 1. Supplementary Table legends 2. Supplementary Figure legends 3. Supplementary Tables 4. Supplementary Figures 5. Supplementary References 1. Supplementary Table legends Suppl.
Supplemental Data. Shin et al. Plant Cell. (2012) /tpc YFP N
 /tpc YFP N") MYC YFP N PIF5 YFP C N-TIC TIC Supplemental Data. Shin et al. Plant Cell. ()..5/tpc..95 Supplemental Figure. TIC interacts with MYC in the nucleus. Bimolecular fluorescence complementation assay using
MYC YFP N PIF5 YFP C N-TIC TIC Supplemental Data. Shin et al. Plant Cell. ()..5/tpc..95 Supplemental Figure. TIC interacts with MYC in the nucleus. Bimolecular fluorescence complementation assay using
Figure S1. Analysis of genomic and cdna sequences of the targeted regions in WT-KI and
 Figure S1. Analysis of genomic and sequences of the targeted regions in and indicated mutant KI cells, with WT and corresponding mutant sequences underlined. (A) cells; (B) K21E-KI cells; (C) D33A-KI cells;
Figure S1. Analysis of genomic and sequences of the targeted regions in and indicated mutant KI cells, with WT and corresponding mutant sequences underlined. (A) cells; (B) K21E-KI cells; (C) D33A-KI cells;
Nature Structural & Molecular Biology: doi: /nsmb Supplementary Figure 1
 Supplementary Figure 1 U1 inhibition causes a shift of RNA-seq reads from exons to introns. (a) Evidence for the high purity of 4-shU-labeled RNAs used for RNA-seq. HeLa cells transfected with control
Supplementary Figure 1 U1 inhibition causes a shift of RNA-seq reads from exons to introns. (a) Evidence for the high purity of 4-shU-labeled RNAs used for RNA-seq. HeLa cells transfected with control
Supplementary Figure 1. ROS induces rapid Sod1 nuclear localization in a dosagedependent manner. WT yeast cells (SZy1051) were treated with 4NQO at
 were treated with 4NQO at") Supplementary Figure 1. ROS induces rapid Sod1 nuclear localization in a dosagedependent manner. WT yeast cells (SZy1051) were treated with 4NQO at different concentrations for 30 min and analyzed for
Supplementary Figure 1. ROS induces rapid Sod1 nuclear localization in a dosagedependent manner. WT yeast cells (SZy1051) were treated with 4NQO at different concentrations for 30 min and analyzed for
Supplementary Materials
 Supplementary Materials 1 Supplementary Table 1. List of primers used for quantitative PCR analysis. Gene name Gene symbol Accession IDs Sequence range Product Primer sequences size (bp) β-actin Actb gi
Supplementary Materials 1 Supplementary Table 1. List of primers used for quantitative PCR analysis. Gene name Gene symbol Accession IDs Sequence range Product Primer sequences size (bp) β-actin Actb gi
CD31 5'-AGA GAC GGT CTT GTC GCA GT-3' 5 ' -TAC TGG GCT TCG AGA GCA GT-3'
 Table S1. The primer sets used for real-time RT-PCR analysis. Gene Forward Reverse VEGF PDGFB TGF-β MCP-1 5'-GTT GCA GCA TGA ATC TGA GG-3' 5'-GGA GAC TCT TCG AGG AGC ACT T-3' 5'-GAA TCA GGC ATC GAG AGA
Table S1. The primer sets used for real-time RT-PCR analysis. Gene Forward Reverse VEGF PDGFB TGF-β MCP-1 5'-GTT GCA GCA TGA ATC TGA GG-3' 5'-GGA GAC TCT TCG AGG AGC ACT T-3' 5'-GAA TCA GGC ATC GAG AGA
Table S1. Oligonucleotides used for the in-house RT-PCR assays targeting the M, H7 or N9. Assay (s) Target Name Sequence (5 3 ) Comments
 Target Name Sequence (5 3 ) Comments") SUPPLEMENTAL INFORMATION 2 3 Table S. Oligonucleotides used for the in-house RT-PCR assays targeting the M, H7 or N9 genes. Assay (s) Target Name Sequence (5 3 ) Comments CDC M InfA Forward (NS), CDC M
SUPPLEMENTAL INFORMATION 2 3 Table S. Oligonucleotides used for the in-house RT-PCR assays targeting the M, H7 or N9 genes. Assay (s) Target Name Sequence (5 3 ) Comments CDC M InfA Forward (NS), CDC M
Supplementary Figure 1 a
 Supplementary Figure a Normalized expression/tbp (A.U.).6... Trip-br transcripts Trans Trans Trans b..5. Trip-br Ctrl LPS Normalized expression/tbp (A.U.) c Trip-br transcripts. adipocytes.... Trans Trans
Supplementary Figure a Normalized expression/tbp (A.U.).6... Trip-br transcripts Trans Trans Trans b..5. Trip-br Ctrl LPS Normalized expression/tbp (A.U.) c Trip-br transcripts. adipocytes.... Trans Trans
Abbreviations: P- paraffin-embedded section; C, cryosection; Bio-SA, biotin-streptavidin-conjugated fluorescein amplification.
 Supplementary Table 1. Sequence of primers for real time PCR. Gene Forward primer Reverse primer S25 5 -GTG GTC CAC ACT ACT CTC TGA GTT TC-3 5 - GAC TTT CCG GCA TCC TTC TTC-3 Mafa cds 5 -CTT CAG CAA GGA
Supplementary Table 1. Sequence of primers for real time PCR. Gene Forward primer Reverse primer S25 5 -GTG GTC CAC ACT ACT CTC TGA GTT TC-3 5 - GAC TTT CCG GCA TCC TTC TTC-3 Mafa cds 5 -CTT CAG CAA GGA
Supplementary Figure 1 MicroRNA expression in human synovial fibroblasts from different locations. MicroRNA, which were identified by RNAseq as most
 Supplementary Figure 1 MicroRNA expression in human synovial fibroblasts from different locations. MicroRNA, which were identified by RNAseq as most differentially expressed between human synovial fibroblasts
Supplementary Figure 1 MicroRNA expression in human synovial fibroblasts from different locations. MicroRNA, which were identified by RNAseq as most differentially expressed between human synovial fibroblasts
Toluidin-Staining of mast cells Ear tissue was fixed with Carnoy (60% ethanol, 30% chloroform, 10% acetic acid) overnight at 4 C, afterwards
 overnight at 4 C, afterwards") Toluidin-Staining of mast cells Ear tissue was fixed with Carnoy (60% ethanol, 30% chloroform, 10% acetic acid) overnight at 4 C, afterwards incubated in 100 % ethanol overnight at 4 C and embedded in
Toluidin-Staining of mast cells Ear tissue was fixed with Carnoy (60% ethanol, 30% chloroform, 10% acetic acid) overnight at 4 C, afterwards incubated in 100 % ethanol overnight at 4 C and embedded in
Supplementary Figures
 Supplementary Figures Supplementary Figure 1. H3F3B expression in lung cancer. a. Comparison of H3F3B expression in relapsed and non-relapsed lung cancer patients. b. Prognosis of two groups of lung cancer
Supplementary Figures Supplementary Figure 1. H3F3B expression in lung cancer. a. Comparison of H3F3B expression in relapsed and non-relapsed lung cancer patients. b. Prognosis of two groups of lung cancer
Citation for published version (APA): Oosterveer, M. H. (2009). Control of metabolic flux by nutrient sensors Groningen: s.n.
: Oosterveer, M. H. (2009). Control of metabolic flux by nutrient sensors Groningen: s.n.") University of Groningen Control of metabolic flux by nutrient sensors Oosterveer, Maaike IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite from it.
University of Groningen Control of metabolic flux by nutrient sensors Oosterveer, Maaike IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite from it.
Culture Density (OD600) 0.1. Culture Density (OD600) Culture Density (OD600) Culture Density (OD600) Culture Density (OD600)
 0.1. Culture Density (OD600) Culture Density (OD600) Culture Density (OD600) Culture Density (OD600)") A. B. C. D. E. PA JSRI JSRI 2 PA DSAM DSAM 2 DSAM 3 PA LNAP LNAP 2 LNAP 3 PAO Fcor Fcor 2 Fcor 3 PAO Wtho Wtho 2 Wtho 3 Wtho 4 DTSB Low Iron 2 4 6 8 2 4 6 8 2 22 DTSB Low Iron 2 4 6 8 2 4 6 8 2 22 DTSB
A. B. C. D. E. PA JSRI JSRI 2 PA DSAM DSAM 2 DSAM 3 PA LNAP LNAP 2 LNAP 3 PAO Fcor Fcor 2 Fcor 3 PAO Wtho Wtho 2 Wtho 3 Wtho 4 DTSB Low Iron 2 4 6 8 2 4 6 8 2 22 DTSB Low Iron 2 4 6 8 2 4 6 8 2 22 DTSB
Plasmids Western blot analysis and immunostaining Flow Cytometry Cell surface biotinylation RNA isolation and cdna synthesis
 Plasmids psuper-retro-s100a10 shrna1 was constructed by cloning the dsdna oligo 5 -GAT CCC CGT GGG CTT CCA GAG CTT CTT TCA AGA GAA GAA GCT CTG GAA GCC CAC TTT TTA-3 and 5 -AGC TTA AAA AGT GGG CTT CCA GAG
Plasmids psuper-retro-s100a10 shrna1 was constructed by cloning the dsdna oligo 5 -GAT CCC CGT GGG CTT CCA GAG CTT CTT TCA AGA GAA GAA GCT CTG GAA GCC CAC TTT TTA-3 and 5 -AGC TTA AAA AGT GGG CTT CCA GAG
BIOLOGY 621 Identification of the Snorks
 Name: Date: Block: BIOLOGY 621 Identification of the Snorks INTRODUCTION: In this simulation activity, you will examine the DNA sequence of a fictitious organism - the Snork. Snorks were discovered on
Name: Date: Block: BIOLOGY 621 Identification of the Snorks INTRODUCTION: In this simulation activity, you will examine the DNA sequence of a fictitious organism - the Snork. Snorks were discovered on
SUPPLEMENTARY INFORMATION
 doi: 10.1038/nature05883 SUPPLEMENTARY INFORMATION Supplemental Figure 1 Prostaglandin agonists and antagonists alter runx1/cmyb expression. a-e, Embryos were exposed to (b) PGE2 and (c) PGI2 (20μM) and
doi: 10.1038/nature05883 SUPPLEMENTARY INFORMATION Supplemental Figure 1 Prostaglandin agonists and antagonists alter runx1/cmyb expression. a-e, Embryos were exposed to (b) PGE2 and (c) PGI2 (20μM) and
A smart acid nanosystem for ultrasensitive. live cell mrna imaging by the target-triggered intracellular self-assembly
 Electronic Supplementary Material (ESI) for Chemical Science. This journal is The Royal Society of Chemistry 2017 A smart ZnO@polydopamine-nucleic acid nanosystem for ultrasensitive live cell mrna imaging
Electronic Supplementary Material (ESI) for Chemical Science. This journal is The Royal Society of Chemistry 2017 A smart ZnO@polydopamine-nucleic acid nanosystem for ultrasensitive live cell mrna imaging
Supplementary Table 2. Conserved regulatory elements in the promoters of CD36.
 Supplementary Table 1. RT-qPCR primers for CD3, PPARg and CEBP. Assay Forward Primer Reverse Primer 1A CAT TTG TGG CCT TGT GCT CTT TGA TGA GTC ACA GAA AGA ATC AAT TC 1B AGG AAA TGA ACT GAT GAG TCA CAG
Supplementary Table 1. RT-qPCR primers for CD3, PPARg and CEBP. Assay Forward Primer Reverse Primer 1A CAT TTG TGG CCT TGT GCT CTT TGA TGA GTC ACA GAA AGA ATC AAT TC 1B AGG AAA TGA ACT GAT GAG TCA CAG
Nature Immunology: doi: /ni.3836
 Supplementary Figure 1 Recombinant LIGHT-VTP induces pericyte contractility and endothelial cell activation. (a) Western blot showing purification steps for full length murine LIGHT-VTP (CGKRK) protein:
Supplementary Figure 1 Recombinant LIGHT-VTP induces pericyte contractility and endothelial cell activation. (a) Western blot showing purification steps for full length murine LIGHT-VTP (CGKRK) protein:
Phylogenetic analysis of human and chicken importins. Only five of six importins were studied because
 Supplementary Figure S1 Phylogenetic analysis of human and chicken importins. Only five of six importins were studied because importin-α6 was shown to be testis-specific. Human and chicken importin protein
Supplementary Figure S1 Phylogenetic analysis of human and chicken importins. Only five of six importins were studied because importin-α6 was shown to be testis-specific. Human and chicken importin protein
Supplementary Figure 1a
 Supplementary Figure 1a Hours: E-cadherin TGF-β On TGF-β Off 0 12 24 36 48 24 48 72 Vimentin βactin Fig. S1a. Treatment of AML12 cells with TGF-β induces EMT. Treatment of AML12 cells with TGF-β results
Supplementary Figure 1a Hours: E-cadherin TGF-β On TGF-β Off 0 12 24 36 48 24 48 72 Vimentin βactin Fig. S1a. Treatment of AML12 cells with TGF-β induces EMT. Treatment of AML12 cells with TGF-β results
Beta Thalassemia Case Study Introduction to Bioinformatics
 Beta Thalassemia Case Study Sami Khuri Department of Computer Science San José State University San José, California, USA sami.khuri@sjsu.edu www.cs.sjsu.edu/faculty/khuri Outline v Hemoglobin v Alpha
Beta Thalassemia Case Study Sami Khuri Department of Computer Science San José State University San José, California, USA sami.khuri@sjsu.edu www.cs.sjsu.edu/faculty/khuri Outline v Hemoglobin v Alpha
*To whom correspondence should be addressed. This PDF file includes:
 www.sciencemag.org/cgi/content/full/science.1212182/dc1 Supporting Online Material for Partial Retraction to Detection of an Infectious Retrovirus, XMRV, in Blood Cells of Patients with Chronic Fatigue
www.sciencemag.org/cgi/content/full/science.1212182/dc1 Supporting Online Material for Partial Retraction to Detection of an Infectious Retrovirus, XMRV, in Blood Cells of Patients with Chronic Fatigue
Supplementary Figure 1
 Supplementary Figure 1 Supplementary Figure 1. Lats1/2 deleted ihbs and ihps showed decreased transcripts of hepatocyte related genes (a and b) Western blots (a) and recombination PCR (b) of control and
Supplementary Figure 1 Supplementary Figure 1. Lats1/2 deleted ihbs and ihps showed decreased transcripts of hepatocyte related genes (a and b) Western blots (a) and recombination PCR (b) of control and
SUPPLEMENTARY DATA. Supplementary Table 1. Primer sequences for qrt-pcr
 Supplementary Table 1. Primer sequences for qrt-pcr Gene PRDM16 UCP1 PGC1α Dio2 Elovl3 Cidea Cox8b PPARγ AP2 mttfam CyCs Nampt NRF1 16s-rRNA Hexokinase 2, intron 9 β-actin Primer Sequences 5'-CCA CCA GCG
Supplementary Table 1. Primer sequences for qrt-pcr Gene PRDM16 UCP1 PGC1α Dio2 Elovl3 Cidea Cox8b PPARγ AP2 mttfam CyCs Nampt NRF1 16s-rRNA Hexokinase 2, intron 9 β-actin Primer Sequences 5'-CCA CCA GCG
BHP 2-7 and Nthy-ori 3-1 cells were grown in RPMI1640 medium (Hyclone) supplemented with 10% fetal bovine serum (Gibco), 2mM L-glutamine, and 100 U/mL
 supplemented with 10% fetal bovine serum (Gibco), 2mM L-glutamine, and 100 U/mL") 1 2 3 4 Materials and Methods Cell culture BHP 2-7 and Nthy-ori 3-1 cells were grown in RPMI1640 medium (Hyclone) 5 supplemented with 10% fetal bovine serum (Gibco), 2mM L-glutamine, and 100 U/mL 6 penicillin-streptomycin.
1 2 3 4 Materials and Methods Cell culture BHP 2-7 and Nthy-ori 3-1 cells were grown in RPMI1640 medium (Hyclone) 5 supplemented with 10% fetal bovine serum (Gibco), 2mM L-glutamine, and 100 U/mL 6 penicillin-streptomycin.
Beta Thalassemia Sami Khuri Department of Computer Science San José State University Spring 2015
 Bioinformatics in Medical Product Development SMPD 287 Three Beta Thalassemia Sami Khuri Department of Computer Science San José State University Hemoglobin Outline Anatomy of a gene Hemoglobinopathies
Bioinformatics in Medical Product Development SMPD 287 Three Beta Thalassemia Sami Khuri Department of Computer Science San José State University Hemoglobin Outline Anatomy of a gene Hemoglobinopathies
 www.lessonplansinc.com Topic: Protein Synthesis - Sentence Activity Summary: Students will simulate transcription and translation by building a sentence/polypeptide from words/amino acids. Goals & Objectives:
www.lessonplansinc.com Topic: Protein Synthesis - Sentence Activity Summary: Students will simulate transcription and translation by building a sentence/polypeptide from words/amino acids. Goals & Objectives:
Supplementary Figure 1
 Supplementary Figure 1 3 3 3 1 1 Bregma -1.6mm 3 : Bregma Ref) Http://www.mbl.org/atlas165/atlas165_start.html Bregma -.18mm Supplementary Figure 1 Schematic representation of the utilized brain slice
Supplementary Figure 1 3 3 3 1 1 Bregma -1.6mm 3 : Bregma Ref) Http://www.mbl.org/atlas165/atlas165_start.html Bregma -.18mm Supplementary Figure 1 Schematic representation of the utilized brain slice
SUPPORTING INFORMATION
 SUPPORTING INFORMATION Biology is different in small volumes: endogenous signals shape phenotype of primary hepatocytes cultured in microfluidic channels Amranul Haque, Pantea Gheibi, Yandong Gao, Elena
SUPPORTING INFORMATION Biology is different in small volumes: endogenous signals shape phenotype of primary hepatocytes cultured in microfluidic channels Amranul Haque, Pantea Gheibi, Yandong Gao, Elena
TetR repressor-based bioreporters for the detection of doxycycline using Escherichia
 Supplementary materials TetR repressor-based bioreporters for the detection of doxycycline using Escherichia coli and Acinetobacter oleivorans Hyerim Hong and Woojun Park * Department of Environmental
Supplementary materials TetR repressor-based bioreporters for the detection of doxycycline using Escherichia coli and Acinetobacter oleivorans Hyerim Hong and Woojun Park * Department of Environmental
Description of Supplementary Files. File Name: Supplementary Information Description: Supplementary Figures and Supplementary Tables
 Description of Supplementary Files File Name: Supplementary Information Description: Supplementary Figures and Supplementary Tables Supplementary Figure 1: (A), HCT116 IDH1-WT and IDH1-R132H cells were
Description of Supplementary Files File Name: Supplementary Information Description: Supplementary Figures and Supplementary Tables Supplementary Figure 1: (A), HCT116 IDH1-WT and IDH1-R132H cells were
Supplemental Information. Th17 Lymphocytes Induce Neuronal. Cell Death in a Human ipsc-based. Model of Parkinson's Disease
 Cell Stem Cell, Volume 23 Supplemental Information Th17 Lymphocytes Induce Neuronal Cell Death in a Human ipsc-based Model of Parkinson's Disease Annika Sommer, Franz Maxreiter, Florian Krach, Tanja Fadler,
Cell Stem Cell, Volume 23 Supplemental Information Th17 Lymphocytes Induce Neuronal Cell Death in a Human ipsc-based Model of Parkinson's Disease Annika Sommer, Franz Maxreiter, Florian Krach, Tanja Fadler,
Supplementary Figure 1
 Metastatic melanoma Primary melanoma Healthy human skin Supplementary Figure 1 CD22 IgG4 Supplementary Figure 1: Immunohisochemical analysis of CD22+ (left) and IgG4 (right), cells (shown in red and indicated
Metastatic melanoma Primary melanoma Healthy human skin Supplementary Figure 1 CD22 IgG4 Supplementary Figure 1: Immunohisochemical analysis of CD22+ (left) and IgG4 (right), cells (shown in red and indicated
Supplemental Information. Cancer-Associated Fibroblasts Neutralize. the Anti-tumor Effect of CSF1 Receptor Blockade
 Cancer Cell, Volume 32 Supplemental Information Cancer-Associated Fibroblasts Neutralize the Anti-tumor Effect of CSF1 Receptor Blockade by Inducing PMN-MDSC Infiltration of Tumors Vinit Kumar, Laxminarasimha
Cancer Cell, Volume 32 Supplemental Information Cancer-Associated Fibroblasts Neutralize the Anti-tumor Effect of CSF1 Receptor Blockade by Inducing PMN-MDSC Infiltration of Tumors Vinit Kumar, Laxminarasimha
Supplementary Figure 1
 Supplementary Figure 1 Supplementary Figure 1: Cryopreservation alters CD62L expression by CD4 T cells. Freshly isolated (left) or cryopreserved PBMCs (right) were stained with the mix of antibodies described
Supplementary Figure 1 Supplementary Figure 1: Cryopreservation alters CD62L expression by CD4 T cells. Freshly isolated (left) or cryopreserved PBMCs (right) were stained with the mix of antibodies described
Supplementary Materials and Methods
 DD2 suppresses tumorigenicity of ovarian cancer cells by limiting cancer stem cell population Chunhua Han et al. Supplementary Materials and Methods Analysis of publicly available datasets: To analyze
DD2 suppresses tumorigenicity of ovarian cancer cells by limiting cancer stem cell population Chunhua Han et al. Supplementary Materials and Methods Analysis of publicly available datasets: To analyze
A basic helix loop helix transcription factor controls cell growth
 A basic helix loop helix transcription factor controls cell growth and size in root hairs Keke Yi 1,2, Benoît Menand 1,3, Elizabeth Bell 1, Liam Dolan 1,4 Supplementary note Low soil phosphate availability
A basic helix loop helix transcription factor controls cell growth and size in root hairs Keke Yi 1,2, Benoît Menand 1,3, Elizabeth Bell 1, Liam Dolan 1,4 Supplementary note Low soil phosphate availability
Formylpeptide receptor2 contributes to colon epithelial homeostasis, inflammation, and tumorigenesis
 Supplementary Data Formylpeptide receptor2 contributes to colon epithelial homeostasis, inflammation, and tumorigenesis Keqiang Chen, Mingyong Liu, Ying Liu, Teizo Yoshimura, Wei Shen, Yingying Le, Scott
Supplementary Data Formylpeptide receptor2 contributes to colon epithelial homeostasis, inflammation, and tumorigenesis Keqiang Chen, Mingyong Liu, Ying Liu, Teizo Yoshimura, Wei Shen, Yingying Le, Scott
Expression of Selected Inflammatory Cytokine Genes in Bladder Biopsies
 Borneo Journal of Resource Science and Technology (2013) 3(2): 15-20 Expression of Selected Inflammatory Cytokine Genes in Bladder Biopsies EDMUND UI-HANG SIM *1, NUR DIANA ANUAR 2, TENG-AIK ONG 3, GUAN-
Borneo Journal of Resource Science and Technology (2013) 3(2): 15-20 Expression of Selected Inflammatory Cytokine Genes in Bladder Biopsies EDMUND UI-HANG SIM *1, NUR DIANA ANUAR 2, TENG-AIK ONG 3, GUAN-
Astaxanthin prevents and reverses diet-induced insulin resistance and. steatohepatitis in mice: A comparison with vitamin E
 Supplementary Information Astaxanthin prevents and reverses diet-induced insulin resistance and steatohepatitis in mice: A comparison with vitamin E Yinhua Ni, 1,2 Mayumi Nagashimada, 1 Fen Zhuge, 1 Lili
Supplementary Information Astaxanthin prevents and reverses diet-induced insulin resistance and steatohepatitis in mice: A comparison with vitamin E Yinhua Ni, 1,2 Mayumi Nagashimada, 1 Fen Zhuge, 1 Lili
Cross-talk between mineralocorticoid and angiotensin II signaling for cardiac
 ONLINE SUPPLEMENT TO Crosstalk between mineralocorticoid and angiotensin II signaling for cardiac remodeling An Di ZHANG,,3, Aurelie NGUYEN DINH CAT*,,3, Christelle SOUKASEUM *,,3, Brigitte ESCOUBET, 4,
ONLINE SUPPLEMENT TO Crosstalk between mineralocorticoid and angiotensin II signaling for cardiac remodeling An Di ZHANG,,3, Aurelie NGUYEN DINH CAT*,,3, Christelle SOUKASEUM *,,3, Brigitte ESCOUBET, 4,
Baseline clinical characteristics for the 81 CMML patients Routine diagnostic testing and statistical analyses... 3
 Next-Generation Sequencing Technology Reveals a Characteristic Pattern of Molecular Mutations in 72.8% of Chronic Myelomonocytic Leukemia (CMML) by Detecting Frequent Alterations in TET2, CBL, RAS, and
Next-Generation Sequencing Technology Reveals a Characteristic Pattern of Molecular Mutations in 72.8% of Chronic Myelomonocytic Leukemia (CMML) by Detecting Frequent Alterations in TET2, CBL, RAS, and
Integration Solutions
 Integration Solutions (1) a) With no active glycosyltransferase of either type, an ii individual would not be able to add any sugars to the O form of the lipopolysaccharide. Thus, the only lipopolysaccharide
Integration Solutions (1) a) With no active glycosyltransferase of either type, an ii individual would not be able to add any sugars to the O form of the lipopolysaccharide. Thus, the only lipopolysaccharide
Journal of Cell Science Supplementary information. Arl8b +/- Arl8b -/- Inset B. electron density. genotype
 J. Cell Sci. : doi:.4/jcs.59: Supplementary information E9. A Arl8b /- Arl8b -/- Arl8b Arl8b non-specific band Gapdh Tbp E7.5 HE Inset B D Control al am hf C E Arl8b -/- al am hf E8.5 F low middle high
J. Cell Sci. : doi:.4/jcs.59: Supplementary information E9. A Arl8b /- Arl8b -/- Arl8b Arl8b non-specific band Gapdh Tbp E7.5 HE Inset B D Control al am hf C E Arl8b -/- al am hf E8.5 F low middle high
Nucleotide Sequence of the Australian Bluetongue Virus Serotype 1 RNA Segment 10
 J. gen. Virol. (1988), 69, 945-949. Printed in Great Britain 945 Key words: BTV/genome segment lo/nucleotide sequence Nucleotide Sequence of the Australian Bluetongue Virus Serotype 1 RNA Segment 10 By
J. gen. Virol. (1988), 69, 945-949. Printed in Great Britain 945 Key words: BTV/genome segment lo/nucleotide sequence Nucleotide Sequence of the Australian Bluetongue Virus Serotype 1 RNA Segment 10 By
ice-cold 70% ethanol with gentle vortexing, incubated at -20 C for 4 hours, and washed with PBS.
 Cell cycle analysis For cell cycle analysis, single cell suspensions of E12.5 fetal liver cells were suspended in 4 ml ice-cold 7% ethanol with gentle vortexing, incubated at -2 C for 4 hours, and washed
Cell cycle analysis For cell cycle analysis, single cell suspensions of E12.5 fetal liver cells were suspended in 4 ml ice-cold 7% ethanol with gentle vortexing, incubated at -2 C for 4 hours, and washed
without LOI phenotype by breeding female wild-type C57BL/6J and male H19 +/.
 Sakatani et al. 1 Supporting Online Material Materials and methods Mice and genotyping: H19 mutant mice with C57BL/6J background carrying a deletion in the structural H19 gene (3 kb) and 10 kb of 5 flanking
Sakatani et al. 1 Supporting Online Material Materials and methods Mice and genotyping: H19 mutant mice with C57BL/6J background carrying a deletion in the structural H19 gene (3 kb) and 10 kb of 5 flanking
HCV Persistence and Immune Evasion in the Absence of Memory T Cell Help.
 SOM Text HCV Persistence and Immune Evasion in the Absence of Memory T Cell Help. Arash Grakoui 1, Naglaa H. Shoukry 2, David J. Woollard 2, Jin-Hwan Han 1, Holly L. Hanson 1, John Ghrayeb 3, Krishna K.
SOM Text HCV Persistence and Immune Evasion in the Absence of Memory T Cell Help. Arash Grakoui 1, Naglaa H. Shoukry 2, David J. Woollard 2, Jin-Hwan Han 1, Holly L. Hanson 1, John Ghrayeb 3, Krishna K.
Detection of 549 new HLA alleles in potential stem cell donors from the United States, Poland and Germany
 HLA ISSN 2059-2302 BRIEF COMMUNICATION Detection of 549 new HLA alleles in potential stem cell donors from the United States, Poland and Germany C. J. Hernández-Frederick 1, N. Cereb 2,A.S.Giani 1, J.
HLA ISSN 2059-2302 BRIEF COMMUNICATION Detection of 549 new HLA alleles in potential stem cell donors from the United States, Poland and Germany C. J. Hernández-Frederick 1, N. Cereb 2,A.S.Giani 1, J.
Supporting Information
 Supporting Information Malapeira et al. 10.1073/pnas.1217022110 SI Materials and Methods Plant Material and Growth Conditions. A. thaliana seedlings were stratified at 4 C in the dark for 3 d on Murashige
Supporting Information Malapeira et al. 10.1073/pnas.1217022110 SI Materials and Methods Plant Material and Growth Conditions. A. thaliana seedlings were stratified at 4 C in the dark for 3 d on Murashige
Mutation Screening and Association Studies of the Human UCP 3 Gene in Normoglycemic and NIDDM Morbidly Obese Patients
 Mutation Screening and Association Studies of the Human UCP 3 Gene in Normoglycemic and NIDDM Morbidly Obese Patients Shuichi OTABE, Karine CLEMENT, Séverine DUBOIS, Frederic LEPRETRE, Veronique PELLOUX,
Mutation Screening and Association Studies of the Human UCP 3 Gene in Normoglycemic and NIDDM Morbidly Obese Patients Shuichi OTABE, Karine CLEMENT, Séverine DUBOIS, Frederic LEPRETRE, Veronique PELLOUX,
Cancer Genetics 204 (2011) 45e52
 45e52") Cancer Genetics 204 (2011) 45e52 Exon scanning by reverse transcriptaseepolymerase chain reaction for detection of known and novel EML4eALK fusion variants in nonesmall cell lung cancer Heather R. Sanders
Cancer Genetics 204 (2011) 45e52 Exon scanning by reverse transcriptaseepolymerase chain reaction for detection of known and novel EML4eALK fusion variants in nonesmall cell lung cancer Heather R. Sanders
CIRCRESAHA/2004/098145/R1 - ONLINE 1. Validation by Semi-quantitative Real-Time Reverse Transcription PCR
 CIRCRESAHA/2004/098145/R1 - ONLINE 1 Expanded Materials and Methods Validation by Semi-quantitative Real-Time Reverse Transcription PCR Expression patterns of 13 genes (Online Table 2), selected with respect
CIRCRESAHA/2004/098145/R1 - ONLINE 1 Expanded Materials and Methods Validation by Semi-quantitative Real-Time Reverse Transcription PCR Expression patterns of 13 genes (Online Table 2), selected with respect
Lezione 10. Sommario. Bioinformatica. Lezione 10: Sintesi proteica Synthesis of proteins Central dogma: DNA makes RNA makes proteins Genetic code
 Lezione 10 Bioinformatica Mauro Ceccanti e Alberto Paoluzzi Lezione 10: Sintesi proteica Synthesis of proteins Dip. Informatica e Automazione Università Roma Tre Dip. Medicina Clinica Università La Sapienza
Lezione 10 Bioinformatica Mauro Ceccanti e Alberto Paoluzzi Lezione 10: Sintesi proteica Synthesis of proteins Dip. Informatica e Automazione Università Roma Tre Dip. Medicina Clinica Università La Sapienza
Frequency of mosaicism points towards mutation prone early cleavage cell divisions.
 Frequency of mosaicism points towards mutation prone early cleavage cell divisions. Chad Harland, Wouter Coppieters, Latifa Karim, Carole Charlier, Michel Georges Germ-line de novo mutations Definition:
Frequency of mosaicism points towards mutation prone early cleavage cell divisions. Chad Harland, Wouter Coppieters, Latifa Karim, Carole Charlier, Michel Georges Germ-line de novo mutations Definition:
Supplementary Information. Bamboo shoot fiber prevents obesity in mice by. modulating the gut microbiota
 Supplementary Information Bamboo shoot fiber prevents obesity in mice by modulating the gut microbiota Xiufen Li 1,2, Juan Guo 1, Kailong Ji 1,2, and Ping Zhang 1,* 1 Key Laboratory of Tropical Plant Resources
Supplementary Information Bamboo shoot fiber prevents obesity in mice by modulating the gut microbiota Xiufen Li 1,2, Juan Guo 1, Kailong Ji 1,2, and Ping Zhang 1,* 1 Key Laboratory of Tropical Plant Resources
Relationship of the APOA5/A4/C3/A1 gene cluster and APOB gene polymorphisms with dyslipidemia
 elationship of the APOA5/A4/C3/A1 gene cluster and APOB gene polymorphisms with dyslipidemia H.J. Ou 1, G. Huang 2, W. Liu 3, X.L. Ma 2, Y. Wei 4, T. Zhou 5 and Z.M. Pan 3 1 Department of Neurology, The
elationship of the APOA5/A4/C3/A1 gene cluster and APOB gene polymorphisms with dyslipidemia H.J. Ou 1, G. Huang 2, W. Liu 3, X.L. Ma 2, Y. Wei 4, T. Zhou 5 and Z.M. Pan 3 1 Department of Neurology, The
Single-Molecule Analysis of Gene Expression Using Two-Color RNA- Labeling in Live Yeast
 Supplemental Figures, Tables and Results Single-Molecule Analysis of Gene Expression Using Two-Color RNA- Labeling in Live Yeast Sami Hocine 1, Pascal Raymond 2, Daniel Zenklusen 2, Jeffrey A. Chao 1 &
Supplemental Figures, Tables and Results Single-Molecule Analysis of Gene Expression Using Two-Color RNA- Labeling in Live Yeast Sami Hocine 1, Pascal Raymond 2, Daniel Zenklusen 2, Jeffrey A. Chao 1 &
Loyer, et al. microrna-21 contributes to NASH Suppl 1/15
 Loyer, et al. microrna-21 contributes to NASH Suppl 1/15 SUPPLEMENTARY MATERIAL: Liver MicroRNA-21 is Overexpressed in Non Alcoholic Steatohepatitis and Contributes to the Disease in Experimental Models
Loyer, et al. microrna-21 contributes to NASH Suppl 1/15 SUPPLEMENTARY MATERIAL: Liver MicroRNA-21 is Overexpressed in Non Alcoholic Steatohepatitis and Contributes to the Disease in Experimental Models
Resistance to Tetracycline Antibiotics by Wangrong Yang, Ian F. Moore, Kalinka P. Koteva, Donald W. Hughes, David C. Bareich and Gerard D. Wright.
 Supplementary Data for TetX is a Flavin-Dependent Monooxygenase Conferring Resistance to Tetracycline Antibiotics by Wangrong Yang, Ian F. Moore, Kalinka P. Koteva, Donald W. Hughes, David C. Bareich and
Supplementary Data for TetX is a Flavin-Dependent Monooxygenase Conferring Resistance to Tetracycline Antibiotics by Wangrong Yang, Ian F. Moore, Kalinka P. Koteva, Donald W. Hughes, David C. Bareich and
SALSA MLPA probemix P315-B1 EGFR
 SALSA MLPA probemix P315-B1 EGFR Lot B1-0215 and B1-0112. As compared to the previous A1 version (lot 0208), two mutation-specific probes for the EGFR mutations L858R and T709M as well as one additional
SALSA MLPA probemix P315-B1 EGFR Lot B1-0215 and B1-0112. As compared to the previous A1 version (lot 0208), two mutation-specific probes for the EGFR mutations L858R and T709M as well as one additional
Isolate Sexual Idiomorph Species
 SUPLEMENTARY TABLE 1. Isolate identification, sexual idiomorph and species of each isolate used for MAT locus distribution in Paracoccidioides species. Isolate Sexual Idiomorph Species Pb01 MAT1-1 P. lutzii
SUPLEMENTARY TABLE 1. Isolate identification, sexual idiomorph and species of each isolate used for MAT locus distribution in Paracoccidioides species. Isolate Sexual Idiomorph Species Pb01 MAT1-1 P. lutzii
Supplementary Information
 Supplementary Information Remodeling of heterochromatin structure slows neuropathological progression and prolongs survival in an animal model of Huntington s disease Junghee Lee, Yu Jin Hwang, Yunha Kim,
Supplementary Information Remodeling of heterochromatin structure slows neuropathological progression and prolongs survival in an animal model of Huntington s disease Junghee Lee, Yu Jin Hwang, Yunha Kim,
Deciphering the Clinical Significance of BRCA Variants. A Senior Honors Thesis
 Deciphering the Clinical Significance of BRCA Variants A Senior Honors Thesis Presented in Partial Fulfillment of the Requirements for graduation with research distinction in Biology in the undergraduate
Deciphering the Clinical Significance of BRCA Variants A Senior Honors Thesis Presented in Partial Fulfillment of the Requirements for graduation with research distinction in Biology in the undergraduate
Supplemental Figures: Supplemental Figure 1
 Supplemental Figures: Supplemental Figure 1 Suppl. Figure 1. BM-DC infection with H. pylori does not induce cytotoxicity and treatment of BM-DCs with H. pylori sonicate, but not heat-inactivated bacteria,
Supplemental Figures: Supplemental Figure 1 Suppl. Figure 1. BM-DC infection with H. pylori does not induce cytotoxicity and treatment of BM-DCs with H. pylori sonicate, but not heat-inactivated bacteria,
Recommended laboratory tests to identify influenza A/H5 virus in specimens from patients with an influenza-like illness
 World Health Organization Recommended laboratory tests to identify influenza A/H5 virus in specimens from patients with an influenza-like illness General information Highly pathogenic avian influenza (HPAI)
World Health Organization Recommended laboratory tests to identify influenza A/H5 virus in specimens from patients with an influenza-like illness General information Highly pathogenic avian influenza (HPAI)
Development of RT-qPCR-based molecular diagnostic assays for therapeutic target selection of breast cancer patients
 Development of RT-qPCR-based molecular diagnostic assays for therapeutic target selection of breast cancer patients Sangjung Park The Graduate School Yonsei University Department of Biomedical Laboratory
Development of RT-qPCR-based molecular diagnostic assays for therapeutic target selection of breast cancer patients Sangjung Park The Graduate School Yonsei University Department of Biomedical Laboratory
PATIENTS AND METHODS. Subjects
 PATIENTS AND METHODS Subjects Twenty-nine morbidly obese subjects involved in a gastric surgery program were enrolled in the study between October 25 and March 21. Bariatric surgery was performed in patients
PATIENTS AND METHODS Subjects Twenty-nine morbidly obese subjects involved in a gastric surgery program were enrolled in the study between October 25 and March 21. Bariatric surgery was performed in patients
What do you think of when you here the word genome?
 What do you think of when you here the word genome? What do you think of when you here the word genome? Personal Genomics Outline Review of pre-lab work Genomics and Medicine Case Overview & Assignment
What do you think of when you here the word genome? What do you think of when you here the word genome? Personal Genomics Outline Review of pre-lab work Genomics and Medicine Case Overview & Assignment
SUPPLEMENTARY RESULTS
 SUPPLEMENTARY RESULTS Supplementary Table 1. hfpr1- Flpln-CHO hfpr2-flpln-cho pec 50 E max (%) Log( /K A) Log( /K A) N pec 50 E max (%) Log( /K A) Log( /K A) n ERK1/2 phosphorylation fmlp 9.0±0.6 80±7
SUPPLEMENTARY RESULTS Supplementary Table 1. hfpr1- Flpln-CHO hfpr2-flpln-cho pec 50 E max (%) Log( /K A) Log( /K A) N pec 50 E max (%) Log( /K A) Log( /K A) n ERK1/2 phosphorylation fmlp 9.0±0.6 80±7
Finding protein sites where resistance has evolved
 Finding protein sites where resistance has evolved The amino acid (Ka) and synonymous (Ks) substitution rates Please sit in row K or forward The Berlin patient: first person cured of HIV Contracted HIV
Finding protein sites where resistance has evolved The amino acid (Ka) and synonymous (Ks) substitution rates Please sit in row K or forward The Berlin patient: first person cured of HIV Contracted HIV
University of Groningen. Vasoregression in incipient diabetic retinopathy Pfister, Frederick
 University of Groningen Vasoregression in incipient diabetic retinopathy Pfister, Frederick IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite from
University of Groningen Vasoregression in incipient diabetic retinopathy Pfister, Frederick IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite from
Supplementary information
 Supplementary information Unique polypharmacology nuclear receptor modulator blocks inflammatory signaling pathways Mi Ra Chang 1, Anthony Ciesla 1, Timothy S. Strutzenberg 1, Scott J. Novick 1, Yuanjun
Supplementary information Unique polypharmacology nuclear receptor modulator blocks inflammatory signaling pathways Mi Ra Chang 1, Anthony Ciesla 1, Timothy S. Strutzenberg 1, Scott J. Novick 1, Yuanjun
The Clinical Performance of Primary HPV Screening, Primary HPV Screening Plus Cytology Cotesting, and Cytology Alone at a Tertiary Care Hospital
 The Clinical Performance of Primary HPV Screening, Primary HPV Screening Plus Cytology Cotesting, and Cytology Alone at a Tertiary Care Hospital Jung-Woo Choi MD, PhD; Younghye Kim MD, PhD; Ju-Han Lee
The Clinical Performance of Primary HPV Screening, Primary HPV Screening Plus Cytology Cotesting, and Cytology Alone at a Tertiary Care Hospital Jung-Woo Choi MD, PhD; Younghye Kim MD, PhD; Ju-Han Lee
SUPPLEMENTAL METHODS Cell culture RNA extraction and analysis Immunohistochemical analysis and laser capture microdissection (LCM)
") SUPPLEMENTAL METHODS Cell culture Human peripheral blood mononuclear cells were isolated from healthy donors by Ficoll density gradient centrifugation. Monocyte differentiation to resting macrophages ()
SUPPLEMENTAL METHODS Cell culture Human peripheral blood mononuclear cells were isolated from healthy donors by Ficoll density gradient centrifugation. Monocyte differentiation to resting macrophages ()
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION doi:10.1038/nature10743 Supplementary Figures and Legends Supplementary Figure 1. CYP17A1 (red boxes) lies at the intersection of steroid hormone biosynthetic pathways. CYP17A1
SUPPLEMENTARY INFORMATION doi:10.1038/nature10743 Supplementary Figures and Legends Supplementary Figure 1. CYP17A1 (red boxes) lies at the intersection of steroid hormone biosynthetic pathways. CYP17A1
CANCER. Inherited Cancer Syndromes. Affects 25% of US population. Kills 19% of US population (2nd largest killer after heart disease)
") CANCER Affects 25% of US population Kills 19% of US population (2nd largest killer after heart disease) NOT one disease but 200-300 different defects Etiologic Factors In Cancer: Relative contributions
CANCER Affects 25% of US population Kills 19% of US population (2nd largest killer after heart disease) NOT one disease but 200-300 different defects Etiologic Factors In Cancer: Relative contributions
Genetic characterisation of invasive breast cancer: a comparison of CGH and PCR based multiplex microsatellite analysis
 836 Gerhard-Domagk- Institute of Pathology, University of Münster, 48149 Münster, Germany H Buerger W Boecker Institute of Clinical Chemistry and Laboratory Medicine, University of Münster H Schmidt A
836 Gerhard-Domagk- Institute of Pathology, University of Münster, 48149 Münster, Germany H Buerger W Boecker Institute of Clinical Chemistry and Laboratory Medicine, University of Münster H Schmidt A
Advanced Subsidiary Unit 1: Lifestyle, Transport, Genes and Health
 Write your name here Surname Other names Edexcel GCE Centre Number Candidate Number Biology Advanced Subsidiary Unit 1: Lifestyle, Transport, Genes and Health Thursday 8 January 2009 Morning Time: 1 hour
Write your name here Surname Other names Edexcel GCE Centre Number Candidate Number Biology Advanced Subsidiary Unit 1: Lifestyle, Transport, Genes and Health Thursday 8 January 2009 Morning Time: 1 hour
MRC-Holland MLPA. Description version 06; 23 December 2016
 SALSA MLPA probemix P417-B2 BAP1 Lot B2-1216. As compared to version B1 (lot B1-0215), two reference probes have been added and two target probes have a minor change in length. The BAP1 (BRCA1 associated
SALSA MLPA probemix P417-B2 BAP1 Lot B2-1216. As compared to version B1 (lot B1-0215), two reference probes have been added and two target probes have a minor change in length. The BAP1 (BRCA1 associated
Mechanistic and functional insights into fatty acid activation in Mycobacterium tuberculosis SUPPLEMENTARY INFORMATION
 Mechanistic and functional insights into fatty acid activation in Mycobacterium tuberculosis Pooja Arora 1, Aneesh Goyal 2, Vivek T atarajan 1, Eerappa Rajakumara 2, Priyanka Verma 1, Radhika Gupta 3,
Mechanistic and functional insights into fatty acid activation in Mycobacterium tuberculosis Pooja Arora 1, Aneesh Goyal 2, Vivek T atarajan 1, Eerappa Rajakumara 2, Priyanka Verma 1, Radhika Gupta 3,
Supplementary Materials for
 www.sciencesignaling.org/cgi/content/full/10/473/eaai7696/dc1 Supplementary Materials for Astrocyte-shed extracellular vesicles regulate the peripheral leukocyte response to inflammatory brain lesions
www.sciencesignaling.org/cgi/content/full/10/473/eaai7696/dc1 Supplementary Materials for Astrocyte-shed extracellular vesicles regulate the peripheral leukocyte response to inflammatory brain lesions
McAlpine PERK-GSK3 regulates foam cell formation. Supplemental Material. Supplementary Table I. Sequences of real time PCR primers.
 Mclpine PERK-GSK3 regulates foam cell formation Supplemental Material Supplementary Table I. Sequences of real time PCR primers. Primer Name Primer Sequences (5-3 ) Product Size (bp) GRP78 (human) Fwd:
Mclpine PERK-GSK3 regulates foam cell formation Supplemental Material Supplementary Table I. Sequences of real time PCR primers. Primer Name Primer Sequences (5-3 ) Product Size (bp) GRP78 (human) Fwd:
Enhanced detection and serotyping of Streptococcus pneumoniae using multiplex polymerase chain reaction
 Original article http://dx.doi.org/10.3345/kjp.2012.55.11.424 Korean J Pediatr 2012;55(11):424-429 eissn 1738-1061 pissn 2092-7258 Enhanced detection and serotyping of Streptococcus pneumoniae using multiplex
Original article http://dx.doi.org/10.3345/kjp.2012.55.11.424 Korean J Pediatr 2012;55(11):424-429 eissn 1738-1061 pissn 2092-7258 Enhanced detection and serotyping of Streptococcus pneumoniae using multiplex
Malignant Amelanotic Melanoma of the Pleura without Primary Skin Lesion: An Autopsy Case Report. a a*
 2009 63 6 379 384 Malignant Amelanotic Melanoma of the Pleura without Primary Skin Lesion: An Autopsy Case Report a b a a a* a b 380 63 6 Chest x ray and computed tomography (CT). A, Chest x ray on admission
2009 63 6 379 384 Malignant Amelanotic Melanoma of the Pleura without Primary Skin Lesion: An Autopsy Case Report a b a a a* a b 380 63 6 Chest x ray and computed tomography (CT). A, Chest x ray on admission
Mutation analysis of a Chinese family with oculocutaneous albinism
 /, 2016, Vol. 7, (No. 51), pp: 84981-84988 Mutation analysis of a Chinese family with oculocutaneous albinism Xiong Wang 1, Yaowu Zhu 1, Na Shen 1, Jing Peng 1, Chunyu Wang 1, Haiyi Liu 2, Yanjun Lu 1
/, 2016, Vol. 7, (No. 51), pp: 84981-84988 Mutation analysis of a Chinese family with oculocutaneous albinism Xiong Wang 1, Yaowu Zhu 1, Na Shen 1, Jing Peng 1, Chunyu Wang 1, Haiyi Liu 2, Yanjun Lu 1
CHAPTER 4 RESULTS. showed that all three replicates had similar growth trends (Figure 4.1) (p<0.05; p=0.0000)
 (p<0.05; p=0.0000)") CHAPTER 4 RESULTS 4.1 Growth Characterization of C. vulgaris 4.1.1 Optical Density Growth study of Chlorella vulgaris based on optical density at 620 nm (OD 620 ) showed that all three replicates had similar
CHAPTER 4 RESULTS 4.1 Growth Characterization of C. vulgaris 4.1.1 Optical Density Growth study of Chlorella vulgaris based on optical density at 620 nm (OD 620 ) showed that all three replicates had similar
Molecular Pathology of Ovarian Carcinoma with Morphological Correlation
 Molecular athology of Ovarian Carcinoma with Morphological Correlation Kathleen R. Cho, M.D. Comprehensive Cancer Center and Departments of athology and Internal Medicine University of Michigan Medical
Molecular athology of Ovarian Carcinoma with Morphological Correlation Kathleen R. Cho, M.D. Comprehensive Cancer Center and Departments of athology and Internal Medicine University of Michigan Medical
Nucleotide diversity of the TNF gene region in an African village
 (2001) 2, 343 348 2001 Nature Publishing Group All rights reserved 1466-4879/01 $15.00 www.nature.com/gene Nucleotide diversity of the TNF gene region in an African village A Richardson 1, F Sisay-Joof
(2001) 2, 343 348 2001 Nature Publishing Group All rights reserved 1466-4879/01 $15.00 www.nature.com/gene Nucleotide diversity of the TNF gene region in an African village A Richardson 1, F Sisay-Joof
Supporting Information
 Supporting Information Molecular Recognition Based DNA Nanoassemblies on the Surfaces of Nanosized Exosomes Shuo Wan,, Liqin Zhang,,, Sai Wang, Yuan Liu,, Cuichen Wu, Cheng Cui, Hao Sun, Muling Shi, Ying
Supporting Information Molecular Recognition Based DNA Nanoassemblies on the Surfaces of Nanosized Exosomes Shuo Wan,, Liqin Zhang,,, Sai Wang, Yuan Liu,, Cuichen Wu, Cheng Cui, Hao Sun, Muling Shi, Ying
Supporting Information. Mutational analysis of a phenazine biosynthetic gene cluster in
 Supporting Information for Mutational analysis of a phenazine biosynthetic gene cluster in Streptomyces anulatus 9663 Orwah Saleh 1, Katrin Flinspach 1, Lucia Westrich 1, Andreas Kulik 2, Bertolt Gust
Supporting Information for Mutational analysis of a phenazine biosynthetic gene cluster in Streptomyces anulatus 9663 Orwah Saleh 1, Katrin Flinspach 1, Lucia Westrich 1, Andreas Kulik 2, Bertolt Gust
Isolation and Genetic Characterization of New Reassortant H3N1 Swine Influenza Virus from Pigs in the Midwestern United States
 JOURNAL OF VIROLOGY, May 2006, p. 5092 5096 Vol. 80, No. 10 0022-538X/06/$08.00 0 doi:10.1128/jvi.80.10.5092 5096.2006 Copyright 2006, American Society for Microbiology. All Rights Reserved. Isolation
JOURNAL OF VIROLOGY, May 2006, p. 5092 5096 Vol. 80, No. 10 0022-538X/06/$08.00 0 doi:10.1128/jvi.80.10.5092 5096.2006 Copyright 2006, American Society for Microbiology. All Rights Reserved. Isolation