A Positive Feedback Loop between Mesenchymal-like Cancer Cells and Macrophages Is Essential to Breast Cancer Metastasis
|
|
|
- Quentin Palmer
- 6 years ago
- Views:
Transcription
1 Cancer Cell Article A Positive Feedback Loop between Mesenchymal-like Cancer Cells and Macrophages Is Essential to Breast Cancer Metastasis Shicheng Su, 1,2,7 Qiang Liu, 1,2,7 Jingqi Chen, 1,2,3 Jianing Chen, 1,2 Fei Chen, 1,2 Chonghua He, 1,2 Di Huang, 1,2 Wei Wu, 1,2 Ling Lin, 4 Wei Huang, 1 Jin Zhang, 5 Xiuying Cui, 1 Fang Zheng, 1 Haiyan Li, 1,2 Herui Yao, 6 Fengxi Su, 1,2 and Erwei Song 1,2, * 1 Guangdong Provincial Key Laboratory of Malignant Tumor Epigenetics and Gene Regulation, Medical Research Center, Sun Yat-Sen Memorial Hospital, Sun Yat-Sen University, Guangzhou , China 2 Breast Tumor Center, Sun Yat-Sen Memorial Hospital, Sun Yat-Sen University, Guangzhou , China 3 Department of Medical Oncology, No. 2 Affiliated Hospital, Guangzhou Medical College, Guangzhou , China 4 Department of Internal Medicine, The First Affiliated Hospital, Shantou University Medical College, Shantou , China 5 Department of Breast Surgery, Tianjin Medical University Cancer Institute and Hospital, Tianjin , China 6 Department of Oncology, Sun Yat-Sen Memorial Hospital, Sun Yat-Sen University, Guangzhou , China 7 Co-first author *Correspondence: songew@mail.sysu.edu.cn SUMMARY The close vicinity of cancer cells undergoing epithelial-mesenchymal transition (EMT) and tumor-associated macrophages (TAMs) at the invasive front of tumors suggests that these two cell type may mutually interact. We show that mesenchymal-like breast cancer cells activate macrophages to a TAM-like phenotype by GM- CSF. Reciprocally, CCL18 from TAMs induces cancer cell EMT, forming a positive feedback loop, in coculture systems and humanized mice. Inhibition of GM-CSF or CCL18 breaks this loop and reduces cancer metastasis. High GM-CSF expression in breast cancer samples is associated with more CCL18 + macrophages, cancer cell EMT, enhanced metastasis, and reduced patient survival. These findings suggest that a positive feedback loop between GM-CSF and CCL18 is important in breast cancer metastasis. INTRODUCTION It is well established that tumor microenvironment plays an important role in cancer development and metastasis (Allavena et al., 2008; Condeelis and Pollard, 2006). Tumor-associated macrophages (TAMs), the most abundant immune-related stromal cells in tumor microenvironment (Allavena et al., 2008), are key orchestrators of tumor microenvironment, directly affecting neoplastic cell growth, neoangiogenesis, and extracellular matrix remodeling (Solinas et al., 2010). TAMs of breast cancer often display an alternatively activated phenotype, promoting tumor invasion and metastasis, and are associated with poor prognosis in cancer patients (Chen et al., 2011; DeNardo et al., 2009). In addition, cancer cells can actively modulate nonmalignant stromal cells, including macrophages (Pollard, 2004), in tumor microenvironment to enhance cancer development and metastasis (Joyce and Pollard, 2009). Cancer is a disease of heterogeneity and plasticity; however, it is not yet known whether different kinds of cancer cells have different ability to modulate tumor microenvironment. Epithelial-mesenchymal transition (EMT) is a process whereby cancer cells lose their epithelial properties to acquire a mesenchymal phenotype and become motile and invasive, which is closely associated with metastasis (Thiery, 2002). EMT has also been connected to induction of cancer stem cells (Mani et al., 2008), drug resistance (Thiery et al., 2009), and Significance Cancer cell epithelial-mesenchymal transition (EMT) and tumor-associated macrophages (TAMs) are associated with increased metastasis and are frequently observed at the invasive front of advanced tumors. However, the interaction between cancer cells undergoing EMT and TAMs remains unknown. In this study, mesenchymal-like cancer cells activate macrophages to a TAM-like phenotype via GM-CSF, suggesting that mesenchymal-like cancer cells may be a previously unappreciated source of TAM activators. In turn, TAMs induce EMT of breast cancer cells through CCL18, forming a positive feedback loop. Furthermore, the interaction between cancer cells and macrophages is essential to cancer metastasis in a humanized mouse model of breast cancer, suggesting that the feedback loop between GM-CSF and CCL18 could be a therapeutic target for cancer metastasis. Cancer Cell 25, , May 12, 2014 ª2014 Elsevier Inc. 605
 Macrophages were obtained by culture of monocytes in DMEM medium supplemented with 10% heat-inactivated human")
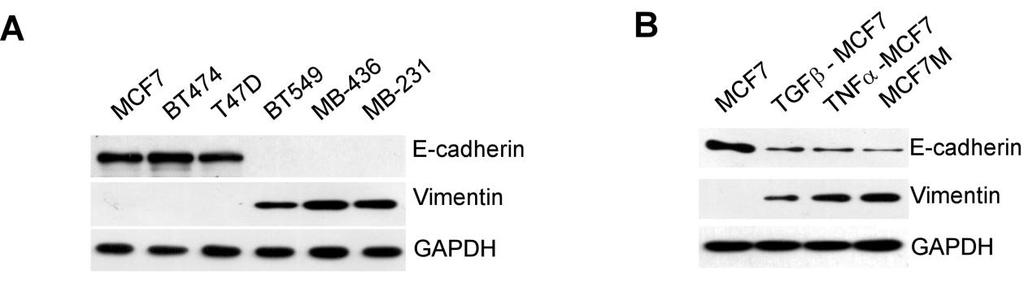
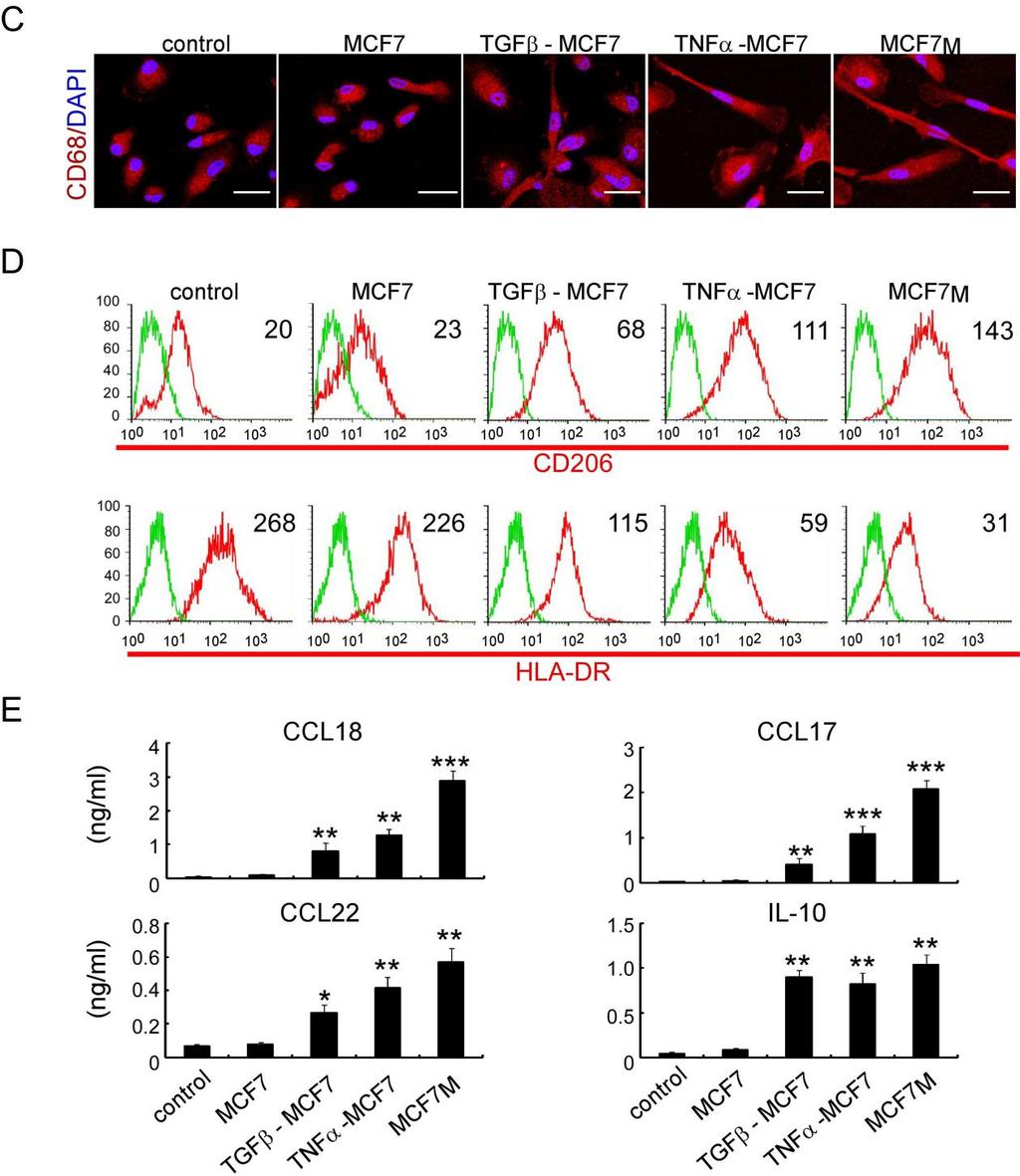
2 Cancer Cell GM-CSF-CCL18 Loop Promotes Breast Cancer Metastasis Figure 1. Mesenchymal-like Breast Cancer Cells Activate Macrophages to a TAM-like Phenotype (A C) Macrophages were obtained by culture of monocytes in DMEM medium supplemented with 10% heat-inactivated human AB serum in the presence or absence of 30% CM from indicated cell lines for 6 days. The primary TAMs were isolated from fresh human breast cancer tissue as positive controls. (A) Fluorescent CD68/DAPI staining in macrophages. The scale bar represents 20 mm. (B) Expression of CD206/HLA-DR in macrophages. The histograms are representatives of five independent experiments of macrophages from five different donors. Numerical values denote the mean fluorescence intensity (MFI). (C) Cytokine levels in the media of macrophages (mean ± SEM, n = 5 independent experiments; *p < 0.05, **p < 0.01, and ***p < compared with control). See also Figure S1. immunosuppression (Kudo-Saito et al., 2009), suggesting that EMT may underlie many biological processes related to cancer development. It has been reported that EMT of cancer cells is frequently observed at the invasive front of advanced tumors (Christofori, 2006; Francí et al., 2006), where TAMs are usually found (Biswas and Mantovani, 2010; Wyckoff et al., 2004). These results suggest that cancer cells that undergo EMT may have a selective advantage in modulating tumor microenvironment. In this study, we investigated the interaction between cancer cells with EMT and TAMs and explored the underlying mechanisms that link these two metastasis-promoting phenomena. RESULTS Mesenchymal-like Breast Cancer Cells Activate Macrophages to a TAM-like Phenotype To investigate whether cancer cells with distinct epithelial or mesenchymal-like phenotypes have different ability to activate macrophages, we cultured freshly isolated human monocytes in culture medium with 30% conditioned medium (CM) from six breast cancer cell lines, including three typical epithelial-like lines (MCF-7, BT-474, and T47D) and three mesenchymal-like lines that express increased vimentin and decreased E-cadherin (BT-549, MDA-MB-436, and MDA-MB-231) (Figure S1A available online), for 6 days to obtain macrophages. As a positive control, we isolated primary TAMs from breast cancer tissue and found that primary TAMs produced high amounts of protumor cytokines (chemokine [C-C motif] ligand [CCL] 18, CCL17, CCL22, and interleukin-10 [IL-10]) (Chen et al., 2011; Dangaj et al., 2011; Mizukami et al., 2008) and exhibited a cluster of differentiation (CD) 206 high /human leukocyte antigen (HLA)-DR low phenotype that is associated with immunosuppression (Dangaj et al., 2011; Kuang et al., 2007). Macrophages treated with CM from mesenchymal-like lines, but not CM from epithelial-like lines, became stretched and elongated (Figure 1A) and exhibited a CD206 high /HLA-DR low phenotype (Figure 1B), similar to primary TAMs. Consistent with the changes in morphology and surface markers, macrophages treated with CM from mesenchymal-like lines produced significantly more tumor-promoting cytokines than untreated macrophages or those treated with CM from epithelial lines (Figure 1C), indicating functional activation of TAMs. Moreover, the level of activation by CM from 606 Cancer Cell 25, , May 12, 2014 ª2014 Elsevier Inc.
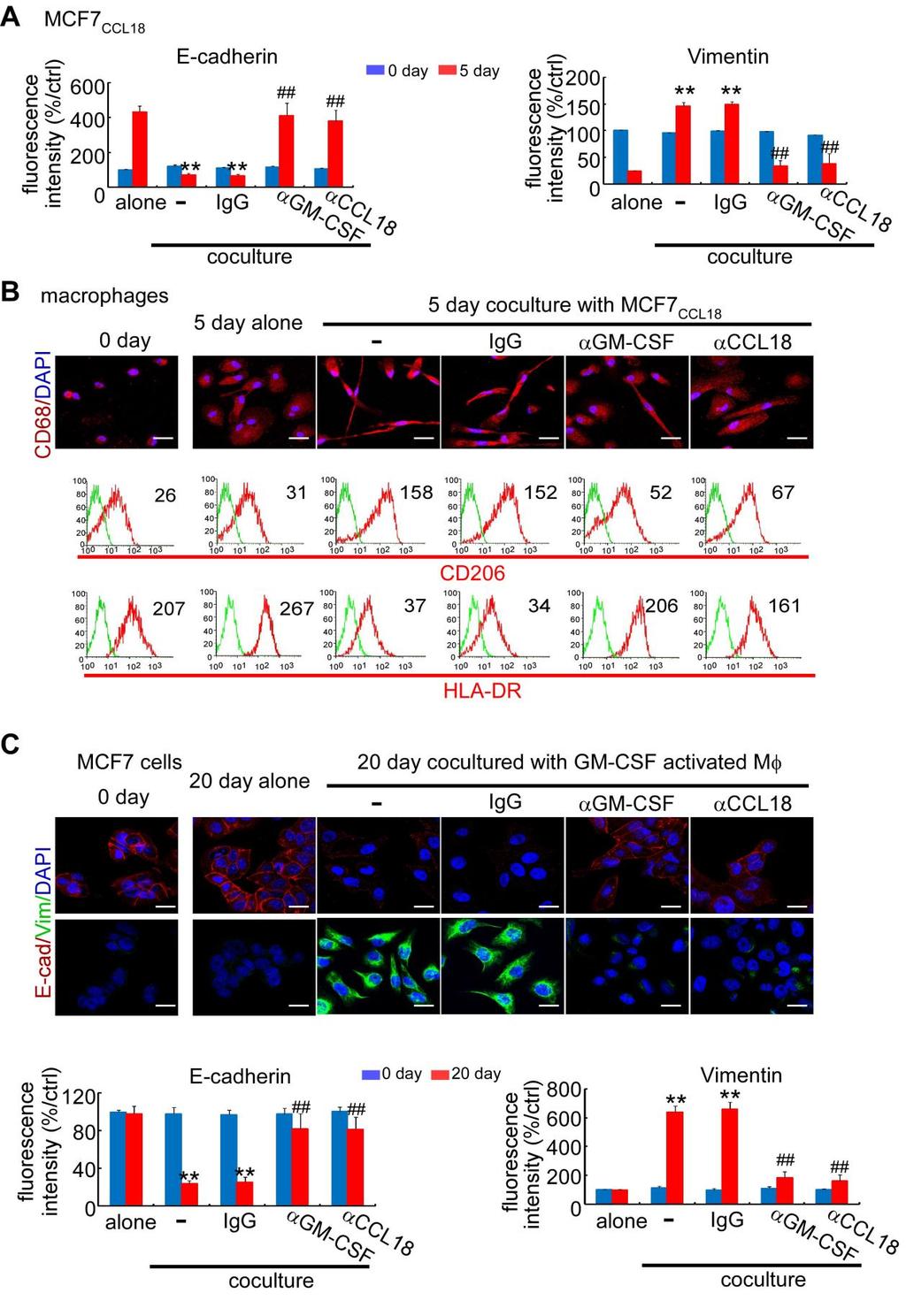
3 Cancer Cell GM-CSF-CCL18 Loop Promotes Breast Cancer Metastasis mesenchymal-like cancer cells, measured by cytokine production and CD206/HLA-DR expression, was comparable with that of primary TAMs, suggesting that CM from mesenchymallike cancer cells is a strong macrophage activator. To confirm whether the ability of cancer cells to activate TAMs is determined by their mesenchymal state, we induced EMT of MCF-7 cells with transforming growth factor b (TGF-b), tumor necrosis factor a (TNF-a) or by mammosphere culture (Guttilla et al., 2012) (Figure S1B). CM from MCF-7 cells that had undergone EMT also induced both morphological and functional TAM-like changes in macrophages, with CM from mammosphere-cultured MCF-7 (MCF-7 M ) having the strongest effects (Figures S1C S1E). BT-474 cells in which EMT was induced also showed similar effects (data not shown). Together, these data indicate that mesenchymal-like cancer cells or cancer cells that have undergone EMT can directly activate macrophages to a TAM-like phenotype and may serve as a previously unappreciated source of TAM activators as important as interleukin-4 (IL-4)-producing CD4 + T cells (DeNardo et al., 2009). Granulocyte-Macrophage Colony-Stimulating Factor Mediates the Activation of Macrophages by Mesenchymal-like Breast Cancer Cells To identify the cytokines secreted by mesenchymal-like cancer cells that activate macrophages, the cytokine profiles of CM from MCF-7, MCF-7 induced to undergo EMT by treatment of TGF-b, TNF-a, and prolonged mammosphere culture, and MDA-MB-231 cells were analyzed using RayBio Human Cytokine Antibody Array. Five cytokines were significantly increased in the CM of MCF-7 cells under EMT and MDA-MB-231 cells compared with CM of MCF-7 cells, which are granulocytemacrophage colony-stimulating factor (GM-CSF), interleukin-8 (IL-8), CCL2, growth-regulated oncogene (GRO; includes GROa, GROb, and GROg), and GROa (Figure 2A). ELISA further confirmed the increases in GM-CSF, IL-8, CCL2, GROa, GROb, and GROg in the CM of mesenchymal-like cells (Figure S2). We then evaluated which of these cytokines is responsible for activating TAMs. We found that only GM-CSF, but not IL-8, CCL2, or GROs, significantly induced the production of TAMrelated cytokines to levels comparable with the CM from MDA- MB-231 cells (Figure 2B). Addition of IL-8, CCL2, or GROs to GM-CSF did not further enhance the activation (data not shown), suggesting that GM-CSF alone is sufficient to activate macrophages. Furthermore, the addition of a neutralizing anti- GM-CSF antibody to the CM of MCF-7 M, MDA-MB-231, and MDA-MB-436 significantly suppressed the induction of the TAM-related surface marker expression (Figure 2C) and cytokines (Figure 2D), indicating that GM-CSF is necessary for mesenchymal-like cancer cells to induce macrophage activation to a TAM-like phenotype. Tumor-Derived Lactate Abrogates the Proinflammatory Cytokine Production from GM-CSF-Activated Macrophages It has been reported that GM-CSF can exhibit proinflammatory or anti-inflammatory activity in different disease models, and it has been hypothesized that conflicting roles of GM-CSF may be due to the concomitant stimulation factors (Hamilton, 2002). We compared the cytokine profiles of macrophages treated with interferon-g (IFN-g) (a standard activator of classically activated macrophages), GM-CSF, or IL-4 (a standard activator of alternatively activated macrophages). We found that GM-CSF alone induced the production of both proinflammatory and antiinflammatory cytokines. As previously reported (Hamilton, 2008), the addition of lipopolysaccharide (LPS) further increased the production of proinflammatory cytokines (Figure S3A). However, LPS is not a common factor found in tumor microenvironment. We then explored the role of lactate, one of the most abundant products of aerobic glycolysis and a widespread concomitant factor in tumor microenvironment (Hirschhaeuser et al., 2011). Breast cancer cell lines produced significantly more lactate ( mm) than macrophages or immortalized breast epithelial cell lines (MCF-10A and HMLE) ( mm) (Figure 3A). The addition of 5 mm lactate abrogated the production of proinflammatory cytokines but did not inhibit or even promoted that of anti-inflammatory cytokines from GM-CSF-activated macrophages (Figure 3B). To further demonstrate the effect of tumor-derived lactate, we treated macrophages with CM of MDA-MB-231 and found that proinflammatory cytokines were significantly decreased, while anti-inflammatory cytokines were significantly increased. The suppression of proinflammatory cytokine production by 231 CM was completely reversed if the 231 cells were pretreated by oxamic acid, an inhibitor of lactate dehydrogenase that blocked the lactate production. The addition of exogenous lactate in oxamic acid-pretreated 231 CM restored the suppression of the proinflammatory cytokines (Figure 3C; Figure S3B), indicating that lactate is responsible for the skew of macrophages toward an anti-inflammatory phenotype. Furthermore, we found that the higher the lactate level, the lower the ph value measured in the cell culture media (Figure 3A). To investigate whether the lactate or the drop in ph level was responsible for the suppression of proinflammatory cytokine production, we adjusted the ph value of the oxamic acid-pretreated 231 CM to the level of untreated 231 CM by 1% hydrochloric acid (HCl) (Figure S3B). We found the acidification by HCl did not suppress the production of proinflammatory cytokines (Figure 3C), suggesting that it is the lactate, not the drop in ph, that is accountable for the immunosuppressive effects by CM of cancer cells. GM-CSF-Activated Macrophages Induce EMT in Breast Cancer Cells via CCL18 Previously, we have reported that TAMs can enhance metastasis of breast cancer cells (Chen et al., 2011; DeNardo et al., 2009), which is often associated with EMT. Thus, we examined whether activated macrophages can induce EMT of cancer cells. We found that MCF-7 cells, when cocultured with GM-CSF-activated macrophages, changed from a rounded shape to an elongated one, with loss of cell-cell contact, decrease of E-cadherin, increase of vimentin, and enhancements of migration and invasiveness (Figures 4A and 4B; Figure S4A), suggesting that GM- CSF-activated macrophages induced EMT of breast cancer cells. Further, anti-ccl18 neutralizing antibody inhibited EMT induced by GM-CSF-activated macrophages, and CCL18 alone induced EMT in MCF-7 (Figures 4A and 4B; Figure S4A), indicating that CCL18 is the cytokine responsible for the EMT of breast cancer cells induced by GM-CSF-activated macrophages. Cancer Cell 25, , May 12, 2014 ª2014 Elsevier Inc. 607
4 Cancer Cell GM-CSF-CCL18 Loop Promotes Breast Cancer Metastasis Figure 2. GM-CSF Mediates the Activation of Macrophages by Mesenchymal-like Breast Cancer Cells (A) Cytokine array of the CM of MCF-7; MCF-7 induced to undergo EMT by treatment of TGF-b, TNF-a, and prolong mammosphere culture; and MDA-MB-231 cells. A table summarizing the relative signal intensity of indicated cytokines is presented in the lower left corner. (B) Cytokines levels in the media of macrophages treated with indicated recombinant human cytokines at indicated concentrations (ng/ml) (mean ± SEM, n=4 independent experiments of macrophages from four different donors). (C) Expression of CD206/HLA-DR in macrophages treated with 30% indicated tumor-cm in the presence or absence of control IgG or a GM-CSF neutralizing antibody. The histograms are representative of five independent experiments of macrophages from five different donors. Numerical values denote the MFI. (D) Cytokine levels in the media of macrophages treated as in (C) (mean ± SEM, n = 5 independent experiments; *p < 0.05, **p < 0.01, and ***p < compared with macrophages treated with cancer cell CM alone; p values were obtained using two-tailed Student s t tests). See also Figure S Cancer Cell 25, , May 12, 2014 ª2014 Elsevier Inc.
5 Cancer Cell GM-CSF-CCL18 Loop Promotes Breast Cancer Metastasis It was recently shown that CCL18 induced EMT in breast cancer cells via the phosphatidylinositol 3-kinase (PI3K)/Akt/ glycogen synthase kinase 3b (GSK3b)/Snail signaling pathway (Zhang et al., 2013). We also observed that CCL18 activated PI3K/Akt pathways (data not shown). NF-kB is an important transcription factor that links the activation of the PI3K/Akt pathway with EMT (Julien et al., 2007). Thus, we explored the role of NF-kB and found that CCL18 induced p65 nuclear translocation and enhanced NF-kB transcriptional activity through membrane-associated phosphatidylinositol transfer protein 3 (PITPNM3) (Figure 4C; Figures S4B and S4C), indicating the activation of the NF-kB pathway. Inhibition of NF-kB by BAY (an IKK inhibitor), 4-methyl-N1-(3-phenyl-propyl)-benzene- 1,2-diamine (JSH-23) (an inhibitor of NF-kB nuclear translocation) or small interfering RNAs (sirnas) to p65 reversed the CCL18-induced EMT in MCF-7 cells (Figure 4D), suggesting that CCL18 activates NF-kB pathway to induce EMT. NF-kB is known to be important in EMT, and it also regulates the expression of various cytokines, including GM-CSF, IL-8, CCL2, and GRO (Barnes and Karin, 1997). We found that inhibition of NF-kB by pharmacological inhibitors or sirna targeting p65 not only led to decreased vimentin and increased E-cadherin in mesenchymal-like breast cancer cell lines (Figure 4D; Figure S4D) but also inhibited their cytokine production in CM (Figure 4E; Figure S4E), suggesting that high NF-kB activity in cancer cells is not only essential to their mesenchymal properties but also important to the ability of secreting cytokines to induce TAMs. A Positive Feedback Loop between GM-CSF and CCL18 In Vitro Because mesenchymal-like cancer cells activate tumor-promoting macrophages that can in turn induce EMT of cancer cells, we investigated the possibility of a positive feedback loop between TAMs and cancer cells with EMT. CCL18-treated MCF-7 cells (MCF-7 CCL18) displayed an elongated shape and mesenchymal markers with increased GM-CSF in media. When these cells were replated without CCL18, the cells gradually reversed to their original round shape, with reexpression of E-cadherin and loss of vimentin by day 5 (Figure 5B; Figure S5A). The GM-CSF in the media also decreased after an initial increase within 48 hr (Figure 5A). However, coculture with macrophages maintained the mesenchymal state of MCF-7 CCL18 cells, and the levels of GM-CSF and CCL18 in media kept increasing through day 5 (Figures 5A and 5B; Figure S5A). The cocultured macrophages became stretched and elongated and displayed a CD206 high /HLA-DR low TAM-like phenotype (Figure S5B). Furthermore, the addition of anti-gm-csf or anti-ccl18 neutralizing antibodies led to regression of the mesenchymal state of MCF-7 CCL18 cells and TAM-like phenotype of macrophages, while decreasing the levels of both GM-CSF and CCL18 in media (Figures 5A and 5B; Figures S5A and S5B), suggesting mutually dependent increases of GM-CSF and CCL18 in the coculture system of MCF-7 CCL18 cells and macrophages. On the other hand, GM-CSF-activated macrophages, when replated without GM-CSF, slowly reversed to a CD206 low /HLA- DR high phenotype (Figure 5D), and the production of CCL18 decreased after the peak at day 4 (Figure 5C). Nevertheless, coculture of GM-CSF-activated macrophages with MCF-7 not only induced EMT of MCF-7 cells (Figure S5C) but also maintained the TAM-like phenotype of the macrophages and increased the levels of GM-CSF and CCL18 through day 20 (Figure 5C and 5D). The addition of anti-gm-csf or anti-ccl18 neutralizing antibodies, but not control immunoglobulin G (IgG), again led to the reversal of the mesenchymal state of MCF-7 cells and the TAM-like phenotype of macrophages, as well as decreasing GM-CSF and CCL18 in media (Figures 5C and 5D; Figure S5C). Collectively, these data suggest that a positive feedback loop between GM-CSF from mesenchymallike cancer cells and CCL18 from TAMs sustains or promotes the mesenchymal state of cancer cells and the TAM-like phenotype of macrophages. Furthermore, similar results were observed when we cocultured MCF-7 cells with macrophages activated by CM of MDA-MB-231, or cocultured macrophages with MDA-MB-231 cells or MCF-7 cells that were induced to EMT by TGF-b or TNF-a (data not shown), corroborating the existence of a GM- CSF-CCL18 positive feedback loop between TAMs and mesenchymal-like cancer cells. The GM-CSF-CCL18 Loop Promotes EMT and Metastasis in a Humanized Mouse Model of Breast Cancer It has been reported that chemokines released by human macrophages are very different from those released by mouse macrophages. Many of the chemokines upregulated in human alternatively activated macrophages either lack murine orthologs or are not upregulated in mice (Martinez et al., 2009). CCL18, the key cytokine released by human macrophages to promote metastasis (Chen et al., 2011), does not yet have a mouse counterpart. Therefore, to study the interaction between human macrophages and cancer cells in vivo, it is necessary to use a humanized mouse model with human hematopoietic cells. To establish such a model, nonobese diabetic (NOD)/severe combined immunodeficiency (SCID) mice were treated with anti-asialo-gm1 antibody to deplete host natural killer (NK) cells, irradiated, transplanted with enriched CD34 + hematopoietic stem cells (HSCs) isolated from fresh human umbilical cord blood (CB) and injected with lentiviral vectors expressing human CSF1 to improve human monocyte reconstitution. This model, modified from a recent protocol (Wang et al., 2012), was reported to increase the engraftment of human monocytes and macrophages (Chen et al., 2009). Engraftment of human HSCs into the mice resulted in the detection of human CD45 + leukocytes, CD14 + monocytes and macrophages, and CD19 + B cells in the peripheral blood and xenografts of humanized mice. Few human CD3 + T cells and CD56 + NK cells were detected (Figures S6A, S6B, and S6E), which is consistent with previous reports (Ito et al., 2012; Wang et al., 2012). Consistent with the findings in vitro, injection of MCF-7 CCL18 cells into the fat pads of humanized mice activated many CCL18 + macrophages nearby, maintained the GM-CSF expression and EMT state of cancer cells (Figure 6A), and significantly increased lung and liver metastasis (Figures 6B 6D), compared with the injection of MCF-7 CCL18 cells into the irradiated mice without HSC transplantation or injection of MCF-7 cells without CCL18 induction into the humanized mice. Anti-GM-CSF Cancer Cell 25, , May 12, 2014 ª2014 Elsevier Inc. 609
6 Cancer Cell GM-CSF-CCL18 Loop Promotes Breast Cancer Metastasis Figure 3. Tumor-Derived Lactate Abrogates the Proinflammatory Cytokine Production from GM-CSF-Activated Macrophages (A) The indicated cells were plated in a 24-well plate at a density of /ml. After 24 hr, lactate production and ph values were measured in the media (mean ± SEM, n = 3; *p < 0.05 and **p < 0.01 compared with MCF10A; p values were obtained using two-tailed Student s t tests). (B) Macrophages were obtained by culture of monocytes in culture medium in the presence or absence of 500 U/ml IFN-g, 50 ng/ml GM-CSF, or 20 ng/ml IL-4 for 6 days. Afterward, macrophages were cultured in culture medium with or without 5 mm L-(+) lactic acid (lactate) for 24 hr. Cytokine levels in the media were (legend continued on next page) 610 Cancer Cell 25, , May 12, 2014 ª2014 Elsevier Inc.
7 Cancer Cell GM-CSF-CCL18 Loop Promotes Breast Cancer Metastasis neutralizing antibody not only decreased the number of CCL18 + cells adjacent to the tumor but also reversed the EMT state of cancer cells and inhibited their metastasis. Furthermore, anti- CCL18 antibody also decreased the expression of GM-CSF in tumor cells, reversed the EMT state of cancer cells, and inhibited metastasis (Figures 6A 6D). Neither GM-CSF nor CCL18 blockade altered human leukocyte reconstitution, the percentages of monocytes and macrophages (Figures S6A S6D), and primary tumor size (Figure 6E) in humanized mice. These results indicate that GM-CSF from cancer cells and CCL18 from TAMs form a positive feedback loop and promote metastasis of cancer cells in vivo. In addition, either anti-gm-csf or anti-ccl18 antibody prolonged the metastasis-free survival of humanized mice injected with MCF-7 CCL18 cells (Figure 6F), suggesting that the GM-CSF-CCL18 loop could be a therapeutic target to inhibit breast cancer metastasis. Similar results were observed in humanized mice injected with MDA-MB-231, a mesenchymal breast cancer cell line, or when GM-CSF was silenced in MCF- 7 CCL18 cells or CCL18 was silenced in HSC-derived macrophages (data not shown) in humanized mice injected with MCF-7 CCL18 cells, substantiating the findings using MCF- 7 CCL18 cells with neutralizing antibodies. The GM-CSF-CCL18 Loop Is Associated with EMT, Triple-Negative Subtype, Metastasis, and Poor Prognosis in Breast Cancer Patients To determine whether our findings are clinically relevant, we examined 1,017 breast cancer cases from three independent breast cancer centers for the expression of GM-CSF. We found that GM-CSF was highly expressed in 293 (28.8%) cancer samples, often observed at the invasive front, with no to weak expression in normal or nonmalignant lesions of the breast (Figure S7A). Double immunostaining of GM-CSF and CCL18 showed significantly more stromal CCL18 + cells in cancers with high GM-CSF expression than in cancers with low GM-CSF expression (Figure 7A). The levels of GM-CSF and CCL18 were also highly correlated in patients serum (Figure S7B). High GM-CSF expression was significantly correlated with lymph node and distant metastasis, more advanced histological grade, triple-negative breast cancer subtype (Table S1). Either GM-CSF or CCL18 was an independent prognostic factor associated with poor prognosis (Figure 7B; Tables S2 and S3). Furthermore, the cancer samples with high GM-CSF expression or more stromal CCL18 + cells nearby expressed significantly less E-cadherin and more vimentin than the cancer samples with less GM-CSF or CCL18 expression (Figures S7C S7E), an indication of EMT. More interestingly, the infiltration of CCL18 + macrophages was often observed at the invasive front of tumors, where cancer cells displayed more mesenchymal features and expressed high levels of GM-CSF (Figure 7C). These data are consistent with the notion that mesenchymal-like cancer cells secret more GM-CSF to activate TAMs that produce CCL18 at the invasive front. To further validate these findings, we searched the Oncomine database for the prognostic value of GM-CSF and CCL18 in human breast cancer. Four data sets showed a correlation between GM-CSF expression and high-grade or triple-negative subtype breast cancer, while one and four data sets showed associations between CCL18 expression and high-grade breast cancer and between CCL18 expression and triple-negative subtype breast cancer, respectively. In addition, four and three data sets confirmed that high levels of GM-CSF and of CCL18 are significantly correlated with poor prognosis in breast cancer patients, respectively. Furthermore, four and two data sets showed significantly inverse correlations between GM-CSF and CCL18 expression and E-cadherin expression, respectively. More important, in The Cancer Genome Atlas data set, with 593 breast cancer cases, GM-CSF was significantly correlated with CCL18 in breast cancer samples (p < ) (Table 1). Collectively, these data suggest that the GM-CSF-CCL18 loop plays an important role in EMT and metastasis of human breast cancer. DISCUSSION In this study, we show that cancer cells that have undergone EMT secrete GM-CSF to activate macrophages to a TAM-like phenotype. In turn, activated macrophages produce CCL18 to induce EMT of cancer cells, forming a positive feedback loop. This GM-CSF-CCL18 loop is essential to promoting the metastasis of breast cancer cells and is associated with poor prognosis in breast cancer patients. EMT of cancer cells results in increased cell motility and is associated with metastasis. Our results indicate that cancer cells that undergo EMT, often located at the invasive front of tumors, have a superior function in activating macrophages to support tumor expansion. Besides metastasis, EMT of cancer cells is also linked with cancer stem cells, drug resistance, and immunosuppression (Kudo-Saito et al., 2009; Mani et al., 2008; Thiery et al., 2009). We report here that the change of secretory cytokine profile is also an important feature for cancer cells that undergo EMT. We previously described that CCL18 is one of the major cytokines released by TAMs and enhances breast cancer cell metastasis (Chen et al., 2011). In line with these results, CCL18 from activated macrophages induced EMT of breast cancer cells through the activation of NF-kB. In addition, a panel of inflammatory cytokines, including GM-CSF, CCL2, IL-8, and GROs, were significantly induced as NF-kB target genes (Barnes measured by ELISA (mean ± SEM, n = 4 independent experiments of macrophages from four different donors; *p < 0.05, **p < 0.01, and ***p < 0.001; p values were obtained using two-tailed Student s t tests). (C) 231 cells were cultured in the presence or absence of 90 mm oxamic acid for 5 days. CM were collected after the cells were cultured for another 24 hr without oxamic acid. In addition, the lactate or ph level in 231 cell CM treated with oxamic acid were adjusted to the levels in untreated 231 cell CM by the addition of lactate or 1% HCl, respectively. Macrophages were obtained by culture of monocytes in culture medium in the presence or absence of indicated 231 CM for 6 days. The cytokine concentrations of the macrophages were measured afterward (mean ± SEM, n = 4 independent experiments of macrophages from four different donors; *p < 0.05, **p < 0.01, and ***p < 0.001; p values were obtained using two-tailed Student s t tests). See also Figure S3. Cancer Cell 25, , May 12, 2014 ª2014 Elsevier Inc. 611
 Morphology and E-cadherin and vimentin expression of MCF-7 cells without or with the coculture of control macrophages (M4),")
8 Cancer Cell GM-CSF-CCL18 Loop Promotes Breast Cancer Metastasis Figure 4. GM-CSF-Activated Macrophages Induce EMT of Cancer Cells through CCL18 (A) Morphology and E-cadherin and vimentin expression of MCF-7 cells without or with the coculture of control macrophages (M4), GM-CSF-activated macrophages alone or with control IgG, anti-ccl18 neutralizing antibody, or recombinant human CCL18. The scale bar represents 20 mm. GAPDH, glyceraldehyde 3-phosphate dehydrogenase. (B) Migration and invasion assays of MCF-7 cells treated as in (A). The scale bar represents 200 mm (mean ± SEM, n = 3; **p < 0.01 and ***p < 0.001; p values were obtained using two-tailed Student s t tests). (legend continued on next page) 612 Cancer Cell 25, , May 12, 2014 ª2014 Elsevier Inc.
9 Cancer Cell GM-CSF-CCL18 Loop Promotes Breast Cancer Metastasis and Karin, 1997) by CCL18. Among the cytokines induced during EMT, GM-CSF is the one responsible for macrophage activation. Other increased cytokines are also known to be important in other aspects of tumor microenvironment, for example, CCL2 in recruiting monocytes (Qian et al., 2011), GROs in neutrophil recruitment (Jinquan et al., 1995), and IL-8 in angiogenesis (Lin et al., 2004). It is possible that the inflammatory factors induced during EMT work together to provide a permissive environment for tumor progression. It has been reported that both breast cancer cells and macrophages comigrate and depend upon each other to be invasive in vivo (Wyckoff et al., 2004). Our findings may extend knowledge of interactions between cancer cells and macrophages by showing that GM-CSF from cancer cells and CCL18 from TAMs form a positive feedback loop to sustain or promote EMT and metastasis of cancer cells. The levels of these two cytokines are mutually dependent, such that neutralizing either of them broke the vicious circle and suppressed breast cancer metastasis in a humanized mouse model. These results, together with the analysis of clinical samples, provide an explanation for the colocalization of TAMs and cancer cells that have undergone EMT at the invasive front of tumors. Our data suggest a protumor role of GM-CSF in cancer. However, GM-CSF has been shown to elicit powerful immune responses and is often used as an immune adjuvant to cancer vaccines (Dranoff, 2002). However, the effect of exogenous GM-CSF on cancer remains controversial. Some recent clinical studies reported that GM-CSF as an adjuvant to different cancer vaccines can lead to adverse outcome in terms of immune response as well as relapse-free and overall survival (Faries et al., 2009; Filipazzi et al., 2007; Slingluff et al., 2009). Careful analysis suggested that GM-CSF may increase the vaccineinduced immune response when administered repeatedly at relatively low doses (range mg for 1 5 days), whereas an opposite effect was often reported at higher dosages of mg (Parmiani et al., 2007). Similar findings were also reported in animal models (Serafini et al., 2004). Furthermore, it has been consistently reported that endogenous GM-CSF from cancer cells is capable of suppressing the immune response (Bronte et al., 1999; Marigo et al., 2010; Tsuchiya et al., 1988) and correlated with increased metastasis (Park et al., 2007; Takeda et al., 1991). The analysis of our clinical samples as well as independent Oncomine data sets also showed that GM-CSF is associated with poor prognosis in breast cancer patients. We observed that LPS, a common factor of infectious disease, skewed GM-CSF-activated macrophages to a proinflammatory phenotype, while lactate, one of the most abundant tumor metabolites, changed GM-CSF-activated macrophages to an anti-inflammatory phenotype. It is possible that longterm proximal exposure to endogenous GM-CSF from cancer cells together with tumor metabolites, unlike the short-term systemic use of low doses of GM-CSF in cancer vaccines, leads to cancer progression. Compared with immortalized breast epithelial cell lines, both epithelial-like and mesenchymal-like breast cancer cell lines produced significantly more lactate. Further, the addition of 5 mm lactate, which was equivalent to or lower than the amount produced by both epithelial-like and mesenchymal-like breast cancer cell lines ( mm), abrogated the production of proinflammatory cytokines but did not inhibit that of anti-inflammatory cytokines from GM-CSF-activated macrophages. These results indicate that lactate produced by epithelial-like breast cancer cells is sufficient to affect GM-CSF-activated macrophage. However, our data also demonstrated that GM-CSF produced by epithelial-like breast cancer cell lines was significantly less than that by mesenchymal-like breast cancer cells and was insufficient to activate macrophages. Thus, both increased lactate level and enhanced GM-CSF production are needed for breast cancer cells to activate macrophages to an anti-inflammatory phenotype. Previous studies have identified the interaction between cancer cells and macrophages, including the colony-stimulating factor 1 (CSF-1)(macrophage colony-stimulating factor [M-CSF])-epidermal growth factor (EGF) loop by using a rat mammary cancer cell line (MTLn3) and mouse macrophages (Goswami et al., 2005; Hernandez et al., 2009). When human breast cancer line MDA-MB-231 was used in an in vivo mouse xenograft model, the proportion of macrophages among the invasive cells was only 6%, much less than the 25% observed with rodent tumors (Patsialou et al., 2009). In addition, while Iressa (an EGF receptor inhibitor) completely inhibited the EGF or CSF-1-driven invasion in rodent tumor, it failed to do so in the MDA-MB-231 xenografts (Patsialou et al., 2009). It is known that the chemokine profiles of human and mouse macrophages are very different. At least five chemokines that are upregulated in human alternatively activated macrophages either lack murine orthologs or are not upregulated in mice (Martinez et al., 2009). We find that GM-CSF is significantly induced in human mesenchymal-like cancer cells and is responsible for the macrophage activation, which then promotes the EMT of cancer cells to form the self-reinforcing loop. We also found that CSF-1 is much lower than GM-CSF and is only mildly increased in human mesenchymal-like cancer cells (data not shown). Moreover, blocking the CSF1-EGF loop has no significant effect on mesenchymal-like cancer cell-induced macrophage activation in vitro and metastasis of cancer cell in vivo (data not shown). More important, our findings using human breast cancer cell lines and human macrophages were validated in a humanized mouse model of breast cancer in vivo and further corroborated in 1,017 clinical samples as well as Oncomine data sets. Nevertheless, it is possible that GM-CSF plays an indispensable role in initiating the positive feedback loop at (C) Confocal fluorescent microscopy of p65/dapi staining in MCF-7 cells treated without or with rhccl20 or rhccl18 together with sirnas targeting GFP or PITPNM3. The scale bar represents 20 mm. (D) Western blotting for E-cadherin and vimentin of CCL18-treated MCF-7 cells pretreated with DMSO, BAY , or JSH-23 or pretransfected with GFPsiRNA or P65-siRNAs. (E) The cytokine levels in the media of cells treated as in (D) (mean ± SEM, n = 3; **p < 0.01 and ***p < 0.001; p values were obtained using two-tailed Student s t tests). See also Figure S4. Cancer Cell 25, , May 12, 2014 ª2014 Elsevier Inc. 613
 Upper: schematic showing that MCF-7CCL18 cells were re-plated alone or cocultured with macrophages in")
10 Cancer Cell GM-CSF-CCL18 Loop Promotes Breast Cancer Metastasis Figure 5. A Positive Feedback Loop between GM-CSF from Cancer Cells and CCL18 from Macrophages In Vitro (A) Upper: schematic showing that MCF-7CCL18 cells were re-plated alone or cocultured with macrophages in transwell apparatus with 0.4 mm pore size with or without control IgG, anti-gm-csf, or anti-ccl18 neutralizing antibody for 5 days. Lower: GM-CSF and CCL18 levels in the indicated media were measured by ELISA at indicated time points. All values are mean ± SEM, n = 4 independent experiments of macrophages from four different donors; *p < 0.05, **p < 0.01, and ***p < compared with cells cultured alone; #p < 0.05, ##p < 0.01, and ###p < compared with cells in the coculture system without antibody addition at the same time points (p values were obtained using two-tailed Student s t tests). (legend continued on next page) 614 Cancer Cell 25, , May 12, 2014 ª2014 Elsevier Inc.
11 Cancer Cell GM-CSF-CCL18 Loop Promotes Breast Cancer Metastasis the early stage of metastasis, while other factors may be crucial at later stages. In summary, our study suggests an important role of the positive feedback loop between cancer cells with EMT and TAMs in cancer metastasis. The GM-CSF-CCL18 loop is essential for cancer cells to maintain or promote their mesenchymal phenotype and could be a potential therapeutic target for cancer metastasis. EXPERIMENTAL PROCEDURES Detailed procedures are provided in Supplemental Experimental Procedures. Patients and Tissue Samples Primary breast carcinomas were obtained from 759 patients at the Sun Yat- Sen Memorial Hospital, Sun Yat-Sen University (Guangzhou, China), 145 patients at the First Affiliated Hospital, Shantou University Medical College (Shantou, China), and 113 patients at Tianjin Medical University Cancer Institute and Hospital (Tianjin, China). Normal breast tissue from 9 patients undergoing breast reduction surgery and benign breast tissue samples from 23 patients with fibroadenoma and 17 patients with cystic fibrosis with or without atypical epithelial hyperplasia were collected at Sun Yat-Sen Memorial Hospital. Additionally, serum levels of GM-CSF and CCL18 were measured in peripheral blood samples from 151 patients with breast cancer before starting therapy at Sun Yat-Sen Memorial Hospital. All samples were collected with informed consent from patients, and all related procedures were performed with the approval of the internal review and ethics boards of the indicated hospitals. Cell Culture and Treatment MDA-MB-231, MDA-MB-436, BT-549, T47D, BT-474, and MCF-7 breast cancer cells were obtained from American Type Culture Collection and grown according to standard protocols. HMLE cells were kindly provided and cultured as recommended by Dr. R.A. Weinberg (Whitehead Institute, Cambridge, MA). When grown to 80% of confluence, cells were washed with PBS, and fresh serum-free media were added. CM were harvested 24 hr later and filtered through a 0.22 mm filter. In some experiments, MCF-7 cells were treated with 5 ng/ml TGF-b, 10 ng/ml TNF-a, or 20 ng/ml CCL18 (PeproTech) for 2 weeks or given prolonged mammosphere culture for 5 weeks as previously described (Guttilla et al., 2012). Afterward, cells were washed in PBS and grown in adherent condition with cytokine/serum-free media, which were harvested 24 hr later. To inhibit respective pathways, cells were incubated with vehicle (DMSO), 5 mm BAY , or 6 mm JSH-23 (Calbiochem) for 1 hr at 37 C prior to the experiments. To inhibit lactate production, cells were treated with 90 mm oxamic acid (Sigma) for 5 days. Primary Human TAM Isolation from Breast Cancer TAMs were isolated from five fresh breast cancer samples as previously described (Chen et al., 2011; Dangaj et al., 2011), with slight modifications. Briefly, the tissues were minced into small (1 to 2 mm) pieces and digested with 5% fetal bovine serum Dulbecco s modified Eagle s medium containing 2 mg/ml collagenase I and 2 mg/ml hyaluronidase (Sigma) at 37 C for 2 hr. The cells were sequentially filtered through 500 mm mesh, 100 mm, and 70 mm cell strainer. The cells were then centrifuged in a Beckman Allegra X-15R centrifuge at 2,500 rpm for 20 min with 1 ml cell suspension above 5 ml 45% Percoll (GE Healthcare) in the middle and 5 ml 60% Percoll at the bottom in a 15 ml tube. Mononuclear cells were collected from the cell layer in the interphase between 45% and 60% Percoll. CD14 + monocytes and macrophages were isolated by a magnetic-activated cell sorting using direct CD14 Isolation Kit (Miltenyi Biotec) according to the manufacturer s instructions. Hematopoietic Stem/Progenitor Cell Isolation Fresh human CB was obtained from Sun Yat-Sen Memorial Hospital, according to guidelines approved by the ethics boards and the Clinical Research Committee at Sun Yat-Sen Memorial Hospital. The HSCs were isolated as previously described (Holt et al., 2010). Briefly, CB mononuclear cells were separated by density gradient of Ficoll-Hypaque. CD34 + HSCs were isolated by using direct CD34 Progenitor Cell Isolation Kit (Miltenyi Biotec). More than 95% of CD34 + cells were positively selected after two rounds of enrichment. The HSCs were transplanted within 24 hr after isolation. Transplantation of CD34 + Cells and MCF-7 into NOD/SCID Mice All animal work was conducted in accordance with a protocol approved by the Institutional Animal Care and Use Committee at the medical college of Sun Yat-Sen University. Female NOD/SCID mice (3 to 4 weeks old) were injected intraperitoneally with 50 ml of anti-asialo-gm1 antibody (Wako) 24 hr before HSC transplantation. The mice were subjected to 200 cgy totalbody irradiation 12 hr before being injected with HSCs in 0.2 ml of medium via the tail vein. After transplantation, treatment with anti-asialo- GM1 antibody was repeated once every 4 days for 24 days. The mice were injected with 0.1 ml of human M-CSF encoding lentiviral vectors at titers ranging from to total TU/ml via the tail vein 1 week after transplantation. At 8 weeks after HSC transplantation, MCF-7 cells with indicated treatment were injected into the mammary fat pads of the mice. To supplement estrogen for MCF-7 growth, each mouse was implanted with a 1.5 mg 17b-estradiol pellet (Innovative Research of America) 3 days before MCF-7 injection. For antibody treatment, mice were injected with CCL18-specific neutralizing antibody (Catalog No. ab9849; abcam) and/or GM-CSF-specific neutralizing antibody (Catalog No. MAB215; R&D Systems) via the tail vein at 10 mg/mouse twice weekly after the xenografts became palpable (about 0.4 cm in diameter). Tumor growth was evaluated by monitoring tumor volume (length 3 width ) every 3 days. The whole-body metastasis burden of animals bearing xenografts that stably express luciferase was monitored using the IVIS Lumina Imaging System (Xenogen) every 2 days as previously described (Oskarsson et al., 2011). The animals were sacrificed when the xenografts reached 1.5 cm in diameter. The plasma, tumor xenografts, lungs, and livers of the sacrificed mice were harvested for further investigation. Sections (4 mm) of the harvested organs were immunostained with antibody specific for human cytokeratin (catalog no. ab756; abcam), and total RNA was extracted for quantitative reverse transcription polymerase chain reaction analysis of human hypoxanthine phosphoribosyltransferase (HPRT) mrna expression. Statistics All statistical analysis was done using SPSS for Windows version 13.0 (SPSS). Pearson s correlation and regression analysis was performed to assess the relationship between GM-CSF and CCL18 in the serum of patients and human breast tissue mrna expression data from Oncomine. Chi-square analysis was applied to analyze the relationship between GM-CSF expression and clinicopathological status. Kaplan-Meier survival curves were plotted, and the logrank test was done. Groups of discrete variables were compared by means (B) Immunofluorescence analysis of E-cadherin (E-cad) and vimentin (Vim) in MCF-7 CCL18 cells alone or in the coculture system as described in (A). The scale bar represents 20 mm. (C) Upper: schematic showing that macrophages obtained by culture of monocytes in culture medium with 50 ng/ml GM-CSF for 6 days were re-plated alone or cocultured with untreated MCF-7 in the absence or presence of control IgG, anti-gm-csf, or anti-ccl18 neutralizing antibody for 20 days. Lower: GM-CSF and CCL18 levels in the indicated media were measured by ELISA at indicated time points. All values are mean ± SEM, n = 4 independent experiments of macrophages from four different donors; *p < 0.05, **p < 0.01, and ***p < compared with cells cultured alone; #p < 0.05, ##p < 0.01, and ###p < compared with cells in the coculture system without antibody addition at the same time points (p values were obtained using two-tailed Student s t tests). (D) Surface marker detection by flow cytometry in GM-CSF-activated macrophages as described in (C). The histograms are representative of four independent experiments of macrophages from four different donors. Numerical values denote the MFI. See also Figure S5. Cancer Cell 25, , May 12, 2014 ª2014 Elsevier Inc. 615

12 Cancer Cell GM-CSF-CCL18 Loop Promotes Breast Cancer Metastasis Figure 6. The GM-CSF-CCL18 Loop Promotes EMT and Metastasis in a Humanized Mouse Model MCF-7CCL18 or MCF-7 cells without CCL18 treatment were injected into the mammary fat pads of NOD/SCID mice with or without human hematopoietic stem cell transplantation. Mice were injected with control IgG, anti-ccl18 neutralizing antibody, and/or anti-gm-csf neutralizing antibody via the tail vein after the xenografts became palpable. (legend continued on next page) 616 Cancer Cell 25, , May 12, 2014 ª2014 Elsevier Inc.
 Left: representative double IHC staining of GMCSF (red) and CCL18 (brown) in breast cancer samples with low")
13 Cancer Cell GM-CSF-CCL18 Loop Promotes Breast Cancer Metastasis Figure 7. The GM-CSF-CCL18 Loop Is Associated with EMT and Metastasis in Breast Cancer Patients (A) Left: representative double IHC staining of GMCSF (red) and CCL18 (brown) in breast cancer samples with low (upper panel; n = 724) or high (lower panel; n = 293) GM-CSF expression. Right: quantification of CCL18+ macrophages in human breast cancer samples (mean ± SEM; ***p < 0.001; p values were determined using Student s t tests). Scale bars represent 50 mm. (B) Kaplan-Meier survival curve of breast cancer patients with low (n = 724) and high (n = 293) GMCSF expression (p < by log-rank test), with a median follow-up period of 68 months. The number of surviving patients stratified to the follow-up periods is indicated below the graph. (C) Representative IHC staining for E-cadherin, vimentin, GM-CSF, and CCL18 in the invasive front and noninvasive front of serial sections from a human breast cancer sample. The sample used was derived from 1,017 breast cancer cases from three independent breast cancer centers. Scale bars represent 50 mm. See also Tables S1 S3 and Figure S7. of the Mann-Whitney U test or Kruskal-Wallis nonparametric analysis of variance. All experiments for cell cultures were performed independently at least three times and in triplicate each time. In all cases, p values < 0.05 were considered statistically significant. ACCESSION NUMBERS The Gene Expression Omnibus database ( accession number for the array data reported in this paper is GSE (A) Upper: representative pictures of double immunohistochemical (IHC) staining of E-cadherin (red) and vimentin (brown) or GM-CSF (red) and CCL18 (brown) in the sections from indicated tumor xenografts. Arrows indicate CCL18+ cells. Insets show CCL18+ cells at higher magnification. The scale bar represents 50 mm. Lower: immunoreactive scores of E-cadherin (red) and vimentin (brown) and immunoreactive scores of GM-CSF (red) and CCL18+ cell number counts (brown) within indicated xenografts (mean ± SEM, n = 8 for each group; ***p < compared with cells in nonhumanized mice; ##p < 0.01 compared with MCF-7CCL18 in humanized mice treated with control antibody; p values were determined using Kruskal-Wallis tests). Abs, antibodies. (B) Representative bioluminescence imaging (upper panel) and lung and liver sections immunostained with human cytokeratin antibody to indicate human cancer cell metastasis (lower panel) in mice treated as in (A). Scale bars represent 50 mm. (C) Bioluminescence signal of lung and liver metastasis in mice with the indicated treatments (mean ± SEM, n = 8 for each group; **p < 0.01 and ***p < 0.001; p values were determined using Student s t tests). (D) Expression of human HPRT mrna relative to mouse 18S rrna in the lungs and livers of the tumor-bearing mice. Data were normalized to the group of nonhumanized mice (mean ± SEM, n = 8 for each group; **p < 0.01 and ***p < 0.001; p values were determined using Student s t tests). (E) Tumor size during the course of each indicated treatment (arrow indicates beginning of the treatment when tumor is palpable). Error bars show ±SEM. (F) Kaplan-Meier analysis of metastasis-free survival in mice with the indicated treatments. Metastasis was determined by bioluminescence (n = 8 for each group; **p < 0.01 and ***p < 0.001; p values were determined using log-rank tests). See also Figure S6. Cancer Cell 25, , May 12, 2014 ª2014 Elsevier Inc. 617
14 Cancer Cell GM-CSF-CCL18 Loop Promotes Breast Cancer Metastasis Table 1. Correlation of CCL18 and GM-CSF Expression and Clinical Features in Breast Cancer in the Oncomine Online Database Fold Change p Value Database (Case Number) Upregulation of GM-CSF in Triple-Negative Breast Cancer Triple-negative breast carcinoma versus others < TCGA Breast (593) Triple-negative breast carcinoma versus others Stickeler Breast (57) Triple-negative breast carcinoma versus others Waddell Breast (85) Triple-negative breast carcinoma versus others Kao Breast (327) Upregulation of GM-CSF in High-Grade Breast Cancer Grade III versus grades I and II < Loi Breast3 (77) Grade III versus grades I and II < Curtis Breast (2136) Grade III versus grades I and II Gluck Breast (158) Grade III versus grades I and II Stickeler Breast (57) Correlation of GM-CSF and Poor Prognosis in Breast Cancer Metastasis at 5 years versus no metastasis at 5 years Kao Breast (327) Metastasis at 5 years versus no metastasis at 5 years Bos Breast (204) Metastasis at 5 years versus no metastasis at 5 years Hatzis Breast (508) Recurrence at 1 year versus no recurrence at 1 year Vandevijver Breast (295) Upregulation of CCL18 in Triple-Negative Breast Cancer Triple-negative breast carcinoma versus others < TCGA Breast (593) Triple-negative breast carcinoma versus others Bild Breast (158) Triple-negative breast carcinoma versus others < Bittner Breast (336) Triple-negative breast carcinoma versus others < Chin Breast (118) Upregulation of CCL18 in High-Grade Breast Cancer Grade III versus grades I and II Sorlie Breast (167) Correlation of CCL18 and Poor Prognosis in Breast Cancer Recurrence at 5 years versus no recurrence at 5 years Ma Breast (60) Dead at 5 years versus alive at 5 years Vandevijver Breast (295) Metastasis at 5 years versus no metastasis at 5 years Hatzis Breast (508) Correlation of GM-CSF and E-cadherin in Breast Cancer Correlation of GM-CSF and E-cadherin in breast cancer < Hatzis Breast (508) Correlation of GM-CSF and E-cadherin in breast cancer TCGA Breast (593) Correlation of GM-CSF and E-cadherin in breast cancer Bittner Breast (336) Correlation of GM-CSF and E-cadherin in breast cancer Kao Breast (327) Correlation of CCL18 and E-cadherin in Breast Cancer Correlation of CCL18 and E-cadherin in breast cancer Hatzis Breast (508) Correlation of CCL18 and E-cadherin in breast cancer TCGA Breast (593) Correlation of CCL18 and GM-CSF in Breast Cancer Correlation of CCL18 and GM-CSF in breast cancer < TCGA Breast (593) SUPPLEMENTAL INFORMATION Supplemental Information includes Supplemental Experimental Procedures, seven figures, and three tables and can be found with this article online at ACKNOWLEDGMENTS This work was supported by grants from 973 (2010CB and 2011CB504203) Projects from the Ministry of Science and Technology of China, the Natural Science Foundation of China ( , , , , and ), the National S&T Major Special Project on New Drug Innovation of China (no. 2011ZX ), the Science Foundation of Guangdong Province (S ), the Translational Medicine Public Platform of Guangdong Province ( ), and the Ministry of Education of China ( ). Received: April 22, 2013 Revised: October 25, 2013 Accepted: March 19, 2014 Published: May 12, 2014 REFERENCES Allavena, P., Sica, A., Solinas, G., Porta, C., and Mantovani, A. (2008). The inflammatory micro-environment in tumor progression: the role of tumor-associated macrophages. Crit. Rev. Oncol. Hematol. 66, 1 9. Barnes, P.J., and Karin, M. (1997). Nuclear factor-kappab: a pivotal transcription factor in chronic inflammatory diseases. N. Engl. J. Med. 336, Biswas, S.K., and Mantovani, A. (2010). Macrophage plasticity and interaction with lymphocyte subsets: cancer as a paradigm. Nat. Immunol. 11, Bronte, V., Chappell, D.B., Apolloni, E., Cabrelle, A., Wang, M., Hwu, P., and Restifo, N.P. (1999). Unopposed production of granulocyte-macrophage 618 Cancer Cell 25, , May 12, 2014 ª2014 Elsevier Inc.
15 Cancer Cell GM-CSF-CCL18 Loop Promotes Breast Cancer Metastasis colony-stimulating factor by tumors inhibits CD8+ T cell responses by dysregulating antigen-presenting cell maturation. J. Immunol. 162, Chen, Q., Khoury, M., and Chen, J. (2009). Expression of human cytokines dramatically improves reconstitution of specific human-blood lineage cells in humanized mice. Proc. Natl. Acad. Sci. U S A 106, Chen, J., Yao, Y., Gong, C., Yu, F., Su, S., Liu, B., Deng, H., Wang, F., Lin, L., Yao, H., et al. (2011). CCL18 from tumor-associated macrophages promotes breast cancer metastasis via PITPNM3. Cancer Cell 19, Christofori, G. (2006). New signals from the invasive front. Nature 441, Condeelis, J., and Pollard, J.W. (2006). Macrophages: obligate partners for tumor cell migration, invasion, and metastasis. Cell 124, Dangaj, D., Abbott, K.L., Mookerjee, A., Zhao, A., Kirby, P.S., Sandaltzopoulos, R., Powell, D.J., Jr., Lamazière, A., Siegel, D.L., Wolf, C., and Scholler, N. (2011). Mannose receptor (MR) engagement by mesothelin GPI anchor polarizes tumor-associated macrophages and is blocked by anti-mr human recombinant antibody. PLoS ONE 6, e DeNardo, D.G., Barreto, J.B., Andreu, P., Vasquez, L., Tawfik, D., Kolhatkar, N., and Coussens, L.M. (2009). CD4(+) T cells regulate pulmonary metastasis of mammary carcinomas by enhancing protumor properties of macrophages. Cancer Cell 16, Dranoff, G. (2002). GM-CSF-based cancer vaccines. Immunol. Rev. 188, Faries, M.B., Hsueh, E.C., Ye, X., Hoban, M., and Morton, D.L. (2009). Effect of granulocyte/macrophage colony-stimulating factor on vaccination with an allogeneic whole-cell melanoma vaccine. Clin. Cancer Res. 15, Filipazzi, P., Valenti, R., Huber, V., Pilla, L., Canese, P., Iero, M., Castelli, C., Mariani, L., Parmiani, G., and Rivoltini, L. (2007). Identification of a new subset of myeloid suppressor cells in peripheral blood of melanoma patients with modulation by a granulocyte-macrophage colony-stimulation factor-based antitumor vaccine. J. Clin. Oncol. 25, Francí, C., Takkunen, M., Dave, N., Alameda, F., Gómez, S., Rodríguez, R., Escrivà, M., Montserrat-Sentís, B., Baró, T., Garrido, M., et al. (2006). Expression of Snail protein in tumor-stroma interface. Oncogene 25, Goswami, S., Sahai, E., Wyckoff, J.B., Cammer, M., Cox, D., Pixley, F.J., Stanley, E.R., Segall, J.E., and Condeelis, J.S. (2005). Macrophages promote the invasion of breast carcinoma cells via a colony-stimulating factor-1/ epidermal growth factor paracrine loop. Cancer Res. 65, Guttilla, I.K., Phoenix, K.N., Hong, X., Tirnauer, J.S., Claffey, K.P., and White, B.A. (2012). Prolonged mammosphere culture of MCF-7 cells induces an EMT and repression of the estrogen receptor by micrornas. Breast Cancer Res. Treat. 132, Hamilton, J.A. (2002). GM-CSF in inflammation and autoimmunity. Trends Immunol. 23, Hamilton, J.A. (2008). Colony-stimulating factors in inflammation and autoimmunity. Nat. Rev. Immunol. 8, Hernandez, L., Smirnova, T., Kedrin, D., Wyckoff, J., Zhu, L., Stanley, E.R., Cox, D., Muller, W.J., Pollard, J.W., Van Rooijen, N., and Segall, J.E. (2009). The EGF/CSF-1 paracrine invasion loop can be triggered by heregulin beta1 and CXCL12. Cancer Res. 69, Hirschhaeuser, F., Sattler, U.G., and Mueller-Klieser, W. (2011). Lactate: a metabolic key player in cancer. Cancer Res. 71, Holt, N., Wang, J., Kim, K., Friedman, G., Wang, X., Taupin, V., Crooks, G.M., Kohn, D.B., Gregory, P.D., Holmes, M.C., and Cannon, P.M. (2010). Human hematopoietic stem/progenitor cells modified by zinc-finger nucleases targeted to CCR5 control HIV-1 in vivo. Nat. Biotechnol. 28, Ito, R., Takahashi, T., Katano, I., and Ito, M. (2012). Current advances in humanized mouse models. Cell. Mol. Immunol. 9, Jinquan, T., Frydenberg, J., Mukaida, N., Bonde, J., Larsen, C.G., Matsushima, K., and Thestrup-Pedersen, K. (1995). Recombinant human growth-regulated oncogene-alpha induces T lymphocyte chemotaxis. A process regulated via IL-8 receptors by IFN-gamma, TNF-alpha, IL-4, IL-10, and IL-13. J. Immunol. 155, Joyce, J.A., and Pollard, J.W. (2009). Microenvironmental regulation of metastasis. Nat. Rev. Cancer 9, Julien, S., Puig, I., Caretti, E., Bonaventure, J., Nelles, L., van Roy, F., Dargemont, C., de Herreros, A.G., Bellacosa, A., and Larue, L. (2007). Activation of NF-kappaB by Akt upregulates Snail expression and induces epithelium mesenchyme transition. Oncogene 26, Kuang, D.M., Wu, Y., Chen, N., Cheng, J., Zhuang, S.M., and Zheng, L. (2007). Tumor-derived hyaluronan induces formation of immunosuppressive macrophages through transient early activation of monocytes. Blood 110, Kudo-Saito, C., Shirako, H., Takeuchi, T., and Kawakami, Y. (2009). Cancer metastasis is accelerated through immunosuppression during Snail-induced EMT of cancer cells. Cancer Cell 15, Lin, Y., Huang, R., Chen, L., Li, S., Shi, Q., Jordan, C., and Huang, R.P. (2004). Identification of interleukin-8 as estrogen receptor-regulated factor involved in breast cancer invasion and angiogenesis by protein arrays. Int. J. Cancer 109, Mani, S.A., Guo, W., Liao, M.J., Eaton, E.N., Ayyanan, A., Zhou, A.Y., Brooks, M., Reinhard, F., Zhang, C.C., Shipitsin, M., et al. (2008). The epithelial-mesenchymal transition generates cells with properties of stem cells. Cell 133, Marigo, I., Bosio, E., Solito, S., Mesa, C., Fernandez, A., Dolcetti, L., Ugel, S., Sonda, N., Bicciato, S., Falisi, E., et al. (2010). Tumor-induced tolerance and immune suppression depend on the C/EBPbeta transcription factor. Immunity 32, Martinez, F.O., Helming, L., and Gordon, S. (2009). Alternative activation of macrophages: an immunologic functional perspective. Annu. Rev. Immunol. 27, Mizukami, Y., Kono, K., Kawaguchi, Y., Akaike, H., Kamimura, K., Sugai, H., and Fujii, H. (2008). CCL17 and CCL22 chemokines within tumor microenvironment are related to accumulation of Foxp3+ regulatory T cells in gastric cancer. Int. J. Cancer 122, Oskarsson, T., Acharyya, S., Zhang, X.H., Vanharanta, S., Tavazoie, S.F., Morris, P.G., Downey, R.J., Manova-Todorova, K., Brogi, E., and Massagué, J. (2011). Breast cancer cells produce tenascin C as a metastatic niche component to colonize the lungs. Nat. Med. 17, Park, B.K., Zhang, H., Zeng, Q., Dai, J., Keller, E.T., Giordano, T., Gu, K., Shah, V., Pei, L., Zarbo, R.J., et al. (2007). NF-kappaB in breast cancer cells promotes osteolytic bone metastasis by inducing osteoclastogenesis via GM-CSF. Nat. Med. 13, Parmiani, G., Castelli, C., Pilla, L., Santinami, M., Colombo, M.P., and Rivoltini, L. (2007). Opposite immune functions of GM-CSF administered as vaccine adjuvant in cancer patients. Ann. Oncol. 18, Patsialou, A., Wyckoff, J., Wang, Y., Goswami, S., Stanley, E.R., and Condeelis, J.S. (2009). Invasion of human breast cancer cells in vivo requires both paracrine and autocrine loops involving the colony-stimulating factor-1 receptor. Cancer Res. 69, Pollard, J.W. (2004). Tumour-educated macrophages promote tumour progression and metastasis. Nat. Rev. Cancer 4, Qian, B.Z., Li, J., Zhang, H., Kitamura, T., Zhang, J., Campion, L.R., Kaiser, E.A., Snyder, L.A., and Pollard, J.W. (2011). CCL2 recruits inflammatory monocytes to facilitate breast-tumour metastasis. Nature 475, Serafini, P., Carbley, R., Noonan, K.A., Tan, G., Bronte, V., and Borrello, I. (2004). High-dose granulocyte-macrophage colony-stimulating factorproducing vaccines impair the immune response through the recruitment of myeloid suppressor cells. Cancer Res. 64, Slingluff, C.L., Jr., Petroni, G.R., Olson, W.C., Smolkin, M.E., Ross, M.I., Haas, N.B., Grosh, W.W., Boisvert, M.E., Kirkwood, J.M., and Chianese-Bullock, K.A. (2009). Effect of granulocyte/macrophage colony-stimulating factor on circulating CD8+ and CD4+ T-cell responses to a multipeptide melanoma vaccine: outcome of a multicenter randomized trial. Clin. Cancer Res. 15, Solinas, G., Schiarea, S., Liguori, M., Fabbri, M., Pesce, S., Zammataro, L., Pasqualini, F., Nebuloni, M., Chiabrando, C., Mantovani, A., and Allavena, P. (2010). Tumor-conditioned macrophages secrete migration-stimulating Cancer Cell 25, , May 12, 2014 ª2014 Elsevier Inc. 619
16 Cancer Cell GM-CSF-CCL18 Loop Promotes Breast Cancer Metastasis factor: a new marker for M2-polarization, influencing tumor cell motility. J. Immunol. 185, Takeda, K., Hatakeyama, K., Tsuchiya, Y., Rikiishi, H., and Kumagai, K. (1991). A correlation between GM-CSF gene expression and metastases in murine tumors. Int. J. Cancer 47, Thiery, J.P. (2002). Epithelial-mesenchymal transitions in tumour progression. Nat. Rev. Cancer 2, Thiery, J.P., Acloque, H., Huang, R.Y., and Nieto, M.A. (2009). Epithelialmesenchymal transitions in development and disease. Cell 139, Tsuchiya, Y., Igarashi, M., Suzuki, R., and Kumagai, K. (1988). Production of colony-stimulating factor by tumor cells and the factor-mediated induction of suppressor cells. J. Immunol. 141, Wang, X., Qi, Z., Wei, H., Tian, Z., and Sun, R. (2012). Characterization of human B cells in umbilical cord blood-transplanted NOD/SCID mice. Transpl. Immunol. 26, Wyckoff, J., Wang, W., Lin, E.Y., Wang, Y., Pixley, F., Stanley, E.R., Graf, T., Pollard, J.W., Segall, J., and Condeelis, J. (2004). A paracrine loop between tumor cells and macrophages is required for tumor cell migration in mammary tumors. Cancer Res. 64, Zhang, B., Yin, C., Li, H., Shi, L., Liu, N., Sun, Y., Lu, S., Liu, Y., Sun, L., Li, X., et al. (2013). Nir1 promotes invasion of breast cancer cells by binding to chemokine (C-C motif) ligand 18 through the PI3K/Akt/GSK3b/Snail signalling pathway. Eur. J. Cancer 49, Cancer Cell 25, , May 12, 2014 ª2014 Elsevier Inc.
17 Cancer Cell, Volume 25 Supplemental Information A Positive Feedback Loop between Mesenchymal-like Cancer Cells and Macrophages Is Essential to Breast Cancer Metastasis Shicheng Su, Qiang Liu, Jingqi Chen, Jianing Chen, Fei Chen, Chonghua He, Di Huang, Wei Wu, Ling Lin, Wei Huang, Jin Zhang, Xiuying Cui, Fang Zheng, Haiyan Li, Herui Yao, Fengxi Su, and Erwei Song
18 Supplementary Data
19 Figure S1, related to Figure 1 Breast cancer cells underwent EMT activate macrophages to a TAM-like phenotype. (A)Western blotting of E-cadherin and vimentin in epithelial-like breast cancer cell lines (MCF7, BT474 and T47D), mesenchymal-like cell lines (BT549, MDA-MB-436 and MDA-MB-231). (B) Western blotting of E-cadherin and vimentin of MCF7 or MCF7 undergone EMT that are induced by 5 ng/ml TGF-β, 10 ng/ml TNF-α or prolonged mammosphere culture. (C-E) Macrophages were obtained by culture of monocytes in DMEM medium supplemented with 10% heat inactivated human AB serum in the presence or absence of 30% CM from indicated cell lines in 24-well culture plates for 6 days. (C) Fluorescent CD68/DAPI staining in macrophages. Scale bar 20μm. (D) Expression of CD206/HLA-DR in macrophages. The histograms are representatives of 5 independent experiments of macrophages from 5 different donors. Numerical values denote the mean fluorescence intensity (MFI). (E) Cytokine levels in the media of macrophages (mean + s.e.m, n=5 independent experiments of macrophages from 5 different donors; *, p<0.05; **, p<0.01; ***, p<0.001, compared with control, p values were obtained using a two-tailed Student s t test).
20 Figure S2, related to Figure 2 Cytokines overexpressed in mesenchymal-like breast cancer cell lines. Levels of GM-CSF, IL-8, CCL2, GROα, GROβ and GROγ in indicated tumor CM were measured by ELISA (mean + s.e.m., n=3; *, p<0.05; **, p<0.01;***, p<0.001, compared with MCF7,p values were obtained using a two-tailed Student s t test).
21 Figure S3, related to Figure 3. Tumor-derived lactate abrogates the pro-inflammatory cytokine production from GM-CSF-activated macrophages. (A) Macrophages were obtained by culture of monocytes in culture medium in the presence or absence of 500 unit/ml IFN-γ, 50ng/ml GM-CSF or 20ng/ml IL-4 in 24-well culture plates for 6 days. Afterward, macrophages were cultured in culture medium in the presence or absence of 100ng/ml LPS in 24-well culture plates for 24h. Cytokine levels in the media were measured by ELISA (mean + s.e.m, n=4 independent experiments of macrophages from 4 different donors; *, p<0.05; **, p<0.01; ***, p<0.001, p values were obtained using a two-tailed Student s t test). (B) 231 cells were cultured in the presence or absence of 90mM oxamic acid for 5 days. Condition media (CM) were collected after the cells were cultured for another 24 hours
22 without oxamic acid. In addition, the lactate or the ph level in oxamic acid treated 231 cell CM were adjusted to the levels in untreated 231 cell CM by addition of lactate or 1% hydrochloric acid (HCl), respectively. Macrophages were obtained by culture of monocytes in culture medium in the presence or absence of 30% indicated 231 CM in 24-well culture plates for 6 days. The lactate levels and ph values of the macrophages culture were measured afterward(mean + s.e.m, n=4).
23 Figure S4, related to Figure 4 GM-CSF-activated macrophages induce EMT through CCL18. (A) Macrophages were obtained by culture of monocytes in culture medium in the presence or absence of 50ng/ml GM-CSF in 24-well culture plates for 6 days. Fluorescence intensity quantitation of E-cadherin and vimentin of MCF7 cells without or with the co-culture of control macrophages (Mφ), GM-CSF-activated macrophages alone or with control IgG,
24 anti-ccl18 neutralizing antibody, or rhccl18. Fluorescence intensity was normalized to control levels (mean + s.e.m, n=3 independent experiments of macrophages from 3 different donors; **, p<0.01;***, p<0.001, p values were obtained using a two-tailed Student s t test). (B) MCF7 cells were treated without or with rhccl20, or rhccl18 together with sirnas targeting GFP or PITPNM3. The nuclear/cytoplasmic ratio of their P65 immunofluorescence intensity was determined (mean + s.e.m, n=3; ***, p<0.001, p values were obtained using a two-tailed Student s t test ). (C) NF-kB luciferase reporter activity of CCL18-treated MCF7 cells pre-transfected with GFP-siRNA or PITPNM3-siRNAs (mean + s.e.m, n=3; *p<0.05,** p<0.01,*** p<0.001, p values were obtained using a two-tailed Student s t test). (D) Western blotting for E-cadherin and vimentin of MDA-MB-231 cells treated without or with DMSO, BAY or JSH-23. (E) The cytokine levels in the media of MDA-MB-231 cells treated as in D (mean + s.e.m., n=3; *** p<0.001 compared to untreated cells).
25
26 Figure S5, related to Figure 5 A positive feedback loop between GM-CSF from cancer cells and CCL18 from macrophages in vitro. (A & B) MCF7 CCL18 cells were re-plated alone or with macrophages in 12-well transwell apparatus with 0.4 m pore size with or without control IgG, anti-gm-csf or anti-ccl18 neutralizing antibody for 5 days. (A)The cells were immunstained for E-cadherin and vimentin. Fluorescence intensity was quantitated and normalized to the levels of cells cultured alone (0 day) (mean + s.e.m., n=4 independent experiments of macrophages from 4 different donors; **, p<0.01 compared with cells cultured alone; ##, p<0.01 compared with cells in the co-culture system without antibody addition at the same time points, p values were obtained using a two-tailed Student s t test). (B) Morphology analysis by CD68 immunofluorescent staining (upper panel) and surface markers detection by flow cytometry (lower panel) in macrophages alone or co-cultured with MCF7 CCL18 cells in the absence or presence of control IgG, anti-gm-csf or anti-ccl18 neutralizing antibody. The histograms are representative of 4 independent experiments of macrophages from 4 different donors. Numerical values denote the MFI. Scale bar, 20μm. (C) Macrophages obtained by culture of monocytes in culture medium with 50ng/ml GM-CSF for 6 days were re-plated alone or with untreated MCF7 in 12-well transwell apparatus with 0.4 m pore size with or without control IgG, anti-gm-csf or anti-ccl18 neutralizing antibody for 20 days. Representative pictures (upper panel) and fluorescence intensity quantitation (lower panel) of immunofluorescence analysis of E-cadherin and vimentin expression in MCF7 cells alone or in the co-culture system. Fluorescence intensity was normalized to the levels of cells cultured alone (0 day) (mean + s.e.m., n=4 independent experiments of macrophages from 4 different donors; **, p<0.01 compared with cells cultured alone; ##, p<0.01 compared with cells in the co-culture system without antibody addition at the same time points, p values were obtained using a two-tailed Student s t test). Scale bar, 20μm.
27
28 Figure S6, related to Figure 6 The GM-CSF-CCL18 loop promotes metastasis in a humanized mouse model. (A) Phenotypes of human immune cells in peripheral blood of mice. Human CD45 (a pan leukocyte marker), human CD14 (a monocyte marker), human CD3 (a T cell marker), human CD19 (a B cell marker) and human CD56 (a NK cell marker) were used to demonstrate the level of human immune cell reconstitution. Representative flow plots in each group are shown. Numbers in top left corner of the plots represent the percentages. (B) Percentages of CD45 + CD14 + human monocytes, CD45 + CD3 + human T cells, CD45 + CD19 + human B cells and CD45 + CD56 + human NK cells in peripheral blood of mice with the indicated treatments. Results shown are mean ± s.e.m. n=8 for each group. (C & D) Numbers of CD45 + total human leukocytes (C) CD45 + CD14 + human monocytes, CD45 + CD3 + human T cells, CD45 + CD19 + human B cells and CD45 + CD56 + human NK cells (D) per microliter in peripheral blood of mice. Results shown are mean ± s.e.m. n=8 for each group. (E) Phenotypes of human immune cells infiltrated in tumor xenografts of mice were evaluated by flow cytometry. Human CD45 (a pan leukocyte marker), human CD14 (a
29 monocyte/macrophage marker), human CD3 (a T cell marker), human CD19 (a B cell marker) and human CD56 (a NK cell marker) were used to demonstrate the level of human immune cell infiltration. Representative flow plots in each group are shown. Numbers in top left corner of the plots represent the percentages.
 Representative IHC staining of GM-CSF in normal breast tissue, breast fibroadenoma and breast invasive ductal carcinoma, showing that GM-CSF")
30 Figure S7, related to Figure 7 The GM-CSF and CCL18 loop is associated with metastasis in breast cancer patients. (A) Representative IHC staining of GM-CSF in normal breast tissue, breast fibroadenoma and breast invasive ductal carcinoma, showing that GM-CSF is highly expressed in the tumor
SUPPLEMENTARY INFORMATION
 DOI: 10.1038/ncb2607 Figure S1 Elf5 loss promotes EMT in mammary epithelium while Elf5 overexpression inhibits TGFβ induced EMT. (a, c) Different confocal slices through the Z stack image. (b, d) 3D rendering
DOI: 10.1038/ncb2607 Figure S1 Elf5 loss promotes EMT in mammary epithelium while Elf5 overexpression inhibits TGFβ induced EMT. (a, c) Different confocal slices through the Z stack image. (b, d) 3D rendering
Supplementary Fig. 1: ATM is phosphorylated in HER2 breast cancer cell lines. (A) ATM is phosphorylated in SKBR3 cells depending on ATM and HER2
 ATM is phosphorylated in SKBR3 cells depending on ATM and HER2") Supplementary Fig. 1: ATM is phosphorylated in HER2 breast cancer cell lines. (A) ATM is phosphorylated in SKBR3 cells depending on ATM and HER2 activity. Upper panel: Representative histograms for FACS
Supplementary Fig. 1: ATM is phosphorylated in HER2 breast cancer cell lines. (A) ATM is phosphorylated in SKBR3 cells depending on ATM and HER2 activity. Upper panel: Representative histograms for FACS
Supplementary Figures
 Supplementary Figures Supplementary Figure 1 DOT1L regulates the expression of epithelial and mesenchymal markers. (a) The expression levels and cellular localizations of EMT markers were confirmed by
Supplementary Figures Supplementary Figure 1 DOT1L regulates the expression of epithelial and mesenchymal markers. (a) The expression levels and cellular localizations of EMT markers were confirmed by
Type of file: PDF Size of file: 0 KB Title of file for HTML: Supplementary Information Description: Supplementary Figures
 Type of file: PDF Size of file: 0 KB Title of file for HTML: Supplementary Information Description: Supplementary Figures Supplementary Figure 1 mir-128-3p is highly expressed in chemoresistant, metastatic
Type of file: PDF Size of file: 0 KB Title of file for HTML: Supplementary Information Description: Supplementary Figures Supplementary Figure 1 mir-128-3p is highly expressed in chemoresistant, metastatic
(a) Significant biological processes (upper panel) and disease biomarkers (lower panel)
 Significant biological processes (upper panel) and disease biomarkers (lower panel)") Supplementary Figure 1. Functional enrichment analyses of secretomic proteins. (a) Significant biological processes (upper panel) and disease biomarkers (lower panel) 2 involved by hrab37-mediated secretory
Supplementary Figure 1. Functional enrichment analyses of secretomic proteins. (a) Significant biological processes (upper panel) and disease biomarkers (lower panel) 2 involved by hrab37-mediated secretory
Supplementary Fig. 1 p38 MAPK negatively regulates DC differentiation. (a) Western blot analysis of p38 isoform expression in BM cells, immature DCs
 Western blot analysis of p38 isoform expression in BM cells, immature DCs") Supplementary Fig. 1 p38 MAPK negatively regulates DC differentiation. (a) Western blot analysis of p38 isoform expression in BM cells, immature DCs (idcs) and mature DCs (mdcs). A myeloma cell line expressing
Supplementary Fig. 1 p38 MAPK negatively regulates DC differentiation. (a) Western blot analysis of p38 isoform expression in BM cells, immature DCs (idcs) and mature DCs (mdcs). A myeloma cell line expressing
ANGPTL2 increases bone metastasis of breast cancer cells through. Tetsuro Masuda, Motoyoshi Endo, Yutaka Yamamoto, Haruki Odagiri, Tsuyoshi
 Masuda et al. Supplementary information for ANGPTL2 increases bone metastasis of breast cancer cells through enhancing CXCR4 signaling Tetsuro Masuda, Motoyoshi Endo, Yutaka Yamamoto, Haruki Odagiri, Tsuyoshi
Masuda et al. Supplementary information for ANGPTL2 increases bone metastasis of breast cancer cells through enhancing CXCR4 signaling Tetsuro Masuda, Motoyoshi Endo, Yutaka Yamamoto, Haruki Odagiri, Tsuyoshi
Effective activity of cytokine-induced killer cells against autologous metastatic melanoma including cells with stemness features
 Effective activity of cytokine-induced killer cells against autologous metastatic melanoma including cells with stemness features Loretta Gammaitoni, Lidia Giraudo, Valeria Leuci, et al. Clin Cancer Res
Effective activity of cytokine-induced killer cells against autologous metastatic melanoma including cells with stemness features Loretta Gammaitoni, Lidia Giraudo, Valeria Leuci, et al. Clin Cancer Res
Supplementary Figures
 Supplementary Figures Supplementary Figure 1. Confirmation of Dnmt1 conditional knockout out mice. a, Representative images of sorted stem (Lin - CD49f high CD24 + ), luminal (Lin - CD49f low CD24 + )
Supplementary Figures Supplementary Figure 1. Confirmation of Dnmt1 conditional knockout out mice. a, Representative images of sorted stem (Lin - CD49f high CD24 + ), luminal (Lin - CD49f low CD24 + )
An epithelial-to-mesenchymal transition-inducing potential of. granulocyte macrophage colony-stimulating factor in colon. cancer
 An epithelial-to-mesenchymal transition-inducing potential of granulocyte macrophage colony-stimulating factor in colon cancer Yaqiong Chen, Zhi Zhao, Yu Chen, Zhonglin Lv, Xin Ding, Renxi Wang, He Xiao,
An epithelial-to-mesenchymal transition-inducing potential of granulocyte macrophage colony-stimulating factor in colon cancer Yaqiong Chen, Zhi Zhao, Yu Chen, Zhonglin Lv, Xin Ding, Renxi Wang, He Xiao,
SHREE ET AL, SUPPLEMENTAL MATERIALS. (A) Workflow for tumor cell line derivation and orthotopic implantation.
 Workflow for tumor cell line derivation and orthotopic implantation.") SHREE ET AL, SUPPLEMENTAL MATERIALS SUPPLEMENTAL FIGURE AND TABLE LEGENDS Supplemental Figure 1. Derivation and characterization of TS1-TGL and TS2-TGL PyMT cell lines and development of an orthotopic
SHREE ET AL, SUPPLEMENTAL MATERIALS SUPPLEMENTAL FIGURE AND TABLE LEGENDS Supplemental Figure 1. Derivation and characterization of TS1-TGL and TS2-TGL PyMT cell lines and development of an orthotopic
Nature Immunology: doi: /ni Supplementary Figure 1. Huwe1 has high expression in HSCs and is necessary for quiescence.
 Supplementary Figure 1 Huwe1 has high expression in HSCs and is necessary for quiescence. (a) Heat map visualizing expression of genes with a known function in ubiquitin-mediated proteolysis (KEGG: Ubiquitin
Supplementary Figure 1 Huwe1 has high expression in HSCs and is necessary for quiescence. (a) Heat map visualizing expression of genes with a known function in ubiquitin-mediated proteolysis (KEGG: Ubiquitin
B16-F10 (Mus musculus skin melanoma), NCI-H460 (human non-small cell lung cancer
, NCI-H460 (human non-small cell lung cancer") Electronic Supplementary Material (ESI) for ChemComm. This journal is The Royal Society of Chemistry 2017 Experimental Methods Cell culture B16-F10 (Mus musculus skin melanoma), NCI-H460 (human non-small
Electronic Supplementary Material (ESI) for ChemComm. This journal is The Royal Society of Chemistry 2017 Experimental Methods Cell culture B16-F10 (Mus musculus skin melanoma), NCI-H460 (human non-small
Supplementary Materials for
 www.sciencesignaling.org/cgi/content/full/8/381/ra59/dc1 Supplementary Materials for Analysis of single-cell cytokine secretion reveals a role for paracrine signaling in coordinating macrophage responses
www.sciencesignaling.org/cgi/content/full/8/381/ra59/dc1 Supplementary Materials for Analysis of single-cell cytokine secretion reveals a role for paracrine signaling in coordinating macrophage responses
Supplemental Figure 1
 Supplemental Figure 1 1a 1c PD-1 MFI fold change 6 5 4 3 2 1 IL-1α IL-2 IL-4 IL-6 IL-1 IL-12 IL-13 IL-15 IL-17 IL-18 IL-21 IL-23 IFN-α Mut Human PD-1 promoter SBE-D 5 -GTCTG- -1.2kb SBE-P -CAGAC- -1.kb
Supplemental Figure 1 1a 1c PD-1 MFI fold change 6 5 4 3 2 1 IL-1α IL-2 IL-4 IL-6 IL-1 IL-12 IL-13 IL-15 IL-17 IL-18 IL-21 IL-23 IFN-α Mut Human PD-1 promoter SBE-D 5 -GTCTG- -1.2kb SBE-P -CAGAC- -1.kb
Tumor Associated Macrophages as a Novel Target for Cancer Therapy
 Tumor mass Tumor Associated Macrophage Tumor Associated Macrophages as a Novel Target for Cancer Therapy This booklet contains forward-looking statements that are based on Amgen s current expectations
Tumor mass Tumor Associated Macrophage Tumor Associated Macrophages as a Novel Target for Cancer Therapy This booklet contains forward-looking statements that are based on Amgen s current expectations
Supplementary Figure 1. HOPX is hypermethylated in NPC. (a) Methylation levels of HOPX in Normal (n = 24) and NPC (n = 24) tissues from the
 Methylation levels of HOPX in Normal (n = 24) and NPC (n = 24) tissues from the") Supplementary Figure 1. HOPX is hypermethylated in NPC. (a) Methylation levels of HOPX in Normal (n = 24) and NPC (n = 24) tissues from the genome-wide methylation microarray data. Mean ± s.d.; Student
Supplementary Figure 1. HOPX is hypermethylated in NPC. (a) Methylation levels of HOPX in Normal (n = 24) and NPC (n = 24) tissues from the genome-wide methylation microarray data. Mean ± s.d.; Student
Supplementary Table S1. Tumor samples used for analysis Tumor size (cm) BNG (grade) ERα PR. pn-
 BNG (grade) ERα PR. pn-") Supplementary Table S1. Tumor samples used for analysis Sample# Age Tumor size (cm) pn- Stage Stage BNG (grade) ERα PR HER2 (FISH) Triple negative T1 46 3 N1a III 2 Pos Neg N T2 58 1 N(i-) I 3 Pos Neg
Supplementary Table S1. Tumor samples used for analysis Sample# Age Tumor size (cm) pn- Stage Stage BNG (grade) ERα PR HER2 (FISH) Triple negative T1 46 3 N1a III 2 Pos Neg N T2 58 1 N(i-) I 3 Pos Neg
Supplementary Materials for
 www.sciencetranslationalmedicine.org/cgi/content/full/8/352/352ra110/dc1 Supplementary Materials for Spatially selective depletion of tumor-associated regulatory T cells with near-infrared photoimmunotherapy
www.sciencetranslationalmedicine.org/cgi/content/full/8/352/352ra110/dc1 Supplementary Materials for Spatially selective depletion of tumor-associated regulatory T cells with near-infrared photoimmunotherapy
Supplementary Materials for
 www.sciencesignaling.org/cgi/content/full/7/310/ra11/dc1 Supplementary Materials for STAT3 Induction of mir-146b Forms a Feedback Loop to Inhibit the NF-κB to IL-6 Signaling Axis and STAT3-Driven Cancer
www.sciencesignaling.org/cgi/content/full/7/310/ra11/dc1 Supplementary Materials for STAT3 Induction of mir-146b Forms a Feedback Loop to Inhibit the NF-κB to IL-6 Signaling Axis and STAT3-Driven Cancer
Supplementary Figure 1. Deletion of Smad3 prevents B16F10 melanoma invasion and metastasis in a mouse s.c. tumor model.
 A B16F1 s.c. Lung LN Distant lymph nodes Colon B B16F1 s.c. Supplementary Figure 1. Deletion of Smad3 prevents B16F1 melanoma invasion and metastasis in a mouse s.c. tumor model. Highly invasive growth
A B16F1 s.c. Lung LN Distant lymph nodes Colon B B16F1 s.c. Supplementary Figure 1. Deletion of Smad3 prevents B16F1 melanoma invasion and metastasis in a mouse s.c. tumor model. Highly invasive growth
Supplementary Figure S1 Expression of mir-181b in EOC (A) Kaplan-Meier
 Kaplan-Meier") Supplementary Figure S1 Expression of mir-181b in EOC (A) Kaplan-Meier curves for progression-free survival (PFS) and overall survival (OS) in a cohort of patients (N=52) with stage III primary ovarian
Supplementary Figure S1 Expression of mir-181b in EOC (A) Kaplan-Meier curves for progression-free survival (PFS) and overall survival (OS) in a cohort of patients (N=52) with stage III primary ovarian
Supplemental Information. CD4 + CD25 + Foxp3 + Regulatory T Cells Promote. Th17 Cells In Vitro and Enhance Host Resistance
 Immunity, Volume 34 Supplemental Information D4 + D25 + + Regulatory T ells Promote Th17 ells In Vitro and Enhance Host Resistance in Mouse andida albicans Th17 ell Infection Model Pushpa Pandiyan, Heather
Immunity, Volume 34 Supplemental Information D4 + D25 + + Regulatory T ells Promote Th17 ells In Vitro and Enhance Host Resistance in Mouse andida albicans Th17 ell Infection Model Pushpa Pandiyan, Heather
mir-509-5p and mir-1243 increase the sensitivity to gemcitabine by inhibiting
 mir-509-5p and mir-1243 increase the sensitivity to gemcitabine by inhibiting epithelial-mesenchymal transition in pancreatic cancer Hidekazu Hiramoto, M.D. 1,3, Tomoki Muramatsu, Ph.D. 1, Daisuke Ichikawa,
mir-509-5p and mir-1243 increase the sensitivity to gemcitabine by inhibiting epithelial-mesenchymal transition in pancreatic cancer Hidekazu Hiramoto, M.D. 1,3, Tomoki Muramatsu, Ph.D. 1, Daisuke Ichikawa,
TEB. Id4 p63 DAPI Merge. Id4 CK8 DAPI Merge
 a Duct TEB b Id4 p63 DAPI Merge Id4 CK8 DAPI Merge c d e Supplementary Figure 1. Identification of Id4-positive MECs and characterization of the Comma-D model. (a) IHC analysis of ID4 expression in the
a Duct TEB b Id4 p63 DAPI Merge Id4 CK8 DAPI Merge c d e Supplementary Figure 1. Identification of Id4-positive MECs and characterization of the Comma-D model. (a) IHC analysis of ID4 expression in the
5K ALDEFLUOR-positive/ CXCR1-negative. 5K ALDEFLUOR-positive/ CXCR1-positive BAAA BAAA CXCR1-APC BAAA BAAA CXCR1-APC
 A +DEAB -DEAB K ALDEFLUOR-positive/ CXCR-negative BAAA BAAA CXCR-APC B +DEAB -DEAB K ALDEFLUOR-positive/ CXCR-positive BAAA BAAA CXCR-APC C Supplemental Figure. Tumorigenicity of the ALDEFLUOR-positive/CXCR-positive
A +DEAB -DEAB K ALDEFLUOR-positive/ CXCR-negative BAAA BAAA CXCR-APC B +DEAB -DEAB K ALDEFLUOR-positive/ CXCR-positive BAAA BAAA CXCR-APC C Supplemental Figure. Tumorigenicity of the ALDEFLUOR-positive/CXCR-positive
Transfer protocol of human HSC into NOG mice
 Transfer protocol of human HSC into NOG mice Mice: Adult NOG mice are aged 8-12 weeks. Newborn mice are 1 2 days old. 8-12 week old NOG mice irradiated with 2.5 Gy Intravenous transfer of 1-0.5 x 10 5
Transfer protocol of human HSC into NOG mice Mice: Adult NOG mice are aged 8-12 weeks. Newborn mice are 1 2 days old. 8-12 week old NOG mice irradiated with 2.5 Gy Intravenous transfer of 1-0.5 x 10 5
Endogenous TNFα orchestrates the trafficking of neutrophils into and within lymphatic vessels during acute inflammation
 SUPPLEMENTARY INFORMATION Endogenous TNFα orchestrates the trafficking of neutrophils into and within lymphatic vessels during acute inflammation Samantha Arokiasamy 1,2, Christian Zakian 1, Jessica Dilliway
SUPPLEMENTARY INFORMATION Endogenous TNFα orchestrates the trafficking of neutrophils into and within lymphatic vessels during acute inflammation Samantha Arokiasamy 1,2, Christian Zakian 1, Jessica Dilliway
COPD lungs show an attached stratified mucus layer that separate. bacteria from the epithelial cells resembling the protective colonic
 COPD lungs show an attached stratified mucus layer that separate bacteria from the epithelial cells resembling the protective colonic mucus SUPPLEMENTARY TABLES AND FIGURES Tables S1 S8, page 1 and separate
COPD lungs show an attached stratified mucus layer that separate bacteria from the epithelial cells resembling the protective colonic mucus SUPPLEMENTARY TABLES AND FIGURES Tables S1 S8, page 1 and separate
Supplementary Figure 1. Characterization of NMuMG-ErbB2 and NIC breast cancer cells expressing shrnas targeting LPP. NMuMG-ErbB2 cells (a) and NIC
 and NIC") Supplementary Figure 1. Characterization of NMuMG-ErbB2 and NIC breast cancer cells expressing shrnas targeting LPP. NMuMG-ErbB2 cells (a) and NIC cells (b) were engineered to stably express either a LucA-shRNA
Supplementary Figure 1. Characterization of NMuMG-ErbB2 and NIC breast cancer cells expressing shrnas targeting LPP. NMuMG-ErbB2 cells (a) and NIC cells (b) were engineered to stably express either a LucA-shRNA
Supplementary Figure (OH) 22 nanoparticles did not affect cell viability and apoposis. MDA-MB-231, MCF-7, MCF-10A and BT549 cells were
 22 nanoparticles did not affect cell viability and apoposis. MDA-MB-231, MCF-7, MCF-10A and BT549 cells were") Supplementary Figure 1. Gd@C 82 (OH) 22 nanoparticles did not affect cell viability and apoposis. MDA-MB-231, MCF-7, MCF-10A and BT549 cells were treated with PBS, Gd@C 82 (OH) 22, C 60 (OH) 22 or GdCl
Supplementary Figure 1. Gd@C 82 (OH) 22 nanoparticles did not affect cell viability and apoposis. MDA-MB-231, MCF-7, MCF-10A and BT549 cells were treated with PBS, Gd@C 82 (OH) 22, C 60 (OH) 22 or GdCl
Suppl Video: Tumor cells (green) and monocytes (white) are seeded on a confluent endothelial
 and monocytes (white) are seeded on a confluent endothelial") Supplementary Information Häuselmann et al. Monocyte induction of E-selectin-mediated endothelial activation releases VE-cadherin junctions to promote tumor cell extravasation in the metastasis cascade
Supplementary Information Häuselmann et al. Monocyte induction of E-selectin-mediated endothelial activation releases VE-cadherin junctions to promote tumor cell extravasation in the metastasis cascade
Supplementary Figure 1: Digitoxin induces apoptosis in primary human melanoma cells but not in normal melanocytes, which express lower levels of the
 Supplementary Figure 1: Digitoxin induces apoptosis in primary human melanoma cells but not in normal melanocytes, which express lower levels of the cardiac glycoside target, ATP1A1. (a) The percentage
Supplementary Figure 1: Digitoxin induces apoptosis in primary human melanoma cells but not in normal melanocytes, which express lower levels of the cardiac glycoside target, ATP1A1. (a) The percentage
Supplementary Figure 1: STAT3 suppresses Kras-induced lung tumorigenesis
 Supplementary Figure 1: STAT3 suppresses Kras-induced lung tumorigenesis (a) Immunohistochemical (IHC) analysis of tyrosine 705 phosphorylation status of STAT3 (P- STAT3) in tumors and stroma (all-time
Supplementary Figure 1: STAT3 suppresses Kras-induced lung tumorigenesis (a) Immunohistochemical (IHC) analysis of tyrosine 705 phosphorylation status of STAT3 (P- STAT3) in tumors and stroma (all-time
well for 2 h at rt. Each dot represents an individual mouse and bar is the mean ±
 Supplementary data: Control DC Blimp-1 ko DC 8 6 4 2-2 IL-1β p=.5 medium 8 6 4 2 IL-2 Medium p=.16 8 6 4 2 IL-6 medium p=.3 5 4 3 2 1-1 medium IL-1 n.s. 25 2 15 1 5 IL-12(p7) p=.15 5 IFNγ p=.65 4 3 2 1
Supplementary data: Control DC Blimp-1 ko DC 8 6 4 2-2 IL-1β p=.5 medium 8 6 4 2 IL-2 Medium p=.16 8 6 4 2 IL-6 medium p=.3 5 4 3 2 1-1 medium IL-1 n.s. 25 2 15 1 5 IL-12(p7) p=.15 5 IFNγ p=.65 4 3 2 1
SUPPLEMENTARY INFORMATION
 DOI: 1.138/ncb3355 a S1A8 + cells/ total.1.8.6.4.2 b S1A8/?-Actin c % T-cell proliferation 3 25 2 15 1 5 T cells Supplementary Figure 1 Inter-tumoral heterogeneity of MDSC accumulation in mammary tumor
DOI: 1.138/ncb3355 a S1A8 + cells/ total.1.8.6.4.2 b S1A8/?-Actin c % T-cell proliferation 3 25 2 15 1 5 T cells Supplementary Figure 1 Inter-tumoral heterogeneity of MDSC accumulation in mammary tumor
Supplemental Table 1. Primer sequences for transcript analysis
 Supplemental Table 1. Primer sequences for transcript analysis Primer Sequence (5 3 ) Primer Sequence (5 3 ) Mmp2 Forward CCCGTGTGGCCCTC Mmp15 Forward CGGGGCTGGCT Reverse GCTCTCCCGGTTTC Reverse CCTGGTGTGCCTGCTC
Supplemental Table 1. Primer sequences for transcript analysis Primer Sequence (5 3 ) Primer Sequence (5 3 ) Mmp2 Forward CCCGTGTGGCCCTC Mmp15 Forward CGGGGCTGGCT Reverse GCTCTCCCGGTTTC Reverse CCTGGTGTGCCTGCTC
MATERIALS AND METHODS. Neutralizing antibodies specific to mouse Dll1, Dll4, J1 and J2 were prepared as described. 1,2 All
 MATERIALS AND METHODS Antibodies (Abs), flow cytometry analysis and cell lines Neutralizing antibodies specific to mouse Dll1, Dll4, J1 and J2 were prepared as described. 1,2 All other antibodies used
MATERIALS AND METHODS Antibodies (Abs), flow cytometry analysis and cell lines Neutralizing antibodies specific to mouse Dll1, Dll4, J1 and J2 were prepared as described. 1,2 All other antibodies used
Blocking antibodies and peptides. Rat anti-mouse PD-1 (29F.1A12, rat IgG2a, k), PD-
, PD-") Supplementary Methods Blocking antibodies and peptides. Rat anti-mouse PD-1 (29F.1A12, rat IgG2a, k), PD- L1 (10F.9G2, rat IgG2b, k), and PD-L2 (3.2, mouse IgG1) have been described (24). Anti-CTLA-4 (clone
Supplementary Methods Blocking antibodies and peptides. Rat anti-mouse PD-1 (29F.1A12, rat IgG2a, k), PD- L1 (10F.9G2, rat IgG2b, k), and PD-L2 (3.2, mouse IgG1) have been described (24). Anti-CTLA-4 (clone
Supplementary Figure 1. The mir-182 binding site of SMAD7 3 UTR and the. mutated sequence.
 Supplementary Figure 1. The mir-182 binding site of SMAD7 3 UTR and the mutated sequence. 1 Supplementary Figure 2. Expression of mir-182 and SMAD7 in various cell lines. (A) Basal levels of mir-182 expression
Supplementary Figure 1. The mir-182 binding site of SMAD7 3 UTR and the mutated sequence. 1 Supplementary Figure 2. Expression of mir-182 and SMAD7 in various cell lines. (A) Basal levels of mir-182 expression
SSM signature genes are highly expressed in residual scar tissues after preoperative radiotherapy of rectal cancer.
 Supplementary Figure 1 SSM signature genes are highly expressed in residual scar tissues after preoperative radiotherapy of rectal cancer. Scatter plots comparing expression profiles of matched pretreatment
Supplementary Figure 1 SSM signature genes are highly expressed in residual scar tissues after preoperative radiotherapy of rectal cancer. Scatter plots comparing expression profiles of matched pretreatment
Nature Genetics: doi: /ng Supplementary Figure 1. Phenotypic characterization of MES- and ADRN-type cells.
 Supplementary Figure 1 Phenotypic characterization of MES- and ADRN-type cells. (a) Bright-field images showing cellular morphology of MES-type (691-MES, 700-MES, 717-MES) and ADRN-type (691-ADRN, 700-
Supplementary Figure 1 Phenotypic characterization of MES- and ADRN-type cells. (a) Bright-field images showing cellular morphology of MES-type (691-MES, 700-MES, 717-MES) and ADRN-type (691-ADRN, 700-
Supplementary Figure 1. Double-staining immunofluorescence analysis of invasive colon and breast cancers. Specimens from invasive ductal breast
 Supplementary Figure 1. Double-staining immunofluorescence analysis of invasive colon and breast cancers. Specimens from invasive ductal breast carcinoma (a) and colon adenocarcinoma (b) were staining
Supplementary Figure 1. Double-staining immunofluorescence analysis of invasive colon and breast cancers. Specimens from invasive ductal breast carcinoma (a) and colon adenocarcinoma (b) were staining
Fang et al. NMuMG. PyVmT unstained Anti-CCR2-PE MDA-MB MCF MCF10A
 A NMuMG PyVmT 16.5+.5 47.+7.2 Fang et al. unstained Anti-CCR2-PE 4T1 Control 37.6+6.3 56.1+.65 MCF1A 16.1+3. MCF-7 3.1+5.4 MDA-M-231 42.1+5.5 unstained Secondary antibody only Anti-CCR2 SUPPLEMENTAL FIGURE
A NMuMG PyVmT 16.5+.5 47.+7.2 Fang et al. unstained Anti-CCR2-PE 4T1 Control 37.6+6.3 56.1+.65 MCF1A 16.1+3. MCF-7 3.1+5.4 MDA-M-231 42.1+5.5 unstained Secondary antibody only Anti-CCR2 SUPPLEMENTAL FIGURE
Journal Club WS 2012/13 Stefanie Nickl
 Journal Club WS 2012/13 Stefanie Nickl Background Mesenchymal Stem Cells First isolation from bone marrow 30 ys ago Isolation from: spleen, heart, skeletal muscle, synovium, amniotic fluid, dental pulp,
Journal Club WS 2012/13 Stefanie Nickl Background Mesenchymal Stem Cells First isolation from bone marrow 30 ys ago Isolation from: spleen, heart, skeletal muscle, synovium, amniotic fluid, dental pulp,
TMA-VARESE COHORT-1 TMA-BERN COHORT-2
 Supplementary Figure 1 TMA-VARESE COHORT-1 TOTAL SAMPLES #5 GLEASON SCORE Number Percentage 6 16 32% = 7 17 34% >7 17 34% TUMOR STAGE T2C 28 56% T3A- 21 42% T3C-T4 1 2% NODE STATUS N 42 84% N1 8 16% PSA
Supplementary Figure 1 TMA-VARESE COHORT-1 TOTAL SAMPLES #5 GLEASON SCORE Number Percentage 6 16 32% = 7 17 34% >7 17 34% TUMOR STAGE T2C 28 56% T3A- 21 42% T3C-T4 1 2% NODE STATUS N 42 84% N1 8 16% PSA
Targeting tumour associated macrophages in anti-cancer therapies. Annamaria Gal Seminar Series on Drug Discovery Budapest 5 January 2018
 Targeting tumour associated macrophages in anti-cancer therapies Annamaria Gal Seminar Series on Drug Discovery Budapest 5 January 2018 Macrophages: Professional phagocytes of the myeloid lineage APC,
Targeting tumour associated macrophages in anti-cancer therapies Annamaria Gal Seminar Series on Drug Discovery Budapest 5 January 2018 Macrophages: Professional phagocytes of the myeloid lineage APC,
CCL18 from Tumor-Associated Macrophages Promotes Breast Cancer Metastasis via PITPNM3
 Article CCL18 from Tumor-Associated Macrophages Promotes Breast Cancer Metastasis via PITPNM3 Jingqi Chen, 1,2,5 Yandan Yao, 1,5 Chang Gong, 1,5 Fengyan Yu, 1,5 Shicheng Su, 1 Jianing Chen, 1 Bodu Liu,
Article CCL18 from Tumor-Associated Macrophages Promotes Breast Cancer Metastasis via PITPNM3 Jingqi Chen, 1,2,5 Yandan Yao, 1,5 Chang Gong, 1,5 Fengyan Yu, 1,5 Shicheng Su, 1 Jianing Chen, 1 Bodu Liu,
CD34+ Cells: A Comparison of Stem and Progenitor Cells in Cord Blood, Peripheral Blood, and the Bone Marrow
 White Paper September 2016 CD34+ Cells: A Comparison of Stem and Progenitor Cells in Cord Blood, Peripheral Blood, and the Bone Marrow Lily C. Trajman, PhD Introduction: Hematopoietic Stem Cells (HSCs)
White Paper September 2016 CD34+ Cells: A Comparison of Stem and Progenitor Cells in Cord Blood, Peripheral Blood, and the Bone Marrow Lily C. Trajman, PhD Introduction: Hematopoietic Stem Cells (HSCs)
Supplementary Figure 1
 Supplementary Figure 1 AAV-GFP injection in the MEC of the mouse brain C57Bl/6 mice at 4 months of age were injected with AAV-GFP into the MEC and sacrificed at 7 days post injection (dpi). (a) Brains
Supplementary Figure 1 AAV-GFP injection in the MEC of the mouse brain C57Bl/6 mice at 4 months of age were injected with AAV-GFP into the MEC and sacrificed at 7 days post injection (dpi). (a) Brains
Material and Methods. Flow Cytometry Analyses:
 Material and Methods Flow Cytometry Analyses: Immunostaining of breast cancer cells for HER2 was performed by incubating cells with anti- HER2/neu APC (Biosciences, Cat# 340554), anti-her2/neu PE (Biosciences,
Material and Methods Flow Cytometry Analyses: Immunostaining of breast cancer cells for HER2 was performed by incubating cells with anti- HER2/neu APC (Biosciences, Cat# 340554), anti-her2/neu PE (Biosciences,
ECM1 controls T H 2 cell egress from lymph nodes through re-expression of S1P 1
 ZH, Li et al, page 1 ECM1 controls T H 2 cell egress from lymph nodes through re-expression of S1P 1 Zhenhu Li 1,4,Yuan Zhang 1,4, Zhiduo Liu 1, Xiaodong Wu 1, Yuhan Zheng 1, Zhiyun Tao 1, Kairui Mao 1,
ZH, Li et al, page 1 ECM1 controls T H 2 cell egress from lymph nodes through re-expression of S1P 1 Zhenhu Li 1,4,Yuan Zhang 1,4, Zhiduo Liu 1, Xiaodong Wu 1, Yuhan Zheng 1, Zhiyun Tao 1, Kairui Mao 1,
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature12215 Supplementary Figure 1. The effects of full and dissociated GR agonists in supporting BFU-E self-renewal divisions. BFU-Es were cultured in self-renewal medium with indicated GR
doi:10.1038/nature12215 Supplementary Figure 1. The effects of full and dissociated GR agonists in supporting BFU-E self-renewal divisions. BFU-Es were cultured in self-renewal medium with indicated GR
(A) Dose response curves of HMLE_shGFP (blue circle), HMLE_shEcad (red square),
 Dose response curves of HMLE_shGFP (blue circle), HMLE_shEcad (red square),") Supplementary Figures and Tables Figure S1. Validation of EMT-selective small molecules (A) Dose response curves of HMLE_shGFP (blue circle), HMLE_shEcad (red square), and HMLE_Twist (black diamond) cells
Supplementary Figures and Tables Figure S1. Validation of EMT-selective small molecules (A) Dose response curves of HMLE_shGFP (blue circle), HMLE_shEcad (red square), and HMLE_Twist (black diamond) cells
Inhibition of TGFβ enhances chemotherapy action against triple negative breast cancer by abrogation of
 SUPPLEMENTAL DATA Inhibition of TGFβ enhances chemotherapy action against triple negative breast cancer by abrogation of cancer stem cells and interleukin-8 Neil E. Bhola 1, Justin M. Balko 1, Teresa C.
SUPPLEMENTAL DATA Inhibition of TGFβ enhances chemotherapy action against triple negative breast cancer by abrogation of cancer stem cells and interleukin-8 Neil E. Bhola 1, Justin M. Balko 1, Teresa C.
Bmi-1 regulates stem cell-like properties of gastric cancer cells via modulating mirnas
 Wang et al. Journal of Hematology & Oncology (2016) 9:90 DOI 10.1186/s13045-016-0323-9 RESEARCH Bmi-1 regulates stem cell-like properties of gastric cancer cells via modulating mirnas Open Access Xiaofeng
Wang et al. Journal of Hematology & Oncology (2016) 9:90 DOI 10.1186/s13045-016-0323-9 RESEARCH Bmi-1 regulates stem cell-like properties of gastric cancer cells via modulating mirnas Open Access Xiaofeng
Extended Mammosphere Culture of Human Breast Cancer Cells
 Extended Mammosphere Culture of Human Breast Cancer Cells Application Note The PromoCell 3D Tumorsphere Medium XF The PromoCell 3D Tumorsphere Medium XF has been designed to meet your requirements for
Extended Mammosphere Culture of Human Breast Cancer Cells Application Note The PromoCell 3D Tumorsphere Medium XF The PromoCell 3D Tumorsphere Medium XF has been designed to meet your requirements for
Contents 1 The Windows of Susceptibility to Breast Cancer 2 The So Called Pre-Neoplastic Lesions and Carcinoma In Situ
 Contents 1 The Windows of Susceptibility to Breast Cancer... 1 1.1 Introduction... 1 1.2 Risk Factor and Etiological Agents... 2 1.3 The Concept of the Windows of Susceptibility to Carcinogenesis... 5
Contents 1 The Windows of Susceptibility to Breast Cancer... 1 1.1 Introduction... 1 1.2 Risk Factor and Etiological Agents... 2 1.3 The Concept of the Windows of Susceptibility to Carcinogenesis... 5
Pathologic Stage. Lymph node Stage
 ASC ASC a c Patient ID BMI Age Gleason score Non-obese PBMC 1 22.1 81 6 (3+3) PBMC 2 21.9 6 6 (3+3) PBMC 3 22 84 8 (4+4) PBMC 4 24.6 68 7 (3+4) PBMC 24. 6 (3+3) PBMC 6 24.7 73 7 (3+4) PBMC 7 23. 67 7 (3+4)
ASC ASC a c Patient ID BMI Age Gleason score Non-obese PBMC 1 22.1 81 6 (3+3) PBMC 2 21.9 6 6 (3+3) PBMC 3 22 84 8 (4+4) PBMC 4 24.6 68 7 (3+4) PBMC 24. 6 (3+3) PBMC 6 24.7 73 7 (3+4) PBMC 7 23. 67 7 (3+4)
Figure S1: Effects on haptotaxis are independent of effects on cell velocity A)
") Supplemental Figures Figure S1: Effects on haptotaxis are independent of effects on cell velocity A) Velocity of MV D7 fibroblasts expressing different GFP-tagged Ena/VASP family proteins in the haptotaxis
Supplemental Figures Figure S1: Effects on haptotaxis are independent of effects on cell velocity A) Velocity of MV D7 fibroblasts expressing different GFP-tagged Ena/VASP family proteins in the haptotaxis
Epstein-Barr virus driven promoter hypermethylated genes in gastric cancer
 RESEARCH FUND FOR THE CONTROL OF INFECTIOUS DISEASES Epstein-Barr virus driven promoter hypermethylated genes in gastric cancer J Yu *, KF To, QY Liang K e y M e s s a g e s 1. Somatostatin receptor 1
RESEARCH FUND FOR THE CONTROL OF INFECTIOUS DISEASES Epstein-Barr virus driven promoter hypermethylated genes in gastric cancer J Yu *, KF To, QY Liang K e y M e s s a g e s 1. Somatostatin receptor 1
Supplementary Figures
 Inhibition of Pulmonary Anti Bacterial Defense by IFN γ During Recovery from Influenza Infection By Keer Sun and Dennis W. Metzger Supplementary Figures d a Ly6G Percentage survival f 1 75 5 1 25 1 5 1
Inhibition of Pulmonary Anti Bacterial Defense by IFN γ During Recovery from Influenza Infection By Keer Sun and Dennis W. Metzger Supplementary Figures d a Ly6G Percentage survival f 1 75 5 1 25 1 5 1
TITLE: Notch in Pathological Angiogenesis and Lymphangiogenesis
 Award Number: W81XWH-10-1-0304 TITLE: Notch in Pathological Angiogenesis and Lymphangiogenesis PRINCIPAL INVESTIGATOR: Minji Kim CONTRACTING ORGANIZATION: Columbia University New York, NY 10032 REPORT
Award Number: W81XWH-10-1-0304 TITLE: Notch in Pathological Angiogenesis and Lymphangiogenesis PRINCIPAL INVESTIGATOR: Minji Kim CONTRACTING ORGANIZATION: Columbia University New York, NY 10032 REPORT
(A) RT-PCR for components of the Shh/Gli pathway in normal fetus cell (MRC-5) and a
 RT-PCR for components of the Shh/Gli pathway in normal fetus cell (MRC-5) and a") Supplementary figure legends Supplementary Figure 1. Expression of Shh signaling components in a panel of gastric cancer. (A) RT-PCR for components of the Shh/Gli pathway in normal fetus cell (MRC-5) and
Supplementary figure legends Supplementary Figure 1. Expression of Shh signaling components in a panel of gastric cancer. (A) RT-PCR for components of the Shh/Gli pathway in normal fetus cell (MRC-5) and
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature11095 Supplementary Table 1. Summary of the binding between Angptls and various Igdomain containing receptors as determined by flow cytometry analysis. The results were summarized from
doi:10.1038/nature11095 Supplementary Table 1. Summary of the binding between Angptls and various Igdomain containing receptors as determined by flow cytometry analysis. The results were summarized from
Supplemental Table S1
 Supplemental Table S. Tumorigenicity and metastatic potential of 44SQ cell subpopulations a Tumorigenicity b Average tumor volume (mm ) c Lung metastasis d CD high /4 8. 8/ CD low /4 6./ a Mice were injected
Supplemental Table S. Tumorigenicity and metastatic potential of 44SQ cell subpopulations a Tumorigenicity b Average tumor volume (mm ) c Lung metastasis d CD high /4 8. 8/ CD low /4 6./ a Mice were injected
SUPPLEMENTARY INFORMATION. CXCR4 inhibitors could benefit to HER2 but not to Triple-Negative. breast cancer patients
 SUPPLEMENTARY INFORMATION CXCR4 inhibitors could benefit to HER2 but not to Triple-Negative breast cancer patients Lefort S. 1,2, Thuleau A. 3, Kieffer Y. 1,2, Sirven P. 1,2, Bieche I. 4, Marangoni E.
SUPPLEMENTARY INFORMATION CXCR4 inhibitors could benefit to HER2 but not to Triple-Negative breast cancer patients Lefort S. 1,2, Thuleau A. 3, Kieffer Y. 1,2, Sirven P. 1,2, Bieche I. 4, Marangoni E.
Fig 1 CD163. CD11b S100A9. Sirius Red. 100μm ** ** CD163. CD11b S100A9 ** Sirius Red (PL) Sirius Red SUM Mo.
 Sirius Red SUM Mo.") T47D T47D + o SU-59 Fig SU-59 + o IHC score (-3) IHC score (-2) CD63 3 2 IHC score (-3) CD63 3 ** 2 CDb CDb * * SA9 SA9 ** * 2 IHC score (-4) αsa αsa 4 ** ** 2 Sirius Red μm IHC score (%) Sirius Red 8
T47D T47D + o SU-59 Fig SU-59 + o IHC score (-3) IHC score (-2) CD63 3 2 IHC score (-3) CD63 3 ** 2 CDb CDb * * SA9 SA9 ** * 2 IHC score (-4) αsa αsa 4 ** ** 2 Sirius Red μm IHC score (%) Sirius Red 8
Nature Neuroscience: doi: /nn Supplementary Figure 1
 Supplementary Figure 1 EGFR inhibition activates signaling pathways (a-b) EGFR inhibition activates signaling pathways (a) U251EGFR cells were treated with erlotinib (1µM) for the indicated times followed
Supplementary Figure 1 EGFR inhibition activates signaling pathways (a-b) EGFR inhibition activates signaling pathways (a) U251EGFR cells were treated with erlotinib (1µM) for the indicated times followed
(A) Cells grown in monolayer were fixed and stained for surfactant protein-c (SPC,
 Cells grown in monolayer were fixed and stained for surfactant protein-c (SPC,") Supplemental Figure Legends Figure S1. Cell line characterization (A) Cells grown in monolayer were fixed and stained for surfactant protein-c (SPC, green) and co-stained with DAPI to visualize the nuclei.
Supplemental Figure Legends Figure S1. Cell line characterization (A) Cells grown in monolayer were fixed and stained for surfactant protein-c (SPC, green) and co-stained with DAPI to visualize the nuclei.
Supplemental Information. T Cells Enhance Autoimmunity by Restraining Regulatory T Cell Responses via an Interleukin-23-Dependent Mechanism
 Immunity, Volume 33 Supplemental Information T Cells Enhance Autoimmunity by Restraining Regulatory T Cell Responses via an Interleukin-23-Dependent Mechanism Franziska Petermann, Veit Rothhammer, Malte
Immunity, Volume 33 Supplemental Information T Cells Enhance Autoimmunity by Restraining Regulatory T Cell Responses via an Interleukin-23-Dependent Mechanism Franziska Petermann, Veit Rothhammer, Malte
Supplementary Information. Tissue-wide immunity against Leishmania. through collective production of nitric oxide
 Supplementary Information Tissue-wide immunity against Leishmania through collective production of nitric oxide Romain Olekhnovitch, Bernhard Ryffel, Andreas J. Müller and Philippe Bousso Supplementary
Supplementary Information Tissue-wide immunity against Leishmania through collective production of nitric oxide Romain Olekhnovitch, Bernhard Ryffel, Andreas J. Müller and Philippe Bousso Supplementary
McAb and rhil-2 activated bone marrow on the killing and purging of leukemia cells
 Effects of McAb and rhil-2 activated bone marrow on the killing and purging of leukemia cells X.C. Wei, D.D. Yang, X.R. Han, Y.A. Zhao, Y.C. Li, L.J. Zhang and J.J. Wang Institute of hematological research,
Effects of McAb and rhil-2 activated bone marrow on the killing and purging of leukemia cells X.C. Wei, D.D. Yang, X.R. Han, Y.A. Zhao, Y.C. Li, L.J. Zhang and J.J. Wang Institute of hematological research,
Functional genomics reveal that the serine synthesis pathway is essential in breast cancer
 Functional genomics reveal that the serine synthesis pathway is essential in breast cancer Results Presented by Stacey Lin Lloyd Lab http://www.amsbio.com/expression-ready-lentiviral-particles.aspx Overview
Functional genomics reveal that the serine synthesis pathway is essential in breast cancer Results Presented by Stacey Lin Lloyd Lab http://www.amsbio.com/expression-ready-lentiviral-particles.aspx Overview
Title: Human breast cancer associated fibroblasts exhibit subtype specific gene expression profiles
 Author's response to reviews Title: Human breast cancer associated fibroblasts exhibit subtype specific gene expression profiles Authors: Julia Tchou (julia.tchou@uphs.upenn.edu) Andrew V Kossenkov (akossenkov@wistar.org)
Author's response to reviews Title: Human breast cancer associated fibroblasts exhibit subtype specific gene expression profiles Authors: Julia Tchou (julia.tchou@uphs.upenn.edu) Andrew V Kossenkov (akossenkov@wistar.org)
In vitro scratch assay: method for analysis of cell migration in vitro labeled fluorodeoxyglucose (FDG)
") In vitro scratch assay: method for analysis of cell migration in vitro labeled fluorodeoxyglucose (FDG) 1 Dr Saeb Aliwaini 13/11/2015 Migration in vivo Primary tumors are responsible for only about 10%
In vitro scratch assay: method for analysis of cell migration in vitro labeled fluorodeoxyglucose (FDG) 1 Dr Saeb Aliwaini 13/11/2015 Migration in vivo Primary tumors are responsible for only about 10%
MicroRNAs Modulate the Noncanonical NF- B Pathway by Regulating IKK Expression During Macrophage Differentiation
 MicroRNAs Modulate the Noncanonical NF- B Pathway by Regulating IKK Expression During Macrophage Differentiation Tao Li 1 *, Michael J. Morgan 1 *, Swati Choksi 1, Yan Zhang 1, You-Sun Kim 2#, Zheng-gang
MicroRNAs Modulate the Noncanonical NF- B Pathway by Regulating IKK Expression During Macrophage Differentiation Tao Li 1 *, Michael J. Morgan 1 *, Swati Choksi 1, Yan Zhang 1, You-Sun Kim 2#, Zheng-gang
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature10134 Supplementary Figure 1. Anti-inflammatory activity of sfc. a, Autoantibody immune complexes crosslink activating Fc receptors, promoting activation of macrophages, and WWW.NATURE.COM/NATURE
doi:10.1038/nature10134 Supplementary Figure 1. Anti-inflammatory activity of sfc. a, Autoantibody immune complexes crosslink activating Fc receptors, promoting activation of macrophages, and WWW.NATURE.COM/NATURE
Supplementary Figure 1. Identification of tumorous sphere-forming CSCs and CAF feeder cells. The LEAP (Laser-Enabled Analysis and Processing)
") Supplementary Figure 1. Identification of tumorous sphere-forming CSCs and CAF feeder cells. The LEAP (Laser-Enabled Analysis and Processing) platform with laser manipulation to efficiently purify lung
Supplementary Figure 1. Identification of tumorous sphere-forming CSCs and CAF feeder cells. The LEAP (Laser-Enabled Analysis and Processing) platform with laser manipulation to efficiently purify lung
Cytokine Arrays Reveal Black Ops Tactics of Tumor-induced Immunosuppression
 Cytokine Arrays Reveal Black Ops Tactics of Tumor-induced Immunosuppression Jarad J Wilson, Ph.D. Technical Support & Marketing Specialist Ruo-Pan Huang, MD, Ph.D. Founder and CEO What are Antibody Arrays?
Cytokine Arrays Reveal Black Ops Tactics of Tumor-induced Immunosuppression Jarad J Wilson, Ph.D. Technical Support & Marketing Specialist Ruo-Pan Huang, MD, Ph.D. Founder and CEO What are Antibody Arrays?
Supplemental Experimental Procedures
 Cell Stem Cell, Volume 2 Supplemental Data A Temporal Switch from Notch to Wnt Signaling in Muscle Stem Cells Is Necessary for Normal Adult Myogenesis Andrew S. Brack, Irina M. Conboy, Michael J. Conboy,
Cell Stem Cell, Volume 2 Supplemental Data A Temporal Switch from Notch to Wnt Signaling in Muscle Stem Cells Is Necessary for Normal Adult Myogenesis Andrew S. Brack, Irina M. Conboy, Michael J. Conboy,
ONCO-HU TM MICE FOR IMMUNOTHERAPEUTIC DRUG DISCOVERY. Brian W. Soper, Ph.D. Senior Technical Information Scientist
 ONCO-HU TM MICE FOR IMMUNOTHERAPEUTIC DRUG DISCOVERY Brian W. Soper, Ph.D. Senior Technical Information Scientist Presentation Outline Capabilities Humanization Hu-NSG TM versus Hu-NSG TM -SGM3 Immuno-oncology
ONCO-HU TM MICE FOR IMMUNOTHERAPEUTIC DRUG DISCOVERY Brian W. Soper, Ph.D. Senior Technical Information Scientist Presentation Outline Capabilities Humanization Hu-NSG TM versus Hu-NSG TM -SGM3 Immuno-oncology
Supporting Information
 Supporting Information Chan et al. 1.173/pnas.9654916 A Patient B Xenograft C * remaining feature of normal lymph node * * * D lymphocytes Infiltrating transitional carcinoma cells E Enlarged axillary
Supporting Information Chan et al. 1.173/pnas.9654916 A Patient B Xenograft C * remaining feature of normal lymph node * * * D lymphocytes Infiltrating transitional carcinoma cells E Enlarged axillary
Cell isolation. Spleen and lymph nodes (axillary, inguinal) were removed from mice
 were removed from mice") Supplementary Methods: Cell isolation. Spleen and lymph nodes (axillary, inguinal) were removed from mice and gently meshed in DMEM containing 10% FBS to prepare for single cell suspensions. CD4 + CD25
Supplementary Methods: Cell isolation. Spleen and lymph nodes (axillary, inguinal) were removed from mice and gently meshed in DMEM containing 10% FBS to prepare for single cell suspensions. CD4 + CD25
Supplementary Data Table of Contents:
 Supplementary Data Table of Contents: - Supplementary Methods - Supplementary Figures S1(A-B) - Supplementary Figures S2 (A-B) - Supplementary Figures S3 - Supplementary Figures S4(A-B) - Supplementary
Supplementary Data Table of Contents: - Supplementary Methods - Supplementary Figures S1(A-B) - Supplementary Figures S2 (A-B) - Supplementary Figures S3 - Supplementary Figures S4(A-B) - Supplementary
Claudin-4 Expression in Triple Negative Breast Cancer: Correlation with Androgen Receptors and Ki-67 Expression
 Claudin-4 Expression in Triple Negative Breast Cancer: Correlation with Androgen Receptors and Ki-67 Expression Mona A. Abd-Elazeem, Marwa A. Abd- Elazeem Pathology department, Faculty of Medicine, Tanta
Claudin-4 Expression in Triple Negative Breast Cancer: Correlation with Androgen Receptors and Ki-67 Expression Mona A. Abd-Elazeem, Marwa A. Abd- Elazeem Pathology department, Faculty of Medicine, Tanta
Supporting Information
 Supporting Information Aldridge et al. 10.1073/pnas.0900655106 Fig. S1. Flow diagram of sublethal (a) and lethal (b) influenza virus infections. (a) Infection of lung epithelial cells by influenza virus
Supporting Information Aldridge et al. 10.1073/pnas.0900655106 Fig. S1. Flow diagram of sublethal (a) and lethal (b) influenza virus infections. (a) Infection of lung epithelial cells by influenza virus
Supplementary Figures
 Supplementary Figures Supplementary Figure 1. NKT ligand-loaded tumour antigen-presenting B cell- and monocyte-based vaccine induces NKT, NK and CD8 T cell responses. (A) The cytokine profiles of liver
Supplementary Figures Supplementary Figure 1. NKT ligand-loaded tumour antigen-presenting B cell- and monocyte-based vaccine induces NKT, NK and CD8 T cell responses. (A) The cytokine profiles of liver
Supplementary Figure 1. Repression of hepcidin expression in the liver of mice treated with
 Supplementary Figure 1. Repression of hepcidin expression in the liver of mice treated with DMN Immunohistochemistry for hepcidin and H&E staining (left). qrt-pcr assays for hepcidin in the liver (right).
Supplementary Figure 1. Repression of hepcidin expression in the liver of mice treated with DMN Immunohistochemistry for hepcidin and H&E staining (left). qrt-pcr assays for hepcidin in the liver (right).
Human and mouse T cell regulation mediated by soluble CD52 interaction with Siglec-10. Esther Bandala-Sanchez, Yuxia Zhang, Simone Reinwald,
 Human and mouse T cell regulation mediated by soluble CD52 interaction with Siglec-1 Esther Bandala-Sanchez, Yuxia Zhang, Simone Reinwald, James A. Dromey, Bo Han Lee, Junyan Qian, Ralph M Böhmer and Leonard
Human and mouse T cell regulation mediated by soluble CD52 interaction with Siglec-1 Esther Bandala-Sanchez, Yuxia Zhang, Simone Reinwald, James A. Dromey, Bo Han Lee, Junyan Qian, Ralph M Böhmer and Leonard
Correlation between serum level of chemokine (C-C motif) ligand 18 and poor prognosis in breast cancer
 ligand 18 and poor prognosis in breast cancer") Correlation between serum level of chemokine (C-C motif) ligand 18 and poor prognosis in breast cancer J.H. Sun 1, N. Fan 2 and Y. Zhang 1 1 Xianyang Hospital of Yan an University Clinical Laboratory,
Correlation between serum level of chemokine (C-C motif) ligand 18 and poor prognosis in breast cancer J.H. Sun 1, N. Fan 2 and Y. Zhang 1 1 Xianyang Hospital of Yan an University Clinical Laboratory,
Pearson r = P (one-tailed) = n = 9
 = n = 9") 8F4-Specific Lysis, % 1 UPN1 UPN3 8 UPN7 6 Pearson r =.69 UPN2 UPN5 P (one-tailed) =.192 4 UPN8 n = 9 2 UPN9 UPN4 UPN6 5 1 15 2 25 8 8F4, % Max MFI Supplementary Figure S1. AML samples UPN1-UPN9 show variable
8F4-Specific Lysis, % 1 UPN1 UPN3 8 UPN7 6 Pearson r =.69 UPN2 UPN5 P (one-tailed) =.192 4 UPN8 n = 9 2 UPN9 UPN4 UPN6 5 1 15 2 25 8 8F4, % Max MFI Supplementary Figure S1. AML samples UPN1-UPN9 show variable
Tumor-associated macrophages drive spheroid formation during early transcoelomic metastasis of ovarian cancer
 Tumor-associated macrophages drive spheroid formation during early transcoelomic metastasis of ovarian cancer Mingzhu Yin, 1 Xia Li, 1 Shu Tan, 1,2 Huanjiao Jenny Zhou, 1,3,4 Weidong Ji, 3,4 Stefania Bellone,
Tumor-associated macrophages drive spheroid formation during early transcoelomic metastasis of ovarian cancer Mingzhu Yin, 1 Xia Li, 1 Shu Tan, 1,2 Huanjiao Jenny Zhou, 1,3,4 Weidong Ji, 3,4 Stefania Bellone,
W/T Itgam -/- F4/80 CD115. F4/80 hi CD115 + F4/80 + CD115 +
 F4/8 % in the peritoneal lavage 6 4 2 p=.15 n.s p=.76 CD115 F4/8 hi CD115 + F4/8 + CD115 + F4/8 hi CD115 + F4/8 + CD115 + MHCII MHCII Supplementary Figure S1. CD11b deficiency affects the cellular responses
F4/8 % in the peritoneal lavage 6 4 2 p=.15 n.s p=.76 CD115 F4/8 hi CD115 + F4/8 + CD115 + F4/8 hi CD115 + F4/8 + CD115 + MHCII MHCII Supplementary Figure S1. CD11b deficiency affects the cellular responses
Supplementary Figure 1
 Supplementary Figure 1 14 12 SEM4C PLXN2 8 SEM4C C 3 Cancer Cell Non Cancer Cell Expression 1 8 6 6 4 log2 ratio Expression 2 1 4 2 2 p value.1 D Supplementary Figure 1. Expression of Sema4C and Plexin2
Supplementary Figure 1 14 12 SEM4C PLXN2 8 SEM4C C 3 Cancer Cell Non Cancer Cell Expression 1 8 6 6 4 log2 ratio Expression 2 1 4 2 2 p value.1 D Supplementary Figure 1. Expression of Sema4C and Plexin2
Supplementary Figure 1. Characterization of basophils after reconstitution of SCID mice
 Supplementary figure legends Supplementary Figure 1. Characterization of after reconstitution of SCID mice with CD4 + CD62L + T cells. (A-C) SCID mice (n = 6 / group) were reconstituted with 2 x 1 6 CD4
Supplementary figure legends Supplementary Figure 1. Characterization of after reconstitution of SCID mice with CD4 + CD62L + T cells. (A-C) SCID mice (n = 6 / group) were reconstituted with 2 x 1 6 CD4
Supplemental Figure 1. Western blot analysis indicated that MIF was detected in the fractions of
 Supplemental Figure Legends Supplemental Figure 1. Western blot analysis indicated that was detected in the fractions of plasma membrane and cytosol but not in nuclear fraction isolated from Pkd1 null
Supplemental Figure Legends Supplemental Figure 1. Western blot analysis indicated that was detected in the fractions of plasma membrane and cytosol but not in nuclear fraction isolated from Pkd1 null
Nature Medicine: doi: /nm.3922
 Title: Glucocorticoid-induced tumor necrosis factor receptor-related protein co-stimulation facilitates tumor regression by inducing IL-9-producing helper T cells Authors: Il-Kyu Kim, Byung-Seok Kim, Choong-Hyun
Title: Glucocorticoid-induced tumor necrosis factor receptor-related protein co-stimulation facilitates tumor regression by inducing IL-9-producing helper T cells Authors: Il-Kyu Kim, Byung-Seok Kim, Choong-Hyun
DISCOVERING ATCC IMMUNOLOGICAL CELLS - MODEL SYSTEMS TO STUDY THE IMMUNE AND CARDIOVASCULAR SYSTEMS
 DISCOVERING ATCC IMMUNOLOGICAL CELLS - MODEL SYSTEMS TO STUDY THE IMMUNE AND CARDIOVASCULAR SYSTEMS James Clinton, Ph.D. Scientist, ATCC February 19, 2015 About ATCC Founded in 1925, ATCC is a non-profit
DISCOVERING ATCC IMMUNOLOGICAL CELLS - MODEL SYSTEMS TO STUDY THE IMMUNE AND CARDIOVASCULAR SYSTEMS James Clinton, Ph.D. Scientist, ATCC February 19, 2015 About ATCC Founded in 1925, ATCC is a non-profit
Title of file for HTML: Supplementary Information Description: Supplementary Figures and Supplementary Table
 Title of file for HTML: Supplementary Information Description: Supplementary Figures and Supplementary Table Title of file for HTML: Peer Review File Description: Innate Scavenger Receptor-A regulates
Title of file for HTML: Supplementary Information Description: Supplementary Figures and Supplementary Table Title of file for HTML: Peer Review File Description: Innate Scavenger Receptor-A regulates