Evaluation of Insulin-like Growth Factor Receptor 1 Inhibition as a Treatment for Prostate Cancer: Preclinical Efficacy and Resistance
|
|
|
- Ira Walters
- 6 years ago
- Views:
Transcription
1 University of Miami Scholarly Repository Open Access Dissertations Electronic Theses and Dissertations Evaluation of Insulin-like Growth Factor Receptor 1 Inhibition as a Treatment for Prostate Cancer: Preclinical Efficacy and Resistance Cale Fahrenholtz University of Miami, cfahrenholtz@gmail.com Follow this and additional works at: Recommended Citation Fahrenholtz, Cale, "Evaluation of Insulin-like Growth Factor Receptor 1 Inhibition as a Treatment for Prostate Cancer: Preclinical Efficacy and Resistance" (2013). Open Access Dissertations This Embargoed is brought to you for free and open access by the Electronic Theses and Dissertations at Scholarly Repository. It has been accepted for inclusion in Open Access Dissertations by an authorized administrator of Scholarly Repository. For more information, please contact repository.library@miami.edu.
2 UNIVERSITY OF MIAMI EVALUATION OF INSULIN-LIKE GROWTH FACTOR RECEPTOR 1 INHIBITION AS A TREATMENT FOR PROSTATE CANCER: PRECLINICAL EFFICACY AND RESISTANCE By Cale Fahrenholtz A DISSERTATION Submitted to the Faculty of the University of Miami in partial fulfillment of the requirements for the degree of Doctor of Philosophy Coral Gables, Florida December 2013
3 2013 Cale Fahrenholtz All Rights Reserved
4 UNIVERSITY OF MIAMI A dissertation submitted in partial fulfillment of the requirements for the degree of Doctor of Philosophy EVALUATION OF INSULIN-LIKE GROWTH FACTOR RECEPTOR 1 INHIBITION AS A TREATMENT FOR PROSTATE CANCER: PRECLINICAL EFFICACY AND RESISTANCE Cale Fahrenholtz Approved: Kerry Burnstein, Ph.D. Professor of Molecular and Cellular Pharmacology Vladlen Slepak, Ph.D. Professor of Molecular and Cellular Pharmacology Charles Luetje, Ph.D. Professor of Molecular and Cellular Pharmacology Zafar Nawaz, Ph.D. Professor of Biochemistry and Molecular Biology M. Brian Blake, Ph.D. Dean of the Graduate School
5 FAHRENHOLTZ, CALE (Ph.D., Molecular and Cellular Pharmacology) Evaluation of Insulin-like Growth Factor Receptor 1 (December 2013) Inhibition as a Treatment for Prostate Cancer: Preclinical Efficacy and Resistance Abstract of a dissertation at the University of Miami. Dissertation supervised by Kerry Burnstein, Ph.D. No. of pages in text. (69) Prostate cancer is the most commonly diagnosed non-cutaneous malignancy in U.S. men. While tumors initially respond to androgen-deprivation therapy, the standard care for advanced or metastatic disease, tumors eventually recur as castration-resistant prostate cancer. Upregulation of the insulin-like growth factor signaling axis drives growth and progression of prostate cancer by promoting proliferation, survival, and angiogenesis. Ganitumab (formerly AMG 479) is a fully human antibody that inhibits binding of IGF-1 and IGF-2 to IGF-1R. Ganitumab decreased IGF-1 induced phosphorylation of the downstream effector AKT and reduced proliferation of multiple androgen-dependent and castrationresistant human prostate cancer cell lines in vitro albeit to varying extents. We evaluated the therapeutic value of ganitumab in several pre-clinical settings including androgen-dependent prostate cancer, castration-resistant (recurrent) prostate cancer, and in combination with androgen-deprivation therapy. Ganitumab inhibited androgen-dependent human VCaP prostate cancer xenograft growth and increased tumor doubling time from 2.3±0.4 weeks to 6.4±0.4 weeks. Ganitumab inhibited growth of castration-resistant VCaP xenografts for over 11.5 weeks of treatment. In contrast, ganitumab did not have appreciable effects on the castration-resistant CWR-22Rv1 xenograft model.
6 Ganitumab was most potent against VCaP xenografts when combined with androgen-deprivation therapy (castration). Tumor volume was reduced by 72% after 4 weeks of treatment and growth suppression was maintained over 16 weeks of treatment. However, a significant proportion of these tumors acquired resistance to ganitumab. To address mechanisms of resistance, we developed an in vitro model of ganitumab resistance, which we termed VCaP/GanR, by prolonged culture of the VCaP human prostate cancer cell in ganitumab. Acquired resistance to ganitumab in VCaP/GanR was a reproducible and stable phenotype. Unlike parental VCaP cells, VCaP/GanR did not undergo apoptosis following ganitumab treatment or serum deprivation. VCaP/GanR did not express increased levels of IGF-1R, insulin receptor, or phospho-akt compared to parental VCaP. Increased IGF-1R, insulin receptor and phospho-akt are associated with acquired resistance to IGF-1R inhibition in other cancers. VCaP/GanR exhibited increased levels of phosphorylated S6 indicative of high mtor activity. However, acquired resistance to ganitumab was not dependent on increased mtor activity in VCaP/GanR as mtor inhibitor rapamycin failed to restore sensitivity to ganitumab. Phospho-proteomic arrays revealed alterations in several signaling cascades in VCaP/GanR compared to parental VCaP cells. Inhibitors were identified that restored the antiproliferative effects of ganitumab in VCaP/GanR. These data implicate a new mechanism of resistance to IGF-1R inhibition and support the judicious use of ganitumab in the treatment of prostate cancer.
7 Dedication This dissertation is dedicated to my two loves, my wife Stephanie, and Bulleit bourbon. iii
8 TABLE OF CONTENTS LIST OF FIGURES...v LIST OF TABLES vii LIST OF ABBREVIATIONS..viii CHAPTER 1 INTRODUCTION..1 Prostate Cancer Background Insulin-like Growth Factor 1 Signaling Axis..2 IGF-1R Signaling Axis and Prostate Cancer 5 IGF-1R Inhibitor Ganitumab (formerly AMG 479) 6 Cell Models of Prostate Cancer..7 Summary of Results.8 2 MATERIALS AND METHODS.10 3 PRECLINICAL EFFICACY OF GANITUMAB 16 Ganitumab inhibits IGF-1R signaling in androgen-dependent and castration-resistant prostate cancer cells in vitro..16 Ganitumab inhibits growth of androgen-dependent and castrationresistant prostate cancer in vitro..18 Ganitumab inhibits androgen-dependent prostate cancer growth in vivo..18 Effect of ganitumab treatment on castration-resistant prostate cancer in vivo 21 Ganitumab combined with androgen-deprivation is a highly effective treatment regimen..26 Reversibility and durability of treatment with ganitumab combined with androgen-deprivation therapy ACQUIRED RESISTANCE TO GANITUMAB...32 Development of a ganitumab resistant prostate cancer cell derivative.32 Evaluation of IGF-1R related signaling pathways in VCaP/GanR..35 Phospho-proteome kinase arrays 37 Elevated mtor activity does not promote acquired resistance to ganitumab 41 Decreased AMPK activity does not contribute to acquired resistance to ganitumab 43 Acquired resistance to ganitumab is calcium dependent 46 Acquired resistance to ganitumab is not dependent on calmodulin or calcineurin signaling DISCUSSION..52 REFERENCES...63 iv
9 LIST OF FIGURES Figure 1. Figure 2. Figure 3. Figure 4. Figure 5. Figure 6. Figure 7. Figure 8. Figure 9. Figure 10. Figure 11. Figure 12. Figure 13. Figure 14. Figure 15. Figure 16. Schematic of insulin-like growth factor receptor Schematic of IGF-1R signaling pathways. 4 Analysis of ganitumab effects on IGF-1R signaling and cell proliferation in vitro.17 Ganitumab inhibits the growth of androgen-dependent VCaP xenografts 20 Ganitumab inhibits the growth of castration-resistant VCaP xenografts 22 Ganitumab does not affect the growth of well-established castration-resistant 22Rv1 xenografts.25 Combining ganitumab with androgen-deprivation therapy most effectively inhibits VCaP xenografts 27 Ganitumab reduces IGF-1R levels when combined with androgen-deprivation therapy..29 Evaluation of durability and reversibility of combined ganitumab and androgen-deprivation therapy on VCaP xenografts..31 Derivation of a ganitumab resistant prostate cancer VCaP line termed VCaP/GanR...32 Derivation of a second ganitumab resistant prostate cancer VCaP cell line termed (VCaP/GanR-2)...33 Acquired resistance to ganitumab in VCaP/GanR is stable...34 Characterization of human prostate cancer VCaP derivative VCaP/GanR.35 Characterization of IGF-1R related signaling pathways...36 Phospho-proteome kinase profiling of VCaP and VCaP/GanR cells 38 Analysis and verification of phsopho-proteomic profiling.39 v
10 Figure 17. Figure 18. Figure 19. Figure 20. Figure 21. Figure 22. Figure 23. Effects of PYK2 inhibition, PLCγ inhibition and 5-FU treatment on VCaP/GanR...40 mtor inhibition does not restore sensitivity to ganitumab in VCaP/GanR.42 Metformin partially restores sensitivity to ganitumab in VCaP/GanR.44 AMPK activation by AICAR does not restore sensitivity to ganitumab in VCaP/GanR.45 Treatment with cell permeable calcium chelators restores sensitivity to ganitumab.48 Reduction of intracellular calcium does not increase sensitivity to ganitumab in CWR-R1, 22Rv1 or LNCaP prostate cancer cells.49 Acquired resistance to ganitumab of VCaP/GanR does not require calcineurin or calmodulin signaling 50 vi
11 LIST OF TABLES Table 1. Table 2. Table 3. Characteristics of prostate cancer cell models 8 Mouse weights in 22Rv1 xenograft studies 24 Mouse weights in VCaP xenograft studies.28 vii
12 LIST OF ABBREVIATIONS AICAR AMPK AR AR LBD BAPTA-AM CSS CRPC CREB CsA EGF-R EGTA-AM FBS IGF-1 IGF-1R IGF-2 IGFBP INSR mtor PARP PI3K PSA PTEN PYK2 PLCγ RTK SCID SD SEM N 1 -(β-d-ribofuranosyl)-5-aminoimidazole-4-carboxamide 5' AMP-activated protein kinase Androgen receptor Androgen receptor lacking the ligand binding domain 1,2-Bis(2-aminophenoxy)ethane-N,N,N',N'-tetraacetic acid tetrakis(acetoxymethyl ester) Charcoal-stripped serum Castration-resistant prostate cancer camp response element-binding protein Cyclosporin A Epidermal growth factor receptor Ethyleneglycol-bis(β-aminoethyl)-N,N,N,N -tetraacetoxymethyl ester Fetal bovine serum Insulin-like growth factor 1 Insulin-like growth factor receptor 1 Insulin-like growth factor 2 Insulin-like growth factor binding protein Insulin receptor Mammalian target of rapamycin Poly (ADP-ribose) polymerase Phosphatidylinositol 3-kinase Prostate specific antigen phosphatidylinositol-3,4,5-trisphosphate 3-phosphatase Protein tyrosine kinase 2 beta Phospholipase C gamma Receptor tyrosine kinase Severe combined immunodeficient Standard deviation Standard error of the mean viii
13 CHAPTER 1: INTRODUCTION Prostate cancer background Prostate cancer is the most commonly diagnosed cancer in men and is the second leading cause of cancer-related deaths in U.S. men each year (1). It is estimated that 1 in 6 men will be diagnosed with prostate cancer within their lifetimes (2). Prostate cancer incidence is highly correlated with increasing age (3). While many great strides have been made in the treatment of prostate cancer, the mortality rate underscores the need for new treatment regimens as the current armamentarium is not sufficient. The prostate gland is a walnut-sized exocrine gland found in males (4). The function of the prostate is to secrete the alkaline fluids that make up 50-75% of the semen protecting sperm from acidity encountered in the urinal and vaginal tracts (5). The normal prostate relies on androgen signaling, through the hormone dihydrotestosterone, for normal growth and maintenance (6). The most common presenting features of adenocarcinoma of the prostate are due to complications from urinary tract obstructions including: urgency, nocturia, frequency, and hesitancy in urination (7). For clinical diagnosis, the prostate gland is manually checked during a digital rectal exam, and serum prostate-specific antigen (PSA) is also assessed for clinical diagnosis. PSA is an androgen regulated gene whose levels can indicate severity and stage of prostate cancer. At this early diagnosis, the prostate tumor, like the normal prostate, relies on androgens for growth, and is termed androgen-dependent. This androgen-dependent localized prostate cancer is treated with a combination 1
14 2 of either radiation therapy, surgical excision of the tumor, or active surveillance. Upon failure of these treatments, the prostate cancer generally progresses to an advanced or metastatic disease. Androgen deprivation therapy is the standard of treatment for advanced or metastatic prostate cancer. This treatment regimen initially results in decreased tumor burden, decreased PSA, and marginally increases life expectancy (8). However, the prostate cancer will inevitably return, within approximately 9 to 23 months (9). At this stage, the disease is termed castration-resistant prostate cancer (CRPC) (10, 11). At this stage, CRPC is incurable and treatment options are very limited. Insulin-like growth factor 1 signaling axis Insulin-like growth factor receptor type I (IGF-1R) is a receptor tyrosine kinase (RTK) that is structurally related to insulin receptor (INSR) sharing 60% sequence identity. IGF-1 and IGF-2 are the native ligands for IGF-1R, both of which are produced in the liver as a consequence of growth hormone signaling (12). The IGF-IR complex contains two disulfide linked extracellular α-chains that bind IGF-1 and IGF-2, and two β-chains that span the membrane into the intracellular space harboring the tyrosine kinase domain (Figure 1) (13, 14). Binding of IGF-1 or IGF-2 induces a conformational change which results in activation of the tyrosine kinase domain through autophosphorylation of intracellular tyrosine residues (15). IGF-1R can also be found as a hybrid receptor, containing one IGF-1R α and β chain, and also one INSR α and β chain (16). These hybrid receptors can be activated by IGF-1, and to a lesser degree
.")
15 3 Figure 1. Schematic of insulin like growth factor receptor 1. The insulin-like growth factor receptor 1 consists of two alpha chains and two beta chains connected by disulfide bonds. The extracellular L2 domain is important for ligand binding of both IGF-1 and IGF-2. Ganitumab binds the L2 domain inhibiting ligand binding. by insulin (16, 17). The role of hybrid receptor signaling in both cancer and normal physiology is currently being investigated. Signaling through the IGF-1R plays an important role in many cellular processes including: mitogenesis, apoptosis, proliferation, and angiogenesis (Figure 2). The downstream effects of IGF-1R activation include increased signaling through the PI3K/AKT and MEK/ERK signaling cascades (18, 19). PTEN is the negative regulator of PI3K that is non-functional (through mutation or deletion) in ~50% of all prostate cancer specimens (20, 21). Lack of functional


16 Figure 2. Schematic of IGF-1R signaling pathways. Under normal conditions, IGF-1 binds IGF-1R, which activates several downstream effector pathways. Major downstream effector pathways of IGF- 1R include, but are not limited to, the PI3K/AKT and MEK/ERK signaling cascades, which result in increased cell proliferation, survival, and protein translation mediated through various downstream effectors. Ganitumab inhibits the binding of IGF-1 and IGF-2 to IGF-1R thereby attenuating signaling through downstream effector pathways. Upon binding of ganitumab to IGF-1R, IGF-1R is subsequently internalized and degraded. 4
17 5 PTEN results is a more active AKT through maintained phosphorylation. In general the PI3K pathway inhibits apoptosis and increases protein synthesis, while the MEK/ERK pathway stimulates proliferation (22), although these effects are not mutually exclusive. AKT activates mammalian target of rapamycin (mtor). mtor is part of a complex that contains RAPTOR, mlst8, and PRAS40 termed mtorc1 (23). Activation of mtorc1 results in increased mrna transcription, and increased protein synthesis both of which promote cell survival and proliferation (24). Increased mtorc1 activity is suggested as a mechanism of resistance to IGF- 1R inhibition in Ewing s Sarcoma (25). Increased AKT-independent mtor activity is also a mechanism of resistance to the inhibition of HER2, another RTK, in breast cancer cells (26). IGF-1R signaling axis and prostate cancer The insulin-like growth factor receptor type I and native ligands, IGF-1 and IGF-2, are dysregulated in a variety of cancers including prostate cancer (25, 27-29). Elevated levels of serum IGF-1, the most potent activator of IGF-1R, have been correlated with an increased risk of prostate cancer (30, 31). IGF-1 and IGF-1R are increased with progression to castration-resistance in vivo (32). In clinical specimens, IGF-1R expression is elevated in primary prostate cancer versus benign prostatic epithelium (33). Studies with an IGF-1R receptor inhibitor, the antibody cixutumumab (formerly IMC-A12), showed positive results against LuCaP 35 prostate cancer xenografts alone and in conjunction with androgen-deprivation therapy or docetaxel (34-36). A recently completed phase
18 6 2 clinical trial with figitumumab (formerly CP-751,871), an IgG2 IGF-1R antibody inhibitor that blocks only IGF-1 binding and promotes receptor internalization, in patients with localized prostate cancer showed a decrease in serum levels of PSA (37, 38). Another clinical trial which combines cixutumumab with androgen deprivation therapy shows significant changes in IGF and glucose homeostasis pathways (39). These changes decreased the IGF-1R signaling axis, which would alter AKT signaling, glucose transport and regulation and may be beneficial against tumor proliferation, maintenance, and progression. These studies support further investigation of inhibition of IGF-1R signaling with ganitumab in robust models of human prostate cancer and suggest that IGF-1R inhibition may be a valuable treatment paradigm for androgen-dependent and castration-resistant prostate cancer. IGF-1R inhibitor ganitumab (formerly AMG 479) Ganitumab (formerly AMG 479) is a fully human antibody (IgG1) against IGF-1R. Ganitumab reduces IGF-1R activation by binding the L2 domain (an extracellular leucine-rich domain that contributes to ligand binding) of IGF-1R thereby preventing both IGF-1 and IGF-2 interaction with the receptor (Figure 1) (14, 40). Binding of ganitumab to IGF-1R also induces internalization and degradation of IGF-1R in vivo (40). Ganitumab does not interact with the closely related insulin receptor (INSR) but does inhibit hybrid IGF-1R/INSR receptors (40). Ganitumab has been studied in phase II trials for a variety of advanced tumors (41, 42). In phase I/Ib escalation studies, ganitumab was generally well-
19 7 tolerated when administered to patients with advanced solid tumors both as a single agent and in combination with either the multi-kinase inhibitor sorafenib, either of the epidermal growth factor receptor (EGF-R) inhibitors panitumumab or erlotinib, or gemcitabine. These combination therapies as well as ganitumab as a single agent displayed promising anti-tumor effects in patients and resulted in few dose-limiting toxicities, and generally mild adverse events including fatigue, nausea, vomiting or chills (43, 44). To date, ganitumab has shown signs of efficacy against pancreatic cancer and Ewing s sarcoma, yet there has been minimal evaluation of ganitumab for prostate cancer. Ganitumab, like cixutumumab, blocks both the high affinity binding of IGF-1 and IGF-2 to IGF-1R, whereas figitumumab only blocks IGF-1 binding (45-47). Cell models of human prostate cancer We employed several prostate cancer cell models that share characteristics with a significant proportion of prostate cancer specimens (Table 1). VCaP is an androgen-dependent human prostate cancer cell line that like ~50% of prostate cancers is wild-type for PTEN (a negative regulator for AKT activator PI3K) (20, 21). As observed in human prostate cancer, VCaP cells progress to castration-resistance in vivo (20, 48, 49). VCaP also expresses the TMPRSS2:ERG fusion gene, which is seen in ~50% of primary prostate cancer specimens and may promote prostate cancer progression (50). Another androgen-dependent human prostate cancer cell line, LNCaP, does not express active PTEN (51), but will progress to castration-resistance in vivo similar to VCaP. CWR-22Rv1 (22Rv1) and CWR-R1 are castration-resistant, wild-type for
 as well as constitutively active AR splice variants, which lack the ligand binding domain (termed AR")
20 8 Table 1. Cell models of human prostate cancer PTEN, and express both full length androgen receptor (AR) as well as constitutively active AR splice variants, which lack the ligand binding domain (termed AR LBDs) and no longer require androgens for activity (52, 53). Summary of results Utilizing established models of prostate cancer, we assessed the effectiveness of ganitumab alone and in conjunction with complete androgendeprivation therapy (castration) as a treatment for androgen-dependent prostate cancer, advanced CRPC, and progression to CRPC. We show that ganitumab inhibits growth of both androgen-dependent and castration-resistant VCaP xenografts. Ganitumab does not affect growth of aggressive castration-resistant 22Rv1 xenografts. We found that ganitumab is highly effective against VCaP
21 9 xenografts when combined with androgen-deprivation therapy, however some instances of acquired resistance to ganitumab occurred. Additionally we developed and characterized an in vitro model of acquired ganitumab resistance, which we termed VCaP/GanR, derived from the VCaP prostate cancer cell line. Using VCaP/GanR cells as a model, we evaluated the mechanism of acquired resistance to ganitumab. Unlike the parental VCaP, VCaP/GanR cells did not undergo apoptosis after ganitumab treatment; additionally, apoptosis was prevented in VCaP/GanR after serum starvation. While VCaP/GanR cells exhibited increased mtor activity, attenuation of mtor signaling was not sufficient to restore sensitivity to ganitumab. Lastly we found that acquired resistance to ganitumab in VCaP/GanR cells was dependent on calcium, outlining a novel resistance mechanism that impacts cell proliferation through cell cycle alteration.
22 CHAPTER 2: MATERIALS AND METHODS Cell culture and chemical reagents The human prostate cancer cell lines LNCaP.FGC (CRL 1740; batch F ) and CWR-22Rv1 (CRL-2505, batch ) were obtained from American Type Culture Collection (ATCC). LNCaP and 22Rv1 cells were authenticated and certified by ATCC. ATCC ensures each cell line is negative for mycoplasma, bacteria, and fungi contamination; confirms species identity; and performs DNA profiling and cytogenetic analysis to authenticate each cell line. RWPE immortalized prostate epithelial cells were also obtained from ATCC. CWR-R1 cells were provided by Dr. Elizabeth Wilson (University of North Carolina, Chapel Hill, NC) in July 2011 and tested for prostate and cell line specific characteristics including AR and AR splice variants (54, 55). VCaP cells were provided in March 2009 by Dr. Kenneth Pienta (University of Michigan, Ann Arbor, MI). VCaP cells were tested for prostate and cell line specific characteristics including AR, TMPRSS:ERG fusion, and PSA. VCaP and 22Rv1 cells were negative for mycoplasma, human T-lymphotropic virus, hepatitis (A, B, and C), and HIV. All cell lines were used within 6 months of resuscitation. LNCaP, CWR-R1 and CWR-22Rv1 were maintained in RPMI (Cellgro) supplemented with 100 IU/mL penicillin, 100 µg/ml streptomycin, 2 mm L- glutamine (Life Technologies), and 10% fetal bovine serum (FBS)(Atlanta Biologicals). VCaP cells were maintained in high glucose DMEM (DMEM-H) supplemented with 1x antimycotic/antimycotic solution (Gibco) and 10% FBS. 10
23 11 RWPE cells were maintained in Keratinocyte Serum Free Medium (K-SFM) (Gibco) supplemented with bovine pituitary extract and epidermal growth factor. Ganitumab and anti-streptavidin control IgG1 antibody were obtained from Amgen, Inc. U73122, rapamycin, metformin, and AICAR were obtained from Cayman Chemicals. PF was obtained from Sigma-Aldrich. BAPTA-AM and EGTA-AM were obtained from Calbiochem. Recombinant human IGF-1 was obtained from Peprotech. Generation of ganitumab-resistant VCaP derivatives VCaP were passaged in 500 nm ganitumab for 12 weeks and were termed VCaP/GanR (ganitumab-resistant). A second ganitumab resistant derivative termed VCaP/GanR-2 was derived in the same manner. Both VCaP/GanR and VCaP/GanR-2 were maintained in the presence of 500 nm ganitumab. VCaP/GanR cells were withdrawn from ganitumab for 8 weeks and this derivative was termed VCaP/GanWD. All parental VCaP cells used in these studies were passage-matched with VCaP/GanR. In vitro AKT phosphorylation studies VCaP and 22Rv1 were plated in medium containing 10% FBS. At ~70% confluence, medium were replaced with fresh medium supplemented with 5% charcoal-stripped serum and cells cultured for an additional 24 hours. Ganitumab ( nm) was added 90 minutes prior to treatment with 1nM IGF-1. Cells were harvested in RIPA buffer 30 minutes after IGF-1 administration and immunoblotted.
24 12 In vitro apoptosis studies 6.5x10 5 VCaP, 22Rv1, CWR-R1, or LnCaP were plated in the appropriate medium containing 10% FBS in 60mm plates. The following day, cell monolayers were washed with PBS, and medium supplemented with 2% FBS containing 500 nm ganitumab or vehicle was added. Plates were incubated for 3 days, harvested in RIPA buffer and immunoblotted. Data represent at least two independent experiments. In vitro proliferation assays LNCaP, 22Rv1, or CWR-R1 cells were seeded in 24-well plates (BD Falcon) (2x10 4 per well) in RPMI/10% FBS. RWPE cells were seeded in 24-well plates (BD Falcon) (1x10 4 per well) in normal growth medium (K-SFM). VCaP, VCaP/GanR, VCaP/GanR-2, or VCaP/GanWD were seeded in 24-well plates (BD Falcon) (4.5x10 4 cells) in DMEM-H (Gibco) supplemented with 10% FBS (Atlanta Biologicals). The following day medium was exchanged for regular or calcium-free medium supplemented with 2% FBS and drug or vehicle as indicated. Cells were incubated for six days, with media exchanged every three days. Cells were trypsinized (Cellgro), mixed with trypan blue (Gibco), and live cells were counted using a hemocytometer. Xenograft studies Studies involving animals were conducted in a manner approved by the University of Miami Animal Care and Use Committee. VCaP cells ( ) were injected subcutaneously with an equal volume of Matrigel (BD Biosciences) into both hind flanks of severe-combined immunodeficient mice (Harlan). 22Rv1 cells
25 13 (2x10 6 ) were injected subcutaneously into both hind flanks of castrated nude mice (Harlan) with Matrigel (BD Biosciences). Tumor volume was measured at least twice weekly with calipers calculated using the formula length width height Mice were euthanized when tumors reached a maximal volume of 1000mm 3 or 10 weeks post-treatment for androgen-replete VCaP xenografts; 1000 mm 3 or 17 weeks post-treatment for castration-resistant VCaP xenografts; and 1000 mm 3 or 7 weeks post-treatment for castration-resistant 22Rv1 xenografts; upon which tumors were excised, and flash frozen. Levels of circulating PSA were quantified from serum samples by an enzyme-linked immunosorbent assay (Biocheck Inc.). Levels of IGF-1R, INSR, phospho-akt and total AKT were measured using either MSD multiplex assays (Meso Scale Discovery) as previously described (25, 40) or western blot followed by densitometry using Photoshop CS3. Ganitumab (Amgen) and control antistreptavidin IgG1 antibody (Amgen) were diluted in sterile PBS and administered via intraperitoneal injections at a dosage of 300µg twice weekly as specified. Western blots 8.5x10 5 VCaP, VCaP/GanR, VCaP/GanR-2, or VCaP/GanWD, were plated in the appropriate medium containing 10% FBS in 60 mm plates. The following day cell monolayers were washed with PBS, and medium supplemented with 2-10% FBS as indicated was added with treatment or vehicle. Plates were incubated for time periods as indicated, harvested in RIPA buffer and cell lysates immunoblotted.
26 14 Samples were run in 10-12% SDS-PAGE, then transferred onto nitrocellulose membranes (Biorad). Membranes were blocked with 5 % milk in tris-buffered saline (TBS) supplemented with 0.1% Tween-20 (Sigma-Aldrich) or 5% bovine serum albumin (BSA) (Sigma-Aldrich) in TBS supplemented with 0.1% Tween-20 for phospho-specific antibodies. Antibodies against phospho- AKT, total AKT, cleaved PARP, IGF-1R, phospho-s6, PRAS40, and total S6 were obtained from Cell Signaling Technologies. The antibody against Rb was obtained from Oncogene Research Products. Antibodies against androgen receptor (N-20), actin, INSR, cyclin A, and all secondary HPRT conjugated antibodies were obtained from Santa Cruz. Both primary and secondary antibodies were diluted in 5% milk in TBS-Tween except for primary phosphospecific antibodies which were diluted in 5% BSA in TBS-Tween. Immunoblots were developed using an enhanced chemiluminescence detection spray (Denville Scientific). Densitometry was performed using Adobe Photoshop CS3. Serum starvation cell proliferation assay VCaP and VCaP/GanR were seeded in 3 separate 24-well plates (BD Falcon) (5x10 4 cells) in DMEM-H (Gibco) supplemented with 10% FBS (Atlanta Biologicals). The following day cells were washed with PBS, and one plate trypsinized and counted as detailed above to serve as Day 0 timepoint. Cells were incubated with medium containing 10% charcoal-stripped serum (CSS), and a plate was counted on Day 3 and Day 6 as detailed above.
27 15 Phospho-proteome kinase array 5x10 6 VCaP or VCaP/GanR cells were plated in 100 mm plates (BD Falcon) in media containing 10% FBS. The following day media was replaced with DMEM-H containing 10% FBS and 500 nmol/l ganitumab or control IgG1 antibodies and incubated for six hours prior to harvest. Proteome Profiler Human Phospho-Kinase Arrays ARY003B were obtained from R&D systems and performed following the manufacturer s protocol. Densitometry was performed using Adobe Photoshop CS3. Subcellular fractionation 5x10 6 VCaP or VCaP/GanR cells were plated in 100 mm plates (BD Falcon) in media containing 10% FBS. The following day media was aspirated and replaced with DMEM-H containing 10% FBS and 500 nm ganitumab or control IgG1 antibody and incubated for six hours prior to harvest. Subcellular fractionation was performed according to the manufacturer s protocol (Thermo Scientific).
28 CHAPTER 3: PRECLINICAL EFFICACY OF GANITUMAB Ganitumab inhibits IGF-1R signaling in androgen-dependent and castration-resistant prostate cancer cells in vitro To assess the potential use of ganitumab for treatment of both androgendependent and castration-resistant prostate cancer, we determined whether androgen-dependent VCaP and castration-resistant 22Rv1 were sensitive to IGF-1 stimulation. Serum deprivation decreased AKT phosphorylation while IGF- 1 treatment induced phosphorylation of AKT, a downstream effector of IGF- 1/IGF-1R signaling in both VCaP and 22Rv1 cell lines (Figure 3a-b). Administration of ganitumab reduced IGF-1-induced phosphorylation of AKT in both cell lines. These results indicate that ganitumab inhibited IGF-1/IGF-1R signaling in both androgen-dependent and castration-resistant prostate cancer cells. Figure 3. Analysis of ganitumab effects on IGF-1R signaling and prostate cancer cell proliferation in vitro. VCaP and 22Rv1 were cultured in serum deprived media for 24 hours, and treated with ganitumab [0 nm, 10 nm, 100 nm, 1000 nm] for 90 minutes prior to treatment with IGF-1(1 nmol/l) for 30 minutes. Lysates were probed by western blot for phospho-akt then total-akt in VCaP (A) and 22Rv1 (B). The effects of six day ganitumab treatment ( nm) on prostate cancer cell proliferation in 2% FBS containing media [VCaP (C), 22Rv1 (D), CWR-R1 (E), and LNCaP (F)] were determined. (G) The effects of six day ganitumab treatment ( nm) on RWPE cells in normal growth medium were assessed. (H) VCaP, 22Rv1, CWR-R1 and LNCaP were cultured in medium containing 2%FBS for 72 hours with ganitumab (500 nmol/l) or control. Lysates were immunoblotted for cleaved PARP and actin. Panels (C-E) are representative of 3 experiments performed in triplicate. (F) Represents 3 combined experiments. (G) is representative of 2 independent experiments performed in triplicate. All data are shown ± SD. (*p<.05, ***<.001, Two-tailed Student s t-test) 16
29 17
30 18 Ganitumab inhibits growth of androgen-dependent and castration-resistant prostate cancer in vitro To assess the effect of ganitumab on prostate cancer cell proliferation, we administered ganitumab to several prostate cancer cell line models in vitro. The proliferation of VCaP, 22Rv1, CWR-R1 and LNCaP cells was inhibited by ganitumab to varying degrees (Figure 3c-f). Immortalized normal prostate epithelial RWPE cells were also sensitive to ganitumab similar to that of 22Rv1 and LNCaP (Figure 3g). VCaP cells were the most sensitive to ganitumab. Ganitumab increased the level of cleaved PARP, a late-stage marker of apoptosis, in VCaP cells, but had no effect on cleaved PARP levels in 22Rv1, CWR-R1 or LNCaP cell lines (Figure 3h). We chose an androgen-dependent (VCaP) and a castration-resistant (22Rv1) model for further studies in vivo. Ganitumab inhibits androgen-dependent prostate cancer growth in vivo We utilized androgen-dependent VCaP xenografts to evaluate ganitumab treatment in a more physiological setting. VCaP human prostate cancer cells were injected subcutaneously into intact mice. When tumors reached an average volume of mm 3, mice were randomized into two groups and treatment with ganitumab or control antibody at a dose of 300µg twice weekly via intraperitoneal injections commenced. The ganitumab dosage was based on safety and efficacy shown in previous mouse xenograft studies (25, 40). Ganitumab inhibition of tumor growth was observed beginning one week after treatment initiation (Figure 4a). Growth inhibition was maintained as shown by
31 19 the tumor doubling time, which was strikingly increased from 2.3±0.4 weeks in control-treated mice to 6.4±0.4 weeks in ganitumab-treated mice (Figure 4b). The clinically relevant biomarker prostate-specific antigen (PSA) was also assessed throughout the duration of the experiment. The PSA gene is regulated by AR and circulating PSA is used to assess biochemical recurrence in patients (56). Serum PSA doubling time was increased from 1.6±0.5 weeks in controltreated mice to 4.1±0.9 weeks in ganitumab-treated mice which paralleled changes in tumor volume (Figure 4c). Levels of IGF-1R were significantly decreased in ganitumab-treated mice compared to control-treated mice, and INSR levels were unaffected by ganitumab treatment (Figure 4d-f). Ganitumab had no effect on levels of AR (Figure 4g,h). These data show that ganitumab slows the growth of this androgen-dependent prostate cancer model. Figure 4. Ganitumab inhibits the growth of androgen-dependent VCaP xenografts. VCaP xenografts were established in intact (androgen-replete) mice. When tumors reached an average volume of 278 mm 3, treatment with ganitumab or control antibody was initiated. (A) Tumor volumes (mm 3 ) are shown (n=3-4 for control; n=4-8 for ganitumab). (B) Tumor doubling times for control and ganitumab treated mice are shown (n=4 for control; n=8 for ganitumab). (C) Serum PSA doubling times are shown (n=4 for control; n=8 for ganitumab). (D) Xenograft lysates were immunoblotted for IGF-1R, INSR, and actin for control- and ganitumab-treated tumors. Western blots were quantified by densitometry and relative IGF-1R (E) and relative INSR (F) normalized to actin are shown. (G) Tumor lysates were immunoblotted for AR (including AR LBD) and actin for control- and ganitumab-treated tumors. (H) Western blots were quantified for AR and AR LBD by densitometry and plotted normalized to actin. All data are displayed ± SEM. (*p<.05, **<.01, Two-tailed Student s t-test)
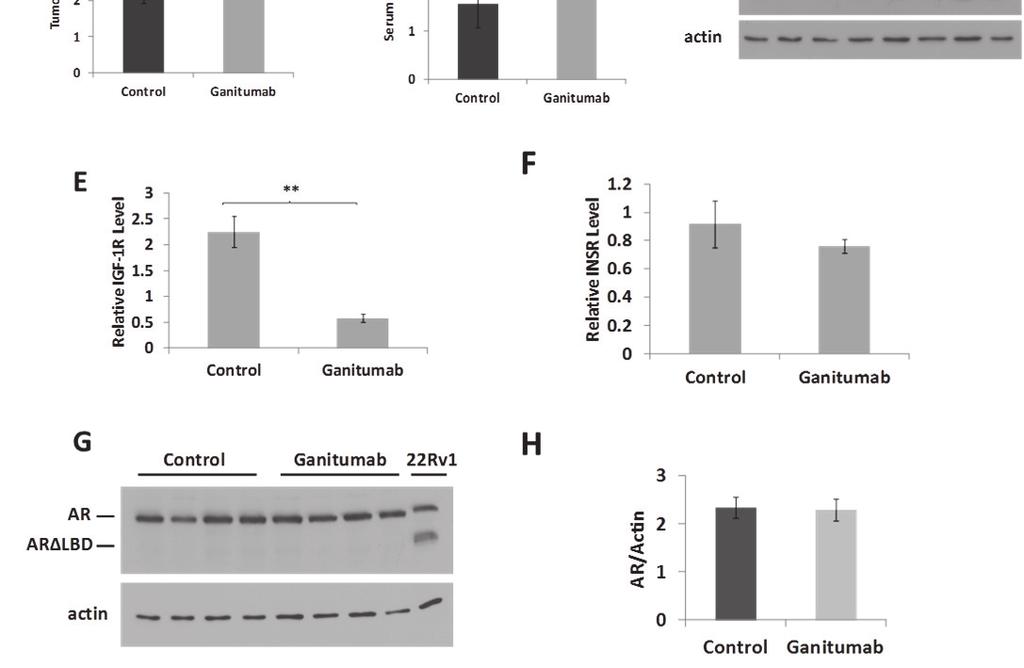
32 20
33 21 Effect of ganitumab treatment on castration-resistant prostate cancer in vivo To test whether ganitumab may be effective for CRPC, we utilized two xenograft models, VCaP and 22Rv1. VCaP xenografts are initially androgendependent, but recur as castration-resistant xenografts after murine castration (49, 57). 22Rv1 is a castration-resistant model of prostate cancer that grows readily in androgen-deprived environments including xenografts in castrated mice (52). VCaP cells were injected into the flanks of intact SCID mice, and all mice were castrated when average tumor volumes reached 260~320 mm 3. Five weeks after castration, mice with tumors averaging 538 mm 3 were randomized into 2 groups and treatment with ganitumab or continued control antibody was initiated. This experimental design allowed evaluation of ganitumab effectiveness for tumors that had recently progressed to castration-resistance. Ganitumab halted tumor growth for the remainder of the experiment (11.5 weeks) whereas the tumors of the control-treated mice continued to grow rapidly (Figure 5a). The ganitumab-treated mice showed decreased serum PSA relative Figure 5. Ganitumab inhibits the growth of castration-resistant VCaP xenografts. Castrationresistant VCaP xenografts were established and mice were treated with control antibody beginning one week after castration. Five weeks after castration with an average tumor volume of 538 mm 3, mice were randomized into 2 groups and treatment with ganitumab was initiated (Control Ganitumab) or control antibody was continued (Control Control). (A) Tumor volumes (mm 3 ) are shown (n=4-5 for control; n=3-4 for ganitumab). (B) Serum PSA (ng/ml) levels are shown (n=18 for pre-treatment; n=3-4 for control; n=3 for ganitumab). (C) Xenograft lysates were immunoblotted for IGF-1R, INSR, and actin for end point Control Control and Control Ganitumab tumors. Blots were quantified by densitometry and relative IGF-1R (D) and relative INSR (E) normalized to actin are shown. (F) Tumor lysates were immunoblotted for AR (including AR LBD) and actin for control- and ganitumab-treated tumors. Western blots were quantified for AR (G) and AR LBD (H) by densitometry and plotted normalized to actin. All data are displayed ± SEM. (*p<.05, **<.01, Two-tailed Student s t-test)
34 22
35 23 to the control-treated mice at all comparable time points (Figure 5b). Although tumor volume did not significantly increase in the ganitumab-treated mice, the serum PSA increased at later time points (15 weeks post-castration). Levels of IGF-1R were significantly decreased in ganitumab-treated mice compared to control-treated mice (Figure 5c,d). The levels of INSR, an RTK related to IGF-1R that has been implicated in resistance to ganitumab (25), was unaffected by ganitumab treatment (Figure 5c,e). Additionally levels of AR and AR splice variants (AR LBDs) were unaffected by ganitumab treatment (Figure 5f-h). AR LBDs lack the ligand binding domain and are considered to be a constitutively androgen receptor. Levels of AR and AR LBDs are important as prostate cancer, even CRPC, still relies on AR transcriptional activity. Increases in AR and AR LBDs could be indicative of increased severity or progression of the disease state. 22Rv1 cells were injected into the hind flanks of castrated nude mice. Once tumors reached an average volume of 260~320 mm 3, treatment with ganitumab or control antibody commenced for 5 weeks. In this highly aggressive model of prostate cancer, ganitumab had no appreciable effect on tumor growth (Figure 6a). There was no difference in weight between control and ganitumabtreated mice (Table 2). Protein levels of IGF-1R were significantly decreased in ganitumab-treated mice compared to control treated mice (Figure 6b,c). INSR, AR, and AR LBD levels were not significantly changed after treatment with ganitumab (Figure 6b,d-g).
36 24 Table 2. Mouse weights in 22Rv1 xenograft studies. Weights of mice (g) ± SEM are shown for 22Rv1 xenoraft studies in nude mice. Since ganitumab showed no effect in castration-resistant 22Rv1 xenograft growth, we assessed the sensitivity of 22Rv1 to IGF-1 treatment in vitro. IGF-1 treatment (1-100 nm) induced cell proliferation (Figure 6h). Additionally, ganitumab treatment abrogated the IGF-1 induced proliferation effects showing that ganitumab does block IGF-1 activation of IGF-1R in 22Rv1 (Figure 6i). Figure 6. Ganitumab does not affect the growth of well-established castration-resistant 22Rv1 xenografts. 22Rv1 xenografts were established in castrated nude mice. When tumors reached an average volume of 265 mm 3 treatment with ganitumab or control antibody was initiated. (A) Tumor volumes (mm 3 ) are shown for ganitumab- and control-treated mice (n=4-14 for control, n=3-14 for ganitumab). (B) Xenograft lysates were immunoblotted for IGF-1R, INSR, and actin. Blots were quantified by densitometry and relative IGF-1R levels (C) and relative INSR levels (D) normalized to actin are shown. (E) Tumor lysates were immunoblotted for AR (including AR LBD) and actin for end point control- and ganitumab-treated tumors. Western blots were quantified for AR (F) and AR LBD (G) by densitometry and plotted normalized to actin. (H) 22Rv1 cells were treated with IGF-1 (0-100 nmol/l) for six days in medium containing 2% FBS containing media and proliferation relative to vehicle treatment is shown. (I) 22Rv1 cells were treated with IGF-1 (1 nmol/l) alone and in combination with ganitumab (500 nmol/l) for six days in medium containing 2% FBS. Panels (A,C,D,F,G) are displayed ± SEM. (H) represents three combined experiments performed in triplicate and is shown ±SD. (I) is representative of three independent experiments performed in triplicate and is shown ±SD. (*p<.05, Two-tailed Student s t-test)
37 25
38 26 Ganitumab combined with androgen-deprivation is a highly effective treatment regimen The standard of care for advanced or metastatic prostate cancer is androgen-deprivation therapy. Since ganitumab showed inhibitory effects on both androgen-dependent and castration-resistant VCaP prostate cancer xenografts, we tested whether combining ganitumab with androgen-deprivation therapy would show added benefits. VCaP cells were implanted into the hind flanks of SCID mice, and all mice were castrated when average tumor volume reached mm 3. One week later, treatment with ganitumab or control antibody was initiated. A single dose of ganitumab resulted in an immediate decrease in tumor volume compared to control-treated mice after castration (Figure 7a). Tumors of ganitumab-treated mice remained significantly smaller than those of control-treated mice throughout the remainder of the experiment (16 weeks). In addition to decreased tumor volume, combining androgendeprivation therapy with ganitumab lengthened time to recurrence in the 17 week experiment from 3.1±0.7 weeks (control-treated) to 16.5±0.7 weeks (ganitumabtreated) with recurrence defined as tumor volume surpassing that at time of castration as determined for each tumor individually (Figure 7b). Serum PSA decreased to a greater extent with androgen-deprivation and ganitumab when compared to androgen-deprivation combined with control antibody (Figure 7c). Between weeks 11 and 15 there was a sharp increase in serum PSA which was not reflected in an increase in tumor volume. Ganitumab-treated mice lost a small proportion of body weight (~10%) whereas control-treated mice did not lose
39 Figure 7. Combining ganitumab with androgen-deprivation therapy most effectively inhibits VCaP xenografts. VCaP xenografts were established in intact mice, and mice were castrated when tumors reached an average volume of 318 mm 3. One week after castration mice were randomized into 2 groups and treatment with ganitumab or control antibody was initiated. (A) Tumor volumes (mm 3 ) are shown relative to time of castration for control and ganitumab treated mice (n=4-14 for control; n=5-16 for ganitumab) (control-treated tumor volumes are those from figure 4a). (B) Weeks observed without recurrence were calculated and are shown above (n=11 for control; n=5 for ganitumab) (C) Serum PSA (ng/ml) is shown (n=3-13 for control; n=5-11 for ganitumab). (*p<.05, ***p<.001, Two-tailed Student s t-test) 27
40 28 weight during the experiment (Table 3). Total levels of IGF-1R protein in xenografts were decreased in the ganitumab-treated mice compared to intact mice (Figure 8a) and also in previous studies in pancreatic cancer and Ewing s sarcoma (25, 40). INSR levels remained unchanged under all conditions tested (Figure 8b). Levels of phosphorylated AKT and total AKT were also unaffected by androgendeprivation, ganitumab treatment, or combined androgen-deprivation and ganitumab treatment (Figure 8c,d). AR and AR LBD variant AR3 (also known as AR-V7) mrna levels were increased in end stage ganitumab-treated Table 3. Mouse weights in VCaP xenograft studies. Weights of mice (g) ± SEM are shown for VCaP tumors (10 weeks postcastration) compared to control-treated tumors (17 weeks xenoraft studies in SCID mice. post-castration) (Figure 8e,f). These data show that combination of ganitumab and androgendeprivation substantially decreased tumor growth and delayed progression to castration resistance.
41 Figure 8. Ganitumab reduces IGF-1R levels when combined with androgen-deprivation therapy. Tumors from Figure 7 were assessed for relative IGF-1R (A), INSR (B), phospho-akt (C), and total AKT (D). Protein levels were detected by MSD multiplex analysis are shown for intact (pre-castrate baseline), control-treated, and ganitumab-treated mice relative to time of castration (n=3-5 per group). Relative mrna levels of (E) AR and (F) AR3 (AR-V7) were assessed by reverse transcriptase realtime PCR for intact (pre-castrate baseline), control- and ganitumab-treated mice relative to time of castration (n=3-4 per group). Data are normalized using HPRT1 and plotted relative to intact (pre-castrate baseline). All data are displayed ± SEM. (*p<.05, ***<.001, Two-tailed Student s t-test) 29
42 30 Reversibility and durability of treatment with ganitumab combined with androgen-deprivation therapy We evaluated the durability of combining androgen-deprivation therapy with ganitumab treatment by examining long-term ganitumab treatment. Reversibility of ganitumab treatment was also assessed. VCaP xenografts were established and mice castrated when tumors reached an average volume of mm 3. Treatment with ganitumab commenced one week after castration. Five weeks after castration, mice were randomized into two groups. One group remained on ganitumab (to evaluate durability of response) and ganitumab was discontinued in the other group (to evaluate reversibility of response). In the ganitumab discontinued cohort, all tumors (n=4) recurred within 11 weeks (Figure 9a). Two of five tumors recurred in the long-term ganitumab treatment arm (Figure 9b). Thus, while combination therapy with androgen-deprivation and ganitumab is highly effective, tumors recur after ganitumab cessation and some tumors recur during long-term ganitumab treatment outlining the possibility of acquired resistance to ganitumab in the clinical setting. Figure 9. Evaluation of durability and reversibility of combined ganitumab and androgen-deprivation therapy on VCaP xenografts. VCaP xenografts were established in intact mice, and mice were castrated when tumors reached an average volume of 318 mm 3. Beginning one week after castration mice were treated with ganitumab for four weeks. Mice were randomized into 2 groups and treatment with control antibody was initiated or ganitumab was continued. (A) Discontinuation of ganitumab after 4 weeks (5 weeks post castration) resulted in recurrence of all VCaP tumors (n=4) (ϯ deceased mouse due to causes unrelated to treatment). (B) Long-term treatment with ganitumab resulted in recurrence in 2 of 5 tumors. (Average tumor volume is reflected in figure 6a.) (C) Quantification of recurrence after ganitumab discontinuation (reversibility) and long-term ganitumab treatment (durability) is shown.
43 31
44 CHAPTER 4: ACQUIRED RESISTANCE TO GANITUMAB Development of a ganitumab resistant prostate cancer cell derivative To develop an in vitro model in which to examine mechanisms of resistance to ganitumab, VCaP cells were passaged in 500 nmol/l ganitumab for 12 weeks at which point significant cell proliferation was evident (Figure 10a). These ganitumab resistant VCaP cells (termed VCaP/GanR) were routinely Figure 10. Derivation of a ganitumab resistant prostate cancer VCaP line termed VCaP/GanR. VCaP prostate cancer cells were passaged in medium supplemented with 10% fetal bovine serum and ganitumab (500 nmol/l) for 12 weeks until proliferation became apparent. (A) Images of VCaP and VCaP/GanR were obtained at a magnification of 10x. Parental VCaP and derivative VCaP/GanR were treated with ganitumab ( nmol/l) (B) or ( nmol/l) (C) in medium containing 2% FBS and relative proliferation is displayed ±SD. Panel (B) represents three combined independent experiments performed in triplicate. Panels (C,D) are representative of two independent experiments performed in triplicate. 32
, VCaP/GanR cells were not substantially growth inhibited by ganitumab (Figure 10c).")
45 33 maintained in 500 nmol/l ganitumab. Treatment of parental, passage-matched VCaP cells with ganitumab significantly decreased cell proliferation compared to VCaP/GanR (Figure 10b). Even at higher concentrations of ganitumab (2000 nmol/l), VCaP/GanR cells were not substantially growth inhibited by ganitumab (Figure 10c). To determine whether the anti-proliferative effects of ganitumab were due to decreased proliferation or increased cell death, ganitumab was administered to parental VCaP and VCaP/GanR cells and levels of cleaved PARP, a marker of apoptosis, and cyclin A, a marker of cell cycle G1 to S progression, were assessed. Ganitumab increased cleaved PARP in parental VCaP cells, but little cleaved PARP was evident in VCaP/GanR cells and there was no change with ganitumab treatment (Figure 10d). In contrast, cyclin A was decreased after ganitumab treatment in both cell lines. An additional ganitumab resistant cell model (termed VCaP/GanR-2) was derived in the same manner as VCaP/GanR and exhibited similar resistance to ganitumab as VCaP/GanR as shown by cleaved PARP and cell proliferation assays (Figure 11a,b). Figure 11. Derivation of a second ganitumab resistant prostate cancer VCaP cell line termed (VCaP/GanR-2). VCaP prostate cancer cells were passaged in medium supplemented with 10% fetal bovine serum and ganitumab (500 nmol/l) for 12 weeks until proliferation became apparent. (A) Parental VCaP and VCaP/GanR were treated with ganitumab (500 nmol/l) or vehicle for 72 hours in medium containing 2% FBS. Lysates were probed for CL-PARP and actin. (B) VCaP/GanR-2 cells were treated with ganitumab (500 nmol/l) or vehicle for six days in medium containing 2%FBS. Fold proliferation is shown relative to vehicle treatment (± SD). Panel (A) is representative of two independent experiments. Panel (B) is representative of three independent experiments performed in triplicate.
 VCaP, VCaP/GanR, and VCaP/GanWD were treated with ganitumab (500 nmol/l) or vehicle for six days in medium containing 2%FBS. Fold proliferation is shown relative to vehicle treatment (± SD).")
46 34 Figure 12. Acquired resistance to ganitumab in VCaP/GanR is stable. VCaP/GanR cells were withdrawn from ganitumab for 8 weeks and now termed VCaP/Gan-WD. (A) VCaP, VCaP/GanR, and VCaP/GanWD were treated with ganitumab (500 nmol/l) or vehicle for six days in medium containing 2%FBS. Fold proliferation is shown relative to vehicle treatment (± SD). (B) VCaP, VCaP/GanR, and VCaP/GanWD were treated with ganitumab (500 nmol/l) or vehicle for 72 hours in medium containing 2% FBS. Lysates were probed for CL- PARP and actin. To determine whether the acquired resistance to ganitumab was a stable change, VCaP/GanR cells were withdrawn from ganitumab for 8 weeks (termed VCaP/GanWD). VCaP/GanWD were rechallenged with ganitumab and found to maintain equal resistance to ganitumab as VCaP/GanR as assessed by cell proliferation assays (Figure 12a). Additionally, ganitumab did not increase levels of cleaved PARP in VCaP/GanWD suggesting that acquired resistance to ganitumab was a stable phenotype (Figure 12b). To evaluate whether the resistance to apoptosis and decreased cell proliferation of VCaP/GanR was specific to ganitumab treatment, VCaP/GanR and parental VCaP cells were cultured in medium containing 10% charcoal stripped serum (CSS), which is androgen- and mitogen-depleted. VCaP/GanR did not undergo apoptosis as evidenced by lack of cleaved PARP and these cells also maintained higher levels of cyclin A (Figure 13a). VCaP/GanR continued to proliferate in mitogen- and steroid-depleted medium whereas VCaP decreased in
.")
47 35 cell number (Figure 13b). VCaP and VCaP/GanR were treated with chemotherapeutic agent 5- fluorouracil. 5-fluorouracil had no effect of cyclin A levels in VCaP or VCaP/GanR and induced apoptosis in VCaP cells but not VCaP/GanR (Figure 13c). 5-fluorouracil did not induce apoptosis in VCaP/GanR, again outlining an increased avoidance to apoptosis. Evaluation of IGF-1R related signaling pathways in VCaP/GanR VCaP and VCaP/GanR cells Figure 13. Characterization of human prostate cancer VCaP derivative VCaP/GanR. (A) VCaP and VCaP/GanR cells were cultured in medium containing 10% charcoal stripped serum, treated for the indicated times, and probed for cleaved PARP, cyclin A and actin. (B) Cell proliferation of VCaP and VCaP/GanR in medium containing 10% CSS for the indicated times was assessed. (C) VCaP and VCaP/GanR were treated with 5-fluorouracil (50 µmol/l) or vehicle 72 hours in medium containing 2% FBS. Lysates were probed for cleaved PARP, cyclin A and actin. Panels (A,C) are representative two independent experiments. Panel (B) is representative of two independent experiments performed in triplicate. harbored similar basal levels of IGF- 1R and treatment with ganitumab reduced IGF-1R protein levels in both VCaP and VCaP/GanR cells (Figure 14a). The closely related RTK insulin receptor (INSR), whose overexpression is a proposed mechanism of resistance to ganitumab in Ewing s Sarcoma, was
.")
48 36 also assessed (25). VCaP and VCaP/GanR cells contained similar levels of INSR. VCaP cells showed an acute increase in INSR after ganitumab treatment, which was not observed in the VCaP/GanR cells (Figure 14b). These results suggest that overexpression of either IGF-1R or INSR to compensate for loss of IGF-1R signaling after ganitumab treatment does not underlie ganitumab resistance in VCaP/GanR. Further, unlike parental VCaP, VCaP/GanR did not upregulate INSR in response to ganitumab-mediated loss of IGF-1R signaling, which may indicate acquisition of alternative growth and survival signaling. Figure 14. Characterization of IGF-1R related signaling pathways. VCaP and VCaP/GanR were cultured in medium containing 2% FBS for 72 hours with 500 nmol/l ganitumab or vehicle. Lysates were immunoblotted for IGF-1R and actin (A), INSR and actin (B), or phosphorylated AKT then probed for total AKT (C). Panels (A- C) are representative of three independent experiments. AKT and ERK are downstream effectors of IGF-1R (18, 19) and phosphorylation status of these signaling molecules was assessed. AKT phosphorylation levels were decreased after ganitumab treatment in both cell lines (Figure 14c). ERK1/2 phosphorylation levels were similar between VCaP and VCaP/GanR cells and were not affected by ganitumab treatment (data not shown). These data suggest that resistance to ganitumab is not due to
49 37 reactivation of downstream IGF-1R signaling pathways through alternative means. Phospho-proteome kinase arrays To simultaneously assess protein phosphorylation changes that might underlie acquired resistance to ganitumab in VCaP/GanR, protein kinase phosphorylation arrays were performed. These arrays permit evaluation of the relative phosphorylation of 42 distinct signaling molecules. To determine the optimal time to examine differences in protein phosphorylation between VCaP and VCaP/GanR cells, a time course of ganitumab treatment (0-24 hours) was performed (Figure 15a). The arrays were conducted following a six hour ganitumab treatment as IGF-1R levels were reduced, but apoptosis had not yet increased in the control VCaP cells. This time frame should permit us to eliminate protein phosphorylation changes that were a consequence of apoptosis. Actual results are displayed in Figure 15b. Arrays were quantified by densitometry and complete results are shown relative to vehicle treatment of VCaP parental cells (Figure 15c). We found changes in phosphorylation of p27, two proteins (AMPK and PRAS40) that modulate activity of mammalian target of rapamycin (mtor), as well as several proteins that are subject to calcium regulation or are mediators of calcium signaling including CREB, PLCγ, PYK2, and c-jun. Alterations in protein phosphorylation were plotted relative to vehicle treatment of VCaP (Figure 16a). The array was also validated by western blot using a phospho-specific antibody against p-pras40 (Figure 16b).
 VCaP and VCaP/GanR were treated with ganitumab (500 nmol/l) for the indicated times in medium containing 10% FBS. Lysates were probed for IGF-1R, cleaved PARP, and actin.")
 displays the raw results obtained from the phosphoproteome profiling array. Each pair of dots represents the relative phosphorylation of a distinct signaling kinase.")
50 Figure 15. Phospho-proteome kinase profiling of VCaP and VCaP/GanR cells. (A) VCaP and VCaP/GanR were treated with ganitumab (500 nmol/l) for the indicated times in medium containing 10% FBS. Lysates were probed for IGF-1R, cleaved PARP, and actin. (B) VCaP and VCaP/GanR were treated with ganitumab (500 nmol/l) or control antibody for six hours followed by a phospho-proteome array. Panel (B) displays the raw results obtained from the phosphoproteome profiling array. Each pair of dots represents the relative phosphorylation of a distinct signaling kinase. (C) Results obtained in (B) were quantified using densitometry and full results are shown relative to VCaP control antibody treatment. 38
51 39 Phosphorylation of PYK2, a critical mediator of cell migration, proliferation and survival (58) was elevated in VCaP/GanR relative to parental VCaP cells (Figure 16a). To assess the possible contribution of increased PYK2 signaling to ganitumab resistance, VCaP and VCaP/GanR cells were treated with ganitumab, the PYK2 inhibitor PF alone, or in combination with ganitumab. Inhibition of PYK2 was not sufficient to restore sensitivity to ganitumab in VCaP/GanR cells (Figure 17a). While PF induced apoptosis in both VCaP and VCaP/GanR, VCaP/GanR displayed less apoptosis compared to VCaP (Figure 17b). Additionally, cyclin A Figure 16. Analysis and verification of phsopho-proteomic profiling. (A) Selected results from Figure 13c are shown relative to VCaP control antibody treatment. (B) VCaP and VCaP/GanR were treated with ganitumab or vehicle for six hours in 10% FBS and lysates were probed for p-pras40 and actin. levels were maintained longer in VCaP/GanR compared to VCaP after PF treatment (Figure 17b). PLCγ phosphorylation was increased in VCaP/GanR compared to
 VCaP and VCaP/GanR were treated for six days with PYK2 inhibitor (50 nmol/l) alone and in combination with ganitumab (500 nmol/l) in medium containing 2% FBS and proliferation relative to vehicle")
52 40 Figure 17. Effects of PYK2 inhibition, PLCγ inhibition and 5-FU treatment on VCaP/GanR. (A) VCaP and VCaP/GanR were treated for six days with PYK2 inhibitor (50 nmol/l) alone and in combination with ganitumab (500 nmol/l) in medium containing 2% FBS and proliferation relative to vehicle treatment is displayed ± SD. (B) VCaP and VCaP/GanR were cultured in medium supplemented with 2% FBS containing PF (100 nmol/l) or vehicle and probed for cleaved PARP, cyclin A, and actin. (C) VCaP and VCaP/GanR were treated for six days with PLCγ inhibitor U73122 (500 nmol/l) alone and in combination with ganitumab (500 nmol/l) in medium containing 2% FBS and proliferation relative to vehicle treatment is displayed ± SD. (D) VCaP and VCaP/GanR were treated with ganitumab (500 nmol/l) or vehicle for six hours in medium containing 10% FBS. Cellular fractionation was performed and fractionated lysates were probed for p27, histone, SOD1, and actin. Panels (A,C) are representative of 2 independent experiments performed in triplicate. parental VCaP cells (Figure 16a). However, inhibition of PLCγ using U73122 was not sufficient to restore sensitivity to the anti-proliferative effects of ganitumab in VCaP/GanR cells (Figure 17c). U73122 administration alone did not induce apoptosis in either VCaP or VCaP/GanR cells at the concentrations tested (data not shown).
53 41 The phospho-proteomic kinase array also showed an increase in p27 phosphorylation at threonine 198 in VCaP/GanR compared to VCaP under both vehicle and ganitumab treatments (Figure 16a). p27 phosphorylation at threonine 198 is known to increase cytoplasmic localization which promotes cellular proliferation and increases cellular motility (59). This change in localization of p27 may contribute to ganitumab resistance through increased progression through the cell cycle. VCaP and VCaP/GanR were treated with ganitumab or vehicle and subjected to subcellular fractionation; there was no apparent difference in the levels or localization of p27 comparing VCaP and VCaP/GanR (Figure 17d). These data suggest that p27 localization does not contribute to acquired resistance to ganitumab in VCaP/GanR. Elevated mtor activity does not promote acquired resistance to ganitumab mtor activity is implicated as a mechanism of resistance to RTK inhibition in several cancers (25, 26). The phospho-proteomic assays showed alterations in two regulators of mtor (AMPK and PRAS40) that have the potential to increase mtor activity in VCaP/GanR. To assess mtor activity, we treated VCaP and VCaP/GanR cells with ganitumab and monitored phosphorylated S6, a marker of mtor activity. S6 was highly phosphorylated in VCaP/GanR cells relative to VCaP cells following vehicle or ganitumab treatment (Figure 18a). Ganitumab treatment decreased levels of phosphorylated S6 in both parental VCaP and VCaP/GanR cells, which was apparent after one hour of ganitumab treatment (Figure 18b). VCaP/GanR-2 and VCaP/GanWD cells also
 VCaP and VCaP/GanR were treated with ganitumab (500 nmol/l) for the times indicated in medium containing 2% FBS.")
 VCaP and VCaP/GanR were treated with rapamycin (50 nmol/l), ganitumab (500 nmol/l) or the combination for six days in medium containing 2% FBS and cell proliferation is shown.")
54 Figure 18. mtor inhibition does not restore sensitivity to ganitumab in VCaP/GanR. (A) VCaP and VCaP/GanR were treated with ganitumab (500 nmol/l) or vehicle for 72 hours in medium containing 2% FBS. Lysates were probed for p-s6, then total S6 and actin. (B) VCaP and VCaP/GanR were treated with ganitumab (500 nmol/l) for the times indicated in medium containing 2% FBS. VCaP and VCaP/GanR-2 (C) or VCaP, VCaP/GanR, and VCaP/GanWD (D) were treated as in panel (A) and lysates probed for p-s6, then total S6 and actin. (E) VCaP and VCaP/GanR were treated with rapamycin (50 nmol/l), ganitumab (500 nmol/l) or the combination for six days in medium containing 2% FBS and cell proliferation is shown. VCaP and VCaP/GanR were treated with rapamycin (50 nmol/l), ganitumab (500 nmol/l) or the combination for 72 hours and probed for (F) phospho-s6 and actin or (G) phospho-akt then AKT. (A) is representative of three independent experiments performed in triplicate and is shown relative to vehicle treatment ± SD. (B,C) are representative of two independent experiments. In (B) image is cropped from the same immunoblot from the same exposure. 42
55 43 exhibited increased phosphorylated S6 (Figure 18c,d). Thus, mtor activity appeared to be elevated in models of ganitumab resistance in a stable (shown by VCaP/GanWD) and reproducible manner (shown by VCaP/GanR-2). To test whether increased mtor activity was responsible for ganitumab resistance, VCaP and VCaP/GanR cells were treated with rapamycin, an mtor inhibitor, alone and in combination with ganitumab. Rapamycin alone inhibited cell proliferation in VCaP but did not significantly decrease VCaP/GanR cell proliferation (Figure 18e). No increase in sensitivity to ganitumab was noted when rapamycin was combined with ganitumab. The concentration of rapamycin used was sufficient to decrease phosphorylated S6 in both VCaP and VCaP/GanR (Figure 18f). These results indicate that mtor activity is likely not responsible for acquired resistance to ganitumab. AKT phosphorylation was not increased after rapamycin treatment (Figure 18g). Thus, activation of compensatory mechanisms that have been shown to increase phospho-akt after rapamycin administration (60), are not likely responsible for the lack of ganitumab-sensitizing effects of rapamycin. Decreased AMPK activity does not contribute to acquired resistance to ganitumab AMPK phosphorylation was found to be decreased in VCaP/GanR cells relative to the parental VCaP (Figure 16a). In order to test whether this decreased phosphorylation of AMPK contributed to ganitumab resistance, VCaP and VCaP/GanR cells were treated with a range of metformin concentrations, an indirect activator of AMPK. Both cell lines were inhibited by metformin treatment,
.")
56 44 with VCaP/GanR being less sensitive than VCaP (Figure 19a). VCaP and VCaP/GanR cells were treated with a suboptimal dose of metformin alone and in combination with ganitumab. Combining metformin with ganitumab increased VCaP/GanR sensitivity to ganitumab, but did not restore full sensitivity (Figure 19b). Combining metformin with ganitumab did not restore the apoptosis- Figure 19. Metformin partially restores sensitivity to ganitumab in VCaP/GanR. (A) VCaP and VCaP/GanR were treated with metformin (0-15 µmol/l) or vehicle for six days in medium containing 2% FBS and fold cell proliferation is shown ± SD. (B) VCaP and VCaP/GanR were treated with metformin (5 µmol/l), ganitumab (500 nmol/l) or the combination for six days in medium containing 2%FBS and cell proliferation is shown ± SD. (C) VCaP and VCaP/GanR were treated with metformin (0-10 µmol/l) with ganitumab (500 nmol/l) or vehicle for 72 hours and probed for cleaved PARP and actin. (D) VCaP and VCaP/GanR were treated with metformin (5 µmol/l) for the times (hrs) indicated and probed for phospho-s6 then total S6. (E) 22Rv1 cells were treated with metformin (0-15 µmol/l) with ganitumab (500 nmol/l) or vehicle for six days in medium containing 2% FBS and proliferation relative to vehicle treatment is shown ± SD. Panels (A,E) represent two independent experiments performed in triplicate. Panel (B) represents three combined independent experiments performed in triplicate. (*<.05, Two-tailed Student s t-test)
.")
57 45 inducing effects of ganitumab in VCaP/GanR (Figure 19c). The suboptimal concentration of metformin was sufficient to activate AMPK as shown by decreased phosphorylation of mtor-regulated S6 phosphorylation (Figure 19d). In contrast, treating 22Rv1 cells with metformin and ganitumab did not increase sensitivity to ganitumab at any concentrations tested (Figure 19e). As metformin is an indirect activator of AMPK with many established effects in addition to AMPK activation (61), we employed an alternative activator of AMPK, AICAR (N 1 -(β-d-ribofuranosyl)-5-aminoimidazole-4-carboxamide). AICAR is a direct allosteric activator of AMPK. VCaP and VCaP/GanR were treated with a range of concentrations of AICAR and a dose dependent decrease in proliferation was noted for both cell lines (Figure 20a). When a Figure 20. AMPK activation by AICAR does not restore sensitivity to ganitumab in VCaP/GanR.. (A) VCaP and VCaP/GanR were treated with AICAR (0-750 µmol/l) or vehicle for six days in medium containing 2% FBS and fold cell proliferation is shown relative to vehicle treatment ± SD. (B) VCaP and VCaP/GanR were treated with AICAR (250 µmol/l), ganitumab (500 nmol/l) or the combination for six days in medium containing 2%FBS and cell proliferation is shown. (A) represents two independent experiments performed in triplicate. (B) represents four combined experiments performed in triplicate. Panels (A,B) are shown ± SD.
58 46 suboptimal dose of AICAR was combined with ganitumab, no increase in sensitivity to ganitumab was seen in VCaP/GanR (Figure 20b). These data suggest that decreased AMPK is not required for resistance to ganitumab and the partial ganitumab-sensitizing effects of metformin may have been due to an AMPK-independent mechanism. Acquired resistance to ganitumab is calcium dependent The phospho-proteome arrays implicated several signaling molecules that are regulated by intracellular calcium including: CREB, PLCγ, PYK2, and c-jun. These proteins showed increased phosphorylation (Figure 16a) in VCaP/GanR relative to VCaP under vehicle and ganitumab treatments. We used BAPTA-AM, a cell-permeable calcium chelator, which decreases intracellular calcium, to evaluate the contribution of calcium to ganitumab resistance. While BAPTA-AM had modest anti-proliferative effects on both VCaP and VCaP/GanR as a single agent, combining ganitumab with BAPTA-AM greatly decreased cell proliferation of VCaP/GanR cells compared to ganitumab alone (Figure 21a). Combining BAPTA-AM and ganitumab did not have an additive inhibitory effect on cell proliferation in passage-matched parental VCaP cells (Figure 21a). Thus, reduction in intracellular calcium sensitized VCaP/GanR to the anti-proliferative effects of ganitumab. We examined the effects of combined BAPTA-AM and ganitumab on apoptosis in VCaP/GanR. While ganitumab and BAPTA-AM substantially decreased cell proliferation, this combination did not result in increased levels of cleaved PARP in VCaP/GanR cells, suggesting that the decrease in cell number
59 47 was not due to apoptosis (Figure 21b). In addition, phospho-s6 was unchanged in VCaP/GanR when BAPTA-AM was administered alone, and no additive effect was apparent when BAPTA-AM was combined with ganitumab (Figure 21b). These results further validate the conclusion that mtor activity was not required for acquired resistance to ganitumab in VCaP/GanR cells. Because the combination of BAPTA-AM and ganitumab inhibited cell proliferation in VCaP/GanR without affecting apoptosis, markers of G1 to S cell cycle progression were examined. Ganitumab treatment decreased cyclin A levels in both VCaP and VCaP/GanR (Figure 21c). However, a larger decrease in cyclin A occurred in VCaP/GanR when ganitumab was combined with BAPTA- AM (Figure 21c). Combining ganitumab and BAPTA-AM also decreased hyperphosphorylated Rb (pp-rb) in both VCaP and VCaP/GanR also indicative of decreased G1 to S cell cycle progression (Figure 21c). These data suggest that BAPTA-AM restores sensitivity of VCaP/GanR cells to ganitumab through Figure 21. Treatment with cell permeable calcium chelators restores sensitivity to ganitumab. (A) VCaP and VCaP/GanR were treated for six days with with BAPTA-AM (1 µmol/l) alone or combined with ganitumab (500 nmol/l) in medium containing 2% FBS and cell proliferation is shown relative to vehicle treatments. VCaP and VCaP/GanR were cultured in medium supplemented with 2% FBS containing BAPTA-AM (1 µmol/l) alone or combined with ganitumab (500 nmol/l) for 72 hours. Lysates were immunoblotted for actin, cleaved PARP, phosphorylated S6, then probed for total S6 (B). (C) Upper Panel: Lysates were immunoblotted for cyclin A and actin or Rb and actin. Lower Panel: Quantification of cyclin A is shown normalized to actin from three independent experiments and shown relative to VCaP vehicle treatment ± SEM. (D) VCaP and VCaP/GanR cells were treated with EGTA-AM (1 µmol/l) alone or combined with ganitumab (500 nmol/l) for six days in medium containing 2%FBS and proliferation is shown ± SD. VCaP and VCaP/GanR were treated with EGTA-AM (1 µmol/l) alone or combined with ganitumab (500 nmol/l) for six days in medium containing 2%FBS for 72 hours. Lysates were probed for cleaved PARP, cyclin A and actin (E). (F) VCaP and VCaP/GanR cells were treated for 72 hours with ganitumab (500 nmol/l) or vehicle in either normal medium or calcium-free medium containing 2% FBS for 72 hours. Panels (A,D) represent 3 combined experiments performed in triplicate (± SD). (F) represents four combined experiments performed in triplicate (± SEM). Panels (B,C,E) are representative of at least 2 independent experiments. (*<.05, ***<.001, Two-tailed Student s t-test)
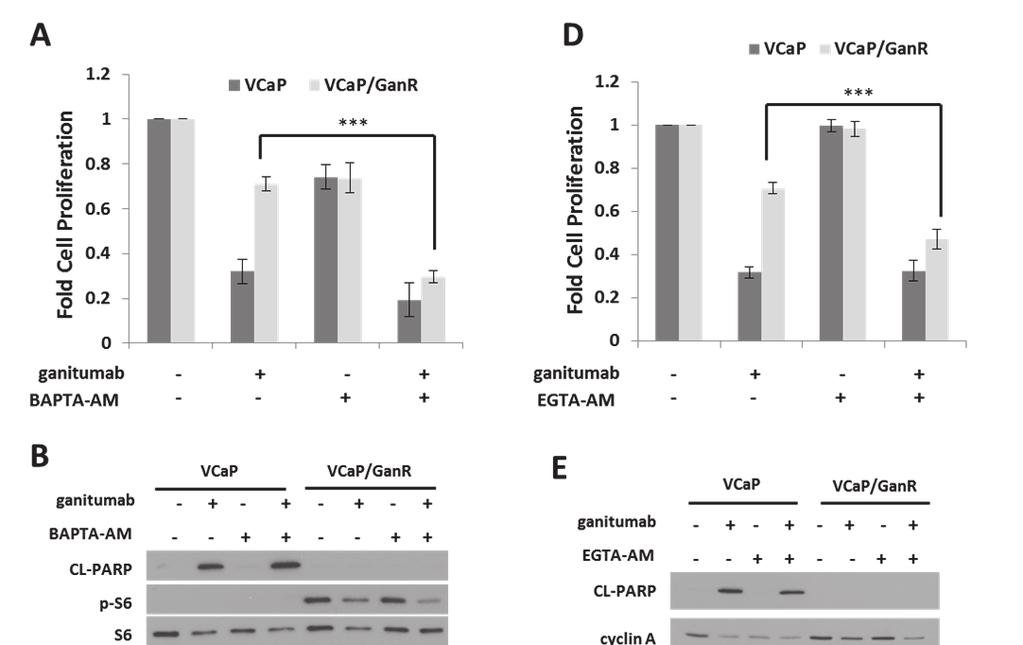
60 48
Supplementary Material
 Supplementary Material The Androgen Receptor is a negative regulator of eif4e Phosphorylation at S209: Implications for the use of mtor inhibitors in advanced prostate cancer Supplementary Figures Supplemental
Supplementary Material The Androgen Receptor is a negative regulator of eif4e Phosphorylation at S209: Implications for the use of mtor inhibitors in advanced prostate cancer Supplementary Figures Supplemental
Diabetes Mellitus and Breast Cancer
 Masur K, Thévenod F, Zänker KS (eds): Diabetes and Cancer. Epidemiological Evidence and Molecular Links. Front Diabetes. Basel, Karger, 2008, vol 19, pp 97 113 Diabetes Mellitus and Breast Cancer Ido Wolf
Masur K, Thévenod F, Zänker KS (eds): Diabetes and Cancer. Epidemiological Evidence and Molecular Links. Front Diabetes. Basel, Karger, 2008, vol 19, pp 97 113 Diabetes Mellitus and Breast Cancer Ido Wolf
Supplemental Data. TGF-β-mediated mir-181a expression promotes breast cancer metastasis by targeting Bim.
 Supplemental Data TGF-β-mediated mir-181a expression promotes breast cancer metastasis by targeting Bim. Molly A. Taylor 1, Khalid Sossey-Alaoui 2, Cheryl L. Thompson 3, David Danielpour 4, and William
Supplemental Data TGF-β-mediated mir-181a expression promotes breast cancer metastasis by targeting Bim. Molly A. Taylor 1, Khalid Sossey-Alaoui 2, Cheryl L. Thompson 3, David Danielpour 4, and William
TITLE: Investigation of the Akt/Pkb Kinase in the Development of Hormone- Independent Prostate Cancer
 AD Award Number: TITLE: Investigation of the Akt/Pkb Kinase in the Development of Hormone- Independent Prostate Cancer PRINCIPAL INVESTIGATOR: Linda A. degraffenried, PhD CONTRACTING ORGANIZATION: University
AD Award Number: TITLE: Investigation of the Akt/Pkb Kinase in the Development of Hormone- Independent Prostate Cancer PRINCIPAL INVESTIGATOR: Linda A. degraffenried, PhD CONTRACTING ORGANIZATION: University
SUPPLEMENTARY INFORMATION
 Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
A novel calcium-dependent mechanism of acquired resistance to IGF-1 receptor inhibition in prostate cancer cells
 /, Vol. 5, No. 19 A novel calcium-dependent mechanism of acquired resistance to IGF-1 receptor inhibition in prostate cancer cells Cale D Fahrenholtz 1, Ann M Greene 1, Pedro J Beltran 2, Kerry L Burnstein
/, Vol. 5, No. 19 A novel calcium-dependent mechanism of acquired resistance to IGF-1 receptor inhibition in prostate cancer cells Cale D Fahrenholtz 1, Ann M Greene 1, Pedro J Beltran 2, Kerry L Burnstein
The PI3K/AKT axis. Dr. Lucio Crinò Medical Oncology Division Azienda Ospedaliera-Perugia. Introduction
 The PI3K/AKT axis Dr. Lucio Crinò Medical Oncology Division Azienda Ospedaliera-Perugia Introduction Phosphoinositide 3-kinase (PI3K) pathway are a family of lipid kinases discovered in 1980s. They have
The PI3K/AKT axis Dr. Lucio Crinò Medical Oncology Division Azienda Ospedaliera-Perugia Introduction Phosphoinositide 3-kinase (PI3K) pathway are a family of lipid kinases discovered in 1980s. They have
A Hepatocyte Growth Factor Receptor (Met) Insulin Receptor hybrid governs hepatic glucose metabolism SUPPLEMENTARY FIGURES, LEGENDS AND METHODS
 Insulin Receptor hybrid governs hepatic glucose metabolism SUPPLEMENTARY FIGURES, LEGENDS AND METHODS") A Hepatocyte Growth Factor Receptor (Met) Insulin Receptor hybrid governs hepatic glucose metabolism Arlee Fafalios, Jihong Ma, Xinping Tan, John Stoops, Jianhua Luo, Marie C. DeFrances and Reza Zarnegar
A Hepatocyte Growth Factor Receptor (Met) Insulin Receptor hybrid governs hepatic glucose metabolism Arlee Fafalios, Jihong Ma, Xinping Tan, John Stoops, Jianhua Luo, Marie C. DeFrances and Reza Zarnegar
Supplementary Information POLO-LIKE KINASE 1 FACILITATES LOSS OF PTEN-INDUCED PROSTATE CANCER FORMATION
 Supplementary Information POLO-LIKE KINASE 1 FACILITATES LOSS OF PTEN-INDUCED PROSTATE CANCER FORMATION X. Shawn Liu 1, 3, Bing Song 2, 3, Bennett D. Elzey 3, 4, Timothy L. Ratliff 3, 4, Stephen F. Konieczny
Supplementary Information POLO-LIKE KINASE 1 FACILITATES LOSS OF PTEN-INDUCED PROSTATE CANCER FORMATION X. Shawn Liu 1, 3, Bing Song 2, 3, Bennett D. Elzey 3, 4, Timothy L. Ratliff 3, 4, Stephen F. Konieczny
hexahistidine tagged GRP78 devoid of the KDEL motif (GRP78-His) on SDS-PAGE. This
 on SDS-PAGE. This") SUPPLEMENTAL FIGURE LEGEND Fig. S1. Generation and characterization of. (A) Coomassie staining of soluble hexahistidine tagged GRP78 devoid of the KDEL motif (GRP78-His) on SDS-PAGE. This protein was expressed
SUPPLEMENTAL FIGURE LEGEND Fig. S1. Generation and characterization of. (A) Coomassie staining of soluble hexahistidine tagged GRP78 devoid of the KDEL motif (GRP78-His) on SDS-PAGE. This protein was expressed
TITLE: Investigation of the Akt/PKB Kinase in the Development of Hormone-Independent Prostate Cancer
 AD Award Number: TITLE: Investigation of the Akt/PKB Kinase in the Development of Hormone-Independent Prostate Cancer PRINCIPAL INVESTIGATOR: Linda A. degraffenried, Ph.D. CONTRACTING ORGANIZATION: The
AD Award Number: TITLE: Investigation of the Akt/PKB Kinase in the Development of Hormone-Independent Prostate Cancer PRINCIPAL INVESTIGATOR: Linda A. degraffenried, Ph.D. CONTRACTING ORGANIZATION: The
Validation & Assay Performance Summary
 Validation & Assay Performance Summary LanthaScreen IGF-1R GripTite Cells Cat. no. K1834 Modification Detected: Phosphorylation of Multiple Tyr Residues on IGF-1R LanthaScreen Cellular Assay Validation
Validation & Assay Performance Summary LanthaScreen IGF-1R GripTite Cells Cat. no. K1834 Modification Detected: Phosphorylation of Multiple Tyr Residues on IGF-1R LanthaScreen Cellular Assay Validation
Phospho-AKT Sampler Kit
 Phospho-AKT Sampler Kit E 0 5 1 0 0 3 Kits Includes Cat. Quantity Application Reactivity Source Akt (Ab-473) Antibody E021054-1 50μg/50μl IHC, WB Human, Mouse, Rat Rabbit Akt (Phospho-Ser473) Antibody
Phospho-AKT Sampler Kit E 0 5 1 0 0 3 Kits Includes Cat. Quantity Application Reactivity Source Akt (Ab-473) Antibody E021054-1 50μg/50μl IHC, WB Human, Mouse, Rat Rabbit Akt (Phospho-Ser473) Antibody
Supplementary data Supplementary Figure 1 Supplementary Figure 2
 Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
PREPARED FOR: U.S. Army Medical Research and Materiel Command Fort Detrick, Maryland
 AD Award Number: W81XWH-14-1-0505 TITLE: Targeting the Neural Microenvironment in Prostate Cancer PRINCIPAL INVESTIGATOR: Michael Ittmann MD PhD CONTRACTING ORGANIZATION: BAYLOR COLLEGE OF MEDICINE HOUSTON,
AD Award Number: W81XWH-14-1-0505 TITLE: Targeting the Neural Microenvironment in Prostate Cancer PRINCIPAL INVESTIGATOR: Michael Ittmann MD PhD CONTRACTING ORGANIZATION: BAYLOR COLLEGE OF MEDICINE HOUSTON,
In Vitro Studies of the Aurora-Kinase Inhibitor MLN8237 in Prostate Cancer Cell Lines
 In Vitro Studies of the Aurora-Kinase Inhibitor MLN837 in Prostate Cancer Cell Lines Nicole V. Julian Mentor: Ana Aparicio, M.D. Special Thanks to Brittany Kleb Department of Genitourinary Medical Oncology
In Vitro Studies of the Aurora-Kinase Inhibitor MLN837 in Prostate Cancer Cell Lines Nicole V. Julian Mentor: Ana Aparicio, M.D. Special Thanks to Brittany Kleb Department of Genitourinary Medical Oncology
HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation
 SUPPLEMENTARY INFORMATION Materials and Methods Human cell lines and culture conditions HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation in exon 20 of BRCA1
SUPPLEMENTARY INFORMATION Materials and Methods Human cell lines and culture conditions HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation in exon 20 of BRCA1
MicroRNA sponges: competitive inhibitors of small RNAs in mammalian cells
 MicroRNA sponges: competitive inhibitors of small RNAs in mammalian cells Margaret S Ebert, Joel R Neilson & Phillip A Sharp Supplementary figures and text: Supplementary Figure 1. Effect of sponges on
MicroRNA sponges: competitive inhibitors of small RNAs in mammalian cells Margaret S Ebert, Joel R Neilson & Phillip A Sharp Supplementary figures and text: Supplementary Figure 1. Effect of sponges on
Receptor mediated Signal Transduction
 Receptor mediated Signal Transduction G-protein-linked receptors adenylyl cyclase camp PKA Organization of receptor protein-tyrosine kinases From G.M. Cooper, The Cell. A molecular approach, 2004, third
Receptor mediated Signal Transduction G-protein-linked receptors adenylyl cyclase camp PKA Organization of receptor protein-tyrosine kinases From G.M. Cooper, The Cell. A molecular approach, 2004, third
injected subcutaneously into flanks of 6-8 week old athymic male nude mice (LNCaP SQ) and body
 and body") SUPPLEMENTAL FIGURE LEGENDS Figure S1: Generation of ENZR Xenografts and Cell Lines: (A) 1x10 6 LNCaP cells in matrigel were injected subcutaneously into flanks of 6-8 week old athymic male nude mice (LNCaP
SUPPLEMENTAL FIGURE LEGENDS Figure S1: Generation of ENZR Xenografts and Cell Lines: (A) 1x10 6 LNCaP cells in matrigel were injected subcutaneously into flanks of 6-8 week old athymic male nude mice (LNCaP
Lecture: CHAPTER 13 Signal Transduction Pathways
 Lecture: 10 17 2016 CHAPTER 13 Signal Transduction Pathways Chapter 13 Outline Signal transduction cascades have many components in common: 1. Release of a primary message as a response to a physiological
Lecture: 10 17 2016 CHAPTER 13 Signal Transduction Pathways Chapter 13 Outline Signal transduction cascades have many components in common: 1. Release of a primary message as a response to a physiological
Figure S1. ERBB3 mrna levels are elevated in Luminal A breast cancers harboring ERBB3
 Supplemental Figure Legends. Figure S1. ERBB3 mrna levels are elevated in Luminal A breast cancers harboring ERBB3 ErbB3 gene copy number gain. Supplemental Figure S1. ERBB3 mrna levels are elevated in
Supplemental Figure Legends. Figure S1. ERBB3 mrna levels are elevated in Luminal A breast cancers harboring ERBB3 ErbB3 gene copy number gain. Supplemental Figure S1. ERBB3 mrna levels are elevated in
AP VP DLP H&E. p-akt DLP
 A B AP VP DLP H&E AP AP VP DLP p-akt wild-type prostate PTEN-null prostate Supplementary Fig. 1. Targeted deletion of PTEN in prostate epithelium resulted in HG-PIN in all three lobes. (A) The anatomy
A B AP VP DLP H&E AP AP VP DLP p-akt wild-type prostate PTEN-null prostate Supplementary Fig. 1. Targeted deletion of PTEN in prostate epithelium resulted in HG-PIN in all three lobes. (A) The anatomy
TFEB-mediated increase in peripheral lysosomes regulates. Store Operated Calcium Entry
 TFEB-mediated increase in peripheral lysosomes regulates Store Operated Calcium Entry Luigi Sbano, Massimo Bonora, Saverio Marchi, Federica Baldassari, Diego L. Medina, Andrea Ballabio, Carlotta Giorgi
TFEB-mediated increase in peripheral lysosomes regulates Store Operated Calcium Entry Luigi Sbano, Massimo Bonora, Saverio Marchi, Federica Baldassari, Diego L. Medina, Andrea Ballabio, Carlotta Giorgi
Supplementary Figure 1. Basal level EGFR across a panel of ESCC lines. Immunoblots demonstrate the expression of phosphorylated and total EGFR as
 Supplementary Figure 1. Basal level EGFR across a panel of ESCC lines. Immunoblots demonstrate the expression of phosphorylated and total EGFR as well as their downstream effectors across a panel of ESCC
Supplementary Figure 1. Basal level EGFR across a panel of ESCC lines. Immunoblots demonstrate the expression of phosphorylated and total EGFR as well as their downstream effectors across a panel of ESCC
G-Protein Signaling. Introduction to intracellular signaling. Dr. SARRAY Sameh, Ph.D
 G-Protein Signaling Introduction to intracellular signaling Dr. SARRAY Sameh, Ph.D Cell signaling Cells communicate via extracellular signaling molecules (Hormones, growth factors and neurotransmitters
G-Protein Signaling Introduction to intracellular signaling Dr. SARRAY Sameh, Ph.D Cell signaling Cells communicate via extracellular signaling molecules (Hormones, growth factors and neurotransmitters
Introduction: 年 Fas signal-mediated apoptosis. PI3K/Akt
 Fas-ligand (CD95-L; Fas-L) Fas (CD95) Fas (apoptosis) 年 了 不 度 Fas Fas-L 力 不 Fas/Fas-L T IL-10Fas/Fas-L 不 年 Fas signal-mediated apoptosis 度降 不 不 力 U-118, HeLa, A549, Huh-7 MCF-7, HepG2. PI3K/Akt FasPI3K/Akt
Fas-ligand (CD95-L; Fas-L) Fas (CD95) Fas (apoptosis) 年 了 不 度 Fas Fas-L 力 不 Fas/Fas-L T IL-10Fas/Fas-L 不 年 Fas signal-mediated apoptosis 度降 不 不 力 U-118, HeLa, A549, Huh-7 MCF-7, HepG2. PI3K/Akt FasPI3K/Akt
Figure S1. Figure S2. Figure S3.
 Figure S1. PSA expression in VCaP was not dependent on the residual androgens in hormonedepleted medium. VCaP or LNCaP cells grown in CSS medium or SFM (serum-free medium) were treated with ethanol (-)
Figure S1. PSA expression in VCaP was not dependent on the residual androgens in hormonedepleted medium. VCaP or LNCaP cells grown in CSS medium or SFM (serum-free medium) were treated with ethanol (-)
Data Sheet TIGIT / NFAT Reporter - Jurkat Cell Line Catalog #60538
 Data Sheet TIGIT / NFAT Reporter - Jurkat Cell Line Catalog #60538 Background: TIGIT is a co-inhibitory receptor that is highly expressed in Natural Killer (NK) cells, activated CD4+, CD8+ and regulatory
Data Sheet TIGIT / NFAT Reporter - Jurkat Cell Line Catalog #60538 Background: TIGIT is a co-inhibitory receptor that is highly expressed in Natural Killer (NK) cells, activated CD4+, CD8+ and regulatory
Supporting Information Table of Contents
 Supporting Information Table of Contents Supporting Information Figure 1 Page 2 Supporting Information Figure 2 Page 4 Supporting Information Figure 3 Page 5 Supporting Information Figure 4 Page 6 Supporting
Supporting Information Table of Contents Supporting Information Figure 1 Page 2 Supporting Information Figure 2 Page 4 Supporting Information Figure 3 Page 5 Supporting Information Figure 4 Page 6 Supporting
Supplementary Table; Supplementary Figures and legends S1-S21; Supplementary Materials and Methods
 Silva et al. PTEN posttranslational inactivation and hyperactivation of the PI3K/Akt pathway sustain primary T cell leukemia viability Supplementary Table; Supplementary Figures and legends S1-S21; Supplementary
Silva et al. PTEN posttranslational inactivation and hyperactivation of the PI3K/Akt pathway sustain primary T cell leukemia viability Supplementary Table; Supplementary Figures and legends S1-S21; Supplementary
Breast Cancer: the interplay of biology, drugs, radiation. Prof. L. Livi Università degli Studi di Firenze. Brescia, October 3rd 4th, 2013
 Breast Cancer: the interplay of biology, drugs, radiation Prof. L. Livi Università degli Studi di Firenze Brescia, October 3rd 4th, 2013 BACKGROUND (1) The complex interactions between tumor-specific signaling
Breast Cancer: the interplay of biology, drugs, radiation Prof. L. Livi Università degli Studi di Firenze Brescia, October 3rd 4th, 2013 BACKGROUND (1) The complex interactions between tumor-specific signaling
EGFR: fundamenteel en klinisch
 EGFR: fundamenteel en klinisch Guido Lammering MAASTRO Clinic Maastricht, NL What is EGFR?? The EGFR some facts 1186 amino acids 170 kda Expressed by all cells of epithelial origin Increased activation
EGFR: fundamenteel en klinisch Guido Lammering MAASTRO Clinic Maastricht, NL What is EGFR?? The EGFR some facts 1186 amino acids 170 kda Expressed by all cells of epithelial origin Increased activation
(A) PCR primers (arrows) designed to distinguish wild type (P1+P2), targeted (P1+P2) and excised (P1+P3)14-
 PCR primers (arrows) designed to distinguish wild type (P1+P2), targeted (P1+P2) and excised (P1+P3)14-") 1 Supplemental Figure Legends Figure S1. Mammary tumors of ErbB2 KI mice with 14-3-3σ ablation have elevated ErbB2 transcript levels and cell proliferation (A) PCR primers (arrows) designed to distinguish
1 Supplemental Figure Legends Figure S1. Mammary tumors of ErbB2 KI mice with 14-3-3σ ablation have elevated ErbB2 transcript levels and cell proliferation (A) PCR primers (arrows) designed to distinguish
ACK1 Tyrosine Kinases: A Critical Regulator of Prostate Cancer
 ACK1 Tyrosine Kinases: A Critical Regulator of Prostate Cancer Nupam Mahajan Moffitt Cancer Center Learners Objectives How Androgen Receptor (AR) signaling is accomplished in absence of androgen What are
ACK1 Tyrosine Kinases: A Critical Regulator of Prostate Cancer Nupam Mahajan Moffitt Cancer Center Learners Objectives How Androgen Receptor (AR) signaling is accomplished in absence of androgen What are
Figure S1. Generation of inducible PTEN deficient mice and the BMMCs (A) B6.129 Pten loxp/loxp mice were mated with B6.
 B6.129 Pten loxp/loxp mice were mated with B6.") Figure S1. Generation of inducible PTEN deficient mice and the BMMCs (A) B6.129 Pten loxp/loxp mice were mated with B6.129-Gt(ROSA)26Sor tm1(cre/ert2)tyj /J mice. To induce deletion of the Pten locus,
Figure S1. Generation of inducible PTEN deficient mice and the BMMCs (A) B6.129 Pten loxp/loxp mice were mated with B6.129-Gt(ROSA)26Sor tm1(cre/ert2)tyj /J mice. To induce deletion of the Pten locus,
STAT3 (py705) (Human/Mouse/Rat) ELISA Kit
 (Human/Mouse/Rat) ELISA Kit") STAT3 (py705) (Human/Mouse/Rat) ELISA Kit Catalog Number KA2175 96 assays Version: 01 Intended for research use only www.abnova.com I. INTRODUCTION STAT3 (py705) (Human/Mouse/Rat) ELISA (Enzyme-Linked
STAT3 (py705) (Human/Mouse/Rat) ELISA Kit Catalog Number KA2175 96 assays Version: 01 Intended for research use only www.abnova.com I. INTRODUCTION STAT3 (py705) (Human/Mouse/Rat) ELISA (Enzyme-Linked
Antitumor Activity of CUDC-305, a Novel Oral HSP90 Inhibitor, in Solid and Hematological Tumor Xenograft Models
 Antitumor Activity of CUDC-5, a Novel Oral HSP Inhibitor, in Solid and Hematological Tumor Xenograft Models Rudi Bao, MD/PhD April 1, 2 AACR 1th Annual Meeting 2 Experimental and Molecular Therapeutics
Antitumor Activity of CUDC-5, a Novel Oral HSP Inhibitor, in Solid and Hematological Tumor Xenograft Models Rudi Bao, MD/PhD April 1, 2 AACR 1th Annual Meeting 2 Experimental and Molecular Therapeutics
SUPPLEMENTAL MATERIAL. Supplementary Methods
 SUPPLEMENTAL MATERIAL Supplementary Methods Culture of cardiomyocytes, fibroblasts and cardiac microvascular endothelial cells The isolation and culturing of neonatal rat ventricular cardiomyocytes was
SUPPLEMENTAL MATERIAL Supplementary Methods Culture of cardiomyocytes, fibroblasts and cardiac microvascular endothelial cells The isolation and culturing of neonatal rat ventricular cardiomyocytes was
Supplementary Figure 1.TRIM33 binds β-catenin in the nucleus. a & b, Co-IP of endogenous TRIM33 with β-catenin in HT-29 cells (a) and HEK 293T cells
 and HEK 293T cells") Supplementary Figure 1.TRIM33 binds β-catenin in the nucleus. a & b, Co-IP of endogenous TRIM33 with β-catenin in HT-29 cells (a) and HEK 293T cells (b). TRIM33 was immunoprecipitated, and the amount of
Supplementary Figure 1.TRIM33 binds β-catenin in the nucleus. a & b, Co-IP of endogenous TRIM33 with β-catenin in HT-29 cells (a) and HEK 293T cells (b). TRIM33 was immunoprecipitated, and the amount of
Award Number: W81XWH TITLE: Dependency on SRC-Family Kinases for Recurrence of Androgen- Independent Prostate Cancer
 AD Award Number: W81XWH-08-1-0026 TITLE: Dependency on SRC-Family Kinases for Recurrence of Androgen- Independent Prostate Cancer PRINCIPAL INVESTIGATOR: Irwin H. Gelman, Ph.D. CONTRACTING ORGANIZATION:
AD Award Number: W81XWH-08-1-0026 TITLE: Dependency on SRC-Family Kinases for Recurrence of Androgen- Independent Prostate Cancer PRINCIPAL INVESTIGATOR: Irwin H. Gelman, Ph.D. CONTRACTING ORGANIZATION:
SUPPLEMENTAL INFORMATION
 SUPPLEMENTAL INFORMATION EXPERIMENTAL PROCEDURES Tryptic digestion protection experiments - PCSK9 with Ab-3D5 (1:1 molar ratio) in 50 mm Tris, ph 8.0, 150 mm NaCl was incubated overnight at 4 o C. The
SUPPLEMENTAL INFORMATION EXPERIMENTAL PROCEDURES Tryptic digestion protection experiments - PCSK9 with Ab-3D5 (1:1 molar ratio) in 50 mm Tris, ph 8.0, 150 mm NaCl was incubated overnight at 4 o C. The
Plasma exposure levels from individual mice 4 hours post IP administration at the
 Supplemental Figure Legends Figure S1. Plasma exposure levels of MKC-3946 in mice. Plasma exposure levels from individual mice 4 hours post IP administration at the indicated dose mg/kg. Data represent
Supplemental Figure Legends Figure S1. Plasma exposure levels of MKC-3946 in mice. Plasma exposure levels from individual mice 4 hours post IP administration at the indicated dose mg/kg. Data represent
Serum Amyloid A3 Gene Expression in Adipocytes is an Indicator. of the Interaction with Macrophages
 Serum Amyloid A3 Gene Expression in Adipocytes is an Indicator of the Interaction with Macrophages Yohei Sanada, Takafumi Yamamoto, Rika Satake, Akiko Yamashita, Sumire Kanai, Norihisa Kato, Fons AJ van
Serum Amyloid A3 Gene Expression in Adipocytes is an Indicator of the Interaction with Macrophages Yohei Sanada, Takafumi Yamamoto, Rika Satake, Akiko Yamashita, Sumire Kanai, Norihisa Kato, Fons AJ van
Part-4. Cell cycle regulatory protein 5 (Cdk5) A novel target of ERK in Carb induced cell death
 A novel target of ERK in Carb induced cell death") Part-4 Cell cycle regulatory protein 5 (Cdk5) A novel target of ERK in Carb induced cell death 95 1. Introduction The process of replicating DNA and dividing cells can be described as a series of coordinated
Part-4 Cell cycle regulatory protein 5 (Cdk5) A novel target of ERK in Carb induced cell death 95 1. Introduction The process of replicating DNA and dividing cells can be described as a series of coordinated
Supplementary Materials for
 www.sciencesignaling.org/cgi/content/full/8/364/ra18/dc1 Supplementary Materials for The tyrosine phosphatase (Pez) inhibits metastasis by altering protein trafficking Leila Belle, Naveid Ali, Ana Lonic,
www.sciencesignaling.org/cgi/content/full/8/364/ra18/dc1 Supplementary Materials for The tyrosine phosphatase (Pez) inhibits metastasis by altering protein trafficking Leila Belle, Naveid Ali, Ana Lonic,
Rationale Design of Combination Therapy in Prostate Cancer: Targeting the AR and PI3K pathways
 Rationale Design of Combination Therapy in Prostate Cancer: Targeting the AR and PI3K pathways Brett S Carver, MD Assistant Attending, Department of Surgery/Urology Prostate Cancer Therapeutics: A Changed
Rationale Design of Combination Therapy in Prostate Cancer: Targeting the AR and PI3K pathways Brett S Carver, MD Assistant Attending, Department of Surgery/Urology Prostate Cancer Therapeutics: A Changed
Xenoestrogen-induced Regulation of EZH2 and Histone Methylation via Non-Genomic Estrogen
 Xenoestrogen-induced Regulation of EZH2 and Histone Methylation via Non-Genomic Estrogen Receptor Signaling to PI3K/AKT Tiffany G. Bredfeldt, Kristen L. Greathouse, Stephen H. Safe, Mien-Chie Hung, Mark
Xenoestrogen-induced Regulation of EZH2 and Histone Methylation via Non-Genomic Estrogen Receptor Signaling to PI3K/AKT Tiffany G. Bredfeldt, Kristen L. Greathouse, Stephen H. Safe, Mien-Chie Hung, Mark
mtor plays critical roles in pancreatic cancer stem cells through specific and stemness-related functions
 Supplementary Information mtor plays critical roles in pancreatic cancer stem cells through specific and stemness-related functions Shyuichiro Matsubara 1, Qiang Ding 1, Yumi Miyazaki 1, Taisaku Kuwahata
Supplementary Information mtor plays critical roles in pancreatic cancer stem cells through specific and stemness-related functions Shyuichiro Matsubara 1, Qiang Ding 1, Yumi Miyazaki 1, Taisaku Kuwahata
General Laboratory methods Plasma analysis: Gene Expression Analysis: Immunoblot analysis: Immunohistochemistry:
 General Laboratory methods Plasma analysis: Plasma insulin (Mercodia, Sweden), leptin (duoset, R&D Systems Europe, Abingdon, United Kingdom), IL-6, TNFα and adiponectin levels (Quantikine kits, R&D Systems
General Laboratory methods Plasma analysis: Plasma insulin (Mercodia, Sweden), leptin (duoset, R&D Systems Europe, Abingdon, United Kingdom), IL-6, TNFα and adiponectin levels (Quantikine kits, R&D Systems
Validation & Assay Performance Summary
 Validation & Assay Performance Summary CellSensor DBE-bla MDA-MB-468 Cell Line Cat. no. K1814 Pathway Description The phosphatidylinositol-3-kinase (PI3K) signaling cascade is essential for cell growth
Validation & Assay Performance Summary CellSensor DBE-bla MDA-MB-468 Cell Line Cat. no. K1814 Pathway Description The phosphatidylinositol-3-kinase (PI3K) signaling cascade is essential for cell growth
Growth and Differentiation Phosphorylation Sampler Kit
 Growth and Differentiation Phosphorylation Sampler Kit E 0 5 1 0 1 4 Kits Includes Cat. Quantity Application Reactivity Source Akt (Phospho-Ser473) E011054-1 50μg/50μl IHC, WB Human, Mouse, Rat Rabbit
Growth and Differentiation Phosphorylation Sampler Kit E 0 5 1 0 1 4 Kits Includes Cat. Quantity Application Reactivity Source Akt (Phospho-Ser473) E011054-1 50μg/50μl IHC, WB Human, Mouse, Rat Rabbit
Supplementary Table 1. Characterization of HNSCC PDX models established at MSKCC
 Supplementary Table 1. Characterization of HNSCC PDX models established at MSKCC Supplementary Table 2. Drug content and loading efficiency estimated with F-NMR and UV- Vis Supplementary Table 3. Complete
Supplementary Table 1. Characterization of HNSCC PDX models established at MSKCC Supplementary Table 2. Drug content and loading efficiency estimated with F-NMR and UV- Vis Supplementary Table 3. Complete
The attached protocol was used for growing RWPE-1 cells per ATCC instructions.
 RWPE-1 (prostate epithelial transformed by HPV) From: Duke/UNC/UT/EBI ENCODE group Date: 7/7/11 ATCC: Catalog #CRL-11609 The attached protocol was used for growing RWPE-1 cells per ATCC instructions. Cell
RWPE-1 (prostate epithelial transformed by HPV) From: Duke/UNC/UT/EBI ENCODE group Date: 7/7/11 ATCC: Catalog #CRL-11609 The attached protocol was used for growing RWPE-1 cells per ATCC instructions. Cell
Cell Signaling part 2
 15 Cell Signaling part 2 Functions of Cell Surface Receptors Other cell surface receptors are directly linked to intracellular enzymes. The largest family of these is the receptor protein tyrosine kinases,
15 Cell Signaling part 2 Functions of Cell Surface Receptors Other cell surface receptors are directly linked to intracellular enzymes. The largest family of these is the receptor protein tyrosine kinases,
Lankenau Institute for Medical Research Annual Progress Report: 2010 Formula Grant
 Lankenau Institute for Medical Research Annual Progress Report: 21 Formula Grant Reporting Period July 1, 211 December 31, 211 Formula Grant Overview The Lankenau Institute for Medical Research received
Lankenau Institute for Medical Research Annual Progress Report: 21 Formula Grant Reporting Period July 1, 211 December 31, 211 Formula Grant Overview The Lankenau Institute for Medical Research received
Supplementary Figure 1 IMQ-Induced Mouse Model of Psoriasis. IMQ cream was
 Supplementary Figure 1 IMQ-Induced Mouse Model of Psoriasis. IMQ cream was painted on the shaved back skin of CBL/J and BALB/c mice for consecutive days. (a, b) Phenotypic presentation of mouse back skin
Supplementary Figure 1 IMQ-Induced Mouse Model of Psoriasis. IMQ cream was painted on the shaved back skin of CBL/J and BALB/c mice for consecutive days. (a, b) Phenotypic presentation of mouse back skin
A549 and A549-fLuc cells were maintained in high glucose Dulbecco modified
 Cell culture and animal model A549 and A549-fLuc cells were maintained in high glucose Dulbecco modified Eagle medium supplemented with 10% fetal bovine serum at 37 C in humidified atmosphere containing
Cell culture and animal model A549 and A549-fLuc cells were maintained in high glucose Dulbecco modified Eagle medium supplemented with 10% fetal bovine serum at 37 C in humidified atmosphere containing
4.2 RESULTS AND DISCUSSION
 phosphorylated proteins on treatment with Erlotinib.This chapter describes the finding of the SILAC experiment. 4.2 RESULTS AND DISCUSSION This is the first global report of its kind using dual strategies
phosphorylated proteins on treatment with Erlotinib.This chapter describes the finding of the SILAC experiment. 4.2 RESULTS AND DISCUSSION This is the first global report of its kind using dual strategies
Phospholipase C γ Prof. Graham Carpenter
 Graham Carpenter, h.d. rofessor of Biochemistry Cornelia Crooke Department of Biochemistry Vanderbilt University School of Medicine, Nashville, TN 1 Receptor Tyrosine Kinases GF Extracellular M Intracellular
Graham Carpenter, h.d. rofessor of Biochemistry Cornelia Crooke Department of Biochemistry Vanderbilt University School of Medicine, Nashville, TN 1 Receptor Tyrosine Kinases GF Extracellular M Intracellular
Supplementary Figure 1
 Supplementary Figure 1 6 HE-50 HE-116 E-1 HE-108 Supplementary Figure 1. Targeted drug response curves of endometrial cancer cells. Endometrial cancer cell lines were incubated with serial dilutions of
Supplementary Figure 1 6 HE-50 HE-116 E-1 HE-108 Supplementary Figure 1. Targeted drug response curves of endometrial cancer cells. Endometrial cancer cell lines were incubated with serial dilutions of
TUMOR M ARKERS MARKERS
 TUMOR MARKERS M.Shekarabi IUMS Definition Many cancers are associated with the abnormal production of some molecules l which h can be measured in plasma. These molecules are known as tumor markers. A good
TUMOR MARKERS M.Shekarabi IUMS Definition Many cancers are associated with the abnormal production of some molecules l which h can be measured in plasma. These molecules are known as tumor markers. A good
William C. Comb, Jessica E. Hutti, Patricia Cogswell, Lewis C. Cantley, and Albert S. Baldwin
 Molecular Cell, Volume 45 Supplemental Information p85 SH2 Domain Phosphorylation by IKK Promotes Feedback Inhibition of PI3K and Akt in Response to Cellular Starvation William C. Comb, Jessica E. Hutti,
Molecular Cell, Volume 45 Supplemental Information p85 SH2 Domain Phosphorylation by IKK Promotes Feedback Inhibition of PI3K and Akt in Response to Cellular Starvation William C. Comb, Jessica E. Hutti,
Kit for assay of thioredoxin
 FkTRX-02-V2 Kit for assay of thioredoxin The thioredoxin system is the major protein disulfide reductase in cells and comprises thioredoxin, thioredoxin reductase and NADPH (1). Thioredoxin systems are
FkTRX-02-V2 Kit for assay of thioredoxin The thioredoxin system is the major protein disulfide reductase in cells and comprises thioredoxin, thioredoxin reductase and NADPH (1). Thioredoxin systems are
MTC-TT and TPC-1 cell lines were cultured in RPMI medium (Gibco, Breda, The Netherlands)
") Supplemental data Materials and Methods Cell culture MTC-TT and TPC-1 cell lines were cultured in RPMI medium (Gibco, Breda, The Netherlands) supplemented with 15% or 10% (for TPC-1) fetal bovine serum
Supplemental data Materials and Methods Cell culture MTC-TT and TPC-1 cell lines were cultured in RPMI medium (Gibco, Breda, The Netherlands) supplemented with 15% or 10% (for TPC-1) fetal bovine serum
Modulating Glucose Uptake in Skeletal Myotubes: Insulin Induction with Bioluminescent Glucose Uptake Analysis
 icell Skeletal Myoblasts Application Protocol Modulating Glucose Uptake in Skeletal Myotubes: Insulin Induction with Bioluminescent Glucose Uptake Analysis Introduction The skeletal muscle is one of the
icell Skeletal Myoblasts Application Protocol Modulating Glucose Uptake in Skeletal Myotubes: Insulin Induction with Bioluminescent Glucose Uptake Analysis Introduction The skeletal muscle is one of the
Revolutionizing the Treatment of Cancer
 Revolutionizing the Treatment of Cancer June 2014 Safe Harbor Statement The statements that follow (including projections and business trends) are forward looking statements. Rexahn's actual results may
Revolutionizing the Treatment of Cancer June 2014 Safe Harbor Statement The statements that follow (including projections and business trends) are forward looking statements. Rexahn's actual results may
Hexagon PSA. Design Verification. Contents
 Design Verification Hexagon PSA Contents 1. Function...2 2. Sensitivity, Dynamic Range and Traceability...2 Description of Control Materials...2 Analytical Sensitivity...2 3. Clinical Evaluation...3 Evaluation
Design Verification Hexagon PSA Contents 1. Function...2 2. Sensitivity, Dynamic Range and Traceability...2 Description of Control Materials...2 Analytical Sensitivity...2 3. Clinical Evaluation...3 Evaluation
SUPPLEMENTARY INFORMATION
 DOI: 10.1038/ncb3311 A B TSC2 -/- MEFs C Rapa Hours WCL 0 6 12 24 36 pakt.s473 AKT ps6k S6K CM IGF-1 Recipient WCL - + - + - + pigf-1r IGF-1R pakt ps6 AKT D 1 st SILAC 2 nd SILAC E GAPDH FGF21 ALKPGVIQILGVK
DOI: 10.1038/ncb3311 A B TSC2 -/- MEFs C Rapa Hours WCL 0 6 12 24 36 pakt.s473 AKT ps6k S6K CM IGF-1 Recipient WCL - + - + - + pigf-1r IGF-1R pakt ps6 AKT D 1 st SILAC 2 nd SILAC E GAPDH FGF21 ALKPGVIQILGVK
Online Data Supplement. Anti-aging Gene Klotho Enhances Glucose-induced Insulin Secretion by Upregulating Plasma Membrane Retention of TRPV2
 Online Data Supplement Anti-aging Gene Klotho Enhances Glucose-induced Insulin Secretion by Upregulating Plasma Membrane Retention of TRPV2 Yi Lin and Zhongjie Sun Department of physiology, college of
Online Data Supplement Anti-aging Gene Klotho Enhances Glucose-induced Insulin Secretion by Upregulating Plasma Membrane Retention of TRPV2 Yi Lin and Zhongjie Sun Department of physiology, college of
Supplementary Fig. 1. GPRC5A post-transcriptionally down-regulates EGFR expression. (a) Plot of the changes in steady state mrna levels versus
 Plot of the changes in steady state mrna levels versus") Supplementary Fig. 1. GPRC5A post-transcriptionally down-regulates EGFR expression. (a) Plot of the changes in steady state mrna levels versus changes in corresponding proteins between wild type and Gprc5a-/-
Supplementary Fig. 1. GPRC5A post-transcriptionally down-regulates EGFR expression. (a) Plot of the changes in steady state mrna levels versus changes in corresponding proteins between wild type and Gprc5a-/-
Low Cell Binding Property of LIPIDURE -COAT
 Technical Note_1 ver.1 Low Cell Binding Property of LIPIDURE -COAT 1. LIPIDURE -COAT MULTI DISH A-6MD (Cat. No. 51011617) 2. Cell; NIH 3T3 (Fibroblast, mouse) 1. 10 %CS-DMEM; DMEM (Dulbecco's Modified
Technical Note_1 ver.1 Low Cell Binding Property of LIPIDURE -COAT 1. LIPIDURE -COAT MULTI DISH A-6MD (Cat. No. 51011617) 2. Cell; NIH 3T3 (Fibroblast, mouse) 1. 10 %CS-DMEM; DMEM (Dulbecco's Modified
SUPPLEMENTARY INFORMATION. Involvement of IL-21 in the epidermal hyperplasia of psoriasis
 SUPPLEMENTARY INFORMATION Involvement of IL-21 in the epidermal hyperplasia of psoriasis Roberta Caruso 1, Elisabetta Botti 2, Massimiliano Sarra 1, Maria Esposito 2, Carmine Stolfi 1, Laura Diluvio 2,
SUPPLEMENTARY INFORMATION Involvement of IL-21 in the epidermal hyperplasia of psoriasis Roberta Caruso 1, Elisabetta Botti 2, Massimiliano Sarra 1, Maria Esposito 2, Carmine Stolfi 1, Laura Diluvio 2,
TITLE: Overcoming Resistance to Inhibitors of the Akt Protein Kinase by Modulation of the Pim Kinase Pathway
 AWARD NUMBER: W81XWH-12-1-0560 TITLE: Overcoming Resistance to Inhibitors of the Akt Protein Kinase by Modulation of the Pim Kinase Pathway PRINCIPAL INVESTIGATOR: Andrew S. Kraft, MD CONTRACTING ORGANIZATION:
AWARD NUMBER: W81XWH-12-1-0560 TITLE: Overcoming Resistance to Inhibitors of the Akt Protein Kinase by Modulation of the Pim Kinase Pathway PRINCIPAL INVESTIGATOR: Andrew S. Kraft, MD CONTRACTING ORGANIZATION:
2.5. AMPK activity
 Supplement Fig. A 3 B phos-ampk 2.5 * Control AICAR AMPK AMPK activity (Absorbance at 45 nm) 2.5.5 Control AICAR Supplement Fig. Effects of AICAR on AMPK activation in macrophages. J774. macrophages were
Supplement Fig. A 3 B phos-ampk 2.5 * Control AICAR AMPK AMPK activity (Absorbance at 45 nm) 2.5.5 Control AICAR Supplement Fig. Effects of AICAR on AMPK activation in macrophages. J774. macrophages were
Chapter 20. Cell - Cell Signaling: Hormones and Receptors. Three general types of extracellular signaling. endocrine signaling. paracrine signaling
 Chapter 20 Cell - Cell Signaling: Hormones and Receptors Three general types of extracellular signaling endocrine signaling paracrine signaling autocrine signaling Endocrine Signaling - signaling molecules
Chapter 20 Cell - Cell Signaling: Hormones and Receptors Three general types of extracellular signaling endocrine signaling paracrine signaling autocrine signaling Endocrine Signaling - signaling molecules
Influenza B Hemagglutinin / HA ELISA Pair Set
 Influenza B Hemagglutinin / HA ELISA Pair Set Catalog Number : SEK11053 To achieve the best assay results, this manual must be read carefully before using this product and the assay is run as summarized
Influenza B Hemagglutinin / HA ELISA Pair Set Catalog Number : SEK11053 To achieve the best assay results, this manual must be read carefully before using this product and the assay is run as summarized
KEY CONCEPT QUESTIONS IN SIGNAL TRANSDUCTION
 Signal Transduction - Part 2 Key Concepts - Receptor tyrosine kinases control cell metabolism and proliferation Growth factor signaling through Ras Mutated cell signaling genes in cancer cells are called
Signal Transduction - Part 2 Key Concepts - Receptor tyrosine kinases control cell metabolism and proliferation Growth factor signaling through Ras Mutated cell signaling genes in cancer cells are called
SUPPLEMENTARY INFORMATION. Supplementary Figures S1-S9. Supplementary Methods
 SUPPLEMENTARY INFORMATION SUMO1 modification of PTEN regulates tumorigenesis by controlling its association with the plasma membrane Jian Huang 1,2#, Jie Yan 1,2#, Jian Zhang 3#, Shiguo Zhu 1, Yanli Wang
SUPPLEMENTARY INFORMATION SUMO1 modification of PTEN regulates tumorigenesis by controlling its association with the plasma membrane Jian Huang 1,2#, Jie Yan 1,2#, Jian Zhang 3#, Shiguo Zhu 1, Yanli Wang
Effects of Second Messengers
 Effects of Second Messengers Inositol trisphosphate Diacylglycerol Opens Calcium Channels Binding to IP 3 -gated Channel Cooperative binding Activates Protein Kinase C is required Phosphorylation of many
Effects of Second Messengers Inositol trisphosphate Diacylglycerol Opens Calcium Channels Binding to IP 3 -gated Channel Cooperative binding Activates Protein Kinase C is required Phosphorylation of many
SOMAPLEX REVERSE PHASE PROTEIN MICROARRAY HUMAN KIDNEY TUMOR & NORMAL TISSUE
 SOMAPLEX REVERSE PHASE PROTEIN MICROARRAY HUMAN KIDNEY TUMOR & NORMAL TISSUE 45 CLINICAL CASES SERIAL DILUTION MULTIPLE PROTEIN CONCENTRATION QUANTITATIVE ASSAY PRODUCT NUMBER: PM1-001-N SOMAPLEX REVERSE
SOMAPLEX REVERSE PHASE PROTEIN MICROARRAY HUMAN KIDNEY TUMOR & NORMAL TISSUE 45 CLINICAL CASES SERIAL DILUTION MULTIPLE PROTEIN CONCENTRATION QUANTITATIVE ASSAY PRODUCT NUMBER: PM1-001-N SOMAPLEX REVERSE
Synthesis of Substituted 2H-Benzo[e]indazole-9-carboxylate as Potent Antihyperglycemic Agent that May Act through IRS-1, Akt and GSK-3β Pathways
![Synthesis of Substituted 2H-Benzo[e]indazole-9-carboxylate as Potent Antihyperglycemic Agent that May Act through IRS-1, Akt and GSK-3β Pathways](/thumbs/96/128474991.jpg "Synthesis of Substituted 2H-Benzo[e]indazole-9-carboxylate as Potent Antihyperglycemic Agent that May Act through IRS-1, Akt and GSK-3β Pathways") Electronic Supplementary Material (ESI) for MedChemComm. This journal is The Royal Society of Chemistry 2016 Supplementary Data Synthesis of Substituted 2H-Benzo[e]indazole-9-carboxylate as Potent Antihyperglycemic
Electronic Supplementary Material (ESI) for MedChemComm. This journal is The Royal Society of Chemistry 2016 Supplementary Data Synthesis of Substituted 2H-Benzo[e]indazole-9-carboxylate as Potent Antihyperglycemic
HIV-1 Virus-like Particle Budding Assay Nathan H Vande Burgt, Luis J Cocka * and Paul Bates
 HIV-1 Virus-like Particle Budding Assay Nathan H Vande Burgt, Luis J Cocka * and Paul Bates Department of Microbiology, Perelman School of Medicine at the University of Pennsylvania, Philadelphia, USA
HIV-1 Virus-like Particle Budding Assay Nathan H Vande Burgt, Luis J Cocka * and Paul Bates Department of Microbiology, Perelman School of Medicine at the University of Pennsylvania, Philadelphia, USA
STAT3 (py705)/ Pan STAT3 (Human/Mouse/Rat) ELISA Kit
/ Pan STAT3 (Human/Mouse/Rat) ELISA Kit") STAT3 (py705)/ Pan STAT3 (Human/Mouse/Rat) ELISA Kit Catalog Number KA2176 96 assays Version: 02 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Principle of the Assay...
STAT3 (py705)/ Pan STAT3 (Human/Mouse/Rat) ELISA Kit Catalog Number KA2176 96 assays Version: 02 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Principle of the Assay...
Multimedia Appendix 6 Educational Materials Table of Contents. Intervention Educational Materials Audio Script (version 1)
") Multimedia Appendix 6 Educational Materials Table of Contents Intervention Educational Materials... 1 Audio Script (version 1)... 1 Text (version 1)... 5 Slides (version 1)... 17 Audio Script (version
Multimedia Appendix 6 Educational Materials Table of Contents Intervention Educational Materials... 1 Audio Script (version 1)... 1 Text (version 1)... 5 Slides (version 1)... 17 Audio Script (version
Supplementary Figure 1. Characterization of NMuMG-ErbB2 and NIC breast cancer cells expressing shrnas targeting LPP. NMuMG-ErbB2 cells (a) and NIC
 and NIC") Supplementary Figure 1. Characterization of NMuMG-ErbB2 and NIC breast cancer cells expressing shrnas targeting LPP. NMuMG-ErbB2 cells (a) and NIC cells (b) were engineered to stably express either a LucA-shRNA
Supplementary Figure 1. Characterization of NMuMG-ErbB2 and NIC breast cancer cells expressing shrnas targeting LPP. NMuMG-ErbB2 cells (a) and NIC cells (b) were engineered to stably express either a LucA-shRNA
STAT1 (ps727) (Human/Mouse) ELISA Kit
 (Human/Mouse) ELISA Kit") STAT1 (ps727) (Human/Mouse) ELISA Kit Catalog Number KA2171 96 assays Version: 01 Intended for research use only www.abnova.com I. INTRODUCTION STAT1 (ps727) (Human/Mouse) ELISA (Enzyme-Linked Immunosorbent
STAT1 (ps727) (Human/Mouse) ELISA Kit Catalog Number KA2171 96 assays Version: 01 Intended for research use only www.abnova.com I. INTRODUCTION STAT1 (ps727) (Human/Mouse) ELISA (Enzyme-Linked Immunosorbent
Supplementary Figure 1: Neuregulin 1 increases the growth of mammary organoids compared to EGF. (a) Mammary epithelial cells were freshly isolated,
 Mammary epithelial cells were freshly isolated,") 1 2 3 4 5 6 7 8 9 10 Supplementary Figure 1: Neuregulin 1 increases the growth of mammary organoids compared to EGF. (a) Mammary epithelial cells were freshly isolated, embedded in matrigel and exposed
1 2 3 4 5 6 7 8 9 10 Supplementary Figure 1: Neuregulin 1 increases the growth of mammary organoids compared to EGF. (a) Mammary epithelial cells were freshly isolated, embedded in matrigel and exposed
CHAPTER VII CONCLUDING REMARKS AND FUTURE DIRECTION. Androgen deprivation therapy is the most used treatment of de novo or recurrent
 CHAPTER VII CONCLUDING REMARKS AND FUTURE DIRECTION Stathmin in Prostate Cancer Development and Progression Androgen deprivation therapy is the most used treatment of de novo or recurrent metastatic PCa.
CHAPTER VII CONCLUDING REMARKS AND FUTURE DIRECTION Stathmin in Prostate Cancer Development and Progression Androgen deprivation therapy is the most used treatment of de novo or recurrent metastatic PCa.
HIV-1 p24 ELISA Pair Set Cat#: orb54951 (ELISA Manual)
") HIV-1 p24 ELISA Pair Set Cat#: orb54951 (ELISA Manual) BACKGROUND Human Immunodeficiency Virus ( HIV ) can be divided into two major types, HIV type 1 (HIV-1) and HIV type 2 (HIV-2). HIV-1 is related to
HIV-1 p24 ELISA Pair Set Cat#: orb54951 (ELISA Manual) BACKGROUND Human Immunodeficiency Virus ( HIV ) can be divided into two major types, HIV type 1 (HIV-1) and HIV type 2 (HIV-2). HIV-1 is related to
(a) Significant biological processes (upper panel) and disease biomarkers (lower panel)
 Significant biological processes (upper panel) and disease biomarkers (lower panel)") Supplementary Figure 1. Functional enrichment analyses of secretomic proteins. (a) Significant biological processes (upper panel) and disease biomarkers (lower panel) 2 involved by hrab37-mediated secretory
Supplementary Figure 1. Functional enrichment analyses of secretomic proteins. (a) Significant biological processes (upper panel) and disease biomarkers (lower panel) 2 involved by hrab37-mediated secretory
RayBio KinaseSTAR TM Akt Activity Assay Kit
 Activity Assay Kit User Manual Version 1.0 March 13, 2015 RayBio KinaseSTAR TM Akt Activity Kit Protocol (Cat#: 68AT-Akt-S40) RayBiotech, Inc. We Provide You With Excellent Support And Service Tel:(Toll
Activity Assay Kit User Manual Version 1.0 March 13, 2015 RayBio KinaseSTAR TM Akt Activity Kit Protocol (Cat#: 68AT-Akt-S40) RayBiotech, Inc. We Provide You With Excellent Support And Service Tel:(Toll
PI3K/mTOR Dual Inhibitor
 PI3K/mTOR Dual Inhibitor LY3023414 Courtney KD, et al 1 Drug Discovery Platform: Cancer Cell Signaling A Double-Blinded, Placebo-Controlled, Randomized Phase II Study of Enzalutamide With or Without the
PI3K/mTOR Dual Inhibitor LY3023414 Courtney KD, et al 1 Drug Discovery Platform: Cancer Cell Signaling A Double-Blinded, Placebo-Controlled, Randomized Phase II Study of Enzalutamide With or Without the
PREPARED FOR: U.S. Army Medical Research and Materiel Command Fort Detrick, Maryland
 Page 1 09/10/2013 AD Award Number: W81XWH-12-1-0453 TITLE: The cytoplasm translocation of the androgen receptor cofactor p44 as a target for prostate cancer treatment PRINCIPAL INVESTIGATOR: Zhengxin Wang
Page 1 09/10/2013 AD Award Number: W81XWH-12-1-0453 TITLE: The cytoplasm translocation of the androgen receptor cofactor p44 as a target for prostate cancer treatment PRINCIPAL INVESTIGATOR: Zhengxin Wang
Essential Medium, containing 10% fetal bovine serum, 100 U/ml penicillin and 100 µg/ml streptomycin. Huvec were cultured in
 Supplemental data Methods Cell culture media formulations A-431 and U-87 MG cells were maintained in Dulbecco s Modified Eagle s Medium. FaDu cells were cultured in Eagle's Minimum Essential Medium, containing
Supplemental data Methods Cell culture media formulations A-431 and U-87 MG cells were maintained in Dulbecco s Modified Eagle s Medium. FaDu cells were cultured in Eagle's Minimum Essential Medium, containing
PSA and the Future. Axel Heidenreich, Department of Urology
 PSA and the Future Axel Heidenreich, Department of Urology PSA and Prostate Cancer EAU Guideline 2011 PSA is a continuous variable PSA value (ng/ml) risk of PCa, % 0 0.5 6.6 0.6 1 10.1 1.1 2 17.0 2.1 3
PSA and the Future Axel Heidenreich, Department of Urology PSA and Prostate Cancer EAU Guideline 2011 PSA is a continuous variable PSA value (ng/ml) risk of PCa, % 0 0.5 6.6 0.6 1 10.1 1.1 2 17.0 2.1 3
Supplementary Figure 1. Normal T lymphocyte populations in Dapk -/- mice. (a) Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk
 Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk") Supplementary Figure 1. Normal T lymphocyte populations in Dapk -/- mice. (a) Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk -/- mice were stained for expression of CD4 and CD8.
Supplementary Figure 1. Normal T lymphocyte populations in Dapk -/- mice. (a) Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk -/- mice were stained for expression of CD4 and CD8.
April 5, :45 AM 1:45 PM MARRIOTT MARQUIS HOTEL & MARINA, MIRAMAR VIGNETTE 1 VIGNETTE 2 VIGNETTE 3* VIGNETTE 4* VIGNETTE 5*
 April 5, 2016 11:45 AM 1:45 PM MARRIOTT MARQUIS HOTEL & MARINA, MIRAMAR CHAIR: DANNY A. MILNER, JR., BRIGHAM & WOMEN S HOSPITAL, BOSTON, MA VIGNETTE 1 VIGNETTE 2 VIGNETTE 3* VIGNETTE 4* VIGNETTE 5* *VIGNETTES
April 5, 2016 11:45 AM 1:45 PM MARRIOTT MARQUIS HOTEL & MARINA, MIRAMAR CHAIR: DANNY A. MILNER, JR., BRIGHAM & WOMEN S HOSPITAL, BOSTON, MA VIGNETTE 1 VIGNETTE 2 VIGNETTE 3* VIGNETTE 4* VIGNETTE 5* *VIGNETTES
BMP6 treatment compensates for the molecular defect and ameliorates hemochromatosis in Hfe knockout mice
 SUPPLEMENTARY MATERIALS BMP6 treatment compensates for the molecular defect and ameliorates hemochromatosis in Hfe knockout mice Elena Corradini, Paul J. Schmidt, Delphine Meynard, Cinzia Garuti, Giuliana
SUPPLEMENTARY MATERIALS BMP6 treatment compensates for the molecular defect and ameliorates hemochromatosis in Hfe knockout mice Elena Corradini, Paul J. Schmidt, Delphine Meynard, Cinzia Garuti, Giuliana
RayBio Human PPAR-alpha Transcription Factor Activity Assay Kit
 RayBio Human PPAR-alpha Transcription Factor Activity Assay Kit Catalog #: TFEH-PPARa User Manual Jan 5, 2018 3607 Parkway Lane, Suite 200 Norcross, GA 30092 Tel: 1-888-494-8555 (Toll Free) or 770-729-2992,
RayBio Human PPAR-alpha Transcription Factor Activity Assay Kit Catalog #: TFEH-PPARa User Manual Jan 5, 2018 3607 Parkway Lane, Suite 200 Norcross, GA 30092 Tel: 1-888-494-8555 (Toll Free) or 770-729-2992,