Rapid changes in polyphosphate content within. acidocalcisomes in response to cell growth, differentiation
|
|
|
- Ruby Jackson
- 6 years ago
- Views:
Transcription
1 JBC Papers in Press. Published on May 22, 2001 as Manuscript M Running Title: Acidocalcisomal polyphosphate in T. cruzi Rapid changes in polyphosphate content within acidocalcisomes in response to cell growth, differentiation and environmental stress in Trypanosoma cruzi* Felix A. Ruiz, Claudia O. Rodrigues, and Roberto Docampo # From the Laboratory of Molecular Parasitology, Department of Pathobiology, University of Illinois at Urbana-Champaign, 2001 South Lincoln Avenue, Urbana, IL 61802, USA *This work was supported by National Institutes of Health Grant AI (to R.D.) #To whom correspondence should be addressed: Laboratory of Molecular Parasitology, Department of Pathobiology, College of Veterinary Medicine, University of Illinois at Urbana-Champaign, 2001 South Lincoln Avenue, Urbana, IL 61802, USA. Tel: ; Fax: (217) ; rodoc@ uiuc.edu. Abbreviations: BCECF, 2,7 -bis(2-carboxyethyl)-5-(and 6)-carboxyfluorescein; fura 2, 1-[2-(5-carboxyoxazol-2-yl)-6-aminobenzofuran-5-oxyl]-2-(2 -amino-5 methylphenoxy]-ethane-n, N, N, N -tetraacetic acid; polyp, polyphosphate; Pi, Copyright 2001 by The American Society for Biochemistry and Molecular Biology, Inc.
2 orthophosphate; PPi, pyrophosphate, DAPI, 4, 6-diamidino-2-phenylindole. Inorganic polyphosphate (polyp) has been identified and measured in different stages of Trypanosoma cruzi. Millimolar levels (in terms of Pi residues) in chains of less than 50 residues long, and micromolar levels in chains of about residues long, were found in different stages of T. cruzi. Analysis of purified T. cruzi acidocalcisomes indicated that polyps were preferentially located in these organelles. This was confirmed by visualization of polyps in the acidocalcisomes using 4, 6-diamidino-2-phenylindole (DAPI). A rapid increase (within 2-4 h) in the levels of short and long-chain polyps was detected during trypomastigote to amastigote differentiation and during the lag phase of growth of epimastigotes (within h). Levels rapidly decreased after the epimastigotes resumed growth. Short- and long-chain polyp levels rapidly decreased upon exposure of epimastigotes to hypo-osmotic or alkaline stresses while levels increased after hyperosmotic stress. Ca2+ release from acidocalcisomes by a combination of ionophores (ionomycin, nigericin) was associated with the hydrolysis of short- and long-chain polyps. In agreement with these results, acidocalcisomes were shown to contain polyphosphate kinase and exopolyphosphatase activities. Together, these results suggest a critical role for these organelles in the adaptation of the parasite to environmental changes. 2
3 There is considerable interest in developing novel chemotherapeutic approaches against Trypanosoma cruzi, the etiologic agent of Chagas disease that remains an important health problem in Mexico, Central and South America (1). Some of these approaches are oriented towards the identification of biochemical pathways that allow survival of the parasite and are absent in the host. An unusual characteristic of T. cruzi in comparison with mammalian cells, is the storage of calcium in acidic organelles which were termed acidocalcisomes (2). Initially identified in intact or permeabilized cells (3), the organelles have been isolated (4, 5) and found to have a high density, a high content of phosphorus, calcium, magnesium, sodium, and zinc (4, 6) and a number of pumps and exchangers in their limiting membrane, among them a Ca 2+ -ATPase, a vacuolar H + -ATPase, and a vacuolar H + -pyrophosphatase (V- H + -PPase) (3-7). Recent studies have shown that the phosphorus in the acidocalcisomes is in the form of pyrophosphate (PPi) (8) and short-chain polyphosphate (polyp) (9). Acidocalcisomes are therefore similar to the volutin granules described in other microorganisms (10-14). Although volutin granules were first described almost one hundred years ago (13), they have not been investigated concerning the presence of proton or calcium pumps in their limiting membrane. This is despite the fact that they were known to be acidic and to contain large amounts of calcium (12). The presence of these organelles in many microorganisms such as bacteria, fungi, algae, and protozoa and their apparent absence in mammalian cells makes them promising targets for chemotherapy. Our previous 31 P-NMR findings of large amounts of inorganic pyrophosphate and 3
4 short-chain polyp in T. cruzi (8, 9) are extended here to report the biochemical identification of the polyps present in extracts of different stages of T. cruzi and isolated acidocalcisomes together with the first report of the presence of long-chain polyp and enzymatic activities involved in its synthesis and degradation, in these organisms. Our results indicate that the concentration of polyps changes drastically during growth and differentiation of these parasites and that polyps are rapidly mobilized under osmotic or alkaline stresses. Also, Ca 2+ release from acidocalcisomes is associated with hydrolysis of polyp. The enzymatic activities required for these rapid changes, polyp kinase and exopolyphosphatase are present in the acidocalcisomes. The rapid mobilization of Ca2+ and polyp from acidocalcisomes suggests a critical role for these organelles in the adaptation of the parasite to environmental changes. EXPERIMENTAL PROCEDURES Culture methods- T. cruzi amastigotes and trypomastigotes (Y strain) were obtained from the culture medium of L6E9 myoblasts as we have described before (15). T. cruzi epimastigotes (Y strain) were grown at 28 oc in liver infusion tryptose (LIT) medium (16) supplemented with 10% newborn calf serum. Protein concentration was determined using the Bio-Rad protein assay. Trypomastigotes were induced to transform into amastigotes axenically as described previously (15). Materials- Leupeptin, trans-epoxysuccinyl-l-leucylamido-(4-guanidino) butane 4
5 (E64), Nα-p-tosyl-L-lysine chloromethyl ketone (TLCK), ATP, ionophores except ionomycin, and reagents for marker enzyme assays, polyphosphates and phosphate glass (also known as sodium insoluble metaphosphate) were purchased from Sigma Chemical Co (St. Louis, MO). Silicon carbide (400 mesh) was bought from Aldrich (Milwaukee, WI). 4-(2- aminoethyl)benzenesulfonyl fluoride (AEBSF) and ionomycin (free acid) were from Calbiochem (San Diego, CA). Pepstatin came from Boehringer Mannheim (Indianapolis, IN). Iodixanol (40% solution [OptiPrep], Nycomed) and Dulbecco s PBS were obtained from Gibco BRL, Life Technology Inc. (Gaithersburg, MD). E. coli strain CA38 ptrcppx1 was kindly provided by Prof. Arthur Kornberg, Stanford University School of Medicine (Stanford, CA). Coomassie Blue protein assay reagent was from Bio-Rad (Hercules, CA). The HiTrapTM desalting columns were from Amersham Pharmacia Biotech AB (Upsala, Sweden). The HisBind column was from Novagen Inc. (Madison, WI). The EnzChek phosphate assay kit, and the tetraacetoxymethyl esters of fura 2 (1-{2-(5-carboxyoxazol-2-yl)-6- aminobenzofuran-5-oxyl]-2-(2 -amino-5 -methylphenoxy)-ethane-n,n,n,n -tetraacetic acid), and BCECF (2,7 -bis-(carboxyethyl)-5(and-6)-carboxyfluorescein), fura 2-AM and BCECF-AM, respectively, were from Molecular Probes, Inc (Eugene, OR). [ 32 P]orthophosphate (8500 Ci/mmol) was obtained from New England Nuclear Life Sciences Product Inc. (Boston, MA). All other reagents were analytical grade. Isolation of acidocalcisomes- Isolation of acidocalcisomes was done exactly as described before (5). Collected gradient fractions were assayed for hexokinase (glycosomal marker), acid phosphatase (lysosomal marker), alanine and aspartate aminotransferases (mitochondrial and cytosolic markers) and vacuolar pyrophosphatase (acidocalcisomal 5
6 marker) as previously described (4, 5). Extraction of long- and short-chain polyp- Cells (1 X x 108) were washed once with Dulbecco s PBS and treated with methods to extract either long-chain, or shortchain polyp. Different samples were used for each method. Long-chain polyp extraction was performed as described by Ault-Riché et al. (17). For short-chain polyp extraction, the cell pellet was resuspended in ice-cold 0.5 M HClO 4 (2 ml/g wet weight of cells). After 30 min incubation on ice, the extracts were centrifuged at 3,000 x g for 5 min. The supernatants were neutralized by the addition of 0.72 M KOH/0.6 M KHCO 3. Precipitated KClO 4 was removed by centrifugation at 3,000 x g for 5 min and the extracted supernatant was used for polyp determination. Purification of recombinant exopolyphophatase (rppx1) from Saccharomyces cerevisiae- E. coli strain CA38 ptrcppx1 is an insertionally inactivated mutant for endogenous polyphosphate kinase and exopolyphophatase (18), containing a plasmid with the His-tagged rppx1 gene from S. cerevisiae (19). This strain was grown to an A 600 of 0.6 in LB medium (1% tryptose, 0.5% yeast extract, 1% NaCl, ph 7.5). Addition of isopropyl β- D-thiogalactopyranoside (IPTG) (0.5 mm final concentration) induced the production of rppx1. After incubation for 6 h, cells were harvested by centrifugation (4,000 x g for 15 min). The cells were resuspended in buffer I (50 mm Tris-HCl, ph 7.5, 0.1 M NaCl and 20 µm phenylmethylsulfonyl fluoride (PMSF) with 2 mg lysozyme/ml) and incubated on ice for 15 min. Then, the cells were sonicated (3 times for 20 sec at 20% intensity in a Branson sonifier, model 102c). The suspension was centrifuged for 30 min at 45,000 x g. The 6
7 supernatant was applied to a HisBind TM column that was equilibrated with buffer I and the column was washed with 3 column volumes of buffer I. Proteins were eluted with sequential additions of buffer I containing 50, 100, 200, 400, 800 mm imidazole and equilibrated with buffer I. Exopolyphosphatase activity was determined as in the analysis of polyp (below), using 0.6 mg/ml of polyp 15. PPX1 was eluted with buffer containing mm imidazole, and the purity of the enzyme, estimated by SDS-PAGE, was over 95%. Analysis of polyp- PolyP levels were determined from the amount of Pi released upon treatment with an excess of rppx1. Aliquots of long- or short-chain polyp extracts (always less than 1.5 nmol) were incubated for 15 min at 37 oc with 60 mm Tris-HCl, ph 7.5, 6.0 mm MgCl 2 and 3,000 to 5,000 Units purified rppx1 in a final volume of 75 µl. One unit corresponds to the release of 1 pmol of Pi per min at 37 ºC. rppx1 has been shown to be a powerful catalyst increasing the hydrolytic activity of a phosphoanhydride bond by fold and is active on both polyp 3 and longer chain polyp, as detected by treatment of radiolabeled polyp (20). Release of Pi was monitored by the method of Lanzetta et al. (21). The intracellular concentrations of polyp in different stages of T. cruzi were calculated from the respective cell volumes reported before (22). These values correspond to 30 µl/10 9 epimastigotes and 12 µl/10 9 amastigotes or trypomastigotes. Electrophoretic analysis of polyp- Urea-polyacrylamide gels were prepared and stained with toluidine blue as previously described (23). Marker polyps were obtained by electrophoresis of phosphate glass in 1.5% agarose gels. Four mm-long gel slices were eluted by centrifugation trough Millipore Ultrafree-MC columns to obtain polyps of 7
8 different sizes. The markers were localized by toluidine blue staining and their size was calculated by calibration with commercial polyps (Sigma). Isolation of [32P]polyP from epimastigotes- Epimastigotes (2 x 107 cells) obtained at the exponential phase of growth were harvested by centrifugation, washed with 116 mm NaCl, 5.4 mm KCl, 0.8 mm Mg SO 4, 5.5 mm glucose, 50 mm Tris-HCl ph 7.4, and resuspended in the same buffer supplemented with [ 32 P]Pi (0.02 mci/ml), and 50 mm NaH 2 PO 4. After incubation for 60 min at 28oC, the parasites were washed twice in the same buffer without [ 32 P]Pi. Total poly P (short- and long-chain) extraction was performed as described by Kumble and Kornberg (23). Fluorescence microscopy- Cells (5 x 107) obtained as described above (5) were washed twice with Dulbecco s PBS. The pellet was resuspended in 2 ml of the same buffer and 45 µl of this suspension was incubated at room temperature with 10 µg 4, 6- diamidino-2-phenylindole (DAPI)/ml. After 10 min, the samples were mounted on a slide and observed with an Olympus model BX-60 epifluorescence microscope. Acidocalcisomal fractions (1 mg protein/ml) were incubated with DAPI as above. Olympus WIG ( nm excitation; >580 nm emission) and Olympus WU ( nm excitation; >420 emission) filters were used for polyp and DNA detection, respectively. The images were recorded with a CCD camera (model CH250; Photometrics Ltd., Tucson, AZ) and IPLab software (Signal Analytics, Vienna, VA) as described previously (24). Cell treatments- Epimastigotes (1 x 10 9 ) were washed once in Dulbecco s PBS, 8
9 resuspended in prewarmed isotonic, hypotonic or hypertonic media, and incubated at 30 o C. At the indicated times, aliquots were withdrawn, quickly transferred to an isotonic buffer pre-equilibrated at 4 o C to stop the reaction, centrifuged, and the polyp content of the pellets was quantified as described above. Isotonic medium (137 mm NaCl, 4 mm KCl, 1.5 mm K 2 HPO 4 and 8.5 mm Na 2 HPO 4 ) was 300 ± 5 mosm, hypotonic medium (half of these salts concentrations) was 150 ± 4 mosm and hypertonic medium (3/2 of these salts concentrations) was 450 ± 6 mosm. All media contained in addition 20 mm Hepes, ph 7.4, 11 mm glucose, 1 mm CaCl2 and 0.8 mm MgSO4. Osmolarity of all solutions were checked with an Osmette S automatic osmometer (Precision Systems Inc; Sudbury, MA). For the alkaline or ionophore treatments epimastigotes or amastigotes (2.5 x 10 8 ) were washed once with Dulbecco s PBS and resuspended in 0.55 ml of 116 mm NaCl, 5.4 mm KCl, 0.8 mm Mg SO4, 5.5 mm glucose, 1 mm EGTA, and 50 mm Hepes ph 7.4 (buffer A). At the times indicated, 40 mm NH4Cl, 1 µm ionomycin or 1 µm nigericin were added. Aliquots of 50 µl were withdrawn at the times indicated and mixed with 500 µl of guanidine isothiocyanate (GITC) lysis buffer (17) for long-chain polyp determination or with 300 µl of ice-cold 0.5 M HClO4 for short-chain polyp determination as described above. Spectrofluorometric determinations of amastigotes loaded with fura 2/AM or BCECF/AM were performed as described previously (3). Data depicted in graphs are of single representative experiments with data points given as means ± SEM. Acidocalcisomal synthesis and degradation of polyp- To investigate polyp synthesis 9
10 by isolated acidocalcisomes, the isolated fraction (100 µg protein) (5) was incubated for 5 min at 37 o C in buffer A containing 0.1 mm PPi. ATP (1 mm) was then added and the preparation incubated for different times at 37 o C, after which 500 µl of GITC lysis buffer (for long-chain polyp determination) or 300 µl of ice-cold 0.5 M HClO4 (for short-chain polyp determination) were added and polyp extracted and quantified as described above. To investigate polyp hydrolysis acidocalcisomes were suspended in a buffer containing 250 mm sucrose, 2 mm MgCl 2 and 50 mm Tris-HCl at the phs indicated under Results. The suspension was divided in two samples and 1 mg/ml digitonin was added to one of them, while the other was kept as control. The suspension was then vortexed for about 1 minute and kept on ice for 5 min to measure tetrapolyphosphatase activity in the presence of 0.1 mm tetrapolyphosphate using a phosphate release assay kit (5). After the digitonin treatment aliquots were centrifuged at 10,000 x g for 5 min, to isolate the soluble content. The supernatant was collected and submitted to a second centrifugation step under the same conditions. Final supernatants and pellets were used for measurement of tetrapolyphosphatase activity at different ph and CaCl 2 concentrations. RESULTS PolyP abundance in different stages of T. cruzi- Long- and short-chain polyp were present in the different stages of T. cruzi; values for short-chain polyp were in the mm range and considerably higher in epimastigotes, which have also mm amounts of long- 10
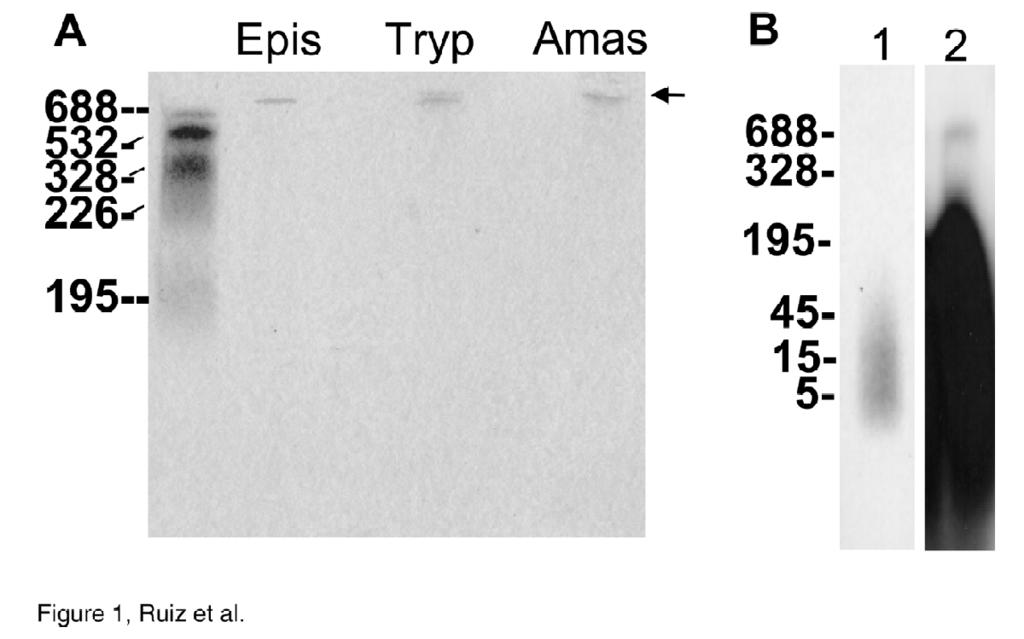
11 chain polyp. Levels of 3.1± 1.4, 25.5 ± 5.1, and 54.3 ± 0.3 mm (in terms of Pi residues and calculated taking into account the cell volumes indicated under Experimental Procedures) in chains of less than 50 residues long, and levels of 82.5 ± 5.75, 130 ± 15, and 2889 ± µm in chains of about residues long, were found in trypomastigotes, amastigotes and epimastigotes, respectively. PolyPs extracted from different stages of T. cruzi were electrophoresed by 6% urea- PAGE to determine their size distribution (Fig. 1A). Only one size class of polyp was detected in the three developmental stages: long-chain polyp of about residues. The lack of detection of other polyps suggests that the short-chain polyps present in the different stages are too small to be recognized by toluidine blue (probably less than 5 residues) (25). In order to investigate the presence of short-chain polyps, we labeled epimastigotes with [ 32 P]Pi and the polyps were extracted and electrophoresed using 6% urea-page. The results are shown in Fig. 1B. Under these conditions labeled compounds that co-eluted with unlabeled commercial standards of polyp of about 5-15 Pi residues were obtained. This is consistent with our previous 31 P NMR work (8, 9), in which high amounts of PPi, tri-, tetra-, and penta-phosphate were detected in epimastigotes and amastigotes of T. cruzi. Addition of a larger amount of material also permitted visualization of long-chain polyp (around residues) (Fig. 1B, lane 2). Accumulation of polyp in acidocalcisomes- The subcellular localization of the large amounts of polyp present in the parasites was investigated using two different methods. First, by subcellular fractionation, and second, by cytochemical techniques. Subcellular fractionation of epimastigotes of T. cruzi revealed that more than 95% of the short- and 11
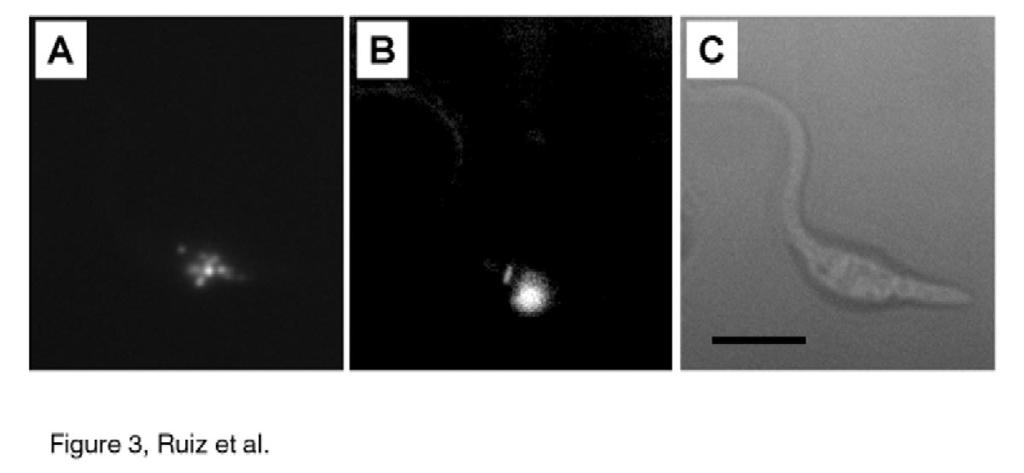
12 long-chain polyps were present in membrane fractions (10,000 x g pellet, Table 1). To investigate whether polyps were present in acidocalcisomes, we isolated these organelles from epimastigotes using an iodixanol (Optiprep) density gradient (5). Short- and longchain polyps were concentrated towards the bottom (dense end) of the gradient (fractions 23-24), with a smaller peak close to the upper part of the gradient (fractions 5-9) (Fig. 2). Markers for other compartments all peaked further up the gradient in the region of fractions 5-9. As previously described, this middle peak also contains acidocalcisomes within ghosts of cells (26). Since the densest fractions (fractions 23 and 24) from the iodixanol gradients contained significant amounts (25% and 33%, respectively) of the total short- and long-chain polyps recovered, which correlated well with the distribution of proton-translocating pyrophoshatase activity, an acidocalcisomal marker (4), the results suggest a preferential acidocalcisomal location of these compounds. The location of polyp in T. cruzi was also investigated using 4,6-diamidino-2- phenylindole (DAPI). DAPI is a useful tool in the fluorometric analysis of DNA but can also be used to study polyps (27, 28). DAPI has a fluorescence emission maximum at 456 nm. PolyP shifts DAPI fluorescence to a higher wavelength with a maximum at about 525 nm (27). This DAPI fluorescence change is specific for polyp and is not produced by PPi or other anions (results not shown and 28). Epimastigotes of T. cruzi incubated in solutions of DAPI (10 µg/ml) were mounted on slides and examined by fluorescence microscopy. When a blue filter was used for DNA staining, the nuclei and kinetoplast were clearly visible (Fig. 3B). In contrast, when a red filter was used for polyp, staining in small spherical bodies corresponding to acidocalcisomes (3-6) was detected (Fig. 3A). No 12
13 staining was detected when DAPI was omitted and similar results were obtained using confocal microscopy (data not shown). To further confirm the acidocalcisomal localization of polyps, acidocalcisomal fractions were incubated with DAPI, mounted on slides and examined by fluorescence microscopy. The isolated acidocalcisomes appeared in clusters and stained with DAPI when a red filter was used (Fig. 4). No fluorescence was detected when DAPI was omitted from the incubation medium or a blue filter was used (data not shown). Changes in polyp levels during cell growth and differentiation- When T. cruzi epimastigotes were passaged into LIT medium, there was an initial lag period of h before growth commenced. During this lag phase a rapid and massive accumulation of short- and long-chain polyp occurred (Fig. 5). Maximal accumulation of short-chain polyp was at about 12 h after inoculation and was followed by a rapid decrease at about 24 h. Maximal accumulation of long-chain polyp was at about 24 h and then rapidly decreased to steady state levels at about 72 h. Levels of both short- and long-chain polyp remained stable during the rest of the logarithmic and stationary phases of growth. These results suggest synthesis of short-chain polyp followed by their decrease simultaneously with the synthesis of long-chain polyp and finally hydrolysis of long-chain polyp once growth was resumed. Since quantitative analysis of polyp indicated a larger amount of short- and longchain polyp in amastigotes than in trypomastigotes, it was of interest to study how rapidly these changes occurred during differentiation. The transformation of T. cruzi trypomastigotes was induced by acidic medium (ph 5.0) and polyp content was 13
14 determined at different times. Short- (Fig. 6A) and long-chain (Fig. 6B) polyp synthesis was induced after only a two-hour incubation in the acidic medium (ph 5.0) and progressed with time. After overnight incubation, amastigote extracts contained amounts equivalent to those of amastigotes obtained from tissue cultures (Figs. 6A and 6B). Trypomastigotes in neutral ph (ph 7.5) medium did not show any significant increase in the amount of either short- or long-chain polyp at any time point (from time zero to overnight incubation) (Figs. 6A and 6B). A parallel transformation into amastigotes with an increase in the number of DAPI-positive vacuoles was also detected (data not shown). These results show that polyp synthesis occurs progressively during the development of amastigotes. Changes in polyp levels under stress conditions- When submitted to hypo-osmotic stress, levels of short- (Fig. 7A) and long-chain (Fig. 7B) polyp decreased within 5-10 min. Long-chain polyp decreased to negligible levels after 20 min incubation under hypoosmotic conditions (Fig. 7B). On the other hand, when epimastigotes were submitted to an hyperosmotic stress, short- (Fig. 7A) and long-chain (Fig. 7B) polyp levels increased within 5-10 min and remained stable until about 20 min. Similarly, epimastigotes submitted to an alkaline stress (incubation with 40 mm NH 4 Cl) showed a progressive decrease in the levels of both short- (Fig. 8A) and long-chain (Fig. 8B) polyp. Association of Ca2+ release and polyp hydrolysis- Hypo-osmotic and alkaline stresses have been shown before to result in increases in the intracellular Ca2+ concentration ([Ca 2+ ] i ) of different trypanosomatids (29-31), including T. cruzi (3). We 14
15 therefore investigated whether there was a correlation between Ca 2+ release from the acidic compartment containing most polyp (acidocalcisome) and polyp hydrolysis. In previous work (3) we showed that addition of nigericin (a H + /K + exchanger), to epimastigotes previously exposed to ionomycin (a Ca2+ ionophore) caused a secondary rise in [Ca 2+ ] i and that similar results were obtained when the order of additions was reversed. This indicated the existence of a Ca 2+ pool, in epimastigotes, that needed ph gradient neutralization for ionomycin-induced Ca 2+ transport to be effective. This is because ionomycin does not bind calcium below ph 7.0 (32) and cannot mobilize Ca 2+ out of acidic compartments. Using a similar protocol we observed that addition of ionomycin to epimastigotes previously exposed to nigericin resulted in significant decreases in short- (Fig. 8E) and long-chain (Fig. 8F) polyp and that similar results could be observed when the order of additions was reversed except that the nigericin effect was slower than the ionomycin effect (Figs. 8C and 8D). Taken together, the results in Fig. 8 suggest that processes that lead to alkalinization of the acidocalcisomes (NH 4 Cl addition or treatment with ionophores) and result in [Ca 2+ ] i increase also result in polyp hydrolysis. Simultaneous measurement of changes in phi, [Ca2+] i, and short- and long-chain polyps in amastigotes are shown in Fig. 9. Addition of ionomycin induced acidification of the cells and Ca 2+ release from intracellular compartments (EGTA was present in the extracellular medium to avoid Ca 2+ entry). This acidification was accompanied by immediate 15
16 hydrolysis of long chain polyp followed later by hydrolysis of short-chain polyp. Subsequent addition of nigericin resulted in a further acidification and Ca 2+ release accompanied by immediate hydrolysis of long-chain polyp and delayed hydrolysis of short-chain polyp. Synthesis and degradation of polyp in acidocalcisomes- The rapid synthesis and hydrolysis of polyp that occurs during growth, differentiation and environmental stress of T. cruzi suggested the presence of enzyme activities for polyp synthesis and degradation in the acidocalcisomes. In agreement with this suggestion, addition of ATP produced a significant increase in short- (SC) and long-chain (LC) polyps in isolated acidocalcisomes within a few minutes (Fig. 10). This increase was time-dependent for ate least 8 (LC) or 10 (SC) min (Figs. 10C and D) and depended on the previous acidification of the acidocalcisome produced by pre-incubation with PPi (Figs. 10A, and B) suggesting a relationship between acidocalcisomal ph and polyp synthesis. This association has also been demonstrated in a S. cerevisiae mutant defective in vacuolar H+-ATPase that fails to accumulate polyp in the vacuole (33). Evidence for a polyp hydrolyzing activity in the acidocalcisomes was also obtained. Fig. 11A shows that a significant increase in tetrapolyp hydrolysis was detected upon treatment of acidocalcisomes with 1 mg/ml digitonin. Supernatants isolated from acidocalcisomes treated with 1 mg/ml digitonin were shown to hydrolyze tetrapolyp at a rate of 0.38 ± µmol/min x mg of protein at ph 7.5 (n = 22). This activity was lower at acidic ph (Fig. 11B) and was inhibited by high Ca2+ concentrations (Fig. 11C), conditions that are prevalent within intact acidocalcisomes (4). 16
17 Interestingly a significant tetrapolyphosphatase activity was retained in the acidocalcisomal pellet after digitonin treatment. The rate of tetrapolyp hydrolysis in the acidocalcisomal pellet was 0.76 ± 0.07 µmol/min x mg of protein (n = 9) at ph 7.5. DISCUSSION In this study we have identified and measured the polyp content of different stages of T. cruzi. The results indicate the presence of high levels of short-chain polyp and lower levels of long-chain polyp. Analysis of purified T. cruzi acidocalcisomes, indicated that polyp was preferentially located in these organelles. This was confirmed by visualization of polyp in the acidocalcisomes using DAPI. The storage of phosphate as polyp appears ideal to reduce the osmotic effect of large pools of this crucial nutrient element. On the other hand its has the potential disadvantage that the cells need to be able to mobilize it under conditions of phosphate starvation or other forms of stress. Based on the total concentration of polyp in different stages of T. cruzi and the relative volume of acidocalcisomes in these cells (0.86%, 2.3%, and 0.26% of the total cell volume of epimastigotes, amastigotes and trypomastigotes, respectively (34)) and assuming that these compounds are essentially concentrated in acidocalcisomes, the calculated concentration in the organelles is in the molar range. Although polyps could attain molar concentration in the acidic (ph 4-5) aqueous environment expected in the acidocalcisome, addition of divalent cations such as calcium or magnesium, present at stochiometric concentrations in the organelles (6), is expected to lead to almost quantitative precipitation of the resulting complexes. We thus 17
18 conclude that polyp in acidocalcisomes is most likely present as a microcrystalline aggregate. This conclusion is consistent with the very high electron density of acidocalcisomes in situ (6) and is further supported by the results of ˆmagic angle sample spinning (MASS) 31 P-NMR experiments of intact parasites and acidocalcisomes (Moreno, Ruiz, Rodrigues, Bailey, Moreno, Urbina, Oldfield and Docampo, unpublished results). A high surface to volume ratio of the microcrystallites may be required for rapid metabolic turnover of polyps accumulated in acidocalcisomes; other components of these organelles, such as carbohydrate (6) or lipids could be involved in maintaining this physical configuration. A rapid increase (within 2-4 h) in the levels of short- and long-chain polyps was detected during trypomastigote to amastigote differentiation and during the lag phase of growth of epimastigotes (within h). Levels rapidly decreased after the epimastigotes resumed growth. These changes are different to those observed in bacteria and in yeast (35, 36). In bacteria, massive accumulations of polyp take place during the exponential phase of growth (35) and it has been proposed that polyp supports survival of stationary-phase E. coli (35). In S. cerevisiae it has been shown that, in glucose medium the mass and total cellular polyp content increased in parallel until glucose was depleted (11 h of culture growth) (36). After glucose depletion, the content of polyps in the cells fell sharply and then increased again in a 24-h culture. The significant decline in the content of intracellular polyps, while Pi was present in the growth medium at high concentrations, was suggested to imply that in this growth phase, polyp are an energy rather that phosphate source (36). Similarly, the changes observed in the content of polyp in T. cruzi epimastigotes and 18
19 amastigotes, which occur before cell division starts, could imply some requirement of these compounds as an energy source for resuming growth. It is important to note that the tissue culture-derived amastigotes assayed were either amastigotes released into the medium after 7 days of culture or after differentiation from trypomastigotes (Fig. 6). In both cases these amastigotes are in a lag phase of growth since they either did not start to divide (when differentiating from trypomastigotes) or they had already finished their intracellular division cycle and because of the fragility of the tissue culture cells were released without differentiating into trypomastigotes. The presence of polyps in various microorganisms is well established and the hypothetical roles of these molecules have been reviewed (10, 12, 36). The localization of these molecules within the cation-rich acidocalcisomes implies that their functional roles could be: (1) energy stores, and/or (2) chelators of metal ions. Short-chain polyps such as PPi could be used in place of ATP as an energy donor in several reactions in trypanosomatids, such as the glycosomal pyruvate, phosphate dikinase (37) and the vacuolar-type proton-translocating pyrophosphatase that has been shown to drive proton uptake into the acidocalcisomal compartment through cleavage of cytosolic pyrophosphate (4). As PPi is a charged and polar molecule, any movement of PPi through a cell membrane is likely to involve a specialized channel or transporter. In this regard, a transmembrane transporter that shuttles PPi between intracellular and extracellular compartments has recently been identified in many mammalian tissues (38). A similar channel in the acidocalcisomal membrane would explain PPi accumulation after its synthesis through anabolic reactions occurring in the cytosol or its release to the cytosol to serve as substrate 19
20 for the V-H + -PPase. On the basis of the fast metabolic turnover of ATP (39) it has been suggested (40) that even highly elevated levels of long-chain polyp, that when expressed in phosphoanhydride bonds might be five times or more the level of ATP in some microorganisms, could supply energy for only a second or two. It has therefore been suggested that a regulatory role for long-chain polyp needs to be considered (40). Long chain polyp, even at relatively low levels, has been shown to be essential for adaptation to various stresses and for survival of bacteria in stationary phase (17, 35, 40). Similar studies have been reported in eukaryotic cells such as yeast (41, 42), fungi (43), and algae (44-46). In the yeast S. cerevisiae and in the alga Dunaliella salina ammonium ions induce hydrolysis of long-chain polyp and the appearance of tripolyp (41, 45). We have reported (47) that influx of ammonia into epimastigotes induces a rapid alkalinization of the cytoplasm followed by recovery of the cytoplasmic ph. This recovery occurs in parallel with massive hydrolysis of polyp (Figs. 8A, and B). In this regard, it has been indicated that H + generation from polyp hydrolysis can neutralize up to a 2.5 ph unit change in S. cerevisiae (42). Two main classes of polyphosphatases have been described. Exopolyphosphatases have been found in prokaryotes and eukaryotes, and remove orthophosphate from the end of the polyphosphate chain. Although in bacteria these enzymes hydrolyse mostly highmolecular weight polyphosphates (48), at least some of the enzymes from yeast are more active hydrolyzing short-chain polyphosphates, such as tripolyphosphate (48). Endopolyphosphatases that act on long-chain polyp, generating tripolyphosphate, have 20
21 also been detected in eukaryotes, including the protist Giardia lamblia (49). Interestingly, the yeast endopolyphosphatase is localized in vacuoles (23). Enzymes that hydrolyze tripolyphosphate have also been reported from different organisms (50). Our results would be consistent with the presence of an exopolyphosphatase in acidocalcisomes that catalyzes the hydrolysis of long- and short-chain polyps to Pi. This exopolyphosphatase is probably tightly associated to the acidocalcisomal membrane as a high activity is still retained in the membrane fraction after detergent treatment of the organelles. It is intriguing that a gene with homology to exopolyphosphatases has recently been found in L. major (51), providing the first evidence for a breakdown pathway for these molecules in trypanosomatids. Our results would also suggest the presence of polyphosphate kinase in acidocalcisomes. Organelle acidification and addition of ATP were necessary to detect polyp synthesis. This would suggest that either the acidocalcisomal membrane is permeable to ATP or the enzyme is located in the acidocalcisomal membrane with its catalytic site oriented towards the cytosolic side of the organelle, and requires an intraorganellar acidic ph for activity. Short- and long-chain polyp levels also rapidly decreased upon exposure of epimastigotes to hypo-osmotic stress while levels increased after hyper-osmotic stress (Fig. 7A and B). This would suggest a role for Pi in the adaptation of the parasites to osmotic stress. This is extremely important for a parasite that lives in environments of widely different osmotic conditions such as the intestine of the insect vector, the bloodstream and the cytosol of host cells. A role for acidocalcisomes in the response of L. major promastigotes to osmotic stress has recently been proposed on the basis of their changes in Na and Cl content after hypo-osmotic stress (52). 21
22 Ca 2+ release from acidocalcisomes by a combination of ionophores (ionomycin, nigericin) was associated with short- and long-chain polyp hydrolysis. Ionomycin is not effective in releasing Ca 2+ from acidic compartments (32). However, acidification of the cytosol (Fig. 9A) could provide some driving force for Ca2+ release through a Ca2+/H+ exchanger, the presence of which has been demonstrated in trypanosomatid acidocalcisomes (53). Further addition of nigericin leads to alkalinization of the acidocalcisomes by K + /H + exchange and further acidification of the cytosol, which would favor further Ca 2+ release (Fig. 9A). Release of Ca 2+ and alkalinization of the acidocalcisomes would result in activation of the polyp hydrolyzing activities in the organelles. It is also currently hypothesized that one of the main roles of the acidocalcisome in T. cruzi is calcium storage for use in intracellular signaling, particularly in the infective stages (2). Enzymes cleaving short- and long-chain polyps to orthophosphate in acidocalcisomes may therefore indirectly regulate intracellular calcium content. In this regard an endogenous Ca2+-inhibited pyrophosphatase activity was postulated to be involved in PPi hydrolysis in the volutin granules of Tetrahymena pyriformis (54). In conclusion, our results indicate that the concentrations of polyp change drastically during growth and differentiation of T. cruzi and that they are rapidly mobilized under osmotic or alkaline stresses. Ca 2+ release from acidocalcisomes is associated with the acidocalcisomal hydrolysis of polyp. The rapid mobilization of Ca 2+ and polyp from 22
23 acidocalcisomes suggests a critical role for these organelles in the adaptation of the parasite to environmental changes. Acknowledgments- We thank Narayana N. Rao and Arthur Kornberg for the gift of E. coli strain CA38 ptrcppx1, David A. Scott for useful comments, and Linda Brown for technical assistance. REFERENCES 1. World Health Organization (1997) In Thirteenth Programme Report, UNDP/World Bank/World Health Organization Special Programme for Research and Training in Tropical Diseases. World Health Organization, Geneva. 2. Docampo, R. and Moreno, S. N. J. (1999) Parasitology Today 15, Docampo, R., Scott, D.A., Vercesi, A.E. and Moreno, S.N.J. (1995) Biochem. J. 310, Scott, D. A., de Souza, W., Benchimol, M., Zhong, L., Lu, H.-G., Moreno, S. N. J. and Docampo, R. (1998) J. Biol. Chem. 273, Scott, D.A. and Docampo, R. (2000) J. Biol. Chem. 275, Scott, D. A., Docampo, R., Dvorak, J. A., Shi, S. and Leapman, R. D. (1997) J. Biol. Chem. 272, Lu, H.-G., Zhong, L., de Souza, W., Benchimol, M., Moreno, S.N.J. and Docampo, R. (1998) Mol. Cell. Biol. 18, Urbina, J. A., Moreno, B., Vierkotter, S., Oldfield, E., Payares, G., Sanoja, C., Bailey, B. N., Yan, 23
24 W., Scott, D. A., Moreno S. N. J. and Docampo, R. (1999) J. Biol Chem. 274, Moreno, B., Urbina, J.A., Oldfield, E., Bailey, B.N., Rodrigues, C.O. and Docampo, R. (2000) J. Biol. Chem. 275, Harold, F.M. (1966) Bacteriol. Rev. 30, Jacobson, L., Halmann, M. and Yariv, J. (1982) Biochem. J. 201, Kornberg, A. (1995) J. Bacteriol. 177, Meyer, A. (1904) Bot. Zeir. 62, Wilkinson, J.F. and Duguid, J.P. (1960) Int. Rev Cytol. 9, Furuya, T., Kashuba, C., Docampo, R., and Moreno, S.N.J. (2000) J. Biol. Chem. 275, Bone, G.J. and Steinert, M. (1956) Nature 178, Ault-Riché, D., Fraley, C.D., Tzeng, C.-M. and Kornberg, A. (1998) J. Bacteriol. 180, Crooke, E., Akiyama, M., Rao, N.N. and Kornberg, A. (1994) J. Biol. Chem. 269, Wurst, H., Shiba, T. and Kornberg, A. (1995) J. Bacteriol Wurst, H. and Kornberg, A. (1994) J. Biol. Chem. 269, Lanzetta, P.A., Alvarez, L.J., Reinach, P.S. and Candia, O.A. (1979) Anal. Biochem. 100, Rangel-Aldao, R., Allende, O., Triana, F., Piras, R., Henriquez, D. and Piras, M. (1987) Mol. Biochem. Parasitol. 22, Kumble, K.D. and Kornberg, A. (1995) J. Biol. Chem. 270,
25 24. Lu, H.-G., Zhong, L., Chang, K.-P. and Docampo, R. (1997) J. Biol. Chem., 272, Lorenz, B., Münkner, J., Oliveira, M.P., Leitão, J.M., Müller, W.E.G. and Schröeder, H.C. (1997) Anal. Biochem. 246, Rodrigues, C., Scott, D.A. and Docampo, R. (1999) Mol. Cell. Biol. 19, Allan, R.A. and Miller, J.J. (1980) Can. J. Microbiol. 26, Tijssen, J.P.F., Beekes, H.W. and Van Steveninck, J. (1982) Biochim. Biophys. Acta 721, Rodrigues, C. O., Scott, D. A. and Docampo, R. (1999) Biochem J. 340, Vercesi, A. E., Moreno, S. and Docampo, R. (1994) Biochem. J. 304, Vieira, L.L., Lafuente, E., Blum, J. and Cabantchik, Z.I. (1997) Mol. Biochem. Parasitol. 90, Liu, C. and Hermannn, T.E. (1978) J. Biol. Chem. 253, Ogawa, N., DeRisi, J., and Brown, P.O. (2000) Mol. Biol. Cell 11, Miranda, K., Benchimol, M., Docampo, R. and de Souza, W. (2000) Parasitol. Res. 86, Rao, N.N., Liu, S. and Kornberg, A. (1998) J. Bacteriol. 180, Kulaev, I.S., and Vagabov, V.M. and Kulakovskaya, T. (1999) J. Biosci. Bioengineer. 88, Bringaud, F., Baltz, D. and Baltz, T. (1998) Proc. Natl. Acad. Sci. USA 95, Ho, A.M., Johnson, M.D. and Kingsley, D.M. (2000) Science 289, Chapman, A.G. and Atkinson, D.E. (1977) Adv. Microbiol. Physiol. 15, Rao, N.N. & Kornberg, A. (1996) J. Bacteriol. 178, Castro, C.D., Koretsky, A.P. and Domach, M.M. (1999) Biotechnol. Prog. 15,
26 42. Castro, C.D., Meehan, A.J., Koretsky, A.P. and Domach, M.M. (1995) Appl. Environm. Microbiol. 61, Yang, Y.C., Bastos, M. and Chen, K.Y. (1993) Biochim. Biophys. Acta 1179, Pick, U. and Weiss, M. (1991) Plant Physiol. 97, Weiss, M., Bental, M. and Pick, U. (1991) Plant Physiol. 97, Pick, U., Zeelon, O. and Weiss, M. (1991) Plant Physiol. 97, VanderHeyden, N., Benaim, G. and Docampo, R. (1996) Biochem. J. 318, Kulaev, I.S., Kulakovskaya, T.C., Andreeva, N.A. and Lichko, L.P. (1999) Prog. Mol. Subcell. Biol. 23, Kumble, K. and Kornberg, A. (1996) J. Biol. Chem. 271, Kornberg, A. (1956) J. Biol. Chem. 218, Myler, P., Audleman, L., DeVos, T., Hixson, G., Kiser, P., Lemley, C., Magness, C., Rickel, E., Sisk, E., Sunkin, S., Swatzell, S., Westlake, T., Bastien, P., Fu, G., Ivens, A. and Stuart, K. (1999) Proc. Natl. Acad. Sci. USA 96, LeFurgey, A., Ingram, P., and Blum, J.J. (2001) Comp. Biochem. Physiol. Part A 128, Vercesi, A.E. and Docampo, R. (1996) Biochem. J. 315, Rosenberg, H. and Munk, N. (1969) Biochim. Biophys. Acta 184,
27 Table I : Distribution of polyp in different fractions Fraction SC PolyP LC PolyP Total protein Purification (µmoles) (µmoles) (mg) -fold (SC/LC).. Lysate ± ± ± 7.8 1/1 10,000g supernatant 0.54 ± ± ± /0.1 10,000g pellet 9.93 ± ± ± /3.93 Isolated Acidocalcisomes 1.69 ± ± ± /114 SC/LC, short-chain and long-chain polyp 27
28 Figure legends: FIG. 1. Urea-PAGE analysis of polyp from different stages of T. cruzi. PolyP extracted from epimastigotes (Epis), trypomastigotes (Tryp), and amastigotes (Amas) was electrophoresed by 6% urea-page. Chain lengths of standards are on the left. The position of migration of long-chain polyp is indicated with an arrow. B, Incorporation of [ 32 P] Pi into short- and long-chain polyp in epimastigotes. Cells (1 x 10 9 epimastigotes) were incubated for 60 min in medium supplemented with [ 32 P]Pi. The polyps were extracted and analyzed by Urea- PAGE. Lane 1 is with 0.2% and lane 2 with 8% of the incorporated [ 32 P]Pi. FIG. 2. Distribution of short- (SC) and long-chain (LC) polyps from epimastigotes on iodixanol gradients. PolyPs are concentrated in a distinct dense fraction. PolyP content was compared with the distribution of established organelle markers, hexokinase (glycosome), acid phosphatase (lysosome), alanine (ALT) and aspartate aminotransferases (AST) (mithochondria and cytosol) and vacuolar pyrophosphatase (acidocalcisome). FIG. 3. Accumulation of DAPI in acidocalcisomes of epimastigotes of T. cruzi. Cells were treated with DAPI for 10 min and observed with a red filter for polyp detection (A) or a blue filter (B) for nucleic acid detection. C, is a bright field micrograph of the same cell. Note the numerous fluorescent vacuoles (acidocalcisomes) in A, and the nucleus and kinetoplast in B. Bar, 10 µm. 28
29 FIG. 4. Accumulation of DAPI in isolated acidocalcisomes. Acidocalcisomes were isolated and treated with DAPI as described under Experimental Procedures. A, acidocalcisomes observed with a red filter for polyp detection. B, bright field micrograph of the same fraction. Bar, 10 µm. FIG. 5. PolyP accumulation during growth of epimastigotes. Short- (open squares) and long-chain (closed squares) polyps accumulate during the lag phase of growth (closed triangles). FIG. 6. PolyP increase during trypomastigote to amastigote transformation. T. cruzi trypomastigotes were incubated at ph 5.0 (closed bars) or ph 7.5 (open bars) for 0, 2, 4, and 12 h and short-chain (A) and long-chain (B) polyp were quantified as described under Experimental Procedures. The average ± S.D. from 3 experiments are shown. The cellular content of short- and long-chain polyp did not vary significantly after 2, 4, and 12 h at ph 7.5, but increased significantly after 2, 4, and 12 h at ph 5.0 with P < 0.05 as determined by the Student s t test. FIG. 7. Effect of osmotic shock on the polyp content of epimastigotes. Epimastigotes were resuspended in isotonic (300 ± 5 mosm, closed diamonds), hypo-osmotic (150 ± 4 mosm, closed squares) or hyperosmotic (450 ± 6 mosm, open squares) medium, as described under Experimental Procedures. At the indicated times, aliquots were withdrawn, quickly transferred to an isotonic buffer pre-equilibrated at 4 o C, to stop the reaction, centrifuged, 29
30 and the polyp content of the pellets was quantified as described under Experimental Procedures. A, shows changes in short-chain polyp and B, changes in long-chain polyp. FIG. 8. The effect of alkaline stress or ionophores on the polyp content of epimastigotes. Epimastigotes (2.5 x 108) were washed once with Dulbecco s PBS and resuspended in 0.55 ml 116 mm NaCl, 5.4 mm KCl, 0.8 mm Mg SO4, 5.5 mm glucose, 1 mm EGTA, 50 mm Tris-HCl ph 7.4. At the times indicated 40 mm NH4Cl, 1 µm ionomycin or 1 µm nigericin were added and shortchain (A, C, E) or long-chain (B, D, F) polyp content was determined as indicated under Experimental Procedures. Dashed lines indicate second addition. Data are of single representative experiments with data points given as means ± SEM. FIG. 9. The effect of ionophores on the intracellular calcium concentration, intracellular ph and polyp content of T. cruzi amastigotes. Amastigotes were loaded with fura 2/AM or BCECF/AM and suspended in 116 mm NaCl, 5.4 mm KCl, 0.8 mm Mg SO 4, 5.5 mm glucose, 1mM EGTA and 50 mm Tris ph 7.4. At the times indicated 1 µm ionomycin or 1 µm nigericin were added. Intracellular calcium (broad line) and ph (narrow line) changes were determined as indicated under Experimental Procedures (A). In parallel experiments short- (B) and long-chain (C) polyp content of the samples was examined as described under Experimental Procedures. Closed squares are controls with no additions. Open squares are after addition of ionomycin and closed circles are after addition of nigericin. Data depicted in panels B and C are of single representative experiments with data points given as means ± SEM. 30
31 FIG. 10. PolyP synthesis in acidocalcisomes: In A and B isolated acidocalcisomes were resuspended in buffer A and incubated for 5 min with or without 0.1 mm PPi. Then, 1 mm ATP (lane 3 and 7) or buffer A (lane 4 and 8) was added. After 5 min at 37 oc, long-chain (LC, A) and short-chain (SC, B) polyp were extracted and quantified as described under Experimental Procedures. C and D show the time-dependent accumulation of long-chain (C) and short-chain (D) polyp in isolated acidocalcisomes after addition of ATP to the samples preincubated with 0.1 mm PPi for 5 min. Data are from a single representative experiments with data points given as mean ± SEM. FIG. 11. Tetrapolyphosphatase activity released from acidocalcisomes upon treatment with 1 mg/ml digitonin. A, Tetrapolyphosphatase activity of acidocalcisomal pellets treated or untreated with 1 mg/ml digitonin (n = 5). B, Range of phs of tetrapolyphosphatase activity present in the supernatant fraction of digitonin-treated acidocalcisomes (n = 6) C, Dosedependent inhibition of tetrapolyphosphatase activity present in the supernatant fraction of digitonin-treated acidocalcisomes, by CaCl 2 (n = 7). 31
32
33
34

35
36
37
38
39
40
41
42
43 Rapid changes in polyphosphate content within acidocalcisomes in response to cell growth, differentiation and environmental stress in Trypanosoma cruzi Felix A. Ruiz, Claudia O. Rodriguez and Roberto Docampo J. Biol. Chem. published online May 22, 2001 Access the most updated version of this article at doi: /jbc.M Alerts: When this article is cited When a correction for this article is posted Click here to choose from all of JBC's alerts This article cites 0 references, 0 of which can be accessed free at
SUPPLEMENTARY MATERIAL
 SUPPLEMENTARY MATERIAL Purification and biochemical properties of SDS-stable low molecular weight alkaline serine protease from Citrullus Colocynthis Muhammad Bashir Khan, 1,3 Hidayatullah khan, 2 Muhammad
SUPPLEMENTARY MATERIAL Purification and biochemical properties of SDS-stable low molecular weight alkaline serine protease from Citrullus Colocynthis Muhammad Bashir Khan, 1,3 Hidayatullah khan, 2 Muhammad
Supporting Information for:
 Supporting Information for: Methylerythritol Cyclodiphosphate (MEcPP) in Deoxyxylulose Phosphate Pathway: Synthesis from an Epoxide and Mechanisms Youli Xiao, a Rodney L. Nyland II, b Caren L. Freel Meyers
Supporting Information for: Methylerythritol Cyclodiphosphate (MEcPP) in Deoxyxylulose Phosphate Pathway: Synthesis from an Epoxide and Mechanisms Youli Xiao, a Rodney L. Nyland II, b Caren L. Freel Meyers
DIDS INHIBITION OF SARCOPLASMIC RETICULUM ANION EFFLUX AND CALCIUM TRANSPORT
 DIDS INHIBITION OF SARCOPLASMIC RETICULUM ANION EFFLUX AND CALCIUM TRANSPORT Kevin P. Campbell and David H. MacLennan Reprinted from ANNALS OF THE NEW YORK ACADEMY OF SCIENCES Volume 358 Pages 328-331
DIDS INHIBITION OF SARCOPLASMIC RETICULUM ANION EFFLUX AND CALCIUM TRANSPORT Kevin P. Campbell and David H. MacLennan Reprinted from ANNALS OF THE NEW YORK ACADEMY OF SCIENCES Volume 358 Pages 328-331
Acidocalcisomes Are Functionally Linked to the Contractile Vacuole of Dictyostelium discoideum*
 THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 277, No. 10, Issue of March 8, pp. 8146 8153, 2002 2002 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. Acidocalcisomes Are
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 277, No. 10, Issue of March 8, pp. 8146 8153, 2002 2002 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. Acidocalcisomes Are
Protocol for Gene Transfection & Western Blotting
 The schedule and the manual of basic techniques for cell culture Advanced Protocol for Gene Transfection & Western Blotting Schedule Day 1 26/07/2008 Transfection Day 3 28/07/2008 Cell lysis Immunoprecipitation
The schedule and the manual of basic techniques for cell culture Advanced Protocol for Gene Transfection & Western Blotting Schedule Day 1 26/07/2008 Transfection Day 3 28/07/2008 Cell lysis Immunoprecipitation
Mammalian Membrane Protein Extraction Kit
 Mammalian Membrane Protein Extraction Kit Catalog number: AR0155 Boster s Mammalian Membrane Protein Extraction Kit is a simple, rapid and reproducible method to prepare cellular protein fractions highly
Mammalian Membrane Protein Extraction Kit Catalog number: AR0155 Boster s Mammalian Membrane Protein Extraction Kit is a simple, rapid and reproducible method to prepare cellular protein fractions highly
20N W81XWH Award Number: Polyphosphate Affects on Breast Cancer Cell Survival TITLE:
 AD Award Number: W81XWH-04-1-0379 TITLE: Polyphosphate Affects on Breast Cancer Cell Survival PRINCIPAL INVESTIGATOR: Christine L. Haakenson CONTRACTING ORGANIZATION: Georgetown University Medical Center
AD Award Number: W81XWH-04-1-0379 TITLE: Polyphosphate Affects on Breast Cancer Cell Survival PRINCIPAL INVESTIGATOR: Christine L. Haakenson CONTRACTING ORGANIZATION: Georgetown University Medical Center
Supplementary material: Materials and suppliers
 Supplementary material: Materials and suppliers Electrophoresis consumables including tris-glycine, acrylamide, SDS buffer and Coomassie Brilliant Blue G-2 dye (CBB) were purchased from Ameresco (Solon,
Supplementary material: Materials and suppliers Electrophoresis consumables including tris-glycine, acrylamide, SDS buffer and Coomassie Brilliant Blue G-2 dye (CBB) were purchased from Ameresco (Solon,
PhosFree TM Phosphate Assay Biochem Kit
 PhosFree TM Phosphate Assay Biochem Kit (Cat. # BK050) ORDERING INFORMATION To order by phone: (303) - 322-2254 To order by Fax: (303) - 322-2257 To order by e-mail: cservice@cytoskeleton.com Technical
PhosFree TM Phosphate Assay Biochem Kit (Cat. # BK050) ORDERING INFORMATION To order by phone: (303) - 322-2254 To order by Fax: (303) - 322-2257 To order by e-mail: cservice@cytoskeleton.com Technical
Figure S1. PMVs from THP-1 cells expose phosphatidylserine and carry actin. A) Flow
 Flow") SUPPLEMENTARY DATA Supplementary Figure Legends Figure S1. PMVs from THP-1 cells expose phosphatidylserine and carry actin. A) Flow cytometry analysis of PMVs labelled with annexin-v-pe (Guava technologies)
SUPPLEMENTARY DATA Supplementary Figure Legends Figure S1. PMVs from THP-1 cells expose phosphatidylserine and carry actin. A) Flow cytometry analysis of PMVs labelled with annexin-v-pe (Guava technologies)
BabyBio IMAC columns DATA SHEET DS
 BabyBio IMAC columns DATA SHEET DS 45 655 010 BabyBio columns for Immobilized Metal Ion Affinity Chromatography (IMAC) are ready-to-use for quick and easy purification of polyhistidine-tagged (His-tagged)
BabyBio IMAC columns DATA SHEET DS 45 655 010 BabyBio columns for Immobilized Metal Ion Affinity Chromatography (IMAC) are ready-to-use for quick and easy purification of polyhistidine-tagged (His-tagged)
A ph-dependent Charge Reversal Peptide for Cancer Targeting
 Supporting Information A ph-dependent Charge Reversal Peptide for Cancer Targeting Naoko Wakabayashi 1, Yoshiaki Yano 1, Kenichi Kawano 1, and Katsumi Matsuzaki 1 1 Graduate School of Pharmaceutical Sciences,
Supporting Information A ph-dependent Charge Reversal Peptide for Cancer Targeting Naoko Wakabayashi 1, Yoshiaki Yano 1, Kenichi Kawano 1, and Katsumi Matsuzaki 1 1 Graduate School of Pharmaceutical Sciences,
Characterization of the DNA-mediated Oxidation of Dps, a Bacterial Ferritin
 SUPPORTING INFORMATION Characterization of the DNA-mediated Oxidation of Dps, a Bacterial Ferritin Anna R. Arnold, Andy Zhou, and Jacqueline K. Barton Division of Chemistry and Chemical Engineering, California
SUPPORTING INFORMATION Characterization of the DNA-mediated Oxidation of Dps, a Bacterial Ferritin Anna R. Arnold, Andy Zhou, and Jacqueline K. Barton Division of Chemistry and Chemical Engineering, California
The Schedule and the Manual of Basic Techniques for Cell Culture
 The Schedule and the Manual of Basic Techniques for Cell Culture 1 Materials Calcium Phosphate Transfection Kit: Invitrogen Cat.No.K2780-01 Falcon tube (Cat No.35-2054:12 x 75 mm, 5 ml tube) Cell: 293
The Schedule and the Manual of Basic Techniques for Cell Culture 1 Materials Calcium Phosphate Transfection Kit: Invitrogen Cat.No.K2780-01 Falcon tube (Cat No.35-2054:12 x 75 mm, 5 ml tube) Cell: 293
GLYCATION OF PROTEINS IN ESCHERICHIA COLI: EFFECT OF NUTRIENT BROTH INGREDIENTS ON GLYCATION
 Industry GLYCATION OF PROTEINS IN ESCHERICHIA COLI: EFFECT OF NUTRIENT BROTH INGREDIENTS ON GLYCATION R. Dimitrova, R. Mironova, I. Ivanov Institute of Molecular biology, Bulgarian Academy of Sciences
Industry GLYCATION OF PROTEINS IN ESCHERICHIA COLI: EFFECT OF NUTRIENT BROTH INGREDIENTS ON GLYCATION R. Dimitrova, R. Mironova, I. Ivanov Institute of Molecular biology, Bulgarian Academy of Sciences
The Polyphosphate Bodies of Chlamydomonas reinhardtii Possess a Proton-pumping Pyrophosphatase and Are Similar to Acidocalcisomes*
 THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 276, No. 49, Issue of December 7, pp. 46196 46203, 2001 2001 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. The Polyphosphate
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 276, No. 49, Issue of December 7, pp. 46196 46203, 2001 2001 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. The Polyphosphate
Adenosine triphosphate (ATP)
") Adenosine triphosphate (ATP) 1 High energy bonds ATP adenosine triphosphate N NH 2 N -O O P O O P O- O- O O P O- O CH 2 H O H N N adenine phosphoanhydride bonds (~) H OH ribose H OH Phosphoanhydride bonds
Adenosine triphosphate (ATP) 1 High energy bonds ATP adenosine triphosphate N NH 2 N -O O P O O P O- O- O O P O- O CH 2 H O H N N adenine phosphoanhydride bonds (~) H OH ribose H OH Phosphoanhydride bonds
Western Immunoblotting Preparation of Samples:
 Western Immunoblotting Preparation of Samples: Total Protein Extraction from Culture Cells: Take off the medium Wash culture with 1 x PBS 1 ml hot Cell-lysis Solution into T75 flask Scrap out the cells
Western Immunoblotting Preparation of Samples: Total Protein Extraction from Culture Cells: Take off the medium Wash culture with 1 x PBS 1 ml hot Cell-lysis Solution into T75 flask Scrap out the cells
Biol110L-Cell Biology Lab Spring Quarter 2012 Module 1-4 Friday April 13, 2012 (Start promptly; work fast; the protocols take ~4 h)
") Biol110L-Cell Biology Lab Spring Quarter 2012 Module 1-4 Friday April 13, 2012 (Start promptly; work fast; the protocols take ~4 h) A. Microscopic Examination of the Plasma Membrane and Its Properties
Biol110L-Cell Biology Lab Spring Quarter 2012 Module 1-4 Friday April 13, 2012 (Start promptly; work fast; the protocols take ~4 h) A. Microscopic Examination of the Plasma Membrane and Its Properties
LANCE Eu-W1024 ITC Chelate & Europium Standard AD0013 Development grade
 AD0017P-4 (en) 1 LANCE Eu-W1024 ITC Chelate & Europium Standard AD0013 Development grade INTRODUCTION Fluorescent isothiocyanato-activated (ITC-activated) Eu-W1024 chelate is optimized for labelling proteins
AD0017P-4 (en) 1 LANCE Eu-W1024 ITC Chelate & Europium Standard AD0013 Development grade INTRODUCTION Fluorescent isothiocyanato-activated (ITC-activated) Eu-W1024 chelate is optimized for labelling proteins
CELLS and TRANSPORT Student Packet SUMMARY CELL MEMBRANES ARE SELECTIVELY PERMEABLE DUE TO THEIR STRUCTURE Hydrophilic head
 CELLS and TRANSPORT Student Packet SUMMARY CELL MEMBRANES ARE SELECTIVELY PERMEABLE DUE TO THEIR STRUCTURE Hydrophilic head Hydrophobic tail Hydrophobic regions of protein Hydrophilic regions of protein
CELLS and TRANSPORT Student Packet SUMMARY CELL MEMBRANES ARE SELECTIVELY PERMEABLE DUE TO THEIR STRUCTURE Hydrophilic head Hydrophobic tail Hydrophobic regions of protein Hydrophilic regions of protein
Chapter 2 Transport Systems
 Chapter 2 Transport Systems The plasma membrane is a selectively permeable barrier between the cell and the extracellular environment. It permeability properties ensure that essential molecules such as
Chapter 2 Transport Systems The plasma membrane is a selectively permeable barrier between the cell and the extracellular environment. It permeability properties ensure that essential molecules such as
Cell Lysis Buffer. Catalog number: AR0103
 Cell Lysis Buffer Catalog number: AR0103 Boster s Cell Lysis Buffer is a ready-to-use Western blot related reagent solution used for efficient extraction of total soluble protein in nondenatured state
Cell Lysis Buffer Catalog number: AR0103 Boster s Cell Lysis Buffer is a ready-to-use Western blot related reagent solution used for efficient extraction of total soluble protein in nondenatured state
Caution: For Laboratory Use. A product for research purposes only. Eu-W1024 ITC Chelate & Europium Standard. Product Number: AD0013
 TECHNICAL DATA SHEET Lance Caution: For Laboratory Use. A product for research purposes only. Eu-W1024 ITC Chelate & Europium Standard Product Number: AD0013 INTRODUCTION: Fluorescent isothiocyanato-activated
TECHNICAL DATA SHEET Lance Caution: For Laboratory Use. A product for research purposes only. Eu-W1024 ITC Chelate & Europium Standard Product Number: AD0013 INTRODUCTION: Fluorescent isothiocyanato-activated
TECHNICAL BULLETIN. MDR1, human recombinant, expressed in Sf9 cells, membrane preparation, for ATPase. Product Number M9194 Storage Temperature 70 C
 MDR1, human recombinant, expressed in Sf9 cells, membrane preparation, for ATPase Product Number M9194 Storage Temperature 70 C TECHNICAL BULLETIN Product Description Multi-drug resistance (MDR) is a major
MDR1, human recombinant, expressed in Sf9 cells, membrane preparation, for ATPase Product Number M9194 Storage Temperature 70 C TECHNICAL BULLETIN Product Description Multi-drug resistance (MDR) is a major
Instructions for Use. APO-AB Annexin V-Biotin Apoptosis Detection Kit 100 tests
 3URGXFW,QIRUPDWLRQ Sigma TACS Annexin V Apoptosis Detection Kits Instructions for Use APO-AB Annexin V-Biotin Apoptosis Detection Kit 100 tests For Research Use Only. Not for use in diagnostic procedures.
3URGXFW,QIRUPDWLRQ Sigma TACS Annexin V Apoptosis Detection Kits Instructions for Use APO-AB Annexin V-Biotin Apoptosis Detection Kit 100 tests For Research Use Only. Not for use in diagnostic procedures.
MEK1 Assay Kit 1 Catalog # Lot # 16875
 MEK1 Assay Kit 1 Kit Components Assay Dilution Buffer (ADB), Catalog # 20-108. Three vials, each containing 1.0ml of assay dilution buffer (20mM MOPS, ph 7.2, 25mM ß-glycerol phosphate, 5mM EGTA, 1mM sodium
MEK1 Assay Kit 1 Kit Components Assay Dilution Buffer (ADB), Catalog # 20-108. Three vials, each containing 1.0ml of assay dilution buffer (20mM MOPS, ph 7.2, 25mM ß-glycerol phosphate, 5mM EGTA, 1mM sodium
Chromatin IP (Isw2) Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles.
 Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles.") Chromatin IP (Isw2) 7/01 Toshi last update: 06/15 Reagents Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles. 2.5 M glycine. TBS:
Chromatin IP (Isw2) 7/01 Toshi last update: 06/15 Reagents Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles. 2.5 M glycine. TBS:
ab CytoPainter Golgi/ER Staining Kit
 ab139485 CytoPainter Golgi/ER Staining Kit Instructions for Use Designed to detect Golgi bodies and endoplasmic reticulum by microscopy This product is for research use only and is not intended for diagnostic
ab139485 CytoPainter Golgi/ER Staining Kit Instructions for Use Designed to detect Golgi bodies and endoplasmic reticulum by microscopy This product is for research use only and is not intended for diagnostic
Kit for assay of thioredoxin
 FkTRX-02-V2 Kit for assay of thioredoxin The thioredoxin system is the major protein disulfide reductase in cells and comprises thioredoxin, thioredoxin reductase and NADPH (1). Thioredoxin systems are
FkTRX-02-V2 Kit for assay of thioredoxin The thioredoxin system is the major protein disulfide reductase in cells and comprises thioredoxin, thioredoxin reductase and NADPH (1). Thioredoxin systems are
TRANSPORT OF AMINO ACIDS IN INTACT 3T3 AND SV3T3 CELLS. Binding Activity for Leucine in Membrane Preparations of Ehrlich Ascites Tumor Cells
 Journal of Supramolecular Structure 4:441 (401)-447 (407) (1976) TRANSPORT OF AMINO ACIDS IN INTACT 3T3 AND SV3T3 CELLS. Binding Activity for Leucine in Membrane Preparations of Ehrlich Ascites Tumor Cells
Journal of Supramolecular Structure 4:441 (401)-447 (407) (1976) TRANSPORT OF AMINO ACIDS IN INTACT 3T3 AND SV3T3 CELLS. Binding Activity for Leucine in Membrane Preparations of Ehrlich Ascites Tumor Cells
BILAYER CHANNEL RECONSTITUTION
 (1) 1% Agar Salt Bridge 1.0 g Agar 3.75g KCl in 100ml distilled water, store at 4 o C. BILAYER CHANNEL RECONSTITUTION (2) Cs solution: (Cesium Methanesulfonate) 1) 50 mm Cs + solution 0.209 MOPS, 10mM
(1) 1% Agar Salt Bridge 1.0 g Agar 3.75g KCl in 100ml distilled water, store at 4 o C. BILAYER CHANNEL RECONSTITUTION (2) Cs solution: (Cesium Methanesulfonate) 1) 50 mm Cs + solution 0.209 MOPS, 10mM
TFEB-mediated increase in peripheral lysosomes regulates. Store Operated Calcium Entry
 TFEB-mediated increase in peripheral lysosomes regulates Store Operated Calcium Entry Luigi Sbano, Massimo Bonora, Saverio Marchi, Federica Baldassari, Diego L. Medina, Andrea Ballabio, Carlotta Giorgi
TFEB-mediated increase in peripheral lysosomes regulates Store Operated Calcium Entry Luigi Sbano, Massimo Bonora, Saverio Marchi, Federica Baldassari, Diego L. Medina, Andrea Ballabio, Carlotta Giorgi
Caution: For Laboratory Use. A product for research purposes only. Eu-W1284 Iodoacetamido Chelate & Europium Standard. Product Number: AD0014
 TECHNICAL DATA SHEET Lance Caution: For Laboratory Use. A product for research purposes only. Eu-W1284 Iodoacetamido Chelate & Europium Standard Product Number: AD0014 INTRODUCTION: Iodoacetamido-activated
TECHNICAL DATA SHEET Lance Caution: For Laboratory Use. A product for research purposes only. Eu-W1284 Iodoacetamido Chelate & Europium Standard Product Number: AD0014 INTRODUCTION: Iodoacetamido-activated
Supplementary Information
 Supplementary Information Supplementary Figure 1. CD4 + T cell activation and lack of apoptosis after crosslinking with anti-cd3 + anti-cd28 + anti-cd160. (a) Flow cytometry of anti-cd160 (5D.10A11) binding
Supplementary Information Supplementary Figure 1. CD4 + T cell activation and lack of apoptosis after crosslinking with anti-cd3 + anti-cd28 + anti-cd160. (a) Flow cytometry of anti-cd160 (5D.10A11) binding
Total Phosphatidic Acid Assay Kit
 Product Manual Total Phosphatidic Acid Assay Kit Catalog Number MET- 5019 100 assays FOR RESEARCH USE ONLY Not for use in diagnostic procedures Introduction Phosphatidic Acid (PA) is a critical precursor
Product Manual Total Phosphatidic Acid Assay Kit Catalog Number MET- 5019 100 assays FOR RESEARCH USE ONLY Not for use in diagnostic procedures Introduction Phosphatidic Acid (PA) is a critical precursor
Introduction. Biochemistry: It is the chemistry of living things (matters).
.") Introduction Biochemistry: It is the chemistry of living things (matters). Biochemistry provides fundamental understanding of the molecular basis for the function and malfunction of living things. Biochemistry
Introduction Biochemistry: It is the chemistry of living things (matters). Biochemistry provides fundamental understanding of the molecular basis for the function and malfunction of living things. Biochemistry
Supporting Information
 Supporting Information Dauvillée et al. 10.1073/pnas.0907424106 Fig. S1. Iodine screening of the C. cohnii mutant bank. Each single colony was grown on rich-medium agar plates then vaporized with iodine.
Supporting Information Dauvillée et al. 10.1073/pnas.0907424106 Fig. S1. Iodine screening of the C. cohnii mutant bank. Each single colony was grown on rich-medium agar plates then vaporized with iodine.
DELFIA Tb-N1 DTA Chelate & Terbium Standard
 AD0029P-1 (en) 1 DELFIA Tb-N1 DTA Chelate & AD0012 Terbium Standard For Research Use Only INTRODUCTION DELFIA Tb-N1 DTA Chelate is optimized for the terbium labeling of proteins and peptides for use in
AD0029P-1 (en) 1 DELFIA Tb-N1 DTA Chelate & AD0012 Terbium Standard For Research Use Only INTRODUCTION DELFIA Tb-N1 DTA Chelate is optimized for the terbium labeling of proteins and peptides for use in
Application Note. Introduction
 Simultaneously Measuring Oxidation of Exogenous and Endogenous Fatty Acids Using the XF Palmitate-BSA FAO Substrate with the Agilent Seahorse XF Cell Mito Stress Test Application Note Introduction The
Simultaneously Measuring Oxidation of Exogenous and Endogenous Fatty Acids Using the XF Palmitate-BSA FAO Substrate with the Agilent Seahorse XF Cell Mito Stress Test Application Note Introduction The
Mammalian Tissue Protein Extraction Reagent
 Mammalian Tissue Protein Extraction Reagent Catalog number: AR0101 Boster s Mammalian Tissue Protein Extraction Reagent is a ready-to-use Western blot related reagent solution used for efficient extraction
Mammalian Tissue Protein Extraction Reagent Catalog number: AR0101 Boster s Mammalian Tissue Protein Extraction Reagent is a ready-to-use Western blot related reagent solution used for efficient extraction
OF LIGHT CHAINS OF CARDIAC MYOSIN ISOZYMES: ATRIAL AND VENTRICULAR MYOSINS
 CROSS-HYBRIDIZATION OF LIGHT CHAINS OF CARDIAC MYOSIN ISOZYMES: ATRIAL AND VENTRICULAR MYOSINS Gabor HOLLGSI*, Sudhir SRIVASTAVA** and Joan WIKMAN-COFFELT University of California, San Francisco Cardiovascular
CROSS-HYBRIDIZATION OF LIGHT CHAINS OF CARDIAC MYOSIN ISOZYMES: ATRIAL AND VENTRICULAR MYOSINS Gabor HOLLGSI*, Sudhir SRIVASTAVA** and Joan WIKMAN-COFFELT University of California, San Francisco Cardiovascular
PRODUCT: RNAzol BD for Blood May 2014 Catalog No: RB 192 Storage: Store at room temperature
 PRODUCT: RNAzol BD for Blood May 2014 Catalog No: RB 192 Storage: Store at room temperature PRODUCT DESCRIPTION. RNAzol BD is a reagent for isolation of total RNA from whole blood, plasma or serum of human
PRODUCT: RNAzol BD for Blood May 2014 Catalog No: RB 192 Storage: Store at room temperature PRODUCT DESCRIPTION. RNAzol BD is a reagent for isolation of total RNA from whole blood, plasma or serum of human
The rabbit femoral artery was prepared and each arterial ring was permeabilized
 Online Supplement Nakmura et al. cgmp-dependent relaxation of smooth muscle Materials and Methods Measurement of tension The rabbit femoral artery was prepared and each arterial ring was permeabilized
Online Supplement Nakmura et al. cgmp-dependent relaxation of smooth muscle Materials and Methods Measurement of tension The rabbit femoral artery was prepared and each arterial ring was permeabilized
BIL 256 Cell and Molecular Biology Lab Spring, Tissue-Specific Isoenzymes
 BIL 256 Cell and Molecular Biology Lab Spring, 2007 Background Information Tissue-Specific Isoenzymes A. BIOCHEMISTRY The basic pattern of glucose oxidation is outlined in Figure 3-1. Glucose is split
BIL 256 Cell and Molecular Biology Lab Spring, 2007 Background Information Tissue-Specific Isoenzymes A. BIOCHEMISTRY The basic pattern of glucose oxidation is outlined in Figure 3-1. Glucose is split
PRODUCT INFORMATION & MANUAL
 PRODUCT INFORMATION & MANUAL Nuclear Extraction Kit NBP2-29447 Research use only. Not for diagnostic or therapeutic procedures. www.novusbio.com - P: 888.506.6887 - technical@novusbio.com Novus kits are
PRODUCT INFORMATION & MANUAL Nuclear Extraction Kit NBP2-29447 Research use only. Not for diagnostic or therapeutic procedures. www.novusbio.com - P: 888.506.6887 - technical@novusbio.com Novus kits are
FOCUS SubCell. For the Enrichment of Subcellular Fractions. (Cat. # ) think proteins! think G-Biosciences
 think proteins! think G-Biosciences") 169PR 01 G-Biosciences 1-800-628-7730 1-314-991-6034 technical@gbiosciences.com A Geno Technology, Inc. (USA) brand name FOCUS SubCell For the Enrichment of Subcellular Fractions (Cat. # 786 260) think
169PR 01 G-Biosciences 1-800-628-7730 1-314-991-6034 technical@gbiosciences.com A Geno Technology, Inc. (USA) brand name FOCUS SubCell For the Enrichment of Subcellular Fractions (Cat. # 786 260) think
SUPPLEMENTARY INFORMATION
 Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
CELLS. Cells. Basic unit of life (except virus)
") Basic unit of life (except virus) CELLS Prokaryotic, w/o nucleus, bacteria Eukaryotic, w/ nucleus Various cell types specialized for particular function. Differentiation. Over 200 human cell types 56%
Basic unit of life (except virus) CELLS Prokaryotic, w/o nucleus, bacteria Eukaryotic, w/ nucleus Various cell types specialized for particular function. Differentiation. Over 200 human cell types 56%
THE QUANTITATIVE GLUCOSE AND MINERAL NUTRIENT REQUIREMENTS OF MOUSE LS (SUSPENSION) CELLS IN CHEMICALLY DEFINED MEDIUM
 CELLS IN CHEMICALLY DEFINED MEDIUM") J. Cell Sci. 8, 693-700 (1971) Printed in Great Britain THE QUANTITATIVE GLUCOSE AND MINERAL NUTRIENT REQUIREMENTS OF MOUSE LS (SUSPENSION) CELLS IN CHEMICALLY DEFINED MEDIUM J. R. BIRCH* AND S. J. PIRT
J. Cell Sci. 8, 693-700 (1971) Printed in Great Britain THE QUANTITATIVE GLUCOSE AND MINERAL NUTRIENT REQUIREMENTS OF MOUSE LS (SUSPENSION) CELLS IN CHEMICALLY DEFINED MEDIUM J. R. BIRCH* AND S. J. PIRT
Human platelet dense granules contain polyphosphate and are similar to acidocalcisomes of bacteria and unicellular eukaryotes
 JBC Papers in Press. Published on August 11, 2004 as Manuscript M406261200 Running title: Polyphosphate in platelet dense granules Human platelet dense granules contain polyphosphate and are similar to
JBC Papers in Press. Published on August 11, 2004 as Manuscript M406261200 Running title: Polyphosphate in platelet dense granules Human platelet dense granules contain polyphosphate and are similar to
(Anderson, 1946) containing sodium chloride, sodium-potassium phosphate. added to this basic medium in a concentration sufficient for maximum growth.
 containing sodium chloride, sodium-potassium phosphate. added to this basic medium in a concentration sufficient for maximum growth.") THE EFFECTS OF A TRYPTOPHAN-HISTIDINE DEFICIENCY IN A MUTANT OF ESCHERICHIA COLI MARGOT K. SANDS AND RICHARD B. ROBERTS Carnegie Institution of Washington, Department of Terrestrial Magnetism, Washington,
THE EFFECTS OF A TRYPTOPHAN-HISTIDINE DEFICIENCY IN A MUTANT OF ESCHERICHIA COLI MARGOT K. SANDS AND RICHARD B. ROBERTS Carnegie Institution of Washington, Department of Terrestrial Magnetism, Washington,
<Supplemental information>
 The Structural Basis of Endosomal Anchoring of KIF16B Kinesin Nichole R. Blatner, Michael I. Wilson, Cai Lei, Wanjin Hong, Diana Murray, Roger L. Williams, and Wonhwa Cho Protein
The Structural Basis of Endosomal Anchoring of KIF16B Kinesin Nichole R. Blatner, Michael I. Wilson, Cai Lei, Wanjin Hong, Diana Murray, Roger L. Williams, and Wonhwa Cho Protein
BIOL 347L Laboratory Three
 Introduction BIOL 347L Laboratory Three Osmosis in potato and carrot samples Osmosis is the movement of water molecules through a selectively permeable membrane into a region of higher solute concentration,
Introduction BIOL 347L Laboratory Three Osmosis in potato and carrot samples Osmosis is the movement of water molecules through a selectively permeable membrane into a region of higher solute concentration,
SUPPLEMENTARY INFORMATION. Bacterial strains and growth conditions. Streptococcus pneumoniae strain R36A was
 SUPPLEMENTARY INFORMATION Bacterial strains and growth conditions. Streptococcus pneumoniae strain R36A was grown in a casein-based semisynthetic medium (C+Y) supplemented with yeast extract (1 mg/ml of
SUPPLEMENTARY INFORMATION Bacterial strains and growth conditions. Streptococcus pneumoniae strain R36A was grown in a casein-based semisynthetic medium (C+Y) supplemented with yeast extract (1 mg/ml of
UV Tracer TM Maleimide NHS ester
 UV Tracer TM Maleimide HS ester Product o.: 1020 Product ame: UV-Tracer TM Maleimide-HS ester Chemical Structure: Chemical Composition: C 41 H 67 5 18 Molecular Weight: 1014.08 Appearance: Storage: Yellow
UV Tracer TM Maleimide HS ester Product o.: 1020 Product ame: UV-Tracer TM Maleimide-HS ester Chemical Structure: Chemical Composition: C 41 H 67 5 18 Molecular Weight: 1014.08 Appearance: Storage: Yellow
Trident Membrane Protein Extraction Kit
 Cat. No. Size Shelf life GTX16373 5/ 20 tests 12 months at the appropriate storage temperatures (see below) Contents Component Storage Amount for 5 tests Amount for 20 tests Buffer A -20 o C 2.5 ml 10
Cat. No. Size Shelf life GTX16373 5/ 20 tests 12 months at the appropriate storage temperatures (see below) Contents Component Storage Amount for 5 tests Amount for 20 tests Buffer A -20 o C 2.5 ml 10
Copyright is owned by the Author of the thesis. Permission is given for a copy to be downloaded by an individual for the purpose of research and
 Copyright is owned by the Author of the thesis. Permission is given for a copy to be downloaded by an individual for the purpose of research and private study only. The thesis may not be reproduced elsewhere
Copyright is owned by the Author of the thesis. Permission is given for a copy to be downloaded by an individual for the purpose of research and private study only. The thesis may not be reproduced elsewhere
Work-flow: protein sample preparation Precipitation methods Removal of interfering substances Specific examples:
 Dr. Sanjeeva Srivastava IIT Bombay Work-flow: protein sample preparation Precipitation methods Removal of interfering substances Specific examples: Sample preparation for serum proteome analysis Sample
Dr. Sanjeeva Srivastava IIT Bombay Work-flow: protein sample preparation Precipitation methods Removal of interfering substances Specific examples: Sample preparation for serum proteome analysis Sample
ab Histone Deacetylase (HDAC) Activity Assay Kit (Fluorometric)
 Activity Assay Kit (Fluorometric)") ab156064 Histone Deacetylase (HDAC) Activity Assay Kit (Fluorometric) Instructions for Use For the quantitative measurement of Histone Deacetylase activity in cell lysates This product is for research
ab156064 Histone Deacetylase (HDAC) Activity Assay Kit (Fluorometric) Instructions for Use For the quantitative measurement of Histone Deacetylase activity in cell lysates This product is for research
2-Deoxyglucose Assay Kit (Colorimetric)
") 2-Deoxyglucose Assay Kit (Colorimetric) Catalog Number KA3753 100 assays Version: 01 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 General Information...
2-Deoxyglucose Assay Kit (Colorimetric) Catalog Number KA3753 100 assays Version: 01 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 General Information...
Pinpoint Slide RNA Isolation System II Catalog No. R1007
 INSTRUCTION MANUAL Pinpoint Slide RNA Isolation System II Catalog No. R1007 Highlights Allows for the isolation of total RNA from paraffin-embedded tissue sections on glass slides Simple procedure combines
INSTRUCTION MANUAL Pinpoint Slide RNA Isolation System II Catalog No. R1007 Highlights Allows for the isolation of total RNA from paraffin-embedded tissue sections on glass slides Simple procedure combines
Regulation of Lamp2a Levels in the Lysosomal Membrane
 Traffic 2000 1: 570 583 Munksgaard International Publishers Regulation of Lamp2a Levels in the Lysosomal Membrane A.M. Cuervo and J.F. Dice* Department of Physiology, Tufts University School of Medicine,
Traffic 2000 1: 570 583 Munksgaard International Publishers Regulation of Lamp2a Levels in the Lysosomal Membrane A.M. Cuervo and J.F. Dice* Department of Physiology, Tufts University School of Medicine,
BIOL 158: BIOLOGICAL CHEMISTRY II
 BIOL 158: BIOLOGICAL CHEMISTRY II Lecture 1: Membranes Lecturer: Christopher Larbie, PhD Introduction Introduction Cells and Organelles have membranes Membranes contain lipids, proteins and polysaccharides
BIOL 158: BIOLOGICAL CHEMISTRY II Lecture 1: Membranes Lecturer: Christopher Larbie, PhD Introduction Introduction Cells and Organelles have membranes Membranes contain lipids, proteins and polysaccharides
MagCapture Exosome Isolation Kit PS Q&A
 MagCapture Exosome Isolation Kit PS Q&A Specifications and performance P.1 Comparison of the conventional method P.2 Operation methods and composition P.4 Amount of starting sample P.5 Analysis after exosomes
MagCapture Exosome Isolation Kit PS Q&A Specifications and performance P.1 Comparison of the conventional method P.2 Operation methods and composition P.4 Amount of starting sample P.5 Analysis after exosomes
ENHANCEMENT BY F-ACTIN OF MGATP-DEPENDENT DOPAMINE UPTAKE INTO ISOLATED CHROMAFFIN GRANULES
 Vol. 4, No. 1, September 1996 BIOCHEMISTRY and MOLECULAR BIOLOGY INTERNATIONAL Pages 61-66 ENHANCEMENT BY F-ACTIN OF MGATP-DEPENDENT DOPAMINE UPTAKE INTO ISOLATED CHROMAFFIN GRANULES Kyoji Morita ~)*,
Vol. 4, No. 1, September 1996 BIOCHEMISTRY and MOLECULAR BIOLOGY INTERNATIONAL Pages 61-66 ENHANCEMENT BY F-ACTIN OF MGATP-DEPENDENT DOPAMINE UPTAKE INTO ISOLATED CHROMAFFIN GRANULES Kyoji Morita ~)*,
Cholesterol determination using protein-templated fluorescent gold nanocluster probes
 Electronic Supplementary Information for Cholesterol determination using protein-templated fluorescent gold nanocluster probes Xi Chen and Gary A. Baker* Department of Chemistry, University of Missouri-Columbia,
Electronic Supplementary Information for Cholesterol determination using protein-templated fluorescent gold nanocluster probes Xi Chen and Gary A. Baker* Department of Chemistry, University of Missouri-Columbia,
Chromatin Immunoprecipitation (ChIPs) Protocol (Mirmira Lab)
 Protocol (Mirmira Lab)") Chromatin Immunoprecipitation (ChIPs) Protocol (Mirmira Lab) Updated 12/3/02 Reagents: ChIP sonication Buffer (1% Triton X-100, 0.1% Deoxycholate, 50 mm Tris 8.1, 150 mm NaCl, 5 mm EDTA): 10 ml 10 % Triton
Chromatin Immunoprecipitation (ChIPs) Protocol (Mirmira Lab) Updated 12/3/02 Reagents: ChIP sonication Buffer (1% Triton X-100, 0.1% Deoxycholate, 50 mm Tris 8.1, 150 mm NaCl, 5 mm EDTA): 10 ml 10 % Triton
Mechanistic Studies of Pentamidine Analogs on Leishmania donovani Promastigotes
 Mechanistic Studies of Pentamidine Analogs on Leishmania donovani Promastigotes Undergraduate Honors Thesis The Ohio State University, College of Pharmacy Division of Medicinal Chemistry and Pharmacognosy
Mechanistic Studies of Pentamidine Analogs on Leishmania donovani Promastigotes Undergraduate Honors Thesis The Ohio State University, College of Pharmacy Division of Medicinal Chemistry and Pharmacognosy
TECHNICAL BULLETIN. R 2 GlcNAcβ1 4GlcNAcβ1 Asn
 GlycoProfile II Enzymatic In-Solution N-Deglycosylation Kit Product Code PP0201 Storage Temperature 2 8 C TECHNICAL BULLETIN Product Description Glycosylation is one of the most common posttranslational
GlycoProfile II Enzymatic In-Solution N-Deglycosylation Kit Product Code PP0201 Storage Temperature 2 8 C TECHNICAL BULLETIN Product Description Glycosylation is one of the most common posttranslational
DELFIA Tb-DTPA ITC Chelate & Terbium Standard
 AD0035P-2 (en) 1 DELFIA Tb-DTPA ITC Chelate & AD0029 Terbium Standard For Research Use Only INTRODUCTION DELFIA Tb-DTPA ITC Chelate is optimized for the terbium labelling of proteins and peptides for use
AD0035P-2 (en) 1 DELFIA Tb-DTPA ITC Chelate & AD0029 Terbium Standard For Research Use Only INTRODUCTION DELFIA Tb-DTPA ITC Chelate is optimized for the terbium labelling of proteins and peptides for use
Nuclear Extraction Kit
 Nuclear Extraction Kit Catalog Number KA1346 50 assays Version: 07 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Principle of the Assay... 3 General Information... 4
Nuclear Extraction Kit Catalog Number KA1346 50 assays Version: 07 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Principle of the Assay... 3 General Information... 4
Loss of protein association causes cardiolipin degradation in Barth syndrome
 SUPPLEMENTARY INFORMATION Loss of protein association causes cardiolipin degradation in Barth syndrome Yang Xu 1, Colin K.L. Phoon 2, Bob Berno 5, Kenneth D Souza 6, Esthelle Hoedt 4, Guoan Zhang 4, Thomas
SUPPLEMENTARY INFORMATION Loss of protein association causes cardiolipin degradation in Barth syndrome Yang Xu 1, Colin K.L. Phoon 2, Bob Berno 5, Kenneth D Souza 6, Esthelle Hoedt 4, Guoan Zhang 4, Thomas
DELFIA Eu-DTPA ITC Chelate & Europium Standard
 AD0026P-3 (en) 1 DELFIA Eu-DTPA ITC Chelate & AD0021 Europium Standard For Research Use Only INTRODUCTION DELFIA Eu-DTPA ITC Chelate is optimized for the europium labelling of proteins and peptides for
AD0026P-3 (en) 1 DELFIA Eu-DTPA ITC Chelate & AD0021 Europium Standard For Research Use Only INTRODUCTION DELFIA Eu-DTPA ITC Chelate is optimized for the europium labelling of proteins and peptides for
AP Biology Cells: Chapters 4 & 5
 AP Biology Cells: Chapters 4 & 5 Multiple Choice Identify the choice that best completes the statement or answers the question. 1. The was the first unifying principle of biology. a. spontaneous generation
AP Biology Cells: Chapters 4 & 5 Multiple Choice Identify the choice that best completes the statement or answers the question. 1. The was the first unifying principle of biology. a. spontaneous generation
6. C-type cytochrome, soluble and membrane protein
 185 6. C-type cytochrome, soluble and membrane protein analysis of Rhodobacter sp SW2 and Rhodopseudomonas palustris TIE-1 ABSTRACT The ability to grown on Fe(II) is thought to be a primitive metabolism
185 6. C-type cytochrome, soluble and membrane protein analysis of Rhodobacter sp SW2 and Rhodopseudomonas palustris TIE-1 ABSTRACT The ability to grown on Fe(II) is thought to be a primitive metabolism
STORE AT 4 o C Version 3
 Instruction Manual Advanced Protein Assay Reagent (Cat. # ADV01) ORDERING INFORMATION To order by phone: (303) - 322-2254 To order by Fax: (303) - 322-2257 To order by e-mail: cserve@cytoskeleton.com Technical
Instruction Manual Advanced Protein Assay Reagent (Cat. # ADV01) ORDERING INFORMATION To order by phone: (303) - 322-2254 To order by Fax: (303) - 322-2257 To order by e-mail: cserve@cytoskeleton.com Technical
ab Membrane Fractionation Kit Instructions for Use For the rapid and simple separation of membrane, cytosolic and nuclear cellular fractions.
 ab139409 Membrane Fractionation Kit Instructions for Use For the rapid and simple separation of membrane, cytosolic and nuclear cellular fractions. This product is for research use only and is not intended
ab139409 Membrane Fractionation Kit Instructions for Use For the rapid and simple separation of membrane, cytosolic and nuclear cellular fractions. This product is for research use only and is not intended
Supplementary Figure 1. Method development.
 Supplementary Figure 1 Method development. Titration experiments to determine standard antibody:lysate concentration. Lysates (~2 mg of total proteins) were prepared from cells expressing FLAG- tagged
Supplementary Figure 1 Method development. Titration experiments to determine standard antibody:lysate concentration. Lysates (~2 mg of total proteins) were prepared from cells expressing FLAG- tagged
MBB 694:407, 115:511. Please use BLOCK CAPITAL letters like this --- A, B, C, D, E. Not lowercase!
 MBB 694:407, 115:511 First Test Severinov/Deis Tue. Sep. 30, 2003 Name Index number (not SSN) Row Letter Seat Number This exam consists of two parts. Part I is multiple choice. Each of these 25 questions
MBB 694:407, 115:511 First Test Severinov/Deis Tue. Sep. 30, 2003 Name Index number (not SSN) Row Letter Seat Number This exam consists of two parts. Part I is multiple choice. Each of these 25 questions
The following protocol describes the isolation of nuclei from tissue. Item. Catalog No Manufacturer
 SOP: Nuclei isolation from tissue and DNaseI treatment Date modified: 090923 Modified by: P. Sabo. (UW) The following protocol describes the isolation of nuclei from tissue. Ordering Information Item.
SOP: Nuclei isolation from tissue and DNaseI treatment Date modified: 090923 Modified by: P. Sabo. (UW) The following protocol describes the isolation of nuclei from tissue. Ordering Information Item.
CytoPainter Lysosomal Staining Kit - Blue Fluorescence
 ab112135 CytoPainter Lysosomal Staining Kit - Blue Fluorescence Instructions for Use For staining Lysosomes in suspension and adherent cells by using our proprietary blue fluorescence probe This product
ab112135 CytoPainter Lysosomal Staining Kit - Blue Fluorescence Instructions for Use For staining Lysosomes in suspension and adherent cells by using our proprietary blue fluorescence probe This product
Recombinant Trypsin, Animal Origin Free
 Recombinant Trypsin, Animal Origin Free PRODUCT INFORMATION: BioGenomics r-trypsin powder is ready to use, animal origin free optimized for cell culture applications. It is derived by r-dna technology.
Recombinant Trypsin, Animal Origin Free PRODUCT INFORMATION: BioGenomics r-trypsin powder is ready to use, animal origin free optimized for cell culture applications. It is derived by r-dna technology.
Monitoring intracellular activity of Arylsulfatase B on its natural substrates in a functional bioassay using LIF-CZE
 Monitoring intracellular activity of Arylsulfatase B on its natural substrates in a functional bioassay using LIF-CZE Erno Pungor Jr; Charles M. Hague; Ginger Chen; Jeffrey F. Lemontt; William S. Prince
Monitoring intracellular activity of Arylsulfatase B on its natural substrates in a functional bioassay using LIF-CZE Erno Pungor Jr; Charles M. Hague; Ginger Chen; Jeffrey F. Lemontt; William S. Prince
Glucose Uptake Colorimetric Assay Kit
 ab136955 Glucose Uptake Colorimetric Assay Kit Instructions for Use For the sensitive and accurate measurement of Glucose uptake in various samples This product is for research use only and is not intended
ab136955 Glucose Uptake Colorimetric Assay Kit Instructions for Use For the sensitive and accurate measurement of Glucose uptake in various samples This product is for research use only and is not intended
Communication. Identification of Methionine N -Acetyltransferase from Saccharomyces cerevisiae
 Communication THE JOURNAL OP BIOLOGICAL CHEMISTRY Vol. 265, No. 7, Issue of March 5, pp. 3603-3606,lSSO 0 1990 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U. S. A. Identification
Communication THE JOURNAL OP BIOLOGICAL CHEMISTRY Vol. 265, No. 7, Issue of March 5, pp. 3603-3606,lSSO 0 1990 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U. S. A. Identification
Instructions. Fuse-It-mRNA easy. Shipping and Storage. Overview. Kit Contents. Specifications. Note: Important Guidelines
 Membrane fusion is a highly efficient method for transfecting various molecules and particles into mammalian cells, even into sensitive and primary cells. The Fuse-It reagents are cargo-specific liposomal
Membrane fusion is a highly efficient method for transfecting various molecules and particles into mammalian cells, even into sensitive and primary cells. The Fuse-It reagents are cargo-specific liposomal
TRACP & ALP Assay Kit
 Cat. # MK301 For Research Use TRACP & ALP Assay Kit Product Manual Table of Contents I. Description...3 II. III. IV. Introduction...3 Components...4 Materials Required but not Provided...4 V. Storage...4
Cat. # MK301 For Research Use TRACP & ALP Assay Kit Product Manual Table of Contents I. Description...3 II. III. IV. Introduction...3 Components...4 Materials Required but not Provided...4 V. Storage...4
Fluorescence Microscopy
 Fluorescence Microscopy Imaging Organelles Mitochondria Lysosomes Nuclei Endoplasmic Reticulum Plasma Membrane F-Actin AAT Bioquest Introduction: Organelle-Selective Stains Organelles are tiny, specialized
Fluorescence Microscopy Imaging Organelles Mitochondria Lysosomes Nuclei Endoplasmic Reticulum Plasma Membrane F-Actin AAT Bioquest Introduction: Organelle-Selective Stains Organelles are tiny, specialized
Endopolyphosphatases for Long Chain Inorganic Polyphosphate in Yeast and Mammals*
 THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 271, No. 43, Issue of October 25, pp. 27146 27151, 1996 1996 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. Endopolyphosphatases
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 271, No. 43, Issue of October 25, pp. 27146 27151, 1996 1996 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. Endopolyphosphatases
Protocol for purification of recombinant protein from 300 ml yeast culture
 Protocol for purification of recombinant protein from 300 ml yeast culture Equipment and reagents needed: Zirconia beads (0.5 mm diameter from BSP, Germany) Paint Shaker (at 4 C) Tube rotator for 15 ml
Protocol for purification of recombinant protein from 300 ml yeast culture Equipment and reagents needed: Zirconia beads (0.5 mm diameter from BSP, Germany) Paint Shaker (at 4 C) Tube rotator for 15 ml
ab Lysosome/Cytotoxicity Dual Staining Kit
 ab133078 Lysosome/Cytotoxicity Dual Staining Kit Instructions for Use For studying lysosome function at the cellular level. This product is for research use only and is not intended for diagnostic use.
ab133078 Lysosome/Cytotoxicity Dual Staining Kit Instructions for Use For studying lysosome function at the cellular level. This product is for research use only and is not intended for diagnostic use.
Materials and Methods , The two-hybrid principle.
 The enzymatic activity of an unknown protein which cleaves the phosphodiester bond between the tyrosine residue of a viral protein and the 5 terminus of the picornavirus RNA Introduction Every day there
The enzymatic activity of an unknown protein which cleaves the phosphodiester bond between the tyrosine residue of a viral protein and the 5 terminus of the picornavirus RNA Introduction Every day there
Product Contents. 1 Specifications 1 Product Description. 2 Buffer Preparation... 3 Protocol. 3 Ordering Information 4 Related Products..
 INSTRUCTION MANUAL Quick-RNA MidiPrep Catalog No. R1056 Highlights 10 minute method for isolating RNA (up to 1 mg) from a wide range of cell types and tissue samples. Clean-Spin column technology allows
INSTRUCTION MANUAL Quick-RNA MidiPrep Catalog No. R1056 Highlights 10 minute method for isolating RNA (up to 1 mg) from a wide range of cell types and tissue samples. Clean-Spin column technology allows
PREPARATION OF IF- ENRICHED CYTOSKELETAL PROTEINS
 TMM,5-2011 PREPARATION OF IF- ENRICHED CYTOSKELETAL PROTEINS Ice-cold means cooled in ice water. In order to prevent proteolysis, make sure to perform all steps on ice. Pre-cool glass homogenizers, buffers
TMM,5-2011 PREPARATION OF IF- ENRICHED CYTOSKELETAL PROTEINS Ice-cold means cooled in ice water. In order to prevent proteolysis, make sure to perform all steps on ice. Pre-cool glass homogenizers, buffers
SensoLyte 520 HDAC Activity Assay Kit *Fluorimetric*
 SensoLyte 520 HDAC Activity Assay Kit *Fluorimetric* Catalog # 72084 Kit Size 100 Assays (96-well plate) Optimized Performance: This kit is optimized to detect HDAC activity. Enhanced Value: It provides
SensoLyte 520 HDAC Activity Assay Kit *Fluorimetric* Catalog # 72084 Kit Size 100 Assays (96-well plate) Optimized Performance: This kit is optimized to detect HDAC activity. Enhanced Value: It provides
User s Manual and Instructions
 User s Manual and Instructions Mitochondria Activity Assay (Cytochrome C Oxidase Activity Assay) Kit Catalog Number: KC310100 Introduction Mitochondria are the eukaryotic subcellular organelles that contain
User s Manual and Instructions Mitochondria Activity Assay (Cytochrome C Oxidase Activity Assay) Kit Catalog Number: KC310100 Introduction Mitochondria are the eukaryotic subcellular organelles that contain
Nature Protocols: doi: /nprot Supplementary Figure 1. Fluorescent titration of probe CPDSA.
 Supplementary Figure 1 Fluorescent titration of probe CPDSA. Fluorescent titration of probe CPDSA (10 um) upon addition of GSH in HEPES (10 mm, ph = 7.4) containing 10% DMSO. Each spectrum was recorded
Supplementary Figure 1 Fluorescent titration of probe CPDSA. Fluorescent titration of probe CPDSA (10 um) upon addition of GSH in HEPES (10 mm, ph = 7.4) containing 10% DMSO. Each spectrum was recorded
Plasma Membrane Protein Extraction Kit
 ab65400 Plasma Membrane Protein Extraction Kit Instructions for Use For the rapid and sensitive extraction and purification of Plasma Membrane proteins from cultured cells and tissue samples. This product
ab65400 Plasma Membrane Protein Extraction Kit Instructions for Use For the rapid and sensitive extraction and purification of Plasma Membrane proteins from cultured cells and tissue samples. This product
Europium Labeling Kit
 Europium Labeling Kit Catalog Number KA2096 100ug *1 Version: 03 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Intended Use... 3 Background... 3 Principle of the Assay...
Europium Labeling Kit Catalog Number KA2096 100ug *1 Version: 03 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Intended Use... 3 Background... 3 Principle of the Assay...