Foot-and-mouth disease virus serotype A vaccine matching: Comparison of test
|
|
|
- Jason Shelton
- 5 years ago
- Views:
Transcription
1 CVI Accepts, published online ahead of print on 12 March 2014 Clin. Vaccine Immunol. doi: /cvi Copyright 2014, American Society for Microbiology. All Rights Reserved. 1 2 Foot-and-mouth disease virus serotype A vaccine matching: Comparison of test methodologies Tesfaalem Tekleghiorghis 1,3, Klaas Weerdmeester 1, Froukje van Hemert-Kluitenberg 1, Rob J.M. Moormann 1,2 and Aldo Dekker 1# 1 Central Veterinary Institute, part of Wageningen UR, Houtribweg 39, 8221 RA, Lelystad, The Netherlands 2 Department of Infectious Diseases & Immunology, Faculty of Veterinary Medicine, Utrecht University, Yalelaan 1, Utrecht 3584 CL, The Netherlands 3 National Veterinary Laboratory, Ministry of Agriculture, Asmara, Eritrea # Corresponding Author: Aldo Dekker, Central Veterinary Institute, Wageningen UR, Houtribweg 39, 8221 RA, Lelystad, The Netherlands, Tele.: , Aldo.Dekker@wur.nl Abstract Vaccination has been one of the most important interventions in disease prevention and control. The impact of vaccination largely depends on the quality and suitability of the chosen vaccine. To determine the suitability of the vaccine strain, antigenic matching is usually studied by in vitro analysis. In this study, we performed three in vitro test methods to determine which one gives the lowest variability and the highest discriminatory capacity. Binary ethylenimine inactivated vaccines, prepared from 10 different FMD virus serotype A strains, were used to 1
2 vaccinate cattle (5 animals for each strain). Antibody titers in 3 weeks post-vaccination (WPV) sera were determined by virus neutralization, neutralization index and liquid phase blocking ELISA. The titers were then used to calculate r 1 -values. These r 1 -values were compared to the genetic lineage using receiver operating characteristics (ROC) analysis. In the two neutralization test methods the median titer observed against the test strains differed considerably, and sera of vaccinated animals did not always show the highest titers against their homologous virus strain. When titers were corrected for test strain effect (scaling) the variability (standard error of the mean per vaccinated group) increased because the results were on a different scale, but the discriminatory capacity improved. ROC analysis of the r 1 -value calculated on both observed and scaled titers showed that only r 1 -values of the liquid phase blocking ELISA gave a consistent statistically significant result. Under the conditions of the present study, the liquid phase blocking ELISA showed less variation and still had a higher discriminatory capacity. INTRODUCTION Foot-and-mouth disease (FMD) is highly contagious infectious disease that affects all clovenhoofed animals and is one of the most economically important diseases of livestock. The virus belongs to the genus Aphthovirus, in the family of the Picornaviridae (1). It has seven different serotypes namely; O, A, C, Asia 1 and Southern African Territories (SAT)-1, 2 and 3 with multiple subtypes within each serotype (2). FMD virus (FMDV) is a highly variable RNA virus (3-5) and, in general, there is little or no cross-protection between serotypes and even between different strains of the same serotype (2, 6, 7). Serotype A is considered to be the most antigenically and genetically diverse of the FMDV serotypes and new antigenic variants emerge frequently (8, 9). 2
3 Inactivated vaccine of FMDV is being used to control FMD. This type of vaccine is commonly used in many parts of the world namely, South America, Asia and Africa where FMD is endemic. Although the quality of the vaccine used is probably the most important factor for the success of a vaccination program, a reasonable antigenic match between the FMDV vaccine and the outbreak virus strains is also considered essential for the effectiveness of the vaccine (6). Initially in vivo cross-protection studies were performed to test for antigenic differences. However, since serological tests became available, they are also being used for antigenic matching on the basis of the assumption/hypothesis that the level of protection is correlated to the antigenic match in the serological tests. In such serological tests the antibody titer of sera collected from vaccinated animals is determined against both the vaccine strain and field strain virus (6, 10). The most used value to express antigenic match is the r 1 -value (relationship coefficient), which is the ratio of the titers obtained in serological tests using the heterologous strains and the vaccine strain (7, 10-12). Serological test methods have been used to quantify antigenic differences between FMDV structural protein antibodies and thereby estimate the vaccine matching between a vaccine strain and a field isolate. Antigenic analysis of field isolates in relation to vaccine strains, based on virus neutralization tests (VNTs) plays a significant role in evaluating the suitability of existing vaccine strains (6, 10, 11), although significant variation has been reported with VNTs (13, 14). The genetic diversity in the P1-coding region within the SAT serotypes are reflected in the antigenic properties of these viruses and therefore have implications for the selection of vaccine strains that would provide the best vaccine match against emerging viruses (15). In the antigenically diverse FMDV serotype A strains, predicting antigenic differences using genetic 3
4 sequences alone does not provide reliable information for vaccine matching (16). Therefore, others have added structural information on the location of the amino acid sequence in the virus to the sequence data and have shown that it was a powerful tool to predict antigenic relationships that has the potential to be applied to a variety of different infectious agents (17). In studies in which cross-protection was related to cross-reactions in serology (10, 18, 19) the relation between cross-protection and cross-reactions varies. In some studies, protection was observed when high potency (type A) vaccines were being used (10) even when the r 1 -values were low (ranging from 0.04 to 0.23). In other studies, no cross-protection was detected after vaccination with A/Iran/05 vaccine and challenge with A/TUR/64/11 (18) while the r 1 -value 0.1 fell within the range of the r 1 -value found in the previous studies (10). For type O Manisa vaccine a reasonable high r 1 -value 0.6, against O Campos was observed, but a vaccine containing 15 µg of O Manisa antigen protect in 99% of the cases against homologous challenge but only in 54% of the cases against a O Campos challenge (19). This shows that a highly potent O Manisa vaccine did not protect sufficiently against a heterologous O Campos challenge even while the r 1 -value indicated it would. This shows that r 1 -values are not always consistent with the challenge results. Since protection against a challenge with FMD is well correlated with serology results (20), variation in r 1 -value determination might have caused this inconsistency. Cross validation studies on r 1 -value determination between laboratories (18, 21) showed huge variation between laboratories and techniques used. The exact cause of variation could not be established as there were too many variables. In a recent study, (22) analyzed the r 1 -value of the VNT using ROC analysis. They showed that the r 1 -value did not predict cross-protection. Other authors have linked genetic information to cross-reactions in serology (15-17) and have showed that the r 1 - value could be predicted from surface exposed amino acid changes. These studies did not 4
5 address the inherent variability of the r 1 -value and it is therefore important to have a more comprehensive view on the value of r 1 -values using results generated with various FMD strains in one laboratory. One of the first questions is to identify which test to be used to test crossreaction in serology. In this study, we analyzed the cross-reactions in serology using sera of cattle vaccinated with 10 different FMDV serotype A strain vaccines using three serological test methods, with the objective to determine which serological test method provides the least variation within a group of vaccinated cattle and provides the best discrimination between vaccines. MATERIALS AND METHODS FMDV serotype A strains. Different serotype A FMDV (n = 10) strains that were isolated in Africa, the Middle East and Europe were selected from various genetic lineages (Table 1). The 10 strains were: A/KEN/12/2005, A/ERI/2/98, A/SUD/2/84, A/ETH/13/2005 and A/MAU/1/2006, which were received from OIE/FAO World Reference Laboratory for FMD, The Pirbright Institute, UK and A 10 Holland/42, A 22 /IRQ/24/64, A/TUR/20/2006, A/TUR/14/98 and A/IRN/2/97 obtained from Central Veterinary Institute (CVI), part of Wageningen UR, Lelystad, the Netherlands. For control on the strain identity we sequenced the VP1 gene of the 10 FMDV serotype A strains and compared them to data in NCBI database. Vaccine (antigen production). The vaccines employed in this study were non-commercially produced at the CVI for this particular study. The vaccines were prepared from the aforementioned 10 different FMDV serotype A strains after growing them on monolayers of BHK-21 cells in 850 cm 2 roller bottles (CORNING, Tissue Culture Treated, Non-Pyrogenic, Corning Incorporated N.Y., USA). The viruses were subsequently inactivated with binary 5
6 ethyleneimine (BEI) (10 mm) (23, 24). The BEI was neutralized by sodium thiosulphate (0.79% w/v) and the inactivated antigens were concentrated with one cycle of polyethylene glycol (PEG) (8.0% w/v) precipitation. The antigens were checked by standard VP1 sequencing (25). The 146S antigen concentration was determined by quantitative sucrose density gradient analysis as previously described (26, 27). A known 146S FMDV antigen reference standard was tested with each run. The aqueous vaccines were formulated using aluminium hydroxide (alhydrogel) 2% (Ph. Eur. Brenntag Biosector) and saponin (10% w/v Quil-A solution in PBS) as adjuvant. The vaccine payload was 10 µg of antigen per 2 ml dose and expected to have 3 PD 50. The antigens were tested for the presence of live FMD virus both in vitro as well as in vivo. Vaccination of cattle. Vaccination was performed in Healthy, unvaccinated, Eritrean local indigenous zebu breed (Bos indicus) cattle, 10 to 18 months of age were used for this study. The cattle used originated from regions known to be free from FMD without vaccination. Before purchase and prior to vaccination, all cattle were ear tagged and bled twice at a two weeks interval and tested for presence of anti-fmdv non-structural protein (NSP) antibodies using the FMD NS-ELISA (PrioCHECK) (28). Only cattle with a negative result in the FMD NS-ELISA were purchased. The cattle were housed in stables at the National Veterinary Laboratory, Asmara, Eritrea. The cattle were randomly allocated to 11 groups of 5 cattle. In total, we used 10 groups for the 10 different FMDV serotype A vaccines and 10 cattle as non-vaccinated controls to monitor incursions of FMD infection which would influence the experiment. Cattle in each vaccine group were vaccinated subcutaneously in the middle of the cervical area. Blood samples were collected at 0, 7, 14, 21 and 28 days post vaccination for serology testing. Rabbit and guinea pig antisera for liquid phase blocking ELISA (LPB-ELISA). In total 20 rabbits and 20 guinea pigs (two for each FMDV serotype A strain antigens) were immunized 6
7 as described by Hamblin (29) using the same semi-purified PEG precipitated antigens as those used to vaccinate the cattle in this study. The vaccine was prepared by mixing an FMD serotype A antigen with an equal volume of Montanide ISA 206VG oil adjuvant to get a final concentration of 5 µg/ml. The rabbits were vaccinated subcutaneously and intramuscularly with 0.5 ml of vaccine on each site. The guinea pigs were vaccinated subcutaneously with 0.5 ml of vaccine. All animals were boosted 2 WPV with the same protocol used for primary vaccination. The animals were bled days after the booster vaccination by exsanguination under general anaesthesia. FMDV NS ELISA. PrioCHECK FMDV NS ELISA (Prionics ) (28) is a blocking ELISA that detects antibodies directed against the non-structural 3ABC protein of FMDV. This ELISA detects FMDV infected animals independent of the serotype causing the infection. Serum samples were tested by PrioCHECK FMDV NS ELISA kit and test results were interpreted as indicated by the producer, i.e.: < 50 percent inhibition (PI) negative to NS antibodies and 50 PI values as positive to NS antibodies. Virus neutralization test. VNT was performed as described in the OIE Manual of Diagnostic Tests and Vaccines for Terrestrial Animals (7). It was performed using BHK-21 cells in flatbottomed tissue culture grade microtiter plates (Greiner Bio-one). Stock viruses for the VNT were grown in cell monolayers, titrated and stored at -70 C until used. The experimental sera were inactivated at 56 C for 30 minutes before testing. Medium (DMEM; Gibco/Invitrogen) supplemented with 5% foetal calf serum (FCS) and 2% antibiotics was used for diluting the sera, the virus and as medium for growing the cells. All 3 WPV sera were tested in duplicate by mixing 50 µl of 2-fold dilutions of the serum with 50 µl virus suspension containing 100 TCID 50 FMDV. This mixture was incubated for 1 hour at 37 C, 5% CO 2 in a humidified atmosphere. 7
8 After 1 hour incubation, 150 µl BHK-21 cell suspension was added to each well and the plates were incubated for 3 days at 37 C, 5% CO 2 in a humidified atmosphere. After 3 days, the monolayers were stained with 50 µl amido-black solution (1 g/l in water containing 88 mm sodium acetate, 10% glycerol and 5.4% acetic acid), and cytopathic effect (CPE) was read macroscopically. The end point titers of the serum samples, tested against all 10 vaccine strains, were expressed as the logarithm (base 10) of the reciprocal of the last dilution of serum that neutralized 100 TCID 50 of the virus in 50% of the wells (30). Sera collected at 0, 1, 2, 3 and 4 WPV were tested by VNT using the homologous test strain to see the response of neutralizing antibodies post-vaccination. For comparison of tests the 3 WPV sera were tested against all 10 virus strains used for vaccination. In the calculation of the mean titers, VNT titers with a titer of < 0.3 at the day of vaccination were considered 0. In the calculations of r 1- values and scaled titers, the titers below the detection limit (< 0.3) were excluded since no titer was measured. Neutralization index test (NIT). In the NIT 50 µl of 10-fold dilutions of the virus (6 wells per dilution) were incubated with 50 µl of a fixed dilution of a 1:25 dilution of test serum for 1 hour at 37 C, 5% CO 2 in a humidified atmosphere. In each test a titration mixed with 50 µl of dilution medium (DMEM with 2% foetal bovine serum and antibiotics) instead of test serum was included. After one hour, 150 µl BHK-21 cell suspension was added to each well and the plates were incubated for 3 days at 37 C, 5% CO 2 in a humidified atmosphere. After 3 days, the plates were washed and the monolayers were stained with 50 µl of amido-black solution (see above), and CPE was read macroscopically. The log 10 titers of the virus with and without serum was calculated (30). The neutralization index log 10 titer is calculated by subtracting the virus log 10 8
9 titer of the strain with test serum from the log 10 titer of the strain without serum. All 3 WPV sera were tested against all 10 FMDV serotype A strains used for vaccination. LPB-ELISA. The LPB-ELISA was performed as described by (29). Specific rabbit sera for coating and the guinea-pig sera for detection were produced for each of the 10 strains. Rabbit and guinea-pig antisera were partially purified by precipitation using saturated aluminium sulphate. The dilutions of coating antibody and detecting antibodies were optimized in the laboratory as well as the rabbit anti-guinea-pig conjugate (Dako P0141). Antigens were prepared from the 10 FMDV type A strains by growing them on monolayers of BHK-21 cells. After freeze/thawing the cell culture and centrifuging for 10 minutes at 250 x g, the un-purified supernatants were titrated and the final dilution that gave an OD (optical density) of approximately 1.2 to 1.5 at 450 nm in the ELISA was used. ELISA plates (Corning Costar) were coated with a predetermined dilution of 100 µl/well rabbit anti FMDV antibodies diluted in bicarbonate buffer ph 9.6 left overnight at room temperature. At the same day in U-bottomed microtiter plates (dummy plate) each test serum was diluted, 2-fold dilution series, in duplicate wells starting with a 1:5 dilution (60 µl/well) using phosphate-buffered saline containing 0.05% Tween 80, 0.5M NaCl and 5% FCS (ELISA buffer). 60 µl of a predetermined concentration of antigen, diluted in ELISA buffer, was added, mixed and incubated overnight at 4 C. The next day, the ELISA plates were washed 6 times with tap water containing 0.05% Tween 80. Then 100 µl of test serum/antigen mixture was transferred from dummy plates into the corresponding wells of the rabbit-serum coated ELISA plates and incubated at 37 C for 1 hour. After washing, 100 µl of the guinea pig antiserum (in a predetermined dilution) homologous to the viral antigen used was added to each well and incubated at 37 C for 1 hour. The plates were washed and 100 µl of a predetermined dilution rabbit anti-guinea pig immunoglobulins conjugated with 9
10 horseradish peroxidase was added to all wells and incubated for 1 hour at 37 C. The guinea pig antiserum and the rabbit anti-guinea pig conjugate were diluted in ELISA buffer. After washing, 100 µl of a substrate/chromogen solution (BioFX TMBS ) was added and the plates were left to develop colour for 10 minutes at room temperature. The reaction was stopped by adding 100 µl 0.5 M sulphuric acid. Plates were read at 450 nm and log 10 antibody titers were expressed as the log 10 of the reciprocal of the final dilution of serum giving 50% of the mean OD value recorded in the maximum signal control wells (two wells with ELISA buffer instead of serum). Data Analysis Mean titers. Mean titers were computed by adding the log 10 titers and divided by the number of observations. Scaled titer (correction for strain effect). Scaled titers were used to minimize variations observed between neutralization tests with different viruses. For each specified serotype A strain the titer was standardized using the following formula: Scaled titer observed titer - mean titer for all sera tested against a specified test strain standard deviation of all sera tested against a specified test strain So for each specified strain the mean scaled titer will be 0 with a standard deviation of 1.0 (31). Determination of r 1- value. One-way antigenic relationship (r 1- value) for each 3 WPV sera were determined using VNT, NIT and LPB-ELISA titers as described in the OIE manual (7) using: r1- value Serum titer against heterologous (field) virus Serum titer against homologous(vaccine) virus 10
11 To test whether the original and scaled titers of the sera from vaccinated cattle were able to discriminate between the various vaccine strains we used an analysis of variance (ANOVA). In the ANOVA we tested whether the titers induced by each vaccine were similar. If the ANOVA showed that one or more of the vaccine groups were significantly different, all possible pairwise comparisons were tested with a t-test using Holme s adjustment for multiple testing. A p-value < 0.05 was considered statistically significant. Statistical analyses were carried out using R version 2.14 (32). Receiver Operating Characteristics (ROC) analysis. A ROC analysis was carried out to determine which serological technique and the r 1 -value derived from that technique had the best sensitivity and specificity in determining differences between test strains. In the first analysis we assumed that the two strains from lineage Iran-96 have the same antigenic make-up (gold standard for the sensitivity and specificity). In this case the amino acid homology in VP1 was 97.5%. In the second, we assumed that strains within both the Iran-96 and the G-IV lineages were antigenically similar. In that case the amino acid homology in VP1 was 93.1%. The ROC analysis was performed using the proc library in R (33). RESULTS FMD vaccine strains. The antigens used for vaccination came from 8 different lineages defined by the World Reference Laboratory in Pirbright (Table 1). In two lineages we had two vaccine candidates. In lineage Iran-96 we selected A/IRN/2/79 and A/TUR/14/98, which were 97.7% similar in VP1 on amino-acid level. And in lineage G-IV we selected A/ERI/2/98 and A/SUD/2/84, which were 93.1% similar in VP1 on amino-acid level. The identity on amino-acid level ranged from % for strains that did not belong to the same lineage. The VP1 11
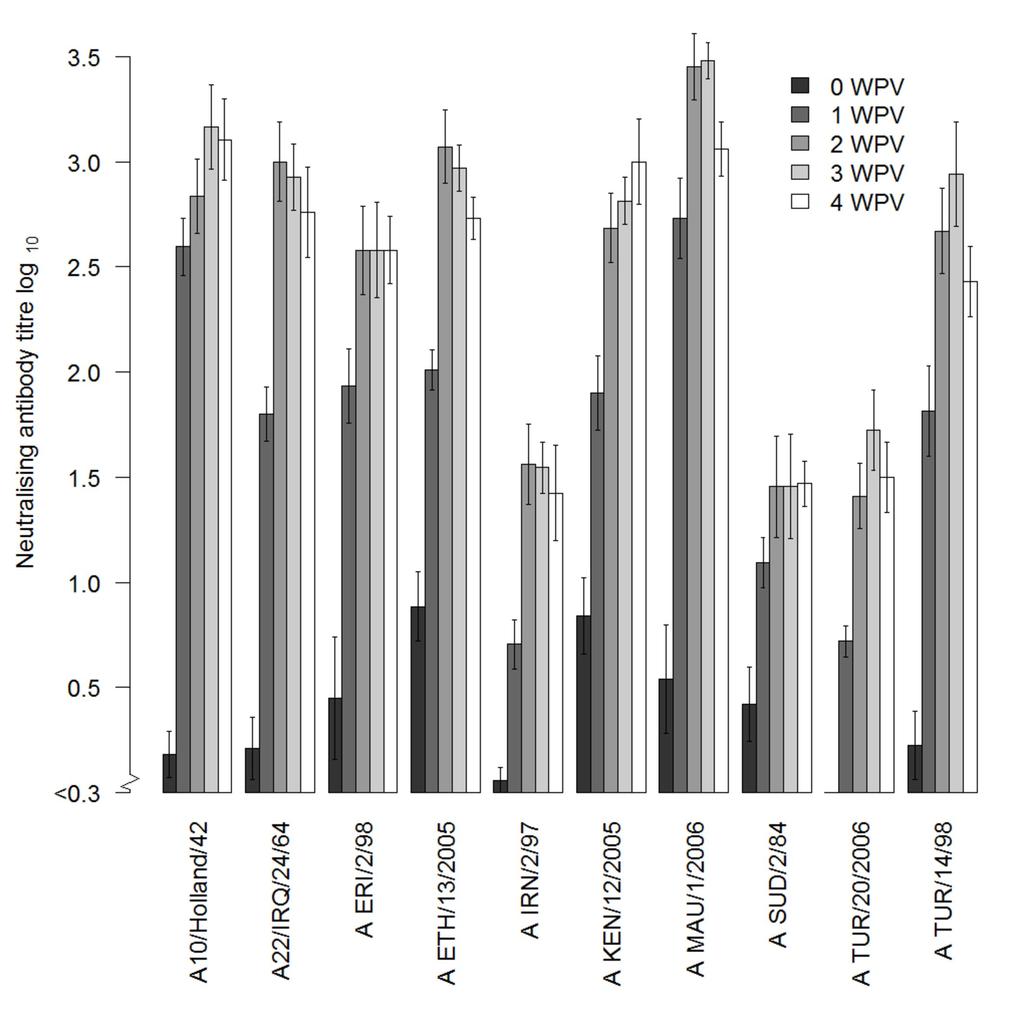
12 sequences were compared to known sequences in the GenBank. For most strains, a match (99-100% nucleotide identity) was found and is shown in Table 1. Only for our A/KEN/12/2005 VP1 sequence, the best match (93% on nucleotide level) was with A/KEN/22/2009 (GenBank accession number KF112912). Both the in vitro and in vivo innocuity tests did not detect live FMDV in the BEI-inactivated antigens produced from these 10 strains. FMDV NS ELISA. All sera collected at the start of the experiment were negative in the Prionics NS ELISA (28). In total 7 cattle were positive in the NS ELISA post-vaccination, in 4 of them only at one time point post-vaccination (1, 3 or 4 WPV) in 2 cattle at two time points, and in one cow at 2, 3 and 4 WPV. The maximum percentage inhibition observed was 58%. Post-vaccination antibody response. Serum samples collected from all cattle at 1, 2, 3 and 4 WPV were tested using VNT against the homologous vaccine test strain. In some cattle a low reaction was detected in the VNT before vaccination (Fig. 1), this was also seen in the sera of the control animals, especially with test strain A 10 /Holland/42. All vaccinated cattle responded to vaccination with an increase in log 10 antibody titer in the first week ranging from 0.3 (2-fold) to 3.0 (1000-fold) with a median of 1.1. At 2 4 WPV the increase was at least 0.6 (4 fold) compared to the log 10 titer before vaccination with a medium increase of 2.1 log 10. The mean 3 WPV homologous log 10 titer in the VNT ranged from (Fig. 1). The mean VNT titer 3 WPV against the homologous and 9 heterologous virus test strains used in the VNT ranged in the different vaccine groups from 1.6 for A/MAU/1/2006 to 2.0 for A/TUR/20/2006 with an overall mean titer of 1.8. Analyzing differences between test strains using different serological test methods. All 3 WPV sera were tested and analyzed by the three serological assays (VNT, NIT and LPB-ELISA) against the homologous and heterologous virus strains. Mean titers and standard error of the 12
13 mean (SEM) were calculated for each vaccine (5 sera per vaccine) (Fig. 2). The correlation between VNT and NIT was higher (0.72) than correlation between NIT and LPB-ELISA (0.55) or VNT and LPB-ELISA (0.49). Some virus test strains were more easily neutralized than others. Consequently, the mean titer against the homologous strain is not always the highest one. With test strains A/IRN/2/97, A/SUD/2/84 and A/TUR/20/2006, low responses were observed with all sera from vaccinated animals in both VNT and NIT, similar to the homologous strain shown in Fig. 1. Because of these inherent variations between the test strains, many sera were found where the homologous titer was lower than some of the heterologous ones. Fig. 2 shows that the mean VNT (in 4 of 10 strains), the mean NIT (in 3 out of 10 strains) and the mean LPB-ELISA (in 2 out of 10 strains) homologous titer was not the highest. Scaled titers (correction for strain effect). Because some virus strains were more easily neutralized than others, as mentioned above, we used scaled titers to correct and standardize for the test strain effect. Using scaled titers the mean homologous response is in general the highest one (Fig. 3). Table 2 shows the range of the standard error of the mean (SEM) determined for each group of 5 vaccinated cattle tested against each of the test strains for both observed and scaled titers. Before scaling the LPB-ELISA had the lowest maximum SEM. After scaling both NIT and LPB-ELISA have the lowest maximum SEM. r 1 -value. One would expect the r 1 -value to range between 0 and 1, but that was only observed in 78%, 92% and 96% for VNT, NIT and LPB-ELISA respectively (Table 3). The maximum r 1 - values were 178, 1148 and 2.8 for VNT, NIT and LPB-ELISA respectively. After scaling a higher proportion of r 1 -values were found between 0 and 1 (Table 3). Discriminating between vaccines. As stated above the mean titers induced by the different vaccines were similar, we therefore used the ANOVA to determine whether there are significant 13
14 differences between vaccines. Using titers and scaled titers obtained by VNT no significant differences between vaccines were found. Using titers and scaled titers obtained by NIT a significant difference between test strains was observed in the ANOVA and in 1 (based on titer) or 3 (based on scaled titers) out of the 45 (Table 4A and 4B) possible pairwise comparisons. Using titers and scaled titers obtained by LPB-ELISA, a significant difference between test strains was observed in the ANOVA, and in 7 (based on titers) or 12 (based on scaled titers) out of the 45 possible pairwise comparisons (Table 4C and 4D). ROC analysis. In the dataset we had 2 genetic lineages (Iran-96 and G-IV) that contained each 2 strains. In the ROC analysis we compared the sensitivity and specificity of the r 1 -values of the different test methods before and after scaling. In the first analysis we assumed that only strains A/IRN/2/97 and A/TUR/14/98 were antigenically similar (Fig. 4A for titers and 4B for scaled titers). In the second analysis we assumed that also strains A/ERI/2/98 and A/SUD/2/84 were antigenically similar (Fig. 4C for titers and 4D for scaled titers). The 95% confidence interval of the area under the curve (AUC) of the ROC curve of the r 1 -value based on the LPB-ELISA titers and scaled titers did not include 0.5, hence showing a statistically significant result. However, for VNT and NIT only when using the assumption that the strains in both lineages are antigenically similar, the AUC of the ROC curve of the r 1 -values based on scaled NIT titers (AUC = 0.67, 95% CI , Fig. 4D) had a 95% confidence interval that did not include 0.5. So the dashed line in Fig. 4D was the only statistically significant result for NIT in the ROC analysis. DISCUSSION The study was performed to determine which serological test method provides the least variation within a group of vaccinated cattle and provides the best discrimination between vaccines. We 14
15 selected 10 FMDV serotype A strains from 8 different lineages (Table 1). Sequencing of the VP1 region of the genome confirmed the identity of the strains used. Although Eritrea is not free from FMD, the cattle used in this study were obtained from a region where no outbreaks of FMD were recorded. All cattle were free of antibodies to non-structural proteins of FMD virus at the start of the study implying that, they had not been exposed to FMDV. Before vaccination the cattle were tested for neutralizing antibodies against 5 other serotypes (O, C, Asia 1, SAT-1 and SAT-2) with a negative result. Some cattle showed low antibody responses to NS protein postvaccination, but no FMDV clinical signs were observed. The observed 58% inhibition is just above the cut-off of 50% in the test. Probably the NS response was due to some remaining NS proteins present in the vaccine, as has been described before (34). The low titers found before vaccination in some of the cattle in both VNT and LPB-ELISA is similar to the results reported for cattle in the Netherlands (35). Using the 10 strains as vaccine antigen, we observed a good homologous neutralizing antibody response with at least a 4-fold increase in titer two WPV. The small differences, mean titers ranging from , observed in the mean response of each group of vaccinated cattle tested against all test strains show that the quality of the vaccines was similar. So this experimental design in which cattle are vaccinated with vaccines containing a similar amount of antigen offers an excellent opportunity to compare the different serological test methods. The major difficulty in interpreting the results with both VNT and NIT was the difference in the level of neutralization observed using different virus test strains. Some virus test strains like A 10 /Holland/42 were neutralized easily with high titers in homologous and heterologous sera. For this reason we also evaluated scaled titers. Even though a difference in titer level was observed the mean response within the groups of 5 vaccinated cattle was consistent, resulting in 15
16 a small range of observed SEM's in all the 3 test methods. The range of observed SEM, however, was lowest in the LPB-ELISA showing that the lowest amount of variation is observed with the LPB-ELISA results. Lower variation in the LPB-ELISA compared to the VNT has also been reported by Van Maanen and Terpstra (36). After scaling the range of the SEM observed in the groups of cattle increased, but still the lowest maximum SEM was found using the LPB-ELISA, but after scaling the results obtained with the NIT were similar. The titer differences between test strains resulted in r 1- values that were often higher than one. This was mainly observed with the seven test strains that had high homologous neutralizing antibody titers. After scaling, lower r 1 -values were calculated, this is consistent with the fact that the strains were obtained from 8 distinct genetic lineages from which we assume that they are also antigenically different. Using the LPB-ELISA more significant differences between vaccines could be observed by ANOVA followed by pairwise t-tests. The four strains from 2 genetic lineages were still similar in the pairwise t-test (Table 4). Not only were the LPB-ELISA results more capable to distinguish between vaccines, they are also more reproducible as they are not influenced by variations in tissue culture susceptibility. Although neutralizing antibodies are often considered in relation to protection, LPB-ELISA titers are also correlated with protection (36, 37). So LPB-ELISA seems to perform better than the neutralization tests to discriminate between vaccines. To further build on the assumption that genetically related strains should also be antigenically related we did an ROC analysis using the r 1 -values derived from the different tests before and after scaling. A ROC curve with an AUC of 0.5 would indicate that the test does not differentiate the differences defined by the gold standard (in our case the sequence data). Before scaling only the r 1 -values based on titers from the LPB-ELISA had an AUC of the ROC curve which was 16
17 significantly different from 0.5. Therefore, without scaling the titers, the LPB-ELISA is the only test that can discriminate between strains that are genetically different, and therefore also assumed to be antigenically different. After scaling also the AUC of the ROC curve of the NIT, when assuming the strains in both the Iran-96 and the G-IV lineage were similar, and was significantly different from 0.5. This shows that scaling can help to improve the NIT for discriminating strains antigenically, but the LPB-ELISA is the best test for this. In our analysis, we used the r 1 -value as a measurement of the antigenic relation, but the reason we do the analysis is to see whether a vaccine can protect. Previous studies have shown that there is a strong relation between antibody response and protection (36-38). The precision of this relation can slightly be improved by the inclusion of other immunological techniques (39, 40). But the height of the titer that relates with protection is not the same in different strains. Therefore, the use of an r 1 -value to estimate protection against challenge is not logical because the difference in titer needed for protection is not taken into consideration. Hence, if the objective is to choose a vaccine that will best protect against infection with a field isolate, it would be better to look for the vaccine that induced the highest titer against the field virus strain instead of looking at the r 1 -value. In a recent study on analysis of the serological results of crossprotection tests (22), it was shown that the r 1 -value of a serum of a vaccinated and heterologous challenged cow did not predict protection, but the serum titer against the challenge strain in the VNT did. In fact it was shown that analysis of the humoral immune response using IgG 1 and IgG 2 ELISAs even improved this prediction (22). This also indicates that VNT or ELISA titers are probably more important for prediction of protection than the r 1 -value. As stated before, the cattle sera used in this study are unique, as they were produced using the same amount of antigen in each vaccine batch. These cattle sera can be very valuable for further 17
18 research in antigenic differences of new serotype A FMDV isolates as they cover a large part of the genetic differences observed in Europe, Middle East and Africa. We conclude that the LPB- ELISA, under the conditions of the present study, is the best test for detecting antigenic differences between FMDV strains. As the variability within a vaccinated group of animals is lower and its discriminatory capacity is higher than with the neutralization tests. However, one should realize that in our study we used strain specific guinea-pig and rabbit antibodies. Nevertheless, also neutralizing antibody tests have their place as VNT titers are very well related with protection (36, 38), but in that case the use of r 1 -values is not feasible and suitability of a vaccine strain should be evaluated on the titer against the outbreak strain. The higher the titer the better the vaccine is suited. ACKNOWLEDGEMENTS This work was supported by the International Atomic Energy agency (IAEA) grant C6/ERI/07010, Vienna, Austria. The production of the guinea-pig and rabbit antisera was supported by the EuFMD. We thank the The Pirbright Institute, UK for providing FMDV type A strains. We are grateful to Tsghe Kfleyesus, Ghebremichael G.Egziabhier and the late Tesfu Yemane, Animal Health staff, (MoA), Asmara, Eritrea for assisting in sample collection during animal experiment. We are grateful to Boonstra Christine for sequencing the FMDV serotype A strains. We would also like to thank the FAO, Representative Office in Eritrea for providing internet access during the study period. REFERENCES 18
19 Pereira HG Foot-and-mouth disease., p In E.P.J. G (ed.), Virus disease of food animals. Academic Press Inc., London. 2. Brooksby JB Portraits of Viruses: Foot-and-Mouth Disease Virus. Intervirology 18: Holland J, Spindler K, Horodyski F, Grabau E, Nichol S, VandePol S Rapid evolution of RNA genomes. Science Holland JJ, Torre JC, Steinhauer DA RNA Virus Populations as Quasispecies, p In Holland J (ed.), Genetic Diversity of RNA Viruses, vol Springer Berlin Heidelberg. 5. longjam N, Tayo T Antigenic variation of Foot and Mouth Disease Virus - An Overview. Vet World 4: Paton DJ, Valarcher JF, Bergmann I, Matlho OG, Zakharov VM, Palma EL Selection of foot and mouth disease vaccine strains a review. Rev. sci. tech. Off. int. Epiz., 24: OIE Manual of Diagnostic Tests and Vaccines for Terrestrial Animals, Chapter Foot-and-mouth disease, 6th ed, vol Knowles NJ, Samuel AR Molecular epidemiology of foot-and-mouth disease virus. Virus Res. 91: Mittal M, Tosh C, Hemadri D, Sanyal A, Bandyopadhyay SK Phylogeny, genome evolution, and antigenic variability among endemic foot-and-mouth disease virus type A isolates from India. Arch. Virol. 150: Brehm KE, Kumar N, Thulke HH, Haas B High potency vaccines induce protection against heterologous challenge with foot-and-mouth disease virus. Vaccine 26: Jangra RK, Tosh C, Sanyal A, Hemadri D, Bandyopadhyay SK Antigenic and genetic analyses of foot-and-mouth disease virus type A isolates for selection of candidate vaccine strain reveals emergence of a variant virus that is responsible for most recent outbreaks in India. Virus Res 112: Mattion N, Goris N, Willems T, Robiolo B, Maradei E, Beascoechea CP, Perez L, Smitsaart E, Fondevila N, Palma E, De Clercq K, Torre JL Some guidelines for determining foot-and-mouth disease vaccine strain matching by serology. Vaccine 27: Rweyemamu MM, Booth JC, Head M, Pay TWF Microneutralization tests for serological typing and subtyping of foot-and-mouth disease virus strains. Epidemiol Infect 81: Rweyemamu MM Antigenic variation in foot-and-mouth disease: studies based on the virus neutralization reaction. J Biol Stand 13: Maree FF, Blignaut B, Esterhuysen JJ, de Beer TAP, Theron J, O'Neill HG, Rieder E Predicting antigenic sites on the foot-and-mouth disease virus capsid of the South African Territories types using virus neutralization data. The Journal of general virology 92: Ludi A, Horton D, Li Y, Mahapatra M, King DP, Knowles NJ, Russell C, Paton D, Wood J, Smith D, Hammond J Antigenic variation of foot-and-mouth disease virus serotype A. J Gen Virol (In Press). 19
20 Reeve R, Blignaut B, Esterhuysen JJ, Opperman P, Matthews L, Fry EE, de Beer TAP, Theron J, Rieder E, Vosloo W, O'Neill HG, Haydon DT, Maree FF Sequence-Based Prediction for Vaccine Strain Selection and Identification of Antigenic Variability in Foot-and-Mouth Disease Virus. PLoS comput biol 6: Anonymous FMD DISCONVAC 3rd periodic report; WP3. Assessment and improvement of heterologous protection by FMD vaccines. 19. Nagendrakumar SB, Srinivasan VA, Madhanmohan M, Yuvaraj S, Parida S, Di Nardo A, Horsington J, Paton DJ Evaluation of cross-protection between O1 Manisa and O1 Campos in cattle vaccinated with foot-and-mouth disease virus vaccine incorporating different payloads of inactivated O1 Manisa antigen. Vaccine 29: Pay TWF, Hingley PJ Correlation of 140S antigen dose with the serum neutralizing antibody response and the level of protection induced in cattle by foot-andmouth disease vaccines. Vaccine 5: Hammond J OIE/FAO FMD Reference Laboratory Network Annual Report. The Pirbright Institute, UK. 22. Brito BP, Perez AM, Capozzo AV Accuracy of traditional and novel serology tests for predicting cross-protection in foot-and-mouth disease vaccinated cattle. Vaccine 32: Bahnemann HG Binary ethylenimine as an inactivant for foot-and-mouth disease virus and its application for vaccine production. Arch Virol 47: Bahnemann HG Inactivation of viral antigens for vaccine preparation with particular reference to the application of binary ethylenimine. Vaccine 8: Knowles NJ, Samuel AR RT-PCR and sequencing protocols for the molecular epidemiology of exotic virus diseases of animals. OIE/FAO World Reference Laboratory for Foot-and-mouth disease, Institute for Animal Health, Pirbright, UK: Doel TR, Baccarini PJ Thermal stability of foot-and-mouth disease virus. Arch Virol 70: Wagner GG, Card JL, Cowan KM Immunochemical studies of foot-and-mouth disease. VII. Characterization of foot-and-mouth disease virus concentrated by polyethylene glycol precipitation. Arch. Gesamte Virusforsch 30: Sørensen KJ, Madsen KG, Madsen ES, Salt JS, Nqindi J, Mackay DKJ Differentiation of infection from vaccination in foot-and-mouth disease by the detection of antibodies to the non-structural proteins 3D, 3AB and 3ABC in ELISA using antigens expressed in baculovirus. Arch. Virol. 143: Hamblin C, Barnett ITR, Hedger RS A new enzyme-linked immunosorbent assay (ELISA) for the detection of antibodies against foot-and-mouth disease virus I. Development and method of ELISA. J Immunol Methods 93: Karber G Beitrag zur kollektiven Behandlung pharmakologischer Reihenversuche. Arch. Exp. Pathol. Pharmakol. 162: Becker RA, Chambers JM, Wilks AR The New S Language, A Programming; Environment for Data Analysis and Graphics. Wadsworth & Brooks/Cole, Computer Science Series, Pacific Grove, CA, USA. 32. R Development Core Team R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. 20
21 Robin X, Turck N, Hainard A, Tiberti N, Lisacek F, Sanchez J-C, Müller M proc: an open-source package for R and S+ to analyze and compare ROC curves. BMC Bioinformatics 12: Doel TR Review FMD vaccines. Virus Res 91: Moonen P, Miedema GKW, van Hemert-Kluitenberg F, Chenard G, Dekker A Comparison of FMDV neutralisation tests using three different cell lines: validation of the new FAO reference sera, Session of the research group of the European commission for the control of foot-and-mouth disease, Borovets, Bulgaria 36. Van Maanen C, Terpstra C Comparison of a liquid-phase blocking sandwich ELISA and a serum neutralization test to evaluate immunity in potency tests of foot-andmouth disease vaccines. J Immunol Methods 124: Maradei E, La Torre J, Robiolo B, Esteves J, Seki C, Pedemonte A, Iglesias M, D Aloia R, Mattion N Updating of the correlation between lpelisa titers and protection from virus challenge for the assessment of the potency of polyvalent aphtovirus vaccines in Argentina. Vaccine 26: Hingley PJ, Pay TWF Sources of variability in foot and mouth disease vaccine potency estimates based on serum neutralizing antibody assay. J Biol Stand 15: McCullough KC, D e Simone F, Brocchi E, Capucci L, Crowther JR, Kihm U Protective immune response against foot-and-mouth disease. J Virol 6: Lavoria MÁ, Di-Giacomo S, Bucafusco D, Franco-Mahecha OL, Pérez-Filgueira DM, Capozzo AV Avidity and subtyping of specific antibodies applied to the indirect assessment of heterologous protection against Foot-and-Mouth Disease Virus in cattle. Vaccine 30: Carrillo C, Tulman ER, Delhon G, Lu Z, Carreno A, Vagnozzi A, Kutish GF, Rock DL Comparative Genomics of Foot-and-Mouth Disease Virus. J Virol Chitray M Investigation potential factors affecting foot-and-mouth disease virus internalisation. Submitted in partial fulfilment of the requirements for the degree of Master of Science. University of Pretoria, Pretoria, South Africa. 43. Ayelet G, Mahapatra M, Gelaye E, Egziabher BG, Rufeal T, Sahle M, Ferris NP, Wadsworth J, Hutchings GH, Knowles NJ Genetic Characterization of Footand-Mouth Disease Viruses, Ethiopia, Emerg Infect Dis. 15: Xu L, Hurtle W, Rowland JM, Casteran KA, Bucko SM, Grau FR, Valdazo- González Bn, Knowles NJ, King DP, Beckham TR, McIntosh MT Development of a universal RT-PCR for amplifying and sequencing the leader and capsid-coding region of foot-and-mouth disease virus. J Virol Methods 189: Habiela M, Ferris NP, Hutchings GH, Wadsworth J, Reid SM, Madi M, Ebert K, Sumption KJ, Knowles NJ, King DP, Paton DJ Molecular Characterization of Foot-and-Mouth Disease Viruses Collected from Sudan. Transbound. Emerg. Dis. 57: Knowles NJ, Nazem Shirazi MH, Wadsworth J, Swabey KG, Stirling JM, Statham RJ, Li Y, Hutchings GH, Ferris NP, Parlak Ü, Özyörük F, Sumption KJ, King DP, Paton DJ Recent Spread of a New Strain (A-Iran-05) of Foot-and-Mouth Disease Virus Type A in the Middle East. Transbound. Emerg. Dis. 56:
22 Klein J, Parlak U, Ozyoruk F, Christensen LS The molecular epidemiology of foot-and-mouth disease virus serotypes A and O from 1998 to 2004 in Turkey. BMC Vet Res 2: TABLE 1 FMDV serotype A strains used in the study. Ten strains came from 8 different lineages (as defined by the World Reference Laboratory). Two lineages, Iran-96 and G-IV contained two strains. Within lineage Iran-96 both strains were 97.7% similar in VP1 on aminoacid level, within lineage G-IV both strains were 93.1% similar in VP1 on amino-acid level. FMDV type A strain Lineage / sub-lineage * Accession no. (VP1) Reference A10/Holland/42 EURO-SA AY (41) A22/IRQ/24/64 ASIA AY (41) A/ERI/2/98 G-IV EU (42) A/ETH/13/2005 G-II FJ (43) A/IRN/2/97 Iran-96 KF (43) A/KEN/12/2005 G-I KF (16) A/MAU/1/2006 G-VI JF (44) A/SUD/2/84 G-IV GU (45) A/TUR/20/2006 Iran-05 FG (46) A/TUR/14/98 Iran-96 DQ (47) *Best matching VP1 sequence in NCBI TABLE 2 Comparison of the 3 serological test methods using titers and scaled titers. Comparison of the range of SEM within groups of 5 cattle. Test method Range of SEM of observed titers Range of SEM of Scaled titers 22
23 VNT NIT LPB-ELISA TABLE 3 The percentage of r1 values within the range 0 1 before and after scaling the titers. Test Before scaling titer After scaling titer VNT 78 % 93 % NIT 83 % 94 % LPBE 96 % 96 % Downloaded from on December 3, 2018 by guest 23
24 TABLE 4 P-values of the pairwise t-test results from the serological tests in which the ANOVA was significant (in bold italics are the p-values lower than 0.05). A: the results of the comparison of the observed titers of the NIT. B: the results of the comparison of the scaled titers of the NIT. C: the results of the comparison of the observed titers of the LPB-ELISA. D: the results of the comparison of the scaled titers of the LPB-ELISA Downloaded from on December 3, 2018 by guest 24
25 A A10/Holland/42 A22/IRQ/24/64 A/ERI/2/98 A/ETH/13/2005 A/IRN/2/97 A/KEN/12/2005 A/MAU/1/2006 A/SUD/2/84 A/TUR/20/2006 A 22 /IRQ/24/ A/ERI/2/ A/ETH/13/ A/IRN/2/ A/KEN/12/ A/MAU/1/ A/SUD/2/ A/TUR/20/ A/TUR/14/ B A 22 /IRQ/24/ A/ERI/2/ A/ETH/13/ A/IRN/2/ A/KEN/12/ A/MAU/1/ A/SUD/2/ A/TUR/20/ A/TUR/14/ C A 22 /IRQ/24/ A/ERI/2/ A/ETH/13/ A/IRN/2/ A/KEN/12/ A/MAU/1/ A/SUD/2/ A/TUR/20/ A/TUR/14/ D A 22 /IRQ/24/ A/ERI/2/ A/ETH/13/ A/IRN/2/ A/KEN/12/ A/MAU/1/ A/SUD/2/ A/TUR/20/ A/TUR/14/
26 FIG 1 VNT homologous mean titers including standard error of the mean of FMDV serotype A strains post-vaccination responses in cattle at 0, 1, 2, 3 and 4 WPV FIG 2 Mean titers and the standard error of the mean (SEM) of the three serological methods, i.e., VNT (black), NIT (grey) and LPB-ELISA (light grey) for each vaccine and tested against the corresponding test strain (equal number means the same strain). The individual graphs show the log 10 mean titers and the standard error of the mean of the sera of each group of vaccinated cattle (5 cattle) tested against the 10 different FMDV type A test strains (X-axis). The homologous responses are those were the number of the test strain (X-axis) is equal to the number in the main title of the separate graph. FIG 3 Mean scaled titers including the standard error of the mean of VNT (black), NIT (grey) and LPBE (light grey). The homologous responses are those were the number of the test strain (X-axis) is equal to the number in the main title of the separate graph. 26
27 FIG 4 ROC curves of the r 1 -value titres of the VNT (solid line), NIT (dashed line) and LPB- ELISA (dotted line). In the graphs, the area under the curve (AUC) is given including the 95% confidence interval, between brackets. Graph A and B: assuming strains A/IRN/2/97 and A/TUR/14/98 are antigenetically the same, using the observed titers (A) and scaled titers (B). Graph C and D: assuming strains A/IRN/2/97 and A/TUR/14/98 are antigenetically the same as well as A/ERI/2/98 and A/SUD/2/84, using the observed titers (C) and scaled titers (D). Downloaded from on December 3, 2018 by guest 27
28
29
Foot-and-mouth disease (FMD) is highly contagious infectious
 is highly contagious infectious") Comparison of Test Methodologies for Foot-and-Mouth Disease Virus Serotype A Vaccine Matching Tesfaalem Tekleghiorghis, a,c Klaas Weerdmeester, a Froukje van Hemert-Kluitenberg, a Rob J. M. Moormann, a,b
Comparison of Test Methodologies for Foot-and-Mouth Disease Virus Serotype A Vaccine Matching Tesfaalem Tekleghiorghis, a,c Klaas Weerdmeester, a Froukje van Hemert-Kluitenberg, a Rob J. M. Moormann, a,b
Appendix 30. Preliminary results to evaluate cross-protection between O 1 Manisa and O 1 Campos in cattle
 Appendix 30 Preliminary results to evaluate cross-protection between O 1 Manisa and O 1 Campos in cattle V.A. Srinivasan 1, S.B.Nagendra Kumar 1, M.Madhan Mohan 1, V.Maroudam 1, P.Santha Kumar 1, S. Parida
Appendix 30 Preliminary results to evaluate cross-protection between O 1 Manisa and O 1 Campos in cattle V.A. Srinivasan 1, S.B.Nagendra Kumar 1, M.Madhan Mohan 1, V.Maroudam 1, P.Santha Kumar 1, S. Parida
A solid-phase competition ELISA for measuring antibody to foot-and-mouth disease virus
 197 Appendix 24 A solid-phase competition ELISA for measuring antibody to foot-and-mouth disease virus N.P. Ferris a, A.N. Bulut b, T. Rendle a, F. Davidson a and D.K.J. Mackay c a b c Institute for Animal
197 Appendix 24 A solid-phase competition ELISA for measuring antibody to foot-and-mouth disease virus N.P. Ferris a, A.N. Bulut b, T. Rendle a, F. Davidson a and D.K.J. Mackay c a b c Institute for Animal
COMPARISON OF DIFFERENT ELISA METHODS FOR THE DETECTION OF ANTIBODIES AGAINST FOOT-AND-MOUTH DISEASE VIRUS (FMDV) TYPE O
 TYPE O") Bull. Vet. Inst. Pulawy 48, 5-9, 24 COMPARISON OF DIFFERENT ELISA METHODS FOR THE DETECTION OF ANTIBODIES AGAINST FOOT-AND-MOUTH DISEASE VIRUS (FMDV) TYPE O WIESŁAW NIEDBALSKI Department of Foot-and-Mouth
Bull. Vet. Inst. Pulawy 48, 5-9, 24 COMPARISON OF DIFFERENT ELISA METHODS FOR THE DETECTION OF ANTIBODIES AGAINST FOOT-AND-MOUTH DISEASE VIRUS (FMDV) TYPE O WIESŁAW NIEDBALSKI Department of Foot-and-Mouth
Cedivac-FMD; Duration of Immunity in cattle, sheep and pigs. 2004, 8203 AA Lelystad, The Netherlands * Corresponding Author
 Appendix 3 Cedivac-FMD; Duration of Immunity in cattle, sheep and pigs. Paulus Selman *, Gilles Chénard and Aldo Dekker Animal Sciences Group, Wageningen UR, P.O. Box 65, 8 AB Lelystad, The Netherlands
Appendix 3 Cedivac-FMD; Duration of Immunity in cattle, sheep and pigs. Paulus Selman *, Gilles Chénard and Aldo Dekker Animal Sciences Group, Wageningen UR, P.O. Box 65, 8 AB Lelystad, The Netherlands
FAO Collaborative Study Phase XVII: Standardisation of FMD Antibody Detection
 Appendix 28 FAO Collaborative Study Phase XVII: Standardisation of FMD Antibody Detection D J Paton, R M Armstrong, L S Turner, P A Hamblin, M Corteyn, D Gibson, J Anderson Institute for Animal Health,
Appendix 28 FAO Collaborative Study Phase XVII: Standardisation of FMD Antibody Detection D J Paton, R M Armstrong, L S Turner, P A Hamblin, M Corteyn, D Gibson, J Anderson Institute for Animal Health,
Longevity of the antibody response in pigs and sheep following a single administration of high potency emergency FMD vaccines
 247 Appendix 31 Longevity of the antibody response in pigs and sheep following a single administration of high potency emergency FMD vaccines S. J Cox and P. V. Barnett Institute for Animal Health, Pirbright
247 Appendix 31 Longevity of the antibody response in pigs and sheep following a single administration of high potency emergency FMD vaccines S. J Cox and P. V. Barnett Institute for Animal Health, Pirbright
Lelystad, The Netherlands
 Appendix 4 Emergency vaccination against FMD in pigs: influence of used vaccine dose and correlation between induced immunological responses and reduction of virus shedding P.L. Eblé 1 *, A. Bouma 2, K.
Appendix 4 Emergency vaccination against FMD in pigs: influence of used vaccine dose and correlation between induced immunological responses and reduction of virus shedding P.L. Eblé 1 *, A. Bouma 2, K.
High potency vaccines induce protection against heterologous challenge with FMD virus
 High potency vaccines induce protection against heterologous challenge with FMD virus Katharina Brehm, Naveen Kumar, Hans-Herman Thulke and Bernd Haas Principle of current production method for FMD vaccines
High potency vaccines induce protection against heterologous challenge with FMD virus Katharina Brehm, Naveen Kumar, Hans-Herman Thulke and Bernd Haas Principle of current production method for FMD vaccines
Vaccine matching. Reliability of foot-and-mouth disease r-values. determination
 Vaccine matching Reliability of foot-and-mouth disease r-values determination Interinstitutional Network for R&D in FMD (RIIDFA)- Argentina Veterinary and Agrochemical Research Centre (VAR) - Belgium Protection
Vaccine matching Reliability of foot-and-mouth disease r-values determination Interinstitutional Network for R&D in FMD (RIIDFA)- Argentina Veterinary and Agrochemical Research Centre (VAR) - Belgium Protection
Key words: foot-and-mouth disease; single emulsion vaccine, saponin, immune response, emergency vaccines.
 255 Appendix 32 Enhancement of the immune response induced by the inclusion of saponin in oil adjuvanted vaccines against foot-and-mouth disease E. Smitsaart (1), N. Mattion (1) ; J.L. Filippi (1) ; B.
255 Appendix 32 Enhancement of the immune response induced by the inclusion of saponin in oil adjuvanted vaccines against foot-and-mouth disease E. Smitsaart (1), N. Mattion (1) ; J.L. Filippi (1) ; B.
Appendix 72 Using NSP ELISA (Chekit-FMD-3ABC Bommeli-Intervet) as a Tool for FMDV Serosurveillance in Bulgaria Abstract: Introduction
 as a Tool for FMDV Serosurveillance in Bulgaria Abstract: Introduction") Appendix 72 Using NSP ELISA (Chekit-FMD-3ABC Bommeli-Intervet) as a Tool for FMDV Serosurveillance in Bulgaria Georgi Georgiev*¹, Emiliya Veleva¹, Liliyana Polihronova¹ and Alessandro Rossi² 1 National
Appendix 72 Using NSP ELISA (Chekit-FMD-3ABC Bommeli-Intervet) as a Tool for FMDV Serosurveillance in Bulgaria Georgi Georgiev*¹, Emiliya Veleva¹, Liliyana Polihronova¹ and Alessandro Rossi² 1 National
Regulation of FMD vaccines within the European Union
 Introduction Regulation of FMD vaccines within the European Union K De Clercq 1 and D K J Mackay 2 Appendix 36 The EUFMD European Pharmacopoeia Working Group made a proposal for revision of the FMD vaccine
Introduction Regulation of FMD vaccines within the European Union K De Clercq 1 and D K J Mackay 2 Appendix 36 The EUFMD European Pharmacopoeia Working Group made a proposal for revision of the FMD vaccine
Received 19 September 2009/Returned for modification 13 October 2009/Accepted 27 October 2009
 CLINICAL AND VACCINE IMMUNOLOGY, Jan. 2010, p. 194 198 Vol. 17, No. 1 1556-6811/10/$12.00 doi:10.1128/cvi.00374-09 Copyright 2010, American Society for Microbiology. All Rights Reserved. Use of a Baculovirus-Expressed
CLINICAL AND VACCINE IMMUNOLOGY, Jan. 2010, p. 194 198 Vol. 17, No. 1 1556-6811/10/$12.00 doi:10.1128/cvi.00374-09 Copyright 2010, American Society for Microbiology. All Rights Reserved. Use of a Baculovirus-Expressed
Materials and Methods
 Appendix 18 Repeated administration of maximum payload emergency vaccines made from inactivated purified antigen concentrates do not induce significant titres of antibodies against non-structural proteins
Appendix 18 Repeated administration of maximum payload emergency vaccines made from inactivated purified antigen concentrates do not induce significant titres of antibodies against non-structural proteins
Engineering Foot-and-Mouth Disease Virus with Improved Properties for the Development of Effective Vaccine Introduction: Materials and methods:
 Engineering Foot-and-Mouth Disease Virus with Improved Properties for the Development of Effective Vaccine Haixue Zheng, Fan Yang, Ye Jin, Jianhong Guo, Jijun He, Kaiqi Lian, Zixiang Zhu, Weijun Cao, Lvlv,
Engineering Foot-and-Mouth Disease Virus with Improved Properties for the Development of Effective Vaccine Haixue Zheng, Fan Yang, Ye Jin, Jianhong Guo, Jijun He, Kaiqi Lian, Zixiang Zhu, Weijun Cao, Lvlv,
Appendix 71 Secretory IgA as an indicator of oropharyngeal FMDV replication Abstract Introduction Materials and methods
 Appendix 71 Secretory IgA as an indicator of oropharyngeal FMDV replication Satya Parida, David Paton*, Sarah Cox, Paul Barnett, John Anderson Pirbright Laboratory, Ititute for Animal Health, Ash Road,
Appendix 71 Secretory IgA as an indicator of oropharyngeal FMDV replication Satya Parida, David Paton*, Sarah Cox, Paul Barnett, John Anderson Pirbright Laboratory, Ititute for Animal Health, Ash Road,
Overview of WRL FMD. Historical perspective. Principal activities. FMD threats. Needs/prospects. David Paton
 Tracking the emergence and global spread of foot-and-mouth disease (FMD) Overview of WRL FMD David Paton Historical perspective Principal activities FMD threats Needs/prospects Historical Perspective FMD
Tracking the emergence and global spread of foot-and-mouth disease (FMD) Overview of WRL FMD David Paton Historical perspective Principal activities FMD threats Needs/prospects Historical Perspective FMD
FMD Vaccine Strain Selection
 FMD Vaccine Strain Selection David Paton, Pirbright, UK New Delhi, 13-15 February 2012 Conclusions Vaccine match is one component of vaccine efficacy Vaccine quality may compensate for imperfect match
FMD Vaccine Strain Selection David Paton, Pirbright, UK New Delhi, 13-15 February 2012 Conclusions Vaccine match is one component of vaccine efficacy Vaccine quality may compensate for imperfect match
COMPARISON OF THREE ELISA KITS FOR THE DETECTION OF ANTIBODIES AGAINST FOOT-AND-MOUTH DISEASE VIRUS NON-STRUCTURAL PROTEINS
 Bull Vet Inst Pulawy 49, 147-151 COMPARISON OF THREE ELISA KITS FOR THE DETECTION OF ANTIBODIES AGAINST FOOT-AND-MOUTH DISEASE VIRUS NON-STRUCTURAL PROTEINS WIESŁAW NIEDBALSKI Department of Foot-and-Mouth
Bull Vet Inst Pulawy 49, 147-151 COMPARISON OF THREE ELISA KITS FOR THE DETECTION OF ANTIBODIES AGAINST FOOT-AND-MOUTH DISEASE VIRUS NON-STRUCTURAL PROTEINS WIESŁAW NIEDBALSKI Department of Foot-and-Mouth
Richard Reeve Boyd Orr Centre for Population and Ecosystem Health University of Glasgow
 Advances and gaps in vaccine modelling Richard Reeve Boyd Orr Centre for Population and Ecosystem Health University of Glasgow 1 Should I vaccinate? In disease-free countries, the decision may be difficult
Advances and gaps in vaccine modelling Richard Reeve Boyd Orr Centre for Population and Ecosystem Health University of Glasgow 1 Should I vaccinate? In disease-free countries, the decision may be difficult
1. Engineering Foot-and-Mouth Disease Viruses with Improved
 Engineering Foot-and-Mouth Disease Virus with Improved Properties for the Development of Effective Vaccine Haixue Zheng, Fan Yang, Ye Jin, Jianhong Guo, Jijun He, Lvlv, Xuepeng Cai, Xiangtao Liu, Hong
Engineering Foot-and-Mouth Disease Virus with Improved Properties for the Development of Effective Vaccine Haixue Zheng, Fan Yang, Ye Jin, Jianhong Guo, Jijun He, Lvlv, Xuepeng Cai, Xiangtao Liu, Hong
Formaldehyde enhances BEI-inactivation rates of Foot-and- Mouth Disease (FMD) virus by at least a ten-fold
 virus by at least a ten-fold") 27 Formaldehyde enhances BEI-inactivation rates of Foot-and- Mouth Disease (FMD) virus by at least a ten-fold Appendix 34 Simon J. Barteling and Nazeem I. Cassim, Onderstepoort Veterinary Institute, Private
27 Formaldehyde enhances BEI-inactivation rates of Foot-and- Mouth Disease (FMD) virus by at least a ten-fold Appendix 34 Simon J. Barteling and Nazeem I. Cassim, Onderstepoort Veterinary Institute, Private
Comparative Studies of the Potency of Foot and Mouth Disease Virus Trivalent Vaccine with Different Concentration of the Antigenic Content (146S).
.") Alexandria Journal of Veterinary Sciences 2015, 45: 33-42 ISSN 1110-2047, www.alexjvs.com DOI: 10.5455/ajvs.177834 Comparative Studies of the Potency of Foot and Mouth Disease Virus Trivalent Vaccine with
Alexandria Journal of Veterinary Sciences 2015, 45: 33-42 ISSN 1110-2047, www.alexjvs.com DOI: 10.5455/ajvs.177834 Comparative Studies of the Potency of Foot and Mouth Disease Virus Trivalent Vaccine with
FMD Carrier state and role of carrier buffalo as source of transboundary spread in Southeast Asia and Eastern Asia Satya Parida
 FMD Carrier state and role of carrier buffalo as source of transboundary spread in Southeast Asia and Eastern Asia Satya Parida, Pirbright, UK Foot-and-mouth disease Highly contagious disease of all the
FMD Carrier state and role of carrier buffalo as source of transboundary spread in Southeast Asia and Eastern Asia Satya Parida, Pirbright, UK Foot-and-mouth disease Highly contagious disease of all the
Manufacturers expected contribution to the progressive control of Foot-and-Mouth Disease in South Asia
 Manufacturers expected contribution to the progressive control of Foot-and-Mouth Disease in South Asia Ph. Dubourget & al. 1 GENERAL PRINCIPLES 2 EPIDEMIO- LOGICALLY RELEVANT STRAINS 3 IMMUNO- DOMINANT
Manufacturers expected contribution to the progressive control of Foot-and-Mouth Disease in South Asia Ph. Dubourget & al. 1 GENERAL PRINCIPLES 2 EPIDEMIO- LOGICALLY RELEVANT STRAINS 3 IMMUNO- DOMINANT
The inhibition of FMD virus excretion from the infected pigs by an antiviral agent, T-1105
 The inhibition of FMD virus excretion from the infected pigs by an antiviral agent, T-1105 Appendix 64 Kenichi Sakamoto 1*, Seiichi Ohashi 1, Reiko Yamazoe 1, Kazumi Takahashi 2, and Yousuke Furuta 2 1
The inhibition of FMD virus excretion from the infected pigs by an antiviral agent, T-1105 Appendix 64 Kenichi Sakamoto 1*, Seiichi Ohashi 1, Reiko Yamazoe 1, Kazumi Takahashi 2, and Yousuke Furuta 2 1
Using chloroform as a preservative for trivalent foot and mouth disease vaccine in comparison to thiomersal
 ORIGINAL ARTICLE Using chloroform as a preservative for trivalent foot and mouth disease vaccine in comparison to thiomersal Assem A Mohamed, Wael Mosad, Mohamed A Gamil, Ehbal M Farouk, Amr I Hassanin,
ORIGINAL ARTICLE Using chloroform as a preservative for trivalent foot and mouth disease vaccine in comparison to thiomersal Assem A Mohamed, Wael Mosad, Mohamed A Gamil, Ehbal M Farouk, Amr I Hassanin,
Predicting antigenic sites on the FMDV. F.F. Maree, R. Reeve, B. Blignaut, J.J. Esterhuysen, E. Fry, T. de Beer, E. Rieder and D.
 Predicting antigenic sites on the FMDV capsid from cross-reactivity reactivity data F.F. Maree, R. Reeve, B. Blignaut, J.J. Esterhuysen, E. Fry, T. de Beer, E. Rieder and D.Haydon Introduction The SAT
Predicting antigenic sites on the FMDV capsid from cross-reactivity reactivity data F.F. Maree, R. Reeve, B. Blignaut, J.J. Esterhuysen, E. Fry, T. de Beer, E. Rieder and D.Haydon Introduction The SAT
The Global control of FMD - Tools, ideas and ideals Erice, Italy October 2008 THE STATUS OF FOOT-AND-MOUTH DISEASE (FMD) IN ETHIOPIA
 IN ETHIOPIA") The Global control of FMD - Tools, ideas and ideals Erice, Italy 14-17 October 2008 Appendix 60 THE STATUS OF FOOT-AND-MOUTH DISEASE (FMD) IN ETHIOPIA G. Ayelet*, E. Gelaye, J. Guitian, M. Sahle, N. J.
The Global control of FMD - Tools, ideas and ideals Erice, Italy 14-17 October 2008 Appendix 60 THE STATUS OF FOOT-AND-MOUTH DISEASE (FMD) IN ETHIOPIA G. Ayelet*, E. Gelaye, J. Guitian, M. Sahle, N. J.
Appendix 53 Addition of saponin to double oil emulsion FMD vaccines enhances specific antibody responses in cattle and pigs Introduction:
 Appendix 53 Addition of saponin to double oil emulsion FMD vaccines enhances specific antibody responses in cattle and pigs Smitsaart E* 1, Espinoza AM 1, Sanguinetti R 2, Filippi J 1, Ham A 1, Bellinzoni
Appendix 53 Addition of saponin to double oil emulsion FMD vaccines enhances specific antibody responses in cattle and pigs Smitsaart E* 1, Espinoza AM 1, Sanguinetti R 2, Filippi J 1, Ham A 1, Bellinzoni
Testing the Antibody Response of Pigs to Foot-and- Mouth Disease Vaccines
 Testing the Antibody Response of Pigs to Foot-and- Mouth Disease Vaccines Final Report APL Project 211/139.45 December 211 Australian Animal Health Laboratory Dr Wilna Vosloo Private Bag 24 Geelong Vic
Testing the Antibody Response of Pigs to Foot-and- Mouth Disease Vaccines Final Report APL Project 211/139.45 December 211 Australian Animal Health Laboratory Dr Wilna Vosloo Private Bag 24 Geelong Vic
FMD in Libya. Dr. Abdunaser Dayhum National Center of Animal Health Libya
 FMD in Libya Dr. Abdunaser Dayhum National Center of Animal Health Libya Foot-and-Mouth Disease Cause: Foot-and-mouth disease virus (FMDV). A total of seven different serologic types are recognized: A,
FMD in Libya Dr. Abdunaser Dayhum National Center of Animal Health Libya Foot-and-Mouth Disease Cause: Foot-and-mouth disease virus (FMDV). A total of seven different serologic types are recognized: A,
Chapter 6. Foot and mouth disease virus transmission during the incubation period of the disease in piglets, lambs, calves, and dairy cows
 Chapter 6 Foot and mouth disease virus transmission during the incubation period of the disease in piglets, lambs, calves, and dairy cows K. Orsel* 1, A. Bouma 1, A. Dekker 2, J.A. Stegeman 1 and M.C.M.
Chapter 6 Foot and mouth disease virus transmission during the incubation period of the disease in piglets, lambs, calves, and dairy cows K. Orsel* 1, A. Bouma 1, A. Dekker 2, J.A. Stegeman 1 and M.C.M.
Importance of cell mediated immunity for protection against Foot and Mouth Disease
 Appendix 34 Importance of cell mediated immunity for protection against Foot and Mouth Disease Yooni Oh 1*, ryan Charleston 1, David J. Paton 1, Jong-Hyun Park 2, Paul V. arnett 1, Yi-Seok Joo 2, Satya
Appendix 34 Importance of cell mediated immunity for protection against Foot and Mouth Disease Yooni Oh 1*, ryan Charleston 1, David J. Paton 1, Jong-Hyun Park 2, Paul V. arnett 1, Yi-Seok Joo 2, Satya
Simultaneous immunization of cattle with FMD and live anthrax vaccines
 Simultaneous immunization of cattle with FMD and live anthrax vaccines Alejandra Capozzo Head of the Applied Veterinary Immunology Lab. Institute of Virology, Buenos Aires. Argentina In collaboration with:
Simultaneous immunization of cattle with FMD and live anthrax vaccines Alejandra Capozzo Head of the Applied Veterinary Immunology Lab. Institute of Virology, Buenos Aires. Argentina In collaboration with:
Chapter 5b. RT-PCR tests on tonsils of foot and mouth disease virus infected piglets at least four weeks after initial challenge
 Chapter 5b RT-PCR tests on tonsils of foot and mouth disease virus infected piglets at least four weeks after initial challenge K. Orsel 1 *, H.I.J. Roest 2, E. Elzinga-van der Linde 2, F. van Hemert-Kluitenberg
Chapter 5b RT-PCR tests on tonsils of foot and mouth disease virus infected piglets at least four weeks after initial challenge K. Orsel 1 *, H.I.J. Roest 2, E. Elzinga-van der Linde 2, F. van Hemert-Kluitenberg
The work of the WRLFMD in relation to regional networks. Reference Laboratory for FMDV
 The work of the WRLFMD in relation to regional networks Reference Laboratory for FMDV WRLFMD - GOODBYE TO: Nigel Ferris January 2012 Geoff Hutchings July 2012 Claudia Doel March 2013 Miki Madi June 2013
The work of the WRLFMD in relation to regional networks Reference Laboratory for FMDV WRLFMD - GOODBYE TO: Nigel Ferris January 2012 Geoff Hutchings July 2012 Claudia Doel March 2013 Miki Madi June 2013
The global control of FMD; challenges and opportunities
 EuFMD-Erice 2008 The global control of FMD; challenges and opportunities Keith Sumption, Secretary, European Commission for the Control of Foot-and-Mouth Disease (EuFMD), Food-and-Agriculture Organization
EuFMD-Erice 2008 The global control of FMD; challenges and opportunities Keith Sumption, Secretary, European Commission for the Control of Foot-and-Mouth Disease (EuFMD), Food-and-Agriculture Organization
FMD in Southern Africa
 Molecular Biological Characteristics of Foot-and- Mouth Disease Virus in the African Buffaloes in Southern Africa Christopher Kasanga 1, Rahana Dwarka 2, Gaothlele Thobokwe 3, Jemma Wadsworth 4, Nick Knowles
Molecular Biological Characteristics of Foot-and- Mouth Disease Virus in the African Buffaloes in Southern Africa Christopher Kasanga 1, Rahana Dwarka 2, Gaothlele Thobokwe 3, Jemma Wadsworth 4, Nick Knowles
Development of a peptide based latex agglutination assay for serotype identification of foot and mouth disease virus
 Indian Journal of Experimental Biology Vol. 51, February 2013, pp. 124-128 Development of a peptide based latex agglutination assay for serotype identification of foot and mouth disease virus Dilpreet
Indian Journal of Experimental Biology Vol. 51, February 2013, pp. 124-128 Development of a peptide based latex agglutination assay for serotype identification of foot and mouth disease virus Dilpreet
Comparison of protective Antibody response in guinea pigs immunized with Wild type (Wt) BoHV-1 and ΔgE BoHV-1 mutant virus vaccine
 BoHV-1 and ΔgE BoHV-1 mutant virus vaccine") Comparison of protective Antibody response in guinea pigs immunized with Wild type (Wt) BoHV-1 and ΔgE BoHV-1 mutant virus vaccine L.R.ANANTHAKRISHNA 1*, MOHINI SAINI 2, PRAVEEN K GUPTA 3 and RIZWANA TABASSUM
Comparison of protective Antibody response in guinea pigs immunized with Wild type (Wt) BoHV-1 and ΔgE BoHV-1 mutant virus vaccine L.R.ANANTHAKRISHNA 1*, MOHINI SAINI 2, PRAVEEN K GUPTA 3 and RIZWANA TABASSUM
Foot and Mouth Disease vaccination is effective: Quantification of FMD transmission with and without vaccination
 Foot and Mouth Disease vaccination is effective: Quantification of FMD transmission with and without vaccination Aldo Dekker Acknowledgments Phaedra Eblé, Karin Orsel, Carla Bravo de Rueda Annemarie Bouma,
Foot and Mouth Disease vaccination is effective: Quantification of FMD transmission with and without vaccination Aldo Dekker Acknowledgments Phaedra Eblé, Karin Orsel, Carla Bravo de Rueda Annemarie Bouma,
Introduction. Ahmadi-Vasmehjani A 1*, Mousavi-Nasab SD 2, Baharlou R 1, Jeirani F 3, Shayestehpour M 4, Yaghoubi S 5, Fazel H 4, Mahravani H 3
 Iranian Journal of Virology 2013;7(1&2): 21-29 2013, Iranian Society of Virology Original Article Diagnosis of Foot-and-Mouth Disease Virus by Real Time Reverse Transcription Polymerase Chain Reaction
Iranian Journal of Virology 2013;7(1&2): 21-29 2013, Iranian Society of Virology Original Article Diagnosis of Foot-and-Mouth Disease Virus by Real Time Reverse Transcription Polymerase Chain Reaction
G. W. WOOD J. C. MUSKETT and D. H. THORNTON MAFF, Central Veterinary Laboratory, New Haw, Weybridge, Surrey, U.K.
 J. Comp. Path. 1986 vol. 96 OBSERVATIONS ON THE ABILITY OF AVIAN REOVIRUS VACCINMATION OF HENS TO PROTECT THEIR PROGENY AGAINST THE EFFECTS OF CHALLENGE WITH HOMOLOGOUS AND HETEROLOGOUS STRAINS By G. W.
J. Comp. Path. 1986 vol. 96 OBSERVATIONS ON THE ABILITY OF AVIAN REOVIRUS VACCINMATION OF HENS TO PROTECT THEIR PROGENY AGAINST THE EFFECTS OF CHALLENGE WITH HOMOLOGOUS AND HETEROLOGOUS STRAINS By G. W.
Development of a predictive model for vaccine matching for serotype O FMDV from serology and capsid sequence
 Development of a predictive model for vaccine matching for serotype O FMDV from serology and capsid sequence D. Borley, S. Upadhyaya, D. Paton, R. Reeve and Mana Mahapatra Pirbright Laboratory United Kingdom
Development of a predictive model for vaccine matching for serotype O FMDV from serology and capsid sequence D. Borley, S. Upadhyaya, D. Paton, R. Reeve and Mana Mahapatra Pirbright Laboratory United Kingdom
Foot and mouth disease situation and control strategies in the People s Republic of China the current situation
 Foot and mouth disease situation and control strategies in the People s Republic of China the current situation Lu Zengjun (Associate researcher) Shang Youjun (Associate researcher) National Foot-and-Mouth
Foot and mouth disease situation and control strategies in the People s Republic of China the current situation Lu Zengjun (Associate researcher) Shang Youjun (Associate researcher) National Foot-and-Mouth
Verification of Efficacy of Donated FMD O Type Vaccines in Laos
 OIE Collaborating Centre Diagnosis and Control of Animal Diseases and Related Veterinary Product Assessment in Asia Activities of OIE/JFT Project in Laos September 25, 2014 Verification of Efficacy of
OIE Collaborating Centre Diagnosis and Control of Animal Diseases and Related Veterinary Product Assessment in Asia Activities of OIE/JFT Project in Laos September 25, 2014 Verification of Efficacy of
Longevity of protection in cattle following vaccination with emergency FMD vaccines from the UK strategic reserve
 Longevity of protection in cattle following vaccination with emergency FMD vaccines from the UK strategic reserve Sarah Cox, Satya Parida, Pip Hamblin, Bartek Bankowski Veronica Carr, David Paton and Paul
Longevity of protection in cattle following vaccination with emergency FMD vaccines from the UK strategic reserve Sarah Cox, Satya Parida, Pip Hamblin, Bartek Bankowski Veronica Carr, David Paton and Paul
Foot and Mouth Disease Middle East situation Summary of Answers to the Questionnaire Beirut, Lebanon, 7 9 April 2009
 Foot and Mouth Disease Middle East situation Summary of Answers to the Questionnaire Beirut, Lebanon, 7 9 April 2009 Dr Pierre Primot OIE for the Middle East Purpose of the Questionnaire The purpose of
Foot and Mouth Disease Middle East situation Summary of Answers to the Questionnaire Beirut, Lebanon, 7 9 April 2009 Dr Pierre Primot OIE for the Middle East Purpose of the Questionnaire The purpose of
IMMUNOGENICITY OF FORMALDYDE INACTIVATED NEWCASTLE DISEASE VIRUS FIELD ISOLATE IN MATERNAL ANTIBODY FREE CHICKENS
 IMMUNOGENICITY OF FORMALDYDE INACTIVATED NEWCASTLE DISEASE VIRUS FIELD ISOLATE IN MATERNAL ANTIBODY FREE CHICKENS Anak Agung Ayu Mirah Adi 1 *, IGusti Agung Arta Putra 2, Nyoman Mantik Astawa 3, I Made
IMMUNOGENICITY OF FORMALDYDE INACTIVATED NEWCASTLE DISEASE VIRUS FIELD ISOLATE IN MATERNAL ANTIBODY FREE CHICKENS Anak Agung Ayu Mirah Adi 1 *, IGusti Agung Arta Putra 2, Nyoman Mantik Astawa 3, I Made
FMD vaccination and postvaccination. guidelines. Samia Metwally (FAO) Animal Production and Health Division FAO of the United Nations Rome, Italy
 Animal Production and Health Division FAO of the United Nations Rome, Italy") 4th Middle East FMD Roadmap Meeting Amman, Jordan 15-19 October 2017 FMD vaccination and postvaccination monitoring guidelines Samia Metwally (FAO) Animal Production and Health Division FAO of the United
4th Middle East FMD Roadmap Meeting Amman, Jordan 15-19 October 2017 FMD vaccination and postvaccination monitoring guidelines Samia Metwally (FAO) Animal Production and Health Division FAO of the United
Shing Faculty of Medicine, The University of Hong Kong, 21 Sassoon Road, Pokfulam, Hong Kong Special Administrative Region, China
 CVI Accepts, published online ahead of print on 4 January 2012 Clin. Vaccine Immunol. doi:10.1128/cvi.05358-11 Copyright 2012, American Society for Microbiology. All Rights Reserved. 1 2 Evaluation of
CVI Accepts, published online ahead of print on 4 January 2012 Clin. Vaccine Immunol. doi:10.1128/cvi.05358-11 Copyright 2012, American Society for Microbiology. All Rights Reserved. 1 2 Evaluation of
Mechanism of FMD spread in East Asia
 Mechanism of FMD spread in East Asia China has the important role of the FMD outbreaks in East Asia 1) Long border with FMD epidemic countries in South East and Central Asia 2) Large numbers of susceptible
Mechanism of FMD spread in East Asia China has the important role of the FMD outbreaks in East Asia 1) Long border with FMD epidemic countries in South East and Central Asia 2) Large numbers of susceptible
Foot and mouth disease control strategies in North Africa and the Middle East the current
 1. Title, names and addresses of authors Foot and mouth disease control strategies in North Africa and the Middle East the current situation G. Yehia (1), P. Primot (1) (1) OIE Regional Representation
1. Title, names and addresses of authors Foot and mouth disease control strategies in North Africa and the Middle East the current situation G. Yehia (1), P. Primot (1) (1) OIE Regional Representation
HIV-1 p24 ELISA Pair Set Cat#: orb54951 (ELISA Manual)
") HIV-1 p24 ELISA Pair Set Cat#: orb54951 (ELISA Manual) BACKGROUND Human Immunodeficiency Virus ( HIV ) can be divided into two major types, HIV type 1 (HIV-1) and HIV type 2 (HIV-2). HIV-1 is related to
HIV-1 p24 ELISA Pair Set Cat#: orb54951 (ELISA Manual) BACKGROUND Human Immunodeficiency Virus ( HIV ) can be divided into two major types, HIV type 1 (HIV-1) and HIV type 2 (HIV-2). HIV-1 is related to
DIFFERENTIATION OF INFECTION FROM VACCINATION BY DETECTION OF ANTIBODIES TO THE NON-STRUCTURAL PROTEIN 3ABC OF FOOT-AND-MOUTH DISEASE VIRUS
 Bull. Vet. Inst. Pulawy 7, 5-6, DIFFERENTIATION OF INFECTION FROM VACCINATION BY DETECTION OF ANTIBODIES TO THE NON-STRUCTURAL PROTEIN ABC OF FOOT-AND-MOUTH DISEASE VIRUS WIESŁAW NIEDBALSKI AND BERND HAAS*
Bull. Vet. Inst. Pulawy 7, 5-6, DIFFERENTIATION OF INFECTION FROM VACCINATION BY DETECTION OF ANTIBODIES TO THE NON-STRUCTURAL PROTEIN ABC OF FOOT-AND-MOUTH DISEASE VIRUS WIESŁAW NIEDBALSKI AND BERND HAAS*
Post-Vaccination Monitoring
 Post-Vaccination Monitoring Samia Metwally, DVM, PhD Senior Animal Health Officer (Virologist) Animal Production and Health Division FAO of UN, Rome, Italy samia.metwally@fao.org Overview Why post-vaccination
Post-Vaccination Monitoring Samia Metwally, DVM, PhD Senior Animal Health Officer (Virologist) Animal Production and Health Division FAO of UN, Rome, Italy samia.metwally@fao.org Overview Why post-vaccination
Appendix 68 3 ABC ELISA for the diagnosis of FMD in Egyptian sheep Abstract Introduction
 Appendix 68 3 ABC ELISA for the diagnosis of FMD in Egyptian sheep Laila E. El-Shehawy, Samira El-Kilany & A.M. Doaud Foot and Mouth Disease Department, Veterinary Serum and Vaccine Research Institute,
Appendix 68 3 ABC ELISA for the diagnosis of FMD in Egyptian sheep Laila E. El-Shehawy, Samira El-Kilany & A.M. Doaud Foot and Mouth Disease Department, Veterinary Serum and Vaccine Research Institute,
EPIDEMIOLOGICAL DEVELOPMENTS AND CONTROL OF FOOT AND MOUTH DISEASE IN ASIA
 EPIDEMIOLOGICAL DEVELOPMENTS AND CONTROL OF FOOT AND MOUTH DISEASE IN ASIA Kenichi Sakamoto National Institute of Animal Health, Japan 1 Original: English Summary: In the Asia region, foot and mouth disease
EPIDEMIOLOGICAL DEVELOPMENTS AND CONTROL OF FOOT AND MOUTH DISEASE IN ASIA Kenichi Sakamoto National Institute of Animal Health, Japan 1 Original: English Summary: In the Asia region, foot and mouth disease
WRLFMD: ~1000 samples from 36 countries January- December 2009 (300% up on 2008)
") Appendix 9 IAH Pirbright FMD Reference Laboratory Report: 81st SESSION OF THE EXECUTIVE COMMITTEE OF THE EuFMD COMMISSION 2nd February 2011 Jef M. Hammond, Donald P. King, Nick J. Knowles, Jemma Wadsworth,
Appendix 9 IAH Pirbright FMD Reference Laboratory Report: 81st SESSION OF THE EXECUTIVE COMMITTEE OF THE EuFMD COMMISSION 2nd February 2011 Jef M. Hammond, Donald P. King, Nick J. Knowles, Jemma Wadsworth,
Clinell Universal Wipes Batch number Client
 Test Report: EN 14476 2013 Chemical disinfectants and antiseptics - Virucidal quantitative suspension test for chemical disinfectants and antiseptics used in human medicine - Test method and requirements
Test Report: EN 14476 2013 Chemical disinfectants and antiseptics - Virucidal quantitative suspension test for chemical disinfectants and antiseptics used in human medicine - Test method and requirements
FOOT AND MOUTH DISEASE DIAGNOSTICS: REQUIREMENTS FOR DEMONSTRATION OF FREEDOM FROM INFECTION
 70 SG/12/CS3 C Original: English FOOT AND MOUTH DISEASE DIAGNOSTICS: REQUIREMENTS FOR DEMONSTRATION OF FREEDOM FROM INFECTION R.P. Kitching Director, Canadian Food Inspection Agency, National Centre for
70 SG/12/CS3 C Original: English FOOT AND MOUTH DISEASE DIAGNOSTICS: REQUIREMENTS FOR DEMONSTRATION OF FREEDOM FROM INFECTION R.P. Kitching Director, Canadian Food Inspection Agency, National Centre for
Results of Seromonitoring in FMD Buffer Zone in the CIS Countries
 Appendix Results of Seromonitoring in FMD Buffer Zone in the CIS Countries T.A.Fomina, V.M.Zakharov, A.A.Gusev, N.E.Kamalova, S.R.Kremenchugskaya, O.A.Schekotova All-Russian Research Institute for Animal
Appendix Results of Seromonitoring in FMD Buffer Zone in the CIS Countries T.A.Fomina, V.M.Zakharov, A.A.Gusev, N.E.Kamalova, S.R.Kremenchugskaya, O.A.Schekotova All-Russian Research Institute for Animal
Foot and Mouth Disease Vaccine Research and Development in India
 Foot and Mouth Disease Vaccine Research and Development in India R.Venkataramanan Indian Veterinary Research Institute, Hebbal, Bangalore 560 024 Foot and Mouth Disease in India Present Status Large Susceptible
Foot and Mouth Disease Vaccine Research and Development in India R.Venkataramanan Indian Veterinary Research Institute, Hebbal, Bangalore 560 024 Foot and Mouth Disease in India Present Status Large Susceptible
Toward the control of Foot-and-Mouth disease in East Asia
 Symposium on Prevention and Control of Foot and Mouth Disease and Highly Pathogenic Avian influenza in East Asia 20 September, 2017, Tokyo National Agriculture and Food Research Organization Toward the
Symposium on Prevention and Control of Foot and Mouth Disease and Highly Pathogenic Avian influenza in East Asia 20 September, 2017, Tokyo National Agriculture and Food Research Organization Toward the
Human Immunodeficiency Virus type 1 (HIV-1) p24 / Capsid Protein p24 ELISA Pair Set
 p24 / Capsid Protein p24 ELISA Pair Set") Human Immunodeficiency Virus type 1 (HIV-1) p24 / Capsid Protein p24 ELISA Pair Set Catalog Number : SEK11695 To achieve the best assay results, this manual must be read carefully before using this product
Human Immunodeficiency Virus type 1 (HIV-1) p24 / Capsid Protein p24 ELISA Pair Set Catalog Number : SEK11695 To achieve the best assay results, this manual must be read carefully before using this product
Fufa Dawo Bari, 1 Satya Parida, 1 Amin S. Asfor, 1 Daniel T. Haydon, 2 Richard Reeve, 1,2 David J. Paton 1 and Mana Mahapatra 1 INTRODUCTION
 Journal of General Virology (2015), 96, 1033 1041 DOI 10.1099/vir.0.000051 Prediction and characterization of novel epitopes of serotype A foot-and-mouth disease viruses circulating in East Africa using
Journal of General Virology (2015), 96, 1033 1041 DOI 10.1099/vir.0.000051 Prediction and characterization of novel epitopes of serotype A foot-and-mouth disease viruses circulating in East Africa using
Importance of foot and mouth disease vaccine purity in interpreting serological surveys
 Rev. Sci. Tech. Off. Int. Epiz., 2015, 34 (3),... -... Importance of foot and mouth disease vaccine purity in interpreting serological surveys This paper (No. 24092015-00060-ES) has been peer-reviewed,
Rev. Sci. Tech. Off. Int. Epiz., 2015, 34 (3),... -... Importance of foot and mouth disease vaccine purity in interpreting serological surveys This paper (No. 24092015-00060-ES) has been peer-reviewed,
Immune Response and Viral Persistence in Indian Buffaloes (Bubalus bubalis) Infected with Foot-and-Mouth Disease Virus Serotype Asia 1
 Infected with Foot-and-Mouth Disease Virus Serotype Asia 1") CLINICAL AND VACCINE IMMUNOLOGY, Dec. 2009, p. 1832 1836 Vol. 16, No. 12 1556-6811/09/$12.00 doi:10.1128/cvi.00302-09 Copyright 2009, American Society for Microbiology. All Rights Reserved. Immune Response
CLINICAL AND VACCINE IMMUNOLOGY, Dec. 2009, p. 1832 1836 Vol. 16, No. 12 1556-6811/09/$12.00 doi:10.1128/cvi.00302-09 Copyright 2009, American Society for Microbiology. All Rights Reserved. Immune Response
Foot-and Mouth Disease Ecological Studies In Endemic Settings: Ongoing Studies in Vietnam and Pakistan
 Foot-and Mouth Disease Ecological Studies In Endemic Settings: Ongoing Studies in Vietnam and Pakistan Luis Rodriguez (Jonathan Arzt) Research leader, USDA-ARS Plum Island 1 PAKISTAN CHARACTERIZATION OF
Foot-and Mouth Disease Ecological Studies In Endemic Settings: Ongoing Studies in Vietnam and Pakistan Luis Rodriguez (Jonathan Arzt) Research leader, USDA-ARS Plum Island 1 PAKISTAN CHARACTERIZATION OF
Studies of the Antigenic relationships between Bluetongue virus serotypes 2, 9 & 15 isolated in Andhra Pradesh, India
 Vet. World, 2011, Vol.4(10): 444-448 RESEARCH Studies of the Antigenic relationships between Bluetongue virus serotypes 2, 9 & 15 isolated in Andhra Pradesh, India 1 1 Deepthi Balam*, Sreenivasulu Daggupati,
Vet. World, 2011, Vol.4(10): 444-448 RESEARCH Studies of the Antigenic relationships between Bluetongue virus serotypes 2, 9 & 15 isolated in Andhra Pradesh, India 1 1 Deepthi Balam*, Sreenivasulu Daggupati,
Cross-Reactivity to Field Isolates of Canine Influenza Virus by a Killed Canine Influenza Virus (H3N8, Iowa05) Vaccine
 Vaccine") Cross-Reactivity to Field Isolates of Canine Influenza Virus by a Killed Canine Influenza Virus (H3N8, Iowa05) Vaccine Nancee Oien, B.S., M.S. a Sally Mattern, B.S a Jaime Brozowski, B.S., M.S. b Janet
Cross-Reactivity to Field Isolates of Canine Influenza Virus by a Killed Canine Influenza Virus (H3N8, Iowa05) Vaccine Nancee Oien, B.S., M.S. a Sally Mattern, B.S a Jaime Brozowski, B.S., M.S. b Janet
Experimental evaluation of foot-and-mouth disease vaccines for emergency use in ruminants and pigs: a review
 Vet. Res. (2009) 40:13 DOI: 10.1051/vetres:2008051 C INRA, EDP Sciences, 2009 www.vetres.org Review article Experimental evaluation of foot-and-mouth disease vaccines for emergency use in ruminants and
Vet. Res. (2009) 40:13 DOI: 10.1051/vetres:2008051 C INRA, EDP Sciences, 2009 www.vetres.org Review article Experimental evaluation of foot-and-mouth disease vaccines for emergency use in ruminants and
FMD VACCINE AND VACCINATION. Ahmad Al-Majali Dean, Faculty of Vet Medicine JUST Jordan
 FMD VACCINE AND VACCINATION Ahmad Al-Majali Dean, Faculty of Vet Medicine JUST Jordan General Considerations on FMD vaccination The currently used FMD vaccines are killed virus preparations that are pure,
FMD VACCINE AND VACCINATION Ahmad Al-Majali Dean, Faculty of Vet Medicine JUST Jordan General Considerations on FMD vaccination The currently used FMD vaccines are killed virus preparations that are pure,
Sequence-Based Prediction for Vaccine Strain Selection and Identification of Antigenic Variability in Foot-and- Mouth Disease Virus
 Sequence-Based Prediction for Vaccine Strain Selection and Identification of Antigenic Variability in Foot-and- Mouth Disease Virus Richard Reeve 1,2 *, Belinda Blignaut 3,4, Jan J. Esterhuysen 3, Pamela
Sequence-Based Prediction for Vaccine Strain Selection and Identification of Antigenic Variability in Foot-and- Mouth Disease Virus Richard Reeve 1,2 *, Belinda Blignaut 3,4, Jan J. Esterhuysen 3, Pamela
OIE Reference Laboratory Reports Activities
 OIE Reference Laboratory Reports Activities Activities in 2016 This report has been submitted : 2017-01-12 10:32:31 Name of disease (or topic) for which you are a designated OIE Reference Laboratory: Foot
OIE Reference Laboratory Reports Activities Activities in 2016 This report has been submitted : 2017-01-12 10:32:31 Name of disease (or topic) for which you are a designated OIE Reference Laboratory: Foot
Bordetella pertussis igg-pt elisa Kit
 Bordetella pertussis IgG-PT ELISA Kit Application Bordetella pertussis IgG-PT ELISA Kit is a quantitative test for the detection of IgG antibodies against pertussis toxin in human serum samples. Background
Bordetella pertussis IgG-PT ELISA Kit Application Bordetella pertussis IgG-PT ELISA Kit is a quantitative test for the detection of IgG antibodies against pertussis toxin in human serum samples. Background
CHAPTER 4 IMMUNOLOGICAL TECHNIQUES
 CHAPTER 4 IMMUNOLOGICAL TECHNIQUES Nitroblue Tetrazolium Chloride (NBT) Reduction test NBT reduction test was evaluated by employing the method described by Hudson and Hay,1989 based upon principle that
CHAPTER 4 IMMUNOLOGICAL TECHNIQUES Nitroblue Tetrazolium Chloride (NBT) Reduction test NBT reduction test was evaluated by employing the method described by Hudson and Hay,1989 based upon principle that
Requirements of the Terrestrial Code for FMD surveillance. Dr David Paton Dr Gideon Brückner
 Requirements of the Terrestrial Code for FMD surveillance Dr David Paton Dr Gideon Brückner 1 Surveillance Close observation Origin of the word: Early 19th century: from French, from sur- over + veiller
Requirements of the Terrestrial Code for FMD surveillance Dr David Paton Dr Gideon Brückner 1 Surveillance Close observation Origin of the word: Early 19th century: from French, from sur- over + veiller
Measles IgM ELISA Kit
 Measles IgM ELISA Kit Catalog Number KA2257 96 assays Version: 03 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Intended Use... 3 Background... 3 Principle of the Assay...
Measles IgM ELISA Kit Catalog Number KA2257 96 assays Version: 03 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Intended Use... 3 Background... 3 Principle of the Assay...
Foot and mouth disease vaccination and post-vaccination monitoring. Guidelines. Editors Samia Metwally Susanne Münstermann
 Foot and mouth disease vaccination and post-vaccination monitoring Guidelines Editors Samia Metwally Susanne Münstermann Authors Giancarlo Ferrari David Paton Sergio Duffy Chris Bartels Theo Knight-Jones
Foot and mouth disease vaccination and post-vaccination monitoring Guidelines Editors Samia Metwally Susanne Münstermann Authors Giancarlo Ferrari David Paton Sergio Duffy Chris Bartels Theo Knight-Jones
Foot and mouth disease vaccination and post-vaccination monitoring. Guidelines. Editors Samia Metwally Susanne Münstermann
 Foot and mouth disease vaccination and post-vaccination monitoring Guidelines Editors Samia Metwally Susanne Münstermann Authors Giancarlo Ferrari David Paton Sergio Duffy Chris Bartels Theo Knight-Jones
Foot and mouth disease vaccination and post-vaccination monitoring Guidelines Editors Samia Metwally Susanne Münstermann Authors Giancarlo Ferrari David Paton Sergio Duffy Chris Bartels Theo Knight-Jones
SWINE VESICULAR DISEASE
 NB: Version adopted by the World Assembly of Delegates of the OIE in May 2013 CHAPTER 2.8.9. SWINE VESICULAR DISEASE SUMMARY Swine vesicular disease (SVD) is a contagious disease of pigs, caused by an
NB: Version adopted by the World Assembly of Delegates of the OIE in May 2013 CHAPTER 2.8.9. SWINE VESICULAR DISEASE SUMMARY Swine vesicular disease (SVD) is a contagious disease of pigs, caused by an
PROGRESS REPORT FOR TURKEY ON FMD SITUATION AND CONTROL MEASURES
 Appendix 4 PROGRESS REPORT FOR TURKEY ON FMD SITUATION AND CONTROL MEASURES Dr H Askaroglu Introduction Foot-and-Mouth Disease (FMD) is endemic in the Anatolia Region of Turkey, due to two serotypes. Types
Appendix 4 PROGRESS REPORT FOR TURKEY ON FMD SITUATION AND CONTROL MEASURES Dr H Askaroglu Introduction Foot-and-Mouth Disease (FMD) is endemic in the Anatolia Region of Turkey, due to two serotypes. Types
SWINE HEALTH INFORMATION CENTER FINAL RESEARCH GRANT REPORT FORMAT
 SWINE HEALTH INFORMATION CENTER FINAL RESEARCH GRANT REPORT FORMAT Project Title and Project identification number: Development of a FMDV 3ABC antibody ELISA for swine oral fluid specimens (#17-191) Investigators:
SWINE HEALTH INFORMATION CENTER FINAL RESEARCH GRANT REPORT FORMAT Project Title and Project identification number: Development of a FMDV 3ABC antibody ELISA for swine oral fluid specimens (#17-191) Investigators:
FMD regional update Gulf States 2017
 FMD regional update Gulf States 2017 Nick Lyons on behalf of Donald King donald.king@pirbright.ac.uk WRLFMD Team: Valerie Mioulet, Nick Knowles, Anna Ludi, Ginette Wilsden, Bryony Armson, Pip Hamblin,
FMD regional update Gulf States 2017 Nick Lyons on behalf of Donald King donald.king@pirbright.ac.uk WRLFMD Team: Valerie Mioulet, Nick Knowles, Anna Ludi, Ginette Wilsden, Bryony Armson, Pip Hamblin,
Prevalence of Antibodies Against Foot-and-Mouth Disease Virus in Cattle in Kasese and Bushenyi Districts in Uganda
 International Journal of Animal and Veterinary Advance 2(3): 89-96, 2010 ISSN: 2041-2908 Maxwell Scientific Organization, 2010 Submitted Date: April 24, 2010 Accepted Date: May 26, 2010 Published Date:
International Journal of Animal and Veterinary Advance 2(3): 89-96, 2010 ISSN: 2041-2908 Maxwell Scientific Organization, 2010 Submitted Date: April 24, 2010 Accepted Date: May 26, 2010 Published Date:
Dr Ghazi Yehia OIE Regional Representative for the Middle East
 Foot and mouth disease control strategies in North Africa and the Middle East The current situation Asuncion, Paraguay, 24-26 June 2009 Dr Ghazi Yehia OIE Regional Representative for the Middle East Acknowledgements
Foot and mouth disease control strategies in North Africa and the Middle East The current situation Asuncion, Paraguay, 24-26 June 2009 Dr Ghazi Yehia OIE Regional Representative for the Middle East Acknowledgements
Appendix 22. Vosloo, W. 1*, Dwarka, R.M. 1, Bastos, A.D.S. 2, Esterhuysen, J.J. 1, Sahle, M. 3, Sangare, O. 4
 Appendix 22 Molecular epidemiological studies of Foot-and-Mouth disease virus in sub-saharan Africa indicate the presence of large numbers of topotypes: implications for local and international control
Appendix 22 Molecular epidemiological studies of Foot-and-Mouth disease virus in sub-saharan Africa indicate the presence of large numbers of topotypes: implications for local and international control
Emergency vaccination of sheep against foot-and-mouth disease: protection against disease and reduction in contact transmission
 Vaccine 17 (1999) 1858±1868 Emergency vaccination of sheep against foot-and-mouth disease: protection against disease and reduction in contact transmission S.J. Cox *, P.V. Barnett, P. Dani 1, J.S. Salt
Vaccine 17 (1999) 1858±1868 Emergency vaccination of sheep against foot-and-mouth disease: protection against disease and reduction in contact transmission S.J. Cox *, P.V. Barnett, P. Dani 1, J.S. Salt
Detection of neuraminidase-inhibiting antibodies for measurement of Influenza vaccine immunogenicity
 Borgis New Med 2015; 19(4): 147-155 DOI: 10.5604/14270994.1191796 Detection of neuraminidase-inhibiting antibodies for measurement of Influenza vaccine immunogenicity *Mónika Rózsa 1, István Jankovics
Borgis New Med 2015; 19(4): 147-155 DOI: 10.5604/14270994.1191796 Detection of neuraminidase-inhibiting antibodies for measurement of Influenza vaccine immunogenicity *Mónika Rózsa 1, István Jankovics
The Global FMD situation: SEACFMD in the Global Context
 The Global FMD situation: SEACFMD in the Global Context Anna Ludi anna.ludi@pirbright.ac.uk WRLFMD Team: Donald King, Valerie Mioulet, Nick Knowles, Anna Ludi, Ginette Wilsden, Bryony Armson, Pip Hamblin,
The Global FMD situation: SEACFMD in the Global Context Anna Ludi anna.ludi@pirbright.ac.uk WRLFMD Team: Donald King, Valerie Mioulet, Nick Knowles, Anna Ludi, Ginette Wilsden, Bryony Armson, Pip Hamblin,
Influenza A H1N1 (Swine Flu 2009) Hemagglutinin / HA ELISA Pair Set
 Hemagglutinin / HA ELISA Pair Set") Influenza A H1N1 (Swine Flu 2009) Hemagglutinin / HA ELISA Pair Set Catalog Number : SEK001 To achieve the best assay results, this manual must be read carefully before using this product and the assay
Influenza A H1N1 (Swine Flu 2009) Hemagglutinin / HA ELISA Pair Set Catalog Number : SEK001 To achieve the best assay results, this manual must be read carefully before using this product and the assay
Influenza A H7N9 (A/Anhui/1/2013) Hemagglutinin / HA ELISA Pair Set
 Hemagglutinin / HA ELISA Pair Set") Influenza A H7N9 (A/Anhui/1/2013) Hemagglutinin / HA ELISA Pair Set Catalog Number : SEK40103 To achieve the best assay results, this manual must be read carefully before using this product and the assay
Influenza A H7N9 (A/Anhui/1/2013) Hemagglutinin / HA ELISA Pair Set Catalog Number : SEK40103 To achieve the best assay results, this manual must be read carefully before using this product and the assay
A COMPETITIVE ELISA FOR THE DETECTION OF GROUP-SPECIFIC ANTIBODY TO EQUINE ENCEPHALOSIS VIRUS
 A COMPETITIVE ELISA FOR THE DETECTION OF GROUP-SPECIFIC ANTIBODY TO EQUINE ENCEPHALOSIS VIRUS J.E. Crafford 1, A.J. Guthrie 2, M Van Vuuren 1, P.P.C Mertens 3, J.N. Burroughs 3, P.G. Howell 2, C. A. Batten
A COMPETITIVE ELISA FOR THE DETECTION OF GROUP-SPECIFIC ANTIBODY TO EQUINE ENCEPHALOSIS VIRUS J.E. Crafford 1, A.J. Guthrie 2, M Van Vuuren 1, P.P.C Mertens 3, J.N. Burroughs 3, P.G. Howell 2, C. A. Batten
INTERNATIONAL STANDARDS FOR VACCINE BANKS
 NB: Version adopted by the World Assembly of Delegates of the OIE in May 2008 CHAPTER 1.1.10. INTERNATIONAL STANDARDS FOR VACCINE BANKS INTRODUCTION Emergency vaccination is one of several measures that
NB: Version adopted by the World Assembly of Delegates of the OIE in May 2008 CHAPTER 1.1.10. INTERNATIONAL STANDARDS FOR VACCINE BANKS INTRODUCTION Emergency vaccination is one of several measures that
Excretion of bluetongue virus in cattle semen: a feature of laboratory-adapted virus
 Vet. Ital., 40 (4), 497-501 Bluetongue virus and disease Excretion of bluetongue virus in cattle semen: a feature of laboratory-adapted virus P.D. Kirkland (1), L.F. Melville (2), N.T. Hunt (2), C.F. Williams
Vet. Ital., 40 (4), 497-501 Bluetongue virus and disease Excretion of bluetongue virus in cattle semen: a feature of laboratory-adapted virus P.D. Kirkland (1), L.F. Melville (2), N.T. Hunt (2), C.F. Williams