Redacted for Privacy
|
|
|
- Joella Shields
- 5 years ago
- Views:
Transcription
1 AN ABSTRACT OF THE THESIS OF Barbara S. Drolet for the degree of Doctor of Philosophy in Genetics presented on September 15, Title: Immunohistochemical Analysis of Infectious Hematopoietic Necrosis Virus in Rainbow Trout (Oncorhynchus mykiss). Redacted for Privacy Abstract approved: Jo-Ann C. Leong Infectious hematopoietic necrosis virus (IHNV) is a rhabdovirus which causes devastating epizootics in trout and salmon hatcheries of the Pacific Northwest, Europe and Asia. The complete infection cycle of IHNV in fish has not been determined. It is known that IHNV primarily infects young fish causing an acute, systemic disease which results in mortality rates of up to 100%. The studies described herein were designed to examine the complete pathogenesis of IHNV. The route of viral entry, progression of the virus through the fish tissues, and the possibility of persistence of the virus within the epizootic survivor is examined and discussed. Two immunochemical assays were used to detect the nucleocapsid protein of IHNV in infected cells. To relate the positive staining intensity to the degree of viral infection, an alkaline phosphatase immunocytochemical (APIC) assay was developed to detect IHNV in fixed tissue culture cells. The assay was published as an alternative to current methods used to confirm IHNV in tissue culture. To follow the IHNV infection in the fish, an alkaline phosphatase immunohistochemical (APIH) assay was developed to detect the nucleocapsid protein of the virus in whole, fixed, paraffin-embedded fish. Infected rainbow trout were tested
2 by APIH before and during the epizootic to examine the route of viral entry and the progression of the virus, respectively. Virus entered through the gill epithelium during normal oxygen uptake, and through the gastrointestinal epithelium during digestion. Virus spread to virtually every organ via the circulatory system. Progression of IHNV within and between organs is discussed. Approximately fifty days after the epizootic, survivors are negative for infectious virus by standard diagnostic methods and are released into the wild. The virus source for IHNV-positive returning adults is unknown and has resulted in a twenty year debate of secondary reservoir versus viral persistence. The last part of this study, describes IHNV-related lesions in the kidney of rainbow trout one year after virus exposure and persistence of the virus as shown by immunohistochemistry, PCR amplification, and immunogold electron microscopy.

3 Copyright by Barbara S. Drolet September 15, 1994
4 IMMUNOHISTOCHEMICAL ANALYSIS OF INFECTIOUS HEMATOPOIETIC NECROSIS VIRUS IN RAINBOW TROUT (ONCORHYNCHUS MYKISS) by Barbara S. Drolet A THESIS submitted to Oregon State University in partial fulfillment of the requirements for the degree of Doctor of Philosophy Completed September 15, 1994 Commencement June 1995
5 Doctor of Philosophy thesis of Barbara S. Drolet presented on September 15, 1994 APPROVED: Redacted for Privacy Professor representing Gene Redacted for Privacy Director of Genetics Program Redacted for Privacy Dean of Gradu School I understand that my thesis will become part of the permanent collection of Oregon State University libraries. My signature below authorizes release of my thesis to any reader upon request. Redacted for Privacy Barbara S. Drolet, Author
6 i ACKNOWLEDGMENTS I thank Dr. Jo-Ann Leong, my mentor, for always having faith in my ability, and pushing me to find the best in myself. I thank her for the kind, but firm words of encouragement and instruction of what it takes for women to survive and grow in this field. I thank Dr. John Rohovec for his advice, guidance, encouragement, humor and friendship. I thank Dr. George Pearson for his guidance my first year, and for always taking the time to listen and advise these past 5 years. I thank my family for their constant support, and for never being further than a phone call away. I thank Dr. Carla Mason Endres, my confidant and touchstone. I thank her for our many hours of verbal PCR, the reality checks, and the unconditional friendship she has given me. I thank Dan Mourich and Jon Piganelli for their friendship and humor, which helped keep my sanity intact throughout the years. Finally, I thank Bill Keleher for making my last year here the best.
7 ii CONTRIBUTION OF AUTHORS Dr. Jo-Ann C. Leong was involved in experimental design, data analysis and interpretation, writing criticism and financial support of the work contained within each manuscript. Dr. John S. Rohovec was involved in experimental design, writing criticism, and financial support of the work contained within Chapters 3 and 4. Dr. Jerry Heidel provided histological expertise, writing criticism, and financial support of the work contained within the manuscript in Chapter 5. Pinwen (Peter) Chiou provided PCR amplification expertise, crucial to the work contained within the manuscript in Chapter 5.
8 iii TABLE OF CONTENTS Chapter Page 1. Thesis Introduction 1 2. Literature Review 6 Infectious Hematopoietic Necrosis Virus Infectious Hematopoietic Necrosis 3. Serological Identification of Infectious Hematopoietic Necrosis Virus in Fixed Tissue Culture Cells by Alkaline Phosphatase Immunocytochemistry Abstract Introduction Materials and Methods Results and Discussion Acknowledgments References The Route of Entry and Progression of Infectious Hematopoietic Necrosis Virus in Oncorhynchus mykiss: A Sequential Immunohistochemical Study Abstract Introduction Materials and Methods Results Discussion Acknowledgments References Persistence of an RNA Virus Infection by Defective Interfering Particles 59 Abstract Introduction Materials and Methods Results Discussion Acknowledgments References Thesis Summary 86 BIBLIOGRAPHY APPENDIX
9 iv LIST OF FIGURES Figure Page 2.1 Clinical signs of IHN in rainbow trout (0. mykiss) Nucleocapsid protein of infectious hematopoietic necrosis virus (IHNV) in infected cells by alkaline phosphatase immunocytochemical staining using monoclonal antibody, 1NDW14D 4.1 Mortality curve of 1 g steelhead trout (0. mykiss) after immersion infection with infectious hematopoietic necrosis virus, type Photomicrographs of tissues from infectious hematopoietic necrosis virus infected steelhead fry during the pre-epizootic period (0-5 days post exposure) and the epizootic period (6-14 days post exposure) 5.1 Kidney tissue from a 1 ype IHN epizootic survivor stained with Masson's Trichrome stain 5.2 Kidney tissue from a 1 ypi IHNV survivor as tested by alkaline phosphatase immunohistochemistry 5.3 Cytoplasmic rhabdoviral inclusions in kidney tissue from a 1 ype IHN epizootic survivor 5.4 a Immunogold labeling of IHNV particles using the anti-nucleocapsid monoclonal antibody, 1NDW14D 5.4 b Immunogold labeling of IHNV particles using anti-glycoprotein monoclonal antibody, 3GH136J 5.5 Immunogold labeled IHNV DI particles from 1 ype IHN epizootic survivor 5.6 a Retrotherm PCR amplification of a 252 by fragment of IHNV N gene from kidney tissue of a 1 year post exposure survivor 5.6 b Southern blot hybridization showing 252 by fragment is of IHNV N gene origin
10 V LIST OF TABLES Table Page 3.1 Virus isolates tested by alkaline phosphatase immunocytochemistry with an anti-nucleocapsid monoclonal antibody, 1NDW14D 4.1 Chronological appearance and staining intensity of infectious hematopoietic necrosis virus by alkaline phosphatase immunohistochemistry in steelhead (0. mykiss) fry 4.2 Virus-positive tissues of fish organs as tested by alkaline phosphatase immunohistochemistry
11 vi DEDICATION I dedicate this thesis to my family. Their love and support have given me a firm base to stand and wings to fly.
12 IMMUNOHISTOCHEMICAL ANALYSIS OF INFECTIOUS HEMATOPOIETIC NECROSIS VIRUS IN RAINBOW TROUT (ONCORHYNCHUS MYKISS) CHAPTER 1 THESIS INTRODUCTION Infectious hematopoietic necrosis virus (IHNV) is a rhabdovirus which causes an economically important disease affecting hatcheries around the world. The lifecycle of this virus is not completely known. The studies described in this thesis address the pathogenesis and persistence of IHNV in rainbow trout. The complete pathogenic pathways are known for very few organisms. The most comprehensive studies have been done with reoviruses (Sharpe and Fei lds 1983; Nibert et al. 1991), influenza (Brown et al. 1992) and LCM virus (Cole et al. 1971). The physical size of the host usually prevents whole-body sequential studies often needed for extensive examination of pathogenic pathways in vivo. Because IHNV infects primarily young fish, a complete, whole-body examination of viral pathogenesis was accomplished. Understanding the complete pathogenic pathway of any infectious agent is imperative for designing effective disease treatments and vaccines. Objectives The objectives of these studies were to 1) determine the route of viral entry, 2) examine the progression of virus through the tissues, and 3) determine if a latent or subclinical form of IHNV persists in epizootic survivors.
13 2 Approach To study the pathway of IHNV entry and progression, virus had to be detected undisturbed in vivo. At the time of this study, in situ hybridization and polymerase chain reaction (PCR) amplification technologies were not available for IHNV in fish tissues. In addition, the instability of RNA viruses and the ubiquitous nature of RNAses would have made in situ hybridization and PCR too labor and time intensive for routine examination of the large number of samples needed for this type of study. The plaque assay (Amos 1985) and immunofluorescence (LaPatra 1989b) are well established and are used routinely to detect infectious IHNV in tissue culture and infected tissues. However, both of these assays would require the separation and processing of individual organs and tissues from the host to examine progression of virus within the fish, which destroys the necessary in vivo nature of the study. In addition, the processing of tissues for tissue culture assays results in dilution of the samples which decreases the assay's sensitivity. Direct, in vivo immunofluorescent staining of tissues was attempted for this study, but not pursued because of the shortlived, unstable reaction product, the necessity of examining all sections under a fluorescent microscope, and the results, similar to those reported by Yamamoto et al. (1989), of autofluorescing materials in fish tissues. Because no existing IHNV detection assay was suitable for this study, an immunohistochemical assay was developed which detected stable viral proteins, in vivo, resulting in a permanent color reaction. There was no minimal sample size, and the reaction was highly specific and sensitive. Fixation of tissue Formaldehyde, a cross-linking fixative, was used to immediately immobilize and stabilize tissue and viral components in their native conformation. Formaldehyde
14 3 stabilizes proteins by forming cross-links between polypeptides both within individual proteins and between neighboring proteins. Amino acids involved in formaldehyde cross-linking are those possessing free amino groups, thiol groups, or aromatic ring systems. Formaldehyde fixation is ph, temperature and time dependent. Alkaline phosphatase immunohistochemistry (APIH) An alkaline phosphatase immunohistochemical (APIH) assay was used to detect viral protein in fixed, paraffin-embedded fish tissues. This assay utilized a universal monoclonal antibody which recognizes the most highly conserved, abundant protein of all IHNV types (Ristow and Arnzen 1989), and an alkaline phosphatase (AP) enzyme detection system from Vector Laboratories (Burlingame, CA). The AP system was chosen over immunoperoxidase (IP) because the staining intensity of AP is not as dense as IP, making morphological examination of the stained cells possible. The immunohistochemical method used was derived from the immunoenzymatic studies of Hsu et al. (1981a; 1981b) using avidin-biotin complexes for amplification of antigens in fixed tissue. The chromogen substrate chosen in this study was Vector Red (Vector Labs) with levamisole (Ponder and Wilkinson 1981) added to eliminate endogenous phosphatase activity in the fish tissues. Immunogold electron microscopy Although infectious virus can not be detected by tissue culture in fish past 50 days post exposure (dpe), virus is detected in spawning adults. Virus in adults is thought to be the result of reinfection during spawning migration. The unexpected detection of viral protein in one year post exposure (ype) survivors by APIH, led to an electron microscopy (EM) study to examine survivor fish for persisting virus. An
15 4 immunogold EM procedure was adapted from previous immunochemical EM studies (Faulk and Taylor 1971; Horisberger and Rosset 1977) and used to examine fresh and archived tissues. Introduction to thesis chapters Chapter 3 is a published manuscript which describes a tissue culture adaptation of the histochemical assay used in the pathogenic study. An alkaline phosphatase immunocytochemical (APIC) assay was used to characterize the stain precipitate of the AP immunochemical reaction. Degrees of staining intensity were rated 1 through 5, weakest to strongest, and were related to the degree of productive infection. This assay is currently the only method available to confirm IHNV in fixed, stained plaque assay plates. Chapter 4 is a published manuscript reporting the sequential, immunohistochemical analysis of the complete pathogenic pathway of IHNV in rainbow trout. The objective of the study was to characterize the pathogenesis of IHNV infection of fish by identifying the route of viral entry and the tissues in which the virus replicates. Sequential virus distribution following immersion exposure had not been previously defined. Virus entered both by ingestion and penetration of the gill epithelium and was disseminated by the circulatory system. As indicated by it's name, it is in the hematopoietic tissue that IHNV is most productive. In fish, that organ is the kidney. Fish kidneys are functionally divided into the anterior, or head kidney and the posterior kidney. The anterior kidney is primarily composed of hematopoietic cells and is thought to be the major site of blood cell formation. The relative proportions of red and white cells in the kidney varies from month to month, but overall there are roughly equal numbers of each cell type present (Tatner and Manning 1983a). The posterior kidney is primarily composed of nephrons
16 5 embedded in hematopoietic tissue and functions to conserve salt and eliminate excess water. The spleen is considered a secondary hematopoietic organ (Grace and Manning 1980; Tatner and Manning 1983a). Infection of the hematopoietic tissue results in slow recovery of infected or destroyed blood cells and may result in damage to the kidney if the fish survives the epizootic. Chapter 5 is a manuscript reporting the pathology and viral persistence observed in IHN epizootic survivors at one ype. The health of survivors has not been closely monitored in the wild and it is uncertain how many survive to sexual maturation. Epizootic survivors are released into the wild and are assumed to be virus-free because they show no clinical signs of IHN, and testing fish by tissue culture methods consistently shows no evidence of virus. However, when adults return to spawn, they are again positive for IHNV. For the past 20 years, fisheries' scientists have been studying the life cycle of IHNV trying to prove or disprove its persistence in survivor populations. In this paper, lesions in the kidney of rainbow trout one year after exposure are examined and evidence for subclinical persistence of IHNV is demonstrated in the detection of viral protein by immunohistochemistry, viral RNA by PCR amplification, and IHNV DI particles by immunogold electron microscopy.
17 6 CHAPTER 2 LITERATURE REVIEW Infectious Hematopoietic Necrosis Virus Virion characteristics Infectious hematopoietic necrosis virus (IHNV) is a member of the Rhabdoviridae family, in the genus Lyssavirus. Rhabdoviruses are widely distributed, causing diseases in plants, insects, fish, amphibians, reptiles, wild and domestic mammals, and humans. The prototypic Lyssavirus is rabies virus. Morphology. Standard particles of IFINV are bullet-shaped, with an average length of 160 nm and a core diameter of nm (Amend and Chambers 1970). Virions have an enveloped, helical nucleocapsid with glycoprotein peplomers projecting 10 nm from the surface giving a final diameter of about nm. Nucleic acid. The nucleic acid of IHNV was shown to be RNA by Wingfield et al. (1969). Subsequent work showed the RNA to be single-stranded negative sense and non-segmented (McCain et al. 1974; McAllister et al. 1974; Hill et al. 1975; McAllister and Wagner 1977). IHNV is a unique member of the Rhabdoviridae family in that its genome contains six genes instead of the characteristic five. The six genes of IHNV have been shown by electrophoresis to encode five structural proteins; a nucleocapsid (N) protein, two matrix (M1, M2) proteins, a glycoprotein (G), and an RNA dependent RNA polymerase (L) (McAllister and Wagner 1975; Hill et al. 1975; Lenoir and de Kinkelin 1975; Leong et al. 1981; Kurath et al. 1985). A sixth protein, which can only be detected in virally infected cells, is designated as the nonvirion (NV) protein (Kurath and Leong 1985). The gene order is 3'-N-M1-M2-G-NV-L-5' as determined by R-loop
18 7 mapping (Kurath et al. 1985). Except for the unique NV gene, the gene order of IHNV is identical to the other members of the Lyssavirus genus and members of the Vesiculovirus Rhabdoviridae genus. Proteins. The nucleocapsid protein is the most abundant and highly conserved protein of IHNV (Gilmore and Leong 1988). The N protein binds to the ssrna genome and provides stability and structure. It is necessary for encapsidation and infectivity. Because of the highly conserved, abundant nature of the N protein, it is the target antigen for many diagnostic assays and pathogenic studies. The glycoprotein has been shown to be the sole viral antigen responsible for inducing protective immunity in fish (Engelking and Leong 1989a; Xu et al. 1991). Antibody produced to the G protein induces protective immunity against all five IHNV types (Engelking and Leong 1989b). The glycoprotein continues to be the focus of vaccine studies for IHNV (de Kinkelin et al. 1984; Leong et al. 1988; Leong and Munn 1991; Oberg et al. 1991; Xu et al. 1991; Leong 1993; Leong and Fryer 1993). Types The IHN virus has been classified into five electropherotypes based on gel migration patterns of the nucleocapsid protein and glycoprotein (Leong et al. 1981; Hsu et al. 1986). Type differences in average plaque diameter (Chen et al. 1990), virulence (LaPatra et al. 1990), antigenicity (Winton et al. 1988; Basurco et al 1993; Ristow and de Avila 1991), and single-step growth curves (Mulcahy et al. 1984b) have been shown. Hsu et al. (1986) reported a geographical predominance for specific isolates in certain areas. The electropherotypes of interest to the studies described herein are Round Butte-83 (Type 1) and Rangen (Type 2) which cause a majority of outbreaks in Oregon and Idaho.
19 8 Cell culture Permissive cell lines. Rhabdoviruses have been shown to infect a diverse group of cell types including plants, insects, reptiles, mammals, birds, amphibians and fish (Clark and Soriano 1974; Wagner 1975; Scott et al. 1980). IHNV was first isolated in primary fish cell cultures from infected salmon tissues in 1980 (Pilcher and Fryer 1980). Permissive fish cell lines used routinely today include carp epithelioma papulosum cyprini EPC cells (Fijan et al. 1983), rainbow trout gonad RTG-2 cells (Wolf and Quimby 1962), chinook salmon embryo CHSE-214 cells (Lannan et al. 1984), steelhead trout embryo STE-137 cells (Fryer et al. 1965; Fendrick et al. 1982), sockeye salmon embryo SSE-5 cells (Nims et al. 1970), Atlantic salmon AS cells (Nicholson and Byrne 1973) and fathead minnow FHM cells (Gravell and Malsberger 1965). Chum salmon embryo (SE) and Yamame 0. masou kidney (YNK) cells (Asakawa et al. 1986) have also shown susceptibility to IHNV. Growth requirements. The survival of IHNV has been evaluated under several environmental conditions such as dessication, ph, temperature, water hardness, and salinity (Pietsch et al. 1977; Kamei et al. 1987; 1988). Virus grows best at temperatures between C (Wingfeild et al. 1969; Yasutake and Amend 1972). The low temperature requirement may be due to the heat liability of the virion polymerase (McAllister and Wagner 1975) or the synthesis of interferon at higher temperatures (de Kinkelin et al. 1982). Infection. In cell culture, the cytopathic effect (CPE) appears initially as thickened nuclear membranes. Cells then become granular and rounded, and eventually detach from the surface of the culture vessel. A virus titer of lx108 pfu/ml can be harvested from the supernatant of CHSE-214 cells at peak CPE. The appearance of
20 9 CPE depends on the isolate being grown and the sensitivity of the cell line being used (Kelly et al. 1978; Fendrick 1982; Mulcahy et al. 1984a; Chen et al. 1990). In one-step growth curves with CHSE-214 cells at 18 C, viral antigens can be detected in the cytoplasm by fluorescent antibody at two hours post infection (hpi), and infectious virions appear by four hpi (McAllister et al. 1974). Stability. The infectivity of II-INV is stable if stored at -70 C or -20 C, but decreases at temperatures above 4 C (Pietsch et al. 1977). Virus is not stable in salt water or fresh water containing high numbers of bacteria. Virus has been shown to be sensitive to iodine. Virally contaminated eggs and water can be effectively disinfected with an iodophor treatment (Amend and Pietsch 1972a; Bans et al. 1991). Although IHNV is sensitive to temperatures above 18 C, raising the water temperature is only considered to be a practical solution in controlling the temperature sensitive isolate of IHNV at Coleman Hatchery where there is a ready source of geothermal water. Elsewhere, rainbow trout infection experiments have shown that the higher the temperature the shorter the survival time of the fish (Hetrick et al. 1979). Infectious Hematopoietic Necrosis Infectious hematopoietic necrosis is an acute, systemic, economically devastating disease in trout and salmon populations. Although originally isolated from feral populations, IHN is primarily a problem in hatcheries where large numbers of fish are grown at high densities. While density does not affect survival from egg to fry, it does effect the level of virus titer and thus results in increased mortality during viral epizootics among fry. Lack of genetic selection for resistance was also suggested as a cause of IHN associated mortality (Mulcahy and Bauersfeld 1983).
21 10 Distribution World-wide. IHNV was first reported in 1953 as the cause of a disease among sockeye salmon in Washington (Rucker et al. 1953) and again in 1967 in sockeye in western Canada (Amend et al. 1969). It is endemic to the Pacific Northwest United States and Canada (Parisot et al. 1965; Grischkowsky and Amend 1976; Traxler 1983). Natural outbreaks have been reported in California, Oregon, Washington, Alaska (Leong et al. 1981; Burke and Grischkowsky 1984; Grischkowsky and Amend 1976) and British Columbia (Williams and Amend 1976). The disease was introduced into Japan at Hokkaido in 1968 with eggs from North America (Kimura and Awakura 1977) and rapidly spread throughout the Japanese mainland (Sano et al. 1977; Yoshimizu et al. 1988; Yoshimizu et al. 1993). IHNV isolates have recently been reported in Korea (Park et al. 1993), Taiwan, Belgium (Hill 1992), France (Laurencin 1987), and Italy (Bovo et al. 1987). Oregon. The IHN virus was the first fish virus to be isolated and identified in the state of Oregon (Wingfield et al. 1969). It is thought that IHNV was introduced into Oregon from Puget Sound, Washington, in the form of unpasteurized sockeye salmon viscera which had been incorporated into the diet of the fish. In Oregon the incidence of IHN continues to increase from 6 hatcheries and holding ponds in 1981 to 43 sites in 1994 (Engelking, personal communication). Host range IHNV commonly infects sockeye (Oncorhynchus nerka), kokanee (0. nerka) chum (0. keta), and chinook (0. tshawytscha) salmon and rainbow (0. mykiss) and steelhead (0. mykiss) trout (Pilcher and Fryer 1980; Wolf 1988). Young coho salmon (0. kisutch) are considered resistant to IHNV infection, yet the virus has been detected
22 11 in adult coho in Oregon ( LaPatra et al. 1989a), Washington (Eaton et al. 1991) and California (Hedrick et al. 1987). Cutthroat (0. clarkii), brook (Salvelinus fontinalis) (Goldes and Mead 1992; LaPatra et al. 1992; Bootland et al. 1994), and brown (Salmo trutta) (LaPatra and Fryer 1990) trout have shown susceptibility to experimental IHNV infections. Experimental infection studies with non-salmonid species have shown whitefish to be susceptible, and squaw fish, suckers and lamprey to be resistant to IHNV infection (Bootland and Lorz, unpublished results). In addition, two marine fish species, seabream (Archosargus rhomboidalis) and turbot (Scophthalmus maximus), have shown susceptibility to IHNV infection by intraperitoneal injection (Castric and Jeffroy 1991). IHNV primarily kills young fish: alevins, fry and early juveniles weighing less than 5 g. It has been suggested that susceptibility decreases with increased size and age of the fish (Kasai et al. 1993). However, kokanee and rainbow trout have been shown to be susceptible to two types of IHNV at weights up to 13 g (LaPatra et al. 1990). Transmission Horizontal. Horizontal transmission of IHNV is well documented (for review, see Wolf 1988). Virus is transmitted through feces, urine, and ovarian or seminal fluid (Pilcher and Fryer 1980; Mulcahy et a. 1983a; 1983b; Mulcahy and Batts 1987; Nishimura et al. 1988). Virus can be detected in effluent hatchery water during large epizootics (Leong and Turner 1979; Yoshimizu et al. 1991), and can be transmitted to fish held down stream (Mulcahy et a. 1983b). Vertical. Evidence for vertical transmission is circumstantial. The strongest evidence comes from the association made between the appearance of the disease and the shipment of eggs from infected adults into geographical areas where the virus was
23 12 not known to occur (Amend 1975). In nature, the greatest danger of an epizootic is in the spring when newly hatched fry become infected with virus excreted in sex products during spawning. Infection of hatching fry is thought to be due to virus contaminating the surface of the egg and not virus within the egg itself. However, neutralizing antibodies have been isolated from eggs of spawning steelhead trout (Shors and Winston 1989a) indicating either an internal active infection or passively acquired maternal antibody. Virus has been shown to pass in sockeye salmon from infected adult to egg through ovarian or seminal fluid (Mulcahy and Pascho 1985; Meyers 1990), and virus has been shown to adsorb to fish sperm (Mulcahy and Pascho 1984). A prevalence study in 1983 reported feral, introduced and hatchery-reared salmonid species in the Pacific Northwest had ovarian fluid IHNV titers of 105 pfu/ml at 25%, 43% and 32% respectively (Mulcahy et al. 1983c). The authors suggested that the success of the feral population was due to IHNV titers high enough to maintain the virus in the population, but low enough to cause little transmission of lethal doses through the eggs. The possibility of vertical transmission was studied by Yoshimizu et al. (1989) using the eggs of masu and chum salmon. Eggs externally contaminated with IHNV showed no virus after one day. Virus injected into just-fertilized eggs could not be detected by one month post injection. However, virus injected into eyed eggs caused infection in 90% of the embryos. Several components of the yolk showed anti-viral activity. It was suggested that vertical transmission does not occur, in that even if virus was internalized it would not survive to the eyed stage. In addition, an extensive fertilization study by LaPatra et al. (1991) showed no vertical transmission in steelhead populations. Thus, vertical transmission of IHNV remains controversial. Covert. A third transmission route, covert transmission, is currently being studied by the Oregon Department of Fish and Wildlife pathology staff (LaPatra
24 b). By differentially marking progeny from carrier (virus-positive) and virus-free parents, they hope to determine whether progeny from carrier adults return to hatcheries as carriers of the virus. Environmental reservoirs Persistence of IHNV in the environment has resulted in several studies examining possible secondary viral reservoirs. Thirty-six environmental sampling trips, consisting of 215 benthos and 36 concentrated water samples (Leong and Turner 1979; Batts and Winton 1989a) have been taken over a three year period from 4 sites in Oregon. Results showed no evidence of virus with the exception of 1 water sample taken from the Metolius River (Lorz and Stevens, unpublished data). Virus has been isolated from a freshwater leech (Piscicola salmositica) and a copepod (Salmincola sp.) taken from the gills of infected, spawning sockeye salmon (Mulcahy et al. 1990). Virus has also been isolated from Mayflies found near DIN epizootic sites (Shors and Winston 1989b). It has been suggested that these organisms may act as a vector in which the virus could overwinter and serve as reservoirs of infection, but IHNV has yet to be shown to replicate within these hosts. Pathogenesis The main target of IHNV is the hematopoietic organ of the fish, which is the kidney. Initial sites of replication have been examined by plaque assay (Yamamoto et al. 1992), immunohistochemistry (Yamamoto et al. 1990; Drolet et al. 1994) and electron microscopy (Yamamoto and Clermont 1990). Several infection studies have implicated the gills as the initial site of IHNV entry and infection. Virus has been isolated by tissue culture in gill tissue early in the infection of sockeye fry (Mulcahy et al. 1983a) and smolts (Burke and Grischkowsky 1984). Virus was also isolated by
25 14 tissue culture and G protein detected by immunoperoxidase histochemistry from gill tissue of adult kokanee and sockeye at 3 days post exposure (dpe) (Yamamoto et al. 1989). Virus was detected in gill tissue of rainbow trout at hours post exposure (hpe) by electron microscopy and at 24 hpe by tissue culture methods (Yamamoto and Clermont 1990), but not until 5 dpe by immunohistochemistry (Yamamoto et al. 1990). Yamamoto et al. (1990) also reported that the skin was the major site of IHNV replication and that the gill played a lesser role in the initial stages of infection. Gills have been shown to be the initial site of replication for two other fish rhabdoviruses, spring viremia of carp virus (Ahne 1978) and viral hemorrhagic septicemia virus (Neukirch 1984). Entry at the gill followed by dissemination of the virus by the blood has been suggested from tissue culture studies (Yasutake and Amend 1972; Mulcahy et al. 1983a). The circulatory system of fish is anatomically simple, compared with that of higher vertebrates. Blood circulates from the heart to the gill arches, where it becomes oxygenated primarily by diffusion through the epithelium of the gill lamellae. It is distributed by way of the dorsal aorta to the arteries and peripheral capillaries and returned to the heart through the venous system (Yasutake and Wales 1983). Viral entry through the gills and progression via the circulatory system was recently confirmed by immunohistochemistry (Drolet et al. 1994). Mulcahy et al. (1983a) also reported that infectious virus was detected in the viscera of naturally infected spawning sockeye in the absence of virus in the gills. Moreover, virus has been detected by immunohistochemistry (Yamamoto et al. 1990) and by electron microscopy (Yamamoto and Clermont 1990) in the visceral organs of rainbow trout before the gills. This suggested an alternate or additional origin of virus infection which was determined to be ingestion by Drolet et al. (1994).
26 15 Clinical signs Typical IHN external signs include abdominal swelling, petechial hemorrhages on the ventral surface and at base of pectoral and pelvic fins, exophthalmia, pale ventral surface, dark dorsal surface and occasionally fecal casts (Amend et al. 1969) (Figure 2.1). Dark reddish areas just behind the head are thought to be due to subcutaneous pooling of extravascated blood from the anterior kidney. Internally, the kidney and liver are anemic, ascitic fluid may fill the peritoneal cavity, hemorrhages may be found in the muscle, fat and liver, and the digestive tract is often devoid of food. Behavioral signs include anorexia, lethargy, with brief episodes of frenzied swimming, avoidance of currents, rolling, swimming vertically or circling and flashing. Microscopically, necrosis of the hematopoietic tissues in the anterior kidney and spleen as well as necrosis of the liver and pancreas is common. Granular cells in the lamina propria, stratum compactum and stratum granulosum of the alimentary tract are also commonly necrotic (Yasutake et al. 1965; Yasutake and Amend 1972). Packed cell volume, hemoglobin, red blood cell count, and plasma bicarbonate are significantly depressed by the forth day after exposure (Amend and Smith 1974). Spinal deformities may occur in epizootic survivors.
27 16 Figure 2.1 Clinical signs of IHN in rainbow trout (0. mykiss). The three fish on top show classic signs of IHN infection including abdominal swelling, pale ventral surface, dark dorsal surface, petechial hemorrhages on ventral surface and at the base of pectoral and pelvic fins (up arrow), and exophthalmia (down arrow). On the bottom is a negative control fish.
28 Figure 2.1
29 18 Diagnostics In the field, diagnoses are based on clinical signs, histopathological changes and the hatchery's IHN epizootic history. In the laboratory, presumptive diagnosis requires the isolation of the virus in cell culture with the development of typical CPE. Plaque assay. For plaque assays, EPC cell monolayers are inoculated with infected fish tissue homogenate (Burke and Mulcahy 1980) or mucus supernatant (LaPatra et al. 1989c), incubated at 17 C, fixed and stained as suggested in the Fish Health Blue Book (Amos 1985). The recommended incubation time for tissue culture plaque assay is days, although this can be shortened if cell monolayers are pretreated with polyethylene glycol Watts and Winton 1989b) or the polycation polybrene (Leong et al. 1982). Infectious virus is quantitated by counting the number of plaques resulting from a focal viral infection. Incubation temperatures (Burke and Mulcahy 1983) and media (Amend and Pietsch 1972b; Nishimura et al. 1985) are well established. Plaque neutralization is commonly used to detect IHNV or antibody to IHNV (Jorgensen et al. 1991), and has recently been used to characterized the humoral immune response of rainbow trout to IHNV infection (LaPatra et al. 1993). Immunoassays. Recent advances in detecting IHNV (for review see Winton 1991) have taken advantage of the numerous IHNV-specific monoclonal antibodies now available (Ristow and de Avila 1991). Immunodot (McAllister and Schill 1986; Eaton et al. 1991; Ristow et a. 1991) and immunoblot (Schultz et al. 1989) assays have been developed to detect IHNV in cell culture fluid. Viral antigen can also be rapidly detected from cell culture or infected fry tissue homogenate supernatants with enzymelinked immunosorbent assays (Dixon and Hill 1984; Way and Dixon 1988; Medina et al. 1992).
30 19 A staphylococcal coagglutination test has been developed by Boot land and Leong (1992) primarily for use in the field. This assay detects all five types of IHNV in cell cultures and infected fish in 15 min, but a titer of 106 pfu/ml is required to obtain a positive reaction. Immunofluorescence is used to rapidly detect IHNV using both monoclonal and polyclonal antisera (LaPatra et al. 1989b; Arnzen et al. 1991; JOrgensen et al. 1991). Immunofluorescence or plaque neutralizations are routinely done to confirm the infectious agent as IHNV. Recently, an alternative to these subculturing confirmatory methods has been developed. An alkaline phosphatase immunocytochemical assay can now be used to confirm IHNV in fixed, stained tissue culture cells, alleviating the necessity of subculturing (Drolet et al. 1993). Immunohistochemical techniques have been developed to detect IHNV in vivo. Yamamoto et al. (1990) used a horse-radish peroxidase technique to detect multiplication sites of IHNV in rainbow trout. Most recently, the route of viral entry and progression of IHNV in rainbow trout has been examined using an alkaline phosphatase immunocytochemical assay (Drolet et al. 1994). Techniques developed to detect the nucleic acid of IHNV include in situ hybridization and PCR amplification. A biotinylated DNA probe has been used to detect IHNV mrna in infected cell cultures (Deering et al. 1991) and rainbow trout brain tissue (Anderson et al. 1991). The N gene has been amplified by PCR from purified genomic RNA of all five IHNV isolate types (Arakawa et al. 1990). Most recently, PCR amplification detected viral RNA in archived tissues fixed in formalin for 2 years or more (Chiou et al. 1994). Although not practical as a diagnostic tool, electron microscopy (EM) has been used to characterize virion structure (Amend and Chambers 1970) and infectivity patterns (Yamamoto and Clermont 1990). Most recently, scanning EM has been used to examine IHNV receptor binding and protein expression (Helmick et al. 1991).
31 20 Persistence of IHNV Tissue culture. Persistence of IHNV has been shown to occur in infected tissue culture cells. Engelking and Leong (1981) developed a cell culture model of a viruscarrier state in chinook cells. High levels of virus were released from these cells into the medium although the cells continued to grow well in culture. Virus production fluctuated with passage level and it was demonstrated that nearly 100% of the cells were producing viral antigen. Furthermore, these cells were resistant to superinfection by homologous virus but sensitive to infection by heterologous viruses. Persistence of IHNV apparently was mediated by several factors including the development of temperature sensitive mutants, small plaque mutants and defective interfering (DI) particles. Interferon did not play a significant role in persistence of the virus in these cells, although interferon was suggested as a factor in persistently infected RTG-2 cells, (Okamoto et al. 1983). Persistent IHNV infections have also been reported in YNK cells (Watanabe 1983). Fish. Persistence of IHNV in fish remains controversial. Infectious virus can only be detected in young fry and adult spawning populations. Fish are negative for infectious virus by tissue culture methods between these two life stages. The source of virus for adult populations returning to spawn is unknown. One theory is that returning adults are reinfected with IHNV from currently infected fish or some secondary reservoir during their migration upstream. The other theory is that the virus persists in the survivor in a latent or subclinical state and is reactivated during stressful spawning conditions. This controversy is at the core of understanding the life cycle of IHNV. Neither theory has been proven, nor have they been shown to be mutually exclusive. Mulcahy et al. (1982) reported that 100% of returning adult sockeye that were examined at the Cedar Creek spawning cannel were infected and virus was present in
32 21 nearly all organs. In addition, there was an overall tendency for the mean concentration to increase in many of the organs over time as the fish progressed to ripeness. Because of the correlation of IHNV appearance in returning adult sockeye and the spawning act, sex hormones have been suggested to have a role in susceptibility to reinfection, or reactivation of a latent form of the virus. However Grischkowsky and Mulcahy (1982) reported that hormones injected into naturally infected spawning adults did not significantly alter the viral infection. Evidence for the latency theory. The mechanism by which the carrier state may be established and maintained in adult fish is unknown. In 1975, Amend reported that rainbow trout that had survived an IHN epizootic and were held in pathogen-free water, became positive for virus upon sexual maturation (Amend 1975). It was suggested that the virus entered a latent state in the survivor and was reactivated during sexual maturity (Amend 1975; Mulcahy et al. 1983a; Mulcahy et al. 1984a). Adult fish appear normal and no virus can be detected in their tissues. However, high concentrations of virus do appear in ovarian and seminal fluids during spawning (Amend 1975; Mulcahy et al 1983c; Mulcahy and Batts 1987; Mulcahy et al. 1987). Low positive antibody titers against IHNV have been reported in returning spawning adults (de Avila et al. 1992), suggesting an immune memory occurring in the immunocompetent fry (Tatner and Manning 1983b) at the time of initial virus exposure. This may decrease the reinfection rate of adults during spawning, which would suggest that virus reactivation was responsible for the reappearance of IHNV in the population. The inability to define a secondary reservoir or overwintering vector has also strengthened the latency theory. It is possible that the 100% infection rate of many spawning populations results from virus activation in lifelong virus carriers and the subsequent horizontal infection of the noninfected members of the returning population (Mulcahy et al. 1982).
33 22 Evidence for the reinfection theory. Since 1975, fish disease laboratories have tried to reisolate IHNV from captive survivor populations to no avail. The inability to defmitively show the reactivation of a latent virus in numerous laboratory infection studies has provided the majority of evidence against latency. Amos et al. (1989) reported that no virus was isolated from adult sockeye salmon, captured as they exited saltwater and held in pathogen-free water until sexual maturation. Simultaneously, sockeye allowed to migrate upstream were % positive for virus by the time they reached the hatchery. It was suggested that the high prevalence of IHNV in spawning populations was the result of horizontal transmission from some source in the stream and not the reappearance of virus in fish harboring a life-long latent infection. Persistence of IHNV by defective interfering particles Defective interfering (DI) particles may also be important in viral persistence in IHN epizootic survivors (Drolet, unpublished results). Rhabdoviral DI particles, and their roles in persistent tissue culture infections, are well established (for reviews, see Reichmann and Schnitzlein 1980; Sekellick and Marcus 1980). For IHNV, defective particles have been described in persistently infected fish cell cultures (Engelking and Leong 1981), and autointerference has been demonstrated by serial passage at high multiplicities of infection (McAllister and Pilcher 1974). Although DI particles and their role in tissue culture infection have been demonstrated for almost every virus, defective particles have not been demonstrated in animals with natural infections. Generation of DI particles. DI particles are generated by aberrant replication (for review, see Perrault 1981) which result in a shortened, defective genome. The mechanism of generation can be affected by cell type (Huang and Baltimore 1970; Holland et al. 1976), route of infection (Huang and Baltimore 1970) or multiplicity of
34 23 infection (Holland et al. 1976). The length of rhabdovirus particles is dependent upon the length of genome encapsidated. Thus, IHNV DI particles, like those of the well documented vesicular stomatitis virus, are round or short bullet-shaped. Role in disease. The production of DI and standard particles is cyclic in nature (Huang and Baltimore 1970). The synthesis of standard and DI particles may be in such balance that only a very few virions are synthesized, thus, the infection would remain subacute or latent until the balance is upset by stress or infections. The stress of spawning and the exposure of fish to IHNV and other pathogens in the stream may result in the resurgence of standard particles and thus, recurrence of infection. Inability to isolate infectious virus from non-spawning survivors may be due to population heterogeneity where excess DI particles inhibit standard virus growth and prevent the immediate cytopathic effects needed to detect virus. Treatment Virally contaminated eggs and water can be disinfected with iodine derivatives (Batts et al. 1991), but there are no antivirals or vaccines available for hatcheries to combat this economically devastating disease. It is hoped that understanding the infection process of IHNV will result in efficacious methods of treatment.
35 24 CHAPTER 3 SEROLOGICAL IDENTIFICATION OF INFECTIOUS HEMATOPOIETIC NECROSIS VIRUS IN FIXED TISSUE CULTURE CELLS BY ALKALINE PHOSPHATASE IMMUNOCYTOCHEMISTRY Barbara S. Drolet Genetics Program, Department of Microbiology Oregon State University, Corvallis Oregon John S. Rohovec and Jo-Ann C. Leong Center for Salmon Disease Research Department of Microbiology Oregon State University, Corvallis, Oregon Published: December 19, 1993 Journal of Aquatic Animal Health 5(4): Publication of this manuscript authorized by American Fisheries Society 5410 Grosvenor Lane, Suite 110, Bethesda, Maryland
36 25 Abstract An alkaline phosphatase immunocytochemical (APIC) assay was adapted for direct detection of infectious hematopoietic necrosis virus (IHNV) in infected tissue culture cells. The APIC assay provided a means of confirming the diagnosis of IHNV after the tissue culture plates had been fixed with formalin and stained with crystal violet. In the event that the original fish tissue samples had been discarded, the APIC assay was very useful and was able to detect IHNV on plates that were more than a year old. The assay used a broadly reactive monoclonal antibody (1NDW14D) to the IHNV nucleoprotein to detect viral antigen in cells infected by virus isolates representing the five known IHNV types. Likewise, the Type II-specific monoclonal antibody (2NH105B) was able to distinguish Type 11 IHNV in plaques that had been previously fixed. No cross-reactivity was seen with six other fish rhabdoviruses nor with a fish birnavirus, infectious pancreatic necrosis virus (IPNV). Immunocytochemical staining was able to distinguish between the cytopathology produced by virus infection and that induced by the toxicity of tissue samples. In the latter case, no staining was observed. The staining was not affected by the age of the fixed and dried plates nor by pre-treatment of cells with polyethylene glycol. The ease, specificity, and sensitivity of the procedure made the APIC assay an attractive alternative to the standard serum neutralization or immunofluorescent identification of IHNV for diagnostic fish disease laboratories. Introduction Infectious hematopoietic necrosis virus (IHNV) is a rhabdovirus that can cause epizootics in wild and hatchery-reared salmon and trout (for review, see Wolf 1988). Young salmonid fish are most susceptible and an IHNV outbreak can result in the complete loss of affected fish. Epizootics of IHNV are presumptively diagnosed in the field on the basis of several criteria, e.g. external disease signs, water temperature, fish
37 26 stock, geographic location, and the disease history of the stock and hatchery. Ultimately, the final diagnosis resides on laboratory confirmation which requires virus isolation and growth in tissue culture followed by serological identification of the virus (Amos 1985). The recommended procedure for the laboratory diagnosis of IHNV is the inoculation of the tissue samples onto susceptible cells followed by incubation at C for no less than 14 days. When the plaque assay method of Burke and Mulcahy (1980) is used, an incubation period of 10 days is required. If virus-induced cell lysis is observed as a plaque or as cytopathic effect (CPE), the accepted procedure for identifying the viral agent is serum neutralization (Amos 1985). However, a number of different procedures for identifying IHNV in tissue culture samples have been reported since these recommendations were drafted. Several immunofluorescence tests incorporating a direct fluorescein-conjugated anti-nucleoprotein monoclonal antibody (Ristow and Arnzen 1989) and an indirect test utilizing a monoclonal antibody to the glycoprotein of IHNV have been developed (LaPatra et al. 1989). More recently, Ristow and colleagues developed a quick direct fluorescent antibody test for IHNV that required only 30 min at 37 C for reacting with the viral antigen (Arnzen et al. 1991). The fluorescent antibody tests provide quick, accurate confirmation of IHNV in tissue culture samples. However, for many diagnostic laboratories where virus assays number in the hundreds per week and there is often no time to screen all wells and plates for viral CPE, a procedure was needed to confirm IHNV diagnosis after the plates had been fixed with formalin and stained with crystal violet. An alkaline phosphatase immunocytochemical assay (APIC) was adapted to detect IHNV antigen in fixed, stained plates. The procedure has been useful when tissue samples had been discarded or a retrospective analysis of a virus assay plate has revealed virus-like plaques and further analysis of a "plaque" was required.
38 27 Materials and Methods Viruses, Cell Lines, and Antibodies The IHNV isolates used in this study were previously described and electropherotyped by Hsu et al. (1986). Subsequently, the same isolates were characterized by monoclonal antibody reactivity (Ristow et al. 1991). The IHNV isolates used in this study were: Round Butte, 1983 (Type I); Rangen, 1983 (Type II); Elk River, 1979 (Type III); Nan Scott Lake, 1971 (Type III); Coleman Hatchery, 1980 (Type IV); Cedar River, 1981 (Type V); Karluk River, 1979 (Type V); Sacramento River chinook virus, 1969 (untyped), and Oregon sockeye virus, 1968 (untyped). The other fish rhabdoviruses that were included were viral hemorrhagic septicemia virus (VHSV), pike fry rhabdovirus (PFRV), snakehead rhabdovirus (SHRV); ulcerative disease rhabdovirus (UDRV); hirame rhabdovirus (HRV), and Rhabdovirus carpio (RC). These viruses were obtained from C. Lannan and J. L. Fryer (Mark 0. Hatfield Marine Science Center, Fish Disease Laboratory, Newport, Oregon). Infectious pancreatic necrosis virus (IPNV) of the Sp serotype obtained from P. de Kinkelin (France) was also included. The cell line, epithelioma papillosum cyprini (EPC), obtained from F. Hetrick (University of Maryland) was originally established by N. Fijan (Fijan et al. 1983). These cells were used for all rhabdovirus assays at the appropriate temperature for each virus. For the IPNV assay, chinook salmon embryo cells (CHSE-214) were used at 15 C. The original cell line was obtained from J. L. Fryer (Mark 0. Hatfield Marine Science Center, Fish Disease Laboratory, Newport, Oregon) (Fryer et al. 1965). The monoclonal antibodies were obtained from Sandra Ristow (Washington State University, Pullman, Washington). The antibody preparations consisted of the tissue culture supernatant fluid taken from hybridoma cell lines.
39 28 Virus Assays The IHNV samples from fish were part of a laboratory experiment to detect IHNV in fish during a viral epizootic. Steelhead trout (Oncorhynchus mykiss) fry weighing 1.2 g were exposed to 103 plaque forming units/ml of IHNV (Round Butte 1983, Type I) by static immersion (13 C) for 12 h at the Salmon Disease Laboratory in Corvallis, Oregon. The fish and tissue samples were processed as described (Amos 1985). Virus assays were carried out by the tissue culture infectious dose assay on 96 well tissue culture plates and by the plaque assay on 24-well tissue culture plates. Plaque assays were performed by a slightly modified procedure of Burke and Mulcahy (1980). Virus from homogenate samples was adsorbed to cells that either had or had not been pre-treated with polyethylene glycol (PEG) as recommended by Batts and Winton (1989) for 1 h at 15 C with gentle rocking. Infected cell monolayers were overlaid with 0.75% methyl cellulose in complete MEM (Eagle's minimum essential medium, Gibco BRL, Grand Island, NY). Plates were incubated for up to 14 days at 15 C. Cell monolayers were then fixed and stained with crystal violet in formalin (25% formalin, 10% ethanol, 5% acetic acid, 1% w/v crystal violet), rinsed with tap water, and allowed to dry. Alkaline phosphatase immunocytochemistry (APIC) Plaque assay plates were destained with several changes of 70% ethanol and allowed to dry. All procedures were carried out at room temperature. Non-specific binding sites were first blocked with 5% nonfat powdered milk (Saco Foods, Madison, WI) in phosphate buffered saline (PBS: 137 mm NaC1, 2.7 mm KC1, 4.3 mm NaHP03.7 H2O, 1.4 mm KHPO4), ph 7.4, with gentle rocking. The blocking solution was removed after 1 h when the murine anti-nucleocapsid protein monoclonal antibody (Mab), 1NDW14D, was used after h when the Mab 2NH105B was used (Ristow
40 29 and Arnzen 1989; Ristow et al. 1991). The antibody was adsorbed to the antigen by gently rocking the plate for 1 h. Unbound antibody was removed by two 5 min washes with PBS. The plates were stained according to instructions for alkaline phosphatase immunohistochemistry in the Vectastain-ABC mouse IgG kit of Vector Laboratory (Burlingame, CA). In brief, the bound anti-n monoclonal antibody (N-Mab) was detected by incubation with a biotinylated goat anti-mouse polyclonal antibody for 30 min with rocking. Unbound goat antibody was removed by washing with PBS as previously described and the plates were treated with phosphatase-conjugated avidin-biotin complexes for 1 h with rocking. The cells were then rinsed with PBS as previously described and incubated with 100 mm Tris-HC1, ph 8.2, for 2 min. Cells were then incubated in the dark without rocking for min with Vector-Red phosphate substrate (Vector, Burlingame, CA, kit III) containing Levamisole (Vector, Burlingame, CA). The substrate reaction was stopped with a distilled water rinse, crystal mount (Biomeda, Foster City, CA) was added, and the cells were allowed to dry. The cells were examined for red (positive) staining with an inverted microscope. The APIC assay was also tested with other IHNV isolates and rhabdoviruses. These virus isolates had been previously grown in tissue culture and the virus containing supernatant fluid had been stored at -70 C. When these assays were performed, the plaques were allowed to develop at the appropriate temperature for each virus (Kimura et al. 1986; Kasornchandra et al. 1991, 1992; Fijan et al. 1983). When IPNV was tested, the plaque assay was performed on CHSE-214 cells. These cells were overlaid with agar and incubated for 7 days at 15 C. Results and Discussion The APIC staining of IHNV plaques resulted in bright red plaques that were visible by eye. Microscopically, the APIC stained plaque appeared as an area which had
41 30 rounded positive cells surrounded by a darkly stained ring of positive cells. The adjacent negative monolayer was clear (no stain) (Figure 3.1). The darkly staining red cells enabled the examiner to detect microscopic virus plaques as early as 4 to 6 days postinfection. Individual or groups of IHNV-infected cells were detected as early as 24 h post-infection by microscopic examination. The APIC assay was specific. It distinguished plaques caused by IHNV from those caused by other fish rhabdoviruses or the fish birnavirus, IPNV. No positive staining was seen for these viruses (Table 3.1). These observations were expected since Ristow and colleagues had shown that the N-Mab reagent, 1NDW14D, recognized a group-specific epitope on the IHNV nucleoprotein. The APIC assay also distinguished IHNV Type II isolates from the other IHNV virus types with the N-Mab, 2NH105B. This N-Mab reagent recognized a type-specific epitope on the virus nucleoprotein (Ristow and Arnzen 1989). These findings indicate that the epitopes recognized by 1NDW14D and 2NH105B were not drastically altered by fixation in 25% formalin, 10% ethanol and 5% acetic acid or by washing with 70% ethanol. Ristow and Arnzen (1989) and Ristow et al. (1991) have previously demonstrated the specificity of the 1NDW14D N-Mab using fluorescence indicators and immunodot assays. The APIC procedure described here confirms their results.
42 Figure 3.1 Nucleocapsid protein of infectious hematopoietic necrosis virus (IFINV) in infected cells by alkaline phosphatase immunocytochemical staining using monoclonal antibody, 1NDW14D. IHNV-positive cells (arrows) in viral plaque (P) stained red. 31

43 Figure
44 33 Table 3.1 Virus isolates tested by alkaline phosphatase immunocytochemistry with an anti-nucleocapsid monoclonal antibody, 1NDW14D. Non-IHNV Non-Rhabdovirus IHNV Isolate Typea APICb Rhabdovirusesc APIC Pathogens APIC Round Butte I + VHSV IPNV Rangen II + PFRV Elk River III + SHRV Nan Scott III + UDRV Coleman 2 IV + HRV Cedar River V + SVCRV Karluk River V + SRCRVC Untyped + OSV Untyped + aas determined by electropherotyping (Hsu et al. 1986) b(+) indicates viral antigen detected, () indicates no viral antigen detected csee Methods for complete viral names.
45 34 No other non-specific positive reactions were seen. The toxicity of some fish tissue homogenates can cause virus-like CPE in cell monolayers and this cytopathology may be falsely interpreted as diagnostic of virus infection. This is especially true for tissue homogenates prepared from digestive organs taken from larger fish. The APIC assay did not stain cells that had been killed by toxicity (data not shown). In addition, plaques on plates up to 2 years old showed no decrease in intensity of staining. Pretreatment of cells with PEG had no apparent affect on N-Mab staining by APIC; whereas, Bootland and Leong (1992) did report that cells resuspended in PEG gave non-specific positive reactions in an IHNV coagglutination assay. Although the APIC procedure was not tested with different Mabs, the procedure could be adapted easily for detecting the IHNV glycoprotein as well. LaPatra et al. (1989) used an anti-g Mab to detect IHNV by fluorescence; the G-Mab used in that study should be able to detect IHNV by APIC if the reactive G epitope is not altered by fixation and staining. The N-Mabs were used because the IHNV nucleoprotein is the earliest and most abundantly expressed protein in the infected cell (Leong et al. 1983). Also, several reports had shown that the IHNV nucleoprotein contains conserved antigenic determinants (Hill et al. 1975; Lenoir and de Kinkelin 1975; McAllister and Wagner 1975; Leong et al. 1981; Hsu et al. 1986). Confirmatory identification of viruses by specific reaction with an antibody reagent is a standard procedure in the virus diagnostic laboratory. The immunological assay can be a virus neutralization assay (McCain et al. 1971; Hsu and Leong 1985; Winton et al. 1988) or an assay that measures the specific reactivity of an antibody with a viral antigen (for review of IHNV assays, see Winton 1991). For IHNV, this specific reactivity may be detected by direct and indirect immunofluorescence as described by Ristow et al. (1991), LaPatra et al. (1989), and JOrgensen et al. (1991). Staphylococcal coagglutination (Boot land and Leong 1992), an immunodot method (McAllister and Schill 1986; Eaton et al. 1991), and an enzyme-linked immunosorbent assay (ELISA)
46 35 (Dixon and Hill 1984) have been described using polyclonal rabbit antisera to IHNV. All of these assays have several advantages and disadvantages. The assay of choice for any laboratory will vary with the convictions and experiences of the investigators as well as their available equipment. In our experience, the APIC procedure was easy to perform, did not require special equipment, and a positive result was easy to detect. Complete plaque formation was not necessary because areas of infected cell monolayers were visible under the microscope as early as 24 h post infection. Thus, the APIC assay might be useful for early detection and identification of IHNV infection. It will probably have its greatest use with virus samples that have been lost as a result of laboratory error or a severe reduction in sample viability. It may also be used when a laboratory has to screen hundreds of samples and the examination of individual plates for plaques before fixation and staining is not possible. The assay is simple and may be adapted for early detection or confirmation of many human and veterinary viral pathogens. Acknowledgments The authors thank Sandra Ristow for providing the monoclonal antibodies 1NDW14D and 2NH105B, and IHNV Type V, and Cathy Lannan for providing several of the virus isolates used in this study. This work was supported by Bonneville Power Administration Contract DE-FG79-88BP92431, Project No ; the United States Department of Agriculture to the Western Regional Aquaculture Consortium under grant no , Project No ; United States Department of Agriculture Grants and ; and Oregon Sea Grant with funds from the National Oceanic and Atmospheric Administration, Office of Sea Grant, Department of Commerce, under grant no. NA89AA-D-SG108, Project No. R/FSD-16. Oregon Agricultural Experiment Station Technical Paper No. 10,056.
47 36 References Amos, K., editor Procedures for the detection and identification of certain fish pathogens, 3rd edition. Fish Health Section, American Fisheries Society, Corvallis, Oregon. Arnzen, J.M., S.S. Ristow, C.P. Hesson and J. Lientz Rapid fluorescent antibody tests for infectious hematopoietic necrosis virus (IHNV) utilizing monoclonal antibodies to the nucleoprotein and glycoprotein. J. Aqua. Anim. Health 3: Batts, W.N. and J.R. Winton Enhanced detection of infectious hematopoietic necrosis virus and other fish viruses by pretreatment of cell monolayers with polyethylene glycol. J. Aquat. Anim. Health 1: Boot land, L.M. and J.C. Leong Staphylococcal coagglutination, a rapid method of identifying infectious hematopoietic necrosis virus. Appl. and Environ. Microbiol. 58:6-13. Burke, J.A. and D. Mulcahy Plaguing procedure for IHNV. Appl. Environ. Microbiol. 39: Dixon, P.F. and B.J. Hill Rapid detection of fish rhabdoviruses by the enzymelinked immunosorbent assay (ELISA). Aquaculture 42:1-12. Eaton, W.D., J. Hulett, R. Brunson and K. True The first isolation in North America of infectious hematopoietic necrosis virus (IHNV) and viral hemorrhagic septicemia virus (VHSV) in coho salmon from the same watershed. J. Aquat. Anim. Health 3: Fijan, N., D. Sulimanovic, M. Bearsotti, D. Musinie, L.D. Zwillengerg, S. Chilmonczyk, J.F. Vantherot and P. de Kinkelin Some properties of the Epithelioma papulosum cyprini (EPC) cell line from carp Cyprinus carpio. Ann. Virol. 134: Fryer, J.L., A. Yusha and K.S. Pilcher The in vitro cultivation of tissue and cells of Pacific salmon and steelhead trout. Ann. N.Y. Acad. Science 126: Hill, B.J., B.O. Underwood, C.J. Smale and F. Brown Physicochemical and serological characterization of five rhabdoviruses infecting fish. J. Gen. Virol. 27: Hsu, Y.L., H.M. Engelking and J.C. Leong Occurrence of different types of infectious hematopoietic necrosis virus in fish. Appl. Environ. Microbiol. 52: Hsu, Y.L., and J.C. Leong A comparison of detection methods for infectious hematopoietic necrosis virus. J. Fish Dis. 9:
 and viral hemorrhagic septicemia (VHS): detection of trout antibodies to the causative viruses by means of plaque neutralization, immunofluorescence, and")
48 37 JOrgensen, P.E.V., N.J. Olesen, N. Lorenzen, J.R. Winton and S.S. Ristow Infectious hematopoietic necrosis (IHN) and viral hemorrhagic septicemia (VHS): detection of trout antibodies to the causative viruses by means of plaque neutralization, immunofluorescence, and enzyme-linked immunosorbent assay. J. Aquat. Anim. Health 3: Kasornchandra, J., H.M. Engelking, C.N. Lannan, J.S. Rohovec and J.L. Fryer Characteristics of three rhabdoviruses from snakehead fish Ophicephalus striatus. Dis. Aquat. Org. 13: Kasornchandra, J., C.N. Lannan, J.S. Rohovec and J.L. Fryer Characterization of a rhabdovirus isolated from the snakehead fish (Ophicephalus striatus). Proc. Sec. Int. Symp. Vir. Low. Verteb. 2: Kimura, T., M. Yoshimizu and S. Gorie A new rhabdovirus isolated in Japan from cultured hirame (Japanese flounder) Paralichthys olivaceus and ayu Plecoglossus altivelis. Dis. Aquat. Org. 1: LaPatra, S.E., K.A. Roberti, J.S. Rohovec and J L. Fryer Fluorescent antibody test for the rapid diagnosis of infectious hematopoietic necrosis. J. Aquat. Anim. Health 1: Lenoir, G. and P. de Kinkelin Fish rhabdoviruses: comparative study of protein structure. J. Virol. 16: Leong, J.C., Y.L. Hsu and H.M. Engelking Synthesis of the structural proteins of infectious hematopoietic necrosis virus, pp In: J. Crosa (ed.), Bacterial and viral diseases of fish. Molecular studies. University of Washington Press, Seattle, Washington. Leong, J.C., Y.L. Hsu, H.M. Engelking and D. Mulcahy Strains of infectious hematopoietic necrosis (IHN) virus may be identified by structural protein differences, pp In: D.P. Anderson and W. Hennessen (eds.), International Symposium on Fish Biologics: Serodiagnostics and Vaccines, Development of Biological Standards. S. Karger, Basel, Switzerland. McAllister, P.E. and W.B. Schill Immunoblot assay: a rapid and sensitive method for identification of salmonid fish viruses. J. Wildl. Dis. 22(4): McAllister, P.E. and R.R. Wagner Structural proteins of two salmonid rhabdoviruses. J. Viro1.15: McCain, B. B., J.L. Fryer and K.S. Pilcher Antigenic relationships in a group of three viruses of salmonid fish by cross-neutralization. Proceedings of the Society of Experimental Biology and Medicine 146: Ristow, S.S., and J.M. Arnzen Development of monoclonal antibodies that recognize a Type-2 specific and a common epitope on the nucleoprotein of infectious hematopoietic necrosis virus. J. Aquat. Anim. Health 1:
 and infectious hematopoietic necrosis virus (IHNV). J. Aquat. Anim. Health 3:176-180.")
49 38 Ristow, S.S., N. Lorenzen and P.E.V. JOrgensen Monoclonal antibodybased immunodot assay distinguishes between viral hemorrhagic septicemia virus (VHSV) and infectious hematopoietic necrosis virus (IHNV). J. Aquat. Anim. Health 3: Winton, J.R Recent advances in detection and control of infectious hematopoietic necrosis virus in aquaculture. Ann. Rev. Fish Dis.1: Winton, J.R., C.K. Arakawa, C.N. Lannan and J.L. Fryer Neutralizing monoclonal antibodies recognize antigenic variants among isolates of infectious hematopoietic necrosis virus. Dis. Aquat. Org.4: Wolf, K., editor Infectious hematopoietic necrosis virus, pp In : Fish viruses and fish viral diseases. Cornell University Press, Ithaca, New York.
50 39 CHAPTER 4 THE ROUTE OF ENTRY AND PROGRESSION OF INFECTIOUS HEMATOPOIETIC NECROSIS VIRUS IN ONCORHYNCHUS MYKISS (WALBAUM): A SEQUENTIAL IMMUNOHISTOCHEMICAL STUDY Barbara S. Drolet Genetics Program, Department of Microbiology Oregon State University, Corvallis Oregon John S. Rohovec and Jo-Ann C. Leong Center for Salmon Disease Research Department of Microbiology Oregon State University, Corvallis, Oregon Published: July, 1994 Journal of Fish Diseases 17(4): Publication of this manuscript authorized by Blackwell Scientific Publications Ltd., Osney Mead, Oxford, OX2 OEL.
51 40 Abstract Steelhead trout (Oncorhynchus mykiss) fry were experimentally infected with infectious hematopoietic necrosis virus (IHNV) Round Butte 1983 (Type 1). Fry were sampled daily, before and during the epizootic. Fish tissues were tested for infectious virus by tissue culture assay and for IHNV nucleocapsid protein by alkaline phosphatase immunohistochemistry (APIH). The progression of virus through the tissues was followed by APIH until the 14th day. Viral infection progressed from two major sites, from the gills into the circulatory system, and from the oral region into the gastrointestinal tract and then into the circulatory system. Once in the blood, virus was disseminated to virtually every organ. Progression of IHNV within and between organs is discussed. Introduction Infectious hematopoietic necrosis virus (IHNV) causes an acute, systemic disease in trout and salmon. The complete infection cycle of IHNV in fish has not been determined. It is known that young fish in the wild, or more commonly in aquaculture, may be lethally infected with IHNV. Virus can be isolated from both viscera and mucus using established tissue culture methods (Burke and Mulcahy 1980; Hsu and Leong 1985; Winton 1988; LaPatra et al. 1989a; LaPatra et al. 1989b). Many questions remain concerning the route of viral entry. Vertical transmission of IHNV is controversial; however, waterborne or horizontal transmission has been reported for several species of salmon and trout, and is used routinely in artificial infection experiments. Several infection studies have implicated the gills as the initial site of IHNV entry and infection. Infectious hematopoietic necrosis virus has been isolated from gill tissue of adult kokanee and sockeye at 3 days post exposure (dpe). Direct staining of viral glycoprotein by immunoperoxidase histochemistry was
52 41 also made at this time (Yamamoto et al. 1989). Virus has also been isolated by tissue culture in gill tissue early in the infection of sockeye fry (Mulcahy et al. 1983) and smolts (Burke and Grischkowsky 1984). Virus has been detected in gill tissue of rainbow trout at hours post exposure (hpe) by electron microscopy and at 24 hpe by tissue culture methods (Yamamoto and Clermont 1990), but not until 5 dpe by immunohistochemistry (Yamamoto et al. 1990). Yamamoto et al. (1990) also reported that the skin was the major site of IHNV replication and that the gill played a lesser role in the initial stages of infection. Gills have been shown to be the initial site of replication for two other fish rhabdoviruses, spring viremia of carp virus (Ahne 1978) and viral hemorrhagic septicemia virus (Neukirch 1984). Progression of IHNV infection through the fish is unknown. It has been suggested that IIINV spreads from the gills to other organs via the blood. Initial infection of the gill followed by viremia was reported by Yasutake and Amend (1972). Mulcahy et al. (1983) reported that a titer 1 x 105 plaque forming units per gram (pfu/g) was required in the gills of artificially infected sockeye before virus was seen in other organs. In the same report, however; infectious virus was detected in the viscera of naturally infected spawning sockeye in the absence of virus in the gills. Moreover, virus has been detected by immunohistochemistry (Yamamoto et al. 1990) and by electron microscopy (Yamamoto and Clermont 1990) in the visceral organs of rainbow trout before the gills. This suggests an alternate or additional origin of virus infection. We report the results of an experiment designed to demonstrate the route(s) of initial viral entry and the progression of LHNV through fish tissues. An alkaline phosphatase immunohistochemical assay (APIH) was used to detect the nucleocapsid protein of IHNV in formalin- fixed, paraffin-embedded lg steelhead trout, Oncorhynchus mykiss, which were sampled before and during an artificially induced IHN epizootic. The nucleocapsid protein was the antigen of choice because it is the most abundant, conserved viral antigen among the five known IHNV types (Hill
53 42 et al. 1975; Lenoir and de Kinkelin 1975; McAllister and Wagner 1975; Leong et al. 1981; Hsu et al. 1986). With this assay, we were able to examine viral pathogenesis in very small tissue samples, in samples with low concentrations of virus, and in specific cell types of each tissue. Materials and Methods Infection One thousand steelhead fry, Oncorhynchus mykiss, 1 g were exposed to 103 pfu/ml IHNV (Round Butte, Type I) by static immersion (13 C) for 12 h at the Salmon Disease Laboratory, Corvallis, OR. Asymptomatic fry were taken during the preepizootic period (0-5 dpe), and moribund and dead fry were taken during the epizootic period (6-14 dpe). Three fish were homogenized for tissue culture and three were fixed for APIH daily. Cell culture Epithelioma papillosum cyprini (EPC) cells (Fijan et al. 1983) were grown to confluency in 24-well plates with MEM containing 10% fetal bovine serum, 2% penicillin-streptomycin (lunit/ug), 2% 200 mm L-glutamine, 2% fungizone, 1% gentamycin (50 mg/ml), buffered with 1M Tris buffer to ph 7.0. For viral assays, cell monolayers were inoculated with 0.1 ml of serially diluted samples. Tissue, mucus samples Whole steelhead fry were diluted 1:10 (w/v) in Hanks' balanced salt solution (Gibco BRL, Gaithersberg, MD) and homogenized in a Stomacher 80 homogenizer
54 43 (Tekmar, Cincinnati, OH). The homogenate was centrifuged at low speed for 10 min to sediment tissue debris. The supernatant was diluted 1:1 (v/v) in Eagle's minimum essential media (MEM; Gibco BRL) containing antibiotic-antimycotic (10 µg/ml penicillin G sodium, 10 µg/m1 streptomycin sulfate and 25 µg/ml amphotericin B as fungizone in 0.85% saline, Gibco BRL) and stored at 4 C overnight (Amend and Pietsch 1972). Mucus samples from infected fish were taken with cotton swabs and put directly in the antibiotic media and stored at 4 C overnight (LaPatra 1989). Plaque assay Virus from tissue and mucus samples was adsorbed to cells for 1 h at 15 C with gentle rocking. Mono layers were overlaid with 0.75% methyl cellulose (in complete MEM) and incubated for up to 14 days at 15 C. Cells were fixed and stained (25% formalin, 10% ethanol, 5% acetic acid, 1% crystal violet), rinsed with tap water and allowed to dry. Plaques were counted and virus titers were calculated as pfu per mucus swab or pfu/g of tissue. Fixation and embedding of fish for APIH Fish were fixed in buffered formalin (10% formalin, 33 mm sodium phosphate monohydrate, 46 mm sodium phosphate dibasic), dehydrated in an alcohol-xylene series and embedded in Paraplast paraffin (60 C) (Oxford Labware, St. Louis, MO). Blocks were sectioned sagittally (6 gm) and fixed (40 C overnight) to coated slides (0.1% gelatin, 0.1% chromic potassium sulfate).
55 44 Alkaline phosphatase immunohistochemistry (APIH) All reagents for the APIH assay were diluted in 5.8 mm phosphate buffered saline, ph 7.4 (PBS) and excess reagent was removed by rinsing with PBS between incubations. All incubations were done at room temperature unless otherwise indicated. Tissue sections were deparaffinized by standard methods and rehydrated for 10 min in distilled water and 20 min in PBS. Nonspecific binding sites were blocked for 1 h with 5% nonfat powdered milk (Saco Foods, Madison, WI) in PBS. Blocking solution was blotted off and sections were incubated for 1 h with a universal antinucleocapsid monoclonal antibody (1NDW14D) which is specific for a highly conserved region of the nucleocapsid protein and has been shown by direct immunofluorescence and immunoblotting to recognize all five IHNV types (Ristow and Arnzen 1989; Ristow et al. 1991). Slides were then incubated for 30 min with biotinylated goat anti-mouse polyclonal antibody, followed by a 1 h incubation with phosphatase-conjugated avidin-biotin complex (Vectastain-ABC mouse IgG kit, Vector Laboratories, Burlingame, CA). Sections were equilibrated with 100 mm Tris-HC1, ph 8.2 (substrate buffer), and incubated with Vector-Red phosphate substrate (Vector) for min in the dark. Levamisole (Vector) was added to the substrate solution to inhibit endogenous phosphatase activity. The substrate reaction was stopped with distilled water and sections were counterstained in hematoxylin for 1-2 min and 0.2% ammonium hydroxide in 70% ethanol solution for 30 s. Sections were covered with Crystal Mount (Biomeda, Foster City, CA), Cytoseal mounting media (Stephens Scientific, Riverdale, NJ) and a coverslip and examined by light microscopy.
56 45 Alkaline phosphatase immunocytochemistry (APIC) Infectious hematopoietic necrosis virus from tissue homogenates was confirmed in viral plaques by APIC (Drolet, et al. 1993). Briefly, formalin-fixed, crystal violetstained cell monolayers were destained with 70% ethanol and cells were incubated with the same reagents and antibodies as described for APIH. Results Infection Deaths in the infected population started 5 dpe, peaked at 6-14 dpe, then decreased with time (Figure 4.1). Cumulative percent mortality at 40 dpe was 42%. Clinical signs of the IHNV infection included distended abdomens, petechial hemorrhages on ventral surface and near the base of fins, darkened body coloration and exophthalmia. Behavioral signs included lethargy interspersed with frenzied swimming, vertical drifting, whirling and anorexia. Tissue culture assays Virus was first detected by plaque assay in both mucus and whole fish homogenates at 5 dpe. During the epizootic (6-14 dpe), swab virus titers ranged from 1x104 to 4x108 pfu per mucus swab and tissue homogenate titers ranged from 4x104 to 5x109 pfu/g. Cells at the periphery of plaques stained positive (red) for IHNV by APIC. Infectious virus was not detected in mucus or tissue homogenates after 54 dpe.
57 46 Detection of IHNV by APIH The degree of IHNV infection in individual cells of tissues was rated by the intensity of staining on a scale of 1 to 5. For those tissues that were intact and stained a light red or pink, the rating of 1-2 was used and interpreted to indicate either early stages of infection or transient infection. Intact tissues that were stained a bright red indicated a productive infection and were given ratings of 3-4. The tissues that were stained bright red and showed obvious signs of damage were given a rating of 5. The relationship between intensity of staining and degree of viral infection was also observed in the immunocytochemical staining of viral plaques (Drolet et al. 1993). Pre- Epizootic Epizootic Post Epizootic 40 co 30 - C 715 a) CI 20 O co 10" 0 I. I Days Post Exposure Figure 4.1 Mortality curve of 1 g steelhead trout (0. mykiss) after immersion infection with infectious hematopoietic necrosis virus, type 1. The pre-epizootic period is from day 0-5 and the epizootic period is from day 6-14.
58 47 Pre-epizootic period Circulatory pathway. Sequential progression of virus to the organs via the circulatory system is shown in Table 4.1. Epithelial cells of gill lamellae were positive by day 2 (Figure 4.2 A). In the kidney, virus was also first seen at day 2 in the anterior portion in the lumen and epithelial cells of the renal tubules (Figure 4.2 C). The surrounding hematopoietic tissue was negative until day 4 (Figure 4.2 D). The Hassel's corpuscles (thymic bodies) of the thymus were positive by 3 dpe, and in the spleen, the connective tissue stroma throughout the organ was positive at 5 dpe, but the hematopoietic cells were negative. In the pancreas, the supporting stroma was positive, and the acinar cells were negative. Muscle cells of the heart ventricle were positive on day 4. In the brain, virus was detected in the meninges of the forebrain, optic lobe, and cerebellum by day 5. Gastrointestinal pathway. The sequential progression of virus in gastrointestinal organs is also shown in Table 4.1. Virus-positive red staining was first observed from 2-4 dpe in the epithelial cells of the oral region, pharynx, esophagus, stomach, pyloric caeca (Figure 4.2 E) and intestine. Beginning on day 5, the sub-mucosal and lamina propria layers of these organs (Figure 4.2f) were positive and the epithelial cells were negative. Virus was first detected in liver hepatocytes on day 3. Other tissues. Epithelial cells of the ventral epidermis and head region were weakly positive 1 dpe. Muscle cells consistently gave low background staining and were not considered positive unless particulate dark red staining could be seen in the connective tissue surrounding the skeletal muscle. By 3 dpe, positive cartilage was seen in the head region and the connective tissue surrounding adipose cells ofthe abdomen was positive.
59 48 Table 4.1 Chronological appearance and staining intensity of infectious hematopoietic necrosis virus by alkaline phosphatase immunohistochemistry in steelhead (0. mykiss) fry. Pre-Epizootic Epizootic Days Post Exposure # Sections Examined Pathway: Circulatory: Gills NN Anterior Kidney NN Posterior Kidney NNN Thymus NN N N Spleen NNNNN Pancreas NNNNN Heart NNNNN Brain NNNNN Gastrointestinal: Oral region NN Pharynx NN Esophagus NN Stomach NN Pyloric Caeca NN Liver NNN Intestine NNNN Other: Skin Ni Muscle NNN Cartilage NNN Adipose NNN N * Relative intensity of positive staining; 1 = weak, 5 = strong, N = no vir us detected.
60 49 Figure 4.2 Photomicrographs of tissues from infectious hematopoietic necrosis virus infected steelhead fry during the pre-epizootic period (0-5 days post exposure) and the epizootic period (6-14 days post exposure). A positive reaction is indicated by arrows and red staining cells. (A) Preepizootic gill with virus-positive lamellar and filament epithelial cells (ep) (40x). (B) Epizootic gill with virus-positive lamellar and filament endothelium (en) (40x). (C) Pre-epizootic anterior kidney with viruspositive tubule (t) and negative hematopoietic cells (40x). (D) Epizootic anterior kidney with negative tubule and virus-positive hematopoietic cells (h) (40x). (E) Pre-epizootic pyloric caeca with virus-positive columnar epithelial cells (ep) (40x). (F) Epizootic intestine with negative epithelia and virus-positive lamina propria (1p) (40x). (G) Epizootic brain with virus-positive glial (g) cells in the granular layer of the optic lobe (100x). (H) Epizootic eye with viruspositive retina (r) (10x).
61
62 51 Circulatory pathway. In the gill, the lamellar vessel endothelia and the connective tissue of the filament were strongly positive, whereas the epithelial cells were negative (Figure 4.2 B) during the epizootic. In the kidney, epithelial cells of the anterior renal tubules were necrotic with positive tubule basement membranes and strongly positive, necrotic hematopoietic tissue (Figure 4.2 D). The posterior kidney showed the same distribution of positive staining; however, the negative epithelial cells of the tubules remained intact and the hematopoietic tissue was strongly positive, but less necrotic. Virus-positive cells in hemorrhaged blood pools were primarily macrophage, with little or no red blood cell involvement. The Hassel's corpuscles of the thymus were positive throughout the epizootic and in the spleen, there was a diffuse positive reaction which spared the majority of lymphocytes. In the pancreas, the supporting stroma was positive and the acinar cells were positive and necrotic. In at least 33% of the fish sampled on each day of the epizootic, virus was detected in glial cells in localized regions of the forebrain, optic lobe (Figure 4.2 G), cerebellum, medulla oblongata and spinal cord and in the retinal cells of the eye (Figure 4.2 H). Gastrointestinal pathway. During the epizootic, the epithelial cells of the oral region, pharynx, esophagus, stomach, pyloric caeca and intestine were negative, but the submucosal and loose connective tissue (lamina propria) of these organs was strongly positive (Table 4.1). The mucosal epithelial cells located at the tips of villi in the pyloric caeca and intestine were often destroyed and infected cell debris could be seen in the lumen (Figure 4.2 F). Epithelial cells at the base of the villi were negative and intact. The basally adjacent submucosa and lamina propria were strongly positive, as were specific areas of muscle of the cardiac and pyloric stomach. Distinct foci of viruspositive hepatocytes were seen throughout the liver.
63 52 Other tissues. During the epizootic, Positive epidermis was primarily seen on the ventral surface and rarely on the head or dorsal surface. Cross section of muscle cells showed positive staining on the inside of the cells as well as in the connective tissue surrounding the cell. Positive regions of cartilage were seen in the head and along the spine. Positive staining was seen in the connective tissue surrounding adipose cells of the abdomen. Discussion The results of this study suggest that IHNV infection progressed from two major sites: the gills and the gastrointestinal tract. Both routes of entry eventually resulted in systemic viremia. While IHNV readily infected epithelial cells of numerous organs, it had a propensity for connective tissue. Included in this broad category were the supportive stroma of the spleen, pancreas, adipose and muscle; the submucosa of the oral cavity, pharynx, esophagus and stomach; the lamina propria of the stomach, pyloric caeca and intestine; and the meninges of the brain. Epithelial cells in virtually every organ were virus-positive and intact during the pre-epizootic period and virus negative and intact during the epizootic period. This indicated a short lived or transient infection. Results of daily samples indicate that the epithelial cells had changed from positive to negative and not that negative epithelial cells were replacements for those cells that had been previously lysed. Specific areas of the pyloric caeca and intestine which had cells that were virus-positive and undergoing lysis during the pre-epizootic period were the same areas where only virus-positive cell debris remained, suggesting lysed cells were not replaced with new negative cells. We suggest that ingested virus transiently infected epithelial cells lining the oral cavity, pharynx, esophagus, stomach, pyloric caeca and intestine; and then productively infected the basally adjacent connective tissue (submucosa and/or lamina propria) of each organ (Table 4.2). Virus
64 53 produced in these highly vascularized connective tissues would have been collected by the abdominal vein and transported to the heart for systemic dissemination. The virus-positive staining was strongest and most persistent in the kidney. Results of the sequential sampling indicate that the virus infecting the kidney, probably originated at the gills. Both the proposed circulatory and ingestion pathways resulted in viremia; however, the circulatory pathway is the most direct route to the kidney. We propose that initial infection of the kidney began with the virus infecting the gill epithelium, then infecting the lamellar and filament endothelium, and being carried by the blood through the dorsal aorta and reaching the kidney tissue via the renal arteries. Positive staining of the kidney tubules prior to infection of vascularized tissues of the gut support this hypothesis. A few strongly positive red blood cells and macrophagelike cells were observed and may indicate that the virus replicated during its transportation. Virally contaminated blood may have been responsible for pre-epizootic infections of the thymus (via the thymic artery), the stroma of the spleen (via the splenic artery), and pancreas (via the parietal artery) and the meninges of the brain (via the circle of Willis). Virus was detected in the anterior kidney one day earlier than in the posterior kidney. This anterior to posterior sequence was also reported by Yasutake and Amend (1972) in histopathological evaluation of IHNV infected sockeye salmon. The APIH results indicate that the kidney was infected by virus from the capillaries surrounding the renal tubules. Virus entered the tubules, transiently infected the epithelial cells, budded out of the basement membrane and productively infected the basally adjacent hematopoietic tissue. A large amount of virus was produced throughout the hematopoietic tissue of the kidney, contributing the majority of infected blood to be disseminated systemically.
65 54 Table 4.2 Virus-positive tissues of fish organs as tested by alkaline phosphatase immunohistochemistry. Pre-epizootic to epizootic progression of virus in specific tissues of each organ examined in Table 4.1. ORGAN Circulatory: Gills Anterior kidney Posterior kidney Thymus Spleen Pancreas Heart Brain Gastrointestinal: Oral region Pharynx Esophagus Stomach Pyloric caeca Liver Intestine Other Skin Muscle Cartilage Adipose PRE-EPIZOOTIC Lamellar epithelium Tubule lumen, epithelium Tubule lumen, epithelium Hassel's corpuscles Connective stroma Connective stroma Ventricle musculature Meninges Mucosal epithelium Mucosal epithelium Mucosal epithelium Lumen, epithelium Lumen, epithelium Hepatocytes Lumen, epithelium Ventral Ventral Head Connective stroma EPIZOOTIC Capillary endothelium Hematopoietic cells Hematopoietic cells Hassel's corpuscles Hematopoietic cells Acinar cells Ventricle musculature Meninges and glial cells Submucosa Submucosa Submucosa Lamina propria, muscle Lamina propria Hepatocytes Lamina propria Anterior ventral, dorsal Anterior, ventral, eye Head, spine Connective stroma
66 55 Yamamoto et al. (1990) reported that skin was the major site of IHNV replication and that gills played a lesser role in the initial stages of infection. In our study, the epithelial cells of the ventral and anterior epidermis were weakly positive by APIH at 1 dpe (one fish), but the location of positive staining in the skin varied greatly among all 18 fish sampled during the pre-epizootic period. Throughout both the preepizootic and epizootic periods, the intensity of the positive signal in the skin was more indicative of a transient infection than a productive infection. During the epizootic, ventral muscle was often positive while the adjacent skin epithelium was negative. A consistent progression of infection from the skin to adjacent muscle or other organs was not seen in the sequential samples examined. Although not the source of initial infection of the kidney, the ingestion or gastrointestinal pathway was a highly significant route of infection. All fish sampled from day 2 to day 4 had virus-positive staining in the lumen and regions of epithelial cells lining the gastrointestinal tract. From 5 to 14 dpe the highly vascularized lamina propria was strongly positive in all gastrointestinal organs. The most distinguishable and consistent organ to organ progression of virus was from the stomach to the pyloric caeca. The strongest positive staining and the most cell destruction was observed nearest the stomach. Both the signal and the destruction decreased as one progressed down to the tip of the caeca. The most restricted infection was in the brain. During the epizootic, the distinct positive foci were seen primarily in the granular layers of the lobes and showed no apparent pattern of infection. Infection of the retinal cells of the eye was always near the layer of connective tissue called the falciform process. The resulting inflammation may be in part the cause of exophthalmia, a prominent symptom of NIN disease.
67 56 Results of the immunohistochemical method used in this study suggest that IHNV exhibits a specific tissue tropism for connective tissue. While IHNV readily infected epithelial cells, the infection appeared transient and not highly productive. The majority of cells undergoing productive viral infection were of connective tissue origin. These results may influence the design of vaccines, and the study of specific cell receptors involved in IHNV infection. Post-epizootic samples are currently being examined by APIH to determine the mechanism of viral clearing and the existence of viral latency. Acknowledgments Sandra Ristow (Washington State University, Pullman, Washington) provided the monoclonal antibody used in this study. Jerry Heidel (Oregon State University, Veterinary Medicine, Corvallis, Oregon) gave invaluable histopathology evaluation. Others involved in the project were John Fryer, Linda Boot land, Harriet Lorz and Pin Wen Chiou. This study was supported by Bonneville Power Administration, project number DE-FG79-88BP92431, Project No ; the United States Department of Agriculture to the Western Regional Aquaculture Consortium under grant no , Project No ; United States Department of Agriculture grants and ; and Oregon Sea Grant with funds from the National Oceanic and Atmospheric Administration, Office of Sea Grant, Department of Commerce, under grant no. NA89AA-D-SG108, Project No. R/FSD-16. Oregon Agricultural Experiment Station Technical Paper No.10,303.
68 57 References Ahne, W Uptake and multiplication of spring viraemia of carp virus in carp, Cyprinus carpio L. J. Fish Dis. 1: Amend, D.F. and J.P. Pietsch An improved method for isolating virus from asymptomatic carrier fish. Trans. Amer. Fish. Soc. 101: Burke, J.A. and R. Grischkowsky An epizootic caused by infectious hematopoietic necrosis virus in an enhanced population of sockeye salmon, Oncorhynchus nerka (Walbaum), smolts at Hidden Creek, Alaska. J. Fish Dis. 7: Burke, J.A. and D. Mulcahy Plaguing procedure for infectious hematopoietic necrosis virus. Appl. Environ. Microbiol. 39: Drolet, B.S., J.S. Rohovec and J.C. Leong Serological identification of infectious hematopoietic necrosis virus in fixed tissue culture cells by alkaline phosphatase immunocytochemistry. J. Aquat. Anim. Health 5(4): Fijan, N., D. Sulimanovic, M. Bearsotti, D. Musinie, L.D. Zwillengerg, S. Chilmonczyk, J.F. Vantherot, and P. de Kinkelin Some properties of the Epithelioma papulosum cyprini (EPC) cell line from carp Cyprinus carpio. Ann. Virol. 134: Hill, B.J., B.O. Underwood, C.J. Smale and F. Brown Physicochemical and serological characterization of five rhabdoviruses infecting fish. J. Gen. Virol. 27: Hsu, Y.L., H.M. Engelking and J.C. Leong Occurrence of different types of infectious hematopoietic necrosis virus in fish. Appl. Environ. Microbiol. 52: Hsu, Y.L. and J.C. Leong A comparison of detection methods for infectious hematopoietic necrosis virus. J. Fish Dis. 9: LaPatra, S.E Strain differentiation and detection of infectious hematopoietic necrosis virus, pp In: Ph.D. thesis, Oregon State University, Corvallis, Oregon. LaPatra, S.E., K.A. Roberti, J.S. Rohovec and J.L. Fryer. 1989a. Fluorescent antibody test for the rapid diagnosis of infectious hematopoietic necrosis. J. Aquat. Anim. Health 1: LaPatra, S.E., J.S. Rohovec and J.L. Fryer. 1989b. Detection of infectious hematopoietic necrosis virus in fish mucus. Fish Pathol. 24: Lenoir, G. and P. de Kinkelin Fish rhabdoviruses: Comparative study of protein structure. J. Virol. 16:
69 58 Leong, J.C., Y.L. Hsu, H.M. Engelking and D. Mulcahy Strains of infectious hematopoietic necrosis (IEN) virus may be identified by structural protein differences, pp In: D.A. Anderson and W. Hennessen (eds.), International Symposium on Fish Biologics: Serodiagnostics and Vaccines, Development of Biological Standards. S. Karger, Basel, Switzerland. McAllister, P.E. and R.R. Wagner Structural proteins of two salmonid rhabdoviruses. J. Virol. 15: Mulcahy, D., R.J. Pascho and C.K. Jenes Detection of infectious hematopoietic necrosis virus in river water and demonstration of waterborne transmission. J. Fish Dis. 6: Neukirch, M An experimental study of the entry and multiplication of viral hemorrhagic septicaemia virus in rainbow trout, Sa lmo gairdneri Richardson, after water-borne infection. J. Fish Dis. 7: Ristow, S.S. and J.M. Arnzen, Development of monoclonal antibodies that recognize a Type-2 specific and a common epitope on the nucleoprotein of infectious hematopoietic necrosis virus. J. Aquat. Anim. Health 1: Ristow, S.S., N. Lorenzen and P.E.V. Jorgensen Monoclonal antibody-based immunodot assay distinguishes between viral hemorrhagic septicemia virus (VHSV) and infectious hematopoietic necrosis virus (IHNV). J. Aquat. Anim. Health 3: Winton, J.R., C.K. Arakawa, C.N. Lannan and J.L. Fryer Neutralizing monoclonal virus. Dis. Aquat. Org. 4: Yamamoto, T., W.N. Batts, C.K. Arakawa and J.R. Winton Comparison of infectious hematopoietic necrosis in natural and experimental infections of spawning salmonids by infectivity and immunohistochemistry, pp In: W. Ahne and E. Kurstak (eds.), Viruses of Lower Vertebrates. Springer-Verlag, Berlin. Yamamoto, T., W.N. Batts, C.K. Arakawa and J.R. Winton Multiplication of infectious hematopoietic necrosis virus in rainbow trout following immersion infection: whole body assay and immunohistochemistry. J. Aquat. Anim. Health 2: Yamamoto, T. and T.J. Clermont Multiplication of infectious hematopoietic necrosis virus in rainbow trout following immersion infection: organ assay and electron microscopy. J. Aquat. Anim. Health 2: Yasutake, W.T. and D.F. Amend Some aspects of pathogenesis of infectious hematopoietic necrosis (IHN). J. Fish Biol. 4:
70 59 CHAPTER 5 PERSISTENCE OF AN RNA VIRUS INFECTION BY DEFECTIVE INTERFERING PARTICLES Barbara S. Drolet Genetics Program, Department of Microbiology Oregon State University, Corvallis Oregon Pinwen P. Chiou Department of Microbiology Oregon State University, Corvallis Oregon Jerry Heidel College of Veterinary Medicine Oregon State University, Corvallis, Oregon Jo-Ann C. Leong Center for Salmon Disease Research Department of Microbiology Oregon State University, Corvallis, Oregon Manuscript in preparation for submission to Journal of Virology
71 60 Abstract Infectious hematopoietic necrosis virus (IHNV) is a rhabdovirus which causes devastating epizootics of trout and salmon fry in hatcheries around the world. Epizootic survivors are negative for infectious virus by plaque assay at about 50 days post exposure. Survivors are considered to be virus-free and suffering no ill effects of the viral infection upon subsequent release into the wild. When adults return to spawn, infectious virus can again be isolated by plaque assay. Two hypotheses have been proposed to account for the source of virus in these adults. One hypothesis contends that virus in the epizootic survivors is cleared and that the adults are being reinfected by IHNV during their migration upstream. The second hypothesis contends that IHNV persists in the survivor in a subclinical or latent form, then reactivates during the stressful conditions of spawning. After 20 years of experiments and debate, there is no conclusive evidence to support either of these hypotheses. In this paper, we describe lesions in hematopoietic tissue of rainbow trout, one year post exposure, and provide the first irrefutable evidence for subclinical persistence of virus in tissues of IHN survivors. We demonstrate the presence of viral protein by immunohistochemistry, viral RNA by PCR amplification, and IHNV defective interfering particles by immunogold electron microscopy. Introduction The rhabdovirus infectious hematopoietic necrosis virus (IHNV) causes devastating epizootics in hatchery-reared trout and salmon species of the Pacific Northwest, Europe and Asia. IHNV primarily infects fry and can result in 100% mortality. The main target of IHNV is the hematopoietic organ of the fish, the kidney. Infectious virus can be detected in the viscera of infected fry by plaque assay until approximately 50 days post exposure (dpe) when the virus appears to clear. It is
72 61 believed that epizootic survivors remain negative for infectious virus until sexual maturity when virus can again be detected by plaque assay. It is not known whether infectious virus isolated from spawning adults is the result of reinfection from secondary reservoirs during their migration upstream, or the reactivation of a latent form of IHNV. With the exception of one study (Amend 1975), the inability to detect "reactivated" latent virus in numerous, carefully monitored, naturally and experimentally infected fish populations has resulted in a majority of evidence against latency. The reinfection theory has not been overwhelmingly accepted, however, because large amounts of virus are required to infect large fish, no secondary viral reservoirs have been found for IHNV, and adult infection studies have shown that IHNV survivors are less susceptible than naive controls to reinfection by IHNV upon sexual maturation (unpublished results). The argument against persistence of IHNV has been based on the continued negative results of plaque assays on IHN survivors held in captivity. This assay detects only infectious virus and routinely has a limit of detection of 200 pfu/g tissue. The possibility of viral persistence, at a level below the detection limit of plaque assays, would help explain the resurgence of virus in adult populations where reinfection from an unknown secondary reservoir seems unlikely as the sole source of virus. The persistent rhabdoviral infections of vesicular stomatitis virus (VSV) and rabies virus have been extensively studied and explained by defective interfering (DI) particles (Holland and Villareal 1974; Kawai et al. 1975; Ramseur and Friedman 1978; Horodyski et al. 1983). Huang and Baltimore (1970) were the first to report DI particles, and proposed that their action was to modulate the yield of infectious virus in a cyclical manner. Over the years, several attempts have been made to elucidate a role for the seemingly ubiquitous DI particles of viral infections (For review, see Holland 1987; Barrett and Dimmock 1986; Lazzarini et al. 1981; Perrault 1981). VSV DI particles have been shown to play an important role in relative virulence of different
73 62 serotypes and in the establishment of persistent infections in cell culture. DI particles of many viruses have been shown to alter the pathogenesis and course of disease in cell culture, and experimental animals given DI particles before or during an infection (Bangham and Kirkwood 1990; Browning et al. 1991; Moscona and Peluso 1993) show an altered disease course. Yet, defective virus has not been confirmed in natural infections in animals and often cannot be reisolated from animals where they have been experimentally administered to examine virus replication effects. Two studies have come close to detecting DI particles in natural infections. Pedley et al. (1984) isolated rotaviruses with altered genomes from chronically infected immunodeficient children, but the particles were not confirmed due to the difficulty in culturing the rotavirus. Bean et al. (1985) found "DI-type" RNAs in an avirulent strain of type A influenza virus after they had been passed in embryonated eggs twice. Although it explained the virulence difference, the possibility that the DI-type RNA was made in the eggs was never ruled out. IHNV DI particles have been described in persistently infected fish (McAllister and Pilcher 1974; Engelking and Leong 1981) and insect cell cultures (Scott et al. 1980). These particles caused the establishment of persistent cell culture infections and inhibited subsequent viral infection in susceptible fish. It is possible that IHN disease is modulated by DI particles in natural infections and this may explain the apparent disappearance and reappearance of the virus during the life cycle of its host. Although IHN epizootic survivors have not been closely monitored in the wild, they are assumed to be virus-free and suffering no ill effects of the viral infection upon release. In this paper we describe the lesions found in the kidney of rainbow trout one year after exposure to IHNV and the evidence for persistence of virus in these survivors as shown by immunohistochemistry, PCR amplification, and immunogold electron microscopy.
74 63 Materials and Methods Fish infection and sampling Thirty rainbow trout (Oncorhynchus mykiss), which had been exposed to 103 pfu/ml (LD50) of IHNV (Rangen, type-2) as fry by static immersion were maintained in pathogen-free water at the Center for Salmon Disease Research fish facility (Corvallis, Oregon). IHN epizootic survivors were lethally sampled at 90 d, 180 d, 1 y and 2 y post exposure (ype). At each sampling time, pieces of kidney, spleen, and liver tissues were pooled for each fish and tested for infectious virus by plaque assay as suggested in the Fish Health Blue Book (Amos 1985) on EPC cells (Fijan et al. 1983). In addition, pieces of 11 different organs from each fish were fixed in 10% neutral buffered formalin (10% formalin, 33 mm sodium phosphate monohydrate, 46 mm sodium phosphate dibasic), dehydrated in an alcohol-xylene series and embedded in Paraplast paraffin (60 C) (Oxford Labware) for immunohistochemistry. Mock-infected fish, which were exposed to non-infected cell culture media, were used as negative controls. For elucidation of EM immunogold staining parameters, fresh positive and negative tissue was used from a second groups of fish which were similarly infected. Fresh, epizootic kidney tissues were cut into 1-2 mm3 blocks, fixed in paraformaldehyde/gluteraldehyde (2% paraformaldehyde, 1% gluteraldehyde in 0.1 M sodium cacodylate buffer) (Williams 1985), dehydrated with an ethanol series and embedded in LR White resin (Ted Pella, Inc.). Kidney tissue from 1 ype survivors which had been fixed in formalin for two years, but never embedded in paraffin were treated as fresh tissue. Kidney tissue from 1 ype survivors which had been previously embedded in paraffin were cut into blocks, deparaffinized and rehydrated in a xylene/ethanol series, and treated as fresh tissue. Ultrathin sections of these tissues were collected on nickel or gold grids (Ted Pella, Inc.).
75 64 Histology Several histological stains were used to examine 7 gm paraffin sections of kidney from 1 ype survivors. Sections were stained with hematoxylin and eosin stain (Lillie and Fullmer 1976; Gill et al. 1974), Masson's trichrome stain (Masson 1929), periodic acid-schiff (PAS) stain (McManus 1948), PAS stain with a diastase predigestion and a PAS stain with phenyl hydrazine HC1 (Leppi and Spicer 1967), mucicarmine stain (Mallory 1961), Perl's iron stain (Highman 1942), and Dunn- Thompson hemoglobin stain (Dunn and Thompson 1945). Immunohistochemistry Immunohistochemistry was done as described by Drolet et al. (1994). Briefly, 7 p.m paraffin-embedded sections were deparaffinized, hydrated, blocked with 5% nonfat milk and incubated with a monoclonal antibody (Mab; 1NDW14D) made to a conserved region of the highly conserved nucleocapsid (N) protein of IHNV. Sections were then incubated with a biotinylated goat anti-mouse antibody, phosphatase conjugated avidin-biotin complexes, and a red phosphate substrate (Vector). Sections were then counterstained with hematoxylin for 1 min and 0.2 % ammonium hydroxide for 1 min. Mock-infected fish tissues were used as negative controls and 8-10 day post exposure infected fish were used as positive controls. Immunogold staining All incubations were done in 0.5 ml microfuge tubes with volumes of 25 IA and all rinses were done in disposable glass dilution tubes. All reagents were filtered through a 0.2 j.tm Acrodisc membrane (Gelman Sciences). Grids were blocked with 10% normal goat serum in Tris buffer (20 mm Tris, 20 mm NaN3, 225 mm NaC1,
76 65 0.1% bovine serum albumin, ph 8.2) for 10 mm, blotted and incubated in 1NDW14D Mab in Tris buffer overnight. Grids were rinsed in 5 changes of Tris buffer, then incubated with 5 nm gold-conjugated goat anti-mouse Ig (H+L) (Ted Pella, Inc.) in Tris buffer for 1 hour. Grids were rinsed in two changes of 10X salt Tris buffer (20 mm Tris, 20 mm NaN3, 2.25 M NaC1, 0.1% bovine serum albumin, ph 8.2) (Giberson and Demeree 1994), five changes of Tris buffer, and five changes of nanopure water. Grids were air dried and stained on droplets of 1% uranyl acetate and 0.2% lead citrate for 5 min each. Grids were examined by transmission electron microscopy (Phillips CM12). PCR sample preparation Eight 7 gm paraffin sections were placed, four each, in 1.5 ml microfuge tubes. Sections were deparaffinized with two changes of xylene and two changes of 100% EtOH, centrifuged for 5 mm between each step and air dried (Wright and Manos 1990). RNA was extracted by RNAzolTm (Tel-Test, Inc.) treatment followed by choroform extraction and isopropanol precipitation. RNA was washed twice in 75% ethanol and air dried. The RNA in the two tubes were combined by dissolving the pellets in the same 10 gl volume of tris EDTA (TE; 50mM Tris, 1mM EDTA, and 0.5% Tween 20, ph 7.5). Mock-infected fish were used as negative controls and mock-infected fish with purified IHNV genomic RNA added were used as positive controls. PCR and southern hybridization Retrotherm PCR was done as described by Chiou et al. (1994) with sense (nucleotides ) and antisense (nucleotides ) N gene primers to obtain a 252 by N gene product (Arakawa et al. 1990). PCR reactions were incubated in an automatic thermal cycler (Coy Laboratory Products), electrophoresed on 1.5% GTG agarose gel (FMC Bioproducts) and blotted to nytran plus (Schleicher & Schuell) by
77 66 standard methods (Sambrook et al. 1989). PCR products were probed with a DIG endlabeled (Schmitz et al. 1991) internal N gene primer (nucleotides ) and detected by chemiluminescence (Holtke et al. 1992). Results Tissue culture Rainbow trout (Oncorhynchus mykiss) fry which had been infected with IHNV were positive throughout the epizootic for infectious virus by plaque assay until 46 days post exposure. Plaque assay results continued to be negative for the 90 d, 180 d, and 1 y samples. Histology Kidneys of 1 ype survivors had significant periglomerular and peritubular fibrosis as well as fibrotic foci within the hematopoietic tissue (Figure 5.1). Survivor kidneys contained greater than 50% fewer hematopoietic cells than those of agematched control fish. Glomeruli showed thickening and splitting of the Bowman's capsule and adherence of the glomerular tuft to the capsule. Renal tubules often contained large numbers of epithelial blebs, indicative of ongoing tissue injury. The PAS carbohydrate stain showed polysaccharide-positive deposits in the kidney tubules and in foci of the hematopoietic tissue. A diastase digestion done before the PAS stain which destroys the staining reaction in glycogen did not alter the PAS positive staining indicating that the deposits were not glycogen. Phenyl hydrazine HC1-PAS which destroys the staining reaction in glycoproteins did clear the positive staining reaction indicating the presence of glycoprotein in the deposits. Hemosiderin deposition and nephrocalcinosis were prominent throughout the kidneys. Kidneys from mock-infected
78 67 Figure 5.1 Kidney tissue from a 1 ype IHN epizootic survivor stained with Masson's Trichrome stain. Fibrotic connective tissue stained blue(arrows). Note the extensive fibrosis surrounding the renal tubule (T) and occluding the hematopoietic tissue (H).
79 ...4 vi a) L. a 4:
80 69 negative controls showed no fibrosis or glomerular lesions. Hematopoietic tissue was dense, few if any epithelial blebs were seen in tubules and PAS and Perl's hemosiderin stains were negative. There was little or no nephrocalcinosis in negative controls. Immunohistochemistry Dense, positive deposits were seen within renal tubules and hematopoietic tissue of survivor kidneys at 90 d, 180 d, 1 y and 2 ype when tested with 1NDW14D (Figure 5.2). No evidence of virus was seen in any of the other tissues, with the exception of one positive liver sample at 90 dpi. Weaker positive results were also seen with a second N-specific Mab (N-Mab; 1NCO27G) and two G-specific Mabs (G-Mab; 3GH135L, 3GH136J). The G-Mab APIH showed positive staining in individual hematopoietic cells, whereas the N Mabs showed positive staining in individual hematopoietic cells as well as a highly positive reaction in deposits located in renal tubules and hematopoietic interstitium. No positive staining was seen in mockinfected, age-matched negative control fish when tested with all 4 monoclonal antibodies. EM examination of 1 ype kidneys Negative staining of survivor kidney tissue previously embedded in paraffin and processed for EM contained intracytoplasmic rhabdoviral inclusions (Ghadially 1988) (Figure 5.3). Negative staining of survivor kidney tissue which had been held in formalin for two years then processed for EM contained nm If-INV-like DI particles in hematopoietic tissue.
81 70 Immunogold Immunogold labeling parameters were defined using kidney tissue samples from IHNV infected rainbow trout 8 dpe (Figure 5.4 a and b). IHNV -like DI particles in 1 ype survivors were confirmed as IHNV in origin by positive labeling with an anti- IHNV immunogold reaction (Figure 5.5). PCR and southern analysis IHNV RNA was detected in the kidney tissue of lypi survivors by PCR amplification. A 252 by band was detected in an ethidium bromide stained gel (Figure 5.6 a). The IHNV specificity of this band was confirmed by hybridization with an N gene specific probe to a region within the 252 by amplified fragment (Fig. 5.6 b). Discussion The histological staining with hematoxylin and eosin showed prominent lesions in MN survivor kidneys at 1 ype. Glycoprotein was detected in deposits within the renal tubules and hematopoietic interstitium by staining with PAS-phenyl hydrazine. Epithelial blebs and hemosiderin deposits were prominent throughout the kidney indicating ongoing renal tissue injury and destruction of red blood cells. These results suggest that renal and possibly hematopoietic function in IHN survivors is compromised. The Dunn-Thompson hemoglobin stain showed no indication of formalin-hemoglobin artifacts which can occur during the fixation of blood-rich tissues. Although no infectious virus was detected in survivors after 46 dpe, intense IHNV positive staining was seen by immunohistochemistry in deposits in renal tubules and hematopoietic interstitium of samples taken at 90 d, 180 d, 1 y and 2 ype. Antigen was seen in fibrotic regions, renal tubules and in close association with hematopoietic
82 71 Figure 5.2 Kidney tissue from a 1 ype IHN epizootic survivor as tested by alkaline phosphatase immunohistochemistry. Note the highly positive deposits in the renal tubules (T) (straight arrows) and in the sparse hematopoietic tissue (H) (curved arrows). Deposits stained red on microscopic slides. Figure 5.3 Cytoplasmic rhabdoviral inclusions in kidney tissue from a 1 ype IHN epizootic survivor. Tissue was negatively stained with uranyl acetate and lead citrate. Figure 5.4 a Immunogold labeling of IHNV particles using the anti-nucleocapsid monoclonal antibody, 1NDW14D. Figure 5.4 b Immunogold labeling of IHNV particles using the anti-glycoprotein monoclonal, 3GH136J. Figure 5.5 Immunogold labeled IHNV DI particles from 1 ype IHN epizootic survivor. Figure 5.6 a Retrotherm PCR amplification of a 252 by fragment of IHNV N gene from kidney tissue of a 1 year post exposure survivor. Lanes from left to right contained 123 by ladder marker, IHNV genomic RNA, and RNA extracted from a 1 ype epizootic survivor. The bright band at the bottom of each lane is excess primers and nucleotides. Figure 5.6 b Southern blot hybridization showing 252 by fragment is of IHNV N gene origin. Lane 1 contained 123 by ladder marker. Lane 2 contained a 10-4 dilution of IHNV genomic RNA. Lane 3 contained a 10-3 dilution of IHNV genomic RNA. Lane 4 contained a linearized plasmid containing the IHNV N gene as a positive control. Lane 5 contained the IHNV N gene digested from the positive control plasmid. Lane 6 contained RNA extracted from a negative control fish. Lane 7 contained RNA extracted from a 1 ype survivor kidney sample. The 500 by band in lanes 2 and 3 is the result of self priming (Arakawa et al. 1990).
83
84 73 loonm Figure 5.3
85 Figure 5.4 a "7 4
86 75 S. 50 nm Figure 5.4 b
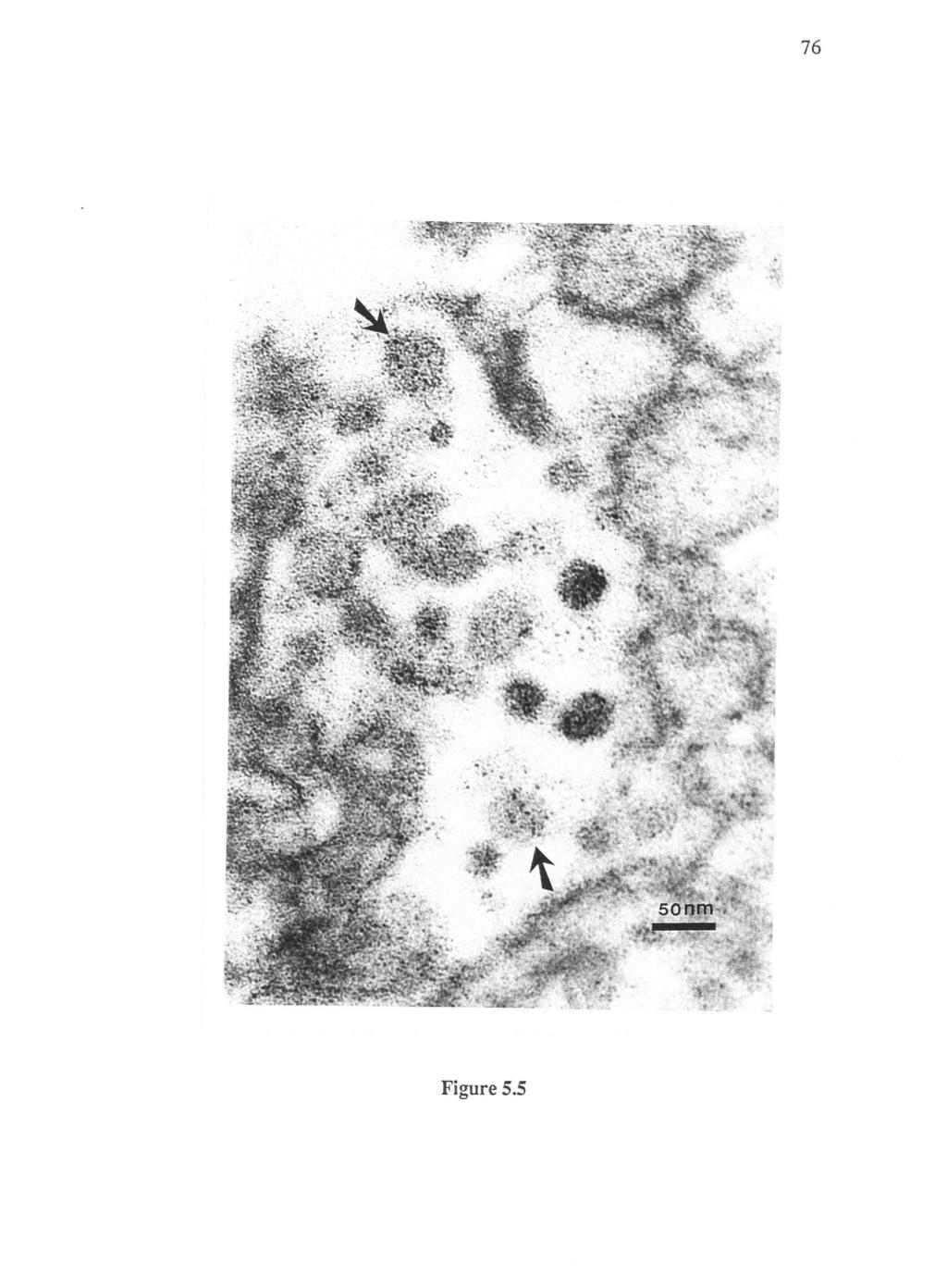
87 76

88 Cr Marker IHNV RNA 1 ype Survivor
89 by 252 by Figure 5.6 b
90 79 cells. As to the possibility of reinfection, survivors showed no signs of IHN infection during their lifetime and were maintained in pathogen-free water. No viral contaminants were detected by tissue culture testing of 11 organs, every 3-6 months, throughout their captivity. Bacterial contaminants, such as Renibacterium salmoninarum the causative agent of bacterial kidney disease, which causes salmonid renal lesions (Sami et al. 1992) were seen in survivor kidneys. Lesions observed were therefore assumed to be due to IHNV. All survivor kidney samples examined had moderate to severe precipitation of calcium complexes in renal tubules. This condition is commonly known as nephrocalcinosis and is associated with excessive carbon dioxide levels in the water (Harrison and Richards 1979; Smart et al. 1979). Age-matched mock-infected negative controls had mild or no nephrocalcinosis. Routine sampling of laboratory rainbow trout populations at the Center for Salmon Disease Research fish facility (Corvallis, Oregon) have shown a 30-60% incidence of nephrocalcinosis in IHN survivors versus 0-10% incidence in age-matched, mock-infected survivors (data not shown). The increased incidence of nephrocalcinosis in fish that survive IHN epizootics suggests that the general impairment of kidney function from IHNV infection predisposes fish to nephrocalcinosis. This, in turn, may result in the death of survivors before sexual maturation. Cytoplasmic rhabdoviral inclusions, like those reported in rabies virus infections (Ghadially 1988), were detected in a kidney sample which had been embedded in paraffin before processing for EM. We were not able to confirm the cytoplasmic rhabdoviral inclusions as IHNV in origin because the tissue was unsuitable for immunogold testing. Rhabdoviral DI particles are typically spherical and one-third the size of standard virus particles (Pattniak and Wertz 1991). Round 50 nm particles, approximately one-third the size of standard IHNV particles (160 x 90 nm; Amend and Chambers 1970) were seen in kidney tissues of 1 ype survivors which had been in
91 80 formalin for 2 years. The DI particles were confirmed as IHNV in origin by immunogold labeling (Figure 5.5). The presence of DI particles suggests that standard infectious virions are present, but kept at levels below tissue culture detection limits. This has been well documented with VSV and rabies virus in tissue culture (for review, see Sekellick and Marcus 1980). Standard and DI particles may be in such balance that only a very few virions are synthesized, resulting in a subacute or latent form of the infection, until the balance is upset by the stressful conditions of spawning and/or exposure of the fish to the same or other viruses. Standard infectious virus would subsequently increase to levels detectable by plaque assay, resulting in an apparently negative survivor becoming positive at sexual maturation. Experimentally or naturally infected fish which are held in captivity do not experience the same stress levels as fish naturally migrating upstream to their natural spawning grounds. Nor are they exposed to the onslaught of pathogens (IHNV and other) which contaminate river waters. The inability to detect infectious virus in experimentally or naturally infected, sexually mature survivors held in captivity may be due to these environmental differences. Until now, there has been no proof that IHNV persists in epizootic survivors past 50 days when the last positive plaque assays are typically seen. We have shown, unequivocally, that IHNV does persist in fish that survive IHN epizootics. IHNVspecific N and G proteins were detected by immunohistochemistry using 4 different Mabs. In addition, a fragment of the N gene was amplified by PCR from survivor kidney and confirmed with southern hybridization (Figure 5.6 a and b). The differential positive staining by G-Mabs versus N-Mabs may be attributed to the G monoclonals binding primarily to currently infected, N and G expressing, hematopoietic cells, and N monoclonals binding primarily to the dense deposits which contained DI particles and nucleoprotein conglomerates upon examination by immunogold EM (Figure 5.5).
92 81 Most, if not all, viruses have been shown to produce DI particles in tissue culture cells. In addition, animals given DI particles before or during viral infections show an altered infection course. Despite these studies, there has been no definite report of DI particles from animals following a natural infection. In this study, animals were infected by adding a moderate dose (103 standard virus particles/ml) of virus directly to the water thereby simulating the horizontal exposure known to occur during natural epizootics. Daily and cumulative mortality was just as expected for experimental exposure of this species to this dose of IHNV, and clinical disease signs were identical to those seen in natural infections. The DI particles seen in these survivors suggest that the persistence of IHNV in salmonid populations is attributable to the subclinical carrier status of survivors caused by DI particles. Standard virions are detected during natural spawning because the DI/standard particle balance is upset and results in a resurgence of standard virus. This finding does not preclude reinfection as a source of virus in the adult. In fact, re-exposure to IHNV could be a factor in upsetting the DI/standard particle balance. Acknowledgments Sandra Ristow (Washington State University, Pullman, Washington) provided the monoclonal antibodies used in this study. Dr. Linda Boot land and Harriet Lorz assisted in fish care, maintenance and sampling. Pat Allison provided technical assistance with histological staining. Julie Duimstra provided technical assistance with immunogold labeling. Al Soeldner provided technical assistance in electron microscopy and photography.
93 82 References Amend, D.F Detection and transmission of infectious hematopoietic necrosis virus in rainbow trout. J. Wildl. Dis. 11: Amend, D.F. and V.C. Chambers Morphology of certain viruses of salmonid fishes. II. In vivo studies of infectious hematopoietic virus. J. Fish. Res. Bd. Canada 27: Amos, K.H., editor Procedures for the detection and identification of certain fish pathogens, 3rd edition. Fish Health Section, American Fisheries Society. Corvallis, Oregon. Arakawa, C.K., R.E. Deering, K.H. Higman, K.H. Oshima, P.J. O'Hara and J.R. Winton Polymerase chain reaction (PCR) amplification of a nucleoprotein gene sequence of infectious hematopoietic necrosis virus. Dis. Aquat. Org. 8(3): Bangham C.R.M. and T.B.L. Kirkwood Defective interfering particles: effects in modulating virus growth and persistence. Virology 179: Barrett, A.D.T. and N.J. Dimmock Defective interfering viruses and infections of animals. Cur.. Top. Microbiol. Immunol. 128: Bean, W.J., Y. Kawaoka, J.M. Wood, J.E. Pearson and R.G. Webster Characterization of virulent and avirulent A/chicken/Pennsylvania/83 influenza A viruses: potential role of defective interfering RNAs in nature. J. Virol. 41: Browning, M.J., B.S. Huneycutt, A.S. Huang and C.S. Reiss Replicationdefective viruses modulate immune responses. J. Immunol. 147: Chiou, P.P., B.S. Drolet and J.C. Leong PCR amplification of IHNV RNA from paraffin embedded fish tissue. J. Aquat. Anim. Health. submitted. Doyle, M. and J.J. Holland Prophylaxis and immunization in mice by use of virus-free defective T particles to protect against intracerebral infection by vesicular stomatitis virus. Proc. Natl. Acad. Sci. (USA) 70: Drolet, B.S., J.S. Rohovec and J.C. Leong The route of entry and progression of infectious hematopoietic necrosis virus in Oncorhynchus mykiss (Walbaum): a sequential immunohistochemical study. J. Fish. Dis. 17(4): Dunn, R.C. and E.C. Thompson A new hemoglobin stain for histologic use. Arch. Pathol. 39: Engelking, H.M. and J.C. Leong IHNV persistently infects chinook salmon embryo cells. Virology, 109(1):47-58.
 cell line from carp Cyprinus carpio. Annals of Virology 134:207-220. Ghadially, F.N. 1988. Cytoplasmic matrix and their inclusions, pp.")
94 83 Fijan, N., D. Sulimanovic, M. Bearsotti, D. Musinie, L.D. Zwillengerg, S. Chilmonczyk, J.F. Vantherot and P. de Kinkelin Some properties of the Epithelioma papulosum cyprini (EPC) cell line from carp Cyprinus carpio. Annals of Virology 134: Ghadially, F.N Cytoplasmic matrix and their inclusions, pp In: A text and atlas of physiological and pathological alterations in the fine structure of cellular and extracellular components, 3rd Ed. Vol. II. Butterworths, London. Giberson, R.T. and R.S. Demeree, Jr The influence of immunogold particle size on labeling density. Microscopy Research and Technique. In Press. Gill, G.W., J.K. Frost and K.A. Miller A new formula for a half-oxidized hematoxylin that neither overstains nor requires differentiation. Acta Cytol. 18(4): Harrison, J.G., R.H. Richards The pathology and histopathology of nephrocalcinosis in rainbow trout (Salmo gairneri) Richardson in fresh water. J. Fish. Dis. 2:1-12. Highman, B A new modification of Perl's reaction for hemosiderin in tissues. Arch. Pathol. 33: Holland, J.J Defective interfering rhabdoviruses, p In: R.R. Wagner (ed.), The Rhabdoviruses. Plenum Press, New York. Holland, J.J. and L.P. Villareal Persistent noncytocidal vesicular stomatitis virus infections mediated by defective T particles that suppress virion transcriptase. Proc. Nat. Acad. Sci. USA 71: Holtke, H.J., G. Sanger, C. Kessler and G. Schmitz Sensitive chemiluminescent detection of digoxigenin-labeled nucleic acids: A fast and simple protocol and its applications. Biotechniques 12: Horodyski, F.M., S.T. Nichol, K.R. Spindler and J.J. Holland Properties of DI particle resistant mutants of vesicular stomatitis virus isolated from persistent infection and from undiluted passages. Cell 33: Huang, A.S. and D. Baltimore Defective viral particles and viral disease processes. Nature 226: Huang, A.S. and D. Baltimore Defective interfering animal viruses, p In H. Fraenkel-Conrat and R.R. Wagner (ed.), Comprehensive Virology, vol. 10. Plenum Press, New York. Kawai, A., S. Matsumoto and K. Tanabe Characterization of rabies viruses recovered from persistently infected BHK cells. Virology 67: Lazzarini, R.A., J.D. Keene and M. Schubert The origins of defective interfering particles of the negative-stranded RNA viruses. Cell 26: Leppi, T.J. and S.S. Spicer The histochemistry of mucins on certain primate salivary glands. Am J. Anat. 118:

95 84 Lillie, R.D. and H.M. Fullmer Histopathological technic and practical histochemistry, ed. 4. McGraw-Hill Book Co., New York. Mallory, F.B Pathological technique. Hafner Publishing Co., New York. Masson, P Trichrome stainings and their preliminary technique. J. Tech. Meth. 12: McAllister, P.E., and K.S. Pilcher Autointerference in infectious hematopoietic necrosis virus of salmonid fish. Proc. Soc. Exp. Biol. Med. 145: McManus, J.F.A The histological and histochemical uses of periodic acid. Stain Technol. 23: Moscona, A. and R.W. Peluso Persistent infection with human parainfluenza virus 3 in CV-1 cells: Analysis of the role of defective interfering particles. Virology 194: Pattniak, A.K. and G.W. Wertz Cells that express all five proteins of vesicular stomatitis virus from cloned cdnas support replication, assembly, and budding of defective interfering particles. PNAS, USA. 88: Perrault, J Origin and replication of defective interfering particles. Cur.. Top. Microbiol. Immunol. 93: Pedley, S., F. Hundley, I. Chrystie, M.A. McCrae and U. Desselberger The genomes of rotaviruses isolated from chronically infected immunodeficient children. J. Gen. Virol. 65: Ramseur, J.M. and R.M. Friedman Prolonged infection of L cells with vesicular stomatitis virus: Defective interfering forms and temperature-sensitive mutants as factors in the infection. Virology 85: Sambrook, J., E.F. Fritsch and T. Maniatis, editors Analysis of genomic DNA by southern hybridization, pp In: Molecular Cloning, A laboratory Manual, Second Edition, volume 2, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York. Sami, S., T. Fischer-Scherl, R.W. Hoffmann, and C. Pfeil-Putzein Immune complex-mediated glomerulonephritis associated with bacterial kidney disease in the rainbow trout (Oncorhynchus mykiss). Vet Pathol. 29: Schmitz, G.G., T. Walter, R. Seibl and C. Kessler Nonradioactive labeling of oligonucleotides in vitro with the hapten digoxigenin by tailing with terminal transferase. Anal. Biochem. 192: Scott, J.L., J.L. Fendrick and J.C. Leong Growth of infectious hematopoietic necrosis virus in mosquito and fish cell lines. Wasmann J. Biol. 38: Sekellick, M.J. and P.I. Marcus Persistent infections by rhabdoviruses, pp In: D.L. Bishop (ed.), Rhabdoviruses. Volume 3. CRC Press, Inc., Boca Raton, Florida.
96 85 Smart G.R., D. Knox, J.G. Harrison, J.A. Ralph, R.H. Richards and C.B. Cowley Nephrocalcinosis in rainbow trout (Salmo gairdneri) Richardson: effect of exposure to elevated CO2 concentrations. J. Fish. Dis. 2: Williams, MA Autoradiography and immunocytochemistry, pp In: A.M. Glauert (ed.), Practical Methods in Electron Microscopy. North-Holland Publishing company, Amsterdam. Wright, D.K. and M.M. Manos Sample preparation from paraffin-embedded tissues, pp In: M.A. Innis, D.H. Gelfand, J.J. Sninsky, and T.J. White (eds.), PCR Protocols. Academic Press, New York.
97 86 CHAPTER 6 THESIS SUMMARY Serological identification of IHNV in fixed tissue culture cells Confirmatory identification of viruses by specific reaction with an antibody reagent is a standard procedure in the virus diagnostic laboratory. The immunological assay can be a virus neutralization assay (McCain et al. 1971; Hsu and Leong 1985; Winton et al. 1988) or an assay that measures the specific reactivity of an antibody with a viral antigen (for review of IHNV assays, see Winton 1991). The APIC procedure described in chapter 3 was easy to perform, did not require special equipment, and a positive result was easy to detect. It will probably have its greatest use with virus samples that have been lost as a result of laboratory error or a severe reduction in sample viability. It may also be used when a laboratory has to screen hundreds of samples and the examination of individual plates for plaques before fixation and staining is not possible. The route of IHNV entry and progression in rainbow trout By carefully examining sequential fish samples taken before and during an epizootic, the route of entry and the progression of IHNV within rainbow trout has been determined. IHNV infection progressed from two major sites: the gills and the gastrointestinal tract. Both routes of entry eventually resulted in systemic viremia. While IHNV readily infected epithelial cells of numerous organs, it had a propensity for connective tissue. While IHNV readily infected epithelial cells, the infection appeared transient and not highly productive. The majority of cells undergoing productive viral infection were of connective tissue origin. These results may influence the design of vaccines, and the study of specific cell receptors involved in IHNV infection.
98 87 Detection of IHNV in epizootic survivors The survivors from the epizootic study described in chapter 4 and survivor rainbow trout from previous infection studies were examined by APIH for IHNV nucleocapsid protein. Intense IHNV positive staining deposits were seen in renal tubules and hematopoietic interstitium of samples taken at 90 d, 180 d, 1 y and 2 ype. This unexpected positivity in fish where no infectious virus had been detected since 46 dpe was the impetus for the survivor study described in chapter 5. In addition to the N protein, glycoprotein was detected in the deposits by staining with PAS-phenyl hydrazine and by APIH. Histological staining with hematoxylin and eosin showed prominent lesions in IHN survivor kidneys at 1 ype. Until now, there has been no proof that IHNV persists in epizootic survivors past 50 days when the last positive plaque assays are typically seen. The amplification of a fragment of the IHNV N gene by PCR and the detection of two IHNV proteins with APIH is proof that IHNV does in fact persist in the epizootic survivor for at least two years. This persistence is in the form of a heterogeneic population of standard and DI particles, resulting in a subclinical carrier status. This finding does not preclude reinfection as a source of virus in the adult. In fact, re-exposure to IHNV could be a factor in upsetting the DI/standard particle balance. Future use of methods The methods described in this thesis have already given new research tools to ongoing IHNV infection studies. The immunocytochemical techniques are currently being used to determine transfection efficiencies of various IHNV gene constructs and to detect the expression of viral proteins in transgenic cells which can then be examined for resistance to IHNV infection (Anderson and Leong 1991). The APIC assay is currently being used by several diagnostic fish laboratories to detect IHNV in fixed
99 88 plaque assay plates as opposed to subculturing for immunofluorescence or plaque neutralizations. Future use of results The detailed description of specific virus-positive cell types and degrees of IHNV infection and progression of the virus in rainbow trout provided preliminary work for a funded grant to characterize the receptor of IHNV. Documentation of the complete pathogenic pathway of a wild type IHNV isolate could be used as a baseline for examining alterations in pathogenesis from protein mutations, protective monoclonal antibodies, or vaccines. The most exciting results of these studies is the irrefutable evidence of persistence of IHNV in epizootic survivors. For the first time, IHNV has been shown to persist after the 50 day positive plaque assay limit. This result allows for the longheld hypothesis that adults returning to spawn are positive for virus because it was never cleared and is somehow reactivated upon sexual maturation. This will inevitably lead to more research on IHNV persistence and the role that it plays in maintaining the endemic level of virus in nature. This may also change the way hatcheries have been dealing with IHN survivors for the past 35 years. In the best scenario, the destruction of IHN survivors will decrease levels of virus in nature to the point where this virus is no longer a significant problem to hatcheries.
100 89 BIBLIOGRAPHY Ahne, W Uptake and multiplication of spring viraemia of carp virus in carp, Cyprinus carpio L. J. Fish Dis. 1: Amend, D.F Detection and transmission of infectious hematopoietic necrosis virus in rainbow trout. J. Wildl. Dis. 11: Amend, D.F. and V.C. Chambers Morphology of certain viruses of salmonid fishes. II. In vivo studies of infectious hematopoietic virus. J. Fish. Res. Bd. Canada 27: Amend D.F. and J.P. Pietsch. 1972a. Virucidal activity of two iodophors to salmonid viruses. J. Fish. Res. Board Can. 29: Amend D.F. and J.P. Pietsch. 1972b. An improved method for isolating viruses from asymptomatic carrier fish. Trans. Amer. Fish. Soc. 2: Amend, D.F. and L. Smith Pathophysiology of infectious hematopoietic necrosis virus disease in rainbow trout (Sa lmo gairdneri): Early changes in blood and aspects of the immune response after injection of IHN virus. J. Fish.. Res. Board Can. 31: Amend, D.F., W.T. Yasutake and R.W. Mead A hematopoietic virus disease of rainbow trout and sockeye salmon. Trans. Am. Fish. Soc. 98: Amos, K.H., (ed.) Procedures for the detection and identification of certain fish pathogens, pp Fish Health Section, American Fisheries Society, 3rd ed. Corvallis, Oregon. Amos, K.H., K.A. Hopper and L. LeVander Absence of infectious hematopoietic necrosis virus in adult sockeye salmon. J. Aquat. Anim. Health. 1(4): Anderson, E.D., C. Cary, J.S. Rohovec and J.C. Leong Detection of infectious hematopoietic necrosis virus mrna by in situ hybridization. Proceedings of the Second International Symposium on Viruses of Lower Vertebrates. Corvallis, Oregon. Anderson, E.D. and J.C. Leong Reduction of IHNV induced CPE by transgene expression of the nucleocapsid gene, pp. 90. Second International Marine Biotechnology Conference, Baltimore, MD. Arakawa, C.K., R.E. Deering, K.H. Higman, K.H. Oshima, P.J. O'Hara and J.R. Winton Polymerase chain reaction (PCR) amplification of a nucleoprotein gene sequence of infectious hematopoietic necrosis virus. Dis. Aquat. Org. 8(3): Arnzen, J.M., S.S. Ristow, C.P. Hesson and J. Lientz Rapid fluorescent antibody tests for infectious hematopoietic necrosis virus (IHNV) utilizing monoclonal antibodies to the nucleoprotein and glycoprotein. J. Aquat. Anim. Health. 3(2):

101 90 Asakawa, H.,Y. Tsuda, M. Sukegawa, M. Nakano, T. Moritomo and T. Watanabe Plaque assay method for fish viruses using the cell line derived from chum salmon embryo. Bull. Coll. Agric. Vet. Med. 43: Bangham C.R.M. and T.B.L. Kirkwood Defective interfering particles: effects in modulating virus growth and persistence. Virology 179: Barrett, A.D.T. and N.J. Dimmock Defective interfering viruses and infections of animals. Cum Top. Microbiol. Immunol. 128: Basurco, B., S. Yun and R.P. Hedrick Comparison of selected strains of infectious hematopoietic necrosis virus (IHNV) using neutralizing trout antisera. Dis. Aquat. Org. 15(3): Batts, W.N., M.L. Landolt and J.R. Winton Inactivation of infectious hematopoietic necrosis virus (IHNV) by low levels of iodine. Appl. Environ. Microbiol. 57(5): Batts, W.N. and J.R. Winton. 1989a. Concentration of infectious hematopoietic necrosis virus from water samples by tangential flow filtration and polyethylene glycol precipitation. Can. J. Fish. Aquat. Sci. 46(6): Batts, W.N. and J.R. Winton. 1989b. Enhanced detection of infectious hematopoietic necrosis virus and other fish viruses by pretreatment of cell monolayers with polyethylene glycol. J. Aquat. Anim. Health. 1(4): Bean, W.J., Y. Kawaoka, J.M. Wood, J.E. Pearson and R.G. Webster Characterization of virulent and avirulent A/chicken/Pennsylvania/83 influenza A viruses: potential role of defective interfering RNAs in nature. J. Virol. 41: Bootland, L. and J.C. Leong Staphylococcal coagglutination, a rapid method of identifying infectious hematopoietic necrosis virus. Appl. Environ. Microbiol. 58(1):6-13. Bootland, L., H.V. Lorz, J.S. Rohovec and J.C. Leong Experimental infection of brook trout with infectious hematopoietic necrosis virus Types 1 and 2. J. Aquat. Anim. Health 6: Bovo, G., G. Giorgetti, P.E.V. JOrgensen and N.J. Nelson Infectious hematopoietic necrosis : first detection in Italy. Bull. Eur. Ass. Fish Pathol. 7:124. Brown, C.C., H.J. Olander and D.A. Senne A pathogenesis study of highly pathogenic avian influenza virus H5N2 in chickens, using immunohistochemistry. J. Compar. Pathol. 107 (3): Browning, M.J., B.S. Huneycutt, A.S. Huang and C.S. Reiss Replicationdefective viruses modulate immune responses. J. Immunol. 147: Burke J. and D. Mulcahy Plaguing procedure for infectious haematopoietic necrosis virus. Appl. Env. Microbiol. 39:
102 91 Burke J. and D. Mulcahy Retention of infectious haematopoietic necrosis virus infectivity in fish tissue homgenates and fluids stored at three temperatures. J. Fish. Dis. 6: Burke, J. and R. Grischkowsky An epizootic caused by infectious hematopoietic necrosis virus in an enhanced population of sockeye salmon, Oncorhynchus nerka (Walbaum), smolts at Hidden Creek, Alaska. J. Fish Dis. 7(5): Castric, J. and J. Jeffroy Experimentally induced diseases in marine fish with IHNV and a rhabdovirus of eel, pp In: N. De Pauw and J. Joyce (eds.), Aquaculture and the environment. EAS Special Publication No. 14. Dublin, Ireland. Chen, M.F., C.M. Aikens, J.L. Fryer and J.S. Rohovec Virulence of four isolates of infectious hematopoietic necrosis virus in salmonid fishes and comparative replication in salmonid fish cell lines. Calif. Fish Game. 76(3): Chen, M.F., J.S. Rohovec and J.L. Fryer Biological properties of large and small plaque variants of infectious hematopoietic necrosis virus. Fish Pathol. 25 (4): Chiou, P.P., B.S. Drolet and J.C. Leong PCR amplification of IHNV RNA from paraffin embedded fish tissue. J. Aquat. Anim. Health. submitted. Clark H.F. and E.Z. Soriano Fish rhabdovirus replication in non-piscine cell culture: New system for the study of rhabdovirus-cell interaction in which the virus and cell have different temparature optima. Infec. Immun. 10: Cole, G.A., D.H. Gilden, A.A. Monjan and N. Nathanson Lymphocytic choriomeningitis virus: Pathogenesis of acute central nervous system disease. Fed. Proc. 30: de Avila, J., S. Steiner, S. Ristow, J. Leintz and C. Hesson The presence of low titers of anti-ihnv antibody in the serum of steelhead trout and chinook salmon broodstock spawned at Dworshak National Fish Hatchery during FHS/AFS Newsletter 20(2):6-8. Deering, R.E., C.K. Arakawa, K.H. Oshima, P.J. O'Hara, M.L. Landolt and J.R. Winton Development of a biotinylated DNA probe for detection and identification of infectious hematopoietic necrosis virus. Dis. Aquat. Org. 11(1): de Kinkelin, P., J. Bernard and A.M. Hattenberger-Baudouy Immunization of fishes against cold water viruses, pp In: P. de Kinkelin (ed.), Symposium on the vaccination of fishes. Theoretical basis and practical results of immunization against infectious diseases. Paris, France. de Kinkelin, P., M. Dorson and A.M. Hattenberger-Baudouy Interferon synthesis in trout and carp after viral infection, pp In: W.B. van Muiswinkel and E.L. Cooper (eds.) Immunology and immunization of fish. Conference, June 22 24, Wageningen, the Netherlands. Dev. Comp. Immunol. Suppl. 2.
 70:2105-2108.")
103 92 Dixon, P.F. and B.J. Hill Rapid detection of fish rhabdoviruses by the enzymelinked immunosorbent assay (ELISA). Aquaculture 42(1):1-12. Doyle, M. and J.J. Holland Prophylaxis and immunization in mice by use of virus-free defective T particles to protect against intracerebral infection by vesicular stomatitis virus. Proc. Natl. Acad. Sci. (USA) 70: Drolet, B.S., J.S. Rohovec and J.C. Leong Serological identification of infectious hematopoietic necrosis virus in fixed tissue culture cells by alkaline phosphatase immunocytochemistry. J. Aquat. Anim.Health 5(4): Drolet, B.S., J.S. Rohovec and J.C. Leong The route of entry and progression of infectious hematopoietic necrosis virus in Oncorhynchus mykiss (Walbaum): a sequential immunohistochemical study. J. Fish. Dis. 17(4): Dunn, R.C. and E.C. Thompson A new hemoglobin stain for histologic use. Arch. Pathol. 39: Eaton, W.D., J. Hulett, R. Brunson and K. True The first isolation in North America of infectious hematopoietic necrosis virus (IHNV) and viral hemorrhagic septicemia virus (VHSV) in the same watershed. J. Aquat. Anim. Health. 3(2): Engelking, H.M. and J.C. Leong IHNV persitently infects chinook salmon embryo cells. Virology, 109(1): Engelking, H.M. and J.C. Leong. 1989a. The glycoprotein of infectious hematopoietic necrosis virus elicits neutralizing antibody and protective responses. Virus Research 13: Engelking, H.M. and J.C. Leong. 1989b. Glycoprotein from infectious hematopoietic necrosis virus (IHNV) induces protective immunity against five IHNV types. J. Aquat. Anim. Health 1: Faulk, W. and G. Taylor An immunocolloid method for the electron microscope. Immunochemistry 8: Fendrick, J.L., W.J. Groberg and J.C. Leong Comparative sensitivity of five fish cell lines to wild type infectious hematopoietic necrosis virus from two Oregon sources. J. Fish. Dis. 5: Fijan, N., D. Sulimanovic, M. Bearsotti, D. Musinie, L.D. Zwillengerg, S. Chilmonczyk, J.F. Vantherot and P. de Kinkelin Some properties of the Epithelioma papulosum cyprini (EPC) cell line from carp Cyprinus carpio. Ann. Virol. 134: Fryer, J.L., A. Yusha and K.S. Pilcher The in vitro cultivation of tissue and cells of Pacific salmon and steelhead trout. Ann. N.Y. Acad. Sci. 126:126: Ghadially, F.N Cytoplasmic matrix and their inclusions, pp In: A text and atlas of physiological and pathological alterations in the fine structure of cellular and extracellular components, 3rd Ed. Vol II. Butterworths, London.
104 93 Giberson, R.T. and R.S. Demeree, Jr The influence of immunogold particle size on labeling density. Microscopy Research and Technique. In Press. Gill, G.W., J.K. Frost and K.A. Miller A new formula for a half-oxidized hematoxyin that neither overstains nor requires differentiation. Acta Cytol. 18(4): Gilmore, R.D. and J.C. Leong The nucleocapsid gene of infectious hematopoietic necrosis virus, a fish rhabdovirus. Virology. 167(2): Goldes, S.A. and S.L. Mead Susceptibility of brook trout Salvelinus fontinalis to infectious hematopoietic necrosis virus. FHS/AFS Spring Newsletter 20(1):4. Grace, M.F. and M.J. Manning Histogenesis of the lymphoid organs in rainbow trout, Salmo gairdneri Rich Dev. Comp. Immunol. 4: Gravell, M. and R.G. Malsberger A permanent cell line from the fathead minnow (Pimephales promelas). Ann. N.Y. Acad. Sci. 126: Grischkowsky, T.S. and D.F. Amend Infectious hematopoietic necrosis virus : Prevalence in certain Alaskan sockeye salmon, Oncorhynchus nerka. J. Fish. Res. Board Can. 33:186. Grischkowsky, T.S. and D. Mulcahy Effects of injection of hormones on the expression of infectious hematopoietic necrosis virus in spawning sockeye salmon (Oncorhynchus nerka), pp In: B.R. Metcalf and R.A. Neve (eds.), Proceedings of the North Pacific Aquaculture Symposium, Alaska University. Harrison, J.G., R.H. Richards The pathology and histopathology of nephrocalcinosis in rainbow trout (Salmo gairneri) Richardson in fresh water. J. Fish. Dis. 2:1-12. Helmick, C.M., J.F. Bailey, J. DeAvila and S.S. Ristow Application of immunogold techniques coupled with quantitative x-ray microanalysis to study infectious hematopoietic necrosis virus. Proc. Int'l. Symp. Vir. Lower Vert. Corvallis, Oregon. 2: Hedrick, R.P., S.E. LaPatra, J.L. Fryer, T. McDowell and W.H. Wingfeild Susceptibility of coho (Oncorhynchus kisutch) and chinook (Oncorhynchus tshawytscha) salmon hybrids to experimental infections with infectious hematopoietic necrosis virus (IHNV). Bull. Eur. Ass. Fish Pathol. 7(4): Hetrick, F.M., J.L. Fryer and M.D. Knittle Effect of water temperature on the infection of rainbow trout Salmo gairdneri (Richardson) with infectious hematopoietic necrosis virus. J. Fish Dis. 2(3): Hill B.J., B.O. Underwood, C.J. Smale and F. Brown Physiochemical and serological characterization of five rhabdoviruses infecting fish. J. Gen. Virol. 27:

105 94 Hill B.J Impact of viral diseases of slmonid fish in the European community, pp In: T. Kimura (ed.), Proceedings of the OJI International Symposium on Salmonid Diseases. Hokkaido University Press, Sapporo, Japan. Highman, B A new modification of Perl's reaction for hemosiderin in tissues. Arch. Path. 33: Holland, J.J Defective interfering rhabdoviruses, pp In R.R.Wagner (ed.), The Rhabdoviruses. Plenum Press, New York. Holland, J.J. and L.P. Villareal Persistent noncytocidal vesicular stomatitis virus infections mediated by defective T particles that suppress virion transcriptase. Proc. Nat. Acad. Sci. USA 71: Holland, J.J., L.P. Villarreal and M. Breindi Factors involved in the generation and replication of rhabdovirus defective T particles. J. Virol. 17: Holtke, H.J., G. Sanger, C. Kessler and G. Schmitz Sensitive chemiluminescent detection of digoxigenin-labeled nucleic acids: A fast and simple protocol and its applications. Biotechniques 12: Horisberger, M. and J. Rosset Colloidal gold, a useful marker for transmission and scanning electron microscopy. J. Histochem. Cytochem. 25(4): Horodyski, F.M., S.T. Nichol, K.R. Spindler and J.J.Holland Properties of DI particle resistant mutants of vesicular stomatitis virus isolated from persistent infection and from undiluted passages. Cell 33: Hsu, Y.L., H.M. Engelking and J.C. Leong Occurrence of different types of infectious hematopoietic necrosis virus in fish. Appl. Environ Microbiol. 52: Hsu, S.M., L. Raine and H. Fanger. 1981a. Use of avidin-biotin-peroxidase complex (ABC) in immunoperoxidase techniques. J. Histochem. Cytochem. 29(4): Hsu, S.M., L. Raine and H. Fanger b. A comparative study of the peroxidaseantiperoxidse method and an avidin-biotin complex method for studying polypeptide hormones with radioimmunoassay antibodies. Am J. Clin. Pathol. 75: Hsu, Y.L. and J.C. Leong A comparison of detection methods for infectious hematopoietic necrosis virus. J. Fish Dis. 9: Huang, A.S. and D. Baltimore Defective viral particles and viral disease processes. Nature 226: Huang, A.S. and D. Baltimore Defective interfering animal viruses, p In H. Fraenkel-Conrat and R.R. Wagner (ed.), Comprehensive Virology, vol. 10. Plenum Press, New York.
 and viral hemorrhagic septicemia (VHS): Detection of trout antibodies to the causative viruses by means of plaque neutralization, immunofluorescence, and")
106 95 JOrgensen, P.E.V., N.J. Olesen, N. Lorenzen, J.R. Winton and S.S. Ristow Infectious hematopoietic necrosis (IHN) and viral hemorrhagic septicemia (VHS): Detection of trout antibodies to the causative viruses by means of plaque neutralization, immunofluorescence, and enzyme-linked immunosorbent assay. J. Aquat. Anim. Health. 3(2): Kamei, Y., M. Yoshimizu, Y. Esura and T. Kimura Effects of estuarine and marine waters on the infectivities of infectious hematopoietic necrosis virus (IHNV) and infectious pancreatic necrosis virus (IPNV). Bull. Fac. Fish. Hokkaido Univ. 38(3): Kamei, Y., M. Yoshimizu, Y. Esura and T. Kimura Effects of environmental water on the infectivities of infectious hematopoietic necrosis virus (IHNV) and infectious pancreatic necrosis virus (IPNV). J. Appl. Ichthyol. 4(1): Kasai, K. J. Yonezawa, A. Ono, A. Hasegawa, T. Honma and H. Fukuda Brood and size dependent variation in susceptibility of rainbow trout, Oncorhynchus mykiss to artificial infection of infectious hematopoietic necrosis virus (IHNV). Fish Pathol. 28(1):35-40 Kasornchandra, J., H.M. Engelking, C.N. Lannan, J.S. Rohovec and J.L. Fryer Characteristics of three rhabdoviruses from snakehead fish Ophicephalus striatus. Dis. Aquat. Org. 13: Kasornchandra, J., C.N. Lannan, J.S. Rohovec and J.L. Fryer Characterization of a rhabdovirus isolated from the snakehead fish (Ophicephalus striatus). Proc. Sec. Int. Symp. Vir. Low. Verteb. 2: Kawai, A., S. Matsumoto and K. Tanabe Characterization of rabies viruses recovered from persistently infected BHK cells. Virology 67: Kelly, R.K., B.W. Souter and H.R. Miller Fish cell lines: Comparisons of CHSE 214, FHM, and RTG-2 in assaying IHN and IPN viruses. J. Fish. Res. Board Can.35(7): Kimura, T. and T. Awakura Studies on viral diseases of Japanese fishes. VI. Infectious hematopoietic necrosis (IHN) of salmonids in the mainland of Japan. J. Tokyo Univ. Fish. 77:63 Kimura, T., M. Yoshimizu and S. Gorie A new rhabdovirus isolated in Japan from cultured hirame (Japanese flounder) Paralichthys olivaceus and ayu Plecoglossus altivelis. Dis. Aquat. Org. 1: Kurath, G., K.G. Ahern, G.D. Pearson and J.C. Leong Molecular cloning of the six mrna species of infectious hematopoietic necrosis virus, a fish rhabdovirus, and gene order determination by R-loop mapping. J. Virol. 53: Kurath, G. and J.C. Leong Characterization of infectious hematopoietic necrosis virus mrna species reveals a nonvirion rhabdoviral protein. J. Virol. 53(2):
107 96 Lannan, C.N., J.R. Winton and J.L. Fryer Fish cell lines: establishment and characterization of nine cell lines from salmonids. In Vitro 20: LaPatra, S.E. 1989a. Strain differentiation and detection of infectious hematopoietic necrosis virus. Ph.D. thesis, Oregon State University, Corvallis, Oregon. pp LaPatra, S.E. 1989b. Infectious hematopoietic necrosis virus (IHNV) transmission studies in Oregon. Northwest Fish Hatchery Newsletter. 5:(2):1-2. LaPatra, S.E. and J.L. Fryer Susceptibility of brown trout (Salmo trutta) to infectious hematopoietic necrosis virus. Bull. Eur. Ass. Fish Pathol. 10(5):125. LaPatra, S.E., J.L. Fryer, W.H. Wingfield and R.P. Hedrick. 1989a. Infectious hematopoietic necrosis virus (IHNV) in coho salmon. J. Aquat. Anim. Health. 1(4): LaPatra, S.E., W.J. Groberg, J.S. Rohovec and J.L. Fryer Size-related susceptibility of salmonids to two strains of infectious hematopoietic necrosis virus. Trans. Am. Fish. Soc. 119(1): LaPatra, S.E., W.G. Groberg, J.S. Rohovec and J.L. Fryer Delayed fertilization of steelhead (Onchorhynchus mykiss) ova to investigate vertical transmission of infectious hematopoietic necrosis virus. Proc. Int'l. Symp. Vir. Low. Vert. Corvallis, Oregon. 2: LaPatra, S.E., J.M. Groff, J.L. Fryer and R.P. Hedrick Comparative pathogenesis of three strains of infectious hematopoietic necrosis virus in rainbow trout Oncorhynchus mykiss. Dis. Aquat. Org. 8(2): LaPatra, S.E., J.E. Parsons, G.R. Jones and W.O. McRoberts Susceptibility of Brook trout and rainbow trout x brook trout hybrids to infectious hematopoietic necrosis virus. FHS/AFS Summer Newsletter 20(2):8-11. LaPatra, S.E., K.A. Roberti, J.S. Rohovec and J.L. Fryer. 1989b. Fluorescent antibody test for the rapid diagnosis of infectious hematopoietic necrosis. J. Aquat. Anim. Health. 1(1): LaPatra, S.E., J.S. Rohovec and J.L. Fryer. 1989c. Detection of infectious hematopoietic necrosis virus in fish mucus. Fish Pathol. 24(4): LaPatra, S.E., T. Turner, K.A. Lauda, G.R. Jones and S. Walker Characterization of the humoral response of rainbow trout to infectious hematopoietic necrosis virus. J. Aquat. Anim. Health 5: Laurencin, F.B IEN in France. Bull. Eur. Ass. Fish Pathol. 7:104. Lazzarini, R.A., J.D. Keene and M. Schubert The origins of defective interfering particles of the negative-stranded RNA viruses. Cell 26: Lenoir, G. and P. dekinkelin Fish rhabdoviruses: Comparative study of protein structure. J. Virol. 16:
108 97 Leong, J.C Molecular and biotechnological approaches to fish vaccines. Curr. Op. Biotech. 4: Leong, J.C., J.L. Fendrick, S. Youngman and A. Lee Effect of polybrene on necrosis virus in tissue culture. J. Fish. Dis. 4: Leong, J.C. and J.L. Fryer Viral vaccines for aquaculture. Ann. Rev. Fish Dis. 93: Leong, J.C., J.L. Fryer and J.R. Winton Vaccination against infectious hematopoietic necrosis, pp In: A.E. Ellis (ed.), Fish vaccination. Academic Press, London. Leong, J.C., Y.L. Hsu and H.M. Engelking Synthesis of the structural proteins of infectious hematopoietic necrosis virus, pp In : J. Crosa (ed.), Bacterial and viral diseases of fish, Molecular studies. University of Washington Press, Seattle, Washington. Leong, J.C., Y.L. Hsu, H.M. Engelking and D. Mulcahy Strains of infectious hematopoietic necrosis (IHN) virus may be identified by structural protein differences. Dev. Biol. Stand. 49: Leong, J.C. and C.B. Munn Potential uses of recombinant DNA in the development of fish vaccines. Bull. Eur. Ass. Fish Pathol. 11(1): Leong, J.C. and S. Turner Isolation of water-borne infectious hematopoietic necrosis virus. Fish Health News. 8(2):6-8. Leppi, T.J.and S.S. Spicer. The histochemistry of mucins on certain primate salivary glands. Am J. Anat. 118: Lillie, R.D. and H.M. Fullmer Histopathological technic and practical histochemistry, ed.4. McGraw-Hill Book Co., New York. Mallory, F.B Pathological technique. Hafner Publishing Co., New York. Masson, P Trichrome stainings and their preliminary technique. J. Tech. Meth. 12: McAllister, P.E., J.L. Fryer and K.S. Pilcher Further characterizarion of infectious hematopoietic necrosis virus of salmonid fish (Oregon strain). Arch. Gesamte Virusforsch. 44: McAllister, P.E. and K.S. Pilcher Autointerference in infectious hematopoietic necrosis virus of salmonid fish. Proc. Soc. Exp. Biol. Med. 145: McAllister, P.E. and W.B. Schill Immunoblot assay: a rapid and sensitive method for identification of salmonid fish viruses. J. Wildl. Dis.22(4): McAllister, P.E. and R.R. Wagner Structural proteins of two salmonid rhabdoviruses. J. Virol. 15(4):
109 98 McAllister, P.E. and R.R. Wagner Virion RNA polymerases of two salmonid rhabdoviruses. J. Virol. 22: McCain, B. B., J.L. Fryer and K.S. Pilcher Antigenic relationships in a group of three viruses of salmonid fish by cross-neutralization. Proceedings of the Society of Experimental Biology and Medicine 146: McCain, B.B., J.L. Fryer and K.S. Pilcher Physiochemical properties of RNA of salmonid hematopoietic necrosis virus (Oregon strain). Proc. Soc. Exp. Biol. Med. 3146: McManus, J.F.A The histological and histochemical uses of periodic acid. Stain Technol. 23: Medina, D.J., P.W. Chang, T.M. Bradley, M.-T. Yeh and E.C. Sadasiv Diagnosis of infectious hematopoietic necrosis virus in Atlantic salmon Salmo salar by enzyme-linked immunosorbent assay. Dis. Aquat. Org. 13(2): Meyers, T.R., J.B. Thomas, J.E. Follett and R.R. Shaft Infectious hematopoietic necrosis virus: Trends in prevalence and risk management approach in Alaskan sockeye salmon culture. J. Aquat. Anim. Health 2: Moscona, A. and R.W. Peluso Persistent infection with human parainfluenza virus 3 in CV-1 cells: Analysis of the role of defective interfering particles. Virology 194: Mulcahy, D. and W.N. Batts Infectious hematopoietic necrosis virus detected by separation and incubation of cells from salmonid cavity fluid. Can. J. Fish. Aquat. Sci. 44: Mulcahy, D. and K. Bauersfeld Effect of loading density of sockeye salmon, Oncorhynchus nerka (Walbaum), eggs in incubation boxes on mortality caused by infectious hematopoietic necrosis. J. Fish Dis. 6(2): Mulcahy, D., J. Burke, R. Pascho and C.K. Jenes Pathogenesis of infectious hematopoietic necrosis virus in adult sockeye salmon (Oncorhynchus nerka). Can. J. Fish. Aquat. Sci. 39(8): Mulcahy, D., C.K. Jenes and R. Pascho. 1984a. Appearance and quantification of infectious hematopoietic necrosis virus in female sockeye salmon (Oncorhynchus nerka) during their spawning migration. Arch. Virol. 80: Mulcahy, D., D. Klaybor and W.N. Batts Isolation of infectious hematopoietic necrosis virus from a leech (Piscicola salmositica) and a copepod (Salmincola sp.), ectoparasites of sockeye salmon Oncorhynchus nerka. Dis. Aquat. Org. 8: Mulcahy, D. and R. Pascho Adsorption to fish sperm of vertically transmitted fish viruses. Science 225(4659):
 and steelhead trout (Salmo gairdneri) for infectious hematopoietic necrosis virus. Can. J.")
110 99 Mulcahy, D.M. and R.J. Pascho Vertical transmission of infectious hematopoietic necrosis virus in sockeye salmon (Oncorhynchus nerka): isolation of virus from dead eggs and fry. J. Fish Dis. 8: Mulcahy, D., R. Pascho and W.N. Batts Testing of male sockeye salmon (Oncorhynchus nerka) and steelhead trout (Salmo gairdneri) for infectious hematopoietic necrosis virus. Can. J. Fish. Aquat. Sci. 44(5): Mulcahy, D., R. Pascho and C.K. Jenes. 1983a. Detection of infectious hematopoietic necrosis virus in river water and demonstration of waterborne transmission. J. Fish Dis. 6: Mulcahy, D., R. Pascho and C.K. Jenes. 1983b. Mortality due to infectious hematopoietic necrosis of sockeye salmon (Oncorhynchus nerka) fry in streamside egg incubation boxes. Can. J. Fish. Aquat. Sci. 40(9): Mulcahy, D., R. Pascho and C.K. Jenes. 1983c. Titre distribution patterns of infectious hematopoietic necrosis virus in ovarian fluids of hatchery and feral salmon populations. J. Fish Dis. 6: Mulcahy, D., R. Pascho and C.K. Jenes. 1984b. Comparison of in vitro growth characteristics of ten isolates of infectious hematopoietic necrosis virus. J. Gen. Virol. 65: Neukirch, M An experimental study of the entry and multiplication of viral hemorrhagic septicaemia virus in rainbow trout, Salmo gairdneri Richardson, after water-borne infection. J. Fish Dis. 7: Nibert, M.L., D.B. Furlong and B.N. Fei lds Mechanisms of viral pathogenesis: Distinct forms of reoviruses and their roles during replication in cells and host. J. Clin. Invest. 88: Nicholson, B.L. and C. Byrne An established cell line from the Atlantic salmon (Salmo salar). J. Fish Res. Board Can. 30: Nims, L., J.L. Fryer and K.S. Pilcher Studies of replication of four selected viruses in two cell lines derived from salmonid fish. Proc. Soc. Exp. Biol. Med. 135:6-12. Nishimura, T., Y. Ishida, S. Yamamoto, H. Fekuda, N. Okamoto and T. Sano Infectious hematopoietic necrosis: Virus titer in the fish bodies, rearing water and feces of artificially infected rainbow trout fry. Fish Pathol. 23(1): Nishimura, T., M. Sano, F. Toyama, Y. Kanai and T. Sano Comparison between EPC and FHM cell lines on IHN virus susceptibility and effect of fetal bovine serum concentration on IHNV CPE appearance. Fish Pathol. 20(1): Oberg, L.A., J. Wirkkula, D. Mourich and J.C. Leong Bacterially expressed nucleoprotein of infectious hematopoietic necrosis virus augments protective immunity induced by the glycoprotein vaccine in fish. J. Virol. 65(8):
111 100 Okamoto, N., N. Shirakura, Y. Nagakura and T. Sano The mechanism of interference with fish viral infection in the RTG-2 cell line. Fish Pathol. 18(1):7-12. Park, M.A., S.G. Sohn, S.D. Lee, S.K. Chun, J.W. Park, J.L. Fryer and Y.C. Yah Infectious haematopoietic necrosis virus from salmonids cultured in Korea. J. Fish Dis. 16: Parisot, R.J., W.T. Yasutake and G.W. Klontz Virus diseases of the Salmonidae in western United States. I. Etiology and Epizootiology. Ann. N.Y. Acad. Sci. 126: Pattniak, A.K. and G.W. Wertz Cells that express all five proteins of vesicular stomatitis virus from cloned cdnas support replication, assembly, and budding of defective interfering particles. PNAS, USA. 88: Perrault, J Origin and replication of defective interfering particles. Curr. Top. Microbiol. Immunol. 93: Pedley, S., F. Hundley, I. Chrystie, M.A. McCrae and U. Desselberger The genomes of rotaviruses isolated from chronically infected immunodeficient children. J. Gen. Virol. 65: Pietsch, J.P., D.F. Amend and S.M. Miller Survival of infectious hematopoietic necrosis virus held under various environmental conditions. J. Fish. Res. Board Can. 34(9): Pilcher, K.S. and J. L. Fryer The viral diseases of fish: A review through Part I: Diseases of proven etiology. CRC Critical Reviews in Microbiology 7: Ponder, B.A. and M.M. Wilkinson Inhibition of endogenous tissue alkaline phosphatase with the use of alkaline phosphatase conjugates in immunohistochemistry. J. Histochem. Cytochem. 29(8): Ramseur, J.M. and R.M. Friedman Prolonged infection of L cells with vesicular stomatitis virus: Defective interfering forms and temperature-sensitive mutants as factors in the infection. Virology 85: Reichmann, M.E. and W.M. Schnitzlein Rhabdovirus defective particles: origin and genome assignments, pp In: D.L. Bishop (ed.), Rhabdoviruses. Volume 2. CRC Press Inc., Boca Raton, Florida. Ristow, S.S. and J.M. Arnzen Development of monoclonal antibodies that recognize a Type-2 specific and a common epitope on the nucleoprotein of infectious hematopoietic necrosis virus. J. Aquat. Anim. Health. 1: Ristow, S.S. and J.A. de Avila Monoclonal antibodies to the glycoprotein and nucleoprotein of infectious hematopoietic necrosis virus (IHNV) reveal differences among isolates of the virus by fluorescence, neutralization and electrophoresis. Dis. Aquat. Org. 11:
 and infectious hematopoietic necrosis virus (IENV). J. Aquat. Anim. Health. 3(3):176-180.")
112 101 Ristow, S.S., N. Lorenzen and P.E.V. Jorgensen Monoclonal-antibody based immunodot assay distinguishes between viral hemorrhagic septicemia virus (VHSV) and infectious hematopoietic necrosis virus (IENV). J. Aquat. Anim. Health. 3(3): Rucker, R.R., W.J. Whipple, J.R. Parvin and C.A. Evans A contagious disease of salmon possibly of virus origin. US Fish and Wildl. Serv. Fish. Bull. 54(76):35. Sambrook, J., E.F. Fritsch and T. Maniatis, editors Analysis of genomic DNA by southern hybridization, pp In Molecular Cloning, A laboratory Manual, Second Edition, volume 2, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York. Sano, T., T. Nishimura, N. Okamoto, T. Yamazaki, H. Hanada and Y. Watanabe Studies on viral diseases of Japanese fishes. Infectious hematopoietic necrosis (IHN) of salmonids in the mainland of Japan. J. Tokyo. Univ. Fish. 63(2): Schmitz, G.G., T. Walter, R. Seibl and C. Kessler Nonradioactive labeling of oligonucleotides in vitro with the hapten digoxigenin by tailing with terminal transferase. Anal. Biochem. 192: Schultz, C.L., P.E. McAllister, W.B. Schill, B.C. Lidgerding and F.M. Hetrick Detection of infectious hematopoietic necrosis virus in cell culture fluid using immunoblot assay and biotinylated monoclonal antibody. Dis. Aquat. Org. 7(1):31-37 Scott, J.L., J.L. Fendrick and J.C. Leong Growth of infectious hematopoietic necrosis virus in mosquito and fish cell lines. Wassman J. Biol. 38: Sekellick, M.J. and P.I. Marcus Persistent infections by rhabdoviruses, pp In: D.L. Bishop (ed.), Rhabdoviruses. Volume 3. CRC Press, Inc., Boca Raton, Florida. Sharpe, A.H. and B.N. Feilds Pathogenesis of reovirus infection, pp In: W.K. Joklik (ed.), The Reoviridae. Plenum Publishing Corp., New York. Shors, S.T. and V. Winston. 1989a. Neutralizing antibodies for infectious hematopoietic necrosis virus in eggs of steelhead trout. Am. J. Vet. Res. 50(2): Shors, S.T. and V. Winston. 1989b. Detection of infectious hematopoietic necrosis virus in an invertebrate (Callibaetis sp.). Am J. Vet. Res. 50(8): Smart G.R., D. Knox, J.G. Harrison, J.A. Ralph, R.H. Richards and C.B. Cowley Nephrocalcinosis in rainbow trout (Salmo gairdneri) Richardson: effect of exposure to elevated CO2 concentrations. J. Fish. Dis. 2: Tatner, M.F. and M.J. Manning. 1983a. Growth of the lymphoid organs in rainbow trout, Salmo gairdneri from one to fifteen months of age. J. Zool. Lond. 199:
 in Great Central Lake, British Columbia. Dept. Fish. Oceans.")
113 102 Tatner, M.F. and M.J. Manning. 1983b. The ontogeny of cellular immunity in the rainbow trout Salmo gairdneri Richardson, in relation to the stage of development of the lymphoid organ. Dev. Comp. Immunol. 7: Traxler, G.S A survey for infectious hematopoietic necrosis virus in adult sockeye salmon (Oncorhynchus nerka) in Great Central Lake, British Columbia. Dept. Fish. Oceans. Nanaimo, B.C. pp. 14. Wagner, R.R Reproduction of rhabdoviruses, pp In: H. Fraenkel-Conrat and R.R. Wagner (eds.), Comprehensive Virology, Vol 4. Plenum Press, New York. Watanabe, T Persistent infection in YNK cells with infectious hematopoietic necrosis virus (IHNV). Bull. Jap. Soc. Sci. Fish. -Nissuishi. 49(2): Way, K. and P.F. Dixon Rapid detection of VHS and IHN viruses by the enzyme-linked immunosorbent assay (ELISA). J. Appl. Ichthyol. 4(4): Williams, MA Autoradiography and immunocytochemistry, pp In: A.M. Glauert (ed.),.practical Methods in Electron Microscopy. North-Holland Publishing company, Amsterdam. Williams, I.V. and D. Amend A natural epizootic of infectious hematopoietic necrosis in fry of sockeye salmon (Oncorhynchus nerka) at Chilko Lake, British Columbia. J. Fish. Res. Board Can. 33: Wingfeild, W.H., J.L. Fryer and K.S. Pilcher Properties of the sockeye salmon virus (Oregon strain). Proc. Soc. Exp. Biol. Med. 130: Winton, J.R Recent advances in detection and control of infectious hematopoietic necrosis virus in aquaculture. Ann. Rev. Fish Dis Winton J.R., C.K. Arakawa, C.N. Lannan and J.L. Fryer Neutralizing monoclonal antibodies recognize antigenic variants among isolates of infectious hematopoietic necrosis virus. Dis. Aquat. Org. 4: Wolf, K., (ed.) Infectious hematopoietic necrosis, pp In: Fish Viruses and fish viral diseases. Cornell University Press, Ithaca, New York. Wolf, K. and M.C. Quimby Established eurythermic line of fish cells in vitro Science 135: Wright, D.K. and M.M. Manos Sample preparation from paraffin-embedded tissues, pp In: M.A. Innis, D.H. Gelfand, J.J. Sninsky, and T.J. White (eds.), PCR Protocols. Academic Press, New York. Xu, L., D.V. Mourich, H.M. Engelking, S. Ristow, J. Arnzen and J.C. Leong Epitope mapping and characterization of the infectious hematopoietic necrosis virus glycoprotein, using fusion proteins synthesized in Escherichia coll. J. Virol. 65(3):
LITERATURE REVIEW Viral Hemorrhagic Septicemia Virus VHSV
 LITERATURE REVIEW Viral Hemorrhagic Septicemia Virus VHSV PELTON ROUND BUTTE HYDROELECTRIC PROJECT FERC No. 2030 Prepared for PGE by: H. Mark Engelking Oregon Department of Fish and Wildlife Fish Pathology
LITERATURE REVIEW Viral Hemorrhagic Septicemia Virus VHSV PELTON ROUND BUTTE HYDROELECTRIC PROJECT FERC No. 2030 Prepared for PGE by: H. Mark Engelking Oregon Department of Fish and Wildlife Fish Pathology
2.3.1 Other Viruses Isolated from Fish
 2.3.1 Other Viruses Isolated from Fish - 1 2.3.1 Other Viruses Isolated from Fish William N. Batts and James R. Winton USGS Western Fisheries Research Center 6505 NE 65th Street, Seattle, WA 98115 Phone:
2.3.1 Other Viruses Isolated from Fish - 1 2.3.1 Other Viruses Isolated from Fish William N. Batts and James R. Winton USGS Western Fisheries Research Center 6505 NE 65th Street, Seattle, WA 98115 Phone:
FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
 3. Salmon and Trout Viral Diseases Hisae Kasai and Mamoru Yoshimizu 3.1. Synopsis Transmissible diseases of socio-economic importance must be controlled within national boundaries. Salmon and trout viral
3. Salmon and Trout Viral Diseases Hisae Kasai and Mamoru Yoshimizu 3.1. Synopsis Transmissible diseases of socio-economic importance must be controlled within national boundaries. Salmon and trout viral
DISEASES OF AQUATIC ORGANISMS Vol. 59: , 2004 Published June 11 Dis Aquat Org
 DISEASES OF AQUATIC ORGANISMS Vol. 59: 211 216, 2004 Published June 11 Dis Aquat Org Susceptibility of captive adult winter-run Chinook salmon Oncorhynchus tshawytscha to waterborne exposures with infectious
DISEASES OF AQUATIC ORGANISMS Vol. 59: 211 216, 2004 Published June 11 Dis Aquat Org Susceptibility of captive adult winter-run Chinook salmon Oncorhynchus tshawytscha to waterborne exposures with infectious
DNA Vaccination Against IHN Virus: A Disease of Trout and Salmon
 DNA Vaccination Against IHN Virus: A Disease of Trout and Salmon Scott E. LaPatra Clear Springs Foods, Inc., Research Division, Buhl, Idaho, U.S.A. Infectious Hematopoietic Necrosis Virus (IHNV) Family:
DNA Vaccination Against IHN Virus: A Disease of Trout and Salmon Scott E. LaPatra Clear Springs Foods, Inc., Research Division, Buhl, Idaho, U.S.A. Infectious Hematopoietic Necrosis Virus (IHNV) Family:
2.2.9 Spring Viremia of Carp
 2.2.9 Spring Viremia of Carp - 1 2.2.9 Spring Viremia of Carp A. E. Goodwin and J. R. Winton* Aquaculture/Fisheries Center University of Arkansas at Pine Bluff 1200 N. University Drive Mail Slot 4912 Pine
2.2.9 Spring Viremia of Carp - 1 2.2.9 Spring Viremia of Carp A. E. Goodwin and J. R. Winton* Aquaculture/Fisheries Center University of Arkansas at Pine Bluff 1200 N. University Drive Mail Slot 4912 Pine
Inactivation of Infectious Hematopoietic Necrosis Virus by Low Levels of Iodine
 APPLIED AND ENVIRONMENTAL MICROBIOLOGY, May 1991, p. 1379-1385 0099-2240/91/051379-07$02.00/0 Copyright ) 1991, American Society for Microbiology Vol. 57, No. 5 Inactivation of Infectious Hematopoietic
APPLIED AND ENVIRONMENTAL MICROBIOLOGY, May 1991, p. 1379-1385 0099-2240/91/051379-07$02.00/0 Copyright ) 1991, American Society for Microbiology Vol. 57, No. 5 Inactivation of Infectious Hematopoietic
Virulence comparison of different electropherotypes of infectious hematopoietic necrosis virus
 Published Vol. 16: 115-120. 1993 DISEASES OF AQUATIC ORGANISMS Dis. aquat. Org. ~ August 5 Virulence comparison of different electropherotypes of infectious hematopoietic necrosis virus 'Oregon Department
Published Vol. 16: 115-120. 1993 DISEASES OF AQUATIC ORGANISMS Dis. aquat. Org. ~ August 5 Virulence comparison of different electropherotypes of infectious hematopoietic necrosis virus 'Oregon Department
Infectious Salmon Anemia
 Infectious Salmon Anemia A paradigm shift for understanding risk of ISAV infection Jill Rolland US Geological Survey Western Fisheries Research Center Seattle, Washington Infectious Salmon Anemia history
Infectious Salmon Anemia A paradigm shift for understanding risk of ISAV infection Jill Rolland US Geological Survey Western Fisheries Research Center Seattle, Washington Infectious Salmon Anemia history
The surveillance and control programme for viral haemorrhagic septicaemia
 Annual Reports 2008 Surveillance and control programmes for terrestrial and aquatic animals in Norway National Veterinary Institute The surveillance and control programme for viral haemorrhagic septicaemia
Annual Reports 2008 Surveillance and control programmes for terrestrial and aquatic animals in Norway National Veterinary Institute The surveillance and control programme for viral haemorrhagic septicaemia
G. Kalayci*, S. Incoglu and B. Özkan
 Bull. Eur. Ass. Fish Pathol., 26(4) 2006, 157 First isolation of viral haemorrhagic septicaemia (VHS) virus from turbot (Scophthalmus maximus) cultured in the Trabzon coastal area of the Black Sea in Turkey
Bull. Eur. Ass. Fish Pathol., 26(4) 2006, 157 First isolation of viral haemorrhagic septicaemia (VHS) virus from turbot (Scophthalmus maximus) cultured in the Trabzon coastal area of the Black Sea in Turkey
Fish Health Management
 Fish Health Management 9 JAMES R. WINTON Introduction Maintaining the health of hatchery fish requires a program of good management and continuous attention to detail. In its most basic sense, disease
Fish Health Management 9 JAMES R. WINTON Introduction Maintaining the health of hatchery fish requires a program of good management and continuous attention to detail. In its most basic sense, disease
Epidemiological characteristics of infectious hematopoietic necrosis virus (IHNV): a review
: a review") DOI 10.1186/s13567-016-0341-1 REVIEW Epidemiological characteristics of infectious hematopoietic necrosis virus (IHNV): a review Peter Dixon 1, Richard Paley 1, Raul Alegria Moran 2 and Birgit Oidtmann
DOI 10.1186/s13567-016-0341-1 REVIEW Epidemiological characteristics of infectious hematopoietic necrosis virus (IHNV): a review Peter Dixon 1, Richard Paley 1, Raul Alegria Moran 2 and Birgit Oidtmann
AN ABSTRACT OF THE THESIS OF VIRUSES INFECTING SALMONID FISHES FROM OREGON PANCREATIC NECROSIS VIRUS THE IMMUNIZATION OF SALMONIDS
 AN ABSTRACT OF THE THESIS OF GUY LEE TEBBIT for the degree of Doctor of Philosophy in Microbiology presented on September 22, 1975 Title: VIRUSES INFECTING SALMONID FISHES FROM OREGON A. THE OCCURRENCE
AN ABSTRACT OF THE THESIS OF GUY LEE TEBBIT for the degree of Doctor of Philosophy in Microbiology presented on September 22, 1975 Title: VIRUSES INFECTING SALMONID FISHES FROM OREGON A. THE OCCURRENCE
Title. Author(s)Yoshimizu, Mamoru. Issue Date Doc URL. Type. File Information. Control Strategy for Viral Diseases of Salmonids and
Yoshimizu, Mamoru. Issue Date Doc URL. Type. File Information. Control Strategy for Viral Diseases of Salmonids and") Title Control Strategy for Viral Diseases of Salmonids and Author(s)Yoshimizu, Mamoru Issue Date 2003 Doc URL http://hdl.handle.net/2115/39590 Type bookchapter (author version) File Information yoshimizu-180i.pdf
Title Control Strategy for Viral Diseases of Salmonids and Author(s)Yoshimizu, Mamoru Issue Date 2003 Doc URL http://hdl.handle.net/2115/39590 Type bookchapter (author version) File Information yoshimizu-180i.pdf
Identification of Microbes Lecture: 12
 Diagnostic Microbiology Identification of Microbes Lecture: 12 Electron Microscopy 106 virus particles per ml required for visualization, 50,000-60,000 magnification normally used. Viruses may be detected
Diagnostic Microbiology Identification of Microbes Lecture: 12 Electron Microscopy 106 virus particles per ml required for visualization, 50,000-60,000 magnification normally used. Viruses may be detected
Pathogenicity of birnaviruses isolated from turbot (Scophthalmus maximus) : comparison with reference serotypes of IPNV
 : comparison with reference serotypes of IPNV") Pathogenicity of birnaviruses isolated from turbot (Scophthalmus maximus) : comparison with reference serotypes of IPNV B. Novoa a, C. Rivas b, A.E. Toranzo b, A. Figueras a,* a Institute de Investigaciones
Pathogenicity of birnaviruses isolated from turbot (Scophthalmus maximus) : comparison with reference serotypes of IPNV B. Novoa a, C. Rivas b, A.E. Toranzo b, A. Figueras a,* a Institute de Investigaciones
2.2.6 Viral Erythrocytic Necrosis
 2.2.7 Viral Erythrocytic Necrosis - 1 2.2.6 Viral Erythrocytic Necrosis J. R. Winton 1 and P. K. Hershberger 2 1 USGS Western Fisheries Research Center 6505 NE 65 th Street Seattle, WA 98115 and 2 USGS
2.2.7 Viral Erythrocytic Necrosis - 1 2.2.6 Viral Erythrocytic Necrosis J. R. Winton 1 and P. K. Hershberger 2 1 USGS Western Fisheries Research Center 6505 NE 65 th Street Seattle, WA 98115 and 2 USGS
Viral structure م.م رنا مشعل
 Viral structure م.م رنا مشعل Viruses must reproduce (replicate) within cells, because they cannot generate energy or synthesize proteins. Because they can reproduce only within cells, viruses are obligate
Viral structure م.م رنا مشعل Viruses must reproduce (replicate) within cells, because they cannot generate energy or synthesize proteins. Because they can reproduce only within cells, viruses are obligate
Phylogeography of infectious haematopoietic necrosis virus in North America
 Journal of General Virology (2003), 84, 803 814 DOI 10.1099/vir.0.18771-0 Phylogeography of infectious haematopoietic necrosis virus in North America Gael Kurath, 1 Kyle A. Garver, 1,2 Ryan M. Troyer,
Journal of General Virology (2003), 84, 803 814 DOI 10.1099/vir.0.18771-0 Phylogeography of infectious haematopoietic necrosis virus in North America Gael Kurath, 1 Kyle A. Garver, 1,2 Ryan M. Troyer,
The Open Access Israeli Journal of Aquaculture Bamidgeh
 The Open Access Israeli Journal of Aquaculture Bamidgeh As from January 2010 The Israeli Journal of Aquaculture - Bamidgeh (IJA) will be published exclusively as an on-line Open Access (OA) quarterly accessible
The Open Access Israeli Journal of Aquaculture Bamidgeh As from January 2010 The Israeli Journal of Aquaculture - Bamidgeh (IJA) will be published exclusively as an on-line Open Access (OA) quarterly accessible
Persistent Infection of MDCK Cells by Influenza C Virus: Initiation and Characterization
 J. gen. Virol. (199), 70, 341-345. Printed in Great Britain 341 Key words: influenza C virus/interferon/persistent infection Persistent Infection of MDCK Cells by Influenza C Virus: Initiation and Characterization
J. gen. Virol. (199), 70, 341-345. Printed in Great Britain 341 Key words: influenza C virus/interferon/persistent infection Persistent Infection of MDCK Cells by Influenza C Virus: Initiation and Characterization
INCLUSION BODY HEPATITIS AND HYDROPERICARDIUM SYNDROME (ADENOVIRUS INFECTIONS)
") INCLUSION BODY HEPATITIS AND HYDROPERICARDIUM SYNDROME (ADENOVIRUS INFECTIONS) AVIAN ADENOVIRUSES (CHICKEN ADENOVIRUSES, FADV) Adenoviruses are common in poultry. Many replicate in healthy birds without
INCLUSION BODY HEPATITIS AND HYDROPERICARDIUM SYNDROME (ADENOVIRUS INFECTIONS) AVIAN ADENOVIRUSES (CHICKEN ADENOVIRUSES, FADV) Adenoviruses are common in poultry. Many replicate in healthy birds without
Infectious hematopoietic necrosis virus antibody profiles in naturally and experimentally infected Atlantic salmon Salmo salar
 DISEASES OF AQUATIC ORGANISMS Vol. 46: 7 14, 2001 Published August 22 Dis Aquat Org Infectious hematopoietic necrosis virus antibody profiles in naturally and experimentally infected Atlantic salmon Salmo
DISEASES OF AQUATIC ORGANISMS Vol. 46: 7 14, 2001 Published August 22 Dis Aquat Org Infectious hematopoietic necrosis virus antibody profiles in naturally and experimentally infected Atlantic salmon Salmo
Detection of virus level in tissues of rainbow trout, Oncoryhinchus mykiss in clinical stage of viral hemorrhagic septicemia
 Downloaded from orbit.dtu.dk on: Nov 08, 2017 Detection of virus level in tissues of rainbow trout, Oncoryhinchus mykiss in clinical stage of viral hemorrhagic septicemia Ghiasi, Farzad; Olesen, Niels
Downloaded from orbit.dtu.dk on: Nov 08, 2017 Detection of virus level in tissues of rainbow trout, Oncoryhinchus mykiss in clinical stage of viral hemorrhagic septicemia Ghiasi, Farzad; Olesen, Niels
Characteristics of three rhabdoviruses from snakehead fish Ophicephalus striatus
 Vol. 13: 89-94, 1992 l DISEASES OF AQUATIC ORGANISMS Dis. aquat. Org. Published July 23 Characteristics of three rhabdoviruses from snakehead fish Ophicephalus striatus 'Department of Microbiology, Oregon
Vol. 13: 89-94, 1992 l DISEASES OF AQUATIC ORGANISMS Dis. aquat. Org. Published July 23 Characteristics of three rhabdoviruses from snakehead fish Ophicephalus striatus 'Department of Microbiology, Oregon
Some living things are made of ONE cell, and are called. Other organisms are composed of many cells, and are called. (SEE PAGE 6)
") Section: 1.1 Question of the Day: Name: Review of Old Information: N/A New Information: We tend to only think of animals as living. However, there is a great diversity of organisms that we consider living
Section: 1.1 Question of the Day: Name: Review of Old Information: N/A New Information: We tend to only think of animals as living. However, there is a great diversity of organisms that we consider living
DISEASES OF AQUATIC ORGANISMS Dis. aquat. Org.
 Vol. 12: 229-233.1992 DISEASES OF AQUATIC ORGANISMS Dis. aquat. Org. Published April 23 NOTE Erythrocytic inclusion body syndrome: a light and electron microscopic study of infected erythrocytes of chinook
Vol. 12: 229-233.1992 DISEASES OF AQUATIC ORGANISMS Dis. aquat. Org. Published April 23 NOTE Erythrocytic inclusion body syndrome: a light and electron microscopic study of infected erythrocytes of chinook
Chapter 6- An Introduction to Viruses*
 Chapter 6- An Introduction to Viruses* *Lecture notes are to be used as a study guide only and do not represent the comprehensive information you will need to know for the exams. 6.1 Overview of Viruses
Chapter 6- An Introduction to Viruses* *Lecture notes are to be used as a study guide only and do not represent the comprehensive information you will need to know for the exams. 6.1 Overview of Viruses
Redacted for Privacy. L. Fryer/ the first reported case of double virus infection in fish. The isolation
 AN ABSTRACT OF THE THESIS OF DANIEL MICHAEL MULCAHY for the degree of Doctor of Philosophy in Microbiology presented on December 9. 1976 Title: DISTRIBUTION AND EPIZOOTIOLOGY OF INFECTIOUS PANCREATIC NECROSIS
AN ABSTRACT OF THE THESIS OF DANIEL MICHAEL MULCAHY for the degree of Doctor of Philosophy in Microbiology presented on December 9. 1976 Title: DISTRIBUTION AND EPIZOOTIOLOGY OF INFECTIOUS PANCREATIC NECROSIS
Coronaviruses cause acute, mild upper respiratory infection (common cold).
.") Coronaviruses David A. J. Tyrrell Steven H. Myint GENERAL CONCEPTS Clinical Presentation Coronaviruses cause acute, mild upper respiratory infection (common cold). Structure Spherical or pleomorphic enveloped
Coronaviruses David A. J. Tyrrell Steven H. Myint GENERAL CONCEPTS Clinical Presentation Coronaviruses cause acute, mild upper respiratory infection (common cold). Structure Spherical or pleomorphic enveloped
Temperature-Sensitive Mutants Isolated from Hamster and
 JOURNAL OF VIROLOGY, Nov. 1975, p. 1332-1336 Copyright i 1975 American Society for Microbiology Vol. 16, No. 5 Printed in U.S.A. Temperature-Sensitive Mutants Isolated from Hamster and Canine Cell Lines
JOURNAL OF VIROLOGY, Nov. 1975, p. 1332-1336 Copyright i 1975 American Society for Microbiology Vol. 16, No. 5 Printed in U.S.A. Temperature-Sensitive Mutants Isolated from Hamster and Canine Cell Lines
Viruse associated gastrointestinal infection
 Viruse associated gastrointestinal infection Dr. Hala Al Daghistani Rotaviruses Rotaviruses are a major cause of diarrheal illness in human (infants), and young animals, including calves and piglets. Infections
Viruse associated gastrointestinal infection Dr. Hala Al Daghistani Rotaviruses Rotaviruses are a major cause of diarrheal illness in human (infants), and young animals, including calves and piglets. Infections
Genetic analyses reveal unusually high diversity of infectious haematopoietic necrosis virus in rainbow trout aquaculture
 Journal of General Virology (2000), 81, 2823 2832. Printed in Great Britain... Genetic analyses reveal unusually high diversity of infectious haematopoietic necrosis virus in rainbow trout aquaculture
Journal of General Virology (2000), 81, 2823 2832. Printed in Great Britain... Genetic analyses reveal unusually high diversity of infectious haematopoietic necrosis virus in rainbow trout aquaculture
Ali Alabbadi. Bann. Bann. Dr. Belal
 31 Ali Alabbadi Bann Bann Dr. Belal Topics to be discussed in this sheet: Particles-to-PFU Single-step and multi-step growth cycles Multiplicity of infection (MOI) Physical measurements of virus particles
31 Ali Alabbadi Bann Bann Dr. Belal Topics to be discussed in this sheet: Particles-to-PFU Single-step and multi-step growth cycles Multiplicity of infection (MOI) Physical measurements of virus particles
Disinfection of unfertilized salmonid eggs: a new method for prevention of vertical transmission of Flavobacterium psychrophilum
 Disinfection of unfertilized salmonid eggs: a new method for prevention of vertical transmission of Flavobacterium psychrophilum Akira Kumagai Miyagi Prefecture Fisheries Technology Institute Freshwater
Disinfection of unfertilized salmonid eggs: a new method for prevention of vertical transmission of Flavobacterium psychrophilum Akira Kumagai Miyagi Prefecture Fisheries Technology Institute Freshwater
INTERNATIONAL CONFERENCE ON HARMONISATION OF TECHNICAL REQUIREMENTS FOR REGISTRATION OF PHARMACEUTICALS FOR HUMAN USE. ICH Considerations
 INTERNATIONAL CONFERENCE ON HARMONISATION OF TECHNICAL REQUIREMENTS FOR REGISTRATION OF PHARMACEUTICALS FOR HUMAN USE ICH Considerations General Principles to Address Virus and Vector Shedding 1.0 Introduction
INTERNATIONAL CONFERENCE ON HARMONISATION OF TECHNICAL REQUIREMENTS FOR REGISTRATION OF PHARMACEUTICALS FOR HUMAN USE ICH Considerations General Principles to Address Virus and Vector Shedding 1.0 Introduction
ODFW Fish Virology: Investigations into Oral Tumors of. Title Grande Ronde Spring Chinook
 ODFW Fish Virology: Investigations into Oral Tumors of Title Grande Ronde Spring Chinook John Kaufman, Fish Health Services, ODFW 220 Nash Hall Department of Microbiology OSU, Corvallis, OR 97331-3804
ODFW Fish Virology: Investigations into Oral Tumors of Title Grande Ronde Spring Chinook John Kaufman, Fish Health Services, ODFW 220 Nash Hall Department of Microbiology OSU, Corvallis, OR 97331-3804
Session 2. TiLV isolation and Koch s Postulates
 Session 2 Win Surachetpong DVM, PhD, CertAqV, DTBVP Kathy Tang-Nelson PhD TiLV isolation and Koch s Postulates Learning objectives Describe how viruses are isolated Apply the appropriate method to the
Session 2 Win Surachetpong DVM, PhD, CertAqV, DTBVP Kathy Tang-Nelson PhD TiLV isolation and Koch s Postulates Learning objectives Describe how viruses are isolated Apply the appropriate method to the
Screening of Bacteria with Antiviral Activity from Fresh Water Salmonid Hatcheries
 Title Screening of Bacteria with Antiviral Activity from W Author(s)Kamei, Yuto; Yoshimizu, Mamoru; Ezura, Yoshio; Kimur CitationMicrobiology and Immunology, 32(1): 6773 Issue Date 1988 Doc URL http://hdl.handle.net/2115/39493
Title Screening of Bacteria with Antiviral Activity from W Author(s)Kamei, Yuto; Yoshimizu, Mamoru; Ezura, Yoshio; Kimur CitationMicrobiology and Immunology, 32(1): 6773 Issue Date 1988 Doc URL http://hdl.handle.net/2115/39493
HEPATITIS VIRUSES. Other causes (not exclusively hepatitis v.)also called sporadic hepatitis: HEPATITIS A(infectious hepatitis)
also called sporadic hepatitis: HEPATITIS A(infectious hepatitis)") Dept.of Microbiology/Virology Assist.prof. Shatha F. Abdullah HEPATITIS VIRUSES Medically important hepatitis v. (liver)are: 1.HAV 2.HBV 3.HCV 4.HDV 5.HEV 6.HGV Other causes (not exclusively hepatitis
Dept.of Microbiology/Virology Assist.prof. Shatha F. Abdullah HEPATITIS VIRUSES Medically important hepatitis v. (liver)are: 1.HAV 2.HBV 3.HCV 4.HDV 5.HEV 6.HGV Other causes (not exclusively hepatitis
Redacted for Privacy
 AN ABSTRACT OF THE THESIS OF Carol A. Shanks for the degree of Master of Science in Microbiology presented on September 11, 1991. Title: Erythrocytic Inclusion Body Syndrome: Salmonid Stock Susceptibility,
AN ABSTRACT OF THE THESIS OF Carol A. Shanks for the degree of Master of Science in Microbiology presented on September 11, 1991. Title: Erythrocytic Inclusion Body Syndrome: Salmonid Stock Susceptibility,
Rabies. By Rosa Reynoso. Rabies; Etiological Agent: Rhabdoviridae family Lyssavirus genus type (4)
") Rabies By Rosa Reynoso Rabies; Etiological Agent: Rhabdoviridae family Lyssavirus genus type (4) Transmision: It is a zoonotic disease transmitted from infected animals to humans (4). Infected animals
Rabies By Rosa Reynoso Rabies; Etiological Agent: Rhabdoviridae family Lyssavirus genus type (4) Transmision: It is a zoonotic disease transmitted from infected animals to humans (4). Infected animals
West Nile Virus. Family: Flaviviridae
 West Nile Virus 1 Family: Flaviviridae West Nile Virus Genus: Flavivirus Japanese Encephalitis Antigenic Complex Complex Includes: Alfuy, Cacipacore, Japanese encephalitis, koutango, Kunjin, Murray Valley
West Nile Virus 1 Family: Flaviviridae West Nile Virus Genus: Flavivirus Japanese Encephalitis Antigenic Complex Complex Includes: Alfuy, Cacipacore, Japanese encephalitis, koutango, Kunjin, Murray Valley
Gastroenteritis and viral infections
 Gastroenteritis and viral infections A Large number of viruses are found in the human gut; these include some that are associated with gastroenteritis Rotaviruses Adenoviruses 40/41 Caliciviruses Norwalk-like
Gastroenteritis and viral infections A Large number of viruses are found in the human gut; these include some that are associated with gastroenteritis Rotaviruses Adenoviruses 40/41 Caliciviruses Norwalk-like
Infection Basics. Lecture 12 Biology W3310/4310 Virology Spring 2016
 Infection Basics Lecture 12 Biology W3310/4310 Virology Spring 2016 Before I came here I was confused about this subject. Having listened to your lecture, I am still confused but at a higher level. ENRICO
Infection Basics Lecture 12 Biology W3310/4310 Virology Spring 2016 Before I came here I was confused about this subject. Having listened to your lecture, I am still confused but at a higher level. ENRICO
Elucidate Traffic Patterns Of Ihn Virus In The Columbia River Basin
 PART I - ADMINISTRATIVE Section 1. General administrative information Title of project Elucidate Traffic Patterns Of Ihn Virus In The Columbia River Basin BPA project number: 20056 Contract renewal date
PART I - ADMINISTRATIVE Section 1. General administrative information Title of project Elucidate Traffic Patterns Of Ihn Virus In The Columbia River Basin BPA project number: 20056 Contract renewal date
Patricia Fitzgerald-Bocarsly
 FLU Patricia Fitzgerald-Bocarsly October 23, 2008 Orthomyxoviruses Orthomyxo virus (ortho = true or correct ) Negative-sense RNA virus (complementary to mrna) Five different genera Influenza A, B, C Thogotovirus
FLU Patricia Fitzgerald-Bocarsly October 23, 2008 Orthomyxoviruses Orthomyxo virus (ortho = true or correct ) Negative-sense RNA virus (complementary to mrna) Five different genera Influenza A, B, C Thogotovirus
CHICKEN INFECTIOUS ANEMIA
 CHICKEN INFECTIOUS ANEMIA Slide study set # 20 Prepared by: Joan A. Smyth Department of Pathobiology and Veterinary Science University of Connecticut 61 North Eagleville Road Storrs, CT 06269-3089, H.
CHICKEN INFECTIOUS ANEMIA Slide study set # 20 Prepared by: Joan A. Smyth Department of Pathobiology and Veterinary Science University of Connecticut 61 North Eagleville Road Storrs, CT 06269-3089, H.
Redacted for Privacy
 AN ABSTRACT OF THE THESIS OF Hamdi Ogut for the degree of Master of Science in Fisheries Sicence presented on December 20. 1995. Title: IN VITRO HOST RANGE OF INFECTIOUS PANCREATIC NECROSIS VIRUS (IPNV)
AN ABSTRACT OF THE THESIS OF Hamdi Ogut for the degree of Master of Science in Fisheries Sicence presented on December 20. 1995. Title: IN VITRO HOST RANGE OF INFECTIOUS PANCREATIC NECROSIS VIRUS (IPNV)
11/15/2011. Outline. Structural Features and Characteristics. The Good the Bad and the Ugly. Viral Genomes. Structural Features and Characteristics
 Chapter 19 - Viruses Outline I. Viruses A. Structure of viruses B. Common Characteristics of Viruses C. Viral replication D. HIV II. Prions The Good the Bad and the Ugly Viruses fit into the bad category
Chapter 19 - Viruses Outline I. Viruses A. Structure of viruses B. Common Characteristics of Viruses C. Viral replication D. HIV II. Prions The Good the Bad and the Ugly Viruses fit into the bad category
Cardiomyopathy Syndrome (CMS) in Atlantic salmon WP4. Litterature review
 in Atlantic salmon WP4. Litterature review") Cardiomyopathy Syndrome (CMS) in Atlantic salmon WP4. Litterature review WP 4. Deliverables Report in Norwegian Fact sheet in Norwegian Scientific review PP- presentation 2 Overview of review CMS - Cardiomyopathy
Cardiomyopathy Syndrome (CMS) in Atlantic salmon WP4. Litterature review WP 4. Deliverables Report in Norwegian Fact sheet in Norwegian Scientific review PP- presentation 2 Overview of review CMS - Cardiomyopathy
Rama Nada. - Malik
 - 2 - Rama Nada - - Malik 1 P a g e We talked about HAV in the previous lecture, now we ll continue the remaining types.. Hepatitis E It s similar to virus that infect swine, so its most likely infect
- 2 - Rama Nada - - Malik 1 P a g e We talked about HAV in the previous lecture, now we ll continue the remaining types.. Hepatitis E It s similar to virus that infect swine, so its most likely infect
Chapter 13 Viruses, Viroids, and Prions. Biology 1009 Microbiology Johnson-Summer 2003
 Chapter 13 Viruses, Viroids, and Prions Biology 1009 Microbiology Johnson-Summer 2003 Viruses Virology-study of viruses Characteristics: acellular obligate intracellular parasites no ribosomes or means
Chapter 13 Viruses, Viroids, and Prions Biology 1009 Microbiology Johnson-Summer 2003 Viruses Virology-study of viruses Characteristics: acellular obligate intracellular parasites no ribosomes or means
Fish Health Assessment of Glass Eels from Canadian Maritime Rivers
 Fish Health Assessment of Glass Eels from Canadian Maritime Rivers D. Groman, R. Threader, D. Wadowska, T. Maynard and L. Blimke Aquatic Diagnostic Services, Atlantic Veterinary College Ontario Power Generation
Fish Health Assessment of Glass Eels from Canadian Maritime Rivers D. Groman, R. Threader, D. Wadowska, T. Maynard and L. Blimke Aquatic Diagnostic Services, Atlantic Veterinary College Ontario Power Generation
Infectious haematopoietic necrosis epidemic (2001 to 2003) in farmed Atlantic salmon Salmo salar in British Columbia
 in farmed Atlantic salmon Salmo salar in British Columbia") DISEASES OF AQUATIC ORGANISMS Vol. 72: 2 22, 2006 Published October 27 Dis Aquat Org Infectious haematopoietic necrosis epidemic (200 to 200) in farmed Atlantic salmon Salmo salar in British Columbia S.
DISEASES OF AQUATIC ORGANISMS Vol. 72: 2 22, 2006 Published October 27 Dis Aquat Org Infectious haematopoietic necrosis epidemic (200 to 200) in farmed Atlantic salmon Salmo salar in British Columbia S.
Medical Virology Immunology. Dr. Sameer Naji, MB, BCh, PhD (UK) Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University
 Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University") Medical Virology Immunology Dr. Sameer Naji, MB, BCh, PhD (UK) Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University Human blood cells Phases of immune responses Microbe Naïve
Medical Virology Immunology Dr. Sameer Naji, MB, BCh, PhD (UK) Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University Human blood cells Phases of immune responses Microbe Naïve
Redacted for Privacy
 AN ABSTRACT OF THE THESIS OF Hamdi OgUt for the degree of Doctor of Philosophy in Fisheries Science presented on January 08, 2001. Title: Biological and Mathematical Modeling of Dynamics of Furunculosis
AN ABSTRACT OF THE THESIS OF Hamdi OgUt for the degree of Doctor of Philosophy in Fisheries Science presented on January 08, 2001. Title: Biological and Mathematical Modeling of Dynamics of Furunculosis
Avian Reo-virus infections. Dr./ Wafaa Abd El-ghany Assistant Professor of poultry dis., Fac. Vet. Med., Cairo Univ.
 Avian Reo-virus infections Dr./ Wafaa Abd El-ghany Assistant Professor of poultry dis., Fac. Vet. Med., Cairo Univ. Economic losses Increased mortality. Diminished weight gains, poor feed conversions.
Avian Reo-virus infections Dr./ Wafaa Abd El-ghany Assistant Professor of poultry dis., Fac. Vet. Med., Cairo Univ. Economic losses Increased mortality. Diminished weight gains, poor feed conversions.
STUDY ON RAINBOW TROUT NODULAR GILL DISEASE DETECTED IN POLAND
 Bull Vet Inst Pulawy 51, 547-551, 2007 STUDY ON RAINBOW TROUT NODULAR GILL DISEASE DETECTED IN POLAND JERZY ANTYCHOWICZ Department of Fish Diseases, National Veterinary Research Institute, 24-100 Pulawy,
Bull Vet Inst Pulawy 51, 547-551, 2007 STUDY ON RAINBOW TROUT NODULAR GILL DISEASE DETECTED IN POLAND JERZY ANTYCHOWICZ Department of Fish Diseases, National Veterinary Research Institute, 24-100 Pulawy,
Isolation of Frog Virus 3 from Pallid Sturgeon (Scaphirhynchus albus) Suggests an Interclass Host Shift
 Suggests an Interclass Host Shift") Isolation of Frog Virus 3 from Pallid Sturgeon (Scaphirhynchus albus) Suggests an Interclass Host Shift Thomas B. Waltzek, Debra L. Miller, Bruce Drecktrah, Jeff T. Briggler, Beth MacConnell, Crystal Hudson,
Isolation of Frog Virus 3 from Pallid Sturgeon (Scaphirhynchus albus) Suggests an Interclass Host Shift Thomas B. Waltzek, Debra L. Miller, Bruce Drecktrah, Jeff T. Briggler, Beth MacConnell, Crystal Hudson,
Aseptic meningitis: inflammation of meninges with sterile CSF (without any causative organisms which can be grown on culture media).
.") You have to refer to the slides, since I have included the extra information only. Slide #1: Both illnesses aseptic meningitis and encephalitis can be caused by the same viruses; that viruses which cause
You have to refer to the slides, since I have included the extra information only. Slide #1: Both illnesses aseptic meningitis and encephalitis can be caused by the same viruses; that viruses which cause
Part I. Content: History of Viruses. General properties of viruses. Viral structure. Viral classifications. Virus-like agents.
 Viruses Part I Content: History of Viruses. General properties of viruses. Viral structure. Viral classifications. Virus-like agents. History Through the 1800s, many scientists discovered that something
Viruses Part I Content: History of Viruses. General properties of viruses. Viral structure. Viral classifications. Virus-like agents. History Through the 1800s, many scientists discovered that something
West Nile Virus. By Frank Riusech
 West Nile Virus By Frank Riusech Disease Etiology: West Nile virus(wnv), genus, flavivirus is positive- stranded RNA arbovirus (arthropod- borne), belonging to the Flaviviridae family. Included in this
West Nile Virus By Frank Riusech Disease Etiology: West Nile virus(wnv), genus, flavivirus is positive- stranded RNA arbovirus (arthropod- borne), belonging to the Flaviviridae family. Included in this
Chapter13 Characterizing and Classifying Viruses, Viroids, and Prions
 Chapter13 Characterizing and Classifying Viruses, Viroids, and Prions 11/20/2017 MDufilho 1 Characteristics of Viruses Viruses Minuscule, acellular, infectious agent having either DNA or RNA Cause infections
Chapter13 Characterizing and Classifying Viruses, Viroids, and Prions 11/20/2017 MDufilho 1 Characteristics of Viruses Viruses Minuscule, acellular, infectious agent having either DNA or RNA Cause infections
Riham. Bann. Bann. Belal
 30 Riham Bann Bann Belal Concepts to be covered in this lecture: 1. The Infectious Cycle 2. Virus cultivation 3. Viruses cytopathic effects 4. Viruses Quantification, Plaque assay, End-point dilution assay
30 Riham Bann Bann Belal Concepts to be covered in this lecture: 1. The Infectious Cycle 2. Virus cultivation 3. Viruses cytopathic effects 4. Viruses Quantification, Plaque assay, End-point dilution assay
A potential vaccine to control bacterial coldwater disease (CWD)
") A potential vaccine to control bacterial coldwater disease (CWD) Ken Cain 1 and Jerry Zinn 2 Northwest Fish Culture Conference, Dec. 6-8 2011 1 Department of Fish and Wildlife and the Aquaculture Research
A potential vaccine to control bacterial coldwater disease (CWD) Ken Cain 1 and Jerry Zinn 2 Northwest Fish Culture Conference, Dec. 6-8 2011 1 Department of Fish and Wildlife and the Aquaculture Research
LEC 2, Medical biology, Theory, prepared by Dr. AYAT ALI
 General Characteristics, Structure and Taxonomy of Viruses Viruses A virus is non-cellular organisms made up of genetic material and protein that can invade living cells. They are considered both a living
General Characteristics, Structure and Taxonomy of Viruses Viruses A virus is non-cellular organisms made up of genetic material and protein that can invade living cells. They are considered both a living
LESSON 4.6 WORKBOOK. Designing an antiviral drug The challenge of HIV
 LESSON 4.6 WORKBOOK Designing an antiviral drug The challenge of HIV In the last two lessons we discussed the how the viral life cycle causes host cell damage. But is there anything we can do to prevent
LESSON 4.6 WORKBOOK Designing an antiviral drug The challenge of HIV In the last two lessons we discussed the how the viral life cycle causes host cell damage. But is there anything we can do to prevent
Lab 3: Pathogenesis of Virus Infections & Pattern 450 MIC PRACTICAL PART SECTION (30397) MIC AMAL ALGHAMDI 1
 MIC AMAL ALGHAMDI 1") Lab 3: Pathogenesis of Virus Infections & Pattern 450 MIC PRACTICAL PART SECTION (30397) 2018 450 MIC AMAL ALGHAMDI 1 Learning Outcomes The pathogenesis of viral infection The viral disease pattern Specific
Lab 3: Pathogenesis of Virus Infections & Pattern 450 MIC PRACTICAL PART SECTION (30397) 2018 450 MIC AMAL ALGHAMDI 1 Learning Outcomes The pathogenesis of viral infection The viral disease pattern Specific
Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics
 Hepadnaviruses Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Hepatitis viruses A group of unrelated pathogens termed hepatitis viruses cause the vast majority
Hepadnaviruses Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Hepatitis viruses A group of unrelated pathogens termed hepatitis viruses cause the vast majority
Lab 2: Diagnostic Tests in Clinical Virology Laboratories 450 MIC PRACTICAL PART SECTION (30397) 2018 (450 MIC) AMAL ALGHAMDI - HUDA ALKHTEEB 1
 2018 (450 MIC) AMAL ALGHAMDI - HUDA ALKHTEEB 1") Lab 2: Diagnostic Tests in Clinical Virology Laboratories 450 MIC PRACTICAL PART SECTION (30397) 2018 (450 MIC) AMAL ALGHAMDI - HUDA ALKHTEEB 1 Diagnostic Virology Virus Isolation and Cultivation Viral
Lab 2: Diagnostic Tests in Clinical Virology Laboratories 450 MIC PRACTICAL PART SECTION (30397) 2018 (450 MIC) AMAL ALGHAMDI - HUDA ALKHTEEB 1 Diagnostic Virology Virus Isolation and Cultivation Viral
Lab Tuesday: Virus Diseases
 Lab Tuesday: Virus Diseases Quiz for Bacterial Pathogens lab (pp 67-73) and Biocontrol of Crown Gall (p. 113-117), Observation of Viral Movement in Plants (p. 119), and Intro section for Viruses (pp. 75-77).
Lab Tuesday: Virus Diseases Quiz for Bacterial Pathogens lab (pp 67-73) and Biocontrol of Crown Gall (p. 113-117), Observation of Viral Movement in Plants (p. 119), and Intro section for Viruses (pp. 75-77).
And Current Situation
 African Swine Fever Research And Current Situation Luis L. Rodriguez Research Leader Foreign Animal Disease Research Unit Agricultural Research Service, Plum Island Animal Disease Center African Swine
African Swine Fever Research And Current Situation Luis L. Rodriguez Research Leader Foreign Animal Disease Research Unit Agricultural Research Service, Plum Island Animal Disease Center African Swine
Biotechnology and DNA vaccines for aquatic animals
 Rev. sci. tech. Off. int. Epiz., 2008, 27 (1), 175-196 Biotechnology and DNA vaccines for aquatic animals G. Kurath Western Fisheries Research Center, Biological Resources Discipline, US Geological Survey,
Rev. sci. tech. Off. int. Epiz., 2008, 27 (1), 175-196 Biotechnology and DNA vaccines for aquatic animals G. Kurath Western Fisheries Research Center, Biological Resources Discipline, US Geological Survey,
numbe r Done by Corrected by Doctor
 numbe r 5 Done by Mustafa Khader Corrected by Mahdi Sharawi Doctor Ashraf Khasawneh Viral Replication Mechanisms: (Protein Synthesis) 1. Monocistronic Method: All human cells practice the monocistronic
numbe r 5 Done by Mustafa Khader Corrected by Mahdi Sharawi Doctor Ashraf Khasawneh Viral Replication Mechanisms: (Protein Synthesis) 1. Monocistronic Method: All human cells practice the monocistronic
SURVEILLANCE TECHNICAL
 CHAPTER 5 SURVEILLANCE TECHNICAL ASPECTS 55 Protect - detect - protect Polio eradication strategies can be summed up as protect and detect protect children against polio by vaccinating them, and detect
CHAPTER 5 SURVEILLANCE TECHNICAL ASPECTS 55 Protect - detect - protect Polio eradication strategies can be summed up as protect and detect protect children against polio by vaccinating them, and detect
Instructions for use
 Title Control strategy for viral diseases and shrimp at hatchery and seed pro Author(s) Yoshimizu, Mamoru Citation 魚病研究 = Fish pathology, 44(1): 9-13 Issue Date 2009-03-15 DOI Doc URLhttp://hdl.handle.net/2115/53211
Title Control strategy for viral diseases and shrimp at hatchery and seed pro Author(s) Yoshimizu, Mamoru Citation 魚病研究 = Fish pathology, 44(1): 9-13 Issue Date 2009-03-15 DOI Doc URLhttp://hdl.handle.net/2115/53211
number Done by Corrected by Doctor Ashraf
 number 4 Done by Nedaa Bani Ata Corrected by Rama Nada Doctor Ashraf Genome replication and gene expression Remember the steps of viral replication from the last lecture: Attachment, Adsorption, Penetration,
number 4 Done by Nedaa Bani Ata Corrected by Rama Nada Doctor Ashraf Genome replication and gene expression Remember the steps of viral replication from the last lecture: Attachment, Adsorption, Penetration,
Infectious Salmon Anemia - ISA. Dagfinn Ulriksen, M.Sc Special Adviser Aquaculture Aon Grieg Norway
 Infectious Salmon Anemia - ISA Dagfinn Ulriksen, M.Sc Special Adviser Aquaculture Aon Grieg Norway 1 Initial comments Since the first confirmed outbreak of ISA in Norway in 1984, ISA has been associated
Infectious Salmon Anemia - ISA Dagfinn Ulriksen, M.Sc Special Adviser Aquaculture Aon Grieg Norway 1 Initial comments Since the first confirmed outbreak of ISA in Norway in 1984, ISA has been associated
Dr. Ahmed K. Ali. Outcomes of the virus infection for the host
 Lec. 9 Dr. Ahmed K. Ali Outcomes of the virus infection for the host In the previous few chapters we have looked at aspects of the virus replication cycle that culminate in the exit of infective progeny
Lec. 9 Dr. Ahmed K. Ali Outcomes of the virus infection for the host In the previous few chapters we have looked at aspects of the virus replication cycle that culminate in the exit of infective progeny
علم األحياء الدقيقة Microbiology Introduction to Virology & Immunology
 علم األحياء الدقيقة Microbiology Introduction to Virology & Immunology What is a virus? Viruses may be defined as acellular organisms whose genomes consist of nucleic acid (DNA or RNA), and which obligatory
علم األحياء الدقيقة Microbiology Introduction to Virology & Immunology What is a virus? Viruses may be defined as acellular organisms whose genomes consist of nucleic acid (DNA or RNA), and which obligatory
Campbell's Biology: Concepts and Connections, 7e (Reece et al.) Chapter 24 The Immune System Multiple-Choice Questions
 Chapter 24 The Immune System Multiple-Choice Questions") Campbell's Biology: Concepts and Connections, 7e (Reece et al.) Chapter 24 The Immune System 24.1 Multiple-Choice Questions 1) The body's innate defenses against infection include A) several nonspecific
Campbell's Biology: Concepts and Connections, 7e (Reece et al.) Chapter 24 The Immune System 24.1 Multiple-Choice Questions 1) The body's innate defenses against infection include A) several nonspecific
Immunity to Fish Rhabdoviruses
 Viruses 2012, 4, 140-166; doi:10.3390/v4010140 Review OPEN ACCESS viruses ISSN 1999-4915 www.mdpi.com/journal/viruses Immunity to Fish Rhabdoviruses Maureen K. Purcell 1, *, Kerry J. Laing 2 and James
Viruses 2012, 4, 140-166; doi:10.3390/v4010140 Review OPEN ACCESS viruses ISSN 1999-4915 www.mdpi.com/journal/viruses Immunity to Fish Rhabdoviruses Maureen K. Purcell 1, *, Kerry J. Laing 2 and James
Susceptibility of Oblong Rockfish Sebastes oblongus to Viral Hemorrhagic Septicemia Virus
 Aquaculture Sci. 58 4 465 471 2010 Susceptibility of Oblong Rockfish Sebastes oblongus to Viral Hemorrhagic Septicemia Virus Tadashi ISSHIKI 1,, Giannakou ELENI 1, Yutaka UETA 2,3 and Taizou NAGANO 2 Abstract:
Aquaculture Sci. 58 4 465 471 2010 Susceptibility of Oblong Rockfish Sebastes oblongus to Viral Hemorrhagic Septicemia Virus Tadashi ISSHIKI 1,, Giannakou ELENI 1, Yutaka UETA 2,3 and Taizou NAGANO 2 Abstract:
Virology Introduction. Definitions. Introduction. Structure of virus. Virus transmission. Classification of virus. DNA Virus. RNA Virus. Treatment.
 DEVH Virology Introduction Definitions. Introduction. Structure of virus. Virus transmission. Classification of virus. DNA Virus. RNA Virus. Treatment. Definitions Virology: The science which study the
DEVH Virology Introduction Definitions. Introduction. Structure of virus. Virus transmission. Classification of virus. DNA Virus. RNA Virus. Treatment. Definitions Virology: The science which study the
Characterization of structural proteins of hirame rhabdovirus, HRV
 Vol. 10: 167-172, 1991 DISEASES OF AQUATIC ORGANISMS Dis. aquat. Org. l Published May 8 Characterization of structural proteins of hirame rhabdovirus, HRV Toyohiko ~ishizawal, Mamoru Yoshimizul, James
Vol. 10: 167-172, 1991 DISEASES OF AQUATIC ORGANISMS Dis. aquat. Org. l Published May 8 Characterization of structural proteins of hirame rhabdovirus, HRV Toyohiko ~ishizawal, Mamoru Yoshimizul, James
Immunodeficiency. (2 of 2)
") Immunodeficiency (2 of 2) Acquired (secondary) immunodeficiencies More common Many causes such as therapy, cancer, sarcoidosis, malnutrition, infection & renal disease The most common of which is therapy-related
Immunodeficiency (2 of 2) Acquired (secondary) immunodeficiencies More common Many causes such as therapy, cancer, sarcoidosis, malnutrition, infection & renal disease The most common of which is therapy-related
The Zombies of the Scientific Community Viruses and Other Acellular Infectious Agents. Acellular Agents
 viruses protein and nucleic acid viroids RNA virusoids RNA prions proteins The Zombies of the Scientific Community Viruses and Other Acellular Infectious Agents Acellular Agents Viruses major cause of
viruses protein and nucleic acid viroids RNA virusoids RNA prions proteins The Zombies of the Scientific Community Viruses and Other Acellular Infectious Agents Acellular Agents Viruses major cause of
The Ebola Virus. By Emilio Saavedra
 The Ebola Virus By Emilio Saavedra Etiological Agents: Ebolavirus is the etiologic agent. [1] There are four main families of viruses that are agents of Ebola (hemorrhagic fever). [1] These four families
The Ebola Virus By Emilio Saavedra Etiological Agents: Ebolavirus is the etiologic agent. [1] There are four main families of viruses that are agents of Ebola (hemorrhagic fever). [1] These four families
Third line of Defense
 Chapter 15 Specific Immunity and Immunization Topics -3 rd of Defense - B cells - T cells - Specific Immunities Third line of Defense Specific immunity is a complex interaction of immune cells (leukocytes)
Chapter 15 Specific Immunity and Immunization Topics -3 rd of Defense - B cells - T cells - Specific Immunities Third line of Defense Specific immunity is a complex interaction of immune cells (leukocytes)
Lifestage Diets for Fish
 Lifestage Diets for Fish % CRUDE PROTEIN % CRUDE FAT DIETS FEED SIZES STARTER CRUMBLES PELLETS (MM) MASH #0 #1 #2 1.2 1.5 2.0 2.5 3.0 4.0 6.0 9.0 PIGMENT FEATURES & FEEDING RECOMMENDATIONS PRODUCT CATALOG
Lifestage Diets for Fish % CRUDE PROTEIN % CRUDE FAT DIETS FEED SIZES STARTER CRUMBLES PELLETS (MM) MASH #0 #1 #2 1.2 1.5 2.0 2.5 3.0 4.0 6.0 9.0 PIGMENT FEATURES & FEEDING RECOMMENDATIONS PRODUCT CATALOG
Lab Tuesday: Virus Diseases
 Lab Tuesday: Virus Diseases Quiz for Bacterial Pathogens lab (pp 69-75) and Biocontrol of Crown Gall (p. 115-119), Observation of Viral Movement in Plants (p. 121), and Intro section for Viruses (pp. 77-79).
Lab Tuesday: Virus Diseases Quiz for Bacterial Pathogens lab (pp 69-75) and Biocontrol of Crown Gall (p. 115-119), Observation of Viral Movement in Plants (p. 121), and Intro section for Viruses (pp. 77-79).
Oregon Hatchery Research Center. Research Plan 2 December 2015 NWFCC
 Oregon Hatchery Research Center Research Plan 2 December 2015 NWFCC OHRC David Noakes Professor Fisheries & Wildlife Department, OSU Director, OHRC david.noakes@oregonstate.edu OHRC GOAL 1: Understand
Oregon Hatchery Research Center Research Plan 2 December 2015 NWFCC OHRC David Noakes Professor Fisheries & Wildlife Department, OSU Director, OHRC david.noakes@oregonstate.edu OHRC GOAL 1: Understand
Nucleic acid: singled stranded, double stranded, RNA, or DNA, linear or circular. Capsid: protein coat that is most of the mass of the virus.
 Viruses General Characteristics of Viruses 1. Depending on view may be regarded as exceptionally complex aggregates of nonliving chemicals or as exceptionally simple living microbes. 2. Contain a single
Viruses General Characteristics of Viruses 1. Depending on view may be regarded as exceptionally complex aggregates of nonliving chemicals or as exceptionally simple living microbes. 2. Contain a single
The Reproductive Patterns of Salmonids: Factors Affecting Male Reproductive Success
 The Reproductive Patterns of Salmonids: Factors Affecting Male Reproductive Success Factors affecting male reproductive success Fertilization success Female choice Male competition Attributes of the female
The Reproductive Patterns of Salmonids: Factors Affecting Male Reproductive Success Factors affecting male reproductive success Fertilization success Female choice Male competition Attributes of the female
Germ Cell Transplantation in Fish
 Larvi 2009 Germ Cell Transplantation in Fish Goro Yoshizaki (Tokyo University of Marine Science and Technology, SORST/JST) Tuna Mackerel Body weight; 300 kg 300 g Body length; 3 m 30 cm Scombridae family
Larvi 2009 Germ Cell Transplantation in Fish Goro Yoshizaki (Tokyo University of Marine Science and Technology, SORST/JST) Tuna Mackerel Body weight; 300 kg 300 g Body length; 3 m 30 cm Scombridae family
Virulence of viral haemorrhagic septicaemia virus (VHSV) genotype III in rainbow trout
 genotype III in rainbow trout") DOI 10.1186/s13567-015-0303-z Veterinary Research RESEARCH ARTICLE Open Access Virulence of viral haemorrhagic septicaemia virus (VHSV) genotype III in rainbow trout Takafumi Ito 1*, Jun Kurita 1, Koh
DOI 10.1186/s13567-015-0303-z Veterinary Research RESEARCH ARTICLE Open Access Virulence of viral haemorrhagic septicaemia virus (VHSV) genotype III in rainbow trout Takafumi Ito 1*, Jun Kurita 1, Koh