CODING EFFICIENCY OF INNER HAIR CELLS AT THE THRESHOLD OF HEARING
|
|
|
- Colleen Todd
- 6 years ago
- Views:
Transcription
1 Computational Models of Auditory Function 5 S. Greenberg and M. Slaney (eds.) IOS Press, 21 CODING EFFICIENCY OF INNER HAIR CELLS AT THE THRESHOLD OF HEARING Ilse C. Gebeshuber 1 and Frank Rattay 2 1 Institut für Allgemeine Physik and 2 TU-BioMed University of Technology, Vienna Wiedner Hauptstraße 8-1/1145 A-14 Wien, Austria 1. Introduction The human hearing threshold curve for pure tones is a nonlinear function of frequency (Figure 1). The minimum sound pressure required for an audible sensation to occur is frequency dependent and spans approximately five orders of magnitude. In the highly sensitive region of the spectrum between 2 and 5 khz an intensity as low as 1-12 W/m 2 is sufficient to evoke an audible sensation. The current modeling study investigates the efficiency with which the mechanical stage of transduction in the inner ear is transformed into a neural form (i.e., mechano electric transduction) for low-intensity sinusoidal signals across a range of frequencies. Our goal is to gain insight into the basic physiological properties that underlie the human hearing threshold curve. The inner hair cells (Figure 2) play a key role in mechano-electric transduction. In the human inner ear are three rows of outer hair cells (OHCs) and one row of inner hair cells (IHCs). A primary function of the OHCs is to amplify the magnitude of low-intensity signals [1][16][24]. IHCs serve as the primary (if not the sole) conduit of frequency-selective information to the brain via their innervation of the auditory nerve. Deflection of the hair cell stereocilia modulates the probability of the cell s transduction channels opening and closing and is responsible for the voltage fluctuations observed within the cell. Such fluctuations provide a low-pass filtered image of the stereociliary displacement (along with additional stochastic components resulting from channel gating). In the active zones, at the base of the cell, sufficient depolarization of the receptor potential results in Ca 2+ -induced neurotransmitter release. This transmitter release, if of sufficient magnitude, results in depolarization of proximal auditory-nerve fibers, resulting in the generation of action potentials that are propagated into the auditory brainstem. The transduction mechanism is so sensitive that displacements resulting from stereociliary Brownian motion contribute significantly to the spontaneous discharge activity observed in highly sensitive (i.e., high spontaneous rate) auditory-nerve fibers [8][1][31]. Acoustically generated displacements less than the thermal motion of the stereocilia are sufficient to cause an audible sensation. The Brownian motion in this instance enhances the detection of weak signals via a mechanism known as stochastic resonance [9][1][31]. Stochastic resonance is based on nonlinear statistical dynamics through which information flow in a multistate system (such as the transduction channel of the inner hair cell or the all-or-none process of spike generation) is enhanced by the presence of optimized random noise [23].

2 6 I. C. Gebeshuber and F. Rattay / Coding Efficiency of Inner Hair Cells at Threshold Figure 1 Region of human audibility (i.e., the range between the threshold of hearing and of pain). The intensity is scaled in db, while the sound pressure is shown in Pascals. Note that the range of pressure variation covers 7 orders of magnitude. Adapted from [33]. Figure 3 shows the number of auditory-nerve fibers innervating an average IHC of a normal human cochlea. The innervation density varies as a function of cochlear-frequency position in a manner comparable to the hearing threshold curve (Figure 1). In the mid-frequency region the innervation density reaches a maximum of ca. 15 fibers per IHC. About sixty percent of these fibers are highly sensitive and exhibit relatively high rates of spontaneous activity (18 12 spikes/s). The highly sensitive nerve fibers change their spiking patterns for low- and mid-frequency signals close to the threshold of hearing as follows: the first sign of influence on the firing of many spontaneously active fibers by a pure tone is phase-locking of the spikes [28] [29]. This may occur at an intensity far below that required to evoke an increase in mean firing rate [12]. This phase-locking effect does not occur for high-frequency signals as a consequence of the jitter associated with interspike times. Afferent fibers may respond differently each time a stimulus of a given amplitude is presented since fluctuations in excitability and latency are directly associated with fluctuations in the membrane resting potential [3]. Endogenous noise in the resting neural membrane potential of nerve fibers decreases with increasing diameter. The noise is on the order of 1 mv root-mean-square (r.m.s.) for myelinated fibers of small diameter and less than 1 mv for larger-diameter myelinated fibers [4]. The mean inner diameter of central axons of human auditory-nerve fibers has an unimodal distribution and ranges between 2.7 and 3.1 µm (with the exception of smaller fibers at the base of the cochlea) [3].
3 I. C. Gebeshuber and F. Rattay / Coding Efficiency of Inner Hair Cells at Threshold 7 Figure 2 Schematic illustration of an inner hair cell. Endolymphatic fluid motion caused by the movement of the middle ear ossicles induces displacement of the stereocilia of the auditory receptor cell. The stereocilia of an inner hair cell are interconnected by links (elastic protein filaments). The open-close kinetics of transduction channels located close to the top of each stereocilium depend on stereociliary deflection (Figure 4). Even in the resting state the transduction channel open probability is about 15%. Due to potential gradients, ion currents (mainly potassium) enter the cell through the transduction channels and leave through ion channels in the cell body membrane, resulting in a resting potential of ca. 4 mv in the unstimulated hair cell and potential changes of several mv following stereociliary displacement. A potential change as low as.1 mv may cause neurotransmitter release and thereby evoke a spike in an auditory-nerve fiber. Note the tapering of the bottom portion of the stereocilia endings. In humans the inner hair cell stereocilia are arranged in a 2 by 3 matrix, with 2 short, 2 intermediate-length and 2 long elements. Each stereocilium behaves like a rigid rod pivoting around its insertion point into the cuticular plate. 2. Materials and Methods 2.1 Brownian Motion The IHC stereocilia are interconnected by tip-links and horizontal links, and act like stiff rods capable of pivoting around their insertion point into the cuticular plate (Figure 2). The Brownian motion of the stereociliary tips is calculated using a reduced version of the stereocilia linear chain model [31]. The r.m.s. value of the modeled intrinsic bundle noise is ca. 2 nm, which is in accordance with experimental data [2]. The small amplitude of the fluctua-
4 8 I. C. Gebeshuber and F. Rattay / Coding Efficiency of Inner Hair Cells at Threshold # of afferents / IHC ,,1,2,3,4,5,6,7,8,9 1, Distance from base / total length Figure 3 Innervation density per inner hair cell in a normal human cochlea (adapted from [7]). Sixty percent of the afferent nerve fibers are highly sensitive. The transformation from normalized distance, d, to the characteristic frequency, f, of the nerve fiber obeys the following relation in the human: f=2(1 2d -.7) [13]. tions due to Brownian motion can be appreciated by comparing them to the dimensions of a single stereocilium (ca..2 µm in diameter) or to the bundle s displacement-response relationship (Figure 4). In vestibular hair cells of the frog, viscous drag acting on the bundle limits Brownian motion to relatively low frequencies (2 8 Hz) [2]. However, theoretical considerations suggest a corner frequency of ca. 4 khz for thermal fluctuations in mammalian hair cells [31], which implies that stochastic resonance may also be effective in the mid-frequency range of audition. The overall displacement of the hair bundle in response to low-intensity signals is the sum of the bundle movements resulting from Brownian motion as well as from the signal-induced displacement. The signal-to-noise ratio is defined as the ratio of the r.m.s. magnitude of the signal and the r.m.s. level of the stereociliary deflections attributable to Brownian motion. Our Hair bundle displacement (nm) Figure 4 The relation of a 3-ms trace of simulated Brownian motion (low-pass filtered, 2 nm r.m.s. white noise) to a cell s displacement-response behavior. This function relates the probability of transduction channels being open (left y-axis) to the hair bundle displacement (x-axis). Note that in this specific case the transduction-channel, resting-open probability is.2. Adapted from [18].
5 I. C. Gebeshuber and F. Rattay / Coding Efficiency of Inner Hair Cells at Threshold Time (ms) Figure 5 Modeled mechanical and electrical fluctuations due to Brownian motion: the intracellular receptor potential changes (bottom trace) are a low-pass filtered version of stereociliary displacements (top trace) with an additional amount of noise resulting from transduction channel kinetics. simulation investigates the effects of signal-to-noise ratio (whose normalized range is between and 1 equivalent to stimulation of the hair bundle with an amplitude between and 2.12 nm r.m.s.). The frequency of the stimulating, deterministic signals ranges between.2 and 2 khz. Figure 5 shows a 2-ms time series of hair-bundle displacements resulting from Brownian motion. 2.2 Endogenous Transduction Channel Noise The receptor potential fluctuations in the IHC are calculated using a model for the mechano electrical transduction in inner hair cells [25]. The model uses equivalent electric circuits for cell membrane and cytoplasm (i.e., RC components and batteries). The kinetics of the transduction channels are modeled as Markov processes without memory: whether the channel stays open or closed depends only on its current open probability and not on the length of time the channel has already been open or closed. For displacements in the range of a few nm, the relation between the stereociliary displacement and the open probability of the transduction channels is linear (Figure 4) [22]. For zero displacement the open probability of the transduction channel is about.15. For small displacements to the lateral side the transduction channel open probability increases, resulting in an influx of potassium ions. This influx causes a depolarization of the receptor potential from its resting state (ca. 4 mv). Displacement to the medial side decreases the open probability, resulting in fewer potassium ions entering the cell and a concomitant hyperpolarization of the membrane potential. Since the model s inner-hair-cell membrane time constant, τ, equals.255 ms [25], the IHC potential can be thought of as a low-pass-filtered version of the stereociliary displacement pattern combined with additional noise resulting from the stochastic components in channel gating (Figure 5).































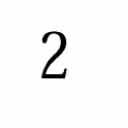
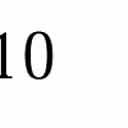
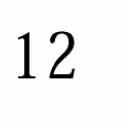
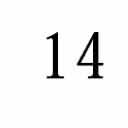
6 1 I. C. Gebeshuber and F. Rattay / Coding Efficiency of Inner Hair Cells at Threshold Figure 6 Simulated receptor potential changes and resulting firing behavior. The noise in the voltage fluctuations evoked by a weak 5-Hz signal alone (thin line, hypothetical case without Brownian motion) is a consequence of the endogenous transduction channel noise. Only in one instance (marked by a dashed arrow at 13.5 ms) are the fluctuations large enough to reach the threshold of spiking at.1 mv. The compound fluctuations caused by the same sinusoidal signal, the endogenous transduction channel noise and the thermal fluctuations with a signal-to-noise ratio of.2 show the enhancing effect of the noise: Seven spikes may occur within 2 ms among associated auditory-nerve fibers. The recovery behavior after spiking is modeled by an exponential decay of the threshold curve. As soon as the voltage fluctuations exceed threshold a new spike can occur. 2.3 Jitter in the Spiking Times: Refractory Period The spike-generation process is modeled in the following way. Whenever the voltage fluctuations of the IHC exceed a threshold of.1 mv (a value sufficient for neurotransmitter release in hair cells [15]), a spike may be generated in an afferent fiber. Because of the stochastic nature of spike generation the probability for spiking is adjusted to obtain a mean spontaneous discharge rate of ca. 1 spikes/s in the resting state [27]. Jitter in the firing pattern is modeled by a single-sided, normally distributed time shift whose standard deviation is 5 µs. Since the absolute refractory period of an auditory-nerve fiber is ca..8 ms (in the cat [19]), the time constant of the exponentially decaying threshold curve is set to.25 ms and the maximum value for the height is set to 2 mv (Figure 6). The spike rate associated with a just-supra-threshold signal does not exceed the spontaneous rate of 1 spikes/s. However, nerve impulses become increasingly phase-locked to the acoustic signal as the signal level increases [1][11][12][14][28][29].
7 I. C. Gebeshuber and F. Rattay / Coding Efficiency of Inner Hair Cells at Threshold khz snr= khz snr= khz snr= khz snr= Figure 7 ISIHs for a 1-kHz signal at several signal-to-noise ratios. With increasing SNR, the proportion of spikes in a specific half of the signal period increases relative to the other half. Signal duration is 1s. The model s output represents the activity of 12 highly sensitive nerve fibers. Histogram binwidth is.5 ms. 3. Results In this section we present an analysis of the frequency information encoded in the interspike interval histograms (ISIHs) of simulated auditory-nerve firing patterns induced by low-intensity, sinusoidal hair bundle deflections. When the histogram binwidth is precisely half the period of the stimulating signal, phase-locking of the interspike times can be readily observed in the ISIHs as an up-downup-down pattern. The distribution of spikes is non-uniform across time, being concentrated in a restricted portion of the stimulus cycle. This phase-locked behavior is manifested in the interspike interval histogram in the form of modes associated with intervals that are integral multiples of the stimulus period. The maximum interspike time considered in our model is 2 ms. Figure 7 shows ISIHs for stereociliary displacements at 1 khz. With increasing SNR, the spikes tend to occur increasingly in the first half of the stimulus period (i.e., the phaselocking effect becomes increasingly apparent). A means to assess the information contained in the ISIH is to measure the ratio of spikes occurring during the positive half-wave of the stimulating signal relative to the total number of spikes. This ratio is a measure of the proportion of informative spikes, and has been used as a metric of phase-locking performance [28]. With decreasing signal frequency the number of bins (and therefore the fine structure information) associated with the ISIH decreases (Figure 8). Although the number of infor-
8 12 I. C. Gebeshuber and F. Rattay / Coding Efficiency of Inner Hair Cells at Threshold Hz snr= Hz snr= Hz snr= Hz snr= mative spikes is still greater than 5% for a 2-Hz signal with an SNR of.2, the number of events in each bin tend to decrease exponentially as occurs when the SNR is. The fine structure in the histogram, with valleys associated with integral multiples of the negative half-wave of the stimulating signal, is lost. In a previous study we had analyzed the information contained in the ISIHs with artificial neural networks [9] [26]. In the mid-frequency range the neural net accurately detected the signal more than 75% of the time for signal-to-noise ratios as low as.1. Taking into consideration the parallel information transfer from several IHCs to the central nervous system considerably reduces the signal duration required to accurately detect the presence of a signal. The information contained in the ISIHs is evaluated by calculating the number of informative spikes over a range of frequencies and signal-to-noise ratios. The resulting frequencyresponse-efficiency tuning curves are illustrated in Figure 9. The curves for low SNRs may be thought of as analogous to human hearing threshold curves for pure tones, as they reflect the combined effects of stereociliary Brownian motion, endogenous hair cell noise, the stochastic nature of neurotransmitter release and the innervation density of primary auditory afferents in various frequency bands. Note that this simulation study models threshold curves as they are reflected in the transduction of small sinusoidal displacements of the stereocilia. Furthermore, any possible effect of inhibitory efferent innervation of afferent nerve fibers (see e.g. [5] [6]) on the spiking pattern is neglected due to an absence of experimental data. Figure 8 ISIHs for a 2-Hz signal at several signal-to-noise ratios. Signal presentation time is 1s. 5 highly sensitive nerve fibers, binwidth is 2.5 ms.
9 I. C. Gebeshuber and F. Rattay / Coding Efficiency of Inner Hair Cells at Threshold 13 Frequency (khz) # of informative spikes (normalised), 2, , 4,4 5, 5,5 5, 6,6 5, 7,7 5, 8,8 5 snr=1 snr=.9 snr=.8 snr=.7 snr=.6 snr=.5 snr=.4 snr=.3 snr=.2 snr=.1 snr= Figure 9 Frequency-response efficiency tuning curves for a multicellular model of peripheral auditory coding, (i.e. normalized number of informative spikes for several frequencies and signal-to-noise ratios). For an SNR of there is no signal present and half of the spikes are phase-locked at chance level (i.e. the normalized number of informative spikes is.5). For frequencies between.2 khz and 2 khz, the phase-locking effect increases with increasing SNR (i.e., the normalized number of informative spikes increases well above.5). In the 5 khz case there is virtually no apparent phase-locking. The 1 khz and 2 khz cases show no effect of phase-locking at all. In such instances only the normalized number of informative spikes for an SNR of 1 is presented. For high signal-to-noise ratios the curves are V-shaped. When the signal is reduced in amplitude and the influence of noise increases, the curves broaden and eventually invert at 1 khz. Stimulus duration for each data point is 1s. For information concerning frequency-dependent innervation density cf. Figure 3. At 1 khz (the frequency which can be encoded and decoded optimally under the present conditions) a reversal of the shape of the curve appears at very low signal-to-noise ratios. When the signal level increases, the curves invert and become sharper. This effect corresponds to experimental results observed in noise-induced tuning-curve changes in mechanoreceptors of the rat foot [17]. Modeling the transduction channel kinetics as a Markov process results in a frequency-dependent peak-to-peak receptor potential. For low and high frequencies, the sub-threshold deterministic stimuli elicit voltage changes further from threshold than ones evoked by mid-frequency stimuli. Therefore, the optimal noise level is also frequency-dependent and the inversion of the tuning curve for low SNR stimuli is directly related to the threshold shift (cf. Figure 9 in [17]). The 2-kHz case is comparable to the 1-kHz case. Increasing the SNR increases the number of informative spikes from approximately (but just higher than) chance level for an SNR of.1 to over 7% for an SNR of 1. For high-frequency signals the jitter in the nerve firing pattern destroys the fine structure in the ISIH. However, statistics of the discharge pattern over a longer period would still contain some temporal information germane to 5 khz, at least for signal-to-noise ratios close to one. For signals in the range of 1 2 khz, increasing the signal does not further increase the number of informative spikes since the jitter completely destroys the phase-locking information. Therefore, the psychophysical hearing threshold data for the high-frequency portion of the spectrum cannot be attributed to phase-locking. This means that for high-frequency signals frequency information must be coded in a different way. The increase in spike rate is the most likely candidate for providing this information.
10 14 I. C. Gebeshuber and F. Rattay / Coding Efficiency of Inner Hair Cells at Threshold 4. Discussion In this study we have shown that a compound model of coding efficiency of inner hair cells at the threshold of hearing accounts for certain properties of the psychophysically measured human hearing threshold curve (Figure 1). Through the mechanism of stochastic resonance the Brownian motion of IHC stereocilia makes otherwise undetectable low-intensity signals audible. The jitter in auditory-nerve fiber spike times accounts for the steep slope in the threshold curve at high frequencies. In the low-frequency portion of the spectrum the long interspike times prevent detection of the signal, especially at low signal-to-noise ratios. Changes in hair-bundle morphology also affect the pattern of thermal fluctuations of the stereocilia and therefore exert some influence on spontaneous activity in auditory-nerve fibers. In milder instances of acoustic trauma, morphological changes are only found in the rootlets of the stereocilia (which appear less dense in electron micrographs) [21]. In more severe instances of trauma (typically resulting in permanent damage) kinks or fractures at the rootlet of the stereocilia, and the packed actin filaments (which impart the stereocilia with their rigidity) are depolymerized [2] [32]. Within the IHC tuft the damage to the tall, outer row of stereocilia is often selective; the shorter rows may remain ultrastructurally normal even when the tallest row is completely missing. Moreover, the tip links remain intact on the shorter stereocilia, suggesting that such IHCs may be capable of transduction, but with reduced sensitivity. Auditory-nerve fibers associated with such IHCs exhibit much lower rates of spontaneous activity [21]. Following acoustic overstimulation, tuning curves with elevated tips and tails are associated with significant decreases in mean spontaneous discharge rate, whereas tuning curves with elevated tips but hypersensitive tails are associated with a clear elevation of the mean spontaneous rates [21]. Our model, in which altered Brownian motion patterns of the stereocilia lead to changes in the spiking pattern, may help to account for the occurrence of such pathological spiking patterns. However, one should bear in mind that in hearing loss of cochlear origin there are other noise-induced changes, such as different steady-state Ca 2+ concentrations, that are the result of altered Ca 2+ pump kinetics. Such changes may also be responsible for the pathological spiking patterns. Future studies of the coding efficiency of inner hair cells at the threshold of hearing should take into consideration the possibility that Brownian motion of the stereocilia changes along the tonotopic axis and may be tuned in such a way as to enhance the audibility of specific frequencies. Such studies should also carefully consider the potential significance of the adaptation process of mammalian-transduction-channel kinetics, as well as the stochastic-resonance phenomena that have recently been demonstrated in transduction channels [18] and in calcium-activated potassium channels in the basolateral IHC membrane (Jaramillo, personal communication). Acknowledgements We thank Tony Wenzelhuemer for critically reading the manuscript and Alice Mladenka, Richardson Naves Leao, Juliana Pontes Pinto and Wolfgang Andreas Svrcek-Seiler for their participation in the preliminary studies. We also thank Jonathan Ashmore, Fernán Jaramillo, Steve Greenberg and Ted Evans for discussions. References [1] Ashmore, J. F. and Kolston, P. J. Hair cell based amplification in the cochlea. Curr. Opin. Neurobiol., 4: 53 8, 1994.
11 I. C. Gebeshuber and F. Rattay / Coding Efficiency of Inner Hair Cells at Threshold 15 [2] Denk, W., Webb, W. W. and Hudspeth, A. J. Mechanical properties of sensory hair bundles are reflected in their Brownian motion measured with a laser differential interferometer. Proc. Nat. Acad. Sci., 16: , [3] Derksen, H. E. Axon membrane voltage fluctuations. Act. Physiol. Pharmacol. Neerl., 13: , [4] Derksen, H. E. and Verveen, A. A. Fluctuations of resting neural membrane potential. Science, 151: , [5] Ehrenberger, K. and Felix, D. Glutamate receptors in afferent cochlear neurotransmission in guinea pigs. Hear. Res., 52: 73 8, [6] Felix, D. and Ehrenberger, K. The efferent modulation of mammalian inner hair cell afferents. Hear. Res., 64: 1 5, [7] Felix, H., Gleeson, M. J., Pollak, A. and Johnsson, L. The cochlear neurons in humans. Progr. Hum. Audit. Vestib. Histopath., S. Iurano and J. E. Veldman (eds.), Amsterdam: Kugler, pp , [8] Gebeshuber, I. C., Mladenka, A., Rattay, F. and Svrcek-Seiler, W. A. Computational demonstration that Brownian motion of inner hair cells stereocilia may enhance the ability to detect low level auditory tones from auditory nerve spiking patterns. J. Physiol. (Lond.), 54P: 127P 128P, [9] Gebeshuber, I. C., Mladenka, A., Rattay, F. and Svrcek-Seiler, W. A. Brownian motion and the ability to detect weak auditory signals. In Chaos and Noise in Biology and Medicine, M. Barbi and S. Chillemi (Eds.), World Scientific: Singapore, pp , [1] Gebeshuber, I. C. The influence of stochastic behavior on the human threshold of hearing. Chaos, Solitons & Fractals, 11: , 2. [11] Gebeshuber, I. C., Pontes Pinto, J., Naves Leao, R., Mladenka, A. and Rattay, F. Stochastic resonance in the inner ear: the effects of endogenous transduction channel noise and stereociliary thermal motions on the human hearing threshold in various frequency bands. ARGESIM Rep. 1: Proc. TU-BioMed Minisymp Brain Modelling F. Rattay (Ed.), pp. 4 44, [12] Gleich, O., Narins, P. M. The phase response of primary auditory afferents in a songbird (Sturnus vulgaris L.). Hear. Res., 32: 81 91,1988. [13] Greenwood, D. D. A cochlear frequency position function for several species 29 years later. J. Acoust. Soc. Am., 87: , 199. [14] Hind, J. E. Physiological correlates of auditory stimulus periodicity. Audiology, 11: 42 57, [15] Hudspeth, A. J. How the ear s works work. Nature, 341: , [16] Hudspeth, A. J. Mechanical amplification of stimuli by hair cells. Curr. Opin. Neurobiol., 7: , [17] Ivey, C., Apkarian, A. V. and Chialvo, D. R. Noise-induced tuning curve changes in mechanoreceptors. J. Neurophysiol. 79: , [18] Jaramillo, F. and Wiesenfeld, K. Mechanoelectrical transduction assisted by Brownian motion: A role for noise in the auditory system. Nature Neurosci., 1: , [19] Javel, E. Acoustic and electrical encoding of temporal information. In Cochlear Implants Models of the Electrically Stimulated Ear, J. M. Miller and F. A. Spelman (eds.), New York: Springer Verlag, pp. 247, 199. [2] Liberman, M. C. Auditory-nerve response from cats raised in a low-noise chamber. J. Acoust. Soc. Am., 63: , [21] Liberman, M. C. and Dodds, L. W. Single-neuron labeling and chronic cochlear pathology. II. Stereocilia damage and alterations of spontaneous discharge rates. Hear. Res., 16: 43 53, [22] Markin, V. S., Jaramillo, F. and Hudspeth, A. J. The three-state model for transduction-channel gating in hair cells. Biophys. J., 64: A93, [23] McNamara, B. and Wiesenfeld, K. The theory of stochastic resonance. Phys. Rev. A, 39: , [24] Nobili, R., Mammano, F. and Ashmore, J. F. How well do we understand the cochlea? Trends Neurosci., 21: , [25] Rattay, F., Gebeshuber, I. C. and Gitter, A. H. The mammalian auditory hair cell: A simple electric circuit model. J. Acoust. Soc. Am., 13: , [26] Rattay, F., Mladenka, A. and Pontes Pinto, J. Classifying auditory nerve patterns with neural nets: A modeling study with low level signals. Sim. Pract. Theor., 6: , 1998.
12 16 I. C. Gebeshuber and F. Rattay / Coding Efficiency of Inner Hair Cells at Threshold [27] Relkin, E. M. and Doucet, J. R. Recovery from prior stimulation. I: Relationship to spontaneous firing rates of primary auditory neurons. Hear. Res., 55: , [28] Rose, J. E., Brugge, J. F., Anderson, D. J., Hind, J. E., Phase-locked response to low-frequency tones in single auditory nerve fibers of the squirrel monkey. J. Neurophysiol., 3: , [29] Rose, J. E., Hind, J. E., Anderson, D. J., Brugge, J. F., Some effects of stimulus intensity on response of auditory nerve fibers in the squirrel monkey. J. Neurophysiol., 34: , [3] Spoendlin, H. and Schrott, A. Analysis of the human auditory nerve. Hear. Res., 43: 25 38, [31] Svrcek-Seiler, W. A., Gebeshuber, I. C., Rattay, F., Biró, T. and Markum, H. Micromechanical models for the Brownian motion of hair cell stereocilia. J. Theor. Biol., 193: , [32] Tilney, L. G., Saunders, J. C., Egelman, E. and DeRosier, D. J. Changes in the organization of actin filaments in the stereocilia of noise-damaged lizard cochleae. Hear. Res., 7: , [33] Zwicker, E. Psychoakustik. Berlin: Springer Verlag, p. 34, 1982.
The in uence of stochastic behavior on the human threshold of hearing
 Chaos, Solitons and Fractals 11 (2000) 1855±1868 www.elsevier.nl/locate/chaos The in uence of stochastic behavior on the human threshold of hearing Ilse Christine Gebeshuber a,b, * a TU-BioMed, University
Chaos, Solitons and Fractals 11 (2000) 1855±1868 www.elsevier.nl/locate/chaos The in uence of stochastic behavior on the human threshold of hearing Ilse Christine Gebeshuber a,b, * a TU-BioMed, University
The mammalian cochlea possesses two classes of afferent neurons and two classes of efferent neurons.
 1 2 The mammalian cochlea possesses two classes of afferent neurons and two classes of efferent neurons. Type I afferents contact single inner hair cells to provide acoustic analysis as we know it. Type
1 2 The mammalian cochlea possesses two classes of afferent neurons and two classes of efferent neurons. Type I afferents contact single inner hair cells to provide acoustic analysis as we know it. Type
What is the effect on the hair cell if the stereocilia are bent away from the kinocilium?
 CASE 44 A 53-year-old man presents to his primary care physician with complaints of feeling like the room is spinning, dizziness, decreased hearing, ringing in the ears, and fullness in both ears. He states
CASE 44 A 53-year-old man presents to his primary care physician with complaints of feeling like the room is spinning, dizziness, decreased hearing, ringing in the ears, and fullness in both ears. He states
Mechanical Properties of the Cochlea. Reading: Yost Ch. 7
 Mechanical Properties of the Cochlea CF Reading: Yost Ch. 7 The Cochlea Inner ear contains auditory and vestibular sensory organs. Cochlea is a coiled tri-partite tube about 35 mm long. Basilar membrane,
Mechanical Properties of the Cochlea CF Reading: Yost Ch. 7 The Cochlea Inner ear contains auditory and vestibular sensory organs. Cochlea is a coiled tri-partite tube about 35 mm long. Basilar membrane,
Cochlear anatomy, function and pathology II. Professor Dave Furness Keele University
 Cochlear anatomy, function and pathology II Professor Dave Furness Keele University d.n.furness@keele.ac.uk Aims and objectives of this lecture Focus (2) on the biophysics of the cochlea, the dual roles
Cochlear anatomy, function and pathology II Professor Dave Furness Keele University d.n.furness@keele.ac.uk Aims and objectives of this lecture Focus (2) on the biophysics of the cochlea, the dual roles
The Structure and Function of the Auditory Nerve
 The Structure and Function of the Auditory Nerve Brad May Structure and Function of the Auditory and Vestibular Systems (BME 580.626) September 21, 2010 1 Objectives Anatomy Basic response patterns Frequency
The Structure and Function of the Auditory Nerve Brad May Structure and Function of the Auditory and Vestibular Systems (BME 580.626) September 21, 2010 1 Objectives Anatomy Basic response patterns Frequency
Auditory System Feedback
 Feedback Auditory System Feedback Using all or a portion of the information from the output of a system to regulate or control the processes or inputs in order to modify the output. Central control of
Feedback Auditory System Feedback Using all or a portion of the information from the output of a system to regulate or control the processes or inputs in order to modify the output. Central control of
The transformation of sound stimuli into electrical signals
 The transformation of sound stimuli into electrical signals Robert Fettiplace 2 1 Introduction Our sense of hearing depends on the correct performance of about 15 000 hair cells in each cochlea that serve
The transformation of sound stimuli into electrical signals Robert Fettiplace 2 1 Introduction Our sense of hearing depends on the correct performance of about 15 000 hair cells in each cochlea that serve
Acoustics, signals & systems for audiology. Psychoacoustics of hearing impairment
 Acoustics, signals & systems for audiology Psychoacoustics of hearing impairment Three main types of hearing impairment Conductive Sound is not properly transmitted from the outer to the inner ear Sensorineural
Acoustics, signals & systems for audiology Psychoacoustics of hearing impairment Three main types of hearing impairment Conductive Sound is not properly transmitted from the outer to the inner ear Sensorineural
Chapter 3: Anatomy and physiology of the sensory auditory mechanism
 Chapter 3: Anatomy and physiology of the sensory auditory mechanism Objectives (1) Anatomy of the inner ear Functions of the cochlear and vestibular systems Three compartments within the cochlea and membranes
Chapter 3: Anatomy and physiology of the sensory auditory mechanism Objectives (1) Anatomy of the inner ear Functions of the cochlear and vestibular systems Three compartments within the cochlea and membranes
Systems Neuroscience Oct. 16, Auditory system. http:
 Systems Neuroscience Oct. 16, 2018 Auditory system http: www.ini.unizh.ch/~kiper/system_neurosci.html The physics of sound Measuring sound intensity We are sensitive to an enormous range of intensities,
Systems Neuroscience Oct. 16, 2018 Auditory system http: www.ini.unizh.ch/~kiper/system_neurosci.html The physics of sound Measuring sound intensity We are sensitive to an enormous range of intensities,
What does it mean to analyze the frequency components of a sound? A spectrogram such as that shown here is the usual display of frequency components
 1 2 3 4 What does it mean to analyze the frequency components of a sound? A spectrogram such as that shown here is the usual display of frequency components as a function of time here during the production
1 2 3 4 What does it mean to analyze the frequency components of a sound? A spectrogram such as that shown here is the usual display of frequency components as a function of time here during the production
Salamanca Study Abroad Program: Neurobiology of Hearing
 Salamanca Study Abroad Program: Neurobiology of Hearing Synaptics and the auditory nerve R. Keith Duncan University of Michigan rkduncan@umich.edu Review Resources Reviews: Safieddine et al., 2012, The
Salamanca Study Abroad Program: Neurobiology of Hearing Synaptics and the auditory nerve R. Keith Duncan University of Michigan rkduncan@umich.edu Review Resources Reviews: Safieddine et al., 2012, The
Representation of sound in the auditory nerve
 Representation of sound in the auditory nerve Eric D. Young Department of Biomedical Engineering Johns Hopkins University Young, ED. Neural representation of spectral and temporal information in speech.
Representation of sound in the auditory nerve Eric D. Young Department of Biomedical Engineering Johns Hopkins University Young, ED. Neural representation of spectral and temporal information in speech.
Improving the diagnostic power of otoacoustic emissions. Arturo Moleti Physics Department University of Roma Tor Vergata
 Improving the diagnostic power of otoacoustic emissions Arturo Moleti Physics Department University of Roma Tor Vergata The human ear Ear canal: resonant cavity Middle ear: impedance adapter and pressure
Improving the diagnostic power of otoacoustic emissions Arturo Moleti Physics Department University of Roma Tor Vergata The human ear Ear canal: resonant cavity Middle ear: impedance adapter and pressure
Auditory System. Barb Rohrer (SEI )
") Auditory System Barb Rohrer (SEI614 2-5086) Sounds arise from mechanical vibration (creating zones of compression and rarefaction; which ripple outwards) Transmitted through gaseous, aqueous or solid medium
Auditory System Barb Rohrer (SEI614 2-5086) Sounds arise from mechanical vibration (creating zones of compression and rarefaction; which ripple outwards) Transmitted through gaseous, aqueous or solid medium
Cochlear anatomy, function and pathology I. Professor Dave Furness Keele University
 Cochlear anatomy, function and pathology I Professor Dave Furness Keele University d.n.furness@keele.ac.uk Aims and objectives of these lectures Introduction to gross anatomy of the cochlea Focus (1) on
Cochlear anatomy, function and pathology I Professor Dave Furness Keele University d.n.furness@keele.ac.uk Aims and objectives of these lectures Introduction to gross anatomy of the cochlea Focus (1) on
Signals, systems, acoustics and the ear. Week 5. The peripheral auditory system: The ear as a signal processor
 Signals, systems, acoustics and the ear Week 5 The peripheral auditory system: The ear as a signal processor Think of this set of organs 2 as a collection of systems, transforming sounds to be sent to
Signals, systems, acoustics and the ear Week 5 The peripheral auditory system: The ear as a signal processor Think of this set of organs 2 as a collection of systems, transforming sounds to be sent to
Sound and Hearing. Decibels. Frequency Coding & Localization 1. Everything is vibration. The universe is made of waves.
 Frequency Coding & Localization 1 Sound and Hearing Everything is vibration The universe is made of waves db = 2log(P1/Po) P1 = amplitude of the sound wave Po = reference pressure =.2 dynes/cm 2 Decibels
Frequency Coding & Localization 1 Sound and Hearing Everything is vibration The universe is made of waves db = 2log(P1/Po) P1 = amplitude of the sound wave Po = reference pressure =.2 dynes/cm 2 Decibels
A computer model of medial efferent suppression in the mammalian auditory system
 A computer model of medial efferent suppression in the mammalian auditory system Robert T. Ferry a and Ray Meddis Department of Psychology, University of Essex, Colchester, CO4 3SQ, United Kingdom Received
A computer model of medial efferent suppression in the mammalian auditory system Robert T. Ferry a and Ray Meddis Department of Psychology, University of Essex, Colchester, CO4 3SQ, United Kingdom Received
Modeling of Mechanoelectrical Transduction of Hair Cells to Action Potentials in the Auditory Nerve
 Modeling of Mechanoelectrical Transduction of Hair Cells to Action Potentials in the Auditory Nerve Anthony Au, Yuchen Wang, Sibai Xie A Term Paper for BENG26 University of California, San Diego San Diego,
Modeling of Mechanoelectrical Transduction of Hair Cells to Action Potentials in the Auditory Nerve Anthony Au, Yuchen Wang, Sibai Xie A Term Paper for BENG26 University of California, San Diego San Diego,
HST 721 Lecture 4: Mechanics, electromotility and the cochlear amplifier
 HST 721 Lecture 4: Mechanics, electromotility and the cochlear amplifier 1 Cochlear Mechanics: Measures of Basilar Membrane Motion 2 Cochlear Mechanics: Measures of Basilar Membrane Motion Bekesy s experiments
HST 721 Lecture 4: Mechanics, electromotility and the cochlear amplifier 1 Cochlear Mechanics: Measures of Basilar Membrane Motion 2 Cochlear Mechanics: Measures of Basilar Membrane Motion Bekesy s experiments
Innervation of the Cochlea. Reading: Yost Ch. 8
 Innervation of the Cochlea Reading: Yost Ch. 8 Fine Structure of the Organ of Corti Auditory Nerve Auditory nerve (AN) is a branch of the VIII th cranial nerve (other branch is vestibular). AN is composed
Innervation of the Cochlea Reading: Yost Ch. 8 Fine Structure of the Organ of Corti Auditory Nerve Auditory nerve (AN) is a branch of the VIII th cranial nerve (other branch is vestibular). AN is composed
Auditory Physiology Richard M. Costanzo, Ph.D.
 Auditory Physiology Richard M. Costanzo, Ph.D. OBJECTIVES After studying the material of this lecture, the student should be able to: 1. Describe the morphology and function of the following structures:
Auditory Physiology Richard M. Costanzo, Ph.D. OBJECTIVES After studying the material of this lecture, the student should be able to: 1. Describe the morphology and function of the following structures:
Auditory nerve model for predicting performance limits of normal and impaired listeners
 Heinz et al.: Acoustics Research Letters Online [DOI 1.1121/1.1387155] Published Online 12 June 21 Auditory nerve model for predicting performance limits of normal and impaired listeners Michael G. Heinz
Heinz et al.: Acoustics Research Letters Online [DOI 1.1121/1.1387155] Published Online 12 June 21 Auditory nerve model for predicting performance limits of normal and impaired listeners Michael G. Heinz
Stimulus Coding in the Auditory Nerve. Neural Coding and Perception of Sound 1
 Stimulus Coding in the Auditory Nerve Neural Coding and Perception of Sound 1 Point of View AUDITORY NERVE SOUND EAR BRAIN BEHAVIOR The auditory nerve conveys information about sound from the ear to the
Stimulus Coding in the Auditory Nerve Neural Coding and Perception of Sound 1 Point of View AUDITORY NERVE SOUND EAR BRAIN BEHAVIOR The auditory nerve conveys information about sound from the ear to the
Electrophysiology. General Neurophysiology. Action Potentials
 5 Electrophysiology Cochlear implants should aim to reproduce the coding of sound in the auditory system as closely as possible, for best sound perception. The cochlear implant is in part the result of
5 Electrophysiology Cochlear implants should aim to reproduce the coding of sound in the auditory system as closely as possible, for best sound perception. The cochlear implant is in part the result of
AUDL GS08/GAV1 Signals, systems, acoustics and the ear. Pitch & Binaural listening
 AUDL GS08/GAV1 Signals, systems, acoustics and the ear Pitch & Binaural listening Review 25 20 15 10 5 0-5 100 1000 10000 25 20 15 10 5 0-5 100 1000 10000 Part I: Auditory frequency selectivity Tuning
AUDL GS08/GAV1 Signals, systems, acoustics and the ear Pitch & Binaural listening Review 25 20 15 10 5 0-5 100 1000 10000 25 20 15 10 5 0-5 100 1000 10000 Part I: Auditory frequency selectivity Tuning
Acoustics Research Institute
 Austrian Academy of Sciences Acoustics Research Institute Modeling Modelingof ofauditory AuditoryPerception Perception Bernhard BernhardLaback Labackand andpiotr PiotrMajdak Majdak http://www.kfs.oeaw.ac.at
Austrian Academy of Sciences Acoustics Research Institute Modeling Modelingof ofauditory AuditoryPerception Perception Bernhard BernhardLaback Labackand andpiotr PiotrMajdak Majdak http://www.kfs.oeaw.ac.at
SOLUTIONS Homework #3. Introduction to Engineering in Medicine and Biology ECEN 1001 Due Tues. 9/30/03
 SOLUTIONS Homework #3 Introduction to Engineering in Medicine and Biology ECEN 1001 Due Tues. 9/30/03 Problem 1: a) Where in the cochlea would you say the process of "fourier decomposition" of the incoming
SOLUTIONS Homework #3 Introduction to Engineering in Medicine and Biology ECEN 1001 Due Tues. 9/30/03 Problem 1: a) Where in the cochlea would you say the process of "fourier decomposition" of the incoming
Comment by Delgutte and Anna. A. Dreyer (Eaton-Peabody Laboratory, Massachusetts Eye and Ear Infirmary, Boston, MA)
") Comments Comment by Delgutte and Anna. A. Dreyer (Eaton-Peabody Laboratory, Massachusetts Eye and Ear Infirmary, Boston, MA) Is phase locking to transposed stimuli as good as phase locking to low-frequency
Comments Comment by Delgutte and Anna. A. Dreyer (Eaton-Peabody Laboratory, Massachusetts Eye and Ear Infirmary, Boston, MA) Is phase locking to transposed stimuli as good as phase locking to low-frequency
Deafness and hearing impairment
 Auditory Physiology Deafness and hearing impairment About one in every 10 Americans has some degree of hearing loss. The great majority develop hearing loss as they age. Hearing impairment in very early
Auditory Physiology Deafness and hearing impairment About one in every 10 Americans has some degree of hearing loss. The great majority develop hearing loss as they age. Hearing impairment in very early
Auditory nerve. Amanda M. Lauer, Ph.D. Dept. of Otolaryngology-HNS
 Auditory nerve Amanda M. Lauer, Ph.D. Dept. of Otolaryngology-HNS May 30, 2016 Overview Pathways (structural organization) Responses Damage Basic structure of the auditory nerve Auditory nerve in the cochlea
Auditory nerve Amanda M. Lauer, Ph.D. Dept. of Otolaryngology-HNS May 30, 2016 Overview Pathways (structural organization) Responses Damage Basic structure of the auditory nerve Auditory nerve in the cochlea
Unit VIII Problem 9 Physiology: Hearing
 Unit VIII Problem 9 Physiology: Hearing - We can hear a limited range of frequency between 20 Hz 20,000 Hz (human hearing acuity is between 1000 Hz 4000 Hz). - The ear is divided into 3 parts. Those are:
Unit VIII Problem 9 Physiology: Hearing - We can hear a limited range of frequency between 20 Hz 20,000 Hz (human hearing acuity is between 1000 Hz 4000 Hz). - The ear is divided into 3 parts. Those are:
Intro to Audition & Hearing
 Intro to Audition & Hearing Lecture 16 Chapter 9, part II Jonathan Pillow Sensation & Perception (PSY 345 / NEU 325) Fall 2017 1 Sine wave: one of the simplest kinds of sounds: sound for which pressure
Intro to Audition & Hearing Lecture 16 Chapter 9, part II Jonathan Pillow Sensation & Perception (PSY 345 / NEU 325) Fall 2017 1 Sine wave: one of the simplest kinds of sounds: sound for which pressure
Theme 2: Cellular mechanisms in the Cochlear Nucleus
 Theme 2: Cellular mechanisms in the Cochlear Nucleus The Cochlear Nucleus (CN) presents a unique opportunity for quantitatively studying input-output transformations by neurons because it gives rise to
Theme 2: Cellular mechanisms in the Cochlear Nucleus The Cochlear Nucleus (CN) presents a unique opportunity for quantitatively studying input-output transformations by neurons because it gives rise to
Fifth Quarterly Progress Report NO1-DC The Neurophysiological Effects of Simulated Auditory Prosthesis Stimulation
 Fifth Quarterly Progress Report NO1-DC-6-2111 The Neurophysiological Effects of Simulated Auditory Prosthesis Stimulation J.T. Rubinstein, P.J. Abbas, C.A. Miller and A.J. Matsuoka Department of Otolaryngology
Fifth Quarterly Progress Report NO1-DC-6-2111 The Neurophysiological Effects of Simulated Auditory Prosthesis Stimulation J.T. Rubinstein, P.J. Abbas, C.A. Miller and A.J. Matsuoka Department of Otolaryngology
SPHSC 462 HEARING DEVELOPMENT. Overview Review of Hearing Science Introduction
 SPHSC 462 HEARING DEVELOPMENT Overview Review of Hearing Science Introduction 1 Overview of course and requirements Lecture/discussion; lecture notes on website http://faculty.washington.edu/lawerner/sphsc462/
SPHSC 462 HEARING DEVELOPMENT Overview Review of Hearing Science Introduction 1 Overview of course and requirements Lecture/discussion; lecture notes on website http://faculty.washington.edu/lawerner/sphsc462/
The mammalian cochlea possesses two classes of afferent neurons and two classes of efferent neurons.
 1 The mammalian cochlea possesses two classes of afferent neurons and two classes of efferent neurons. Type I afferents contact single inner hair cells to provide acoustic analysis as we know it. Type
1 The mammalian cochlea possesses two classes of afferent neurons and two classes of efferent neurons. Type I afferents contact single inner hair cells to provide acoustic analysis as we know it. Type
NEURONS Chapter Neurons: specialized cells of the nervous system 2. Nerves: bundles of neuron axons 3. Nervous systems
 NEURONS Chapter 12 Figure 12.1 Neuronal and hormonal signaling both convey information over long distances 1. Nervous system A. nervous tissue B. conducts electrical impulses C. rapid communication 2.
NEURONS Chapter 12 Figure 12.1 Neuronal and hormonal signaling both convey information over long distances 1. Nervous system A. nervous tissue B. conducts electrical impulses C. rapid communication 2.
Lecture 6 Hearing 1. Raghav Rajan Bio 354 Neurobiology 2 January 28th All lecture material from the following links unless otherwise mentioned:
 Lecture 6 Hearing 1 All lecture material from the following links unless otherwise mentioned: 1. http://wws.weizmann.ac.il/neurobiology/labs/ulanovsky/sites/neurobiology.labs.ulanovsky/files/uploads/purves_ch12_ch13_hearing
Lecture 6 Hearing 1 All lecture material from the following links unless otherwise mentioned: 1. http://wws.weizmann.ac.il/neurobiology/labs/ulanovsky/sites/neurobiology.labs.ulanovsky/files/uploads/purves_ch12_ch13_hearing
ENT 318 Artificial Organs Physiology of Ear
 ENT 318 Artificial Organs Physiology of Ear Lecturer: Ahmad Nasrul Norali The Ear The Ear Components of hearing mechanism - Outer Ear - Middle Ear - Inner Ear - Central Auditory Nervous System Major Divisions
ENT 318 Artificial Organs Physiology of Ear Lecturer: Ahmad Nasrul Norali The Ear The Ear Components of hearing mechanism - Outer Ear - Middle Ear - Inner Ear - Central Auditory Nervous System Major Divisions
A truly remarkable aspect of human hearing is the vast
 AUDITORY COMPRESSION AND HEARING LOSS Sid P. Bacon Psychoacoustics Laboratory, Department of Speech and Hearing Science, Arizona State University Tempe, Arizona 85287 A truly remarkable aspect of human
AUDITORY COMPRESSION AND HEARING LOSS Sid P. Bacon Psychoacoustics Laboratory, Department of Speech and Hearing Science, Arizona State University Tempe, Arizona 85287 A truly remarkable aspect of human
Required Slide. Session Objectives
 Auditory Physiology Required Slide Session Objectives Auditory System: At the end of this session, students will be able to: 1. Characterize the range of normal human hearing. 2. Understand the components
Auditory Physiology Required Slide Session Objectives Auditory System: At the end of this session, students will be able to: 1. Characterize the range of normal human hearing. 2. Understand the components
STRUCTURAL ELEMENTS OF THE NERVOUS SYSTEM
 STRUCTURAL ELEMENTS OF THE NERVOUS SYSTEM STRUCTURE AND MAINTENANCE OF NEURONS (a) (b) Dendrites Cell body Initial segment collateral terminals (a) Diagrammatic representation of a neuron. The break in
STRUCTURAL ELEMENTS OF THE NERVOUS SYSTEM STRUCTURE AND MAINTENANCE OF NEURONS (a) (b) Dendrites Cell body Initial segment collateral terminals (a) Diagrammatic representation of a neuron. The break in
Chapter 11: Sound, The Auditory System, and Pitch Perception
 Chapter 11: Sound, The Auditory System, and Pitch Perception Overview of Questions What is it that makes sounds high pitched or low pitched? How do sound vibrations inside the ear lead to the perception
Chapter 11: Sound, The Auditory System, and Pitch Perception Overview of Questions What is it that makes sounds high pitched or low pitched? How do sound vibrations inside the ear lead to the perception
Ameen Alsaras. Ameen Alsaras. Mohd.Khatatbeh
 9 Ameen Alsaras Ameen Alsaras Mohd.Khatatbeh Nerve Cells (Neurons) *Remember: The neural cell consists of: 1-Cell body 2-Dendrites 3-Axon which ends as axon terminals. The conduction of impulse through
9 Ameen Alsaras Ameen Alsaras Mohd.Khatatbeh Nerve Cells (Neurons) *Remember: The neural cell consists of: 1-Cell body 2-Dendrites 3-Axon which ends as axon terminals. The conduction of impulse through
Chapter 7 Nerve Cells and Electrical Signaling
 Chapter 7 Nerve Cells and Electrical Signaling 7.1. Overview of the Nervous System (Figure 7.1) 7.2. Cells of the Nervous System o Neurons are excitable cells which can generate action potentials o 90%
Chapter 7 Nerve Cells and Electrical Signaling 7.1. Overview of the Nervous System (Figure 7.1) 7.2. Cells of the Nervous System o Neurons are excitable cells which can generate action potentials o 90%
HCS 7367 Speech Perception
 Long-term spectrum of speech HCS 7367 Speech Perception Connected speech Absolute threshold Males Dr. Peter Assmann Fall 212 Females Long-term spectrum of speech Vowels Males Females 2) Absolute threshold
Long-term spectrum of speech HCS 7367 Speech Perception Connected speech Absolute threshold Males Dr. Peter Assmann Fall 212 Females Long-term spectrum of speech Vowels Males Females 2) Absolute threshold
THE INTERPLAY BETWEEN ACTIVE HAIR BUNDLE MECHANICS AND ELECTROMOTILITY IN THE COCHLEA
 451 THE INTERPLAY BETWEEN ACTIVE HAIR BUNDLE MECHANICS AND ELECTROMOTILITY IN THE COCHLEA DÁIBHID Ó MAOILÉIDIGH, FRANK JÜLICHER Max Planck Institute für Physik komplexer Systeme, Nöthnitzerstr. 38, 01187
451 THE INTERPLAY BETWEEN ACTIVE HAIR BUNDLE MECHANICS AND ELECTROMOTILITY IN THE COCHLEA DÁIBHID Ó MAOILÉIDIGH, FRANK JÜLICHER Max Planck Institute für Physik komplexer Systeme, Nöthnitzerstr. 38, 01187
Neurophysiology of Nerve Impulses
 M52_MARI0000_00_SE_EX03.qxd 8/22/11 2:47 PM Page 358 3 E X E R C I S E Neurophysiology of Nerve Impulses Advance Preparation/Comments Consider doing a short introductory presentation with the following
M52_MARI0000_00_SE_EX03.qxd 8/22/11 2:47 PM Page 358 3 E X E R C I S E Neurophysiology of Nerve Impulses Advance Preparation/Comments Consider doing a short introductory presentation with the following
Thursday, January 22, Nerve impulse
 Nerve impulse Transmembrane Potential caused by ions moving through cell membrane at different rates Two main ions of concern Na + - Sodium K + - potassium Cell membrane not freely permeable therefore
Nerve impulse Transmembrane Potential caused by ions moving through cell membrane at different rates Two main ions of concern Na + - Sodium K + - potassium Cell membrane not freely permeable therefore
HST 721 Efferent Control Lecture October 2004
 HST 721 Efferent Control Lecture October 2004 1 Stapedius Muscle Central Circuitry 2 Hypotheses for MEM Function A. Stapedius 1. Extend Dynamic Range - a gain control system 2. Protect the Inner Ear from
HST 721 Efferent Control Lecture October 2004 1 Stapedius Muscle Central Circuitry 2 Hypotheses for MEM Function A. Stapedius 1. Extend Dynamic Range - a gain control system 2. Protect the Inner Ear from
Neurobiology Biomed 509 Sensory transduction References: Luo , ( ), , M4.1, M6.2
, , M4.1, M6.2") Neurobiology Biomed 509 Sensory transduction References: Luo 4.1 4.8, (4.9 4.23), 6.22 6.24, M4.1, M6.2 I. Transduction The role of sensory systems is to convert external energy into electrical signals
Neurobiology Biomed 509 Sensory transduction References: Luo 4.1 4.8, (4.9 4.23), 6.22 6.24, M4.1, M6.2 I. Transduction The role of sensory systems is to convert external energy into electrical signals
Spectrograms (revisited)
") Spectrograms (revisited) We begin the lecture by reviewing the units of spectrograms, which I had only glossed over when I covered spectrograms at the end of lecture 19. We then relate the blocks of a
Spectrograms (revisited) We begin the lecture by reviewing the units of spectrograms, which I had only glossed over when I covered spectrograms at the end of lecture 19. We then relate the blocks of a
Structure, Energy Transmission and Function. Gross Anatomy. Structure, Function & Process. External Auditory Meatus or Canal (EAM, EAC) Outer Ear
 Outer Ear") Gross Anatomy Structure, Energy Transmission and Function IE N O ME 1 Structure, Function & Process 4 External Auditory Meatus or Canal (EAM, EAC) Outer third is cartilaginous Inner 2/3 is osseous Junction
Gross Anatomy Structure, Energy Transmission and Function IE N O ME 1 Structure, Function & Process 4 External Auditory Meatus or Canal (EAM, EAC) Outer third is cartilaginous Inner 2/3 is osseous Junction
to vibrate the fluid. The ossicles amplify the pressure. The surface area of the oval window is
 Page 1 of 6 Question 1: How is the conduction of sound to the cochlea facilitated by the ossicles of the middle ear? Answer: Sound waves traveling through air move the tympanic membrane, which, in turn,
Page 1 of 6 Question 1: How is the conduction of sound to the cochlea facilitated by the ossicles of the middle ear? Answer: Sound waves traveling through air move the tympanic membrane, which, in turn,
Lecture 3: Perception
 ELEN E4896 MUSIC SIGNAL PROCESSING Lecture 3: Perception 1. Ear Physiology 2. Auditory Psychophysics 3. Pitch Perception 4. Music Perception Dan Ellis Dept. Electrical Engineering, Columbia University
ELEN E4896 MUSIC SIGNAL PROCESSING Lecture 3: Perception 1. Ear Physiology 2. Auditory Psychophysics 3. Pitch Perception 4. Music Perception Dan Ellis Dept. Electrical Engineering, Columbia University
Spatiotemporal coding of sound in the auditory nerve for cochlear implants
 Spatiotemporal coding of sound in the auditory nerve for cochlear implants Ian Christopher Bruce BE (Melb) Submitted in total fulfilment of the requirements of the degree of Doctor of Philosophy December
Spatiotemporal coding of sound in the auditory nerve for cochlear implants Ian Christopher Bruce BE (Melb) Submitted in total fulfilment of the requirements of the degree of Doctor of Philosophy December
Lauer et al Olivocochlear efferents. Amanda M. Lauer, Ph.D. Dept. of Otolaryngology-HNS
 Lauer et al. 2012 Olivocochlear efferents Amanda M. Lauer, Ph.D. Dept. of Otolaryngology-HNS May 30, 2016 Overview Structural organization Responses Hypothesized roles in hearing Olivocochlear efferent
Lauer et al. 2012 Olivocochlear efferents Amanda M. Lauer, Ph.D. Dept. of Otolaryngology-HNS May 30, 2016 Overview Structural organization Responses Hypothesized roles in hearing Olivocochlear efferent
Temporal coding in the sub-millisecond range: Model of barn owl auditory pathway
 Temporal coding in the sub-millisecond range: Model of barn owl auditory pathway Richard Kempter* Institut fur Theoretische Physik Physik-Department der TU Munchen D-85748 Garching bei Munchen J. Leo van
Temporal coding in the sub-millisecond range: Model of barn owl auditory pathway Richard Kempter* Institut fur Theoretische Physik Physik-Department der TU Munchen D-85748 Garching bei Munchen J. Leo van
Nature Biotechnology: doi: /nbt Supplementary Figure 1. Analysis of hair bundle morphology in Ush1c c.216g>a mice at P18 by SEM.
 Supplementary Figure 1 Analysis of hair bundle morphology in Ush1c c.216g>a mice at P18 by SEM. (a-c) Heterozygous c.216ga mice displayed normal hair bundle morphology at P18. (d-i) Disorganized hair bundles
Supplementary Figure 1 Analysis of hair bundle morphology in Ush1c c.216g>a mice at P18 by SEM. (a-c) Heterozygous c.216ga mice displayed normal hair bundle morphology at P18. (d-i) Disorganized hair bundles
FIFTH INTERNATIONAL CONGRESS ON SOUND AND VIBRATION DECEMBER 15-18, 1997 ADELAIDE, SOUTH AUSTRALIA. Invited Paper LINEAR RESPONSE OF THE COCHLEA
 FIFTH INTERNATIONAL CONGRESS ON SOUND AND VIBRATION DECEMBER 15-18, 1997 ADELAIDE, SOUTH AUSTRALIA Invited Paper LINEAR RESPONSE OF THE COCHLEA David Alan Bies Visiting Research Fellow Department of Mechanical
FIFTH INTERNATIONAL CONGRESS ON SOUND AND VIBRATION DECEMBER 15-18, 1997 ADELAIDE, SOUTH AUSTRALIA Invited Paper LINEAR RESPONSE OF THE COCHLEA David Alan Bies Visiting Research Fellow Department of Mechanical
Temporal Adaptation. In a Silicon Auditory Nerve. John Lazzaro. CS Division UC Berkeley 571 Evans Hall Berkeley, CA
 Temporal Adaptation In a Silicon Auditory Nerve John Lazzaro CS Division UC Berkeley 571 Evans Hall Berkeley, CA 94720 Abstract Many auditory theorists consider the temporal adaptation of the auditory
Temporal Adaptation In a Silicon Auditory Nerve John Lazzaro CS Division UC Berkeley 571 Evans Hall Berkeley, CA 94720 Abstract Many auditory theorists consider the temporal adaptation of the auditory
Sound waves from the auditory environment all combine in the ear canal to form a complex waveform. This waveform is deconstructed by the cochlea with
 1 Sound waves from the auditory environment all combine in the ear canal to form a complex waveform. This waveform is deconstructed by the cochlea with respect to time, loudness, and frequency and neural
1 Sound waves from the auditory environment all combine in the ear canal to form a complex waveform. This waveform is deconstructed by the cochlea with respect to time, loudness, and frequency and neural
9.01 Introduction to Neuroscience Fall 2007
 MIT OpenCourseWare http://ocw.mit.edu 9.01 Introduction to Neuroscience Fall 2007 For information about citing these materials or our Terms of Use, visit: http://ocw.mit.edu/terms. 9.01 Recitation (R02)
MIT OpenCourseWare http://ocw.mit.edu 9.01 Introduction to Neuroscience Fall 2007 For information about citing these materials or our Terms of Use, visit: http://ocw.mit.edu/terms. 9.01 Recitation (R02)
So now to The Ear. Drawings from Max Brodel, an Austrian artist who came to Johns Hopkins in the 1920s. My point in showing this figure is to
 So now to The Ear. Drawings from Max Brodel, an Austrian artist who came to Johns Hopkins in the 1920s. My point in showing this figure is to emphasize the intricate and well-protected structure of the
So now to The Ear. Drawings from Max Brodel, an Austrian artist who came to Johns Hopkins in the 1920s. My point in showing this figure is to emphasize the intricate and well-protected structure of the
Information Processing During Transient Responses in the Crayfish Visual System
 Information Processing During Transient Responses in the Crayfish Visual System Christopher J. Rozell, Don. H. Johnson and Raymon M. Glantz Department of Electrical & Computer Engineering Department of
Information Processing During Transient Responses in the Crayfish Visual System Christopher J. Rozell, Don. H. Johnson and Raymon M. Glantz Department of Electrical & Computer Engineering Department of
A functional point-neuron model simulating cochlear nucleus ideal onset responses
 Downloaded from orbit.dtu.dk on: Dec 16, 2017 A functional point-neuron model simulating cochlear nucleus ideal onset responses Dicke, Ulrike; Dau, Torsten Published in: Journal of Computational Neuroscience
Downloaded from orbit.dtu.dk on: Dec 16, 2017 A functional point-neuron model simulating cochlear nucleus ideal onset responses Dicke, Ulrike; Dau, Torsten Published in: Journal of Computational Neuroscience
Auditory Physiology PSY 310 Greg Francis. Lecture 29. Hearing
 Auditory Physiology PSY 310 Greg Francis Lecture 29 A dangerous device. Hearing The sound stimulus is changes in pressure The simplest sounds vary in: Frequency: Hertz, cycles per second. How fast the
Auditory Physiology PSY 310 Greg Francis Lecture 29 A dangerous device. Hearing The sound stimulus is changes in pressure The simplest sounds vary in: Frequency: Hertz, cycles per second. How fast the
PSY 310: Sensory and Perceptual Processes 1
 Auditory Physiology PSY 310 Greg Francis Lecture 29 A dangerous device. Hearing The sound stimulus is changes in pressure The simplest sounds vary in: Frequency: Hertz, cycles per second. How fast the
Auditory Physiology PSY 310 Greg Francis Lecture 29 A dangerous device. Hearing The sound stimulus is changes in pressure The simplest sounds vary in: Frequency: Hertz, cycles per second. How fast the
Chapter 17, Part 2! The Special Senses! Hearing and Equilibrium!
 Chapter 17, Part 2! The Special Senses! Hearing and Equilibrium! SECTION 17-5! Equilibrium sensations originate within the inner ear, while hearing involves the detection and interpretation of sound waves!
Chapter 17, Part 2! The Special Senses! Hearing and Equilibrium! SECTION 17-5! Equilibrium sensations originate within the inner ear, while hearing involves the detection and interpretation of sound waves!
Chapter 17, Part 2! Chapter 17 Part 2 Special Senses! The Special Senses! Hearing and Equilibrium!
 Chapter 17, Part 2! The Special Senses! Hearing and Equilibrium! SECTION 17-5! Equilibrium sensations originate within the inner ear, while hearing involves the detection and interpretation of sound waves!
Chapter 17, Part 2! The Special Senses! Hearing and Equilibrium! SECTION 17-5! Equilibrium sensations originate within the inner ear, while hearing involves the detection and interpretation of sound waves!
Chapter 1: Introduction to digital audio
 Chapter 1: Introduction to digital audio Applications: audio players (e.g. MP3), DVD-audio, digital audio broadcast, music synthesizer, digital amplifier and equalizer, 3D sound synthesis 1 Properties
Chapter 1: Introduction to digital audio Applications: audio players (e.g. MP3), DVD-audio, digital audio broadcast, music synthesizer, digital amplifier and equalizer, 3D sound synthesis 1 Properties
Linguistic Phonetics. Basic Audition. Diagram of the inner ear removed due to copyright restrictions.
 24.963 Linguistic Phonetics Basic Audition Diagram of the inner ear removed due to copyright restrictions. 1 Reading: Keating 1985 24.963 also read Flemming 2001 Assignment 1 - basic acoustics. Due 9/22.
24.963 Linguistic Phonetics Basic Audition Diagram of the inner ear removed due to copyright restrictions. 1 Reading: Keating 1985 24.963 also read Flemming 2001 Assignment 1 - basic acoustics. Due 9/22.
An active cochlear model showing sharp tuning and high sensitivity
 Hearing Research, 9 (1983) 123-13 Elsevier Biomedical Press 123 An active cochlear model showing sharp tuning and high sensitivity Stephen T. Neely * and D.O. Kim Box 811, Washington Unitlersit}', St.
Hearing Research, 9 (1983) 123-13 Elsevier Biomedical Press 123 An active cochlear model showing sharp tuning and high sensitivity Stephen T. Neely * and D.O. Kim Box 811, Washington Unitlersit}', St.
J Jeffress model, 3, 66ff
 Index A Absolute pitch, 102 Afferent projections, inferior colliculus, 131 132 Amplitude modulation, coincidence detector, 152ff inferior colliculus, 152ff inhibition models, 156ff models, 152ff Anatomy,
Index A Absolute pitch, 102 Afferent projections, inferior colliculus, 131 132 Amplitude modulation, coincidence detector, 152ff inferior colliculus, 152ff inhibition models, 156ff models, 152ff Anatomy,
Sense system. Introduction The visual system Hearing. Introduction to sensory mechanisms
 Sense system Introduction The visual system Hearing Introduction to sensory mechanisms Sensory receptors & sense organs Sensory neurons & Receptor cells Adequate stimulus threshold Transduction Receptor
Sense system Introduction The visual system Hearing Introduction to sensory mechanisms Sensory receptors & sense organs Sensory neurons & Receptor cells Adequate stimulus threshold Transduction Receptor
Processing of sounds in the inner ear
 Processing of sounds in the inner ear Sripriya Ramamoorthy Associate Professor, IIT Bombay WiSSAP 2018 Cochlea converts sound into electrical signals [Picture courtesy of Northwestern University] von Bekesy
Processing of sounds in the inner ear Sripriya Ramamoorthy Associate Professor, IIT Bombay WiSSAP 2018 Cochlea converts sound into electrical signals [Picture courtesy of Northwestern University] von Bekesy
Effects of Remaining Hair Cells on Cochlear Implant Function
 Effects of Remaining Hair Cells on Cochlear Implant Function 2 nd Quarterly Progress Report Neural Prosthesis Program Contract N1-DC-2-15 (Quarter spanning Oct-Dec, 22) C.A. Miller, P.J. Abbas, N. Hu,
Effects of Remaining Hair Cells on Cochlear Implant Function 2 nd Quarterly Progress Report Neural Prosthesis Program Contract N1-DC-2-15 (Quarter spanning Oct-Dec, 22) C.A. Miller, P.J. Abbas, N. Hu,
211MDS Pain theories
 211MDS Pain theories Definition In 1986, the International Association for the Study of Pain (IASP) defined pain as a sensory and emotional experience associated with real or potential injuries, or described
211MDS Pain theories Definition In 1986, the International Association for the Study of Pain (IASP) defined pain as a sensory and emotional experience associated with real or potential injuries, or described
Emissions are low-intensity sounds that may be detected in the external ear canal by a microphone
 OAEs Emissions are low-intensity sounds that may be detected in the external ear canal by a microphone OAE is a pre-neural phenomenon They can be measured even when the 8 th cranial nerve is severely damaged
OAEs Emissions are low-intensity sounds that may be detected in the external ear canal by a microphone OAE is a pre-neural phenomenon They can be measured even when the 8 th cranial nerve is severely damaged
Trajectory of the Aging Cochlea
 Trajectory of the Aging Cochlea Sumitrajit (Sumit) Dhar Professor & Chair Roxelyn & Richard Pepper Department of Communication Sciences and Disorders Fellow, Hugh Knowles Center for Hearing Science Northwestern
Trajectory of the Aging Cochlea Sumitrajit (Sumit) Dhar Professor & Chair Roxelyn & Richard Pepper Department of Communication Sciences and Disorders Fellow, Hugh Knowles Center for Hearing Science Northwestern
Hearing Lectures. Acoustics of Speech and Hearing. Auditory Lighthouse. Facts about Timbre. Analysis of Complex Sounds
 Hearing Lectures Acoustics of Speech and Hearing Week 2-10 Hearing 3: Auditory Filtering 1. Loudness of sinusoids mainly (see Web tutorial for more) 2. Pitch of sinusoids mainly (see Web tutorial for more)
Hearing Lectures Acoustics of Speech and Hearing Week 2-10 Hearing 3: Auditory Filtering 1. Loudness of sinusoids mainly (see Web tutorial for more) 2. Pitch of sinusoids mainly (see Web tutorial for more)
An Auditory-Model-Based Electrical Stimulation Strategy Incorporating Tonal Information for Cochlear Implant
 Annual Progress Report An Auditory-Model-Based Electrical Stimulation Strategy Incorporating Tonal Information for Cochlear Implant Joint Research Centre for Biomedical Engineering Mar.7, 26 Types of Hearing
Annual Progress Report An Auditory-Model-Based Electrical Stimulation Strategy Incorporating Tonal Information for Cochlear Implant Joint Research Centre for Biomedical Engineering Mar.7, 26 Types of Hearing
Chapter 4 Neuronal Physiology
 Chapter 4 Neuronal Physiology V edit. Pg. 99-131 VI edit. Pg. 85-113 VII edit. Pg. 87-113 Input Zone Dendrites and Cell body Nucleus Trigger Zone Axon hillock Conducting Zone Axon (may be from 1mm to more
Chapter 4 Neuronal Physiology V edit. Pg. 99-131 VI edit. Pg. 85-113 VII edit. Pg. 87-113 Input Zone Dendrites and Cell body Nucleus Trigger Zone Axon hillock Conducting Zone Axon (may be from 1mm to more
Third Quarterly Progress Report NO1-DC The Neurophysiological Effects of Simulated Auditory Prosthesis Stimulation
 Third Quarterly Progress Report NO1-DC-6-2111 The Neurophysiological Effects of Simulated Auditory Prosthesis Stimulation C.A. Miller, P.J. Abbas, J.T. Rubinstein, and A.J. Matsuoka Department of Otolaryngology
Third Quarterly Progress Report NO1-DC-6-2111 The Neurophysiological Effects of Simulated Auditory Prosthesis Stimulation C.A. Miller, P.J. Abbas, J.T. Rubinstein, and A.J. Matsuoka Department of Otolaryngology
Intro. Comp. NeuroSci. Ch. 9 October 4, The threshold and channel memory
 9.7.4 The threshold and channel memory The action potential has a threshold. In figure the area around threshold is expanded (rectangle). A current injection that does not reach the threshold does not
9.7.4 The threshold and channel memory The action potential has a threshold. In figure the area around threshold is expanded (rectangle). A current injection that does not reach the threshold does not
EMANATIONS FROM RESIDUUM OSCILLATIONS IN HUMAN AUDITORY SYSTEM
 EMANATIONS FROM RESIDUUM OSCILLATIONS IN HUMAN AUDITORY SYSTEM V.S. Balaji, N.R.Raajan, S. Rakesh Kumar, Har Narayan Upadhyay School of Electrical & Electronics Engineering, SASTRA University Thanjavur,
EMANATIONS FROM RESIDUUM OSCILLATIONS IN HUMAN AUDITORY SYSTEM V.S. Balaji, N.R.Raajan, S. Rakesh Kumar, Har Narayan Upadhyay School of Electrical & Electronics Engineering, SASTRA University Thanjavur,
Outer Hair Cells in the Cochlea
 Discrete Dynamics in Nature and Society, Vol. 2, pp. 161-165 Reprints available directly from the publisher Photocopying permitted by license only (C) 1998 OPA (Overseas Publishers Association) N.V. Published
Discrete Dynamics in Nature and Society, Vol. 2, pp. 161-165 Reprints available directly from the publisher Photocopying permitted by license only (C) 1998 OPA (Overseas Publishers Association) N.V. Published
Hearing Sound. The Human Auditory System. The Outer Ear. Music 170: The Ear
 Hearing Sound Music 170: The Ear Tamara Smyth, trsmyth@ucsd.edu Department of Music, University of California, San Diego (UCSD) November 17, 2016 Sound interpretation in the auditory system is done by
Hearing Sound Music 170: The Ear Tamara Smyth, trsmyth@ucsd.edu Department of Music, University of California, San Diego (UCSD) November 17, 2016 Sound interpretation in the auditory system is done by
Music 170: The Ear. Tamara Smyth, Department of Music, University of California, San Diego (UCSD) November 17, 2016
 November 17, 2016") Music 170: The Ear Tamara Smyth, trsmyth@ucsd.edu Department of Music, University of California, San Diego (UCSD) November 17, 2016 1 Hearing Sound Sound interpretation in the auditory system is done by
Music 170: The Ear Tamara Smyth, trsmyth@ucsd.edu Department of Music, University of California, San Diego (UCSD) November 17, 2016 1 Hearing Sound Sound interpretation in the auditory system is done by
Effects of Remaining Hair Cells on Cochlear Implant Function
 Effects of Remaining Hair Cells on Cochlear Implant Function 5th Quarterly Progress Report Neural Prosthesis Program Contract N01-DC-2-1005 (Quarter spanning July-Sept, 2003) K.V. Nourski, P.J. Abbas,
Effects of Remaining Hair Cells on Cochlear Implant Function 5th Quarterly Progress Report Neural Prosthesis Program Contract N01-DC-2-1005 (Quarter spanning July-Sept, 2003) K.V. Nourski, P.J. Abbas,
Spectro-temporal response fields in the inferior colliculus of awake monkey
 3.6.QH Spectro-temporal response fields in the inferior colliculus of awake monkey Versnel, Huib; Zwiers, Marcel; Van Opstal, John Department of Biophysics University of Nijmegen Geert Grooteplein 655
3.6.QH Spectro-temporal response fields in the inferior colliculus of awake monkey Versnel, Huib; Zwiers, Marcel; Van Opstal, John Department of Biophysics University of Nijmegen Geert Grooteplein 655
Coding of Sensory Information
 Coding of Sensory Information 22 November, 2016 Touqeer Ahmed PhD Atta-ur-Rahman School of Applied Biosciences National University of Sciences and Technology Sensory Systems Mediate Four Attributes of
Coding of Sensory Information 22 November, 2016 Touqeer Ahmed PhD Atta-ur-Rahman School of Applied Biosciences National University of Sciences and Technology Sensory Systems Mediate Four Attributes of
Carlson (7e) PowerPoint Lecture Outline Chapter 7: Audition, the Body Senses, and the Chemical Senses
 PowerPoint Lecture Outline Chapter 7: Audition, the Body Senses, and the Chemical Senses") Carlson (7e) PowerPoint Lecture Outline Chapter 7: Audition, the Body Senses, and the Chemical Senses This multimedia product and its contents are protected under copyright law. The following are prohibited
Carlson (7e) PowerPoint Lecture Outline Chapter 7: Audition, the Body Senses, and the Chemical Senses This multimedia product and its contents are protected under copyright law. The following are prohibited
Thresholds for different mammals
 Loudness Thresholds for different mammals 8 7 What s the first thing you d want to know? threshold (db SPL) 6 5 4 3 2 1 hum an poodle m ouse threshold Note bowl shape -1 1 1 1 1 frequency (Hz) Sivian &
Loudness Thresholds for different mammals 8 7 What s the first thing you d want to know? threshold (db SPL) 6 5 4 3 2 1 hum an poodle m ouse threshold Note bowl shape -1 1 1 1 1 frequency (Hz) Sivian &
Binaurally-coherent jitter improves neural and perceptual ITD sensitivity in normal and electric hearing
 Binaurally-coherent jitter improves neural and perceptual ITD sensitivity in normal and electric hearing M. Goupell 1 (matt.goupell@gmail.com), K. Hancock 2 (ken_hancock@meei.harvard.edu), P. Majdak 1
Binaurally-coherent jitter improves neural and perceptual ITD sensitivity in normal and electric hearing M. Goupell 1 (matt.goupell@gmail.com), K. Hancock 2 (ken_hancock@meei.harvard.edu), P. Majdak 1
Introduction to Neurobiology
 Biology 240 General Zoology Introduction to Neurobiology Nervous System functions: communication of information via nerve signals integration and processing of information control of physiological and
Biology 240 General Zoology Introduction to Neurobiology Nervous System functions: communication of information via nerve signals integration and processing of information control of physiological and
Auditory System & Hearing
 Auditory System & Hearing Chapters 9 and 10 Lecture 17 Jonathan Pillow Sensation & Perception (PSY 345 / NEU 325) Spring 2015 1 Cochlea: physical device tuned to frequency! place code: tuning of different
Auditory System & Hearing Chapters 9 and 10 Lecture 17 Jonathan Pillow Sensation & Perception (PSY 345 / NEU 325) Spring 2015 1 Cochlea: physical device tuned to frequency! place code: tuning of different