Molecular Medicine. Brown adipose tissue responds to cold and adrenergic
|
|
|
- Magdalen Nash
- 6 years ago
- Views:
Transcription
1 Brown adipose tissue responds to cold and adrenergic stimulation by induction of FGF21 Dionysios V. Chartoumpekis, 1, Ioannis G. Habeos, 1, Panos G. Ziros, 1 Agathoklis I. Psyrogiannis, 1 Venetsana E. Kyriazopoulou, 1 and Athanasios G. Papavassiliou 2 1 Department of Internal Medicine, Division of Endocrinology, Medical School, University of Patras, Patras, Greece; and 2 Department of Biological Chemistry, University of Athens Medical School, Athens, Greece equal contribution Address correspondence and reprint requests to Athanasios G. Papavassiliou, MD, PhD, Professor & Head, Department of Biological Chemistry, Medical School, University of Athens, 75, M. Asias Street, Athens, Greece. Phone: /9; Fax: ; papavas@med.uoa.gr. Running head: Cold induces FGF21 in BAT
2 ABSTRACT Fibroblast growth factor-21 (FGF21) is a pleiotropic protein involved in glucose, lipid metabolism and energy homeostasis with main tissues of expression the liver and adipose tissue. Brown adipose tissue (BAT) is responsible for cold-induced thermogenesis in rodents. The role of FGF21 in BAT biology has not been investigated. In the present study wild-type C57BL/6J mice as well as a brown adipocyte cell line were employed to explore the potential role of cold exposure and β3-adrenergic stimulation in the expression of FGF21 in BAT. Our results demonstrate that short-term exposure to cold, as well as β3-adrenergic stimulation causes a significant induction of FGF21 mrna levels in BAT, without a concomitant increase in FGF21 plasma levels. This finding opens new routes for the potential use of pharmaceuticals that could induce FGF21 hence activate BAT thermogenesis. Key words: FGF21, brown adipose tissue, thermogenesis, PGC-1a, PPARa, obesity
3 INTRODUCTION Fibroblast growth factor-21 (FGF21) belongs to the family of atypical FGFs which lack the conventional heparin-binding domain (1,2) and can thus diffuse away from their tissues of origin to function as hormones. FGF21 is abundantly expressed in liver, pancreas, and white adipose tissue (WAT) (3,4). It signals through cellsurface complexes of FGF receptors with the transmembrane protein β-klotho (5-8). The limited β-klotho expression in metabolically competent liver, pancreas, and adipose tissue permits FGF21 to selectively target these tissues, thus allowing this factor to influence glucose, lipid, and bodyweight homeostasis (8,9). FGF21 is involved in the adaptation of the body to starvation and acts to regulate fatty acid oxidation and ketone formation. The transcription factor peroxisome proliferator-activated receptor-alpha (PPARa) is a critical regulator of FGF21 (10). FGF21-infused mice have higher energy expenditure and elevated core body temperature, suggesting that FGF21 has catabolic functions (11,12). Centrally administered FGF21 induces energy expenditure and insulin sensitivity (13). Systemically administered FGF21 triggers a sustained lowering of blood glucose and triglycerides, improved insulin sensitivity, enrichment in brown adipocytes (14), preservation of β-cell function and mass (15), amelioration of obesity, and hepatosteatosis (10,12). In adipose tissue, FGF21 simultaneously induces lipid accumulation, uncoupling, biogenesis and inhibition of lipolysis, indicative of a state of futile cycling (11,16). Brown adipose tissue (BAT) burns fatty acids for heat production to defend the body against cold and has recently been shown to be present in humans (17).
4 Upon cold-triggered activation, BAT increases its energy demand and burns carbohydrates and lipids to produce heat using uncoupling protein-1 (UCP1) (18). β3- adrenergic receptors are expressed abundantly and predominantly in brown adipocytes and selective agonists of this receptor have been synthesized. Treatment of mice with such agonists doubles oxygen consumption, demonstrating the remarkable capacity of this thermogenic mechanism (19). PPAR-gamma co-activator-1a (PGC- 1a) is highly expressed in brown but not white fat, with marked and rapid induction in brown fat and skeletal muscle upon exposure of mice to cold. This cold induction of PGC-1a is largely due to sympathetic nervous system input through β-adrenergic receptors and camp action (20,21). There are reports that FGF21 is expressed in BAT but its role in metabolism has not been investigated (11,14,22). Instead, FGF21 produced in the liver promotes thermogenic activation of brown fat and is dispensable during starvation-induced torpor (23,24). Here we demonstrate that BAT of mice not only responds to FGF21 produced in the liver, but also over-expresses FGF21 after cold exposure or selective β3- adrenergic stimulation. This FGF21 might act as an autocrine factor. MATERIALS AND METHODS Mice Mice were bred and housed in the animal facility of the University of Patras Medical School at 22 C with ad libitum access to standard laboratory chow diet.we used male, age-matched (24 weeks) C57BL/6J (Jackson Laboratory) wild-type mice. For the cold experiments, mice were individually housed, fasted for 12 h and during the last 4 h of fasting they were exposed to either control (22 C) or low temperature
5 (4 C). At the end of the cold exposure, blood was collected and interscapular BAT, epididymal WAT and liver were harvested in RNA later solution. Similarly, the selective β3-adrenergic receptor agonist CL (Sigma, Germany; 2 mg/kg body weight) was given by intraperitoneal (ip) injection 4 h before the end of the experiment. All animal procedures were approved by the institutional review board of the University of Patras Medical School and were in accordance with EC Directive 86/609/EEC. Measurements of Hormones and Metabolites Plasma was collected using heparin as anticoagulant and was centrifuged at 2000 g for 20 min at 4 C. Plasma measurements were conducted following the manufacturer s instruction for each kit. ELISA kits were used for plasma leptin (ALPCO, Salem, NH) and FGF21 (R&D, Minneapolis, MN). Cholesterol and triglycerides were measured using an Olympus AU640 analyzer (Hamburg, Germany). Quantification of Gene Expression Levels Liver, BAT and WAT were submerged immediately after collection in RNA later solution (Ambion, Foster City, CA). Total RNA was isolated using Trizol reagent (Invitrogen) and further purified using the RNeasy mini kit (Qiagen, Hilden, Germany). A DNAse (TurboDNAse, Ambion) digestion step was included to prevent genomic DNA contamination. cdna was synthesized using the Superscript first strand synthesis system (Invitrogen) and real-time PCR (RT-PCR) reactions were performed in triplicate on a Step One Plus instrument (Applied Biosystems, Foster City, CA) using Taqman Gene Expression assays on demand (Applied Biosystems): FGF21, Mm _g1; PGC-1a, Mm _m1; PPARa, Mm _m1; GAPDH (glyceraldehyde-3-phosphate dehydrogenase), E. Relative mrna
6 levels were calculated by the comparative threshold cycle method using GAPDH as the housekeeping gene. Cell Culture and Treatments SV40T-immortalized brown adipocytes from the C57BL/6J strain of mice were kindly provided by Prof. Johannes Klein, Lübeck, German (25). Pre-adipocytes were grown to confluence in Dulbecco s modified Eagle s medium (DMEM; Life Technologies, Paisley, Strathclyde, UK) supplemented with 20% fetal bovine serum, 4.5 g/l glucose, 20 nm insulin, 1 nm triiodothyronine ( differentiation medium ), and penicillin/streptomycin. Adipocyte differentiation was induced by complementing the medium further with 250 µm indomethacin, 500 µm isobutylmethylxanthine and 2 µg/ml dexamethasone for 24 h when confluence was reached. Following this induction period, cells were changed back to differentiation medium. Cell culture was continued for five more days before cells were starved for 24 h with serum-free medium prior to carrying out the experiments. Maximally differentiated cells were treated, when indicated, with 50 μm of the β3-adrenergic receptor agonist CL for 6 h or with 10 μm of the PPARa antagonist GW6471 (Sigma, Germany) for 16 h before harvesting the cells. Statistical Analyses Experiments were performed three times using at least triplicate samples per group. Data were expressed as the mean ± SEM. Student s t-test or one-way ANOVA followed by Tukey s test were performed using GraphPad Prism 5 (GraphPad Software, La Jolla, CA). P<0.05 was considered significant. RESULTS
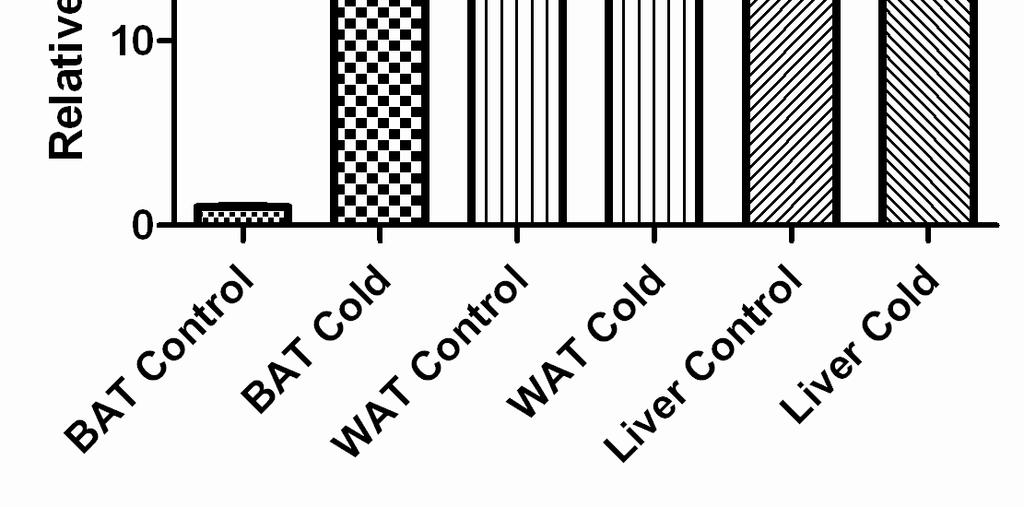
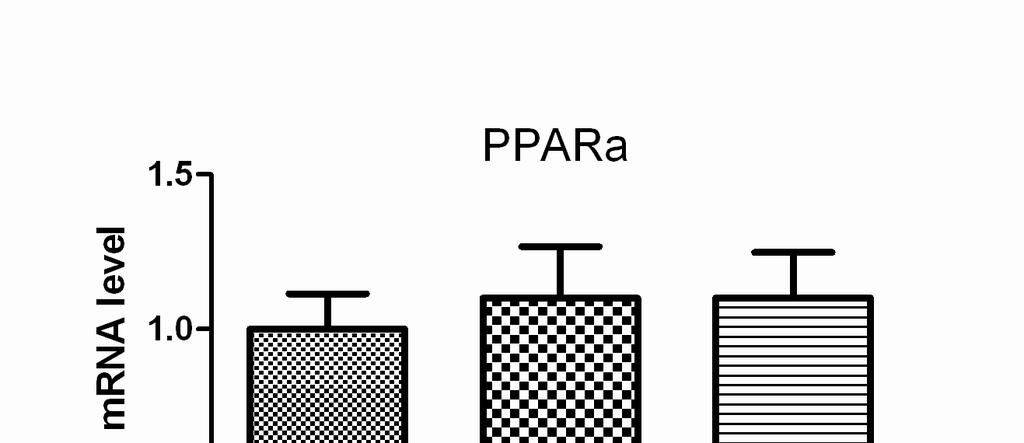
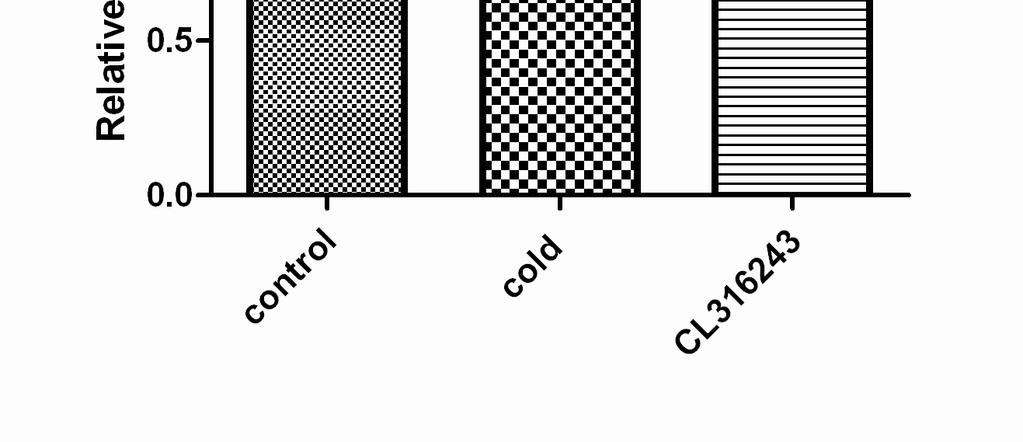
7 Cold Exposure Induces FGF21 Gene Expression in BAT Exposure of wild-type C57BL/6J mice to 4 ºC for 4 h caused a significant fold increase in FGF21 mrna levels in BAT over the base line (25.6±1.3, p<0.01, Figure 1A). It is known that after cold exposure PGC-1a mrna levels of BAT are induced as part of the cold-induced thermogenesis program. In the same experiment, PGC-1a mrna levels indeed increased by 15 fold, as expected (20). In contrast, cold did not change FGF21 mrna levels in neither the liver nor the WAT tissues where FGF21 is known to be mainly expressed (Figure 1A). Bearing in mind that the induction of FGF21 with fasting is PPARa dependent (10), PPARa mrna levels were tested in BAT. No difference however was found between control and cold-exposed animals (Figure 1B). Plasma Chemistries after Exposure to Cold Taking into account that FGF21 might be secreted and act as a hormone, plasma FGF21 levels were measured after exposure of mice to cold for 4 h. No differences were found between the two states. By contrast, plasma leptin and triglyceride levels dropped in accordance with other studies (Table 1) (26,27). β3-adrenergic Receptor Stimulation Induces FGF21 Gene Expression in BAT But not in the Liver Since it is known that cold activates BAT via sympathetic nervous system, a selective β3-agonist (CL316243) was tested for its ability to reproduce the above findings (28). The intraperitoneal administration of CL led to a significant fold increase in FGF21 mrna levels in BAT over the base line (19±0.8, p<0.01, Figure 2A). As a control, PGC-1a mrna levels were measured and found to be induced by
8 both cold and CL316243, as expected (Figure 2B). No differences in the expression of PPARa were found (Figure 2C). Treatment of Differentiated Mouse Brown Adipocytes With the β3-agonist Increases FGF21 mrna Levels In order to discriminate between cell autonomous or non autonomous effect of β3-adrenergic stimulation on FGF21 expression, a brown fat cell line able to differentiate into mature brown adipocytes and respond to the β3-agonist was used (25). Differentiated brown adipocytes were treated with CL (50 μm) for 6 h. The levels of FGF21 mrna were increased more than 5 fold (5.1±0.8, p<0.001, Figure 3). No change in FGF21 mrna was observed in undifferentiated brown preadipocytes (data not shown) since they do not express β3-adrenergic receptors (18). Treatment of the adipocytes with the PPARa antagonist GW6471 did not affect the response of these cells to the β3 agonist. DISCUSSION Ιn the present study we have shown that short-term exposure of mice to 4 ºC induced FGF21 mrna levels in brown fat. This induction is restricted to BAT, while no change is observed in liver and WAT. The baseline expression level of FGF21 in BAT is lower than that in WAT, but after cold exposure it is induced well above that in the latter. At the same time, plasma FGF21 was not found to increase accordingly by this short-term exposure to cold. BAT is heavily innervated by sympathetic nerves, and is responsible for thermogenesis during cold exposure. β3-adrenergic receptor agonists cause an increase in energy expenditure which is comparable to that induced by cold in both rodents and humans (19). In our experiments, stimulation of either mice or a brown
9 adipocyte cell line with the β3 agonist CL had the same affect on FGF21 expression as exposure to cold. This points to a novel mechanism of action of β3 agonists in brown fat through induction of FGF21 and suggests that some of the favourable effects of β3 agonists might be mediated via induction of FG21 (11-13,19). PGC-1a, a known target of β3-adrenergic receptor stimulation in brown fat (19), was also induced in our experimental system. It is noteworthy that FGF21 induces PGC-1a and regulates carbohydrate and fatty acid metabolism during the adaptive starvation response in liver (29). It remains to be seen if this applies to BAT as well. Leptin levels fall after cold exposure and this might be a signal for initiating a broad program of adaptation to starvation (30). FGF21 is considered a starvation factor as well (31). In this vein, the reduction of leptin levels observed after exposure to cold could be a permissive factor for the observed induction of FGF21 in BAT, although this could not explain the selectivity of the induction only in BAT. In liver, FGF21 has been shown to be induced after fasting in a PPARadependent manner. In our in vivo or in vitro experiments no differences were found in the expression of PPARa that could account for the aformentioned increase of FGF21. Nevertheless, activation of PPARa without any change in its expression levels could lead to elevation of FGF21. This is not supported by our findings since the PPARa antagonist GW6471 did not change the response of brown adipocytes to β3 agonist regarding FGF21 induction. The fact that FGF21 plasma levels did not change could be attributed to the small contribution of brown fat to the overall production of FGF21. In this case it is conceivable that FGF21 could be induced in BAT after cold exposure and act in an autocrine fashion. A general model for this kind of action of FGF21 is favoured by a recent review (31). Alternatively, the duration of cold exposure might have not been
10 sufficient enough to reveal any differences. Direct action of sympathetic nervous system on BAT and induction of FGF21 remains as the most plausible explanation. Hepatic FGF21 expression is induced at birth via PPARa in response to milk intake and contributes to thermogenic activation of neonatal brown fat (23). It can be envisioned that medications that augment the expression of FGF21 in brown fat could activate BAT thermogenesis and be used in the treatment of obesity. Experiments are under way in our laboratory to delineate the mechanisms of FGF21 induction by cold/ β3-adrenergic stimulation, as well as the significance of this induction in the thermogenic program of the brown fat adipocyte. We note that during finalizing this manuscript a paper addressing the same issue was published online, corroborating our main finding of FGF21 induction by cold in BAT (32). DISCLOSURE The authors declare that they have no competing interests as defined by Molecular Medicine, or other interests that might be perceived to influence the results and discussion reported in this paper.
11 REFERENCES 1. Itoh N, Ornitz DM. (2008) Functional evolutionary history of the mouse Fgf gene family. Dev. Dyn. 237: Goetz R, et al. (2007) Molecular insights into the klotho-dependent, endocrine mode of action of fibroblast growth factor 19 subfamily members. Mol. Cell. Biol. 27: Nishimura T, Nakatake Y, Konishi M, Itoh N. (2000) Identification of a novel FGF, FGF-21, preferentially expressed in the liver. Biochim. Biophys. Acta 1492: Muise ES, et al. (2008) Adipose fibroblast growth factor 21 is up-regulated by peroxisome proliferator-activated receptor gamma and altered metabolic states. Mol. Pharmacol. 74: Kharitonenkov A, et al. (2008) FGF-21/FGF-21 receptor interaction and activation is determined by betaklotho. J. Cell. Physiol. 215: Kurosu H, et al. (2007) Tissue-specific expression of betaklotho and fibroblast growth factor (FGF) receptor isoforms determines metabolic activity of FGF19 and FGF21. J. Biol. Chem. 282: Suzuki M, et al. (2008) betaklotho is required for fibroblast growth factor (FGF) 21 signaling through FGF receptor (FGFR) 1c and FGFR3c. Mol. Endocrinol. 22: Ogawa Y, et al. (2007) BetaKlotho is required for metabolic activity of fibroblast growth factor 21. Proc. Natl. Acad. Sci. U. S. A. 104: Ito S, et al. (2000) Molecular cloning and expression analyses of mouse betaklotho, which encodes a novel Klotho family protein. Mech. Dev. 98:115-9.
12 10. Badman MK, et al. (2007) Hepatic fibroblast growth factor 21 is regulated by PPARalpha and is a key mediator of hepatic lipid metabolism in ketotic states. Cell Metab. 5: Coskun T, et al. (2008) Fibroblast growth factor 21 corrects obesity in mice. Endocrinology 149: Xu J, et al. (2009) Fibroblast growth factor 21 reverses hepatic steatosis, increases energy expenditure, and improves insulin sensitivity in diet-induced obese mice. Diabetes 58: Sarruf DA, et al. (2010) Fibroblast growth factor 21 action in the brain increases energy expenditure and insulin sensitivity in obese rats. Diabetes 59: Kharitonenkov A, et al. (2005) FGF-21 as a novel metabolic regulator. J. Clin. Invest. 115: Wente W, et al. (2006) Fibroblast growth factor-21 improves pancreatic betacell function and survival by activation of extracellular signal-regulated kinase 1/2 and Akt signaling pathways. Diabetes 55: Xu J, et al. (2009) Acute glucose-lowering and insulin-sensitizing action of FGF21 in insulin resistant mouse models----association with liver and adipose tissue effects. Am. J. Physiol. Endocrinol. Metab. [Aug 25; Epub ahead of print] 17. van Marken Lichtenbelt WD, et al. (2009) Cold-activated brown adipose tissue in healthy men. N. Engl. J. Med. 360: Cannon B, Nedergaard J. (2004) Brown adipose tissue: function and physiological significance. Physiol. Rev. 84:
13 19. Lowell BB, Spiegelman BM. (2000) Towards a molecular understanding of adaptive thermogenesis. Nature 404: Puigserver P, et al. (1998) A cold-inducible coactivator of nuclear receptors linked to adaptive thermogenesis. Cell 92: Boss O, et al. (1999) Role of the beta(3)-adrenergic receptor and/or a putative beta(4)-adrenergic receptor on the expression of uncoupling proteins and peroxisome proliferator-activated receptor-gamma coactivator-1. Biochem. Biophys. Res. Commun. 261: Fon Tacer K, et al. (2010) Research resource: Comprehensive expression atlas of the fibroblast growth factor system in adult mouse. Mol. Endocrinol. 24: Hondares E, et al. (2010) Hepatic FGF21 expression is induced at birth via PPARalpha in response to milk intake and contributes to thermogenic activation of neonatal brown fat. Cell Metab. 11: Oishi K, et al. (2010) FGF21 is dispensable for hypothermia induced by fasting in mice. Neuro Endocrinol. Lett. 31: Klein J, et al. (1999) beta(3)-adrenergic stimulation differentially inhibits insulin signaling and decreases insulin-induced glucose uptake in brown adipocytes. J. Biol. Chem. 274: Bartelt A, et al. (2011) Brown adipose tissue activity controls triglyceride clearance. Nat. Med. 17: Trayhurn P, Duncan JS, Rayner DV. (1995) Acute cold-induced suppression of ob (obese) gene expression in white adipose tissue of mice: mediation by the sympathetic system. Biochem. J. 311(Pt 3):
14 28. Gomez-Ambrosi J, Fruhbeck G, Martinez JA. (2001) Rapid in vivo PGC-1 mrna upregulation in brown adipose tissue of Wistar rats by a beta(3)- adrenergic agonist and lack of effect of leptin. Mol. Cell. Endocrinol. 176: Potthoff MJ, et al. (2009) FGF21 induces PGC-1alpha and regulates carbohydrate and fatty acid metabolism during the adaptive starvation response. Proc. Natl. Acad. Sci. U. S. A. 106: Ahima RS, et al. (1996) Role of leptin in the neuroendocrine response to fasting. Nature 382: Kharitonenkov A, Larsen P. (2010) FGF21 reloaded: challenges of a rapidly growing field. Trends Endocrinol. Metab. [Dec 29; Epub ahead of print] 32. Hondares E, et al. (2011) Thermogenic activation induces FGF21 expression and release in brown adipose tissue. J. Biol. Chem. [Feb 13; Epub ahead of print]
15 FIGURE LEGENDS Figure 1. FGF21 mrna levels are induced in BAT of wild-type C57BL/6J mice after exposure to cold for 4 h. (A) FGF21 mrna levels were measured by quantitative RT-PCR in BAT, WAT and liver obtained from wild-type C57BL/6J mice. Bars show means ± SEM of 8 mice per group. Data were analyzed by one-way Anova. Statistically significant differences between control mice and mice exposed to 4 C for the same tissue are shown as * (p<0.001). Statistically significant differences between control tissues are depicted as **. (B) PPARa and PGC-1a mrna levels were measured by quantitative RT-PCR in BAT of mice. Data were analyzed by t-test for each gene separately. Statistically significant differences between control mice and mice exposed to 4 C are shown as *. Figure 2. FGF21 mrna levels are induced in BAT of wild-type C57BL/6J mice after stimulation with the β3-adrenergic agonist CL Mice were either exposed to 4 C for 4 h or injected ip (2 mg/kg body weight) with CL316243, 4 h before harvesting BAT. mrna levels for FGF21 (A), PGC-1a (B) and PPARa (C) were measured by quantitative RT-PCR. Bars show means ± SEM of 8 mice per group. Data were analyzed by one-way Anova. Statistically significant differences between control mice and mice exposed to 4 C or CL are shown as * (p<0.001). Figure 3. Induction of FGF21 mrna levels with the β3-adrenergic agonist CL is not hampered by co-administration of a PPARa antagonist. Differentiated brown adipocyte cells were treated with CL for 6 h in the presence or absence of GW6471, and FGF21 mrna levels were measured by quantitative RT-PCR. Data were analyzed by one-way Anova. Bars show means ±
16 D.V. Chartoumpekis et al. Molecular Medicine SEM of 3 independent experiments. Statistically significant differences between control and treated cells are denoted with * (p<0.01). 16
17 Table 1. Plasma metabolic parameters of C57BL/6J mice maintained at 22 C or after exposure to 4 C for 4 h. 22 C 4 C Triglycerides (mg/dl) 92±8 74±6* Cholesterol (mg/dl) 94±6 96±6 Leptin (pg/ml) 5200± ±70** FGF21 (pg/ml) 3620± ±173.4 Data represent mean ± SEM; n=10, * p<0.05, ** p<0.001
18
19
20
21
22
23
Brown Adipose Tissue Responds to Cold and Adrenergic Stimulation by Induction of FGF21
 Brown Adipose Tissue Responds to Cold and Adrenergic Stimulation by Induction of FGF21 Dionysios V Chartoumpekis, 1* Ioannis G Habeos, 1* Panos G Ziros, 1 Agathoklis I Psyrogiannis, 1 Venetsana E Kyriazopoulou,
Brown Adipose Tissue Responds to Cold and Adrenergic Stimulation by Induction of FGF21 Dionysios V Chartoumpekis, 1* Ioannis G Habeos, 1* Panos G Ziros, 1 Agathoklis I Psyrogiannis, 1 Venetsana E Kyriazopoulou,
GPR120 *** * * Liver BAT iwat ewat mwat Ileum Colon. UCP1 mrna ***
 a GPR120 GPR120 mrna/ppia mrna Arbitrary Units 150 100 50 Liver BAT iwat ewat mwat Ileum Colon b UCP1 mrna Fold induction 20 15 10 5 - camp camp SB202190 - - - H89 - - - - - GW7647 Supplementary Figure
a GPR120 GPR120 mrna/ppia mrna Arbitrary Units 150 100 50 Liver BAT iwat ewat mwat Ileum Colon b UCP1 mrna Fold induction 20 15 10 5 - camp camp SB202190 - - - H89 - - - - - GW7647 Supplementary Figure
BEIGE AND BROWN FAT: BASIC BIOLOGY AND NOVEL THERAPEUTICS Dr. Carl Ascoli
 BEIGE AND BROWN FAT: BASIC BIOLOGY AND NOVEL THERAPEUTICS Dr. Carl Ascoli Symposium Co-Chairs: Bruce M. Spiegelman (Harvard/Dana Farber) and Sven Enerbäck (U.Gothenburg) April 17-23, 2015 Snowbird Resort,
BEIGE AND BROWN FAT: BASIC BIOLOGY AND NOVEL THERAPEUTICS Dr. Carl Ascoli Symposium Co-Chairs: Bruce M. Spiegelman (Harvard/Dana Farber) and Sven Enerbäck (U.Gothenburg) April 17-23, 2015 Snowbird Resort,
Fibroblast growth factor 21 (FGF21) belongs to the. Nrf2 Represses FGF21 During Long-Term High-Fat Diet Induced Obesity in Mice
 belongs to the. Nrf2 Represses FGF21 During Long-Term High-Fat Diet Induced Obesity in Mice") ORIGINAL ARTICLE Nrf2 Represses FGF21 During Long-Term High-Fat Diet Induced Obesity in Mice Dionysios V. Chartoumpekis, 1 Panos G. Ziros, 1 Agathoklis I. Psyrogiannis, 1 Athanasios G. Papavassiliou, 2
ORIGINAL ARTICLE Nrf2 Represses FGF21 During Long-Term High-Fat Diet Induced Obesity in Mice Dionysios V. Chartoumpekis, 1 Panos G. Ziros, 1 Agathoklis I. Psyrogiannis, 1 Athanasios G. Papavassiliou, 2
SUPPLEMENTARY INFORMATION
 doi: 1.138/nature7221 Brown fat selective genes 12 1 Control Q-RT-PCR (% of Control) 8 6 4 2 Ntrk3 Cox7a1 Cox8b Cox5b ATPase b2 ATPase f1a1 Sirt3 ERRα Elovl3/Cig3 PPARα Zic1 Supplementary Figure S1. stimulates
doi: 1.138/nature7221 Brown fat selective genes 12 1 Control Q-RT-PCR (% of Control) 8 6 4 2 Ntrk3 Cox7a1 Cox8b Cox5b ATPase b2 ATPase f1a1 Sirt3 ERRα Elovl3/Cig3 PPARα Zic1 Supplementary Figure S1. stimulates
Fibroblast growth factor 21 (FGF21) belongs to the. Nrf2 Represses FGF21 During Long-Term High-Fat Diet Induced Obesity in Mice
 belongs to the. Nrf2 Represses FGF21 During Long-Term High-Fat Diet Induced Obesity in Mice") ORIGINAL ARTICLE Nrf2 Represses FGF21 During Long-Term High-Fat Diet Induced Obesity in Mice Dionysios V. Chartoumpekis, 1 Panos G. Ziros, 1 Agathoklis I. Psyrogiannis, 1 Athanasios G. Papavassiliou, 2
ORIGINAL ARTICLE Nrf2 Represses FGF21 During Long-Term High-Fat Diet Induced Obesity in Mice Dionysios V. Chartoumpekis, 1 Panos G. Ziros, 1 Agathoklis I. Psyrogiannis, 1 Athanasios G. Papavassiliou, 2
A Tryptophan Hydroxylase Inhibitor Increases Hepatic FGF21 Production and Decreases Hepatic Gluconeogenesis Independently of Insulin in db/db Mice
 A Tryptophan Hydroxylase Inhibitor Increases Hepatic FGF21 Production and Decreases Hepatic Gluconeogenesis Independently of Insulin in db/db Mice Katsunori Nonogaki, Takao Kaji, Mari Murakami ABSTRACT
A Tryptophan Hydroxylase Inhibitor Increases Hepatic FGF21 Production and Decreases Hepatic Gluconeogenesis Independently of Insulin in db/db Mice Katsunori Nonogaki, Takao Kaji, Mari Murakami ABSTRACT
ab Adipogenesis Assay Kit (Cell-Based)
") ab133102 Adipogenesis Assay Kit (Cell-Based) Instructions for Use For the study of induction and inhibition of adipogenesis in adherent cells. This product is for research use only and is not intended
ab133102 Adipogenesis Assay Kit (Cell-Based) Instructions for Use For the study of induction and inhibition of adipogenesis in adherent cells. This product is for research use only and is not intended
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature12652 Supplementary Figure 1. PRDM16 interacts with endogenous EHMT1 in brown adipocytes. Immunoprecipitation of PRDM16 complex by flag antibody (M2) followed by Western blot analysis
doi:10.1038/nature12652 Supplementary Figure 1. PRDM16 interacts with endogenous EHMT1 in brown adipocytes. Immunoprecipitation of PRDM16 complex by flag antibody (M2) followed by Western blot analysis
A synergistic anti-obesity effect by a combination of capsinoids and cold temperature through the promotion of beige adipocyte biogenesis
 A synergistic anti-obesity effect by a combination of capsinoids and cold temperature through the promotion of beige adipocyte biogenesis Kana Ohyama, 1,2 Yoshihito Nogusa, 1 Kosaku Shinoda, 2 Katsuya
A synergistic anti-obesity effect by a combination of capsinoids and cold temperature through the promotion of beige adipocyte biogenesis Kana Ohyama, 1,2 Yoshihito Nogusa, 1 Kosaku Shinoda, 2 Katsuya
GLUCOSE CONCENTRATION INCREASES IGF EXPRESSION FROM SYNOVIAL MEMBRANE
 GLUCOSE CONCENTRATION INCREASES IGF EXPRESSION FROM SYNOVIAL MEMBRANE Final Report Aug 17 2009 Darryl D'Lima, MD, PhD Shiley Center for Orthopaedic Research and Education at Scripps Clinic La Jolla, California
GLUCOSE CONCENTRATION INCREASES IGF EXPRESSION FROM SYNOVIAL MEMBRANE Final Report Aug 17 2009 Darryl D'Lima, MD, PhD Shiley Center for Orthopaedic Research and Education at Scripps Clinic La Jolla, California
Implications of mitochondrial skeletal muscle metabolism on diabetes and obesity before and after weight loss
 GG2 Implications of mitochondrial skeletal muscle metabolism on diabetes and obesity before and after weight loss Dr Giacomo Gastaldi CHRU Montpellier Folie 1 GG2 19.10.2009 GG_PC; 12.10.2009 Plan Introduction
GG2 Implications of mitochondrial skeletal muscle metabolism on diabetes and obesity before and after weight loss Dr Giacomo Gastaldi CHRU Montpellier Folie 1 GG2 19.10.2009 GG_PC; 12.10.2009 Plan Introduction
In The Name Of God. In The Name Of. EMRI Modeling Group
 In The Name Of God In The Name Of God EMRI Modeling Group Cells work together in functionally related groups called tissues Types of tissues: Epithelial lining and covering Connective support Muscle movement
In The Name Of God In The Name Of God EMRI Modeling Group Cells work together in functionally related groups called tissues Types of tissues: Epithelial lining and covering Connective support Muscle movement
control kda ATGL ATGLi HSL 82 GAPDH * ** *** WT/cTg WT/cTg ATGLi AKO/cTg AKO/cTg ATGLi WT/cTg WT/cTg ATGLi AKO/cTg AKO/cTg ATGLi iwat gwat ibat
 body weight (g) tissue weights (mg) ATGL protein expression (relative to GAPDH) HSL protein expression (relative to GAPDH) ### # # kda ATGL 55 HSL 82 GAPDH 37 2.5 2. 1.5 1..5 2. 1.5 1..5.. Supplementary
body weight (g) tissue weights (mg) ATGL protein expression (relative to GAPDH) HSL protein expression (relative to GAPDH) ### # # kda ATGL 55 HSL 82 GAPDH 37 2.5 2. 1.5 1..5 2. 1.5 1..5.. Supplementary
Mamofillin New aesthetic perspective
 New aesthetic perspective info@ White adipose tissue (WAT) White adipose tissue (WAT) is the prevalent type in human adults functioning as the major storage site for the lipids absorbed from daily intake
New aesthetic perspective info@ White adipose tissue (WAT) White adipose tissue (WAT) is the prevalent type in human adults functioning as the major storage site for the lipids absorbed from daily intake
A Tryptophan Hydroxylase Inhibitor Decreases Hepatic FGF21 Expression and Circulating FGF21 in Mice Fed A High-Fat Diet
 A Tryptophan Hydroxylase Inhibitor Decreases Hepatic FGF21 Expression Circulating FGF21 in Mice Fed A High-Fat Diet Katsunori Nonogaki, Takao Kaji, Mari Murakami Abstract Background: Fibroblast growth
A Tryptophan Hydroxylase Inhibitor Decreases Hepatic FGF21 Expression Circulating FGF21 in Mice Fed A High-Fat Diet Katsunori Nonogaki, Takao Kaji, Mari Murakami Abstract Background: Fibroblast growth
Supplemental Information. Intermittent Fasting Promotes. White Adipose Browning and Decreases Obesity. by Shaping the Gut Microbiota
 Cell Metabolism, Volume 26 Supplemental Information Intermittent Fasting Promotes White Adipose Browning and Decreases Obesity by Shaping the Gut Microbiota Guolin Li, Cen Xie, Siyu Lu, Robert G. Nichols,
Cell Metabolism, Volume 26 Supplemental Information Intermittent Fasting Promotes White Adipose Browning and Decreases Obesity by Shaping the Gut Microbiota Guolin Li, Cen Xie, Siyu Lu, Robert G. Nichols,
Supplementary Information
 Supplementary Information GADD34-deficient mice develop obesity, nonalcoholic fatty liver disease, hepatic carcinoma and insulin resistance Naomi Nishio and Ken-ichi Isobe Department of Immunology, Nagoya
Supplementary Information GADD34-deficient mice develop obesity, nonalcoholic fatty liver disease, hepatic carcinoma and insulin resistance Naomi Nishio and Ken-ichi Isobe Department of Immunology, Nagoya
Supplementary Figure 1. DJ-1 modulates ROS concentration in mouse skeletal muscle.
 Supplementary Figure 1. DJ-1 modulates ROS concentration in mouse skeletal muscle. (a) mrna levels of Dj1 measured by quantitative RT-PCR in soleus, gastrocnemius (Gastroc.) and extensor digitorum longus
Supplementary Figure 1. DJ-1 modulates ROS concentration in mouse skeletal muscle. (a) mrna levels of Dj1 measured by quantitative RT-PCR in soleus, gastrocnemius (Gastroc.) and extensor digitorum longus
Pelagia Research Library
 Available online at www.pelagiaresearchlibrary.com Der Pharmacia Sinica, 2014, 5(3): 32-36 ISSN: 0976-8688 CODEN (USA): PSHIBD Study of the effect of ethanolic extract of Solanum xanthocarpum schrad &
Available online at www.pelagiaresearchlibrary.com Der Pharmacia Sinica, 2014, 5(3): 32-36 ISSN: 0976-8688 CODEN (USA): PSHIBD Study of the effect of ethanolic extract of Solanum xanthocarpum schrad &
Fig. S1. Dose-response effects of acute administration of the β3 adrenoceptor agonists CL316243, BRL37344, ICI215,001, ZD7114, ZD2079 and CGP12177 at
 Fig. S1. Dose-response effects of acute administration of the β3 adrenoceptor agonists CL316243, BRL37344, ICI215,001, ZD7114, ZD2079 and CGP12177 at doses of 0.1, 0.5 and 1 mg/kg on cumulative food intake
Fig. S1. Dose-response effects of acute administration of the β3 adrenoceptor agonists CL316243, BRL37344, ICI215,001, ZD7114, ZD2079 and CGP12177 at doses of 0.1, 0.5 and 1 mg/kg on cumulative food intake
Supplemental Information. Increased 4E-BP1 Expression Protects. against Diet-Induced Obesity and Insulin. Resistance in Male Mice
 Cell Reports, Volume 16 Supplemental Information Increased 4E-BP1 Expression Protects against Diet-Induced Obesity and Insulin Resistance in Male Mice Shih-Yin Tsai, Ariana A. Rodriguez, Somasish G. Dastidar,
Cell Reports, Volume 16 Supplemental Information Increased 4E-BP1 Expression Protects against Diet-Induced Obesity and Insulin Resistance in Male Mice Shih-Yin Tsai, Ariana A. Rodriguez, Somasish G. Dastidar,
Hormonal regulation of. Physiology Department Medical School, University of Sumatera Utara
 Hormonal regulation of nutrient metabolism Physiology Department Medical School, University of Sumatera Utara Homeostasis & Controls Successful compensation Homeostasis reestablished Failure to compensate
Hormonal regulation of nutrient metabolism Physiology Department Medical School, University of Sumatera Utara Homeostasis & Controls Successful compensation Homeostasis reestablished Failure to compensate
SUPPLEMENTARY DATA. Supplementary Table 1. Primers used in qpcr
 Supplementary Table 1. Primers used in qpcr Gene forward primer (5'-3') reverse primer (5'-3') β-actin AGAGGGAAATCGTGCGTGAC CAATAGTGATGACCTGGCCGT Hif-p4h-2 CTGGGCAACTACAGGATAAAC GCGTCCCAGTCTTTATTTAGATA
Supplementary Table 1. Primers used in qpcr Gene forward primer (5'-3') reverse primer (5'-3') β-actin AGAGGGAAATCGTGCGTGAC CAATAGTGATGACCTGGCCGT Hif-p4h-2 CTGGGCAACTACAGGATAAAC GCGTCCCAGTCTTTATTTAGATA
Metabolic Solutions Development Company, Kalamazoo, USA.
 New Insulin Sensitizers Produce Differentiation of Brown-like Adipose Cells from a Subcutaneous Fat Depot and Increase Secretion of Adiponectin in vitro William G. McDonald, Serena L. Cole, Danielle D.
New Insulin Sensitizers Produce Differentiation of Brown-like Adipose Cells from a Subcutaneous Fat Depot and Increase Secretion of Adiponectin in vitro William G. McDonald, Serena L. Cole, Danielle D.
ENHANCEMENT OF BROWN ADIPOSE TISSUE DEVELOPMENT IN VIVO BY A NOVEL INSULIN SENSITIZER
 ENHANCEMENT F BRWN ADIPSE TISSUE DEVELPMENT IN VIV BY A NVEL INSULIN SENSITIZER William G. McDonald 1, Serena L. Cole 1, Brian N. Finck 2, Danielle D. Holewa 1, Angela S. Brightwell-Conrad 1, Charles Mackenzie
ENHANCEMENT F BRWN ADIPSE TISSUE DEVELPMENT IN VIV BY A NVEL INSULIN SENSITIZER William G. McDonald 1, Serena L. Cole 1, Brian N. Finck 2, Danielle D. Holewa 1, Angela S. Brightwell-Conrad 1, Charles Mackenzie
SUPPLEMENTARY INFORMATION
 -. -. SUPPLEMENTARY INFORMATION DOI: 1.1/ncb86 a WAT-1 WAT- BAT-1 BAT- sk-muscle-1 sk-muscle- mir-133b mir-133a mir-6 mir-378 mir-1 mir-85 mir-378 mir-6a mir-18 mir-133a mir- mir- mir-341 mir-196a mir-17
-. -. SUPPLEMENTARY INFORMATION DOI: 1.1/ncb86 a WAT-1 WAT- BAT-1 BAT- sk-muscle-1 sk-muscle- mir-133b mir-133a mir-6 mir-378 mir-1 mir-85 mir-378 mir-6a mir-18 mir-133a mir- mir- mir-341 mir-196a mir-17
MBB317. Dr D MANGNALL OBESITY. Lecture 2
 MBB317 Dr D MANGNALL OBESITY Lecture 2 When the structure of the insulin receptor was first discovered it was assumed that the active beta subunit tyrosine kinase would phosphorylate some intracellular
MBB317 Dr D MANGNALL OBESITY Lecture 2 When the structure of the insulin receptor was first discovered it was assumed that the active beta subunit tyrosine kinase would phosphorylate some intracellular
Non-shivering thermogenesis is found in tissues other than brown adipose tissue during cold exposure. By: Jessica Chan & Dayna Weststeyn
 Non-shivering thermogenesis is found in tissues other than brown adipose tissue during cold exposure By: Jessica Chan & Dayna Weststeyn Hypothesis The hypothesis for this point presentation is that non-shivering
Non-shivering thermogenesis is found in tissues other than brown adipose tissue during cold exposure By: Jessica Chan & Dayna Weststeyn Hypothesis The hypothesis for this point presentation is that non-shivering
Human Leptin ELISA Kit
 Product Manual Human Leptin ELISA Kit Catalog Numbers MET-5057 MET-5057-5 96 assays 5 x 96 assays FOR RESEARCH USE ONLY Not for use in diagnostic procedures Introduction Leptin is a polypeptide hormone
Product Manual Human Leptin ELISA Kit Catalog Numbers MET-5057 MET-5057-5 96 assays 5 x 96 assays FOR RESEARCH USE ONLY Not for use in diagnostic procedures Introduction Leptin is a polypeptide hormone
Control 7 d cold 7 d CL
 Control 7 d cold 7 d ibat iwat gwat Supplementary Figure 1. Histology of adipose tissues after cold or 3-adrenergic receptor stimulation. C57BL/6J wild-type mice were housed at 4 C or injected daily with
Control 7 d cold 7 d ibat iwat gwat Supplementary Figure 1. Histology of adipose tissues after cold or 3-adrenergic receptor stimulation. C57BL/6J wild-type mice were housed at 4 C or injected daily with
Pathogenesis of Diabetes Mellitus
 Pathogenesis of Diabetes Mellitus Young-Bum Kim, Ph.D. Associate Professor of Medicine Harvard Medical School Definition of Diabetes Mellitus a group of metabolic diseases characterized by hyperglycemia
Pathogenesis of Diabetes Mellitus Young-Bum Kim, Ph.D. Associate Professor of Medicine Harvard Medical School Definition of Diabetes Mellitus a group of metabolic diseases characterized by hyperglycemia
Overall Energy metabolism: Integration and Regulation
 Overall Energy metabolism: Integration and Regulation We have discussed various fuels which are oxidized via different catabolic pathways to generate ATP, or reducing equivalents required to carry out
Overall Energy metabolism: Integration and Regulation We have discussed various fuels which are oxidized via different catabolic pathways to generate ATP, or reducing equivalents required to carry out
Supplementary Table 1. Primer Sequences Used for Quantitative Real-Time PCR
 Supplementary Table 1. Primer Sequences Used for Quantitative Real-Time PCR Gene Forward Primer (5-3 ) Reverse Primer (5-3 ) cadl CTTGGGGGCGCGTCT CTGTTCTTTTGTGCCGTTTCG cyl-coenzyme Dehydrogenase, very
Supplementary Table 1. Primer Sequences Used for Quantitative Real-Time PCR Gene Forward Primer (5-3 ) Reverse Primer (5-3 ) cadl CTTGGGGGCGCGTCT CTGTTCTTTTGTGCCGTTTCG cyl-coenzyme Dehydrogenase, very
3-Thia Fatty Acids A New Generation of Functional Lipids?
 Conference on Food Structure and Food Quality 3-Thia Fatty Acids A New Generation of Functional Lipids? Rolf K. Berge rolf.berge@med.uib.no Fatty acids- Essential cellular metabolites Concentrations must
Conference on Food Structure and Food Quality 3-Thia Fatty Acids A New Generation of Functional Lipids? Rolf K. Berge rolf.berge@med.uib.no Fatty acids- Essential cellular metabolites Concentrations must
Supporting Information Table of content
 Supporting Information Table of content Supporting Information Fig. S1 Supporting Information Fig. S2 Supporting Information Fig. S3 Supporting Information Fig. S4 Supporting Information Fig. S5 Supporting
Supporting Information Table of content Supporting Information Fig. S1 Supporting Information Fig. S2 Supporting Information Fig. S3 Supporting Information Fig. S4 Supporting Information Fig. S5 Supporting
Naohito AOKI, Erina ARAKAWA and Miyuki ITO. Department of Life Science, Graduate School of Bioresources, Mie University Tsu ABSTRACT
 Naohito AOKI, Erina ARAKAWA and Miyuki ITO Department of Life Science, Graduate School of Bioresources, Mie University Tsu 514-857 ABSTRACT C57BL/6J mice (male, 4wk old) were fed low fat diet (LF), high
Naohito AOKI, Erina ARAKAWA and Miyuki ITO Department of Life Science, Graduate School of Bioresources, Mie University Tsu 514-857 ABSTRACT C57BL/6J mice (male, 4wk old) were fed low fat diet (LF), high
Therapeutic strategy to reduce Glucagon secretion
 Clinical focus on glucagon: α-cell as a companion of β-cell Therapeutic strategy to reduce Glucagon secretion Sunghwan Suh Dong-A University Conflict of interest disclosure None Committee of Scientific
Clinical focus on glucagon: α-cell as a companion of β-cell Therapeutic strategy to reduce Glucagon secretion Sunghwan Suh Dong-A University Conflict of interest disclosure None Committee of Scientific
Supplemental Information Supplementary Table 1. Tph1+/+ Tph1 / Analyte Supplementary Table 2. Tissue Vehicle LP value
 Supplemental Information Supplementary Table. Urinary and adipose tissue catecholamines in Tph +/+ and Tph / mice fed a high fat diet for weeks. Tph +/+ Tph / Analyte ewat ibat ewat ibat Urine (ng/ml)
Supplemental Information Supplementary Table. Urinary and adipose tissue catecholamines in Tph +/+ and Tph / mice fed a high fat diet for weeks. Tph +/+ Tph / Analyte ewat ibat ewat ibat Urine (ng/ml)
THE ENDOCANNABINOID SYSTEM IN ADIPOSE TISSUE. Key Points
 December 2008 (Vol. 1, Issue 3, pages 31-35) THE ENDOCANNABINOID SYSTEM IN ADIPOSE TISSUE By Uberto Pagotto, MD, PhD Endocrinology Unit and Center of Applied Biomedical Research (C.R.B.A.), Department
December 2008 (Vol. 1, Issue 3, pages 31-35) THE ENDOCANNABINOID SYSTEM IN ADIPOSE TISSUE By Uberto Pagotto, MD, PhD Endocrinology Unit and Center of Applied Biomedical Research (C.R.B.A.), Department
Analysis of AVP functions via V1a and V1b receptors with knockout mice. Akito Tanoue
 Analysis of AVP functions via V1a and V1b receptors with knockout mice Akito Tanoue Department of Pharmacology, National Research Institute for Child Health and Development Arginine-Vasopressin (AVP) is
Analysis of AVP functions via V1a and V1b receptors with knockout mice Akito Tanoue Department of Pharmacology, National Research Institute for Child Health and Development Arginine-Vasopressin (AVP) is
General Laboratory methods Plasma analysis: Gene Expression Analysis: Immunoblot analysis: Immunohistochemistry:
 General Laboratory methods Plasma analysis: Plasma insulin (Mercodia, Sweden), leptin (duoset, R&D Systems Europe, Abingdon, United Kingdom), IL-6, TNFα and adiponectin levels (Quantikine kits, R&D Systems
General Laboratory methods Plasma analysis: Plasma insulin (Mercodia, Sweden), leptin (duoset, R&D Systems Europe, Abingdon, United Kingdom), IL-6, TNFα and adiponectin levels (Quantikine kits, R&D Systems
Supplementary Figure S1. Effect of Glucose on Energy Balance in WT and KHK A/C KO
 Supplementary Figure S1. Effect of Glucose on Energy Balance in WT and KHK A/C KO Mice. WT mice and KHK-A/C KO mice were provided drinking water containing 10% glucose or tap water with normal chow ad
Supplementary Figure S1. Effect of Glucose on Energy Balance in WT and KHK A/C KO Mice. WT mice and KHK-A/C KO mice were provided drinking water containing 10% glucose or tap water with normal chow ad
Hormones. Prof. Dr. Volker Haucke Institut für Chemie-Biochemie Takustrasse 6
 Hormones Prof. Dr. Volker Haucke Institut für Chemie-Biochemie Takustrasse 6 Tel. 030-8385-6920 (Sekret.) 030-8385-6922 (direkt) e-mail: vhaucke@chemie.fu-berlin.de http://userpage.chemie.fu-berlin.de/biochemie/aghaucke/teaching.html
Hormones Prof. Dr. Volker Haucke Institut für Chemie-Biochemie Takustrasse 6 Tel. 030-8385-6920 (Sekret.) 030-8385-6922 (direkt) e-mail: vhaucke@chemie.fu-berlin.de http://userpage.chemie.fu-berlin.de/biochemie/aghaucke/teaching.html
Metabolic Syndrome. DOPE amines COGS 163
 Metabolic Syndrome DOPE amines COGS 163 Overview - M etabolic Syndrome - General definition and criteria - Importance of diagnosis - Glucose Homeostasis - Type 2 Diabetes Mellitus - Insulin Resistance
Metabolic Syndrome DOPE amines COGS 163 Overview - M etabolic Syndrome - General definition and criteria - Importance of diagnosis - Glucose Homeostasis - Type 2 Diabetes Mellitus - Insulin Resistance
Supporting Information
 Supporting Information Charalambous et al. 10.1073/pnas.1406119111 SI Experimental Procedures Serum and Tissue Biochemistry. Enzymatic assay kits were used for determination of plasma FFAs (Roche), TAGs
Supporting Information Charalambous et al. 10.1073/pnas.1406119111 SI Experimental Procedures Serum and Tissue Biochemistry. Enzymatic assay kits were used for determination of plasma FFAs (Roche), TAGs
A microrna-34a/fgf21 Regulatory Axis and Browning of White Fat
 A microrna-34a/fgf21 Regulatory Axis and Browning of White Fat Jongsook Kim Kemper, Ph.D Department of Molecular and Integrative Physiology, University of Illinois at Urbana-Champaign, USA 213 International
A microrna-34a/fgf21 Regulatory Axis and Browning of White Fat Jongsook Kim Kemper, Ph.D Department of Molecular and Integrative Physiology, University of Illinois at Urbana-Champaign, USA 213 International
Online Data Supplement. Anti-aging Gene Klotho Enhances Glucose-induced Insulin Secretion by Upregulating Plasma Membrane Retention of TRPV2
 Online Data Supplement Anti-aging Gene Klotho Enhances Glucose-induced Insulin Secretion by Upregulating Plasma Membrane Retention of TRPV2 Yi Lin and Zhongjie Sun Department of physiology, college of
Online Data Supplement Anti-aging Gene Klotho Enhances Glucose-induced Insulin Secretion by Upregulating Plasma Membrane Retention of TRPV2 Yi Lin and Zhongjie Sun Department of physiology, college of
Method of leptin dosing, strain, and group housing influence leptin sensitivity in high-fat-fed weanling mice
 Am J Physiol Regul Integr Comp Physiol 284: R87 R100, 2003; 10.1152/ajpregu.00431.2002. Method of leptin dosing, strain, and group housing influence leptin sensitivity in high-fat-fed weanling mice HEATHER
Am J Physiol Regul Integr Comp Physiol 284: R87 R100, 2003; 10.1152/ajpregu.00431.2002. Method of leptin dosing, strain, and group housing influence leptin sensitivity in high-fat-fed weanling mice HEATHER
Males- Western Diet WT KO Age (wks) Females- Western Diet WT KO Age (wks)
 Females- Western Diet WT KO Age (wks)") Relative Arv1 mrna Adrenal 33.48 +/- 6.2 Skeletal Muscle 22.4 +/- 4.93 Liver 6.41 +/- 1.48 Heart 5.1 +/- 2.3 Brain 4.98 +/- 2.11 Ovary 4.68 +/- 2.21 Kidney 3.98 +/-.39 Lung 2.15 +/-.6 Inguinal Subcutaneous
Relative Arv1 mrna Adrenal 33.48 +/- 6.2 Skeletal Muscle 22.4 +/- 4.93 Liver 6.41 +/- 1.48 Heart 5.1 +/- 2.3 Brain 4.98 +/- 2.11 Ovary 4.68 +/- 2.21 Kidney 3.98 +/-.39 Lung 2.15 +/-.6 Inguinal Subcutaneous
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION doi:10.1038/nature11464 Supplemental Figure S1. The expression of Vegfb is increased in obese and diabetic mice as compared to lean mice. a-b, Body weight and postprandial blood
SUPPLEMENTARY INFORMATION doi:10.1038/nature11464 Supplemental Figure S1. The expression of Vegfb is increased in obese and diabetic mice as compared to lean mice. a-b, Body weight and postprandial blood
BIOL212 Biochemistry of Disease. Metabolic Disorders - Obesity
 BIOL212 Biochemistry of Disease Metabolic Disorders - Obesity Obesity Approx. 23% of adults are obese in the U.K. The number of obese children has tripled in 20 years. 10% of six year olds are obese, rising
BIOL212 Biochemistry of Disease Metabolic Disorders - Obesity Obesity Approx. 23% of adults are obese in the U.K. The number of obese children has tripled in 20 years. 10% of six year olds are obese, rising
Supplementary Information. Glycogen shortage during fasting triggers liver-brain-adipose. neurocircuitry to facilitate fat utilization
 Supplementary Information Glycogen shortage during fasting triggers liver-brain-adipose neurocircuitry to facilitate fat utilization Supplementary Figure S1. Liver-Brain-Adipose neurocircuitry Starvation
Supplementary Information Glycogen shortage during fasting triggers liver-brain-adipose neurocircuitry to facilitate fat utilization Supplementary Figure S1. Liver-Brain-Adipose neurocircuitry Starvation
Obesity in aging: Hormonal contribution
 Obesity in aging: Hormonal contribution Hormonal issues in obesity and aging Hormonal role in regulation of energy balance Genetic component in hormonal regulation Life style contribution to hormonal changes
Obesity in aging: Hormonal contribution Hormonal issues in obesity and aging Hormonal role in regulation of energy balance Genetic component in hormonal regulation Life style contribution to hormonal changes
Supplemental Fig. 1. Relative mrna Expression. Relative mrna Expression WT KO WT KO RT 4 0 C
 Supplemental Fig. 1 A 1.5 1..5 Hdac11 (ibat) n=4 n=4 n=4 n=4 n=4 n=4 n=4 n=4 WT KO WT KO WT KO WT KO RT 4 C RT 4 C Supplemental Figure 1. Hdac11 mrna is undetectable in KO adipose tissue. Quantitative
Supplemental Fig. 1 A 1.5 1..5 Hdac11 (ibat) n=4 n=4 n=4 n=4 n=4 n=4 n=4 n=4 WT KO WT KO WT KO WT KO RT 4 C RT 4 C Supplemental Figure 1. Hdac11 mrna is undetectable in KO adipose tissue. Quantitative
Understanding the Physiology of FGF21
 ANNUAL REVIEWS Further Click here to view this article's online features: Download figures as PPT slides Navigate linked references Download citations Explore related articles Search keywords Annu. Rev.
ANNUAL REVIEWS Further Click here to view this article's online features: Download figures as PPT slides Navigate linked references Download citations Explore related articles Search keywords Annu. Rev.
Identification of Binding Partners of Fibroblast Gro wth Factor221 in Cell Membrane of 3T32L1 Cells
 ISSN 100727626 CN 1123870ΠQ Chinese Journal of Biochemistry and Molecular Biology 2008 7 24 (7) :667 673 3T32L1 FGF221 1), 1),, 3 (, 150030) (fibroblast growth factor,fgf)221 1, 2.,FGF221.,, 3T32L1, FGF221,
ISSN 100727626 CN 1123870ΠQ Chinese Journal of Biochemistry and Molecular Biology 2008 7 24 (7) :667 673 3T32L1 FGF221 1), 1),, 3 (, 150030) (fibroblast growth factor,fgf)221 1, 2.,FGF221.,, 3T32L1, FGF221,
University of Groningen. Adipose tissue Nies, Vera
 University of Groningen Adipose tissue Nies, Vera IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite from it. Please check the document version below.
University of Groningen Adipose tissue Nies, Vera IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite from it. Please check the document version below.
Leptin Intro/Signaling. ATeamP: Angelo, Anthony, Charlie, Gabby, Joseph
 Leptin Intro/Signaling ATeamP: Angelo, Anthony, Charlie, Gabby, Joseph Overview Intro to Leptin Definition & Sources Physiology Bound vs. Free Receptors Signaling JAK/STAT MAPK PI3K ACC Experimental findings
Leptin Intro/Signaling ATeamP: Angelo, Anthony, Charlie, Gabby, Joseph Overview Intro to Leptin Definition & Sources Physiology Bound vs. Free Receptors Signaling JAK/STAT MAPK PI3K ACC Experimental findings
HORMONES AND CELL SIGNALLING
 HORMONES AND CELL SIGNALLING TYPES OF CELL JUNCTIONS CHEMICAL SIGNALS AND MODES OF ACTION Endocrine system produces chemical messages = hormones that are transported from endocrine gland to target cell
HORMONES AND CELL SIGNALLING TYPES OF CELL JUNCTIONS CHEMICAL SIGNALS AND MODES OF ACTION Endocrine system produces chemical messages = hormones that are transported from endocrine gland to target cell
HIV VPR alters fat metabolism. Dorothy E Lewis PhD/Ashok Balasubramanyam MD
 HIV VPR alters fat metabolism Dorothy E Lewis PhD/Ashok Balasubramanyam MD Old Dogma for HIV associated lipodystrophy Differentiation Block (PI) Lipoatrophy Apoptosis (NRTI) Stem cell Preadipocyte Adipocyte
HIV VPR alters fat metabolism Dorothy E Lewis PhD/Ashok Balasubramanyam MD Old Dogma for HIV associated lipodystrophy Differentiation Block (PI) Lipoatrophy Apoptosis (NRTI) Stem cell Preadipocyte Adipocyte
Defective Hepatic Autophagy in Obesity Promotes ER Stress and Causes Insulin Resistance
 Cell Metabolism, Volume 11 Supplemental Information Defective Hepatic Autophagy in Obesity Promotes ER Stress and Causes Insulin Resistance Ling Yang, Ping Li, Suneng Fu, Ediz S. Calay, and Gökhan S. Hotamisligil
Cell Metabolism, Volume 11 Supplemental Information Defective Hepatic Autophagy in Obesity Promotes ER Stress and Causes Insulin Resistance Ling Yang, Ping Li, Suneng Fu, Ediz S. Calay, and Gökhan S. Hotamisligil
THE GLUCOSE-FATTY ACID-KETONE BODY CYCLE Role of ketone bodies as respiratory substrates and metabolic signals
 Br. J. Anaesth. (1981), 53, 131 THE GLUCOSE-FATTY ACID-KETONE BODY CYCLE Role of ketone bodies as respiratory substrates and metabolic signals J. C. STANLEY In this paper, the glucose-fatty acid cycle
Br. J. Anaesth. (1981), 53, 131 THE GLUCOSE-FATTY ACID-KETONE BODY CYCLE Role of ketone bodies as respiratory substrates and metabolic signals J. C. STANLEY In this paper, the glucose-fatty acid cycle
Central injection of fibroblast growth factor 1 induces sustained remission of diabetic hyperglycemia in rodents
 Central injection of fibroblast growth factor 1 induces sustained remission of diabetic hyperglycemia in rodents Jarrad M Scarlett 1,,1, Jennifer M Rojas 1,1, Miles E Matsen 1, Karl J Kaiyala 3, Darko
Central injection of fibroblast growth factor 1 induces sustained remission of diabetic hyperglycemia in rodents Jarrad M Scarlett 1,,1, Jennifer M Rojas 1,1, Miles E Matsen 1, Karl J Kaiyala 3, Darko
Supplementary Figure 1.
 Supplementary Figure 1. FGF21 does not exert direct effects on hepatic glucose production. The liver explants from C57BL/6J mice (A, B) or primary rat hepatocytes (C, D) were incubated with rmfgf21 (2
Supplementary Figure 1. FGF21 does not exert direct effects on hepatic glucose production. The liver explants from C57BL/6J mice (A, B) or primary rat hepatocytes (C, D) were incubated with rmfgf21 (2
18. PANCREATIC FUNCTION AND METABOLISM. Pancreatic secretions ISLETS OF LANGERHANS. Insulin
 18. PANCREATIC FUNCTION AND METABOLISM ISLETS OF LANGERHANS Some pancreatic functions have already been discussed in the digestion section. In this one, the emphasis will be placed on the endocrine function
18. PANCREATIC FUNCTION AND METABOLISM ISLETS OF LANGERHANS Some pancreatic functions have already been discussed in the digestion section. In this one, the emphasis will be placed on the endocrine function
Crosstalk between Adiponectin and IGF-IR in breast cancer. Prof. Young Jin Suh Department of Surgery The Catholic University of Korea
 Crosstalk between Adiponectin and IGF-IR in breast cancer Prof. Young Jin Suh Department of Surgery The Catholic University of Korea Obesity Chronic, multifactorial disorder Hypertrophy and hyperplasia
Crosstalk between Adiponectin and IGF-IR in breast cancer Prof. Young Jin Suh Department of Surgery The Catholic University of Korea Obesity Chronic, multifactorial disorder Hypertrophy and hyperplasia
Beige fat: A New Hope for Metabolic Disorder
 Young Biomedical Scientists Forum(YBSF) Beige fat: A New Hope for Metabolic Disorder Harim Kim(2) 1 Ewha University, School of Medicine, 52, Ewhayeodae-gil, Seodaemun-gu, Seoul 120-750 Korea k3541729@ybsf21.org
Young Biomedical Scientists Forum(YBSF) Beige fat: A New Hope for Metabolic Disorder Harim Kim(2) 1 Ewha University, School of Medicine, 52, Ewhayeodae-gil, Seodaemun-gu, Seoul 120-750 Korea k3541729@ybsf21.org
UNIVERSITY OF PNG SCHOOL OF MEDICINE AND HEALTH SCIENCES DIVISION OF BASIC MEDICAL SCIENCES DISCIPLINE OF BIOCHEMISTRY AND MOLECULAR BIOLOGY
 1 UNIVERSITY OF PNG SCHOOL OF MEDICINE AND HEALTH SCIENCES DIVISION OF BASIC MEDICAL SCIENCES DISCIPLINE OF BIOCHEMISTRY AND MOLECULAR BIOLOGY GLUCOSE HOMEOSTASIS An Overview WHAT IS HOMEOSTASIS? Homeostasis
1 UNIVERSITY OF PNG SCHOOL OF MEDICINE AND HEALTH SCIENCES DIVISION OF BASIC MEDICAL SCIENCES DISCIPLINE OF BIOCHEMISTRY AND MOLECULAR BIOLOGY GLUCOSE HOMEOSTASIS An Overview WHAT IS HOMEOSTASIS? Homeostasis
Fat and HIV persistence, role of fat metabolism and inflammation
 NIDDK Workshop Obesity and Fat Metabolism in HIV infected individuals Rockville, MD May 22-23, 2018 Centro de Investigación Biomédica En Red Fisiopatología de la Obesidad y Nutrición Fat and HIV persistence,
NIDDK Workshop Obesity and Fat Metabolism in HIV infected individuals Rockville, MD May 22-23, 2018 Centro de Investigación Biomédica En Red Fisiopatología de la Obesidad y Nutrición Fat and HIV persistence,
Cells and reagents. Synaptopodin knockdown (1) and dynamin knockdown (2)
 and dynamin knockdown (2)") Supplemental Methods Cells and reagents. Synaptopodin knockdown (1) and dynamin knockdown (2) podocytes were cultured as described previously. Staurosporine, angiotensin II and actinomycin D were all obtained
Supplemental Methods Cells and reagents. Synaptopodin knockdown (1) and dynamin knockdown (2) podocytes were cultured as described previously. Staurosporine, angiotensin II and actinomycin D were all obtained
Yiying Zhang, PhD Research Scientist. Research Summary:
 Yiying Zhang, PhD Research Scientist Research Summary: Address: Naomi Berrie Diabetes Center at Columbia University Medical Center Russ Berrie Medical Science Pavilion 1150 St. Nicholas Avenue New York,
Yiying Zhang, PhD Research Scientist Research Summary: Address: Naomi Berrie Diabetes Center at Columbia University Medical Center Russ Berrie Medical Science Pavilion 1150 St. Nicholas Avenue New York,
For pair feeding, mice were fed 2.7g of HFD containing tofogliflozin
 Materials and Methods Pair Feeding Experiment For pair feeding, mice were fed 2.7g of HFD containing tofogliflozin (0.005%), which is average daily food intake of mice fed control HFD ad libitum at week
Materials and Methods Pair Feeding Experiment For pair feeding, mice were fed 2.7g of HFD containing tofogliflozin (0.005%), which is average daily food intake of mice fed control HFD ad libitum at week
WY14643 combined with all-trans retinoic acid acts via p38 MAPK to induce browning of white adipocytes in mice
 WY14643 combined with all-trans retinoic acid acts via p38 MAPK to induce browning of white adipocytes in mice J. Wang 1, G.J. Sun 1, J. Ding 2, J.X. Zhang 3, Y. Cui 1, H.R. Li 1 and S.J. Wang 1 1 Department
WY14643 combined with all-trans retinoic acid acts via p38 MAPK to induce browning of white adipocytes in mice J. Wang 1, G.J. Sun 1, J. Ding 2, J.X. Zhang 3, Y. Cui 1, H.R. Li 1 and S.J. Wang 1 1 Department
Intracellular signalling pathways activated by leptin by Gema FRUHBECK. Presentation by Amnesiacs Anonymous
 Intracellular signalling pathways activated by leptin by Gema FRUHBECK Presentation by Amnesiacs Anonymous Introduction to Leptin By Ahrial Young Why is Leptin important? Pleiotropic = it controls the
Intracellular signalling pathways activated by leptin by Gema FRUHBECK Presentation by Amnesiacs Anonymous Introduction to Leptin By Ahrial Young Why is Leptin important? Pleiotropic = it controls the
PXL770, a novel direct AMPK activator, improves metabolic disorders in a diet-induced mice model of obesity and diabetes
 PXL770, a novel direct AMPK activator, improves metabolic disorders in a diet-induced mice model of obesity and diabetes Sébastien Bolze 1 ; Sophie Hallakou-Bozec 1 ; Michael Roden 2, 3,4 ; Julien Roux
PXL770, a novel direct AMPK activator, improves metabolic disorders in a diet-induced mice model of obesity and diabetes Sébastien Bolze 1 ; Sophie Hallakou-Bozec 1 ; Michael Roden 2, 3,4 ; Julien Roux
MTC-TT and TPC-1 cell lines were cultured in RPMI medium (Gibco, Breda, The Netherlands)
") Supplemental data Materials and Methods Cell culture MTC-TT and TPC-1 cell lines were cultured in RPMI medium (Gibco, Breda, The Netherlands) supplemented with 15% or 10% (for TPC-1) fetal bovine serum
Supplemental data Materials and Methods Cell culture MTC-TT and TPC-1 cell lines were cultured in RPMI medium (Gibco, Breda, The Netherlands) supplemented with 15% or 10% (for TPC-1) fetal bovine serum
Interplay between FGF21 and insulin action in the liver regulates metabolism
 Research article Interplay between FGF21 and insulin action in the liver regulates metabolism Brice Emanuelli, 1 Sara G. Vienberg, 1 Graham Smyth, 1 Christine Cheng, 2 Kristin I. Stanford, 1 Manimozhiyan
Research article Interplay between FGF21 and insulin action in the liver regulates metabolism Brice Emanuelli, 1 Sara G. Vienberg, 1 Graham Smyth, 1 Christine Cheng, 2 Kristin I. Stanford, 1 Manimozhiyan
FGF21 Resistance in Adipose Tissues as a Cause of Insulin Resistance
 Resistance in Adipose Tissues as a Cause of Insulin Resistance The ICDM 213 & 5 th AASD Scientific Meeting Seoul, Korea, Nov 8, 213 Aimin Xu Dept of Medicine & Dept of Pharmacology and Pharmacy The University
Resistance in Adipose Tissues as a Cause of Insulin Resistance The ICDM 213 & 5 th AASD Scientific Meeting Seoul, Korea, Nov 8, 213 Aimin Xu Dept of Medicine & Dept of Pharmacology and Pharmacy The University
Regulation of adipose tissue remodeling by peripheral serotonin
 Regulation of adipose tissue remodeling by peripheral serotonin Sangkyu Park Catholic Kwandong University College of Medicine Department of Biochemistry Serotonin (5-HT) is a signaling molecule Hemostasis
Regulation of adipose tissue remodeling by peripheral serotonin Sangkyu Park Catholic Kwandong University College of Medicine Department of Biochemistry Serotonin (5-HT) is a signaling molecule Hemostasis
Adipose Tissue as an Endocrine Organ. Abdel Moniem Ibrahim, MD Professor of Physiology Cairo University
 Adipose Tissue as an Endocrine Organ Abdel Moniem Ibrahim, MD Professor of Physiology Cairo University Functions of Adipose Tissue Adipose tissue expresses and secretes a variety of bioactive peptides,
Adipose Tissue as an Endocrine Organ Abdel Moniem Ibrahim, MD Professor of Physiology Cairo University Functions of Adipose Tissue Adipose tissue expresses and secretes a variety of bioactive peptides,
Inflammation & Type 2 Diabetes Prof. Marc Y. Donath
 Inflammation & Type 2 Diabetes 1, MD Chief Endocrinology, Diabetes & Metabolism University Hospital Basel Petersgraben 4 CH-431 Basel, Switzerland MDonath@uhbs.ch Innate immunity as a sensor of metabolic
Inflammation & Type 2 Diabetes 1, MD Chief Endocrinology, Diabetes & Metabolism University Hospital Basel Petersgraben 4 CH-431 Basel, Switzerland MDonath@uhbs.ch Innate immunity as a sensor of metabolic
NT-PGC-1α activation attenuates high-fat diet-induced obesity by enhancing brown fat thermogenesis and adipose tissue oxidative metabolism
 Page 1 of 37 NT-PGC-1α activation attenuates high-fat diet-induced obesity by enhancing brown fat thermogenesis and adipose tissue oxidative metabolism Hee-Jin Jun, Yagini Joshi, Yuvraj Patil, Robert C.
Page 1 of 37 NT-PGC-1α activation attenuates high-fat diet-induced obesity by enhancing brown fat thermogenesis and adipose tissue oxidative metabolism Hee-Jin Jun, Yagini Joshi, Yuvraj Patil, Robert C.
The Epigenetics of Obesity: Individual, Social, and Environmental Influences. K. J. Claycombe, Ph.D.
 The Epigenetics of Obesity: Individual, Social, and Environmental Influences K. J. Claycombe, Ph.D. What can happen to our gene(s) that would cause obesity? Modification via Epigenetic alterations C
The Epigenetics of Obesity: Individual, Social, and Environmental Influences K. J. Claycombe, Ph.D. What can happen to our gene(s) that would cause obesity? Modification via Epigenetic alterations C
People Living with And Inspired by Diabetes
 People Living with And Inspired by Diabetes IMPERFECTLY MOVING TOWARDS & FAINTING GOATS Vol. 2 No. 2 Fall 2016 Vol. 2 No. 2 Fall 2016 1 Review PARTNERING WITH SCIENCE: New Hope for the Effective Treatment
People Living with And Inspired by Diabetes IMPERFECTLY MOVING TOWARDS & FAINTING GOATS Vol. 2 No. 2 Fall 2016 Vol. 2 No. 2 Fall 2016 1 Review PARTNERING WITH SCIENCE: New Hope for the Effective Treatment
SUPPLEMENTARY INFORMATION
 DOI: 10.1038/ncb3461 In the format provided by the authors and unedited. Supplementary Figure 1 (associated to Figure 1). Cpeb4 gene-targeted mice develop liver steatosis. a, Immunoblot displaying CPEB4
DOI: 10.1038/ncb3461 In the format provided by the authors and unedited. Supplementary Figure 1 (associated to Figure 1). Cpeb4 gene-targeted mice develop liver steatosis. a, Immunoblot displaying CPEB4
Effects of growth hormone secretagogue receptor agonist and antagonist in nonobese type 2 diabetic MKR mice
 Effects of growth hormone secretagogue receptor agonist and antagonist in nonobese type 2 diabetic MKR mice Rasha Mosa (MBCHC, M.D, PhD candidate) School of Biomedical Sciences University of Queensland
Effects of growth hormone secretagogue receptor agonist and antagonist in nonobese type 2 diabetic MKR mice Rasha Mosa (MBCHC, M.D, PhD candidate) School of Biomedical Sciences University of Queensland
Low ambient temperature lowers cholecystokinin and leptin plasma concentrations in adult men
 ISPUB.COM The Internet Journal of Gastroenterology Volume 7 Number 2 Low ambient temperature lowers cholecystokinin and leptin plasma concentrations in adult men M Pizon, P Tomasic, K Sztefko, Z Szafran
ISPUB.COM The Internet Journal of Gastroenterology Volume 7 Number 2 Low ambient temperature lowers cholecystokinin and leptin plasma concentrations in adult men M Pizon, P Tomasic, K Sztefko, Z Szafran
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION FOR Liver X Receptor α mediates hepatic triglyceride accumulation through upregulation of G0/G1 Switch Gene 2 (G0S2) expression I: SUPPLEMENTARY METHODS II: SUPPLEMENTARY FIGURES
SUPPLEMENTARY INFORMATION FOR Liver X Receptor α mediates hepatic triglyceride accumulation through upregulation of G0/G1 Switch Gene 2 (G0S2) expression I: SUPPLEMENTARY METHODS II: SUPPLEMENTARY FIGURES
TITLE: Adipose Estrogen and Increased Breast Cancer Risk in Obesity: Regulation by Leptin and Insulin
 AD Award Number: W81XWH-05-1-0497 TITLE: Adipose Estrogen and Increased Breast Cancer Risk in Obesity: Regulation by Leptin and Insulin PRINCIPAL INVESTIGATOR: Fahumiya Samad CONTRACTING ORGANIZATION:
AD Award Number: W81XWH-05-1-0497 TITLE: Adipose Estrogen and Increased Breast Cancer Risk in Obesity: Regulation by Leptin and Insulin PRINCIPAL INVESTIGATOR: Fahumiya Samad CONTRACTING ORGANIZATION:
Mfn2 deletion in brown adipose tissue protects from insulin resistance and impairs thermogenesis
 Article Mfn2 deletion in brown adipose tissue protects from insulin resistance and impairs thermogenesis Kiana Mahdaviani 1,2, Ilan Y Benador 1,2, Shi Su 1, Raffi A Gharakhanian 1, Linsey Stiles 1,2, Kyle
Article Mfn2 deletion in brown adipose tissue protects from insulin resistance and impairs thermogenesis Kiana Mahdaviani 1,2, Ilan Y Benador 1,2, Shi Su 1, Raffi A Gharakhanian 1, Linsey Stiles 1,2, Kyle
AMPK. Tomáš Kučera.
 AMPK (AMP- ACTIVATED PROTEIN KINASE ) Tomáš Kučera tomas.kucera@lfmotol.cuni.cz Department of Medical Chemistry and Clinical Biochemistry 2nd Faculty of Medicine, Charles University in Prague and Motol
AMPK (AMP- ACTIVATED PROTEIN KINASE ) Tomáš Kučera tomas.kucera@lfmotol.cuni.cz Department of Medical Chemistry and Clinical Biochemistry 2nd Faculty of Medicine, Charles University in Prague and Motol
Role of fatty acids in the development of insulin resistance and type 2 diabetes mellitus
 Emerging Science Role of fatty acids in the development of insulin resistance and type 2 diabetes mellitus George Wolf Insulin resistance is defined as the reduced responsiveness to normal circulating
Emerging Science Role of fatty acids in the development of insulin resistance and type 2 diabetes mellitus George Wolf Insulin resistance is defined as the reduced responsiveness to normal circulating
Chapter 41. Lecture 14. Animal Hormones. Dr. Chris Faulkes
 Chapter 41 Lecture 14 Animal Hormones Dr. Chris Faulkes Animal Hormones Aims: To appreciate the variety and roles of hormones in the body To understand the basic types of hormones To understand how hormones
Chapter 41 Lecture 14 Animal Hormones Dr. Chris Faulkes Animal Hormones Aims: To appreciate the variety and roles of hormones in the body To understand the basic types of hormones To understand how hormones
Lack of TRPV2 impairs thermogenesis in mouse brown adipose tissue
 Manuscript EMBO-2015-40819 Lack of TRPV2 impairs thermogenesis in mouse brown adipose tissue Wuping Sun, Kunitoshi Uchida, Yoshiro Suzuki, Yiming Zhou, Minji Kim, Yasunori Takayama, Nobuyuki Takahashi,
Manuscript EMBO-2015-40819 Lack of TRPV2 impairs thermogenesis in mouse brown adipose tissue Wuping Sun, Kunitoshi Uchida, Yoshiro Suzuki, Yiming Zhou, Minji Kim, Yasunori Takayama, Nobuyuki Takahashi,
Integrative Metabolism: Significance
 Integrative Metabolism: Significance Energy Containing Nutrients Carbohydrates Fats Proteins Catabolism Energy Depleted End Products H 2 O NH 3 ADP + Pi NAD + NADP + FAD + Pi NADH+H + NADPH+H + FADH2 Cell
Integrative Metabolism: Significance Energy Containing Nutrients Carbohydrates Fats Proteins Catabolism Energy Depleted End Products H 2 O NH 3 ADP + Pi NAD + NADP + FAD + Pi NADH+H + NADPH+H + FADH2 Cell
Supplementary Materials for
 www.sciencesignaling.org/cgi/content/full/8/407/ra127/dc1 Supplementary Materials for Loss of FTO in adipose tissue decreases Angptl4 translation and alters triglyceride metabolism Chao-Yung Wang,* Shian-Sen
www.sciencesignaling.org/cgi/content/full/8/407/ra127/dc1 Supplementary Materials for Loss of FTO in adipose tissue decreases Angptl4 translation and alters triglyceride metabolism Chao-Yung Wang,* Shian-Sen
Molecular Mechanisms associated with the Cancer-Cachexia Syndrome
 Molecular Mechanisms associated with the Cancer-Cachexia Syndrome Prof. Dr. Josep M. Argilés Department of Biochemistry & Molecular Biology University of Barcelona, Spain Disclosures: DANONE (Scientific
Molecular Mechanisms associated with the Cancer-Cachexia Syndrome Prof. Dr. Josep M. Argilés Department of Biochemistry & Molecular Biology University of Barcelona, Spain Disclosures: DANONE (Scientific
Supplementary Table 2. Plasma lipid profiles in wild type and mutant female mice submitted to a HFD for 12 weeks wt ERα -/- AF-1 0 AF-2 0
 Supplementary Table 1. List of specific primers used for gene expression analysis. Genes Primer forward Primer reverse Hprt GCAGTACAGCCCCAAAATGG AACAAAGTCTGGCCTGTATCCA Srebp-1c GGAAGCTGTCGGGGTAGCGTC CATGTCTTCAAATGTGCAATCCAT
Supplementary Table 1. List of specific primers used for gene expression analysis. Genes Primer forward Primer reverse Hprt GCAGTACAGCCCCAAAATGG AACAAAGTCTGGCCTGTATCCA Srebp-1c GGAAGCTGTCGGGGTAGCGTC CATGTCTTCAAATGTGCAATCCAT
PEPCK. The Regulation of Eukaryotic Gene Expression. Why choose PEPCK? PEPCK. PEPCK overexpression in muscle. The Supermouse.
 PEPK The Regulation of Eukaryotic Gene Expression..using the example of PEPK This is an acronym for an enzyme PhosphoEnol Pyruvate arboxykinase This enzyme is NLY regulated by gene expression! No allosteric
PEPK The Regulation of Eukaryotic Gene Expression..using the example of PEPK This is an acronym for an enzyme PhosphoEnol Pyruvate arboxykinase This enzyme is NLY regulated by gene expression! No allosteric