Modeling the Primary Visual Cortex
|
|
|
- Daisy Sanders
- 6 years ago
- Views:
Transcription
1 Modeling the Primary Visual Cortex David W. McLaughlin Courant Institute & Center for Neural Science New York University Ohio - MBI Oct 02
2 Input Layer of Primary Visual Cortex (V1) for Macaque Monkey Modeled at : Courant Institute of Math. Sciences & Center for Neural Science, NYU In collaboration with: Robert Shapley (Neural Sci) Michael Shelley Louis Tao Jacob Wielaard But today, I ll describe work in collaboration with David Cai David Lorentz Louis Tao
3 Today, I ll Briefly set the background for our modeling study; Jump to a description of some of my current focus; (Bob and Mike will summarize our current results.)
4 We re studying the front end of the visual system -- the primary visual cortex
5 Visual Pathway: Retina --> LGN --> V1 --> Beyond
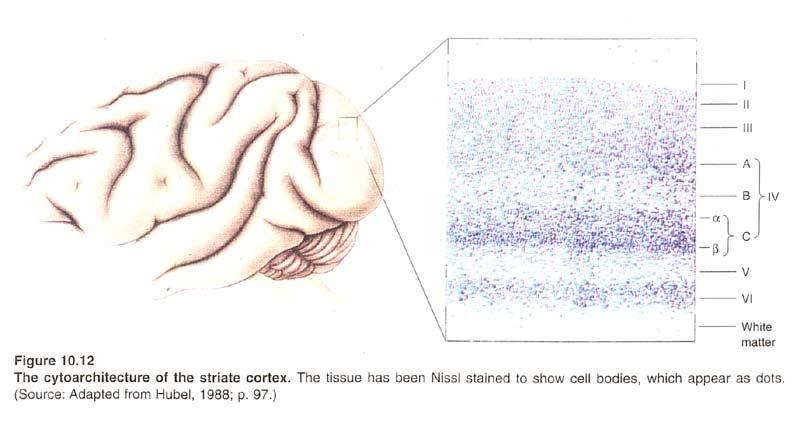
6
7
8 Elementary Feature Detectors Individual neurons in V1 respond preferentially to elementary features of the visual scene (color, direction of motion, speed of motion, spatial wave-length). Two important features: Spatial phase φ (relative to receptive field center) Orientation θ of edges.

9 Grating Stimuli Standing & Drifting Two Angles: Angle of orientation -- θ Angle of spatial phase -- φ (relevant for standing gratings)
10 Orientation Tuning Curves (Firing Rates Vs Angle of Orientation) Spikes/sec Terminology: Orientation Preference Orientation Selectivity Measured by Half-Widths or Peak-to-Trough

11 Cortical Map of Orientation Preference Optical Imaging Blasdel, 1992 Outer layers (2/3) of V µ ---- Color coded for angle of orientation preference right eye left eye
12 Our Model A detailed, fine scale model of a local patch of input layer of Primary Visual Cortex; Realistically constrained by experimental data; Refs: McLaughlin, Shapley, Shelley & Wielaard --- PNAS (July, 2000) --- J Neural Science (July, 2001) --- J Comp Neural Sci (2002) --- J Comp Neural Sci (2002) ---
13 Two Extreme Classes: Classes of Models I. Feed forward filtering -- Hubel & Wiesel ( 62) II. Recurrent networks -- with strong excitatory feedback, and nonlinear attractor states Our model is in the middle -- recurrent network filtering with cortico-cortical inhibition dominating for simple (linear) cells, and with cortico-cortical excitation more dominant for complex (nonlinear) cells
14 Pinwheel Centers
15 Equations of the Model v j σ -- membrane potential -- σ = Exc, Inhib -- j = 2 dim label of location on cortical layer neurons per sq mm (12000 Exc, 4000 Inh) σ = E,I V E & V I -- Exc & Inh Reversal Potentials (Constants) Integrate & Fire, with voltage reset at spiking

16 Ordered and Disordered Maps Convergent LGN input confer the maps of orientation & spatial phase preference (Reid & Alonso, 1995) Regular Map of Orientation in Pinwheels (Optical Imaging: Bonhoeffer & Grinvald, 1991; Blasdel, 1992; Maldonado et al., 1997 Random Map of Spatial Phase (DeAngelis et al., 1999)

17 -- excitation -- inhibition Circles of Influence (Monosynaptic coupling)
18 Characteristic Features of our Recurrent Network Model Local (< 500 µm) lateral connectivity -- nonspecific & isotropic Ordered map of orientation preference Disordered map of spatial phase preference Dominance of cortico-cortical inhibition for simple cells Stronger cortico-cortical excitation for complex cells Each of which supported by experimental measurements
19 But remember, our model only local

20 Pinwheel Centers

21 Lateral Connections and Orientation -- Tree Shrew Bosking, Zhang, Schofield & Fitzpatrick J. Neuroscience, 1997

22
23 Coarse-Grained Representations Using the spatial regularity of cortical maps (such as orientation preference), we coarse grain the cortical layer into local cells or placquets.
24
25
26 Coarse-grained Representations: V1 Average firing rate models (Shelley & McLaughlin) m σ (x,t), σ = E,I PDF representations ( Cai, Tao & McLaughlin) ρ σ (v,g; x,t), σ = E,I Sub-network of embedded point neurons -- in a coarse-grained, dynamical background (Cai & McLaughlin)
27 Outline of Rest of Talk I. NMDA Behavior & Integrate and Fire Modeling II. Within a coarse-grained cell (or plaquet) IIa. NMDA & Oscillations and Synchrony IIb. Embedded point neurons
28 I. NMDA --Background In our network, slow excitation reduces synchrony and lessens oscillations. NMDA is a primary cause of slow excitation in V1; it is prevalent throughout the cortex, including V1 and layer 4c. NMDA channels can be blocked by Mg ions, leading to a voltage dependent conductance as mapped by Jahr & Stevens, How does this voltage dependence effect I&F modeling? Equation : I NMDA g t V Trans Volt = ( ) ( ) NMDA NMDA g [ V E ] NMDA
29 Realistic Voltage Behavior A basis for comparison. By X.J. Wang, Based upon Pinsky & Rinzel, Compartments: Soma and Dendrite. A number of ion channels, with Hodgkin-Huxley type mechanics: g Na,g K,g Ca, and g K AHP. AMPA-only synaptic makeup has been replaced by NMDA+AMPA. Significant g K AHP, slowdecaying. Potential (mv) Time (msec) Dendritic Potential Somatic Potential
30 Significant K-AHP conductance 0.2 g K AHP Other Conductances Conductance (ms) Time (msec) Can be approximated in an IF model by an Adaptive Conductance step function. 350
31 Integrate-and-Fire (IF) Modeling A neuron s voltage threshold can be found experimentally. An all or none event, the action potential (AP) or spike occurs if and only if V Th is crossed. IF modeling truncates spikes instead of modeling them, only recording spike times. Potential (mv) Dendritic Potential Somatic Potential V Th Time (msec)
32 1 NMDA Voltage-Dependence g NMDA Volt (Normalized) V Th E NMDA V Peak Jahr and Stevens, V R Voltage (mv) A Problem for IF Modeling: Superthreshold potential, the majority of the voltagedependence curve, is truncated in IF models.
33 Pasting Spikes I Everything is integrated as usual. NMDA behavior is determined by a pasted-in spike, added to the standard IF potential, following threshold crossing. Equation : Trans Volt NMDA= g () t g ( V+ V ) NMDA NMDA Paste [ V+ V E ] Paste NMDA Potential (mv) V + V Paste V Spike Times Time (msec)
34 40 Detail of Pasted Spikes Potential (mv) Time (msec) Potential (mv) Detail Time (msec) This model encourages overexcitation: there is no refractoriness.

35
36 Comparison of Firing Statistics
37
38 40 20 ISI Distributions: Light Stimulus (mean input ISI = 5 msec) Wang Model IF Model without Pasted Spike IF Model with Pasted Spike Miller Model with O riginal Parameterization
39 Why so little effect from spikes?
40 I NMDA = g NMDA (v - E NMDA )
41 Effects of Spike Height Spike Width
42 Result: Pasting Spike has little effect. Voltage (mv) Voltage (mv) Spikes 8 Spikes 9 Spikes Time (msec) Time (msec) 8 Spikes With Pasted Spike Without Pasted Spike: Identical Spike Rate Causes: (i) Reversal of current at reversal potential crossing; (ii) Short duration of each spike. Insensitivity of NMDA conductance to spikes also shown, very convincingly, by clipping spikes in Wang model
43 40 20 Regular-Spiking Model: 20 Spikes With NMDA at all times Potential (mv) Potential (mv) Time (ms) No NMDA when V D >= Spikes Time (ms)
44 Effective Reversal Potential & NMDA Modeling An alternative approach, using an effective reversal or shadow potential Krukowski & Miller (Nature Neuroscience 01; To describe this approach, we begin from the concept of an effective reversal potential (Shelley, McLaughlin, Shapley & Wielaard; J. Comp. Neurosci 02)
45 Conductance Based Model dv/dt = - g T (t) [ v - V Eff (t) ], σ = E,I where g T (t) denotes the total conductance, gj T (t) = g T + g j σe (t) + g j σi (t), and V Eff (t) = [V E g EE (t) - V I g EI (t) ] [g T (t)] -1 If [g T (t)] -1 << τ syn v V Eff (t)
46 Active Cortex - Causes High Conductances, which in turn cause Sub-threshold membrane potential ``instantaneously tracks conductances on the synaptic time scale. V(t) ~ V Eff (t) = [V E g EE (t) - V I g EI (t) ] [g T (t)] -1 where g T (t) denotes the total conductance
47 Active Background High Conductances in Active Cortex Membrane Potential Tracks Instantaneously Effective Reversal Potential

48 Effective Reversal Potential & NMDA Wang s Model Krukowski & Millers Original Model Reparameterized Model [Mg] = 1.0 mm & Stronger KAHP
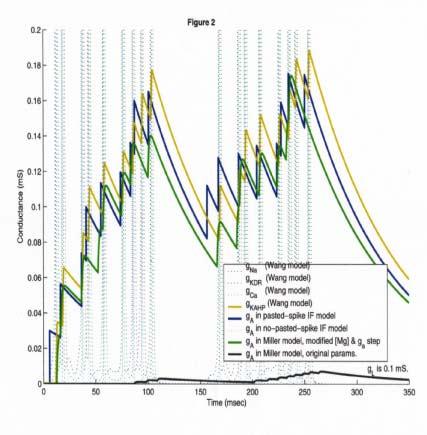
49 gkahp
50 Can we linearize g NV? (Normalized) volt g NMDA NMDA Voltage Dependence Voltage (mv) (Normalized) volt g NMDA Jahr Equation Linearized Voltage (mv) This yields nearly identical behavior.
51 Can we make g NV constant? A good constant (horizontal) value can be chosen computationally, to fit typical stimulation intensities. However, the actual value of the constant must be fit for each different numerical run (change of intensity of stimulus, etc.) Probably best to use linear representation
52
53 Conclusions (for this NMDA part of the talk) NMDA s can be realistically incorporated into traditional IF models of regularly spiking neurons, with no special modifications (pasted spike, shadow potential) necessary. That is, spikes themselves have little effect. The voltage dependence can be linearized to fit the subthreshold region. A few remarks:
54 Remark 1: Calcium Transport While voltage and spiking (for regularly firing neurons) not affected by the nonlinearity of the NMDA conductance, Not so for calcium transport because the calcium current does not average to zero over a spike. Could be important for longer time effects, such as plasticity, LTP, etc for which calcium plays a major role.
55 Remark 2: Bursting Neurons The nonlinearity could be more important for bursting neurons, which fire on top of a voltage plateau.
56 1 NMDA Voltage-Dependence g NMDA Volt (Normalized) V Th E NMDA V Peak Jahr and Stevens, V R Voltage (mv)
57 Bursting Model: With NMDA at all times 20 Potential (mv) Spikes Time (ms) No NMDA when V D >= -50 Potential (mv) Spikes Time (ms)
58 40 20 Regular-Spiking Model: 20 Spikes With NMDA at all times Potential (mv) Potential (mv) Time (ms) No NMDA when V D >= Spikes Time (ms)
59 Outline I. NMDA Behavior & Integrate and Fire Modeling II. Within a coarse-grained cell (or plaquet) IIa. NMDA & Oscillations and Synchrony IIb. Embedded point neurons
60 II. Within One Coarse-Grained Cell IIa. NMDA s Effects
61
62 Effects of NMDA Within CG Cell NMDA lessons oscillations and synchrony Increases firing rates With respect to removing synchrony, NMDA works in the same direction as synaptic failure -- as seen in an all AMPA network by (Cai, Tao & Shelley, 02) We illustrate these effects in a toy, all excitatory network
63 Driver frequency = 5Hz, no synaptic failure, 0% NMDA on Complex ps = S = 0.24, p = 1, efracs = 0., efracc = 0., n = 120 /NMDA5Hz1 Firing rate = 73/sec for SIMPLE 80 = 13/sec for Complex
64 Driver frequency = 5Hz, 1% synaptic failure, 0% NMDA on Complex ps = S = , p = 0.99, efracs = 0., efracc = 0., n = 120 /NMDA5Hz1.1 Firing rate = 74/sec for SIMPLE 80 = 14/sec for Complex
65 Driver frequency = 5Hz, no synaptic failure, 50% NMDA on Complex ps = S = 0.24, p = 1, efracs = 0., efracc = 0.5, n = 120 /NMDA5Hz2 Firing rate = 86/sec for SIMPLE 80 = 56/sec for Complex
66 Driver frequency = 5Hz, 1% synaptic failure, 50% NMDA on Complex ps = S = , p = 0.99, efracs = 0., efracc = 0.5, n = 120 /NMDA5Hz1.2 Firing rate = 88/sec for SIMPLE 80 = 65/sec for Complex
67 Driver frequency = 5Hz, no synaptic failure, 100% NMDA on Complex ps = 0.24 S = 0.24, p = 1, efracs = 0., efracc = 1., n = 120 /NMDA5Hz3 120 Firing rate = 110/sec for SIMPLE 100 = 122/sec for Complex 80 Note: Firing rate is higher for COMPLEX
68 A Realistic Network Within a CG Cell Two types of inhibition KAHP current AMPA & NMDA Very preliminary
69
70
71 Outline I. NMDA Behavior & Integrate and Fire Modeling II. Within a coarse-grained cell (or plaquet) IIa. NMDA & Oscillations and Synchrony IIb. Embedded point neurons
72 IIb. Embedded Point Neurons Point neurons -- embedded in, and fully interacting with, coarse-grained firing rate representation For scale-up computer efficiency Yet maintaining firing properties of individual neurons -- for spike coding, coincidence detection, etc. Today, I ll show some preliminary consistency studies
73 Embedded Network All excitatory network both AMPA & NMDA; Within the CG cell 200 neurons (100 simple ; 100 complex ) Compared with embedded network, with 80 of the complex neurons replaced by a CG firing rate representation, leaving 20 embedded complex neurons; In the embedded network, CG firing rate drives 80 modulated Poisson spikers.
74 Mixed Representation, SIMPLE: 100 I&F, COMPLEX: 20 I&F + MF vs 100 SIMPLE I&F and 100 COMPLEX I&F Firing rate = /sec for SIMPLE; 45-75/sec for Complex All I&F: 120 efracs = 0, efracc = 0.2 /MF6.7 I&F COMPLEX: 20% NMDA, SIMPLE: all AMPA, INPUT: all AMPA
75 Mixed Representation, SIMPLE: 100 I&F, COMPLEX: 20 I&F + MF vs 100 SIMPLE I&F and 100 COMPLEX I&F Firing rate = /sec for SIMPLE; 45-75/sec for Complex Mixed: efracs = 0, efracc = 0.2 /MF6.6 MeanField Firing rate = 126/sec for SIMPLE = 66/sec for Complex COMPLEX: 20% NMDA, SIMPLE: all AMPA, INPUT: all AMPA
76 Membrane Potential of a representative COMPLEX neuron ( all I&F vs Mixed): Voltage vs time 1 I & F 0.8 Neuron in full network Same neuron, in embedded network Mean Field Mixed
77 Embedded Network Computational efficiency -- scales as N 2 where N = # of point neurons; (i.e. 100! 20 yields 10000!400)
78 Conclusion I. NMDA Behavior & Integrate and Fire Modeling [For regularly spiking neurons, NMDA nonlinearity (and spikes) have little effect.] II. Within a coarse-grained cell (or plaquet) IIa. NMDA & Oscillations and Synchrony [Lessoned by NMDA (and synaptic failure)] IIb. Embedded point neurons [Consistency, within 10%, easily achievable.]
79
80
81
82 400 ISI Distributions: Heavy Stimulus (mean input ISI = 1.5 msec) Wang Model IF Model without Pasted Spike IF Model with Pasted Spike Miller Model with O riginal Parameterization
Dependence of Orientation Tuning on Recurrent Excitation and Inhibition in a Network Model of V1
 Dependence of Orientation Tuning on Recurrent Excitation and Inhibition in a Network Model of V1 Klaus Wimmer 1 *, Marcel Stimberg 1 *, Robert Martin 1, Lars Schwabe 2, Jorge Mariño 3, James Schummers
Dependence of Orientation Tuning on Recurrent Excitation and Inhibition in a Network Model of V1 Klaus Wimmer 1 *, Marcel Stimberg 1 *, Robert Martin 1, Lars Schwabe 2, Jorge Mariño 3, James Schummers
Basics of Computational Neuroscience: Neurons and Synapses to Networks
 Basics of Computational Neuroscience: Neurons and Synapses to Networks Bruce Graham Mathematics School of Natural Sciences University of Stirling Scotland, U.K. Useful Book Authors: David Sterratt, Bruce
Basics of Computational Neuroscience: Neurons and Synapses to Networks Bruce Graham Mathematics School of Natural Sciences University of Stirling Scotland, U.K. Useful Book Authors: David Sterratt, Bruce
Inhibition: Effects of Timing, Time Scales and Gap Junctions
 Inhibition: Effects of Timing, Time Scales and Gap Junctions I. Auditory brain stem neurons and subthreshold integ n. Fast, precise (feed forward) inhibition shapes ITD tuning. Facilitating effects of
Inhibition: Effects of Timing, Time Scales and Gap Junctions I. Auditory brain stem neurons and subthreshold integ n. Fast, precise (feed forward) inhibition shapes ITD tuning. Facilitating effects of
Prof. Greg Francis 7/31/15
 s PSY 200 Greg Francis Lecture 06 How do you recognize your grandmother? Action potential With enough excitatory input, a cell produces an action potential that sends a signal down its axon to other cells
s PSY 200 Greg Francis Lecture 06 How do you recognize your grandmother? Action potential With enough excitatory input, a cell produces an action potential that sends a signal down its axon to other cells
Cell Responses in V4 Sparse Distributed Representation
 Part 4B: Real Neurons Functions of Layers Input layer 4 from sensation or other areas 3. Neocortical Dynamics Hidden layers 2 & 3 Output layers 5 & 6 to motor systems or other areas 1 2 Hierarchical Categorical
Part 4B: Real Neurons Functions of Layers Input layer 4 from sensation or other areas 3. Neocortical Dynamics Hidden layers 2 & 3 Output layers 5 & 6 to motor systems or other areas 1 2 Hierarchical Categorical
Computational cognitive neuroscience: 2. Neuron. Lubica Beňušková Centre for Cognitive Science, FMFI Comenius University in Bratislava
 1 Computational cognitive neuroscience: 2. Neuron Lubica Beňušková Centre for Cognitive Science, FMFI Comenius University in Bratislava 2 Neurons communicate via electric signals In neurons it is important
1 Computational cognitive neuroscience: 2. Neuron Lubica Beňušková Centre for Cognitive Science, FMFI Comenius University in Bratislava 2 Neurons communicate via electric signals In neurons it is important
The extraordinary power of the brain is apparent from the
 Images courtesy of Daniel Einstein Multiscale Modeling of the Primary Visual Cortex Modeling of Neuronal Network Dynamics MULTISCALE MODELING PART 2 BY AADITYA V. RANGAN, LOUIS TAO, GREGOR KOVACIC, AND
Images courtesy of Daniel Einstein Multiscale Modeling of the Primary Visual Cortex Modeling of Neuronal Network Dynamics MULTISCALE MODELING PART 2 BY AADITYA V. RANGAN, LOUIS TAO, GREGOR KOVACIC, AND
Intro. Comp. NeuroSci. Ch. 9 October 4, The threshold and channel memory
 9.7.4 The threshold and channel memory The action potential has a threshold. In figure the area around threshold is expanded (rectangle). A current injection that does not reach the threshold does not
9.7.4 The threshold and channel memory The action potential has a threshold. In figure the area around threshold is expanded (rectangle). A current injection that does not reach the threshold does not
Plasticity of Cerebral Cortex in Development
 Plasticity of Cerebral Cortex in Development Jessica R. Newton and Mriganka Sur Department of Brain & Cognitive Sciences Picower Center for Learning & Memory Massachusetts Institute of Technology Cambridge,
Plasticity of Cerebral Cortex in Development Jessica R. Newton and Mriganka Sur Department of Brain & Cognitive Sciences Picower Center for Learning & Memory Massachusetts Institute of Technology Cambridge,
Perceptual Grouping in a Self-Organizing Map of Spiking Neurons
 Perceptual Grouping in a Self-Organizing Map of Spiking Neurons Yoonsuck Choe Department of Computer Sciences The University of Texas at Austin August 13, 2001 Perceptual Grouping Group Two! Longest Contour?
Perceptual Grouping in a Self-Organizing Map of Spiking Neurons Yoonsuck Choe Department of Computer Sciences The University of Texas at Austin August 13, 2001 Perceptual Grouping Group Two! Longest Contour?
Transitions between dierent synchronous ring modes using synaptic depression
 Neurocomputing 44 46 (2002) 61 67 www.elsevier.com/locate/neucom Transitions between dierent synchronous ring modes using synaptic depression Victoria Booth, Amitabha Bose Department of Mathematical Sciences,
Neurocomputing 44 46 (2002) 61 67 www.elsevier.com/locate/neucom Transitions between dierent synchronous ring modes using synaptic depression Victoria Booth, Amitabha Bose Department of Mathematical Sciences,
Different Roles for Simple-Cell and Complex-Cell Inhibition in V1
 The Journal of Neuroscience, November 12, 2003 23(32):10201 10213 10201 Behavioral/Systems/Cognitive Different Roles for Simple-Cell and Complex-Cell Inhibition in V1 Thomas Z. Lauritzen 1,4 and Kenneth
The Journal of Neuroscience, November 12, 2003 23(32):10201 10213 10201 Behavioral/Systems/Cognitive Different Roles for Simple-Cell and Complex-Cell Inhibition in V1 Thomas Z. Lauritzen 1,4 and Kenneth
Models of visual neuron function. Quantitative Biology Course Lecture Dan Butts
 Models of visual neuron function Quantitative Biology Course Lecture Dan Butts 1 What is the neural code"? 11 10 neurons 1,000-10,000 inputs Electrical activity: nerve impulses 2 ? How does neural activity
Models of visual neuron function Quantitative Biology Course Lecture Dan Butts 1 What is the neural code"? 11 10 neurons 1,000-10,000 inputs Electrical activity: nerve impulses 2 ? How does neural activity
Input-speci"c adaptation in complex cells through synaptic depression
 0 0 0 0 Neurocomputing }0 (00) } Input-speci"c adaptation in complex cells through synaptic depression Frances S. Chance*, L.F. Abbott Volen Center for Complex Systems and Department of Biology, Brandeis
0 0 0 0 Neurocomputing }0 (00) } Input-speci"c adaptation in complex cells through synaptic depression Frances S. Chance*, L.F. Abbott Volen Center for Complex Systems and Department of Biology, Brandeis
Computing with Spikes in Recurrent Neural Networks
 Computing with Spikes in Recurrent Neural Networks Dezhe Jin Department of Physics The Pennsylvania State University Presented at ICS Seminar Course, Penn State Jan 9, 2006 Outline Introduction Neurons,
Computing with Spikes in Recurrent Neural Networks Dezhe Jin Department of Physics The Pennsylvania State University Presented at ICS Seminar Course, Penn State Jan 9, 2006 Outline Introduction Neurons,
Theory of correlation transfer and correlation structure in recurrent networks
 Theory of correlation transfer and correlation structure in recurrent networks Ruben Moreno-Bote Foundation Sant Joan de Déu, Barcelona Moritz Helias Research Center Jülich Theory of correlation transfer
Theory of correlation transfer and correlation structure in recurrent networks Ruben Moreno-Bote Foundation Sant Joan de Déu, Barcelona Moritz Helias Research Center Jülich Theory of correlation transfer
M Cells. Why parallel pathways? P Cells. Where from the retina? Cortical visual processing. Announcements. Main visual pathway from retina to V1
 Announcements exam 1 this Thursday! review session: Wednesday, 5:00-6:30pm, Meliora 203 Bryce s office hours: Wednesday, 3:30-5:30pm, Gleason https://www.youtube.com/watch?v=zdw7pvgz0um M Cells M cells
Announcements exam 1 this Thursday! review session: Wednesday, 5:00-6:30pm, Meliora 203 Bryce s office hours: Wednesday, 3:30-5:30pm, Gleason https://www.youtube.com/watch?v=zdw7pvgz0um M Cells M cells
Analysis of in-vivo extracellular recordings. Ryan Morrill Bootcamp 9/10/2014
 Analysis of in-vivo extracellular recordings Ryan Morrill Bootcamp 9/10/2014 Goals for the lecture Be able to: Conceptually understand some of the analysis and jargon encountered in a typical (sensory)
Analysis of in-vivo extracellular recordings Ryan Morrill Bootcamp 9/10/2014 Goals for the lecture Be able to: Conceptually understand some of the analysis and jargon encountered in a typical (sensory)
Primary Visual Pathways (I)
") Primary Visual Pathways (I) Introduction to Computational and Biological Vision CS 202-1-5261 Computer Science Department, BGU Ohad Ben-Shahar Where does visual information go from the eye? Where does
Primary Visual Pathways (I) Introduction to Computational and Biological Vision CS 202-1-5261 Computer Science Department, BGU Ohad Ben-Shahar Where does visual information go from the eye? Where does
Information Processing During Transient Responses in the Crayfish Visual System
 Information Processing During Transient Responses in the Crayfish Visual System Christopher J. Rozell, Don. H. Johnson and Raymon M. Glantz Department of Electrical & Computer Engineering Department of
Information Processing During Transient Responses in the Crayfish Visual System Christopher J. Rozell, Don. H. Johnson and Raymon M. Glantz Department of Electrical & Computer Engineering Department of
Introduction to Computational Neuroscience
 Introduction to Computational Neuroscience Lecture 7: Network models Lesson Title 1 Introduction 2 Structure and Function of the NS 3 Windows to the Brain 4 Data analysis 5 Data analysis II 6 Single neuron
Introduction to Computational Neuroscience Lecture 7: Network models Lesson Title 1 Introduction 2 Structure and Function of the NS 3 Windows to the Brain 4 Data analysis 5 Data analysis II 6 Single neuron
2/3/17. Visual System I. I. Eye, color space, adaptation II. Receptive fields and lateral inhibition III. Thalamus and primary visual cortex
 1 Visual System I I. Eye, color space, adaptation II. Receptive fields and lateral inhibition III. Thalamus and primary visual cortex 2 1 2/3/17 Window of the Soul 3 Information Flow: From Photoreceptors
1 Visual System I I. Eye, color space, adaptation II. Receptive fields and lateral inhibition III. Thalamus and primary visual cortex 2 1 2/3/17 Window of the Soul 3 Information Flow: From Photoreceptors
Thalamo-Cortical Relationships Ultrastructure of Thalamic Synaptic Glomerulus
 Central Visual Pathways V1/2 NEUR 3001 dvanced Visual Neuroscience The Lateral Geniculate Nucleus () is more than a relay station LP SC Professor Tom Salt UCL Institute of Ophthalmology Retina t.salt@ucl.ac.uk
Central Visual Pathways V1/2 NEUR 3001 dvanced Visual Neuroscience The Lateral Geniculate Nucleus () is more than a relay station LP SC Professor Tom Salt UCL Institute of Ophthalmology Retina t.salt@ucl.ac.uk
Nonlinear processing in LGN neurons
 Nonlinear processing in LGN neurons Vincent Bonin *, Valerio Mante and Matteo Carandini Smith-Kettlewell Eye Research Institute 2318 Fillmore Street San Francisco, CA 94115, USA Institute of Neuroinformatics
Nonlinear processing in LGN neurons Vincent Bonin *, Valerio Mante and Matteo Carandini Smith-Kettlewell Eye Research Institute 2318 Fillmore Street San Francisco, CA 94115, USA Institute of Neuroinformatics
Neuromorphic computing
 Neuromorphic computing Robotics M.Sc. programme in Computer Science lorenzo.vannucci@santannapisa.it April 19th, 2018 Outline 1. Introduction 2. Fundamentals of neuroscience 3. Simulating the brain 4.
Neuromorphic computing Robotics M.Sc. programme in Computer Science lorenzo.vannucci@santannapisa.it April 19th, 2018 Outline 1. Introduction 2. Fundamentals of neuroscience 3. Simulating the brain 4.
Modeling Depolarization Induced Suppression of Inhibition in Pyramidal Neurons
 Modeling Depolarization Induced Suppression of Inhibition in Pyramidal Neurons Peter Osseward, Uri Magaram Department of Neuroscience University of California, San Diego La Jolla, CA 92092 possewar@ucsd.edu
Modeling Depolarization Induced Suppression of Inhibition in Pyramidal Neurons Peter Osseward, Uri Magaram Department of Neuroscience University of California, San Diego La Jolla, CA 92092 possewar@ucsd.edu
Beyond Vanilla LTP. Spike-timing-dependent-plasticity or STDP
 Beyond Vanilla LTP Spike-timing-dependent-plasticity or STDP Hebbian learning rule asn W MN,aSN MN Δw ij = μ x j (v i - φ) learning threshold under which LTD can occur Stimulation electrode Recording electrode
Beyond Vanilla LTP Spike-timing-dependent-plasticity or STDP Hebbian learning rule asn W MN,aSN MN Δw ij = μ x j (v i - φ) learning threshold under which LTD can occur Stimulation electrode Recording electrode
Investigation of Physiological Mechanism For Linking Field Synapses
 Investigation of Physiological Mechanism For Linking Field Synapses Richard B. Wells 1, Nick Garrett 2, Tom Richner 3 Microelectronics Research and Communications Institute (MRCI) BEL 316 University of
Investigation of Physiological Mechanism For Linking Field Synapses Richard B. Wells 1, Nick Garrett 2, Tom Richner 3 Microelectronics Research and Communications Institute (MRCI) BEL 316 University of
Basics of Computational Neuroscience
 Basics of Computational Neuroscience 1 1) Introduction Lecture: Computational Neuroscience, The Basics A reminder: Contents 1) Brain, Maps,, Networks,, and The tough stuff: 2,3) Membrane Models 3,4) Spiking
Basics of Computational Neuroscience 1 1) Introduction Lecture: Computational Neuroscience, The Basics A reminder: Contents 1) Brain, Maps,, Networks,, and The tough stuff: 2,3) Membrane Models 3,4) Spiking
Early Stages of Vision Might Explain Data to Information Transformation
 Early Stages of Vision Might Explain Data to Information Transformation Baran Çürüklü Department of Computer Science and Engineering Mälardalen University Västerås S-721 23, Sweden Abstract. In this paper
Early Stages of Vision Might Explain Data to Information Transformation Baran Çürüklü Department of Computer Science and Engineering Mälardalen University Västerås S-721 23, Sweden Abstract. In this paper
Photoreceptors Rods. Cones
 Photoreceptors Rods Cones 120 000 000 Dim light Prefer wavelength of 505 nm Monochromatic Mainly in periphery of the eye 6 000 000 More light Different spectral sensitivities!long-wave receptors (558 nm)
Photoreceptors Rods Cones 120 000 000 Dim light Prefer wavelength of 505 nm Monochromatic Mainly in periphery of the eye 6 000 000 More light Different spectral sensitivities!long-wave receptors (558 nm)
CS294-6 (Fall 2004) Recognizing People, Objects and Actions Lecture: January 27, 2004 Human Visual System
 Recognizing People, Objects and Actions Lecture: January 27, 2004 Human Visual System") CS294-6 (Fall 2004) Recognizing People, Objects and Actions Lecture: January 27, 2004 Human Visual System Lecturer: Jitendra Malik Scribe: Ryan White (Slide: layout of the brain) Facts about the brain:
CS294-6 (Fall 2004) Recognizing People, Objects and Actions Lecture: January 27, 2004 Human Visual System Lecturer: Jitendra Malik Scribe: Ryan White (Slide: layout of the brain) Facts about the brain:
The Integration of Features in Visual Awareness : The Binding Problem. By Andrew Laguna, S.J.
 The Integration of Features in Visual Awareness : The Binding Problem By Andrew Laguna, S.J. Outline I. Introduction II. The Visual System III. What is the Binding Problem? IV. Possible Theoretical Solutions
The Integration of Features in Visual Awareness : The Binding Problem By Andrew Laguna, S.J. Outline I. Introduction II. The Visual System III. What is the Binding Problem? IV. Possible Theoretical Solutions
The Role of Mitral Cells in State Dependent Olfactory Responses. Trygve Bakken & Gunnar Poplawski
 The Role of Mitral Cells in State Dependent Olfactory Responses Trygve akken & Gunnar Poplawski GGN 260 Neurodynamics Winter 2008 bstract Many behavioral studies have shown a reduced responsiveness to
The Role of Mitral Cells in State Dependent Olfactory Responses Trygve akken & Gunnar Poplawski GGN 260 Neurodynamics Winter 2008 bstract Many behavioral studies have shown a reduced responsiveness to
How has Computational Neuroscience been useful? Virginia R. de Sa Department of Cognitive Science UCSD
 How has Computational Neuroscience been useful? 1 Virginia R. de Sa Department of Cognitive Science UCSD What is considered Computational Neuroscience? 2 What is considered Computational Neuroscience?
How has Computational Neuroscience been useful? 1 Virginia R. de Sa Department of Cognitive Science UCSD What is considered Computational Neuroscience? 2 What is considered Computational Neuroscience?
Lecture 22: A little Neurobiology
 BIO 5099: Molecular Biology for Computer Scientists (et al) Lecture 22: A little Neurobiology http://compbio.uchsc.edu/hunter/bio5099 Larry.Hunter@uchsc.edu Nervous system development Part of the ectoderm
BIO 5099: Molecular Biology for Computer Scientists (et al) Lecture 22: A little Neurobiology http://compbio.uchsc.edu/hunter/bio5099 Larry.Hunter@uchsc.edu Nervous system development Part of the ectoderm
Mechanisms of stimulus feature selectivity in sensory systems
 Mechanisms of stimulus feature selectivity in sensory systems 1. Orientation and direction selectivity in the visual cortex 2. Selectivity to sound frequency in the auditory cortex 3. Feature selectivity
Mechanisms of stimulus feature selectivity in sensory systems 1. Orientation and direction selectivity in the visual cortex 2. Selectivity to sound frequency in the auditory cortex 3. Feature selectivity
Bursting dynamics in the brain. Jaeseung Jeong, Department of Biosystems, KAIST
 Bursting dynamics in the brain Jaeseung Jeong, Department of Biosystems, KAIST Tonic and phasic activity A neuron is said to exhibit a tonic activity when it fires a series of single action potentials
Bursting dynamics in the brain Jaeseung Jeong, Department of Biosystems, KAIST Tonic and phasic activity A neuron is said to exhibit a tonic activity when it fires a series of single action potentials
Theta sequences are essential for internally generated hippocampal firing fields.
 Theta sequences are essential for internally generated hippocampal firing fields. Yingxue Wang, Sandro Romani, Brian Lustig, Anthony Leonardo, Eva Pastalkova Supplementary Materials Supplementary Modeling
Theta sequences are essential for internally generated hippocampal firing fields. Yingxue Wang, Sandro Romani, Brian Lustig, Anthony Leonardo, Eva Pastalkova Supplementary Materials Supplementary Modeling
Synaptic plasticityhippocampus. Neur 8790 Topics in Neuroscience: Neuroplasticity. Outline. Synaptic plasticity hypothesis
 Synaptic plasticityhippocampus Neur 8790 Topics in Neuroscience: Neuroplasticity Outline Synaptic plasticity hypothesis Long term potentiation in the hippocampus How it s measured What it looks like Mechanisms
Synaptic plasticityhippocampus Neur 8790 Topics in Neuroscience: Neuroplasticity Outline Synaptic plasticity hypothesis Long term potentiation in the hippocampus How it s measured What it looks like Mechanisms
Lateral Geniculate Nucleus (LGN)
") Lateral Geniculate Nucleus (LGN) What happens beyond the retina? What happens in Lateral Geniculate Nucleus (LGN)- 90% flow Visual cortex Information Flow Superior colliculus 10% flow Slide 2 Information
Lateral Geniculate Nucleus (LGN) What happens beyond the retina? What happens in Lateral Geniculate Nucleus (LGN)- 90% flow Visual cortex Information Flow Superior colliculus 10% flow Slide 2 Information
Dopamine modulation of prefrontal delay activity - Reverberatory activity and sharpness of tuning curves
 Dopamine modulation of prefrontal delay activity - Reverberatory activity and sharpness of tuning curves Gabriele Scheler+ and Jean-Marc Fellous* +Sloan Center for Theoretical Neurobiology *Computational
Dopamine modulation of prefrontal delay activity - Reverberatory activity and sharpness of tuning curves Gabriele Scheler+ and Jean-Marc Fellous* +Sloan Center for Theoretical Neurobiology *Computational
Working models of working memory
 Working models of working memory Omri Barak and Misha Tsodyks 2014, Curr. Op. in Neurobiology Referenced by Kristjan-Julius Laak Sept 16th 2015 Tartu Working models of working memory Working models of
Working models of working memory Omri Barak and Misha Tsodyks 2014, Curr. Op. in Neurobiology Referenced by Kristjan-Julius Laak Sept 16th 2015 Tartu Working models of working memory Working models of
Reading Assignments: Lecture 5: Introduction to Vision. None. Brain Theory and Artificial Intelligence
 Brain Theory and Artificial Intelligence Lecture 5:. Reading Assignments: None 1 Projection 2 Projection 3 Convention: Visual Angle Rather than reporting two numbers (size of object and distance to observer),
Brain Theory and Artificial Intelligence Lecture 5:. Reading Assignments: None 1 Projection 2 Projection 3 Convention: Visual Angle Rather than reporting two numbers (size of object and distance to observer),
Synaptic Transmission: Ionic and Metabotropic
 Synaptic Transmission: Ionic and Metabotropic D. Purves et al. Neuroscience (Sinauer Assoc.) Chapters 5, 6, 7. C. Koch. Biophysics of Computation (Oxford) Chapter 4. J.G. Nicholls et al. From Neuron to
Synaptic Transmission: Ionic and Metabotropic D. Purves et al. Neuroscience (Sinauer Assoc.) Chapters 5, 6, 7. C. Koch. Biophysics of Computation (Oxford) Chapter 4. J.G. Nicholls et al. From Neuron to
SUPPLEMENTARY INFORMATION. Supplementary Figure 1
 SUPPLEMENTARY INFORMATION Supplementary Figure 1 The supralinear events evoked in CA3 pyramidal cells fulfill the criteria for NMDA spikes, exhibiting a threshold, sensitivity to NMDAR blockade, and all-or-none
SUPPLEMENTARY INFORMATION Supplementary Figure 1 The supralinear events evoked in CA3 pyramidal cells fulfill the criteria for NMDA spikes, exhibiting a threshold, sensitivity to NMDAR blockade, and all-or-none
OPTO 5320 VISION SCIENCE I
 OPTO 5320 VISION SCIENCE I Monocular Sensory Processes of Vision: Color Vision Mechanisms of Color Processing . Neural Mechanisms of Color Processing A. Parallel processing - M- & P- pathways B. Second
OPTO 5320 VISION SCIENCE I Monocular Sensory Processes of Vision: Color Vision Mechanisms of Color Processing . Neural Mechanisms of Color Processing A. Parallel processing - M- & P- pathways B. Second
Cognitive Modelling Themes in Neural Computation. Tom Hartley
 Cognitive Modelling Themes in Neural Computation Tom Hartley t.hartley@psychology.york.ac.uk Typical Model Neuron x i w ij x j =f(σw ij x j ) w jk x k McCulloch & Pitts (1943), Rosenblatt (1957) Net input:
Cognitive Modelling Themes in Neural Computation Tom Hartley t.hartley@psychology.york.ac.uk Typical Model Neuron x i w ij x j =f(σw ij x j ) w jk x k McCulloch & Pitts (1943), Rosenblatt (1957) Net input:
Relative contributions of cortical and thalamic feedforward inputs to V2
 Relative contributions of cortical and thalamic feedforward inputs to V2 1 2 3 4 5 Rachel M. Cassidy Neuroscience Graduate Program University of California, San Diego La Jolla, CA 92093 rcassidy@ucsd.edu
Relative contributions of cortical and thalamic feedforward inputs to V2 1 2 3 4 5 Rachel M. Cassidy Neuroscience Graduate Program University of California, San Diego La Jolla, CA 92093 rcassidy@ucsd.edu
Part 11: Mechanisms of Learning
 Neurophysiology and Information: Theory of Brain Function Christopher Fiorillo BiS 527, Spring 2012 042 350 4326, fiorillo@kaist.ac.kr Part 11: Mechanisms of Learning Reading: Bear, Connors, and Paradiso,
Neurophysiology and Information: Theory of Brain Function Christopher Fiorillo BiS 527, Spring 2012 042 350 4326, fiorillo@kaist.ac.kr Part 11: Mechanisms of Learning Reading: Bear, Connors, and Paradiso,
Spontaneous Cortical Activity Reveals Hallmarks of an Optimal Internal Model of the Environment. Berkes, Orban, Lengyel, Fiser.
 Statistically optimal perception and learning: from behavior to neural representations. Fiser, Berkes, Orban & Lengyel Trends in Cognitive Sciences (2010) Spontaneous Cortical Activity Reveals Hallmarks
Statistically optimal perception and learning: from behavior to neural representations. Fiser, Berkes, Orban & Lengyel Trends in Cognitive Sciences (2010) Spontaneous Cortical Activity Reveals Hallmarks
Signal detection in networks of spiking neurons with dynamical synapses
 Published in AIP Proceedings 887, 83-88, 7. Signal detection in networks of spiking neurons with dynamical synapses Jorge F. Mejías and Joaquín J. Torres Dept. of Electromagnetism and Physics of the Matter
Published in AIP Proceedings 887, 83-88, 7. Signal detection in networks of spiking neurons with dynamical synapses Jorge F. Mejías and Joaquín J. Torres Dept. of Electromagnetism and Physics of the Matter
Beyond bumps: Spiking networks that store sets of functions
 Neurocomputing 38}40 (2001) 581}586 Beyond bumps: Spiking networks that store sets of functions Chris Eliasmith*, Charles H. Anderson Department of Philosophy, University of Waterloo, Waterloo, Ont, N2L
Neurocomputing 38}40 (2001) 581}586 Beyond bumps: Spiking networks that store sets of functions Chris Eliasmith*, Charles H. Anderson Department of Philosophy, University of Waterloo, Waterloo, Ont, N2L
Neural response time integration subserves. perceptual decisions - K-F Wong and X-J Wang s. reduced model
 Neural response time integration subserves perceptual decisions - K-F Wong and X-J Wang s reduced model Chris Ayers and Narayanan Krishnamurthy December 15, 2008 Abstract A neural network describing the
Neural response time integration subserves perceptual decisions - K-F Wong and X-J Wang s reduced model Chris Ayers and Narayanan Krishnamurthy December 15, 2008 Abstract A neural network describing the
Rule-based firing for network simulations
 Neurocomputing 69 (2006) 1160 1164 www.elsevier.com/locate/neucom Rule-based firing for network simulations William W. Lytton, Mark Stewart Departments of Physiology, Pharmacology & Neurology, SUNY Downstate,
Neurocomputing 69 (2006) 1160 1164 www.elsevier.com/locate/neucom Rule-based firing for network simulations William W. Lytton, Mark Stewart Departments of Physiology, Pharmacology & Neurology, SUNY Downstate,
Spectrograms (revisited)
") Spectrograms (revisited) We begin the lecture by reviewing the units of spectrograms, which I had only glossed over when I covered spectrograms at the end of lecture 19. We then relate the blocks of a
Spectrograms (revisited) We begin the lecture by reviewing the units of spectrograms, which I had only glossed over when I covered spectrograms at the end of lecture 19. We then relate the blocks of a
Lecture overview. What hypothesis to test in the fly? Quantitative data collection Visual physiology conventions ( Methods )
") Lecture overview What hypothesis to test in the fly? Quantitative data collection Visual physiology conventions ( Methods ) 1 Lecture overview What hypothesis to test in the fly? Quantitative data collection
Lecture overview What hypothesis to test in the fly? Quantitative data collection Visual physiology conventions ( Methods ) 1 Lecture overview What hypothesis to test in the fly? Quantitative data collection
Cellular Bioelectricity
 ELEC ENG 3BB3: Cellular Bioelectricity Notes for Lecture 24 Thursday, March 6, 2014 8. NEURAL ELECTROPHYSIOLOGY We will look at: Structure of the nervous system Sensory transducers and neurons Neural coding
ELEC ENG 3BB3: Cellular Bioelectricity Notes for Lecture 24 Thursday, March 6, 2014 8. NEURAL ELECTROPHYSIOLOGY We will look at: Structure of the nervous system Sensory transducers and neurons Neural coding
Afferent specificity, feature specific connectivity influence orientation selectivity: A computational study in mouse primary visual cortex
 Afferent specificity, feature specific connectivity influence orientation selectivity: A computational study in mouse primary visual cortex arxiv:131.996v1 [q-bio.nc] 6 Jan 13 Dipanjan Roy *, Yenni Tjandra
Afferent specificity, feature specific connectivity influence orientation selectivity: A computational study in mouse primary visual cortex arxiv:131.996v1 [q-bio.nc] 6 Jan 13 Dipanjan Roy *, Yenni Tjandra
TEMPORAL PRECISION OF SENSORY RESPONSES Berry and Meister, 1998
 TEMPORAL PRECISION OF SENSORY RESPONSES Berry and Meister, 1998 Today: (1) how can we measure temporal precision? (2) what mechanisms enable/limit precision? A. 0.1 pa WHY SHOULD YOU CARE? average rod
TEMPORAL PRECISION OF SENSORY RESPONSES Berry and Meister, 1998 Today: (1) how can we measure temporal precision? (2) what mechanisms enable/limit precision? A. 0.1 pa WHY SHOULD YOU CARE? average rod
The Emergence of Contrast-Invariant Orientation Tuning in Simple Cells of Cat Visual Cortex
 Article The Emergence of Contrast-Invariant Orientation Tuning in Simple Cells of Cat Visual Cortex Ian M. Finn, 1,2 Nicholas J. Priebe, 1,2 and David Ferster 1, * 1 Department of Neurobiology and Physiology,
Article The Emergence of Contrast-Invariant Orientation Tuning in Simple Cells of Cat Visual Cortex Ian M. Finn, 1,2 Nicholas J. Priebe, 1,2 and David Ferster 1, * 1 Department of Neurobiology and Physiology,
Modeling synaptic facilitation and depression in thalamocortical relay cells
 College of William and Mary W&M ScholarWorks Undergraduate Honors Theses Theses, Dissertations, & Master Projects 5-2011 Modeling synaptic facilitation and depression in thalamocortical relay cells Olivia
College of William and Mary W&M ScholarWorks Undergraduate Honors Theses Theses, Dissertations, & Master Projects 5-2011 Modeling synaptic facilitation and depression in thalamocortical relay cells Olivia
Models of Attention. Models of Attention
 Models of Models of predictive: can we predict eye movements (bottom up attention)? [L. Itti and coll] pop out and saliency? [Z. Li] Readings: Maunsell & Cook, the role of attention in visual processing,
Models of Models of predictive: can we predict eye movements (bottom up attention)? [L. Itti and coll] pop out and saliency? [Z. Li] Readings: Maunsell & Cook, the role of attention in visual processing,
Synfire chains with conductance-based neurons: internal timing and coordination with timed input
 Neurocomputing 5 (5) 9 5 www.elsevier.com/locate/neucom Synfire chains with conductance-based neurons: internal timing and coordination with timed input Friedrich T. Sommer a,, Thomas Wennekers b a Redwood
Neurocomputing 5 (5) 9 5 www.elsevier.com/locate/neucom Synfire chains with conductance-based neurons: internal timing and coordination with timed input Friedrich T. Sommer a,, Thomas Wennekers b a Redwood
Resonant synchronization of heterogeneous inhibitory networks
 Cerebellar oscillations: Anesthetized rats Transgenic animals Recurrent model Review of literature: γ Network resonance Life simulations Resonance frequency Conclusion Resonant synchronization of heterogeneous
Cerebellar oscillations: Anesthetized rats Transgenic animals Recurrent model Review of literature: γ Network resonance Life simulations Resonance frequency Conclusion Resonant synchronization of heterogeneous
Memory Systems II How Stored: Engram and LTP. Reading: BCP Chapter 25
 Memory Systems II How Stored: Engram and LTP Reading: BCP Chapter 25 Memory Systems Learning is the acquisition of new knowledge or skills. Memory is the retention of learned information. Many different
Memory Systems II How Stored: Engram and LTP Reading: BCP Chapter 25 Memory Systems Learning is the acquisition of new knowledge or skills. Memory is the retention of learned information. Many different
How Neurons Do Integrals. Mark Goldman
 How Neurons Do Integrals Mark Goldman Outline 1. What is the neural basis of short-term memory? 2. A model system: the Oculomotor Neural Integrator 3. Neural mechanisms of integration: Linear network theory
How Neurons Do Integrals Mark Goldman Outline 1. What is the neural basis of short-term memory? 2. A model system: the Oculomotor Neural Integrator 3. Neural mechanisms of integration: Linear network theory
Simulating inputs of parvalbumin inhibitory interneurons onto excitatory pyramidal cells in piriform cortex
 Simulating inputs of parvalbumin inhibitory interneurons onto excitatory pyramidal cells in piriform cortex Jeffrey E. Dahlen jdahlen@ucsd.edu and Kerin K. Higa khiga@ucsd.edu Department of Neuroscience
Simulating inputs of parvalbumin inhibitory interneurons onto excitatory pyramidal cells in piriform cortex Jeffrey E. Dahlen jdahlen@ucsd.edu and Kerin K. Higa khiga@ucsd.edu Department of Neuroscience
File name: Supplementary Information Description: Supplementary Figures, Supplementary Table and Supplementary References
 File name: Supplementary Information Description: Supplementary Figures, Supplementary Table and Supplementary References File name: Supplementary Data 1 Description: Summary datasheets showing the spatial
File name: Supplementary Information Description: Supplementary Figures, Supplementary Table and Supplementary References File name: Supplementary Data 1 Description: Summary datasheets showing the spatial
Winter 2017 PHYS 178/278 Project Topics
 Winter 2017 PHYS 178/278 Project Topics 1. Recurrent Neural Integrator Network Model for Horizontal Eye Position Stability of the Memory of Eye Position in a Recurrent Network of Conductance- Based Model
Winter 2017 PHYS 178/278 Project Topics 1. Recurrent Neural Integrator Network Model for Horizontal Eye Position Stability of the Memory of Eye Position in a Recurrent Network of Conductance- Based Model
Embryological origin of thalamus
 diencephalon Embryological origin of thalamus The diencephalon gives rise to the: Thalamus Epithalamus (pineal gland, habenula, paraventricular n.) Hypothalamus Subthalamus (Subthalamic nuclei) The Thalamus:
diencephalon Embryological origin of thalamus The diencephalon gives rise to the: Thalamus Epithalamus (pineal gland, habenula, paraventricular n.) Hypothalamus Subthalamus (Subthalamic nuclei) The Thalamus:
Why is our capacity of working memory so large?
 LETTER Why is our capacity of working memory so large? Thomas P. Trappenberg Faculty of Computer Science, Dalhousie University 6050 University Avenue, Halifax, Nova Scotia B3H 1W5, Canada E-mail: tt@cs.dal.ca
LETTER Why is our capacity of working memory so large? Thomas P. Trappenberg Faculty of Computer Science, Dalhousie University 6050 University Avenue, Halifax, Nova Scotia B3H 1W5, Canada E-mail: tt@cs.dal.ca
LISC-322 Neuroscience Cortical Organization
 LISC-322 Neuroscience Cortical Organization THE VISUAL SYSTEM Higher Visual Processing Martin Paré Assistant Professor Physiology & Psychology Most of the cortex that covers the cerebral hemispheres is
LISC-322 Neuroscience Cortical Organization THE VISUAL SYSTEM Higher Visual Processing Martin Paré Assistant Professor Physiology & Psychology Most of the cortex that covers the cerebral hemispheres is
Rolls,E.T. (2016) Cerebral Cortex: Principles of Operation. Oxford University Press.
 Cerebral Cortex: Principles of Operation. Oxford University Press.") Digital Signal Processing and the Brain Is the brain a digital signal processor? Digital vs continuous signals Digital signals involve streams of binary encoded numbers The brain uses digital, all or none,
Digital Signal Processing and the Brain Is the brain a digital signal processor? Digital vs continuous signals Digital signals involve streams of binary encoded numbers The brain uses digital, all or none,
The Eye. Cognitive Neuroscience of Language. Today s goals. 5 From eye to brain. Today s reading
 Cognitive Neuroscience of Language 5 From eye to brain Today s goals Look at the pathways that conduct the visual information from the eye to the visual cortex Marielle Lange http://homepages.inf.ed.ac.uk/mlange/teaching/cnl/
Cognitive Neuroscience of Language 5 From eye to brain Today s goals Look at the pathways that conduct the visual information from the eye to the visual cortex Marielle Lange http://homepages.inf.ed.ac.uk/mlange/teaching/cnl/
Self-Organization and Segmentation with Laterally Connected Spiking Neurons
 Self-Organization and Segmentation with Laterally Connected Spiking Neurons Yoonsuck Choe Department of Computer Sciences The University of Texas at Austin Austin, TX 78712 USA Risto Miikkulainen Department
Self-Organization and Segmentation with Laterally Connected Spiking Neurons Yoonsuck Choe Department of Computer Sciences The University of Texas at Austin Austin, TX 78712 USA Risto Miikkulainen Department
Neural circuits PSY 310 Greg Francis. Lecture 05. Rods and cones
 Neural circuits PSY 310 Greg Francis Lecture 05 Why do you need bright light to read? Rods and cones Photoreceptors are not evenly distributed across the retina 1 Rods and cones Cones are most dense in
Neural circuits PSY 310 Greg Francis Lecture 05 Why do you need bright light to read? Rods and cones Photoreceptors are not evenly distributed across the retina 1 Rods and cones Cones are most dense in
Consciousness The final frontier!
 Consciousness The final frontier! How to Define it??? awareness perception - automatic and controlled memory - implicit and explicit ability to tell us about experiencing it attention. And the bottleneck
Consciousness The final frontier! How to Define it??? awareness perception - automatic and controlled memory - implicit and explicit ability to tell us about experiencing it attention. And the bottleneck
Neural Coding. Computing and the Brain. How Is Information Coded in Networks of Spiking Neurons?
 Neural Coding Computing and the Brain How Is Information Coded in Networks of Spiking Neurons? Coding in spike (AP) sequences from individual neurons Coding in activity of a population of neurons Spring
Neural Coding Computing and the Brain How Is Information Coded in Networks of Spiking Neurons? Coding in spike (AP) sequences from individual neurons Coding in activity of a population of neurons Spring
PHY3111 Mid-Semester Test Study. Lecture 2: The hierarchical organisation of vision
 PHY3111 Mid-Semester Test Study Lecture 2: The hierarchical organisation of vision 1. Explain what a hierarchically organised neural system is, in terms of physiological response properties of its neurones.
PHY3111 Mid-Semester Test Study Lecture 2: The hierarchical organisation of vision 1. Explain what a hierarchically organised neural system is, in terms of physiological response properties of its neurones.
Multi compartment model of synaptic plasticity
 Multi compartment model of synaptic plasticity E. Paxon Frady We introduce a biophysical model of a neuronal network that can accurately replicate many classical plasticity experiments. The model uses
Multi compartment model of synaptic plasticity E. Paxon Frady We introduce a biophysical model of a neuronal network that can accurately replicate many classical plasticity experiments. The model uses
Emanuel Todorov, Athanassios Siapas and David Somers. Dept. of Brain and Cognitive Sciences. E25-526, MIT, Cambridge, MA 02139
 A Model of Recurrent Interactions in Primary Visual Cortex Emanuel Todorov, Athanassios Siapas and David Somers Dept. of Brain and Cognitive Sciences E25-526, MIT, Cambridge, MA 2139 Email: femo,thanos,somersg@ai.mit.edu
A Model of Recurrent Interactions in Primary Visual Cortex Emanuel Todorov, Athanassios Siapas and David Somers Dept. of Brain and Cognitive Sciences E25-526, MIT, Cambridge, MA 2139 Email: femo,thanos,somersg@ai.mit.edu
Neurons! John A. White Dept. of Bioengineering
 Neurons! John A. White Dept. of Bioengineering john.white@utah.edu What makes neurons different from cardiomyocytes? Morphological polarity Transport systems Shape and function of action potentials Neuronal
Neurons! John A. White Dept. of Bioengineering john.white@utah.edu What makes neurons different from cardiomyocytes? Morphological polarity Transport systems Shape and function of action potentials Neuronal
Ube3a is required for experience-dependent maturation of the neocortex
 Ube3a is required for experience-dependent maturation of the neocortex Koji Yashiro, Thorfinn T. Riday, Kathryn H. Condon, Adam C. Roberts, Danilo R. Bernardo, Rohit Prakash, Richard J. Weinberg, Michael
Ube3a is required for experience-dependent maturation of the neocortex Koji Yashiro, Thorfinn T. Riday, Kathryn H. Condon, Adam C. Roberts, Danilo R. Bernardo, Rohit Prakash, Richard J. Weinberg, Michael
Competing Frameworks in Perception
 Competing Frameworks in Perception Lesson II: Perception module 08 Perception.08. 1 Views on perception Perception as a cascade of information processing stages From sensation to percept Template vs. feature
Competing Frameworks in Perception Lesson II: Perception module 08 Perception.08. 1 Views on perception Perception as a cascade of information processing stages From sensation to percept Template vs. feature
Competing Frameworks in Perception
 Competing Frameworks in Perception Lesson II: Perception module 08 Perception.08. 1 Views on perception Perception as a cascade of information processing stages From sensation to percept Template vs. feature
Competing Frameworks in Perception Lesson II: Perception module 08 Perception.08. 1 Views on perception Perception as a cascade of information processing stages From sensation to percept Template vs. feature
What do you notice? Edited from
 What do you notice? Edited from https://www.youtube.com/watch?v=ffayobzdtc8&t=83s How can a one brain region increase the likelihood of eliciting a spike in another brain region? Communication through
What do you notice? Edited from https://www.youtube.com/watch?v=ffayobzdtc8&t=83s How can a one brain region increase the likelihood of eliciting a spike in another brain region? Communication through
The Visual System. Cortical Architecture Casagrande February 23, 2004
 The Visual System Cortical Architecture Casagrande February 23, 2004 Phone: 343-4538 Email: vivien.casagrande@mcmail.vanderbilt.edu Office: T2302 MCN Required Reading Adler s Physiology of the Eye Chapters
The Visual System Cortical Architecture Casagrande February 23, 2004 Phone: 343-4538 Email: vivien.casagrande@mcmail.vanderbilt.edu Office: T2302 MCN Required Reading Adler s Physiology of the Eye Chapters
Active Control of Spike-Timing Dependent Synaptic Plasticity in an Electrosensory System
 Active Control of Spike-Timing Dependent Synaptic Plasticity in an Electrosensory System Patrick D. Roberts and Curtis C. Bell Neurological Sciences Institute, OHSU 505 N.W. 185 th Avenue, Beaverton, OR
Active Control of Spike-Timing Dependent Synaptic Plasticity in an Electrosensory System Patrick D. Roberts and Curtis C. Bell Neurological Sciences Institute, OHSU 505 N.W. 185 th Avenue, Beaverton, OR
Synaptic Plasticity and Memory
 Synaptic Plasticity and Memory Properties and synaptic mechanisms underlying the induction of long-term potentiation (LTP) The role of calcium/calmodulin-dependent kinase II (CamKII) in the induction,
Synaptic Plasticity and Memory Properties and synaptic mechanisms underlying the induction of long-term potentiation (LTP) The role of calcium/calmodulin-dependent kinase II (CamKII) in the induction,
Spatial coding of position and orientation in primary visual cortex
 and displacement of population activity for small changes in stimulus position, factors that would limit reliable population coding of stimulus position. Second, how uniform is the size of the population
and displacement of population activity for small changes in stimulus position, factors that would limit reliable population coding of stimulus position. Second, how uniform is the size of the population
Dynamics of Hodgkin and Huxley Model with Conductance based Synaptic Input
 Proceedings of International Joint Conference on Neural Networks, Dallas, Texas, USA, August 4-9, 2013 Dynamics of Hodgkin and Huxley Model with Conductance based Synaptic Input Priyanka Bajaj and Akhil
Proceedings of International Joint Conference on Neural Networks, Dallas, Texas, USA, August 4-9, 2013 Dynamics of Hodgkin and Huxley Model with Conductance based Synaptic Input Priyanka Bajaj and Akhil
THE DISCHARGE VARIABILITY OF NEOCORTICAL NEURONS DURING HIGH-CONDUCTANCE STATES
 Neuroscience 119 (2003) 855 873 THE DISCHARGE VARIABILITY OF NEOCORTICAL NEURONS DURING HIGH-CONDUCTANCE STATES M. RUDOLPH* AND A. DESTEXHE Unité de Neuroscience Intégratives et Computationnelles, CNRS,
Neuroscience 119 (2003) 855 873 THE DISCHARGE VARIABILITY OF NEOCORTICAL NEURONS DURING HIGH-CONDUCTANCE STATES M. RUDOLPH* AND A. DESTEXHE Unité de Neuroscience Intégratives et Computationnelles, CNRS,
A neural circuit model of decision making!
 A neural circuit model of decision making! Xiao-Jing Wang! Department of Neurobiology & Kavli Institute for Neuroscience! Yale University School of Medicine! Three basic questions on decision computations!!
A neural circuit model of decision making! Xiao-Jing Wang! Department of Neurobiology & Kavli Institute for Neuroscience! Yale University School of Medicine! Three basic questions on decision computations!!
Modulators of Spike Timing-Dependent Plasticity
 Modulators of Spike Timing-Dependent Plasticity 1 2 3 4 5 Francisco Madamba Department of Biology University of California, San Diego La Jolla, California fmadamba@ucsd.edu 6 7 8 9 10 11 12 13 14 15 16
Modulators of Spike Timing-Dependent Plasticity 1 2 3 4 5 Francisco Madamba Department of Biology University of California, San Diego La Jolla, California fmadamba@ucsd.edu 6 7 8 9 10 11 12 13 14 15 16
Dual Inhibitory Mechanisms for Definition of Receptive Field Characteristics in Cat Striate Cortex
 Dual Inhibitory Mechanisms for Definition of Receptive Field Characteristics in Cat Striate Cortex A. B. Bonds Dept. of Electrical Engineering Vanderbilt University N ashville, TN 37235 Abstract In single
Dual Inhibitory Mechanisms for Definition of Receptive Field Characteristics in Cat Striate Cortex A. B. Bonds Dept. of Electrical Engineering Vanderbilt University N ashville, TN 37235 Abstract In single
Department of Numerical Analysis and Computer Science, Royal Institute of Technology, S Stockholm, Sweden, 2
 The Journal of Neuroscience, February, 2002, 22(3):08 097 Simulations of the Role of the Muscarinic-Activated Calcium- Sensitive Nonspecific Cation Current I NCM in Entorhinal Neuronal Activity during
The Journal of Neuroscience, February, 2002, 22(3):08 097 Simulations of the Role of the Muscarinic-Activated Calcium- Sensitive Nonspecific Cation Current I NCM in Entorhinal Neuronal Activity during
Lab 4: Compartmental Model of Binaural Coincidence Detector Neurons
 Lab 4: Compartmental Model of Binaural Coincidence Detector Neurons Introduction The purpose of this laboratory exercise is to give you hands-on experience with a compartmental model of a neuron. Compartmental
Lab 4: Compartmental Model of Binaural Coincidence Detector Neurons Introduction The purpose of this laboratory exercise is to give you hands-on experience with a compartmental model of a neuron. Compartmental
Informationsverarbeitung im zerebralen Cortex
 Informationsverarbeitung im zerebralen Cortex Thomas Klausberger Dept. Cognitive Neurobiology, Center for Brain Research, Med. Uni. Vienna The hippocampus is a key brain circuit for certain forms of memory
Informationsverarbeitung im zerebralen Cortex Thomas Klausberger Dept. Cognitive Neurobiology, Center for Brain Research, Med. Uni. Vienna The hippocampus is a key brain circuit for certain forms of memory
Electrophysiological and firing properties of neurons: categorizing soloists and choristers in primary visual cortex
 *Manuscript Click here to download Manuscript: Manuscript revised.docx Click here to view linked Referenc Electrophysiological and firing properties of neurons: categorizing soloists and choristers in
*Manuscript Click here to download Manuscript: Manuscript revised.docx Click here to view linked Referenc Electrophysiological and firing properties of neurons: categorizing soloists and choristers in