Two new lipid-dependent Malassezia species from domestic animals. Universitat Autònoma de Barcelona, Bellaterra, Barcelona, E Spain.
|
|
|
- Clara Hardy
- 5 years ago
- Views:
Transcription
1 Two new lipid-dependent Malassezia species from domestic animals. F. Javier Cabañes 1*, Bart Theelen 2, Gemma Castellá 1 and Teun Boekhout 2. 1 Grup de Micologia Veterinària. Departament de Sanitat i d Anatomia Animals, Universitat Autònoma de Barcelona, Bellaterra, Barcelona, E Spain. 2 Centraalbureau voor Schimmelcultures, Utrecht, The Netherlands *corresponding author Abstract During a study on the occurrence of lipid-dependent Malassezia spp. in domestic animals some atypical strains, phylogenetically related to Malassezia sympodialis Simmons et Guého, were revealed to represent novel species. In the present study we describe two new taxa, Malassezia caprae sp. nov. (type strain MA383 = CBS 10434) isolated mainly from goats and Malassezia equina sp. nov. (type strain MA146 = CBS 9969) isolated mainly from horses, including their morphological and physiological characteristics. The validation of these new taxa is further supported by analysis of the D1/D2 regions of 26S rdna, the ITS1-5.8S-ITS2 rdna, the RNA polymerase subunit 1 (RPB1) and chitin synthase nucleotide sequences and by analysis of the amplified fragment length polymorphism (AFLP) patterns, which were all consistent in separating these new species from the other species of the genus, and the M. sympodialis species cluster specifically. Keywords: Malassezia caprae, Malassezia equina, taxonomy, yeast, domestic animals, skin, asexual speciation 1
2 Introduction Since the genus Malassezia was created by Baillon in 1889, its taxonomy has been a matter of controversy. The genus remained limited to M. furfur and M. pachydermatis for a long time (Batra et al., 2005). Traditionally, the lipid-dependent species M. furfur (sensu lato) was thought to occur only on human skin, while the lipophilic, but non lipid-dependent, species M. pachydermatis was restricted to animal skin. Malassezia sympodialis, a lipid-dependent species described in 1990 (Simmons & Guého, 1990) was the third species accepted in the genus, a century after of the description of M. furfur. Afterwards, the genus Malassezia was revised on the basis of morphological, physiological and rrna sequencing studies and four new lipid-dependent species were described: M. globosa, M. obtusa, M. restricta and M. slooffiae (Guého et al., 1996). More recently, another four new lipid-dependent Malassezia species have been described, namely M. dermatis (Sugita et al., 2002), M. japonica (Sugita et al., 2003), M. nana (Hirai et al., 2004) and M. yamatoensis (Sugita et al., 2004). Malassezia pachydermatis is frequently found on wild and domestic carnivores and rarely on humans (Guillot & Bond, 1999). Lipid-dependent Malassezia yeasts have also been isolated from healthy dogs and cats (Bond et al., 1996; Bond et al., 1997; Crespo et al., 1999; Crespo et al., 2002a) and from the healthy skin of horses and different domestic ruminants, being the major component of the lipophilic mycobiota in these later animals (Crespo et al., 2002b). Some of these isolates from horses and ruminants could not be identified because the different physiological tests results and their morphological characteristics precluded fitting them into any of the previously 2
3 described species of the genus. A new species, tentatively named M. equi, was reported from normal equine skin (Nell et al., 2002), but without including a valid description, nor a type specimen. It was identified by 26S rdna D1/D2 sequence analysis as a member of the genus Malassezia, and was found to be most closely related to M. sympodialis. Unfortunately, the only strain that was deposited in the NCYC yeast collection (Norwich, UK) is not alive anymore (C. Bond, personal communication). Crespo et al. (Crespo et al., 2000a) reported for the first time lipid-dependent yeasts associated with otitis externa in cats having similar morphological characteristics and some shared physiological characteristics with the type strain of M. sympodialis. Recently, Hirai et al. (Hirai et al., 2004) described M. nana, a novel species from otic discharges of a cat and cows, which are also closely related to M. sympodialis. However, the difficulty to obtain a high level of certainty in the identification of some of these lipid-dependent strains using physiological tests has been also reported (Crespo et al., 2002b; Gupta et al., 2004; Batra et al., 2005). The speciation of lipid-dependent isolates from animals by means of physiological tests presents some difficulties and some of them can not even be identified (Duarte et al., 1999; Crespo et al., 2000a; Crespo et al., 2002b Duarte et al., 2002). Recently, some lipid-dependent strains similar to the M. sympodialis type strain and isolated from various domestic animal species were studied using DNA sequence analysis and their phylogenetic relationships with the M. sympodialis related species, M. dermatis and M. nana, were discussed (Cabañes et al., 2005). Phylogenetic analysis of both the D1/D2 regions of 26S rdna and ITS-5.8S rdna sequences showed 4 distinct clades. One cluster included isolates from different domestic animal species and the type culture of M. sympodialis that originated from 3
4 humans. The remaining three clusters included isolates from cats, grouping together with the M. nana AB sequence and isolates from horses and goats, respectively. Based on a polyphasic approach, we describe here two new lipid-dependent species in the genus Malassezia, Malassezia caprae sp. nov and Malassezia equina sp. nov., isolated mainly from healthy skin of goats and horses, respectively. Material and methods Strains The strains examined corresponding to the new species are listed in Table 1. Each strain was isolated from a single animal and mainly from healthy skin of the ears from goats and from the healthy skin of the anus from horses. They are from a survey carried out in the Autonomous University of Barcelona (Spain) in the years 1997 and 1998 (Crespo et al. 2002b). Type strains and other strains included in this study are listed also in this table. The strains were stored at 80ºC (Crespo et al., 2000b). Morphological and physiological characterization The characterization of lipid-dependent yeasts was based on the inability to grow on Sabouraud glucose agar (SGA) and on the ability to use certain polyoxyethylene sorbitanesters (Tweens 20, 40, 60 and 80), following the current identification scheme of species described by Guého et al. (Guého et al., 1996) and the Tween diffusion test proposed by Guillot et al. (Guillot et al., 1996). The Cremophor EL assimilation test 4
5 (Mayser et al., 1997) and the splitting of esculin (β-glucosidase activity) (Mayser et al., 1997; Guého et al., 1998) were used as additional key characters. Other tests, such as the catalase reaction, growth at different temperatures (32ºC, 37ºC and 40ºC) on modified Dixon agar (mda) (36 g malt extract; 6 g peptone, 20 g desiccated ox-bile; 10 ml Tween 40; 2 ml glycerol; 2 ml oleic acid and 12 g agar per litre, ph 6.0) and the morphological characteristics after incubation at 32ºC for 7 days in the same culture medium were also performed (Guého et al., 1996). D1/D2 26S rdna and ITS-5.8S rdna sequencing and analysis Methods to isolate the DNA and sequencing of the D1/D2 domain of the 26S rdna and the ITS regions and the 5.8S rdna were similar to those described previously (Cabañes et al., 2005). Cells were harvested from 4- to 5-day-old cultures in modified Dixon s medium. The cells were incubated for 1h at 65 C in 500 µl of extraction buffer (50 mm Tris-HCl, 50 mm EDTA, 3% sodium dodecyl sulfate, and 1% 2-mercaptoethanol). The lysate was extracted with phenol-chloroform-isoamyl alcohol (25:24:1, v/v/v). Then 65 µl of 3Msodium acetate and 75 µl of 1M NaCl were added to 350 µl of the supernatant, and the resulting volume was incubated at 4 C for 30min. DNA was recovered by isopropanol precipitation and washed with 70% (v/v) ethanol, dried under a vacuum, and resuspended in TE buffer (10 mm Tris-HCl, 1 mm EDTA, ph 8). DNA was cleaned with the Geneclean kit II (BIO 101, Inc., La Jolla, Calif.) according to the manufacturer s instructions. ITS rdna and 5.8S rdna were amplified using a Perkin Elmer 2400 thermal cycler and primer pairs ITS5 and ITS4 (White et al., 1990). PCR consisted of a pre- 5
6 denaturation step at 94ºC for 5 min, followed by 35 cycles of denaturation at 95ºC for 30 s, annealing at 50ºC for 1 min and extension at 72ºC for 1 min, plus a final extension of 7 min at 72ºC. The molecular masses of the amplified DNA were estimated by comparison with a 100-bp DNA ladder (Bio-Rad Laboratories S.A., Barcelona, Spain). The PCR product was purified with the GFX PCR DNA and gel band purification kit (Amersham Pharmacia Biotech, Uppsala, Sweden), following the supplier s protocol and purified PCR products were used as a template for sequencing. The protocol BigDye Terminator v3.1 Cycle Sequencing kit (Applied Biosystems, Nieuwerkerk aan de IJsel, The Netherlands) was used for sequencing. The primers ITS5 and ITS4 described by White et al. (White et al., 1990) were used in the sequencing reaction and an Applied Biosystems 3100 sequencer was used to obtain the DNA sequences. The sequences were aligned by using the software program Clustal X (1.81). The Mega package, version 2.1, was used to perform a neighbor joining analysis of a distance matrix (Kimura 2-parameter model, transition to tranversion rate: 2.0) with 1000 bootstrap replicates and a maximum parsimony analysis. Chitin synthase and RNA polymerase subunit 1 (RPB1) sequence analysis The chitin synthase gene was amplified using the primers ChiSyn2f (5 - CTG AAG CTT ACN ATG TAY AAY GAR GAY) and ChiSyn2r (5 -GTT CTC GAG YTT RTA YTC RAA RTT YTG) (Aizawa et al., 1999) in 50 μl reaction volumes containing 3 mm MgCl 2, 200 μm of each dntp, 1 μm of each primer and 1U DNA Taq polymerase (Bioline, Gentaur, Brussels, Belgium) and 1 µl of isolated genomic DNA. The 6
7 following PCR conditions were used: initial denaturation of 5 min at 96ºC, followed by 35 cycles each with a denaturation step of 45 sec at 96ºC, annealing of 1 min at 54ºC, an elongation step of 2 min at 72ºC and a final elongation step of 6 min at 72ºC. For the RPB1 gene, primers RPB1-Af (5 GAR TGY CCD GGD CAY TTY GG) and RPB1-Cr (5 CC NGC DAT NTC RTT RTC CAT RTA) (see were used in a reaction mixture as described (Matheny et al., 2002). The gene fragment was amplified using the following conditions: initial denaturation of 5 min at 96ºC, followed by 35 cycles each with a denaturation step of 30 sec at 96ºC, annealing of 30 sec at 59ºC, an elongation step of 2 min at 72ºC and a final elongation step of 6 min at 72ºC. Amplicons were purified using the GFX Tm PCR DNA purification kit (Amersham Pharmacia Biotech, Roosendaal, The Netherlands). One to ten ng of the purified PCR products were used in the cycle sequencing reaction in a total volume of 10 μl, containing 3 μl 5x sequencing buffer and 1 μl BigDye Tm terminator mix, v3.1 (both from Applied Biosystems) and 400 nm primer. The sequencing primers were the same as for the PCR-reactions. Sequence amplicons were purifed using the MultiScreen Tm Filtration System (Millipore, Etten- Leur, The Netherlands) in combination with Sephadex Tm G-50 Super fine (Amersham Pharmacia Biotech). The sequences were size fractionated on an ABI 3700 capillary sequencer (Applied Biosystems) and were analysed using the Lasergene software package (DNASTAR Inc., Madison, Wisconsin, U.S.A.). Phylogenetic trees were generated using PAUP * version 4.0b10 for Macintosh (Swofford, 2002). Neighbour joining analysis was performed with the uncorrected ( p ) substitution model, alignment gaps were treated as missing data 7
8 and all characters were unordered and of equal weight. For parsimony analysis gaps were treated as missing data and all characters were unordered and of equal weight. The heuristic search was performed with 1000 random taxa additions and tree bisection and reconstruction (TBR) as the branch-swapping algorithm. Branches of zero length were collapsed and all equally parsimonious trees were saved. The robustness of the obtained trees was evaluated by 1000 bootstrap replications. Other statistic measures included tree length, consistency index, retention index and rescaled consistence index (TL, CI, RI and RC). Amplified fragment length polymorphism analysis. AFLP analysis was performed according to the manufacturer s instructions in the AFLP microbial fingerprinting protocol (Applied Biosystems), with some modifications (Gupta et al., 2004). Restriction and ligation were performed simultaneously on 10 ng of genomic DNA by using 1 U of MseI, 5 U of EcoRI, and 3 U of T4 DNAligase (Biolabs, Westburg, The Netherlands). The sequences of the primers EcoRI and MseI were 5 -GACTGCGTACCAATTCAC-3 and 5 -GATGAGTCCTGAGTAAC-3, respectively. The adaptors used were EcoRI (5 -CTCGTAGACTGCGTACC-3, forward; 3 -CATCTGACGCATGGTTAA-5, reverse) and MseI (5 - GACGATGAGTCCTGAG-3, forward; 3 -CTACTCAGGACTCAT-5, reverse). The reaction took place in a total volume of 5.5 µl with the following constituents: a 0.36 µm concentration of the EcoRI adaptor and a 3.64 µm concentration of the MseI adaptor from the AFLP microbial fingerprinting kit, 0.1 M NaCl, 0.91 mm Tris-HCl (ph 7.8), 0.18 mm MgCl 2, 0.18 mm dithiothreitol, 18 µm ATP, and µg of bovine serum albumin ml -1. The restriction ligation mixture was incubated for 2 h at 37 C and 8
9 later diluted by adding 25 µl of sterile double-distilled water. The first PCR was performed with two preselective primers (EcoRI core sequence and MseI core sequence) and the AFLP amplification core mix from the AFLP microbial fingerprinting kit, according to the manufacturer s manual, under the following conditions: 2 min at 72 C, followed by 20 cycles of 20 s at 94 C, 30 s at 56 C, and 2 min at 72 C each. The PCR product was diluted by adding 25 µl of sterile doubledistilled water. The second PCR used more-selective primers, EcoRI-A FAM and MseI- G. The conditions were 2 min at 94 C; 10 cycles consisting of 20 s at 94 C, 30 s at 66 C (decreasing 1 C every step of the cycle), and 2 min at 72 C; and then 25 cycles consisting of 20 s at 94 C, 30 s at 56 C, and 2 min at 72 C. The samples were prepared for acrylamide capillary electrophoresis with the following loading mix: 2.0 µl of selective amplification product, 24 µl of deionised formamide, and 1 µl of GeneScan- 500 labelled with 6-carboxy-X-rhodamine (Applied Biosystems) as an internal size standard. After incubation for 5 min at 95 C, the samples were run on an ABI 310 genetic analyser for 30 min each. Data were analysed with the Bionumerics software package (version 2.5; Applied Maths, Kortrijk, Belgium), by using (i) Pearson correlation based on similarities of the densitometric curves and (ii) the unweighted pair group method with arithmetic means analysis (UPGMA). Results and discussion Morphology and physiology Malassezia isolates belonging to the new species were characterized by using the current morphological and physiological identification scheme (Table 2). The 9
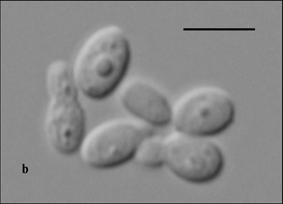
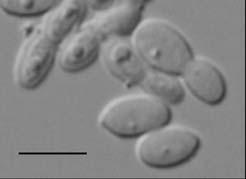
10 phenotypical characteristics of the new species, M. caprae and M. equina, and the other described Malassezia species are summarized in Table 3. The isolates belonging to the new species did not growth in SGA without any lipid supplementation. In general, they grew very slowly and formed small colonies (< mm in diameter; average diameter of M. caprae =1 mm; average diameter of M. equina =1.3 mm ) on mda at 32 ºC after 7 days of incubation. After 21 days of incubation at the same temperature, colonies reached 3-6 mm of diameter. All the isolates of M. equina and two isolates belonging to M. caprae grew slowly at 37º C. None of these isolates grew at 40 ºC, thus differing from other M. sympodialis related species, such M. dermatis, M. nana or M. sympodialis, which can grow at this temperature. Cells were ellipsoidal to subglobose in M. caprae (Fig. 1, Table 2) and mainly ovoidal in M. equina (Fig. 2, Table 2). Special micromorphological characteristics have been cited for some Malassezia spp. In the case of M. furfur the micromorphology appears to be variable in size and shape, including oval, cylindrical or spherical cells, with buds formed on a broad base (Guého et al., 1996). On the contrary, M. globosa has spherical cells with buds formed on a narrow base. M. sympodialis-related species are known to have a small cell size in comparison to other Malassezia spp. (Crespo et al., 2000a; Hirai et al., 2004) and buds formed on a narrow base (Simmons & Guého, 1990; Guého et al., 1996; Crespo et al., 2000a; Hirai et al., 2004). Occasionally, sympodial budding (Simmons & Guého, 1990; Guého et al., 1996; Crespo et al., 2000a) has been reported. However, the separation of Malassezia species based on morphological characteristics may be considered to be subjective (Guého et al., 1996) or unreliable (Guillot & Guého, 1995). 10
11 Although the two new species had similar Tween assimilation profiles to M. sympodialis and M. nana, the isolates analysed in the present study did not completely fit the assimilation profiles of any described species and, hence, could not be identified (Table 2). Following the Tween dilution test proposed by Guého et al. (Guého et al., 1996) the isolates grew poorly on glucose-peptone agar with 0.5% Tween 40, 0.5% Tween 60, 0.1% Tween 80, and they did not grow on 10% Tween 20. In the Tween diffusion test proposed by Guillot et al. (Guillot et al., 1996) most of the isolates showed inhibition areas around the Tweens 40, 60 and 80. These inhibition areas were wider around the Tween 20 wells and in most cases the isolates did not grow around this compound. These inhibition areas are related to the toxic effects of these compounds at higher concentrations. In fact, the initial poor growth that these isolates showed on culture media for lipid-dependent species, containing different Tweens or other lipidic sources, such as mda, may be related to their fungistatic properties. None of the isolates grew around Cremophor EL. All, except one (MA 125) of the M. caprae isolates showed a strong β-glucosidase activity, which was revealed by the splitting of esculin. On the contrary, most of the M. equina isolates were β-glucosidase negative. Among other differences, the isolates belonging to the new species can be distinguished from M. pachydermatis by their inability to grow in SGA; from M. dermatis, M. furfur, M. slooffiae and M. sympodialis by their inability to grow at 40º C; from M. japonica by their ability to assimilate Tween 80; from M. yamatoensis by their inability to assimilate Tween 20; from M. obtusa and M. globosa by their ability to assimilate Tween 40 and tween 60; from M. restricta by their catalase activity and from M. nana by their inability to assimilate Tween 20, by their inability to grow at 40º C and because they showed poor or no growth at 37º C. 11
12 Molecular analysis Based on the sequence divergence observed in the D1/D2 domains of the 26S rdna (Fig. 3), the ITS regions and the 5.8S rdna (Fig. 4), as well as the chitin synthase (Fig. 5) and RPB1 genes (Fig. 6) we concluded that M. sympodialis represents a species complex. Full concordance was observed with clustering of the isolates using the above mentioned partial genome sequences as well as the AFLP analysis (Fig. 7). Here we formally describe two of these species. Molecular sequences and AFLP data for species included in Figures 3-7 were compared to confirm that the isolates studied were distinct from the other species of the genus and represent undescribed species. Figure 3 shows the molecular phylogenetic tree based on the D1/D2 regions of the 26S rdna sequences constructed by the neighbor-joining method. Figure 4 shows the molecular phylogenetic tree based on the ITS1-5.8S-ITS2 sequences. The isolates belonging to the new proposed species formed a cluster with M. sympodialis, M. dermatis and M. nana. The isolates belonging to the novel proposed species Malassezia caprae had identical D1/D2 sequences. Dissimilarities between M. caprae strains and M. sympodialis CBS 7222 T, M. dermatis CBS 9169 T and M. nana CBS 9557 T in their D1/D2 sequences sequences were 1.5%,1.8% and 2.8%, respectively. Their ITS1-5.8S-ITS2 rdna sequences were also identical having dissimilarities between M. caprae strains and M. sympodialis CBS 7222 T, M. dermatis CBS 9169 T and M. nana CBS 9557 T of 6.5%, 3.4% and 9.9%, respectively. 12
13 Isolates from the novel proposed species Malassezia equina showed nearly identical D1/D2 and ITS sequences, thus indicating that these are conspecific strains. Dissimilarities between M. equina CBS 9969 T and the M. sympodialis, M. dermatis and M. nana type strains in the D1/D2 regions of 26S and ITS1-5.8S-ITS2 were 1.3% and 9.1%, 1.3% and 6.7%, and 3.5% and 12.2%, respectively. The sequences of M. equi AJ (Nell et al., 2002) and M. equina CBS 9969 T were identical, but unfortunately, we were not able to analyse M. equi ITS1-5.8S-ITS2 sequences, because there is no such sequence deposited in the GenBank and, furthermore, no M. equi type strain is preserved in culture collections. In each novel species, the strains were found to be closely related to each other. Phylogenetic analysis of sequences from these novel species showed that they were clearly distinct from the other eleven described Malassezia species, exceeding the variation generally observed to occur between species (Scorzetti et al., 2002). The clades obtained with the analysed strains of M. caprae and M. equina using chitin synthase (Fig. 5) and RPB1 (Fig. 6) sequences are also close to those of M. sympodialis. These sequences showed the following dissimilarity between M. sympodialis and those from M. caprae: 1.6% and 9.4% respectively and from M. equina: 19.7% and 12.9%, respectively). The same sequences showed that M. dermatis differs from M. caprae by 7.5% and 18% respectively, and from M. equina by 12.2% and 4.5%, respectively. Those from M. nana differs from M. caprae by 12.2% and 14.2% respectively and from M. equina by 17% and 14%, respectively. Therefore, these data clearly support the distinction of our new species from the remaining species of the genus Malassezia. 13
14 The UPGMA dendrogram (Fig. 7) calculated from the AFLP fingerprints obtained from the different Malassezia strains, clearly differentiated the strains belonging to the proposed novel species M. caprae and M. equina from the rest of the species belonging to the genus Malassezia. The similarities in the AFLP profile among the analyzed M. caprae and M. equina strains were 89.8 % and 95.7 %, respectively. On the other hand, the similarity of these two novel species in comparison with other species in the genus Malassezia was 62.7% for M. caprae with its closest relative M. sympodialis, and 13.0% between M. equina and all other Malassezia species. Mechanisms of divergence All four targeted genome regions supported the sympodialis-lineage within Malassezia with high statistical support, thus indicating the reliability of our analysis. The five species, M. nana, M. dermatis, M. sympodialis, M. equina and M. caprae, formed all separate and well supported clades in the analysis of each molecular marker, as well as in the AFLP analysis. However, within the sympodialis-lineage, the topology of the species was not concordant between the four markers investigated. Three main topologies were observed, with ITS-5.8S rdna and RPB1 supporting the same topology, and both D1/D2 and chitin synthase supporting alternative topologies. Interestingly, all these topologies received high nodal support. In case of speciation through clonal divergence and genetic drift, probably followed by some host adaptation, one would expect concordance between the phylogenetic patterns of each individual gene. This clearly is not the case, and the lack of concordance may indicate that probably recombination has played a role in the divergence of these species. This is particularly interesting as sexual reproduction is unknown in Malassezia. However, 14
15 recombination has been suggested to occur in M. pachydermatis based on isozyme analyses (Midreuil et al., 1999) and in the M. furfur complex a putative hybrid genotype has been observed (R. Batra and T. Boekhout, unpubl. observ.), thus suggesting that cell fusion, karyogamy and meiosis may be possible within the genus. Interestingly, within the sympodialis-lineage, the cat and cow-associated M. nana was found to be a basal species in all cases. The subsequent ingroup lineage was found to be M. equina (ITS-5.8S rdna) or M. caprae (D1/D2), and these analyses placed the two species from human hosts, M. sympodialis and M. dermatis, together with isolates from goats (M. caprae). The D1/D2 analysis, in contrast, placed the two human-associated species with M. equina, a horse-associated species, whereas the chitin synthase, placed M. sympodialis as a sister group to M. caprae, and M. dermatis to M. equina. Therefore, all our data support that a cat and cow-associated species (i.e. M. nana) formed a basal lineage to the other species. Moreover, the data also support that host shifts from animals to humans may have occurred more than once. In order to better understand the mechanism of speciation in relation to host jumps in this interesting asexual and clinically important group of yeasts, a considerable effort is needed to sequence more loci across the known biodiversity of the genus. This is even more true as one has to include the other lineages known to exist within Malassezia (e.g. the furfur-lineage, the globosa-lineage, and M. slooffiae) as the exact infrageneric relationships among these lineages is not yet clear from our data. To summarize this molecular analysis we point out that members of these new species form two well-supported clades using comparative analysis of five molecular markers. However, the differences found in the different genes analysed among the strains under 15
16 study support the recognition of two distinct species, for which the names Malassezia caprae sp. nov and Malassezia equina sp. nov., are proposed. Latin diagnosis of Malassezia caprae Cabañes et Boekhout, sp. nov. Cultura in agaro Dixonii post 7 dies ad 32º C albida vel cremea, glabra, lucida aut hebetata, butyracea, moderate convexa, margine expresso (1 mm). Cellulae ovoidae aut globosae, x μm, e base angusta gemmantes. In agaro glucoso-peptonico Tween 40 (0.5%), Tween 60 (0.5%) et Tween 80 (0.1%) addito paulum crescit. In agaro glucoso-peptonico Tween 20 (10%) addito non crescit. 37ºC non vel paulum crescit neque 40ºC. Teleomorphis ignota. Typus CBS (MA383 = JCM 14561); isolatus ex cute caprina; depositus in collectione zymotica Centraalbureau voor Schimmelcultures, Utrecht, The Netherlands. Description of Malassezia caprae Cabañes & Boekhout, sp. nov. Malassezia caprae (caprae- this Latin derived species epithet refers to the the host animal from which the yeast was first isolated [ca prae. L.fem. n. capra goat; L. fem. gen. n. caprae of a goat] On mda, after 7 days at 32ºC, colonies are small (average diameter 1 mm, < mm), whitish to cream-coloured, smooth, glistening or dull, butyrous and moderately convex with entire margins. Cells are ovoidal to spherical, x μm, with buds formed monopolarly on a narrow base. No growth is obtained on SGA. Catalase reaction is positive and β-glucosidase activity is usually positive, except for isolate MA 16
17 125. No growth occurs on glucose-peptone agar with 10% Tween 20. Poor growth is observed on glucose-peptone agar with Tween 40 (0.5%), Tween 60 (0.5%) and Tween 80 (0.1%). No growth is observed on glucose-peptone agar with Cremophor EL. No or weak growth appears at 37ºC and no growth occurs at 40ºC. The teleomorph is unknown. The type strain CBS (= JCM 14561; originally strain MA383) was isolated from healthy skin of the ear of a goat in Barcelona, Spain. The strains were deposited in the Collection of the Centraalbureau voor Schimmelcultures, Utrecht, The Netherlands and in the Japan Collection of Microorganisms, Saitama, Japan, as CBS and JCM 14561, respectively. Latin diagnosis of Malassezia equina Cabañes et Boekhout, sp. nov. Cultura in agaro Dixonii post 7 dies ad 32º C albida vel cremea, glabra, lucida aut hebetata, butyracea, moderate convexa, margine expresso (1.3 mm). Cellulae ovoidae x μm, e base angusta gemmantes. In agaro glucoso-peptonico Tween 40 (0.5%), Tween 60 (0.5%) et Tween 80 (0.1%) addito paulum crescit. In agaro glucosopeptonico Tween 20 (10%) addito non crescit. 37ºC paulum crescit. 40ºC non crescit. Teleomorphis ignota. Typus CBS 9969 (MA146 = JCM 14562); isolatus ex cute equina; depositus in collectione zymotica Centraalbureau voor Schimmelcultures, Utrecht, The Netherlands. Description of Malassezia equina Cabañes & Boekhout, sp. nov. 17
18 Malassezia equina (equina- this Latin derived species epithet refers to the the host animal from which the yeast was first isolated [e.quin a. L.adj. equina of horses] On mda, after 7 days at 32ºC, colonies are small (average diameter 1.3 mm, range <0.5-2 mm), whitish to cream-coloured, smooth, glistening to dull, butyrous and moderately convex with an entire margin. Cells are ovoidal, x μm, with buds formed monopolarly on a narrow base. No growth is obtained on SGA. Catalase reaction is positive and the β-glucosidase activity is usually negative. No growth occurs on glucose-peptone agar with 10% Tween 20. Poor growth is observed on glucosepeptone agar with Tween 40 (0.5%), Tween 60 (0.5%) and Tween 80 (0.1%). No growth is observed on glucose-peptone agar with Cremophor EL. Poor growth appears at 37ºC and no growth occurs at 40ºC. The teleomorph is unknown. The type strain CBS 9969 (=JCM 14562; originally strain MA146) was isolated from healthy skin of the anus of a horse in Barcelona, Spain. The strains were deposited in the Collection of the Centraalbureau voor Schimmelcultures, Utrecht, The Netherlands and in the Japan Collection of Microorganisms, Saitama, Japan, as CBS 9969 and JCM 14562, respectively. Nell et al. (Nell et al., 2002) reported the presence of a novel Malassezia species from normal equine skin, which they tentatively named Malassezia equi, but without including a valid description. It was identified by 26S rdna D1/D2 sequence analysis as a member of the genus Malassezia, and was found to be most closely related to M. sympodialis. The D1/D2 sequences of M. equi (AJ305330) (Nell et al., 2002) and of the type species of M. equina (CBS 9969 T = MA146) are identical (Cabañes et al., 2005), so they are very related organisms, and probably conspecific. We were not able 18
19 to analyze the M. equi ITS sequences or any other gene sequences because there is no other sequence deposited in GenBank. Moreover, M. equi was not formally been described (e.g. no latin diagnosis and no type strain indicated) and data on morphological and physiological description, such as growth on the various Tweens, esculin and cremophor EL, were not provided in the description. Furthermore, no strain has been preserved for this taxon. For these reasons this species (Nell et al., 2002) is an invalidly described species that, consequently does not exist. Therefore the name M. equi Nell et al. can be considered as non-existent. To avoid any future confusion we decided to provide our species the epithet equina. ACKNOWLEDGMENTS This work was supported by grants NL-TAF-864, SYNTHESYS project of the European Commission s Research Infrastructure Action (EU) and 2005SGR00684 from the DURSI, Generalitat de Catalunya, Spain. References Aizawa T, Kano R, Nakamura Y, Watanabe S, Hasegawa A (1999) Molecular heterogeneity in clinical isolates of Malassezia pachydermatis from dogs. Vet Microbiol 70: Batra R, Boekhout T, Guého E, Cabañes FJ, Dawson TL & Gupta AK (2005) Malassezia Baillon, emerging clinical yeasts. FEMS Yeast Res 5:
20 Bond R, Anthony RM, Dodd M & Lloyd DH (1996) Isolation of Malassezia sympodialis from feline skin. J Med Vet Mycol 34: Bond R, Howell SA, Haywood PJ & Lloyd DH (1997) Isolation of Malassezia sympodialis and Malassezia globosa from healthy pet cats. Vet Rec 141: Cabañes FJ, Hernández JJ & Castellá G (2005) Molecular analysis of Malassezia sympodialis related strains from domestic animals. J Clin Microbiol 43: Crespo MJ, Abarca ML & Cabanes FJ (1999) Isolation of Malassezia furfur from a cat. J Clin Microbiol 37: Crespo MJ, Abarca ML & Cabanes FJ (2000a) Otitis externa associated with Malassezia sympodialis in two cats. J Clin Microbiol 38: Crespo MJ, Abarca ML & Cabanes FJ (2000b) Evaluation of different preservation and storage methods for Malassezia spp. J Clin Microbiol 38: Crespo MJ, Abarca ML & Cabanes FJ (2002a) Occurrence of Malassezia spp. in the external ear canals of dogs and cats with and without otitis externa. Med Mycol 40: Crespo MJ, Abarca ML & Cabanes FJ (2002b) Occurrence of Malassezia spp. in horses and domestic ruminants. Mycoses 45: Duarte ER, Melo MM, Hahn RC & Hamdan JS (1999) Prevalence of Malassezia spp. in the ears of asymptomatic cattle and cattle with otitis in Brazil. Med Mycol 37: Duarte ER, Lachance MA & Hamdan JS (2002) Identification of atypical strains of Malassezia spp. from cattle and dog. Can J Microbiol 48:
21 Guého E, Midgley G & Guillot J (1996) The genus Malassezia with the description of four new species. Antonie van Leeuwenhoek 69: Guého E, Boekhout T, Ashbee HR, Guillot J, van Belkum A & Faergemann J (1998) The role of Malassezia species in the ecology of human skin and as pathogens. Med. Mycol. 36, Guillot J & Bond R (1999) Malassezia pachydermatis: a review. Med Mycol 37: Guillot J & Guého E (1995) The diversity of Malassezia yeasts confirmed by rrna sequence and nuclear DNA comparisons. Antonie van Leeuwenhoek 67: Guillot J, Guého E, Lesourd M, Midgley G, Chevrier G & Dupont B (1996). Identification of Malassezia species, a practical approach. J Mycol Méd 6: Gupta AK, Boekhout T, Theelen B, Summerbell RC & Batra R (2004) Identification and typing of Malassezia species by amplified fragment length polymorphism (AFLP) and sequence analyses of the internal transcribed spacer (ITS) and large subunit (LSU) regions of ribosomal DNA. J Clin Microbiol 42: Hirai A, Kano R, Makimura K et al. (2004) Malassezia nana sp. nov., a novel lipiddependent yeast species isolated from animals. Int J Syst Evol Microbiol 54: Matheny PB, Liu YJ, Ammirati JF & Hall BD (2002) Using RPB1 sequences to improve phylogenetic inference among mushrooms (Inocybe, Agaricales). Am J Bot 89:
22 Mayser P, Haze P, Papavassilis C, Pickel M, Gruender K & Guého E (1997) Differentiation of Malassezia species: selectivity of cremophor EL, castor oil and ricinoleic acid for M. furfur. Br J Dermatol 137: Midreuil F, Guillot J, Guého E, Renaud F, Mallie M & Bastide JM (1999) Genetic diversity in the yeast species Malassezia pachydermatis analysed by multilocus enzyme electrophoresis. Int J Syst Bacteriol 49: Nell A, James SA, Bond CJ, Hunt B & Herrtage ME (2002) Identification and distribution of a novel Malassezia species yeast on normal equine skin. Veter Rec 150: Scorzetti G, Fell JW, Fonseca A & Statzell-Tallman, A (2002) Systematics of basidiomycetous yeasts: a comparison of large subunit D1/D2 and internal transcribed spacer rdna regions. FEMS Yeasts Res 2: Simmons RB & Guého E (1990) A new species of Malassezia. Mycol Res 94: Sugita T, Takashima M, Shinoda T et al. (2002) New yeast species Malassezia dermatis isolated from patients with atopic dermatitis. J Clin Microbiol 40: Sugita T, Takashima M, Kodama M, Tsuboi R & Nishikawa A (2003) Description of a new yeast species, Malassezia japonica, and its detection in patients with atopic dermatitis and healthy subjects. J Clin Microbiol 41: Sugita T, Tajima M, Takashima M, Amaya M, Saito M, Tsuboi R & Nishikawa A (2004) A new yeast, Malassezia yamatoensis, isolated from a patient with seborrheic 22
23 dermatitis, and its distribution in patients and healthy subjects. Microbiol Immunol 48: Swofford, D.L. PAUP *. Phylogenetic Analysis Using Parsimony ( * And Other Methods). Version 4.0b10, (2002), Sinauer Associates, Sunderland, MA. USA. White TJ, Bruns T, Lee S & Taylor J (1990) Amplification and direct sequencing of fungi ribosomal RNA genes for phylogenetics, PCR protocols. A guide to methods and applications (Innis MA, Gelfand DH, Sninsky JJ & White TJ, eds), pp Academic Press, San Diego, CA. USA. 23
24 Figure 1. Cells of M. caprae a) CBS T (MA383) and b) CBS 9973 (MA400) cultured on mda at 32ºC for 7 days. Bar, 4 μm. Figure 2. Cells of M. equina CBS 9969 T (MA146) cultured on mda at 32ºC for 7 days. Bar, 4 μm. Figure 3. Molecular phylogenetic tree constructed using the sequences of D1/D2 26S rdna of members of the genus Malassezia species. The numbers at branch point are the percentages of 1,000 bootstrapped data sets that supported the specific internal branches. Outgroup: Filobasidiella neoformans CBS 132 T. Species with GenBank numbers represent sequences obtained from GenBank. Figure 4. Molecular phylogenetic tree constructed using the sequences of ITS-5.8S rdna gene sequences of members of the genus Malassezia. The numbers at branch point are the percentages of 1,000 bootstrapped data sets that supported the specific internal branches. Outgroup: Cryptococcus neoformans CBS 132 T. Species with GenBank numbers represent sequences obtained from GenBank. Figure 5. Molecular phylogenetic tree constructed using the sequences of chitin synthase gene sequences of members of the genus Malassezia. The numbers at branch point are the percentages of 1,000 bootstrapped data sets that supported the specific internal branches. Outgroup: Cryptococcus neoformans JEC 21. Species with GenBank numbers represent sequences obtained from GenBank. Figure 6. Molecular phylogenetic tree constructed using the sequences of RNA polymerase subunit 1 gene sequences of members of the genus Malassezia. The numbers at branch point are the percentages of 1,000 bootstrapped data sets that supported the specific internal branches. Outgroup: Cryptococcus neoformans JEC 20. Species with GenBank numbers represent sequences obtained from GenBank. Figure 7. UPGMA dendrogram assessed from the comparison of AFLP fingerprints of Malassezia species. 24
25 Table 1. Strains analyzed and their hosts. Strain Host M. caprae CBS T (MA383) Goat M. caprae CBS 9967 (MA80) Goat M. caprae CBS 9973 (MA400) Goat M. caprae MA125 Horse M. caprae MA333 Goat M. dermatis CBS 9169 T Human M. furfur CBS 1878 a Human M. furfur CBS 7019 NT Human M. equina CBS 9969 T (MA146) Horse M. equina CBS 9986 (MA88) Cow M. equina MA250 Horse M. equina MA461 Horse M. equina MA470 Horse M. globosa CBS 7966 T Human M. japonica CBS 9431 T Human M. japonica CBS 9432 Human M. nana CBS 9557 T Cat M. nana CBS 9558 Cow M. nana CBS 9561 Cow M. obtusa CBS 7876 T Human M. obtusa CBS 7968 Human M. pachydermatis CBS 1879 NT Dog M. pachydermatis CBS 1919 Dog M. pachydermatis CBS 4165 Dog M. restricta CBS 7877 T Human M. slooffiae CBS 7956 T Pig M. sympodialis CBS 7222 T Human M. sympodialis CBS 7978 Human M. sympodialis CBS 7979 Human M. sympodialis CBS 8740 Human M. sympodialis CBS 9968 (MA73) Sheep M. sympodialis CBS 9970 Horse M. sympodialis CWB1 Human M. yamatoensis CBS 9725 T Human a M. furfur CBS 1878 is the neotype of Pityrosporum ovale 25

26 Figure 1. Figure 2. 26
Tween 40-based precipitate production observed on modified chromogenic agar and development of biological identification kit for Malassezia species
 Medical Mycology May 2006, 44, 227231 Tween 40-based precipitate production observed on modified chromogenic agar and development of biological identification kit for Malassezia species TAKAMASA KANEKO*$,
Medical Mycology May 2006, 44, 227231 Tween 40-based precipitate production observed on modified chromogenic agar and development of biological identification kit for Malassezia species TAKAMASA KANEKO*$,
Malassezia, 2. Malassezia
 Jpn. J. Med. Mycol. Vol. 46, 163 167, 2005 ISSN 0916 4804 Malassezia Malassezia,,, SD, AD, Malassezia. SD, AD Malassezia PCR. AD, SD M. globosa, M. restricta,. AD 20, 2, Malassezia., 2 Malassezia 90, 70.
Jpn. J. Med. Mycol. Vol. 46, 163 167, 2005 ISSN 0916 4804 Malassezia Malassezia,,, SD, AD, Malassezia. SD, AD Malassezia PCR. AD, SD M. globosa, M. restricta,. AD 20, 2, Malassezia., 2 Malassezia 90, 70.
Physiological and Molecular Characterization of Atypical Isolates of Malassezia ACCEPTED
 JCM Accepts, published online ahead of print on October 00 J. Clin. Microbiol. doi:./jcm.01-0 Copyright 00, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.
JCM Accepts, published online ahead of print on October 00 J. Clin. Microbiol. doi:./jcm.01-0 Copyright 00, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.
Molecular Characterization of Malassezia Species Isolated from Dog with and Without Otitis and Seborrhoeic Dermatitis
 World Journal of Zoology 6 (2): 134-141, 2011 ISSN 1817-3098 IDOSI Publications, 2011 Molecular Characterization of Malassezia Species Isolated from Dog with and Without Otitis and Seborrhoeic Dermatitis
World Journal of Zoology 6 (2): 134-141, 2011 ISSN 1817-3098 IDOSI Publications, 2011 Molecular Characterization of Malassezia Species Isolated from Dog with and Without Otitis and Seborrhoeic Dermatitis
New Yeast Species, Malassezia dermatis, Isolated from Patients with Atopic Dermatitis
 JOURNAL OF CLINICAL MICROBIOLOGY, Apr. 2002, p. 1363 1367 Vol. 40, No. 4 0095-1137/02/$04.00 0 DOI: 10.1128/JCM.40.4.1363 1367.2002 Copyright 2002, American Society for Microbiology. All Rights Reserved.
JOURNAL OF CLINICAL MICROBIOLOGY, Apr. 2002, p. 1363 1367 Vol. 40, No. 4 0095-1137/02/$04.00 0 DOI: 10.1128/JCM.40.4.1363 1367.2002 Copyright 2002, American Society for Microbiology. All Rights Reserved.
Comparison of Nested PCR and RFLP for Identification and Classification of Malassezia Yeasts from Healthy Human Skin
 Ann Dermatol Vol. 21, No. 4, 2009 ORIGINAL ARTICLE Comparison of Nested PCR and RFLP for Identification and Classification of Malassezia Yeasts from Healthy Human Skin Byung Ho Oh, M.D., Young Chan Song,
Ann Dermatol Vol. 21, No. 4, 2009 ORIGINAL ARTICLE Comparison of Nested PCR and RFLP for Identification and Classification of Malassezia Yeasts from Healthy Human Skin Byung Ho Oh, M.D., Young Chan Song,
Identi cation of Malassezia species isolated from patients with seborrhoeic dermatitis, atopic dermatitis, pityriasis versicolor and normal subjects
 Medical Mycology 2000, 38, 337 341 Accepted 28 February 2000 Identi cation of Malassezia species isolated from patients with seborrhoeic dermatitis, atopic dermatitis, pityriasis versicolor and normal
Medical Mycology 2000, 38, 337 341 Accepted 28 February 2000 Identi cation of Malassezia species isolated from patients with seborrhoeic dermatitis, atopic dermatitis, pityriasis versicolor and normal
Molecular Analysis of Malassezia Microflora on the Skin of the Patients with Atopic Dermatitis
 Ann Dermatol Vol. 22, No. 1, 2010 DOI: 10.5021/ad.2010.22.1.41 ORIGINAL ARTICLE Molecular Analysis of Malassezia Microflora on the Skin of the Patients with Atopic Dermatitis Seon Mi Yim, M.D. 1, Ji Young
Ann Dermatol Vol. 22, No. 1, 2010 DOI: 10.5021/ad.2010.22.1.41 ORIGINAL ARTICLE Molecular Analysis of Malassezia Microflora on the Skin of the Patients with Atopic Dermatitis Seon Mi Yim, M.D. 1, Ji Young
Occurrence of Malassezia spp. in the external ear canals of dogs and cats with and without otitis externa
 Ó Medical Mycology 2002, 40, 115 121 Accepted 21 February 2001 Occurrence of Malassezia spp. in the external ear canals of dogs and cats with and without otitis externa M. J. CRESPO, M. L. ABARCA & F.
Ó Medical Mycology 2002, 40, 115 121 Accepted 21 February 2001 Occurrence of Malassezia spp. in the external ear canals of dogs and cats with and without otitis externa M. J. CRESPO, M. L. ABARCA & F.
NOTES. Received 8 December 2004/Returned for modification 4 March 2005/Accepted 14 April 2005
 JOURNAL OF CLINICAL MICROBIOLOGY, Aug. 2005, p. 4147 4151 Vol. 43, No. 8 0095-1137/05/$08.00 0 doi:10.1128/jcm.43.8.4147 4151.2005 Copyright 2005, American Society for Microbiology. All Rights Reserved.
JOURNAL OF CLINICAL MICROBIOLOGY, Aug. 2005, p. 4147 4151 Vol. 43, No. 8 0095-1137/05/$08.00 0 doi:10.1128/jcm.43.8.4147 4151.2005 Copyright 2005, American Society for Microbiology. All Rights Reserved.
Vital growth factors of Malassezia species on modified CHROMagar Candida
 Medical Mycology December 2005, 43, 699 /704 Vital growth factors of Malassezia species on modified CHROMagar Candida TAKAMASA KANEKO*$, KOICHI MAKIMURA$*, MASANOBU ONOZAKI*$, KUMIKO UEDA$, YOHKO YAMADA$,
Medical Mycology December 2005, 43, 699 /704 Vital growth factors of Malassezia species on modified CHROMagar Candida TAKAMASA KANEKO*$, KOICHI MAKIMURA$*, MASANOBU ONOZAKI*$, KUMIKO UEDA$, YOHKO YAMADA$,
Received 4 February 2002/Returned for modification 3 April 2002/Accepted 2 June 2002
 JOURNAL OF CLINICAL MICROBIOLOGY, Sept. 2002, p. 3350 3357 Vol. 40, No. 9 0095-1137/02/$04.00 0 DOI: 10.1128/JCM.40.9.3350 3357.2002 Copyright 2002, American Society for Microbiology. All Rights Reserved.
JOURNAL OF CLINICAL MICROBIOLOGY, Sept. 2002, p. 3350 3357 Vol. 40, No. 9 0095-1137/02/$04.00 0 DOI: 10.1128/JCM.40.9.3350 3357.2002 Copyright 2002, American Society for Microbiology. All Rights Reserved.
Seborrheic dermatitis: predisposing factors and ITS2 secondary structure for Malassezia phylogenic analysis
 Medical Mycology November 2013, 51, 868 875 Seborrheic dermatitis: predisposing factors and ITS2 secondary structure for Malassezia phylogenic analysis YULIEN AMADO *, ANELVI PATI Ñ O-UZC Á TEGUI *, MARIA
Medical Mycology November 2013, 51, 868 875 Seborrheic dermatitis: predisposing factors and ITS2 secondary structure for Malassezia phylogenic analysis YULIEN AMADO *, ANELVI PATI Ñ O-UZC Á TEGUI *, MARIA
Molecular Biological Identification of Malassezia Yeasts Using Pyrosequencing
 Ann Dermatol Vol. 25, No. 1, 2013 http://dx.doi.org/10.5021/ad.2013.25.1.73 ORIGINAL ARTICLE Molecular Biological Identification of Malassezia Yeasts Using Pyrosequencing Ji Young Kim, Hyung Jin Hahn,
Ann Dermatol Vol. 25, No. 1, 2013 http://dx.doi.org/10.5021/ad.2013.25.1.73 ORIGINAL ARTICLE Molecular Biological Identification of Malassezia Yeasts Using Pyrosequencing Ji Young Kim, Hyung Jin Hahn,
Tomasz Jagielski 1*,Elżbieta Rup 2, Aleksandra Ziółkowska 1, Katarzyna Roeske 1, Anna B Macura 2 and Jacek Bielecki 1
 Jagielski et al. BMC Dermatology 2014, 14:3 RESEARCH ARTICLE Open Access Distribution of Malassezia species on the skin of patients with atopic dermatitis, psoriasis, and healthy volunteers assessed by
Jagielski et al. BMC Dermatology 2014, 14:3 RESEARCH ARTICLE Open Access Distribution of Malassezia species on the skin of patients with atopic dermatitis, psoriasis, and healthy volunteers assessed by
Evaluation of Expression of Lipases and Phospholipases of Malassezia restricta in Patients with Seborrheic Dermatitis
 Ann Dermatol Vol. 25, No. 3, 2013 http://dx.doi.org/10.5021/ad.2013.25.3.310 ORIGINAL ARTICLE Evaluation of Expression of Lipases and Phospholipases of Malassezia restricta in Patients with Seborrheic
Ann Dermatol Vol. 25, No. 3, 2013 http://dx.doi.org/10.5021/ad.2013.25.3.310 ORIGINAL ARTICLE Evaluation of Expression of Lipases and Phospholipases of Malassezia restricta in Patients with Seborrheic
Update on the genus Malassezia
 Medical Mycology June 2007, 45, 287303 Review Update on the genus Malassezia H. R. ASHBEE Mycology Reference Centre, Department of Microbiology, Leeds General Infirmary, Leeds, UK Malassezia yeasts are
Medical Mycology June 2007, 45, 287303 Review Update on the genus Malassezia H. R. ASHBEE Mycology Reference Centre, Department of Microbiology, Leeds General Infirmary, Leeds, UK Malassezia yeasts are
Shivaprakash M Rudramurthy Additional Professor, Department of Medical Microbiology, PGIMER, Chandigarh
 Shivaprakash M Rudramurthy Additional Professor, Department of Medical Microbiology, PGIMER, Chandigarh Commensal Unipolar budding Lipophilic Resides in area rich in sebaceous gland Early difficulty in
Shivaprakash M Rudramurthy Additional Professor, Department of Medical Microbiology, PGIMER, Chandigarh Commensal Unipolar budding Lipophilic Resides in area rich in sebaceous gland Early difficulty in
Table S1. Primers and PCR protocols for mutation screening of MN1, NF2, KREMEN1 and ZNRF3.
 Table S1. Primers and PCR protocols for mutation screening of MN1, NF2, KREMEN1 and ZNRF3. MN1 (Accession No. NM_002430) MN1-1514F 5 -GGCTGTCATGCCCTATTGAT Exon 1 MN1-1882R 5 -CTGGTGGGGATGATGACTTC Exon
Table S1. Primers and PCR protocols for mutation screening of MN1, NF2, KREMEN1 and ZNRF3. MN1 (Accession No. NM_002430) MN1-1514F 5 -GGCTGTCATGCCCTATTGAT Exon 1 MN1-1882R 5 -CTGGTGGGGATGATGACTTC Exon
Susceptibility pattern of Malassezia species to selected plant extracts and antifungal agents
 Original Article Susceptibility pattern of Malassezia species to selected plant extracts and antifungal agents G. Sibi, Md. Ansar Alam, Jhanvi Shah, Masna Razak Department of Biotechnology, Indian Academy
Original Article Susceptibility pattern of Malassezia species to selected plant extracts and antifungal agents G. Sibi, Md. Ansar Alam, Jhanvi Shah, Masna Razak Department of Biotechnology, Indian Academy
New genomic typing method MLST
 New genomic typing method MLST Bon KIMURA fingerprinting PFGE DNA multilocus sequence typingmlst alleles PFGE MLST 1990 PCR 1 PCR DNA PFGE 1 PFGE RAPDrandomly amplified polymorphic DNA 3 AFLPAmplified
New genomic typing method MLST Bon KIMURA fingerprinting PFGE DNA multilocus sequence typingmlst alleles PFGE MLST 1990 PCR 1 PCR DNA PFGE 1 PFGE RAPDrandomly amplified polymorphic DNA 3 AFLPAmplified
in the Gastrointestinal and Reproductive Tracts of Quarter Horse Mares
 Influence of Probiotics on Microflora in the Gastrointestinal and Reproductive Tracts of Quarter Horse Mares Katie Barnhart Research Advisors: Dr. Kimberly Cole and Dr. John Mark Reddish Department of
Influence of Probiotics on Microflora in the Gastrointestinal and Reproductive Tracts of Quarter Horse Mares Katie Barnhart Research Advisors: Dr. Kimberly Cole and Dr. John Mark Reddish Department of
Product Manual. Omni-Array Sense Strand mrna Amplification Kit, 2 ng to 100 ng Version Catalog No.: Reactions
 Genetic Tools and Reagents Universal mrna amplification, sense strand amplification, antisense amplification, cdna synthesis, micro arrays, gene expression, human, mouse, rat, guinea pig, cloning Omni-Array
Genetic Tools and Reagents Universal mrna amplification, sense strand amplification, antisense amplification, cdna synthesis, micro arrays, gene expression, human, mouse, rat, guinea pig, cloning Omni-Array
Malassezia furfur in Infantile Seborrheic Dermatitis
 ASIAN PACIFIC JOURNAL OF ALLERGY AND IMMUNOLOGY (2005) 23: 101-105 Malassezia furfur in Infantile Seborrheic Dermatitis Siriwan Wananukul 1, Ariya Chindamporn 2, Poomjit Yumyourn 2, Sunchai Payungporn
ASIAN PACIFIC JOURNAL OF ALLERGY AND IMMUNOLOGY (2005) 23: 101-105 Malassezia furfur in Infantile Seborrheic Dermatitis Siriwan Wananukul 1, Ariya Chindamporn 2, Poomjit Yumyourn 2, Sunchai Payungporn
Hepatitis B Virus Genemer
 Product Manual Hepatitis B Virus Genemer Primer Pair for amplification of HBV Viral Specific Fragment Catalog No.: 60-2007-10 Store at 20 o C For research use only. Not for use in diagnostic procedures
Product Manual Hepatitis B Virus Genemer Primer Pair for amplification of HBV Viral Specific Fragment Catalog No.: 60-2007-10 Store at 20 o C For research use only. Not for use in diagnostic procedures
OVERVIEW OF CURRENT IDENTIFICATION SYSTEMS AND DATABASES
 OVERVIEW OF CURRENT IDENTIFICATION SYSTEMS AND DATABASES EVERY STEP OF THE WAY 1 EVERY STEP OF THE WAY MICROBIAL IDENTIFICATION METHODS DNA RNA Genotypic Sequencing of ribosomal RNA regions of bacteria
OVERVIEW OF CURRENT IDENTIFICATION SYSTEMS AND DATABASES EVERY STEP OF THE WAY 1 EVERY STEP OF THE WAY MICROBIAL IDENTIFICATION METHODS DNA RNA Genotypic Sequencing of ribosomal RNA regions of bacteria
Relation of Malassezia spp. with steroids acne and folliculitis
 Relation of Malassezia spp. with steroids acne and folliculitis Azhar A. F. Al-Attraqhchi 1 Abbas M. M. Al-Ammari 2 and Mona Al-Jibouri 2 College of Medicine/Al-Nahrain University/Dep. of Medical Microbiology
Relation of Malassezia spp. with steroids acne and folliculitis Azhar A. F. Al-Attraqhchi 1 Abbas M. M. Al-Ammari 2 and Mona Al-Jibouri 2 College of Medicine/Al-Nahrain University/Dep. of Medical Microbiology
Multi-clonal origin of macrolide-resistant Mycoplasma pneumoniae isolates. determined by multiple-locus variable-number tandem-repeat analysis
 JCM Accepts, published online ahead of print on 30 May 2012 J. Clin. Microbiol. doi:10.1128/jcm.00678-12 Copyright 2012, American Society for Microbiology. All Rights Reserved. 1 2 Multi-clonal origin
JCM Accepts, published online ahead of print on 30 May 2012 J. Clin. Microbiol. doi:10.1128/jcm.00678-12 Copyright 2012, American Society for Microbiology. All Rights Reserved. 1 2 Multi-clonal origin
Identification of Yeasts. Medical Mycology Training Network 15 November 2018 Dr Tan Ai Ling Department of Microbiology, Singapore General Hospital
 Identification of Yeasts Medical Mycology Training Network 15 November 2018 Dr Tan Ai Ling Department of Microbiology, Singapore General Hospital Definition of Yeasts Eukaryote cells have defined nucleus
Identification of Yeasts Medical Mycology Training Network 15 November 2018 Dr Tan Ai Ling Department of Microbiology, Singapore General Hospital Definition of Yeasts Eukaryote cells have defined nucleus
ITS accuracy at GenBank. Conrad Schoch Barbara Robbertse
 ITS accuracy at GenBank Conrad Schoch Barbara Robbertse Improving accuracy Barcode tag in GenBank Barcode submission tool Standards RefSeq Targeted Loci Well validated sequences already in GenBank Bacteria
ITS accuracy at GenBank Conrad Schoch Barbara Robbertse Improving accuracy Barcode tag in GenBank Barcode submission tool Standards RefSeq Targeted Loci Well validated sequences already in GenBank Bacteria
IL-1, IL-6, IL-8, MCP-1, TNF- ELISA. 1 Candida albicans ,,,, C. albicans., Clonetics BioWhittaker 5% CO 2. TLD-0142, TLD-0143, 37 C, 3,
 Jpn. J. Med. Mycol. Vol. 45, 131 136, 2004 ISSN 0916 4804,,.,,.,,,, IL-1, IL-6, IL-8, MCP-1, TNF- ELISA., IL-8, IL-8, TNF-, IL-1, IL-6, IL-8, TNF-.,,,,,.,. key word: cytokine, keratinocyte, ELISA enzyme-linked
Jpn. J. Med. Mycol. Vol. 45, 131 136, 2004 ISSN 0916 4804,,.,,.,,,, IL-1, IL-6, IL-8, MCP-1, TNF- ELISA., IL-8, IL-8, TNF-, IL-1, IL-6, IL-8, TNF-.,,,,,.,. key word: cytokine, keratinocyte, ELISA enzyme-linked
a) SSR with core motif > 2 and repeats number >3. b) MNR with repeats number>5.
 SSR with core motif > 2 and repeats number >3. b) MNR with repeats number>5.") 1 2 APPENDIX Legends to figures 3 4 5 Figure A1: Distribution of perfect SSR along chromosome 1 of V. cholerae (El-Tor N191). a) SSR with core motif > 2 and repeats number >3. b) MNR with repeats number>5.
1 2 APPENDIX Legends to figures 3 4 5 Figure A1: Distribution of perfect SSR along chromosome 1 of V. cholerae (El-Tor N191). a) SSR with core motif > 2 and repeats number >3. b) MNR with repeats number>5.
First Report of Penicillium adametzioides from Decayed Grapes (Vitis vinifera) in Pakistan
 in Pakistan") International Journal of Current Microbiology and Applied Sciences ISSN: 2319-7706 Volume 5 Number 12 (2016) pp. 316-320 Journal homepage: http://www.ijcmas.com Original Research Article http://dx.doi.org/10.20546/ijcmas.2016.512.034
International Journal of Current Microbiology and Applied Sciences ISSN: 2319-7706 Volume 5 Number 12 (2016) pp. 316-320 Journal homepage: http://www.ijcmas.com Original Research Article http://dx.doi.org/10.20546/ijcmas.2016.512.034
Role of Paired Box9 (PAX9) (rs ) and Muscle Segment Homeobox1 (MSX1) (581C>T) Gene Polymorphisms in Tooth Agenesis
 (rs ) and Muscle Segment Homeobox1 (MSX1) (581C>T) Gene Polymorphisms in Tooth Agenesis") EC Dental Science Special Issue - 2017 Role of Paired Box9 (PAX9) (rs2073245) and Muscle Segment Homeobox1 (MSX1) (581C>T) Gene Polymorphisms in Tooth Agenesis Research Article Dr. Sonam Sethi 1, Dr. Anmol
EC Dental Science Special Issue - 2017 Role of Paired Box9 (PAX9) (rs2073245) and Muscle Segment Homeobox1 (MSX1) (581C>T) Gene Polymorphisms in Tooth Agenesis Research Article Dr. Sonam Sethi 1, Dr. Anmol
INSTRUCTION MANUAL. RNA Clean & Concentrator -5 Catalog Nos. R1015 & R1016. Highlights. Contents
 INSTRUCTION MANUAL Catalog Nos. R1015 & R1016 Highlights Quick (5 minute) method for cleaning and concentrating RNA. Ideal for purification of RNA from aqueous phase following an acid phenol extraction.
INSTRUCTION MANUAL Catalog Nos. R1015 & R1016 Highlights Quick (5 minute) method for cleaning and concentrating RNA. Ideal for purification of RNA from aqueous phase following an acid phenol extraction.
Molecular Analysis of Fungal Microbiota in Samples from Healthy Human Skin and Psoriatic Lesions
 JOURNAL OF CLINICAL MICROBIOLOGY, Aug. 2006, p. 2933 2941 Vol. 44, No. 8 0095-1137/06/$08.00 0 doi:10.1128/jcm.00785-06 Copyright 2006, American Society for Microbiology. All Rights Reserved. Molecular
JOURNAL OF CLINICAL MICROBIOLOGY, Aug. 2006, p. 2933 2941 Vol. 44, No. 8 0095-1137/06/$08.00 0 doi:10.1128/jcm.00785-06 Copyright 2006, American Society for Microbiology. All Rights Reserved. Molecular
Molecular Identification of Lipase Producing Bacteria based on 16S rdna Sequencing
 International Journal of Current Microbiology and Applied Sciences ISSN: 2319-7706 Volume 6 Number 5 (2017) pp. 2067-2071 Journal homepage: http://www.ijcmas.com Original Research Article https://doi.org/10.20546/ijcmas.2017.605.230
International Journal of Current Microbiology and Applied Sciences ISSN: 2319-7706 Volume 6 Number 5 (2017) pp. 2067-2071 Journal homepage: http://www.ijcmas.com Original Research Article https://doi.org/10.20546/ijcmas.2017.605.230
Reassortment of influenza A virus genes linked to PB1 polymerase gene
 International Congress Series 1263 (2004) 714 718 Reassortment of influenza A virus genes linked to PB1 polymerase gene Jean C. Downie* www.ics-elsevier.com Centre for Infectious Diseases and Microbiology,
International Congress Series 1263 (2004) 714 718 Reassortment of influenza A virus genes linked to PB1 polymerase gene Jean C. Downie* www.ics-elsevier.com Centre for Infectious Diseases and Microbiology,
CHAPTER 7: REAGENTS AND SOLUTIONS
 7.1. ANALYSIS OF MODULATION OF SOD ENZYME Acetic acid (cat. no. 11007, Glaxo Qualigen, India): Bovine Serum Albumin stock solution (BSA, 1mg/ml): 1 mg of standard bovine serum albumin (cat. no. A2153,
7.1. ANALYSIS OF MODULATION OF SOD ENZYME Acetic acid (cat. no. 11007, Glaxo Qualigen, India): Bovine Serum Albumin stock solution (BSA, 1mg/ml): 1 mg of standard bovine serum albumin (cat. no. A2153,
Malassezia Species Associated With Dermatitis in Dogs and Their Antifungal Susceptibility
 International Journal of Current Microbiology and Applied Sciences ISSN: 2319-7706 Volume 7 Number 06 (2018) Journal homepage: http://www.ijcmas.com Original Research Article https://doi.org/10.20546/ijcmas.2018.706.236
International Journal of Current Microbiology and Applied Sciences ISSN: 2319-7706 Volume 7 Number 06 (2018) Journal homepage: http://www.ijcmas.com Original Research Article https://doi.org/10.20546/ijcmas.2018.706.236
Chromatin IP (Isw2) Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles.
 Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles.") Chromatin IP (Isw2) 7/01 Toshi last update: 06/15 Reagents Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles. 2.5 M glycine. TBS:
Chromatin IP (Isw2) 7/01 Toshi last update: 06/15 Reagents Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles. 2.5 M glycine. TBS:
Psoriasis and infestation with Malassezia
 Original Research Medical Journal of the Islamic Republic of Iran.Vol. 21, No.1, May 2007. pp. 11-16 Psoriasis and infestation with Malassezia Downloaded from mjiri.iums.ac.ir at 6:14 IRDT on Monday July
Original Research Medical Journal of the Islamic Republic of Iran.Vol. 21, No.1, May 2007. pp. 11-16 Psoriasis and infestation with Malassezia Downloaded from mjiri.iums.ac.ir at 6:14 IRDT on Monday July
Kit Components Product # EP42720 (24 preps) MDx 2X PCR Master Mix 350 µl Cryptococcus neoformans Primer Mix 70 µl Cryptococcus neoformans Positive
 MDx 2X PCR Master Mix 350 µl Cryptococcus neoformans Primer Mix 70 µl Cryptococcus neoformans Positive") 3430 Schmon Parkway Thorold, ON, Canada L2V 4Y6 Phone: 866-667-4362 (905) 227-8848 Fax: (905) 227-1061 Email: techsupport@norgenbiotek.com Cryptococcus neoformans End-Point PCR Kit Product# EP42720 Product
3430 Schmon Parkway Thorold, ON, Canada L2V 4Y6 Phone: 866-667-4362 (905) 227-8848 Fax: (905) 227-1061 Email: techsupport@norgenbiotek.com Cryptococcus neoformans End-Point PCR Kit Product# EP42720 Product
For in vitro Veterinary Diagnostics only. Kylt Rotavirus A. Real-Time RT-PCR Detection.
 For in vitro Veterinary Diagnostics only. Kylt Rotavirus A Real-Time RT-PCR Detection www.kylt.eu DIRECTION FOR USE Kylt Rotavirus A Real-Time RT-PCR Detection A. General Kylt Rotavirus A products are
For in vitro Veterinary Diagnostics only. Kylt Rotavirus A Real-Time RT-PCR Detection www.kylt.eu DIRECTION FOR USE Kylt Rotavirus A Real-Time RT-PCR Detection A. General Kylt Rotavirus A products are
The hair strand testa new method for testing antifungal effects of antidandruff preparations
 j. Cosmet. Sci., 54, 263-270 (May/June 2003) The hair strand testa new method for testing antifungal effects of antidandruff preparations PETER MAYSER, HORST ARGEMBEAUX, and FRANK RIPPKE, Zentrum fiir
j. Cosmet. Sci., 54, 263-270 (May/June 2003) The hair strand testa new method for testing antifungal effects of antidandruff preparations PETER MAYSER, HORST ARGEMBEAUX, and FRANK RIPPKE, Zentrum fiir
7/13/09. Definition. Infections Due to Malassezia. Case Report 1. Case Report 1 (cont.) Case Report 1 (cont.)
 Case Report 1 (cont.)") Definition Infections Due to Malassezia Various species of Malassezia cause both opportunistic, superficial infections and occasionally systemic infections Common superficial infections include: Pityriasis
Definition Infections Due to Malassezia Various species of Malassezia cause both opportunistic, superficial infections and occasionally systemic infections Common superficial infections include: Pityriasis
Protocol for Gene Transfection & Western Blotting
 The schedule and the manual of basic techniques for cell culture Advanced Protocol for Gene Transfection & Western Blotting Schedule Day 1 26/07/2008 Transfection Day 3 28/07/2008 Cell lysis Immunoprecipitation
The schedule and the manual of basic techniques for cell culture Advanced Protocol for Gene Transfection & Western Blotting Schedule Day 1 26/07/2008 Transfection Day 3 28/07/2008 Cell lysis Immunoprecipitation
Isolation and identification of Mycoplasma gallisepticum in chickensbn from industrial farms in Kerman province
 Available online at http://www.ijabbr.com International journal of Advanced Biological and Biomedical Research Volume 2, Issue 1, 2014: 100-104 Isolation and identification of Mycoplasma gallisepticum
Available online at http://www.ijabbr.com International journal of Advanced Biological and Biomedical Research Volume 2, Issue 1, 2014: 100-104 Isolation and identification of Mycoplasma gallisepticum
Cytochalasins from an Australian marine sediment-derived Phomopsis sp. (CMB-M0042F): Acid-mediated intra-molecular cycloadditions enhance
: Acid-mediated intra-molecular cycloadditions enhance") SUPPORTING INFORMATION Cytochalasins from an Australian marine sediment-derived Phomopsis sp. (CMB-M42F): Acid-mediated intra-molecular cycloadditions enhance chemical diversity Zhuo Shang, Ritesh Raju,
SUPPORTING INFORMATION Cytochalasins from an Australian marine sediment-derived Phomopsis sp. (CMB-M42F): Acid-mediated intra-molecular cycloadditions enhance chemical diversity Zhuo Shang, Ritesh Raju,
Oligo Sequence* bp %GC Tm Hair Hm Ht Position Size Ref. HIVrt-F 5 -CTA-gAA-CTT-TRA-ATg-CAT-ggg-TAA-AAg-TA
 Human immunodeficiency virus (HIV) detection & quantitation by qrt-pcr (Taqman). Created on: Oct 26, 2010; Last modified by: Jul 17, 2017; Version: 3.0 This protocol describes the qrt-pcr taqman based
Human immunodeficiency virus (HIV) detection & quantitation by qrt-pcr (Taqman). Created on: Oct 26, 2010; Last modified by: Jul 17, 2017; Version: 3.0 This protocol describes the qrt-pcr taqman based
Human Immunodeficiency Virus-1 (HIV-1) Genemer. Primer Pair for amplification of HIV-1 Specific DNA Fragment
 Genemer. Primer Pair for amplification of HIV-1 Specific DNA Fragment") Product Manual Human Immunodeficiency Virus-1 (HIV-1) Genemer Primer Pair for amplification of HIV-1 Specific DNA Fragment Catalog No.: 60-2002-10 Store at 20 o C For research use only. Not for use in
Product Manual Human Immunodeficiency Virus-1 (HIV-1) Genemer Primer Pair for amplification of HIV-1 Specific DNA Fragment Catalog No.: 60-2002-10 Store at 20 o C For research use only. Not for use in
Principles of phylogenetic analysis
 Principles of phylogenetic analysis Arne Holst-Jensen, NVI, Norway. Fusarium course, Ås, Norway, June 22 nd 2008 Distance based methods Compare C OTUs and characters X A + D = Pairwise: A and B; X characters
Principles of phylogenetic analysis Arne Holst-Jensen, NVI, Norway. Fusarium course, Ås, Norway, June 22 nd 2008 Distance based methods Compare C OTUs and characters X A + D = Pairwise: A and B; X characters
Phylogenetic Methods
 Phylogenetic Methods Multiple Sequence lignment Pairwise distance matrix lustering algorithms: NJ, UPM - guide trees Phylogenetic trees Nucleotide vs. amino acid sequences for phylogenies ) Nucleotides:
Phylogenetic Methods Multiple Sequence lignment Pairwise distance matrix lustering algorithms: NJ, UPM - guide trees Phylogenetic trees Nucleotide vs. amino acid sequences for phylogenies ) Nucleotides:
The Schedule and the Manual of Basic Techniques for Cell Culture
 The Schedule and the Manual of Basic Techniques for Cell Culture 1 Materials Calcium Phosphate Transfection Kit: Invitrogen Cat.No.K2780-01 Falcon tube (Cat No.35-2054:12 x 75 mm, 5 ml tube) Cell: 293
The Schedule and the Manual of Basic Techniques for Cell Culture 1 Materials Calcium Phosphate Transfection Kit: Invitrogen Cat.No.K2780-01 Falcon tube (Cat No.35-2054:12 x 75 mm, 5 ml tube) Cell: 293
iplex genotyping IDH1 and IDH2 assays utilized the following primer sets (forward and reverse primers along with extension primers).
.") Supplementary Materials Supplementary Methods iplex genotyping IDH1 and IDH2 assays utilized the following primer sets (forward and reverse primers along with extension primers). IDH1 R132H and R132L Forward:
Supplementary Materials Supplementary Methods iplex genotyping IDH1 and IDH2 assays utilized the following primer sets (forward and reverse primers along with extension primers). IDH1 R132H and R132L Forward:
Supplementary Information
 Supplementary Information Methods Growth Characterisation Total Adenosine Triphosphate (ATP) was determined using a Molecular Probes ATP Determination Kit (Life Technologies, USA). 0.5 ml of sample was
Supplementary Information Methods Growth Characterisation Total Adenosine Triphosphate (ATP) was determined using a Molecular Probes ATP Determination Kit (Life Technologies, USA). 0.5 ml of sample was
Detection of equine infectious anemia nucleic acid in asymptomatic carrier horses by nested PCR
 Asian Biomedicine Vol. 4 No. 6 December 2010; 971-975 Brief communication (Original) Detection of equine infectious anemia nucleic acid in asymptomatic carrier horses by nested PCR Sunutcha Suntrarachun
Asian Biomedicine Vol. 4 No. 6 December 2010; 971-975 Brief communication (Original) Detection of equine infectious anemia nucleic acid in asymptomatic carrier horses by nested PCR Sunutcha Suntrarachun
HIV-1 Genemer Detection Kit Ready to Use Amplification Kit for HIV-1 Specific DNA Fragment Analysis
 Product Manual HIV-1 Genemer Detection Kit Ready to Use Amplification Kit for HIV-1 Specific DNA Fragment Analysis For research use only. Not for use in diagnostic procedures for clinical purposes Catalog
Product Manual HIV-1 Genemer Detection Kit Ready to Use Amplification Kit for HIV-1 Specific DNA Fragment Analysis For research use only. Not for use in diagnostic procedures for clinical purposes Catalog
Molecular phylogeny of Australian isolates of Sporothrix schenckii sensu lato. David New Microbiology Registrar, PathWest
 Molecular phylogeny of Australian isolates of Sporothrix schenckii sensu lato David New Microbiology Registrar, PathWest Background Sporothrix schenckii causes a cutaneous mycosis Sporotrichosis (aka rose
Molecular phylogeny of Australian isolates of Sporothrix schenckii sensu lato David New Microbiology Registrar, PathWest Background Sporothrix schenckii causes a cutaneous mycosis Sporotrichosis (aka rose
Plasmids Western blot analysis and immunostaining Flow Cytometry Cell surface biotinylation RNA isolation and cdna synthesis
 Plasmids psuper-retro-s100a10 shrna1 was constructed by cloning the dsdna oligo 5 -GAT CCC CGT GGG CTT CCA GAG CTT CTT TCA AGA GAA GAA GCT CTG GAA GCC CAC TTT TTA-3 and 5 -AGC TTA AAA AGT GGG CTT CCA GAG
Plasmids psuper-retro-s100a10 shrna1 was constructed by cloning the dsdna oligo 5 -GAT CCC CGT GGG CTT CCA GAG CTT CTT TCA AGA GAA GAA GCT CTG GAA GCC CAC TTT TTA-3 and 5 -AGC TTA AAA AGT GGG CTT CCA GAG
Prevalence Rate of Malassezia Fungus in Sheep Dogs and Sheep Owners
 World Journal of Zoology 6 (4): 390-394, 2011 ISSN 1817-3098 IDOSI Publications, 2011 Prevalence Rate of Malassezia Fungus in Sheep Dogs and Sheep Owners 1 2 3 Mansoor Khakpoor, Saeid Safarmashaei and
World Journal of Zoology 6 (4): 390-394, 2011 ISSN 1817-3098 IDOSI Publications, 2011 Prevalence Rate of Malassezia Fungus in Sheep Dogs and Sheep Owners 1 2 3 Mansoor Khakpoor, Saeid Safarmashaei and
ab E3 Ligase Auto- Ubiquitilylation Assay Kit
 ab139469 E3 Ligase Auto- Ubiquitilylation Assay Kit Instructions for Use For testing ubiquitin E3 ligase activity through assessment of their ability to undergo auto-ubiquitinylation This product is for
ab139469 E3 Ligase Auto- Ubiquitilylation Assay Kit Instructions for Use For testing ubiquitin E3 ligase activity through assessment of their ability to undergo auto-ubiquitinylation This product is for
Keywords: PRRSV, wild boar, seroprevalence, phylogenetic analyses
 54 EUROPRRS2011, Novi Sad, Serbia PORCINE REPRODUCTIVE AND RESPIRATORY SYNDROME VIRUS (PRRSV) INFECTION IN LITHUANIAN WILD BORS (SUS SCROFA) POPULATION Arunas Stankevicius 1, Jurate Buitkuviene 1, Jurgita
54 EUROPRRS2011, Novi Sad, Serbia PORCINE REPRODUCTIVE AND RESPIRATORY SYNDROME VIRUS (PRRSV) INFECTION IN LITHUANIAN WILD BORS (SUS SCROFA) POPULATION Arunas Stankevicius 1, Jurate Buitkuviene 1, Jurgita
Application of Pressure Cycling Technology (PCT) in Differential Extraction
 in Differential Extraction") Application of Pressure Cycling Technology (PCT) in Differential Extraction Deepthi Nori, MFS; Dr. Bruce R. McCord, PhD Department of Chemistry International Forensic Research Institute Florida International
Application of Pressure Cycling Technology (PCT) in Differential Extraction Deepthi Nori, MFS; Dr. Bruce R. McCord, PhD Department of Chemistry International Forensic Research Institute Florida International
Williams Lab Recipes ANTIBIOTICS
 Williams Lab Recipes ANTIBIOTICS 1000x Ampicillin (sodium salt) 100mg/ml recipe 1. Measure out 1 g of Ampicillin tri hydrate 2. Add Milli-Q H2O to 10 ml 3. Add ~.1 g of NaOH pellets (half pellet or more
Williams Lab Recipes ANTIBIOTICS 1000x Ampicillin (sodium salt) 100mg/ml recipe 1. Measure out 1 g of Ampicillin tri hydrate 2. Add Milli-Q H2O to 10 ml 3. Add ~.1 g of NaOH pellets (half pellet or more
Phosphate buffered saline (PBS) for washing the cells TE buffer (nuclease-free) ph 7.5 for use with the PrimePCR Reverse Transcription Control Assay
 for washing the cells TE buffer (nuclease-free) ph 7.5 for use with the PrimePCR Reverse Transcription Control Assay") Catalog # Description 172-5080 SingleShot Cell Lysis Kit, 100 x 50 µl reactions 172-5081 SingleShot Cell Lysis Kit, 500 x 50 µl reactions For research purposes only. Introduction The SingleShot Cell Lysis
Catalog # Description 172-5080 SingleShot Cell Lysis Kit, 100 x 50 µl reactions 172-5081 SingleShot Cell Lysis Kit, 500 x 50 µl reactions For research purposes only. Introduction The SingleShot Cell Lysis
DELFIA Eu-DTPA ITC Chelate & Europium Standard
 AD0026P-3 (en) 1 DELFIA Eu-DTPA ITC Chelate & AD0021 Europium Standard For Research Use Only INTRODUCTION DELFIA Eu-DTPA ITC Chelate is optimized for the europium labelling of proteins and peptides for
AD0026P-3 (en) 1 DELFIA Eu-DTPA ITC Chelate & AD0021 Europium Standard For Research Use Only INTRODUCTION DELFIA Eu-DTPA ITC Chelate is optimized for the europium labelling of proteins and peptides for
For the 5 GATC-overhang two-oligo adaptors set up the following reactions in 96-well plate format:
 Supplementary Protocol 1. Adaptor preparation: For the 5 GATC-overhang two-oligo adaptors set up the following reactions in 96-well plate format: Per reaction X96 10X NEBuffer 2 10 µl 10 µl x 96 5 -GATC
Supplementary Protocol 1. Adaptor preparation: For the 5 GATC-overhang two-oligo adaptors set up the following reactions in 96-well plate format: Per reaction X96 10X NEBuffer 2 10 µl 10 µl x 96 5 -GATC
Figure S1. Schematic presentation of genomic replication of idsiv after transfection and infection. After transfection of idsiv plasmid DNA into 293T
 Figure S1. Schematic presentation of genomic replication of idsiv after transfection and infection. After transfection of idsiv plasmid DNA into 293T cells, the RNA genomes with all modifications are generated
Figure S1. Schematic presentation of genomic replication of idsiv after transfection and infection. After transfection of idsiv plasmid DNA into 293T cells, the RNA genomes with all modifications are generated
Annexure III SOLUTIONS AND REAGENTS
 Annexure III SOLUTIONS AND REAGENTS A. STOCK SOLUTIONS FOR DNA ISOLATION 0.5M Ethylene-diamine tetra acetic acid (EDTA) (ph=8.0) 1M Tris-Cl (ph=8.0) 5M NaCl solution Red cell lysis buffer (10X) White cell
Annexure III SOLUTIONS AND REAGENTS A. STOCK SOLUTIONS FOR DNA ISOLATION 0.5M Ethylene-diamine tetra acetic acid (EDTA) (ph=8.0) 1M Tris-Cl (ph=8.0) 5M NaCl solution Red cell lysis buffer (10X) White cell
Evolution of hepatitis C virus in blood donors and their respective recipients
 Journal of General Virology (2003), 84, 441 446 DOI 10.1099/vir.0.18642-0 Short Communication Correspondence Jean-François Cantaloube jfc-ets-ap@gulliver.fr Evolution of hepatitis C virus in blood donors
Journal of General Virology (2003), 84, 441 446 DOI 10.1099/vir.0.18642-0 Short Communication Correspondence Jean-François Cantaloube jfc-ets-ap@gulliver.fr Evolution of hepatitis C virus in blood donors
Gastric Carcinoma with Lymphoid Stroma: Association with Epstein Virus Genome demonstrated by PCR
 Gastric Carcinoma with Lymphoid Stroma: Association with Epstein Virus Genome demonstrated by PCR Pages with reference to book, From 305 To 307 Irshad N. Soomro,Samina Noorali,Syed Abdul Aziz,Suhail Muzaffar,Shahid
Gastric Carcinoma with Lymphoid Stroma: Association with Epstein Virus Genome demonstrated by PCR Pages with reference to book, From 305 To 307 Irshad N. Soomro,Samina Noorali,Syed Abdul Aziz,Suhail Muzaffar,Shahid
For Research Use Only Ver
 INSTRUCTION MANUAL Quick-RNA Miniprep Kit Catalog Nos. R1054 & R1055 Highlights High-quality total RNA (including small RNAs) from a wide range of samples. You can opt to isolate small and large RNAs in
INSTRUCTION MANUAL Quick-RNA Miniprep Kit Catalog Nos. R1054 & R1055 Highlights High-quality total RNA (including small RNAs) from a wide range of samples. You can opt to isolate small and large RNAs in
Caution: For Laboratory Use. A product for research purposes only. Eu-W1284 Iodoacetamido Chelate & Europium Standard. Product Number: AD0014
 TECHNICAL DATA SHEET Lance Caution: For Laboratory Use. A product for research purposes only. Eu-W1284 Iodoacetamido Chelate & Europium Standard Product Number: AD0014 INTRODUCTION: Iodoacetamido-activated
TECHNICAL DATA SHEET Lance Caution: For Laboratory Use. A product for research purposes only. Eu-W1284 Iodoacetamido Chelate & Europium Standard Product Number: AD0014 INTRODUCTION: Iodoacetamido-activated
For Research Use Only Ver
 INSTRUCTION MANUAL Quick-RNA Miniprep Kit Catalog Nos. R1054 & R1055 Highlights High-quality total RNA (including small RNAs) from a wide range of samples. You can opt to isolate small and large RNAs in
INSTRUCTION MANUAL Quick-RNA Miniprep Kit Catalog Nos. R1054 & R1055 Highlights High-quality total RNA (including small RNAs) from a wide range of samples. You can opt to isolate small and large RNAs in
DELFIA Tb-N1 DTA Chelate & Terbium Standard
 AD0029P-1 (en) 1 DELFIA Tb-N1 DTA Chelate & AD0012 Terbium Standard For Research Use Only INTRODUCTION DELFIA Tb-N1 DTA Chelate is optimized for the terbium labeling of proteins and peptides for use in
AD0029P-1 (en) 1 DELFIA Tb-N1 DTA Chelate & AD0012 Terbium Standard For Research Use Only INTRODUCTION DELFIA Tb-N1 DTA Chelate is optimized for the terbium labeling of proteins and peptides for use in
Construction of a hepatocellular carcinoma cell line that stably expresses stathmin with a Ser25 phosphorylation site mutation
 Construction of a hepatocellular carcinoma cell line that stably expresses stathmin with a Ser25 phosphorylation site mutation J. Du 1, Z.H. Tao 2, J. Li 2, Y.K. Liu 3 and L. Gan 2 1 Department of Chemistry,
Construction of a hepatocellular carcinoma cell line that stably expresses stathmin with a Ser25 phosphorylation site mutation J. Du 1, Z.H. Tao 2, J. Li 2, Y.K. Liu 3 and L. Gan 2 1 Department of Chemistry,
Supplementary material: Materials and suppliers
 Supplementary material: Materials and suppliers Electrophoresis consumables including tris-glycine, acrylamide, SDS buffer and Coomassie Brilliant Blue G-2 dye (CBB) were purchased from Ameresco (Solon,
Supplementary material: Materials and suppliers Electrophoresis consumables including tris-glycine, acrylamide, SDS buffer and Coomassie Brilliant Blue G-2 dye (CBB) were purchased from Ameresco (Solon,
IDENTIFICATION OF CRYPTOSPORIDIUM PARVUM GENOTYPE FROM HIV AND NON-HIV FECAL SAMPLES BY PCR
 IDENTIFICATION OF CRYPTOSPORIDIUM PARVUM GENOTYPE FROM HIV AND NON-HIV FECAL SAMPLES BY PCR Zulhainan Hamzah 1, Songsak Petmitr 2, Mathirut Mungthin 3 and Porntip Chavalitshewinkoon-Petmitr 1 1 Department
IDENTIFICATION OF CRYPTOSPORIDIUM PARVUM GENOTYPE FROM HIV AND NON-HIV FECAL SAMPLES BY PCR Zulhainan Hamzah 1, Songsak Petmitr 2, Mathirut Mungthin 3 and Porntip Chavalitshewinkoon-Petmitr 1 1 Department
PRODUCT: RNAzol BD for Blood May 2014 Catalog No: RB 192 Storage: Store at room temperature
 PRODUCT: RNAzol BD for Blood May 2014 Catalog No: RB 192 Storage: Store at room temperature PRODUCT DESCRIPTION. RNAzol BD is a reagent for isolation of total RNA from whole blood, plasma or serum of human
PRODUCT: RNAzol BD for Blood May 2014 Catalog No: RB 192 Storage: Store at room temperature PRODUCT DESCRIPTION. RNAzol BD is a reagent for isolation of total RNA from whole blood, plasma or serum of human
Immunology of Diseases Associated with Malassezia Species
 CLINICAL MICROBIOLOGY REVIEWS, Jan. 2002, p. 21 57 Vol. 15, No. 1 0893-8512/02/$04.00 0 DOI: 10.1128/CMR.15.1.21 57.2002 Copyright 2002, American Society for Microbiology. All Rights Reserved. Immunology
CLINICAL MICROBIOLOGY REVIEWS, Jan. 2002, p. 21 57 Vol. 15, No. 1 0893-8512/02/$04.00 0 DOI: 10.1128/CMR.15.1.21 57.2002 Copyright 2002, American Society for Microbiology. All Rights Reserved. Immunology
Product Contents. 1 Specifications 1 Product Description. 2 Buffer Preparation... 3 Protocol. 3 Ordering Information 4 Related Products..
 INSTRUCTION MANUAL Quick-RNA MidiPrep Catalog No. R1056 Highlights 10 minute method for isolating RNA (up to 1 mg) from a wide range of cell types and tissue samples. Clean-Spin column technology allows
INSTRUCTION MANUAL Quick-RNA MidiPrep Catalog No. R1056 Highlights 10 minute method for isolating RNA (up to 1 mg) from a wide range of cell types and tissue samples. Clean-Spin column technology allows
aM (modules 1 and 10 are required)
") This form should be used for all taxonomic proposals. Please complete all those modules that are applicable (and then delete the unwanted sections). For guidance, see the notes written in blue and the
This form should be used for all taxonomic proposals. Please complete all those modules that are applicable (and then delete the unwanted sections). For guidance, see the notes written in blue and the
Islet viability assay and Glucose Stimulated Insulin Secretion assay RT-PCR and Western Blot
 Islet viability assay and Glucose Stimulated Insulin Secretion assay Islet cell viability was determined by colorimetric (3-(4,5-dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide assay using CellTiter
Islet viability assay and Glucose Stimulated Insulin Secretion assay Islet cell viability was determined by colorimetric (3-(4,5-dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide assay using CellTiter
The role of mycological investigations in the diagnosis of Pityriasis Versicolor and Seborrheic Dermatitis
 Original article: The role of mycological investigations in the diagnosis of Pityriasis Versicolor and Seborrheic Dermatitis 1Saurabh Ajit Deshpande, 2 Dr. Sunita Deshpande, 3 Dr. Sangeeta Amladi, 4 Dr.
Original article: The role of mycological investigations in the diagnosis of Pityriasis Versicolor and Seborrheic Dermatitis 1Saurabh Ajit Deshpande, 2 Dr. Sunita Deshpande, 3 Dr. Sangeeta Amladi, 4 Dr.
2,6,9-Triazabicyclo[3.3.1]nonanes as overlooked. amino-modification products by acrolein
![2,6,9-Triazabicyclo[3.3.1]nonanes as overlooked. amino-modification products by acrolein](/thumbs/86/94743397.jpg "2,6,9-Triazabicyclo[3.3.1]nonanes as overlooked. amino-modification products by acrolein") Supplementary Information 2,6,9-Triazabicyclo[3.3.1]nonanes as overlooked amino-modification products by acrolein Ayumi Tsutsui and Katsunori Tanaka* Biofunctional Synthetic Chemistry Laboratory, RIKEN
Supplementary Information 2,6,9-Triazabicyclo[3.3.1]nonanes as overlooked amino-modification products by acrolein Ayumi Tsutsui and Katsunori Tanaka* Biofunctional Synthetic Chemistry Laboratory, RIKEN
Signaling in the Nitrogen Assimilation Pathway of Arabidopsis Thaliana
 Biochemistry: Signaling in the Nitrogen Assimilation Pathway of Arabidopsis Thaliana 38 CAMERON E. NIENABER ʻ04 Abstract Long recognized as essential plant nutrients and metabolites, inorganic and organic
Biochemistry: Signaling in the Nitrogen Assimilation Pathway of Arabidopsis Thaliana 38 CAMERON E. NIENABER ʻ04 Abstract Long recognized as essential plant nutrients and metabolites, inorganic and organic
SUPPLEMENTARY INFORMATION. Divergent TLR7/9 signaling and type I interferon production distinguish
 SUPPLEMENTARY INFOATION Divergent TLR7/9 signaling and type I interferon production distinguish pathogenic and non-pathogenic AIDS-virus infections Judith N. Mandl, Ashley P. Barry, Thomas H. Vanderford,
SUPPLEMENTARY INFOATION Divergent TLR7/9 signaling and type I interferon production distinguish pathogenic and non-pathogenic AIDS-virus infections Judith N. Mandl, Ashley P. Barry, Thomas H. Vanderford,
Product Contents. 1 Specifications 1 Product Description. 2 Buffer Preparation... 3 Protocol. 3 Ordering Information 4
 INSTRUCTION MANUAL Quick-RNA Midiprep Kit Catalog No. R1056 Highlights 10 minute method for isolating RNA (up to 1 mg) from a wide range of cell types and tissue samples. Clean-Spin column technology allows
INSTRUCTION MANUAL Quick-RNA Midiprep Kit Catalog No. R1056 Highlights 10 minute method for isolating RNA (up to 1 mg) from a wide range of cell types and tissue samples. Clean-Spin column technology allows
PNGase F Instruction Manual
 PNGase F Instruction Manual Catalog Number 170-6883 Bio-Rad Laboratories, 2000 Alfred Nobel Dr., Hercules, CA 94547 4006094 Rev A Table of Contents Section 1 Introduction...1 Section 2 Kit Components and
PNGase F Instruction Manual Catalog Number 170-6883 Bio-Rad Laboratories, 2000 Alfred Nobel Dr., Hercules, CA 94547 4006094 Rev A Table of Contents Section 1 Introduction...1 Section 2 Kit Components and
AN ELECTROPHORETIC COMPARISON OF ENZYMES IN STRAINS OF SPECIES IN THE FISSION YEAST GENERA SCHJZOSACCHAROMYCES, 0CTOSPOROMYCES, AND HA SEGA WAEA1
 J. Gen. Appl. Microbiol., 33, 363-369 (1987) AN ELECTROPHORETIC COMPARISON OF ENZYMES IN STRAINS OF SPECIES IN THE FISSION YEAST GENERA SCHJZOSACCHAROMYCES, 0CTOSPOROMYCES, AND HA SEGA WAEA1 YUZO YAMADA,
J. Gen. Appl. Microbiol., 33, 363-369 (1987) AN ELECTROPHORETIC COMPARISON OF ENZYMES IN STRAINS OF SPECIES IN THE FISSION YEAST GENERA SCHJZOSACCHAROMYCES, 0CTOSPOROMYCES, AND HA SEGA WAEA1 YUZO YAMADA,
Product # Kit Components
 3430 Schmon Parkway Thorold, ON, Canada L2V 4Y6 Phone: (905) 227-8848 Fax: (905) 227-1061 Email: techsupport@norgenbiotek.com Pneumocystis jirovecii PCR Kit Product # 42820 Product Insert Background Information
3430 Schmon Parkway Thorold, ON, Canada L2V 4Y6 Phone: (905) 227-8848 Fax: (905) 227-1061 Email: techsupport@norgenbiotek.com Pneumocystis jirovecii PCR Kit Product # 42820 Product Insert Background Information
SUPPLEMENTARY MATERIAL
 SUPPLEMENTARY MATERIAL Purification and biochemical properties of SDS-stable low molecular weight alkaline serine protease from Citrullus Colocynthis Muhammad Bashir Khan, 1,3 Hidayatullah khan, 2 Muhammad
SUPPLEMENTARY MATERIAL Purification and biochemical properties of SDS-stable low molecular weight alkaline serine protease from Citrullus Colocynthis Muhammad Bashir Khan, 1,3 Hidayatullah khan, 2 Muhammad
MagCapture Exosome Isolation Kit PS Q&A
 MagCapture Exosome Isolation Kit PS Q&A Specifications and performance P.1 Comparison of the conventional method P.2 Operation methods and composition P.4 Amount of starting sample P.5 Analysis after exosomes
MagCapture Exosome Isolation Kit PS Q&A Specifications and performance P.1 Comparison of the conventional method P.2 Operation methods and composition P.4 Amount of starting sample P.5 Analysis after exosomes
Quality control of Saccharomyces yeasts: differentiation of species level and strain grouping using COX 2 gene analysis and MALDI-TOF-MS analysis
 Quality control of Saccharomyces yeasts: differentiation of species level and strain grouping using COX 2 gene analysis and MALDI-TOF-MS analysis Hiroko Kawasaki 1, Shinya Ninomiya 1, Kanae Teramoto 2,
Quality control of Saccharomyces yeasts: differentiation of species level and strain grouping using COX 2 gene analysis and MALDI-TOF-MS analysis Hiroko Kawasaki 1, Shinya Ninomiya 1, Kanae Teramoto 2,
(A) PCR primers (arrows) designed to distinguish wild type (P1+P2), targeted (P1+P2) and excised (P1+P3)14-
 PCR primers (arrows) designed to distinguish wild type (P1+P2), targeted (P1+P2) and excised (P1+P3)14-") 1 Supplemental Figure Legends Figure S1. Mammary tumors of ErbB2 KI mice with 14-3-3σ ablation have elevated ErbB2 transcript levels and cell proliferation (A) PCR primers (arrows) designed to distinguish
1 Supplemental Figure Legends Figure S1. Mammary tumors of ErbB2 KI mice with 14-3-3σ ablation have elevated ErbB2 transcript levels and cell proliferation (A) PCR primers (arrows) designed to distinguish
MasterPure RNA Purification Kit
 Cat. No. MCR85102 The MasterPure RNA Purification Kit pro vides all of the reagents necessary to recover RNA from a wide variety of biological sources. This kit uses a rapid desalting process 1 to remove
Cat. No. MCR85102 The MasterPure RNA Purification Kit pro vides all of the reagents necessary to recover RNA from a wide variety of biological sources. This kit uses a rapid desalting process 1 to remove