JBC Papers in Press. Published on November 10, 2005 as Manuscript M
|
|
|
- Augustine Bishop
- 6 years ago
- Views:
Transcription
1 JBC Papers in Press. Published on November 10, 2005 as Manuscript M The latest version is at Evidence that Ser 87 of Bim EL is phosphorylated by Akt and regulates Bim EL apoptotic function. Xiao-Jun Qi 1, Gary M. Wildey 1, and Philip H. Howe* Department of Cell Biology, Cleveland Clinic Lerner College of Medicine, Cleveland Clinic Foundation, Cleveland, OH These authors contributed equally to this work. *Address correspondence to: Philip H. Howe, Ph.D., Department of Cell Biology, NC-1, Cleveland Clinic Lerner College of Medicine, Cleveland Clinic Foundation, Cleveland, OH Tel: (216) ; Fax: (216) ; howep@ccf.org Running Title: Ser 87 phosphorylation of Bim Bim, the Bcl-2 interacting mediator of cell death, is a member of the BH3-only family of pro-apoptotic proteins. Recent studies have demonstrated that the apoptotic activity of Bim can be regulated through a post-translational mechanism whereby ERK phosphorylation serves as a signal for Bim ubiquitination and proteasomal degradation. In this report, we investigated the signaling pathways leading to Bim phosphorylation in Ba/F3 cells, an IL-3-dependent B-cell line. IL-3 stimulation induced phosphorylation of Bim EL, one of the predominant isoforms of Bim expressed in cells, at multiple sites, as evidenced by the formation of at least 3-4 bands by Western blotting that were sensitive to phosphatase digestion. The appearance of multiple, phosphorylated species of Bim EL correlated with Akt, and not ERK, activation. The PI3-K inhibitor, LY294002, blocked IL-3-stimulated Akt activity and partially blocked Bim EL phosphorylation. In vitro kinase assays showed that recombinant Akt could directly phosphorylate a Gst-Bim EL fusion protein and identified the Akt phosphorylation site in the Bim EL domain as Ser 87. Further, we demonstrated that cytokine stimulation promotes Bim EL binding to proteins. Finally, we show that mutation of Ser 87 dramatically increases the apoptotic potency of Bim EL. We propose that Ser 87 of Bim EL is an important regulatory site that is targeted by Akt to attenuate the proapoptotic function of Bim EL, thereby promoting cell survival. Bim, the Bcl-2 interacting mediator of cell death, is a member of the BH3-only family of pro-apoptotic proteins that also includes Bad and Bid (1-3). Bim proteins are expressed by a wide variety of tissues, but are most prominently expressed by cells of hematopoietic origin (4). Although multiple Bim mrna transcripts have been described that are thought to be generated by alternative splicing (1,5-8), most tissues express one predominant isoform of Bim as determined by Western blotting analysis, termed Bim EL. Under conditions that promote cell growth, Bim is bound to dynein light chain (LC8), of the microtubular motor complex, and is sequestered away from other Bcl-2 family members (9). Following a pro-apoptotic stimulus, however, Bim localizes to the mitochondria, where it initiates the mitochondrial cell death pathway by either directly activating Bax-like proteins (7,10) or by binding to pro-survival Bcl-2 family members and thereby releasing Bax-like proteins (11-13). The apoptotic function of Bim is regulated by both pro-survival and pro-apoptotic cytokines. Withdrawal of IL-2 (14) or IL-3 (15-17) from cytokine-dependent hematopoietic cells results in an upregulation of Bim expression with an associated induction of apoptosis. Similar results are obtained when NGF is withdrawn from cultured neuronal cells (18-21). However, as opposed to negative regulation of Bim expression levels by these pro-survival cytokines, we have recently demonstrated that addition of a pro-apoptotic cytokine, TGFβ, results in the up-regulation of Bim expression levels in two different B-cell Copyright 2005 by The American Society for Biochemistry and Molecular Biology, Inc.
2 2 lines, WEHI 231 and Ba/F3 (22). We further demonstrated that stimulation of the prosurvival CD40 receptor is capable of inhibiting TGFβ-mediated Bim expression and cell death in WEHI 231 cells concomitant with its rescue of the cells from apoptosis. Several cell signaling pathways have been implicated in the control of Bim expression levels. In Ba/F3 cells, selective activation of either the Ras/MAPK or the Ras/PI3-K pathway by inducible Ras mutants suppresses the increase in Bim expression normally observed during IL-3 deprivation (16). The inhibition of Bim expression by Ras/PI3-K activation could be blocked by either the PI3-K inhibitor wortmannin or the mtor inhibitor rapamycin, but not by overexpression of a dominant negative form of Akt. Taken together, these results suggest that while both Akt and mtor are downstream effectors of PI3-K activation, only mtor regulates Bim expression levels. Other studies have also demonstrated an inhibitory effect of PI3-K signaling on Bim expression. The PI3-K inhibitor LY induces Bim expression in B cells, (Ba/F3)(15) and T cells (CTLL2)(14), concomitant with an increase in cell death. Thus, the biological effects of LY were identical with those of cytokine withdrawal in these model systems. In these studies, however, it was proposed that an important downstream effect of LY was to reduce the amount of Akt in its active, phosphorylated form. The link between reduced Akt activity and increased Bim expression levels was shown to be FOXO3 (previously termed FKHR-L1), a member of the forkhead family of transcriptional regulators that can directly elevate Bim expression levels and induce apoptosis in Ba/F3 and CTLL2 cells when overexpressed. FOXO3 is phosphorylated by Akt, leading to its cytoplasmic retention by proteins and loss of target gene activation (23). Thus, the upregulation of Bim expression induced by cytokine withdrawal or LY addition correlated with a loss of the active, phosphorylated form of Akt and an increase in the active, non-phosphorylated form of FOXO3. More recent studies have demonstrated that the apoptotic function of Bim is regulated by phosphorylation, followed by ubiquitination and degradation. Phosphorylated forms of Bim EL and Bim L were initially reported in Ba/F3 cells grown in the presence, but not the absence, of IL-3 (16). Subsequently, Bim EL phosphorylation has been demonstrated in NGF-stimulated PC12 cells (21) and superior cervical ganglion (SCG) cells (24), serum-stimulated CCL39 fibroblasts (25), IgM-stimulated Ramos B cells (26), IL-2-maintained HT-2 T lymphocytes (27), and M-CSF-stimulated osteoclasts (28). In the majority of these studies Bim phosphorylation could be blocked by the MEK/ERK pathway inhibitors PD98059 or U0126. Inducible expression of Raf, an upstream activator of the MEK/ERK pathway, in CCL39 fibroblasts (29), MCF10A mammary cells (30), and 293 cells (26) also resulted in Bim EL phosphorylation that could be blocked by U0126 and not by LY A specific role for ERK in Bim phosphorylation was established using in vitro kinase assays, where it was demonstrated that purified ERK could phosphorylate only Bim EL, and not Bim L or Bim S (25,26). Inducible-phosphorylation of Bim EL by Raf was also associated with a timedependent loss in total Bim protein. The loss of Bim protein was blocked by lactacystin and MG-132 (26, 28-31), implicating the proteasomal pathway in Bim degradation. Furthermore, multiple ubiquitinated forms of Bim protein could be demonstrated in the presence of proteasomal inhibitors (28,31). Thus, these studies outline an additional mechanism to control the apoptotic function of Bim via its phosphorylation by ERK and subsequent ubiquitination and degradation. In this report we investigate the IL-3-induced signaling pathways leading to Bim phosphorylation in Ba/F3 cells. IL-3 stimulation leads to the appearance of multiple phosphorylated isoforms of Bim EL in Ba/F3 cells. The PI3-K inhibitor LY blocks IL-3-stimulated Akt activity and, surprisingly, partially blocks Bim EL
3 3 phosphorylation. We further demonstrate that Gst-fusion proteins containing the protein domain unique to Bim EL can be phosphorylated in vitro by Akt and identified the Akt phosphorylation site in the Bim EL domain as Ser 87. Akt activation by survival cytokines promotes Bim EL phosphorylation and association with proteins. Finally, we show that mutation of the Akt phosphorylation site at Ser 87 dramatically increases the apoptotic potency of Bim EL. Materials and Methods Reagents- TGFβ was a generous gift from Genzyme Inc. (Cambridge, MA) and was used at a final concentration of 5 ng/ml. Rabbit anti-bim antibody was obtained from BD Pharmingen (San Diego, CA). Rabbit antiphospho-akt (Ser-473) and rabbit anti-total Akt were purchased from Cell Signaling Technology (Beverly, MA). Mouse antiphospho-erk (E4), rabbit anti-erk 1 (C-16), mouse anti β (H-8), rabbit anti-hsp 90 (H-114) and normal rabbit IgG were obtained from Santa Cruz Biotechnology (Santa Cruz, CA). Active recombinant mouse MAP Kinase/ERK 2, recombinant human Akt1/PKBα kinase, recombinant Gsk-3β and RSK1/MAPKAP kinase 1α were purchased from Upstate (Charlottesville, VA). Secondary antibodies were purchased from the following vendors: anti-mouse-igg-hrp from Accurate Antibodies (San Diego, CA) and anti-rabbit- IgG-HRP from Bio-Rad (Hercules, CA). MG- 132, LY294002, PD98059, SP600125, SB and wortmannin were obtained from Calbiochem (La Jolla, CA). U0126 was purchased from Promega (Madison, WI). Murine recombinant IL-3 was purchased from BioSource International (Camarillo, CA) and human recombinant insulin from Sigma Chemical Co. (St. Louis, MO). Protease inhibitor cocktail tablets were purchased from Roche Diagnostics (Indianapolis, IN). TBScasein for Western blotting was obtained from Pierce Chemical Co. (Rockford, IL) and Rainbow low molecular weight markers from Amersham BioSciences Corp (Piscataway, NJ). Calf alkaline phosphatase was purchased from New England Biolabs (Beverly, MA). Lipofectamine Plus transfection reagent was obtained from InVitrogen (Grand Island, NY). prl-tk- Renilla luciferase and the Dual-Luciferase Reporter Assay system were purchased from Promega (Madison, WI). Reagent chemicals and sodium vanadate were obtained from Sigma. Cell Culture- Ba/F3 cells were maintained in T75 flasks at a density of 2 x 10 4 cells/ml (45 ml total) in DMEM/F-12 medium supplemented with 10% fetal calf serum, 30 µm 2-β-mercaptoethanol and antibiotics/ antimycotics (100 units/ml of penicillin G, 100 mg/ml of streptomycin and 0.25 µg/ml amphotericin B; InVitrogen). Conditioned medium from WEHI 3B cells was added to a final concentration of 10% to provide the IL-3 essential for Ba/F3 survival (32). Conditioned medium was prepared from WEHI 3B cells seeded at an initial density of 10 x 10 4 cells/ml (45 ml total) and grown for 24 h in Ba/F3 medium. To induce Bim expression, Ba/F3 cells were seeded at an initial density of 5-10 x 10 4 cells/ml (45 ml total) and grown overnight in the absence of conditioned medium (IL-3- or cytokine-deprived). The next day, conditioned medium (10-20% final concentration) or recombinant mouse IL-3 ( ng/ml, as indicated) was added for the indicated time period to induce Bim phosphorylation. At the end of the experiment, the cells were collected by centrifugation, washed twice with PBS and resuspended in the appropriate lysis buffer. Ba/F3 cells were kept at 4 o C during all the above procedures. Where indicated LY (50 µm), wortmannin (100 nm), U0126 (10 µm), PD98059 (10 µm), SP (10 µm), SB (15 µm) or MG-132 (50 µm) were added to the culture medium min before IL-3 stimulation and remained present during the IL-3 treatment. Human Hep3B and rat FAO hepatoma cells were cultured in DMEM containing 10% FCS and antibiotics/antimycotics. Where indicated, Hep3B cells were serum-starved for 48 h prior to the addition of insulin (10 µg/ml). MG-
4 4 132 (50 µm) was added to the culture medium 60 min before insulin stimulation and remained present during the insulin treatment. Western Blot Analysis- Western blot analysis was performed by standard SDS-PAGE, as described previously (22). Whole cell lysates were prepared in TNMG lysis buffer (20 mm Tris, ph 8.0, 150 mm NaCl, 5.0 mm MgCl 2, 0.5 % NP-40 and 10% glycerol) containing protease inhibitors and 1 mm Na 3 V0 4. Lysates were sonicated and clarified by centrifugation at 4 o C for 10 min in a Beckman tabletop microcentrifuge at maximum speed. Protein concentration of the extracts was determined using Bradford s reagent (Pierce, Rockford, IL). Typically, µg of whole cell lysate was separated on 10 or 12% acrylamide minigels and transferred to Immobilon-P membrane (Millipore, Bedford, MA). The membrane was blocked for 1 h in wash buffer (TBS with 0.05% Tween 20) containing 5% nonfat dry milk, followed by a 2 h incubation with primary antibody diluted in the same blocking buffer. For phosphospecific antibodies, the primary antibody incubation was carried out in TBS-casein solution. After extensive washing, the blot was incubated with secondary antibody for 1 h in blocking buffer, washed and processed using the ECL + Western Blotting Detection System (Amersham Bioscience Corp, Piscataway, NJ). Primary antibodies were employed at a 1:500 to 1:5,000 dilution and secondary antibodies were used at a 1:5,000 to 1:10,000 dilution. Western blotting was performed at room temperature. Alkaline Phosphatase Digestion of Bim- Triplicate samples of cell lysates (150 µg) were individually digested with either a low (20 U) or high (50 U) dose of calf alkaline phosphatase for 1 h at 37 o C in buffer supplied by the manufacturer. For one set of digests, the calf alkaline phosphatase was heat inactivated at 95 o C for 5 min prior to adding the cell lysate. For the third set of digests, the phosphatase inhibitor Na 3 V0 4 (5 mm) was added to the digestion buffer. As a control, a sample of the lysate was incubated with buffer alone for 1 h at 37 o C. Gst-Bim Fusion Proteins- A Gst-Bim fusion protein was produced that contained only the amino acid sequence unique to human Bim EL (aa ). A Bim EL fragment was amplified by PCR using full-length human Bim EL cloned into the pflag-cmv2 expression vector as a template (11). The top primer was 5 - GGATCCGGATCCGGTAATCCTGAAGGCA AT-3 and the bottom primer was 5 - GAATTCGAATTCCTGTGTCAAAAGAGAAA T A-3. The Bim EL PCR fragment was digested with Bam H1 and Eco R1, purified and cloned into similarly-digested pgex-2t (Amersham Pharmacia Biotech, Piscataway, NJ). The expression of Gst-Bim fusion protein was induced for 4 h in BL21(DE3)pLysS cells (Novagen, Madison, WI) with 1 mm IPTG. The fusion protein was purified by affinity chromatography using Glutathione Sepharose 4B beads (Amersham Pharmacia Biotech). When necessary, recombinant protein was eluted from the beads with 50 mm Tris, ph 7.4 containing 10 mm reduced glutathione. The concentration of Gst-Bim EL fusion protein was estimated from Coomassie Blue-stained SDS-PAGE gels. Recombinant Gst protein was produced in parallel with recombinant Gst-Bim EL protein for use in control experiments. A Gst-Bim EL fusion protein containing a 16- amino acid truncation in the Bim EL carboxy (C)-terminal sequence (Gst-Bim EL -16 ) was produced in a similar manner. The truncated Bim EL fragment was amplified by PCR using the same top primer as described above and 5 - GAATTCGAATTCCTCTTCTCATAAAGATGA A-3 as the bottom primer and ligated into pgex-2t to obtain the truncated Bim EL Gst fusion protein. Gst-Bim EL fusion proteins containing single serine to alanine substitutions were produced using a QuikChange Site-Directed Mutagenesis kit from Stratagene (La Jolla, CA) according to the manufacturer s directions. The template for mutations was Bim EL ligated into pgex- 2T. The serine 87 to alanine substitution (Gst-Bim EL S87A ) was made using the top p r i m e r 5 - CTTTATGAGAAGATCCGCCCTGCTGTCTC
5 5 GATC- 3 and its complement in PCR reactions. The serine 94 to alanine substitution (Gst-Bim EL S94A ) was made using the top primer 5 - CTGCTGTCTCGATCCTCCGCCGGGTATTT CTCTTTTGACAC-3 and its complement in PCR reactions. In Vitro Kinase Assays- In vitro kinase assays were performed using both purified enzymes and kinases immunoprecipitated from whole cell lysates. Whole cell lysates for immunoprecipitation (IP) were prepared in IP lysis buffer (20 mm Tris, ph 7.5, 150 mm NaCl, 1 mm EDTA, 1 mm EDTA, 1% Triton X-100) containing protease inhibitors and 1 mm Na 3 V0 4. Lysates were sonicated and clarified by centrifugation at 4 o C for 10 min in a Beckman tabletop microcentrifuge at maximum speed. Cell lysates (250 µg) were incubated for 4 h at 4 o C with 2.5 µg of either rabbit anti-p-akt antibody or anti-akt antibody in 200 µl of IP lysis buffer containing Protein G agarose (Amersham/Pharmacia, Piscataway, NJ). The immune complexes were collected by centrifugation, washed five times in IP lysis buffer and resuspended in 30 µl of kinase buffer (25 mm Tris, ph 7.5, 5 mm β-glycerolphosphate, 10 µm ATP, 2 mm DTT, 0.1 mm Na 3 VO 4, 10 mm MgCl 2 ). Kinase reactions were initiated by adding 5 µg of substrate protein and 0.5 µci of [ 32 P]-γ-ATP (3000 Ci/mmol; PerkinElmer Life Sciences, Boston, MA) in 20 µl of kinase buffer. The model substrate proteins utilized in kinase assays were dephosphorylated bovine myelin basic protein (MBP; UpState) for ERK, Gsk-3 fusion protein (Cell Signaling Technology) for Akt, CREB fusion protein (UpState) for RSK and GSM peptide (UpState) for Gsk-3β. In addition, Gst and Gst-Bim EL fusion proteins were used as substrates in kinase assays. The reactions were stopped after a 20 min incubation at 30 o C by adding SDS-PAGE loading buffer and heating to 95 o C for 2 min. The samples were run on a 12% SDS-PAGE gel and analyzed by autoradiography. In those experiments using pure enzymes, equivalent units ( U) of kinase activity was added to reaction mixtures in 30 µl of kinase buffer. Co-Immunoprecipitation of endogenous Bim and proteins- Whole cell lysates ( µg) were incubated for 4 h at 4 o C with 2.5 µg of either rabbit anti-bim antibody or normal rabbit IgG in 200 µl of IP lysis buffer containing Protein G agarose. The immune complexes were collected by centrifugation and washed five times in IP lysis buffer. The presence of proteins in the immune complexes was detected by Western blotting. Transient transfection of FAO cells- Fulllength human Bim EL was cut out of pflag- CMV2 by XhoI/EcoRI double digestion and ligated into pcdna3.1/hisc (InVitrogen). Serine to alanine mutations at residues 69, 87 and 94 were obtained as described above for Gst-Bim EL fusion proteins. The top PCR primer to produce the S69A mutation was 5 - GCTGGCCCCACCTGCCGCCCCTGGCCCT TTTGC-3 and the bottom primer was 5 - GCAAAAGGGCCAGGGGCGGCAGGTGGG GCCAGC-3. Transient transfection of FAO cells was accomplished using Lipofectamine Plus reagent (InVitrogen) under serum-free conditions following the manufacturer s instructions. After 5-6 h of transfection, the medium was replaced with complete medium. For reporter assays, 150,000 cells/well were seeded into 12-well plates and transfected with 200 ng prl-tk-renilla luciferase and varying amounts of pcdna-bim EL. GFP was used as a negative control and to ensure that all cells received equal amounts of DNA. Approximately 24 h after the start of transfection, the cells were lysed in 200 µl and a 20 µl aliquot was assayed for luciferase activity using Stop & Glo reagent (Promega). For Western blot and DNA ladder assays, 4 x 10 6 cells were seeded into 100 mm dishes and transfected with either 5 or 15 µg of pcdna-bim EL or GFP. Cells were collected 24 h later for analyses. DNA ladder formation was assayed as described previously (22). Results Bim EL is phosphorylated and proteasomallydegraded in Ba/F3 cells- Ba/F3 cells require IL-3 in the tissue culture medium to survive
6 6 and proliferate and withdrawal of IL-3 leads to increased Bim expression and DNA ladder formation, a hallmark of apoptosis (15,16). Because of the recent interest in Bim as a target for ERK phosphorylation and proteosomal degradation, it was of interest to determine whether or not the suppression of Bim expression in Ba/F3 cells by IL-3 involved Bim phosphorylation. To this end, Ba/F3 cells were incubated overnight in the presence or absence of WEHI 3B-conditioned medium, which contains IL-3 (32). The next day, conditioned medium was added to the culture medium of starved cells for 30 min prior to collecting the cells. Western blot analysis of the cell lysates with anti-bim antibody revealed the presence of kda, high molecular weight forms of Bim, presumably Bim EL (Fig. 1A, top panel). As expected, withdrawal of conditioned medium dramatically increased Bim EL protein expression in Ba/F3 cells (lane 2, -CM) relative to cells grown in the presence of conditioned medium (lane 1, +CM). Addition of conditioned medium led to the formation of at least three species of Bim EL protein (lanes 3-4). To prove that it was the IL-3 in the WEHI 3B conditioned medium that stimulated Bim EL phosphorylation, starved Ba/F3 cells were incubated for 30 min with recombinant IL-3. IL-3 stimulation led to a dose-dependent increase in the appearance of multiple, phosphorylated Bim EL bands (Fig. 1A, lanes 5-9) and this stimulation was greater than that observed with conditioned medium (lanes 3 and 4). There also appeared to be a loss of immunoreactive Bim EL protein at higher doses (1-2 ng/ml) of IL-3. Taken together, these observations suggest that IL-3 induces a dose-dependent phosphorylation of Bim EL at multiple sites in Ba/F3 cells, leading to its degradation. A time-course study of IL-3 stimulation of Ba/F3 cells showed that the apparent phosphorylation and degradation of Bim EL was rapid in onset (Fig. 1B, top panel). The results of Fig. 1A and 1B are consistent with the idea that the addition of IL-3 to Ba/F3 cells leads to the production of multiple forms of phosphorylated Bim EL. To prove that the observed mobility shift in Bim EL species was due to phosphorylation, cell lysates from Ba/F3 cells stimulated for 15 min with conditioned medium were digested with calf alkaline phosphatase (CIP)(Fig. 1C). Digestion of the cell lysates with a low (lane 3, L) or high (lane 6, H) dose of CIP resulted in a dose-dependent increase in the mobility of Bim EL species compared with the undigested control sample (lane 1, control). Blocking CIP activity by either heat inactivation of the enzyme (lanes 4 and 7, CIP/Heat) or addition of the phosphatase inhibitor sodium vanadate (lanes 5 and 8, CIP/NaV) prevented the CIP-induced increase in Bim EL mobility. Incubation of the lysate with buffer in the absence of CIP had no effect (lane 2, buffer). Thus, these results demonstrate that the multiple species of Bim EL are produced by phosphorylation. If the addition of IL-3 to Ba/F3 cells leads to the phosphorylation of Bim EL, then the apparent loss of Bim EL protein observed in Fig. 1A and 1B may be due to proteasomal degradation of phosphorylated Bim EL. To address this question, Ba/F3 cells were starved overnight and treated with conditioned medium the next morning for 1 h in the presence or absence of the proteasomal inhibitor MG-132 (Fig. 1D). As expected, Bim EL protein levels were increased by withdrawal of the conditioned medium (lane 2, -CM versus lane 1, +CM). Once again, addition of conditioned medium stimulated Bim EL phosphorylation and loss of Bim EL total protein in the absence of MG-132 (lane 3). In the presence of MG-132, however, a higher molecular weight, phosphorylated species of Bim EL accumulated (lane 4). Taken together, the results of Fig. 1 demonstrate that IL-3 stimulates Bim EL phosphorylation in starved Ba/F3 cells and signals the proteasomal degradation of Bim EL. IL-3 stimulates Bim EL phosphorylation by Akt- The results in Fig. 1A suggest that IL-3 stimulates Bim EL phosphorylation at multiple sites, particularly at higher (0.5-2 ng/ml) doses of IL-3. Since Bim EL is a known target of ERK phosphorylation, then it is possible
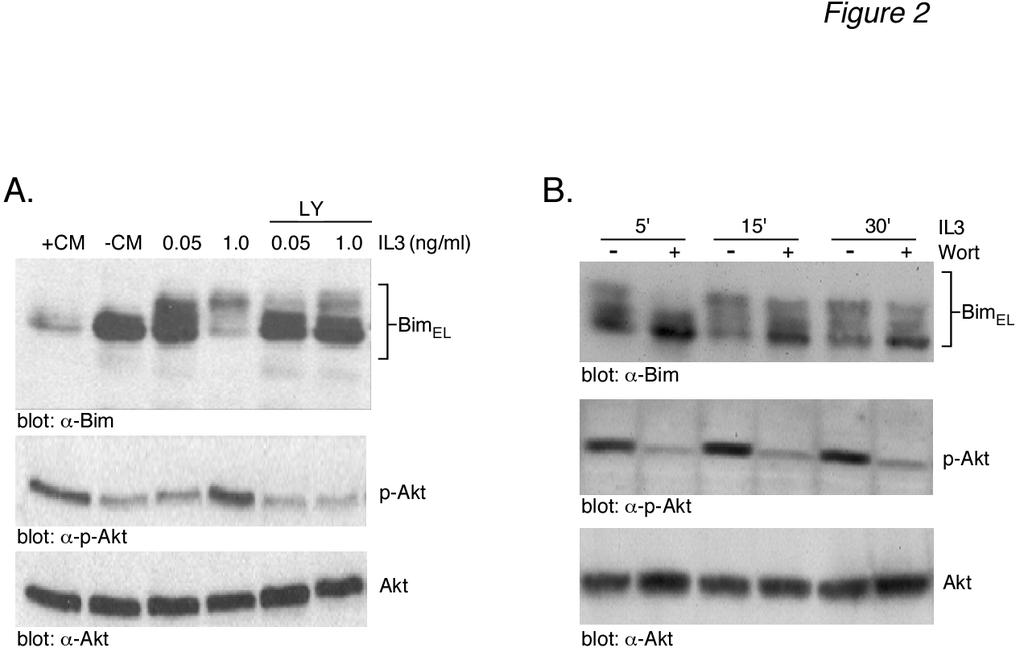
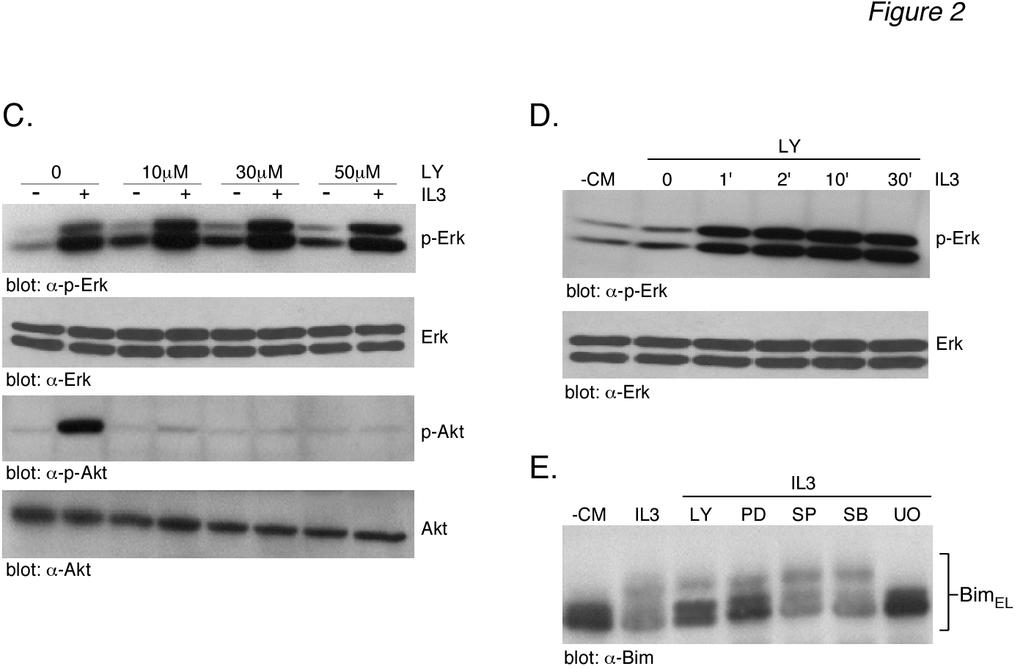
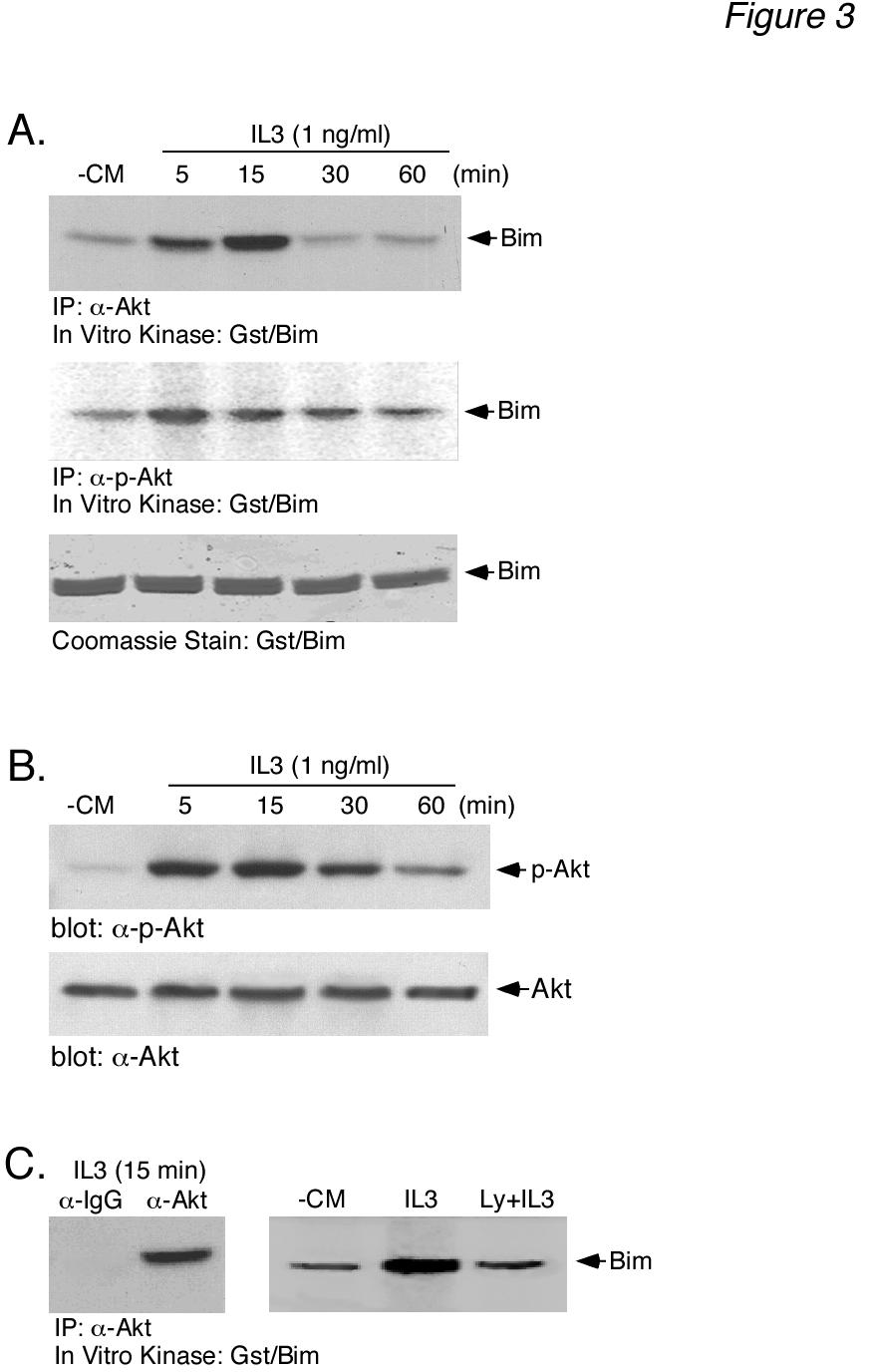
7 7 that the different levels of Bim EL phosphorylation observed in Fig. 1A resulted from different levels of IL-3-induced ERK activation. To address this question, the level of ERK activation was determined using an anti-active phospho-erk antibody. The results (Fig. 1A, middle panels) demonstrated that most doses of IL-3 and CM gave a robust stimulation of ERK activity, as measured by anti-phospho-erk immunoreactivity, with little change in total ERK protein. We next considered whether or not other kinases might be responsible for the multiple phosphorylated forms of Bim EL induced by IL- 3 stimulation. We initially chose to investigate a potential role for Akt in IL-3-induced BimEL phosphorylation because of numerous studies demonstrating the ability the PI3-K inhibitor LY to induce Bim expression and apoptosis in Ba/F3 cells (15,17). The level of Akt activation was determined using anti-active phospho-akt (Ser473) antibody. The results (Fig. 1A, bottom panels) demonstrated a dose-dependent increase in phospho-akt immunoreactivity with both IL-3 and CM that paralleled the dose-dependent increase in Bim EL phosphorylation. The level of total Akt protein was equal in all lanes. This result indicated that Akt may be involved in IL-3-induced Bim EL phosphorylation. If IL-3 stimulates Bim EL phosphorylation by Akt, then LY294002, a PI3K inhibitor that acts as an upstream inhibitor of Akt activation, should block IL-3-induced Bim EL phosphorylation. Cytokine-deprived Ba/F3 cells were stimulated with two different doses of IL-3 for 30 min in the absence or presence of LY and analyzed for Bim EL phosphorylation (Fig. 2A, top panel). As expected, IL-3 induced Bim EL phosphorylation in a dose-dependent manner (lanes 3 and 4). The addition of LY inhibited the phosphorylation of Bim EL to higher molecular weight forms (lanes 5 and 6). Control blots demonstrated that LY blocked the formation of active, phospho-akt (Fig. 2A, lanes 5 and 6, lower panels). Further experiments demonstrated that 100 nm wortmannin, another PI3-K inhibitor, was also capable of inhibiting IL-3-stimulated Bim EL phosphorylation and Akt activation, similar to LY (Fig. 2B). There are reports that LY can inhibit ERK activation in Ba/F3 cells (33). Because this would complicate the interpretation of our Bim EL phosphorylation results, control experiments were performed to investigate the effect of LY on IL-3 stimulated ERK activity (Fig. 2C). The results demonstrated that LY specifically blocked the IL-3 stimulated formation of active, phospho-akt (Fig. 2C, bottom panels) and had no effect on the formation of active, phospho-erk (Fig. 2C, top panels). Time course analysis also indicated that LY had no effect on the rapid kinetics of ERK activation by IL-3 (Fig. 2D). Bim has also been reported to be a target for JNK phosphorylation (34). Therefore, other MAPK pathways, besides ERK, may play a role in IL-3 induced Bim EL phosphorylation. To explore this possibility, specific inhibitors of the ERK (PD98059, U0126), JNK (SP600125) and p38 (SB202190) pathways were tested to determine their effect on Bim EL phosphorylation (Fig. 2E). The results showed that neither SP or SB inhibited IL-3 induced Bim EL phosphorylation. As expected, LY294002, PD98059 and U0126 all inhibited Bim EL phosphorylation, with U0126 being the most effective inhibitor. Taken together, our results suggested that the ERK and Akt pathways were involved in IL-3 induced Bim EL phosphorylation. Akt can phosphorylate Gst-Bim EL - The results of Figs. 1 and 2 suggest that Akt is involved in IL-3-stimulated Bim EL phosphorylation. To directly demonstrate the ability of Akt to phosphorylate Bim EL, in vitro kinase assays were performed using Akt immunoprecipitated from IL-3-stimulated Ba/F3 cells and a Gst-fusion protein containing the protein domain unique to human Bim EL as a substrate (see Fig. 4B for schematic of Gst-Bim construct). Akt activity was immunoprecipitated using either antitotal Akt antibody (Fig. 3A, top panels) or anti-phospho-akt antibody (Fig. 3A, middle
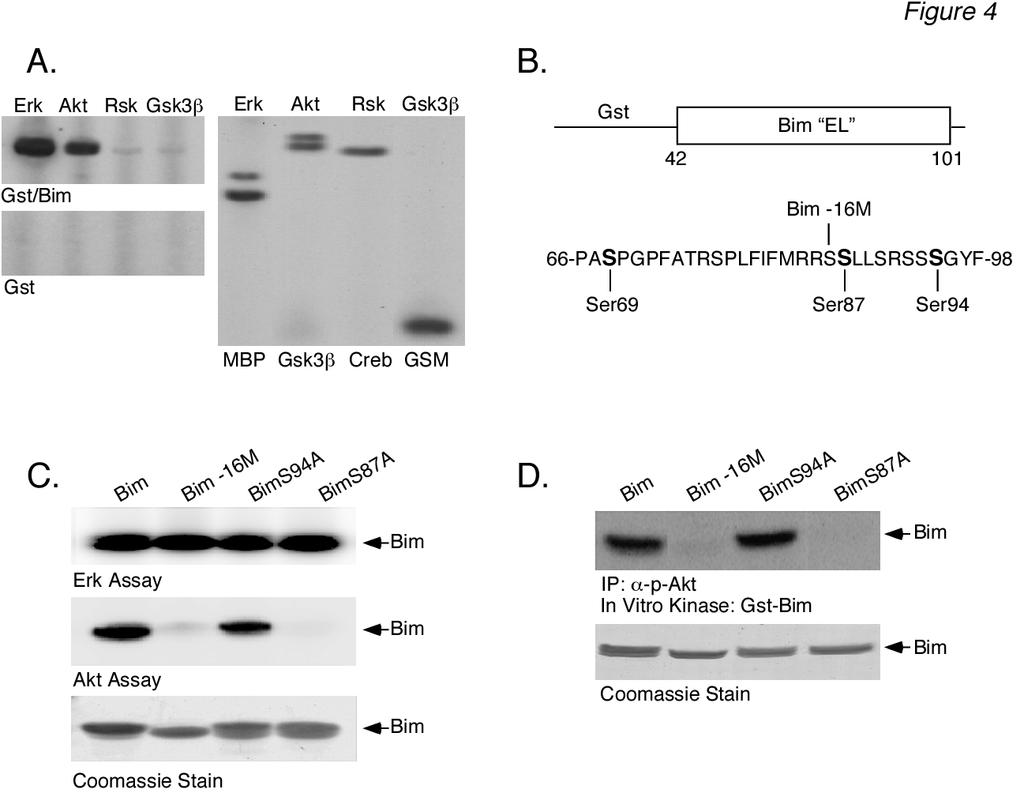
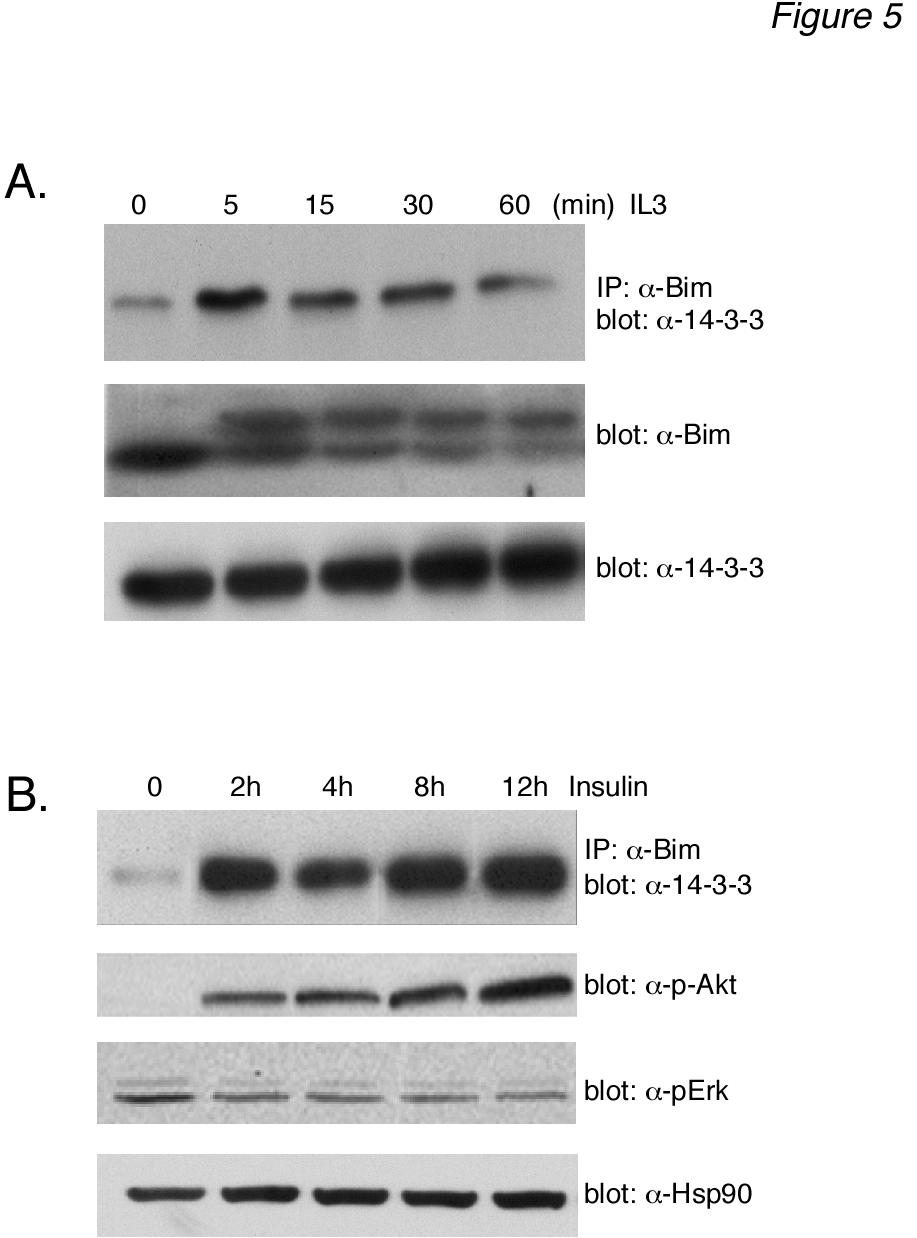
8 8 panel). The results demonstrated that immunoprecipitated Akt activity was able to phosphorylate Gst-Bim EL in vitro. Similar results were obtained with immunoprecipitates using either one of the anti-akt antibodies. Control experiments demonstrated that non-immune anti-rabbit IgG antibody was unable to immunoprecipitate any Gst-Bim EL kinase activity (Fig. 3C, left panel) and that LY treatment in vivo blocked the Akt activity measured in vitro (Fig. 3C, right panel). In parallel, the samples used for the in vitro Gst-Bim EL kinase assay were assayed for Akt activation using the anti-phospho-akt antibody (Fig. 3B, top panel). The results demonstrated that the time-course of IL-3- induced Akt activity measured with the in vitro Gst-Bim EL kinase assay paralleled that measured by using the anti-phospho-akt antibody. Collectively, these data support the idea that Ba/F3 stimulation by IL-3 induces the phosphorylation of Bim EL by Akt. Akt phosphorylates Bim EL on Ser 87 - To confirm that Akt phosphorylates Bim EL, we examined the ability of recombinant Akt to phosphorylate Gst-Bim EL using the in vitro kinase assay (Fig. 4A). As a positive control, we also tested equal units of activity of pure ERK. As controls, we used pure Rsk, a member of the AGC kinase family (PKA, PKG, PKC families) like Akt, as well as pure Gsk-3β, a member of the CMGC kinase family (CDK, MAPK, Gsk-3, CLK families) like ERK, in the in vitro kinase assay. Recombinant Akt phosphorylated Gst-Bim EL, but to a lesser extent than ERK (Fig. 4A, top left panel). The phosphorylation of Gst-Bim EL by recombinant Akt was dose-dependent (data not shown). In contrast, both Rsk and Gsk-3β demonstrated little phosphorylation of Gst-Bim EL (Fig. 4A, top left panel). All of the pure kinases phosphorylated their model substrates (Fig. 4A, right panel). Control experiments demonstrated that none of the purified kinases phosphorylated Gst by itself (Fig. 4A, left bottom panel) and that Coomassie Blue staining revealed equal amounts of substrate used in all of these experiments (data not shown). We next wanted to determine the Akt phosphorylation site(s) of Gst-Bim EL. The reported consensus site for Akt phosphorylation is R-X-R-X-X-S/T (35-38). Although this exact site is not found in the protein domain specific to human Bim EL, there are two copies of the sequence R-X-X-S found in its carboxy-terminal region (see schematic Fig. 4B). We therefore generated two mutants of Gst-Bim EL that contained serine to alanine substitutions at positions 87 and 94, termed Gst-Bim S87A EL and Gst- Bim S94A EL, respectively. In addition, we generated a mutant Gst-Bim EL protein from which the last 16 carboxy-terminal amino acids (Ser 86 to Thr 101 ) were deleted, termed Gst-Bim -16 EL. We then tested the ability of recombinant Akt to phosphorylate these three mutant Gst-Bim EL fusion proteins using the in vitro kinase assay (Fig. 4C, middle panel). Akt phosphorylation of Gst-Bim S94A EL (lane 3) was similar to that of wild-type Gst-Bim EL (lane 1). Akt was unable, however, to phosphorylate either Gst-Bim -16 EL (lane 2) or Gst-Bim S87A EL (lane 4). As a control, all three mutant Gst-Bim EL fusion proteins were tested as substrates for in vitro phosphorylation by recombinant ERK (Fig. 4C, top panel) because the reported ERK phosphorylation site, Ser 69 (25,26), is preserved in all of the Gst-Bim EL mutants (see schematic Fig. 4B). As expected, all three Gst-Bim EL mutants were phosphorylated by recombinant ERK (Fig. 4C). Identical results were obtained when the Akt used in the in vitro kinase assay was obtained by immunoprecipitation from IL- 3-stimulated Ba/F3 lysates (Fig. 4D). These results, therefore, identify Ser 87 as the Akt phosphorylation site. Phosphorylated Bim EL binds proteins in vivo- It was of interest to determine if Bim EL phosphorylation by Akt served some regulatory function. Several studies have demonstrated that Akt phosphorylates Bad on Ser 136, resulting in a down-regulation in the apoptotic function of Bad via sequestration by proteins (39-41). Therefore, we explored whether or not Akt phosphorylation of Bim promoted binding in coimmunoprecipitation experiments (Fig. 5A,
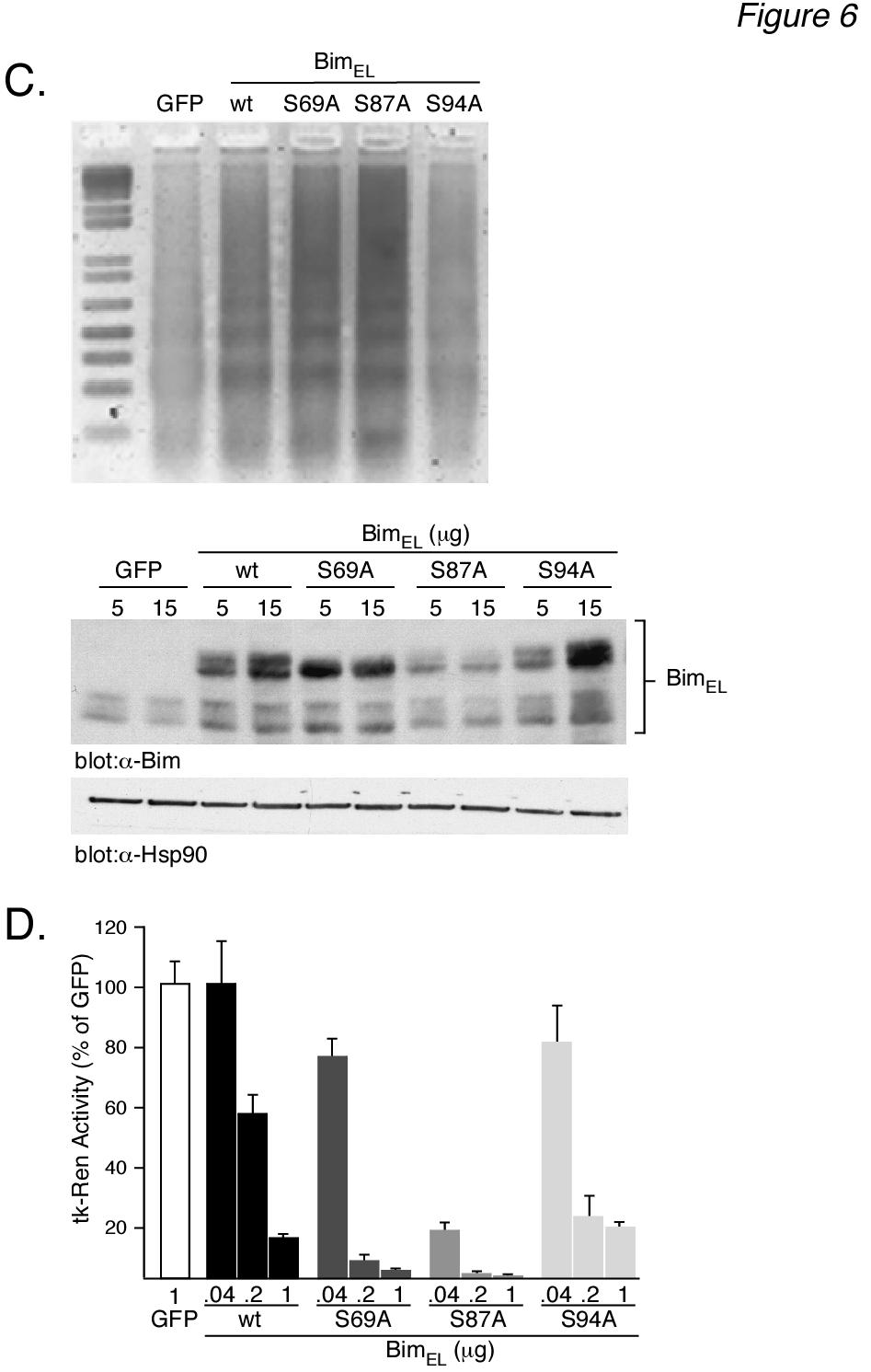
9 9 top panel). Our results demonstrated that IL-3 stimulated a rapid and sustained increase in the amount of protein that coimmunoprecipitated with Bim EL (lanes 2-5) compared to untreated, control levels (lane 1). Control, rabbit IgG was not able to immunoprecipitate protein (data not shown), demonstrating the specificity of the Bim EL / interaction. Although this association could be demonstrated in the absence of MG-132 (data not shown), the results were more consistent in the presence of proteasomal inhibition. This probably is related to the increased stability of Bim EL protein levels in the presence of MG-132 (Fig. 5A, middle panel) protein levels did not change with IL-3-stimulation (Fig. 5A, bottom panel). These results are consistent with the idea that IL-3-stimulated Akt phosphorylation of Bim EL promotes its binding and sequestration by proteins. We next tested whether or not Akt stimulation promoted Bim EL / association in other cell types. For this purpose we used Hep3B cells, a human hepatoma cell line where specific activation of the PI3K/Akt pathway by either IL6 (42) or insulin (43,44) was shown to be required for their pro-survival activity. Hep3B cells were serum-starved for 48 h then stimulated with insulin for 2-12 h in the presence of MG-132. Under these conditions insulin specifically stimulated Akt, and not ERK, as measured by Western blotting using anti-active phospho-akt (Fig. 5B, second panel) and anti-phospho-erk antibodies (Fig. 5B, third panel). Western blotting also showed no change in the levels of total ERK or total Akt (data not shown). The amount of protein loaded in each lane was similar, as judged by Western blotting for a heat shock protein, HSP-90 (Fig. 5B, bottom panel). Of particular interest, however, was the sustained increase in the amount of protein that co-immunoprecipitated with Bim EL that was observed after Akt activation by insulin (Fig. 5B, top panel, lanes 2-5) compared to control levels (lane 1). The specific activation of Akt, and not ERK, by insulin in Hep3B cells under serum-free conditions supports the idea that Akt phosphorylation of Bim EL is responsible for its association with proteins. Mutation of Ser 87 to alanine increases the apoptotic potency of Bim EL - We next wanted to determine whether or not the Akt phosphorylation of Bim EL and its subsequent sequestration by proteins downregulated the apoptotic function of Bim EL, similar to the regulation of Bad. Our strategy was to mutate the phosphorylation sites in Bim EL and determine what effect this had on the ability of Bim EL to induce apoptosis in over-expression studies. For this purpose we used FAO rat hepatoma cells because they are easily transfectable and exhibit low resting levels of Bim EL compared to other cell lines (unpublished observation). Initially, we tested the ability of over-expressed, wild-type Bim EL to induce apoptosis in FAO cells. As shown in Fig. 6A, transient transfection of Bim EL into FAO cells resulted in the dosedependent appearance of a high molecular weight form of immuoreactive Bim that was not observed in cells transfected with GFP (top right panel). This result is consistent with the fact that over-expressed Bim EL contains an epitope tag. Interestingly, there appeared to be multiple bands of over-expressed Bim EL. Western blotting for HSP-90 demonstrated equal protein loading (bottom right panel). As expected, over-expression of Bim EL led to a dose-dependent increase in cell death in FAO cells as measured by DNA ladder formation (Fig. 6A, left panel). The extent of cell death was also demonstrated more quantitatively as a loss of TK-Renilla activity with increasing concentrations of co-expressed Bim EL (Fig. 6B). This type of cell survival assay has been used previously to determine the relative apoptotic potency of Bim EL mutants using either CMV-luciferase (26) or GFP fluorescence (21, 24) as a readout assay. We next tested the apoptotic potency of three mutants of Bim EL compared to wild-type Bim EL. The mutants contained single serine to alanine substitutions at either residue 69, the ERK phosphorylation site, residue 87, the Akt phosphorylation site, or residue 94, a nonphosphorylated site. Western blotting demonstrated that all mutants could be
10 10 expressed, but their patterns of expression were different (Fig. 6C, bottom panels). The expression of the S94A mutant was similar to wild-type Bim EL. Interestingly, neither the S69A or S87A mutant showed a dosedependent increase in expression with increased DNA, in contrast to wild-type Bim EL and the S94A mutant. The S87A mutant demonstrated the lowest expression level of all Bim EL proteins. The S69A mutant was the only Bim EL protein that did not demonstrate multiple immunoreactive bands. Western blotting for HSP-90 demonstrated equal protein loading. When the ability of the four Bim EL proteins to induce apoptosis was measured (Figs. 6C top panel and 6D), we found that the S69A mutant was more potent than wild-type Bim EL, as expected. However, we were surprised to find that the S87A mutant was clearly more potent than the S69A mutant. The S94A mutant was about equal to wild-type Bim EL in its ability to induce cell death. Taken together, the results of Fig. 6 suggest that the low expression of the S87A mutant is likely because it is a very potent inducer of apoptosis and cannot be over-expressed without killing the cell. Discussion Expression levels of BH3-only proteins are key determinants of cellular survival and are therefore under stringent control. Transcriptional and post-translational regulatory mechanisms have previously been described in the control of Bim expression levels. Cytokine-regulated phosphorylation of Bim and its ubiquitin-mediated degradation through the proteasomal pathway has recently emerged as a key regulatory mechanism controlling cellular Bim levels. In this study we show that the kinases responsible for IL-3-induced Bim EL phosphorylation include not only ERK, which has previously been documented, but also the Akt kinase. We provide evidence that Akt phosphorylation of Bim EL, and not ERK, acts as a signal for Bim EL binding to, and likely sequestration by, proteins in that binding correlates with Akt activation and not that of ERK (Fig. 5B). Finally, we demonstrate that mutation of the Akt phosphorylation site in Bim EL, Ser 87, to alanine dramatically increases the apoptotic potency of Bim EL. Our results demonstrate that IL-3 stimulation of Ba/F3 cells induced phosphorylation of Bim EL at multiple sites, as evidenced by the formation of at least 3-4 bands by Western blotting that were sensitive to phosphatase digestion. This finding is in agreement with the results of two previous studies in which the number of Bim EL species was examined by 2-D gel electrophoresis (25,27). For example, it was shown that Bim EL immunoprecipitated from [ 32 P]-metabolicallylabeled murine HT-2 or Bal17 lymphocytes, separated into at least four [ 32 P]-labeled forms (27). Furthermore, some spots appeared to contain more than one form of [ 32 P]-labeled Bim EL. Phospho-amino acid analysis revealed only [ 32 P]-labeled serine, and not threonine or tyrosine residues. After IL-2 deprivation, at least two of the [ 32 P]- labeled forms of Bim EL were decreased in HT- 2 cells (27). Similar results were obtained in CCL39 fibroblasts, where addition of FBS to serum-starved cells increased the appearance of three, presumably phosphorylated, forms of immunoreactive Bim EL (25). The results of both of these studies provide strong evidence that Bim EL is highly phosphorylated and that this phosphorylation is tightly regulated by cytokines. The role of the ERK pathway in Bim-mediated phosphorylation has attracted the most investigation. The FBS-induced appearance of multiple forms of Bim EL in CCL39 cells was shown to be sensitive to the ERK1/2 pathway inhibitor, PD184352, suggesting that multiple ERK phosphorylation sites exist in Bim EL (25). A more recent study, in the IL-3-dependent murine pro-b cell line, FL5.12, demonstrates that when IL-3 is added back to cytokinedeprived cells there is a rapid shift in the mobility of Bim EL to an apparently higher molecular weight form, consistent with its phosphorylation (45). Further, six Bim EL mutants were generated by alanine
11 11 substitution of individual, putative ERK phosphorylation sites and tested for their ability to be phosphorylated when transfected into IL-3-stimulated FL5.12 cells. Three mutants, S55A, S65A and S100A, failed to undergo a mobility shift, indicating that ERK could potentially phosphorylate Bim EL at these three sites. Western blotting with anti- Bim EL antibodies that recognize only the phosphorylated S55, S65 and S100 sites confirmed that all of these sites were phosphorylated in cells overexpressing wildtype Bim EL after IL-3 stimulation and were sensitive to the MEK/ERK pathway inhibitor, U0126. Despite the above evidence that ERK phosphorylates Bim EL at multiple sites, two studies have demonstrated that a single mutation in Gst-Bim EL fusion proteins at Ser 65 of murine Bim EL (25), or the equivalent Ser 69 of human Bim EL (26), is sufficient to completely abrogate phosphorylation by recombinant ERK in an in vitro kinase assay. ERK was shown not to phosphorylate the Bim L or Bim S isoforms of Bim in vitro (25). Thus, it is unlikely that the multiple phosphorylated forms of Bim EL observed by us, as well as others, can be accounted for by only ERK phosphorylation. In this regard, Bim has also been reported to be a substrate for JNK-mediated phosphorylation (34). However, all of the three potential JNK phosphorylation sites identified were found using the Bim L isoform as substrate, and are not present in the EL-specific domain of Bim. In our study, we did not observe any IL-3- induced mobility shift in Bim L (Fig. 1D) and a specific JNK inhibitor, SP600125, had no effect on IL-3-induced Bim EL phosphorylation (Fig. 2E). Furthermore, the p38 inhibitor SB also had no effect on IL-3-induced Bim EL phosphorylation, whereas the ERK pathway inhibitors PD98059 and U0126 inhibited phosphorylation, as expected (Fig. 2E). Therefore, it is likely that only the ERK MAPK pathway is involved in IL-3 stimulated Bim EL phosphorylation in Ba/F3 cells. Based on these previous studies, we reasoned that kinases other than ERK and JNK may mediate the IL-3-induced phosphorylation of Bim EL observed in our studies of Ba/F3 cells. We decided to explore a potential role for Akt in Bim EL phosphorylation because of the wellrecognized role of the Akt pathway in cell survival in general (36-38), and Ba/F3 cells in particular (15,17,46), although there are contradictory reports that the PI3K-Akt pathway plays no role in Ba/F3 apoptosis (47,48). Our results demonstrate that higher doses of IL-3 stimulate activation of the Akt pathway, in addition to the ERK pathway, which correlate with Bim EL phosphorylation at multiple sites. Furthermore, the PI3K/Akt pathway inhibitor LY consistently blocked both IL-3-stimulated Akt activation and Bim EL phosphorylation, supporting a role for Akt in Bim EL phosphorylation. The blockade of Bim EL phosphorylation by LY was of note, since it has been reported that this inhibitor does not block acute (<1 h) Bim phosphorylation induced by NGF in PC12 cells (21), Raf overexpression or PMA treatment in 293 cells (26), or IL-3- stimulation in FL5.12 cells (45). LY also had no effect in more long-term studies (6-18 h) of Bim EL phosphorylation induced by FBS in CCL39 cells (29) or Raf overexpression in MCF-10A cells (30). The reason for these apparent contradictory results with the LY inhibitor may be related to several factors, including cell type, culture conditions or strength of stimulus. In addition, we routinely observe more phosphorylated Bim EL forms by Western blot analysis than do these other studies, and thus effects on phosphorylated species may be more easily detected. We also obtained identical inhibition of IL-3-stimulated Bim EL phosphorylation with wortmannin, another PI3K inhibitor structurally unrelated to LY294002, further supporting a role of the PI3K/Akt pathway. Our data demonstrating that both recombinant Akt and Akt immunoprecipitated from IL-3-stimulated Ba/F3 cells could phosphorylate a Gst-Bim EL fusion protein supports a role for Akt in IL-3-stimulated Bim EL phosphorylation in vivo. The Akt phosphorylation site we identified in Gst-
12 12 Bim EL by mutagenesis, Arg-Arg-Ser-Ser 87, is conserved in both human and mouse species, suggesting it plays an important biological role. Although this sequence does not constitute the full Akt consensus phosphorylation site often cited, R-X-R-X-X- S/T (35-38), other Akt substrates containing only an R-X-X-S/T phosphorylation site have been reported, such as CREB (37). In addition, the Akt phosphorylation site in Bim EL, Ser 87, is immediately followed by the amino acid leucine in both the human and mouse protein sequence. It is thought that the presence of a bulky, hydrophobic residue, such as phenylalanine or leucine, carboxyterminal to serine is favorable for Akt phosphorylation (35,36). It is interesting to note that there are no other potential Akt phosphorylation sites in the Bim protein sequence besides the two R-X-X-S sites located in the carboxy-terminus of the Bim EL domain. Thus, only Bim isoforms containing this unique domain will be subject to Akt phosphorylation and regulation. This may be another reason why Bim EL is the weakest inducer of apoptosis compared with Bim L and Bim S when overexpressed (1,6,21). Recently it has been shown that the ERK docking domain of Bim maps between amino acids Ala 70 and Thr 97 using Gst pull-down experiments (25). More recently, this domain has been identified as a DEF domain containing the amino acid sequence FSF (49). The DEF domain is located at the extreme carboxy-terminal end of the EL domain of Bim EL, after the ERK phosphorylation site. Interestingly, the Akt phosphorylation site that we have identified, Ser 87, maps between the ERK phosphorylation and docking sites. It is unclear, however, whether or not Akt phosphorylation of Bim EL would affect the ability of ERK to dock with and phosphorylate Bim EL because it was also demonstrated that Gst-Bim EL fusion proteins that contained the ERK phosphorylation site, but either lacked the ERK docking site or from which the DEF domain was deleted, could still be phosphorylated by ERK (25,49). In the present study we also explored the effect of Bim EL phosphorylation on binding. Our finding that Akt stimulation in Ba/F3 and Hep3B cells promoted increased Bim EL binding to proteins is similar to results of previous studies demonstrating that Akt phosphorylation of another BH3-only protein, Bad, also results in binding (41). It is postulated that one effect of binding to proteins is to sequester a protein away from its normal binding partners, thereby causing a biological effect. In the case of Bim EL, sequestration may prevent its dimerization with and neutralization of the pro-survival proteins Bcl- 2 and Bcl-X, leading to increased cell survival. Therefore, Bim EL sequestration by proteins may represent a secondary mechanism to rapidly neutralize excess, toxic Bim EL proteins that escape ERK-induced phosphorylation and degradation. This idea is supported by our finding that hyperphosphorylated forms of Bim EL are always observed, even after prolonged incubation with high doses of IL-3 (Fig. 1A and 1B using 1.0 ng/ml IL-3). This pool appears to represent only a small percentage of the total immunoreactive Bim EL detected prior to IL-3 treatment (-CM); therefore, it is likely that ERK-induced degradation of Bim EL represents the primary cellular mechanism to attenuate the apoptotic effects of Bim EL. In this regard, we also consistently noticed that the amount of immunoreactive Bim EL observed after LY treatment was always greater than that detected in corresponding controls, even though the level of ERK activation, as measured by p-erk Western blotting, was always greater in LY treated samples (Fig. 2C). This suggests that Akt phosphorylation of Bim EL may contribute to Bim EL degradation through an as yet undescribed mechanism, which we are currently investigating. Several studies have shown that LY increases Bim protein levels in Ba/F3 cells (15), as well as other cells (14,31). It is now clear that this effect of LY is due to its downstream inhibition of Akt activity. Akt phosphorylates forkhead transcription factors, causing them to be retained in the cytoplasm
13 13 via binding to proteins (23). Inhibition of Akt activity by LY allows forkhead proteins to translocate into the nucleus, where they are known to be direct activators of Bim gene expression (50-52). Therefore, Akt activation can down-regulate Bim transcription via its phosphorylation of forkhead proteins. The direct phosphorylation and sequestration of Bim EL by Akt described herein would appear to complement this longterm effect of Akt on Bim transcription by immediately neutralizing pre-existing Bim within a cell. The biological importance of the Akt phosphorylation site, Ser 87, was clearly demonstrated by the increased apoptotic potency in transient transfection studies of the S87A Bim EL mutant compared to either wild-type Bim EL or, unexpectedly, the S69A ERK phosphorylation site Bim EL mutant (Fig. 6C and 6D). Although many studies have made Ser to Ala mutations of Bim in order to study their effect on either Bim phosphorylation or apoptotic function (21,24-26,30,34,46), this is the first study to examine the biological importance of the Ser 87 site. The most striking feature of this mutant was our inability to over-express it, even at earlier time points (data not shown). Although weakly expressed, the S87A Bim EL mutant demonstrated two bands by Western blotting, suggesting that it was still phosphorylated, presumably by ERK. The apoptotic potency of the Ser 87 Bim EL mutant indicates that this site may regulate more than just binding and we are exploring this idea. The existence of multiple signaling pathways regulating apoptosis is likely to be critical for cell survival. In the present study, we have provided evidence that the apoptotic function of Bim is regulated by Akt, in addition to ERK. Thus, Bim would represent another Bcl-2 protein, in addition to MCL-1 (53) and Bad (54-56), that is targeted by multiple kinase signaling pathways to regulate its function. It will be of interest to determine in future studies whether or not proapoptotic cytokines such as TGFβ, which we have previously shown to increase Bim expression levels (22), can enhance the apoptotic function of Bim by down-regulating ERK and Akt activity. Acknowledgements We thank Dr. Steve Ledbetter at Genzyme Inc. for the generous provision of TGFβ. Human Bim EL cloned into the pflag-cmv2 expression vector was a kind gift of Hong- Gang Wang (University of South Florida College of Medicine, Tampa, FL). This work was supported by grants CA80095 and CA55536 from the National Cancer Institute to PHH. Figure Legends Figure 1: IL-3-induced Akt activation in Ba/F3 cells correlates with Bim EL hyperphosphorylation and proteasomal degradation. (A) Ba/F3 cells were incubated overnight in the presence or absence of conditioned medium. The next day starved cells were stimulated with either 10% or 20% conditioned medium (+CM, 10% and 20%) or with increasing doses of recombinant IL-3 (IL- 3, ng/ml), as indicated. After a 30 min incubation, cells were collected, lysed and analyzed by Western blotting. Cells were also collected at the beginning (+CM and CM) of the experiment to serve as controls. (B) Ba/F3 cells were starved overnight and the next day were stimulated with IL-3 (1.0 ng/ml) for the indicated times. Cell lysates were analyzed by Western blotting. Cells were also collected at the beginning ( CM) of the experiment to serve as a control. (C) Cell lysates were prepared from Ba/F3 cells stimulated with CM for 15 min and divided into eight aliquots. Three aliquots were digested with either a low (L, 20 U) or high (H, 50 U) dose of calf alkaline phosphatase for 1 h. One set of alkaline phosphatase digestions was heat-inactivated (CIP/Heat) prior to digestion while another set was digested in the presence of the phosphatase inhibitor sodium vanadate (CIP/NaV). As controls, one aliquot of cell lysate was untreated and the other was incubated for 1 h in buffer only. The cell lysates were analyzed by Western blotting. (D) Ba/F3 cells were incubated overnight in the presence or
14 14 absence of conditioned medium. The next day conditioned medium was added back to the cytokine-deprived cells for 1 h. Prior to this, cells were incubated for 1 h in the absence (+CM, -MG) or presence (+CM, +MG) of MG-132. Cell lysates were prepared and analyzed by Western blotting. Cells were also collected at the beginning (+CM and CM) of the experiment to serve as controls. The location of molecular weight markers is shown at the left for all panels. Figure 2: LY inhibits Akt activation and Bim EL phosphorylation induced by IL-3. (A) Ba/F3 cells were incubated overnight in the presence or absence of conditioned medium. The next day starved cells were stimulated for 30 min with either of two doses of recombinant IL-3, as indicated. Stimulated cells were preincubated for 30 min with either 50 µm LY (LY) or DMSO prior to the addition of IL-3. The LY remained present throughout the IL-3 stimulation. Cell lysates were analyzed by Western blotting. Cells were also collected at the beginning (+CM and CM) of the experiment to serve as controls. (B) Similar to panel A except that cells were preincubated in the absence or presence of 100 nm wortmannin (Wort) prior to IL-3 stimulation for 5, 15 or 30 min. (C) Similar to panel A except that cells were preincubated in the absence or presence of 10, 30 or 50 µm LY (LY) prior to being stimulated for 30 min with IL-3. (D) Similar to panel A except that cells were preincubated with 30 µm LY (LY) prior to being stimulated with IL-3 for 1-30 min. (E) Ba/F3 cells were stimulated for 30 min with 1.0 ng/ml of recombinant IL-3. Stimulated cells were preincubated for 30 min with either 50 µm LY (LY), 10 µm PD98509 (PD), 10 µm SP (SP), 15 µm SB (SB), 10 µm U0126 (U0) or DMSO prior to the addition of IL-3. The inhibitors remained present throughout the IL-3 stimulation. Cell lysates were analyzed by Western blotting. Cells were also collected at the beginning ( CM) of the experiment to serve as controls. Figure 3: Gst-Bim EL fusion protein can be phosphorylated in vitro by immunoprecipitated Akt. (A) Ba/F3 cells were starved overnight and the next day were stimulated with IL3 (1.0 ng/ml) for the indicated times. Cells were also collected at the beginning ( CM) of the experiment. Akt was immunoprecipitated from cell lysates using either anti-total-akt (top panel) or anti-phospho-akt (middle panel) antibody. Akt activity was measured in an in vitro kinase assay using Gst-Bim EL as a substrate. The amount of Gst-Bim EL substrate used in each kinase assay was shown by Coomassie Blue staining of the SDS-PAGE gel prior to autoradiography (bottom panel). (B) Aliquots of the samples used in panel A for immunoprecipitations were analyzed by Western blotting using anti-akt antibodies. (C) Left Panel: Similar to panel A except that the cell lysate prepared from IL-3-stimulated cells (15 min) was immunoprecipitated with equal amounts of either anti-total-akt antibody or non-immune rabbit IgG antibody and analyzed for Akt kinase activity. Right Panel: Similar to panel A except that Akt activity was measured in lysates prepared from either starved Ba/F3 cells (-CM) or starved cells that were incubated in the presence or absence of 50 µm LY (LY) prior to IL3 stimulation (1.0 ng/ml) for 30 min Figure 4: Akt phosphorylates Gst-Bim EL fusion protein at Ser 87 in vitro. (A) In vitro kinase assays were performed using equal units of activity of either pure ERK, Akt, Rsk or Gsk-3β using Gst-Bim EL (top left panel) or Gst (bottom left panel) as substrate. As positive controls, the kinase activity of each pure enzyme was tested using its model substrate protein (right panel). (B) Schematic representation of the Gst- Bim EL fusion protein. The fusion protein consisted of amino acids Gly 42 through Thr 101 of human Bim EL fused to the carboxyterminus of Gst (top). Listed below is the amino acid sequence of human Bim EL from Pro 66 through Phe 98. The location of putative Erk (Ser 69 ) and Akt (Ser 87 and Ser 94 ) phosphorylation sites are shown in bold. The location of the carboxy-terminal end of the
15 15 Gst-Bim EL -16 fusion protein at Arg 85 is also indicated. (C) Equal amounts of Gst-Bim EL fusion proteins containing a wild-type Bim EL domain (Bim), a truncated Bim EL domain (Bim-16M), or a single serine to alanine mutation in the Bim EL domain (BimS94A and BimS87A) were used as substrates for in vitro kinase assays employing recombinant Erk (top panel) or Akt (middle panel) as the kinase. The amount of Gst-Bim EL substrate used in each kinase assay was shown by Coomassie Blue staining of the SDS-PAGE gel prior to autoradiography (bottom panel). (D) Results of in vitro kinase assays using the same four Gst-Bim EL proteins as in part C above except that the Akt was immunoprecipitated from Ba/F3 cell lysate prepared from cells stimulated with IL-3 for 15 min (top panel). The amount of Gst-Bim EL substrate used in each kinase assay is shown in bottom panel. Figure 5: Akt stimulation promotes Bim EL binding to proteins. (A) Ba/F3 cells were starved overnight and the next day were stimulated with IL3 (1.0 ng/ml) for the times indicated. Bim was immunoprecipitated from cell lysates and the amount of protein coimmunoprecipitated with Bim was determined by Western blotting (top panel). The amount of Bim (middle panel) and (bottom panel) protein in the original cell lysates was also determined by Western blotting. (B) Hep3B cells were serum-starved for 48 h prior to the addition of insulin for the times indicated. Bim was immunoprecipitated from cell lysates and the amount of protein co-immunoprecipitated with Bim was determined by Western blotting (top panel). The amount of phospho-akt (second panel), phospho-erk (third panel) and Hsp-90 (bottom panel) protein in the original Hep3B cell lysates was also determined by Western blotting. Figure 6: The S87A mutant of Bim EL is a more potent inducer of cell death compared to wildtype Bim EL. (A) Duplicate plates of FAO cells were transiently transfected with either 5 or 15 µg of wild-type Bim EL vector or 15 µg of GFP vector using Lipofectamine Plus. After 24 h, the cells were harvested and lysates prepared for analysis of either DNA ladder formation (left panel) or Bim EL expression by Western blotting (right panel). In the left panel, lane 1 contains a standard 100 bp DNA ladder as a size marker. (B) FAO cells were co-transfected with TK-Renilla vector and increasing concentrations of wild-type Bim EL or GFP vector, as indicated. After 24 h, the cells were lysed and assayed for luciferase activity. The values shown represent the average of quadruplicate wells + SD of a representative experiment. (C) Similar to panel A except that four different Bim EL vector constructs were used in transfections: wild-type and S69A, S87A and S94A mutants. Only 5 µg of vector was used in transfections for DNA ladder experiments (top panel). Western blotting results are shown in the bottom panels. (D) Similar to panel B except that the four different Bim EL vector constructs mentioned above were tested. References 1. O Conner, L., Strasser, A., O Reilly, L. A., Hausmann, G., Adams, J. A., Cory, S., and Huang, D. C. S. (1998) EMBO J 17, Puthalakath, H., and Strasser, A. (2002) Cell Death Differentiation 9, Gross, A., McDonnell, J.M., and Korsmeyer, S.J. (1999) Genes & Development 13, O Reilly, L. A., Cullen, L., Visvader, J., Lindeman, G. J., Print, C., Bath, M. L., Huang, D. C. S., and Strasser, A. (2000) Am J Pathol 157, Bouillet, P., Zhand, L. C., Huang, D. C. S., Webb, G. C., Bottema, C. D. K., Shore, P., Eyre, H. J., Sutherland, G. R., and Adams, J.M. (2001) Mammalian Genome 12, U, M., Miyashita, T., Shikama, Y., Tadokoro, K., and Yamada, M. (2001) FEBS Lett 509, Marani, M., Tenev, T., Hancock, D., Downward, J. and Lemoine, N. R. (2002) Mol Cell Biol 22,
16 16 8. Liu, J.-W., Chandra, D., Tang, S.-H., Chopra, D., and Tang, D. G. (2002) Cancer Res 62, Puthalath, H., Huang, D. C. S., O Reilly, L. A., King, S. M., and Strasser, A. (1999) Mol Cell 3, Liu, X., Dai, S., Zhu, Y., Marrack, P., and Kappler, J.W. (2003) Immunity 19, Yamaguchi, H., and Wang, H.-G. (2002) J Biol Chem 277, Terradillos, O., Montessuit, S., Huang, D.C.S., and Martinou, J.-C. (2002) FEBS Lett 522, Wilson-Annan, J., O Reilly, L.A., Crawford, S.A., Hausmann, G., Beaumont, J.G., Parma, L.P., Chen, L., Lackmann, M., Lithgow, T., Hinds, M.G., Day, C.L., Adams, J.M., and Huang, D.C. (2003) J Cell Biol 162, Stahl, M., Dijkers, P.F., Kops, G.J.P.L., Lens, S.M.A., Coffer, P.J., Burgering, B.M.T., and Medema, R.H. (2002) J Immunol 168, Dijkers, P.F., Medema, R.H., Lammers, J.J., Koenderman, L., and Coffer, P.J. (2000) Curr Biol 10, Shinjyo, T., Kuribara, R., Inukai, T., Hosoi, H., Kinoshita, T., Miyajima, A., Houghton, P., Look, A.T., Ozawa, K., and Inaba, T. (2001) Mol Cell Biol 21, Dijkers, P.F., Birkenkamp, K.U., Lam, E.W.-F., Thomas, N.S.B., Lammers, J.- W.J., Koenderman, L., and Coffer, P.J. (2002) J Cell Biol 156, Putcha, G.V., Moulder, K.L., Golden, J.P., Bouillet, P., Adams, J.A., Strasser, A., and Johnson, E.M., (2001) Neuron 29, Whitfield, J., Neame, S.J., Paquet, L., Bernard, O., and Ham, J. (2001) Neuron 29, Harris, C.A., and Johnson, E.M. (2001) J Biol Chem 276, Biswas, S.C., and Greene, L.A. (2002) J Biol Chem 277, Wildey, G.M., Patil, S., and Howe, P.H. (2003) J Biol Chem 278, Brunet, A., Bonni, A., Zigmond, M.J., Lin, M.Z., Juo, P., Hu, L.S., Anderson, M.J., Arden, K.C., Blenis, J., and Greenberg, M.E. (1999) Cell 96, Putcha, G.V., Le, S., Frank, S., Besirli, C.G., Clark, K., Chu, B., Alix, S., Youle, R.J., LaMarche, A., Maroney, A.C., and Johnson, E.M. (2003) Neuron 38, Ley, R., Ewings, K.E., Hadfield, K., Howes, E., Balmanno, K., and Cook, S.J. (2004) J Biol Chem 279, Luciano, F., Jacquel, A., Colosetti, P., Herrant, M., Cagnol, S., Pages, G., and Auberger, P. (2003) Oncogene 22, Seward, R.J., von Haller, R.D., Aebersold, R., Huber, B.T. (2003) Mol Immunol 39, Akiyama, T., Bouillet, P., Miyazaki, T., Kadono, Y., Chikuda, H., Chung, U., Fukuda, A., Hikita, A., Seto, H., Okada, T., Inaba, T., Sanjay, A., Baron, R., Kawaguchi, H., Oda, H., Nakamura, K., Strasser, A., and Tanaka, S. (2003) EMBO 22, Weston, C.R., Balmanno, K., Chalmers, C., Hadfield, K., Molton, S.A., Ley, R., Wagner, E.F., and Cook, S.J. (2003) Oncogene 22, Marani, M., Hancock, D., Lopes, R., Tenev, T., Downward, J., and Lemoine, N.R. (2003) Oncogene 23, Ley, R., Balmanno, K., Hadfield, K., Weston, C., and Cook, S.J. (2003) J Biol Chem 278, Ymer, S., Tucker, W.Q.J., Sanderson, C.J., Hapel, A.J., Campbell, H.D., and Young, I.G. (1985) Nature 317, Craddock, B.L., Hobbs, J., Edmead, C.E., and Welham, M.J. (2001) J Biol Chem 276, Lei, K., and Davis, R.J. (2003) Proc Natl Acad Sci USA 100, Alessi, D.R., Caudwell, F.B., Andjelkovic, M., Hemmings, B.A., and Cohen, P. (1996) FEBS Letters 399, Lawlor, M.A., and Alessi, D.R. (2001) J Cell Sci 114, Brazil, D.P., and Hemmings, B.A. TRENDS Biochem 26,
17 Datta, S.R., Brunet, A., and Greenberg, M.E. (1999) Genes & Devel 13, Datta, S.R., Dudek, H., Tao, X., Masters, S., Fu, H., Gotoh, Y., and Greenberg, M.E. (1997) Cell 91, del Peso, L., Gonzalez-Garcia, M., Page, C., Herrera, R., and Nunez, G. (1997) Science 278, Masters, S.C., Yang, H., Datta, S.R., Greenberg, M.E., and Fu, H. (2001) Mol Pharm 60, Chen, R.-H., Chang, M.-C., Su, Y.-H., Tsai, Y.-T., and Kuo, M.-L. (1999) J BiolChem 274, Chuang, L.-Y., Hung, W.-C., Chang, C.- C., and Tsai, J.-H. (1994) Anticancer Res 14, Chen, R.-H., Su, Y.-H., Chuang, R.L.C., and Chang, T.-Y. (1998) Oncogene 17, Harada, H., Quearry, B., Ruiz-Vela, A., and Korsmeyer, S.J. (2004) Proc Natl Acad Sci USA 101, Dijkers, P.F., Medema, R.H., Pals, C., Banerji, L., Thomas, N.S.B., Lam, E.W.- F., Burgering, B.M.T., Raaijmakers, J.A.M., Lammers, J.-W.J., Koenderman, L., and Coffer, P.J. (2000) Mol Cell Biol 20, Craddock, B.L., Orchiston, E.A., Hinton, H.J., and Welham, M.J. (1999) J Biol Chem 274, Hinton, H.J., and Welham, M.J. (1999) J Immunol 162, Ley, R., Hadfield, K., Howes, E., and Cook, S.J. (2005) J Biol Chem 280, Gilley, J., Coffer, P.J., and Ham, J. (2003) J Cell Biol 162, Sunters, A., Fernandez de Mattos, S., Stahl, M., Brosens, J.J., Zoumpoulidou, G., Saunders, C.A., Coffer, P.J., Medema, R.H., Coombes, R.C., and Lam, E.W.-F. (2003) J Biol Chem 278, Essafi, A., Fernandez de Mattos, S., Hassen, T.A.M., Soeiro, I., Mufti, G.J., Thomas, N.S.B., Medema, R.H., and Lam, E.W.-F. (2005) Oncogene 24, Domina, A.M., Vrana, J.A., Gregory, M.A., Hann, S.R., and Craig, R.W. (2004) Oncogene 23, Bonni, A., Brunet, A., West, A.E., Datta, S.R., Takasu, M.A., and Greenberg, M.E. (1999) Science 286, Bertolotto, C., Maulin, L., Filippa, N., Baier, G., and Auberger, P. (2000) J Biol Chem 275, Tan, Y., Demeter, M.R., Ruan, H., and Comb, M.J. (2000) J Biol Chem 275,

18
19
20
21
22
23
24
25
SUPPLEMENTARY INFORMATION
 Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
Supplementary data Supplementary Figure 1 Supplementary Figure 2
 Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
MEK1 Assay Kit 1 Catalog # Lot # 16875
 MEK1 Assay Kit 1 Kit Components Assay Dilution Buffer (ADB), Catalog # 20-108. Three vials, each containing 1.0ml of assay dilution buffer (20mM MOPS, ph 7.2, 25mM ß-glycerol phosphate, 5mM EGTA, 1mM sodium
MEK1 Assay Kit 1 Kit Components Assay Dilution Buffer (ADB), Catalog # 20-108. Three vials, each containing 1.0ml of assay dilution buffer (20mM MOPS, ph 7.2, 25mM ß-glycerol phosphate, 5mM EGTA, 1mM sodium
Supporting Online Material Material and Methods References Supplemental Figures S1, S2, and S3
 Supporting Online Material Material and Methods References Supplemental Figures S1, S2, and S3 Sarbassov et al. 1 Material and Methods Materials Reagents were obtained from the following sources: protein
Supporting Online Material Material and Methods References Supplemental Figures S1, S2, and S3 Sarbassov et al. 1 Material and Methods Materials Reagents were obtained from the following sources: protein
FOXO Reporter Kit PI3K/AKT Pathway Cat. #60643
 Data Sheet FOXO Reporter Kit PI3K/AKT Pathway Cat. #60643 Background The PI3K/AKT signaling pathway is essential for cell growth and survival. Disruption of this pathway or its regulation has been linked
Data Sheet FOXO Reporter Kit PI3K/AKT Pathway Cat. #60643 Background The PI3K/AKT signaling pathway is essential for cell growth and survival. Disruption of this pathway or its regulation has been linked
RayBio KinaseSTAR TM Akt Activity Assay Kit
 Activity Assay Kit User Manual Version 1.0 March 13, 2015 RayBio KinaseSTAR TM Akt Activity Kit Protocol (Cat#: 68AT-Akt-S40) RayBiotech, Inc. We Provide You With Excellent Support And Service Tel:(Toll
Activity Assay Kit User Manual Version 1.0 March 13, 2015 RayBio KinaseSTAR TM Akt Activity Kit Protocol (Cat#: 68AT-Akt-S40) RayBiotech, Inc. We Provide You With Excellent Support And Service Tel:(Toll
Phospho-AKT Sampler Kit
 Phospho-AKT Sampler Kit E 0 5 1 0 0 3 Kits Includes Cat. Quantity Application Reactivity Source Akt (Ab-473) Antibody E021054-1 50μg/50μl IHC, WB Human, Mouse, Rat Rabbit Akt (Phospho-Ser473) Antibody
Phospho-AKT Sampler Kit E 0 5 1 0 0 3 Kits Includes Cat. Quantity Application Reactivity Source Akt (Ab-473) Antibody E021054-1 50μg/50μl IHC, WB Human, Mouse, Rat Rabbit Akt (Phospho-Ser473) Antibody
a b G75 G60 Sw-2 Sw-1 Supplementary Figure 1. Structure predictions by I-TASSER Server.
 a b G75 2 2 G60 Sw-2 Sw-1 Supplementary Figure 1. Structure predictions by I-TASSER Server. a. Overlay of top 10 models generated by I-TASSER illustrates the potential effect of 7 amino acid insertion
a b G75 2 2 G60 Sw-2 Sw-1 Supplementary Figure 1. Structure predictions by I-TASSER Server. a. Overlay of top 10 models generated by I-TASSER illustrates the potential effect of 7 amino acid insertion
Protocol for Gene Transfection & Western Blotting
 The schedule and the manual of basic techniques for cell culture Advanced Protocol for Gene Transfection & Western Blotting Schedule Day 1 26/07/2008 Transfection Day 3 28/07/2008 Cell lysis Immunoprecipitation
The schedule and the manual of basic techniques for cell culture Advanced Protocol for Gene Transfection & Western Blotting Schedule Day 1 26/07/2008 Transfection Day 3 28/07/2008 Cell lysis Immunoprecipitation
SUPPLEMENTARY INFORMATION. Supplementary Figures S1-S9. Supplementary Methods
 SUPPLEMENTARY INFORMATION SUMO1 modification of PTEN regulates tumorigenesis by controlling its association with the plasma membrane Jian Huang 1,2#, Jie Yan 1,2#, Jian Zhang 3#, Shiguo Zhu 1, Yanli Wang
SUPPLEMENTARY INFORMATION SUMO1 modification of PTEN regulates tumorigenesis by controlling its association with the plasma membrane Jian Huang 1,2#, Jie Yan 1,2#, Jian Zhang 3#, Shiguo Zhu 1, Yanli Wang
SUPPLEMENTARY MATERIAL
 SUPPLEMENTARY MATERIAL Table S1. Primers and fluorescent probes used for qrt-pcr analysis of relative expression levels of PPP family phosphatases. gene name forward primer, 5-3 probe, 5-3 reverse primer,
SUPPLEMENTARY MATERIAL Table S1. Primers and fluorescent probes used for qrt-pcr analysis of relative expression levels of PPP family phosphatases. gene name forward primer, 5-3 probe, 5-3 reverse primer,
Supplemental material for Hernandez et al. Dicoumarol downregulates human PTTG1/Securin mrna expression. through inhibition of Hsp90
 Supplemental material for Hernandez et al. Dicoumarol downregulates human PTTG1/Securin mrna expression through inhibition of Hsp90 Dicoumarol-Sepharose co-precipitation. Hsp90 inhibitors can co-precipitate
Supplemental material for Hernandez et al. Dicoumarol downregulates human PTTG1/Securin mrna expression through inhibition of Hsp90 Dicoumarol-Sepharose co-precipitation. Hsp90 inhibitors can co-precipitate
Nature Methods: doi: /nmeth Supplementary Figure 1
 Supplementary Figure 1 Subtiligase-catalyzed ligations with ubiquitin thioesters and 10-mer biotinylated peptides. (a) General scheme for ligations between ubiquitin thioesters and 10-mer, biotinylated
Supplementary Figure 1 Subtiligase-catalyzed ligations with ubiquitin thioesters and 10-mer biotinylated peptides. (a) General scheme for ligations between ubiquitin thioesters and 10-mer, biotinylated
supplementary information
 Figure S1 Nucleotide binding status of RagA mutants. Wild type and mutant forms of MycRagA was transfected into HEK293 cells and the transfected cells were labeled with 32 Pphosphate. MycRagA was immunoprecipitated
Figure S1 Nucleotide binding status of RagA mutants. Wild type and mutant forms of MycRagA was transfected into HEK293 cells and the transfected cells were labeled with 32 Pphosphate. MycRagA was immunoprecipitated
Supplementary Figure 1 Role of Raf-1 in TLR2-Dectin-1-mediated cytokine expression
 Supplementary Figure 1 Supplementary Figure 1 Role of Raf-1 in TLR2-Dectin-1-mediated cytokine expression. Quantitative real-time PCR of indicated mrnas in DCs stimulated with TLR2-Dectin-1 agonist zymosan
Supplementary Figure 1 Supplementary Figure 1 Role of Raf-1 in TLR2-Dectin-1-mediated cytokine expression. Quantitative real-time PCR of indicated mrnas in DCs stimulated with TLR2-Dectin-1 agonist zymosan
Supplementary Figure 1. Normal T lymphocyte populations in Dapk -/- mice. (a) Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk
 Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk") Supplementary Figure 1. Normal T lymphocyte populations in Dapk -/- mice. (a) Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk -/- mice were stained for expression of CD4 and CD8.
Supplementary Figure 1. Normal T lymphocyte populations in Dapk -/- mice. (a) Normal thymic development in Dapk -/- mice. Thymocytes from WT and Dapk -/- mice were stained for expression of CD4 and CD8.
p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO
 Supplementary Information p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO Yuri Shibata, Masaaki Oyama, Hiroko Kozuka-Hata, Xiao Han, Yuetsu Tanaka,
Supplementary Information p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO Yuri Shibata, Masaaki Oyama, Hiroko Kozuka-Hata, Xiao Han, Yuetsu Tanaka,
Growth and Differentiation Phosphorylation Sampler Kit
 Growth and Differentiation Phosphorylation Sampler Kit E 0 5 1 0 1 4 Kits Includes Cat. Quantity Application Reactivity Source Akt (Phospho-Ser473) E011054-1 50μg/50μl IHC, WB Human, Mouse, Rat Rabbit
Growth and Differentiation Phosphorylation Sampler Kit E 0 5 1 0 1 4 Kits Includes Cat. Quantity Application Reactivity Source Akt (Phospho-Ser473) E011054-1 50μg/50μl IHC, WB Human, Mouse, Rat Rabbit
SUPPLEMENT. Materials and methods
 SUPPLEMENT Materials and methods Cell culture and reagents Cell media and reagents were from Invitrogen unless otherwise indicated. Antibiotics and Tet-certified serum were from Clontech. In experiments
SUPPLEMENT Materials and methods Cell culture and reagents Cell media and reagents were from Invitrogen unless otherwise indicated. Antibiotics and Tet-certified serum were from Clontech. In experiments
A Hepatocyte Growth Factor Receptor (Met) Insulin Receptor hybrid governs hepatic glucose metabolism SUPPLEMENTARY FIGURES, LEGENDS AND METHODS
 Insulin Receptor hybrid governs hepatic glucose metabolism SUPPLEMENTARY FIGURES, LEGENDS AND METHODS") A Hepatocyte Growth Factor Receptor (Met) Insulin Receptor hybrid governs hepatic glucose metabolism Arlee Fafalios, Jihong Ma, Xinping Tan, John Stoops, Jianhua Luo, Marie C. DeFrances and Reza Zarnegar
A Hepatocyte Growth Factor Receptor (Met) Insulin Receptor hybrid governs hepatic glucose metabolism Arlee Fafalios, Jihong Ma, Xinping Tan, John Stoops, Jianhua Luo, Marie C. DeFrances and Reza Zarnegar
The Schedule and the Manual of Basic Techniques for Cell Culture
 The Schedule and the Manual of Basic Techniques for Cell Culture 1 Materials Calcium Phosphate Transfection Kit: Invitrogen Cat.No.K2780-01 Falcon tube (Cat No.35-2054:12 x 75 mm, 5 ml tube) Cell: 293
The Schedule and the Manual of Basic Techniques for Cell Culture 1 Materials Calcium Phosphate Transfection Kit: Invitrogen Cat.No.K2780-01 Falcon tube (Cat No.35-2054:12 x 75 mm, 5 ml tube) Cell: 293
TRAF6 ubiquitinates TGFβ type I receptor to promote its cleavage and nuclear translocation in cancer
 Supplementary Information TRAF6 ubiquitinates TGFβ type I receptor to promote its cleavage and nuclear translocation in cancer Yabing Mu, Reshma Sundar, Noopur Thakur, Maria Ekman, Shyam Kumar Gudey, Mariya
Supplementary Information TRAF6 ubiquitinates TGFβ type I receptor to promote its cleavage and nuclear translocation in cancer Yabing Mu, Reshma Sundar, Noopur Thakur, Maria Ekman, Shyam Kumar Gudey, Mariya
Supplementary Information
 Supplementary Information Supplementary Figure 1. CD4 + T cell activation and lack of apoptosis after crosslinking with anti-cd3 + anti-cd28 + anti-cd160. (a) Flow cytometry of anti-cd160 (5D.10A11) binding
Supplementary Information Supplementary Figure 1. CD4 + T cell activation and lack of apoptosis after crosslinking with anti-cd3 + anti-cd28 + anti-cd160. (a) Flow cytometry of anti-cd160 (5D.10A11) binding
SUPPLEMENTAL INFORMATION
 SUPPLEMENTAL INFORMATION EXPERIMENTAL PROCEDURES Tryptic digestion protection experiments - PCSK9 with Ab-3D5 (1:1 molar ratio) in 50 mm Tris, ph 8.0, 150 mm NaCl was incubated overnight at 4 o C. The
SUPPLEMENTAL INFORMATION EXPERIMENTAL PROCEDURES Tryptic digestion protection experiments - PCSK9 with Ab-3D5 (1:1 molar ratio) in 50 mm Tris, ph 8.0, 150 mm NaCl was incubated overnight at 4 o C. The
Serum Amyloid A3 Gene Expression in Adipocytes is an Indicator. of the Interaction with Macrophages
 Serum Amyloid A3 Gene Expression in Adipocytes is an Indicator of the Interaction with Macrophages Yohei Sanada, Takafumi Yamamoto, Rika Satake, Akiko Yamashita, Sumire Kanai, Norihisa Kato, Fons AJ van
Serum Amyloid A3 Gene Expression in Adipocytes is an Indicator of the Interaction with Macrophages Yohei Sanada, Takafumi Yamamoto, Rika Satake, Akiko Yamashita, Sumire Kanai, Norihisa Kato, Fons AJ van
Construction of a hepatocellular carcinoma cell line that stably expresses stathmin with a Ser25 phosphorylation site mutation
 Construction of a hepatocellular carcinoma cell line that stably expresses stathmin with a Ser25 phosphorylation site mutation J. Du 1, Z.H. Tao 2, J. Li 2, Y.K. Liu 3 and L. Gan 2 1 Department of Chemistry,
Construction of a hepatocellular carcinoma cell line that stably expresses stathmin with a Ser25 phosphorylation site mutation J. Du 1, Z.H. Tao 2, J. Li 2, Y.K. Liu 3 and L. Gan 2 1 Department of Chemistry,
Supplementary Figure 1.TRIM33 binds β-catenin in the nucleus. a & b, Co-IP of endogenous TRIM33 with β-catenin in HT-29 cells (a) and HEK 293T cells
 and HEK 293T cells") Supplementary Figure 1.TRIM33 binds β-catenin in the nucleus. a & b, Co-IP of endogenous TRIM33 with β-catenin in HT-29 cells (a) and HEK 293T cells (b). TRIM33 was immunoprecipitated, and the amount of
Supplementary Figure 1.TRIM33 binds β-catenin in the nucleus. a & b, Co-IP of endogenous TRIM33 with β-catenin in HT-29 cells (a) and HEK 293T cells (b). TRIM33 was immunoprecipitated, and the amount of
Mammalian Tissue Protein Extraction Reagent
 Mammalian Tissue Protein Extraction Reagent Catalog number: AR0101 Boster s Mammalian Tissue Protein Extraction Reagent is a ready-to-use Western blot related reagent solution used for efficient extraction
Mammalian Tissue Protein Extraction Reagent Catalog number: AR0101 Boster s Mammalian Tissue Protein Extraction Reagent is a ready-to-use Western blot related reagent solution used for efficient extraction
Islet viability assay and Glucose Stimulated Insulin Secretion assay RT-PCR and Western Blot
 Islet viability assay and Glucose Stimulated Insulin Secretion assay Islet cell viability was determined by colorimetric (3-(4,5-dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide assay using CellTiter
Islet viability assay and Glucose Stimulated Insulin Secretion assay Islet cell viability was determined by colorimetric (3-(4,5-dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide assay using CellTiter
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION doi:1.138/nature9814 a A SHARPIN FL B SHARPIN ΔNZF C SHARPIN T38L, F39V b His-SHARPIN FL -1xUb -2xUb -4xUb α-his c Linear 4xUb -SHARPIN FL -SHARPIN TF_LV -SHARPINΔNZF -SHARPIN
SUPPLEMENTARY INFORMATION doi:1.138/nature9814 a A SHARPIN FL B SHARPIN ΔNZF C SHARPIN T38L, F39V b His-SHARPIN FL -1xUb -2xUb -4xUb α-his c Linear 4xUb -SHARPIN FL -SHARPIN TF_LV -SHARPINΔNZF -SHARPIN
SUPPLEMENTARY INFORMATION
 Supplementary Table 1. Cell sphingolipids and S1P bound to endogenous TRAF2. Sphingolipid Cell pmol/mg TRAF2 immunoprecipitate pmol/mg Sphingomyelin 4200 ± 250 Not detected Monohexosylceramide 311 ± 18
Supplementary Table 1. Cell sphingolipids and S1P bound to endogenous TRAF2. Sphingolipid Cell pmol/mg TRAF2 immunoprecipitate pmol/mg Sphingomyelin 4200 ± 250 Not detected Monohexosylceramide 311 ± 18
The functional investigation of the interaction between TATA-associated factor 3 (TAF3) and p53 protein
 and p53 protein") THESIS BOOK The functional investigation of the interaction between TATA-associated factor 3 (TAF3) and p53 protein Orsolya Buzás-Bereczki Supervisors: Dr. Éva Bálint Dr. Imre Miklós Boros University of
THESIS BOOK The functional investigation of the interaction between TATA-associated factor 3 (TAF3) and p53 protein Orsolya Buzás-Bereczki Supervisors: Dr. Éva Bálint Dr. Imre Miklós Boros University of
Introduction: 年 Fas signal-mediated apoptosis. PI3K/Akt
 Fas-ligand (CD95-L; Fas-L) Fas (CD95) Fas (apoptosis) 年 了 不 度 Fas Fas-L 力 不 Fas/Fas-L T IL-10Fas/Fas-L 不 年 Fas signal-mediated apoptosis 度降 不 不 力 U-118, HeLa, A549, Huh-7 MCF-7, HepG2. PI3K/Akt FasPI3K/Akt
Fas-ligand (CD95-L; Fas-L) Fas (CD95) Fas (apoptosis) 年 了 不 度 Fas Fas-L 力 不 Fas/Fas-L T IL-10Fas/Fas-L 不 年 Fas signal-mediated apoptosis 度降 不 不 力 U-118, HeLa, A549, Huh-7 MCF-7, HepG2. PI3K/Akt FasPI3K/Akt
HEK293FT cells were transiently transfected with reporters, N3-ICD construct and
 Supplementary Information Luciferase reporter assay HEK293FT cells were transiently transfected with reporters, N3-ICD construct and increased amounts of wild type or kinase inactive EGFR. Transfections
Supplementary Information Luciferase reporter assay HEK293FT cells were transiently transfected with reporters, N3-ICD construct and increased amounts of wild type or kinase inactive EGFR. Transfections
Part-4. Cell cycle regulatory protein 5 (Cdk5) A novel target of ERK in Carb induced cell death
 A novel target of ERK in Carb induced cell death") Part-4 Cell cycle regulatory protein 5 (Cdk5) A novel target of ERK in Carb induced cell death 95 1. Introduction The process of replicating DNA and dividing cells can be described as a series of coordinated
Part-4 Cell cycle regulatory protein 5 (Cdk5) A novel target of ERK in Carb induced cell death 95 1. Introduction The process of replicating DNA and dividing cells can be described as a series of coordinated
hexahistidine tagged GRP78 devoid of the KDEL motif (GRP78-His) on SDS-PAGE. This
 on SDS-PAGE. This") SUPPLEMENTAL FIGURE LEGEND Fig. S1. Generation and characterization of. (A) Coomassie staining of soluble hexahistidine tagged GRP78 devoid of the KDEL motif (GRP78-His) on SDS-PAGE. This protein was expressed
SUPPLEMENTAL FIGURE LEGEND Fig. S1. Generation and characterization of. (A) Coomassie staining of soluble hexahistidine tagged GRP78 devoid of the KDEL motif (GRP78-His) on SDS-PAGE. This protein was expressed
Supplementary Information POLO-LIKE KINASE 1 FACILITATES LOSS OF PTEN-INDUCED PROSTATE CANCER FORMATION
 Supplementary Information POLO-LIKE KINASE 1 FACILITATES LOSS OF PTEN-INDUCED PROSTATE CANCER FORMATION X. Shawn Liu 1, 3, Bing Song 2, 3, Bennett D. Elzey 3, 4, Timothy L. Ratliff 3, 4, Stephen F. Konieczny
Supplementary Information POLO-LIKE KINASE 1 FACILITATES LOSS OF PTEN-INDUCED PROSTATE CANCER FORMATION X. Shawn Liu 1, 3, Bing Song 2, 3, Bennett D. Elzey 3, 4, Timothy L. Ratliff 3, 4, Stephen F. Konieczny
HIV-1 Virus-like Particle Budding Assay Nathan H Vande Burgt, Luis J Cocka * and Paul Bates
 HIV-1 Virus-like Particle Budding Assay Nathan H Vande Burgt, Luis J Cocka * and Paul Bates Department of Microbiology, Perelman School of Medicine at the University of Pennsylvania, Philadelphia, USA
HIV-1 Virus-like Particle Budding Assay Nathan H Vande Burgt, Luis J Cocka * and Paul Bates Department of Microbiology, Perelman School of Medicine at the University of Pennsylvania, Philadelphia, USA
Online Data Supplement. Anti-aging Gene Klotho Enhances Glucose-induced Insulin Secretion by Upregulating Plasma Membrane Retention of TRPV2
 Online Data Supplement Anti-aging Gene Klotho Enhances Glucose-induced Insulin Secretion by Upregulating Plasma Membrane Retention of TRPV2 Yi Lin and Zhongjie Sun Department of physiology, college of
Online Data Supplement Anti-aging Gene Klotho Enhances Glucose-induced Insulin Secretion by Upregulating Plasma Membrane Retention of TRPV2 Yi Lin and Zhongjie Sun Department of physiology, college of
For the rapid, sensitive and accurate quantification of Ras in various samples
 ab128504 Ras Assay Kit Instructions for Use For the rapid, sensitive and accurate quantification of Ras in various samples This product is for research use only and is not intended for diagnostic use.
ab128504 Ras Assay Kit Instructions for Use For the rapid, sensitive and accurate quantification of Ras in various samples This product is for research use only and is not intended for diagnostic use.
Supplementary Figure 1 CD4 + T cells from PKC-θ null mice are defective in NF-κB activation during T cell receptor signaling. CD4 + T cells were
 Supplementary Figure 1 CD4 + T cells from PKC-θ null mice are defective in NF-κB activation during T cell receptor signaling. CD4 + T cells were isolated from wild type (PKC-θ- WT) or PKC-θ null (PKC-θ-KO)
Supplementary Figure 1 CD4 + T cells from PKC-θ null mice are defective in NF-κB activation during T cell receptor signaling. CD4 + T cells were isolated from wild type (PKC-θ- WT) or PKC-θ null (PKC-θ-KO)
A Novel mtor-regulated Phosphorylation Site in Elongation Factor 2 Kinase Modulates the Activity of the Kinase and Its Binding to Calmodulin
 REFERENCES CONTENT ALERTS A Novel mtor-regulated Phosphorylation Site in Elongation Factor 2 Kinase Modulates the Activity of the Kinase and Its Binding to Calmodulin Gareth J. Browne and Christopher G.
REFERENCES CONTENT ALERTS A Novel mtor-regulated Phosphorylation Site in Elongation Factor 2 Kinase Modulates the Activity of the Kinase and Its Binding to Calmodulin Gareth J. Browne and Christopher G.
Cell Lysis Buffer. Catalog number: AR0103
 Cell Lysis Buffer Catalog number: AR0103 Boster s Cell Lysis Buffer is a ready-to-use Western blot related reagent solution used for efficient extraction of total soluble protein in nondenatured state
Cell Lysis Buffer Catalog number: AR0103 Boster s Cell Lysis Buffer is a ready-to-use Western blot related reagent solution used for efficient extraction of total soluble protein in nondenatured state
Supplementary Figure 1
 Supplementary Figure 1 Supplementary Figure 1. Neither the activation nor suppression of the MAPK pathway affects the ASK1/Vif interaction. (a, b) HEK293 cells were cotransfected with plasmids encoding
Supplementary Figure 1 Supplementary Figure 1. Neither the activation nor suppression of the MAPK pathway affects the ASK1/Vif interaction. (a, b) HEK293 cells were cotransfected with plasmids encoding
Validation & Assay Performance Summary
 Validation & Assay Performance Summary LanthaScreen IGF-1R GripTite Cells Cat. no. K1834 Modification Detected: Phosphorylation of Multiple Tyr Residues on IGF-1R LanthaScreen Cellular Assay Validation
Validation & Assay Performance Summary LanthaScreen IGF-1R GripTite Cells Cat. no. K1834 Modification Detected: Phosphorylation of Multiple Tyr Residues on IGF-1R LanthaScreen Cellular Assay Validation
Problem Set 8 Key 1 of 8
 7.06 2003 Problem Set 8 Key 1 of 8 7.06 2003 Problem Set 8 Key 1. As a bright MD/PhD, you are interested in questions about the control of cell number in the body. Recently, you've seen three patients
7.06 2003 Problem Set 8 Key 1 of 8 7.06 2003 Problem Set 8 Key 1. As a bright MD/PhD, you are interested in questions about the control of cell number in the body. Recently, you've seen three patients
HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation
 SUPPLEMENTARY INFORMATION Materials and Methods Human cell lines and culture conditions HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation in exon 20 of BRCA1
SUPPLEMENTARY INFORMATION Materials and Methods Human cell lines and culture conditions HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation in exon 20 of BRCA1
Supplementary Figure 1. PD-L1 is glycosylated in cancer cells. (a) Western blot analysis of PD-L1 in breast cancer cells. (b) Western blot analysis
 Western blot analysis of PD-L1 in breast cancer cells. (b) Western blot analysis") Supplementary Figure 1. PD-L1 is glycosylated in cancer cells. (a) Western blot analysis of PD-L1 in breast cancer cells. (b) Western blot analysis of PD-L1 in ovarian cancer cells. (c) Western blot analysis
Supplementary Figure 1. PD-L1 is glycosylated in cancer cells. (a) Western blot analysis of PD-L1 in breast cancer cells. (b) Western blot analysis of PD-L1 in ovarian cancer cells. (c) Western blot analysis
Data Sheet PD-1 / NFAT Reporter - Jurkat Cell Line Catalog #: 60535
 Data Sheet PD-1 / NFAT Reporter - Jurkat Cell Line Catalog #: 60535 Product Description Recombinant Jurkat T cell expressing firefly luciferase gene under the control of NFAT response elements with constitutive
Data Sheet PD-1 / NFAT Reporter - Jurkat Cell Line Catalog #: 60535 Product Description Recombinant Jurkat T cell expressing firefly luciferase gene under the control of NFAT response elements with constitutive
Signal Transduction Pathways Involved in Phosphorylation and
 JBC Papers in Press. Published on March 28, 2001 as Manuscript M009047200 Signal Transduction Pathways Involved in Phosphorylation and Activation of p70 S6K Following Exposure to UVA Irradiation* Yiguo
JBC Papers in Press. Published on March 28, 2001 as Manuscript M009047200 Signal Transduction Pathways Involved in Phosphorylation and Activation of p70 S6K Following Exposure to UVA Irradiation* Yiguo
Supplemental Data. TGF-β-mediated mir-181a expression promotes breast cancer metastasis by targeting Bim.
 Supplemental Data TGF-β-mediated mir-181a expression promotes breast cancer metastasis by targeting Bim. Molly A. Taylor 1, Khalid Sossey-Alaoui 2, Cheryl L. Thompson 3, David Danielpour 4, and William
Supplemental Data TGF-β-mediated mir-181a expression promotes breast cancer metastasis by targeting Bim. Molly A. Taylor 1, Khalid Sossey-Alaoui 2, Cheryl L. Thompson 3, David Danielpour 4, and William
MicroRNA sponges: competitive inhibitors of small RNAs in mammalian cells
 MicroRNA sponges: competitive inhibitors of small RNAs in mammalian cells Margaret S Ebert, Joel R Neilson & Phillip A Sharp Supplementary figures and text: Supplementary Figure 1. Effect of sponges on
MicroRNA sponges: competitive inhibitors of small RNAs in mammalian cells Margaret S Ebert, Joel R Neilson & Phillip A Sharp Supplementary figures and text: Supplementary Figure 1. Effect of sponges on
MTC-TT and TPC-1 cell lines were cultured in RPMI medium (Gibco, Breda, The Netherlands)
") Supplemental data Materials and Methods Cell culture MTC-TT and TPC-1 cell lines were cultured in RPMI medium (Gibco, Breda, The Netherlands) supplemented with 15% or 10% (for TPC-1) fetal bovine serum
Supplemental data Materials and Methods Cell culture MTC-TT and TPC-1 cell lines were cultured in RPMI medium (Gibco, Breda, The Netherlands) supplemented with 15% or 10% (for TPC-1) fetal bovine serum
(A) PCR primers (arrows) designed to distinguish wild type (P1+P2), targeted (P1+P2) and excised (P1+P3)14-
 PCR primers (arrows) designed to distinguish wild type (P1+P2), targeted (P1+P2) and excised (P1+P3)14-") 1 Supplemental Figure Legends Figure S1. Mammary tumors of ErbB2 KI mice with 14-3-3σ ablation have elevated ErbB2 transcript levels and cell proliferation (A) PCR primers (arrows) designed to distinguish
1 Supplemental Figure Legends Figure S1. Mammary tumors of ErbB2 KI mice with 14-3-3σ ablation have elevated ErbB2 transcript levels and cell proliferation (A) PCR primers (arrows) designed to distinguish
Supplemental Information
 Supplemental Information Tobacco-specific Carcinogen Induces DNA Methyltransferases 1 Accumulation through AKT/GSK3β/βTrCP/hnRNP-U in Mice and Lung Cancer patients Ruo-Kai Lin, 1 Yi-Shuan Hsieh, 2 Pinpin
Supplemental Information Tobacco-specific Carcinogen Induces DNA Methyltransferases 1 Accumulation through AKT/GSK3β/βTrCP/hnRNP-U in Mice and Lung Cancer patients Ruo-Kai Lin, 1 Yi-Shuan Hsieh, 2 Pinpin
T H E J O U R N A L O F C E L L B I O L O G Y
 T H E J O U R N A L O F C E L L B I O L O G Y Supplemental material Krenn et al., http://www.jcb.org/cgi/content/full/jcb.201110013/dc1 Figure S1. Levels of expressed proteins and demonstration that C-terminal
T H E J O U R N A L O F C E L L B I O L O G Y Supplemental material Krenn et al., http://www.jcb.org/cgi/content/full/jcb.201110013/dc1 Figure S1. Levels of expressed proteins and demonstration that C-terminal
Protein Dephosphorylation Methods
 Protein Dephosphorylation Methods Phosphospecific antibodies are designed to differentiate between the phosphorylated and the non-phosphorylated states of a protein. The method to determine if or how well
Protein Dephosphorylation Methods Phosphospecific antibodies are designed to differentiate between the phosphorylated and the non-phosphorylated states of a protein. The method to determine if or how well
Electron micrograph of phosphotungstanic acid-stained exosomes derived from murine
 1 SUPPLEMENTARY INFORMATION SUPPLEMENTARY FIGURES Supplementary Figure 1. Physical properties of murine DC-derived exosomes. a, Electron micrograph of phosphotungstanic acid-stained exosomes derived from
1 SUPPLEMENTARY INFORMATION SUPPLEMENTARY FIGURES Supplementary Figure 1. Physical properties of murine DC-derived exosomes. a, Electron micrograph of phosphotungstanic acid-stained exosomes derived from
Validation & Assay Performance Summary
 Validation & Assay Performance Summary CellSensor DBE-bla MDA-MB-468 Cell Line Cat. no. K1814 Pathway Description The phosphatidylinositol-3-kinase (PI3K) signaling cascade is essential for cell growth
Validation & Assay Performance Summary CellSensor DBE-bla MDA-MB-468 Cell Line Cat. no. K1814 Pathway Description The phosphatidylinositol-3-kinase (PI3K) signaling cascade is essential for cell growth
Receptor mediated Signal Transduction
 Receptor mediated Signal Transduction G-protein-linked receptors adenylyl cyclase camp PKA Organization of receptor protein-tyrosine kinases From G.M. Cooper, The Cell. A molecular approach, 2004, third
Receptor mediated Signal Transduction G-protein-linked receptors adenylyl cyclase camp PKA Organization of receptor protein-tyrosine kinases From G.M. Cooper, The Cell. A molecular approach, 2004, third
Effec<ve Use of PI3K and MEK Inhibitors to Treat Mutant K Ras G12D and PIK3CA H1047R Murine Lung Cancers
 Effec
Effec
Chromatin IP (Isw2) Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles.
 Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles.") Chromatin IP (Isw2) 7/01 Toshi last update: 06/15 Reagents Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles. 2.5 M glycine. TBS:
Chromatin IP (Isw2) 7/01 Toshi last update: 06/15 Reagents Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles. 2.5 M glycine. TBS:
Cells and reagents. Synaptopodin knockdown (1) and dynamin knockdown (2)
 and dynamin knockdown (2)") Supplemental Methods Cells and reagents. Synaptopodin knockdown (1) and dynamin knockdown (2) podocytes were cultured as described previously. Staurosporine, angiotensin II and actinomycin D were all obtained
Supplemental Methods Cells and reagents. Synaptopodin knockdown (1) and dynamin knockdown (2) podocytes were cultured as described previously. Staurosporine, angiotensin II and actinomycin D were all obtained
Data Sheet TIGIT / NFAT Reporter - Jurkat Cell Line Catalog #60538
 Data Sheet TIGIT / NFAT Reporter - Jurkat Cell Line Catalog #60538 Background: TIGIT is a co-inhibitory receptor that is highly expressed in Natural Killer (NK) cells, activated CD4+, CD8+ and regulatory
Data Sheet TIGIT / NFAT Reporter - Jurkat Cell Line Catalog #60538 Background: TIGIT is a co-inhibitory receptor that is highly expressed in Natural Killer (NK) cells, activated CD4+, CD8+ and regulatory
Human PKA (Protein Kinase A) Activity Assay Kit
 Activity Assay Kit") Human PKA (Protein Kinase A) Activity Assay Kit CATALOG NO: IRAAKT2532 LOT NO: SAMPLE INTENDED USE The PKA (Protein Kinase A) Activity kit is designed to quantitatively measure PKA activity in a variety
Human PKA (Protein Kinase A) Activity Assay Kit CATALOG NO: IRAAKT2532 LOT NO: SAMPLE INTENDED USE The PKA (Protein Kinase A) Activity kit is designed to quantitatively measure PKA activity in a variety
RAW264.7 cells stably expressing control shrna (Con) or GSK3b-specific shrna (sh-
 or GSK3b-specific shrna (sh-") 1 a b Supplementary Figure 1. Effects of GSK3b knockdown on poly I:C-induced cytokine production. RAW264.7 cells stably expressing control shrna (Con) or GSK3b-specific shrna (sh- GSK3b) were stimulated
1 a b Supplementary Figure 1. Effects of GSK3b knockdown on poly I:C-induced cytokine production. RAW264.7 cells stably expressing control shrna (Con) or GSK3b-specific shrna (sh- GSK3b) were stimulated
Essential Medium, containing 10% fetal bovine serum, 100 U/ml penicillin and 100 µg/ml streptomycin. Huvec were cultured in
 Supplemental data Methods Cell culture media formulations A-431 and U-87 MG cells were maintained in Dulbecco s Modified Eagle s Medium. FaDu cells were cultured in Eagle's Minimum Essential Medium, containing
Supplemental data Methods Cell culture media formulations A-431 and U-87 MG cells were maintained in Dulbecco s Modified Eagle s Medium. FaDu cells were cultured in Eagle's Minimum Essential Medium, containing
Characterization of the role of EGF-A of low density lipoprotein receptor in PCSK9 binding
 Characterization of the role of EGF-A of low density lipoprotein receptor in PCSK9 binding Hong-mei Gu, 1 Ayinuer Adijiang, 1 Matthew Mah, and Da-wei Zhang 2 Departments of Pediatrics and Biochemistry,
Characterization of the role of EGF-A of low density lipoprotein receptor in PCSK9 binding Hong-mei Gu, 1 Ayinuer Adijiang, 1 Matthew Mah, and Da-wei Zhang 2 Departments of Pediatrics and Biochemistry,
Supplementary Material for
 Supplementary Material for Parathyroid Hormone Signaling through Low-density-lipoprotein-related Protein 6 Mei Wan, Chaozhe Yang, Jun Li, Xiangwei Wu, Hongling Yuan, Hairong Ma, Xi He, Shuyi Nie, Chenbei
Supplementary Material for Parathyroid Hormone Signaling through Low-density-lipoprotein-related Protein 6 Mei Wan, Chaozhe Yang, Jun Li, Xiangwei Wu, Hongling Yuan, Hairong Ma, Xi He, Shuyi Nie, Chenbei
On Line Data Supplement
 On Line Data Supplement Chemicals and Other Materials [γ- 32 P]ATP, L-[ 35 S]methionine, L-[ 3 H]leucine, m 7 GTP-Sepharose, glutathione- Sepharose 4B and ECL reagents were purchased from Amersham Pharmacia
On Line Data Supplement Chemicals and Other Materials [γ- 32 P]ATP, L-[ 35 S]methionine, L-[ 3 H]leucine, m 7 GTP-Sepharose, glutathione- Sepharose 4B and ECL reagents were purchased from Amersham Pharmacia
Electrical Stimulation Control Nerve Regeneration via the p38 Mitogen-activated Protein Kinase and CREB
 Electrical Stimulation Control Nerve Regeneration via the p38 Mitogen-activated Protein Kinase and CREB Kenji Kawamura, Yoshio Kano. Kibi International University, Takahashi-city, Japan. Disclosures: K.
Electrical Stimulation Control Nerve Regeneration via the p38 Mitogen-activated Protein Kinase and CREB Kenji Kawamura, Yoshio Kano. Kibi International University, Takahashi-city, Japan. Disclosures: K.
Rabbit Polyclonal antibody to NFkB p65 (v-rel reticuloendotheliosis viral oncogene homolog A (avian))
)") Datasheet GeneTex, Inc : Toll Free 1-877-GeneTex (1-877-436-3839) Fax:1-949-309-2888 info@genetex.com GeneTex International Corporation : Tel:886-3-6208988 Fax:886-3-6208989 infoasia@genetex.com Date :
Datasheet GeneTex, Inc : Toll Free 1-877-GeneTex (1-877-436-3839) Fax:1-949-309-2888 info@genetex.com GeneTex International Corporation : Tel:886-3-6208988 Fax:886-3-6208989 infoasia@genetex.com Date :
Supplementary Information Supplementary Fig. 1. Elevated Usp9x in melanoma and NRAS mutant melanoma cells are dependent on NRAS for 3D growth.
 Supplementary Information Supplementary Fig. 1. Elevated Usp9x in melanoma and NRAS mutant melanoma cells are dependent on NRAS for 3D growth. a. Immunoblot for Usp9x protein in NRAS mutant melanoma cells
Supplementary Information Supplementary Fig. 1. Elevated Usp9x in melanoma and NRAS mutant melanoma cells are dependent on NRAS for 3D growth. a. Immunoblot for Usp9x protein in NRAS mutant melanoma cells
Luminescent platforms for monitoring changes in the solubility of amylin and huntingtin in living cells
 Electronic Supplementary Material (ESI) for Molecular BioSystems. This journal is The Royal Society of Chemistry 2016 Contents Supporting Information Luminescent platforms for monitoring changes in the
Electronic Supplementary Material (ESI) for Molecular BioSystems. This journal is The Royal Society of Chemistry 2016 Contents Supporting Information Luminescent platforms for monitoring changes in the
Cyclin I activates Cdk5 and regulates expression of Bcl-2 and Bcl-XL in postmitotic mouse cells
 Research article Cyclin I activates Cdk5 and regulates expression of Bcl-2 and Bcl-XL in postmitotic mouse cells Paul T. Brinkkoetter, 1 Paul Olivier, 2 Jimmy S. Wu, 1 Scott Henderson, 1 Ronald D. Krofft,
Research article Cyclin I activates Cdk5 and regulates expression of Bcl-2 and Bcl-XL in postmitotic mouse cells Paul T. Brinkkoetter, 1 Paul Olivier, 2 Jimmy S. Wu, 1 Scott Henderson, 1 Ronald D. Krofft,
Supplementary Fig. 1. GPRC5A post-transcriptionally down-regulates EGFR expression. (a) Plot of the changes in steady state mrna levels versus
 Plot of the changes in steady state mrna levels versus") Supplementary Fig. 1. GPRC5A post-transcriptionally down-regulates EGFR expression. (a) Plot of the changes in steady state mrna levels versus changes in corresponding proteins between wild type and Gprc5a-/-
Supplementary Fig. 1. GPRC5A post-transcriptionally down-regulates EGFR expression. (a) Plot of the changes in steady state mrna levels versus changes in corresponding proteins between wild type and Gprc5a-/-
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION FOR Liver X Receptor α mediates hepatic triglyceride accumulation through upregulation of G0/G1 Switch Gene 2 (G0S2) expression I: SUPPLEMENTARY METHODS II: SUPPLEMENTARY FIGURES
SUPPLEMENTARY INFORMATION FOR Liver X Receptor α mediates hepatic triglyceride accumulation through upregulation of G0/G1 Switch Gene 2 (G0S2) expression I: SUPPLEMENTARY METHODS II: SUPPLEMENTARY FIGURES
Supporting Information
 Supporting Information Identification of Novel ROS Inducers: Quinone Derivatives Tethered to Long Hydrocarbon Chains Yeonsun Hong,, Sandip Sengupta,, Wooyoung Hur, *, Taebo Sim,, * KU-KIST Graduate School
Supporting Information Identification of Novel ROS Inducers: Quinone Derivatives Tethered to Long Hydrocarbon Chains Yeonsun Hong,, Sandip Sengupta,, Wooyoung Hur, *, Taebo Sim,, * KU-KIST Graduate School
Data Sheet. NFAT Reporter (Luc) Jurkat Cell line Catalog #: 60621
 Jurkat Cell line Catalog #: 60621") Data Sheet NFAT Reporter (Luc) Jurkat Cell line Catalog #: 60621 Background The nuclear factor of activator T cells (NFAT) family of transcription factors plays an important role in immune response. T
Data Sheet NFAT Reporter (Luc) Jurkat Cell line Catalog #: 60621 Background The nuclear factor of activator T cells (NFAT) family of transcription factors plays an important role in immune response. T
CAN Resubmission SUPPLEMENTAL INFORMATION
 SUPPLEMENTAL INFORMATION CAN-05-0686 Resubmission Protein Immunoblotting. Primary antibodies used in this study include: Casp-9, Casp-3 and Survivin from Novus Biologicals (Littleton, CO); Casp-8 (Ab-3)
SUPPLEMENTAL INFORMATION CAN-05-0686 Resubmission Protein Immunoblotting. Primary antibodies used in this study include: Casp-9, Casp-3 and Survivin from Novus Biologicals (Littleton, CO); Casp-8 (Ab-3)
MEK Kinase 1, a Substrate for DEVD-Directed Caspases, Is Involved in Genotoxin-Induced Apoptosis
 See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/51321268 MEK Kinase 1, a Substrate for DEVD-Directed Caspases, Is Involved in Genotoxin-Induced
See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/51321268 MEK Kinase 1, a Substrate for DEVD-Directed Caspases, Is Involved in Genotoxin-Induced
AMPK Phosphorylation Assay Kit
 AMPK Phosphorylation Assay Kit Catalog Number KA3789 100 assays Version: 02 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Intended Use... 3 Background... 3 Principle
AMPK Phosphorylation Assay Kit Catalog Number KA3789 100 assays Version: 02 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Intended Use... 3 Background... 3 Principle
NF-κB p65 (Phospho-Thr254)
") Assay Biotechnology Company www.assaybiotech.com Tel: 1-877-883-7988 Fax: 1-877-610-9758 NF-κB p65 (Phospho-Thr254) Colorimetric Cell-Based ELISA Kit Catalog #: OKAG02015 Please read the provided manual
Assay Biotechnology Company www.assaybiotech.com Tel: 1-877-883-7988 Fax: 1-877-610-9758 NF-κB p65 (Phospho-Thr254) Colorimetric Cell-Based ELISA Kit Catalog #: OKAG02015 Please read the provided manual
Impact of hyper-o-glcnacylation on apoptosis and NF-κB activity SUPPLEMENTARY METHODS
 SUPPLEMENTARY METHODS 3D culture and cell proliferation- MiaPaCa-2 cell culture in 3D was performed as described previously (1). Briefly, 8-well glass chamber slides were evenly coated with 50 µl/well
SUPPLEMENTARY METHODS 3D culture and cell proliferation- MiaPaCa-2 cell culture in 3D was performed as described previously (1). Briefly, 8-well glass chamber slides were evenly coated with 50 µl/well
Regulation of Fas Expression by STAT3 and c-jun Is Mediated by Phosphatidylinositol 3-Kinase-AKT Signaling*
 THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 277, No. 7, Issue of February 15, pp. 4932 4944, 2002 2002 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. Regulation of
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 277, No. 7, Issue of February 15, pp. 4932 4944, 2002 2002 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. Regulation of
Data Sheet. Notch Pathway Reporter Kit Catalog # 60509
 Data Sheet Notch Pathway Reporter Kit Catalog # 60509 6042 Cornerstone Court W, Ste B Background The Notch signaling pathway controls cell fate decisions in vertebrate and invertebrate tissues. NOTCH signaling
Data Sheet Notch Pathway Reporter Kit Catalog # 60509 6042 Cornerstone Court W, Ste B Background The Notch signaling pathway controls cell fate decisions in vertebrate and invertebrate tissues. NOTCH signaling
Supplementary Data Table of Contents:
 Supplementary Data Table of Contents: - Supplementary Methods - Supplementary Figures S1(A-B) - Supplementary Figures S2 (A-B) - Supplementary Figures S3 - Supplementary Figures S4(A-B) - Supplementary
Supplementary Data Table of Contents: - Supplementary Methods - Supplementary Figures S1(A-B) - Supplementary Figures S2 (A-B) - Supplementary Figures S3 - Supplementary Figures S4(A-B) - Supplementary
Supplementary Data 1. Alanine substitutions and position variants of APNCYGNIPL. Applied in
 Supplementary Data 1. Alanine substitutions and position variants of APNCYGNIPL. Applied in Supplementary Fig. 2 Substitution Sequence Position variant Sequence original APNCYGNIPL original APNCYGNIPL
Supplementary Data 1. Alanine substitutions and position variants of APNCYGNIPL. Applied in Supplementary Fig. 2 Substitution Sequence Position variant Sequence original APNCYGNIPL original APNCYGNIPL
Supplementary Material
 Supplementary Material Nuclear import of purified HIV-1 Integrase. Integrase remains associated to the RTC throughout the infection process until provirus integration occurs and is therefore one likely
Supplementary Material Nuclear import of purified HIV-1 Integrase. Integrase remains associated to the RTC throughout the infection process until provirus integration occurs and is therefore one likely
Supporting Information. FADD regulates NF-кB activation and promotes ubiquitination of cflip L to induce. apoptosis
 1 2 Supporting Information 3 4 5 FADD regulates NF-кB activation and promotes ubiquitination of cflip L to induce apoptosis 6 7 Kishu Ranjan and Chandramani Pathak* 8 9 Department of Cell Biology, School
1 2 Supporting Information 3 4 5 FADD regulates NF-кB activation and promotes ubiquitination of cflip L to induce apoptosis 6 7 Kishu Ranjan and Chandramani Pathak* 8 9 Department of Cell Biology, School
TSH Receptor Monoclonal Antibody (49) Catalog Number MA3-218 Product data sheet
 Catalog Number MA3-218 Product data sheet") Website: thermofisher.com Customer Service (US): 1 800 955 6288 ext. 1 Technical Support (US): 1 800 955 6288 ext. 441 TSH Receptor Monoclonal Antibody (49) Catalog Number MA3-218 Product data sheet Details
Website: thermofisher.com Customer Service (US): 1 800 955 6288 ext. 1 Technical Support (US): 1 800 955 6288 ext. 441 TSH Receptor Monoclonal Antibody (49) Catalog Number MA3-218 Product data sheet Details
Notch Signaling Pathway Notch CSL Reporter HEK293 Cell line Catalog #: 60652
 Notch Signaling Pathway Notch CSL Reporter HEK293 Cell line Catalog #: 60652 Background The Notch signaling pathway controls cell fate decisions in vertebrate and invertebrate tissues. Notch signaling
Notch Signaling Pathway Notch CSL Reporter HEK293 Cell line Catalog #: 60652 Background The Notch signaling pathway controls cell fate decisions in vertebrate and invertebrate tissues. Notch signaling
MAPK Pathway
 MAPK Pathway Mitogen-activated protein kinases (MAPK) are proteins that are serine/ threonine specific kinases which are activated by a wide range of stimuli including proinflammatory cytokines, growth
MAPK Pathway Mitogen-activated protein kinases (MAPK) are proteins that are serine/ threonine specific kinases which are activated by a wide range of stimuli including proinflammatory cytokines, growth
Supplementary Figure 1: si-craf but not si-braf sensitizes tumor cells to radiation.
 Supplementary Figure 1: si-craf but not si-braf sensitizes tumor cells to radiation. (a) Embryonic fibroblasts isolated from wildtype (WT), BRAF -/-, or CRAF -/- mice were irradiated (6 Gy) and DNA damage
Supplementary Figure 1: si-craf but not si-braf sensitizes tumor cells to radiation. (a) Embryonic fibroblasts isolated from wildtype (WT), BRAF -/-, or CRAF -/- mice were irradiated (6 Gy) and DNA damage
Supplementary Figure 1
 Supplementary Figure 1 Constitutive EGFR signaling does not activate canonical EGFR signals (a) U251EGFRInd cells with or without tetracycline exposure (24h, 1µg/ml) were treated with EGF for 15 minutes
Supplementary Figure 1 Constitutive EGFR signaling does not activate canonical EGFR signals (a) U251EGFRInd cells with or without tetracycline exposure (24h, 1µg/ml) were treated with EGF for 15 minutes
Supplementary Figure S1. Venn diagram analysis of mrna microarray data and mirna target analysis. (a) Western blot analysis of T lymphoblasts (CLS)
 Western blot analysis of T lymphoblasts (CLS)") Supplementary Figure S1. Venn diagram analysis of mrna microarray data and mirna target analysis. (a) Western blot analysis of T lymphoblasts (CLS) and their exosomes (EXO) in resting (REST) and activated
Supplementary Figure S1. Venn diagram analysis of mrna microarray data and mirna target analysis. (a) Western blot analysis of T lymphoblasts (CLS) and their exosomes (EXO) in resting (REST) and activated
2,6,9-Triazabicyclo[3.3.1]nonanes as overlooked. amino-modification products by acrolein
![2,6,9-Triazabicyclo[3.3.1]nonanes as overlooked. amino-modification products by acrolein](/thumbs/86/94743397.jpg "2,6,9-Triazabicyclo[3.3.1]nonanes as overlooked. amino-modification products by acrolein") Supplementary Information 2,6,9-Triazabicyclo[3.3.1]nonanes as overlooked amino-modification products by acrolein Ayumi Tsutsui and Katsunori Tanaka* Biofunctional Synthetic Chemistry Laboratory, RIKEN
Supplementary Information 2,6,9-Triazabicyclo[3.3.1]nonanes as overlooked amino-modification products by acrolein Ayumi Tsutsui and Katsunori Tanaka* Biofunctional Synthetic Chemistry Laboratory, RIKEN
Apoptosis Mediated Cytotoxicity of Curcumin Analogues PGV-0 and PGV-1 in WiDr Cell Line
 Apoptosis Mediated Cytotoxicity of Curcumin Analogues PGV-0 and PGV-1 in WiDr Cell Line Endah Puji Septisetyani, Muthi Ikawati, Barinta Widaryanti and Edy Meiyanto* ) Cancer Chemoprevention Research Center,
Apoptosis Mediated Cytotoxicity of Curcumin Analogues PGV-0 and PGV-1 in WiDr Cell Line Endah Puji Septisetyani, Muthi Ikawati, Barinta Widaryanti and Edy Meiyanto* ) Cancer Chemoprevention Research Center,
Apoptosis is a cell-death program that plays an important
 Phosphatidylinositol 3-Kinase (PI-3K)/Akt but Not PI-3K/p70 S6 Kinase Signaling Mediates IGF-1-Promoted Lens Epithelial Cell Survival Gudiseva Chandrasekher and Dasetty Sailaja From the Department of Ophthalmology
Phosphatidylinositol 3-Kinase (PI-3K)/Akt but Not PI-3K/p70 S6 Kinase Signaling Mediates IGF-1-Promoted Lens Epithelial Cell Survival Gudiseva Chandrasekher and Dasetty Sailaja From the Department of Ophthalmology
For the 5 GATC-overhang two-oligo adaptors set up the following reactions in 96-well plate format:
 Supplementary Protocol 1. Adaptor preparation: For the 5 GATC-overhang two-oligo adaptors set up the following reactions in 96-well plate format: Per reaction X96 10X NEBuffer 2 10 µl 10 µl x 96 5 -GATC
Supplementary Protocol 1. Adaptor preparation: For the 5 GATC-overhang two-oligo adaptors set up the following reactions in 96-well plate format: Per reaction X96 10X NEBuffer 2 10 µl 10 µl x 96 5 -GATC