Human platelet dense granules contain polyphosphate and are similar to acidocalcisomes of bacteria and unicellular eukaryotes
|
|
|
- Tracey Nelson
- 6 years ago
- Views:
Transcription
1 JBC Papers in Press. Published on August 11, 2004 as Manuscript M Running title: Polyphosphate in platelet dense granules Human platelet dense granules contain polyphosphate and are similar to acidocalcisomes of bacteria and unicellular eukaryotes Felix A. Christopher R. Lea^, Eric Oldfield^, and Roberto From Laboratory of Molecular Parasitology, Department of Pathobiology and Center for Zoonoses Research, University of Illinois at Urbana-Champaign, 2001 S. Lincoln Avenue, Urbana, IL and the ^Department of Chemistry, University of Illinois at Urbana-Champaign, Urbana, IL Present address: Unidad de Investigacion, Hospital Universitario, Puerta del Mar, Avenida Ana de Viya 21, Cadiz, Spain. *This work was supported in part by the U.S. National Institutes of Health (grants AI23259 to R.D. and GM50694 to E.O.) To whom correspondence should be addressed at: Laboratory of Molecular Parasitology, Department of Pathobiology and Center for Zoonoses Research, College of Veterinary Medicine, University of Illinois at Urbana-Champaign, 2001 S. Lincoln Avenue, Urbana, Illinois 61802, Tel: ; Fax: ; rodoc@uiuc.edu 1 Copyright 2004 by The American Society for Biochemistry and Molecular Biology, Inc.
2 Inorganic polyphosphate (polyp) has been identified and measured in human platelets. Millimolar levels (in terms of P i residues) of short chain polyp were found. The presence of polyp of about phosphate units was identified by 31 P NMR and by ureapolyacrylamide gel electrophoresis of platelet extracts. Analysis of human platelet dense granules, purified using metrizamide gradient centrifugation, indicated that polyp was preferentially located in these organelles. This was confirmed by visualization of polyp in the dense granules using 4,6-diamidino-2-phenylindole and by its release together with pyrophosphate (PP i ), and serotonin upon thrombin stimulation of intact platelets. Dense granules were also shown to contain large amounts of calcium, and potassium, and both bafilomycin A 1 -sensitive ATPase, and pyrophosphatase activities. In agreement with these results, when human platelets were loaded with the fluorescent calcium indicator fura-2 acetoxymethyl ester to measure their intracellular Ca 2+ concentration ([Ca 2+ ] i ) they were shown to possess a significant amount of Ca 2+ stored in an acidic compartment. This was indicated by: (1) the increase in [Ca 2+ ] i induced by nigericin, monensin, or the weak base, NH 4 Cl, in the nominal absence of extracellular Ca 2, and (2) the effect of ionomycin, which cannot take Ca 2+ out of acidic organelles and was more effective after alkalinization of this compartment by the previous addition of nigericin, monensin, or NH 4 Cl. All these characteristics of the platelet dense granules, together with their known acidity and high density (both by weight and by electron microscopy), are similar to those of acidocalcisomes (volutin granules, polyp bodies) of bacteria and unicellular eukaryotes. The results suggest that acidocalcisomes have been conserved during evolution from bacteria to humans. 2
3 Inorganic polyphosphate (polyp) is a ubiquitous polymer formed by phosphate (P i ) residues linked by high-energy phosphoanhydride bonds. Although the presence of polyp in bacteria, fungi, algae, and protozoa has been widely noted, the distribution and abundance of polyp in more complex eukaryotic forms has remained uncertain for many years (1). The very low levels of polyp in animal cells and subcellular compartments and the lack of sensitive methods have left its metabolic and functional roles entirely obscure (1). The recent introduction of novel quantitative enzymatic analytical methods (2) has permitted the detection of levels of 20 to 120 µm (in terms of P i residues), in chains 50 to 800 residues long in rodent tissues (brain, heart, kidneys, liver, and lungs) and subcellular fractions (nuclei, mitochondria, plasma membrane, and microsomes) (3) but no studies have been reported with human tissues. In bacteria (4) as well as in several unicellular eukaryotes, like trypanosomatid and apicomplexan parasites (5), the green algae Chlamydomonas reinhardtii (6), and the slime mold Dictyostelium discoideum (7), polyp is accumulated in acidic granules known as acidocalcisomes where it can reach millimolar or molar levels. Acidocalcisomes have been shown to possess calcium- and proton-accumulating activities in their limiting membranes, have very high density (both in weight and by electron microscopy), and are also rich in pyrophosphate (PP i ), calcium, and other cations (5). Human platelets possess an organelle, the dense granule, with morphological and biochemical similarities to acidocalcisomes. Platelet dense granules store extremely high concentrations of calcium, together with ATP, ADP, serotonin, and PP i (8). Holmsen and Weiss (9) calculated the intragranular calcium concentration to be 2.2 M. The internal matrix ph of the dense granules is about 5.4 (10), and this acidic ph is maintained by the function of an H + -ATPase, which is different from F 1 F 0 - and P-type ATPases (11). Since acridine orange uptake in dense granules was inhibited by bafilomycin A 1 (12), which is a 3
4 specific inhibitor of vacuolar H + -ATPases (13), it was suggested that this proton pump is a V-H + -ATPase. In addition, calcium release from intracellular acidic stores by the K + /H + ionophore nigericin has been observed in a number of unicellular eukaryotes that posses acidocacisomes (5) as well as from platelets (14, 15). Because acidocalcisomes in bacteria and unicellular eukaryotes are morphologically very similar to platelet dense granules, we investigated whether these granules have similar characteristics to acidocalcisomes, also known as volutin granules or polyp bodies (5). In this report, we describe the isolation and biochemical characterization of dense granules from human platelets and demonstrate that, as the acidocalcisomes, they are rich in polyp, which is secreted upon thrombin stimulation. In addition, we report experiments using intact fura 2-loaded platelets that demonstrate that a considerable portion of the releasable Ca 2+ is located in these organelles. Our results suggest that acidocalcisome-like organelles are the only ones described to date that are present from bacteria to humans. EXERIMENTAL PROCEDURES Chemicals-Leupeptin, trans-epoxysuccinyl-l-leucylamido-(4-guanidino) butane (E64), N-p-tosyl-L-lysine chloromethyl ketone, ATP, ionophores, metrizamide, reagents for marker enzyme assays, apyrase, anti-human von Willebrand factor IgG fraction antibodies, apyrase and polyps were purchased from Sigma Chemicals Co. (St. Louis, MO). 4-(2- aminoethyl)benzenesulfonyl fluoride and ionomycin (free acid) were from Calbiochem (La Jolla, CA). Pepstatin came from Roche Molecular Biochemicals (Indianapolis, IN). Coomassie Blue protein assay reagent was from Bio-Rad (Hercules, CA). Escherichia coli strain CA38 ptrcppx1 was kindly provided by Prof. Arthur Kornberg, Stanford University School of Medicine (Stanford, CA). Fresh Human platelets were obtained from the 4
5 Community Blood Services of Illinois, (Urbana, IL). Aminomethylenediphosphonate was synthesized by Michael Martin, Department of Chemistry, University of Illinois at Urbana- Champaign. All other reagents were analytical grade. Platelet Dense Granule Isolation-Dense granules were isolated using metrizamide gradient centrifugation by a method previously described (16) with minor modifications. A unit of platelet rich plasma was centrifuged twice at 200 x g for 10 min, to eliminate contaminant red blood cells and leukocytes. Then, the supernatant was centrifuged at 1,000 x g for 15 min, and the resulting pellet was resuspended at 2 x 10 9 platelet/ml in lysis buffer (25 mm Hepes, 0.25 M sucrose, 12 mm sodium citrate, 1 mm EDTA, 0.1 mm 4-(2- aminoethyl) benzenesulfonyl fluoride, 10 mm pepstatin, 10 mm leupeptin, 10 mm transepoxysuccinyl-l-leucylamido-(4-guanidino) butane and 10 mm N-a -tosyl-l-lysine chloromethyl ketone, ph. 6.5). Platelets were sonicated twice (5 sec at 20% intensity in a Branson sonifier, model 102c) and unbroken cells were separated by centrifugation (1,000 x g for 15 min). Sonication and centrifugation were repeated twice with the pellets resuspended in a similar volume of lysis buffer. The three supernatants were combined and centrifuged for 20 min at 19,000 x g at 4 o C. The pellet was resuspended with lysis buffer up to 1.5 ml and mixed with 1.5 ml of metrizamide solution (40% w/v in nanopure water). This mixture was applied at the top of a discontinuous gradient of metrizamide, with 3-ml steps of 35, 38, and 40% metrizamide (prepared by dilution of the 40% solution in lysis buffer). The gradient was centrifuged at 100,000 x g using a Beckman SW 41 rotor for 60 min. The dense granule rich fraction pelleted at the bottom of the tube and was resuspended in lysis buffer. To determine serotonin incorporation into gradient fractions, platelet rich plasma lacking red blood cells and leukocytes, were incubated for 15 min at 37 o C with 1 µci/ml of 5
6 1 mm 5-[2-14 C]-hydroxytryptamine binoxalate (PerkinElmer Life Sciences) and the fractionation procedure was performed as described above. Von Willebrand factor levels in the gradient fractions were measured by ELISA using Anti-human von Willebrand factor IgG fraction antibodies. Gradient fractions were assayed also for succinate-cytochrome C reductase (mitochondrial marker) and acid phosphatase (lysosome marker) (17), short and long chain chain polyp and PP i (18), bafilomycin A 1 (0.5 µm)-sensitive ATPase (V-H + - ATPase), and pyrophosphatase (7). The construction of normalized density distribution histograms was carried out as described before (19). Electrophoretic analysis of polyp-urea-polyacrylamide gels were prepared and stained with toluidine blue as previously described (18). Marker polyps were obtained by electrophoresis of phosphate glass in 1.5% agarose gels. Four mm-long gel slices were eluted by centrifugation trough Millipore Ultrafree-MC columns to obtain polyps of different sizes. The markers were localized by toluidine blue staining and their size was calculated by calibration with commercial polyps. Electron Microscopy and X-ray Microanalysis For imaging whole cells, platelets were washed first in Tris citrate buffer (63 mm Tris, 95 mm NaCl, 5 mm KCl, 12 mm citric acid, ph 6.5) and then with 0.25 M sucrose. Dense granule fractions were washed with 0.25 M sucrose. Samples were placed on a Formvar-coated 200-mesh copper grid, allowed to adhere for 10 min at room temperature, blotted dry, and observed directly with a Hitachi 600 transmission electron microscope operating at 100 kv (20). Energy-dispersive x-ray analysis was done at the Electron Microscopy Center, Southern Illinois University (Carbondale, IL). Specimen grids were examined in a Hitachi H-7100FA transmission electron microscope at an accelerating voltage of 50 kv. Fine probe sizes were adjusted to cover the dense granules (or a similar area of the background), and x-rays were collected for 100 s by utilizing a thin window (Norvar) detector. Analysis was performed by using a 6
7 Noran Voyager III analyzer with a standardless analysis identification program. Conventional electron microscopy of the dense granule fraction was done as described before (20). Fluorescence Microscopy-Platelets (~10 8 ) were washed with New Tyrode's buffer (137 mm NaCl, 2.7 mm KCl, 12 mm NaHCO 3, 1 mm MgCl 2, 5.5 mm glucose, 0.35% BSA, ph 7.35). The pellet was resuspended in 2 ml of the same buffer, and 45 µl of this suspension was incubated at room temperature with 10 µg of 4',6-diamidino-2-phenylindole (DAPI)/ml. After 10 min, the samples were mounted on a slide and observed with an Olympus model BX-60 epifluorescence microscope. An Olympus WU filter (excitation: nm; emission: > 420) was used. The images were recorded with a charge-coupled device camera (model CH250; Photometrics Ltd., Tucson, AZ) and IPLab software (Signal Analytics, Vienna, VA) as described previously (21). Intracellular Calcium Measurements-Platelet rich plasma was centrifuged at 700 x g for 5 min and 100 mm aspirin, and 40ug/ml apyrase were added to the supernatant. Platelets were incubated at 37 o C with 1 mm fura-2/am for 1h. Labeled platelets were centrifuged at 1,000 x g 10 min and washed in New Tyrode's buffer. Then, cells were resuspended in the same buffer at 2 x 10 8 platelets/ml and kept at 37 o C. For fluorescence measurements, 1.25 x 10 8 cells/ml (final density) were suspended in a cuvette containing New Tyrode's buffer and 1 mm EGTA. Fluorescence emission at 510 nm was measured with 340 and 380nm excitation in a thermostated Hitachi F-2000 spectrofluorometer at 37 o C with agitation. Calibration was performed as described before (22). Perchloric acid extracts-for NMR, platelets were washed twice with New Tyrode s buffer, then extracted with ice-cold 0.5 M HClO4 (2 ml/g wet weight cells). After 30 min incubation on ice, the extracts were centrifuged at 3,000 x g for 5 min. The supernatants 7
8 were neutralized by the addition of 0.72 M KOH/0.6 M KHCO3 (18). Precipitated KClO4 was removed by centrifugation at 12,000 x g for 10 min., the supernatant separated and EDTA added to a final concentration of 400 µm prior to adjusting to ph 7. All extracts contained 10% D 2 O (v/v) to provide a field-frequency lock. NMR spectroscopy-phosphorus NMR spectra were acquired at MHz using a Varian INOVA NMR spectrometer equipped with a 17.6 Tesla Oxford Instruments magnet. For perchloric acid extracts, 8,192 transients were collected at room temperature using 25 µs (90 ) pulse excitation, a 20 khz spectral width, 32,768 data points, and a 5 sec recycle time. Inverse gate proton decoupling was used to remove NOE and J-coupling effects. The chemical shifts of all 31 P spectra were referenced to 0 ppm using an 85% phosphoric acid external standard (23). The specific assignments of individual resonances were based on published chemical shifts and 31 P- 31 P scalar couplings (24). NMR spectra were processed using the VNMR 6.1B software package (Varian Inc., Palo Alto, CA) running on a Sun Ultra 5 (Sun Microsystems, Santa Clara, CA) workstation, and included baseline correction, zero-filling, and a 3 Hz exponential line broadening prior to Fourier transformation. Analysis of Platelet Released Material-Platelet rich plasma lacking red blood cells and leukocytes, were incubated for 15 min at 37 o C with 1 µci/ml of 1 mm 5-[2-14 C]- hydroxytryptamine binoxalate. Platelets were washed and resuspended in New Tyrode's buffer at 10 8 cell/ml. The platelet suspension was placed in a cuvette with agitation. Thrombin was added at a final concentration of 1.2 U/ml and aliquots were taken at different times. Samples were directly filtered trough a 0.2 µm filter to separate the secreted material from the suspension. Content of PP i, short chain polyp, and serotonin in the samples were analyzed as described above. 8
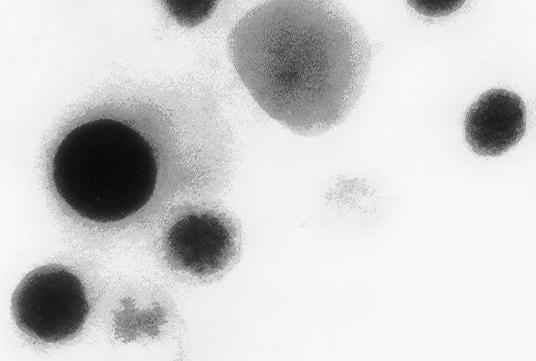
9 H + Transport Assays-H + uptake into platelet dense granules was assayed using acridine orange as described before (6) using a standard buffer containing 120 mm KCl, 2 mm MgCl 2, 50 mm K-Hepes, 50 µm EGTA, ph 7.2. RESULTS Elemental Analysis and Isolation of Human Platelet Dense Granules-Human platelet dense granules are known to be acidic (10) and to accumulate calcium (9). They are characterized, as the acidocalcisomes (5), by their high electron-density. They were first described as electron-opaque organelles in air-dried whole mounts by Bull (25), White (26) and Costa et al. (27). Several dense granules with high electron density of varying diameter (average 161 ± 51 nm) were seen when whole human platelets were observed by transmission electron microscopy without fixation and staining (Fig. 1A). 5 to 20 granules of various shapes and sizes were observed per platelet (n = 20). This is in agreement with previous reports of the presence a variable number of dense bodies per platelet ranging from none to more than 20 (28). X-ray microanalysis was performed on these granules (Fig. 1B). All ten spectra taken from different platelets were qualitatively similar, the P/Ca ratio being 1.76 as it has been reported before (29). Potassium and oxygen were also detected. Carbon, silicon and copper arose from the formvar-coated grids, and were also detected in spectra taken from the background. To purify the dense granules and investigate their chemical and enzymatic content, we adapted the purification procedure used by Rendu et al. (16). Examination of the densest fraction 12 (see below) by transmission electron microscopy without fixation and staining showed round electron-dense granules of variable size, up to 200 nm diameter (Fig. 2A). When submitted to the electron beam, changes in their internal structure led to 9
10 the appearance of a sponge-like structure (Fig. 2A, inset), which has been described before in acidocalcisomes of T. cruzi (30), T. brucei (31), C. reinhardtii (6) and Agrobacterium tumefaciens (4). Examination of the fraction using conventional transmission electron microscopy with fixation, dehydration and staining procedures showed membrane-bound organelles of similar size that appeared empty (Fig. 2B). Marker enzymes were used to characterize the different fractions (Fig. 3). Serotonin incorporation (26) was used as a marker for dense granules. Its yield in the dense granule fraction (fraction 12) was 25%, whereas the yield of protein in the same fraction was only 6%, a 4.2-fold purification. Although the incorporation of serotonin was greater in fraction 3, which consisted of whole platelets and large fragments, the purest dense granule fraction was the densest fraction (fraction 12). Mitochondria (marked by succinate cytochrome c reductase), lysosomes (marked by acid phosphatase), and alpha granules (marked by the presence of von Willebrand factor) were not enriched in the densest fraction. The dense granule fraction contained about 20% of the total PP i and short chain polyphosphate (polyp; see below) (Fig. 3). PolyP in human platelets-platelet extracts yielded levels of polyp of 0.74 ± 0.08 nmol per 10 8 platelets (n = 8) and no detectable long chain polyp (data not shown). If we assume that the platelet volume is 6.8 mm 3 (9), the concentration of polyp in platelets is around 1.1 mm. Thus, platelet polyp concentration is 10 to 20 times higher than that measured in rodent tissues such as brain, heart, kidney, lung, and liver (3). Interestingly, in contrast to other cell types studied, all the polyp detected was less than 100 residues long. PolyP levels were in the same order of magnitude as the PP i (0.92 ± 0.19 nmol per 10 8 platelets; n = 8) measured by its hydrolysis with yeast pyrophosphatase. Excess of pyrophosphatase in our assay degraded less than 2% of the polyp, demonstrating the reliability of the obtained values for the different populations of phosphate polymers. We can estimate the 10
11 concentration of polyp in the dense granule from the total amount of polyp in the whole platelet (see above). If we assume that the polyp in the dense granule is about 60% of the total (as occurs with calcium, ADP, ATP and PP i ), the volume of each granule is 0.1% of the platelet volume (9), and there are an average of 5 dense granules per platelet (8, 28), the intra-granular concentration of polyp is about 130 mm. Further evidence for the localization of polyp in the dense granules-we also investigated the location of polyp using DAPI (Fig. 4). DAPI has been shown to shift its emission fluorescence to a maximum wavelength of 525 nm in the presence of polyp, this change being specific for polyp and not produced by PP i or other anions (18, 32). Human platelets incubated in solutions of DAPI (0.2 mg/ml) were mounted on slides and examined by confocal fluorescence microscopy. We detected staining of numerous intracellular vacuoles. No staining was detected when DAPI was omitted (data not shown). Identification of polyp by 31 P NMR and urea-page analysis-fig. 5 shows the MHz ( 1 H decoupled) 31 P NMR spectra of perchloric acid extracts of human platelets. Resonance assignments for these spectra are given in Table 1. The predominant peak in the spectrum is inorganic phosphate (A). The region between 6 and 8 ppm is shown in inset magnified 2.5X respective to the overall spectra and contains peaks for the terminal phosphates of adenosine di- (B) and triphosphate (C) in addition to peaks for the terminal phosphates of polyps (D) and a large peak for PP i (E). The region from 10 to 12 ppm contains peaks for the a-phosphates of adenosine di- (F) and triphosphate (G). The region from 20 to 23 ppm is again shown in inset magnified 2.5X respective to the overall spectra and contains peaks for the central phosphate of adenosine triphosphate (H) and the g-phosphates (central) of polyps (I). 11
12 PolyPs extracted from human platelets were electrophoresed by 6% urea-page to determine their size distribution (Fig. 6). Only one size class of polyp was detected in the three samples analyzed: short-chain polyp of about residues. PolyP release after stimulation of platelets with thrombin-since 80% of the total PP i is secreted by platelets treated with thrombin with a time course similar to secretion of ATP, ADP and serotonin from the platelet dense granules (33) we investigated whether thrombin had a similar effect on the release of polyp. Fig. 7 shows that this was the case, confirming that polyp is in the same compartment containing PP i and serotonin. Intracellular Ca 2+ Concentration [Ca 2+ ] i in Human Platelets: Effect of Thapsigargin and Nigericin-The concentration of cytosolic calcium in human platelets was 22 ± 2.8 nm (n = 6) in the absence of extracellular Ca 2+ (1 mm EGTA added). This concentration is in the range observed in previous studies (34). Addition of 1 µm thapsigargin, a specific inhibitor of the endoplasmic reticulum Ca 2+ -ATPase when used at low concentrations (35), increased the [Ca 2+ ] i (Fig. 8A, upper trace). Subsequent addition of nigericin (a K + /H + ionophore; 2 µm) caused a second rise in [Ca 2+ ] i to a level that was significantly higher to that in the absence of nigericin. Similar results were observed when the order of additions was reversed (Fig. 8A, lower trace), suggesting the presence of a thapsigargin-sensitive, and an acidic thapsigargin-insensitive Ca 2+ compartment. Synergistic Effect of Ionomycin and Ammonium Chloride on Ca 2+ Release from Intracellular Compartments-To provide more evidence for the presence of an acidic Ca 2+ store in human platelets, we performed experiments using sequential additions of ionomycin and ammonium chloride. It has been demonstrated (36) that ionomycin can only mobilize Ca 2+ from neutral or alkaline compartments, but releases more Ca 2+ after alkalinizing agents have elevated the ph of acidic compartments. Accordingly, adding ionomycin to human platelets previously exposed to NH 4 Cl (10 mm) caused an increase in [Ca 2+ ] i to a level 12
13 which was significantly higher to that in the absence of NH 4 Cl (Fig. 8B, upper trace). Similar results were obtained when the order of additions was reversed (Fig. 8B, upper trace) and when nigericin (2 µm) or monensin (1 µm) were used as alkalinizing agents (data not shown). Taken together these results indicate the presence of an acidic compartment, sensitive to a K + /H + ionophore (nigericin), and a Na + /H + ionophore (monensin), and insensitive to thapsigargin, that contains a significant amount of Ca 2+ in human platelets and which is physiologically similar to that described as the acidocalcisome in trypanosomatids and apicomplexan parasites (4). V-H + -ATPase and Pyrophosphatase Activities in the Isolated Dense Granules- Bafilomycin A 1 -sensitive ATPase activity (measured by P i release from ATP) was enriched in the dense granule fraction (Fig. 9A). Although isolated dense granules could take up acridine orange, demonstrating that they were still intact and acidic, neither ATP nor PP i were able to stimulate this process. Alkalinization of dense granules by addition of NH 4 Cl resulted in release of the acridine orange accumulated (Fig. 9B). A pyrophosphatase activity was also enriched in the dense granule fraction (Fig. 9A). Interestingly, when this activity was measured in the presence of aminomethylenediphosphonate (AMDP), an inhibitor of vacuolar proton translocating pyrophosphatases (V-H + -PPases) (37), its yield in the dense granule fraction was higher than in any other fraction (30%). DISCUSSION We report here that human platelets are rich in polyp. PolyP was detected in platelet extracts by biochemical methods and 31 P NMR and shown to be present at millimolar 13
14 levels, 10 to 20 times higher than in rodent tissues previously investigated (3). The presence of polyp of about phosphate units was identified by urea-polyacrylamide gel electrophoresis of platelet extracts. Most polyp was found in the dense granules, purified using metrizamide gradient centrifugation. This was confirmed by visualization of polyp in the dense granules using 4,6-diamidino-2-phenylindole and by its release together with pyrophosphate (PP i ), and serotonin upon thrombin stimulation of intact platelets. Dense granules were also shown to contain large amounts of calcium, and potassium, and both bafilomycin A 1 -sensitive ATPase, and pyrophosphatase activities. In agreement with these results, when human platelets were loaded with the fluorescent calcium indicator fura-2 acetoxymethyl ester to measure their intracellular Ca 2+ concentration ([Ca 2+ ] i ) they were shown to possess a significant amount of Ca 2+ stored in an acidic compartment. Numerous studies have defined the role of dense granules in platelets. Activation of platelets leads to the secretion of dense granule components such as ATP, ADP, PP i, calcium, and serotonin. ATP could act on P2Y receptors on endothelial cells to release prostacyclin and nitric oxide, which in turn cause vasodilation (8). ADP acts as a platelet agonist, and is important for the activation of additional platelets and their recruitment to the site of injury (8). Serotonin acts to activate additional platelets and thus recruit them into the aggregate and also has a vasoconstrictive action, which reduces flow at the site of injury and thereby limits blood loss (8). Calcium could be important for the binding of adhesive proteins to their platelet receptors (8). The functions of PP i and polyp are unknown. However, polyp, as heparin, is a negatively charged polymer that has been postulated to stabilize fibroblast growth factor (FGF) and facilitate FGF-2 binding to its receptor tyrosine kinase promoting signaling via the formation of receptor dimers, and to prevent FGF degradation in tissues that have been injured (38). Since polyp is released 14
15 from platelets after thrombin stimulation (Fig. 7), it could contribute to the homeostatic functions of platelets and further work will be needed to investigate its role. Calcium is important for many physiological processes leading to platelet activation, e.g. shape change, eicosanoid formation, adhesion, aggregation and granule secretion. Platelet agonist such as ADP and thrombin increase cytosolic Ca 2+ by initially releasing it from one or more internal storage sites. Two different Ca 2+ stores had been reported in human platelets (14, 39). Thapsigargin, a sarcoplasmic-endoplasmic reticulum (SERCA) Ca 2+ -ATPase inhibitor (35), was shown to release Ca 2+ from an inositol 1,4,5-trisphosphate (InsP 3 )-sensitive intracellular store (14) (possibly the dense tubular system, a structure derived from the megakaryocyte endoplasmic reticulum; ref. 40). Addition of thrombin to thapsigargin-treated platelets resulted in further Ca 2+ release, and this Ca 2+ release was diminished by previous treatment with nigericin (14), suggesting that Ca 2+ was being released from an acidic compartment. The addition of a combination of ionomycin with alkalinizing agents, such as nigericin, monensin, or NH 4 Cl, which has been previously used to identify acidocalcisomes in trypanosomatids and apicomplexan parasites (5), confirmed the presence of an acidic Ca 2+ pool in human platelets (Fig. 8). In this regard, it has been shown that platelets express both SERCA 2b and SERCA 3 Ca 2+ -ATPase isoforms suggesting that the two isoforms may be located in functionally distinct Ca 2+ storage pools of the cells (41, 42). Platelet dense granules have, therefore, many characteristics in common to acidocalcisomes: [1] they are acidic due to the operation of a bafilomycin A 1 -sensitive proton ATPase (Fig. 9A; and ref. 12), and are able to accumulate acidophilic dyes such as acridine orange (Fig. 9B, and refs. 12, and 43) or mepacrine (quinacrine) (44); [2] they can store extremely high concentrations of calcium, and other cations, such as potassium (Fig. 1), or lithium (45), and can release Ca 2+ in the presence of ionophores such as nigericin or 15
16 monensin (Fig. 8); [3] they contain very high concentrations of phosphorus in the form of PP i (33) and polyp (Figs. 3-5); [4] they have very high density both by electron microscopy (Fig. 1), and by weight (Fig. 3); [5] they have a diameter of about 200 nm, and are predominantly spherical, although they can also show elongated or irregular shapes. Acidocalcisomes described in bacteria and unicellular eukaryotes have some peculiarities not found in dense granules such as the presence of a vacuolar-type H + - pyrophosphatase in their surrounding membrane (5). As occurs with acidocalcisomes of Trypanosoma brucei (46) a pyrophosphatase activity was enriched in the dense granule fraction of human platelets, and this activity was inhibited by AMDP, an inhibitor of V-H + - PPases (37) (Fig. 9A). Although one study reported PP i -induced acidification of trans cisternal elements of rat liver Golgi membranes (47), there has been no reports of the presence of genes with similarity to those encoding V-H + -PPases of bacteria, plants, and protists. Since no PP i -dependent H + - transport could be measured in dense granule fractions (Fig. 9B), and AMDP has also been shown to inhibit soluble pyrophosphatases at micromolar levels (37), we can rule out that the activity detected is due to a V-H + -PPase. Although a bafilomycin A 1 -sensitive ATPase activity could be measured by P i release from ATP (Fig. 9A), H + -transport in isolated dense granules could not be detected (Fig. 9B). This has been observed with other acidocalcisomes that are known to posses the V-H + -ATPase and it has been proposed that the V-H + -ATPase complex dissociates, losing its peripheral subunits, or otherwise becomes inactive in H + transport during purification (48), as has been observed in other cases (49, 50). On the other hand, platelet dense granules accumulate high concentrations of ATP, ADP, and serotonin, which have not been reported to be present in acidocalcisomes. However, acidocalcisomes of trypanosomatids have been found to contain the biogenic amine g-aminobutyric acid (GABA; Rohloff and Docampo, unpublished results), which has 16
17 been shown to accumulate in platelets (51), and porcine dense granules possess histamine (8, 9). Having a homolog enzyme (H + -ATPase) and similar chemical composition (PP i, polyp, Ca, K, etc) in a similar organelle is a strong argument of the common origin of the two organelles. Acidocalcisomes have beeen shown previously (5) to be similar to volutin granules or metachromatic granules found in bacteria and in a number of unicellular eukaryotes (5). Volutin granules were discovered 100 years ago (52), and as dense granules, were easily identified in different microorganisms because of their density in whole mount electron microscopy (53). They were originally thought to be devoid of a limiting membrane. In recent years they have been found to be surrounded by a membrane containing a number of pumps and exchangers and were named acidocalcisomes (5). In conclusion, our results suggest that human platelet dense granules belong to the same class of organelles as volutin granules or acidocalcisomes and are therefore the only known organelle group that has been conserved during evolution from bacteria to humans. Acknowledgments-We thank Arthur Kornberg for E. coli CA38 ptrcppx1, Michael Martin for the synthesis of AMDP, and John Bozzola and Steve Schmitt for help with the X- ray microanalysis. REFERENCES 1. Kornberg, A. (1995) J. Bacteriol. 177, Kornberg, A., Rao, N. N., and Ault-Riche, D. (1999) Annu Rev. Biochem. 68, Kumble, K. D., and Kornberg, A. (1995) J. Biol. Chem. 270,
18 4. Seufferheld, M., Vieira, M.C.F., Ruiz, F.A., Rodrigues, C.O., Moreno, S.N.J., and Docampo, R. (2003) J. Biol. Chem. 278, Docampo, R., and Moreno, S.N.J. (2001) Mol. Biochem. Parasitol. 33, Ruiz, F.A., Marchesini, N., Seufferheld, M., Govindjee, and Docampo, R. (2001) J. Biol. Chem. 276, Marchesini, N., Ruiz, F.A., Vieira, M., and Docampo, R. (2002) J. Biol. Chem. 277, McNicol, A., and Israels, S. J. (1999) Thrombosis Res. 95, Holmsen, H., and Weiss, H. J. (1979) Ann. Rev. Med. 30, Carty, S.E., Johnson, R.G., and Scarpa, A. (1981) J. Biol. Chem. 256, Dean, G. E., Fishkes, H., Nelson, P. J., and Rudnick, G. (1984) J. Biol. Chem. 259, Turetta, L., Bazzan, E., Bertagno, K., Musacchio, E., and Deana, R. (2002) Cell Calcium 31, Bowman, E. J., Siebers, A., and Altendorf, K. (1988) Proc. Natl. Acad. Sci. U. S. A. 85, Brüne, B., and Ulrich, V. (1991) J. Biol. Chem. 266, Tsunoda, Y., Matsuno, K., and Tashiro, Y. (1991) Exp. Cell Res. 193, Rendu, F., Lebret, M., Nurden, A. T., Caen, J. P. (1982) Br. J. Haematol. 52, Fukami, M. H., Bauer, J. S., Stewart, G. J., and Salganicoff, L. (1978) J. Cell Biol. 77, Ruiz, F.A., Rodrigues, C.O., and Docampo, R. (2001) J. Biol. Chem. 276, Leighton, F., Poole, B., Beaufay, H., Baudhin, P., Coffey, J.W., Fowler, S., and De Duve, C. (1968) J. Cell Biol. 37,
19 20. Scott, D. A., de Souza, W., Benchimol, M., Zhong, L., Lu, H.-G., Moreno, S. N. J., and Docampo, R. (1998) J. Biol. Chem. 273, Lu, H.-G., Zhong, L., Chang, K.-P., and Docampo, R. (1997) J. Biol. Chem. 272, Docampo, R., Scott, D.S., Vercesi, A.E., and Moreno, S.N.J. (1995) Biochem. J., 310, Van Wazer, J. R., and Ditchfield, R. (1987) in Phosphorus NMR in Biology (Burt, C. T., ed) pp. 1-23, CRC Press, Boca Raton, FL. 24. Moreno, B., Urbina, J. A., Oldfield, E., Bailey, B. N., Rodrigues, C. O., and Docampo, R. (2000) J. Biol. Chem. 275, Bull, B. S. (1966) Blood 28, White, J. G. (1969) Blood 33, Costa, J. L., Reese, T. S., and Murphy, D. L. (1974) Science 183, Costa, J.L., Detwiler, T. C., Feinman, R. D., Murphy, D. L., Patlak, D. S., and Pettigrew, K. D. (1977) J. Physiol. 264, Skaer, R.J., Peters, P. D., and Emmines, J. P. (1976) J. Cell Sci. 20, Lu, H.-G., Zhong, L., de Souza, W., Benchimol, M., Moreno, S. N. J., and Docampo, R. (1998) Mol. Cell. Biol. 18, Rodrigues, C. O., Scott, D. A., and Docampo, R. (1999) Mol. Cell. Biol. 19, Allan, R.A. and Miller, J. J. (1980) Can J. Microbiol. 26, Fukami, M.H., Dangelmaier, C.A., Bauer, J.S., and Holmsen, H. (1980) Biochem. J. 192, Sun, B., and Kamayashi, J.-i. (2000) J. Biomed. Sci. 7, Thastrup, O., Cullen, P. J., Drobak, B. K., Hanley, M. R., Dawson, A. P. (1990) Proc. Natl. Acad. Sci. U. S. A. 87,
20 36. Liu, C., and Hermann, T.E. (1978) J. Biol. Chem. 253, Zhen, R.-G., Baykov, A. A., Bakuleva, N. P., and Rea, P. A. (1994) Plant Physiol. 104, Shiba, T., Nishimura, D., Kawazoe, Y., Onodera, Y., Tsutsumi, K., Nakamura, R., and Ohshiro, M. (2003) J. Biol. Chem. 278, Authi, K. S., Bokkala, S., Patel, Y., Kakkar, V.V., and Munkonge, F. (1993) Biochem. J. 294, Brass, L. F., and Joseph, S. K. (1985) J. Biol. Chem. 260, Papp, B., Enyedi, A., Paszty, K., Kovacs, T., Sakardi, B., Gardos, G., Magnier, C., Wuytack, F., and Enouf, J. (1992) Biochem. J. 288, Engelender, S., Wolosker, H., and de Meis, L. (1995) J. Biol. Chem. 270, Popov, E. G., Gabbasov, Z.A., Gavilov, I.Y., Pozin, E.Y., Mejlumina, A.G. (1987) Thromb. Res. 47, Skaer, R. J., Flemans, R. J., and McQuilkan, S. (1981) Brit. J. Haematol. 49, Costa, J. L, Fay, D. D., Nurnberger, J. I., and Murphy, D. L. (1982) Biochem Pharmacol. 31, Lemercier, G., Espiau, B., Ruiz, F. A., Vieira, M., Luo, S., Baltz, T., Docampo, R., and Bakalara, N. (2004) J. Biol. Chem. 279, Brightman, A. O., Navas, P., Minnifield, N. M., and Morré, D. J. (1992) Biochim. Biophys. Acta 1104, Scott, D. A., and Docampo, R. (2000) J. Biol. Chem. 275, Merzendorfer, H,m Gräf, R., Huss, M., Harvey, W. R., and Wieczorek, H. (1997) J. Exp. Biol. 200, Moriyama, Y., and Nelson, N. (1989) J. Biol. Chem. 264, Boullin, D. J., Green, A. R. (1972) Br J Pharmacol. 45,
21 52. Meyer, A. (1904) Bot. Zeit. 62, Jensen, T. E. (1993) In Ultrastructure of Microalgae (Berner, T., Ed) Chapter 1, pp. 7-50, CRC Press Boca Raton, Florida. 21
22 Table P NMR resonance assignments for perchloric acid extracts of human platelets. Peak Assignment Chemical shift (ppm) A Inorganic phosphate 1.22 B g-p of ATP C b-p ADP D a-p of polyphosphates E Pyrophosphate F a-p of ADP G a-p of ATP H b-p of ATP I Central P of polyphosphates
23 FIGURE LEGENDS FIG. 1. Electron microscopy and X-ray microanalysis of whole platelets. A, Transmission electron microscopy of an unfixed and unstained platelet preparation. Dense granules are identified by arrows. Bar, 1 µm. B, Typical X-ray microanalysis spectrum of dense granules in whole platelets. C, X-ray microanalysis spectrum of the background of whole platelets. FIG. 2. Electron microscopy of the dense granule fraction. A, direct observation of unfixed and unstained dense granules air-dried directly onto microscope grids. The inset shows, at higher magnification, the sponge-like structure of the dense granules after submission to the electron beam. B, Fixed and sectioned dense granule fraction. Bars: 0.25 µm FIG. 3. Distribution of different markers from human platelets on metrizamide gradients. Serotonin, PP i and short chain polyp are all present in the dense granule fraction (fraction 12). This distribution was compared with that of established platelet organelle markers, acid phosphatase (lysosomes), von Willebrand factor (alpha granules) and cytochrome c reductase (mitochondria). FIG. 4. Fluorescence microscopy showing the localization of polyp using DAPI. A, Platelets were treated with DAPI as described under Experimental Procedures. Note the accumulation of DAPI in small organelles. Arrow in B (bright field microscopy) shows an activated platelet. Bar: 10 µm. FIG MHz ( 1 H decoupled) 31 P NMR spectra of perchloric acid extracts of human platelets. Resonance assignments are given in Table 1. 23
24 FIG. 6. Urea-PAGE analysis of polyp from human platelets from three different donors. PolyP extracted from platelets was electrophoresed by 6% urea-page. Chain lengths of standards are on the left. An arrowhead on the right shows the position of migration of samples from three different donors. FIG. 7. Time course of thrombin-induced serotonin release compared to PP i and polyp. Release of serotonin (triangles), PP i (squares), and short chain polyp (circles) in platelets after the addition of 1.2 U/ml thrombin as described under Experimental Procedures. Results are expressed as percentage of the material released at 300 sec (representative of 3 experiments). FIG. 8. Effect of ionophores and NH 4 Cl on human platelets [Ca 2+ ] i. Platelets were loaded with fura-2/am as described under Experimental Procedures and resuspended in New Tyrode's buffer containing 1 mm EGTA. Thapsigargin (TG, 1 µm), nigericin (NIG, 2 µm), ionomycin (ION, 1 µm), or NH 4 Cl (20 mm) were added where indicated. FIG. 9. Distribution of bafilomycin A 1 -sensitive ATPase and pyrophosphatase activities on metrizamide gradients and acridine orange uptake by isolated dense granule fraction. A, bafilomycin A 1 (1 µm)-sensitive ATPase, pyrophosphatase, and AMDP (20 µm)-sensitive pyrophosphatase activities are all enriched in the dense granule fraction (Fraction 12). Other conditions were as described in the legend to Fig. 3, and under Experimental Proceures. B, Dense granule fraction (45 µg protein) was added to 2.5 ml of a reaction medium described under Experimental Procedures. Acridine orange (3 µm), ATP (1 mm), 24
25 PP i (0.1 mm), of NH 4 Cl (20 mm) were added where indicated. Changes in absorbance were followed at nm. 25
26 A B C 400 Counts 70 C O P Cu Si Ca K Cu Cu 10 KeV Counts C Cu Si Ruiz et al., Fig KeV

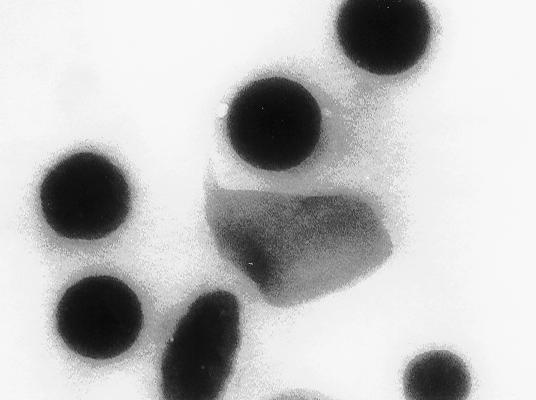
27 A B Ruiz et al., Fig. 2
28 % SC PolyP %PPi % Serotonin incorporation % von Willebrand factor %CytC reductase activity Density (g/cm 3 ) Fraction % Acid Phosphatase activity Proteins (mg/ml) Fraction Ruiz et al., Fig. 3

29 A B Ruiz et al., Fig. 4
30 Ruiz et al., Fig. 5

31 PolyP 470 PolyP 300 PolyP 75 PolyP 45 PolyP 25 PolyP 15 PolyP 5 Samples Ruiz et al., Fig. 6
32 120 % Total secreted Time (sec) Ruiz et al., Fig. 7
33 Intracellular Ca +2 concentrations TG 50 NIG TG A B NIG NH 4 Cl 50 0 ION NH ION 4 Cl Time (s). Ruiz et al., Fig. 8
34 A % Total Pyrophosphatase activity % PPase (AMDP-sensitive) activity % ATPase (BAF-sensitive) activity B Fraction Ruiz et al., Fig. 9 Downloaded from by guest on April 7, 2018
35 Human platelet dense granules contain polyphosphate and are similar to acidocalcisomes of bacteria and unicellular eukaryotes Felix A. Ruiz, Christopher R. Lea, Eric Oldfield and Roberto Docampo J. Biol. Chem. published online August 11, 2004 Access the most updated version of this article at doi: /jbc.M Alerts: When this article is cited When a correction for this article is posted Click here to choose from all of JBC's alerts
Acidocalcisomes Are Functionally Linked to the Contractile Vacuole of Dictyostelium discoideum*
 THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 277, No. 10, Issue of March 8, pp. 8146 8153, 2002 2002 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. Acidocalcisomes Are
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 277, No. 10, Issue of March 8, pp. 8146 8153, 2002 2002 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. Acidocalcisomes Are
The Polyphosphate Bodies of Chlamydomonas reinhardtii Possess a Proton-pumping Pyrophosphatase and Are Similar to Acidocalcisomes*
 THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 276, No. 49, Issue of December 7, pp. 46196 46203, 2001 2001 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. The Polyphosphate
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 276, No. 49, Issue of December 7, pp. 46196 46203, 2001 2001 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. The Polyphosphate
Heparin Sodium ヘパリンナトリウム
 Heparin Sodium ヘパリンナトリウム Add the following next to Description: Identification Dissolve 1 mg each of Heparin Sodium and Heparin Sodium Reference Standard for physicochemical test in 1 ml of water, and
Heparin Sodium ヘパリンナトリウム Add the following next to Description: Identification Dissolve 1 mg each of Heparin Sodium and Heparin Sodium Reference Standard for physicochemical test in 1 ml of water, and
SUPPLEMENTARY MATERIAL
 SUPPLEMENTARY MATERIAL Purification and biochemical properties of SDS-stable low molecular weight alkaline serine protease from Citrullus Colocynthis Muhammad Bashir Khan, 1,3 Hidayatullah khan, 2 Muhammad
SUPPLEMENTARY MATERIAL Purification and biochemical properties of SDS-stable low molecular weight alkaline serine protease from Citrullus Colocynthis Muhammad Bashir Khan, 1,3 Hidayatullah khan, 2 Muhammad
ab CytoPainter Golgi/ER Staining Kit
 ab139485 CytoPainter Golgi/ER Staining Kit Instructions for Use Designed to detect Golgi bodies and endoplasmic reticulum by microscopy This product is for research use only and is not intended for diagnostic
ab139485 CytoPainter Golgi/ER Staining Kit Instructions for Use Designed to detect Golgi bodies and endoplasmic reticulum by microscopy This product is for research use only and is not intended for diagnostic
Supporting Information for:
 Supporting Information for: Methylerythritol Cyclodiphosphate (MEcPP) in Deoxyxylulose Phosphate Pathway: Synthesis from an Epoxide and Mechanisms Youli Xiao, a Rodney L. Nyland II, b Caren L. Freel Meyers
Supporting Information for: Methylerythritol Cyclodiphosphate (MEcPP) in Deoxyxylulose Phosphate Pathway: Synthesis from an Epoxide and Mechanisms Youli Xiao, a Rodney L. Nyland II, b Caren L. Freel Meyers
User s Manual and Instructions
 User s Manual and Instructions Mitochondria Activity Assay (Cytochrome C Oxidase Activity Assay) Kit Catalog Number: KC310100 Introduction Mitochondria are the eukaryotic subcellular organelles that contain
User s Manual and Instructions Mitochondria Activity Assay (Cytochrome C Oxidase Activity Assay) Kit Catalog Number: KC310100 Introduction Mitochondria are the eukaryotic subcellular organelles that contain
Supplementary Material (ESI) for Chemical Communications This journal is (c) The Royal Society of Chemistry 2008
 for Chemical Communications This journal is (c) The Royal Society of Chemistry 2008") Experimental Details Unless otherwise noted, all chemicals were purchased from Sigma-Aldrich Chemical Company and were used as received. 2-DOS and neamine were kindly provided by Dr. F. Huang. Paromamine
Experimental Details Unless otherwise noted, all chemicals were purchased from Sigma-Aldrich Chemical Company and were used as received. 2-DOS and neamine were kindly provided by Dr. F. Huang. Paromamine
Rapid changes in polyphosphate content within. acidocalcisomes in response to cell growth, differentiation
 JBC Papers in Press. Published on May 22, 2001 as Manuscript M102402200 Running Title: Acidocalcisomal polyphosphate in T. cruzi Rapid changes in polyphosphate content within acidocalcisomes in response
JBC Papers in Press. Published on May 22, 2001 as Manuscript M102402200 Running Title: Acidocalcisomal polyphosphate in T. cruzi Rapid changes in polyphosphate content within acidocalcisomes in response
Thursday, October 16 th
 Thursday, October 16 th Good morning. Those of you needing to take the Enzymes and Energy Quiz will start very soon. Students who took the quiz Wednesday: Please QUIETLY work on the chapter 6 reading guide.
Thursday, October 16 th Good morning. Those of you needing to take the Enzymes and Energy Quiz will start very soon. Students who took the quiz Wednesday: Please QUIETLY work on the chapter 6 reading guide.
Chromatin IP (Isw2) Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles.
 Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles.") Chromatin IP (Isw2) 7/01 Toshi last update: 06/15 Reagents Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles. 2.5 M glycine. TBS:
Chromatin IP (Isw2) 7/01 Toshi last update: 06/15 Reagents Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles. 2.5 M glycine. TBS:
ENDOPLASMIC RETICULUM MEMBRANE ISOLATED FROM SMALL-INTESTINAL EPITHELIAL CELLS: ENZYME AND PROTEIN COMPONENTS
 J. Cell Sci. 5a, 215-222 (1981) 21 c Printed in Great Britain Company of Biologist! Limited 1981 ENDOPLASMIC RETICULUM MEMBRANE ISOLATED FROM SMALL-INTESTINAL EPITHELIAL CELLS: ENZYME AND PROTEIN COMPONENTS
J. Cell Sci. 5a, 215-222 (1981) 21 c Printed in Great Britain Company of Biologist! Limited 1981 ENDOPLASMIC RETICULUM MEMBRANE ISOLATED FROM SMALL-INTESTINAL EPITHELIAL CELLS: ENZYME AND PROTEIN COMPONENTS
Supplementary material: Materials and suppliers
 Supplementary material: Materials and suppliers Electrophoresis consumables including tris-glycine, acrylamide, SDS buffer and Coomassie Brilliant Blue G-2 dye (CBB) were purchased from Ameresco (Solon,
Supplementary material: Materials and suppliers Electrophoresis consumables including tris-glycine, acrylamide, SDS buffer and Coomassie Brilliant Blue G-2 dye (CBB) were purchased from Ameresco (Solon,
A ph-dependent Charge Reversal Peptide for Cancer Targeting
 Supporting Information A ph-dependent Charge Reversal Peptide for Cancer Targeting Naoko Wakabayashi 1, Yoshiaki Yano 1, Kenichi Kawano 1, and Katsumi Matsuzaki 1 1 Graduate School of Pharmaceutical Sciences,
Supporting Information A ph-dependent Charge Reversal Peptide for Cancer Targeting Naoko Wakabayashi 1, Yoshiaki Yano 1, Kenichi Kawano 1, and Katsumi Matsuzaki 1 1 Graduate School of Pharmaceutical Sciences,
LANCE Eu-W1024 ITC Chelate & Europium Standard AD0013 Development grade
 AD0017P-4 (en) 1 LANCE Eu-W1024 ITC Chelate & Europium Standard AD0013 Development grade INTRODUCTION Fluorescent isothiocyanato-activated (ITC-activated) Eu-W1024 chelate is optimized for labelling proteins
AD0017P-4 (en) 1 LANCE Eu-W1024 ITC Chelate & Europium Standard AD0013 Development grade INTRODUCTION Fluorescent isothiocyanato-activated (ITC-activated) Eu-W1024 chelate is optimized for labelling proteins
PhosFree TM Phosphate Assay Biochem Kit
 PhosFree TM Phosphate Assay Biochem Kit (Cat. # BK050) ORDERING INFORMATION To order by phone: (303) - 322-2254 To order by Fax: (303) - 322-2257 To order by e-mail: cservice@cytoskeleton.com Technical
PhosFree TM Phosphate Assay Biochem Kit (Cat. # BK050) ORDERING INFORMATION To order by phone: (303) - 322-2254 To order by Fax: (303) - 322-2257 To order by e-mail: cservice@cytoskeleton.com Technical
Mitochondrial DNA Isolation Kit
 Mitochondrial DNA Isolation Kit Catalog Number KA0895 50 assays Version: 01 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 General Information... 4 Materials
Mitochondrial DNA Isolation Kit Catalog Number KA0895 50 assays Version: 01 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 General Information... 4 Materials
Deep Oscillation EFFECTS ON BLOOD PARAMETERS (EXPERIMENTAL STUDY)
") Deep Oscillation EFFECTS ON BLOOD PARAMETERS (EXPERIMENTAL STUDY) I. EFFECTS OF Deep Oscillation ON THE WHOLE BLOOD AND WHITE BLOOD CELL CHEMILUMINESCENCE The stimulation of oxygen radical production by
Deep Oscillation EFFECTS ON BLOOD PARAMETERS (EXPERIMENTAL STUDY) I. EFFECTS OF Deep Oscillation ON THE WHOLE BLOOD AND WHITE BLOOD CELL CHEMILUMINESCENCE The stimulation of oxygen radical production by
ASSAY OF SPHINGOMYELINASE ACTIVITY
 ASSAY OF SPHINGOMYELINASE ACTIVITY Protocol for Protein Extraction Stock Solution 1. Leupeptin/hydrochloride (FW 463.0,
ASSAY OF SPHINGOMYELINASE ACTIVITY Protocol for Protein Extraction Stock Solution 1. Leupeptin/hydrochloride (FW 463.0,
Figure S1. PMVs from THP-1 cells expose phosphatidylserine and carry actin. A) Flow
 Flow") SUPPLEMENTARY DATA Supplementary Figure Legends Figure S1. PMVs from THP-1 cells expose phosphatidylserine and carry actin. A) Flow cytometry analysis of PMVs labelled with annexin-v-pe (Guava technologies)
SUPPLEMENTARY DATA Supplementary Figure Legends Figure S1. PMVs from THP-1 cells expose phosphatidylserine and carry actin. A) Flow cytometry analysis of PMVs labelled with annexin-v-pe (Guava technologies)
BIOL 347L Laboratory Three
 Introduction BIOL 347L Laboratory Three Osmosis in potato and carrot samples Osmosis is the movement of water molecules through a selectively permeable membrane into a region of higher solute concentration,
Introduction BIOL 347L Laboratory Three Osmosis in potato and carrot samples Osmosis is the movement of water molecules through a selectively permeable membrane into a region of higher solute concentration,
Caution: For Laboratory Use. A product for research purposes only. Eu-W1024 ITC Chelate & Europium Standard. Product Number: AD0013
 TECHNICAL DATA SHEET Lance Caution: For Laboratory Use. A product for research purposes only. Eu-W1024 ITC Chelate & Europium Standard Product Number: AD0013 INTRODUCTION: Fluorescent isothiocyanato-activated
TECHNICAL DATA SHEET Lance Caution: For Laboratory Use. A product for research purposes only. Eu-W1024 ITC Chelate & Europium Standard Product Number: AD0013 INTRODUCTION: Fluorescent isothiocyanato-activated
Caution: For Laboratory Use. A product for research purposes only. Eu-W1284 Iodoacetamido Chelate & Europium Standard. Product Number: AD0014
 TECHNICAL DATA SHEET Lance Caution: For Laboratory Use. A product for research purposes only. Eu-W1284 Iodoacetamido Chelate & Europium Standard Product Number: AD0014 INTRODUCTION: Iodoacetamido-activated
TECHNICAL DATA SHEET Lance Caution: For Laboratory Use. A product for research purposes only. Eu-W1284 Iodoacetamido Chelate & Europium Standard Product Number: AD0014 INTRODUCTION: Iodoacetamido-activated
Nature Protocols: doi: /nprot Supplementary Figure 1. Fluorescent titration of probe CPDSA.
 Supplementary Figure 1 Fluorescent titration of probe CPDSA. Fluorescent titration of probe CPDSA (10 um) upon addition of GSH in HEPES (10 mm, ph = 7.4) containing 10% DMSO. Each spectrum was recorded
Supplementary Figure 1 Fluorescent titration of probe CPDSA. Fluorescent titration of probe CPDSA (10 um) upon addition of GSH in HEPES (10 mm, ph = 7.4) containing 10% DMSO. Each spectrum was recorded
Work-flow: protein sample preparation Precipitation methods Removal of interfering substances Specific examples:
 Dr. Sanjeeva Srivastava IIT Bombay Work-flow: protein sample preparation Precipitation methods Removal of interfering substances Specific examples: Sample preparation for serum proteome analysis Sample
Dr. Sanjeeva Srivastava IIT Bombay Work-flow: protein sample preparation Precipitation methods Removal of interfering substances Specific examples: Sample preparation for serum proteome analysis Sample
CELLS and TRANSPORT Student Packet SUMMARY CELL MEMBRANES ARE SELECTIVELY PERMEABLE DUE TO THEIR STRUCTURE Hydrophilic head
 CELLS and TRANSPORT Student Packet SUMMARY CELL MEMBRANES ARE SELECTIVELY PERMEABLE DUE TO THEIR STRUCTURE Hydrophilic head Hydrophobic tail Hydrophobic regions of protein Hydrophilic regions of protein
CELLS and TRANSPORT Student Packet SUMMARY CELL MEMBRANES ARE SELECTIVELY PERMEABLE DUE TO THEIR STRUCTURE Hydrophilic head Hydrophobic tail Hydrophobic regions of protein Hydrophilic regions of protein
Instructions for Use. APO-AB Annexin V-Biotin Apoptosis Detection Kit 100 tests
 3URGXFW,QIRUPDWLRQ Sigma TACS Annexin V Apoptosis Detection Kits Instructions for Use APO-AB Annexin V-Biotin Apoptosis Detection Kit 100 tests For Research Use Only. Not for use in diagnostic procedures.
3URGXFW,QIRUPDWLRQ Sigma TACS Annexin V Apoptosis Detection Kits Instructions for Use APO-AB Annexin V-Biotin Apoptosis Detection Kit 100 tests For Research Use Only. Not for use in diagnostic procedures.
FOCUS SubCell. For the Enrichment of Subcellular Fractions. (Cat. # ) think proteins! think G-Biosciences
 think proteins! think G-Biosciences") 169PR 01 G-Biosciences 1-800-628-7730 1-314-991-6034 technical@gbiosciences.com A Geno Technology, Inc. (USA) brand name FOCUS SubCell For the Enrichment of Subcellular Fractions (Cat. # 786 260) think
169PR 01 G-Biosciences 1-800-628-7730 1-314-991-6034 technical@gbiosciences.com A Geno Technology, Inc. (USA) brand name FOCUS SubCell For the Enrichment of Subcellular Fractions (Cat. # 786 260) think
DELFIA Tb-N1 DTA Chelate & Terbium Standard
 AD0029P-1 (en) 1 DELFIA Tb-N1 DTA Chelate & AD0012 Terbium Standard For Research Use Only INTRODUCTION DELFIA Tb-N1 DTA Chelate is optimized for the terbium labeling of proteins and peptides for use in
AD0029P-1 (en) 1 DELFIA Tb-N1 DTA Chelate & AD0012 Terbium Standard For Research Use Only INTRODUCTION DELFIA Tb-N1 DTA Chelate is optimized for the terbium labeling of proteins and peptides for use in
Tunable Hydrophobicity in DNA Micelles Anaya, Milena; Kwak, Minseok; Musser, Andrew J.; Muellen, Klaus; Herrmann, Andreas; Müllen, Klaus
 University of Groningen Tunable Hydrophobicity in DNA Micelles Anaya, Milena; Kwak, Minseok; Musser, Andrew J.; Muellen, Klaus; Herrmann, Andreas; Müllen, Klaus Published in: Chemistry DOI: 10.1002/chem.201001816
University of Groningen Tunable Hydrophobicity in DNA Micelles Anaya, Milena; Kwak, Minseok; Musser, Andrew J.; Muellen, Klaus; Herrmann, Andreas; Müllen, Klaus Published in: Chemistry DOI: 10.1002/chem.201001816
Supporting Information
 Electronic Supplementary Material (ESI) for ChemComm. This journal is The Royal Society of Chemistry 2015 Supporting Information Enzyme-activatable Probe with a Self-immolative Linker for Rapid and Sensitive
Electronic Supplementary Material (ESI) for ChemComm. This journal is The Royal Society of Chemistry 2015 Supporting Information Enzyme-activatable Probe with a Self-immolative Linker for Rapid and Sensitive
Plasmonic blood glucose monitor based on enzymatic. etching of gold nanorods
 Plasmonic blood glucose monitor based on enzymatic etching of gold nanorods Xin Liu, Shuya Zhang, Penglong Tan, Jiang Zhou, Yan Huang, Zhou Nie* and Shouzhuo Yao State Key Laboratory of Chemo/Biosensing
Plasmonic blood glucose monitor based on enzymatic etching of gold nanorods Xin Liu, Shuya Zhang, Penglong Tan, Jiang Zhou, Yan Huang, Zhou Nie* and Shouzhuo Yao State Key Laboratory of Chemo/Biosensing
Cholesterol determination using protein-templated fluorescent gold nanocluster probes
 Electronic Supplementary Information for Cholesterol determination using protein-templated fluorescent gold nanocluster probes Xi Chen and Gary A. Baker* Department of Chemistry, University of Missouri-Columbia,
Electronic Supplementary Information for Cholesterol determination using protein-templated fluorescent gold nanocluster probes Xi Chen and Gary A. Baker* Department of Chemistry, University of Missouri-Columbia,
BILAYER CHANNEL RECONSTITUTION
 (1) 1% Agar Salt Bridge 1.0 g Agar 3.75g KCl in 100ml distilled water, store at 4 o C. BILAYER CHANNEL RECONSTITUTION (2) Cs solution: (Cesium Methanesulfonate) 1) 50 mm Cs + solution 0.209 MOPS, 10mM
(1) 1% Agar Salt Bridge 1.0 g Agar 3.75g KCl in 100ml distilled water, store at 4 o C. BILAYER CHANNEL RECONSTITUTION (2) Cs solution: (Cesium Methanesulfonate) 1) 50 mm Cs + solution 0.209 MOPS, 10mM
MEK1 Assay Kit 1 Catalog # Lot # 16875
 MEK1 Assay Kit 1 Kit Components Assay Dilution Buffer (ADB), Catalog # 20-108. Three vials, each containing 1.0ml of assay dilution buffer (20mM MOPS, ph 7.2, 25mM ß-glycerol phosphate, 5mM EGTA, 1mM sodium
MEK1 Assay Kit 1 Kit Components Assay Dilution Buffer (ADB), Catalog # 20-108. Three vials, each containing 1.0ml of assay dilution buffer (20mM MOPS, ph 7.2, 25mM ß-glycerol phosphate, 5mM EGTA, 1mM sodium
Protocol for Gene Transfection & Western Blotting
 The schedule and the manual of basic techniques for cell culture Advanced Protocol for Gene Transfection & Western Blotting Schedule Day 1 26/07/2008 Transfection Day 3 28/07/2008 Cell lysis Immunoprecipitation
The schedule and the manual of basic techniques for cell culture Advanced Protocol for Gene Transfection & Western Blotting Schedule Day 1 26/07/2008 Transfection Day 3 28/07/2008 Cell lysis Immunoprecipitation
AMPK Assay. Require: Sigma (1L, $18.30) A4206 Aluminum foil
 A4206 Aluminum foil") AMPK Assay Require: Acetone Sigma (1L, $18.30) A4206 Aluminum foil Ammonium sulfate Fisher BP212R-1 AMP Sigma A1752 ATP Sigma A6144 (alt. use A7699) Beta-mercaptoethanol Sigma M6250 (alt. use M7154) Bio-Rad
AMPK Assay Require: Acetone Sigma (1L, $18.30) A4206 Aluminum foil Ammonium sulfate Fisher BP212R-1 AMP Sigma A1752 ATP Sigma A6144 (alt. use A7699) Beta-mercaptoethanol Sigma M6250 (alt. use M7154) Bio-Rad
1. endoplasmic reticulum This is the location where N-linked oligosaccharide is initially synthesized and attached to glycoproteins.
 Biology 4410 Name Spring 2006 Exam 2 A. Multiple Choice, 2 pt each Pick the best choice from the list of choices, and write it in the space provided. Some choices may be used more than once, and other
Biology 4410 Name Spring 2006 Exam 2 A. Multiple Choice, 2 pt each Pick the best choice from the list of choices, and write it in the space provided. Some choices may be used more than once, and other
Blood Cells. Dr. Sami Zaqout. Dr. Sami Zaqout Faculty of Medicine IUG
 Blood Cells Dr. Sami Zaqout Blood Blood Blood cells (45%) Erythrocytes Platelets Leukocytes Plasma (55%) Hematocrit tubes with blood Composition of Plasma Plasma Aqueous solution (90%) Substances (10%)
Blood Cells Dr. Sami Zaqout Blood Blood Blood cells (45%) Erythrocytes Platelets Leukocytes Plasma (55%) Hematocrit tubes with blood Composition of Plasma Plasma Aqueous solution (90%) Substances (10%)
Mammalian Membrane Protein Extraction Kit
 Mammalian Membrane Protein Extraction Kit Catalog number: AR0155 Boster s Mammalian Membrane Protein Extraction Kit is a simple, rapid and reproducible method to prepare cellular protein fractions highly
Mammalian Membrane Protein Extraction Kit Catalog number: AR0155 Boster s Mammalian Membrane Protein Extraction Kit is a simple, rapid and reproducible method to prepare cellular protein fractions highly
BIL 256 Cell and Molecular Biology Lab Spring, Tissue-Specific Isoenzymes
 BIL 256 Cell and Molecular Biology Lab Spring, 2007 Background Information Tissue-Specific Isoenzymes A. BIOCHEMISTRY The basic pattern of glucose oxidation is outlined in Figure 3-1. Glucose is split
BIL 256 Cell and Molecular Biology Lab Spring, 2007 Background Information Tissue-Specific Isoenzymes A. BIOCHEMISTRY The basic pattern of glucose oxidation is outlined in Figure 3-1. Glucose is split
DELFIA Eu-DTPA ITC Chelate & Europium Standard
 AD0026P-3 (en) 1 DELFIA Eu-DTPA ITC Chelate & AD0021 Europium Standard For Research Use Only INTRODUCTION DELFIA Eu-DTPA ITC Chelate is optimized for the europium labelling of proteins and peptides for
AD0026P-3 (en) 1 DELFIA Eu-DTPA ITC Chelate & AD0021 Europium Standard For Research Use Only INTRODUCTION DELFIA Eu-DTPA ITC Chelate is optimized for the europium labelling of proteins and peptides for
TFEB-mediated increase in peripheral lysosomes regulates. Store Operated Calcium Entry
 TFEB-mediated increase in peripheral lysosomes regulates Store Operated Calcium Entry Luigi Sbano, Massimo Bonora, Saverio Marchi, Federica Baldassari, Diego L. Medina, Andrea Ballabio, Carlotta Giorgi
TFEB-mediated increase in peripheral lysosomes regulates Store Operated Calcium Entry Luigi Sbano, Massimo Bonora, Saverio Marchi, Federica Baldassari, Diego L. Medina, Andrea Ballabio, Carlotta Giorgi
Europium Labeling Kit
 Europium Labeling Kit Catalog Number KA2096 100ug *1 Version: 03 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Intended Use... 3 Background... 3 Principle of the Assay...
Europium Labeling Kit Catalog Number KA2096 100ug *1 Version: 03 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Intended Use... 3 Background... 3 Principle of the Assay...
BabyBio IMAC columns DATA SHEET DS
 BabyBio IMAC columns DATA SHEET DS 45 655 010 BabyBio columns for Immobilized Metal Ion Affinity Chromatography (IMAC) are ready-to-use for quick and easy purification of polyhistidine-tagged (His-tagged)
BabyBio IMAC columns DATA SHEET DS 45 655 010 BabyBio columns for Immobilized Metal Ion Affinity Chromatography (IMAC) are ready-to-use for quick and easy purification of polyhistidine-tagged (His-tagged)
STORE AT 4 o C Version 3
 Instruction Manual Advanced Protein Assay Reagent (Cat. # ADV01) ORDERING INFORMATION To order by phone: (303) - 322-2254 To order by Fax: (303) - 322-2257 To order by e-mail: cserve@cytoskeleton.com Technical
Instruction Manual Advanced Protein Assay Reagent (Cat. # ADV01) ORDERING INFORMATION To order by phone: (303) - 322-2254 To order by Fax: (303) - 322-2257 To order by e-mail: cserve@cytoskeleton.com Technical
Characterization of the DNA-mediated Oxidation of Dps, a Bacterial Ferritin
 SUPPORTING INFORMATION Characterization of the DNA-mediated Oxidation of Dps, a Bacterial Ferritin Anna R. Arnold, Andy Zhou, and Jacqueline K. Barton Division of Chemistry and Chemical Engineering, California
SUPPORTING INFORMATION Characterization of the DNA-mediated Oxidation of Dps, a Bacterial Ferritin Anna R. Arnold, Andy Zhou, and Jacqueline K. Barton Division of Chemistry and Chemical Engineering, California
E.Z.N.A. SQ Blood DNA Kit II. Table of Contents
 E.Z.N.A. SQ Blood DNA Kit II Table of Contents Introduction and Overview...2 Kit Contents/Storage and Stability...3 Blood Storage and DNA Yield...4 Preparing Reagents...5 100-500 μl Whole Blood Protocol...6
E.Z.N.A. SQ Blood DNA Kit II Table of Contents Introduction and Overview...2 Kit Contents/Storage and Stability...3 Blood Storage and DNA Yield...4 Preparing Reagents...5 100-500 μl Whole Blood Protocol...6
ab Membrane Fractionation Kit Instructions for Use For the rapid and simple separation of membrane, cytosolic and nuclear cellular fractions.
 ab139409 Membrane Fractionation Kit Instructions for Use For the rapid and simple separation of membrane, cytosolic and nuclear cellular fractions. This product is for research use only and is not intended
ab139409 Membrane Fractionation Kit Instructions for Use For the rapid and simple separation of membrane, cytosolic and nuclear cellular fractions. This product is for research use only and is not intended
CytoPainter Lysosomal Staining Kit - Blue Fluorescence
 ab112135 CytoPainter Lysosomal Staining Kit - Blue Fluorescence Instructions for Use For staining Lysosomes in suspension and adherent cells by using our proprietary blue fluorescence probe This product
ab112135 CytoPainter Lysosomal Staining Kit - Blue Fluorescence Instructions for Use For staining Lysosomes in suspension and adherent cells by using our proprietary blue fluorescence probe This product
Molecular Cell Biology Problem Drill 16: Intracellular Compartment and Protein Sorting
 Molecular Cell Biology Problem Drill 16: Intracellular Compartment and Protein Sorting Question No. 1 of 10 Question 1. Which of the following statements about the nucleus is correct? Question #01 A. The
Molecular Cell Biology Problem Drill 16: Intracellular Compartment and Protein Sorting Question No. 1 of 10 Question 1. Which of the following statements about the nucleus is correct? Question #01 A. The
DIDS INHIBITION OF SARCOPLASMIC RETICULUM ANION EFFLUX AND CALCIUM TRANSPORT
 DIDS INHIBITION OF SARCOPLASMIC RETICULUM ANION EFFLUX AND CALCIUM TRANSPORT Kevin P. Campbell and David H. MacLennan Reprinted from ANNALS OF THE NEW YORK ACADEMY OF SCIENCES Volume 358 Pages 328-331
DIDS INHIBITION OF SARCOPLASMIC RETICULUM ANION EFFLUX AND CALCIUM TRANSPORT Kevin P. Campbell and David H. MacLennan Reprinted from ANNALS OF THE NEW YORK ACADEMY OF SCIENCES Volume 358 Pages 328-331
VaTx1 VaTx2 VaTx3. VaTx min Retention Time (min) Retention Time (min)
 Retention Time (min)") a Absorbance (mau) 5 2 5 3 4 5 6 7 8 9 6 2 3 4 5 6 VaTx2 High Ca 2+ Low Ca 2+ b 38.2 min Absorbance (mau) 3 2 3 4 5 3 2 VaTx2 39.3 min 3 4 5 3 2 4. min 3 4 5 Supplementary Figure. Toxin Purification For
a Absorbance (mau) 5 2 5 3 4 5 6 7 8 9 6 2 3 4 5 6 VaTx2 High Ca 2+ Low Ca 2+ b 38.2 min Absorbance (mau) 3 2 3 4 5 3 2 VaTx2 39.3 min 3 4 5 3 2 4. min 3 4 5 Supplementary Figure. Toxin Purification For
Mitochondrial Trifunctional Protein (TFP) Protein Quantity Microplate Assay Kit
 Protein Quantity Microplate Assay Kit") PROTOCOL Mitochondrial Trifunctional Protein (TFP) Protein Quantity Microplate Assay Kit DESCRIPTION Mitochondrial Trifunctional Protein (TFP) Protein Quantity Microplate Assay Kit Sufficient materials
PROTOCOL Mitochondrial Trifunctional Protein (TFP) Protein Quantity Microplate Assay Kit DESCRIPTION Mitochondrial Trifunctional Protein (TFP) Protein Quantity Microplate Assay Kit Sufficient materials
Organization of ATPases
 The Primary Active Transporter II: The ATPase Objectives: Organization P type with NPA domains Proton pumps of the rotary V type ATPase 1 Organization of P type, solute transport, found in plasma membranes
The Primary Active Transporter II: The ATPase Objectives: Organization P type with NPA domains Proton pumps of the rotary V type ATPase 1 Organization of P type, solute transport, found in plasma membranes
SensoLyte pnpp Alkaline Phosphatase Assay Kit *Colorimetric*
 SensoLyte pnpp Alkaline Phosphatase Assay Kit *Colorimetric* Catalog # 72146 Kit Size 500 Assays (96-well plate) Optimized Performance: This kit is optimized to detect alkaline phosphatase activity Enhanced
SensoLyte pnpp Alkaline Phosphatase Assay Kit *Colorimetric* Catalog # 72146 Kit Size 500 Assays (96-well plate) Optimized Performance: This kit is optimized to detect alkaline phosphatase activity Enhanced
Laboratory 8 Succinate Dehydrogenase Activity in Cauliflower Mitochondria
 BIO354: Cell Biology Laboratory 1 I. Introduction Laboratory 8 Succinate Dehydrogenase Activity in Cauliflower Mitochondria In eukaryotic cells, specific functions are localized to different types of organelles.
BIO354: Cell Biology Laboratory 1 I. Introduction Laboratory 8 Succinate Dehydrogenase Activity in Cauliflower Mitochondria In eukaryotic cells, specific functions are localized to different types of organelles.
Chapter 5 MITOCHONDRIA AND RESPIRATION 5-1
 Chapter 5 MITOCHONDRIA AND RESPIRATION All organisms must transform energy. This energy is required to maintain a dynamic steady state, homeostasis, and to insure continued survival. As will be discussed
Chapter 5 MITOCHONDRIA AND RESPIRATION All organisms must transform energy. This energy is required to maintain a dynamic steady state, homeostasis, and to insure continued survival. As will be discussed
Nitro-Grela-type complexes containing iodides. robust and selective catalysts for olefin metathesis
 Supporting Information for Nitro-Grela-type complexes containing iodides robust and selective catalysts for olefin metathesis under challenging conditions. Andrzej Tracz, 1,2 Mateusz Matczak, 1 Katarzyna
Supporting Information for Nitro-Grela-type complexes containing iodides robust and selective catalysts for olefin metathesis under challenging conditions. Andrzej Tracz, 1,2 Mateusz Matczak, 1 Katarzyna
PosterREPRINT A NOVEL APPROACH TO MALDI-TOF-MS SAMPLE PREPARATION. Presented at ABRF 2002, Austin, Texas, USA, 9th - 12th March 2002.
 Introduction A NOVEL APPROACH TO MALDI-TOF-MS SAMPLE PREPARATION Ed Bouvier 2, Jeff Brown 1, Emmanuelle Claude 1, John L. Gebler 2, Weibin Chen 2, *Dominic Gostick 1, Kevin Howes 1, James Langridge 1,
Introduction A NOVEL APPROACH TO MALDI-TOF-MS SAMPLE PREPARATION Ed Bouvier 2, Jeff Brown 1, Emmanuelle Claude 1, John L. Gebler 2, Weibin Chen 2, *Dominic Gostick 1, Kevin Howes 1, James Langridge 1,
Fig. S4. Current-voltage relations of iglurs. A-C: time courses of currents evoked by 100 ms pulses
 Fig. S1. Immunohistochemical detection of iglur2 protein in single islet cells. A: α cells identified using glucagon-specific antibody express the iglur2 subtype of AMPA receptor. 24 out of 26 identified
Fig. S1. Immunohistochemical detection of iglur2 protein in single islet cells. A: α cells identified using glucagon-specific antibody express the iglur2 subtype of AMPA receptor. 24 out of 26 identified
FIRST MIDTERM EXAMINATION
 FIRST MIDTERM EXAMINATION 1. True or false: because enzymes are produced by living organisms and because they allow chemical reactions to occur that would not otherwise occur, enzymes represent an exception
FIRST MIDTERM EXAMINATION 1. True or false: because enzymes are produced by living organisms and because they allow chemical reactions to occur that would not otherwise occur, enzymes represent an exception
DELFIA Tb-DTPA ITC Chelate & Terbium Standard
 AD0035P-2 (en) 1 DELFIA Tb-DTPA ITC Chelate & AD0029 Terbium Standard For Research Use Only INTRODUCTION DELFIA Tb-DTPA ITC Chelate is optimized for the terbium labelling of proteins and peptides for use
AD0035P-2 (en) 1 DELFIA Tb-DTPA ITC Chelate & AD0029 Terbium Standard For Research Use Only INTRODUCTION DELFIA Tb-DTPA ITC Chelate is optimized for the terbium labelling of proteins and peptides for use
ELECTRON MICROSCOPIC STUDIES ON EQUINE ENCEPHALOSIS VIRUS
 Onderstepoort]. vet. Res. 40 (2), 53-58 (1973) ELECTRON MICROSCOPIC STUDIES ON EQUINE ENCEPHALOSIS VIRUS G. LECATSAS, B. J. ERASMUS and H. J. ELS, Veterinary Research Institute, Onderstepoort ABSTRACT
Onderstepoort]. vet. Res. 40 (2), 53-58 (1973) ELECTRON MICROSCOPIC STUDIES ON EQUINE ENCEPHALOSIS VIRUS G. LECATSAS, B. J. ERASMUS and H. J. ELS, Veterinary Research Institute, Onderstepoort ABSTRACT
1. This is the location where N-linked oligosaccharide is initially synthesized and attached to glycoproteins.
 Biology 4410 Name Spring 2006 Exam 2 A. Multiple Choice, 2 pt each Pick the best choice from the list of choices, and write it in the space provided. Some choices may be used more than once, and other
Biology 4410 Name Spring 2006 Exam 2 A. Multiple Choice, 2 pt each Pick the best choice from the list of choices, and write it in the space provided. Some choices may be used more than once, and other
Experiment 1. Isolation of Glycogen from rat Liver
 Experiment 1 Isolation of Glycogen from rat Liver Figure 35: FIG-2, Liver, PAS, 100x. Note the presence of a few scattered glycogen granules (GG). Objective To illustrate the method for isolating glycogen.
Experiment 1 Isolation of Glycogen from rat Liver Figure 35: FIG-2, Liver, PAS, 100x. Note the presence of a few scattered glycogen granules (GG). Objective To illustrate the method for isolating glycogen.
Annexin V-PE Apoptosis Detection Kit
 Annexin V-PE Apoptosis Detection Kit Catalog Number KA0716 100 assays Version: 02 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 General Information...
Annexin V-PE Apoptosis Detection Kit Catalog Number KA0716 100 assays Version: 02 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 General Information...
Electronic Supplementary Information
 Electronic Supplementary Material (ESI) for Chemical Communications. This journal is The Royal Society of Chemistry 2015 Electronic Supplementary Information Enzymatic Synthesis and Post-Functionalization
Electronic Supplementary Material (ESI) for Chemical Communications. This journal is The Royal Society of Chemistry 2015 Electronic Supplementary Information Enzymatic Synthesis and Post-Functionalization
Kit for assay of thioredoxin
 FkTRX-02-V2 Kit for assay of thioredoxin The thioredoxin system is the major protein disulfide reductase in cells and comprises thioredoxin, thioredoxin reductase and NADPH (1). Thioredoxin systems are
FkTRX-02-V2 Kit for assay of thioredoxin The thioredoxin system is the major protein disulfide reductase in cells and comprises thioredoxin, thioredoxin reductase and NADPH (1). Thioredoxin systems are
CytoPainter LysoGreen Indicator Reagent
 ab176826 CytoPainter LysoGreen Indicator Reagent Instructions for Use For staining lysosomes in live cells with our proprietary Green probe. This product is for research use only and is not intended for
ab176826 CytoPainter LysoGreen Indicator Reagent Instructions for Use For staining lysosomes in live cells with our proprietary Green probe. This product is for research use only and is not intended for
Supplementary Information: Liquid-liquid phase coexistence in lipid membranes observed by natural abundance 1 H 13 C solid-state NMR
 Electronic Supplementary Material (ESI) for Physical Chemistry Chemical Physics. This journal is the wner Societies 28 Supplementary Information: Liquid-liquid phase coexistence in lipid membranes observed
Electronic Supplementary Material (ESI) for Physical Chemistry Chemical Physics. This journal is the wner Societies 28 Supplementary Information: Liquid-liquid phase coexistence in lipid membranes observed
2,6,9-Triazabicyclo[3.3.1]nonanes as overlooked. amino-modification products by acrolein
![2,6,9-Triazabicyclo[3.3.1]nonanes as overlooked. amino-modification products by acrolein](/thumbs/86/94743397.jpg "2,6,9-Triazabicyclo[3.3.1]nonanes as overlooked. amino-modification products by acrolein") Supplementary Information 2,6,9-Triazabicyclo[3.3.1]nonanes as overlooked amino-modification products by acrolein Ayumi Tsutsui and Katsunori Tanaka* Biofunctional Synthetic Chemistry Laboratory, RIKEN
Supplementary Information 2,6,9-Triazabicyclo[3.3.1]nonanes as overlooked amino-modification products by acrolein Ayumi Tsutsui and Katsunori Tanaka* Biofunctional Synthetic Chemistry Laboratory, RIKEN
Chapter 6: A Tour of the Cell. 1. Studying Cells 2. Intracellular Structures 3. The Cytoskeleton 4. Extracellular Structures
 Chapter 6: A Tour of the Cell 1. Studying Cells 2. Intracellular Structures 3. The Cytoskeleton 4. Extracellular Structures 1. Studying Cells Concepts of Microscopy MAGNIFICATION factor by which the image
Chapter 6: A Tour of the Cell 1. Studying Cells 2. Intracellular Structures 3. The Cytoskeleton 4. Extracellular Structures 1. Studying Cells Concepts of Microscopy MAGNIFICATION factor by which the image
1. Studying Cells. Concepts of Microscopy 11/7/2016. Chapter 6: A Tour of the Cell
 Electron microscope Light microscope Unaided eye 11/7/2016 Chapter 6: A Tour of the Cell 1. Studying Cells 2. Intracellular Structures 3. The Cytoskeleton 4. Extracellular Structures 1. Studying Cells
Electron microscope Light microscope Unaided eye 11/7/2016 Chapter 6: A Tour of the Cell 1. Studying Cells 2. Intracellular Structures 3. The Cytoskeleton 4. Extracellular Structures 1. Studying Cells
PRODUCT INFORMATION & MANUAL
 PRODUCT INFORMATION & MANUAL Mitochondrial Extraction Kit NBP2-29448 Research use only. Not for diagnostic or therapeutic procedures www.novusbio.com P: 303.760.1950 P: 888.506.6887 F: 303.730.1966 technical@novusbio.com
PRODUCT INFORMATION & MANUAL Mitochondrial Extraction Kit NBP2-29448 Research use only. Not for diagnostic or therapeutic procedures www.novusbio.com P: 303.760.1950 P: 888.506.6887 F: 303.730.1966 technical@novusbio.com
BIOL 158: BIOLOGICAL CHEMISTRY II
 BIOL 158: BIOLOGICAL CHEMISTRY II Lecture 1: Membranes Lecturer: Christopher Larbie, PhD Introduction Introduction Cells and Organelles have membranes Membranes contain lipids, proteins and polysaccharides
BIOL 158: BIOLOGICAL CHEMISTRY II Lecture 1: Membranes Lecturer: Christopher Larbie, PhD Introduction Introduction Cells and Organelles have membranes Membranes contain lipids, proteins and polysaccharides
High resolution structural evidence suggests the Sarcoplasmic Reticulum forms microdomains with Acidic Stores (lyososomes) in the heart.
 in the heart.") High resolution structural evidence suggests the Sarcoplasmic Reticulum forms microdomains with Acidic Stores (lyososomes) in the heart. Daniel Aston, Rebecca A. Capel, Kerrie L. Ford, Helen C. Christian,
High resolution structural evidence suggests the Sarcoplasmic Reticulum forms microdomains with Acidic Stores (lyososomes) in the heart. Daniel Aston, Rebecca A. Capel, Kerrie L. Ford, Helen C. Christian,
Western Immunoblotting Preparation of Samples:
 Western Immunoblotting Preparation of Samples: Total Protein Extraction from Culture Cells: Take off the medium Wash culture with 1 x PBS 1 ml hot Cell-lysis Solution into T75 flask Scrap out the cells
Western Immunoblotting Preparation of Samples: Total Protein Extraction from Culture Cells: Take off the medium Wash culture with 1 x PBS 1 ml hot Cell-lysis Solution into T75 flask Scrap out the cells
Supporting information
 Supporting information Figure legends Supplementary Table 1. Specific product ions obtained from fragmentation of lithium adducts in the positive ion mode comparing the different positional isomers of
Supporting information Figure legends Supplementary Table 1. Specific product ions obtained from fragmentation of lithium adducts in the positive ion mode comparing the different positional isomers of
Loss of protein association causes cardiolipin degradation in Barth syndrome
 SUPPLEMENTARY INFORMATION Loss of protein association causes cardiolipin degradation in Barth syndrome Yang Xu 1, Colin K.L. Phoon 2, Bob Berno 5, Kenneth D Souza 6, Esthelle Hoedt 4, Guoan Zhang 4, Thomas
SUPPLEMENTARY INFORMATION Loss of protein association causes cardiolipin degradation in Barth syndrome Yang Xu 1, Colin K.L. Phoon 2, Bob Berno 5, Kenneth D Souza 6, Esthelle Hoedt 4, Guoan Zhang 4, Thomas
Sulfate Radical-Mediated Degradation of Sulfadiazine by CuFeO 2 Rhombohedral Crystal-Catalyzed Peroxymonosulfate: Synergistic Effects and Mechanisms
 Supporting Information for Sulfate Radical-Mediated Degradation of Sulfadiazine by CuFeO 2 Rhombohedral Crystal-Catalyzed Peroxymonosulfate: Synergistic Effects and Mechanisms Submitted by Yong Feng, Deli
Supporting Information for Sulfate Radical-Mediated Degradation of Sulfadiazine by CuFeO 2 Rhombohedral Crystal-Catalyzed Peroxymonosulfate: Synergistic Effects and Mechanisms Submitted by Yong Feng, Deli
Supplementary Figure 1. Overview of steps in the construction of photosynthetic protocellular systems
 Supplementary Figure 1 Overview of steps in the construction of photosynthetic protocellular systems (a) The small unilamellar vesicles were made with phospholipids. (b) Three types of small proteoliposomes
Supplementary Figure 1 Overview of steps in the construction of photosynthetic protocellular systems (a) The small unilamellar vesicles were made with phospholipids. (b) Three types of small proteoliposomes
For the quantitative measurement of ATP Synthase Specific activity in samples from Human, Rat and Cow
 ab109716 ATP Synthase Specific Activity Microplate Assay Kit Instructions for Use For the quantitative measurement of ATP Synthase Specific activity in samples from Human, Rat and Cow This product is for
ab109716 ATP Synthase Specific Activity Microplate Assay Kit Instructions for Use For the quantitative measurement of ATP Synthase Specific activity in samples from Human, Rat and Cow This product is for
Supplementary Figure S1. Venn diagram analysis of mrna microarray data and mirna target analysis. (a) Western blot analysis of T lymphoblasts (CLS)
 Western blot analysis of T lymphoblasts (CLS)") Supplementary Figure S1. Venn diagram analysis of mrna microarray data and mirna target analysis. (a) Western blot analysis of T lymphoblasts (CLS) and their exosomes (EXO) in resting (REST) and activated
Supplementary Figure S1. Venn diagram analysis of mrna microarray data and mirna target analysis. (a) Western blot analysis of T lymphoblasts (CLS) and their exosomes (EXO) in resting (REST) and activated
Fluorescent Carbon Dots as Off-On Nanosensor for Ascorbic Acid
 Electronic Supplementary Material (ESI) for RSC Advances. This journal is The Royal Society of Chemistry 2014 Fluorescent Carbon Dots as Off-On Nanosensor for Ascorbic Acid Jun Gong, Xin Lu, Xueqin An*
Electronic Supplementary Material (ESI) for RSC Advances. This journal is The Royal Society of Chemistry 2014 Fluorescent Carbon Dots as Off-On Nanosensor for Ascorbic Acid Jun Gong, Xin Lu, Xueqin An*
THE ROLE OF ALTERED CALCIUM AND mtor SIGNALING IN THE PATHOGENESIS OF CYSTINOSIS
 Research Foundation, 18 month progress report THE ROLE OF ALTERED CALCIUM AND mtor SIGNALING IN THE PATHOGENESIS OF CYSTINOSIS Ekaterina Ivanova, doctoral student Elena Levtchenko, MD, PhD, PI Antonella
Research Foundation, 18 month progress report THE ROLE OF ALTERED CALCIUM AND mtor SIGNALING IN THE PATHOGENESIS OF CYSTINOSIS Ekaterina Ivanova, doctoral student Elena Levtchenko, MD, PhD, PI Antonella
Item Catalog Number Manufacturer 1,4-Dithioerythritol (1 g) D9680 Sigma-Aldrich
 D9680 Sigma-Aldrich") SOP: Nuclei isolation from fresh mouse tissues and DNaseI treatment Date modified: 01/12/2011 Modified by: E. Giste/ T. Canfield (UW) The following protocol describes the isolation of nuclei and subsequent
SOP: Nuclei isolation from fresh mouse tissues and DNaseI treatment Date modified: 01/12/2011 Modified by: E. Giste/ T. Canfield (UW) The following protocol describes the isolation of nuclei and subsequent
MagCapture Exosome Isolation Kit PS Q&A
 MagCapture Exosome Isolation Kit PS Q&A Specifications and performance P.1 Comparison of the conventional method P.2 Operation methods and composition P.4 Amount of starting sample P.5 Analysis after exosomes
MagCapture Exosome Isolation Kit PS Q&A Specifications and performance P.1 Comparison of the conventional method P.2 Operation methods and composition P.4 Amount of starting sample P.5 Analysis after exosomes
Chapter 2 Transport Systems
 Chapter 2 Transport Systems The plasma membrane is a selectively permeable barrier between the cell and the extracellular environment. It permeability properties ensure that essential molecules such as
Chapter 2 Transport Systems The plasma membrane is a selectively permeable barrier between the cell and the extracellular environment. It permeability properties ensure that essential molecules such as
Supplementary Information
 Supplementary Information Supplementary Figure 1. CD4 + T cell activation and lack of apoptosis after crosslinking with anti-cd3 + anti-cd28 + anti-cd160. (a) Flow cytometry of anti-cd160 (5D.10A11) binding
Supplementary Information Supplementary Figure 1. CD4 + T cell activation and lack of apoptosis after crosslinking with anti-cd3 + anti-cd28 + anti-cd160. (a) Flow cytometry of anti-cd160 (5D.10A11) binding
Trypsin Mass Spectrometry Grade
 058PR-03 G-Biosciences 1-800-628-7730 1-314-991-6034 technical@gbiosciences.com A Geno Technology, Inc. (USA) brand name Trypsin Mass Spectrometry Grade A Chemically Modified, TPCK treated, Affinity Purified
058PR-03 G-Biosciences 1-800-628-7730 1-314-991-6034 technical@gbiosciences.com A Geno Technology, Inc. (USA) brand name Trypsin Mass Spectrometry Grade A Chemically Modified, TPCK treated, Affinity Purified
PMT. Contains ribosomes attached to the endoplasmic reticulum. Genetic material consists of linear chromosomes. Diameter of the cell is 1 µm
 1. (a) Complete each box in the table, which compares a prokaryotic and a eukaryotic cell, with a tick if the statement is correct or a cross if it is incorrect. Prokaryotic cell Eukaryotic cell Contains
1. (a) Complete each box in the table, which compares a prokaryotic and a eukaryotic cell, with a tick if the statement is correct or a cross if it is incorrect. Prokaryotic cell Eukaryotic cell Contains
Handheld Fluorometers
 s Portable and light-weight, our handheld fluorometers provide exceptional sensitivity, rapid-reading, and reliability at a much lower cost than other portable fluorometers on the market. These convenient
s Portable and light-weight, our handheld fluorometers provide exceptional sensitivity, rapid-reading, and reliability at a much lower cost than other portable fluorometers on the market. These convenient
Manual. Precision Red Advanced Protein Assay Reagent. Cat. # ADV02. cytoskeleton.com. Cytoskeleton, Inc.
 The Protein Experts Manual Cytoskeleton, Inc. V. 6.0 Precision Red Advanced Protein Assay Reagent Cat. # ADV02 cytoskeleton.com Phone: (303) 322.2254 Fax: (303) 322.2257 Customer Service: cserve@cytoskeleton.com
The Protein Experts Manual Cytoskeleton, Inc. V. 6.0 Precision Red Advanced Protein Assay Reagent Cat. # ADV02 cytoskeleton.com Phone: (303) 322.2254 Fax: (303) 322.2257 Customer Service: cserve@cytoskeleton.com
HEPARIN SODIUM. Heparinum natricum
 Pharmeuropa 25.1 1 Reference: PA/PH/Exp. 6/T (12) 37 ANP NOTE ON THE MONOGRAPH Definition. It is proposed to restrict the scope to heparin material of porcine origin since some of the latest requirements
Pharmeuropa 25.1 1 Reference: PA/PH/Exp. 6/T (12) 37 ANP NOTE ON THE MONOGRAPH Definition. It is proposed to restrict the scope to heparin material of porcine origin since some of the latest requirements
Supplementary Material
 Supplementary Material Materials and methods Enzyme assay The enzymatic activity of -glucosidase toward salicin was measured with the Miller method (Miller, 1959) using glucose as the standard. A total
Supplementary Material Materials and methods Enzyme assay The enzymatic activity of -glucosidase toward salicin was measured with the Miller method (Miller, 1959) using glucose as the standard. A total
Structural Characterization of Prion-like Conformational Changes of the Neuronal Isoform of Aplysia CPEB
 Structural Characterization of Prion-like Conformational Changes of the Neuronal Isoform of Aplysia CPEB Bindu L. Raveendra, 1,5 Ansgar B. Siemer, 2,6 Sathyanarayanan V. Puthanveettil, 1,3,7 Wayne A. Hendrickson,
Structural Characterization of Prion-like Conformational Changes of the Neuronal Isoform of Aplysia CPEB Bindu L. Raveendra, 1,5 Ansgar B. Siemer, 2,6 Sathyanarayanan V. Puthanveettil, 1,3,7 Wayne A. Hendrickson,
bio-mof-1 DMASM Wavenumber (cm -1 ) Supplementary Figure S1 FTIR spectra of bio-mof-1, DMASMI, and bio-mof-1 DMASM.
 Supplementary Figure S1 FTIR spectra of bio-mof-1, DMASMI, and bio-mof-1 DMASM.") bio-mof-1 Transmittance bio-mof-1 DMASM DMASMI 2000 1500 1000 500 Wavenumber (cm -1 ) Supplementary Figure S1 FTIR spectra of bio-mof-1, DMASMI, and bio-mof-1 DMASM. Intensity (a.u.) bio-mof-1 DMASM as
bio-mof-1 Transmittance bio-mof-1 DMASM DMASMI 2000 1500 1000 500 Wavenumber (cm -1 ) Supplementary Figure S1 FTIR spectra of bio-mof-1, DMASMI, and bio-mof-1 DMASM. Intensity (a.u.) bio-mof-1 DMASM as
Green Cathepsin B Kit. For Research Use Only
 Green Cathepsin B Kit For Research Use Only ICT9151-25 Tests Component Storage Conditions Quantity 1 vial Rhodamine 110-(RR) 2-20 o C 25 tests 1 vial Hoechst 33342 (200 µg) 2-8 o C 1 ml 1 bottle 10x Cellular
Green Cathepsin B Kit For Research Use Only ICT9151-25 Tests Component Storage Conditions Quantity 1 vial Rhodamine 110-(RR) 2-20 o C 25 tests 1 vial Hoechst 33342 (200 µg) 2-8 o C 1 ml 1 bottle 10x Cellular
Supplementary Table 1. Properties of lysates of E. coli strains expressing CcLpxI point mutants
 Supplementary Table 1. Properties of lysates of E. coli strains expressing CcLpxI point mutants Species UDP-2,3- diacylglucosamine hydrolase specific activity (nmol min -1 mg -1 ) Fold vectorcontrol specific
Supplementary Table 1. Properties of lysates of E. coli strains expressing CcLpxI point mutants Species UDP-2,3- diacylglucosamine hydrolase specific activity (nmol min -1 mg -1 ) Fold vectorcontrol specific