Published in: Neuropsychologia. DOI: /j.neuropsychologia Document Version Peer reviewed version
|
|
|
- Molly Arnold
- 6 years ago
- Views:
Transcription


1 Do I know you? Examining face and object memory in frontotemporal dementia Kumfor, F., Hutchings, R., Irish, M., Hodges, J. R., Rhodes, G., Palermo, R., & Piguet, O. (2015). Do I know you? Examining face and object memory in frontotemporal dementia. Neuropsychologia, 71, DOI: /j.neuropsychologia Published in: Neuropsychologia DOI: /j.neuropsychologia Document Version Peer reviewed version Link to publication in the UWA Research Repository Rights statement 2015, Elsevier. Licensed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International General rights Copyright owners retain the copyright for their material stored in the UWA Research Repository. The University grants no end-user rights beyond those which are provided by the Australian Copyright Act Users may make use of the material in the Repository providing due attribution is given and the use is in accordance with the Copyright Act Take down policy If you believe this document infringes copyright, raise a complaint by contacting repository-lib@uwa.edu.au. The document will be immediately withdrawn from public access while the complaint is being investigated. Download date: 26. Jun. 2017
2 Title: Do I know you? Examining face and object memory in frontotemporal dementia Authors: Kumfor, F. 1,2,3, Hutchings, R. 1, Irish, M. 1,3,4, Hodges, JR. 1,2,3, Rhodes, G. 3,5, Palermo, R. 3,5 & Piguet, O. 1,2,3 Affiliations: 1. Neuroscience Research Australia, Sydney, Australia 2. The School of Medical Sciences, the University of New South Wales, Sydney, Australia 3. ARC Centre of Excellence in Cognition and its Disorders 4. School of Psychology, the University of New South Wales, Sydney, Australia 5. School of Psychology, University of Western Australia, Perth, Australia Corresponding author: Dr Fiona Kumfor Neuroscience Research Australia PO Box 1165, Randwick, Sydney, NSW, 2031 Australia Ph: Fax: f.kumfor@neura.edu.au Word Count: 5999 words Tables: 2 Figures: 6 Target journal: Neuropsychologia 1
3 Abstract The ability to perceive, learn and recognise faces is a complex ability, which is key to successful social interactions. This ability is proposed to be coordinated by neural regions in the occipital and temporal lobes, specialised for face perception and memory. While previous studies have suggested that memory for faces is compromised in some dementia syndromes, it remains unclear whether this simply reflects more generalised memory deficits. Here, we examined basic face perception (Identity-Matching), face recognition (Cambridge Face Memory Task) and object recognition (Cambridge Car Memory Task) in 11 semantic dementia (SD) patients (8 left-lateralised, 3 right-lateralised) and 13 behaviouralvariant frontotemporal dementia (bvftd) patients, compared with 11 controls. On the Identity-Matching task, bvftd were impaired compared to controls, with a similar trend observed in the SD group. Importantly, both bvftd and SD also demonstrated impaired face recognition. In contrast, only bvftd showed impaired object recognition, with SD performing within normal limits on this task. Voxel-based morphometry analyses revealed that Identity-Matching and face recognition were associated with partly dissociable regions including the fusiform cortex and anterior temporal lobe. Object-memory was associated with thalamic integrity in the bvftd group only. These results reveal that face perception and face memory deficits are common in bvftd and SD, and have been previously underestimated. These deficits are due to neurodegeneration of key regions within the core and extended face processing system, providing convergent evidence of the neural regions supporting face perception. From a clinical perspective, impaired ability to recognise faces is common in bvftd and SD and therefore strategies to improve face perception and memory may be beneficial for these patients. 2
4 Keywords: face perception, frontotemporal dementia, memory, fusiform cortex, temporal lobe 3
5 1 Introduction The ability to perceive and recognise faces is arguably one of the most highly specialised skills in humans. It enables us to recognise friends or family members, as well as acquaintances or even famous individuals that we have not met before in real life. This capacity occurs with ease and requires minimal cognitive effort. In addition, the ability to learn and remember new faces appears to be virtually unlimited. Despite this ostensible simplicity, the cognitive and neural mechanisms supporting face perception and recognition are complex. Stages involved in face perception include early perception of facial features supported by the inferior occipital gyri (Haxby et al. 2000). This is followed by extraction of invariant facial features (i.e., facial identity), while simultaneously disregarding any changeable facial information (e.g., emotional expression, age, lip movements during speech) (Bruce & Young 1986; Calder & Young 2005; Haxby, et al. 2000). This stage is supported by the inferotemporal cortex, including the lateral fusiform face area. Other neural regions including the superior temporal sulcus, purportedly involved in processing changeable face features, have also been implicated (for reviews see Calder & Young 2005; Haxby, et al. 2000). Retrieval of biographical/semantic information related to the face then involves the anterior temporal lobe (for review see Haxby, et al. 2000). Finally, successful face recognition, as well as (re-)encoding of facial information and its related contextual information may recruit additional memory systems including the medial temporal lobe and prefrontal regions (e.g., Bernstein et al. 2002). Breakdown at any of these cognitive or neural stages will compromise face perception and memory for faces. 4
6 Impaired recognition of familiar or famous faces (i.e., prosopagnosia) has received considerable interest from a cognitive and neural perspective, with a large number of studies reporting impairment, in several neurodegenerative disorders (e.g., Gefen et al. 2013; Josephs et al. 2008; Mendez et al. 1992; Snowden et al. 2004). The extent that impaired face perception and recognition of novel faces is present in dementia, however, is less clear. Frontotemporal dementia (FTD) is an umbrella term, referring to a group of progressive, neurodegenerative brain disorders that affect the frontal and temporal lobes (Gorno- Tempini et al. 2011; Rascovsky et al. 2011). Three clinical subtypes are typically recognised: behavioural-variant frontotemporal dementia (bvftd), semantic dementia (SD) and progressive nonfluent aphasia (PNFA). This study focuses on the bvftd and SD subtypes. Individuals with bvftd present with a marked disturbance of behaviour and personality, which is associated with degeneration of the orbitofrontal and medial prefrontal cortices (Kipps et al. 2009; Seeley et al. 2008). Episodic memory performance is often impaired, and in many cases individuals with bvftd are as impaired as disease-matched individuals with Alzheimer s disease on standard neuropsychological tests of episodic memory (Hornberger & Piguet 2012; Hornberger et al. 2010; Irish et al. 2014b). In contrast, individuals with SD present with a multimodal loss of general conceptual/ semantic knowledge, which is usually associated with atrophy in the left anterior temporal lobe (Chan et al. 2001; Galton et al. 2001; Mion et al. 2010). Despite this profound loss of 5
7 semantic memory, non-verbal episodic memory is usually well-preserved (Hodges et al. 1992). Of relevance here, a proportion of patients (~ 30%) present with right- rather than left-lateralised temporal lobe degeneration, referred to as right SD (Chan et al. 2009). Clinically, right SD is often characterised by a change in behaviour and personality, with reports of prosopagnosia also common (Irish et al. 2013; Josephs et al. 2009; Kamminga et al. in press; Snowden, et al. 2004; Thompson et al. 2004). Although few studies have systematically examined episodic memory performance in right SD, some evidence suggests that non-verbal episodic memory and spatial navigation is compromised in this phenotype (Chan, et al. 2009; Irish, et al. 2013). FTD provides a unique model to study the neurobiological basis of face perception and memory. While a large number of studies have reported impaired memory for famous faces in FTD, with divergent profiles in left SD compared to right SD (e.g., Gefen, et al. 2013; Josephs, et al. 2008; Snowden, et al. 2004; Thompson, et al. 2004), far fewer studies have examined memory for novel faces, with mixed findings. Omar et al. (2010) used the Benton Facial Recognition Test (Benton 1994), which assesses face perception by asking participants to match a target face to those in a simultaneously presented set. Patients with SD (12 left SD; 1 right SD) and bvftd (n = 19) all performed within normal limits. Many healthy individuals, however, score in the normal range on the Benton Facial Recognition Test when only the eyebrows are visible, suggesting a piecemeal face matching approach is sufficient to complete this task successfully (Duchaine & Weidenfeld 2003). Indirect evidence from emotion processing tasks using caricatures further suggests that face perception is 6
8 abnormal in bvftd (Kumfor et al. 2011), although this has not been studied systematically. Thus, the integrity of basic face perception in SD and bvftd is unknown. One of the earliest studies to examine recognition memory for novel faces in FTD used the Warrington Recognition Memory Test for Faces (Warrington 1984), an old/new test of face memory (Simons et al. 2001). Consistent with clinical reports, only the group with predominant right-lateralised SD (n = 4) were impaired, while the left-lateralised SD group (n = 4) performed within normal limits. The SD group with relatively bilateral atrophy (n = 5) was more impaired than the left-lateralised patients, but less impaired than the rightlateralised patients (Simons, et al. 2001). In contrast, no significant differences were observed between left and right SD on standard neuropsychological tests of episodic memory (e.g., Rey Complex Figure, Warrington Recognition Memory Test for Words). These results led the authors to propose that the right temporal lobe is specialised for memory for faces (Simons, et al. 2001) and therefore, apart from right SD, this function should be relatively intact in other FTD phenotypes. Importantly, however, the stimuli in the Warrington Recognition Memory Test for Faces contain substantial non-face information (e.g., hairstyles, clothing) that can be used to support recognition memory. Indeed, healthy individuals are able to score within the normal range on this test, even when all facial information is occluded (Duchaine & Weidenfeld 2003). Moreover, other evidence suggests that face memory, at least for familiar faces, is compromised in the other FTD subtypes. Individuals with left SD tend to be impaired on tests of memory for famous faces, even when only recognition and not naming is required (Gefen, et al. 2013). Thus, face recognition deficits in left SD and bvftd may have been previously underestimated. Further, 7
9 no studies to date have directly compared faces recognition with recognition of visual nonface stimuli of similar complexity. It is therefore unclear whether any observed face memory deficits in FTD are due to the degradation of face perception, a specific recognition memory deficit, or a combination of both. Systematic investigation of the neural correlates of face memory in FTD is also currently lacking. Existing studies investigating famous face recognition, and clinical reports of prosopagnosia in SD, highlighted the importance of the right anterior temporal lobe as a key region supporting recognition of known faces (Gefen, et al. 2013; Hsieh et al. 2011; Josephs, et al. 2008). To our knowledge, however, no studies have directly investigated the neural correlates of basic face identity perception and memory for novel faces in this population. Further, the extent that common or divergent neural correlates contribute to face memory disturbance across FTD subtypes is unknown. Thus, the current study had two broad aims: (i) To investigate face perception and face memory in bvftd and SD using more sensitive tasks than used in previous studies, by employing both a face memory task and a well-matched object memory task and; (ii) to identify the neural correlates underlying face perception, face memory and object memory in bvftd and SD. Given the degradation to regions within the frontal and temporal lobes in bvftd and SD, we hypothesised that face perception and face memory deficits would be observed in both FTD subtypes. In addition, we predicted bvftd would show impaired object memory performance, in line with previous reports of episodic memory deficits in this syndrome. 8
10 2 Methods 2.1 Participants Thirteen bvftd and 11 SD patients (8 SD with predominantly left-sided atrophy, 3 SD with predominantly right-sided atrophy) were consecutively recruited from FRONTIER, the frontotemporal dementia research group in Sydney, Australia and compared with 11 healthy age- and education-matched controls. All patients were assessed by an experienced behavioural neurologist, underwent comprehensive neuropsychological assessment (Table 1) and neuroimaging, and a reliable informant was interviewed. Based on this information, patients were classified according to current consensus diagnostic criteria for bvftd or SD (Gorno-Tempini, et al. 2011; Rascovsky, et al. 2011). In line with previous studies, individuals with SD were classified by the clinical team as either left- or right-lateralised based on their coronal MRI brain scan of the temporal lobes and their clinical presentation (e.g., Kamminga, et al. in press). The proportion of left and right SD cases in the current study is consistent with previous reports in the literature (e.g., Chan, et al. 2009). Control participants were recruited from the Neuroscience Research Australia Volunteer database and local community clubs. For all participants, exclusion criteria were: history of significant neurological or psychological disorder, use of centrally-acting medication and limited proficiency in English. In addition, all controls were required to score > 88/100 on the Addenbrooke s Cognitive Examination-Revised (ACE-R), or the recently updated Addenbrooke s Cognitive Examination-III (ACE-III), which are general screening measures of cognition (Hsieh et al. 9
11 2013b; Mioshi et al. 2006). All control participants also scored 0 on the Clinical Dementia Rating sum of boxes (Morris 1993). All participants or their Person Responsible provided informed consent according to the Declaration of Helsinki. The South Eastern Sydney Local Health District and the University of New South Wales ethics committees approved the study. 2.2 Background demographics and neuropsychological assessment Disease duration was measured in months from the first onset of symptoms. The Frontotemporal Dementia Functional Rating Scale (FRS) was used to determine functional severity in bvftd and SD (Mioshi et al. 2010). In addition, all participants underwent a neuropsychological assessment to measure attention and working memory (Digit Span (Wechsler 1997), Trail Making Test (Tombaugh 2004)), visuoconstruction and non-verbal episodic memory (Rey Complex Figure Test (Rey 1941)) and language (SYDBAT (Savage et al. 2013)). 2.3 Face Perception Aspects of basic face perception were assessed using two short non-verbal tasks, which are depicted in Figure 1 (Kumfor et al. 2014; Miller et al. 2012). In brief, on the Face-Perception task, individuals are presented with 40 face pairs, one pair at a time, showing a neutral expression, and are asked to determine whether the two images are identical or different. 10
12 For the Identity-Matching task, participants are shown pairs of faces simultaneously, which differ in emotional expression, and are asked to determine whether the two pictures are of the same person (i.e., same identity). The task consists of 42 trials. Participants are verbally asked Is this the same picture? or Is this the same person?, depending on the task, and required to respond verbally either yes or no. For both tasks, faces were selected from the NimStim stimulus set ( Faces were cropped so that all non-facial features including hair were removed and all images are presented in greyscale. Responding was untimed and no feedback was provided. [INSERT FIGURE 1 ABOUT HERE] 2.4 Cambridge Face Memory Test (CFMT) (Duchaine & Nakayama 2006) Memory for faces was assessed using the upright version of the Cambridge Face Memory Test (CFMT), shown in Figure 2 (Bowles et al. 2009; Duchaine & Nakayama 2006). During the task, participants learn to recognise six individuals, presented in grey scale, with hair and other identifying facial features (e.g., facial blemishes) removed. On each trial, the participant is required to identify the target face from two similar distractor faces. Several of the distractor faces are repeated throughout the task, so the participant cannot make a judgement based on familiarity alone. In Section 1, the target face is the identical image as shown during encoding (same images, 18 trials). In Section 2, faces are shown from different viewpoints or under different lighting conditions than seen during encoding (novel images, 30 trials). Finally, in Section 3, participants are again shown the target face from different viewpoints or lighting conditions, with additional Gaussian noise (novel image + noise, 24 trials). On each trial, participants are asked to point to the target face. Responding is untimed and no feedback is provided. Following each section, participants review the 6 11
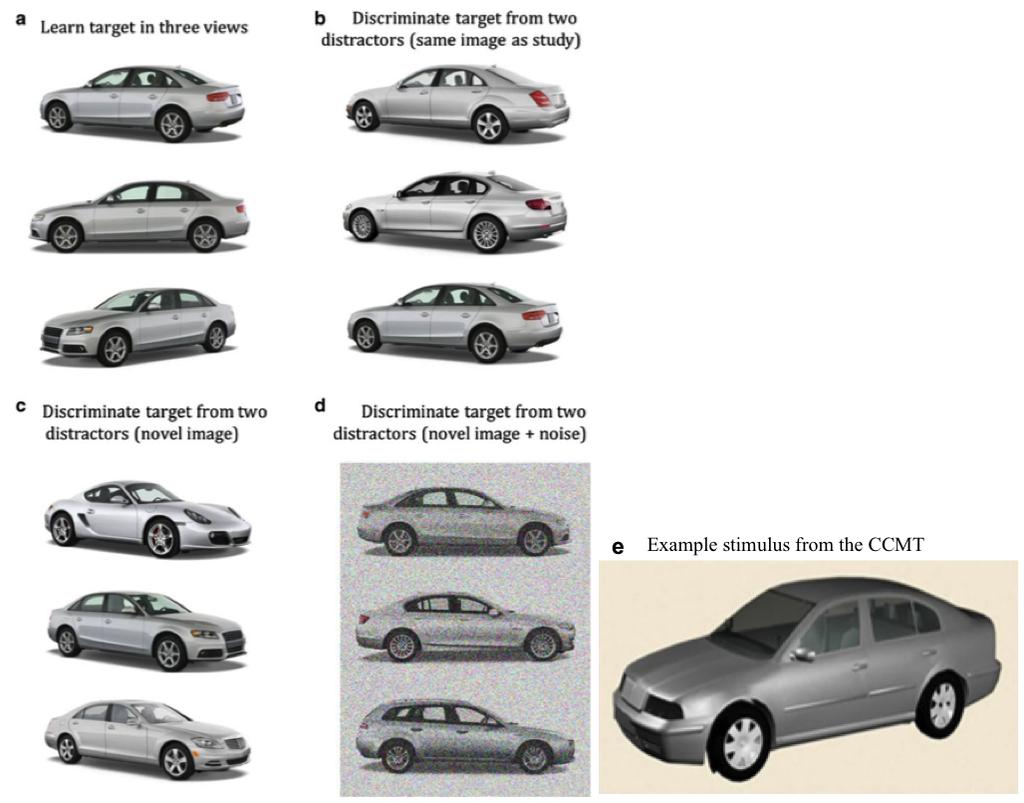
13 target images for 20 seconds. A percent correct overall score for the 72 trials was calculated. Prior to testing, participants completed 3 practice trials and participants only progressed to the testing session after achieving 100% correct on the practice trials. [INSERT FIGURE 2 ABOUT HERE] 2.5 Cambridge Car Memory Test (CCMT) (Dennett et al. 2012) To assess memory for objects, we used the Cambridge Car Memory Test (CCMT), which was designed to be analogous to the CFMT, shown in Figure 3 (Dennett, et al. 2012). Similar to the CFMT, participants learn to recognise 6 target items (cars) and then recognition memory is assessed across 3 sections: Section 1 (same images, 18 trials); Section 2 (novel images, 30 trials); Sections 3 (novel images + noise, 24 trials). On each trial, participants are asked to point to the target car. The task is untimed and no feedback is provided. A percent correct overall score (/72) was calculated for subsequent analyses. [INSERT FIGURE 3 ABOUT HERE] 2.6 Behavioural Statistical Analyses Data were analysed using IBM SPSS Statistics version Univariate analyses of variance (ANOVA) were conducted to examine continuous demographic and general cognitive performance variables. Chi-square analyses were used to analyse categorical variables. For the experimental variables (Face-Perception, Identity-Matching, CFMT and CCMT) univariate ANOVAs were conducted. Prior to the analyses, the suitability of variables for parametric analyses was determined using Kolmogorov-Smirnov tests. The Face-Perception and Identity-Matching tasks, did not meet the assumption of homogeneity of variance, based on Levene s test. Therefore we reanalysed the data using the non-parametric equivalent. The 12
14 pattern of results remained the same and for this reason we have reported the parametric analyses throughout, to maintain consistency. Follow-up post hoc analyses with Sidak correction for multiple comparisons were employed to investigate between group differences. For the analyses investigating the experimental tasks of interest (Face- Perception, Identity-Matching, CFMT and CCMT) we first compared SD with bvftd and controls. Then, given the small number of right SD cases, we conducted the same analyses in left SD only, to investigate whether any differences in patterns of performance were observed after excluding right SD cases. 2.7 Neuroimaging MRI Acquisition Participants underwent whole-brain structural magnetic resonance imaging (MRI) with a 3 T Phillips scanner. High resolution T1-images were obtained using the following protocol: 256 x 256, 200 slices, 1mm 2 in-plane resolution, 1mm slice thickness, echo time/repetition time = 2.6/5.8 ms, flip angle = 19. Brain scans were available for 30 participants (10 bvftd, 10 SD (8 left SD, 2 right SD), 10 controls) Data preprocessing FSL voxel-based morphometry (VBM), part of the FMRIB software library package (Smith et al. 2004) was used to analyse the MRI data (Ashburner & Friston 2000; Mechelli et al. 2005; Woolrich et al. 2009). First, structural images were brain-extracted using BET and tissue segmentation was undertaken using automatic segmentation (FAST) (Zhang et al. 2001). Then, grey matter partial volume 13
15 maps were aligned to Montreal Neurological Institute standard space (MNI152) using nonlinear registration (FNIRT), which uses a b-spline representation of the registration warp field (Rueckert et al. 1999). A study-specific template was created and the native grey matter images were non-linearly re-registered. Modulation of the registered partial volume maps was carried out, by dividing them by the Jacobian of the warp field. The modulated, segmented images were smoothed with an isotropic Gaussian kernel (sigma = 3mm, fullwidth at half maximum (FWHM) = 8mm) VBM analyses A voxel-wise general linear model was applied to investigate grey matter intensity differences using permutation-based, non-parametric statistics with 5000 permutations per contrast (Nichols & Holmes 2002). In the first set of analyses, differences in grey matter integrity between patient groups and controls were investigated using t-tests (Supplementary material). Next, correlations between Identity-Matching, CFMT and CCMT performance and grey matter intensity were conducted in all participants combined. Total scores on either the Identity-Matching task, the CFMT or the CCMT were entered as covariates into separate design matrices and a positive [1] correlation analysis was performed (Supplementary Material). Finally, the same covariate analysis was performed separately for each patient group combined with controls in order to identify the neural correlates subtending Identity- Matching, Face Memory and Object Memory, distinct to each patient group. This approach increases the variance in behavioural scores, without contributing towards atrophy. 14
16 Inclusion of controls therefore increases the statistical power to detect an effect, which is particularly useful in clinical populations where large sample sizes are not available (Irish et al. 2012; Kumfor et al. 2013; Sollberger et al. 2009). As for the behavioural analyses, we then repeated the same analyses excluding the 2 right SD cases, to ensure effects were not driven by this subset of SD patients (Supplementary Material). VBM analyses were conducted across the whole brain, including the cerebellum. For all comparisons voxel-wise analyses are reported and the statistical threshold was set at p <.001 uncorrected. In addition, we applied a conservative cluster-extent threshold of 50 voxels to reduce the likelihood of false positives (Lieberman & Cunningham 2009). Anatomical locations of significant results were overlaid on the Montreal Neurological Institute (MNI) standard brain, with maximum voxel coordinates provided in stereotaxic space. Anatomic labels were determined with reference to the Harvard-Oxford probabilistic cortical and subcortical atlases. 3 Results 3.1 Participants All groups were matched for age, sex and education (all p values >.05). No significant difference in disease duration was observed between patient groups; however, on the Frontotemporal Dementia Functional Rating Scale (FRS) the bvftd group was rated as more functionally impaired than the SD group (Table 1). 15
17 Examination of the cognitive performance across groups revealed profiles consistent with the clinical features typically reported (Table 1). Both bvftd and SD patients performed worse than controls on the Addenbrooke s Cognitive Examination (ACE) measure of general cognition (both p values <.001). The SD group also performed significantly worse than the bvftd group, reflecting the high language demands of this task (p =.02). In addition, the SD group was impaired on tasks assessing naming (p <.001) and comprehension (p <.001), performing significantly worse than both bvftd and control participants. In contrast, bvftd were impaired on tasks assessing attention and working memory (Trail Making Test A (p =.008), Trail Making Test B (p =.001) Digit Span Forwards (p =.042) Digit Span Backwards (p =.020)) and also performed significantly worse on the Rey Complex Figure copy (p =.005), likely indicating reduced planning and organisation when approaching the task. Both bvftd (p <.001) and SD (p =.015) were impaired on the Rey Complex Figure percent retention. Given that non-verbal episodic memory is typically reported to be intact in left SD, we reanalysed performance on this task after removing the three right SD cases. These analyses indicated that left SD cases performed within normal limits (p =.110), suggesting that nonverbal memory is relatively preserved in left SD but is more variable in right SD, consistent with previous findings. [INSERT TABLE 1 ABOUT HERE] 3.2 Face Perception As shown in Figure 4, group performance did not significantly differ on the Face-Perception task (F(2,32) = 1.175, p =.322). In contrast, a significant main effect of Diagnosis was present on the Identity-Matching task (F(2,33) = 8.263, p =.001). Post hoc tests revealed 16
18 that the bvftd group performed significantly worse than controls on this task (p =.001) with a trend also observed in the SD group (right and left SD patients combined) (p =.055). No difference was observed between patient groups on this task (p =.441) (Figure 4). Next we conducted the same analyses, to determine whether the pattern of results remained after excluding the right SD cases. On the Face-Perception test, again no effect of Diagnosis was observed (F(2,30) = 0.997, p =.389). On the Identity-Matching task, a significant effect of Diagnosis was again present (F(2,30) = 7.686, p =.002). Post hoc analyses indicated that bvftd performed significantly worse than controls (p =.002), whereas performance in left SD was not significantly different from controls (p =.275). Once again, no difference was observed between patient groups on this task (p >.05). [INSERT FIGURE 4 ABOUT HERE] 3.3 Cambridge Face and Car Memory Test Performance As shown in Figure 5, on the CFMT a significant effect of Diagnosis was observed (F(2,34) = , p <.001) with bvftd (p <.001) and SD (p <.001) performing worse than controls. No difference was observed between patient groups (p =.805). On the CCMT, again a significant effect of Diagnosis was present (F(2,34) = 3.742, p =.035). Here, however, only the bvftd group were impaired compared to controls (p =.032), with no significant difference in performance between SD and controls (p =.255). The difference between bvftd and SD was not significant (p =.750) (Figure 5). 17
19 To investigate whether the findings were driven by the 3 right SD cases, we reanalysed the data including left SD cases only. Importantly, the significant effect of Diagnosis previously observed was again present on the CFMT (F(2,31) = , p <.001), with both bvftd (p <.001) and SD (p =.001) performing worse than controls. Results on the CCMT also showed the same pattern as the initial analyses with an overall effect of Diagnosis (F(2,31) = 4.481, p =.020). Again the bvftd group only, performed worse than controls (p =.021), with left SD demonstrating relatively intact object memory on the CCMT (p =.856). Individual patient profiles are reported in the Supplementary Material. [INSERT FIGURE 5 ABOUT HERE] 3.4 VBM Analyses Patterns of grey matter intensity changes Compared with controls, the SD group demonstrated striking changes in the anterior temporal lobe, encompassing the temporal pole, temporal fusiform gyrus, amygdala and parahippocampal gyrus which extended into the orbitofrontal cortex. Although the grey matter intensity decrease was present bilaterally, it was somewhat more extensive in the left than the right hemisphere (See Supplementary Material for figure). When we reanalysed the pattern of change in left SD cases only, we again found significant changes encompassing the left temporal pole, extending into the left orbitofrontal cortex, amygdala, hippocampus and left fusiform cortex. A smaller cluster was also identified in the right hemisphere including the right inferior temporal gyrus, and right temporal pole, confirming 18
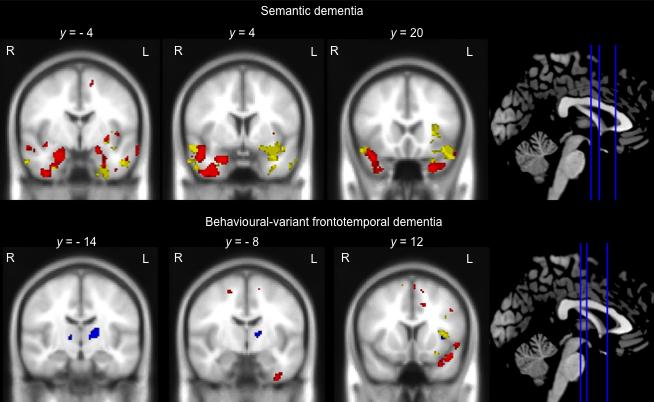
20 the disproportionate left-lateralisation of brain changes in these patients. The bvftd group showed relatively circumscribed grey matter intensity decrease compared with controls. Significant clusters were identified in the frontal pole, as well as the left orbitofrontal cortex and small regions in the left insula and temporal pole (See Supplementary Material for figures depicting patterns of atrophy in each group) Neural correlates of face processing, face memory and object memory As shown in Figure 6, in the SD group, Identity-Matching performance (shown in red) was predominantly associated with integrity of the right temporal lobe. This cluster encompassed the right anterior portion of the fusiform cortex and extended into the right inferior temporal gyrus, amygdala and temporal pole. In addition, Identity-Matching performance was associated with integrity of the left temporal pole, left inferior temporal gyrus and left parahippocampal gyrus, extending into the amygdala (Table 2). Neural correlates of face memory performance in the SD group (shown in yellow) tended to be left lateralised and were associated with integrity of the left fusiform cortex and temporal pole as well as the anterior portion of the left middle temporal gyrus. In addition, a cluster was identified in the right temporal lobe which encompassed the anterior portion of the middle temporal gyrus, extending into the right temporal pole and the right orbitofrontal cortex. No regions significantly correlated with object memory in SD (Figure 6, Table 2). When we reanalysed the data excluding the 2 right SD cases with available MRI scans, the results remained largely the same (Supplementary Material). 19
21 In contrast, in bvftd, Identity-Matching performance (shown in red) was associated with integrity of the left orbitofrontal cortex extending into the left temporal pole, which is shown in Figure 6. In addition, performance was significantly associated with integrity of the left fusiform gyrus. Memory for faces (CFMT performance) (shown in yellow) was associated with integrity of the left temporal pole, extending into the left orbitofrontal cortex, with a second region identified in the left insula. Notably, object memory (CCMT performance) (shown in blue) in this group was associated with a distinct set of brain regions including the right frontal opercular cortex extending into the insula, together with the left thalamus (Figure 6, Table 2). VBM analyses for the Face-Perception task were not conducted because both patient groups performed within normal limits. VBM analyses in all participants combined are reported in the Supplementary Material. [INSERT FIGURE 6 ABOUT HERE] 4 Discussion In this study we used novel experimental tasks, in combination with voxel-based morphometry, to investigate face perception, face memory and object memory in individuals with SD and bvftd. Our results revealed that patients with SD showed circumscribed deficits on tasks assessing face memory, even in individuals with disproportionate left-lateralised temporal atrophy. Crucially, these deficits were associated 20
22 with degradation of regions within the temporal lobes bilaterally, brain regions that are thought to play a key role in face processing. In contrast, individuals with bvftd showed deficits in face perception, as well as face and object memory. Notably, these impairments were associated with degradation of distinct regions involved in face processing (i.e., the fusiform cortex, anterior temporal lobe) and memory (i.e., the thalamus). In the following sections we consider the current findings in light of our understanding of these FTD phenotypes, and current models of face processing. SD is typically characterised as a language disorder, with prominent anomia and loss of general conceptual knowledge. Increasingly, however, it is recognised that deficits in social cognition and emotional functioning are also present in this syndrome (Hsieh et al. 2013a; Irish et al. 2014a; Kumfor, et al. 2013; Kumfor, et al. 2011; Kumfor & Piguet 2012; Rankin et al. 2009; Rosen et al. 2002). The precise mechanisms and neural substrates subtending these deficits, however, are relatively poorly understood. Our results provide new evidence that face recognition is disproportionately affected in SD. Consistent with previous studies (e.g., Simons, et al. 2001), qualitative examination of patient profiles of individuals with predominantly right-lateralised temporal atrophy suggest that these patients do indeed show worse face recognition performance than those with the typical left-lateralised profile. Importantly, using a well-matched object recognition task, we have demonstrated circumscribed face recognition deficits are present in SD, even in cases with predominantly left hemisphere damage. 21
23 Our neuroimaging analyses revealed both common and distinct brain regions subtending face perception and memory in SD. Basic face perception, as measured by the Identity- Matching task, was associated with a predominantly right-lateralised network including the fusiform cortex, inferior temporal gyrus, amygdala and temporal pole. Haxby (2000) proposed that the fusiform cortex is the key node involved in identity recognition within the core face processing system. Our results provide additional support for this model using a neuropsychological paradigm to elucidate brain-behaviour relationships. Interestingly, regions within the extended face processing system, including the anterior temporal pole, were also significantly associated with identity recognition. The anterior temporal pole is commonly implicated in aspects of biographical retrieval, and thus its association with identity perception of novel rather than familiar faces, was somewhat unexpected. Recent evidence, however, indicates that the anterior temporal lobe is also involved in aspects of face recognition. For example, two individuals who had undergone unilateral anterior temporal lobe resection, were found to experience impaired performance on a face matching task similar to the identity matching task we used here (Olson et al. 2014). The authors proposed that these deficits were due to disruption in the role of the anterior temporal lobe, including the perirhinal cortex, in high-level face processing, specifically the coding of face patches to enable identification across individuals. This proposition is in line with fmri findings demonstrating sensitivity of the ventral anterior temporal lobe for processing identity in both healthy adults and macaques (reviewed by Von Der Heide et al. 2013). In addition, recent studies have revealed that abnormal functioning of the anterior temporal lobe node of the extended face processing system is present in congenital prosopagnosia (Avidan et al. 2014; Rivolta et al. 2014). This emerging evidence, together 22
24 with the current findings, therefore provides important support for the role of the anterior temporal lobe in face perception and recognition. Memory for faces in SD was also associated with regions within the temporal lobe, although the neural correlates identified appeared more bilateral than those associated with identity matching. These regions included the left anterior fusiform gyrus, extending into the left temporal lobe as well as the left middle temporal gyrus. Atrophy of the left superior temporal sulcus/medial temporal gyrus has also been previously associated with poor face identity recognition in developmental prosopagnosia (Choi et al. 2012), indicating that this brain region contributes to abnormal face recognition across different clinical populations. In addition to these left-lateralised structures, a region within the right middle temporal gyrus, extending into the orbitofrontal cortex and right temporal pole was associated with face memory performance in the SD group. Evidence supporting the lateralisation of face memory to the right temporal lobe is mixed. Some researchers have suggested that recognition of non-verbal information, including recognition of familiar/famous faces, is right-lateralised whereas recognition of familiar names and similar verbal information appears left-lateralised (Gainotti 2013). Others, however, have proposed that recognition of familiar faces is left-lateralised, while recognition of novel faces only, is right-lateralised (Von Der Heide, et al. 2013). Our findings indicate that both the left and right temporal lobes are involved in recognition of novel faces. We were unable to investigate whether different mechanisms contribute to face memory deficits in left and right SD, given the small number of right SD cases. Nevertheless, 23
25 it is plausible that divergent neurocognitive mechanisms subtend face memory in left and right SD. Finally, it should be explicitly noted that deficits in face memory in SD were observed despite object memory remaining relatively intact. Although non-verbal memory is variable in right SD (Kamminga, et al. in press; Thompson et al. 2003), it is typically well preserved in left SD (Tan et al. 2014). The finding of intact object memory in our SD cohort therefore provides corroborative evidence that deficits in face recognition in these patients, is specific to faces, rather than just a general episodic memory deficit. The behavioural and neuroimaging findings in the bvftd group were distinct from those observed in SD. Individuals with bvftd showed reduced ability to accurately discriminate facial identity across pairs of faces. Crucially, this impairment in identity recognition was correlated with integrity of the anterior portion of the left fusiform gyrus, a region which is known to be critical for face perception, specifically the perception of unique facial identity (Haxby, et al. 2000). In addition to the fusiform gyrus, reduced identity recognition correlated with the left orbitofrontal cortex extending into the left temporal pole, as well as a region within the anterior cingulate. These regions have been previously implicated in bvftd with regards to social cognition more generally (Ibañez & Manes 2012). Our findings suggest that like other aspects of social cognition, including emotion processing and theory of mind, face processing is also affected in bvftd, and this appears to be due to degradation to key nodes within the Social Context Network (Couto et al. 2013; Ibañez & Manes 2012) as well as core brain regions in the face processing system (Haxby, et al. 2000). 24
26 Unlike in SD, individuals with bvftd were impaired on both the face and object memory tasks. Interestingly, however, divergent neural correlates were associated with performance on these tasks. Similar to SD, performance on the face memory task was associated with integrity of the left anterior temporal pole in bvftd, confirming the role of this region in face recognition. Conversely, performance on the object memory task was associated with integrity of the left thalamus. The thalamus is a key relay station with white matter projections to the prefrontal, temporal, premotor, primary motor, sensory, occipital and posterior parietal cortices (Behrens et al. 2003). Notably, individuals who have suffered a thalamic stroke frequently present with an amnesic syndrome, together with executive dysfunction and behavioural change (reviewed by Carlesimo et al. 2011), a profile which is remarkably consistent with that seen in bvftd. The role of the thalamus in supporting memory in bvftd has only recently been recognised, with disproportionate atrophy in this region evident on both neuroimaging and at post-mortem in bvftd compared to Alzheimer s disease and healthy controls (Hornberger et al. 2012). Moreover, bvftd patients with memory impairment show greater anterior thalamic atrophy than those with less memory impairment (Hornberger, et al. 2012). Additional examination of the potential role of the thalamus in cognition in bvftd is certainly warranted. Importantly, our findings demonstrate that non-verbal object memory is impaired in bvftd and this is due to degradation of the thalamus, a key node within the episodic memory network. The identification of divergent neural correlates underpinning face and object memory, together with the impaired Identity-Matching performance in bvftd, provides important evidence that face perception, rather than more general cognitive or memory deficits, 25
27 contribute to the identity recognition deficits observed behaviourally in this clinical syndrome. Studies examining face memory have reported variable results to date, with several studies suggesting that this ability is relatively preserved in bvftd and left SD (e.g., Omar et al. 2010). Nevertheless, indirect evidence using caricatured facial expressions has indicated that abnormal face processing may in part contribute to the facial emotion recognition deficits in this group (Kumfor, et al. 2011). Importantly, the CFMT task employed here precludes the potential for participants to use a piecemeal approach or non-facial features to successfully perform the task (Duchaine & Nakayama 2006; Duchaine & Weidenfeld 2003). Under these conditions we have demonstrated that both individuals with bvftd and SD show deficits in aspects of face and identity processing, which impact on the ability to learn and recognise novel faces. Given the current findings, systematic examination of face perception ability, such as whether holistic face processing is preserved in bvftd and SD, is warranted. From a clinical perspective, the CFMT and CCMT appear to offer novel, well-validated and psychometrically sound tasks to assess face and object memory in neurodegenerative syndromes. Notably, in other clinical populations (e.g., developmental prosopagnosia), face perception ability predicts face memory performance (Huis in 't Veld et al. 2012). While we were not able to investigate this potential relationship here, due to the limited sample size, this will be an important area for future studies to address. Nevertheless, the impaired ability to recognise and process faces as demonstrated here, likely contributes to the pervasive social deficits observed in bvftd and SD, although to our knowledge this has not yet been investigated. This relationship warrants attention, as strategies to improve face 26
28 perception and face memory may help to improve the quality of life of patients and their interpersonal relationships with their carers and families. In summary, our results demonstrate that both face perception and memory is abnormal to various degrees in SD and bvftd, and is due to the degradation of key regions within the core and extended face processing system. Individuals with bvftd exhibit additional deficits in object memory that is due to degeneration of memory processing brain regions, namely the thalamus. Importantly, these results highlight that abnormalities in face perception and memory are not exclusive to right SD. Finally, while our results provide additional evidence from a neurobiological perspective supporting current models of face processing, they raise important questions about the laterality of face processing, which will be crucial for future studies to consider. 27
29 5 Figure Captions Figure 1: Example trial for the Face-Perception and Identity-Matching tasks. Note. For both examples, the correct answer is yes. Figure 2: Examples of stimuli and design of the CFMT. Note. Panel A shows study views of a target face. These are presented one at a time for 3 seconds each. Panel B shows a test panel from Section 1 (face 3 is the target) Panel C shows a test panel from Section 2 (face 1 is the target). Panel D shows a test panel from Section 3 (face 3 is the target). Reprinted from: Neuropsychologia, 44, Duchaine & Nakayama, The Cambridge Face Memory Test: Results for neurologically intact individuals and an investigation of its validity using inverted face stimuli and prosopagnosic participants, p (2006), with permission from Elsevier. Figure 3: Format and design of the CCMT. Note. Panel A shows study views of a target car. These are presented one at a time for 3 seconds each. Panel B shows a test panel from Section 1 Panel C shows a test panel from Section 2. Panel D shows a test panel from Section 3. Panel E shows an example of the type of actual stimuli used. Reprinted from: Behaviour Research Methods, 44, Dennett et al., The Cambridge Car Memory Test: A task matched in format to the Cambridge Face Memory Test, with norms, reliability, sex differences, dissociations from face memory, and expertise effects. p (2011) with permission from Springer. Figure 4. Performance on the Face-Perception and Identity-Matching tasks in bvftd and SD compared with healthy controls. Note. Markers represent individual scores. Left SD cases represented with filled circles, right SD represented with open circles. Lines represent the mean and standard deviation. Missing 28
30 data for 1 bvftd case on the Identity-Matching task and 1 left SD case on the Face- Perception and Identity-Matching tasks. * p <.05 compared to controls, T p =.055 compared to controls. Figure 5. Performance on the CFMT (face memory) and CCMT (object memory) tasks in bvftd and SD compared with healthy controls. Note. Markers represent individual scores. Left SD cases represented with filled circles, right SD represented with open circles. Lines represent the mean and standard deviation. * p <.05 compared to controls. Figure 6. Neural correlates of Identity-matching (red), CFMT (face memory) (yellow) and CCMT (object memory) (blue) in semantic dementia and behavioural-variant frontotemporal dementia, combined with controls. Note. Coloured voxels indicate regions that are significant at p <.001 uncorrected for multiple comparisons. L = left, R = right. y values are MNI coordinates 29
31 6 Acknowledgements We are grateful to the research participants involved with the ForeFront research studies. This work was supported by funding to Forefront, a collaborative research group dedicated to the study of frontotemporal dementia and motor neurone disease, from the National Health and Medical Research Council (NHMRC) of Australia program grant (# ) and a cross program grant from the Australian Research Council (ARC) Centre of Excellence in Cognition and its Disorders (#CE ). MI is supported by an ARC Discovery Early Career Research Award (DE ). GR is supported by an ARC Discovery Outstanding Researcher Award (DP ). OP is supported by an NHMRC Career Development Fellowship (APP ). 30
32 7 References Ashburner, J, & Friston, KJ. (2000). Voxel-based morphometry - the methods. Neuroimage, 11, Avidan, G, Tanzer, M, Hadj-Bouziane, F, Liu, N, Ungerleider, LG, & Behrmann, M. (2014). Selective Dissociation Between Core and Extended Regions of the Face Processing Network in Congenital Prosopagnosia. Cerebral Cortex, 24, Behrens, TEJ, Johansen-Berg, H, Woolrich, MW, Smith, SM, Wheeler-Kingshott, CAM, Boulby, PA, et al. (2003). Non-invasive mapping of connections between human thalamus and cortex using diffusion imaging. [ /nn1075]. Nat Neurosci, 6, Benton, AL. (1994). Contributions to neuropsychological assessment: A clinical manual: Oxford University Press. Bernstein, LJ, Beig, S, Siegenthaler, AL, & Grady, CL. (2002). The effect of encoding strategy on the neural correlates of memory for faces. Neuropsychologia, 40, Bowles, DC, McKone, E, Dawel, A, Duchaine, B, Palermo, R, Schmalzl, L, et al. (2009). Diagnosing prosopagnosia: Effects of ageing, sex, and participant stimulus ethnic match on the Cambridge Face Memory Test and Cambridge Face Perception Test. Cognitive Neuropsychology, 26, Bruce, V, & Young, A. (1986). Understanding face recognition. British Journal of Psychology, 77, Calder, AJ, & Young, AW. (2005). Understanding the recognition of facial identity and facial expression. Nature reviews neuroscience, 6, Carlesimo, GA, Lombardi, MG, & Caltagirone, C. (2011). Vascular thalamic amnesia: A reappraisal. Neuropsychologia, 49, Chan, D, Anderson, V, Pijenburg, Y, Whitwell, JL, Barnes, J, Scahill, R, et al. (2009). The clinical profile of right temporal lobe atrophy. Brain, 132, Chan, D, Fox, NC, Scahill, R, Crum, WR, Whitwell, JL, Leschziner, G, et al. (2001). Patterns of temporal lobe atrophy in semantic dementia and Alzheimer's disease. Annals of Neurology, 49, Choi, EY, Yeo, BTT, & Buckner, RL. (2012). The organization of the human striatum estimated by intrinsic functional connectivity. Journal of Neurophysiology, 108, Couto, B, Manes, F, Montañés, P, Matallana, D, Reyes, P, Velaszquez, M, et al. (2013). Structural neuroimaging of social cognition in PNFA and bvftd. [Original Research]. Frontiers in Human Neuroscience, 7, Dennett, H, McKone, E, Tavashmi, R, Hall, A, Pidcock, M, Edwards, M, et al. (2012). The Cambridge Car Memory Test: A task matched in format to the Cambridge Face Memory Test, with norms, reliability, sex differences, dissociations from face memory, and expertise effects. Behavior Research Methods, 44, Duchaine, B, & Nakayama, K. (2006). The Cambridge Face Memory Test: Results for neurologically intact individuals and an investigation of its validity using inverted face stimuli and prosopagnosic participants. Neuropsychologia, 44, Duchaine, BC, & Weidenfeld, A. (2003). An evaluation of two commonly used tests of unfamiliar face recognition. Neuropsychologia, 41,
33 Gainotti, G. (2013). Laterality effects in normal subjects' recognition of familiar faces, voices and names. Perceptual and representational components. Neuropsychologia, 51, Galton, CJ, Patterson, K, Graham, K, Lambon-Ralph, M, Williams, G, Antoun, N, et al. (2001). Differing patterns of temporal atrophy in Alzheimer s disease and semantic dementia. Neurology, 57, Gefen, T, Wieneke, C, Martersteck, A, Whitney, K, Weintraub, S, Mesulam, MM, et al. (2013). Naming vs knowing faces in primary progressive aphasia: A tale of 2 hemispheres. Neurology, 81, Gorno-Tempini, ML, Hillis, A, Weintraub, S, Kertesz, A, Mendez, M, Cappa, SF, et al. (2011). Classification of primary progressive aphasia and its variants. Neurology, 76, Haxby, JV, Hoffman, EA, & Gobbini, MI. (2000). The distributed human neural system for face perception. Trends in Cognitive Neurosciences, 4, Hodges, JR, Patterson, K, Oxbury, S, & Funnell, E. (1992). Semantic dementia: Progressive fluent aphasia with temporal lobe atrophy. Brain, 115, Hornberger, M, & Piguet, O. (2012). Episodic memory in frontotemporal dementia: a critical review. Brain, 135, Hornberger, M, Piguet, O, Graham, AJ, Nestor, PJ, & Hodges, JR. (2010). How preserved is episodic memory in behavioural variant frontotemporal dementia. Neurology, 74, Hornberger, M, Wong, S, Tan, R, Irish, M, Piguet, O, Kril, J, et al. (2012). In vivo and postmortem memory circuit integrity in frontotemporal dementia and Alzheimer s disease. Brain, 135, Hsieh, S, Hornberger, M, Piguet, O, & Hodges, JR. (2011). Neural basis of music knowledge: Evidence from the dementias. Brain, 134, Hsieh, S, Irish, M, Daveson, N, Hodges, JR, & Piguet, O. (2013a). When One Loses Empathy: Its Effect on Carers of Patients With Dementia. Journal of Geriatric Psychiatry and Neurology, 26, Hsieh, S, Schubert, S, Hoon, C, Mioshi, E, & Hodges, JR. (2013b). Validation of the Addenbrooke's Cognitive Examination III in Frontotemporal Dementia and Alzheimer's Disease. Dementia and Geriatric Cognitive Disorders, 36, Huis in 't Veld, E, Van den Stock, J, & de Gelder, B. (2012). Configuration perception and face memory, and face context effects in developmental prosopagnosia. Cognitive Neuropsychology, 29, Ibañez, A, & Manes, F. (2012). Contextual social cognition and the behavioral variant of frontotemporal dementia. Neurology, 78, Irish, M, Addis, DR, Hodges, JR, & Piguet, O. (2012). Considering the role of semantic memory in episodic future thinking: evidence from semantic dementia. Brain, 135, Irish, M, Hodges, JR, & Piguet, O. (2014a). Right anterior temporal lobe dysfunction underlies theory of mind impairments in semantic dementia. Brain, 137, Irish, M, Kumfor, F, Hodges, JR, & Piguet, O. (2013). A tale of two hemispheres: Contrasting patterns of socioemotional dysfunction in left- versus right-lateralised semantic dementia. Dementia and Neuropsychologia, 7,
34 Irish, M, Piguet, O, Hodges, JR, & Hornberger, M. (2014b). Common and unique gray matter correlates of episodic memory dysfunction in frontotemporal dementia and alzheimer's disease. Human brain mapping, 35, Josephs, KA, Whitwell, JL, Knopman, DS, Boeve, BF, Vemuri, P, Senjem, ML, et al. (2009). Two distinct subtypes of right temporal variant frontotemporal dementia. Neurology, 73, Josephs, KA, Whitwell, JL, Vemuri, P, Senjem, ML, Boeve, BF, Knopman, DS, et al. (2008). The anatomic correlate of prosopagnosia in semantic dementia. Neurology, 71, Kamminga, J, Kumfor, F, Burrell, JR, Piguet, O, Hodges, JR, & Irish, M. (in press). Differentiating between right-lateralised semantic dementia and behavioural-variant frontotemporal dementia: An examination of clinical characteristics and emotion processing. Journal of Neurology, Neurosurgery and Psychiatry. Kipps, C, Hodges, J, Fryer, T, & Nestor, P. (2009). Combined magnetic resonance imaging and positron emission tomography brain imaging in behavioural variant frontotemporal degeneration: refining the clinical phenotype. Brain, awp077. Kumfor, F, Irish, M, Hodges, JR, & Piguet, O. (2013). The orbitofrontal cortex is involved in emotional enhancement of memory: Evidence from the dementias. Brain, 136, Kumfor, F, Miller, L, Lah, S, Hsieh, S, Savage, S, Hodges, JR, et al. (2011). Are you really angry? The effect of intensity on emotion recognition in frontotemporal dementia. Social Neuroscience, 6, Kumfor, F, & Piguet, O. (2012). Disturbance of emotion processing in frontotemporal dementia: A synthesis of cognitive and neuroimaging findings. Neuropsychology Review, 22, Kumfor, F, Sapey-Triomphe, L-A, Leyton, CE, Burrell, JR, Hodges, JR, & Piguet, O. (2014). Degradation of emotion processing ability in corticobasal syndrome and Alzheimer's disease. Brain, 137, Lieberman, MD, & Cunningham, WA. (2009). Type I and Type II error concerns in fmri research: re-balancing the scale. Social Cognitive and Affective Neuroscience, 4, Mechelli, A, Price, CJ, Friston, KJ, & Ashburner, J. (2005). Voxel-based morphometry of the human brain: Methods and applications. Current Medical Imaging Reviews 1, Mendez, MF, Martin, RJ, Smyth, KA, & Whitehouse, PJ. (1992). Disturbances of person identification in Alzheimer's disease: A retrospective study. The Journal of nervous and mental disease, 180, Miller, L, Hsieh, S, Lah, S, Savage, S, Hodges, JR, & Piguet, O. (2012). One size does not fit all: Face emotion processing impairments in semantic dementia, behavioural-variant frontotemporal dementia and Alzheimer's disease are mediated by distinct cognitive deficits. Behavioural Neurology, 25, Mion, M, Patterson, K, Acosta-Cabronero, J, Pengas, G, Izquierdo-Garcia, D, Hong, YT, et al. (2010). What the left and right anterior fusiform gyri tell us about semantic memory. Brain, 133, Mioshi, E, Dawson, K, Mitchell, J, Arnold, R, & Hodges, JR. (2006). The Addenbrooke's Cognitive Examination Revised (ACE-R): A brief cognitive test battery for dementia screening. International Journal of Geriatric Psychiatry, 21,
35 Mioshi, E, Hsieh, S, Savage, S, Hornberger, M, & Hodges, JR. (2010). Clinical staging and disease progression in frontotemporal dementia. Neurology, 74, Morris, JC. (1993). The Clinical Dementia Rating (CDR): current version and scoring rules. Neurology; Neurology. Nichols, TE, & Holmes, AP. (2002). Nonparametric permutation tests for functional neuroimaging: A primer with examples. Human Brain Mapping, 15, Olson, IR, Ezzyat, Y, Plotzker, A, & Chatterjee, A. (2014). The end point of the ventral visual stream: face and non-face perceptual deficits following unilateral anterior temporal lobe damage. Neurocase, 1-9. Omar, R, Rohrer, JD, Hailstone, JC, & Warren, J. (2010). Structural neuroanatomy of face processing in frontotemporal dementia. Journal of Neurology, Neurosurgery & Psychiatry, 82, Rankin, KP, Salazar, A, Gorno-Tempini, ML, Sollberger, M, Wilson, SM, Pavlic, D, et al. (2009). Detecting sarcasm from paralinguistic cues: Anatomic and cognitive correlates in neurodegenerative disease. Neuroimage, 47, Rascovsky, K, Hodges, JR, Knopman, DS, Mendez, MF, Kramer, JH, Neuhaus, JM, et al. (2011). Sensitivity of revised diagnostic criteria for the behavioural variant of frontotemporal dementia. Brain, 134, Rey, A. (1941). L'examen psychologique dans les cas d'encéphalopathie traumatique. (Les problems.). [The psychological examination in cases of traumatic encepholopathy. Problems.]. Archives de Psychologie, 28, Rivolta, D, Woolgar, A, Palermo, R, Butko, M, Schmalzl, L, & Williams, MA. (2014). Multivoxel pattern analysis (MVPA) reveals abnormal fmri activity in both the core and extended face network in congenital prosopagnosia. [Original Research]. Frontiers in Human Neuroscience, 8. Rosen, HJ, Perry, RJ, Murphy, J, Kramer, JH, Mychack, P, Schuff, N, et al. (2002). Emotion comprehension in the temporal variant of frontotemporal dementia. Brain, 125, Rueckert, D, Sonoda, LI, Hayes, C, Hill, DL, Leach, MO, & Hawkes, DJ. (1999). Nonrigid registration using free-form deformations: Application to breast MR images. IEEE Transactions on Medical Imaging, 18, Savage, S, Hsieh, S, Leslie, F, Foxe, D, Piguet, O, & Hodges, JR. (2013). Distinguishing subtypes in primary progressive aphasia: Application of the Sydney Language Battery. Dementia and Geriatric Cognitive Disorders, 35, Seeley, WW, Crawford, R, Rascovsky, K, Kramer, JH, Weiner, M, Miller, BL, et al. (2008). Frontal paralimbic network atrophy in very mild behavioural variant frontotemporal dementia. Archives of Neurology, 65, Simons, JS, Graham, KS, Galton, CJ, Patterson, K, & Hodges, JR. (2001). Semantic knowledge and episodic memory for faces in semantic dementia. Neuropsychology, 15, 101. Smith, SM, Jenkinson, M, Woolrich, MW, Beckmann, CF, Behrens, TEJ, Johansen-Berg, H, et al. (2004). Advances in functional and structural MR image analysis and implementation as FSL. Neuroimage, 23, Snowden, J, Thompson, J, & Neary, D. (2004). Knowledge of famous faces and names in semantic dementia. Brain, 127, Sollberger, M, Stanley, CM, Wilson, SM, Gyurak, A, Beckman, V, Growden, ME, et al. (2009). Neural basis of interpersonal traits in neurodegenerative diseases. Neuropsychologia, 47,
36 Tan, RH, Wong, S, Kril, JJ, Piguet, O, Hornberger, M, Hodges, JR, et al. (2014). Beyond the temporal pole: limbic memory circuit in the semantic variant of primary progressive aphasia. Brain, awu118. Thompson, SA, Graham, KS, Williams, G, Patterson, K, Kapur, N, & Hodges, JR. (2004). Dissociating person-specific from general semantic knowledge: roles of the left and right temporal lobes. Neuropsychologia, 42, Thompson, SA, Patterson, K, & Hodges, JR. (2003). Left/right asymmetry of atrophy in semantic dementia. Neurology, 61, Tombaugh, TN. (2004). Trail making test A and B: Normative data stratified by age and education. Archives of Clinical Neuropsychology, 19, Von Der Heide, RJ, Skipper, LM, & Olson, IR. (2013). Anterior temporal face patches: a metaanalysis and empirical study. Frontiers in Human Neuroscience, 7. Warrington, E. (1984). Recognition memory test. NFER-Nelson, Windsor. Wechsler, D. (1997). WAIS-III administration and scoring manual. San Antonio: TX: The Psychological Corporation. Woolrich, MW, Jbabdi, S, Patenaude, B, Chappell, M, Makni, S, Behren, T, et al. (2009). Bayesian analysis of neuroimaging data in FSL. Neuroimage, 45, S Zhang, Y, Brady, M, & Smith, S. (2001). Segmentation of brain MR images though a hidden Markov random field model and the expectation-maximisation algorithm. IEEE Transactions on Medical Imaging, 20,

37

38
39
40
41
42
Supplementary Online Content
 Supplementary Online Content Devenney E, Bartley L, Hoon C, et al. Progression in behavioral variant frontotemporal dementia: a longitudinal study. JAMA Neurol. Published online October 26, 2015. doi:10.1001/jamaneurol.2015.2061.
Supplementary Online Content Devenney E, Bartley L, Hoon C, et al. Progression in behavioral variant frontotemporal dementia: a longitudinal study. JAMA Neurol. Published online October 26, 2015. doi:10.1001/jamaneurol.2015.2061.
Piano playing skills in a patient with frontotemporal dementia: A longitudinal case study
 International Symposium on Performance Science ISBN 978-94-90306-01-4 The Author 2009, Published by the AEC All rights reserved Piano playing skills in a patient with frontotemporal dementia: A longitudinal
International Symposium on Performance Science ISBN 978-94-90306-01-4 The Author 2009, Published by the AEC All rights reserved Piano playing skills in a patient with frontotemporal dementia: A longitudinal
On the right side? A longitudinal study of left- versus right-lateralized semantic dementia
 doi:10.1093/brain/awv387 BRAIN 2016: 139; 986 998 986 On the right side? A longitudinal study of left- versus right-lateralized semantic dementia Fiona Kumfor, 1,2,3 Ramon Landin-Romero, 1,3 Emma Devenney,
doi:10.1093/brain/awv387 BRAIN 2016: 139; 986 998 986 On the right side? A longitudinal study of left- versus right-lateralized semantic dementia Fiona Kumfor, 1,2,3 Ramon Landin-Romero, 1,3 Emma Devenney,
FAILURES OF OBJECT RECOGNITION. Dr. Walter S. Marcantoni
 FAILURES OF OBJECT RECOGNITION Dr. Walter S. Marcantoni VISUAL AGNOSIA -damage to the extrastriate visual regions (occipital, parietal and temporal lobes) disrupts recognition of complex visual stimuli
FAILURES OF OBJECT RECOGNITION Dr. Walter S. Marcantoni VISUAL AGNOSIA -damage to the extrastriate visual regions (occipital, parietal and temporal lobes) disrupts recognition of complex visual stimuli
Dr Fiona Kumfor University of Sydney
 Social Cognition in Dementia: Informing Diagnosis, Prognosis and Management Dr Fiona Kumfor University of Sydney CHANGES IN DEMENTIA Memory Social cognition Language ELEMENTS OF SOCIAL COGNITION Face processing
Social Cognition in Dementia: Informing Diagnosis, Prognosis and Management Dr Fiona Kumfor University of Sydney CHANGES IN DEMENTIA Memory Social cognition Language ELEMENTS OF SOCIAL COGNITION Face processing
Supplementary Information Methods Subjects The study was comprised of 84 chronic pain patients with either chronic back pain (CBP) or osteoarthritis
 or osteoarthritis") Supplementary Information Methods Subjects The study was comprised of 84 chronic pain patients with either chronic back pain (CBP) or osteoarthritis (OA). All subjects provided informed consent to procedures
Supplementary Information Methods Subjects The study was comprised of 84 chronic pain patients with either chronic back pain (CBP) or osteoarthritis (OA). All subjects provided informed consent to procedures
Resistance to forgetting associated with hippocampus-mediated. reactivation during new learning
 Resistance to Forgetting 1 Resistance to forgetting associated with hippocampus-mediated reactivation during new learning Brice A. Kuhl, Arpeet T. Shah, Sarah DuBrow, & Anthony D. Wagner Resistance to
Resistance to Forgetting 1 Resistance to forgetting associated with hippocampus-mediated reactivation during new learning Brice A. Kuhl, Arpeet T. Shah, Sarah DuBrow, & Anthony D. Wagner Resistance to
Final Report 2017 Authors: Affiliations: Title of Project: Background:
 Final Report 2017 Authors: Dr Gershon Spitz, Ms Abbie Taing, Professor Jennie Ponsford, Dr Matthew Mundy, Affiliations: Epworth Research Foundation and Monash University Title of Project: The return of
Final Report 2017 Authors: Dr Gershon Spitz, Ms Abbie Taing, Professor Jennie Ponsford, Dr Matthew Mundy, Affiliations: Epworth Research Foundation and Monash University Title of Project: The return of
Frontotemporal Dementia: Towards better diagnosis. Frontotemporal Dementia. John Hodges, NeuRA & University of New South Wales, Sydney.
 I.1 I.2 II.1 II.2 II.3 II.4 II.5 II.6 III.1 III.2 III.3 III.4 III.5 III.6 III.7 III.8 III.9 III.10 III.11 III.12 IV.1 IV.2 IV.3 IV.4 IV.5 Frontotemporal Dementia: Towards better diagnosis Frontotemporal
I.1 I.2 II.1 II.2 II.3 II.4 II.5 II.6 III.1 III.2 III.3 III.4 III.5 III.6 III.7 III.8 III.9 III.10 III.11 III.12 IV.1 IV.2 IV.3 IV.4 IV.5 Frontotemporal Dementia: Towards better diagnosis Frontotemporal
O Connor 1. Appendix e-1
 O Connor 1 Appendix e-1 Neuropsychiatric assessment The Cambridge Behavioural Inventory Revised (CBI-R) 1, 2 is a proxy behavioural questionnaire that has been extensively used in studies involving FTD
O Connor 1 Appendix e-1 Neuropsychiatric assessment The Cambridge Behavioural Inventory Revised (CBI-R) 1, 2 is a proxy behavioural questionnaire that has been extensively used in studies involving FTD
Excellent Network Courses. Department of Neurology Affiliated hospital of Jiangsu University
 Excellent Network Courses Department of Neurology Affiliated hospital of Jiangsu University Agnosia Visual Agnosia Lissauer (1890) described 2 types: a) Apperceptive Cannot see objects b) Associative Does
Excellent Network Courses Department of Neurology Affiliated hospital of Jiangsu University Agnosia Visual Agnosia Lissauer (1890) described 2 types: a) Apperceptive Cannot see objects b) Associative Does
Title:Atypical language organization in temporal lobe epilepsy revealed by a passive semantic paradigm
 Author's response to reviews Title:Atypical language organization in temporal lobe epilepsy revealed by a passive semantic paradigm Authors: Julia Miro (juliamirollado@gmail.com) Pablo Ripollès (pablo.ripolles.vidal@gmail.com)
Author's response to reviews Title:Atypical language organization in temporal lobe epilepsy revealed by a passive semantic paradigm Authors: Julia Miro (juliamirollado@gmail.com) Pablo Ripollès (pablo.ripolles.vidal@gmail.com)
Do women with fragile X syndrome have problems in switching attention: Preliminary findings from ERP and fmri
 Brain and Cognition 54 (2004) 235 239 www.elsevier.com/locate/b&c Do women with fragile X syndrome have problems in switching attention: Preliminary findings from ERP and fmri Kim Cornish, a,b, * Rachel
Brain and Cognition 54 (2004) 235 239 www.elsevier.com/locate/b&c Do women with fragile X syndrome have problems in switching attention: Preliminary findings from ERP and fmri Kim Cornish, a,b, * Rachel
Supplementary Online Content
 Supplementary Online Content Green SA, Hernandez L, Tottenham N, Krasileva K, Bookheimer SY, Dapretto M. The neurobiology of sensory overresponsivity in youth with autism spectrum disorders. Published
Supplementary Online Content Green SA, Hernandez L, Tottenham N, Krasileva K, Bookheimer SY, Dapretto M. The neurobiology of sensory overresponsivity in youth with autism spectrum disorders. Published
Text to brain: predicting the spatial distribution of neuroimaging observations from text reports (submitted to MICCAI 2018)
") 1 / 22 Text to brain: predicting the spatial distribution of neuroimaging observations from text reports (submitted to MICCAI 2018) Jérôme Dockès, ussel Poldrack, Demian Wassermann, Fabian Suchanek, Bertrand
1 / 22 Text to brain: predicting the spatial distribution of neuroimaging observations from text reports (submitted to MICCAI 2018) Jérôme Dockès, ussel Poldrack, Demian Wassermann, Fabian Suchanek, Bertrand
Supplementary Information
 Supplementary Information The neural correlates of subjective value during intertemporal choice Joseph W. Kable and Paul W. Glimcher a 10 0 b 10 0 10 1 10 1 Discount rate k 10 2 Discount rate k 10 2 10
Supplementary Information The neural correlates of subjective value during intertemporal choice Joseph W. Kable and Paul W. Glimcher a 10 0 b 10 0 10 1 10 1 Discount rate k 10 2 Discount rate k 10 2 10
Procedia - Social and Behavioral Sciences 159 ( 2014 ) WCPCG 2014
 WCPCG 2014") Available online at www.sciencedirect.com ScienceDirect Procedia - Social and Behavioral Sciences 159 ( 2014 ) 743 748 WCPCG 2014 Differences in Visuospatial Cognition Performance and Regional Brain Activation
Available online at www.sciencedirect.com ScienceDirect Procedia - Social and Behavioral Sciences 159 ( 2014 ) 743 748 WCPCG 2014 Differences in Visuospatial Cognition Performance and Regional Brain Activation
Define functional MRI. Briefly describe fmri image acquisition. Discuss relative functional neuroanatomy. Review clinical applications.
 Dr. Peter J. Fiester November 14, 2012 Define functional MRI. Briefly describe fmri image acquisition. Discuss relative functional neuroanatomy. Review clinical applications. Briefly discuss a few examples
Dr. Peter J. Fiester November 14, 2012 Define functional MRI. Briefly describe fmri image acquisition. Discuss relative functional neuroanatomy. Review clinical applications. Briefly discuss a few examples
A possible mechanism for impaired joint attention in autism
 A possible mechanism for impaired joint attention in autism Justin H G Williams Morven McWhirr Gordon D Waiter Cambridge Sept 10 th 2010 Joint attention in autism Declarative and receptive aspects initiating
A possible mechanism for impaired joint attention in autism Justin H G Williams Morven McWhirr Gordon D Waiter Cambridge Sept 10 th 2010 Joint attention in autism Declarative and receptive aspects initiating
MULTI-CHANNEL COMMUNICATION
 INTRODUCTION Research on the Deaf Brain is beginning to provide a new evidence base for policy and practice in relation to intervention with deaf children. This talk outlines the multi-channel nature of
INTRODUCTION Research on the Deaf Brain is beginning to provide a new evidence base for policy and practice in relation to intervention with deaf children. This talk outlines the multi-channel nature of
Cover Page. The handle holds various files of this Leiden University dissertation
 Cover Page The handle http://hdl.handle.net/1887/32078 holds various files of this Leiden University dissertation Author: Pannekoek, Nienke Title: Using novel imaging approaches in affective disorders
Cover Page The handle http://hdl.handle.net/1887/32078 holds various files of this Leiden University dissertation Author: Pannekoek, Nienke Title: Using novel imaging approaches in affective disorders
Automated detection of abnormal changes in cortical thickness: A tool to help diagnosis in neocortical focal epilepsy
 Automated detection of abnormal changes in cortical thickness: A tool to help diagnosis in neocortical focal epilepsy 1. Introduction Epilepsy is a common neurological disorder, which affects about 1 %
Automated detection of abnormal changes in cortical thickness: A tool to help diagnosis in neocortical focal epilepsy 1. Introduction Epilepsy is a common neurological disorder, which affects about 1 %
Supplementary Online Content
 Supplementary Online Content Redlich R, Opel N, Grotegerd D, et al. Prediction of individual response to electroconvulsive therapy via machine learning on structural magnetic resonance imaging data. JAMA
Supplementary Online Content Redlich R, Opel N, Grotegerd D, et al. Prediction of individual response to electroconvulsive therapy via machine learning on structural magnetic resonance imaging data. JAMA
The Role of Working Memory in Visual Selective Attention
 Goldsmiths Research Online. The Authors. Originally published: Science vol.291 2 March 2001 1803-1806. http://www.sciencemag.org. 11 October 2000; accepted 17 January 2001 The Role of Working Memory in
Goldsmiths Research Online. The Authors. Originally published: Science vol.291 2 March 2001 1803-1806. http://www.sciencemag.org. 11 October 2000; accepted 17 January 2001 The Role of Working Memory in
CCD Memory Program Inaugural Symposium
 CCD Memory Program Inaugural Symposium THURSDAY 31 st OCTOBER Neuroscience Research Australia This Symposium provides an opportunity to hear eminent researchers, postdoctoral fellows and PhD students present
CCD Memory Program Inaugural Symposium THURSDAY 31 st OCTOBER Neuroscience Research Australia This Symposium provides an opportunity to hear eminent researchers, postdoctoral fellows and PhD students present
Theory of mind skills are related to gray matter volume in the ventromedial prefrontal cortex in schizophrenia
 Theory of mind skills are related to gray matter volume in the ventromedial prefrontal cortex in schizophrenia Supplemental Information Table of Contents 2 Behavioral Data 2 Table S1. Participant demographics
Theory of mind skills are related to gray matter volume in the ventromedial prefrontal cortex in schizophrenia Supplemental Information Table of Contents 2 Behavioral Data 2 Table S1. Participant demographics
fmri and Voxel-based Morphometry in Detection of Early Stages of Alzheimer's Disease
 fmri and Voxel-based Morphometry in Detection of Early Stages of Alzheimer's Disease Andrey V. Sokolov 1,3, Sergey V. Vorobyev 2, Aleksandr Yu. Efimtcev 1,3, Viacheslav S. Dekan 1,3, Gennadiy E. Trufanov
fmri and Voxel-based Morphometry in Detection of Early Stages of Alzheimer's Disease Andrey V. Sokolov 1,3, Sergey V. Vorobyev 2, Aleksandr Yu. Efimtcev 1,3, Viacheslav S. Dekan 1,3, Gennadiy E. Trufanov
Hippocampal brain-network coordination during volitionally controlled exploratory behavior enhances learning
 Online supplementary information for: Hippocampal brain-network coordination during volitionally controlled exploratory behavior enhances learning Joel L. Voss, Brian D. Gonsalves, Kara D. Federmeier,
Online supplementary information for: Hippocampal brain-network coordination during volitionally controlled exploratory behavior enhances learning Joel L. Voss, Brian D. Gonsalves, Kara D. Federmeier,
Andy C.H. Lee a,b,, Trevor W. Robbins b, Stephen Smith c, Gemma A. Calvert c, Irene Tracey c, Paul Matthews c, Adrian M. Owen a. 1.
 Neuropsychologia 40 (2002) 2420 2437 Evidence for asymmetric frontal-lobe involvement in episodic memory from functional magnetic resonance imaging and patients with unilateral frontal-lobe excisions Andy
Neuropsychologia 40 (2002) 2420 2437 Evidence for asymmetric frontal-lobe involvement in episodic memory from functional magnetic resonance imaging and patients with unilateral frontal-lobe excisions Andy
Brain Imaging Applied to Memory & Learning
 Brain Imaging Applied to Memory & Learning John Gabrieli Department of Brain & Cognitive Sciences Institute for Medical Engineering & Sciences McGovern Institute for Brain Sciences MIT Levels of Analysis
Brain Imaging Applied to Memory & Learning John Gabrieli Department of Brain & Cognitive Sciences Institute for Medical Engineering & Sciences McGovern Institute for Brain Sciences MIT Levels of Analysis
11, in nonhuman primates, different cell populations respond to facial identity and expression12, and
 UNDERSTANDING THE RECOGNITION OF FACIAL IDENTITY AND FACIAL EXPRESSION Andrew J. Calder* and Andrew W. Young Abstract Faces convey a wealth of social signals. A dominant view in face-perception research
UNDERSTANDING THE RECOGNITION OF FACIAL IDENTITY AND FACIAL EXPRESSION Andrew J. Calder* and Andrew W. Young Abstract Faces convey a wealth of social signals. A dominant view in face-perception research
Supplementary materials. Appendix A;
 Supplementary materials Appendix A; To determine ADHD diagnoses, a combination of Conners' ADHD questionnaires and a semi-structured diagnostic interview was used(1-4). Each participant was assessed with
Supplementary materials Appendix A; To determine ADHD diagnoses, a combination of Conners' ADHD questionnaires and a semi-structured diagnostic interview was used(1-4). Each participant was assessed with
Face Aftereffects Predict Individual Differences in Face Recognition Ability
 Psychological Science OnlineFirst, published on October 16, 2012 as doi:10.1177/0956797612446350 Research Report Face Aftereffects Predict Individual Differences in Face Recognition Ability Psychological
Psychological Science OnlineFirst, published on October 16, 2012 as doi:10.1177/0956797612446350 Research Report Face Aftereffects Predict Individual Differences in Face Recognition Ability Psychological
Comparing event-related and epoch analysis in blocked design fmri
 Available online at www.sciencedirect.com R NeuroImage 18 (2003) 806 810 www.elsevier.com/locate/ynimg Technical Note Comparing event-related and epoch analysis in blocked design fmri Andrea Mechelli,
Available online at www.sciencedirect.com R NeuroImage 18 (2003) 806 810 www.elsevier.com/locate/ynimg Technical Note Comparing event-related and epoch analysis in blocked design fmri Andrea Mechelli,
VIII. 10. Right Temporal-Lobe Contribution to the Retrieval of Family Relationships in Person Identification
 CYRIC Annual Report 2009 VIII. 10. Right Temporal-Lobe Contribution to the Retrieval of Family Relationships in Person Identification Abe N. 1, Fujii T. 1, Ueno A. 1, Shigemune Y. 1, Suzuki M. 2, Tashiro
CYRIC Annual Report 2009 VIII. 10. Right Temporal-Lobe Contribution to the Retrieval of Family Relationships in Person Identification Abe N. 1, Fujii T. 1, Ueno A. 1, Shigemune Y. 1, Suzuki M. 2, Tashiro
Title of file for HTML: Supplementary Information Description: Supplementary Figures, Supplementary Tables and Supplementary References
 Title of file for HTML: Supplementary Information Description: Supplementary Figures, Supplementary Tables and Supplementary References Supplementary Information Supplementary Figure 1. The mean parameter
Title of file for HTML: Supplementary Information Description: Supplementary Figures, Supplementary Tables and Supplementary References Supplementary Information Supplementary Figure 1. The mean parameter
Supporting Online Material for
 www.sciencemag.org/cgi/content/full/324/5927/646/dc1 Supporting Online Material for Self-Control in Decision-Making Involves Modulation of the vmpfc Valuation System Todd A. Hare,* Colin F. Camerer, Antonio
www.sciencemag.org/cgi/content/full/324/5927/646/dc1 Supporting Online Material for Self-Control in Decision-Making Involves Modulation of the vmpfc Valuation System Todd A. Hare,* Colin F. Camerer, Antonio
Cognitive Neuroscience of Memory
 Cognitive Neuroscience of Memory Types and Structure of Memory Types of Memory Type of Memory Time Course Capacity Conscious Awareness Mechanism of Loss Sensory Short-Term and Working Long-Term Nondeclarative
Cognitive Neuroscience of Memory Types and Structure of Memory Types of Memory Type of Memory Time Course Capacity Conscious Awareness Mechanism of Loss Sensory Short-Term and Working Long-Term Nondeclarative
Experimental Design. Outline. Outline. A very simple experiment. Activation for movement versus rest
 Experimental Design Kate Watkins Department of Experimental Psychology University of Oxford With thanks to: Heidi Johansen-Berg Joe Devlin Outline Choices for experimental paradigm Subtraction / hierarchical
Experimental Design Kate Watkins Department of Experimental Psychology University of Oxford With thanks to: Heidi Johansen-Berg Joe Devlin Outline Choices for experimental paradigm Subtraction / hierarchical
Supplemental Information. Triangulating the Neural, Psychological, and Economic Bases of Guilt Aversion
 Neuron, Volume 70 Supplemental Information Triangulating the Neural, Psychological, and Economic Bases of Guilt Aversion Luke J. Chang, Alec Smith, Martin Dufwenberg, and Alan G. Sanfey Supplemental Information
Neuron, Volume 70 Supplemental Information Triangulating the Neural, Psychological, and Economic Bases of Guilt Aversion Luke J. Chang, Alec Smith, Martin Dufwenberg, and Alan G. Sanfey Supplemental Information
Learning Objectives.
 Emilie O Neill, class of 2016 Learning Objectives 1. Describe the types of deficits that occur with lesions in association areas including: prosopagnosia, neglect, aphasias, agnosia, apraxia 2. Discuss
Emilie O Neill, class of 2016 Learning Objectives 1. Describe the types of deficits that occur with lesions in association areas including: prosopagnosia, neglect, aphasias, agnosia, apraxia 2. Discuss
Frontal Contributions to Memory Encoding Before and After Unilateral Medial Temporal Lobectomy
 Frontal Contributions to Memory Encoding Before and After Unilateral Medial Temporal Lobectomy Jeff Ojemann, MD Department of Neurological Surgery University of Washington Children s Hospital & Regional
Frontal Contributions to Memory Encoding Before and After Unilateral Medial Temporal Lobectomy Jeff Ojemann, MD Department of Neurological Surgery University of Washington Children s Hospital & Regional
The Frontal Lobes. Anatomy of the Frontal Lobes. Anatomy of the Frontal Lobes 3/2/2011. Portrait: Losing Frontal-Lobe Functions. Readings: KW Ch.
 The Frontal Lobes Readings: KW Ch. 16 Portrait: Losing Frontal-Lobe Functions E.L. Highly organized college professor Became disorganized, showed little emotion, and began to miss deadlines Scores on intelligence
The Frontal Lobes Readings: KW Ch. 16 Portrait: Losing Frontal-Lobe Functions E.L. Highly organized college professor Became disorganized, showed little emotion, and began to miss deadlines Scores on intelligence
Remembering the Past to Imagine the Future: A Cognitive Neuroscience Perspective
 MILITARY PSYCHOLOGY, 21:(Suppl. 1)S108 S112, 2009 Copyright Taylor & Francis Group, LLC ISSN: 0899-5605 print / 1532-7876 online DOI: 10.1080/08995600802554748 Remembering the Past to Imagine the Future:
MILITARY PSYCHOLOGY, 21:(Suppl. 1)S108 S112, 2009 Copyright Taylor & Francis Group, LLC ISSN: 0899-5605 print / 1532-7876 online DOI: 10.1080/08995600802554748 Remembering the Past to Imagine the Future:
Neuroimaging methods vs. lesion studies FOCUSING ON LANGUAGE
 Neuroimaging methods vs. lesion studies FOCUSING ON LANGUAGE Pioneers in lesion studies Their postmortem examination provided the basis for the linkage of the left hemisphere with language C. Wernicke
Neuroimaging methods vs. lesion studies FOCUSING ON LANGUAGE Pioneers in lesion studies Their postmortem examination provided the basis for the linkage of the left hemisphere with language C. Wernicke
Henry Molaison. Biography. From Wikipedia, the free encyclopedia
 Henry Molaison From Wikipedia, the free encyclopedia Henry Gustav Molaison (February 26, 1926 December 2, 2008), known widely as H.M., was an American memory disorder patient who had a bilateral medial
Henry Molaison From Wikipedia, the free encyclopedia Henry Gustav Molaison (February 26, 1926 December 2, 2008), known widely as H.M., was an American memory disorder patient who had a bilateral medial
Twelve right-handed subjects between the ages of 22 and 30 were recruited from the
 Supplementary Methods Materials & Methods Subjects Twelve right-handed subjects between the ages of 22 and 30 were recruited from the Dartmouth community. All subjects were native speakers of English,
Supplementary Methods Materials & Methods Subjects Twelve right-handed subjects between the ages of 22 and 30 were recruited from the Dartmouth community. All subjects were native speakers of English,
Neural Correlates of Episodic Memory in Behavioral Variant Frontotemporal Dementia
 Journal of Alzheimer s Disease 24 (2011) 261 268 DOI 10.3233/JAD-2010-101668 IOS Press 261 Neural Correlates of Episodic Memory in Behavioral Variant Frontotemporal Dementia Catherine Pennington a, John
Journal of Alzheimer s Disease 24 (2011) 261 268 DOI 10.3233/JAD-2010-101668 IOS Press 261 Neural Correlates of Episodic Memory in Behavioral Variant Frontotemporal Dementia Catherine Pennington a, John
Functional topography of a distributed neural system for spatial and nonspatial information maintenance in working memory
 Neuropsychologia 41 (2003) 341 356 Functional topography of a distributed neural system for spatial and nonspatial information maintenance in working memory Joseph B. Sala a,, Pia Rämä a,c,d, Susan M.
Neuropsychologia 41 (2003) 341 356 Functional topography of a distributed neural system for spatial and nonspatial information maintenance in working memory Joseph B. Sala a,, Pia Rämä a,c,d, Susan M.
SUPPLEMENTARY MATERIAL. Table. Neuroimaging studies on the premonitory urge and sensory function in patients with Tourette syndrome.
 SUPPLEMENTARY MATERIAL Table. Neuroimaging studies on the premonitory urge and sensory function in patients with Tourette syndrome. Authors Year Patients Male gender (%) Mean age (range) Adults/ Children
SUPPLEMENTARY MATERIAL Table. Neuroimaging studies on the premonitory urge and sensory function in patients with Tourette syndrome. Authors Year Patients Male gender (%) Mean age (range) Adults/ Children
Fundamentals of Cognitive Psychology, 3e by Ronald T. Kellogg Chapter 2. Multiple Choice
 Multiple Choice 1. Which structure is not part of the visual pathway in the brain? a. occipital lobe b. optic chiasm c. lateral geniculate nucleus *d. frontal lobe Answer location: Visual Pathways 2. Which
Multiple Choice 1. Which structure is not part of the visual pathway in the brain? a. occipital lobe b. optic chiasm c. lateral geniculate nucleus *d. frontal lobe Answer location: Visual Pathways 2. Which
Supplementary Online Material Supplementary Table S1 to S5 Supplementary Figure S1 to S4
 Supplementary Online Material Supplementary Table S1 to S5 Supplementary Figure S1 to S4 Table S1: Brain regions involved in the adapted classification learning task Brain Regions x y z Z Anterior Cingulate
Supplementary Online Material Supplementary Table S1 to S5 Supplementary Figure S1 to S4 Table S1: Brain regions involved in the adapted classification learning task Brain Regions x y z Z Anterior Cingulate
Voxel-based morphometry in clinical neurosciences
 Voxel-based morphometry in clinical neurosciences Ph.D. Thesis Ádám Feldmann Department of Behavioural Sciences Leader of Doctoral School: Prof. Dr.Sámuel Komoly, D.Sc. Program leader: Prof. Dr.Sámuel
Voxel-based morphometry in clinical neurosciences Ph.D. Thesis Ádám Feldmann Department of Behavioural Sciences Leader of Doctoral School: Prof. Dr.Sámuel Komoly, D.Sc. Program leader: Prof. Dr.Sámuel
Chapter 2 Test. 1. Evolutionary structures within the are the most primitive. *a. hindbrain b. thalamus c. forebrain d. midbrain e.
 Cognitive Psychology In and Out of the Laboratory 5th Edition Galotti TEST BANK Full clear download (no formatting errors) at: https://testbankreal.com/download/cognitive-psychology-laboratory-5thedition-galotti-test-bank/
Cognitive Psychology In and Out of the Laboratory 5th Edition Galotti TEST BANK Full clear download (no formatting errors) at: https://testbankreal.com/download/cognitive-psychology-laboratory-5thedition-galotti-test-bank/
Neural activity to positive expressions predicts daily experience of schizophrenia-spectrum symptoms in adults with high social anhedonia
 1 Neural activity to positive expressions predicts daily experience of schizophrenia-spectrum symptoms in adults with high social anhedonia Christine I. Hooker, Taylor L. Benson, Anett Gyurak, Hong Yin,
1 Neural activity to positive expressions predicts daily experience of schizophrenia-spectrum symptoms in adults with high social anhedonia Christine I. Hooker, Taylor L. Benson, Anett Gyurak, Hong Yin,
correlates with social context behavioral adaptation.
 REVIEW OF FRONTAL LOBE STRUCTURES Main organization of frontal cortex: 1. Motor area (precentral gyrus). 2. Premotor & supplementary motor areas (immediately anterior to motor area). Includes premotor,
REVIEW OF FRONTAL LOBE STRUCTURES Main organization of frontal cortex: 1. Motor area (precentral gyrus). 2. Premotor & supplementary motor areas (immediately anterior to motor area). Includes premotor,
Supplementary Digital Content
 Supplementary Digital Content Contextual modulation of pain in masochists: involvement of the parietal operculum and insula Sandra Kamping a, Jamila Andoh a, Isabelle C. Bomba a, Martin Diers a,b, Eugen
Supplementary Digital Content Contextual modulation of pain in masochists: involvement of the parietal operculum and insula Sandra Kamping a, Jamila Andoh a, Isabelle C. Bomba a, Martin Diers a,b, Eugen
MSc Neuroimaging for Clinical & Cognitive Neuroscience
 MSc Neuroimaging for Clinical & Cognitive Neuroscience School of Psychological Sciences Faculty of Medical & Human Sciences Module Information *Please note that this is a sample guide to modules. The exact
MSc Neuroimaging for Clinical & Cognitive Neuroscience School of Psychological Sciences Faculty of Medical & Human Sciences Module Information *Please note that this is a sample guide to modules. The exact
FRONTAL LOBE. Central Sulcus. Ascending ramus of the Cingulate Sulcus. Cingulate Sulcus. Lateral Sulcus
 FRONTAL LOBE Central Ascending ramus of the Cingulate Cingulate Lateral Lateral View Medial View Motor execution and higher cognitive functions (e.g., language production, impulse inhibition, reasoning
FRONTAL LOBE Central Ascending ramus of the Cingulate Cingulate Lateral Lateral View Medial View Motor execution and higher cognitive functions (e.g., language production, impulse inhibition, reasoning
Supplemental Information. Direct Electrical Stimulation in the Human Brain. Disrupts Melody Processing
 Current Biology, Volume 27 Supplemental Information Direct Electrical Stimulation in the Human Brain Disrupts Melody Processing Frank E. Garcea, Benjamin L. Chernoff, Bram Diamond, Wesley Lewis, Maxwell
Current Biology, Volume 27 Supplemental Information Direct Electrical Stimulation in the Human Brain Disrupts Melody Processing Frank E. Garcea, Benjamin L. Chernoff, Bram Diamond, Wesley Lewis, Maxwell
Supplemental information online for
 Supplemental information online for Sleep contributes to the strengthening of some memories over others, depending on hippocampal activity at learning. Géraldine Rauchs (1,2), Dorothée Feyers (1), Brigitte
Supplemental information online for Sleep contributes to the strengthening of some memories over others, depending on hippocampal activity at learning. Géraldine Rauchs (1,2), Dorothée Feyers (1), Brigitte
CISC 3250 Systems Neuroscience
 CISC 3250 Systems Neuroscience Levels of organization Central Nervous System 1m 10 11 neurons Neural systems and neuroanatomy Systems 10cm Networks 1mm Neurons 100μm 10 8 neurons Professor Daniel Leeds
CISC 3250 Systems Neuroscience Levels of organization Central Nervous System 1m 10 11 neurons Neural systems and neuroanatomy Systems 10cm Networks 1mm Neurons 100μm 10 8 neurons Professor Daniel Leeds
doi: /brain/awu246 Brain 2014: 137; Degradation of emotion processing ability in corticobasal syndrome and Alzheimer s disease
 doi:10.1093/brain/awu246 Brain 2014: 137; 3061 3072 3061 BRAIN A JOURNAL OF NEUROLOGY Degradation of emotion processing ability in corticobasal syndrome and Alzheimer s disease Fiona Kumfor, 1,2,3, * Laurie-Anne
doi:10.1093/brain/awu246 Brain 2014: 137; 3061 3072 3061 BRAIN A JOURNAL OF NEUROLOGY Degradation of emotion processing ability in corticobasal syndrome and Alzheimer s disease Fiona Kumfor, 1,2,3, * Laurie-Anne
Neural correlates of memory for object identity and object location: effects of aging
 Neuropsychologia 40 (2002) 1428 1442 Neural correlates of memory for object identity and object location: effects of aging Alessandra Schiavetto a, Stefan Köhler a, Cheryl L. Grady a, Gordon Winocur a,c,
Neuropsychologia 40 (2002) 1428 1442 Neural correlates of memory for object identity and object location: effects of aging Alessandra Schiavetto a, Stefan Köhler a, Cheryl L. Grady a, Gordon Winocur a,c,
Functional MRI Mapping Cognition
 Outline Functional MRI Mapping Cognition Michael A. Yassa, B.A. Division of Psychiatric Neuro-imaging Psychiatry and Behavioral Sciences Johns Hopkins School of Medicine Why fmri? fmri - How it works Research
Outline Functional MRI Mapping Cognition Michael A. Yassa, B.A. Division of Psychiatric Neuro-imaging Psychiatry and Behavioral Sciences Johns Hopkins School of Medicine Why fmri? fmri - How it works Research
Top-Down Activation of Fusiform Cortex without Seeing Faces in Prosopagnosia
 Cerebral Cortex August 2010;20:1878--1890 doi:10.1093/cercor/bhp254 Advance Access publication November 25, 2009 Top-Down Activation of Fusiform Cortex without Seeing Faces in Prosopagnosia Ruthger Righart
Cerebral Cortex August 2010;20:1878--1890 doi:10.1093/cercor/bhp254 Advance Access publication November 25, 2009 Top-Down Activation of Fusiform Cortex without Seeing Faces in Prosopagnosia Ruthger Righart
Voxel-based Lesion-Symptom Mapping. Céline R. Gillebert
 Voxel-based Lesion-Symptom Mapping Céline R. Gillebert Paul Broca (1861) Mr. Tan no productive speech single repetitive syllable tan Broca s area: speech production Broca s aphasia: problems with fluency,
Voxel-based Lesion-Symptom Mapping Céline R. Gillebert Paul Broca (1861) Mr. Tan no productive speech single repetitive syllable tan Broca s area: speech production Broca s aphasia: problems with fluency,
doi: /brain/aws024 Brain 2012: 135; Altered brain mechanisms of emotion processing in pre-manifest Huntington s disease
 doi:10.1093/brain/aws024 Brain 2012: 135; 1165 1179 1165 BRAIN A JOURNAL OF NEUROLOGY Altered brain mechanisms of emotion processing in pre-manifest Huntington s disease Marianne J. U. Novak, 1 Jason D.
doi:10.1093/brain/aws024 Brain 2012: 135; 1165 1179 1165 BRAIN A JOURNAL OF NEUROLOGY Altered brain mechanisms of emotion processing in pre-manifest Huntington s disease Marianne J. U. Novak, 1 Jason D.
Cover Page. The handle holds various files of this Leiden University dissertation
 Cover Page The handle http://hdl.handle.net/1887/39720 holds various files of this Leiden University dissertation Author: Hafkemeijer, Anne Title: Brain networks in aging and dementia Issue Date: 2016-05-26
Cover Page The handle http://hdl.handle.net/1887/39720 holds various files of this Leiden University dissertation Author: Hafkemeijer, Anne Title: Brain networks in aging and dementia Issue Date: 2016-05-26
Classification and Statistical Analysis of Auditory FMRI Data Using Linear Discriminative Analysis and Quadratic Discriminative Analysis
 International Journal of Innovative Research in Computer Science & Technology (IJIRCST) ISSN: 2347-5552, Volume-2, Issue-6, November-2014 Classification and Statistical Analysis of Auditory FMRI Data Using
International Journal of Innovative Research in Computer Science & Technology (IJIRCST) ISSN: 2347-5552, Volume-2, Issue-6, November-2014 Classification and Statistical Analysis of Auditory FMRI Data Using
UNDERSTANDING THE RECOGNITION OF FACIAL IDENTITY AND FACIAL EXPRESSION
 UNDERSTANDING THE RECOGNITION OF FACIAL IDENTITY AND FACIAL EXPRESSION Andrew J. Calder* and Andrew W. Young Abstract Faces convey a wealth of social signals. A dominant view in face-perception research
UNDERSTANDING THE RECOGNITION OF FACIAL IDENTITY AND FACIAL EXPRESSION Andrew J. Calder* and Andrew W. Young Abstract Faces convey a wealth of social signals. A dominant view in face-perception research
Social Neuroscience Publication details, including instructions for authors and subscription information:
 This article was downloaded by: [Olivier Piguet] On: 30 September 11, At: 15:49 Publisher: Psychology Press Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer
This article was downloaded by: [Olivier Piguet] On: 30 September 11, At: 15:49 Publisher: Psychology Press Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer
I do not have any disclosures
 Alzheimer s Disease: Update on Research, Treatment & Care Clinicopathological Classifications of FTD and Related Disorders Keith A. Josephs, MST, MD, MS Associate Professor & Consultant of Neurology Mayo
Alzheimer s Disease: Update on Research, Treatment & Care Clinicopathological Classifications of FTD and Related Disorders Keith A. Josephs, MST, MD, MS Associate Professor & Consultant of Neurology Mayo
Perception of Faces and Bodies
 CURRENT DIRECTIONS IN PSYCHOLOGICAL SCIENCE Perception of Faces and Bodies Similar or Different? Virginia Slaughter, 1 Valerie E. Stone, 2 and Catherine Reed 3 1 Early Cognitive Development Unit and 2
CURRENT DIRECTIONS IN PSYCHOLOGICAL SCIENCE Perception of Faces and Bodies Similar or Different? Virginia Slaughter, 1 Valerie E. Stone, 2 and Catherine Reed 3 1 Early Cognitive Development Unit and 2
Involvement of both prefrontal and inferior parietal cortex. in dual-task performance
 Involvement of both prefrontal and inferior parietal cortex in dual-task performance Fabienne Collette a,b, Laurence 01ivier b,c, Martial Van der Linden a,d, Steven Laureys b, Guy Delfiore b, André Luxen
Involvement of both prefrontal and inferior parietal cortex in dual-task performance Fabienne Collette a,b, Laurence 01ivier b,c, Martial Van der Linden a,d, Steven Laureys b, Guy Delfiore b, André Luxen
Structural MRI in Frontotemporal Dementia: Comparisons between Hippocampal Volumetry, Tensor- Based Morphometry and Voxel-Based Morphometry
 : Comparisons between Hippocampal Volumetry, Tensor- Based Morphometry and Voxel-Based Morphometry Miguel Ángel Muñoz-Ruiz 1,Päivi Hartikainen 1,2, Juha Koikkalainen 3, Robin Wolz 4, Valtteri Julkunen
: Comparisons between Hippocampal Volumetry, Tensor- Based Morphometry and Voxel-Based Morphometry Miguel Ángel Muñoz-Ruiz 1,Päivi Hartikainen 1,2, Juha Koikkalainen 3, Robin Wolz 4, Valtteri Julkunen
Impaired face discrimination in acquired prosopagnosia is associated with abnormal response to individual faces in the right middle fusiform gyrus
 Impaired face discrimination in acquired prosopagnosia is associated with abnormal response to individual faces in the right middle fusiform gyrus Christine Schiltz Bettina Sorger Roberto Caldara Fatima
Impaired face discrimination in acquired prosopagnosia is associated with abnormal response to individual faces in the right middle fusiform gyrus Christine Schiltz Bettina Sorger Roberto Caldara Fatima
Auditory Processing Of Schizophrenia
 Auditory Processing Of Schizophrenia In general, sensory processing as well as selective attention impairments are common amongst people with schizophrenia. It is important to note that experts have in
Auditory Processing Of Schizophrenia In general, sensory processing as well as selective attention impairments are common amongst people with schizophrenia. It is important to note that experts have in
Regional Grey Matter Atrophy is associated with Social Cognition impairment in Progressive Multiple Sclerosis (MS).
.") Escuela de Medicina Regional Grey Matter Atrophy is associated with Social Cognition impairment in Progressive Multiple Sclerosis (MS). Escuela de Medicina Tomas Labbe(1), Ethel Ciampi(2), Juan Pablo Cruz(3),
Escuela de Medicina Regional Grey Matter Atrophy is associated with Social Cognition impairment in Progressive Multiple Sclerosis (MS). Escuela de Medicina Tomas Labbe(1), Ethel Ciampi(2), Juan Pablo Cruz(3),
Brain activity related to integrative processes in visual object recognition: bottom-up integration and the modulatory influence of stored knowledge
 Neuropsychologia 40 (2002) 1254 1267 Brain activity related to integrative processes in visual object recognition: bottom-up integration and the modulatory influence of stored knowledge C. Gerlach a,,
Neuropsychologia 40 (2002) 1254 1267 Brain activity related to integrative processes in visual object recognition: bottom-up integration and the modulatory influence of stored knowledge C. Gerlach a,,
Ch 8. Learning and Memory
 Ch 8. Learning and Memory Cognitive Neuroscience: The Biology of the Mind, 2 nd Ed., M. S. Gazzaniga, R. B. Ivry, and G. R. Mangun, Norton, 2002. Summarized by H.-S. Seok, K. Kim, and B.-T. Zhang Biointelligence
Ch 8. Learning and Memory Cognitive Neuroscience: The Biology of the Mind, 2 nd Ed., M. S. Gazzaniga, R. B. Ivry, and G. R. Mangun, Norton, 2002. Summarized by H.-S. Seok, K. Kim, and B.-T. Zhang Biointelligence
Supplementary Materials for
 Supplementary Materials for Folk Explanations of Behavior: A Specialized Use of a Domain-General Mechanism Robert P. Spunt & Ralph Adolphs California Institute of Technology Correspondence may be addressed
Supplementary Materials for Folk Explanations of Behavior: A Specialized Use of a Domain-General Mechanism Robert P. Spunt & Ralph Adolphs California Institute of Technology Correspondence may be addressed
Group-Wise FMRI Activation Detection on Corresponding Cortical Landmarks
 Group-Wise FMRI Activation Detection on Corresponding Cortical Landmarks Jinglei Lv 1,2, Dajiang Zhu 2, Xintao Hu 1, Xin Zhang 1,2, Tuo Zhang 1,2, Junwei Han 1, Lei Guo 1,2, and Tianming Liu 2 1 School
Group-Wise FMRI Activation Detection on Corresponding Cortical Landmarks Jinglei Lv 1,2, Dajiang Zhu 2, Xintao Hu 1, Xin Zhang 1,2, Tuo Zhang 1,2, Junwei Han 1, Lei Guo 1,2, and Tianming Liu 2 1 School
Ch 8. Learning and Memory
 Ch 8. Learning and Memory Cognitive Neuroscience: The Biology of the Mind, 2 nd Ed., M. S. Gazzaniga,, R. B. Ivry,, and G. R. Mangun,, Norton, 2002. Summarized by H.-S. Seok, K. Kim, and B.-T. Zhang Biointelligence
Ch 8. Learning and Memory Cognitive Neuroscience: The Biology of the Mind, 2 nd Ed., M. S. Gazzaniga,, R. B. Ivry,, and G. R. Mangun,, Norton, 2002. Summarized by H.-S. Seok, K. Kim, and B.-T. Zhang Biointelligence
Supplementary information Detailed Materials and Methods
 Supplementary information Detailed Materials and Methods Subjects The experiment included twelve subjects: ten sighted subjects and two blind. Five of the ten sighted subjects were expert users of a visual-to-auditory
Supplementary information Detailed Materials and Methods Subjects The experiment included twelve subjects: ten sighted subjects and two blind. Five of the ten sighted subjects were expert users of a visual-to-auditory
Reading Words and Non-Words: A Joint fmri and Eye-Tracking Study
 Reading Words and Non-Words: A Joint fmri and Eye-Tracking Study A N N - M A R I E R A P H A I L S R E B C S, S K I D M O R E C O L L E G E D R. J O H N H E N D E R S O N U N I V E R S I T Y O F S O U
Reading Words and Non-Words: A Joint fmri and Eye-Tracking Study A N N - M A R I E R A P H A I L S R E B C S, S K I D M O R E C O L L E G E D R. J O H N H E N D E R S O N U N I V E R S I T Y O F S O U
Right lateral prefrontal cortex Specificity for inhibition or strategy use?
 Right lateral prefrontal cortex Specificity for inhibition or strategy use? M. Hornberger 1 & M. Bertoux 1 1 Department of Clinical Neurosciences, University of Cambridge, Cambridge, UK The specific functions
Right lateral prefrontal cortex Specificity for inhibition or strategy use? M. Hornberger 1 & M. Bertoux 1 1 Department of Clinical Neurosciences, University of Cambridge, Cambridge, UK The specific functions
Brain gray matter volume changes associated with motor symptoms in patients with Parkinson s disease
 Kang et al. Chinese Neurosurgical Journal (2015) 1:9 DOI 10.1186/s41016-015-0003-6 RESEARCH Open Access Brain gray matter volume changes associated with motor symptoms in patients with Parkinson s disease
Kang et al. Chinese Neurosurgical Journal (2015) 1:9 DOI 10.1186/s41016-015-0003-6 RESEARCH Open Access Brain gray matter volume changes associated with motor symptoms in patients with Parkinson s disease
Functional Elements and Networks in fmri
 Functional Elements and Networks in fmri Jarkko Ylipaavalniemi 1, Eerika Savia 1,2, Ricardo Vigário 1 and Samuel Kaski 1,2 1- Helsinki University of Technology - Adaptive Informatics Research Centre 2-
Functional Elements and Networks in fmri Jarkko Ylipaavalniemi 1, Eerika Savia 1,2, Ricardo Vigário 1 and Samuel Kaski 1,2 1- Helsinki University of Technology - Adaptive Informatics Research Centre 2-
Brain Advance Access published November 3, doi: /brain/awp271 Brain 2009: Page 1 of 13 1
 Brain Advance Access published November 3, 2009 doi:10.1093/brain/awp271 Brain 2009: Page 1 of 13 1 BRAIN A JOURNAL OF NEUROLOGY Voxel-based morphometry reveals reduced grey matter volume in the temporal
Brain Advance Access published November 3, 2009 doi:10.1093/brain/awp271 Brain 2009: Page 1 of 13 1 BRAIN A JOURNAL OF NEUROLOGY Voxel-based morphometry reveals reduced grey matter volume in the temporal
Supporting online material. Materials and Methods. We scanned participants in two groups of 12 each. Group 1 was composed largely of
 Placebo effects in fmri Supporting online material 1 Supporting online material Materials and Methods Study 1 Procedure and behavioral data We scanned participants in two groups of 12 each. Group 1 was
Placebo effects in fmri Supporting online material 1 Supporting online material Materials and Methods Study 1 Procedure and behavioral data We scanned participants in two groups of 12 each. Group 1 was
Human Paleoneurology and the Evolution of the Parietal Cortex
 PARIETAL LOBE The Parietal Lobes develop at about the age of 5 years. They function to give the individual perspective and to help them understand space, touch, and volume. The location of the parietal
PARIETAL LOBE The Parietal Lobes develop at about the age of 5 years. They function to give the individual perspective and to help them understand space, touch, and volume. The location of the parietal
5th Mini-Symposium on Cognition, Decision-making and Social Function: In Memory of Kang Cheng
 5th Mini-Symposium on Cognition, Decision-making and Social Function: In Memory of Kang Cheng 13:30-13:35 Opening 13:30 17:30 13:35-14:00 Metacognition in Value-based Decision-making Dr. Xiaohong Wan (Beijing
5th Mini-Symposium on Cognition, Decision-making and Social Function: In Memory of Kang Cheng 13:30-13:35 Opening 13:30 17:30 13:35-14:00 Metacognition in Value-based Decision-making Dr. Xiaohong Wan (Beijing
Accepted Manuscript. Behavioural-variant frontotemporal dementia: a unique window into the disrupted self Reply to Genon & Salmon
 Accepted Manuscript Behavioural-variant frontotemporal dementia: a unique window into the disrupted self Reply to Genon & Salmon Stephanie Wong, Muireann Irish, Michael Hornberger PII: S0010-9452(18)30060-1
Accepted Manuscript Behavioural-variant frontotemporal dementia: a unique window into the disrupted self Reply to Genon & Salmon Stephanie Wong, Muireann Irish, Michael Hornberger PII: S0010-9452(18)30060-1
Theories of memory. Memory & brain Cellular bases of learning & memory. Epileptic patient Temporal lobectomy Amnesia
 Cognitive Neuroscience: The Biology of the Mind, 2 nd Ed., M. S. Gazzaniga, R. B. Ivry, and G. R. Mangun, Norton, 2002. Theories of Sensory, short-term & long-term memories Memory & brain Cellular bases
Cognitive Neuroscience: The Biology of the Mind, 2 nd Ed., M. S. Gazzaniga, R. B. Ivry, and G. R. Mangun, Norton, 2002. Theories of Sensory, short-term & long-term memories Memory & brain Cellular bases
Sensorimotor Functioning. Sensory and Motor Systems. Functional Anatomy of Brain- Behavioral Relationships
 Sensorimotor Functioning Sensory and Motor Systems Understanding brain-behavior relationships requires knowledge of sensory and motor systems. Sensory System = Input Neural Processing Motor System = Output
Sensorimotor Functioning Sensory and Motor Systems Understanding brain-behavior relationships requires knowledge of sensory and motor systems. Sensory System = Input Neural Processing Motor System = Output
Hallucinations and conscious access to visual inputs in Parkinson s disease
 Supplemental informations Hallucinations and conscious access to visual inputs in Parkinson s disease Stéphanie Lefebvre, PhD^1,2, Guillaume Baille, MD^4, Renaud Jardri MD, PhD 1,2 Lucie Plomhause, PhD
Supplemental informations Hallucinations and conscious access to visual inputs in Parkinson s disease Stéphanie Lefebvre, PhD^1,2, Guillaume Baille, MD^4, Renaud Jardri MD, PhD 1,2 Lucie Plomhause, PhD
Visual Rating Scale Reference Material. Lorna Harper Dementia Research Centre University College London
 Visual Rating Scale Reference Material Lorna Harper Dementia Research Centre University College London Background The reference materials included in this document were compiled and used in relation to
Visual Rating Scale Reference Material Lorna Harper Dementia Research Centre University College London Background The reference materials included in this document were compiled and used in relation to
QUANTIFYING CEREBRAL CONTRIBUTIONS TO PAIN 1
 QUANTIFYING CEREBRAL CONTRIBUTIONS TO PAIN 1 Supplementary Figure 1. Overview of the SIIPS1 development. The development of the SIIPS1 consisted of individual- and group-level analysis steps. 1) Individual-person
QUANTIFYING CEREBRAL CONTRIBUTIONS TO PAIN 1 Supplementary Figure 1. Overview of the SIIPS1 development. The development of the SIIPS1 consisted of individual- and group-level analysis steps. 1) Individual-person
Online appendices are unedited and posted as supplied by the authors. SUPPLEMENTARY MATERIAL
 Appendix 1 to Sehmbi M, Rowley CD, Minuzzi L, et al. Age-related deficits in intracortical myelination in young adults with bipolar SUPPLEMENTARY MATERIAL Supplementary Methods Intracortical Myelin (ICM)
Appendix 1 to Sehmbi M, Rowley CD, Minuzzi L, et al. Age-related deficits in intracortical myelination in young adults with bipolar SUPPLEMENTARY MATERIAL Supplementary Methods Intracortical Myelin (ICM)