UNIQUE PHARMACODYNAMIC PROFILE OF METHYLPHENIDATE: SELF- ADMINISTRATION-INDUCED DOPAMINE SYSTEM ALTERATIONS ERIN SUE CALIPARI
|
|
|
- Benedict Malone
- 5 years ago
- Views:
Transcription
1 UNIQUE PHARMACODYNAMIC PROFILE OF METHYLPHENIDATE: SELF- ADMINISTRATION-INDUCED DOPAMINE SYSTEM ALTERATIONS BY ERIN SUE CALIPARI A Dissertation Submitted to the Graduate Faculty of WAKE FOREST UNIVERSITY GRADUATE SCHOOL OF ARTS AND SCIENCES In Partial Fulfillment of the Requirements for the Degree of DOCTOR OF PHILOSOPHY Program in Neuroscience December 2013 Winston-Salem, North Carolina Approved by: Sara R. Jones, Ph.D., Advisor Thomas J. Martin, Ph.D., Chairman Rong Chen, Ph.D. Brian McCool, Ph.D. Steven Childers, Ph.D
2 ii DEDICATION AND ACKNOWLEDGEMENTS First and foremost, I want to thank my advisor, Sara R. Jones, Ph. D. She gave me the freedom to explore many of my own interests while still providing important and insightful input to improve my projects and my writing. She gave her full support to my training at all times and helped me to develop as both a scientist and a person. I truly enjoyed working with her and admire her work ethic and dedication. The example that she has set for me will have a lasting impact beyond my scientific career. Second, I would like to thank the members of the Jones lab. I could not possibly thank them enough for all of the help and support that they have given me over the years. Mark J. Ferris, Ph. D, was instrumental in my training and often put as much effort into my projects as he did into his own. He was always there when I needed advice and although I was difficult to work with at times he always supported me. Jordan Yorgason was there with me every step of the way and helped me figure everything out, even when it was inconvenient for him. Jordan has the ability to fix anything that is broken, and I could not have done any of my work without his programming, soldering, or electronics skills. Mark and Jordan have become colleagues and friends (whether they like it or not) that I hope to continue to work with throughout my career. I would also like to thank Joanne Konstantopoulos and Jason Locke for their training and help with many of my projects. Without their willingness to cover for me, even on weekends, none of this work would have been possible. Also, I would like to thank the other members of the Jones lab, Anushree N. Karkhanis, Ph. D, Jamie Rose, James Melchior, and Cody Siciliano. Because of them, I enjoyed coming to work every day. The lab has been like my second
3 iii family, first at times, and there is not anywhere else where I could have had a better graduate experience. I would also like to thank the members of my dissertation committee Drs. Martin, McCool, Chen, and Childers. They have been a source of advice and support throughout my graduate career and played a large role in the development of this dissertation. Additionally, their comments on my proposal contributed greatly to the National Research Service Award that I was awarded from the National Institute on Drug Abuse, and for that I am very grateful. Special thanks go to the Department of Physiology and Pharmacology and the Neuroscience program. The people within this department contributed greatly to this dissertation in many ways. The collaborative environment at Wake Forest University School of Medicine allowed me to learn numerous techniques and skills, many of which are included within this dissertation. I was able to form a network of collaborators that I will keep in touch with throughout my career. Additionally, the advice and support that I was given when applying for grants and postdoctoral positions was paramount to my success. I would also like to thank my family. My mother, Ellen, and my father, John, have given me their full support throughout my entire life, but especially throughout my graduate career. They were constant sources of encouragement that kept me going during difficult times. Also, I would like to thank my sister, Megan, and brother, Brad. They are two of the smartest people that I know and they have motivated me to work hard and dedicate myself completely to anything and everything that I do.
4 iv And finally, I would like to thank the extracurricular groups and activities I was a part of in my spare time. The Phys/Pharm softball team allowed me to display my athletic prowess and make friends within the department. It was a consistent time each week for me to spend time with Alison Arter and Sarah Kromrey, and for that I am particularly grateful, although it is unlikely that anyone else is.
5 v TABLE OF CONTENTS LIST OF TABLES AND FIGURES... viii LIST OF ABBREVIATIONS... xiv ABSTRACT... xviii PREFACE... xxi Chapters CHAPTER I NEUROCHEMICAL AND BEHAVIORAL CONSEQUENCES OF METHYLPHENIDATE (MPH) SELF-ADMINISTRATION: MPH IS UNIQUE IN ITS ACTIONS AT THE DOPAMINE TRANSPORTER... 1 Manuscript prepared for submission in Brain Research Reviews, 2013 CHAPTER II EXTENDED ACCESS COCAINE SELF-ADMINISTRATION RESULTS IN TOLERANCE TO THE DOPAMINE-ELEVATING AND LOCOMOTOR STIMULATING EFFECTS OF COCAINE Published in The Journal of Neurochemistry, in press, 2013 CHAPTER III WITHDRAWAL FROM EXTENDED-ACCESS COCAINE SELF- ADMINISTRATION RESULTS IN DYSREGULATED FUNCTIONAL ACTIVITY AND ALTERED LOCOMOTOR ACTIVITY IN RATS Published in European Journal of Neuroscience, in press, 2013 CHAPTER IV TEMPORAL PATTERN OF COCAINE INTAKE DETERMINES TOLERANCE VS SENSITIZATION OF COCAINE EFFECTS AT THE DOPAMINE TRANSPORTER Published in Neuropsychopharmacology, in press, 2013 CHAPTER V METHYLPHENIDATE AND COCAINE SELF-ADMINISTRATION PRODUCE DISTINCT DOPAMINE TERMINAL ALTERATIONS
6 vi Published in Addiction Biology, in press, 2013 CHAPTER VI METHYLPHENIDATE AMPLIFIES THE POTENCY AND REINFORCING EFFECTS OF AMPHETAMINES BY INCREASING DOPAMINE TRASNPORTER EXPRESSION Published in Nature Communications, in press, 2013 CHAPTER VII UNIQUE PROFILE OF METHYLPHENIDATE-INDUCED DOPAMINE SYSTEM KINETICS: RELEASE PROFILE OF A BLOCKER AND UPTAKE PROFILE OF A RELEASER Manuscript prepared for submission in PLoS One CHAPTER VIII INTERMITTENT ACCESS METHYLPHENIDATE SELF-ADMINISTRATION RESULTS IN A SENSITIZED DOPAMINE SYSTEM Manuscript prepared for submission in Neuroscience, 2013 CHAPTER IX DISCUSSION APPENDIX I CONSERVED DORSAL-VENTRAL GRADIENT OF DOPAMINE RELEASE AND UPTAKE RATE IN MICE, RATS, AND RHESUS MACAQUES Published in Neurochemistry International, 61(7):986-9, 2012 APPENDIX II AMPHETAMINE MECHANISMS AND ACTIONS AT THE DOPAMINE TERMINAL REVISITED Published in The Journal of Neuroscience, 33(21):8923-5, 2013 APPENDIX III AMERICAN CHEMICAL SOCIETY: COPYRIGHT CLEARANCE FORM
7 vii APPENDIX IV NEUROPSYCHOPHARMACOLOGY: COPYRIGHT CLEARANCE FORM APPENDIX V ADDICTION BIOLOGY: COPYRIGHT CLEARANCE FORM APPENDIX VI NEUROCHEMISTRY INTERNATIONAL: COPYRIGHT CLEARANCE FORM CHAPTER X CURRICULUM VITA
8 viii LIST OF TABLES AND FIGURES TABLES Chapter III 1 Effect of 48 h withdrawal from a five-day binge cocaine selfadministration on rates of local cerebral glucose utilization in rat brain...99 FIGURES Chapter I 1 Animals acquire methylphenidate (MPH) self-administration at a faster rate than cocaine (COC) self-administration Methylphenidate (MPH) self-administration increases the preferred intake level and reinforcing efficacy of MPH 18 3 Increased dopamine transporter (DAT) levels increase the potency of psychostimulant releasers..42 Chapter II 1 Cocaine self-administration results in escalation of first-hour cocaine intake.67 2 Reduced dopamine (DA) release and uptake following cocaine self-administration.68 3 Cocaine self-administration results in tolerance to the dopamine transporter (DAT)-inhibiting and dopamine (DA)-elevating effects of cocaine Cocaine self-administration results in tolerance to the locomotor
9 ix activating effects of acute cocaine administration.73 Chapter III 1 Cocaine self-administration results in escalation of rate of cocaine intake Cocaine self-administration results in reduced locomotor response to a novel environment Cocaine self-administration does not alter baseline locomotion Altered locomotor activity following cocaine self- administration 97 5 Cocaine self-administration results in reduced functional activity in a number of brain regions..98 Chapter IV 1 Intermittent (IntA), long-access (LgA), and short access (ShA) cocaine self-administration result in different patterns of selfadministration Differential effects of intermittent (IntA), short-access (ShA) and long-access (LgA) self-administration on presynaptic dopamine system kinetics Intermittent access (IntA) self-administration results in sensitization to the neurochemical effects of cocaine while longaccess (LgA) results in tolerance.130
10 x 4 Supplementary Figure 1, Intermittent access (IntA) selfadministration results increased potency, while long-access (LgA) results in decreased potency versus short-access (ShA) Cocaine-induced increases in DA did not differ across groups Chapter V 1 Escalation in rate of cocaine and methylphenidate self- administration Baseline dopamine system kinetics following methylphenidate and cocaine self-administration Western blot hybridization for the dopamine transporter after cocaine or methylphenidate self-administration Tissue content of dopamine and metabolites in the striatum of cocaine and methylphenidate self-administration groups Quantification of tyrosine hydroxylase mrna levels, protein levels, and phosphorylation by western blot hybridization and quantitative polymerase chain reaction Significantly greater dopamine uptake inhibition by methylphenidate, but not cocaine after methylphenidate selfadministration Significant reduction in dopamine uptake inhibition by cocaine, but not methylphenidate, after cocaine self-administration 171
11 xi Chapter VI 1 Methylphenidate (MPH) is self-administered, increases dopamine uptake (V max ), and MPH potency Methylphenidate (MPH) self-administration (SA) increases the potency and motivation to administer releasers, but not blockers Overexpression of the dopamine transporter (DAT) increases the behavioral and neurochemical potency of methylphenidate (MPH) and releasers Increased releaser, but not blocker, potency following methylphenidate (MPH) self-administration Methylphenidate (MPH) self-administration increases motivational effects of releasers and MPH, but not blockers Increased releaser and methylphenidate (MPH), but not blockerinduced DA overflow following MPH self-administration and dopamine transporter (DAT) overexpression Oral methylphenidate (MPH) administration does not affect DA kinetics or the potency of psychostimulants 205 Chapter VII 1 Maximal rates of dopamine (DA) uptake (V max ) are positively correlated with psychostimulant potency Increases in dopamine transporter (DAT) levels by
12 xii methylphenidate (MPH) self-administration result in enhanced potency of releasers and MPH, but not blockers Although maximal rate of dopamine uptake (V max ) and release are positively correlated in DAT-tg mice, dopamine release is not correlated with drug potency Dopamine release is not correlated with psychostimulant potency in methylphenidate (MPH) self-administration animals Drug-induced dopamine release does not affect dopamine uptake inhibition following methylphenidate (MPH) self-administration or DAT over-expression Methylphenidate (MPH) has the release profile of a blocker..238 Chapter VIII 1 Animals self-administer methylphenidate (MPH) at high rates when given intermittent access Intermittent methylphenidate (MPH) self-administration leads to increased stimulated dopamine (DA) release and uptake Intermittent methylphenidate (MPH) self-administration causes D2 autoreceptor subsensitivity Sensitization of the neurochemical effects of methylphenidate (MPH)..264
13 xiii 5 Enhanced amphetamine (AMPH) potency following methylphenidate (MPH) self-administration Cocaine potency is unaffected by a history of methylphenidate (MPH) self-administration Stimulated DA release profile of cocaine, methylphenidate (MPH), and amphetamine (AMPH) following intermittent MPH selfadministration..272 Chapter X 1 Representative voltammetric dopamine traces highlighting the effects of cocaine, amphetamine (AMPH), and methylphenidate (MPH) self-administration (SA) on cocaine potency 288 Appendix I 1 Location of voltammetric recordings in striatal slices from rat, mouse, and rhesus macaque Dopamine release from five striatal regions in monkey, rat, and mouse Maximal dopamine uptake rate as measured by fast scan cyclic voltammetry from dorsal striatum through ventral striatum in each species Ratio of dopamine release to uptake differs across species.336
14 xiv LIST OF ABBREVIATIONS [DA]p, dopamine per pulse 2-DG, [ 14 C]-2-deoxyglucose ACC, anterior cingulate cortex acsf, artificial cerebrospinal fluid ADHD, attention deficit/hyperactivity disorder Akt, protein kinase B AMPA, 2-amino-3-(3-hydroxy-5-methyl-isoxazol-4-yl) propanoic acid AMPH, amphetamine ANOVA, analysis of variance App. K m, apparent affinity of drug for the dopamine transporter CFT, 2 beta-carbomethoxy-3-beta-(4-fluorophenyl)-n-[3h]methyltropane COC, cocaine CPP, conditioned place preference CPu, caudate putamen C-terminal, carboxy terminal Ctrl, control DA, dopamine DAT, dopamine transporter DAT-KO, dopamine transporter knock-out DAT-tg, dopamine transporter overexpressing mice DC, dorsolateral caudate putamen ERK, extracellular signal-regulated kinases
15 xv FSCV, fast scan cyclic voltammetry GABA, Gamma-Aminobutryic Acid hdat, human dopamine transporter HEK, human embryonic kidney cells IM, intramuscular IntA, intermittent-access IP, intraperitoneal IV, intravenous K i, inhibition constant K m, Michaelis-Menten constant LCGU, local cerebral glucose utilization LgA, long-access MAPK, mitogen-activated protein kinases MC, medial caudate putamen mepsc, miniature excitatory post-synaptic current METH, methamphetamine MPH, methylphenidate mrna, messenger ribonucleic acid NAc, nucleus accumbens Na(+)/Cl(-)-dependent transporters, neurotransmitter-sodium-symporter NET, norepinephrine transporter NMDA, N-methyl-D-aspartate NOM, nomifensine
16 xvi N-terminal, amino terminal PCR, polymerase chain reaction PFC, prefrontal cortex PI3k, phosphoinositide 3-kinase PKA, protein kinase A PKC, protein kinase C P max, maximal price paid PR, progressive ratio PTSD, posttraumatic stress disorder PVDF, Polyvinylidene fluoride SA, self-administration SAMHSA, The Substance Abuse and Methal Health Services Administration S.E.M., standard error of the mean SERT, serotonin transporter ShA, short-access SHR, spontaneously hypertensive rat SLC6, solute carrier six family of transporters SUD, substance use disorder TH, threshold procedure TH, tyrosine hydroxylase Thr, threonine VM, ventromedial caudate putamen VMAT2, neuronal vesicular monoamine transporter
17 xvii V max, maximal rate of uptake VTA, ventral tegmental area
18 xviii ABSTRACT Calipari, Erin S. UNIQUE PHARMACODYNAMIC PROFILE OF METHYLPHENIDATE: SELF- ADMINISTRATION-INDUCED DOPAMINE SYSTEM ALTERATIONS Dissertation under the direction of Sara R. Jones, Ph.D., Tenured Professor of Physiology and Pharmacology Methylphenidate (MPH), the active compound in the medication Ritalin, is commonly diverted for off-label use. It is abused at high rates in both the adolescent and adult population; however, until recently, the neurochemical effects of MPH abuse had not been explored in depth. Pharmacologically MPH is a dopamine transporter (DAT) blocker with the same mechanism of action and equivalent potency at the DAT as cocaine. Although MPH is categorized as a DAT blocker, its pharmacological actions are distinct from both blockers and releasers in regards to both its acute effects and the compensatory alterations associated with self-administration of the compound. This work has elucidated the neurochemical consequences of MPH self-administration, whereby MPH administration by a number of paradigms results in consistent neurochemical adaptations characterized by increased maximal rates of uptake (V max ), increased DAT levels, increased evoked dopamine (DA) release, and an increased potency to inhibit the DAT for MPH and releasers, but not blockers. These changes are opposite from the documented changes that occur following cocaine self-administration, further suggesting that the neurochemical consequences of MPH are unique, and cannot be predicted based on previous literature of compounds of the same psychostimulant class, such as cocaine. The changes directly at the DAT following MPH selfadministration are consistent with microdialysis and behavioral analysis that demonstrate increased MPH and amphetamine (AMPH)-induced DA overflow, but not cocaine, and
19 xix increased reinforcing efficacy of AMPH and MPH, but not cocaine. The increases in the reinforcing efficacy of MPH and releasers such as AMPH, suggests that these compounds may having significant abuse liability following MPH abuse in the adolescent and adult populations. Further, this work has determined the neurochemical mechanism underlying the effects of MPH self-administration. This work shows that MPH self-administration increases DAT levels, driving enhanced potency and drug-seeking for MPH and AMPH, without altering cocaine potency. To corroborate these effects, genetic over-expression of the DAT increased AMPH and MPH potencies, but not cocaine potency as measured by voltammetry, microdialysis, and locomotor analysis. Utilizing transgenic DAT overexpressing mice to study changes in psychostimulant potency, this work has determined a basic mechanism that may predict vulnerability to abuse amphetamines and other dopamine releasers. Namely, increases in dopamine transporter levels are responsible for the increases in AMPH and DA releaser potency. This is important because it suggests that known variability in human DAT expression, particularly in disorders such as attention deficit/hyperactivity disorder (ADHD), post traumatic stress disorder (PTSD), and early life stress, may serve as a marker for vulnerability for substance abuse disorder / addiction. Because these drugs are readily available and are used commonly in the clinic, these results should be considered when treating at risk individuals. Importantly, the effects of MPH resemble those of amphetamines in regards to the effects of DAT levels on its potency, whereas other traditional blockers are unaffected. This is particularly important when making inferences about the consequences of MPH
20 xx administration, both therapeutic and abuse relevant, from previous work using other compounds of the same class. Importantly, this work suggests a revision of the current two-category model for psychostimulants beyond the classification as purely blocker or releaser, as compounds such as MPH can exhibit properties of both classes.
21 xxi PREFACE This thesis contains work on the neurochemical consequences of methylphenidate abuse as it relates to the larger body of cocaine abuse literature. This thesis was prepared by Erin S. Calipari as a partial fulfillment of the requirements for the degree of doctor of philosophy. This thesis was made under supervision of Sara R. Jones. The works contained within have been either submitted for publication or published previously, and are republished with permission. Publications have been reformatted to fit the guidelines of thesis preparation at Wake Forest School of Medicine. Stylistic variations within the chapters are due to specific requirements for each journal.
22 1 CHAPTER I NEUROCHEMICAL AND BEHAVIORAL CONSEQUENCES OF METHYLPHENIDATE (MPH) SELF-ADMINISTRATION: MPH IS UNIQUE IN ITS ACTIONS AT THE DOPAMINE TRANSPORTER Erin S. Calipari, Mark J. Ferris, and Sara R. Jones The following manuscript is prepared for submission at Brain Research Reviews. Stylistic variations are due to the requirements of the journal. Erin S. Calipari prepared the manuscript. Drs. Mark J. Ferris and Dr. Sara R. Jones acted in an advisory and editorial capacity.
23 2 Abstract: Methylphenidate (MPH), the active compound in the medication Ritalin, is commonly diverted for off-label use. It is abused at high rates in both the adolescent and adult population; however, until recently, the neurochemical effects of MPH abuse had not been explored in depth. Here we outline the pre-clinical literature on MPH abuse focusing on rodent self-administration models and the behavioral and neurochemical consequences of MPH self-administration as they relate to the dopamine system. Pharmacologically, MPH is a dopamine transporter (DAT) blocker with the same mechanism of action and roughly equivalent potency at the DAT as cocaine. Although MPH is categorized as a DAT blocker, its pharmacological actions are distinct from both blockers and releasers in regards to both its acute effects and the compensatory alterations associated with its self-administration. Importantly, MPH is affected similarly to amphetamines in regards to the effects of DAT levels on its potency, whereas other traditional blockers are not. This is particularly important when making inferences about the consequences of MPH administration, which are different from previous findings using other compounds of the same class.
24 3 1. Introduction: 1.1. Methylphenidate (MPH) use and abuse After marijuana, prescription stimulants are the most commonly abused drugs that are used to get high in teenagers (Johnston et al., 2009, 2010). Methylphenidate (MPH), the active ingredient in medications (Ritalin and Methylin ), used in the treatment of attention deficit/hyperactivity disorder (ADHD) and narcolepsy, is prone to diversion for non-medical use. In the adolescent and adult populations, MPH abuse in non-adhd individuals has become increasingly prevalent, with oral, intranasal or intravenous (i.v.) routes of administration being most common. 5-17% of college students are estimated to take 2-10 times the prescribed doses (Teter et al, 2003, 2005, 2006, 2010; McCabe et al, 2004, 2005; Barrett et al, 2005; Klein-Schwartz, 2002; Sepúlveda et al., 2011; Fone et al., 2005). Recently, increases in MPH prescriptions, and as a result, increases in both acute and chronic MPH abuse, have been documented (Setlik et al, 2009). The drug is used for cognitive enhancement and to get high, causing effects indistinguishable from cocaine when taken i.v. or intranasally (Morton and Stockton, 2000; Sherman et al, 1987). The emergence of intravenous (i.v.) MPH use is outlined by a number of case studies; however, currently the neurochemical and behavioral effects of i.v. MPH use are understudied (Gautschi & Zellweger, 2006; Levine et al, 1986; Parran & Jasinski, 1991; Sherman et al, 1987; Shaw et al, 2008; Teter et al, 2006). The prevalence of MPH use as a therapeutic agent in adolescents coupled with the high rates of abuse makes it critical to understand the pharmacology of the compound.
25 Relevance MPH is a unique drug that exerts its rewarding and reinforcing effects via increasing dopamine (DA) levels in brain regions such as the dorsal and ventral striatum. Although MPH is categorized as a DAT blocker, it has a unique profile that is not like other psychostimulants in regards to both its acute and chronic effects. Because of MPH s unique pharmacological profile, it is difficult to make predictions about the neurochemical consequences of MPH use and abuse based on the current literature describing the neurochemical effects of other compounds of the same structural or functional class. Currently, there are a plethora of reviews on the therapeutic effects of MPH as it relates to ADHD or the treatment of cocaine dependence (Mariani and Levin, 2013; Grunblatt et al., 2013; Morris et al., 2012; Prommer 2012; Del Campo et al., 2011); however, there is no review on the effects of abused MPH on the DA system and related behaviors. This review will comprehensively outline the current pre-clinical studies on the behavioral and neurochemical consequences of MPH administration as a model of abuse as it relates to the DA system. 2. MPH use, abuse, and addiction potential 2.1. MPH may facilitate the development of a substance use disorder Some suggest that MPH use may facilitate the development of a substance use disorder (SUD) (Lambert and Hartsough, 1998), while others suggest that treatment of ADHD sufferers with stimulants during childhood and adolescence decreases the probability of subsequent psychostimulant abuse (Willens, 2004). However, the studies
26 5 which suggest that therapeutic intervention with MPH reduces the likelihood of future drug abuse often fail to take into consideration the role of increased academic and social success in treated vs. untreated ADHD sufferers, which may be protective against later stimulant abuse (Biederman, 2003; Wilens et al, 2003; Mannuzza et al, 2003). However, although individuals may be less likely to seek drugs due to improved social interactions, self-esteem, and scholastic performance, it is possible that neurochemical changes following chronic low dose MPH treatment may increase the abuse potential of MPH and other psychostimulants. Even with the improvement of a variety of social factors, there still may be a subset of individuals treated with MPH who may be susceptible to addiction if they are in an environment where drugs are readily available. Congruent with human studies, literature examining the effects of long-term MPH exposure in rodents is inconsistent, prompting the need to determine the effects of multiple MPH dosing paradigms on neurochemical outcomes. Brandon et al (2001) showed that MPH administration during adolescence augments the reinforcing properties of other psychostimulants, as seen by increases in cocaine self-administration in adulthood. However a larger body of preclinical literature suggests that therapeutic MPH administration does not ehance the addiction potential of psychostimulants such as cocaine. Carlezon et al (2003) found that chronic adolescent MPH administration in rats caused low doses of cocaine to be aversive and high doses less rewarding in adulthood. Additionally, a number of epidemiological studies as well as rodent studies have shown that oral MPH administration at therapeutically relevant doses does not result in changes to the DA system or to the reinforcing effects of psychostimulants (Calipari et al., 2013a; Soto et al., 2012; Gill et al., 2012, 2013; Carlezon et al., 2003).
27 6 Even though MPH is used in the adolescent population for the treatment of ADHD, the abuse of MPH by normal individuals may be a greater cause for concern. When abused, individuals take 2-10 times the prescribed doses, as well as often administering the compound intranasally or intravenously (i.v.), which may result in different neurobiological alterations as compared to low-dose therapeutic usage. Many studies and reviews have outlined the behavioral and neurochemical consequences of therapeutically relevant dosing of MPH; however fewer studies have focused on the effects of high-dose MPH abuse in non-adhd models (Calipari et al., 2013a, b; Wooters & Bardo, 2011; Wooters et al., 2011). Because of epidemiological reports that indicate that MPH administration does not result in increased abuse of psychostimulants, many people believe that MPH does not have abuse potential and is safe. However, there is increasing evidence that MPH abuse is occurring at high rates (Teter et al, 2003, 2005, 2006, 2010; McCabe et al, 2004, 2005; Barrett et al, 2005; Klein-Schwartz, 2002; Sepúlveda et al., 2011; Fone et al., 2005) and may have significant adverse neurobiological consequences when administered at high doses (Calipari et al., 2013a, b; Marusich et al, 2010; Botly et al, 2008; Burton et al, 2010; Collins et al, 1984) Differential effects of therapeutic and abuse relevant dosing of MPH When determining the neurochemical effects of MPH, an important distinction must be made between abuse and therapeutic dosing, due to factors such as contingent versus non-contingent administration and route of administration. The route of administration, which dictates the speed of the onset of drug effects, is the most important
28 7 factor when considering the abuse potential of drugs as different routes of administration can cause vastly different subjective effects (Volkow et al., 1999). The ability of psychostimulants such as MPH and cocaine to elicit a high is associated with their ability to increase DA in striatal sub-regions (Volkow et al., 1999). These drugs act by blocking the DAT, which is the protein responsible for clearing DA from the synapse and terminating DA signals. DAT blockade leads to elevated DA levels as well as a prolonged half-life of DA within the extracellular space, which results in increased signaling of postsynaptic receptors and enhanced behavioral outputs. It has been demonstrated that the rate at which a drug enters the brain and inhibits the DAT to increase DA levels is the primary factor associated with the perception of high, and not just the ability to increase striatal DA levels alone. Volkow et al (1999) demonstrate that while the level of DAT inhibition is important to determine the intensity of the high, where greater than 50% inhibition is needed to produce high, the rate at which the transporters are occupied determines whether the high is perceived or not. This is particularly important when thinking about therapeutic administration, as even when more than 60% of DATs are occupied by drug, if the rate at which they are occupied is slow there is no perception of high. Thus, it is likely that in therapeutic cases, MPH not only does not result in a high, but also results in different neurochemical alterations. Because of this, it is important not to use therapeutic studies to predict the neurochemical consequences of MPH abuse.
29 MPH Use in ADHD and normal individuals Although MPH abuse in all individuals is of concern, MPH use and abuse may have differential effects in non-adhd and ADHD sufferers. Sakrikar et al., 2012 found that a particular coding variant in the DAT which is prevalent in the ADHD population results not only in differences in normal DAT trafficking but also in differential responsivity to psychostimulant drugs such as AMPH. Because MPH has been shown in a number of voltammetry studies to exhibit the pharmacological properties of releasers (Ferris et al., 2012; Calipari et al., 2013a), it is possible that this mutation also results in differential sensitivity to the effects of MPH in the different populations. This is particularly important, because it suggests that there may be significant differences in the consequences of MPH use/abuse on the DA system in normal populations and ADHD sufferers. Further, this may suggest that while MPH is safe and effective in the ADHD population, it may have neurochemical and behavioral effects that promote the abuse of other psychostimulants in normal people. Additionally, imaging studies using a radiolabeled antagonist of the DAT have shown that DAT binding is higher in subjects with ADHD and this elevation is decreased to control levels after subjects are treated with methylphenidate (Dougherty et al., 1999; Dresel et al., 2000; Krause et al., 2000; Cheon et al., 2003). This further highlights the importance of choosing a relevant model to study the effects of MPH abuse, as ADHD individuals are likely affected differently by MPH administration than normal individuals.
30 MPH has different effects in animal models of ADHD: implications for human use Consistent with the differences in DA system function and psychostimulant effects between ADHD and non-adhd humans, rodent models have shown similar disparities making it critical to use models that accurately mimic the target human population for translational studies. It has been shown repeatedly that different strains of rats exhibit differential acute behavioral activating effects of MPH as well as differential self-administration behaviors (Dafny & Yang, 2006; Yang et al., 2003; Amini et al., 2004; dela Peña et al., 2012). The spontaneously hypertensive rat (SHR) is a widely accepted model of ADHD, and exhibits much of the inattentive phenotype that is observe in ADHD individuals. These animals have different responses to self-administration in general, where SHR have increased acquisition and responding for sucrose reinforcement than both their inbred counter parts (Wistar-Kyoto) and out-bred control strains (Sprague- Dawley) (Marusich et al., 2011). In addition, MPH self-administration has differential effects in the strains, where prior MPH exposure enhanced fixed ratio 3 MPH selfadministration in Wistar rats, but not SHR. Further, MPH pretreatment reduced progressive ratio responding in SHR, but not Wistar rats (dela Peña et al., 2012). Reinstatement of drug seeking is a measure that is used to model relapse in rodents, and interestingly, MPH exposure greatly increases reinstated responding in Wistar rats as compared to SHR (dela Peña et al., 2012). Taken together, these data suggest that the SHR rats (ADHD model) may be less susceptible to the rewarding and reinforcing effects of MPH, where MPH treatment either does not affect or reduces responding depending on paradigm. Although there are clear differences in responses to MPH between ADHD
31 10 and non-adhd models, the main focus of this review will be on non-adhd models because MPH abuse most often occurs in normal, non-adhd individuals. In addition to the differential behavioral effects, the neurochemical effects of MPH are also divergent between SHR and other rats. Russel et al., (1998) demonstrated that SHR have reduced stimulated DA release in both prefrontal and striatal slices, as well as significantly reduced MPH potency in SHR animals as compared to Wistar-Kyoto controls. Roessner et al (2010) demonstrated that, at baseline, SHR have increased DAT levels as compared to Wistar-Kyoto and Wistar rats, a difference that was reversed by MPH treatment. Although the differential DA system dynamics, reinforcement behaviors, and responses to MPH treatment in SHR compared to out-bred strains make SHR a good model for studying the therapeutic use of the MPH, SHR rats are likely not the optimal strain for modeling MPH abuse. This review will focus on the effects of MPH abuse on non-adhd models in order to model MPH abuse in the non-adhd human population Subjective effects Growing evidence suggests that MPH has abuse potential, especially via non-oral routes, which produce subjective effects indistinguishable from those of cocaine (Rush and Baker, 2001; Setlik et al, 2009; Teter et al, 2006). In discrimination experiments, MPH produces full substitution for both cocaine and amphetamine (AMPH) (Rosen et al, 1985; Silverman and Ho, 1980), indicating that the subjective effects of these compounds are indistinguishable. These behavioral similarities are likely due to their shared ability
32 11 to acutely elevate extracellular DA levels in reward related brain areas, resulting in similar levels of activation of post-synaptic receptors. The indistinguishable subjective effects suggest that MPH may be administered in place of stimulants such as AMPH and cocaine, as prescription drugs are becoming easier to obtain than illicit drugs, and because people view these drugs as much safer. In addition, a number of measures such as tolerance, withdrawal and sensitization are associated with drugs that may have abuse/addiction potential such as cocaine and AMPH. A number of reports using a range of MPH doses have demonstrated all of these phenomena, suggesting that MPH may pose significant abuse potential and possibly addiction potential when taken via similar routes of administration as cocaine and AMPH (Eckermann et al., 2001; Yang et al., 2000, 2007a, b; Lee et al., 2008; Gaytan et al., 1997). Additionally, comparison studies with other psychostimulants such as cocaine suggest that MPH may have serious abuse, and possibly addiction potential. Comparing MPH and cocaine at the most reinforcing doses, which produce similar selfadministration behavior (Figure 1), animals acquire MPH self-administration faster than cocaine. In the cocaine literature, faster acquisition rates have been associated with animals that are drug-use prone (Deroche-Gamonet et al., 2004, 2008; Belin et al., 2009, 2011), suggesting that MPH may have significant abuse liability. Currently, in the human population cocaine and MPH are often taken via different routes of administration, with cocaine being primarily intranasal, intravenous, or smoked and MPH being primarily oral or intranasal (Teter et al., 2006). However, reports of intravenous use of MPH are emerging, suggesting that MPH abusers are transitioning to routes of
33 12
34 13 Figure 1. Animals Acquire Methylphenidate (MPH) Self-Administration at Faster Rates than Cocaine (COC) Self-Administration. (A) Representative traces demonstrating that MPH (0.56mg/kg/inj) and cocaine (1.5mg/kg/inj) result in similar self-administration behavior. Tick marks represent infusions earned on a fixed ratio one schedule of reinforcement. Adapted from Calipari et al., 2013, Addiction Biology. (B) Escalation of rate of intake does not differ between cocaine and MPH demonstrating that 1.5mg/kg/inj cocaine and 0.56 mg/kg/inj MPH produce comparable self-administration behavior. Adapted from Calipari et al., 2013, Addiction Biology. (C) Group data plotting average time (in sessions) to reach acquisition criteria (>35 injections) with behaviorally equivalent doses of MPH and cocaine. On average, it takes fewer sessions for animals to acquire MPH self-administration as compared to cocaine (D) Group data plotting the percentage of animals that have met acquisition criteria over sessions.
35 14 administration comparable to cocaine (Gautschi & Zellweger, 2006; Levine et al, 1986; Parran & Jasinski, 1991; Sherman et al, 1987; Shaw et al, 2008). Conditioned place preference (CPP), which is often used as a measure of the rewarding effects of drugs is produced by MPH in both rats and mice (Chen et al., 2006; Wooters, Walton and Bardo, 2012). In rats, conditioned preference for MPH occurred when it was administered (intraperitoneal) i.p. (3 mg/kg); however when administered orally MPH did not result in preference for the drug-paired chamber (Wooters, Walton, and Bardo, 2012). The lack of CPP using therapeutic routes of administration (oral) in rodents is consistent with the human literature demonstrating that when administered orally at therapeutic doses, that human subjects do not perceive a high. Alternatively, in rodents, i.p. dosing caused preference indicating that the route of administration is an important factor in influencing the subjective effects associated with a particular drug. This is particularly important to note when considering the clinical implications of the MPH abuse literature as compared to the therapeutic literature and highlights the importance of using relevant routes of administration in order to model the high associated with MPH abuse. 3. Behavioral models of MPH abuse 3.1. Self-administration The gold standard for preclinical drug abuse research is self-administration, where animals respond on a lever in order to contingently administer drug. Most drugs that are abused in the human population are readily self-administered by rodents, and a
36 15 number of studies have demonstrated that MPH maintains i.v. self-administration in rats (Marusich et al, 2010; Botly et al, 2008; Burton et al, 2010; Collins et al, 1984). Further, MPH supports high breakpoints on a progressive ratio (PR) schedule, which are comparable to cocaine, in both rats and humans that are (Botly et al, 2008; Rush et al, 2001). These studies indicate that MPH is a potent reinforcer, especially when taken i.v., and given that most, if not all, drugs of abuse are self-administered by rodents, there is some cause for concern for MPH abuse in the human population. Giving further support to the idea that MPH has significant abuse/ addiction potential is the cue-induced augmentation of MPH self-administration. Associations made between drugs and drug-associated cues have often been implicated in relapse, where cues, in the absence of drugs, can trigger drug seeking and drug taking behavior. Marusich et al. (2011) conducted a study to determine how pairing cues with selfadministration affected self-administration behavior. They found that, in the absence of paired cues, MPH functioned as a reinforcer, however, when delivery of MPH was paired with a cue light, animals pressed significantly more to obtain the drug. This demonstrates that MPH is a reinforcer in the absence of environmental stimuli and cues; however, the interplay between environmental cues and MPH reinforcement is synergistic. Because of this strong enhancement of MPH self-administration in the presence of cues, it is possible that once individuals abstain from MPH for extended periods, the cues may trigger drugseeking behavior and lead to relapse to abuse of the compound. Botly et al., (2008) demonstrated that MPH itself can reinstate extinguished drug seeking; however, it would be interesting to determine if the drug-paired cues were capable of doing so as well.
37 16 In addition to normal baseline responding for MPH under a number of schedules, MPH also results in escalation of fixed ratio 1 (FR1) intake over sessions. This escalation is highlighted by an overall increase in total drug intake over sessions as well as an increase in intake within the first hour of each session (Marusich et al., 2010; Figure 1). An important feature of dependence and addiction is the switch from moderate drug taking to excessive and uncontrolled intake (Koob and Le Moal, 1997). Humans exhibit escalation of intake in extended-access situations (Koob and Kreek, 2007; Gawin and Ellinwood, 1989) and this has been modeled in rats using long access (LgA, 6 hr/day) self-administration sessions. While daily short access sessions (ShA, 1hr/day) result in stable drug intake, it has been repeatedly shown that LgA produces an escalation in intake for a variety of addictive drugs including COC, AMPH, and MPH (Ahmed and Koob, 1998, 1999; Johansen et al, 1976; Marusich et al., 2010). It is well accepted that the phenomenon of escalation is a predictor of abuse/dependence potential of drugs in the human population, which suggests that because MPH escalates that when taken via certain routes it may have serious abuse potential. Marusich et al. (2010) demonstrated that long access MPH resulted in escalation of intake at 0.1 mg/kg/infusion as well as at 0.3 mg/kg/infusion. A short-access group (1hr/session) showed no increase in intake over sessions. This demonstrates that short, controlled access, to MPH, as seen with therapeutic use, does not result in increases in intake over a session, and may not have abuse potential. It is likely that longer access and intake of these drugs, as seen with abuse will more likely result in a dependence/addiction phenotype, which is dangerous, particularly in the adolescent population, where use in this pattern has been observed.
38 MPH escalation is due to increased reinforcing efficacy of MPH Because escalation experiments are done on an FR-1 schedule where one lever response results in one infusion of drug, it is impossible to determine using this paradigm if escalation in intake is due to tolerance or an increased motivation to administer the compound. Koob and colleagues argue that the escalation that occurs with cocaine selfadministration is due to a shift in hedonic set point (Ahmed & Koob, 1998; Koob and Le Moal, 1997; Koob 1996a, b). Because it was only recently determined that MPH results in escalation, little research has focused on the mechanism of such intake. However, using a similar LgA paradigm, Calipari et al. (2013a) demonstrated that the reinforcing efficacy of MPH was increased following LgA self-administration that resulted in first hour escalation, using two independent measures of reinforcing efficacy (Figure 2). As little as five days of extended-access MPH self-administration resulted in increased progressive ratio responding at 0.56mg/kg MPH, the most reinforcing dose of the drug. This alone indicates that the increase in first hour administration of MPH is not due to tolerance, as tolerance would likely result in decreased PR responding as animals are less motivated to work for the compound when it has less of an effect. Further, prior MPH self-administration results in increased responding and maximal price paid for MPH, using a threshold procedure (Zimmer et al., 2012; Oleson et al., 2009, 2012; Figure 2). The within-session threshold procedure is a behavioral economics approach to assessing drug-taking and drug-seeking by performing a within session dose-response curve. The doses are administered in min bins over a session and the doses decrease logarithmically from high to low over the session. Overall shifts in the entire dose-response curve can be determined by plotting the number of
39 18
40 19 Figure 2. Methylphenidate (MPH) Self-Administration Increases the Preferred Intake Level and Reinforcing Efficacy of MPH. MPH reinforcement was assessed using a within session threshold procedure. This procedure is a within session dose response curve where doses start high and are reduced throughout the session (every 15 min). Using this procedure the cost [measured as number of responses required to obtain 1mg MPH (resp/mg)] can be assessed for each dose, and the maximal price (P max ) that an animal is willing to pay to obtain drug can be assessed. (A) Demand curve plotting the average intake at each of the doses administered. At high doses (where cost is low) animals with a history of MPH self-administration administer more MPH. (B) Group data plotted as responses versus dose. At low doses (where cost is high) animals with a history of MPH self-administration expend greater effort to maintain MPH blood levels. Adapted from Calipari et al., 2013, Nature Communications (C) The P max is determined from the peak of the MPH dose-response curve. Boxes highlight the price at which control and MPH self-administration animals respond most for MPH. MPH selfadministration increased P max for MPH. (D) Group data indicating that the P max for MPH is increased following MPH self-administration. Adapted from Calipari et al., 2013, Nature Communications.
41 20 responses that are emitted at each dose, much like a PR dose-response. Second, a measure of the maximal price paid (P max ) for a drug can be determined before or after a treatment. The price is expressed as the number of responses necessary to obtain one mg of drug; thus, as the dose is reduced over the session the price is increased because animals must respond more to maintain the same level of drug self-administration. The P max is the price at which the animal responds most, and is the apex of the dose-response curve. Using this paradigm Calipari et al. (2013a, b) found that, following extendedaccess MPH self-administration, there was a shift in the dose-response curve to indicate that MPH was more potent. In addition, the P max for MPH was significantly increased, indicating that animals were willing to work harder to obtain MPH following selfadministration. Further, when plotting the data as a demand curve (intake versus responses), it can be determined if the animals are titrating their drug intake to a different preferred blood level. Figure 2A demonstrates that, following MPH self-administration animals take more drug when the cost is low (higher doses). This is in line with escalation experiments, which show that on an FR1 schedule, animals administer more drug in subsequent sessions. Taken together, these data demonstrate that extended-access MPH self-administration does not cause behavioral tolerance and that animals are, in fact, more motivated to self-administer following prior MPH exposure. This is particularly alarming as it may promote a cycle of drug use in the human population within which individuals that abuse MPH become more motivated to abuse MPH subsequently, which is a cycle that could potentially lead to addiction of the compound. In addition, it is important to determine the effects of MPH administration, not only on MPH, but also on
42 21 other psychostimulants as they are often used in combination in the adolescent population (Sepúlveda et al., 2011; See section 3. C.) Cross-sensitization Locomotor sensitization experiments are traditionally used to study the neurochemical alterations that are involved in the addiction process by determining the behavioral consequences of repeated psychostimulant administration. Repeated MPH administration results in robust locomotor sensitization (Wooters and Bardo, 2009); this effect is also characteristic of other commonly abused psychostimulants including cocaine and AMPH (Kalivas and Duffy, 1990, 1993; Parsons and Justice, 1993). It has been demonstrated using a number of paradigms that MPH cross-sensitizes with other psychostimulants, suggesting that MPH use/abuse could affect not only itself, but also the propensity for abuse for these other compounds as well. The locomotor effects of MPH cross-sensitize with AMPH, demonstrating that AMPH-induced behavioral effects can be altered by MPH (Yang et al, 2003, 2011; Burton et al, 2010). This is of interest because MPH abusers are more likely to abuse other addictive substances (Barrett et al, 2005; Teter et al, 2006; Wilens et al, 2008). Because sensitization is thought to involve a number of neural circuits including DA and glutamate, this also suggests that MPH exposure may alter other neurotransmitter systems in addition to DA and may have behavioral implications that extend beyond reward and reinforcement (see section ).
43 MPH affects the reinforcing efficacy of other psychostimulants In addition to cross-sensitization, MPH self-administration has been shown to affect the reinforcing efficacy of not only MPH itself, but also other psychostimulants as well. Using two paradigms of self-administration, extended-access (Calipari et al., 2013a, b) and intermittent access (Calipari et al., unpublished observations), the reinforcing efficacy of MPH, a prototypical releaser (AMPH), and a prototypical blocker (cocaine) were determined following self-administration using the threshold procedure of reinforcement, described above. The effects of both self-administration paradigms were similar, with MPH self-administration augmenting only AMPH and MPH, but not cocaine reinforcement (Calipari et al., 2013c). These data demonstrate a number of important concepts. First, MPH is capable of altering the reinforcing efficacy of other psychostimulants when taken via multiple patterns of self-administration, an effect that may lead to increased abuse/addiction of these compounds in the human population following MPH abuse. Second, it demonstrates that MPH can class specifically affect the potency of certain compounds. And finally, that MPH, although considered a DAT blocker, may be unique, as its self-administration affects only a releaser and not another compound of the same class. The unique behavioral properties of MPH may be explained by its acute and long-term neurochemical effects, which seem to be unique in comparison to other psychostimulant blockers and releasers.
44 23 4. Acute neurochemical effects 4.1. The dopamine system Although behavior is of high relevance in determining the abuse potential of drugs, it is also important to understand the neurochemical processes underling a drug s behavioral effects. The DA system plays a critical role in motivated behaviors, goal oriented movement, attention, reward and reinforcement (Carlsson, 1987; Robbins, 2003). Because of this, mutations and dysfunction in the DA system results in a number of pathologies such as Parkinson s disease (Chase et al., 1998), ADHD (Mazei-Robison et al., 2005), and addiction (Ritz et al., 1987). Dopaminergic innervation of the nucleus accumbens (NAc) is an important mediator of motivated reward-related behaviors (Di Chiara, 1995; Peoples et al, 1998; Koob, 1996); accordingly, MPH, AMPH, and COC, elevate DA in the rat NAc and enhance dopaminergic signaling (DiChiara and Imperato, 1988; Carboni et al, 1989; Woods and Meyer, 1991; Gerasimov et al, 2000). MPH, a DA and norepinephrine transporter inhibitor, increases accumbal DA levels via DA uptake inhibition directly at the DAT (Volkow et al, 2002; Heal et al, 2009; Han & Gu 2006). Increased NAc DA levels mediate the reinforcing and rewarding effects of MPH as well as the high associated with MPH abuse. MPH rewarding effects have been directly linked to the DAT in studies, which demonstrate that MPH-induced CPP is abolished in mice with mutations of the DAT which reduce the affinity of MPH (Chen et al, 2005; Tilly & Gu, 2008). Because of the necessary role for the DAT in MPH-induced reward, reinforcement, and therapeutic effects, much of the work on the neurochemical effects of MPH has focused primarily on the DA system.
45 MPH pharmacology Although behaviorally similar, COC, AMPH, and MPH are markedly different in the way in which they acutely interact with the DA system. DAT inhibiting compounds can act by either simply blocking (e.g., cocaine) the transporter or by acting as releasers (e.g. AMPH), however the net acute effect is increased DA levels in the synapse (Rothman and Bauman, 2003). Blockers act by binding to the DAT and inhibiting the uptake of DA from the synaptic cleft, while releasers bind to the DAT and are transported into the terminals. Traditionally, releasers act via three major mechanisms: 1) inhibition of DA uptake by competitively binding to the DA DAT, 2) modulation of the ph with in vesicles resulting in the movement of DA into the cytoplasm, and 3) action potential independent DAT-mediated reverse transport of DA into the synapse (Fleckenstein et al., 2007). Because most releasers must be transported for their effects to occur, blockers can often inhibit the actions of releasers (Rothman and Bauman, 2003; Rudnick and Clark, 1993). MPH is traditionally classified as an uptake blocker, as cell culture studies have demonstrated that it is not transported into the cell via the DAT, and thus cannot interact with vesicles to cause action-potential independent DA release (Sonders et al, 1997). Even though MPH is categorized as a blocker, it has been shown to have releasing capabilities at 0.1 mm, suggesting that MPH may share some properties with both blockers and releasers (Sproson et al, 2001; Russell et al, 1998; Heal et al, 1996; Dyck et al., 1980). Since MPH is not a substrate for the DAT, similar to the prototypical blocker cocaine, yet it has release capabilities at high doses, similar to the prototypical releaser AMPH, MPH may function as an atypical DAT inhibitor, exhibiting pharmacological
46 25 characteristics similar to AMPH as well as cocaine. The exact molecular mechanisms responsible for MPH s unique actions are not yet understood, however. In addition to MPH being unique in its mechanisms of action, binding studies demonstrate that MPH interacts with the DAT in a way that is distinct from both AMPH and cocaine (Wayment et al., 1999; Dar et al., 2005). These studies show that MPH interacts with the transporter in a manner similar to benztropine, as measured by displacement of radiolabled CFT (2 beta-carbomethoxy-3-beta-(4-fluorophenyl)-n- [3H]methyltropane) binding and uptake. In these studies both MPH and benztropine displace radiolabled CFT in a manner that was dissimilar from both blockers and releasers and led the researchers to discuss the interaction of these compounds with the DAT as a separate pharmacological class of psychostimulants (Dar et al., 2005). This demonstrates that, although MPH is traditionally classified as a blocker, similar to cocaine, MPH is different in the way in which it interacts with the transporter. This difference in DAT-MPH interactions may not have an effect on the observed acute effects of MPH, specifically the ability of MPH to elevate DA; however, the differences in binding properties of MPH may lead to, and explain, some of the observed differences in neurochemical adaptations that occur as a result of chronic MPH administration as compared to traditional blockers. MPH has been shown to have multiple effects on presynaptic DA terminals. It mobilizes docked vesicles, causing vesicles to be redistributed from membrane-bound to free pools, which may cause the vesicles to be more likely to be released. A 40 mg/kg subcutaneous dose of MPH one hour prior to synaptosomal preparation results in a redistribution of the vesicular monoamine transporter (VMAT)-2 protein from the
47 26 membrane compartment to the cytosolic compartment. Much work has been done on releasable DA pools, and it is possible that since MPH is capable of redistributing vesicles from membrane-bound forms into the readily releasable pools, it may lead to greater DA release per stimulation. This suggests that although MPH does not enter the cell, it may be able to elevate DA levels in the synapse indirectly, by promoting greater DA release per action potential (Volz et al., 2007, 2009; Sandoval et al., 2002; Chadchankar et al., 2012). In addition to altering vesicle distributions within the terminals, acute MPH injection (40 mg/kg, 1hr prior) results in increased vesicular DA uptake, suggesting that the system is more efficient at repackaging DA, an effect that, like vesicular redistribution, could lead to greater DA release upon each stimulation of the terminal. Not only is uptake changed, but also release, where the speed and magnitude of release from striatal synaptosomes is increased, lending further support to the fact that MPH may increase DA release independent of its DAT effects (Volz et al., 2009). Because of these unique properties of MPH, it is difficult to make predictions about the neurochemical consequences of use and abuse, based on previous work with other psychostimulants. As a result, much work needs to be done, not only to further elucidate the mechanism of acute MPH actions on the DA system, but also to determine the effects of chronic MPH abuse.
48 27 5. Long-term consequences of MPH administration 5.1. Chronic non-contingent effects Because of the role of the DA system in reward and reinforcement, and the DAT s role as the primary site of action for the euphorigenic effects of psychostimulants, much focus has been placed on the effects of chronic exposure of these compounds on the DA system and the DAT. MPH self-administration has previously been shown to be mediated by the DA system, because both D1 and D2 antagonism lead to increased FR1, but decreased PR responding (Botly et al., 2008). These changes are consistent with reduced subjective effects of MPH in the presence of the antagonists, leading to a reduction in reinforcing effects. The same phenomenon is seen with behavioral tolerance, where the reduced effects of a drug lead to an increase in FR1 responding in order to overcome the reduced effects of that drug. Further, animals are not willing to expend as much effort to obtain MPH, which leads to decreases in reinforcing efficacy and a concomitant decrease in PR responding. Studies, such as this, point to the importance of the DA system in the reinforcing effects of MPH. The different interactions with the DA system, more specifically the DAT, are most important in determining the compensatory changes that occur after long-term repeated exposure to the compound. Thus drugs like MPH, which interact with the DAT in a way that is unique from other psychostimulants, must be assessed individually to determine their neurochemical effects. Most work aimed at assessing the neurochemical consequences of MPH has focused on non-contingent administration of MPH in rodent models. The effects of repeated non-contingent MPH administration on DAT function are unclear, as numerous studies using different dosing paradigms have shown different, sometimes
49 28 opposite, effects of exposure. It was initially reported that acute 4-day (21 or 50 mg/kg/day) MPH failed to cause changes in tissue DA levels, metabolites, or transporter numbers (Zaczek et al, 1989). Later studies, however, showed that repeated 7-day (44 mg/kg/day) MPH exposure decreased DAT numbers in the caudate putamen, but not the nucleus accumbens or the substantia nigra (Izenwasser et al, 1999). Moreover, administration of MPH at pre- and post-pubertal stages decreased DA transporter density by 25% at day 45 and 50% at adulthood (day 70; Moll et al, 2001). Further, MPH administration results in increased VMAT-2 immunoreactivity, VMAT-2-mediated DA uptake, and increased DA content in a cytoplasmic vesicular fractions in the striatum, (Volz et al., 2007). Although these studies were important in laying the groundwork for MPH research on the DA system, the results are unclear and often conflicting. In addition, although these studies use high doses that would constitute abused doses of MPH, they are administered non-contingently. Non-contingent drug administration can give us insight into the basic neurochemical effects of drugs, however, in regards to drug abuse, in which the neurochemical consequences are dictated by a number of factors, it is particularly important to model the contingency with which humans administer drugs. A number of studies have demonstrated that contingent and non-contingent administration of drug can have vastly different neurochemical effects, thus when attempting to accurately model the effects of drug abuse it is important to use contingent drug administration.
50 Neurochemical consequences of MPH self-administration Self-administration is an ideal method for determining the neurochemical effects of drug abuse, as animals, like humans, choose to administer drug. Because the administration of drug is contingent, this is currently the best working model of human drug abuse that we have in rodents. Although primate work likely more accurately models human drug use, rodents allow for more in-depth probes of the neurochemical and behavioral consequences of drug abuse. Currently, little work has been done to determine the effects of MPH self-administration on DA system neurochemistry. LgA self-administration has been useful for documenting the neurochemical adaptations that occur during the addiction process, and studies have demonstrated that ShA and LgA paradigms differentially alter DA system function as measured by changes in uptake and DAT density, particularly for drugs such as COC and AMPH (Oleson et al, 2009; Ben- Shahar et al, 2006). Long access self-administration of 0.56 mg/kg/infusion MPH for five days resulted in significant DA system alterations. Extended-access self-administration of MPH results in escalation of first-hour intake, an effect that is thought to signify the switch from abuse to addiction of drugs of abuse (Ahmed & Koob, 1998). In these animals DA system changes were assessed using fast scan cyclic voltammetry (FSCV), an electrochemical technique that allows for real-time recording of DA signals with subsecond time resolution allows for the determination of a number of kinetic parameters. FSCV in slices containing the NAc core from animals that had previously selfadministered MPH exhibited significantly higher stimulated release and maximal rate of uptake (V max ), indicating significant changes in DA signaling within these animals.
51 30 These changes are not due to alterations in tissue content levels of DA, or DA synthesis, as tissue content levels were not elevated following MPH self-administration, and the levels of tyrosine hydroxylase (TH), the rate limiting enzyme for DA synthesis, remained unchanged. In addition to the lack of difference in TH levels, there was also no difference in TH phosphorylation (Calipari et al., 2013b). Therefore, it is likely that changes in release reflect changes in releasable pools of DA, something that MPH has been shown to acutely affect by redistributing vesicular pools from membrane-bound into the readily releasable pool (Volz et al., 2009). Changes in uptake rates in this case were determined to be due to increases in DAT levels, which have previously been shown to alter the potency of psychostimulants (Chen and Reith 2007; Salahpour et al., 2008). Because two major factors that govern normal DA signaling are DA release and reuptake, the changes in these measures alone at baseline indicate that the DA system may be functioning differently in these animals independent of drug effects. Because DA release is increased, it is possible that phasic activity of DA neurons may be increased as well. Because phasic firing of DA neurons is associated with goal-reward learning, it is possible that these changes may affect subsequent drug administration and drug-seeking through an enhancement of goal-reward associations. In addition to assaying the effects of MPH self-administration in an LgA model, we also determined the neurochemical consequences of MPH self-administration using an intermittent-access paradigm that does not result in escalation over sessions. Intermittent-access was used because human administration of psychostimulant drugs is often not continuous, but rather follows a spiking pattern where people take a bolus of drug and wait a period of time before administering the compound again. Within each
52 31 session, animals were given unlimited access to MPH (0.56 mg/kg) for 5 minutes with no timeouts, following a 25-minute time-out period during which the lever is retracted and the animals cannot respond to obtain drug. This pattern of self-administration repeats for the six-hour session, resulting in 12 5-minute access periods. Following 14 days of intermittent MPH self-administration, FSCV was used to determine the DA system kinetics in NAc core containing brain slices from these animals. Surprisingly, this different paradigm of self-administration resulted in the same adaptations to DA system as previous LgA models, characterized by a significant increase in stimulated DA release as well as an increase in uptake rate. The increased DA release is not due to changes in D2 auto-receptors, as the functional activity of the receptors was unchanged (Calipari et al., submitted). The fact that two different MPH self-administration paradigms resulted in the same DA changes, suggests that the effects of contingent MPH administration are the same, regardless of paradigm, however, further research would have to be conducted to determine the validity of that hypothesis. 6. Psychostimulant self-administration-induced changes in drug potencies MPH self-administration effects on psychostimulant potency The use of FSCV for the assessment of DA signaling, allows not only for the determination of DA system kinetics and dynamics, but also for determining the potency of DAT-inhibitor compounds. Using kinetic modeling, we are able to determine the extent to which drugs such as cocaine, AMPH, and MPH inhibit DA uptake, which can give us a measure of drug potency following MPH self-administration (Yorgason et al.,
53 ; Ferris et al., 2013). Because other psychostimulants are often used in combination with MPH, it is important to not only determine the potency of MPH. but other compounds as well (Sepúlveda et al., 2011). Following extended-access MPH selfadministration, MPH potency was increased, which suggests that MPH may be more rewarding / reinforcing. This theory is supported by the behavioral data discussed previously that indicates that the reinforcing efficacy of MPH is also increased; suggesting that changes in psychostimulant potency within the NAc have behavioral correlates (Calipari et al., 2013a). This is important, as the self-administration other of psychostimulant blockers, such as cocaine, has been shown to result in tolerance, not sensitization, to its neurochemical effects at the DAT, characterized by a reduced DA uptake-inhibition as measured by both FSCV and microdialysis (Mateo et al., 2005; Ferris et al., 2011, 2012, 2013; Calipari et al., 2013b, c, d, e). In addition to determining changes in MPH potency following MPH selfadministration, MPH self-administration resulted in an increased potency for the releasers AMPH and methamphetamine (METH), but had no effect on the blockers cocaine and nomifensine. Therefore, MPH self-administration selectively increases the potency of releasers and MPH, and leaves blockers unaffected. Not only was this phenomenon observed following LgA self-administration, but the same trend was observed following intermittent access MPH where AMPH and MPH potency were increased and cocaine potency remained unaffected. This class specific increase in drug potency is important as certain drugs may be more susceptible to abuse in individuals that abuse MPH, while others remain unaffected.
54 33 Although the previous work was done in vitro using fast FSCV, the results have also been validated using in vivo microdialysis techniques. Following MPH selfadministration, as predicted by voltammetry studies, there is an increase in MPH- and AMPH-, but not cocaine-, induced DA overflow, indicating that the effects at the DAT are directly related to the elevations in DA that are observed following the administration of these psychostimulants (Calipari et al., 2013a). Further, the increases in neurochemical potency are accompanied by increases in reinforcement for these compounds as well, where a history of MPH self-administration results in increased reinforcing efficacy for AMPH and MPH, but not cocaine, as measured by using the within session threshold procedure (Described in section 3.1.). The increase in reinforcement for releaser drugs suggests that these compounds may have an increase in abuse liability, which is particularly important given the co-abuse of MPH with other compounds such as amphetamines A mechanism for MPH self-administration effects The changes in psychostimulant potency following MPH self-administration have been shown to be due to increases in DAT levels (Calipari et al., 2013b). This was confirmed using transgenic DAT over-expressing mice (DAT-tg) to show that increases in DAT levels alone, independent of pharmacological manipulations, are capable of causing the same neurochemical and behavioral effects as MPH self-administration (Calipari et al., 2013a). DAT-tg mice show the same kinetic and dynamic changes, characterized by increased evoked DA release and increased uptake rates. Further,
55 34 increasing DAT levels results in increased potency for releasers and MPH, but not blockers. Additionally the increase in MPH and AMPH potency directly at the DAT are accompanied by increased AMPH- and MPH-induced DA overflow and increased locomotor activating effects of the compounds (Salahpour et al., 2008; Calipari et al., 2013a). These data demonstrate that increasing DAT levels is sufficient replicate the effects that are observed following MPH self-administration, validating this as a mechanism for the neurochemical and behavioral effects. Further, this demonstrates that the acute effects of MPH are affected by DAT levels in a similar fashion to releaser compounds, but not blockers, further demonstrating that MPH is unique in regards to its behavioral effects and its interactions with the DA system. Increased potency of releasers and MPH in the presence of high V max /DAT levels may be important in clinical treatment settings, as individuals with ADHD have welldocumented increases in DAT levels ranging from 17-70% (Madras et al., 2005). These findings may explain in part the high frequency of drug abuse in untreated ADHD sufferers (Biederman et al., 1999), and these individuals may be more susceptible to the neurochemical, behavioral, and neurotoxic effects of MPH, AMPH and other releasers like methamphetamine and bath salts. Also, models of PTSD (Hoexter et al., 2012), early life stress (Yorgason et al., 2012), and repeated stress (Kohut et al., 2012) all result in similar increases in DAT levels; therefore, many individuals fall into this category, making the relevance of these results extend far beyond the drug abuse field.
56 The effects of MPH self-administration are different from AMPH and cocaine self-administration Cocaine self-administration Calipari et al., (2013b) demonstrated that although MPH self-administration caused an increase in the measures of DA release and uptake, and an increase in MPH potency, the effects of cocaine self-administration were very different. Cocaine selfadministration resulted in decreased DA release and uptake, effects that were both opposite from MPH self-administration. In addition, the effect of the self-administration of each drug on potency was different, where cocaine self-administration resulted in decreased cocaine potency, but no affect on MPH potency, and MPH self-administration resulted in increased MPH potency, without effecting cocaine potency. The opposite compensatory changes that occur in the DA system following the self-administration of two traditional DAT blockers are surprising, but highlight the unique properties of MPH, not only acutely, but also in long-term self-administration paradigms Amphetamine self-administration It was initially hypothesized that the effects of MPH self-administration were opposite of cocaine because MPH is structurally similar to releasers, thus, its effects would likely resemble the changes that occur following AMPH self-administration. However, MPH effects were different from the previous reports of the effects of AMPH self-administration. AMPH self-administration resulted in no change in baseline DA kinetics or on psychostimulant potency of either blockers or releasers (Ferris et al., 2011).
57 36 This demonstrates that although MPH effects are different from traditional blockers, they are also different from releasers, suggesting that MPH may have properties of both classes of drugs. This further supports the idea that inferences on the effects of MPH on the DA system cannot be made based on previously published work on either blockers or releasers Pattern of self-administration alters the compensatory changes associated with cocaine, but not MPH self-administration Further supporting the idea that the effects of MPH self-administration are unique is data indicating that MPH is unaffected by pattern of self-administration, which is different from the literature on cocaine self-administration. The cocaine literature demonstrates that LgA and intermittent access cocaine administration result in different changes in DA system kinetics and psychostimulant potency (Calipari et al., 2013c, d, e). Specifically, tolerance to the DAT-inhibiting effects of cocaine is dictated by intake pattern, where continuous high-dose cocaine intake results in tolerance to cocaine effects, while intermittent cocaine intake results in sensitization where cocaine is more potent at inhibiting the DAT (Calipari et al., 2013c). This is different from MPH selfadministration, which results in the same DA kinetic and MPH potency effects for both intermittent and continuous access paradigms (Calipari et al., submitted).
58 What is different about MPH? The unique properties of MPH in regards to its chronic effects make it impossible to make generalizations based on the effects of other drugs. Although the compensatory changes associated with MPH self-administration do not resemble those of blockers, they also do not resemble those of a releaser, amphetamine, which had no effect on DA dynamics or kinetics. Although in vitro studies have shown that MPH exhibits the properties of both blockers and releasers, it is clear that MPH does not fit neatly into either category. Additionally, this work highlights that it is likely that the compensatory changes that occur within the DA system following repeated administration of MPH are driven by the way in which it interacts with the DAT, and not just its ability to elevate DA, as both cocaine and AMPH elevate DA to a similar extent. The fact that MPH causes compensatory changes that are different from both a prototypical blocker and a prototypical releaser demonstrates that not only does MPH have functional properties of both classes, but rather that it belongs in a class of its own MPH has limited serotonin transporter affinity In addition to the unique acute effects of MPH in regards to binding to the DAT, MPH is different than cocaine in regards to its affinity for the serotonin transporter. While cocaine binds with high affinity to the norepinephrine, DA, and serotonin transporters, MPH binds with high affinity only to the norepinephrine and DATs (John and Jones, 2007). MPH has low affinity for the serotonin transporter, so that at physiologically relevant concentrations, MPH does not elevate serotonin levels.
59 38 Although the acute effects of the drug at the DAT are independent of these differences, it is possible that the differences between the chronic effects of MPH and cocaine, as assayed following self-administration studies, are due to the lack of affinity of MPH for the serotonin transporter. In mice is has been shown that repeated MPH administration results in changes in the connectivity between the serotonin and dopamine systems, whereby, following MPH administration, serotonin reuptake inhibitors such as fluoxetine are able to increase DA-mediated behaviors (Brookshire & Jones, 2012). Thus, it is possible that changes such as this are happening within the self-administering animal, which leads to differential compensatory changes as compared to cocaine selfadministration MPH modulates the glutamate system Extensive work from the Dafny group has demonstrated that many of MPH s effects, especially in regards to behavioral sensitization and modulation of VTA receptor excitability, depend on activation of glutamate receptors (Yang et al., 2000a, b; Gaytan et al., 2000; Sripada et al., 1998; Gaytan et al., 2001; Prieto-Gómez et al., 2004; Prieto- Gómez et al., 2005). Gaytan et al., (2000) showed that the non-selective NMDA receptor antagonist, MK-801, is able to block MPH-induced locomotor sensitization demonstrating that the glutamate system is necessary for the development of sensitization. This is particularly relevant to the differential effects observed with MPH self-administration as MPH and AMPH have differential effects on glutamatergic transmission in VTA DA neurons (Prieto-Gómez et al., 2005). Pretreatment with AMPH
60 39 increased both the frequency and amplitude of miniature excitatory post-synaptic currents (mepscs), while pre-treatment with MPH increased only the frequency (Prieto-Gómez et al., 2005). Additionally, MPH and AMPH had differential acute effects on the type of mepsc with MPH only modulating NMDA receptor mediated mepscs and AMPH modulating both NMDA and AMPA receptor mediated mepscs. The differential effects of MPH on the glutamate system could potentially play a role, at least in part, in the differential effects of AMPH, cocaine, and MPH self-administration on the DA system. 7. MPH is a unique drug 7.1. MPH does not act like other blockers following cocaine self-administration MPH does not fit cleanly into either the blocker or releaser class, as cocaine selfadministration studies have demonstrated that MPH is affected in a manner that is significantly different from other psychostimulant blockers. Extended-access cocaine self-administration has been demonstrated repeatedly to result in pharmacodynamic tolerance, whereby cocaine is less effective at inhibiting DA uptake in the NAc core (Mateo et al., 2005; Calipari et al., 2013b, c, d, e; Ferris et al., 2011, 2012, 2013). In a recent publication, Ferris and colleagues (2012) determined the potency of a number of blocker and releaser compounds following cocaine self-administration. Of all the blockers tested, MPH, was the only one that was unaffected by cocaine selfadministration. Even more surprising, was the fact that none of the releasers tested were affected by cocaine self-administration, therefore, in this study, MPH seemed to fit into the same category as releaser compounds.
61 DAT levels affect MPH potency in a similar fashion to releasers MPH is unique not only in structure or function but also in the neurochemical alterations that influence its effects. Previous work by Salahpour et al., (2008) has demonstrated that over-expression of the DAT results in an increase in AMPH, but not blocker, potency as measured by microdialysis or locomotor behavior. This was further confirmed using FSCV, where AMPH, but not cocaine, potency was increased following transgenic DAT over-expression (Calipari et al., 2013a). Because MPH is a DAT blocker, one would hypothesize that it would be unaffected by changes in DAT levels, however, MPH s potency is increased with increasing DAT levels, both in uptake inhibition and in locomotor activity. One caveat is that Salahpour et al (2008) showed no difference in MPH-induced locomotor responding in the DAT-tg animals at the 20 mg/kg dose. Calipari et al. (2013a) also show no difference at this dose; however 20 mg/kg dose is on the descending limb of the dose-response curve for MPH, thus the groups are no longer significantly different. Regardless, this illustrates both behaviorally and neurochemically that MPH is unique as it is a DAT blocker that has pharmacological actions that mimic those of releasers. In addition, many neurochemical manipulations that increase transporter levels at baseline as measured by western blot and functionally by FSCV also increase the potency of MPH. This has been illustrated with a number of models including DAT-tg mice, MPH self-administration, social isolation (Yorgason et al., in preparation) and intermittent access cocaine self-administration (Calipari et al., 2013c). These paradigms result in increases in V max at baseline that results in increases in releaser and MPH potency. One advantage to using V max measurements as a functional measure of DAT
62 41 levels, is that dose response curves for drugs can be preformed within the same subject, thus, allowing for correlations between uptake rates and drug potency. Previously we correlated our measure of uptake inhibition (app. Km) with a functional measure of DAT levels (V max ). Correlation analysis of data from both MPH SA and DAT-tg show that DAT levels are positively correlated with potency measurements for AMPH, MPH, and to a lesser extent cocaine. Although there was a positive correlation between uptake rates and cocaine potency, the shift in cocaine potency across increasing V max values was substantially less robust than MPH and AMPH, and the effect size for cocaine was so small that no difference in app. K m was found between fast and slow V max based on a median split of V max values (Calipari et al., 2013a). This further supports the idea that DAT density increases affect MPH in a similar fashion to releasers. The proposed mechanism for the increased potency of releasers in scenarios where there are elevated DAT levels is outlined in Figure 3. It is possible that because releasers enter the cell through the DAT that when there are more transporters that more drug can enter the cell leading to enhanced drug effects and increased releaser potency. Further, because the release of DA into the cytoplasm is via DAT-mediated reverse transport, when DAT levels are elevated more DA can be released into the synapse. It has been shown previously that MPH does not enter the cell (Sonders et al., 1997); however, it is possible that MPH is causing release effects via a DAT mediated mechanism. For example, a number of kinases have been shown to participate in DAT signaling cascades (Beaulieu et al., 2005; Beom et al., 2004; Brami-Cherrier et al., 2002; Burgering and Coffer, 1995; Chen et al., 2010; Giros and Caron, 1993; Guptaroy et al.,

63 42
64 43 Figure 3. Increased Dopamine Transporter (DAT) Levels Increase the Potency of Psychostimulant Releasers. Schematic demonstrating the proposed mechanism for elevated releaser potency in dopamine (DA) neurons when DAT levels are elevated. (Left) The naïve state when there are normal DAT levels. Amphetamine (AMPH) enters the cell via the DAT (blue arrow), followed by entry into the vesicle via the vesicular monoamine transporter (VMAT). AMPH releases DA into the cytoplasm, which is released into the synapse via DAT-mediated reverse transport (red arrow). (Right) Elevated DAT levels following methylphenidate (MPH) self-administration (SA) or in DAT over-expressing mice. Because DAT levels are elevated, more AMPH can enter the cell and enter vesicles. AMPH releases more DA into the cytoplasm, which is released into the synapse via DAT-mediated reverse transport. Because DAT levels are elevated more DA can be released into the synapse from the cytoplasm, which leads to elevated synaptic DA levels and augmented post-synaptic receptor signaling as compared to the naïve state.
65 , 2010; Neve et al., 2004), and it is possible that MPH is causing changes to these signaling pathways, which leads to enhanced DAT-mediated release (Ferris et al., 2013). 8. Conclusions A high potential for MPH abuse in the human population is predicted by the behavioral and neurochemical profile of MPH in translational studies, which have demonstrated high rates of self-administration, escalation of intake over sessions, and high break-points on a progressive ratio schedule of reinforcement (Marusich et al., 2010; Calipari et al., 2013b). Relatively little work has been conducted on the neurochemical consequences of MPH abuse, however, it has been shown that MPH self-administration, under a number of reinforcement schedules, increases the potency of amphetamines and MPH, both in drug effects at the DAT and in reinforcement behavior (Calipari et al., 2013a). Although there are many similarities between the effects of MPH and cocaine both behaviorally and neurochemically, indicating that MPH may have abuse/addiction potential, the neurochemical consequences of binge self-administration of the two compounds are vastly different, making it impossible to make inferences on the neurochemical consequences of MPH based on the previous cocaine literature. Because MPH abuse in both the adolescent and adult population has been on the rise over the past decade, and will likely continue to increase, further understanding of the effects of high dose MPH on cognition, impulsivity, as well MPH effects on many brain systems is important to predict the clinical outcomes of MPH abuse.
66 45 9. Summary In conclusion, this dissertation uses psychostimulant (cocaine and MPH) selfadministration in combination with a variety of behavioral and neurochemical techniques including locomotor analysis, FSCV, and microdialysis to 1.) determine the consequences of cocaine self-administration on the DA system (Chapters 1-4), 2.) compare the effects of MPH self-administration on the DA system to previous work using cocaine (Chapter 5) 3.) determine the consequences of MPH self-administration on DA system function and psychostimulant reward and reinforcement (Chapters 5-8), and 4.) determine a mechanism for the MPH self-administration-induced increases in releaser, but not blocker, neurochemical and behavioral potency using DAT transgenic overexpressing mice (Chapters 6-7).
67 46 References: Ahmed SH, Koob GF (1998) Transition from moderate to excessive drug intake: change in hedonic set point. Science 282(5387): Ahmed SH, Koob GF (1999) Long-lasting increase in the set point for cocaine selfadministration after escalation in rats. Psychopharmacology Berl 146(3): Amini B, Yang PB, Swann AC, Dafny N. (2004) Differential locomotor responses in male rats from three strains to acute methylphenidate. Int J Neurosci114(9): Barrett SP, Darredeau C, Bordy LE, Pihl RO (2005) Characteristics of methylphenidate misuse in a university student sample. Can J Psychiatry 50: Beaulieu JM, Sotnikova TD, Marion S, Lefkowitz RJ, Gainetdinov RR, Caron MG. (2005) An Akt/beta-arrestin 2/PP2A signaling complex mediates dopaminergic neurotransmission and behavior. Cell. 122(2): Belin D, Balado E, Piazza PV, Deroche-Gamonet V. (2009) Pattern of intake and drug craving predict the development of cocaine addiction-like behavior in rats. Biol Psychiatry. 65(10): Belin D, Berson N, Balado E, Piazza PV, Deroche-Gamonet V. (2011) High-noveltypreference rats are predisposed to compulsive cocaine self-administration. Neuropsychopharmacology. 36(3): Ben-Shahar O, Moscarello JM, Ettenberg A (2006) One hour, but not six hours, of daily access to self-administered cocaine results in elevated levels of the dopamine transporter. Brain Res 1095(1): Beom S, Cheong D, Torres G, Caron MG, Kim KM. (2004) Comparative studies of molecular mechanisms of dopamine D2 and D3 receptors for the activation of extracellular signal-regulated kinase. J Biol Chem. 279(27): Biederman J, Quinn D, Weiss M, Markabi S, Weidenman M, Edson K, Karlsson G, Pohlmann H, Wigal S. (2003) Efficacy and safety of Ritalin LA, a new, once daily, extended-release dosage form of methylphenidate, in children with attention deficit hyperactivity disorder. Paediatr Drugs. 5(12): Biederman J, Spencer T (2002) Methylphenidate in treatment of adults with attentiondeficit/hyperactivity disorder. J Atten Disord 6:S Biederman J, Wilens T, Mick E, Spencer T, Faraone SV. (1999) Pharmacotherapy of attention-deficit/hyperactivity disorder reduces risk for substance use disorder. Pediatrics 104, e20 Botly LC, Burton CL, Rizos Z, Fletcher PJ (2008) Characterization of methylphenidate self-administration and reinstatement in the rat. Psychopharmacology (Berl). 199(1):55-66.
68 47 Bouffard R, Hechtman L, Minde K, Iaboni-Kassab F (2003) The efficacy of 2 different dosages of methylphenidate in treating adults with attention-deficit hyperactivity disorder. Can J Psychiatry 48: Brami-Cherrier K, Valjent E, Garcia M, Pagès C, Hipskind RA, Caboche J. (2002) Dopamine induces a PI3-kinase-independent activation of Akt in striatal neurons: a new route to camp response element-binding protein phosphorylation. J Neurosci. 22(20): Brandon CL, Marinelli M, Baker LK, White FJ. (2001) Enhanced reactivity and vulnerability to cocaine following methylphenidate treatment in adolescent rats. Neuropsychopharmacology. 25(5): Brookshire BR, Jones SR. (2012) Chronic methylphenidate administration in mice produces depressive-like behaviors and altered responses to fluoxetine. Synapse. 66(9): Burgering BM, Coffer PJ. (1995) Protein kinase B (c-akt) in phosphatidylinositol-3-oh kinase signal transduction. Nature. 376(6541): Burton CL, Nobrega JN, Fletcher PJ (2010) The effects of adolescent methylphenidate self-administration on responding for a conditioned reward, amphetamine-induced locomotor activity, and neuronal activation. Psychopharmacology (Berl) 208(3): Calipari ES, Ferris MJ, Salahpour A, Caron MG, Roberts DCS, Jones SR (2013a) Methylphenidate amplifies the potency and reinforcing effects of amphetamines by increasing dopamine transporter expression. Nature Communications. In press. Calipari ES, Ferris MJ, Melchior JR, Bermejo K, Salahpour A, Roberts DC et al (2013b). Methylphenidate and cocaine self-administration produce distinct dopamine terminal alterations. Addict Biol (in press). Calipari ES, Ferris MJ, Zimmer BA, Roberts DC, Jones SR. (2013c) Temporal pattern of cocaine intake determines tolerance vs sensitization of cocaine effects at the dopamine transporter. Neuropsychopharmacology. In press. Calipari ES, Beveridge TJ, Jones SR, Porrino LJ (2013d) Withdrawal from extended access cocaine self-administration results in dysregulated functional activity and altered locomotor activity in rats. Eur. J. Neurosci. In press. Calipari ES, Ferris MJ, Jones SR (2013e) Extended access cocaine self-administration results in tolerance to the dopamine-elevating and locomotor-stimulating effects of cocaine. J. Neurochem. In press. Carboni E, Imperato A, Perezzani L, Di Chiara G (1989) Amphetamine, cocaine, phencyclidine and nomifensine increase extracellular dopamine concentrations
69 48 preferentially in the nucleus accumbens of freely moving rats. Neuroscience 28(3): Carlezon WA Jr, Mague SD, Andersen SL.(2003) Enduring behavioral effects of early exposure to methylphenidate in rats. Biol Psychiatry. 54(12): Carlsson A. (1987) Perspectives on the discovery of central monoaminergic neurotransmission. Annu Rev Neurosci.10: Chadchankar H, Ihalainen J, Tanila H, Yavich L. (2012) Methylphenidate modifies overflow and presynaptic compartmentalization of dopamine via an α-synucleindependent mechanism. J Pharmacol Exp Ther. 341(2): Chase TN, Oh JD, Blanchet PJ. (1998) Neostriatal mechanisms in Parkinson's disease. Neurology. 51(2 Suppl 2):S30-5. Chen R, Furman CA, Gnegy ME. (2010) Dopamine transporter trafficking: rapid response on demand. Future Neurol. 5(1):123. Chen N, Reith ME. (2007) Substrates and inhibitors display different sensitivity to expression level of the dopamine transporter in heterologously expressing cells. J Neurochem. 101(2): Chen R, Han DD, Gu HH (2005) A triple mutation in the second transmembrane domain of mouse dopamine transporter markedly decreases sensitivity to cocaine and methylphenidate. J Neurochem 94(2): Chen R, Tilley MR, Wei H, Zhou F, Zhou FM, Ching S, Quan N, Stephens RL, Hill ER, Nottoli T, Han DD, Gu HH. (2006) Abolished cocaine reward in mice with a cocaineinsensitive dopamine transporter. Proc Natl Acad Sci U S A. 103(24): Cheon KA, Ryu YH, Kim YK, Namkoong K, Kim CH, Lee JD. (2003) Dopamine transporter density in the basal ganglia assessed with [123I]IPT SPET in children with attention deficit hyperactivity disorder. Eur J Nucl Med Mol Imaging. 30(2): Collins JR, Weeks JR, Cooper MM, Good PI, Russell RR (1984) Prediction of abuse liability of drugs using IV self-administration by rats. Psychopharmacology 82: Dafny N, Yang PB. (2006) The role of age, genotype, sex, and route of acute and chronic administration of methylphenidate: a review of its locomotor effects. Brain Res Bull. 68(6): Dar DE, Mayo C, Uhl GR. (2005) The interaction of methylphenidate and benztropine with the dopamine transporter is different than other substrates and ligands. Biochem Pharmacol. 70, Del Campo N, Chamberlain SR, Sahakian BJ, Robbins TW. (2011) The roles of dopamine and noradrenaline in the pathophysiology and treatment of attentiondeficit/hyperactivity disorder. Biol Psychiatry. 69(12):e
70 49 dela Peña I, Yoon SY, Lee JC, dela Peña JB, Sohn AR, Ryu JH, Shin CY, Cheong JH. (2012) Methylphenidate treatment in the spontaneously hypertensive rat: influence on methylphenidate self-administration and reinstatement in comparison with Wistar rats. Psychopharmacology (Berl). 221(2): Deroche-Gamonet V, Belin D, Piazza PV. (2004) Evidence for addiction-like behavior in the rat. Science. 305(5686): Deroche-Gamonet, V. (2008) Psychobiological features associated with cocaine addiction-like behavior in rats. Int. J. Psychol., 43, 181. Di Chiara G (1995) The role of dopamine in drug abuse viewed from the perspective of its role in motivation. Drug Alcohol Depend 38(2): Di Chiara G, Imperato A (1988) Drugs abused by humans preferentially increase synaptic dopamine concentrations in the mesolimbic system of freely moving rats. Proc Natl Acad Sci U S A 85(14): Dougherty DD, Bonab AA, Spencer TJ, Rauch SL, Madras BK, Fischman AJ. (1999) Dopamine transporter density in patients with attention deficit hyperactivity disorder. Lancet. 354(9196): Dresel S, Krause J, Krause KH, LaFougere C, Brinkbäumer K, Kung HF, Hahn K, Tatsch K. (2000) Attention deficit hyperactivity disorder: binding of [99mTc]TRODAT-1 to the dopamine transporter before and after methylphenidate treatment. Eur J Nucl Med. 27(10): Dyck LE, Boulton AA, Jones RS. (1980) A comparison of the effects of methylphenidate and amphetamine on the simultaneous release of radiolabelled dopamine and p- or m- tyramine from rat striatal slices. Eur J Pharmacol. 68(1): Eckermann K, Beasley A, Yang P, Gaytan O, Swann A, Dafny N. (2001) Methylphenidate sensitization is modulated by valproate. Life Sci. 69(1): Ferris MJ, Calipari ES, Mateo Y, Melchior JR, Roberts DC, Jones SR (2012). Cocaine self-administration produces pharmacodynamic tolerance: differential effects on the potency of dopamine transporter blockers, releasers, and methylphenidate. Neuropsychopharmacology 37: Ferris MJ, Calipari ES, Melchior JR, Roberts DC, España RA, Jones SR. (2013) Paradoxical tolerance to cocaine after initial supersensitivity in drug-use-prone animals. Eur J Neurosci. In press. Ferris MJ, Mateo Y, Roberts DC, Jones SR (2011) Cocaine-insensitive dopamine transporters with intact substrate transport produced by self-administration. Biol Psychiatry 69(3):201-7.
71 50 Ferris MJ, Calipari ES, Yorgason JT, Jones SR. (2013) Examining the complex regulation and drug-induced plasticity of dopamine release and uptake using voltammetry in brain slices. ACS Chem Neurosci. 4(5): Fleckenstein AE, Volz TJ, Riddle EL, Gibb JW, Hanson GR (2007) New insights into the mechanism of action of amphetamines. Annu Rev Pharmacol Toxicol. 47: Fone KC, Nutt DJ. (2005) Stimulants: use and abuse in the treatment of attention deficit hyperactivity disorder. Curr Opin Pharmacol. 5 (1):87 93 Frauger E, Moracchini C, Le Boisselier R, Braunstein D, Thirion X, Micallef J. (2013) OPPIDUM surveillance program: 20 years of information on drug abuse in France. Fundam Clin Pharmacol. In press. Gautschi OP, Zellweger R (2006) Necrotising myositis after intravenous methylphenidat (Ritalin) injection. Emerg Med J 23(9):739. Gawin FH, Ellinwood EH Jr. (1989) Cocaine dependence. Annu Rev Med. 40: Gaytan O, al-rahim S, Swann A, Dafny N. (1997) Sensitization to locomotor effects of methylphenidate in the rat. Life Sci. 61(8):PL Gaytan O, Nason R, Alagugurusamy R, Swann A, Dafny N. (2000) MK-801 blocks the development of sensitization to the locomotor effects of methylphenidate. Brain Res Bull. 51(6): Gaytan O, Sripada S, Swann A, Dafny N. (2001) Blockade of sensitization to methylphenidate by MK-801: partial dissociation from motor effects. Neuropharmacology. 40(2): Gerasimov MR, Franceschi M, Volkow ND, Gifford A, Gatley SJ, Marsteller D, Molina PE, Dewey SL (2000) Comparison between intraperitoneal and oral methylphenidate administration: A microdialysis and locomotor activity study. J Pharmacol Exp Ther. 295(1):51-7. Gill KE, Beveridge TJ, Smith HR, Porrino LJ. (2013) The effects of rearing environment and chronic methylphenidate administration on behavior and dopamine receptors in adolescent rats. Brain Res. 1527: Gill KE, Pierre PJ, Daunais J, Bennett AJ, Martelle S, Gage HD, Swanson JM, Nader MA, Porrino LJ. (2012) Chronic treatment with extended release methylphenidate does not alter dopamine systems or increase vulnerability for cocaine self-administration: a study in nonhuman primates. Neuropsychopharmacology. 37(12): Giros B, Caron MG. (1993) Molecular characterization of the dopamine transporter. Trends Pharmacol Sci. 2:43-9. Grünblatt E, Bartl J, Marinova Z, Walitza S. (2013) In vitro study methodologies to investigate genetic aspects and effects of drugs used in attention-deficit hyperactivity
72 51 disorder.j Neural Transm. 120(1): Guptaroy B, Fraser R, Desai A, Zhang M, Gnegy ME. (2010) Site directed mutations near transmembrane domain 1 (TM1) alter conformation and function of norepinephrine and dopamine transporters. Mol Pharmacol. 79(3):5ss20-32 Guptaroy B, Zhang M, Bowton E, Binda F, Shi L, Weinstein H, Galli A, Javitch JA, Neubig RR, Gnegy ME. (2009) A juxtamembrane mutation in the N terminus of the dopamine transporter induces preference for an inward-facing conformation. Mol Pharmacol. 75(3): Han DD, Gu HH (2006) Comparison of the monoamine transporters from human and mouse in their sensitivities to psychostimulant drugs. BMC Pharmacol. 3(6):6. Heal DJ, Cheetham SC, Smith SL (2009) The neuropharmacology of ADHD drugs in vivo: insights on efficacy and safety. Neuropharmacology 57(7-8): Heal DJ, Prow MR, Hearson M, Buckett WR (1996) Efflux of [3H]dopamine from superfused rat striatal slices: predictive value for detecting stimulant drugs of abuse. Br. J. Pharmacol 117:325P. Hoexter MQ, Fadel G, Felício AC, Calzavara MB, Batista IR, Reis MA, Shih MC, Pitman RK, Andreoli SB, Mello MF, Mari JJ, Bressan RA. (2012) Higher striatal dopamine transporter density in PTSD: an in vivo SPECT study with [(99m)Tc]TRODAT-1. Psychopharmacology (Berl). 224(2): Izenwasser S, Coy AE, Ladenheim B, Loeloff RJ, Cadet JL and French D (1999) Chronic methylphenidate alters locomotor activity and dopamine transporters differently from cocaine. Eur J Pharmacol 373: Johanson CE, Balster RL, Bonese K (1976) Self-administration of psychomotor stimulant drugs: the effects of unlimited access. Pharmacol Biochem Behav. 4(1): John CE, Jones SR. (2007) Voltammetric characterization of the effect of monoamine uptake inhibitors and releasers on dopamine and serotonin uptake in mouse caudateputamen and substantia nigra slices. Neuropharmacology. 52(8): Johnston, L. D., O'Malley, P. M., Bachman, J. G., & Schulenberg, J. E. (2010). Monitoring the Future national results on adolescent drug use: Overview of key findings, 2009 (NIH Publication No ). Bethesda, MD: National Institute on Drug Abuse. Johnston, L. D., O'Malley, P. M., Bachman, J. G., & Schulenberg, J. E. (2010). Monitoring the Future national results on adolescent drug use: Overview of key findings, 2009 (NIH Publication No ). Bethesda, MD: National Institute on Drug Abuse. Kalivas PW, Duffy P (1990). Effect of acute and daily cocaine treatment on extracellular dopamine in the nucleus accumbens. Synapse 5: Kalivas PW, Duffy P (1993). Time course of extracellular dopamine and behavioral sensitization to cocaine. I. Dopamine axon terminals. J Neurosci 13:
73 52 Klein-Schwartz W. (2002) Abuse and toxicity of methylphenidate. Curr Opin Pediatr. 14(2): Kohut SJ, Decicco-Skinner KL, Johari S, Hurwitz ZE, Baumann MH, Riley AL. (2012) Differential modulation of cocaine's discriminative cue by repeated and variable stress exposure: relation to monoamine transporter levels. Neuropharmacology. 63(2): Koob G, Kreek MJ (2007) Stress, dysregulation of drug reward pathways, and the transition to drug dependence. Am J Psychiatry. 164(8): Koob GF, Le Moal M. (1997) Drug abuse: hedonic homeostatic dysregulation. Science. 278(5335):52-8. Koob GF. (1996a) Hedonic valence, dopamine and motivation. Mol Psychiatry. 1(3): Koob GF. (1996b) Drug addiction: the yin and yang of hedonic homeostasis. Neuron. 16(5): Krause KH, Dresel SH, Krause J, Kung HF, Tatsch K. (2000) Increased striatal dopamine transporter in adult patients with attention deficit hyperactivity disorder: effects of methylphenidate as measured by single photon emission computed tomography. Neurosci Lett. 285(2): Lambert NM, Hartsough CS. (1998) Prospective study of tobacco smoking and substance dependencies among samples of ADHD and non-adhd participants. J Learn Disabil. 31(6): Lee MJ, Swann AC, Dafny N. (2008) Methylphenidate sensitization is prevented by prefrontal cortex lesion. Brain Res Bull. 76(1-2): Levine B, Caplan YH, Kauffman G (1986) Fatality resulting from methylphenidate overdose. J Anal Toxicol. 10(5): Madras BK, Miller GM, Fischman AJ. (2005) The dopamine transporter and attentiondeficit/hyperactivity disorder. Biol Psychiatry. 57, Mannuzza S, Klein RG, Moulton JL 3rd. (2003) Does stimulant treatment place children at risk for adult substance abuse? A controlled, prospective follow-up study. J Child Adolesc Psychopharmacol Fall;13(3): Mariani JJ, Levin FR. Psychostimulant treatment of cocaine dependence. (2012) Psychiatr Clin North Am. 35(2): Marusich JA, Beckmann JS, Gipson CD, Bardo MT (2010) Methylphenidate as a reinforcer for rats: contingent delivery and intake escalation. Exp Clin Psychopharmacol 18(3):
74 53 Marusich JA, Beckmann JS, Gipson CD, Bardo MT. (2011) Cue effects on methylphenidate self-administration in rats. Behav Pharmacol. 22(7): Marusich JA, McCuddy WT, Beckmann JS, Gipson CD, Bardo MT. (2011) Strain differences in self-administration of methylphenidate and sucrose pellets in a rat model of attention-deficit hyperactivity disorder. Behav Pharmacol. 22(8): Mateo Y, Lack CM, Morgan D, Roberts DC, Jones SR (2005) Reduced dopamine terminal function and insensitivity to cocaine following cocaine binge self-administration and deprivation. Neuropsychopharm 30(8): Mazei-Robison MS, Couch RS, Shelton RC, Stein MA, Blakely RD. (2005) Sequence variation in the human dopamine transporter gene in children with attention deficit hyperactivity disorder. Neuropharmacology. 249(6): McCabe SE, Knight JR, Teter CJ, Wechsler H. (2005) Non-medical use of prescription stimulants among US college students: prevalence and correlates from a national survey. Addiction.100(1): McCabe SE, Teter CJ, Boyd CJ, Guthrie SK. (2004) Prevalence and correlates of illicit methylphenidate use among 8th, 10th, and 12th grade students in the United States, J Adolesc Health. 35(6): Moll GH, Hause S, Rüther E, Rothenberger A, Huether G (2001) Early methylphenidate administration to young rats causes a persistent reduction in the density of striatal dopamine transporters. J Child Adolesc Psychopharmacol 11(1): Morris SM, Petibone DM, Lin WJ, Chen JJ, Vitiello B, Witt KL, Mattison DR. (2012) The genetic toxicity of methylphenidate: a review of the current literature.j Appl Toxicol. 32(10): Morton WA, Stockton GG (2000) Methylphenidate Abuse and Psychiatric Side Effects. Prim Care Companion J Clin Psychiatry. 2(5): Neve KA, Seamans JK, Trantham-Davidson H. (2004) Dopamine receptor signaling. J Recept Signal Transduct Res. 24(3): Oleson EB, Talluri S, Childers SR, Smith JE, Roberts DC, Bonin KD, Budygin EA (2009) Dopamine uptake changes associated with cocaine self-administration. Neuropsychopharmacology. 34(5): Oleson EB, Roberts DC. (2012) Cocaine self-administration in rats: threshold procedures. Methods Mol Biol. 829: Oleson EB, Roberts DC. (2009) Behavioral economic assessment of price and cocaine consumption following self-administration histories that produce escalation of either final ratios or intake. Neuropsychopharmacology. 34(3):
75 54 Parran TV Jr, Jasinski DR (1991) Intravenous methylphenidate abuse. Prototype for prescription drug abuse. Arch Intern Med. 151(4): Parsons LH, Justice JBJr (1993). Serotonin and dopamine sensitization in the nucleus accumbens, ventral tegmental area, and dorsal raphe nucleus following repeated cocaine administration. J Neurochem 61: Peoples LL, Gee F, Bibi R, West MO (1998) Phasic firing time locked to cocaine selfinfusion and locomotion: dissociable firing patterns of single nucleus accumbens neurons in the rat. J Neurosci. 18(18): Prieto-Gomez B, Benitez MT, Vazquez-Alvarez AM, Yang PB, Reyes Vazquez C, Dafny N. (2004) Dopaminergic ventral tegmental neurons modulated by methylphenidate. Life Sci. 74(13): Prieto-Gomez B, Benitez MT, Vazquez-Alvarez AM, Yang PB, Reyes Vazquez C, Dafny N. (2004) Dopaminergic ventral tegmental neurons modulated by methylphenidate. Life Sci. 74(13): Prieto-Gómez B, Vázquez-Alvarez AM, Martínez-Peña JL, Reyes-Vázquez C, Yang PB, Dafny N. (2005) Methylphenidate and amphetamine modulate differently the NMDA and AMPA glutamatergic transmission of dopaminergic neurons in the ventral tegmental area. Life Sci. 77(6): Prommer E. (2012) Methylphenidate: established and expanding roles in symptom management. Am J Hosp Palliat Care. 29(6): Ritz MC, Lamb RJ, Goldberg SR, Kuhar MJ (1987). Cocaine receptors on dopamine transporters are related to self-administration of cocaine. Science 237: Robbins TW. (2003) Dopamine and cognition. Curr Opin Neurol. 16 Suppl 2:S1-2. Roessner V, Sagvolden T, Dasbanerjee T, Middleton FA, Faraone SV, Walaas SI, Becker A, Rothenberger A, Bock N. (2010) Methylphenidate normalizes elevated dopamine transporter densities in an animal model of the attention-deficit/hyperactivity disorder combined type, but not to the same extent in one of the attention-deficit/hyperactivity disorder inattentive type. Neuroscience. 167(4): Rosen JB, Young AM, Beuthin FC, Louis-Ferdinand RT (1985) Discriminative stimulus properties of amphetamine and other stimulants in lead-exposed and normal rats, Pharmacol, Biochem Behav 24: Rothman RB, Baumann MH. (2003) Monoamine transporters and psychostimulant drugs. Eur J Pharmacol. 479(1-3): Rudnick G, Clark J. (1993) From synapse to vesicle: the reuptake and storage of biogenic amine neurotransmitters. Biochim Biophys Acta. 1144(3): Rush CR, Baker RW (2001) Behavioral pharmacological similarities between methylphenidate and cocaine in cocaine abusers. Exp Clin Psychopharmacol. 9(1):59-73.
76 55 Rush CR, Essman WD, Simpson CA, Baker RW (2001) Reinforcing and subject-rated effects of methylphenidate and d-amphetamine in non-drug-abusing humans. J Clin Psychopharmacol. 21(3): Russell V, de Villiers A, Sagvolden T, Lamm M, Taljaard J (1998) Differences between electrically-, ritalin- and D-amphetamine-stimulated release of [3H]dopamine from brain slices suggest impaired vesicular storage of dopamine in an animal model of Attention- Deficit Hyperactivity Disorder. Behav Brain Res. 94(1): Sakrikar D, Mazei-Robison MS, Mergy MA, Richtand NW, Han Q, Hamilton PJ, Bowton E, Galli A, Veenstra-Vanderweele J, Gill M, Blakely RD. (2012) Attention deficit/hyperactivity disorder-derived coding variation in the dopamine transporter disrupts microdomain targeting and trafficking regulation. J Neurosci. 32(16): Salahpour A, Ramsey AJ, Medvedev IO, Kile B, Sotnikova TD, Holmstrand E, Ghisi V, Nicholls PJ, Wong L, Murphy K, Sesack SR, Wightman RM, Gainetdinov RR, Caron MG. (2008) Increased amphetamine-induced hyperactivity and reward in mice overexpressing the dopamine transporter. Proc Natl Acad Sci U S A (11): Sandoval V, Riddle EL, Hanson GR, Fleckenstein AE. (2002) Methylphenidate redistributes vesicular monoamine transporter-2: role of dopamine receptors. J Neurosci. 22(19): Sepúlveda DR, Thomas LM, McCabe SE, Cranford JA, Boyd CJ, Teter CJ. (2011) Misuse of prescribed stimulant medication for ADHD and associated patterns of substance use: preliminary analysis among college students. J Pharm Pract. 24(6): Setlik J, Bond GR, Ho M (2009) Adolescent Prescription ADHD Medication Abuse Is Rising Along With Prescriptions for These Medications. Pediatrics [Epub ahead of print]. Shaw SY, Shah L, Jolly AM, Wylie JL (2008) Identifying heterogeneity among injection drug users: a cluster analysis approach. Am J Public Health. 98(8): Sherman CB, Hudson LD, Pierson DJ (1987) Severe precocious emphysema in intravenous methylphenidate (Ritalin) abusers. Chest. 92(6): Silverman PB, Ho BT (1980) The discriminative stimulus properties of 2,5-dimethoxy-4- methylamphetamine (DOM): differentiation from amphetamine. Psychopharmacology (Berl). 68(3): Sonders MS, Zhu SJ, Zahniser NR, Kavanaugh MP, Amara SG (1997) Multiple ionic conductances of the human dopamine transporter: the actions of dopamine and psychostimulants. J Neurosci 17(3): Soto PL, Wilcox KM, Zhou Y, Kumar A, Ator NA, Riddle MA, Wong DF, Weed MR. (2012) Long-term exposure to oral methylphenidate or dl-amphetamine mixture in periadolescent rhesus monkeys: effects on physiology, behavior, and dopamine system development. Neuropsychopharmacology. 37(12):
77 56 Sproson EJ, Chantrey J, Hollis C, Marsden CA, Fonel KC (2001) Effect of repeated methylphenidate administration on presynaptic dopamine and behaviour in young adult rats. J Psychopharmacol 15(2): Sripada S, Gaytan O, Al-rahim S, Swann A, Dafny N. (1998) Dose-related effects of MK-801 on acute and chronic methylphenidate administration. Brain Res. 814(1-2): Substance Abuse and Mental Health Services Administration (SAMHSA) (2008). National survey on drug use and health. Retrieved from Teter CJ, Falone AE, Cranford JA, Boyd CJ, McCabe SE. (2010) Nonmedical use of prescription stimulants and depressed mood among college students: frequency and routes of administration. J Subst Abuse Treat. 38(3): Teter CJ, McCabe SE, Boyd CJ, Guthrie SK. (2003) Illicit methylphenidate use in an undergraduate student sample: prevalence and risk factors. Pharmacotherapy. 23(5): Teter CJ, McCabe SE, Cranford JA, Boyd CJ, Guthrie SK. (2005) Prevalence and motives for illicit use of prescription stimulants in an undergraduate student sample. J Am Coll Health. 53(6): Teter CJ, McCabe SE, LaGrange K, Cranford JA, Boyd CJ (2006) Illicit use of specific prescription stimulants among college students: prevalence, motives, and routes of administration. Pharmacotherapy 26(10): Tilley MR, Gu HH (2008) The effects of methylphenidate on knockin mice with a methylphenidate-resistant dopamine transporter. J Pharmacol Exp Ther. 327(2): Volkow ND, Fowler JS, Gatley SJ, Dewey SL, Wang GJ, Logan J, Ding YS, Franceschi D, Gifford A, Morgan A, Pappas N, King P. (1999) Comparable changes in synaptic dopamine induced by methylphenidate and by cocaine in the baboon brain. Synapse. 31(1): Volkow ND, Fowler JS, Wang G, Ding Y, Gatley SJ. (2002) Mechanism of action of methylphenidate: insights from PET imaging studies. J Atten Disord 6 Suppl 1:S Volkow ND, Fowler JS, Wang GJ. (1999) Imaging studies on the role of dopamine in cocaine reinforcement and addiction in humans. J Psychopharmacol.13(4): Volz TJ, Farnsworth SJ, Hanson GR, Fleckenstein AE. (2009) Methylphenidate-Induced Alterations in Synaptic Vesicle Trafficking and Activity: Functional Consequences and Therapeutic Implications. Ann N Y Acad Sci. 1139: Volz TJ, Farnsworth SJ, King JL, Riddle EL, Hanson GR, Fleckenstein AE. (2007) Methylphenidate administration alters vesicular monoamine transporter-2 function in cytoplasmic and membrane-associated vesicles. J Pharmacol Exp Ther. 323(2):
78 57 Wayment HK, Deutsch H, Schweri MM, Schenk JO. (1999) Effects of methylphenidate analogues on phenethylamine substrates for the striatal dopamine transporter: potential as amphetamine antagonists? J Neurochem. 72, Wilens TE, Adler LA, Adams J, Sgambati S, Rotrosen J, Sawtelle R, Utzinger L, Fusillo S (2008) Misuse and diversion of stimulants prescribed for ADHD: a systematic review of the literature. J Am Acad Child Adolesc Psychiatry. 47(1): Wilens TE. (2003) Does the medicating ADHD increase or decrease the risk for later substance abuse? Rev Bras Psiquiatr. 25(3): Willens TE (2004) Impact of ADHD and its treatment on substance abuse in adults. J. Clin. Psychiatry. 65: Woods SK, Meyer JS (1991) Exogenous tyrosine potentiates the methylphenidateinduced increase in extracellular dopamine in the nucleus accumbens: a microdialysis study. Brain Res. 560(1-2): Wooters TE, Bardo MT, Dwoskin LP, Midde NM, Gomez AM, Mactutus CF, Booze RM, Zhu J. (2011) Effect of environmental enrichment on methylphenidate-induced locomotion and dopamine transporter dynamics. Behav Brain Res. 219(1): Wooters TE, Bardo MT. (2009) Nicotinic receptors differentially modulate the induction and expression of behavioral sensitization to methylphenidate in rats. Psychopharmacology (Berl). 204(3): Wooters TE, Bardo MT. (2011) Methylphenidate and fluphenazine, but not amphetamine, differentially affect impulsive choice in spontaneously hypertensive, Wistar-Kyoto and Sprague-Dawley rats. Brain Res. 1396: Wooters TE, Walton MT, Bardo MT. (2011) Oral methylphenidate establishes a conditioned place preference in rats. Neurosci Lett. 487(3): Yang P, Beasley A, Swann A, Dafny N. (2000) Valproate modulates the expression of methylphenidate (ritalin) sensitization. Brain Res. 874(2): Yang P, Beasley A, Swann A, Dafny N. (2000) Valproate modulates the expression of methylphenidate (ritalin) sensitization. Brain Res. 874(2): Yang P, Swann A, Dafny N. (2000) NMDA receptor antagonist disrupts acute and chronic effects of methylphenidate. Physiol Behav. 71(1-2): Yang PB, Amini B, Swann AC, Dafny N. (2003) Strain differences in the behavioral responses of male rats to chronically administered methylphenidate. Brain Res. 971(2): Yang PB, Atkins KD, Dafny N. (2011) Behavioral sensitization and cross-sensitization between methylphenidate amphetamine, and 3,4-methylenedioxymethamphetamine (MDMA) in female SD rats. Eur J Pharmacol. 661(1-3):72-85.
79 58 Yang PB, Swann AC, Dafny N. (2003) Chronic pretreatment with methylphenidate induces cross-sensitization with amphetamine. Life Sci. 73(22): Yang PB, Swann AC, Dafny N. (2007) Chronic administration of methylphenidate produces neurophysiological and behavioral sensitization. Brain Res. 1145: Yang PB, Swann AC, Dafny N. (2007) Methylphenidate treated at the test cage--dosedependent sensitization or tolerance depend on the behavioral assay used. Crit Rev Neurobiol. 19(1): Yorgason JT, España RA, Jones SR (2012) Demon Voltammetry and Analysis software: Analysis of cocaine-induced alterations in dopamine signaling using multiple kinetic measures. J Neurosci Methods. In press. Zaczek R, Battaglia G, Contrera JF, Culp S, De Souza EB (1989) Methylphenidate and pemoline do not cause depletion of rat brain monoamine markers similar to that observed with methamphetamine. Toxicol Appl Pharmacol 100: Zimmer BA, Oleson EB, Roberts DCS (2012). The motivation to self-administer is increased after a history of spiking brain levels of cocaine. Neuropsychopharmacology 37:
80 59 CHAPTER II EXTENDED ACCESS COCAINE SELF-ADMINISTRATION RESULTS IN TOLERANCE TO THE DOPAMINE-ELEVATING AND LOCOMOTOR STIMULATING EFFECTS OF COCAINE Erin S. Calipari, Mark J. Ferris and Sara R. Jones The following manuscript was published in The Journal of Neurochemistry, in press, 2013, and is reprinted with permission. Stylistic variations are due to the requirements of the journal. Erin S. Calipari performed all self-administration, microdialysis and behavioral experiments, data analysis, and prepared the manuscript. Dr. Mark J. Ferris performed select voltammetry experiments. Drs. Sara R. Jones and Mark J. Ferris acted in an advisory and editorial capacity.
81 60 Abstract: Tolerance to the neurochemical and behavioral effects of cocaine after repeated use is a hallmark of cocaine addiction in humans. However, comprehensive studies on tolerance to the behavioral and neurochemical effects of cocaine following contingent administration in rodents are lacking. We outlined the consequences of extended access cocaine self-administration as it related to tolerance to the psychomotor, dopamine (DA) elevating, and DA transporter (DAT) inhibiting effects of cocaine. Cocaine selfadministration (1.5 mg/kg/inj; 40 inj; 5 days), which resulted in escalation of first hour intake, caused reductions in evoked DA release and reduced maximal rates of uptake through the DAT. Further, we report reductions in cocaine-induced uptake inhibition as measured by fast scan cyclic voltammetry, and a corresponding increase in the dose of cocaine required for 50% inhibition of DA uptake (K i ) at the DAT. Cocaine tolerance at the DAT translated to reductions in cocaine-induced DA overflow as measured by microdialysis. Additionally, cocaine-induced elevations in locomotor activity were reduced. Here we demonstrate both neurochemical and behavioral cocaine tolerance in an extended-access rodent model of cocaine abuse, which allows for a better understanding of the neurochemical and psychomotor tolerance that develops to cocaine during the human addiction process.
82 61 Introduction: Cocaine is a DAT blocker that inhibits the uptake of dopamine (DA), thus increasing extracellular DA levels and augmenting postsynaptic DA receptor activation. The ability of cocaine to inhibit the DAT is essential for its rewarding effects, which is highlighted by the fact that transgenic mice with cocaine-insensitive DATs do not develop conditioned place preference for the drug (Chen et al., 2008). Further, the potency of stimulants for inhibiting the DAT is strongly correlated with selfadministration behavior (Ritz et al., 1987, Roberts et al. 1977), suggesting that changes in cocaine potency at the DAT have important behavioral implications. In rodents, tolerance to the pharmacological DAT-inhibiting effects of cocaine has been reported previously following a variety of fixed-ratio and progressive-ratio cocaine selfadministration paradigms (Calipari et al., 2012, 2013; Ferris et al., 2011, 2012, 2013; Mateo et al., 2005). Tolerance has also been reported to the DA elevating and locomotor activating effects of cocaine (Hurd et al., 1989; Lack et al., 2008). Although tolerance has been demonstrated to these different aspects of cocaine effects, the results are from a wide range of different self-administration paradigms, and a comprehensive understanding of the tolerance induced by the extended-access, defined-intake (5 days, 40 inj/day, 1.5 mg/kg/inj) model is lacking. Tolerance to the euphorigenic and DA elevating effects of cocaine has been reported consistently in human cocaine addicts (Dackis and O Brien, 2001; Mendelson et al., 1998; Reed et al., 2009; Volkow et al., 1997a, b; Volkow et al., 1996), and is thought to mediate some aspects of continued drug taking. Tolerance has been suggested to modulate cocaine intake behavior, specifically escalation of intake that occurs after
83 62 repeated use that has been reported in both human addicts and rodent self-administration studies (Ahmed et al., 2002, 2003; Ahmed and Koob, 2005; Barrett et al., 2004; Dackis and O Brien, 2001; Koob and Le Moal, 2001). Therefore, we aimed to define cocaine self-administration induced tolerance in rodents. We found that the extended-access, defined-intake self-administration model (Calipari et al., 2012; Ferris et al., 2011, 2012) mimics important aspects of the tolerance reported in humans, which is characterized by deficits in baseline DA functioning, reduced cocaine potency, escalated selfadministration behavior, and reduced cocaine-induced locomotor behaviors. Here we show that a history of cocaine self-administration resulted in reduced DA system function characterized by reductions in baseline DA release and uptake, as well as reduced psychomotor and neurochemical potencies of cocaine. These data highlight that self-administration results in tolerance to DA uptake inhibition by cocaine at the DAT that is concomitant with reduced cocaine-induced DA elevations as well as reduced behavioral activation. In addition, because of the integral role that the DA system plays in learning and motivated behaviors, it is possible that the documented changes could have implications for many different types of behavior such as learning and the execution of goal oriented behaviors in the absence of drug. Materials and Methods Animals: Male, Sprague-Dawley rats ( g; Harlan Laboratories, Frederick, MD) were maintained according to the National Institutes of Health guidelines in Association for Assessment and Accreditation of Laboratory Animal Care accredited facilities. The
84 63 experimental protocol was approved by the Institutional Animal Care and Use Committee at Wake Forest School of Medicine. Self-Administration: Rats were anesthetized with ketamine (100 mg/kg) and xylazine (10 mg/kg), implanted with chronic indwelling jugular catheters, and trained for i.v. selfadministration as previously described (Calipari et al., 2013). Following surgery, animals were singly housed, and all self-administration sessions took place in the home cage. Each animal was maintained on a reversed light cycle (3:00 am lights off; 3:00 pm lights on), and all self-administration procedures occurred during the active/dark cycle. Sessions were six hours in length and were terminated at the end of the six hours or after 40 injections of drug. Animals self-administered cocaine (1.5 mg/kg/inj over 4 sec) on a fixed-ratio 1 schedule of administration. Concurrent with the start of each injection, the lever retracted and a stimulus light was activated for 20 seconds to signal a time-out period. Under these conditions, animals acquired a stable pattern of intake within 1 to 5 days. For self-administering animals, acquisition (Day 1) was counted when the animal reached 35 or more responses. Following acquisition, the animals were given access to 40 injections per day for a period of 5 consecutive days. In this study all animals took the maximum number of injections (40) on each of the self-administration days. Control animals were naïve rats housed under the same reversed light-dark light cycle for at least one week prior to neurochemical analysis. Fast Scan Cyclic Voltammetry in Brain Slices: Fast scan cyclic voltammetry was used to characterize presynaptic DA signaling in the NAc. Voltammetry experiments were conducted during the dark phase of the light cycle, 24 hours after commencement of the final self-administration session. 400 µm thick coronal brain sections containing the NAc
85 64 core were cut using a vibrating tissue slicer (Leica Microsystems Inc.; Buffalo Grove, IL). A carbon fiber electrode was placed approximately 75 µm below the surface of the slice in close proximity to a bipolar stimulating electrode in the NAc core. DA release was evoked by a single electrical pulse (300 μa, 4 msec, monophasic) applied every 5 minutes. Extracellular DA was recorded by applying a triangular waveform ( 0.4 to +1.2 to 0.4 V vs. silver/silver chloride, 400 V/sec) to the electrode every 100 msec. Locomotor Analysis: Locomotor analysis was performed, as previously described (Lack et al ), the day following completion of cocaine self-administration. On the test day, prior to locomotor recording, animals were allowed to habituate in the testing room, in their home cages, for 60 minutes. Following habituation to the room, rats were placed in the locomotor chamber (MedAssociates; St. Albans, VT) and baseline activity recorded for 30 min. Rats then received an IP injection of saline, and activity was recorded for 90 min. Lastly, rats were injected with 15 mg/kg cocaine i.p., and locomotor response was measured again. Locomotor recordings were performed in two separate groups (control and cocaine self-administration) for between-subject comparisons. Microdialysis: Microdialysis surgery was performed the day following locomotor activity analysis. Microdialysis guide cannulae (CMA/Microdialysis AB, Stockholm, Sweden) were stereotaxically implanted above the NAc core (anteroposterior, mm; lateral, 2.0 mm; ventral, 6.0 mm). Concentric microdialysis probes (2 mm membrane length, CMA/Microdialysis) were inserted the day before recording and approximately 16 h prior to the beginning of sample collection. Probes were implanted in animals the day following locomotor testing and then microdialysis experiments were conducted the following day. Thus, rats were tested 72 h following their final SA session. The probes
86 65 were continuously perfused at 0.8 μl/min with artificial cerebrospinal fluid (acsf; ph 7.4): NaCl 148 mm, KCl 2.7 mm, CaCl mm, MgCl mm. Once stable baselines were established, animals received a cocaine (15 mg/kg) i.p. challenge and DA was monitored at 15 minute intervals until DA levels returned to baseline. High Performance Liquid Chromatography: The HPLC (ESA/Thermo Scientific, Chelmsford, MA) consisted of a syringe pump, a glassy carbon working electrode, a reference electrode, and an electrochemical detector. A 2 50 mm (3 µm particle) reverse-phase column (Luna, Phenomenex, Torrance, CA) was used to separate compounds. The applied potential was +650 mv as referenced to an Ag/AgCl electrode. The mobile phase (75 mm NaH 2 PO 4, 1.7 mm 1-octanesulfonic acid sodium salt, 100 µl/l triethylamine, 25 µm EDTA, 10% acetonitrile v/v, ph=3.0) was pumped at a rate of 170 µl/min, with a detection limit for DA of 10 pm. DA quantification was achieved by comparing dialysate samples with DA standards of known concentration. Statistics: Graph Pad Prism (version 5, La Jolla, CA, USA) was used to statistically analyze data sets and create graphs. Self-administration data was subjected to a one-way analysis of variance (ANOVA), and differences across sessions were tested using a Tukey post hoc test. Baseline voltammetry data was compared using a two-tailed Student s t-test. Locomotor data was subjected to a two-way ANOVA with experimental group and time as the factors. Differences between groups were tested using a Bonferroni post hoc test.
87 66 Results: Cocaine self-administration results in escalation of first-hour intake over sessions. Animals self-administered cocaine (1.5 mg/kg/inj) for five consecutive days in 6 hour sessions with a maximum of 40 total injections. Injections were limited in order to ensure that animals all had the same intake over the 5 self-administration sessions, to control for total intake as a factor in differences in neurochemical effects. Because of the limit on the total number of injections, animals did not escalate in total intake over the entire session. However, traditional long-access self-administration paradigms use increases in first-hour intake as a marker of escalation (Ahmed and Koob, 1998, 1999; Marusich et al., 2010). ANOVA revealed a main effect of session on first hour intake (F 4, 10 = 9.515, p < , Fig. 1). Tukey post hoc analysis revealed a significant increase in rate of intake on sessions 3 (p < 0.01), 4 (p < 0.001), and 5 (p < 0.001) versus session 1. Stimulated DA release and uptake are reduced following cocaine selfadministration. Using fast scan cyclic voltammetry, we assessed presynaptic DA signaling parameters in the NAc core to determine the effects of cocaine self-administration on DA release and uptake when drug is not present. We found significant reductions in DA release elicited by single pulse electrical stimulations after cocaine self-administration (Fig. 2A). Stimulated release was reduced by 44% versus controls (t 45 = 2.915, p < 0.001; Fig. 2B). In addition to stimulated release, we also found reductions in the maximal rate of DA uptake (V max ; Fig 2C). V max was reduced by 32% versus controls (t 42

88 67 Figure 1. Cocaine self-administration results in escalation of first-hour cocaine intake. Group data presented as cocaine injections within the first hour of the selfadministration session over consecutive sessions. The graph demonstrates that first-hour cocaine intake increases over session, indicating escalation over days. **, p < 0.01; ***, p < 0.001
89 68 Figure 2. Reduced dopamine (DA) release and uptake following cocaine selfadministration. (A) Representative data indicating that stimulated DA release is reduced following cocaine self-administration in the nucleus accumbens core. Plots are represented as DA concentration over time. (B) Group changes in μm DA released upon stimulation of the terminal. Cocaine self-administration results in a decrease in stimulated DA release relative to control animals. (C) Representative traces, overlaid to match for concentration, to highlight differences in maximal rate of DA uptake in the nucleus accumbens core. (D) Group data indicate that cocaine self-administration results in reduced maximal rate of uptake as compared to control animals. **, p < 0.01; SA, self-administration; V max, maximal rate of dopamine uptake
90 69 = 3.123, p < 0.001; Fig. 2D). These reductions in presynaptic DA measures demonstrate that DA signaling and DAT function is reduced following cocaine self-administration. DAT sensitivity to cocaine is reduced following self-administration. Because the DAT is the main site of action of cocaine for elevating DA, and for producing reinforcement, we aimed to determine if the effects of cocaine at the DAT were altered following extended-access, defined-intake cocaine self-administration. We found a reduction in cocaine-induced DA uptake inhibition in the NAc core region by a number of potency measures (Fig. 3). A two-way repeated measures ANOVA revealed a main effect of dose (F 4, 14 = 231.3, p < ) indicating that cocaine dose-dependently inhibits DA uptake (Fig. 3A). In addition there was a main effect of group (F 1, 14 = 38.35, p < ; Fig. 3B) indicating that the DAT was less sensitive to cocaine-induced DA uptake inhibition following cocaine self-administration. Bonferroni post hoc analysis revealed significant effects at the 10µM (p < 0.01) and 30 µm (p < 0.001) concentrations. K i is the drug concentration that results in 50% uptake inhibition. This measurement can determine relative potencies between drugs in naïve animals as well as potency changes following drug self-administration. The K i for cocaine was significantly higher in animals with a history of cocaine self-administration (t 14 = 5.312, p < ; Fig. 3C), indicating that cocaine potency was reduced. Cocaine-induced DA overflow was reduced following a history of cocaine selfadministration. In order to determine if the changes in DA kinetics following cocaine self-administration translated to reductions in extracellular DA levels, we performed microdialysis in freely-
91 70 moving animals approximately 72 hrs following the last self-administered cocaine infusion (Fig. 3D). There were no statistically significant differences in basal DA levels across groups. Systemic administration of a challenge dose of cocaine (15 mg/kg, i.p.) markedly increased DA levels in naïve rats (t 7 = 5.654, p < 0.001) but failed to produce significant changes in DA in cocaine self-administering rats. In addition, a two-way repeated measures ANOVA revealed a main effect of cocaine self-administration cocaine-induced DA overflow as compared to control animals (F 1, 12 = 16.39, p < 0.01). Bonferroni post hoc analysis revealed significant differences at the 40 (p < 0.001), 60 (p < 0.001), 80 (p < 0.01), 100 (p < 0.01), and 120 (p < 0.05) minute time points. These differences indicate that extended-access cocaine self-administration results in neurochemical tolerance to the DA elevating effects of systemically administered cocaine. Behavioral analysis showed no differences in basal locomotor activity, but tolerance to cocaine effects following self-administration. We used locomotor analysis to determine the effects of prior cocaine self-administration on the psychomotor activating effects of a cocaine challenge (15 mg/kg i.p.; Fig. 4). Figure 5A illustrates the time course of locomotor activity, with habituation to the chamber (30 min) followed by saline-induced locomotion (90min) and finally cocaineinduced locomotion. Saline-induced locomotion was not significantly different across groups (Fig. 4 A, inset), demonstrating that differences in cocaine-induced locomotor activity are not due to differences in response to injection. As expected, cocaine resulted in increased locomotion in all groups [Cocaine self-administration (F 6, 18 = 5.114, p < ); Control (F 6, 18 = 6.641, p < )]. A repeated measures ANOVA revealed a
92 71
93 72 Figure 3. Cocaine self-administration results in tolerance to the dopamine transporter (DAT)-inhibiting and dopamine (DA)-elevating effects of cocaine. (A) Representative traces normalized to peak height and plotted over time highlighting differences in 10μ cocaine-induced DA uptake inhibition in the nucleus accumbens core. Data indicate that cocaine is less effective at inhibiting DA uptake following a history of cocaine self-administration. (B) Group data of cocaine potency as plotted as apparent K m over cocaine concentration. Cocaine potency is reduced over a dose-response ( μm) following cocaine self-administration. (C) The K i for cocaine was significantly increased following cocaine self-administration. Increased K i values indicate that higher concentrations of drug are needed, relative to controls, to achieve the same degree of DA uptake inhibition. (D) DA overflow in the NAc was measured by freely moving microdialysis. Pre-drug baselines were determined, animals were injected with 15 mg/kg cocaine i.p., and cocaine-induced elevations in DA overflow were monitored. *, p < 0.05; **, p < 0.01; ***, p < 0.001; SA, self-administration
 Group data of locomotor activity as measured in cm over time plotted in five-minute bins. Data indicate that a history of cocaine self-administration results in reduced cocaine (15mg/kg; i.p.)-induced locomotor activity.")
94 73 Figure 4. Cocaine self-administration results in tolerance to the locomotor activating effects of acute cocaine administration. (A) Group data of locomotor activity as measured in cm over time plotted in five-minute bins. Data indicate that a history of cocaine self-administration results in reduced cocaine (15mg/kg; i.p.)-induced locomotor activity. (A, inset) Saline-induced locomotion was not different between controls and animals with a history of cocaine self-administration. (B) Cocaine-induced locomotor activity as represented as percent control. (C) Stereotypy as represented as average stereotypy counts per five-minute bin over the 90-minute session. (D) Vertical activity as represented as average vertical counts per five-minute bin over the 90-minute session. *, p < 0.05; **, p < 0.01; ***, p < 0.001; SA, self-administration
95 74 main effect of cocaine self-administration on cocaine-induced locomotor activity (F 1, 12 = 27.49, p < Fig. 4 A, B) and a significant Time Self-Administration interaction (F 17, 12 = 4.055, p < ). Bonferroni post hoc analysis revealed significant differences at time points minutes post-injection (Fig. 4 A). Further, locomotor activity was significantly different between groups when it was presented as total distance traveled in the first 30 min (t 12 = 2.665, p < 0.05), 90 min (t 12 = 1.925, p < 0.05) or as percent baseline (t 12 = 3.388, p < 0.01; Fig. 5 B). Thus, cocaine is less effective at stimulating locomotor activity following cocaine self-administration, which is indicative of tolerance to the psychomotor activating effects of cocaine. In order to assess changes in activity, in addition to distance traveled we assessed stereotypy behavior and vertical activity following a cocaine challenge. Student s t-test revealed that cocaine self-administration animals had lower stereotypy counts as compared to controls (t 12 = 5.206, p < ; Fig. 5 C). In addition, cocaine selfadministration animals had higher vertical counts as compared to controls (t 12 = 7.640, p < ; Fig. 5 D). These results, taken with the lack of difference in distance traveled, highlight a change in spontaneous locomotor activity, distinct from forward activity. Discussion: Here we demonstrate that a history of extended-access, defined-intake cocaine selfadministration results in robust tolerance to the neurochemical and psychomotor activating effects of cocaine. In the absence of drug, at baseline, DA system deficits, characterized by reductions in DA release and uptake in the NAc were also observed. The sensitivity of the DAT to cocaine-induced DA uptake inhibition was reduced, which translated to reduced cocaine-induced overflow, indicating neurochemical tolerance. The
96 75 neurochemical data was supported by behavioral data that demonstrated an attenuated cocaine-induced locomotor activating response. Psychomotor effects are hypothesized to predict the rewarding effects of drugs, suggesting that cocaine-induced reward may be reduced following cocaine self-administration (Wise and Bozarth, 1987). Because of the critical role of the DA system in reward, reinforcement, goal-reward associations, and motivated behaviors, the changes observed here could have profound inhibitory effects on goal-directed behavior in the absence of cocaine as well as increased cocaine-related behaviors, such as cocaine seeking/intake in order to normalize DA deficits. Because of the critical role of the NAc core in cue-reward association, assignment of motivational value, goal oriented behaviors, and the reinforcing properties of psychostimulant drugs, it was important to determine the effects of cocaine selfadministration on this region (Willuhn et al., 2010). Here we show decreased DA function, expressed as reduced DA release and uptake. These DA deficits could result in reduced emphasis on, and engagement in, normal behaviors associated with obtaining and processing rewards as well as an increased focus on drug seeking in order to normalize DA levels. Although previous work has demonstrated reduced basal DA levels as measured by microdialysis following cocaine self-administration (Ferris et al., 2011; Hurd et al., 1989; Mateo et al., 2005; Maisonneuve et al., 1995; Weiss et al., 1992) here we found no difference in basal DA concentrations. This may be explained by differences in the time of microdialysis measurements relative to the final cocaine selfadministration session, where measurements for the current study occurred 72 hours post self-administration as compared to 24 hours for previous work. This suggests that within this time period changes in baseline DA are normalized; however, tolerance to cocaine,
97 76 both neurochemical and behavioral, remains. This further demonstrates that differences in baseline DA functioning are not driving the cocaine tolerance, but rather they are two completely separate phenomena, which could both potentially affect drug-seeking behavior in cocaine addicts. Only recently was it discovered that the effects of cocaine on the DA transporter were reduced following extended access cocaine self-administration (Calipari et al., 2012, 2013; Ferris et al., 2011, 2012, 2013). Here we replicate the reduced sensitivity of the DAT to cocaine, and demonstrate that this neurochemical tolerance at the DAT is also observed with DA overflow as measured by microdialysis, where cocaine failed to significantly increase DA levels following cocaine self-administration. This is consistent with previous work demonstrating reductions in cocaine-induced DA release as a function of cocaine experience under various paradigms (Ferris et al, 2011; Hurd et al, 1989; Mateo et al., 2005; Meil et al., 1995). The decreased DA overflow elicited by cocaine administration is likely due to the decreased ability of the compound to block the DAT. This is consistent with human studies, which have demonstrated a reduced ability of cocaine to elevate DA levels following a history of cocaine abuse (Dackis and O Brien, 2001; Volkow et al., 1997a, b; Volkow et al., 1996). The ability of cocaine to elevate DA in the NAc is essential for its rewarding and reinforcing effects, thus reductions in cocaine-induced DA increases following this paradigm would be hypothesized to reduce the subjective effects of cocaine. There are numerous human reports supporting this idea, where there are reduced positive subjective effects of cocaine in heavy users (Mendelson et al., 1998; Reed et al., 2009). In addition, human cocaine addicts often increase their intake in order to compensate for reductions in the
98 77 rewarding effects of cocaine (Dackis and O Brien, 2001). These clinical findings are consistent with the current study, where we find increased intake rates for cocaine over sessions and decreased cocaine-induced locomotor activity. We report here that animals escalate the rate of cocaine intake over sessions. Although this paradigm is different from traditional long-access models (LgA) due to the fixed injection maximum, animals still escalate in first hour intake, which is the same escalation that is often reported in LgA studies (Ahmed and Koob, 1998, 1999; Marusich et al., 2010). Escalation is thought to be associated with the switch from abuse to addiction in humans (Ahmed and Koob, 1998, 1999), and has been suggested to be due to tolerance to the rewarding effects of cocaine (Ahmed et al., 2002, 2003; Ahmed and Koob, 2005; Barrett et al., 2004; Koob and Le Moal, 2001). The observed changes, where animals consume more drug in shorter periods of time, mimic changes in the patterns of self-administration when a lower dose of cocaine is substituted for a higher one, which would be consistent with reduced cocaine effects, or tolerance (Carelli and Deadwyler, 1996). A similar escalation in cocaine dosing also commonly occurs during bingeing in humans (Dackis and O Brien, 2001), and humans self-report tolerance to the euphorigenic effects of cocaine following repeated use (Mendelson et al., 1998; Reed et al., 2009). It is possible that the DAT tolerance, and concomitant reduction in cocaineinduced DA overflow, results in reduced rewarding effects, leading to an increase of cocaine intake to compensate. Further, locomotor analysis also revealed an altered pattern of cocaine-induced activity, with decreases in forward locomotion and stereotypical behaviors and increases in vertical activity. Elevations in DA levels mediate forward locomotion, thus, decreases
99 78 are consistent with the reductions in cocaine potency at the DAT as well as cocaineinduced DA overflow. Additionally, D2 receptor activation is critical for stereotyped behaviors, thus it is possible that D2 activity is reduced following cocaine selfadministration. Indeed, Mateo et al., (2005) demonstrated that D2 receptors are subsensitive following 10 days of cocaine self-administration. In addition, the reduction in D2 activity was observed at both one and 7 days after self-administration, indicating that some of the changes that occur following this self-administration paradigm may persist well into the withdrawal period. In addition to DA system changes, serotonin levels are inversely related to vertical activity (Brookshire and Jones, 2009); thus, it is possible that the serotonin system may also be affected by self-administration. Studies of whole brain functional activity after this self-administration paradigm have observed reduced dorsal raphe activity (Calipari et al., 2013). This is not surprising as cocaine has direct effects on the serotonin transporter, so it is possible that tolerance to the serotonin elevating effects also develops. Both tolerance and sensitization can develop to the neurochemical effects of cocaine. The development of tolerance is dependent on cocaine intake, where high sustained cocaine levels over a session result in reduced cocaine potency. Alternatively, cocaine sensitization at the DA terminal occurs following intermittent patterns of selfadministration and seems to be independent of intake (Calipari et al., 2013). Much work has been aimed at outlining the consequences and causes of neurochemical sensitization as a model of cocaine addiction (Brown et al., 2011; Cornish and Kalivas 2001; Thomas et al., 2008), and yielded important information, especially on changes in glutamate neurotransmission (Cornish and Kalivas 2001). However, sensitization of cocaine
100 79 pharmacology has been difficult to demonstrate in humans. Only recently was sensitization in humans demonstrated with amphetamine administration; however, it only occurs in drug naïve individuals and is not present in heavy users (Bolieau et al., 2006). It is possible that sensitization occurs with intermittent exposure to drug during the early stages of the addiction process, but later stages, with higher and more continuous drug intake, result in tolerance. As a result, animal models which produce tolerance have more face validity for studying the neurochemical changes that occur within addicted individuals. In fact, similar extended access paradigms, which result in neurochemical tolerance, still show measures of sensitized drug seeking and relapse which are characteristics of human addicts (Koob 1996; Koob and Le Moal, 1997; Orio et al, 2009; Patterson and Markou, 2003). As a result, when studying the DA system, it is important to choose a model that has translational validity, such as extended access paradigms that result in DA system deficits and cocaine tolerance. Acknowledgements: This work was funded by NIH grants R01 DA021325, R01 DA030161, P50 DA06634 (SRJ), K99 DA (MJF), T32 DA and F31 DA (ESC).
101 80 References Ahmed SH, Koob GF. (1998) Transition from moderate to excessive drug intake: change in hedonic set point. Science, 282(5387): Ahmed SH, Koob GF. (1999) Long-lasting increase in the set point for cocaine selfadministration after escalation in rats. Psychopharmacology (Berl)., 146(3): Ahmed SH, Kenny PJ, Koob GF, Markou A. (2002) Neurobiological evidence for hedonic allostasis associated with escalating cocaine use. Nat Neurosci. 5(7): Ahmed SH, Lin D, Koob GF, Parsons LH. (2003) Escalation of cocaine selfadministration does not depend on altered cocaine-induced nucleus accumbens dopamine levels. J Neurochem. 86(1): Ahmed SH, Koob GF. (2005) Transition to drug addiction: a negative reinforcement model based on an allostatic decrease in reward function. Psychopharmacology (Berl). 180(3): Barrett AC, Miller JR, Dohrmann JM, Caine SB. (2004) Effects of dopamine indirect agonists and selective D1-like and D2-like agonists and antagonists on cocaine selfadministration and food maintained responding in rats. Neuropharmacology. 47 Suppl 1: Boileau I, Dagher A, Leyton M, Gunn RN, Baker GB, Diksic M, Benkelfat C. (2006) Modeling sensitization to stimulants in humans: an [11C]raclopride/positron emission tomography study in healthy men. Arch Gen Psychiatry. 63(12): Brookshire BR, Jones SR. (2009) Direct and indirect 5-HT receptor agonists produce gender-specific effects on locomotor and vertical activities in C57 BL/6J mice. Pharmacol Biochem Behav., 94(1): Brown TE, Lee BR, Mu P, Ferguson D, Dietz D, Ohnishi YN, Lin Y, Suska A, Ishikawa M, Huang YH, Shen H, Kalivas PW, Sorg BA, Zukin RS, Nestler EJ, Dong Y, Schlüter OM. (2011) A silent synapse-based mechanism for cocaine-induced locomotor sensitization. J Neurosci. 31(22): Calipari ES, Ferris MJ, Melchior JR, Bermejo K, Salahpour A, Roberts DC, Jones SR. (2012) Methylphenidate and cocaine self-administration produce distinct dopamine terminal alterations. Addict Biol. In press. Calipari ES, Ferris MJ, Zimmer BA, Roberts DCS, Jones SR. (2013) Temporal pattern of cocaine intake determines tolerance versus sensitization of cocaine effects at the dopamine transporter. Neuropsychopharm. In press. Calipari ES, Beveridge TJR, Jones SR, Porrino LJ (2013) Withdrawal from extended access cocaine self-administration results in dysregulated functional activity and altered locomotor activity in rats. European J. Neurosci. In press.
102 81 Carelli RM, Deadwyler SA. (1996) Dose-dependent transitions in nucleus accumbens cell firing and behavioral responding during cocaine self-administration sessions in rats. J Pharmacol Exp Ther. 277(1): Chen R, Tilley MR, Wei H, Zhou F, Zhou FM, Ching S, Quan N, Stephens RL, Hill ER, Nottoli T, Han DD, Gu HH. (2006) Abolished cocaine reward in mice with a cocaineinsensitive dopamine transporter. Proc Natl Acad Sci U S A. 103(24): Cornish JL, Kalivas PW. (2001) Cocaine sensitization and craving: differing roles for dopamine and glutamate in the nucleus accumbens. J Addict Dis. 20(3): Dackis CA, O'Brien CP. (2001) Cocaine dependence: a disease of the brain's reward centers. J Subst Abuse Treat. 21(3): Ferris MJ, Mateo Y, Roberts DC, Jones SR. (2011) Cocaine-insensitive dopamine transporters with intact substrate transport produced by self-administration. Biol Psychiatry., 69(3): Ferris MJ, Calipari ES, Mateo Y, Melchior JR, Roberts DC, Jones SR. (2012) Cocaine self-administration produces pharmacodynamic tolerance: differential effects on the potency of dopamine transporter blockers, releasers, and methylphenidate. Neuropsychopharmacology, 37(7): Ferris MJ, Calipari ES, Melchior JR, Roberts DCS, Espana RA, Jones SR. (2013) Paradoxical tolerance to cocaine after initial supersensitivity in drug-use-prone animals. Eur J Neurosci. In press. Hurd YL, Weiss F, Koob GF, And NE, Ungerstedt U. (1989) Cocaine reinforcement and extracellular dopamine overflow in rat nucleus accumbens: an in vivo microdialysis study. Brain Res. 498(1): Koob GF. (1996) Hedonic valence, dopamine and motivation. Mol Psychiatry. 1(3): Koob GF, Le Moal M. (1997) Drug abuse: hedonic homeostatic dysregulation. Science, 278:52-8. Koob GF, Le Moal M. (2001) Drug addiction, dysregulation of reward, and allostasis. Neuropsychopharmacology. 24(2): Lack CM, Jones SR, Roberts DC. (2008) Increased breakpoints on a progressive ratio schedule reinforced by IV cocaine are associated with reduced locomotor activation and reduced dopamine efflux in nucleus accumbens shell in rats. Psychopharmacology (Berl)., 195(4): Maisonneuve IM, Ho A, Kreek MJ. (1995) Chronic administration of a cocaine "binge" alters basal extracellular levels in male rats: an in vivo microdialysis study. J Pharmacol Exp Ther. 272(2):652-7.
103 82 Marusich JA, Beckmann JS, Gipson CD, Bardo MT. (2010) Methylphenidate as a reinforcer for rats: contingent delivery and intake escalation.exp Clin Psychopharmacol. 18(3): Mateo Y, Lack CM, Morgan D, Roberts DC, Jones SR. (2005) Reduced dopamine terminal function and insensitivity to cocaine following cocaine binge self-administration and deprivation. Neuropsychopharmacology, 30(8): Meil WM, Roll JM, Grimm JW, Lynch AM, See RE. (1995) Tolerance-like attenuation to contingent and noncontingent cocaine-induced elevation of extracellular dopamine in the ventral striatum following 7 days of withdrawal from chronic treatment. Psychopharmacology (Berl) Apr;118(3): Erratum in: Psychopharmacology (Berl) 121(2):285. Mendelson JH, Sholar M, Mello NK, Teoh SK, Sholar JW. (1998) Cocaine tolerance: behavioral, cardiovascular, and neuroendocrine function in men. Neuropsychopharmacology.18(4): Orio L, Edwards S, George O, Parsons LH, Koob GF. (2009) A role for the endocannabinoid system in the increased motivation for cocaine in extended-access conditions. J Neurosci. 29(15): Paterson NE, Markou A. (2003) Increased motivation for self-administered cocaine after escalated cocaine intake. Neuroreport. 14(17): Reed SC, Haney M, Evans SM, Vadhan NP, Rubin E, Foltin RW. (2009) Cardiovascular and subjective effects of repeated smoked cocaine administration in experienced cocaine users. Drug Alcohol Depend. 102(1-3): Ritz MC, Lamb RJ, Goldberg SR, Kuhar MJ. (1987) Cocaine receptors on dopamine transporters are related to self-administration of cocaine. Science. 237(4819): Roberts DC, Corcoran ME, Fibiger HC. (1977) On the role of ascending catecholaminergic systems in intravenous self-administration of cocaine. Pharmacol Biochem Behav. 6(6): Thomas MJ, Kalivas PW, Shaham Y. (2008) Neuroplasticity in the mesolimbic dopamine system and cocaine addiction. Br J Pharmacol. 154(2): Volkow ND, Wang GJ, Fowler JS, Logan J, Hitzemannn R, Gatley SJ, MacGregor RR, Wolf AP. (1996) Cocaine uptake is decreased in the brain of detoxified cocaine abusers. Neuropsychopharmacology. 14(3): Volkow ND, Wang GJ, Fowler JS, Logan J, Gatley SJ, Hitzemann R, Chen AD, Dewey SL, Pappas N. (1997a) Decreased striatal dopaminergic responsiveness in detoxified cocaine-dependent subjects. Nature. 386(6627): Volkow ND, Wang GJ, Fowler JS. (1997b) Imaging studies of cocaine in the human brain and studies of the cocaine addict. Ann N Y Acad Sci. 820:41-54; discussion 54-5.
104 83 Weiss F, Markou A, Lorang MT, Koob GF. (1992) Basal extracellular dopamine levels in the nucleus accumbens are decreased during cocaine withdrawal after unlimited-access self-administration. Brain Res. 593(2): Willuhn I, Wanat MJ, Clark JJ, Phillips PE. (2010) Dopamine signaling in the nucleus accumbens of animals self-administering drugs of abuse. Curr Top Behav Neurosci. 3: Wise RA, Bozarth MA. (1987) A psychomotor stimulant theory of addiction. Psychol Rev. 94(4):
105 84 CHAPTER III WITHDRAWAL FROM EXTENDED-ACCESS COCAINE SELF- ADMINISTRATION RESULTS IN DYSREGULATED FUNCTIONAL ACTIVITY AND ALTERED LOCOMOTOR ACTIVITY IN RATS Erin S. Calipari, Thomas J.R. Beveridge, Sara R. Jones and Linda J. Porrino The following manuscript was published in European Journal of Neuroscience, in press, 2013, and is reprinted with permission. Stylistic variations are due to the requirements of the journal. Erin S. Calipari performed all self-administration experiments, data analysis for self-administration studies, and prepared the manuscript. Dr. Thomas Beveridge performed all functional activity experiments. Drs. Sara R. Jones and Linda Porrino acted in an advisory and editorial capacity.
106 85 Abstract: Much work has been focused on determining the consequences of cocaine selfadministration on specific neurotransmitter systems, thus neglecting the global changes that occur. Previous imaging studies have focused on the effects of cocaine selfadministration in the presence of high blood levels of cocaine, and do not determine the effects of cocaine self-administration on baseline functional activity. Extended access cocaine self-administration, where animals administer cocaine for 6 hours each day, results in escalation in the rate of cocaine intake and is believed to model the transition from recreational use to addiction in humans. We aimed to determine the functional changes following acute (48 hours) withdrawal from animals that have undergone extended access self-administration for 5 days, a time point when behavioral changes are present. Using the 2-[ 14 C]deoxyglucose method to measure rates of local cerebral glucose metabolism, an indicator of functional activity, we found reductions in circuits related to learning and memory, attention, sleep, and reward processing, which have important clinical implications for cocaine addiction. Additionally, lower levels of functional activity were found in the dorsal raphe and locus coeruleus, suggesting that cocaine self-administration may have broader effects on brain function than previously noted. These widespread neurochemical reductions were concomitant with substantial behavioral differences in these animals, highlighted by increased vertical activity and decreased stereotypy, which would be predicted based on the alterations in the dopaminergic and serotonergic nuclei. These data demonstrate that behavioral and neurochemical impairments following cocaine self-administration are present at baseline and persist after cocaine has been cleared.
107 86 Introduction: The neuroadaptations that occur following cocaine administration have been studied extensively in an effort to determine both the consequences of cocaine abuse and to find potential targets for addiction treatment. Previous work using non-contingent cocaine exposure has shown significant neuroadaptations in gene expression, protein function, neurotransmitter release and uptake, and concomitant behavioral changes (Vanderschuren & Pierce, 2010; Mu et al., 2010). However it is important to choose a model that accurately mimics human drug taking (Hemby et al., 1994, 1997; Porrino 1993; Lecca et al., 2007). Rodent self-administration is a translational model of human cocaine abuse, and much of the current literature on cocaine self-administration has focused on extended access self-administration, during which animals have access to cocaine self-administration for 6 hours (h) per day (Ahmed & Koob, 1998). This paradigm allows animals to administer consistent high levels of cocaine over the session and has been reported to accurately model the pattern by which human addicts administer cocaine (Dackis and O Brien, 2001). Extended access cocaine self-administration has been shown previously to have many of the neurochemical hallmarks of cocaine addicts and is characterized by reduced basal dopamine levels (Mateo et al., 2005; Ferris et al., 2011), behavioral and neurochemical tolerance to cocaine (Ferris et al., 2011, 2012: Calipari et al., 2012), and increased motivation to administer cocaine (Zimmer et al., 2012). Most of the current literature on the effects of chronic cocaine self-administration on the brain has focused on the striatal dopamine system, thus neglecting the contribution from other neurotransmitters and circuits. Neuroimaging techniques allow for a global
108 87 view of the effects of cocaine self-administration on the brain. Previous work has demonstrated that immediately following 21 days of self-administration, while blood levels of cocaine are still high, there are reductions in functional activity in a number of brain regions, including striatal regions (Macey et al., 2004); however, the question of whether these changes persisted beyond the self-administration session remained. Most functional activity studies determine the effects of a drug challenge; however, the present study focused on rates of local cerebral glucose utilization in the absence of drug, in order to determine the effects on brain function. These data are important because determining the persistent effects of cocaine self-administration on baseline functional activity can point to changes in specific brain regions and circuits which may be predictive of behavioral deficits in cocaine-addicted individuals. We show that cocaine self-administration results in functional reductions in brain regions involved in reward, learning, and memory that are present 48 h after the final cocaine session. We also see reduced function of the dorsal raphe and locus coeruleus, which has implications for global brain function as these nuclei have an extensive network of projections. Using behavioral activity analysis after cocaine selfadministration we report alterations which would be predicted based on decreased serotonergic and dopaminergic functioning, demonstrating that these neural changes have behavioral implications. The reduced functional activity in selected regions suggests that a 5-day cocaine binge is capable of producing reductions in regional brain activity that may have adverse consequences for normal functioning.
109 88 Materials and Methods Animals: Male, Sprague-Dawley rats ( g; Harlan Laboratories, Frederick, MD) were maintained according to the National Institutes of Health guidelines in Association for Assessment and Accreditation of Laboratory Animal Care accredited facilities. The experimental protocol was approved by the Institutional Animal Care and Use Committee at Wake Forest School of Medicine. Self-Administration: Rats (n = 14) were anesthetized with ketamine (100 mg/kg) and xylazine (10 mg/kg), implanted with chronic indwelling jugular catheters, and trained for i.v. self-administration as previously described (Liu et al., 2005). Following surgery, animals were singly housed, and all self-administration sessions took place in the home cage. Each animal was maintained on a reverse light cycle (3:00 am lights off; 3:00 pm lights on), and all self-administration procedures occurred during the active/dark cycle. Sessions were six hours in length and were terminated at the end of the six hours or after 40 injections of drug. Animals self-administered cocaine (1.5mg/kg/inj over 4 sec) on a fixed-ratio 1 schedule of administration. Concurrent with the start of each injection, the lever retracted and a stimulus light was activated for 20 seconds to signal a time-out period. Under these conditions, animals acquired a stable pattern of cocaine selfadministration within 1 to 5 days. For self-administering animals, acquisition (Day 1) was counted when the animal reached 35 or more responses. Following acquisition, the animals were given access to 40 injections per day for a period of 5 consecutive days. Control animals were naïve rats housed under the same reverse light-dark light cycle for at least one week prior to all experimental manipulations.
110 89 Locomotor Analysis: Locomotor analysis was performed as previously described (Läck et al. 2008) the day following completion of cocaine self-administration. Locomotor analysis was performed on a different group of animals from the functional activity experiments. On the test day, prior to locomotor recording animals were allowed to habituate in the testing room, in their home cages for 60 minutes. Following habituation to the room, rats (control, n=7; cocaine self-administration, n=7) were placed in the locomotor chamber (MedAssociates) and baseline activity recorded for 30 min. Rats then received an intraperitoneal (IP) injection of saline, and activity was recorded for 90 min. Locomotor recordings were performed in two separate groups (control and cocaine selfadministration) and data was compared across groups. Measurement of local cerebral glucose utilization: Twenty-four h after their final selfadministration session, animals underwent femoral artery catheterization surgeries, as previously described (Macey et al., 2004). Animals were allowed 24 h to recover from surgery. Rates of local cerebral glucose utilization (LCGU) in rat brain were quantified 48 h after their last cocaine self-administration session according to the method of Sokoloff et al. (1977), as adapted for use in freely moving animals (Crane and Porrino 1989). As part of a separate study, both cocaine self-administration animals (n=7) and controls (n=6) were administered saline (1 ml/kg, IP) 30 min prior to initiation of the [ 14 C]-2-deoxyglucose (2DG) procedure. One control animal was dropped from analysis due to an occluded femoral catheter. Control animals consisted of both naïve controls and animals that had undergone surgery but did not self-administer cocaine. Three of the six controls were animals that failed to self-administer/ acquire, thus, they underwent exactly the same surgical procedures as the cocaine self-administering animals, as well as
111 90 housing conditions, light/ dark cycle and handling. Rates of local cerebral glucose utilization in the brains of these three animals in response to saline injections were no different from the other controls, which were housed in similar conditions and handled in the same fashion. The 2DG procedure was conducted in the animal s home cage and was initiated by means of an intravenous infusion of a pulse of 2-[ 14 C]DG (75 μci/kg; specific activity 55 mci/mmol; New England Nuclear, Boston, MA) through the jugular venous catheter (via which self-administration had previously occurred). Timed femoral arterial blood samples were collected over the next 45 minutes and immediately centrifuged. Rates of LCGU in cocaine self-administering rats were compared with those obtained from drug-naïve rats. These control animals underwent jugular vein and femoral artery catheterization surgeries in the same fashion as self-administering rats, 24 h prior to the 2DG procedure. Plasma concentrations of 2-[ 14 C]DG were determined by liquid scintillation counting (Beckman Instruments, CA), while plasma glucose levels were determined by means of a Beckman Glucose Analyzer II (Beckman Instruments, CA). Immediately after the 45-minute sample, animals were killed by administration of intravenous pentobarbital (100 mg/kg). Brains were rapidly removed, frozen in isopentane at -45 C, and stored at -80 C until sectioned. Brains were sectioned coronally (20μm) in a cryostat maintained at -20 C, collected on glass coverslips and immediately transferred to a hot plate (60 C) to dry. Coverslips were opposed to Kodak EMC film for days along with a set of calibrated [ 14 C]methylmethacrylate standards (Amersham, IL) previously calibrated for their equivalent wet weight 14 C concentrations. Films were developed in GBX developer (Kodak, NY).
112 91 Autoradiograms were analyzed by quantitative densitometry with a computerized image analysis system (MCID, Imaging Research, Ontario). Tissue 14 C concentrations were determined from densitometric analysis of autoradiograms of the calibrated standards. Rates of glucose utilization were then calculated using the optical densities and a calibration curve obtained from local 14 C tissue concentrations, time-courses of the plasma glucose and 14 C concentrations, and the constants according to the operational equation of the 2DG method (Sokoloff et al. 1977). Glucose utilization measurements were determined for 20 discrete brain regions according to the rat brain atlas of Paxinos and Watson (Paxinos and Watson 1998). Each brain region was analyzed bilaterally in a minimum of five brain sections per animal. Statistics: Graph Pad Prism (version 4, La Jolla, CA, USA) was used to statistically analyze data sets and create graphs. Locomotor data was subjected to a two-way ANOVA with experimental group and time as the factors. Differences between groups were tested using a Bonferroni post hoc test. Rates of local cerebral glucose utilization were analyzed in specific functional groups of brain regions (cortex, basal ganglia, limbic-related and brainstem) using 2-way analyses of variance (ANOVA), brain region group X treatment (control vs. cocaine self-administration) with brain region group as a repeated measure. These were followed by planned Bonferroni s tests for multiple comparisons. Statistical significance was considered as p < Results: Extended access to cocaine self-administration results in escalation in the rate of daily intake. Animals self-administered cocaine (1.5 mg/kg/inj) for five consecutive days
113 92 with a maximum of 40 total injections (all animals self-administered the maximum each day). Injections were limited to ensure that all animals had the same intake over each self-administration session in order to control for total intake as a factor in the functional effects of cocaine. Because of the limit on total injections, therefore, animals cannot escalate their total intake, but rather escalate the rate of intake over the sessions (Figure 1), similar to previous studies (Mateo et al., 2005; Calipari et al., 2012; Ferris et al., 2012). One way ANOVA revealed a main effect of session (F 4, 14 = 14.93, p < 0.001). Tukey post hoc analysis revealed a increase in rate versus the first session on sessions two (q = 4.216, p < 0.05), three (q = 5.843, p < 0.01), four (q = 8.734, p < 0.001), and five (q = 9.673, p < ). Behavioral analysis shows a differential baseline locomotor profile following 48 h withdrawal from cocaine self-administration. Following cocaine self-administration, behavioral activity was assessed using automated monitors. We ran the analysis based on a priori hypothesis that locomotor activity in response to a novel environment would be reduced following cocaine self-administration. Two-way ANOVA revealed a main effect of cocaine self-administration on the response to a novel environment (F( 1, 13 ) = 4.04, p < 0.05). In addition, there was a main effect of time (F( 5, 13 ) = 73.53, p < ) as animals habituated to the activity chamber. Bonferroni post hoc analysis was performed to compare across treatment group, and a reduction in locomotor activity was observed at the 15 minute time point in animals that had undergone cocaine self-administration (t = 3.219, p <0.05; Figure 2). Alternatively, we found no effect of treatment or time on saline-induced locomotion indicating that normal forward locomotion was not impaired by a prior cocaine self-administration history (Figure 3).
114 93 Figure 1. Cocaine self-administration results in escalation of rate of cocaine intake. (A) Event record of a representative cocaine self-administration animal. Each line represents a daily self-administration (SA) session. Each tick mark represents an injection of cocaine (1.5 mg/kg/inj). Animals are allowed 40 injections or 6 hours, whichever comes first. Animals demonstrate regular inter-injection intervals, and while injections stay constant, they take less time over sessions to complete the 40 injections. (B) The animals showed escalation of drug intake by self-administering 40 injections each day in successively shorter time periods. (*p < 0.05, **p < 0.01, ***p < 0.001; SA, selfadministration)
115 94 Figure 2. Cocaine self-administration results in reduced locomotor response to a novel environment. Animals were placed in automated locomotor activity monitors and their behavior on the first exposure to the environment and the subsequent habituation was monitored for 30 minutes. (A) Locomotor activity, represented as distance traveled over time. Cocaine self-administration animals (open circles) habituated to the novel environment faster than their control counterparts (closed circles). (B) Total locomotor distance traveled in the novelty session. Animals with a history of cocaine selfadministration have significantly reduced responses to a novel environment. (*p < 0.05; SA, self-administration)
116 95 Figure 3. Cocaine self-administration does not alter baseline locomotion. Animals were placed in automated locomotor activity monitors and their behavior following a saline injection to determine the effects of cocaine self-administration on forward locomotor activity. (A) Locomotor activity, represented as distance traveled over time. Cocaine self-administration animals (open circles) were not significantly different in their response to a saline injection than control animals (closed circles). (B) Total locomotor distance traveled during the saline session. No differences were observed in normal baseline forward locomotion following cocaine self-administration. (SA, selfadministration)
117 96 In order to assess changes in locomotor behavior, in addition to distance traveled we assessed stereotypy behavior and vertical activity. Student s t-test revealed that cocaine self-administration animals had higher vertical counts as compared to controls (t 12 = 7.604, p < ; Figure 4). In addition, cocaine self-administration animals had lower stereotypy counts as compared to controls (t 12 = 3.988, p < ; Figure 4). These results, taken with the lack of difference in distance traveled, highlight a change in spontaneous locomotor activity, distinct from forward activity. Rates of local cerebral glucose utilization are lower following 48 h withdrawal from cocaine self-administration. Baseline plasma glucose concentrations prior to the initiation of the 2DG procedure were not different between drug-naïve controls and cocaine-experienced animals (controls, ± 6.7 mg/ml; 48 h withdrawal from cocaine, ± 17.0 mg/ml). Rates of local cerebral glucose metabolism were measured in 20 brain regions and data are shown in Table 1. These rates were globally lower in animals with a history of cocaine self-administration measured 48 h after the final self-administration session as compared to drug-naïve controls (84.5 ± 4.7 μmol 100 g min controls versus 74.6 ± 4.4 μmol 100 g min cocaine-withdrawal, p < 0.05). This pattern was observed in all 20 of the regions in which glucose utilization rates were measured. In the cortex, two-way analysis of variance revealed a main effect of treatment (F( 1, 11 ) = 5.95, p <0.05) and brain region (F( 2, 20 ) = 151.9, p <0.001), but no interaction. In the basal ganglia, there was a main effect of treatment (F( 1, 11 ) = 8.10 p <0.05) and brain region (F( 5, 55 ) = , p < 0.001), but no interaction. In limbic brain areas, there was a main effect of treatment (F( 1, 11 ) = 6.10 p <0.05) and brain region (F( 7,

118 97 Figure 4. Altered locomotor activity following cocaine self-administration. Animals were placed in automated locomotor activity monitors and their behavior following a saline injection to determine the effects of cocaine self-administration on stereotypy and vertical activity (A) Stereotypy as represented as average stereotypy counts per five minute bin over the 90 minute session. Cocaine self-administration animals (blue) had reduced stereotypy counts as compared to controls (black). (B) Vertical activity as represented as average stereotypy counts per five minute bin over the 90 minute session. Cocaine self-administration animals (blue) had increased vertical counts as compared to controls (black). (*** p < 0.001; SA, self-administration)
 and hippocampus (bottom row) of a representative control rat")
119 98 Figure 5. Cocaine self-administration results in reduced functional activity in a number of brain regions. Pseudocolor enhanced autoradiograms showing rates of local cerebral glucose metabolism in the striatum (top row) and hippocampus (bottom row) of a representative control rat (panel A) and a rat 48 h following cocaine self-administration (panel B).
120 99 Table 1. Effect of 48 h withdrawal from a five-day binge cocaine self-administration on rates of local cerebral glucose utilization in rat brain. Brain Region Cortex Drug-naïve Controls (n = 6) Cocaine-exposed (n = 7) Medial prefrontal 76.2 ± ± 3.2 Anterior cingulate ± ± 4.2 * Motor ± ± 5.7 Basal Ganglia Dorsal caudate ± ± 4.0 * Ventral caudate 96.8 ± ± 4.7 Nucleus accumbens 94.4 ± ± 3.8 ** Globus pallidus 53.8 ± ± 2.8 Substantia nigra pars compacta 65.2 ± ± 4.0 Substantia nigra pars reticulata 56.2 ± ± 2.7 Limbic Basolateral amygdala 81.3 ± ± 3.4 * Central amygdala 42.9 ± ± 2.1 Lateral thalamus 95.1 ± ± 5.9 Medial thalamus 97.7 ± ± 3.7 * Ventromedial hypothalamus 56.6 ± ± 3.0 Hippocampal CA1 region 78.6 ± ± 3.9 * Hippocampal CA3 region 81.2 ± ± 3.7 Dentate gyrus 64.5 ± ± 2.4 Brainstem
121 100 Dorsal raphe ± ± 7.5 * Locus coeruleus 73.8 ± ± 2.8 * Cerebellum 70.6 ± ± 2.4 * Values are local rates of glucose utilization expressed as mean (μmol/100g/min) ± S.E.M. * P<0.05, ** P<0.001 versus drug-naïve controls.
122 101 77) = 110.3, p < 0.001), and an interaction (F( 7,70 ) = p <0.05). Finally, in the brainstem, there was a main effect of treatment (F( 1, 11 ) = p <0.01) and brain region (F( 2, 22 ) = 75.21, p < 0.001), but no interaction. Planned multiple comparisons showed that 48 h after cocaine self-administration functional activity was lower in the anterior cingulate cortex ( 12%), dorsal caudate putamen ( 16%), nucleus accumbens ( 16%), basolateral amygdala ( 16%), medial nucleus of the thalamus ( 12%), hippocampal CA1 region ( 24%), dorsal raphe ( 18%), locus coeruleus ( 13%) and cerebellum ( 15%), when compared to controls. Discussion: Here we demonstrate that there are functional and behavioral reductions present 48 h after 5-day cocaine self-administration. The functional alterations were characterized by reduced brain activity, as indicated by lower rates of cerebral glucose utilization, in circuits involved in learning and memory, attention, sleep, and reward processing. These data are consistent with human studies that have demonstrated marked reductions in functional brain activity, in particular prefrontal cortical and striatal regions, following cocaine abuse (Volkow et al., 1992, 1993). Previously, we have shown that cocaine self-administration resulted in reductions in functional activity, but these effects were measured immediately following the final infusion at a time when cocaine levels were still high (Macey et al., 2004). The present study extended these findings and demonstrated that there were functional alterations 48 hours following the final selfadministration session, indicating that neuroadaptations associated with cocaine administration may persist and result in reduced activity. In addition, we found that there
123 102 were also behavioral changes, as indicated by increased vertical and reduced stereotypical behavior, suggesting that these functional alterations may have direct behavioral consequences even in the absence of drug. These results highlight the fact that even a relatively short (five day) exposure to cocaine, which resulted in escalation of the rate of daily intake, can lead to neurochemical (Mateo et al., 2005; Calipari et al., 2012; Ferris et al., 2012), functional and behavioral changes that can be detected during withdrawal. While previous reports have focused on the neurochemical changes that occur when drug is still present or in response to a challenge dose of cocaine (Macey et al., 2004; Kornetsky, Huston-Lyons, & Porrino, 1991; Zocchi, Conti, & Orzi, 2001), the present study focused on measuring functional activity 48 hours after the last selfadministration session. At this time point, we found that functional activity was lower than controls in the nucleus accumbens, anterior cingulate cortex, basolateral amygdala, hippocampus, dorsal caudate, medial thalamus, dorsal raphe, and locus coeruleus brain areas involved in vital processes such as learning, memory, reward and reinforcement. In contrast to these data, our previous work has examined the effects of five days of cocaine self-administration on functional activity, while cocaine was present, and found no changes in any of these regions. In fact, most of the regional differences from controls were higher, not lower, levels of functional activity (Macey et al., 2004). Macy et al., (2004) also compared the five-day group to a 30-day self-administration group that was different in duration from the current study, but similar in intake. In the current study, animals administered 100mg cocaine over the five days, while animals in the 30 -day group administered 150mg cocaine. The animals with the 30 -day self-administration history had similar changes to what was observed in the present study, suggesting that
124 103 intake, not duration, determines the adaptations that occur. This observation is consistent with previous work from our lab indicating that total intake, not the length of the selfadministration history, is responsible for neurochemical changes that occur following cocaine self-administration (Calipari et al., 2013). Because the 30-day self-administration group from Macey et al., (2004) was imaged with cocaine present, it suggests that not only are the circuits depressed in the absence of cocaine, but cocaine does not produce sufficient effects to normalize circuits. Continued drug-taking in addicted individuals has been suggested to occur as compensatory behavior to normalize a baseline dysregulated state (Koob, 2009; Koob and Le Moal, 1997). It is important to differentiate the effects of cocaine-induced alterations of neural networks with those of cocaine self-administration on functioning in the absence of drug, as they have very different implications for the functioning of the brain at baseline. The mesocorticolimbic DA system mediates many of the reinforcing and rewarding effects of cocaine (Pierce and Kumaresan, 2006), and because neuroadaptations resulting from chronic drug exposure are often opposite from the acute effects, it is not surprising that there were reductions in the activity of these regions following cocaine self-administration. Previous work has demonstrated reduced function of the striatal DA system at a similar time point following cocaine self-administration, as well as the development of tolerance to the neurochemical effects of cocaine (Ferris et al., 2011). Further, these functional reductions are present 18 hours following the final cocaine self-administration session (Mateo et al., 2005; Ferris et al., 2011, 2012; Calipari et al., 2012), and persist for up to two weeks following cocaine exposure (Ferris et al., 2012). These reductions in DA function have been observed using both fast scan cyclic
125 104 voltammetry and microdialysis where it was found that hours of withdrawal from cocaine self-administration resulted in reduced DA release and uptake, as well as reduced baseline DA overflow, respectively (Ferris et al., 2011, 2012; Calipari et al., 2012, 2013; Maisonneuve et al., 1995; Weiss et al., 1992; but see, Hooks et al., 1994; Meil et al., 1995). The current data agree with these observations in that functional activity was significantly reduced in the terminal fields of the VTA, namely the nucleus accumbens and caudate putamen (Koeltzow and White, 2003). Moreover, the neurochemical changes previously reported concur with the behavioral data, which demonstrated a decreased locomotor response to a novel environment following withdrawal from cocaine self-administration. Previous studies have demonstrated the influential role of striatal DA levels on the locomotor response to a novel environment; for example, animals can be separated into two groups (high and low response) according to their locomotor activity in reaction to a novel environment. Ferris et al., (2013) recently demonstrated that high and low response to novelty can predict both the tolerance that develops to cocaine directly at the dopamine transporter as well as the rate of acquisition of cocaine self-administration. Low novelty responders have been shown to have lower extracellular DA levels, in line with the present data where we observed reduced functional activity in dopaminergic regions following withdrawal from cocaine self-administration (Verheij and Cools, 2008; Verheij et al., 2008). Previous work on withdrawal from cocaine self-administration has found depression of locomotor activity during similar time-points, an effect that is indicative of reductions in DA levels and VTA cell firing (Koeltzow and White, 2003; Gauvin et al., 1997). The lower locomotor activity then would be predicted based on the
126 105 reduced functional activity in dopaminergic nuclei. These alterations suggest that there could be changes in reward processing at baseline, which could play an important role in the reinstatement of cocaine-seeking (Schmidt and Pierce, 2010). Because these regions are involved in the processing of salient stimuli, these data also suggest that the processing of other alternate rewards, such as food, may also be impaired at baseline (Carelli 2002; Schultz, 2010). In addition, there may be differential effects of cocaine on dopaminergic systems involved in motor and reward processing, an effect that was highlighted in the behavioral data indicating that there was no difference in baseline forward locomotion following cocaine self-administration. It is important to note that there were reductions in stereotypic behaviors, indicating that although forward locomotion does not differ between groups, there may be inherent differences in motor control following cocaine self-administration, although it is not clear as to the meaning of these results. Together, these data suggest that the alterations in functional activity are not general changes that occur in all dopaminergic terminal fields, but rather are specific to those associated with reward and reinforcement and specific aspects of motor control. In line with reductions in functional activity in dopaminergic regions 48 hours following cocaine self-administration, electrophysiological recordings have demonstrated reduced action potential firing in nucleus accumbens neurons both in vitro and in vivo after withdrawal from cocaine self-administration (Dong et al., 2006; Ishikawa et al., 2009; Kourrich and Thomas, 2009; Mu et al., 2010; White et al., 1995). The lower rates of glucose metabolism in the brain regions reported here, together with the reductions in cell firing could suggest a reduced/depressive state following acute withdrawal from selfadministration. Indeed, Markou and Koob (1991) have demonstrated elevations in
127 106 intracranial self-stimulation thresholds in rats, indicating a depressed or anhedonic state in animals following self-administration. The elevated thresholds were reversed by a DA D2 receptor agonist, suggesting that the effects were due, at least in part, to reduced DA system activity following a cocaine self-administration history (Markou & Koob, 1992). A similar phenomenon has also been demonstrated in mice, where withdrawal from cocaine delivered via osmotic mini-pump also resulted in elevated reward thresholds 72 hours following treatment (Stoker & Markou, 2011). Because functional activity measurements were made 48 hours following the final cocaine self-administration session, these modifications may be indicative of the changes that occur during early withdrawal periods and may coincide with the alterations in reward thresholds. Further, it is well documented that, similar to the anhedonic behavior seen in rodents, human addicts going through early withdrawal from cocaine exposure also report depression and anhedonia (Markou & Kenney, 2002). It is possible, therefore, that the widespread decreased functional activity may be associated with depression and anhedonia during the early withdrawal periods. Although it is possible that these changes may also underlie the neuroadaptations that occur during later stages of withdrawal, the time points that we measured in this study can only confirm that this state is present 48 hours following cocaine self-administration. Future studies that look at the time course of both the functional and behavioral effects of cocaine withdrawal are necessary to confirm whether the effects observed here are persistent. Although reward and reinforcement are an essential part of the early stages of the addiction process, drug addiction is dependent on neural plasticity associated with druginduced reward learning mechanisms (Jones and Bonci, 2005). Here we observe
128 107 functional reductions in brain regions involved in this type of learning. Specifically, the reductions in the prefrontal cortex (PFC) and nucleus accumbens activity suggest that there may be suppression of cortico-striatal loops. Goal-directed learning is reliant on the dorsomedial striatum through loops that project from the cortex to the striatum (Alexander et al. 1986; Lawrence et al. 1998; Haber and Calzavara 2009; McFarland and Haber 2002). Here we show reductions in functional activity in these areas, implying that this type of learning may also be impaired. These data suggest that 1.) individuals may be less able to learn new goal-directed behaviors, and 2.) that they also may be less able to replace already formed associations. Replacing associations that occurred during the development of drug addiction is a process that is essential for continued abstinence and the prevention of relapse in abstinent individuals. In addition, the motivational loop, comprised of the ventral striatum, orbitofrontal and anterior cingulate cortex, hippocampus, and amygdala (Lawrence et al. 1998), seemed to be particularly affected. These reductions in regional functional activity may also potentially lead to drug-taking in order to restore these brain areas to the functional state that was present before the drug-taking was initiated (Koob and Le Moal, 1997). In additions to reductions in areas involved in reward learning and motivational behaviors, there were also reductions in regions involved in learning and memory. Reductions in functional activity were observed in the hippocampus, medial thalamus, and basolateral amygdala. Reduced activity in these regions has important implications for normal functioning and the learning capacity at baseline after the cessation of drug consumption. Even more important for cocaine abusers is the role that learning plays in cue-reinforcement pairings during drug abuse. It is well established that cue conditioning
129 108 plays a role in the effects of drugs and on relapse, where cues alone are sufficient to reinstate drug taking/seeking after periods of prolonged abstinence (Lu et al., 2004; Shaham et al., 2003; Schmidt and Pierce, 2010). The basolateral amygdala has also been shown to be a major modulator of the extinction of conditioned place preference, further suggesting that the reductions in functional activity, and perhaps learning, may lead to a decreased ability to replace associations between drugs and cues (Schroeder & Packard, 2003, 2004). In addition to extinction of conditioned preference, the basolateral amygdala has been shown to be integral to the neural circuits involved in fear conditioning (Berlau & McGaugh, 2006; LaLumiere et al., 2003; Sacchetti et al, 1999; Vazdarjanova & McGaugh, 1999), thus reductions in the region have implications for associative learning in general, and not just reward based learning. Here we demonstrate that there are large reductions in the functional activity of the dorsal raphe and locus coeruleus following withdrawal from cocaine selfadministration. Our previous study, which investigated the effect of cocaine selfadministration while cocaine was still on board, found no differences in the locus coeruleus and actually found higher levels of metabolism in the dorsal raphe (Macey et al., 2004). Because these areas are cell body nuclei for catecholamine projections that are widespread throughout the brain, these data demonstrate that cocaine self-administration affects a broad expanse of the brain, certainly well beyond the DA system that is typically investigated. Our data of alterations of functional activity in the dorsal raphe are particularly intriguing, since the 5-HT system has been shown to play a role in locomotor activity. Specifically, 5-HT levels have been shown to be inversely related to vertical activity (Brookshire and Jones, 2009); thus, it is tempting to speculate that reduced
130 109 serotonergic activity (as indicated by the lower levels of functional activity in the dorsal raphe) may have had direct behavioral consequences (increased vertical activity at baseline). In addition, if the alterations in raphe activity that we see in rodents are also present in human users, these may account for some of the sleep disturbances that are often reported by addicts following cocaine abuse (Morgan & Malison, 2007; Schierenbeck et al., 2008). Also, dysfunction of both the dorsal raphe and locus coeruleus has been directly related to anxiety and depression during withdrawal (Carrasco and Van de Kar, 2003; Graeff et al., 1996; Itoi and Sugimoto, 2010). Further, the locus coeruleus system has been shown to mediate shifts in attention, thus in addition to stress and anxiety, these reductions could have effects on basic attention, which could in turn lead to additional learning and memory deficits (Aston-Jones et al., 1999; Rajkowski et al., 1994; Usher et al., 1999). These functional alterations could be due to the ability of cocaine to inhibit both the norepinephrine and serotonin transporters (Rothman and Baumann, 2003), thus, these changes during withdrawal may be compensatory effects due to the sustained elevated levels of these transmitters during cocaine self-administration. Taken together, these data demonstrate that the alterations that occur following cocaine self-administration are clearly not limited to just the DA system, but include other neurotransmitter systems as well, and suggest that cocaine has a widespread influence on the brain even when the drug is no longer present. In summary, the present study demonstrated that cocaine self-administration resulted in robust alterations in both functional and behavioral activity long after cocaine has been cleared. These data suggest that there are reductions in the functionality of a number of critical circuits involved in reward processing, memory, attention, sleep, and
131 110 stress processing. The reductions in these areas have important implications for individuals that abuse cocaine as these data indicate that even a short (five-day) selfadministration history can result in functional reductions in activity that are present up to 48 h later. These deficits were accompanied by behavioral changes as well, indicating that these metabolic changes are functionally relevant in the behaving animal. It is important to determine the functioning of neural networks after cocaine selfadministration, as it is possible that the reductions in some of these regions persist for longer periods of time and could facilitate the continued use of drugs in the face of negative consequences, and facilitate continued drug administration in the face of robust tolerance, as well as potentiate drug seeking after periods of prolonged abstinence. Acknowledgements: The authors wish to thank Mr. Mack Miller for his assistance conducting the 2-DG experiments. This work was funded by NIH grants R01 DA09085 (LJP), P50 DA06634, R01 DA (SRJ), R01 DA03016 (SRJ), T32 DA and F31 DA (ESC). The authors have no conflicts to declare. Abbreviations: LCGU, local cerebral glucose utilization; 2-DG, [ 14 C]-2-deoxyglucose; ACC, anterior cingulate cortex; NAc, nucleus accumbens; DA, dopamine; IP, intraperitoneal
132 111 References: Ahmed SH, Koob GF. (1998) Transition from moderate to excessive drug intake: change in hedonic set point. Science, 282(5387): Ahmed SH, Koob GF. (1999) Long-lasting increase in the set point for cocaine selfadministration after escalation in rats. Psychopharmacology (Berl)., 146(3): Alexander GE, DeLong MR, Strick PL. (1986) Parallel organization of functionally segregated circuits linking basal ganglia and cortex. Annu Rev Neurosci., 9: Aston-Jones G, Rajkowski J, Cohen J. (1999) Role of locus coeruleus in attention and behavioral flexibility. Biol Psychiatry, 46(9): Berlau DJ, McGaugh JL. (2006) Enhancement of extinction memory consolidation: the role of the noradrenergic and GABAergic systems within the basolateral amygdala. Neurobiol Learn Mem,. 86(2): Brookshire BR, Jones SR. (2009) Direct and indirect 5-HT receptor agonists produce gender-specific effects on locomotor and vertical activities in C57 BL/6J mice. Pharmacol Biochem Behav., 94(1): Calipari ES, Ferris MJ, Melchior JR, Bermejo K, Salahpour A, Roberts DC, Jones SR. (2012) Methylphenidate and cocaine self-administration produce distinct dopamine terminal alterations. Addict Biol. In press. Calipari ES, Ferris MJ, Zimmer BA, Roberts DCS, Jones SR. (2013) Temporal pattern of cocaine intake determines tolerance versus sensitization of cocaine effects at the dopamine transporter. Neuropsychopharm. In press. Carelli RM. (2002) The nucleus accumbens and reward: neurophysiological investigations in behaving animals. Behav Cogn Neurosci Rev., 1(4): Carrasco GA, Van de Kar LD. (2003) Neuroendocrine pharmacology of stress. Eur J Pharmacol., 463(1-3): Crane AM, Porrino LJ. (1989) Adaptation of the quantitative 2-[14C]deoxyglucose method for use in freely moving rats. Brain Res. 499(1): Dong Y, Green T, Saal D, Marie H, Neve R, Nestler EJ, Malenka RC. (2006) CREB modulates excitability of nucleus accumbens neurons. Nat Neurosci., 9(4): Ferris MJ, Calipari ES, Mateo Y, Melchior JR, Roberts DC, Jones SR. (2012) Cocaine self-administration produces pharmacodynamic tolerance: differential effects on the potency of dopamine transporter blockers, releasers, and methylphenidate. Neuropsychopharmacology, 37(7):
133 112 Ferris MJ, Calipari ES, Melchior JR, Roberts DCS, Espana RA, Jones SR. (2013) Paradoxical tolerance to cocaine after initial supersensitivity in drug-use-prone animals. Eur J Neurosci. In press. Ferris MJ, Mateo Y, Roberts DC, Jones SR. (2011) Cocaine-insensitive dopamine transporters with intact substrate transport produced by self-administration. Biol Psychiatry., 69(3): Gauvin DV, Briscoe RJ, Baird TJ, Vallett M, Carl KL, Holloway FA. (1997). Physiological and subjective effects of acute cocaine withdrawal (crash) in rats. Pharmacology Biochemistry and Behavior, 57: Graeff FG, Guimarães FS, De Andrade TG, Deakin JF. (1996) Role of 5-HT in stress, anxiety, and depression. Pharmacol Biochem Behav., 54(1): Haber SN, Calzavara R. (2009) The cortico-basal ganglia integrative network: the role of the thalamus. Brain Res Bull. 78(2-3): Hemby SE, Co C, Koves TR, Smith JE, Dworkin SI. (1997) Differences in extracellular dopamine concentrations in the nucleus accumbens during response-dependent and response-independent cocaine administration in the rat. Psychopharmacology (Berl)., 133(1):7-16. Hemby SE, Jones GH, Hubert GW, Neill DB, Justice JB. (1994) Assessment of the relative contribution of peripheral and central components in cocaine place conditioning. Pharmacol Biochem Behav., 47(4): Hooks MS, Duffy P, Striplin C, Kalivas PW. (1994) Behavioral and neurochemical sensitization following cocaine self-administration. Psychopharmacology (Berl). 115(1-2): Ishikawa M, Mu P, Moyer JT, Wolf JA, Quock RM, Davies NM, Hu XT, Schlüter OM, Dong Y. (2009) Homeostatic synapse-driven membrane plasticity in nucleus accumbens neurons. J Neurosci., 29(18): Itoi K, Sugimoto N. (2010) The brainstem noradrenergic systems in stress, anxiety and depression. J Neuroendocrinol., 22(5): Jones S, Bonci A. (2005) Synaptic plasticity and drug addiction. Curr Opin Pharmacol., 5(1):20-5. Koeltzow TE, White FJ. (2003) Behavioral depression during cocaine withdrawal is associated with decreased spontaneous activity of ventral tegmental area dopamine neurons. Behav Neurosci. 117(4): Koob GF, Le Moal M. (1997) Drug abuse: hedonic homeostatic dysregulation. Science, 278:52-8.
134 113 Koob GF. (2009) Neurobiological substrates for the dark side of compulsivity in addiction. Neuropharmacology, 1: Kornetsky C, Huston-Lyons D, Porrino LJ. (1991) The role of the olfactory tubercle in the effects of cocaine, morphine and brain-stimulation reward. Brain Res., 541(1): Kourrich S, Thomas MJ. (2009) Similar neurons, opposite adaptations: psychostimulant experience differentially alters firing properties in accumbens core versus shell. J Neurosci., 29(39): Lack CM, Jones SR, Roberts DC. (2008) Increased breakpoints on a progressive ratio schedule reinforced by IV cocaine are associated with reduced locomotor activation and reduced dopamine efflux in nucleus accumbens shell in rats. Psychopharmacology (Berl)., 195(4): LaLumiere RT, Buen TV, McGaugh JL. (2003) Post-training intra-basolateral amygdala infusions of norepinephrine enhance consolidation of memory for contextual fear conditioning. J Neurosci., 23(17): Lawrence AD, Sahakian BJ, Robbins TW. (1998) Cognitive functions and corticostriatal circuits: insights from Huntington's disease. Trends Cogn Sci., 2(10): Lecca D, Cacciapaglia F, Valentini V, Acquas E, Di Chiara G. (2007) Differential neurochemical and behavioral adaptation to cocaine after response contingent and noncontingent exposure in the rat. Psychopharmacology (Berl),. 191: Liu Y, Roberts DCS, Morgan D. (2005) Sensitization of the reinforcing effects of selfadministered cocaine in rats: effects of dose and intravenous injection speed. Eur J Neurosci., 22: Lu L, Grimm JW, Hope BT, Shaham Y. (2004) Incubation of cocaine craving after withdrawal: a review of preclinical data. Neuropharmacology, 1: Macey DJ, Rice WN, Freedland CS, Whitlow CT, Porrino LJ. (2004) Patterns of functional activity associated with cocaine self-administration in the rat change over time. Psychopharmacology (Berl)., 172(4): Maisonneuve IM, Ho A, Kreek MJ. (1995) Chronic administration of a cocaine "binge" alters basal extracellular levels in male rats: an in vivo microdialysis study. J Pharmacol Exp Ther. 272(2): Markou A, Kenny PJ. (2002) Neuroadaptations to chronic exposure to drugs of abuse: relevance to depressive symptomatology seen across psychiatric diagnostic categories. Neurotox Res., 4(4):
135 114 Markou A, Koob GF. (1992) Bromocriptine reverses the elevation in intracranial selfstimulation thresholds observed in a rat model of cocaine withdrawal. Neuropsychopharmacology, 7(3): Mateo Y, Lack CM, Morgan D, Roberts DC, Jones SR. (2005) Reduced dopamine terminal function and insensitivity to cocaine following cocaine binge self-administration and deprivation. Neuropsychopharmacology, 30(8): McFarland NR, Haber SN. (2002) Thalamic relay nuclei of the basal ganglia form both reciprocal and nonreciprocal cortical connections, linking multiple frontal cortical areas. J Neurosci. 22(18): Meil WM, Roll JM, Grimm JW, Lynch AM, See RE. (1994) Tolerance-like attenuation to contingent and noncontingent cocaine-induced elevation of extracellular dopamine in the ventral striatum following 7 days of withdrawal from chronic treatment. Psychopharmacology (Berl). 118(3): Morgan PT, Malison RT. (2007) Cocaine and sleep: early abstinence. Scientific World Journal, 7: Mu P, Moyer JT, Ishikawa M, Zhang Y, Panksepp J, Sorg BA, Schlüter OM, Dong Y. (2010) Exposure to cocaine dynamically regulates the intrinsic membrane excitability of nucleus accumbens neurons. J Neurosci., 30(10): Paxinos G, Watson C (1998) The rat brain in stereotaxic coordinates, 4 th edn. Academic Press, New York. Pierce RC, Kumaresan V. (2006) The mesolimbic dopamine system: the final common pathway for the reinforcing effect of drugs of abuse? Neurosci Biobehav Rev., 30(2): Porrino LJ. (1993) Functional consequences of acute cocaine treatment depend on route of administration. Psychopharmacology (Berl)., 112(2-3): Rajkowski J, Kubiak P, Aston-Jones G. (1994) Locus coeruleus activity in monkey: phasic and tonic changes are associated with altered vigilance. Brain Res Bull., 35(5-6): Rothman RB, Baumann MH. (2003) Monoamine transporters and psychostimulant drugs. Eur J Pharmacol. 479(1-3): Sacchetti B, Lorenzini CA, Baldi E, Tassoni G, Bucherelli C. (1999) Auditory thalamus, dorsal hippocampus, basolateral amygdala, and perirhinal cortex role in the consolidation of conditioned freezing to context and to acoustic conditioned stimulus in the rat. J Neurosci., 19(21):
136 115 Schierenbeck T, Riemann D, Berger M, Hornyak M.(2008) Effect of illicit recreational drugs upon sleep: cocaine, ecstasy and marijuana.sleep Med Rev., 12(5): Schmidt HD, Pierce RC. (2010) Cocaine-induced neuroadaptations in glutamate transmission: potential therapeutic targets for craving and addiction. Ann N Y Acad Sci., 1187: Schroeder JP, Packard MG. (2003) Systemic or intra-amygdala injections of glucose facilitate memory consolidation for extinction of drug-induced conditioned reward. Eur J Neurosci., 17(7): Schroeder JP, Packard MG. (2004) Facilitation of memory for extinction of drug-induced conditioned reward: role of amygdala and acetylcholine. Learn Mem., 11(5): Schultz W. (2010) Subjective neuronal coding of reward: temporal value discounting and risk. Eur J Neurosci., 31(12): Shaham Y, Shalev U, Lu L, De Wit H, Stewart J. (2003) The reinstatement model of drug relapse: history, methodology and major findings. Psychopharmacology (Berl)., 168:3-20. Sokoloff L, Reivich M, Kennedy C, Des Rosiers MH, Patlak CS, Pettigrew KD, Sakurada O, Shinohara M. (1977) The [14C]deoxyglucose method for the measurement of local cerebral glucose utilization: theory, procedure, and normal values in the conscious and anesthetized albino rat. J Neurochem., 28(5): Stoker AK, Markou A. (2011) Withdrawal from chronic cocaine administration induces deficits in brain reward function in C57BL/6J mice. Behav Brain Res., 223(1): Usher M, Cohen JD, Servan-Schreiber D, Rajkowski J, Aston-Jones G. (1999) The role of locus coeruleus in the regulation of cognitive performance. Science, 283(5401): Vanderschuren LJ, Pierce RC. (2010) Sensitization processes in drug addiction. Curr Top Behav Neurosci., 3: Vazdarjanova A, McGaugh JL. (1999) Basolateral amygdala is involved in modulating consolidation of memory for classical fear conditioning. J Neurosci., 19(15): Verheij MM, Cools AR. (2008) Twenty years of dopamine research: individual differences in the response of accumbal dopamine to environmental and pharmacological challenges. Eur J Pharmacol., 585(2-3): Verheij MMM, de Mulder ELW, De Leonibus E, van Loo KMJ, Cools AR (2008) Rats that differentially respond to cocaine differ in their dopaminergic storage capacity of the nucleus accumbens. J. Neurochem., 105:
137 116 Volkow ND, Fowler JS, Wang GJ, Hitzemann R, Logan J, Schlyer DJ, Dewey SL, Wolf AP. (1993) Decreased dopamine D2 receptor availability is associated with reduced frontal metabolism in cocaine abusers. Synapse. 14(2): Volkow ND, Hitzemann R, Wang GJ, Fowler JS, Wolf AP, Dewey SL, Handlesman L. (1992) Long-term frontal brain metabolic changes in cocaine abusers. Synapse. 11(3): Weiss F, Markou A, Lorang MT, Koob GF. (1992) Basal extracellular dopamine levels in the nucleus accumbens are decreased during cocaine withdrawal after unlimited-access self-administration. Brain Res. 593(2): White SR, Harris GC, Imel KM, Wheaton MJ. (1995) Inhibitory effects of dopamine and methylenedioxymethamphetamine (MDMA) on glutamate-evoked firing of nucleus accumbens and caudate/putamen cells are enhanced following cocaine selfadministration. Brain Res., 681(1-2): Zocchi A, Conti G, Orzi F. (2001) Differential effects of cocaine on local cerebral glucose utilization in the mouse and in the rat. Neurosci Lett., 306(3): Dackis CA, O'Brien CP. (2001) Cocaine dependence: a disease of the brain's reward centers. J Subst Abuse Treat. 21(3):111-7.
138 117 CHAPTER IV TEMPORAL PATTERN OF COCAINE INTAKE DETERMINES TOLERANCE VS SENSITIZATION AT THE DOPAMINE TRANSPORTER Erin S. Calipari, Mark J. Ferris, Benjamin A. Zimmer, David C. S. Roberts, and Sara R. Jones The following manuscript was published in Neuropsychopharmacology, in press, 2013, and is reprinted with permission. Stylistic variations are due to the requirements of the journal. Erin S. Calipari performed most self-administration and voltammetry experiments, data analysis, and prepared the manuscript. Dr. Mark J. Ferris performed select voltammetry experiments. Benjamin A. Zimmer generated a small number of selfadministration animals. Dr. Sara R. Jones acted in an advisory and editorial capacity. Dr. David C. S. Roberts acted in an editorial capacity.
139 118 Abstract: The dopamine transporter (DAT) is responsible for terminating dopamine (DA) signaling and is the primary site of cocaine s reinforcing actions. Cocaine self-administration has been shown previously to result in changes in cocaine potency at the DAT. To determine whether the DAT changes associated with self-administration are due to differences in intake levels or temporal patterns of cocaine-induced DAT inhibition, we manipulated cocaine access to produce either continuous or intermittent elevations in cocaine brain levels. Long-access (LgA, 6 hr) and short-access (ShA, 2 hr) continuous selfadministration produced similar temporal profiles of cocaine intake that were sustained throughout the session; however, LgA had greater intake. ShA and Intermittent-access (IntA, 6 hr) produced the same intake, but different temporal profiles, with spiking brain levels in IntA compared to constant levels in ShA. IntA consisted of 5 min access periods alternating with 25 min time-outs, which resulted in bursts of high responding followed by periods of no responding. DA release and uptake, as well as the potency of cocaine for DAT inhibition, were assessed by voltammetry in nucleus accumbens slices following control, IntA, ShA, and LgA self-administration. Continuous access protocols (LgA and ShA) did not change DA parameters, but the spiking protocol (IntA) increased both release and uptake of DA. In addition, high continuous intake (LgA) produced tolerance, while spiking (IntA) produced sensitization, relative to ShA and naïve controls. Thus, intake and pattern can both influence cocaine potency, and tolerance seems to be produced by high intake, while sensitization is produced by intermittent temporal patterns of intake.
140 119 Introduction A substantial literature has documented the effects of cocaine on the dopamine (DA) system and its involvement in the addiction process (Roberts et al., 1977; Izenwasser 2004; Stuber et al., 2011; Blum et al., 2012). Cocaine exerts its reinforcing and neurochemical effects by inhibiting the dopamine transporter (DAT), which leads to augmented DA signaling (Ritz et al., 1987; Kristensen et al., 2011). DA elevations in the nucleus accumbens (NAc) core region are critically involved in self-administration behaviors, reinstatement, cue-reward association, assignment of motivational value, and goal oriented behaviors (for review see Wheeler et al., 2005; Kalivas and McFarland, 2003; Aragona et al., 2009; Willuhn et al., 2010). Both tolerance and sensitization can develop to the neurochemical effects of cocaine; however, the nature of these changes appears to depend on the dosing parameters used. An extensive literature, primarily using daily experimenteradministered intraperitoneal (IP) injections has documented neurochemical sensitization associated with the mesolimbic DA system (Kalivas and Duffy, 1990, 1993; Parsons and Justice, 1993; Addy et al., 2010). Sensitization of cocaine-induced increases in extracellular DA has been linked not only to alterations in the regulation of DA release and reuptake (Jones et al., 1996, Addy et al. 2010), but also to influences such as glutamate (for review see Vanderschuren and Kalivas, 2000), GABA (for review see Steketee, 2005; Filip et al., 2006) and serotonin (Neumaier et al., 2002; Filip et al., 2010). By contrast, studies using self-administration procedures have generally shown tolerance to cocaine s DA-elevating and DAT-inhibiting effects of cocaine, as measured by microdialysis (Hurd et al., 1989; Mateo et al., 2005; Lack et al., 2008; Ferris et al., 2011).
141 120 It is possible that cocaine tolerance is due to a reduced ability of cocaine to inhibit the DAT, which has been reported in a number of paradigms where cocaine levels are high and sustained over daily extended-access self-administration sessions (Mateo et al., 2005; Ferris et al., 2011, 2012; Calipari et al., 2012). Therefore, experimenter-delivered and self-administration protocols produce distinctly different effects on the DA system. There are many differences between these protocols including contingency, temporal pattern of intake, total consumption, and route of administration, which may influence the divergent neurochemical consequences. For example, experimenter-administered injections result in an intermittent pattern of drug administration, whereas intravenous (IV) self-administration procedures result in high rates of responding and high levels of administration for many hours (i.e. for the duration of the self-administration session). In the present study, we investigated the contribution of 1) pattern of consumption and 2) total intake of cocaine on rapid DA signals in the NAc and the ability of selfadministered cocaine to either sensitize or induce tolerance to the pharmacological effects of cocaine on the DAT. Self-administration paradigms were used in order to control for the contribution of contingency and route of administration on the effects of the DA system. A long-access (LgA) procedure was used in which cocaine levels were sustained for 6 hours each day (Ahmed et al., 2003; Martin et al, 2011; Peraile et al., 2010). This was compared to an intermittent access (IntA) self-administration procedure in which cocaine was available during a 6 hr session but only during 5 min trials, which were separated by 25 min time-outs. This protocol allows animals to "load up" during each trial but ensures that cocaine levels cannot be maintained. The IntA procedure, which
142 121 results in a spiking pattern of intake, has recently been shown to produce a greater augmentation of the reinforcing effects of cocaine compared to the long access (LgA) procedure (Zimmer et al., 2012). We also included a short access (ShA) group in which cocaine levels were matched to the IntA group but were maintained during a two-hour session similar to LgA. Here we demonstrate that intake and pattern can both influence cocaine potency at the DAT; while tolerance seems to be dictated by total intake, sensitization appears to be determined by pattern. Methods and Materials: Animals: Male Sprague-Dawley rats ( g; Harlan Laboratories, Frederick, Maryland), maintained on a 12:12 hour reverse light/dark cycle (3:00 am lights off; 3:00 pm lights on) with food and water ad libitum. All animals were maintained according to the National Institutes of Health guidelines in Association for Assessment and Accreditation of Laboratory Animal Care accredited facilities. The experimental protocol was approved by the Institutional Animal Care and Use Committee at Wake Forest School of Medicine. Self-Administration: Rats were anesthetized and implanted with chronic indwelling jugular catheters as previously described (Liu et al., 2007). Animals were singly housed, and all sessions took place in the home cage during the active/dark cycle (9:00 am 3:00 pm). After a 2-day recovery period, animals underwent a training paradigm within which animals were given access on a fixed ratio one (FR1) schedule to a cocaine-paired lever, whereby a single lever press initiated an intravenous injection of cocaine (0.75mg/kg, infused over 4s). After each response/infusion, the lever was retracted and a
143 122 stimulus light was illuminated for a 20 second timeout period. Training sessions were terminated after a maximum of 20 infusions or 6h, whichever occurred first. Acquisition occurred when an animal responded for 20 injections for two consecutive days and a stable pattern of infusion intervals was present. Following training, animals were assigned to either ShA, IntA, or LgA groups. All self-administration was 14 consecutive sessions, after which animals were sacrificed and brains were prepared for voltammetric recordings. Control: Controls were naïve rats housed in the same conditions as animals undergoing self-administration. LgA Group: Subjects completed daily 6h sessions during which they had unlimited access to cocaine (0.75 mg/kg; infused over 4s) on an FR1 schedule for 14 consecutive days. Upon each infusion, the lever was retracted, and a stimulus light signaled a 20-s timeout period. ShA Group: Subjects were given access to cocaine (0.75 mg/kg; infused over 4s) on an FR1 schedule during 2-h daily sessions for 14 consecutive days. At the start of each infusion, a stimulus light signaled a 20-s timeout period during which the lever was retracted. IntA Group: Subjects were given access to cocaine on an intermittent schedule of administration described previously (Zimmer et al., 2012). Briefly here, during each 6h session animals had access to cocaine for 12 five minute trails separated by 25-minute timeout periods. Within each five-minute session, there were no timeouts other than
144 123 during each infusion, and the animal could press the lever on an FR1 schedule to receive a 1-sec infusion of cocaine (0.375 mg/kg/inf). In Vitro Voltammetry: Fast scan cyclic voltammetry (FSCV) was used to characterize presynaptic DA system kinetics and the ability of psychostimulants to inhibit DA uptake in the NAc. Voltammetry experiments were conducted during the dark phase of the light/dark cycle 18 hours after commencement of the final self-administration session. A vibrating tissue slicer was used to prepare 400 µm thick coronal brain sections containing the NAc. The tissue was immersed in oxygenated artificial cerebrospinal fluid (acsf) containing (in mm): NaCl (126), KCl (2.5), NaH 2 PO 4 (1.2), CaCl 2 (2.4), MgCl 2 (1.2), NaHCO 3 (25), glucose (11), L-ascorbic acid (0.4) and ph was adjusted to 7.4. Once sliced, the tissue was transferred to the testing chambers containing bath acsf (32 C), which flowed at 1 ml/min. A carbon fiber microelectrode ( μm length, 7 μm radius) and bipolar stimulating electrode were placed into the core of the NAc, which was selected because of its role in the reinforcing and rewarding actions of cocaine. DA release was evoked by a single electrical pulse (300 μa, 4 msec, monophasic) applied to the tissue every 5 minutes. Extracellular DA was recorded by applying a triangular waveform ( 0.4 to +1.2 to 0.4V vs Ag/AgCl, 400 V/s). Once the extracellular DA response was stable cocaine ( μmol/l) was applied cumulatively to the brain slice. Data Analysis: Demon Voltammetry and Analysis Software was used for all analyses of FSCV data (Yorgason et al., 2011). To evaluate DA kinetics and drug potency, evoked levels of DA were modeled using Michaelis-Menten kinetics. Apparent Km (app. K m ) was used as a measure of the ability of cocaine to inhibit DA clearance, and to evaluate changes in cocaine potency. As app. K m increases, the affinity of DA for the DAT
145 124 decreases. Increasing concentrations of cocaine linearly decrease the affinity of DA for the DAT, such that shifts in app. K m across treatment groups indicate shifts in the ability of cocaine to inhibit DA uptake. Recording electrodes were calibrated by recording responses (in electrical current; na) to a known concentration of DA (3 μm) using a flow-injection system. Calculating K i Values: Inhibition constants (K i ) were determined by plotting the linear concentration-effect profiles and determining the slope of the linear regression. The K i was calculated by the equation K m /slope. K i values are reported in µm and are a measure of the drug concentration that is necessary to produce 50% uptake inhibition. K i is a measure of the drug affinity for the DAT, reported in concentration of drug, while app. K m is a DA related parameter that estimates DA uptake inhibition. Statistics: Graph Pad Prism (version 5, La Jolla, CA, USA) was used to statistically analyze data and create graphs. Baseline voltammetry data and K m values were compared using a one-way analysis of variance (ANOVA). When main effects were obtained (P < 0.05), differences between groups were tested using Tukey post hoc tests. Release data and data obtained after perfusion of cocaine were subjected to a two-way ANOVA with experimental group and concentration of drug as the factors. Differences between groups were tested using a Bonferroni post hoc test. Given the lack of a Gaussian distribution for K i values, non-parametric tests (Kruskal-Wallis and Neuman Keuls) were used to compare inhibition constants.
146 125 Results: ShA and LgA result in similar pattern of self-administration that is different from IntA (Fig 1A). Figure 1 A contains representative behavioral plots demonstrating the different patterns of self-administration behavior between ShA, LgA, and IntA. We demonstrated that ShA results in high and consistent rates of responding for cocaine over a 2hr self-administration session (Fig 1A, top panel). IntA results in an intermittent pattern of responding for cocaine, characterized by high rates of responding followed by time-out periods within which no responding occurs (Fig 1A, middle panel). LgA results in high and consistent rates of responding, similar to responding for ShA, for cocaine over a 6hr self-administration session (Fig 1A, bottom panel). LgA results in greater cocaine intake as compared to ShA and IntA (Fig 1B). Oneway ANOVA revealed a main effect of intake between groups (F 2, 11 = 39.28, p < ). Tukey post hoc analysis indicated that there was a significant increase in intake for the LgA group versus both the ShA group (p < ) and IntA group (p < ). Further, although the pattern of self-administration differed, the total cocaine intake between the ShA and IntA groups did not significantly differ. Because ShA results in similar intake to the IntA group and similar pattern of estimated cocaine brain levels to the LgA group, it was used as a comparator group to determine if effects were due to total intake or temporal pattern of cocaine self-administration. IntA, but not LgA or ShA, increases stimulated DA release and V max. FSCV was used to assess presynaptic baseline DA system kinetics. ANOVA indicated a main effect of
147 126
148 127 Figure 1. Intermittent (IntA), long-access (LgA), and short access (ShA) cocaine selfadministration result in different patterns of self-administration. (A) Plots demonstrating representative self-administration behavior throughout each session for individual rats self-administering cocaine. Tick marks represent infusions/responses on the lever earned on a fixed ratio one schedule of reinforcement. (A, Top) ShA results in high, consistent rates of responding over the two hour session. (A, Middle) IntA is achieved by giving 5 minutes access followed by 25 minute forced time-outs. This results in bursts of high responding followed by no responding. The spacing between clusters of tick marks is due to a forced time-out period. (A, Bottom) LgA results in high consistent rates of responding over the six hour session. (B) Group data plotting total intake over the 14 days of self-administration for each paradigm. ***, p <0.001 vs ShA; ###, p <0.001 vs IntA.
 Representative traces from control (left), IntA (center, left), LgA (center, right), and ShA (right) animals. Traces are represented as concentration (µm) in dopamine (DA) over time.")
149 128 Figure 2. Differential effects of intermittent (IntA), short-access (ShA) and longaccess (LgA) self-administration on presynaptic dopamine system kinetics. (A) Representative traces from control (left), IntA (center, left), LgA (center, right), and ShA (right) animals. Traces are represented as concentration (µm) in dopamine (DA) over time. (B) Stimulated DA release in µm across LgA, IntA, ShA, and control groups. (C) Maximal rate of DA uptake (V max ) across groups. (D) Group data indicating that the Km of DA for the dopamine transporter is unchanged following all self-administration paradigms. **, p < 0.01 vs control; ***, p < vs control; &&&, p < vs LgA.
150 129 paradigm on stimulated DA release (F 3, 63 = 4.020, p < 0.05; Fig 2A, B) and V max (F 3, 63 = 8.397, p < ; Fig 2 A, C). Tukey post hoc analysis revealed that stimulated release (p < 0.05; Fig 2 B) and V max (p < 0.05; Fig 2 C) were both elevated in the IntA group as compared to controls. In addition, V max was also elevated as compared to LgA (p < 0.001; Fig 2 B). Both LgA and ShA resulted in no significant change in either DA release or V max as compared to control animals. There were no changes in any groups in regards to the affinity of DA for the DAT (K m ; Fig 2D). This indicates that pattern of cocaine interaction with the DAT has differential effects on the compensatory mechanisms associated with baseline functioning. Differences in intake (LgA versus ShA) results in decreased cocaine potency, while differences in temporal pattern of cocaine intake (IntA versus ShA) results in increased cocaine potency at the DAT. FSCV in brain slices was used to determine cocaine potency over a concentration-response curve. ANOVA revealed a main effect of paradigm on cocaine potency (F 3,68 = 3.21, p < 0.01; Fig 3). ShA resulted in no changes in cocaine potency as compared to controls (Fig 3A, C, black vs. green lines). When temporal pattern of cocaine self-administration was kept the same, but there were differences in intake, cocaine potency was reduced (ShA vs LgA). This was expressed as a significant reduction in cocaine potency as measured by apparent K m at the 30 µm concentration in LgA (p <0.001) versus ShA (Fig 3C, Green vs. Red curve). Conversely, when intake was kept the same, but there were differences in temporal pattern of selfadministration, cocaine potency was increased (IntA vs ShA). This was expressed as an increase in cocaine s ability to inhibit the DAT in the IntA group (p < ) as
151 130
152 131 Figure 3. Intermittent access (IntA) self-administration results in sensitization to the neurochemical effects of cocaine while long-access (LgA) results in tolerance. (A) Representative traces highlighting the uptake inhibition induced by 10 µm cocaine in control (Black) and short-access (ShA, Green) animals. Traces are represented as concentration (µm) in dopamine (DA) over time and smaller signals were shifted to the right in order to match the peak-height of the small signal to equivalent concentration of the larger signal. This allows for direct comparison of dopamine uptake in signals with different peak-heights. (B) Representative traces highlighting the uptake inhibition induced by 10 µm cocaine in control (Black), IntA (blue) and LgA (Red) animals. (B) Cumulative cocaine ( µm) dose response curves in slices containing the nucleus accumbens core. Cocaine potency is decreased following LgA, unchanged following short-access (ShA), and increased following IntA. (D) Group data of K i values for cocaine in control, LgA, ShA, and IntA groups. K i values are a measure of the concentration of drug at which 50% inhibition is achieved. *, p < 0.5 vs control **, p < 0.01 vs control; # p < 0.01 vs ShA.
153 132 compared to ShA (Fig 3C, Green vs. Blue curve). A similar trend was present when groups were compared to control animals where LgA resulted in decreased cocaine potency at the 30 µm concentration (p < 0.01), and IntA resulted in enhanced potency for cocaine at the same concentration (p < 0.01) (Fig 3C, Red and Blue curves vs. Black curve). K i is a measure of the drug concentration that results in 50% uptake inhibition. This measurement can determine relative potencies between drugs at baseline as well as potency changes following drug self-administration. Kruskal-Wallis non-parametric analysis revealed a significant main effect of paradigm on K i (Chi 2 4=10.94, p < 0.05 = 10.53; Fig 3 D; Supplementary Fig 1). Mann Whitney analysis revealed that K i was increased in LgA animals (p < 0.05) and reduced following IntA (p < 0.05) when compared to ShA. Further, K i was increased in LgA animals (p < 0.05) and reduced following IntA (p < 0.05) when compared to control animals. Cocaine-induced increases in peak-height was assessed across all three groups to determine if this aspect of cocaine effects on DA was changed by self-administration. There were no significant differences between any of the groups tested (Fig 4). Discussion The present results demonstrate that the compensatory changes in DAT sensitivity to cocaine produced by self-administration are influenced by both the temporal pattern of cocaine consumption and the total amount of drug intake. When there was a sustained consumption pattern and intake was high (LgA), tolerance to cocaine s effects at the
154 133 Supplementary Figure 1. Intermittent access (IntA) self-administration results increased potency, while long-access (LgA) results in decreased potency versus short-access (ShA). Linear concentration-response curves for cocaine. The slope of the concentration-response curve for cocaine was significantly increased following IntA, and decreased following LgA, as compared to both control and ShA. K i values across groups were calculated based off of the linear dose-response curve for cocaine in each group. ***, p < vs control.
155 134 Figure 4. Cocaine-induced increases in DA did not differ across groups. Stimulated DA release was measured across a dose-response curve for cocaine, and expressed as a percent pre-cocaine stimulated DA release. There were no differences in cocaine-induced DA elevations between control, intermittent-access (IntA), long-access (LgA), or shortaccess (ShA) cocaine self-administration groups.
156 135 DAT developed. Conversely, when intake was low and patterns of consumption were intermittent (IntA) sensitization of the DAT to cocaine occurred. Differences were also observed in baseline (pre-drug) DA measures, wherein IntA cocaine administration resulted in increased V max and stimulated DA release, and LgA and ShA did not produce any changes. This is the first comparison of the effects ShA and LgA on cocaine potency directly at the DAT. Previous work showed increased measures of drug seeking, motivation to administer cocaine, and craving in LgA versus ShA animals (Koob 1996; Koob and LeMoal 1997; Orio et al., 2009; Patterson and Markou 2003); however, these measures are probably not due to the pharmacological effects of cocaine at the DAT, which we demonstrate are reduced. Further, we found that IntA cocaine selfadministration produced a sensitized cocaine response at the DAT, demonstrating that pattern of self-administration plays an integral role in the compensatory effects of the DAT to cocaine effects following extended use. Here we show that LgA results in reduced cocaine potency, indicative of a neurochemical tolerance to cocaine at the DAT, in comparison to both ShA and naïve control animals. The decreased cocaine potency following LgA is consistent with previous reports demonstrating similar changes following a number of different extended-access paradigms. In our hands and others, LgA results in neurobiological compensations characterized by decreased cocaine effects and blunted DA system function in the NAc (Hurd et al., 1989; Meil et al., 1995; Macey et al., 2004; Ferris et al., 2011, 2012; Calipari et al., 2012; but see Hooks et al., 1994; Zapata et al., 2003). Also, immediately following 21-day LgA cocaine self-administration while cocaine is still present, there are decreases in functional activity in the ventral and dorsal striatum, as
157 136 well as a number of other brain regions, indicating that cocaine is less efficacious at activating these circuits (Macey et al., 2004). Previously we examined a number of selfadministration protocols, manipulating both frequency and duration of access, and regardless time, animals given continuous cocaine access exhibited robust behavioral and neurochemical tolerance. Animals that administered cocaine for only 1 day of 40 injections and animals that underwent extended-access self-administration for five days (40 inj. per day) both exhibited the same effects as LgA self-administration (6 hrs/day, 14 days, Ferris et al., 2011, 2012; Calipari et al., 2012). All of the previously tested procedures shared the property that animals could administer large amounts of cocaine for the duration of the session. Regardless of duration of testing, each of these groups that maintained high rates of responding over 5-6 hr session showed neurochemical tolerance at the DAT when tested 24 hrs after the last self administration session (Ferris et al., 2011; 2012; Calipari et al., 2012). Although ShA animals have a similar pattern of self-administration, and can administer large amounts of drug, they can only do so for 2 hr. This results in significantly less drug consumption, leading to no change in cocaine potency. This suggests that total intake within sessions, and not pattern, is what is driving the tolerance to the pharmacological effects of cocaine at the DAT. In contrast, here we show that the IntA cocaine self-administration procedure enhances cocaine potency, indicative of a sensitized neurochemical response to cocaine, at the DAT relative to both ShA and naïve controls. The IntA results are opposite to the well-documented decrease in cocaine potency after LgA shown here and in previous research (Hurd et al., 1989; Ferris et al., 2011, 2012; Calipari et al., 2012). Although behavioral sensitization to cocaine has been reported previously (Schmidt and Pierce,
158 ), to our knowledge, this is the first demonstration of a sensitized cocaine response directly at the DAT following cocaine self-administration. It is possible that the increased potency may promote increased rewarding and reinforcing effects of cocaine as well as an increase in motivation to self-administer the drug, potentially leading to greater risk of compulsive or addictive-like cocaine intake. Indeed, Zimmer et al. (2012) showed that motivation to self-administer cocaine is significantly increased following IntA as compared to LgA and ShA cocaine self-administration. The marked differences between ShA and IntA cannot be attributed to intake, as the animals administer similar amounts of drug over sessions. In addition, the session length was different between IntA (360 min) and ShA (120 min); however, we do not think that effects are due to session length differences, because LgA and IntA both had 360 min sessions. These groups exhibited opposite effects on DAT pharmacology, therefore it seems that session time is not a contributing factor to the neurochemical effects. The increased effects of cocaine at the DAT seem to be due to the intermittent intake pattern of cocaine that likely leads to intermittent cocaine-dat interactions. The LgA and IntA groups differ substantially in the total cocaine self-administered over the course of the current experiment; however, we would predict that LgA would still produce tolerance when total intake is limited to equal the IntA condition. In fact, we have shown that a single day of LgA (60 mg total), which approximates the IntA group s total intake of cocaine (70 mg total), produced robust tolerance (Ferris et al., 2011). Thus, the sensitized response was only seen in the group with intermittent cocaine access (IntA) and not in the matched intake groups that experienced sustained drug levels examined here (i.e., ShA) and in previous work (i.e. LgA, Ferris et al., 2011).
159 138 Cocaine has been demonstrated to result in changes in DA release that are dependent on vesicular exocytosis (Venton et al., 2006), therefore we wanted to confirm that changes in this process were not driving the observed changes in cocaine effects. DA release over the concentration-response curve for the compound was not changed in any of the groups, suggesting that it was cocaine-induced DAT inhibition, not increased/decreased DA release, causing the change in potency following IntA/LgA. It is possible that allosteric modifications to the DAT protein itself are mediating the decreased potency following LgA and the increased potency following IntA. Indeed, the DAT has been shown to undergo glycosylation and phosphorylation (Foster et al, 2008; Chen et al, 2009). Therefore, it is possible that changes in the interaction of the DAT protein with downstream effectors responsible for these post-translational modifications are altered following self-administration in a way that changes the ability of ligands to bind to the DAT and inhibit the clearance of DA. Additionally, the DAT has been shown to form oligomer complexes (Hastrup et al., 2003) and be modulated by receptors such as the D2 autoreceptor (Chen et al., 2013), which are two additional mechanisms that could play a role in the observed changes in cocaine potency. There appear to be many neurochemical adaptations that occur following cocaine exposure, and how they interact to influence cocaine-induced DA overflow and behavior is as yet unclear. In addition to the sensitized effect at the DAT reported here, other systems appear to play a role in DA system sensitization, including glutamate (for review see Vanderschuren and Kalivas, 2000), GABA (for review see Steketee, 2005; Filip et al., 2006) and serotonin (Neumaier et al., 2002; Filip et al., 2010). Due to the convergence of these systems and factors, which lead to the expression of behavioral sensitization, it
160 139 remains to be determined whether increased DA overflow and behavioral effects, such as locomotor sensitization, would be seen after IntA self-administration. Further, demonstrations of sensitized DA overflow are more robust after 1-2 weeks of drug abstinence (Kalivas and Duffy, 1990, 1993) thus it is possible that the changes observed here could be more robust following acute and prolonged withdrawal. Here we demonstrate that whether tolerance or sensitization develops to specific effects of cocaine appears to depend on access conditions and the intake or pattern of administration, respectively. Pattern, total dose, and abstinence periods need to be taken into account when modeling the behavioral and neurochemical processes involved in addiction. Cocaine sensitization has been difficult to demonstrate in humans, suggesting that either sensitization does not occur in humans, or that it occurs during a part of the addiction process that has not been studied extensively (Leyton & Vezina, 2013). It is possible that early in the drug abuse process administration of cocaine is intermittent, leading to a sensitized cocaine response that facilitates continued drug use, while after long-term use there is tolerance to acute drug effects, an effect that has been reported in human subjects (Mendelson et al, 1998; Reed et al, 2009). It is suggested that in humans, drug use may be based on limited availability, which may lead individuals to administer drug in an intermittent pattern (Ahmed et al., 2013). However, it has also been suggested that LgA self-administration is a model of the escalation that is reported with long-term abuse of the compound in humans (Dackis and O Brien, 2001). Here we demonstrate tolerance to the neurochemical effects of cocaine following LgA, suggesting that LgA may result in neurochemical changes consistent with human studies. In our previous work and currently accepted cocaine self-administration models, much emphasis has
161 140 been placed on maximizing an animal s intake, with the thought that more intake results in greater neurochemical effects. Here we demonstrate that greater intake is not necessary to produce robust neurochemical effects, and in fact, continuous high intake early in the animal s self-administration history may produce marked tolerance to the drug. Thus, we highlight the importance of mimicking human patterns of administration in rodent models, as compensatory mechanisms associated with drug exposure are not only dependent on the drug and total intake, but also the pattern in which the drug is taken. Acknowledgements: This work was funded by NIH grants R01 DA024095, R01 DA03016 (SRJ), R01 DA14030 (DCSR), P50DA (SRJ and DCSR), T32 DA and F31 DA (ESC), K99 DA (MJF).
162 141 References: Addy NA, Daberkow DP, Ford JN, Garris PA, Wightman RM (2010) Sensitization of rapid dopamine signaling in the nucleus accumbens core and shell after repeated cocaine in rats. J Neurophysiol. 104(2): Ahmed SH, Lenoir M, Guillem K (2013) Neurobiology of addiction versus drug use driven by lack of choice. Curr Opin Neurobiol. 23: 1-7. Ahmed SH, Lin D, Koob GF, Parsons LH (2003). Escalation of cocaine selfadministration does not depend on altered cocaine-induced nucleus accumbens dopamine levels. J Neurochem 86(1): Aragona BJ, Day JJ, Roitman MF, Cleaveland NA, Wightman RM, Carelli RM (2009) Regional specificity in the real-time development of phasic dopamine transmission patterns during acquisition of a cue-cocaine association in rats. Eur J Neurosci 30(10): Blum K, Chen AL, Giordano J, Borsten J, Chen TJ, Hauser M, Simpatico T, Femino J, Braverman ER, Barh D (2012) The addictive brain: all roads lead to dopamine. J Psychoactive Drugs. 44(2): Calipari ES, Ferris MJ, Melchior JR, Bermejo K, Salahpour A, Roberts DC, Jones SR. Methylphenidate and cocaine self-administration produce distinct dopamine terminal alterations. Addict Biol. In Press. Chen R, Daining CP, Sun H, Fraser R, Stokes SL, Leitges M, Gnegy ME (2013) Protein kinase Cβ is a modulator of the dopamine D2 autoreceptor-activated trafficking of the dopamine transporter. J Neurochem. In Press. Chen R, Furman CA, Zhang MJ, Kim MN, Gereau RW, Leitges M et al (2009). Protein kinase C beta is a critical regulator of dopamine transporter trafficking and regulates the behavioral response to amphetamine in mice. J Pharmacol Exp Therap 328: Dackis CA, O'Brien CP (2001) Cocaine dependence: a disease of the brain's reward centers. J Subst Abuse Treat. 21(3): Ferris MJ, Calipari ES, Mateo Y, Melchior JR, Roberts DC, Jones SR (2012). Cocaine self-administration produces pharmacodynamic tolerance: differential effects on the potency of dopamine transporter blockers, releasers, and methylphenidate. Neuropsychopharm 37(7): Ferris MJ, Mateo Y, Roberts DC, Jones SR (2011). Cocaine-insensitive dopamine transporters with intact substrate transport produced by self-administration. Biol Psychiatry 69(3):201-7.
163 142 Filip M, Alenina N, Bader M, Przegaliński E (2010) Behavioral evidence for the significance of serotoninergic (5-HT) receptors in cocaine addiction. Addict Biol. 15(3): Filip M, Frankowska M, Gołda A, Zaniewska M, Vetulani J, Przegaliński E (2006) Various GABA-mimetic drugs differently affect cocaine-evoked hyperlocomotion and sensitization. Eur J Pharmacol. 541(3): Foster JD, Adkins SD, Lever JR, Vaughan RA (2008). Phorbol ester induced traffickingindependent regulation and enhanced phosphorylation of the dopamine transporter associated with membrane rafts and cholesterol. J Neurochem 105: Hastrup H, Sen N, Javitch JA (2003) The human dopamine transporter forms a tetramer in the plasma membrane: cross-linking of a cysteine in the fourth transmembrane segment is sensitive to cocaine analogs. J Biol Chem. 278(46): Hooks MS, Duffy P, Striplin C, Kalivas PW (1994). Behavioral and neurochemical sensitization following cocaine self-administration. Psychopharmacology (Berl) 115(1-2): Hurd YL, Weiss F, Koob GF, And NE, Ungerstedt U (1989). Cocaine reinforcement and extracellular dopamine overflow in rat nucleus accumbens an in vivo Microdialysis Study. Brain Res 498: Izenwasser S (2004). The role of the dopamine transporter in cocaine abuse. Neurotox Res 6(5): Review. Jones SR, Lee TH, Wightman RM, Ellinwood EH (1996). Effects of intermittent and continuous cocaine administration on dopamine release and uptake regulation in the striatum: in vitro voltammetric assessment. Psychopharmacology (Berl). 126(4): Kalivas PW, Duffy P (1990) Effect of acute and daily cocaine treatment on extracellular dopamine in the nucleus accumbens. Synapse. 5(1): Kalivas PW, Duffy P (1993) Time course of extracellular dopamine and behavioral sensitization to cocaine. I. Dopamine axon terminals. J Neurosci. 13(1): Kalivas PW, McFarland K (2003) Brain circuitry and the reinstatement of cocaineseeking behavior. Psychopharmacology (Berl). 168(1-2): Koob GF (1996). Hedonic valence, dopamine and motivation. Mol Psychiatry 1(3): Koob GF, Le Moal M (1997). Drug abuse: hedonic homeostatic dysregulation. Science 278(5335):52-8.
164 143 Kristensen AS, Andersen J, Jørgensen TN, Sørensen L, Eriksen J, Loland CJ, et al. (2011). SLC6 neurotransmitter transporters: structure, function, and regulation. Pharmacol Rev. 63(3): Review. Lack CM, Jones SR, Roberts DC (2008). Increased breakpoints on a progressive ratio schedule reinforced by IV cocaine are associated with reduced locomotor activation and reduced dopamine efflux in nucleus accumbens shell in rats. Psychopharmacology (Berl) 195(4): Leyton M, Vezina P (2013). Striatal ups and downs: Their roles in vulnerability to addictions in humans. Neurosci Biobehav Rev. In press. Liu Y, Morgan D, Roberts DCS (2007). Cross-sensitization of the reinforcing effects of cocaine and amphetamine in rats. Psychopharmacology (Berl) 195: Macey DJ, Rice WN, Freedland CS, Whitlow CT, Porrino LJ (2004). Patterns of functional activity associated with cocaine self-administration in the rat change over time. Psychopharmacology (Berl) 172(4): Martin BJ, Naughton BJ, Thirtamara-Rajamani K, Yoon DJ, Han DD, Devries AC, Gu HH (2011). Dopamine transporter inhibition is necessary for cocaine-induced increases in dendritic spine density in the nucleus accumbens. Synapse 65(6): Mateo Y, Lack CM, Morgan D, Roberts DC, Jones SR (2005). Reduced dopamine terminal function and insensitivity to cocaine following cocaine binge self-administration and deprivation. Neuropsychopharm 30(8): Meil WM, Roll JM, Grimm JW, Lynch AM, See RE (1995). Tolerance-like attenuation to contingent and noncontingent cocaine-induced elevation of extracellular dopamine in the ventral striatum following 7 days of withdrawal from chronic treatment. Psychopharmacology (Berl) 118(3): Mendelson JH, Sholar M, Mello NK, Teoh SK, Sholar JW (1998). Cocaine tolerance: behavioral, cardiovascular, and neuroendocrine function in men. Neuropsychopharmacology 18: Neumaier JF, Vincow ES, Arvanitogiannis A, Wise RA, Carlezon WA Jr (2002) Elevated expression of 5-HT1B receptors in nucleus accumbens efferents sensitizes animals to cocaine. J Neurosci. 22(24): Orio L, Edwards S, George O, Parsons LH, Koob GF (2009). A role for the endocannabinoid system in the increased motivation for cocaine in extended-access conditions. J Neurosci. 29(15):
165 144 Parsons LH, Justice JB Jr (1993) Serotonin and dopamine sensitization in the nucleus accumbens, ventral tegmental area, and dorsal raphe nucleus following repeated cocaine administration. J Neurochem. 61(5): Paterson NE, Markou A (2003). Increased motivation for self-administered cocaine after escalated cocaine intake. Neuroreport 14 (17): Peraile I, Torres E, Mayado A, Izco M, Lopez-Jimenez A, Lopez-Moreno JA, et al. (2010). Dopamine transporter down-regulation following repeated cocaine: implications for 3,4-methylenedioxymethamphetamine-induced acute effects and long-term neurotoxicity in mice. Br J Pharmacol 159(1): Reed SC, Haney M, Evans SM, Vadhan NP, Rubin E, Foltin RW (2009). Cardiovascular and subjective effects of repeated smoked cocaine administration in experienced cocaine users. Drug Alcohol Depend 102: Ritz MC, Lamb RJ, Goldberg SR, Kuhar MJ (1987) Cocaine receptors on dopamine transporters are related to self-administration of cocaine. Science. 237(4819): Roberts DC, Corcoran ME, Fibiger HC (1977) On the role of ascending catecholaminergic systems in intravenous self-administration of cocaine. Pharmacol Biochem Behav. 6(6): Schmidt HD, Pierce RC (2010). Cocaine-induced neuroadaptations in glutamate transmission: potential therapeutic targets for craving and addiction. Ann N Y Acad Sci. 1187: Steketee JD (2005) Cortical mechanisms of cocaine sensitization. Crit Rev Neurobiol. 17(2): Stuber GD, Britt JP, Bonci A (2011) Optogenetic modulation of neural circuits that underlie reward seeking. Biol Psychiatry. 71(12): Vanderschuren LJ, Kalivas PW (2000) Alterations in dopaminergic and glutamatergic transmission in the induction and expression of behavioral sensitization: a critical review of preclinical studies. Psychopharmacology (Berl), 151: Venton BJ, Seipel AT, Phillips PE, Wetsel WC, Gitler D, Greengard P, Augustine GJ, Wightman RM (2006) Cocaine increases dopamine release by mobilization of a synapsin-dependent reserve pool. J Neurosci. 26(12): Wheeler RA, Carelli RM (2009) Dissecting motivational circuitry to understand substance abuse. Neuropharmacology. 56 Suppl 1:
166 145 Willuhn I, Wanat MJ, Clark JJ, Phillips PE (2010). Dopamine signaling in the nucleus accumbens of animals self-administering drugs of abuse. Curr Top Behav Neurosci 3: Yorgason JT, España RA, Jones SR. (2011) Demon voltammetry and analysis software: analysis of cocaine-induced alterations in dopamine signaling using multiple kinetic measures. J Neurosci Methods202(2): Zapata A, Chefer VI, Ator R, Shippenberg TS, Rocha BA (2003). Behavioural sensitization and enhanced dopamine response in the nucleus accumbens after intravenous cocaine self-administration in mice. Eur J Neurosci 17(3): Zimmer BA, Oleson EB, Roberts DCS (2012). The Motivation to Self-Administer is Increased after a History of Spiking Brain Levels of Cocaine. Neuropsychopharmacology 37(8):
167 146 CHAPTER V METHYLPHENIDATE AND COCAINE SELF-ADMINISTRATION PRODUCE DISTINCT DOPAMINE TERMINAL ALTERATIONS Erin S. Calipari, Mark J. Ferris, James R. Melchoir, Kristel Bermejo, Ali Salahpour, David C.S. Roberts and Sara. R. Jones The following manuscript was published in Addiction Biology, in press, 2012, and is reprinted with permission. Stylistic variations are due to the requirements of the journal. Erin S. Calipari performed all voltammetry, tissue content, and self-administration experiments, as well as western blot hybridization for the dopamine transporter. Kristel Burmejo performed western blot hybridization for tyrosine hydroxylase as well as the polymerase chain reaction experiments. Erin S. Calipari prepared the manuscript and Drs. Mark J. Ferris, Ali Salahpour, and Sara R. Jones acted in an advisory and editorial capacity.
168 147 Abstract Methylphenidate (MPH) is a commonly abused psychostimulant prescribed for the treatment of attention deficit hyperactivity disorder. MPH has a mechanism of action similar to cocaine (COC) and is commonly characterized as a dopamine transporter (DAT) blocker. While there has been extensive work aimed at understanding dopamine (DA) nerve terminal changes following COC self-administration, very little is known about the effects of MPH self-administration on the DA system. We used fast scan cyclic voltammetry in nucleus accumbens core slices from animals with a five-day selfadministration history of 40 injections/day of either MPH (0.56 mg/kg) or COC (1.5 mg/kg) to explore alterations in baseline DA release and uptake kinetics as well as alterations in the interaction of each compound with the DAT. Although MPH and COC have similar behavioral effects, the consequences of self-administration on DA system parameters were found to be divergent. We show that COC self-administration reduced DAT levels and maximal rates of DA uptake, as well as reducing electrically stimulated release, suggesting decreased DA terminal function. In contrast, MPH self-administration increased DAT levels, DA uptake rates, and DA release, suggesting enhanced terminal function, which was supported by findings of increased metabolite/da tissue content ratios. Tyrosine hydroxylase mrna, protein and phosphorylation levels were also assessed in both groups. Additionally, COC self-administration reduced COC-induced DAT inhibition, while MPH self-administration increased MPH-induced DAT inhibition, suggesting opposite pharmacodynamic effects of these two drugs. These findings suggest that the factors governing DA system adaptations are more complicated than simple DA uptake blockade.
169 148 Introduction Methylphenidate (MPH), the active ingredient in medications (Ritalin, Methylin, Focalin ) used in the treatment of attention deficit/hyperactivity disorder (ADHD) and narcolepsy, is prone to diversion for non-medical use. MPH abuse in non- ADHD individuals has become increasingly prevalent through oral, intranasal or intravenous (i.v.) routes of administration (Teter et al., 2006; Sherman et al, 1987). The drug is used for cognitive enhancement and to get high, causing effects indistinguishable from cocaine (COC) when taken i.v. or intranasally (Morton and Stockton, 2000; Sherman et al, 1987; Teter et al., 2006; Rush and Baker, 2001). In 2008, 4.8 million people reported abusing MPH in the past year, compared to 5.3 million for COC, suggesting that both of these drugs pose a significant threat to public health (SAMHSA, National Survey on Drug Use and Health, 2008). From a behavioral standpoint, MPH and COC share a very similar preclinical profile. In rats, the discriminative stimulus effects of MPH and COC are nearly identical as seen by MPH s ability to fully substitute for COC (Li et al., 2006). Several i.v. selfadministration studies in rats have demonstrated that COC and MPH have similar reinforcing effects. Both compounds maintain high rates of operant responding, and extended access to MPH results in an escalation of intake similar to COC selfadministration (Ahmed & Koob, 1998; Marusich et al, 2010). With regard to reinforcing efficacy, doses of MPH or COC at the peak of their respective progressive ratio dose response curves engender similar break-points, indicating that the motivation to take each
170 149 drug is comparable (Marusich et al., 2010; Roberts et al., 2007; Calipari et al. unpublished). Like many psychostimulants, COC and MPH exert their rewarding and reinforcing properties primarily via their ability to bind to dopamine transporters (DATs) and inhibit dopamine (DA) uptake, elevating DA in the nucleus accumbens (DiChiara and Imperato, 1988; Woods and Meyer, 1991; Gerasimov et al., 2000). For example, a triple mutation in the second transmembrane domain of the DAT, which reduces affinity for MPH and COC, also abolishes conditioned place preference (Chen et al, 2005, 2006; Tilley et al, 2008a, b), highlighting the role of DAT inhibition in their rewarding properties. Acute inhibition of DA uptake by psychostimulants consistently results in enhanced DA signaling, but repeated or prolonged exposure to such drugs can lead to compensatory changes in presynaptic DA terminal function as well as alterations in the pharmacological actions of the drugs themselves (Biederman and Spender, 2002; Bouffard, 2003; Cadet et al., 2009). The neurobiological effects of MPH self-administration on the DA system remain to be elucidated; however, COC effects on the DA system have been well characterized and we hypothesized that MPH, a DAT blocker with a similar mechanism of action, would produce similar changes. In previous studies, COC self-administration resulted in lower basal levels of DA, decreased stimulated DA release and modestly decreased maximal rate of uptake (V max ) (Mateo et al., 2005; Ferris et al., 2011). In addition to baseline DA system alterations, COC self-administration also resulted in a decreased potency of COC in the nucleus accumbens, producing a marked tolerance to its DA-
171 150 elevating and DAT-inhibiting effects (Hurd et al., 1989; Mateo et al., 2005; Ferris et al., 2011). These data, taken together, suggest an overall hypodopaminergic and less responsive DA state following COC self-administration. Because MPH is increasingly becoming a drug of abuse (Marusich et al, 2009, 2010; Burton et al., 2010; Botly et al, 2008), but the neurochemical alterations following contingent administration have yet to be characterized, we chose to compare the behavioral and neurochemical effects of MPH self-administration with selfadministration of an extensively studied DA uptake blocker, COC. The purpose of this study was to systematically evaluate the neurochemical consequences of MPH and COC self-administration by evaluating baseline presynaptic DA system parameters such as release and uptake rate, as well as the ability of COC and MPH to inhibit the reuptake of endogenous DA. Surprisingly, MPH produced a distinctly different pattern of results from COC, suggesting that MPH does not behave as a prototypical dopamine uptake blocker. The present results suggest that the factors governing DA system adaptations are more complicated than simple DA uptake blockade. Materials and Methods Animals: Male, Sprague-Dawley rats ( g; Harlan Laboratories, Frederick, MD) were maintained according to the National Institutes of Health guidelines in Association for Assessment and Accreditation of Laboratory Animal Care accredited facilities. The experimental protocol was approved by the Institutional Animal Care and Use Committee at Wake Forest School of Medicine.
172 151 Self-Administration: Rats were anesthetized with ketamine (100 mg/kg) and xylazine (10 mg/kg), implanted with chronic indwelling jugular catheters, and trained for i.v. selfadministration as previously described (Roberts and Goeders 1989). Following surgery, animals were singly housed, and all self-administration sessions took place in the home cage. Each animal was maintained on a reverse light cycle (3:00 am lights off; 3:00 pm lights on), and all self-administration procedures occurred during the active/dark cycle. Sessions were six hours in length and were terminated at the end of the six hours or after 40 injections of drug. Animals self-administered either COC (1.5mg/kg/inj over 4 sec) or MPH (0.56 mg/kg/inj over 4 sec) on a fixed-ratio 1 schedule of administration. Concurrent with the start of each injection, the lever retracted and a stimulus light was activated for 20 seconds to signal a time-out period. Under these conditions, animals acquired a stable pattern of COC or MPH self-administration within 1 to 5 days. For self-administering animals, acquisition (Day 1) was counted when the animal reached 35 or more responses. There was no significant difference between groups in the injections received before acquisition criteria was met. Following acquisition, the animals were given access to 40 injections per day for a period of 5 consecutive days for both the COC and MPH self-administration experiments. Fixed ratio self-administration allows animals to titrate drug intake by spacing injections to maintain a preferred brain concentration of drug (Norman & Tsibulski, 2006). We compared doses of MPH and COC that were behaviorally equivalent. Due to the significant difference in half-life of MPH, which is approximately two times longer than COC (Huff and Davies, 2002; Weikop et al, 2004), we compared 0.56 mg/kg MPH and 1.5 mg/kg COC, doses which produce maximal responding on a progressive ratio
173 152 schedule of reinforcement (Richardson and Roberts 1996; Marusich et al., 2010). To confirm behavioral equivalence we also measured the time to complete session, interdose interval and rate of intake. Control animals were naïve rats housed under the same reverse light-dark light cycle for at least one week prior to neurochemical analysis. Fast Scan Cyclic Voltammetry in brain slices: Animals were sacrificed 24 hours after commencement of the last session, during the dark phase of the light cycle. 400 µm thick coronal brain sections containing the nucleus accumbens core were cut using a vibrating tissue slicer. A carbon fiber electrode was placed approximately 75 µm below the surface of the slice in close proximity to a bipolar stimulating electrode in the nucleus accumbens core. DA release was evoked by a single electrical pulse (300 μa, 4 msec, monophasic) applied every 5 minutes. Extracellular DA was recorded using fast-scan cyclic voltammetry (FSCV) by applying a triangular waveform 0.4 ( to +1.2 to 0.4 V vs. silver/silver chloride, 400 V/sec) to the electrode every 100 msec. Once the extracellular DA response was stable for three consecutive stimulations, individual brain slices were treated with either COC or MPH ( μm), with increasing concentrations cumulatively added after stable responses were recorded, approximately every 45 minutes. Western Blot Hybridization: Tissue sections were dissected 24 hours after commencement of the last self-administration session, and homogenized in ice-cold lysis buffer (10mM Tris, 2% SDS, 1mM EDTA) and spun in a centrifuge at 13,000g for 20
174 153 minutes to remove cellular debris. Bicinchoninic acid (BCA) protein assays (Thermo Scientific, Rockford, IL) were used to determine the protein content of each sample. Proteins were separated on 10% reducing SDS-PAGE gels (Invitrogen, Carlsbad, CA) and transferred to hydrophobic polyvinylidene difluoride membranes (Bio-Rad, Hercules, CA). For estimation of the molecular weight of the protein bands, Precision Plus protein standards ( kda, Bio-Rad) were run in parallel with the samples on gels. Nonspecific binding was blocked by incubation with blocking buffer (5% non-fat milk in tris-buffered saline with tween-20) for 30 minutes before incubation with primary antibodies: rabbit anti-dat antibody (Millipore, Temecula, CA; 1:5,000 dilution), rabbit anti-phosphorylated tyrosine hydroxylase (TH) Ser19, Ser31, Ser40 (PhosphoSolutions, Aurora, CO; 1:2000) and mouse anti-th (Millipore; 1:5000) at 4 C overnight. Membranes were then incubated with either secondary goat anti-rabbit HRP-conjugated antibody (Millipore; 1:10,000, Rockland, Gilbertsville, PA; 1:5000) or donkey antimouse antibody (Invitrogen; 1:5000). Immunoreactive products were visualized by either chemiluminescence (Pico chemiluminescent substrate, Thermo Scientific) or by infrared detection using the Odyssey LI-COR Infrared Imaging System and quantified using Image J (NIH). Protein loading was visualized by incubation with either a monoclonal antibody to β-actin (Abcam, Cambridge, MA; 1:10,000) or mouse anti- GAPDH (Sigma; 1:6000). Tissue Content: The striatum was dissected, snap-frozen, and samples (10-30 mg/sample wet weight) were homogenized in 250 μl of 0.1 M HClO 4 and analyzed for protein concentration by the BCA method (Thermo Scientific). Extracts were centrifuged and the supernatants removed and analyzed for DA and its metabolites 3,4-
175 154 dihydroxyphenylacetic acid (DOPAC) and homovanillic acid (HVA) using high performance liquid chromatography (HPLC) coupled to electrochemical detection at +220 mv (ESA Inc., Chelmsford, MA) and separated on a Luna 100 x 3.0 mm C 18 3 µm HPLC column (Phenomenex, Torrance, CA). The mobile phase consisted of 50 mm citric acid, 90 mm sodium dihydrogen phosphate, mm 1-octanesulfonic acid, 50 μm ethylenediaminetetracetic acid, % acetonitrile and 0.3 % triethylamine in a volume of 1 L (ph 3.0). Analytes were quantified using PowerChrom software (edaq Inc, Colorado Spring, CO) and a calibration curve. Quantitative Polymerase Chain Reaction (PCR): To obtain RNA from tissues, snapfrozen ventral tegmental area (VTA) tissue sections were homogenized in cold TRI- Reagent (BioShop Canada Inc) as described previously (Hu et al., 2009). Optical density readings at 260/280 nm were used to determine the quality and concentration of the resuspended RNA. 1µg of RNA was used for reverse-transcriptase PCR (RT-PCR) to obtain cdna using SuperScript III Reverse Transcriptase (Invitrogen) following the manufacturer s instructions. Relative quantification of gene targets was obtained using the GoTaq qpcr Master Mix (Promega, Fitchburg, Wisconsin) on the Applied Biosystems 7500 Real-Time PCR System. Phosphogycerlate kinase 1 (PGK1) was used as the reference gene (forward primer: ACCAAAGGATCAAGGCTGCTGTC; reverse primer: GACGGCCCAGGTGGCTCATA). Dopamine transporter (DAT) (forward primer: CCTGGTTCTACGGCGTCCAGC; reverse primer: GCCGCCAGTACAGGTTGGGT) and TH (forward primer: TTGAAGGAGCGGACTGGCTTCCA; reverse primer: TGGCCAGAAAATCACGGGCGG) were used as the target genes. Target genes were
176 155 relatively quantified using the ΔΔCt method, and are represented as % control normalized to PGK-1. Data Analysis: DA current was converted to concentration by calibration with 3 μm DA at the end of each experiment. For all analyses of FSCV data, DEMON voltammetry software was used (Yorgason et al., 2011). To evaluate the effects of MPH and COC self-administration on baseline DA system kinetics, evoked levels of DA were modeled using Michaelis-Menten kinetics, as a balance between release and uptake (Yorgason et al., 2011). For COC and MPH dose-response curves, apparent K m, a measure of apparent affinity of DA for the DAT, was used to determine changes in ability of the psychostimulants to inhibit DA uptake in the nucleus accumbens core relative to baseline. Statistics: Graph Pad Prism (version 4, La Jolla, CA) was used to statistically analyze data sets and create graphs. Baseline voltammetry, western blot hybridization, and PCR data were compared across groups using a two-tailed Student s t-test. Data obtained after perfusion of COC or MPH was subjected to a two-way analysis of variance with experimental group and concentration of COC or MPH as the factors. When significant interactions or main effects were obtained (p <.05), differences between groups were tested using Bonferroni post hoc tests. Behavioral data were subjected to a two-way analysis of variance with experimental group and hours to complete self-administration session as the factors.
177 156 Results COC and MPH intake increases over time. Each self-administration session was six hours in length and consisted of 40 injections per session. Time to complete 40 injections of COC (n = 8) significantly decreased over the five sessions (F (4, 7) = 8.858, p < 0.01,) (Fig. 1A, Top). In addition, the inter-infusion interval was also significantly decreased across sessions, demonstrating an escalation in rate of intake over sessions (F (4, 7) = 8.180, p < 0.01). MPH self-administration resulted in nearly identical changes in behavior. MPH self-administration (n = 11) engendered an increase in rate of lever pressing over selfadministration sessions (F (4, 10) = 7.956, p < 0.01) (Fig. 1A, Bottom). The same trend was observed with inter-infusion interval, demonstrating that the rate of intake also escalates across MPH self-administration sessions (F (4, 10) = 7.041, p < 0.01). Thus, the effects of MPH (n = 11) and COC (n = 8) self-administration on behavioral responding for drug were not significantly different as the two compounds produced the same inter-dose intervals and the same escalation (decreases in time to complete sessions) over days (Fig. 1B). Opposite effect of MPH and COC self-administration on baseline DA system kinetics. Baseline DA system kinetics were measured using FSCV and DAT levels were determined using western blot hybridization. COC self-administration (n=11) engendered a decrease in electrically stimulated DA release as compared to naïve control
178 157
179 158 Figure 1. Escalation in rate of cocaine (COC) and methylphenidate (MPH) selfadministration. (A) Representative self-administration plots from individual animals; each tick mark represents an infusion that was obtained. Five sessions with a maximum of 40 injections of either COC (1.5 mg/kg/inj) or MPH (0.56 mg/kg/inj) resulted in significant increases in rate of intake in over sessions. (B) The increase in rate of intake of was not significantly different between COC ( ) and MPH ( ).
180 159 animals (n=22) (t 31 = 2.348, p < 0.05, Fig. 2A, Center; Fig. 2B). Also, after COC selfadministration there was a significant decrease in maximal rate of DA uptake (t 30 = 2.719, p < 0.05) (Fig. 2A, Center; Fig. 2C). This decrease in maximal rate of uptake was accompanied by a decrease in DAT density in the COC group (n=5) compared to controls (n=3), as measured by western blot hybridization (t 6 = 2.182, p < 0.05) (Fig. 3A, Center; Fig. 3B). Relative expression levels of DAT mrna as measured by quantitative PCR in the VTA (n = 12) were not significantly different from controls (n = 18). Conversely, MPH self-administration resulted in increases in all DA system measurements. Stimulated DA release in the MPH group (n = 11) was increased compared to controls (n = 22) (t 31 = 2.076, p < 0.05) (Fig. 2A, Right; Fig. 2B). Maximal rate of DA uptake was also significantly increased in MPH (n = 7) versus control animals (n = 22) (t 24 = 2.719, p < 0.05) (Fig. 2A, Right; Fig. 2B). Total DAT levels were significantly increased following MPH self-administration (n = 4) compared to controls (n = 5) (t 7 = 3.532, p < 0.01) (Fig. 3A, Right; Fig. 3B). In addition, DAT mrna levels were assessed in the VTA. MPH group levels (n = 18) were not significantly different than control animals (n = 18), suggesting that the increases in protein expression are not due to increased synthesis of new protein. Differential Effects of COC and MPH SA on Tissue Content of DA and Metabolites. Post mortem tissue content of DA and its metabolites DOPAC and HVA were assessed in the striatum following COC and MPH SA (Fig. 4A). COC SA (n = 5) had no effect on DA or any of its metabolites as compared to controls (n = 6) (Fig. 4A). MPH SA (n = 9)
181 160 Figure 2. Baseline dopamine (DA) system kinetics following methylphenidate (MPH) and cocaine (COC) self-administration. (A) Representative traces of electricallyevoked DA signals in nucleus accumbens core slices from control, MPH selfadministration or COC self-administration animals. Traces show decreased maximal rate of uptake (rate of return to baseline) and DA release (peak height max) following COC self-administration and increased uptake and release following MPH self-administration. Insets: Background-subtracted cyclic voltammograms indicate signal is DA. (B) Grouped data showing that stimulated DA release is reduced after COC selfadministration and increased after MPH self-administration. (C) Grouped data showing that the maximal rate of DA uptake was decreased after COC self-administration and increased after MPH self-administration. *p < 0.05 versus control animals.
182 161 resulted in a significant decrease in DA tissue content as compared to control animals (n = 6) (t 13 = 3.598, p < 0.01) (Fig 4A). While DA tissue content was significantly decreased, there was no significant difference between control and MPH SA in the metabolites HVA or DOPAC (Fig. 4A). Metabolite to monoamine ratios were assessed in the striatum to determine changes in functional activity of the DA neurons in this region (Fig. 4B). COC SA (n = 5) did not produce significant differences in either DOPAC/DA or HVA/DA ratios as compared to controls (n = 6) (Fig. 4B). MPH SA (n = 9) resulted in a significant increase in the DOPAC/DA ratio as compared to control animals (n = 6) (t 13 = 2.852, p < 0.01), and although not significant, there was a trend towards an increased HVA/DA ratio (t 13 = 2.144, p = ) (Fig. 4B). No effect of MPH or COC SA on Tyrosine Hydroxylase Protein, mrna, or Phosphorylation Levels. To determine if changes in stimulated DA release could be due to differences in TH, mrna levels of TH were assessed in the VTA of COC and MPH groups using quantitative PCR. COC SA (n = 18) resulted in a significant increase in TH mrna levels as compared to controls (n = 12) (t 28 = 5.704, p < ) (Fig 5A). MPH SA (n=18) also significantly increased levels of TH mrna in the VTA versus controls (n = 18) (t 34 = 2.480, p < 0.05) (Fig 5A), although the levels were higher in COC (n = 12) than MPH groups (n = 18) (t 28 = 4.475, p < ) (Fig. 5A). However, this difference was not reflected in protein levels (see below; Fig. 5 B, C, D), highlighting the well known complex translational regulation of this protein (Tank et al., 2008).
 Representative photographs of Western blots on tissue from the nucleus accumbens core region of control, COC self-administration, and MPH self-administration groups.")
183 162 Figure 3. Western blot hybridization for the dopamine transporter (DAT) after cocaine (COC) or methylphenidate (MPH) self-administration. (A) Representative photographs of Western blots on tissue from the nucleus accumbens core region of control, COC self-administration, and MPH self-administration groups. (B) COC selfadministration reduced DAT levels while MPH self-administration increased DAT levels in the nucleus accumbens. Protein expression levels were determined as the ratio of DAT to the level of β-actin and are reported as percent control values. *p < 0.05 versus control group.
184 163
185 164 Figure 4. Tissue content of dopamine (DA) and metabolites (A) in the striatum of cocaine (COC) and methylphenidate (MPH) self-administration groups. The MPH group had significantly lower DA tissue content levels with no differences in metabolite levels. Tissue content levels of DA and metabolites were unaffected by COC selfadministration. (B) Metabolite/monoamine ratios, a measure of functional activity, in MPH and COC groups as compared to controls. COC self-administration did not affect ratios, while MPH increased DOPAC/DA ratio. *p < 0.05 versus control group.
186 165
187 166 Figure 5. Quantification of tyrosine hydroxylase (TH) mrna levels, protein levels, and phosphorylation by western blot hybridization and quantitative PCR. (A) TH mrna expression levels in the ventral tegmental area (VTA) of methylphenidate (MPH) and cocaine (COC) self-administering rats. TH mrna levels were significantly increased in both groups as compared to controls. The COC group had significantly higher expression levels as compared to the MPH group. TH mrna levels were normalized to phosphogycerlate kinase 1 (PGK-1) and expressed as a percent of control group. (B) Representative western blot images for total TH and two phosphorylation sites (Serine 19, 40) in the striatum (left) and VTA (right). (C) There were no differences in TH or its phosphorylation sites in the striatum or (D) VTA in any of the groups tested. Protein levels were determined as the ratio of DAT to the level of GAPDH and are reported as percent control values. *** p < versus control group, # p < versus MPH group.
188 167 TH protein levels as well as two phosphorylation sites were quantified via western blot hybridization (Fig 5B). COC SA (n = 5) did not change total TH levels or phosphorylation at either serine 40 (THp Ser40) or 19 (THp Ser19) in the striatum (Fig. 5B, left; 5C) or VTA (Fig. 5B right; 5D) as compared to controls (n = 6). Serine 31 was not quantifiable in any samples. Similarly, MPH SA (n = 7) also resulted in no changes in TH, THp Ser40, or THp Ser19, in either the striatum (Fig. 5B, left; 5C) or the VTA (Fig. 5B; 5D) as compared to controls (n = 6). MPH self-administration resulted in increased MPH potency, while COC potency remained unchanged. Cumulative dose-response curves of COC and MPH were collected to determine changes in the compounds ability to inhibit DA uptake following MPH self-administration. The Michaelis-Menten measure of DA affinity for the transporter, apparent K m, was used to measure changes in the ability of a drug to inhibit DA uptake relative to baseline. Following MPH self-administration (n = 4), there was a significant main effect of MPH treatment as compared to controls (n = 4) (F (4, 1) = 23.74, p < 0.001), with significant increases in the maximal apparent K m of MPH selfadministration groups at 30uM (p < 0.001) (Fig. 6D). MPH self-administration resulted in a leftward shift in the dose-response curve, indicating that MPH potency is increased following MPH self-administration. Cumulative COC dose response curves were performed to determine if there was a shift in the potency of COC following MPH self-administration. There was no change in the dose response curve for COC (n = 3) as compared to controls (n = 3), indicating
189 168 that COC s ability to inhibit DA uptake in the nucleus accumbens core remained unchanged after MPH self-administration (Fig. 6E). COC self-administration resulted in decreased COC potency, while MPH potency remained unchanged. COC self-administration resulted in an overall group effect (F (4, 1) = 13.85, p < 0.01) and a rightward shift in the dose response curve for COC (n = 3) as compared to controls (n = 4), signifying a significant decrease in COC-induced DA uptake inhibition (Fig. 7E). Bonferroni analysis demonstrated a significant effect at 30uM (p < 0.001). Conversely, following 5 days of COC self-administration, there was no significant change in MPH s DA uptake-inhibiting effects (Fig. 7D), indicating that MPH (n = 5) potency remained unaltered as compared to controls (n = 3) following repeated administration of COC. Discussion MPH and COC are both DAT inhibitors and produce similar acute DA-elevating effects. Despite the close similarities in self-administration behavior, the current study demonstrates that MPH and COC intake can produce strikingly different effects on DA system kinetics and drug potency. Consistent with previous studies, 5 days of high-dose COC self-administration resulted in decreased DAT density, maximal rate of uptake (V max ), and stimulated DA release (Mateo et al., 2005; Ferris et al., 2011). In contrast,
190 169
191 170 Figure 6. Significantly greater dopamine (DA) uptake inhibition by methylphenidate (MPH), but not cocaine (COC), after MPH self-administration. The effect of COC and MPH applied to nucleus accumbens core slices on representative electrically-evoked (one pulse) DA signals is shown in (A) Pre-drug (B) 10µM MPH and (C) 10 µm COC from control and MPH self-administering rats. Representative plots are shown as percent peak height; control and MPH self-administration animals are overlaid. MPH-induced DA uptake inhibition is present in naive animals but significantly increased in MPH self-administering rats as depicted by a longer time course of the signal to return to baseline; there is no effect of MPH self-administration on COC uptake inhibition. Group data showed that MPH inhibited DA uptake to a greater extent (D), and that COC uptake is unaffected (E) following 5 days of MPH self-administration ( ) compared with control rats ( ),with uptake inhibition measured as apparent K m (***p <.001
192 171
193 172 Figure 7. Significant reduction in dopamine (DA) uptake inhibition by cocaine (COC), but not methylphenidate (MPH), after COC self-administration. The effect of COC and MPH on representative electrically evoked (one pulse) DA signals in nucleus accumbens core slices is shown. (A) Pre-drug (B) 10µM MPH and (C) 10 µm COC from control and cocaine self-administering rats. Representative plots are represented as percent peak height; control and COC self-administration animals are overlaid. COCinduced DA uptake inhibition is significantly reduced in cocaine self-administering rats as evidenced by a shorter time course of the signal s return to baseline; there is no effect of COC self-administration on MPH uptake inhibition. Group data showed that MPH uptake is unaffected (D) but DA uptake inhibition by COC is reduced (E) following 5 days of COC self-administration ( ) compared with control rats ( ), measured as apparent K m (***p <.001).
194 173 MPH self-administration increased DAT density, V max, and stimulated DA release. In addition to robust differences in baseline DA nerve-terminal kinetics, COC self-administration resulted in a decreased potency of COC to inhibit DA uptake, with no change in potency of MPH. Conversely, MPH self-administration resulted in increased potency of MPH and no change in COC potency. Overall, COC self-administration seems to result in sub-sensitive DA terminals while MPH self-administration produces super-sensitive terminals. These profound differences in the neurobiological effects of two DA uptake inhibitors provide evidence that the DA-elevating effects of psychostimulants are not solely, or even primarily, responsible for altered DA system function after repeated exposure. The goal of this study was to compare DA system alterations that occurred following 5 days of COC or MPH self-administration. One challenge was to select comparable doses of these two drugs with different time courses and binding affinities. The doses we chose for MPH (0.56 mg/kg) and COC (1.5 mg/kg) are at the peak of the respective PR dose response curves, indicating that these are the maximally reinforcing dose of each drug. The selected doses of these two drugs produced similar inter-injection intervals within a session and produced escalation in rates of intake across sessions, indicating that these doses are behaviorally equivalent. Therefore, although there are differences in half-life and potency, we were able to equate the behavioral responses to these drugs in order to accurately compare the neurochemical consequences of MPH and COC self-administration.
195 174 Despite almost identical behavioral profiles, baseline changes in maximal DA uptake rate (V max ) and stimulated DA release following MPH and COC selfadministration were found to be opposite. COC self-administration resulted in a decreased V max and decreased stimulated release, effects that have been observed previously with the same paradigm (Ferris et al., 2011). DAT density was found to be decreased following COC self-administration, suggesting that the observed decrease in uptake rate was due to reduced numbers of DA transporters. The alterations in DAT density following self-administration are most likely due to differences in protein processing as there were no observed changes in mrna levels in COC or MPH selfadministration groups. In the literature, there are reports of increased, decreased and unaltered DA uptake rates following COC self-administration (Ferris et al., 2011; Ramamoorthy et al., 2010; Mateo et al., 2005; Wilson and Kish, 1996), and clearly, different self-administration paradigms produce different neurobiological effects, emphasizing the need for equating doses as well as injection frequency, timing and duration. Self-administration of MPH resulted in an increase in V max and DAT density as well as an increase in stimulated DA release in the nucleus accumbens core, demonstrating that two drugs which cause identical self-administration behavior are capable of causing opposite neurobiological adaptations. To address the differences in stimulated DA release between COC and MPH groups, various markers of presynaptic function were evaluated. DA tissue content was found to be decreased in the MPH group, with corresponding increases in metabolite/da ratios, indicating increased functional activity, consistent with the increased DA release in this group. No changes were found in the COC group. A number of factors could be
196 175 responsible for the observed difference between COC and MPH, for example, MPH has been shown to redistribute vesicular DA pools (Volz et al., 2008) and may alter both release and tissue content directly. Also, MPH has a longer half-life of uptake inhibition than COC, which in combination with the increased release and ability to mobilize DA vesicles, could lead to a reduced ability to refill vesicular stores and explain the difference in tissue content between the two groups. These findings further emphasize the differential impact of these two psychostimulant blockers on the DA system. We initially postulated that the MPH-induced DA release and tissue content changes might be explained by alterations in TH, the rate-limiting enzyme in DA synthesis, but no differences were found in protein or phosphorylation levels, indicating that TH is most likely not driving the changes. There were, however, increases in TH mrna levels in both self-administration groups, indicating that either post-translational processing or protein degradation may be altered. With regard to the interaction of stimulants with the DAT, previous work found reduced pharmacological effects of COC in the nucleus accumbens after high-dose COC self-administration (Hurd et al., 1989; Mateo et al., 2005; Ferris et al., 2011). In order to determine if the effects of COC self-administration were unique to cocaine or common to all psychostimulants, Ferris et al. (2011) determined that the effects of amphetamine (AMPH) on DA uptake in nucleus accumbens core slices remained unchanged following COC self-administration. The same study also showed that self-administration of AMPH had no effect on either COC or AMPH potency in slices, suggesting that COC selfadministration produced COC-specific pharmacological changes. We hypothesized that
197 176 AMPH did not alter the pharmacology of the DAT because it was recognized as a substrate/releaser, while COC, a pure uptake blocker, induced allosteric modifications resulting in altered pharmacology. Next, it was reasonable to test whether selfadministration of a structurally dissimilar uptake inhibitor, MPH, would also produce altered DAT pharmacology. The current results demonstrate that MPH selfadministration produces opposite effects from COC self-administration, with increased MPH potency on slices but no change in COC effects. Thus, it appears that DAT pharmacology is altered according to individual drug interactions with the DAT and not according to drug class, such as blocker or releaser. While MPH and COC are both classified as uptake blockers, they have different chemical structures, which may account for the disparate neurochemical changes that occur following self-administration. MPH is structurally related to amphetamine, while COC is a tropane (Uhl et al., 2002; Volkow et al, 2002; Heal et al, 2009; Han & Gu 2006). MPH, although typically categorized as a DAT blocker, has unique properties that are distinct from both releasers and blockers. Studies using cell culture and striatal tissue preparations have shown that MPH exhibits binding properties of both blockers and releasers (Dar et al, 2005; Wayment et al, 1999). MPH is not a DAT substrate, as it is not transported into cells (Sonders et al, 1997); however, some studies have shown that MPH has the ability to promote reverse-transport of DA through the DAT at very high doses (Sproson et al, 2001; Russell et al, 1998; Heal et al, 1996). It is clear that DAT inhibition-induced increases in extracellular DA levels do not explain the distinct adaptations produced by the two drugs studied here, and we hypothesize that drug interactions with specific regions of the DAT may explain the differences observed.
198 177 Changes in DAT density and V max do not explain the potency changes found with MPH and COC. With regard to COC self-administration, in the present study we show decreases in DAT protein levels and V max for DA uptake after COC self-administration. However, in previous studies with COC self-administration, V max was either increased or unchanged, with the same reduction in potency for DA uptake inhibition in slices (Mateo et al., 2005; Ferris et al., 2011). With regard to MPH self-administration, to our knowledge this is the first study examining neurochemical changes and thus we cannot compare to other regimens; however, the Michaelis-Menten based kinetic model used to fit the DA release and uptake profiles takes into account any changes in baseline uptake rate using the pre-drug electrically stimulated DA overflow curves, and this allows the effects of uptake inhibition to be separated from changes in maximal uptake rate. This indicates that the maximal DA uptake rate (V max ), which is correlated with DA transporter number, cannot account for the observed alterations in DA uptake inhibition by both COC and MPH. Thus, these effects are most likely due to allosteric alterations in the DA transporter, making it more or less sensitive to inhibition by specific drugs. These DA system alterations induced by self-administration of COC or MPH have important implications for the subsequent abuse or co-abuse of other psychostimulants. MPH and COC are dependent upon their actions at the DAT for their reinforcing effects, and changes that occur to the DA system would be expected to affect the reinforcing potency or efficacy of other psychostimulant drugs (Chen et al, 2005, 2006; Tilley et al, 2008 a, b). Because psychostimulants that are DA releasers rely on DATs to have their reverse-transport effects, the present results from MPH selfadministering animals would predict greater behavioral potency. Consistent with this
199 178 prediction, previous studies have already demonstrated that MPH self-administration, as well as increasing DAT levels by transgenic overexpression, results in increased AMPHinduced locomotor activity (Burton et al., 2010; Salahpour et al., 2008; Yang et al., 2003). In addition, epidemiological studies have provided evidence that individuals who abuse MPH are more likely to abuse other substances and to have problems with drug use (Barrett et al, 2005; Teter et al, 2006; Wilens et al, 2008). Given the widespread abuse of MPH and the potential risk for greater stimulant effects of other drugs, further studies of the consequences of MPH self-administration will be important. Acknowledgements This work was funded by NIH grants R01DA024095, RO1DA03016 (SRJ), R01DA (DCSR), T32DA (MJF), Institutional NRSA T32DA and Individual NRSA F31 DA (ESC), and a Canadian Institutes of Health Research (CIHR) grant (AS).
200 179 References Ahmed SH, Koob GF (1998) Transition from moderate to excessive drug intake: change in hedonic set point. Science 282(5387): Barrett SP, Darredeau C, Bordy LE, Pihl RO (2005) Characteristics of methylphenidate misuse in a university student sample. Can J Psychiatry 50: Biederman J, Spencer T (2002) Methylphenidate in treatment of adults with attentiondeficit/hyperactivity disorder. J Atten Disord 6:S Botly LC, Burton CL, Rizos Z, Fletcher PJ (2008) Characterization of methylphenidate self-administration and reinstatement in the rat. Psychopharmacology (Berl). 199(1): Bouffard R, Hechtman L, Minde K, Iaboni-Kassab F (2003) The efficacy of 2 different dosages of methylphenidate in treating adults with attention-deficit hyperactivity disorder. Can J Psychiatry 48: Burton CL, Nobrega JN, Fletcher PJ (2010) The effects of adolescent methylphenidate self-administration on responding for a conditioned reward, amphetamine-induced locomotor activity, and neuronal activation. Psychopharmacology (Berl) 208(3): Cadet JL, McCoy MT, Cai NS, Krasnova IN, Ladenheim B, Beauvais G, Wilson N, Wood W, Becker KG, Hodges AB (2009) Methamphetamine preconditioning alters midbrain transcriptional responses to methamphetamine-induced injury in the rat striatum. PLoS One. 12;4(11):e7812. Chen R, Han DD, Gu HH (2005) A triple mutation in the second transmembrane domain of mouse dopamine transporter markedly decreases sensitivity to cocaine and methylphenidate. J Neurochem 94(2): Chen R, Tilley MR, Wei H, Zhou F, Zhou FM, Ching S, Quan N, Stephens RL, Hill ER, Nottoli T, Han DD, Gu HH (2006) Abolished cocaine reward in mice with a cocaineinsensitive dopamine transporter. Proc Natl Acad Sci U S A.103(24): Dar DE, Mayo C, Uhl GR (2005) The interaction of methylphenidate and benztropine with the dopamine transporter is different than other substrates and ligands. Biochem Pharmacol.70(3): Di Chiara G, Imperato A (1988) Drugs abused by humans preferentially increase synaptic dopamine concentrations in the mesolimbic system of freely moving rats. Proc Natl Acad Sci U S A 85(14):
201 180 Ferris MJ, Mateo Y, Roberts DC, Jones SR (2011) Cocaine-insensitive dopamine transporters with intact substrate transport produced by self-administration. Biol Psychiatry 69(3): Gerasimov MR, Franceschi M, Volkow ND, Gifford A, Gatley SJ, Marsteller D, Molina PE, Dewey SL (2000) Comparison between intraperitoneal and oral methylphenidate administration: A microdialysis and locomotor activity study. J Pharmacol Exp Ther. 295(1):51-7. Han DD, Gu HH (2006) Comparison of the monoamine transporters from human and mouse in their sensitivities to psychostimulant drugs. BMC Pharmacol. 3(6):6. Heal DJ, Cheetham SC, Smith SL (2009) The neuropharmacology of ADHD drugs in vivo: insights on efficacy and safety. Neuropharmacology 57(7-8): Heal DJ, Prow MR, Hearson M, Buckett WR (1996) Efflux of [3H]dopamine from superfused rat striatal slices: predictive value for detecting stimulant drugs of abuse. Br. J. Pharmacol 117:325P. Hu YF, Caron MG, Sieber-Blum M (2009) Norepinephrine transport-mediated gene expression in noradrenergic neurogenesis. BMC Genomics. 10:151. Huff JK, Davies MI (2002) Microdialysis monitoring of methylphenidate in blood and brain correlated with changes in dopamine and rat activity. J Pharm Biomed Anal. 29(5): Hurd YL, Weiss F, Koob GF, And NE, Ungerstedt U (1989) Cocaine reinforcement and extracellular dopamine overflow in rat nucleus accumbens: an in vivo microdialysis study. U. Brain Res. 498(1): Li SM, Campbell BL, Katz JL (2006) Interactions of cocaine with dopamine uptake inhibitors or dopamine releasers in rats discriminating cocaine. J Pharmacol Exp Ther. 317(3): Marusich JA, Bardo MT (2009) Differences in impulsivity on a delay-discounting task predict self-administration of a low unit dose of methylphenidate in rats. Behav Pharmacol. 20(5-6): Marusich JA, Beckmann JS, Gipson CD, Bardo MT (2010) Methylphenidate as a reinforcer for rats: contingent delivery and intake escalation. Exp Clin Psychopharmacol 18(3): Mateo Y, Lack CM, Morgan D, Roberts DC, Jones SR (2005) Reduced dopamine terminal function and insensitivity to cocaine following cocaine binge self-administration and deprivation. Neuropsychopharm 30(8):
202 181 Morton WA, Stockton GG (2000) Methylphenidate Abuse and Psychiatric Side Effects. Prim Care Companion J Clin Psychiatry. 2(5): Norman AB, Tsibulsky VL (2006) The compulsion zone: a pharmacological theory of acquired cocaine self-administration. Brain Res. 1116(1): Ramamoorthy S, Samuvel DJ, Balasubramaniam A, See RE, Jayanthi LD (2010) Altered dopamine transporter function and phosphorylation following chronic cocaine selfadministration and extinction in rats. Biochem Biophys Res Commun. 391(3): Richardson NR, Roberts DC (1996) Progressive ratio schedules in drug selfadministration studies in rats: a method to evaluate reinforcing efficacy. J Neurosci Methods. 66(1):1-11. Ritz MC, Lamb RJ, Goldberg SR, Kuhar MJ (1987) Cocaine receptors on dopamine transporters are related to self-administration of cocaine. Science. 237(4819): Roberts DC, Goeders NE (1989) Drug self-administration: Experimental methods and determiants. Boalton AA, Baker GB, Greenshaw AS (eds). In: Neuromethods, vol. 13. Psychopharmacology. Humana press: Clifton, NJ. pp Rush CR, Baker RW (2001) Behavioral pharmacological similarities between methylphenidate and cocaine in cocaine abusers. Exp Clin Psychopharmacol. 9(1): Russell V, de Villiers A, Sagvolden T, Lamm M, Taljaard J (1998) Differences between electrically-, ritalin- and D-amphetamine-stimulated release of [3H]dopamine from brain slices suggest impaired vesicular storage of dopamine in an animal model of Attention- Deficit Hyperactivity Disorder. Behav Brain Res. 94(1): Sherman CB, Hudson LD, Pierson DJ (1987) Severe precocious emphysema in intravenous methylphenidate (Ritalin) abusers. Chest 92(6): Sonders MS, Zhu SJ, Zahniser NR, Kavanaugh MP, Amara SG (1997) Multiple ionic conductances of the human dopamine transporter: the actions of dopamine and psychostimulants. J Neurosci 17(3): Sproson EJ, Chantrey J, Hollis C, Marsden CA, Fonel KC (2001) Effect of repeated methylphenidate administration on presynaptic dopamine and behaviour in young adult rats. J Psychopharmacol 15(2): Substance Abuse and Mental Health Services Administration (SAMHSA) (2008). National survey on drug use and health. Retrieved from
203 182 Tank AW, Xu L, Chen X, Radcliffe P, Sterling CR (2008) Post-transcriptional regulation of tyrosine hydroxylase expression in adrenal medulla and brain. Ann N Y Acad Sci. 1148: Teter CJ, McCabe SE, LaGrange K, Cranford JA, Boyd CJ (2006) Illicit use of specific prescription stimulants among college students: prevalence, motives, and routes of administration. Pharmacotherapy 26(10): Tilley MR, Gu HH (2008a) The effects of methylphenidate on knockin mice with a methylphenidate-resistant dopamine transporter. J Pharmacol Exp Ther. 327(2): Tilley MR, Gu HH (2008b)Dopamine transporter inhibition is required for cocaineinduced stereotypy. Neuroreport. 19(11): Uhl GR, Hall FS, Sora I (2002) Cocaine, reward, movement and monoamine transporters. Mol Psychiatry.7(1):21-6. Volkow ND, Fowler JS, Wang G, Ding Y, Gatley SJ (2002) Mechanism of action of methylphenidate: insights from PET imaging studies. J Atten Disord 6 Suppl 1:S31-43 Volkow ND, Wang GJ, Fowler JS, Fischman M, Foltin R, Abumrad NN, Gatley SJ, Logan J, Wong C, Gifford A, Ding YS, Hitzemann R, Pappas N (1999) Methylphenidate and cocaine have a similar in vivo potency to block dopamine transporters in the human brain. Life Sci. 65(1):PL7-12. Volz TJ, Farnsworth SJ, Hanson GR, Fleckenstein AE (2008) Methylphenidate-induced alterations in synaptic vesicle trafficking and activity. Ann N Y Acad Sci. 1139: Wayment HK, Deutsch H, Schweri MM, Schenk JO (1999) Effects of methylphenidate analogues on phenethylamine substrates for the striatal dopamine transporter: potential as amphetamine antagonists? J Neurochem. 72(3): Weikop P, Egestad B, Kehr J (2004) Application of triple-probe microdialysis for fast pharmacokinetic/pharmacodynamic evaluation of dopamimetic activity of drug candidates in the rat brain.j Neurosci Methods.140 (1-2): Wilens TE, Adler LA, Adams J, Sgambati S, Rotrosen J, Sawtelle R, Utzinger L, Fusillo S (2008) Misuse and diversion of stimulants prescribed for ADHD: a systematic review of the literature. J Am Acad Child Adolesc Psychiatry. 47(1): Wilson JM, Kish SJ (1996) The vesicular monoamine transporter, in contrast to the dopamine transporter, is not altered by chronic cocaine self-administration in the rat. J Neurosci. 16(10):
204 183 Woods SK, Meyer JS (1991) Exogenous tyrosine potentiates the methylphenidateinduced increase in extracellular dopamine in the nucleus accumbens: a microdialysis study. Brain Res. 560(1-2): Yang PB, Swann AC, Dafny N (2003) Chronic pretreatment with methylphenidate induces cross-sensitization with amphetamine. Life Sci. 73(22): Yorgason JT, España RA, Jones SR (20110) Demon Voltammetry and Analysis software: Analysis of cocaine-induced alterations in dopamine signaling using multiple kinetic measures. J Neurosci Methods. 202(2):
205 184 CHAPTER VI METHYLPHENIDATE AMPLIFIES THE POTENCY AND REINFORCING EFFECTS OF AMPHETAMINES BY INCREASING DOPAMINE TRASNPORTER EXPRESSION Erin S. Calipari, Mark J. Ferris, Ali Salahpour, Marc G. Caron and Sara. R. Jones The following manuscript was published in Nature Communications, in press, 2013, and is reprinted with permission. Stylistic variations are due to the requirements of the journal. Erin S. Calipari performed the experiments and prepared the manuscript and Drs. Mark J. Ferris, Ali Salahpour, and Sara R. Jones acted in an advisory and editorial capacity.
206 185 Methylphenidate (MPH) is commonly diverted for non-clinical use, but the neurobiological consequences of exposure to MPH at high doses are not well defined. Here we show that MPH self-administration increases dopamine transporter (DAT) levels, driving enhanced potency and drug-seeking for MPH and amphetamine (AMPH), without altering cocaine potency. To corroborate these effects, genetic over-expression of the DAT increased AMPH and MPH potencies, but not cocaine potency, suggesting that individuals with elevated DAT levels, such as ADHD sufferers, may be more susceptible to the addictive effects of amphetamine-like drugs. Methylphenidate (MPH), the active compound in Ritalin, is commonly used offlabel, with up to 17% of college students reporting abusing MPH, alone and in combination with other stimulants, for its cognitive enhancing or euphoric effects 1. MPH is taken orally, intranasally or intravenously, and is one of the prescribed drugs most diverted onto the illicit market worldwide 2. There are a limited number of studies of MPH self-administration in rats, and little is known about the neurobiological consequences of exposure to high doses. While comparable cocaine self-administration paradigms have produced marked tolerance to cocaine s effects 3, here we show sensitizing effects of MPH. These experiments reveal a mechanism whereby increased DAT levels drive enhanced potency and motivation to administer MPH and dopamine (DA) releaser drugs such as AMPH. One implication of these findings is that individual variations in DAT levels (e.g. ADHD sufferers 4 ) may predict susceptibility to abuse of compounds such as MPH, AMPH and methamphetamine. Rats self-administered MPH (0.56 mg/kg/infusion, i.v.) on a fixed-ratio one schedule for five days. This resulted in escalation of first-hour intake (F (4, 10) = 20.00, p
207 186 < 0.001) as well as overall rate of intake (F (4, 10) = 7.956, p < 0.001; Fig 1 A, B). To examine the motivation to administer MPH, we measured reinforcing efficacy using a progressive ratio paradigm and found an increase in breakpoint (t 12 = 2.068, p < 0.05; Fig 1 C) following MPH self-administration. The reinforcing and rewarding effects of many drugs of abuse are related to their ability to elevate DA in the nucleus accumbens (NAc) 5. While the NAc shell is involved in the acute rewarding effects of drugs and acquisition of self-administration, the NAc core plays a critical role in cue-reward learning as well as continued responding for drugs after repeated administration as in the current paradigm 6. We found DA uptake in the NAc core to be supersensitive to MPH inhibition following MPH self-administration (F (1, 9) = 25.11, p < 0.05; Fig 1 D, E). The proposed mechanism for augmented MPH potency is increased DAT levels, which has been suggested to modulate the potency of DA releasers, although not MPH 7. After MPH selfadministration, the maximal rate of DA uptake was increased (V max ; t60 = 2.719, p < 0.01) (Fig 1 G, H), due to increased total DAT levels 8. To further understand the impact of MPH self-administration on psychostimulant potencies, we examined a number of DAT blockers and DA releasers. We found that MPH self-administration had no effect on blocker potency, but enhanced the potency of releasers. Thus, the ability of the blockers cocaine (Fig 2 A, B) and nomifensine (Sup. Fig 1 C, D) to inhibit DA uptake was unaffected by MPH self-administration. In contrast, the potency of AMPH (F (1, 4) = 76.81, p < 0.001; Fig 2 D, E) and methamphetamine (F (1, 10) = 33.51, p < 0.001; Sup. Fig 2 A, B) was increased. Given the

208 187
209 188 Figure 1. Methylphenidate (MPH) is self-administered, increases dopamine uptake (V max ), and MPH potency. A, Event record from a representative animal. B, increased rate of responding for MPH on a fixed ratio 1 schedule. Session 1 refers to the first session following acquisition. C, progressive ratio responding for MPH in minimally trained (CTRL; black) and MPH self-administration (SA; red) animals. D, E, increased ability of MPH to inhibit the dopamine transporter (DAT) following MPH SA. F, doseresponse curve using the threshold SA procedure in MPH SA and CTRL to assess motivation to administer MPH. G, H, I, increased V max and stimulated dopamine release ([DA]/pulse) following MPH SA. *p < 0.05
210 189
211 190 Figure 2. Methylphenidate (MPH) self-administration (SA) increases the potency and motivation to administer releasers, but not blockers. A, B, the ability of cocaine (COC) to inhibit the dopamine transporter (DAT) following MPH SA is unchanged. C, motivation to administer cocaine is unchanged following MPH. D, E, increased AMPH potency, and motivation to administer AMPH (F) following MPH SA. ***p < 0.001, **p < 0.01, *p < 0.05
212 191 enhanced potency of DA releasers, it is possible that these compounds will have increased abuse liability following MPH abuse in the human population. Therefore, motivation to take drugs was measured using a threshold procedure to determine the maximal price (P max ) an animal is willing to pay to receive drug. After a history of MPH self-administration, there was increased responding (F (1, 13) = 35.16, p < 0.001; Fig 1F) and P max for MPH (t 14 = 2.196, p < 0.05; Sup. Fig 2 A) and AMPH (responding, F (1, 11) = 51.60, p < 0.001; Fig 2 F; P max, t 10 = 3.136, p < 0.01; Sup. Fig 2 B), but not cocaine (Fig 2 C; Sup. Fig 2 C). This shows that the increased potency of MPH and AMPH correlates with increased drug-seeking behavior. We hypothesized that the drug class-specific increase in releaser potency was due to increased DAT levels. To test this hypothesis we used DAT over-expressing mice (DAT-tg), which have increased levels of native, non-drug altered DAT 7, and increased V max (t 31 = 3.787, p < 0.001; Fig 3 A, B). In DAT-Tg mice, there was an increased ability of MPH (F (1, 9) = 21.44, p < 0.001; Fig 3 D), and AMPH (F (1, 9) = 16.05, p < 0.001; Fig 3 E), but not cocaine (Fig 3 F), to inhibit DA uptake. Further, DAT-tg animals showed increased MPH- (F (1, 10) = 4.093, p < 0.05; Fig 3 G) and AMPH- (F (1, 12) = 9.934, p < 0.01; Fig 3 H), but not cocaine- (Fig 3 I) induced locomotion. Increased potency of releasers and MPH in the presence of high V max /DAT levels may be important in clinical treatment settings, as individuals with ADHD have welldocumented increases in DAT levels ranging from 17-70% 4. These findings may explain in part the high frequency of drug abuse in untreated ADHD sufferers 9, and these individuals may be more susceptible to the neurochemical, behavioral, and neurotoxic effects of MPH, AMPH and other releasers like methamphetamine and bath salts.
213 192
214 193 Figure 3. Overexpression of the dopamine transporter (DAT) increases the behavioral and neurochemical potency of methylphenidate (MPH) and releasers. A, B, C increased rate of dopamine uptake (V max ) and increased stimulated dopamine release ([DA]/pulse) in DAT overexpressing mice (DAT-tg; blue) relative to wild-type (WT; green). D, E, F, concentration-response curves for MPH, cocaine, and amphetamine (AMPH) for inhibiting DA uptake. Enhanced locomotor activating effects of MPH (G) and AMPH (H), but not cocaine (I), in DAT-tg mice. ***p < 0.001, *p < 0.05
215 194 Acknowledgements: This work was funded by NIH grants R01 DA021325, R01 DA (SRJ), R01 DA (DCSR), P50 DA06634 (SRJ & DCSR), K99 DA (MJF), T32 DA and F31 DA (ESC).
216 195 References: 1. Teter CJ, McCabe SE, LaGrange K, Cranford JA, Boyd CJ. Illicit use of specific prescription stimulants among college students: prevalence, motives, and routes of administration. Pharmacotherapy 2, (2006) 2. Frauger E, Moracchini C, Le Boisselier R, Braunstein D, Thirion X, Micallef J. OPPIDUM surveillance program: 20 years of information on drug abuse in France. Fundam Clin Pharmacol. In press (2013) 3. Ferris MJ, Calipari ES, Mateo Y, Melchior JR, Roberts DC, Jones SR. Cocaine selfadministration produces pharmacodynamic tolerance: differential effects on the potency of dopamine transporter blockers, releasers, and methylphenidate. Neuropsychopharmacology 37, (2012) 4. Madras BK, Miller GM, Fischman AJ. The dopamine transporter and attentiondeficit/hyperactivity disorder. Biol Psychiatry. 57, (2005) 5. Di Chiara G, Imperato A. Drugs abused by humans preferentially increase synaptic dopamine concentrations in the mesolimbic system of freely moving rats. Proc Natl Acad Sci USA 85, (1988) 6. Di Chiara G. Nucleus accumbens shell and core dopamine: differential role in behavior and addiction. Behav Brain Res 137, (2002) 7. Salahpour A. et al. Increased amphetamine-induced hyperactivity and reward in mice overexpressing the dopamine transporter. Proc Natl Acad Sci USA 105, (2008) 8. Calipari ES et al. Methylphenidate and cocaine self-administration produce distinct dopamine terminal alterations. Addict Biol. In Press (2012) 9. Biederman J, Wilens T, Mick E, Spencer T, Faraone SV. Pharmacotherapy of attention-deficit/hyperactivity disorder reduces risk for substance use disorder. Pediatrics 104, e20 (1999)
217 196 Supplementary Methodology: Animals: Male Sprague-Dawley rats ( g; Harlan Laboratories, Frederick, Maryland) were used for all self-administration experiments. DAT over-expressing mice were used for locomotor analysis and voltammetry. Animals were maintained according to the National Institutes of Health guidelines in Association for Assessment and Accreditation of Laboratory Animal Care accredited facilities on a 12:12 hour light-dark cycle with food and water ad libitum. The experimental protocol was approved by the Institutional Animal Care and Use Committee at Wake Forest School of Medicine. All animals were randomly assigned to experimental groups. There was no blinding done in the current study. Self-Administration: Rats were anesthetized and implanted with chronic indwelling jugular catheters. Following surgery, animals were singly housed, and all sessions took place in the home cage during the active/dark cycle (9:00 am 3:00 pm). All animals underwent a training paradigm where they were given access to a methylphenidate (MPH)-paired lever on a fixed ratio one (FR1) schedule. Training sessions were terminated after a maximum of 20 infusions or 6h, whichever occurred first. Acquisition occurred when an animal responded for 20 injections for two consecutive days. Typically, animals acquired a stable pattern of MPH SA within 1 to 5 days. A small percentage of animals did not acquire self-administration behavior, but this is typical of self-administration studies. We did not consider non-acquiring animals as being part of this study. Control animals for progressive ratio (PR) and threshold (TH) self-administration experiments underwent the training paradigm, and then were
218 197 immediately switched to PR or TH tests of reinforcing efficacy. We have confirmed that the control group (2x20 MPH) is not neurochemically different from naïve controls. Treatment animals underwent five consecutive days of MPH self-administration. Sessions were six hours long and were terminated after the animal responded for 40 injections of drug or at the end of the session. The dose of 0.56 mg/kg/injection MPH was chosen because it was the dose that produced maximal responding on a dose response curve measuring reinforcing efficacy. It is important to note that this method is different from previously described extended-access paradigms as animals do not escalate in total intake as the injection limit (40 inj per day) was set in order to eliminate variability in total consumption of cocaine. Progressive Ratio Procedure: Response requirements were increased through the following ratio progression: 1, 2, 4, 6, 9, 12, 15, 20, 25, 32, 40, 50, 62, 77, 95, 118, 145, etc. Breakpoints were defined as the number of completed ratios before 1 h elapsed without completion of the next ratio. Data from the MPH SA group was compared to a control group (2 days, 20 injections). Differences between groups cannot be due to differences in task learning or experience, as there was no difference in the cocaine TH task, indicating that there was not an overall increase in task performance with longer experience. Threshold Procedure: In order to determine the reinforcing efficacy of psychostimulants, subjects underwent a within-session threshold procedure. We switched from a progressive ratio procedure because the threshold procedure can assess both drug taking (seen in the early bins in the session where the dose is high) and drug seeking (seen near
219 198 the latter bins where the dose is low and the final bin where animals are responding for saline). Rats were given access to a descending series of 12 unit doses of cocaine, MPH, or AMPH (421, 237, 133, 75, 41, 24, 13, 7.5, 4.1, 2.4, 1.3, and 0μg/injection) on an FR1 schedule during consecutive 10 or 15-min bins within a daily session. Because MPH and AMPH are not metabolized as rapidly as cocaine, each bin was 15 minutes for those compounds, while it was 10 min for cocaine. The lever was not retracted at any time during the session. Doses were manipulated by holding the concentration constant and adjusting the pump duration. Oral Methylphenidate Administration: MPH was administered using an oral dosing procedure in which subjects are readily trained, in less than 3 days, to voluntarily consume MPH in a 5 ml volume of 10% sucrose. This procedure results in a pharmacokinetic profile of MPH distribution close to that observed in humans. MPH was administered twice daily (7:00 am and 2:00 pm) at a dose of 5 mg/kg for two weeks (14 days). This dosing regimen takes into account the fact that MPH metabolism is more rapid in rats than in humans and yields blood serum levels near the upper end of the therapeutic range observed in children (5-40 ng/ml). In Vitro Voltammetry: Fast scan cyclic voltammetry was used to characterize DA system kinetics, as well as the ability of psychostimulants to inhibit DA uptake in the NAc. In vitro voltammetry was chosen because it allows for the accurate assessment of psychostimulant potency by running within-subject concentration response curves for each drug. Voltammetry experiments were conducted during the dark phase of the light cycle, beginning 18 hours after commencement of the final SA or oral MPH session. A
220 199 carbon fiber electrode and a stimulating electrode were placed on the surface of the slice in close proximity to each other in the NAc. Endogenous DA release was evoked by a single electrical pulse (300 μa, 4 msec, monophasic) applied to the tissue every 5 minutes. Extracellular DA was recorded by applying a triangular waveform ( 0.4 to +1.2 V). Once the extracellular DA response was stable for three consecutive stimulations, blockers (cocaine, nomifensine), MPH and releasers [AMPH and methamphetamine (METH)] were applied to the brain slice. DA current was converted to concentration by electrode calibration with 3 μm DA at the end of each experiment. DEMON voltammetry software was used for analysis. To evaluate the effects of drugs, evoked levels of DA were modeled using Michaelis Menten kinetics, as a balance between release and uptake (Wightman et al, 1988). Michaelis Menten modeling provides parameters that describe the amount of DA released following stimulation, the maximal rate of DA uptake (V max ), and inhibition of the ability of DA to bind to the DAT, or apparent K m. For pre-drug modeling, we followed standard voltammetric modeling procedures by setting the apparent K m parameter to 160 nm based on the affinity of DA for the DAT, whereas baseline V max values were allowed to vary as the baseline measure of the rate of DA uptake. Following drug application, apparent K m was allowed to vary to account for changes in drug-induced DA uptake inhibition while the respective V max value determined for that subject at baseline was held constant. The apparent K m parameter models the amount of DA uptake inhibition following a particular dose of drug. Locomotor Analysis: Locomotor activity was assessed in automated locomotor activity monitors (MedAscociates). Mice were placed into the activity monitor chamber (20 cm
221 cm) for 60 min, then injected with saline or the drug in 0.1 ml total volume, returned to the chamber, and monitored for 60 min after injection. Locomotor activity was measured as horizontal distance covered as a percent of each animal s saline baseline. Dose response curves were run for each drug. Microdialysis: Animals were anesthetized with ketamine/xylazine, placed in a stereotaxic frame, and concentric microdialysis probes (2 mm (rat) or 1mm (mouse) membrane length; cut off 6000 Daltons; CMA-11, CMA/Microdialysis, Solna, Sweden) were implanted into the NAc core in both rats and mice the day before recording. Appropriate placement of probes was verified by histological examination after the experiments. For rats, microdialysis surgery was performed the day following the final session of MPH self-administration. Probes were perfused with artificial CSF at 1.0 µl/min. Samples were collected every 20 min and analyzed for DA by high-performance liquid chromatography (HPLC, Bionalytical Systems, Mt. Vernon, IN). Once stable baselines were established, the inlet line was switched to a syringe containing the same artificial CSF with AMPH (10µM), MPH (30µM), or cocaine (30µM). HPLC: A 2 50 mm (3 µm particle) reverse-phase column (Luna, Phenomenex, Torrance, CA) was used and the applied potential was +650 mv as referenced to an Ag/AgCl electrode. The mobile phase (75 mm NaH 2 PO 4, 1.7 mm 1-octanesulfonic acid sodium salt, 100 µl/l triethylamine, 25 µm EDTA, 10% acetonitrile v/v, ph=3.0) was pumped at a rate of 170 µl/min, with a detection limit for DA of 10 pm. DA quantification was achieved by comparing dialysate samples with DA standards of known concentration.
222 201 Statistics: Graph Pad Prism (version 5; La Jolla, CA) was used to analyze data sets and compose graphs. Baseline voltammetry, progressive-ratio, and P max data were compared across groups using a two-tailed Student s t-test. Data obtained from MPH SA/DAT-tg animals were subjected to a two-way analysis of variance with experimental group and concentration of drug as the factors. TH and locomotor data were subjected to a two-way analysis of variance with experimental group and dose as the factors. When significant interactions or main effects were obtained (p <.05), differences between groups were tested using Bonferroni post hoc tests. Parametric statistics were used for all analysis. Data met the assumptions of normality and the variance between groups was similar for all experiments. Power analyses were utilized for every measure, with expected effect sizes and standard deviations based on hundreds of previous experiments performed in the laboratory and from published manuscripts from experts in the field. Exclusion of data points only occurred if the points were significant outliers as determined by statistical analysis.
, a releaser, to inhibit the dopamine transporter (DAT) is enhanced following MPH self-administration.")
223 202 Supplementary Material: Supplementary Figure 1. Increased releaser, but not blocker, potency following methylphenidate (MPH) self-administration. A, B, the ability of methamphetamine (METH), a releaser, to inhibit the dopamine transporter (DAT) is enhanced following MPH self-administration. C, D, the potency of nomifensine, a DAT blocker, is unchanged following MPH self-administration. **p < 0.01, ***p < 0.001
 for both MPH and amphetamine (AMPH), but not cocaine (COC) in MPH SA animals. ***p < 0.")
224 203 Supplementary Figure 2. Methylphenidate (MPH) self-administration increases motivational effects of releasers and MPH, but not blockers. Panels A, B, C, demonstrate significantly higher maximal price paid (P max ) for both MPH and amphetamine (AMPH), but not cocaine (COC) in MPH SA animals. ***p < 0.001, **p < 0.01, *p < 0.05
225 204 Supplementary Figure 3. Increased releaser and methylphenidate (MPH), but not blocker-induced DA overflow following MPH self-administration and dopamine transporter (DAT) overexpression. A, B, C Group data showing enhanced MPH-, amphetamine (AMPH)-, but not cocaine (COC)-, induced DA overflow in response to a local drug infusion into the nucleus accumbens in MPH self-administration animals. C, D, E, Enhanced MPH-, amphetamine (AMPH)-, but not cocaine (COC)-, induced DA overflow in DAT-tg mice.
226 205 Supplementary Figure 4. Oral methylphenidate (MPH) administration does not affect DA kinetics or the potency of psychostimulants. A, B Group data from animals treated with oral MPH (5mg/kg, 2x daily; blue) as compared to controls (white), demonstrating no difference in dopamine system kinetics. Panels C, D, and E show the potency of cocaine, MPH, and amphetamine (AMPH) in controls (black) and animals that underwent oral MPH self-administration (blue).
227 206 Supplementary Discussion: 1. Methylphenidate (MPH) self-administration as a model of MPH abuse: MPH abuse occurs at rates comparable to cocaine abuse in the United States; however, there is a paucity of data outlining neurochemical consequences of MPH abuse. A survey of recent drug use indicates that 4.8 million people reported abusing MPH, which is similar to the 5.2 million for cocaine 1. When taken via the same route of administration, the subjective effects of MPH are indistinguishable from cocaine or AMPH, two commonly abused and highly addictive drugs 2-5. In order to determine the neurochemical consequences of MPH abuse, we used MPH self-administration to approximate human abuse. Traditional long-access paradigms of psychostimulant selfadministration, including MPH, result in escalation of intake over sessions, which have been suggested to model the switch from recreational use to abuse 6. Because of the fixed injection maximum, which we imposed to reduce variability in intake, animals do not escalate in total intake. This makes this paradigm different more traditional long-access models 6, however, first hour intake is increased, which is a hallmark of traditional escalation. Further, recent studies have demonstrated that the changes that occur in the DA system following traditional long-access paradigms and the current model (5 days, 40 inj/day) do not differ Changes in drug potency at the DAT are consistent with changes in drug-induced DA overflow in vivo. The reported increase in drug potency for DA uptake inhibition for MPH and releasers following MPH self-administration is accompanied by increases in druginduced DA overflow as measured using in vivo microdialysis. MPH (F (1, 5) = 20.80, p <
228 ; Supp Fig 3 A), and AMPH (F (1, 5) = 11.51, p < ; Supp Fig 3 B), but not cocaine (Supp Fig 3 C), resulted increased drug-induced DA overflow in the NAc. This confirms that the DAT inhibition changes correspond to similar changes in both DA levels and reward/reinforcement-related behaviors. We hypothesized that alterations in DAT levels/function could underlie the psychostimulant potency changes that we observed following MPH self-administration. Indeed, the changes seen in DAT-tg animals are similar to what was observed following MPH self-administration, where DAT-tg mice had increased MPH- (F (1, 5) = p < 0.001; Supp Fig 3 D) and AMPH- (F (1, 5) = 68.84, p < 0.001; Supp Fig 3 E), but not cocaine- (Supp Fig 3 F), induced DA overflow in the NAc. Here we demonstrate that increased MPH potency in the NAc core, increased MPH-induced DA overflow, and increased MPH-induced locomotion are associated with increased DAT levels. These changes are similar to AMPH potency changes and significantly different from cocaine potency changes, indicating that MPH shares some characteristics with releasers. 3. Oral Intake studies indicate that high (abused) and low (therapeutic) doses of MPH have different neurochemical consequences. Varying the dose and route of administration of a drug can produce variable effects, and here we demonstrate that oral administration of low, therapeutic doses and i.v. administration of high, abuse-relevant doses have completely different neurochemical consequences. Data from our lab is consistent with published work 8, 9, and here MPH (5 mg/kg; 2xday) delivered orally for a period of 14 days resulted in no changes in DA kinetics or psychostimulant potencies (Supp Fig 4). These data suggest that although MPH could enhance the reinforcing efficacy of MPH and releasers when taken via non-
229 208 oral routes, and in larger doses than prescribed, the therapeutic use of MPH, taken as prescribed, does not result in any changes in psychostimulant potencies. MPH has differential effects at high and low doses, which is likely due to the fact that low-dose MPH preferentially increases DA in the PFC, while higher doses increase DA in both the PFC and nucleus accumbens 10, 11. The selective increase in the PFC could be involved in the cognitive enhancing effects of MPH, but is likely independent of its reinforcing effects 12. The lack of effects of MPH in striatal regions at therapeutic doses could explain the lack of effects of oral MPH administration on DA terminals and psychostimulant potencies measured here. Together these data further highlight the dissociation between the effects of therapeutic and abused doses of MPH. Currently, there aren t consistent findings in regards to prior MPH exposure on the propensity to abuse cocaine These data do provide important information indicating that prior MPH exposure does not increase the reinforcing efficacy of cocaine at either abuse relevant, or therapeutic dosing levels. Although high dose MPH exposure does increase the potency of releaser compounds, this study suggests that therapeutic MPH use will not increase the likelihood of developing a cocaine abuse disorder. This is particularly relevant due to the large number of children that are prescribed MPH for therapeutic use. This is consistent with previous rodent work demonstrating that MPH administration during the peri-adolescent period does not influence cocaine reinforcement. Although this points to MPH being safe as it relates to cocaine abuse disorders, the propensity of individuals to have a substance use disorder may be significantly higher for other compounds.
230 MPH potency as it relates to blockers and releasers: In this study MPH effects seem to resemble those of DA releasers, even though it is traditionally categorized as a DAT blocker. A number of other studies have found similar effects following cocaine self-administration as well as in regards to MPH binding to the DAT It has been suggested that MPH can function as a releaser at high doses 24, 25. However, more recent studies using oocytes consistently classify MPH as a blocker 26. The distinction between blocker and releaser is important because the two classes of psychostimulants have very different mechanisms of action. Blockers bind to the DAT and inhibit DA uptake. Releasers have at least two main mechanisms: 1) they compete as a substrate for the DA binding site 2) are transported into the cell and into vesicles where they cause DA to be translocated into the cytoplasm and promote release via DAT-mediated reverse transport. MPH, an AMPH analog that is categorized as a DAT blocker, is unique since it is not transported into cells 26 but has been shown to promote reverse transport of DA at high doses 24, 25. These release capabilities, independent of being transported into the cell, could be due to a number of factors. 1) MPH binds to the DAT in a manner that is distinct from either prototypical blockers or releasers 20, 21, with significant overlap between the binding sites for both cocaine and AMPH. It is possible that although MPH is not transported into the cell that the interaction with the AMPH site results in conformational changes in the DAT which promote reverse transport in the same way as AMPH. 2) MPH has been demonstrated to result in the mobilization of vesicles into the releasable pool. It is possible that this effect of MPH is dependent on the DAT, thus when there are increased DAT levels, MPH is better able to induce exocytotic release from vesicles.
231 210 Although not a traditional releaser, MPH effects have been demonstrated previously to be similar to releasers. Ferris et al., (2012) demonstrated that cocaine selfadministration resulted in tolerance to the ability of all blockers to inhibit the DAT 22. However, MPH and all releasers tested were unaffected by cocaine self-administration. Although MPH is categorized with releasers in this study and following cocaine selfadministration, the effects of MPH self-administration are different from selfadministration of AMPH. AMPH self-administration resulted in no changes in drug potency 22. Here MPH self-administration causes neuroadaptations that are different from both cocaine and AMPH self-administration, further demonstrating that it is unique. The similarities of MPH to AMPH could be due to the unique functional properties of MPH. As mentioned previously, MPH has been shown to promote reverse transport of DA at high concentrations 24, 25, thus it is possible, although it is not transported into the cell, that MPH is releasing DA in a DAT-dependent fashion and its ability to increase DA is elevated when there are elevated transporter levels. 5. DAT cell surface expression as it relates to psychostimulant potency: To confirm that the changes in drug potency were due directly to changes in DAT levels, we correlated our measure of uptake inhibition (app. Km) with a functional measure of DAT levels (V max ). Correlation analysis of data from both MPH SA and DAT-tg show that DAT levels are positively correlated with potency measurements for AMPH (MPH SA: r = 0.73, p < 0.05; DAT-tg: r = 0.93, p < 0.001), MPH (MPH SA: r = 0.88, p < 0.001; DAT-tg: r = 0.92, p < 0.001), and cocaine (MPH SA: r = 0.54, ns; DAT-tg: r = 0.84, p < 0.01). Although there was a positive correlation between uptake rates and cocaine
232 211 potency, the shift in cocaine potency across increasing V max was substantially less robust than MPH and AMPH. Indeed, the slope of the regression line (cocaine, MPH SA: β = 1.54 ± 0.37; DAT-tg: β = ± 1.1) was significantly attenuated relative to both MPH (MPH SA: β = ± 1.59; DAT-tg: β = ± 1.467, vs. cocaine: p < ) and AMPH (β = 8.90 ± 1.38; DAT-tg: β = ± 3.716, vs. cocaine p < ), and not significantly different than a slope of zero for both MPH SA and DAT-tg animals. Additionally, the effect size for cocaine was so small that no difference in app. Km is observed between fast and slow V max based on a median split of V max values. A two-fold increase in uptake rates only resulted in 10 percent change in cocaine potency. MPH and AMPH potencies were increased with increasing DAT levels over a larger range, and the slopes of the regression lines did not significantly differ from one another. Additionally, a two-fold change in uptake rates resulted in a greater than 200 percent increase in measures of uptake inhibition. This further supports the idea that releaser compounds are affected in a different way from blockers, whereby increasing DAT levels results in increased neurochemical and behavioral potency. This is important to understand when trying to identify factors in the human population that may make certain individuals susceptible to psychostimulant abuse. This work uses a comprehensive approach to look at the effects of DAT levels on psychostimulant potency in neurochemistry and behavior, and highlights that current theories on DAT levels influencing blocker potency may need revision. The current consensus is that increases in DAT levels lead to a reduction in cocaine potency, and that lower DAT levels results in increased cocaine potency. This theory originates from two main sources. 1) cell culture work shows that DAT overexpression in cells results in a
233 212 decrease in cocaine potency with no change in AMPH potency 27. 2) Historically, the ability of cocaine to elevate DA in the NAc shell, where DAT levels are low, is much greater than that of the dorsal striatum, where DAT levels are high. Our results contradict this generally accepted view on psychostimulant potency. Here we demonstrate both behaviorally and neurochemically that increasing DAT levels in vivo does not change cocaine potency but does alter AMPH potency. Thus, we are proposing a novel idea by which cell surface expression dictates the potency of releasers, but not blockers. With respect to discrepancies with cell culture work, it is possible that our use of endogenous DA may produce different results than cell culture studies that use [ 3 H]-DA in order to measure uptake and uptake inhibition 27. It is possible that [ 3 H]-DA is sequestered into intracellular compartments differently than endogenous DA and that releaser compounds don t have access to this [ 3 H]-DA, thus any differences that may actually be present in release may not be observed. An important advantage of voltammetry is that it measures endogenous DA to look at uptake and release kinetics. Therefore, voltammetry provides a behaviorally relevant representation of the potency changes that occur after drug treatment. The increased potency of cocaine in the shell relative to the caudate has only been demonstrated in vivo with microdialysis, where a number of factors could be influencing the results in addition to the DAT, including different inhibitory autoreceptor function and afferent inputs to the regions. In fact, previous work using voltammetry in freelymoving animals has confirmed that brain regional variations in the potency of cocaine are due to D2 differences and not DAT levels 29.
234 213 Regardless of possible differences in cocaine potency due to other factors, we demonstrate that the changes observed using voltammetry are predictive of behavioral outcomes in regards to both locomotor activity and reinforcing efficacy. Previously, Salahpour et al. (2008) demonstrated that there was no difference in cocaine responses at 20mg/kg in DAT-tg mice. We also found no change in cocaine-induced locomotor activity over a full dose-response curve for cocaine in these mice 30. In addition, we show that DAT-tg animals have significantly greater locomotor responses to AMPH and MPH, which is supported by voltammetry data. Further, measures of drug-seeking and reinforcement in animals with elevated DAT levels corresponded to the neurochemical effects of these compounds where MPH and AMPH were enhances, and cocaine was unchanged. 6. D2 autoreceptors and DA uptake: One caveat to consider when determining the effects of uptake-inhibition by stimulants at the DAT is the role of D2 autoreceptors. D2 autoreceptors have been shown in numerous studies to modulate the function of the DAT, with D2 agonists increasing surface DAT expression and uptake rates, and D2 inhibition increases the potency of stimulants 31. It is possible that this is one mechanism driving the increased DAT levels following MPH self-administration. Sustained D2 activation over the six-hour selfadministration session due to MPH-induced DA elevations could lead to an increase in DAT surface expression that remains following the cessation of MPH self-administration. In addition to modulating baseline DA system kinetics, D2 autoreceptors have also been demonstrated to affect psychostimulant potencies 31. DA tone is absent in a slice preparation. Therefore, these changes may not have implications for blocker compounds
235 214 such as cocaine and MPH as their ability to increase synaptic DA levels is actionpotential dependent and only occurs following electrical stimulation of terminals. However it is possible, that the application of AMPH results in DA tone due to its ability to actively release DA from terminals independent of DA terminal stimulation. This release could activate D2 receptors and alter electrically-stimulated DA release. However, here we show a 267% increase in the ability of AMPH to inhibit DA uptake following MPH SA. Previous work has shown that complete removal of D2 receptors maximally increases uptake inhibition up to 48% 31. Thus, subsensitive D2 receptors following MPH SA cannot account for the large effect size demonstrated in the current study.
236 215 References 1. Substance Abuse and Mental Health Services Administration (SAMHSA) (2008) National Survey on Drug Use and Health. Retrieved at Accessed 15 May Parran TV Jr, Jasinski DR. Intravenous methylphenidate abuse. Prototype for prescription drug abuse. Arch Intern Med. 151, (1991) 3. Sherman CB, Hudson LD, Pierson DJ. Severe precocious emphysema in intravenous methylphenidate (Ritalin) abusers. Chest 92, (1987) 4. Rosen JB, Young AM, Beuthin FC, Louis-Ferdinand RT. Discriminative stimulus properties of amphetamine and other stimulants in lead-exposed and normal rats, Pharmacol, Biochem Behav 24, (1985) 5. Silverman PB, Ho BT. The discriminative stimulus properties of 2,5-dimethoxy-4- methylamphetamine (DOM): differentiation from amphetamine. Psychopharmacology (Berl) (1980) 6. Koob GF, Le Moal M. Drug abuse: hedonic homeostatic dysregulation. Science 278, 52-8 (1997) 7. Calipari ES, Ferris MJ, Zimmer BA, Roberts DCS, Jones SR. Temporal pattern of cocaine intake determines tolerance versus sensitization of cocaine effects at the dopamine transporter. Neuropsychopharm. In press (2013) 8. Gill KE, Pierre PJ, Daunais J, Bennett AJ, Martelle S, Gage HD, Swanson JM, Nader MA, Porrino LJ. Chronic Treatment with Extended Release Methylphenidate Does Not Alter Dopamine Systems or Increase Vulnerability for Cocaine Self- Administration: A Study in Nonhuman Primates. Neuropsychopharmacology in press (2012) 9. Humphreys KL, Eng T, Lee SS. Stimulant Medication and Substance Use Outcomes: A Meta-analysis. JAMA Psychiatry 29:1-9 (2013) 10. Berridge CW, Devilbiss DM, Andrzejewski ME, Arnsten AF, Kelley AE, Schmeichel B, Hamilton C, Spencer RC. Methylphenidate preferentially increases catecholamine neurotransmission within the prefrontal cortex at low doses that enhance cognitive function. Biol Psychiatry 60, (2006) 11. Devilbiss DM, Berridge CW. Low-dose methylphenidate actions on tonic and phasic locus coeruleus discharge. J Pharmacol Exp Ther 319, (2006) 12. Devilbiss DM, Berridge CW. Cognition-enhancing doses of methylphenidate preferentially increase prefrontal cortex neuronal responsiveness. Biol Psychiatry 64, (2008) 13. Wetzell B, Riley AL. Adolescent exposure to methylphenidate has no effect on the
237 216 aversive properties of cocaine in adulthood. Pharmacol Biochem Behav 101, (2012) 14. Warren BL, Iñiguez SD, Alcantara LF, Wright KN, Parise EM, Weakley SK, Bolaños-Guzmán CA. Juvenile administration of concomitant methylphenidate and fluoxetine alters behavioral reactivity to reward- and mood-related stimuli and disrupts ventral tegmental area gene expression in adulthood. J Neurosci 31, (2011) 15. Harvey RC, Sen S, Deaciuc A, Dwoskin LP, Kantak KM. Methylphenidate treatment in adolescent rats with an attention deficit/hyperactivity disorder phenotype: cocaine addiction vulnerability and dopamine transporter function. Neuropsychopharm 36, (2011) 16. Crawford CA, Baella SA, Farley CM, Herbert MS, Horn LR, Campbell RH, Zavala AR. Early methylphenidate exposure enhances cocaine self-administration but not cocaine-induced conditioned place preference in young adult rats. Psychopharm (Berl) 213, (2011) 17. Burton CL, Nobrega JN, Fletcher PJ. The effects of adolescent methylphenidate selfadministration on responding for a conditioned reward, amphetamine-induced locomotor activity, and neuronal activation. Psychopharm (Berl) 208, (2010) 18. Mague SD, Andersen SL, Carlezon WA Jr. Early developmental exposure to methylphenidate reduces cocaine-induced potentiation of brain stimulation reward in rats. Biol Psychiatry 57, (2005) 19. Schenk S, Izenwasser S. Pretreatment with methylphenidate sensitizes rats to the reinforcing effects of cocaine. Pharmacol Biochem Behav 72, (2002) 20. Wayment HK, Deutsch H, Schweri MM, Schenk JO. Effects of methylphenidate analogues on phenethylamine substrates for the striatal dopamine transporter: potential as amphetamine antagonists? J Neurochem 72, (1999) 21. Dar DE, Mayo C, Uhl GR. The interaction of methylphenidate and benztropine with the dopamine transporter is different than other substrates and ligands. Biochem Pharmacol. 70, (2005) 22. Ferris MJ, Mateo Y, Roberts DC, Jones SR. Cocaine-insensitive dopamine transporters with intact substrate transport produced by self-administration. Biol Psychiatry 69, (2011) 23. Ferris MJ, Calipari ES, Mateo Y, Melchior JR, Roberts DC, Jones SR. Cocaine selfadministration produces pharmacodynamic tolerance: differential effects on the potency of dopamine transporter blockers, releasers, and methylphenidate. Neuropsychopharmacology. 37, (2012)
238 Russell V, de Villiers A, Sagvolden T, Lamm M, Taljaard J. Differences between electrically-, ritalin- and D-amphetamine-stimulated release of [3H]dopamine from brain slices suggest impaired vesicular storage of dopamine in an animal model of Attention-Deficit Hyperactivity Disorder. Behav Brain Res. 94, (1998) 25. Dyck LE, Boulton AA, Jones RS. A comparison of the effects of methylphenidate and amphetamine on the simultaneous release of radiolabelled dopamine and p- or m- tyramine from rat striatal slices. Eur J Pharmacol. 68, (1980) 26. Sonders MS, Zhu SJ, Zahniser NR, Kavanaugh MP, Amara SG. Multiple ionic conductances of the human dopamine transporter: the actions of dopamine and psychostimulants. J Neurosci. 17, (1997) 27. Chen N, Reith ME. Substrates and inhibitors display different sensitivity to expression level of the dopamine transporter in heterologously expressing cells. J Neurochem 101, (2007) 28. Jones SR, Garris PA, Wightman RM. Different effects of cocaine and nomifensine on dopamine uptake in the caudate-putamen and nucleus accumbens. J Pharmacol Exp Ther. 274, (1995) 29. Aragona BJ, Cleaveland NA, Stuber GD, Day JJ, Carelli RM, Wightman RM. Preferential enhancement of dopamine transmission within the nucleus accumbens shell by cocaine is attributable to a direct increase in phasic dopamine release events. J Neurosci. 28, (2008) 30. Salahpour A. et al. Increased amphetamine-induced hyperactivity and reward in mice overexpressing the dopamine transporter. Proc Natl Acad Sci USA 105, (2008) 31. Benoit-Marand M, Ballion B, Borrelli E, Boraud T, Gonon F. Inhibition of dopamine uptake by D2 antagonists: an in vivo study. J Neurochem. 116, (2011)
239 218 CHAPTER VII UNIQUE PROFILE OF METHYLPHENIDATE-INDUCED DOPAMINE SYSTEM KINETICS: RELEASE PROFILE OF A BLOCKER AND UPTAKE PROFILE OF A RELEASER Erin S. Calipari, Mark J. Ferris and Sara. R. Jones The following manuscript is prepared for submission to PLoS ONE. Stylistic variations are due to the requirements of the journal. Erin S. Calipari performed the experiments and prepared the manuscript. Dr. Mark J. Ferris helped with statistical analyses and acted in an advisory capacity and Sara R. Jones acted in an advisory and editorial capacity.
240 219 Abstract: Although the dopamine transporter (DAT) is the main site of action of psychostimulant drugs, little work has focused on the effects of fluctuations in DAT levels on psychostimulant potency. Using fast scan cyclic voltammetry, we measured the effects of elevated DAT levels on pre-drug dopamine (DA) kinetics as well as the potency of the blockers (cocaine, MPH) and releasers [amphetamine (AMPH)] to inhibit DA uptake in the nucleus accumbens core (NAc). Here we find that increases in DAT levels, resulting from both transgenic over-expression (DAT-tg) and MPH self-administration, result in an increased maximal rate of uptake (V max ). V max a functional measure of DAT levels, is positively correlated with uptake inhibition for all drugs tested; however, AMPH and MPH were more affected by these changes, with a one-fold change in V max resulting in a 200% increase in potency as compared to a 20% increase in potency for cocaine. We demonstrate that V max and stimulated DA release are correlated in all groups; however, release did not correlate with uptake inhibition for any of the drugs, indicating that V max changes, and not DA release, are likely what are driving the changes in potency. The regression line for the effects of V max on MPH potency in both DAT-tg and MPH selfadministration groups was not significantly different from AMPH, but was significantly different from cocaine, indicating that MPH more closely resembled a releaser. Conversely, MPH-induced DA release more closely resembled that of cocaine, with increases in release over a dose-response curve for MPH, and was very different from the release profile of AMPH, which depleted terminals at high doses, indicating that MPH is not a releaser. This data indicates that although MPH is a DAT blocker it is affected by DAT changes in a similar manner to releasers.
241 220 Introduction: There has been some debate in the literature as to how the potency of psychostimulant compounds is altered by changes in dopamine transporter (DAT) levels (Chen and Reith, 2007; Salahpour et al., 2008; Yamamoto et al., 2013; Rao et al., 2013; Nelson et al., 2009). The dopamine transporter (DAT) is the main protein responsible for clearing DA from the synaptic cleft and terminating dopamine (DA) signaling (Hitri et al., 1994). In addition, it is the primary site of action for the euphorigenic, rewarding, and reinforcing properties of psychostimulant compounds such as cocaine, methylphenidate (MPH), and amphetamine (AMPH) (Chen et al., 2005, 2006; Tilley & Gu, 2008; Ritz et al., 1989). These drugs exert their rewarding and reinforcing effects via elevating DA in the nucleus accumbens (NAc), and they are categorized as either blockers or releasers (Di Chiara 1995; DiChiara and Imperato, 1988; Carboni et al, 1989; Woods and Meyer, 1991; Gerasimov et al, 2000). Blockers, like cocaine and MPH, act via inhibiting the ability of DAT to take up DA, thus augmenting post-synaptic DA signaling. Alternatively, releasers, like AMPH, act via two main mechanisms: first, they competitively inhibit DA uptake by competing for the DA binding site with DA, where they are transported into the cell. Second, they interact with vesicles to release DA into the cytoplasm where it can be actively released into the synapse via DAT-mediated reverse transport (Rothman and Bauman, 2003; Rudnick and Clark, 1993). There are conflicting theories as to how DAT levels affect psychostimulant potency, where cell culture studies demonstrate that increasing DAT levels reduces cocaine potency (Chen & Reith, 2008) and in vivo studies show that it has no affect on cocaine potency (Salahpour et al., 2008). Interestingly, both cell culture and in vivo reports demonstrate that DAT levels differentially affect the potency of blockers and releasers, although it is unclear as
242 221 to which class is more affected by the alterations. Here we determine the relationship between maximal rate of uptake (V max ), a functional measure of DAT levels, and the potency of AMPH, MPH, and cocaine to determine which class of drug is more affected by DAT levels and how specifically they are affected. Although MPH is characterized as a DAT blocker, a number of studies have demonstrated that it is unique in its interaction with the DAT, where it is distinct from both blockers and releasers (Wayment et al., 1999; Dar et al., 2005). Because of this, one aim of this study was to determine if DAT levels alter MPH potency in a similar fashion to blockers or releasers. MPH is categorized as a DAT blocker as studies have demonstrated that it is not actively transported into the cell, and thus, cannot directly interact with vesicles (Sonders et al., 1997). However, at higher concentrations, MPH has been shown to have release capabilities (Russel et al., 1998; Dyck et al., 1980). In addition, in binding experiments, MPH has been shown to interact with the DAT in a manner that is dissimilar from both blockers and releasers (Dar et al., 2005, Wayment et al., 1999). Current experiments using slice voltammetry have demonstrated that MPH is unique in the way in which it is affected by a prior cocaine self-administration history, where it classes with releasers, but not blockers (Ferris et al., 2012), and that the compensatory alterations that occur following MPH self-administration are distinct from both cocaine and AMPH (Calipari et al., 2013a, b; Ferris et al., 2011, 2012, 2013). Due to the unique properties of MPH as compared to other compounds of the same class, we aimed to determine if MPH was affected more similarly to releasers or blockers by altered DAT levels.
243 222 It has been demonstrated previously that MPH self-administration results in increased maximal rate of uptake (V max ), which is due to increases in DAT levels (Calipari et al., 2012). MPH self-administration also leads to an increase in AMPH and MPH, but not cocaine potency demonstrating that MPH self-administration affects the subsequent potency of drugs of abuse differently (Calipari et al., 2013a, b). Here we use correlations to determine the extent to which the increase in baseline V max could be responsible for the drug potency following MPH self-administration. In addition, we utilized DAT-tg mice, which have 4 additional copies of the DAT gene and exhibit increased DAT surface expression. The increased DAT levels lead to changes, similar to MPH self-administration, are characterized by increased MPH and AMPH, but not cocaine potency as measured by voltammetry, microdialysis, and locomotor behavior (Salahpour et al., 2008; Calipari et al., 2013a). The aim of the current study was to determine if blocker or releaser potency was positively correlated with functional measures of DAT levels following MPH self-administration or transgenic overexpression (DAT-tg). Additionally, we aimed to determine if MPH-induced uptake inhibition and MPH-induced DA release effects are affected in a way that is more similar to AMPH or cocaine. Method Animals: Male Sprague-Dawley rats ( g; Harlan Laboratories, Frederick, Maryland) were used for all self-administration experiments. Rats were maintained on a 12:12 hour reverse light-dark cycle (3:00 am lights off; 3:00 pm lights on) with food and water ad libitum. Transgenic DAT over-expressing mice were maintained on a 12:12
244 223 hour light cycle with food and water ad libitum. All animals were maintained according to the National Institutes of Health guidelines in Association for Assessment and Accreditation of Laboratory Animal Care accredited facilities. The experimental protocol was approved by the Institutional Animal Care and Use Committee at Wake Forest University School of Medicine. Self-Administration: Rats were anesthetized and implanted with chronic indwelling jugular catheters and trained for i.v. self-administration as previously described (Calipari et al., 2013b, c). Following surgery, animals were singly housed. Self-administration sessions took place in the home cage during the active/dark cycle (9:00 am 3:00 pm). Sessions were six hours in length and were terminated after the animal responded for 40 injections of drug or at the end of the six-hour session. Animals self-administered MPH (0.56 mg/kg/injection over 4 sec) on a fixed-ratio 1 schedule of administration (described previously, Calipari et al., 2013d). The dose of 0.56 mg/kg/injection MPH was chosen because it was the dose that produced maximal responding on a dose response curve measuring reinforcing efficacy (Marusich et al., 2010). For self-administering animals, acquisition (Day 1) was counted when the animal reaches 35 or more responses, and animals normally reached acquisition criteria in 0-3 days. Following establishment of stable responding, the animals were allowed to self-administer 40 injections per day for a period of 5 consecutive days. In Vitro Voltammetry: Fast scan cyclic voltammetry was used to characterize baseline DA system kinetics, and the ability of psychostimulants to inhibit DA uptake in the NAc. We selected the NAc core given dense innervations of DA nerve terminals and because it is a critical locus for the reinforcing and rewarding actions of cocaine. Further, our
245 224 previous research has concentrated on plasticity of DATs in both the core and shell and demonstrated similar results (Mateo et al., 2005; Ferris et al., 2011, 2012, 2013; Calipari et al., 2013b, c, d, e). Voltammetry experiments were conducted during the dark phase of the light cycle 18 hours after commencement of the final self-administration session in rats. Voltammetry in mice were conducted at the same time point, however, no previous procedures had been run. A vibrating tissue slicer was used to prepare 400 µm thick coronal brain sections containing the NAc core. The tissue was immersed in oxygenated artificial cerebrospinal fluid (acsf) containing (in mm): NaCl (126), KCl (2.5), NaH 2 PO 4 (1.2), CaCl 2 (2.4), MgCl 2 (1.2), NaHCO 3 (25), glucose (11), L-ascorbic acid (0.4) and ph adjusted to 7.4. Once sliced, the tissue was transferred to the testing chambers containing bath acsf (32 C), which flowed at 1 ml/min. After a 30-min equilibration period, a cylindrical carbon fiber microelectrode ( μm length, 7 μm radius) and a bipolar stimulating electrode were placed into the core of the NAc. Endogenous dopamine (DA) release was evoked by a single electrical pulse (300 μa, 4 msec, monophasic) applied to the tissue every 5 minutes. Extracellular DA was recorded by applying a triangular waveform 0.4 ( to +1.2 to 0.4V vs Ag/AgCl, 400 V/s) (Kennedy et al., 1992). Once the extracellular DA response was stable for three consecutive stimulations (within 10% variability), cocaine ( μmol/l), MPH ( μmol/l) and (AMPH) ( μmol/l) were applied cumulatively to the brain slice to determine the effects of MPH self-administration and transgenic over-expression on drug-induced uptake inhibition. Data Analysis: Demon Voltammetry and Analysis Software was used for data acquisition and analysis (Yorgason et al., 2011). To evaluate the effects of drugs, evoked
246 225 levels of DA were modeled using Michaelis Menten kinetics, as a balance between release and uptake (Wightman et al, 1988). Michaelis Menten modeling provides parameters that describe the amount of DA released following stimulation, the maximal rate of DA uptake (V max ), and alterations in the ability of DA to bind to the DAT, or apparent K m. For pre-drug modeling, we followed standard voltammetric modeling procedures by setting the baseline K m parameter to 160 nm based on the affinity of DA for the DAT (Wu et al., 2001), whereas V max values were allowed to vary as the pre-drug measure of the rate of DA uptake. Following drug application, apparent K m was allowed to vary to account for changes in drug-induced DA uptake inhibition while the respective V max value determined for that subject at baseline was held constant. The apparent K m parameter models the amount of DA uptake inhibition following a particular dose of drug. For cocaine, MPH, and AMPH dose-response curves, K m, a measure of apparent affinity for the DAT, was used to determine changes in ability of the psychostimulants to inhibit DA uptake in the NAc relative to baseline. Statistics: Graph Pad Prism (version 5, La Jolla, CA, USA) was used to statistically analyze data sets and create graphs. Data are presented as mean ± standard deviation and percentage unless otherwise stated. Correlational analyses were used to assess the association uptake rates (V max ) and release with cocaine, AMPH and MPH-induced DA uptake inhibition. Pearson s correlation coefficients were used to measure the strength of correlation between V max and release, V max and drug potency, and release and drug potency. Regression analysis was used to compare the extent to which V max altered drug
247 226 potency between MPH, cocaine and AMPH. All p values of < 0.05 were considered to be statistically significant. Results AMPH and MPH potency is similarly predicted by uptake rate in DAT-tg mice. Using voltammetry in slices from DAT-tg and wild type (WT) mice we determined DA kinetics by modeling stimulated DA release and uptake with Michaelis- Menten modeling. DAT-tg mice have been shown previously to have increased DAT expression and V max (Salahpour et al., 2008; Calipari et al., 2013a). To confirm that the changes in drug potency were due directly to changes in DAT levels, we correlated our measure of uptake inhibition (app. K m ) with a functional measure of DAT levels (maximal rate of uptake; V max ). Correlations were run with baseline V max and druginduced uptake inhibition for equivalent doses (based on DAT affinity) for MPH (30µM), cocaine (30µM), and AMPH (10µM). Because AMPH is more potent at inhibiting DA uptake, the lower dose was chosen to equate the extent of uptake inhibition between the three compounds tested. These doses were based on previous data from our lab in which full dose-response curves were run from each compound and compared to each other (John & Jones, 2007). Correlation analysis of data from DAT-tg show that DAT levels are positively correlated with potency measurements for AMPH (r = 0.93, p < 0.001), MPH (r = 0.92, p < 0.001), and cocaine (r = 0.84, p < 0.01). Although there was a positive correlation between uptake rates and cocaine potency, the shift in cocaine potency across increasing V max was substantially less robust than MPH and AMPH (Fig. 1 A).
248 227
249 228 Figure 1. Maximal rates of dopamine (DA) uptake (V max ) are positively correlated with psychostimulant potency. Dopamine transporter transgenic over-expressing mice (DAT-tg) were used in order to increase DAT levels. The functional measure of DAT levels, V max, was used to determine the effects of increased DAT on psychostimulant potency within subject. (A) Correlational analysis was run to compare the pre-drug measure of V max with the apparent K m (app. K m ) of DA uptake inhibition for methylphenidate (MPH; red; 30μM), amphetamine (AMPH; green; 10μM) and cocaine (COC; blue; 30μM). Differential doses were used to equate the potency of the drug at the DAT. V max and DAT levels were correlated for all drugs. (B) The regression lines of AMPH and MPH were compared to show that the range over which increases in V max increased drug potency was similar. Although MPH is a DAT blocker it resembles the profile of a releaser. (C) The extent to which V max was correlated with uptake inhibition was significantly different between MPH and COC.
250 229 In order to determine how MPH compared to a prototypical blocker (cocaine) and a prototypical releaser (AMPH), MPH was compared to the other compounds tested by comparing the slopes of the regression lines. The regression lines for MPH and AMPH were not significantly different (Fig. 1 B), indicating that the extent to which increased V max results in increased uptake inhibition for both MPH and AMPH does not differ. Alternatively, The slope of the regression line for cocaine (β = ± 1.1) was significantly attenuated relative to both MPH (β = ± 1.467, vs. cocaine: p < ) and AMPH (β = ± 3.716, vs. cocaine p < ), and not significantly different than a slope of zero for both MPH self-administration and DAT-tg animals (Fig. 1 C), as demonstrated by significantly different regression lines. This indicates that MPHinduced uptake inhibition is affected by baseline changes in V max in a fashion that is not significantly different from releasers (AMPH), but is significantly different from blocker compounds (cocaine) of the same class in DAT-tg mice. MPH self-administration-induced increases in DAT levels alter AMPH and MPH, but not cocaine potency. In order to determine if any increase in DAT levels/v max at baseline was capable of altering the potency of psychostimulant compounds we used MPH self-administration to elicit a drug-induced increase in V max /DAT levels. MPH self-administration has been demonstrated previously to result in increases in DAT levels and V max at baseline (Calipari et al, 2013d). Here we show that increasing DAT levels either by transgenic overexpression or by MPH self-administration results in similar effects (Fig. 2).
251 230
252 231 Figure 2. Increases in dopamine transporter (DAT) levels by methylphenidate (MPH) self-administration result in enhanced potency of releasers and MPH, but not blockers. MPH self-administration was used in order pharmacologically increase DAT levels. The functional measure of DAT levels, V max, was used to determine the effects of increased DAT on psychostimulant potency within subject. (A) Correlational analysis was run to compare the pre-drug measure of V max with the apparent K m (app. K m ) of DA uptake inhibition for methylphenidate (MPH; red; 30μM), amphetamine (AMPH; green; 10μM) and cocaine (COC; blue; 30μM). Differential doses were used to equate the potency of the drug at the DAT. V max and DAT levels were correlated for MPH and AMPH, but not COC. (B) The range over which increases in V max increased drug potency was similar for AMPH and MPH. Although MPH is a DAT blocker it resembles the profile of a releaser. (C) The extent to which V max was correlated with uptake inhibition was significantly different between MPH and COC.
253 232 Correlation analysis of data from MPH self-administration animals show that DAT levels are positively correlated with potency measurements for AMPH (r = 0.73, p < 0.05) and MPH (r = 0.88, p < 0.001), but not cocaine (r = 0.54, ns) potency (Fig. 2 A). In order to determine if MPH was more similar to AMPH or cocaine the slope of the regression line for MPH was compared to the regression lines for both cocaine and AMPH (Fig. 2 B, C). Similar to the data from DAT-tg mice, the regression lies for AMPH and MPH were not significantly different (Fig. 2 B), indicating that MPH and AMPH potencies are affected by baseline changes in DAT levels/v max to a similar extent. Conversely, the slope of the regression line for cocaine (β = 1.54 ± 0.37) was significantly attenuated relative to both MPH (β = ± 1.59, vs. cocaine: p < ) and AMPH (β = 8.90 ± 1.38, vs. cocaine p < ), and not significantly different than a slope of zero for MPH self-administration animals (Fig. 2C). This demonstrates that baseline DAT levels/v max are capable of altering the potency of psychostimulants, however, the extent to which uptake inhibition is affected by V max differs across drugs. V max and DA release are correlated at baseline, but DA release does not predict drug potency. In DAT-tg mice there is an increase in stimulated DA release in addition to increases in uptake rate; therefore, we aimed to determine if the drug effects were also correlated with pre-drug stimulated DA release. Determining if these two measures are dissociable allows for the determination that one of them alone is responsible for the overall K m effects. Stimulated DA release and V max were positively correlated in DAT-tg
254 233 Figure 3. Although maximal rate of dopamine uptake (V max ) and release are positively correlated in DAT-tg mice, dopamine (DA) release is not correlated with drug potency. (A) V max and pre-drug evoked DA release were positively correlated in all animals (B) Correlational analysis of the relationship between V max and evoked DA release in the methylphenidate (MPH; red), amphetamine (AMPH, green), and cocaine (COC, blue) groups. (C) There is no significant relationship for any of the drugs tested between pre-drug evoked DA release and uptake inhibition. (D) There is no significant relationship for any of the drugs tested between V max and evoked DA release in the presence of MPH (red; 30μM), AMPH (green, 10μM), or COC (blue; 30μM). Differential doses were used to equate the potency of the drug at the DAT. V max and DAT levels were correlated for MPH and AMPH, but not COC.
255 234 mice (r = 0.71, p < ) (Fig. 3 A). To demonstrate that pre-drug baseline differences across groups were not responsible for the differences in correlations across drugs, we split up the groups to demonstrate that stimulated DA release and uptake was correlated to the same extent and over approximately the same range before drug was added (Fig. 3 B). Following the determination of equivalent baseline measurements, we ran correlations to determine if baseline DA release, like V max, was correlated with the druginduced uptake inhibition with MPH, AMPH, and cocaine. We found that although V max and stimulated release are correlated at baseline, and that V max is correlated with druginduced uptake inhibition, there was not a significant correlation between baseline stimulated release and drug-induced uptake inhibition for any of the drugs (Fig. 3 C). In addition, V max is not correlated with post-drug stimulated release (Fig 3 D) indicating that the correlation between V max and stimulated release is only present at baseline and that drug-induced release is not affected by V max. In order to determine if these correlations held true following drug-induced DAT upregulation these correlations were run in control animals and animals that had a history of MPH self-administration. Not surprisingly, stimulated DA release and V max were positively correlated in both groups (r = 0.77, p < 0.001) (Fig. 4 A) with the MPH selfadministration group having significantly higher V max values overall. Congruent with the DAT-tg group, stimulated DA release and uptake were correlated to the same extent and over approximately the same range before drug was added in the MPH, AMPH, and cocaine groups (Fig. 4 B). To determine if baseline DA release, like V max, was correlated with the drug-induced uptake inhibition we correlated baseline DA release with MPH,
256 235 Figure 4. Dopamine release is not correlated with psychostimulant potency in methylphenidate (MPH) self-administration animals. (A) V max and pre-drug evoked DA release were positively correlated in all animals (B) Correlational analysis of the relationship between V max and evoked DA release in the methylphenidate (MPH; red), amphetamine (AMPH, green), and cocaine (COC, blue) groups. (C) There is no significant relationship for any of the drugs tested between pre-drug evoked DA release and uptake inhibition. (D) There is no significant relationship for any of the drugs tested between V max and evoked DA release in the presence of MPH (red; 30μM), AMPH (green, 10μM), or COC (blue; 30μM). Differential doses were used to equate the potency of the drug at the DAT. V max and DAT levels were correlated for MPH and AMPH, but not COC.
257 236 AMPH, and cocaine-induced uptake inhibition. We show that although V max and stimulated release are correlated at baseline, and V max and uptake inhibition are correlated, baseline stimulated release is not significantly correlated with uptake inhibition of any of the drugs tested (Fig 4 C). V max is not correlated with post-drug stimulated release (Fig 4 D) indicating that the correlation between V max and stimulated release is only present at baseline. Stimulated DA release and uptake inhibition are not correlated for any drugs. In addition, to further confirm that stimulated release, and the peak height of the signal, does not affect the uptake-inhibition induced by drugs, we correlated stimulated DA release in the presence of drug with drug-induced uptake inhibition in both the MPH self-administration (Fig. 5 A) and DAT-tg (Fig. 5 B) groups. We found that stimulated release after drug did not correlate with uptake inhibition for any of the drugs tested, indicating that release, and the peak height of the signal, is not a significant factor that influences uptake inhibition (Fig. 5). Increasing V max results in reduced AMPH-induced stimulated release; however, the release profile of MPH resembles cocaine. Because MPH had the uptake inhibition profile of a releaser where its potency was changed to a similar extend in regards to V max changes, but MPH is traditionally categorized as blocker, we analyzed the release profile of MPH as compared to cocaine and AMPH in our two animal models. Not surprisingly, in groups that have significantly higher V max, which is likely mediated by increased DAT levels, AMPH peak height is
258 237 Figure 5. Drug-induced dopamine release does not affect dopamine uptake inhibition following methylphenidate (MPH) self-administration or DAT overexpression. Correlational analysis was run in order to confirm that evoked DA release was not influencing drug induced uptake inhibition. Evoked DA release in the presence of drug was not correlated with uptake inhibition for any of the drugs tested (A) following MPH self-administration or (B) in transgenic DAT over-expressing mice (DAT-tg).
259 238
260 239 Figure 6. Methylphenidate (MPH) has the release profile of a blocker. To determine if MPH, a blocker, had the release profile of a blocker or releaser, full concentrationresponse curves for MPH, amphetamine (AMPH) and cocaine were run in DAT overexpressing mice (DAT-tg; left) and following MPH self-administration (right). (A) AMPH, a prototypical DA releaser results in reductions in DA release over a concentration-response curve for the compound. The AMPH-induced reductions in release were greater in DAT-tg mice. (B) Cocaine, a prototypical DAT blocker, results in an inverted U shape release curve. Release was not different between wild type and DAT-tg mice. (C) MPH has the release profile of a blocker. Evoked release in the presence of MPH was unchanged between groups. (D) Following MPH selfadministration AMPH-induced reductions in release were exacerbated. (E) Evoked DA release in the presence of cocaine was unchanged between MPH self-administration and control groups. (F) The release profile of MPH resembles cocaine and is unchanged following MPH self-administration.
261 240 significantly reduced (Fig. 6 A, D). ANOVA revealed a main effect of group on stimulated DA release over a dose-response curve for both DAT-tg (F 1,40 = 14.79, p < 0.001) (Fig. 6 A) and MPH self-administration (F 1,39 = 16.32, p < 0.001) (Fig. 6 D) groups, indicating that AMPH was more potent at reducing DA release in animals that had elevated DAT levels. Because cocaine is a blocker, and as such does not actively release DA from terminals, we expected no depletion of stimulated DA signals as with AMPH. The pattern of stimulated DA release across a dose-response curve for cocaine was not significantly different between DAT-tg and tintype mice (Fig. 6 B). The same was true for MPH SA with no significant difference being present between stimulated DA release across a cocaine dose-response curve (Fig. 6 E). In addition the pattern of stimulated release with cocaine present was an inverted U shaped curve, with the downward portion of the curve likely being due to prolonged autoreceptor activation, which leads to reduced release. This pattern was very different from the releaser AMPH, highlighting the distinction between the two compounds mechanisms of action. In order to compare MPH to the other compounds tested we assessed stimulated DA release in the presence of a MPH dose-response curve in both DAT-tg and MPH selfadministration animals (Fig. 6 C, F). We found that there was no significant difference between DAT-tg and wild type animals in stimulated release effects of MPH (Fig. 6 C). In addition, there was also no difference between MPH self-administration and control animals in stimulated DA release in the presence of MPH (Fig. 6 F). This indicates in two different models that changes in V max as seen with both DAT-tg and MPH selfadministration that increases in DAT levels do not affect stimulated release. In addition,
262 241 the release profile of MPH more closely resembles the release profile of cocaine, another transporter blocker, which exhibit an inverted U shaped release curve. Although MPH potency as measured by apparent K m is affected by transporter level in a manner more similar to releasers, its release profile resembles that of blockers. Discussion Here we show that although MPH is traditionally characterized as a DAT blocker, its mechanism is more complicated, having characteristics of both blockers and releasers. MPH is affected by changes in V max in a similar manner to releasers, whereby increases in V max result in large increases in MPH potency. Conversely, although MPH potency is affected by V max, more similarly to releasers, the release profile, as measured by stimulated release across a dose-response curve of the compound, more closely mimics that of the blocker cocaine, indicating that MPH is not actually a releaser. In addition, although V max and stimulated DA release are correlated at baseline, they are not correlated with the MPH effects, indicating that the factor that is actually driving the differences in drug potency is the uptake at baseline, which is controlled by DAT levels. These data indicate that MPH, a drug commonly used in the clinic, is a unique compound that has characteristics of both blockers and releasers and its potency can be affected purely by DAT level changes at baseline. MPH is characterized as a DAT blocker because it is not actively transported into cells, thus it cannot interact with vesicles and actively release DA (Sonders et al., 1997). Because of this, much of the MPH literature has focused on MPH as a blocker and has considered it to be similar to cocaine in both acute and chronic pharmacological effects.
263 242 Here we show that MPH resembles a releaser in its acute uptake inhibition profile where its potency is affected by V max changes and a blocker in its release profile. Presently, it is unclear as to how MPH can be affected by DAT levels in a manner similar to releasers, but not release from vesicles. One possible explanation is that the effects are related to the binding site on the DAT, where MPH and AMPH have been shown to share some similarities (Dar et al., 2005; Wayment et al., 1999). Another possible explanation is that MPH has the ability to release DA at high concentrations, which has been seen in cell culture studies, thus making it a blocker at lower concentrations and a releaser at higher concentrations (Russel et al., 1998; Dyck et al., 1980). Additionally, it is possible that MPH-DAT interactions signal second messenger cascades that promote shifts in vesicles from membrane-bound to releasable pools, thus increasing MPH s effects (Chadchankar et al., 2012; Volz et al., 2007, 2009). MPH has been previously been shown to mobilize vesicles, the mechanism of which is not clear, but could be through its interaction with the DAT, thus leading to increased MPH effects in situations where there are increased DAT levels (Chadchankar et al., 2012; Volz et al., 2007, 2009). Although it is unclear as to how transporter levels affect MPH potency, the fact that this phenomenon occurs suggests that care should be taken when administering this compound to individuals that may have differences in DAT levels, such as ADHD sufferers (Madras et al., 2005), as the potency/efficacy of this drug may be increased in these individuals. Although MPH potency is affected by DAT levels in a similar manner to releasers, its release profile demonstrated that it is not a pure releaser. In the groups that had increased V max the releaser AMPH was more efficacious at reducing peak height, due to its ability to deplete vesicles, an effect not seen with MPH. The reduction in peak
264 243 height in the MPH self-administration and DAT-tg groups is likely due to an increase in AMPH-induced DA efflux and depletion of the terminals leading to reduced stimulated release (Jones et al., 1998). This is not surprising as a number of studies have demonstrated that releasers, like AMPH, are more neurotoxic in scenarios where DAT function/levels are elevated, indicating that these compounds are likely more potent (Xie et al., 2000; Fumagalli et al., 1998). Here we show that stimulated DA release in the presence of AMPH is significantly reduced in two animal models that have increases in DAT levels at baseline. It has been demonstrated that the depleting actions of AMPH are dependent on the presence of the DAT (Jones et al., 1998), and it is likely that AMPH is better able to release DA into the synapse when there are more transporters present, leading to the enhanced depletion of terminals observed in the present study. Because MPH is not transported into cells, it cannot release actively from terminals, an effect that is highlighted by the increase, not decrease, in DA release when MPH is applied to the brain slice. This indicated that although MPH potency at the DAT is increased in a similar way to releasers, it is not functioning as a releaser and cannot deplete DA from terminals. This would also indicate that even if MPH is capable of releasing from terminals, it is far less potent than AMPH as highlighted by the ability of AMPH to reduce release with MPH having no effect. One important factor to consider is the contribution of stimulated DA release, or the peak height of the DA signal, on drug-induced uptake inhibition. It is difficult to dissociate the contribution of differences in pre-drug DA release or uptake on the potency of drug. Release and uptake are often correlated, but have been shown to be dissociated in a number of studies (Mateo et al., 2005; Calipari et al., 2013e), demonstrating that the
265 244 correlation between the two measures is physiological. Here we extensively analyzed the extent to which V max and release are correlated and demonstrated that although release and V max are related, that V max is the primary factor dictating changes in psychostimulant potencies. Further, differences in evoked DA release in the presence of drug could potentially influence potency measurements. This is particularly important in the case of AMPH, which affects the peak-height of the DA signal differently across groups. In the AMPH groups there were significant differences between MPH self-administration and control animals as well as between DAT-tg and wild type animals. To demonstrate that the peak height of the signal has no effect on the measured uptake inhibition correlations were run with post-drug stimulated release and uptake inhibition, which revealed that the two measures were not correlated. Additionally, we determined that the reduction in peak height in the MPH and DAT-tg groups did not significantly contribute to the measures of uptake inhibition, indicating that the reduction in release was a separate phenomenon from the potency of AMPH at the DAT. This also was true for the other groups tested, although it was not as pertinent, as there was no significant different in release between groups following MPH or cocaine. These results are particularly important as they contradict the cell-culture theories on psychostimulant potency; however, differences between the current literature and this study can be accounted for by differences between cell culture and vivo models. We are proposing a theory by which cell surface expression dictates the potency of releasers in an opposite direction from which would be supported by in vitro cell studies. Cell culture studies have demonstrated that cocaine, but not AMPH potency, is altered by increasing DAT levels. The differences between cell studies and this particular study are numerous.
266 245 Cell studies often use [ 3 H]-DA in order to look at uptake and uptake inhibition (Chen & Reith, 2008). It is possible that [ 3 H]-DA is sequestered into intracellular compartments differently than endogenous DA and that releaser compounds do not have access to this [ 3 H]-DA, thus any differences that may actually be present in release may not be observed. An important advantage of FSCV is that it uses endogenous DA to look at kinetics and functioning of the system, which may better elucidate potency changes that occur after drug treatment or in transgenic animal models. In addition, the behavioral data supports enhanced releaser potency following DAT over-expression, which gives greater support to this neurochemical model (Calipari et al., 2013a, Salahpour et al., 2008). Because the DA system is responsible for mediating the reward and reinforcement of drugs, it is important to know how the behavioral and neurochemical effects of psychostimulants are altered by differences in DA system functioning. In addition, because MPH is uniquely affected by DA changes that affect releasers, but still has the release profile of a blocker, it makes it difficult to make predictions about MPH use and abuse without conducting studies on the compound itself. This data demonstrates 1.) the positive relationship between DAT levels and psychostimulant releaser, but not blocker potency, and 2.) that predictions on the effects of MPH based on previous work using similar compounds may not give an accurate representation of what is actually happening in vivo. Acknowledgements: This work was funded by NIH grants R01DA024095, RO1DA T32DA and F31 DA (ESC).
267 246 References: Calipari ES, Ferris MJ, Salahpour A, Caron MG, Roberts DCS, Jones SR (2013a) Methylphenidate amplifies the potency and reinforcing effects of amphetamines by increasing dopamine transporter expression. Nature Communications. In press. Calipari ES, Beveridge TJ, Jones SR, Porrino LJ (2013b) Withdrawal from extended access cocaine self-administration results in dysregulated functional activity and altered locomotor activity in rats. European J. Neurosci. In press. Calipari ES, Ferris MJ, Jones SR (2012c) Extended Access Cocaine Self-Administration Results in Tolerance to the Dopamine-Elevating and Locomotor-Stimulating Effects of Cocaine. J. Neurochem. In press. Calipari ES, Ferris MJ, Melchior JR, Bermejo K, Salahpour A, Roberts DC et al (2012d). Methylphenidate and cocaine self-administration produce distinct dopamine terminal alterations. Addict Biol. In press. Calipari ES, Ferris MJ, Zimmer BA, Roberts DC, Jones SR. (2013e) Temporal Pattern of Cocaine Intake Determines Tolerance vs Sensitization of Cocaine Effects at the Dopamine Transporter. Neuropsychopharmacology. In press. Carboni E, Imperato A, Perezzani L, Di Chiara G (1989) Amphetamine, cocaine, phencyclidine and nomifensine increase extracellular dopamine concentrations preferentially in the nucleus accumbens of freely moving rats. Neuroscience 28(3): Chadchankar H, Ihalainen J, Tanila H, Yavich L. (2012) Methylphenidate modifies overflow and presynaptic compartmentalization of dopamine via an α-synucleindependent mechanism. J Pharmacol Exp Ther. 341(2): Chen N, Reith ME. (2007) Substrates and inhibitors display different sensitivity to expression level of the dopamine transporter in heterologously expressing cells. J Neurochem Apr;101(2): Epub 2007 Jan 22. Chen R, Han DD, Gu HH (2005) A triple mutation in the second transmembrane domain of mouse dopamine transporter markedly decreases sensitivity to cocaine and methylphenidate. J Neurochem 94(2): Chen R, Tilley MR, Wei H, Zhou F, Zhou FM, Ching S, Quan N, Stephens RL, Hill ER, Nottoli T, Han DD, Gu HH. (2006) Abolished cocaine reward in mice with a cocaineinsensitive dopamine transporter. Proc Natl Acad Sci U S A. 103(24): Dar DE, Mayo C, Uhl GR. (2005) The interaction of methylphenidate and benztropine with the dopamine transporter is different than other substrates and ligands. Biochem Pharmacol. 70,
268 247 Di Chiara G (1995) The role of dopamine in drug abuse viewed from the perspective of its role in motivation. Drug Alcohol Depend 38(2): Di Chiara G, Imperato A (1988) Drugs abused by humans preferentially increase synaptic dopamine concentrations in the mesolimbic system of freely moving rats. Proc Natl Acad Sci U S A 85(14): Dyck LE, Boulton AA, Jones RS. (1980) A comparison of the effects of methylphenidate and amphetamine on the simultaneous release of radiolabelled dopamine and p- or m- tyramine from rat striatal slices. Eur J Pharmacol. 68(1): Ferris MJ, Calipari ES, Mateo Y, Melchior JR, Roberts DC, Jones SR (2012). Cocaine self-administration produces pharmacodynamic tolerance: differential effects on the potency of dopamine transporter blockers, releasers, and methylphenidate. Neuropsychopharmacology 37: Ferris MJ, Calipari ES, Melchior JR, Roberts DC, España RA, Jones SR. (2013) Paradoxical tolerance to cocaine after initial supersensitivity in drug-use-prone animals. Eur J Neurosci. In press. Ferris MJ, Mateo Y, Roberts DC, Jones SR (2011) Cocaine-insensitive dopamine transporters with intact substrate transport produced by self-administration. Biol Psychiatry 69(3): Fumagalli F, Gainetdinov RR, Valenzano KJ, Caron MG. (1998) Role of dopamine transporter in methamphetamine-induced neurotoxicity: evidence from mice lacking the transporter. J Neurosci. 18(13): Gerasimov MR, Franceschi M, Volkow ND, Gifford A, Gatley SJ, Marsteller D, Molina PE, Dewey SL (2000) Comparison between intraperitoneal and oral methylphenidate administration: A microdialysis and locomotor activity study. J Pharmacol Exp Ther. 295(1):51-7. Hitri A, Hurd YL, Wyatt RJ, Deutsch SI. (1994) Molecular, functional and biochemical characteristics of the dopamine transporter: regional differences and clinical relevance. Clin Neuropharmacol. 17(1):1-22. John CE, Jones SR. (2007) Voltammetric characterization of the effect of monoamine uptake inhibitors and releasers on dopamine and serotonin uptake in mouse caudateputamen and substantia nigra slices. Neuropharmacology. 52(8): Jones SR, Gainetdinov RR, Wightman RM, Caron MG. (1998) Mechanisms of amphetamine action revealed in mice lacking the dopamine transporter. J Neurosci. 18(6): Kennedy RT, Jones SR, Wightman RM (1992). Dynamic observation of dopamine autoreceptor effects in rat striatal slices. J Neurochem 59:
269 248 Madras BK, Miller GM, Fischman AJ. (2005) The dopamine transporter and attentiondeficit/hyperactivity disorder. Biol Psychiatry. 57, Marusich JA, Beckmann JS, Gipson CD, Bardo MT (2010) Methylphenidate as a reinforcer for rats: contingent delivery and intake escalation. Exp Clin Psychopharmacol 18(3): Mateo Y, Lack CM, Morgan D, Roberts DC, Jones SR (2005) Reduced dopamine terminal function and insensitivity to cocaine following cocaine binge self-administration and deprivation. Neuropsychopharm 30(8): Nelson AM, Larson GA, Zahniser NR. (2009) Low or high cocaine responding rats differ in striatal extracellular dopamine levels and dopamine transporter number. J Pharmacol Exp Ther. 331(3): Rao A, Sorkin A, Zahniser NR. (2013) Mice expressing markedly reduced striatal dopamine transporters exhibit increased locomotor activity, dopamine uptake turnover rate, and cocaine responsiveness. Synapse. In press. Ritz MC, Lamb RJ, Goldberg SR, Kuhar MJ (1987). Cocaine receptors on dopamine transporters are related to self-administration of cocaine. Science 237: Rothman RB, Baumann MH. (2003) Monoamine transporters and psychostimulant drugs. Eur J Pharmacol. 479(1-3): Rudnick G, Clark J. (1993) From synapse to vesicle: the reuptake and storage of biogenic amine neurotransmitters. Biochim Biophys Acta. 1144(3): Russell V, de Villiers A, Sagvolden T, Lamm M, Taljaard J (1998) Differences between electrically-, ritalin- and D-amphetamine-stimulated release of [3H]dopamine from brain slices suggest impaired vesicular storage of dopamine in an animal model of Attention- Deficit Hyperactivity Disorder. Behav Brain Res. 94(1): Salahpour A, Ramsey AJ, Medvedev IO, Kile B, Sotnikova TD, Holmstrand E, Ghisi V, Nicholls PJ, Wong L, Murphy K, Sesack SR, Wightman RM, Gainetdinov RR, Caron MG. (2008) Increased amphetamine-induced hyperactivity and reward in mice overexpressing the dopamine transporter. Proc Natl Acad Sci U S A (11): Sonders MS, Zhu SJ, Zahniser NR, Kavanaugh MP, Amara SG (1997) Multiple ionic conductances of the human dopamine transporter: the actions of dopamine and psychostimulants. J Neurosci 17(3): Tilley MR, Gu HH (2008) The effects of methylphenidate on knockin mice with a methylphenidate-resistant dopamine transporter. J Pharmacol Exp Ther. 327(2): Volz TJ, Farnsworth SJ, Hanson GR, Fleckenstein AE. (2009) Methylphenidate-Induced Alterations in Synaptic Vesicle Trafficking and Activity: Functional Consequences and Therapeutic Implications. Ann N Y Acad Sci. 1139:
270 249 Volz TJ, Farnsworth SJ, King JL, Riddle EL, Hanson GR, Fleckenstein AE. (2007) Methylphenidate administration alters vesicular monoamine transporter-2 function in cytoplasmic and membrane-associated vesicles. J Pharmacol Exp Ther. 323(2): Wayment HK, Deutsch H, Schweri MM, Schenk JO. (1999) Effects of methylphenidate analogues on phenethylamine substrates for the striatal dopamine transporter: potential as amphetamine antagonists? J Neurochem. 72, Wightman RM, Amatore C, Engstrom RC, Hale PD, Kristensen EW, Kuhr WG et al (1988). Real-time characterization of dopamine overflow and uptake in the rat striatum. Neuroscience 25: Woods SK, Meyer JS (1991) Exogenous tyrosine potentiates the methylphenidateinduced increase in extracellular dopamine in the nucleus accumbens: a microdialysis study. Brain Res. 560(1-2): Wu Q, Reith MEA, Wightman RM, Kawagoe KT, Garris PA (2001). Determination of release and uptake parameters from electrically evoked dopamine dynamics measured by real-time voltammetry. J Neurosci Methods 112: Xie T, McCann UD, Kim S, Yuan J, Ricaurte GA. (2000) Effect of temperature on dopamine transporter function and intracellular accumulation of methamphetamine: implications for methamphetamine-induced dopaminergic neurotoxicity. J Neurosci. 20(20): Yamamoto DJ, Nelson AM, Mandt BH, Larson GA, Rorabaugh JM, Ng CM, Barcomb KM, Richards TL, Allen RM, Zahniser NR. (2013) Rats classified as low or high cocaine locomotor responders: A unique model involving striatal dopamine transporters that predicts cocaine addiction-like behaviors. Neurosci Biobehav Rev. 37(8): Yorgason JT, España RA, Jones SR (2011) Demon voltammetry and analysis software: analysis of cocaine-induced alterations in dopamine signaling using multiple kinetic measures. J Neurosci Methods. 202(2):
271 250 CHAPTER VIII INTERMITTENT ACCESS METHYLPHENIDATE SELF-ADMINISTRATION RESULTS IN A SENSITIZED DOPAMINE SYSTEM Erin S. Calipari and Sara. R. Jones The following manuscript is prepared for submission at Neuropharmacology. Stylistic variations are due to the requirements of the journal. Erin S. Calipari performed the experiments and prepared the manuscript and Dr. Sara R. Jones acted in an advisory and editorial capacity.
272 251 Abstract: Although methylphenidate (MPH), the active compound in Ritalin, is categorized as a dopamine transporter (DAT) blocker, similar to cocaine, the effects of MPH, both acute and following repeated administration, are unique. Previous work has demonstrated that MPH classes with releasers following MPH self-administration, where extended-access self-administration results in enhanced MPH and releaser, but not blocker potency at the DAT. Extended-access self-administration results in escalation of MPH intake, an effect that has been suggested to mediate the neurochemical changes that occur following extended-access self-administration. Here we used intermittent access of MPH (0.56 mg/kg; 14 days), which results in consistent MPH intake over sessions, in order to determine if escalation is necessary to cause the increased MPH and releaser potencies at the DAT. Further, recently, it was determined that the temporal pattern of cocaine selfadministration dictates the DA system changes that occur, with extended-access resulting in tolerance to cocaine s effects and intermittent access resulting in sensitization. Thus, we aimed to determine if the compensatory changes that occur following MPH selfadministration are dependent on the temporal pattern of intake. Intermittent access MPH self-administration resulted in enhanced releaser and MPH potency, but no change in cocaine potency, demonstrating that the effects are not due to escalation, and, unlike cocaine, MPH effects are not affected by pattern of intake. Further, although MPH s effects are enhanced similar to releasers following MPH self-administration, its release profile more similarly resembles a blocker, whereby MPH increases release and does not deplete DA terminals. Taken together, this data demonstrates that MPH exhibits properties of both blockers and releasers, and that the compensatory effects of MPH may increase the abuse liability of amphetamines independent of pattern on administration.
273 252 Introduction: Methylphenidate (MPH), the active compound in Ritalin, is commonly diverted for non-medical use, with up to 17% of college students reporting abusing MPH for its cognitive enhancing effects or euphoric effects to get high (Teter et al., 2006). MPH is taken orally, intranasally or intravenously (IV), and is one of the prescribed drugs most diverted onto the illicit market worldwide (Frauger et al., 2013); however, little work has focused on understanding the acute actions of MPH or determining the neurochemical consequences of MPH abuse. A survey of drug use indicates that 4.8 million people reported abusing MPH, which is similar to the 5.2 million for cocaine (SAMHSA, 2008). Even more alarming, when taken via the same route of administration, the subjective effects of MPH are indistinguishable from cocaine or AMPH, two commonly abused and highly addictive drugs (Rosen et al., 1985; Silverman & Ho, 1980). Further, a number of case studies demonstrate numerous instances of IV MPH administration in the human population, which is alarming given the paucity of empirical data on the behavioral and neurochemical consequences of IV administration of the compound (Sherman, Hudson, and Pierson, 1987; Parran &Jasinski, 1991). MPH, like other stimulants, exerts its rewarding and reinforcing effects by inhibiting the dopamine transporter (DAT), which leads to elevation in dopamine (DA) in the nucleus accumbens (NAc) (Tilley & Gu, 2008). As a result, much work has focused on the compensatory effects that occur within the DA system following MPH selfadministration, as they are predictive of changes in the rewarding and reinforcing potential of psychostimulants (Calipari et al., 2013a). Previous work has used extended access MPH self-administration demonstrating that abuse-relevant MPH exposure results
274 253 in enhanced MPH and releaser, but not blocker potency and reinforcing efficacy (Calipari et al., 2013a, b). Work using fixed-intake extended access paradigms has also showed escalation in first-hour intake and rate of intake over sessions (Calipari et al., 2013 a, b), an effect that has been suggested to model the switch from recreational use to abuse (Koob & Le Moal, 1997). Currently, all of the neurochemical studies on MPH selfadministration use paradigms that result in escalation of intake, making it unclear as to whether the effects are due to escalation or MPH administration. Here we use intermittent access (Calipari et al., 2013c; Zimmer et al., 2012), which does not result in escalation, to determine if administration of the compound independent of temporal pattern of intake is responsible for the compensatory changes that occur following MPH self-administration. Additionally, it was recently demonstrated that the compensatory effects that occur with cocaine at the DAT are different when cocaine is administered intermittently versus continuously, with sensitization and tolerance occurring, respectively (Calipari et al., 2013 c). Thus, we also aimed to determine how intermittent MPH administration influenced the compensatory changes that occur within the DA system as it compares to previous work with extended-access conditions. The compensatory effects of MPH self-administration as they relate to the DA system are different from both cocaine and amphetamine (AMPH) self-administration (Mateo et al., 2005; Ferris et al., 2011, 2012, 2013; Calipari et al., 2013 a, b, c, d, e) suggesting that MPH is unique in its actions at the DAT. Indeed, MPH has been shown to interact with the DAT in a way that is unique, where it binds to the DAT in a way that is different from both AMPH and cocaine (Wayment et al., 1999; Dar et al., 2005). Further, while the effects of blockers at the DAT are reduced following cocaine self-
275 254 administration, MPH is unchanged like releasers, suggesting that it may have properties of both classes of psychostimulants (Ferris et al., 2012). MPH is traditionally categorized as a DAT blocker because it is not transported into the cell; however, it has been demonstrated to have release capabilities at high concentrations (Russel et al., 1998; Dyck et al., 1980; Sonders et al., 1997). Using voltammetry, blockers and releasers have distinct profiles with releasers resulting in a depletion of stimulated DA release from terminals with increasing concentrations and blockers increasing DA release (Ferris et al., 2012). Here we used the release profiles of cocaine, AMPH, and MPH to determine if MPH changes release in a fashion that is similar to blockers or releasers. Here we find that MPH intermittent access MPH self-administration resulted in enhanced releaser and MPH potency, but no change in cocaine potency. These effects have been demonstrated previously using extend-access MPH self-administration, demonstrating that the effects are not due to escalation (Calipari et al., 2013a, b). Further unlike cocaine, MPH effects are not affected by pattern of intake, demonstrating that MPH self-administration results in a sensitized response to releasers and MPH regardless of how it is administered, an effect that could be predictive of significant abuse/addiction potential for the compound. Additionally, although MPH s effects are enhanced similar to releasers following MPH self-administration, its release profile more similarly resembles a blocker, whereby MPH increases release and does not deplete DA terminals, suggesting that MPH may represent a new class of psychostimulants.
276 255 Method: Animals: Male Sprague-Dawley rats ( g; Harlan Laboratories, Frederick, Maryland) were used for all self-administration experiments. Rats were maintained on a 12:12 hour reverse light/dark cycle (3:00 am lights off; 3:00 pm lights on) with food and water ad libitum. All animals were maintained according to the National Institutes of Health guidelines in Association for Assessment and Accreditation of Laboratory Animal Care accredited facilities. The experimental protocol was approved by the Institutional Animal Care and Use Committee at Wake Forest University School of Medicine. Self-Administration: Rats were anesthetized and implanted with chronic indwelling jugular catheters and trained for i.v. self-administration as previously described (Liu et al., 2007). Following surgery, animals were singly housed, and all self-administration sessions took place in the home cage during the active/dark cycle (9:00 am 3:00 pm). After a 2-day recovery period, animals underwent a training paradigm within which animals were given access on a fixed ratio one (FR1) schedule to a MPH-paired lever, which, upon responding, initiated an intravenous injection of MPH (0.56 mg/kg, infused over 4 s). After each response/infusion, the lever was retracted and a stimulus light was illuminated for a 20 second timeout period, during which the animal had no access to the lever, and thus could not respond for drug. During training, sessions were terminated after a maximum of 20 infusions or 6 hours, whichever occurred first. An animal was considered to have acquired upon responding for 20 injections for two consecutive days and a stable pattern infusion intervals was present. Following training, animals were assigned intermittent access after which the dopaminergic alterations that resulted were assessed.
277 256 Intermittent Access Group: Animals were given access to MPH on an intermittent schedule of administration described previously (Calipari et al., 2013c). During each 6h session animals had access to MPH for 12 five minute trails separated by 25-minute timeout periods. Within each five-minute session, there were no timeouts other than during each infusion, and the animal could press the lever on an FR1 schedule to receive a 1-sec infusion of MPH (0.140 mg/kg/inf). Upon responding a stimulus light illuminated concurrently with the 1-sec infusion of drug. In Vitro Voltammetry: Fast scan cyclic voltammetry was used to characterize baseline DA system kinetics, D2 autoreceptor activity, and the ability of psychostimulants to inhibit DA uptake in the nucleus accumbens (NAc) core. Voltammetry experiments were conducted during the dark phase of the light cycle 18 hours after commencement of the final self-administration session. A vibrating tissue slicer was used to prepare 400 µm thick coronal brain sections containing the NAc. The tissue was immersed in oxygenated artificial cerebrospinal fluid (acsf) containing (in mm): NaCl (126), KCl (2.5), NaH 2 PO 4 (1.2), CaCl 2 (2.4), MgCl 2 (1.2), NaHCO 3 (25), glucose (11), L-ascorbic acid (0.4) and ph was adjusted to 7.4. Once sliced, the tissue was transferred to the testing chambers containing bath acsf (32 C), which flowed at 1 ml/min. After a 30-min equilibration period, a cylindrical carbon fiber microelectrode ( μm length, 7 μm radius) and a bipolar stimulating electrode were placed into the core of the NAc. The NAc was selected because of the important role in the reinforcing and rewarding actions of cocaine. Further, our previous research has concentrated on plasticity of DATs in the core and demonstrated that these alterations occur due to self-administration history of a number of drugs (Calipari et al., 2013a, b, c, d; Ferris et al., 2011, 2012, 2013).
278 257 Endogenous DA release was evoked by a single electrical pulse (300 μa, 4 msec, monophasic) applied to the tissue every 5 minutes. Extracellular DA was recorded by applying a triangular waveform ( 0.4 to +1.2 to 0.4V vs Ag/AgCl, 400 V/s). Once the extracellular DA response was stable for three consecutive stimulations, cocaine ( μmol/l), MPH ( μmol/l) and (AMPH μmol/l) were applied cumulatively to the brain slice to determine the effects of cocaine self-administration on drug-induced uptake inhibition. In order to determine D2 autoreceptor sensitivity following these selfadministration paradigms, we also ran dose-response curves for quinpirole ( μmol/l), a D2 receptor agonist, and reversed the effects with sulpiride (2 μmol/l), a D2 antagonist. Data Analysis: For all analysis of FSCV data DEMON voltammetry software was used (Yorgason et al., 2011). To evaluate the effects of MPH self-administration on DA system kinetics, evoked levels of DA were modeled using Michaelis-Menten kinetics as described previously (Calipari et al., 2013a). Immediately following the completion of each concentration response curve, recording electrodes were calibrated by recording their response (in electrical current; na) to a known concentration of DA in acsf (3 μm) using a flow-injection system. This value was then used to convert electrical current to DA concentration. For cocaine, MPH, and AMPH dose-response curves, K m, a measure of apparent affinity for the DAT, was used to determine changes in ability of the psychostimulants to inhibit DA uptake in the NAc relative to baseline. Because quinpirole is a D2 agonist it concentration-dependently reduces the peak height of the DA signal. As a result we can assess the functional sensitivity of D2 autoreceptors by
279 258 determining if there is a shift in the concentration-response curve for quinpirole. All curves for D2 function are expressed as percent baseline DA release. Calculating K i values: As described by Jones et al. (1995), inhibition constants (K i ) were determined by plotting the linear concentration-effect profiles and determining the slope of the linear regression. The K i was calculated by the equation K m /slope. K i values are reported in µm and are a measure of the drug concentration that is necessary to produce 50% uptake inhibition. Statistics: Graph Pad Prism (version 5, La Jolla, CA, USA) was used to statistically analyze data sets and create graphs. Baseline voltammetry data and K i values were compared using a one-way analysis of variance (ANOVA). When main effects were obtained (P < 0.05), differences between groups were tested using a Tukey post hoc tests. Release data and data obtained after perfusion of COC, MPH, AMPH, or quinpirole were subjected to a two-way ANOVA with experimental group and concentration of drug as the factors. Differences between groups were tested using a Bonferroni post hoc test. Results: IntA MPH self-administration does not result in escalation. Long-access selfadministration of MPH has been previously been shown to result in escalation of intake over sessions (Marusich et al., 2010). Here we find that intermittent self-administration of MPH results in stable intake over sessions (Fig 1 A). Regression analysis indicates that the slope of intake over sessions is not significantly different from zero indicating that escalation does not occur in these animals with this model (Fig 1 B).

280 259 Figure 1: Animals self-administer methylphenidate (MPH) at high rates when given intermittent access. Rats were given five minute unlimited access followed by a 25 minute time-out period to induce an intermittent pattern of self-administration within each six hour sessions. (A) Representative data showing the pattern of selfadministration over the 14 sessions. Tick marks represent earned infusions. (B) Group data plotting total infusions (0.140 mg/kg/inf) in each session over the 14 total sessions. The slope of the regression line (red) is not significant indicating that animals do not escalate their intake.
281 260 Intermittent MPH self-administration results in increased stimulated DA release and V max at baseline. Pre-synaptic baseline DA system kinetics were assessed using FSCV in MPH self-administration and control groups. MPH self-administration resulted in an increase in maximal rate of uptake (Fig 2A) and stimulated DA release (Fig 2B). A two-tailed student s t-test revealed a significant increase in stimulated DA release following intermittent MPH self-administration (t 36 = 2.191, p < 0.05). In addition, a two-tailed student s t-test also revealed a significant increase in V max following MPH self-administration (t 33 = 2.919, p < 0.01; Fig 2C). D2 autoreceptors are subsensitive following MPH self-administration. In order to determine if the differential DA kinetic effects could be due to differences in D2 autoreceptor activity, we assessed the sensitivity of the DA system to quinpirole, a D2 autoreceptor agonist. ANOVA revealed a significant interaction (F 1, 28 = 4.079, p < 0.01) (Fig 3A). Bonferroni post-hoc analysis revealed that quinpirole was significantly less effective at reducing DA release at the 30 nm dose following MPH self-administration (p < 0.05), indicating that D2 autoreceptor activity is reduced. The subsensitive D2 autoreceptors that are present following MPH self-administration could explain, at least in part, the increase in stimulated DA release. Sulpiride reversal was run to confirm that the reduction in peak height of DA release was due to D2 receptors and not another mechanism (Fig 3B). Sulpiride reversed quinpirole effects, and there was no significant difference between groups. Intermittent MPH self-administration results in increased MPH potency. To investigate how intermittent MPH-DAT interactions affect MPH potency, we ran
 Representative plot of DA release and uptake at baseline in control and MPH self-administration groups.")
 Group data showing that stimulated release is elevated following intermittent MPH selfadministration.")
282 261 Figure 2: Intermittent methylphenidate (MPH) self-administration leads to increased stimulated dopamine (DA) release and uptake. Baseline DA system kinetics assessed by fast scan cyclic voltammetry. (A) Representative plot of DA release and uptake at baseline in control and MPH self-administration groups. Data is normalized to peak height to highlight differences in uptake between groups. Color plots which show DA (green) over time as measured in current (z-axis). (B) Group data showing that stimulated release is elevated following intermittent MPH selfadministration. (C) Group data demonstrating that maximal rate of DA uptake (V max ) is elevated at baseline following MPH self-administration. IntA, intermittent access; *, p < 0.05; **, p < 0.01


283 262 Figure 3. Intermittent methylphenidate (MPH) self-administration causes D2 autoreceptor subsensitivity. D2 autoreceptors were assessed by quinpirole doseresponse curves. (A) Quinpirole dose-response curve represented as percent of baseline DA peak height. Intermittent MPH self-administration resulted in subsensitive D2 autoreceptors as compared to controls. (B) Sulpiride reversal was used to confirm that the reduction in peak height was due to D2 autoreceptors. IntA, intermittent access; *, p < 0.05 vs control
284 263 cumulative dose-response curves for MPH following intermittent MPH selfadministration. Using FSCV in brain slices, MPH effects on DA uptake inhibition were recorded over a five-point dose-response curve for the compound. MPH was found to be more effective at inhibiting DA uptake inhibition following MPH self-administration (Fig 4A). ANOVA revealed a significant interaction of MPH self-administration on the doseresponse curve for MPH (F 1, 28 = 3.238, p < 0.05; Fig 4B). Bonferroni post hoc analysis revealed that MPH-induced uptake inhibition was augmented at the 30 μm dose following MPH self-administration (p < 0.05). A linear dose-response curve was plotted to calculate the K i of MPH in control and self-administration animals (Fig 4C). The slopes of the regression lines of the linear dose-response curve for MPH-induced uptake inhibition were significantly different (p < 0.01), indicating that MPH was more potent following MPH self-administration. The K i is the dose of drug at which 50% uptake inhibition is achieved, and is calculated from the slope of the linear dose-response curve for the compound. A Student s t-test revealed that MPH potency as measured by K i was increased (t 7 = 2.30, p < 0.05; Fig 4D), with a lower concentration of MPH resulting in 50% uptake inhibition following MPH self-administration as compared to controls. AMPH, potency is increased following IntA MPH self-administration. Because MPH is structurally similar to the releaser AMPH, but functionally similar to the blocker cocaine, we assessed the effects of MPH self-administration on AMPH and cocaine potencies to determine if the neurochemical are conferred to compounds of a similar structural or functional class. If the effects are structure specific AMPH potency should be similar to the effects of MPH self-administration on MPH potency. AMPH was found


285 264
286 265 Figure 4. Sensitization of the neurochemical effects of methylphenidate (MPH). Fast scan cyclic voltammetry was used to assess changes in the potency of MPH at the DAT following MPH self-administration. (A) Representative data showing MPH-induced uptake inhibition at the 10 µm dose. Traces demonstrate that MPH is more effective at inhibiting uptake, as demonstrated by the increased time to return to baseline. Data was normalized to peak height to highlight differences in uptake across groups. Color plots which show DA (green) over time as measured in current (z-axis). (B) Group data showing that MPH self-administration results in increased potency of MPH over a doseresponse curve. (C) K i values across groups were calculated based off of the linear doseresponse curve for MPH in each group. (D) Bar graph of K i values for cocaine in control and MPH self-administration groups. K i values are a measure of the concentration of drug at which 50% inhibition is achieved. IntA, intermittent access; *, p < 0.05 vs control; **, p < 0.01 vs control
287 266 to be more effective at inhibiting DA uptake inhibition following MPH selfadministration (Fig 5A), an effect that was similar to that of MPH. ANOVA revealed a significant interaction of MPH self-administration on the dose-response curve for AMPH (F 1, 32 = 2.744, p < 0.05; Fig 5B). Bonferroni post hoc analysis revealed that AMPHinduced uptake inhibition was augmented at the 10 μm dose following MPH selfadministration (p < 0.05). In addition, the linear dose-response curve was plotted to calculate the K i of AMPH in control and self-administration animals (Fig 5C). The slopes of the regression lines of the linear dose-response curve for AMPH-induced uptake inhibition were significantly different (p < 0.05), indicating that AMPH was more potent following MPH self-administration. A Student s t-test revealed that MPH potency as measured by K i was increased (t 7 = 2.7, p < 0.05; Fig 5D) as well, with a lower concentration of AMPH resulting in 50% uptake inhibition following MPH selfadministration as compared to controls. Because AMPH effects are similar to MPH following MPH self-administration it is likely that these effects are specific to the structure of these compounds rather than their function as blocker or releaser. Cocaine potency is unchanged following IntA MPH self-administration. Because AMPH and MPH behaved similarly after MPH self-administration, we hypothesized that these effects were due to the structure of the compound; however, to confirm this, we used cocaine, a compound structurally dissimilar from MPH, but functionally the same. Cocaine was found to be equivalently effective at inhibiting DA uptake inhibition following MPH self-administration (Fig 6A), an effect that was dissimilar to that of MPH. There was no significant effect of MPH self-administration on the dose-response


288 267
289 268 Figure 5. Enhanced amphetamine (AMPH) potency following methylphenidate (MPH) self-administration. Fast scan cyclic voltammetry was used to assess changes in the potency of AMPH following MPH self-administration. (A) Representative data showing AMPH-induced uptake inhibition at the 1 µm dose. AMPH is more effective at inhibiting uptake, as demonstrated by the increased time to return to baseline in the traces. Data was normalized to peak height to highlight differences in uptake across groups. Color plots which show DA (green) over time as measured in current (z-axis). (B) Group data showing that MPH self-administration results in increased potency of AMPH. (C) K i values across groups were calculated based off of the linear dose-response curve for AMPH in each group. (D) Bar graph of K i values for AMPH in control and MPH self-administration groups. IntA, intermittent access; *, p < 0.05 vs control


290 269
291 270 Figure 6. Cocaine potency is unaffected by a history of methylphenidate (MPH) selfadministration. Fast scan cyclic voltammetry was used to assess changes in the potency of MPH at the DAT following MPH self-administration. (A) Representative data showing cocaine (10 µm) -induced uptake inhibition. Data was normalized to peak height. Color plots which show DA (green) over time as measured in current (z-axis). (B) Group data showing no change in cocaine potency following MPH selfadministration. (C) K i values across groups were calculated based off of the linear doseresponse curve for cocaine in each group. (D) Bar graph of K i values for cocaine in control and MPH self-administration groups. K i values are a measure of the concentration of drug at which 50% inhibition is achieved. IntA, intermittent access; *, p < 0.05 vs control curve for cocaine (Fig 6B). In addition, the linear dose-response curve was plotted to calculate the K i of AMPH in control and self-administration animals (Fig 6C). The slopes of the regression lines of the linear dose-response curve for cocaineinduced uptake inhibition were not significantly different. In addition, cocaine potency as measured by K i was unchanged (Fig 6D).
292 271 Stimulated DA release profiles for MPH, cocaine, and AMPH are differentially affected by MPH self-administration. To determine how MPH affected DA release we measured the release effects of MPH, AMPH, and cocaine across a dose-response curve for each of these drugs. First, we assessed the release profile of MPH to determine how it was affected by a prior MPH self-administration history. MPH exhibits an inverted U shaped profile that is indicative of a DAT blocker (Fig 7A). Curves are expressed as percent peak height at baseline, and peak height is a measure of both release and uptake as uptake occurs continuously in the presence of DA. Because of this, blockers result in inverted-u shaped dose-response curves, where they increase peak height at the early doses, due to their uptake-inhibition affects. At the higher doses, the peak height of the signal is reduced, due to more uptake-inhibition resulting in D2 autoreceptor activation. In addition, ANOVA revealed a significant main effect of MPH self-administration (F 1, 33 = 6.929, p < 0.05), indicating that MPH-induced DA release was increased as compared to control animals. To determine if the release effects were common to all drugs or specific to a certain class/structure, cocaine was also run. As with MPH, cocaine exhibited an inverted U shaped release profile indicative of blockers (Fig 7B). Although the release profile was similar to MPH, like with drug potency, the release effects of cocaine were unaffected by MPH self-administration. This gives further support to the idea that the effects of MPH self-administration on subsequent psychostimulant effects are driven by the structure of the compounds and are not specific to function as blocker or releaser. To further characterize the release effects of MPH self-administration, the release profile over a dose-response curve for AMPH was assessed. AMPH has a different

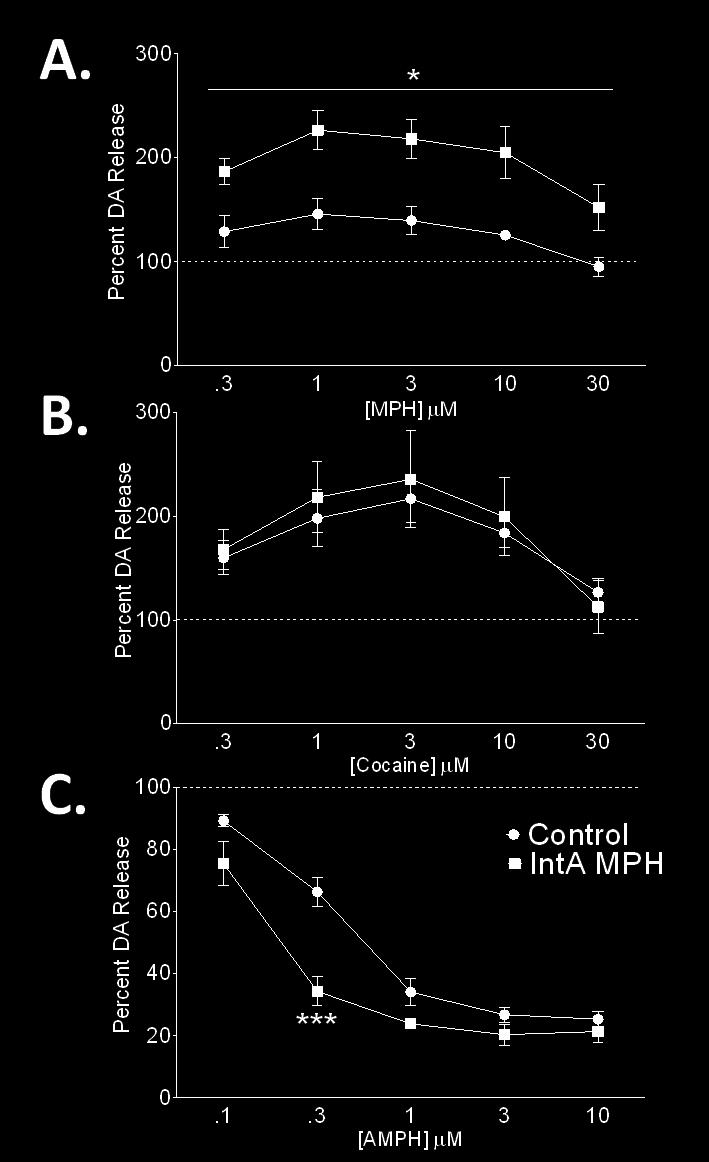
293 272
294 273 Figure 7. Stimulated DA release profile of cocaine, methylphenidate (MPH), and amphetamine (AMPH) following intermittent MPH self-administration. Stimulated release was determined by the peak height of the signal throughout the dose-response curve for each respective drug. Data was then reported as percent baseline peak height. (A) AMPH dose-response curve demonstrating that AMPH-induced release is reduced following MPH self-administration. Profile of releasers is characterized by decreased DA, due to depleted terminals, as doses are increased. (B) MPH dose-response curve that indicates that MPH-induced DA release is augmented following MPH selfadministration. The release profile of MPH indicates that it is a DAT blocker. Profile of DAT blockers is characterized by an inverted U shaped curve with increased DA at lower concentrations and reduced DA at higher concentrations. (C) Cocaine doseresponse curve that indicates that cocaine-induced DA release is unchanged following MPH self-administration.
295 274 profile due to its mechanism of action as a DA releaser. Because AMPH is releasing DA at all times, independent of stimulated release, it dose-dependently depletes DA releasable pools terminals leading to decreased DA release over the dose-response curve (Fig 7C). ANOVA revealed a significant main effect of MPH self-administration on AMPH release (F 1, 28 = 13.96, p < 0.01). Bonferroni post hoc analysis revealed that AMPH release was significantly reduced at the 300nM dose (p < 0.001). The increase in AMPH-induced DA reduction indicates an increase in the potency of AMPH to release DA in these animals. In addition, the fact that both MPH and AMPH are more potent in their DA releasing effects, although it is through different release profiles, gives further support to the idea that the effects of MPH self-administration on subsequent psychostimulant effects are driven by structure. Discussion: Here we find that intermittent MPH self-administration results in DA system changes that may increase the abuse liability of some psychostimulants. In regards to DA kinetics, we found increased maximal rate of uptake and increased stimulated DA release as well as subsensitive D2-like autoreceptors. Further, the potency of AMPH and MPH for DA uptake inhibition, but not cocaine, was increased as was the K i of AMPH and MPH for the DAT. AMPH and MPH, but not cocaine, release effects were also sensitized, whereby, MPH-induced DA release was enhanced and AMPH-induced reductions in evoked DA release were augmented. Although MPH release effects were sensitized, the release profile of MPH resembled that of cocaine, confirming that MPH is not a releaser, even though it classes with releasers in regards to its potency effects. The
296 275 similarities between the MPH self-administration-induced DA system adaptations in the current study as compared to previous work with extended access administration (Calipari et al., 2013 a, b) further highlights the unique properties of MPH, as cocaine self-administration when administered intermittently versus continuously results in divergent effects (Calipari et al., 2013 c). Taken together, this suggests that MPH is a unique compound and the effects cannot be generalized to other psychostimulants. Further, prior MPH abuse may increase the neurochemical potency of releasers and MPH, which could lead to increases in reinforcement and abuse of those compounds. The NAc core region is involved in reward learning, goal oriented behaviors, and continued self-administration after acquisition has already occurred (Willuhn et al, 2010). As a result, changes in release and uptake in this region can have implications, not only for drug effects, but also for learning and the execution of goal-oriented behaviors in the absence of drug. Here we see an increase in evoked DA release, which is likely due to subsensitive D2-like autoreceptors that are not as effective at reducing DA release. The increase in evoked release could lead to enhanced goal-reward associations, which could, in turn, lead to increased associations between drugs and their rewarding/reinforcing effects (Flagel et al., 2011; Wanant et al., 2009). It is important to note that although there are increases in evoked DA release following intermittent MPH self-administration, uptake rates are also increased, which could be a compensatory mechanism for the increased DA release. The increase in uptake rate is likely caused by an increase in DAT levels, which has been demonstrated previously following MPH self-administration (Calipari et al., 2013a). The increase in DAT levels are particularly important, as increasing DAT levels alone, by transgenic over-expression has been shown to increase
297 276 the behavioral and neurochemical potency of releasers such as AMPH (Salahpour et al., 2008; Calipari et al a), highlighting that these baseline changes alone may increase the susceptibility of individuals to abuse/addiction of psychostimulants. In addition to differences in baseline evoked DA release between the groups, there was a facilitation of MPH and AMPH-induced release effects. At increasing concentrations, AMPH leads to depletion of DA terminals, observed as reduced stimulated DA release (Jones et al., 1998), an effect that is enhanced by intermittent MPH self-administration. This release profile is shared by releaser compounds and can be used to determine the function of psychostimulants, specifically if they have releasing properties (Ferris et al., 2011). Although MPH-induced DA release effects are enhanced, MPH exhibits the release profile of a blocker, which is characterized by increased DA release with increasing concentrations. This demonstrates that although MPH classes with releasers in regards to its potency effects at the DAT it is not a pure DA releaser, but rather exhibits properties of both classes of psychostimulants. This is in line with previous work demonstrating that although MPH is not transported into cells, and thus, cannot release from vesicles, it binds to the DAT in a way that is similar to releasers (Sonders et al., 1997; Dar et al., 2005; Wayment et al., 1999). Importantly, the difference between the release profile of MPH and AMPH highlights that the release effects of the compounds are not responsible for the enhanced potency of both of the compounds, as release is decreased for AMPH and increased for MPH but they both have enhanced uptake inhibition effects. Additionally, the D2 subsensitivity is also unlikely to drive the enhanced release of AMPH and MPH, as cocaine release was unaffected by MPH selfadministration.
298 277 A potential mechanism for the increased potency of AMPH and MPH for causing both uptake inhibition and release effects is via increased DAT levels. Previous work has demonstrated that the enhanced potency of AMPH and MPH following extended access MPH self-administration are due to elevated DAT levels, and here we observe the same neurochemical effects, suggesting that these changes may occur via the same mechanism (Calipari et al., 2013a, b). It has been shown previously that both the behavioral and neurochemical effects of both AMPH and MPH, but not cocaine, are enhanced in mice over-expressing the DAT (Calipari et al., 2013a; Salahpour et al., 2008). Although, this is the first study to show enhanced MPH-induced DA release following MPH selfadministration, it is possible that DAT levels dictate these effects as well. MPH selfadministration results in AMPH depleting terminals faster, this could be due to the increased DAT levels, which allow more AMPH into the cell, resulting in enhanced vesicular DA depletion. Although MPH is not a releaser, it has been shown to redistribute vesicles from storage pools into the readily releasable pool (Chadchankar et al., 2012; Volz et al., 2008). This could be a potential mechanism for its release effects and it is possible that these effects could be dependent to MPH binding to the DAT. Behaviorally, these neurochemical changes have important implications, as psychostimulant reinforcement is mediated by the ability of the compound to elevate DA and inhibit the DAT (Ritz et al., 1989; Roberts et al., 1977). Further, changes in drug potency in the NAc core, as measured by voltammetry, have been shown to be associated with concomitant changes in reinforcement behavior, with increases in potency resulting in increases in reinforcing efficacy (Calipari et al., 2013 b). This suggests that intermittent administration of MPH may lead to an increase in reward and reinforcement
299 278 behavior for amphetamines and other releasers, which could increase the abuse liability/addiction potential of these compounds. This is of particular importance, given the high increasing levels of MPH abuse in the adult and adolescent populations in recent years, as well as the co-abuse of MPH with other drugs such as amphetamines (Teter et al., 2006). Both the kinetic changes and the drug potency changes are the same as have been seen previously with an extended access model of MPH administration, demonstrating that the neurochemical effects are due to MPH administration and not the escalation that occurs in extended-access situations (Calipari et al., 2013 a, b). Extended-access selfadministration of MPH, and a number of other psychostimulants such as methamphetamine, nicotine, and cocaine, results in escalation of first-hour intake, and is thought to model the switch from abuse to addiction (Marusich et al., 2010; Ahmed & Koob, 2003; Kitamura et al., 2006; Cohen et al., 2012). As such, much work has focused on the effects of escalation on reward, reinforcement, and DA neurochemistry (Ahmed & Koob, 2003; Ben-Shahar et al., 2006). In previous work it was unclear if the effects were due to MPH administration itself, or the pattern of self-administration, which resulted in escalation. In the current intermittent access model, escalation does not occur; as a result, the neurochemical changes that occur cannot be due to escalation. Increased drug potency in the NAc core region has been shown previously to be predictive of similar changes in reinforcement and reward behaviors (Calipari et al., 201a), suggesting that MPH administration may result in changes that could increase the abuse liability of amphetamines independent of escalation of intake. Further, this suggests that any temporal pattern of MPH intake can cause these effects, although the total intake per day
300 279 is a factor in determining if any changes occur (Calipari et al., 2013a). The changes outlined in this study seem to be characteristic of abuse relevant doses, as therapeutic doses seem to have no effect on DA system neurochemistry in the NAc core region. Previous work using oral administration of MPH at low doses (5mg/kg, 2x per day) resulted in no change in any DA system measures in the NAc. This is not surprising, as at low doses MPH has been shown to preferentially increase DA in the prefrontal cortex, an effect, which is thought to mediate is therapeutic cognitive enhancing effects. It is only until higher doses that MPH effectively enhances DA release in the NAc (Berridge et al., 2006; Devilbiss et al., 2008). The lack of difference between previous work with an extended-access model and the current intermittent access model highlights that MPH is unique in the compensatory changes that occur following self-administration as they relate to other psychostimulants (Calipari et al., 2013a, b). In regards to cocaine self-administration, intermittent-access and long-access self-administration have different neurochemical effects, with longaccess causing cocaine tolerance and intermittent access sensitization, demonstrating the pattern of intake plays an important role in determining the compensatory changes that occur following self-administration (Calipari et al., 2013c). Previous work using extended-access MPH self-administration has shown the same compensatory changes as intermittent self-administration, suggesting that for MPH self-administration, pattern of administration is not a factor in determine the neurochemical changes that occur. This is particularly relevant as MPH causes a sensitized effect when given both continuously and intermittently, while previous work with many other compounds has shown tolerance with continuous administration and sensitization with intermittent administration (Jones
301 280 et al., 1996; Griggs et al., 2010). The fact that sensitization, which is used as an addiction model, occurs when MPH is taken in any pattern of administration, suggests that MPH may be a particularly dangerous compound, especially when taken via non-oral routes of administration. Taken together, this work further highlights that MPH is a unique compound, both in its acute effects as well as the compensatory effects that occur following selfadministration. Acutely, MPH exhibits the uptake-inhibition profile of a releaser but the release profile of a blocker suggesting that MPH is part of a unique class of compounds that does not fit into either the releaser or blocker category. Further, the compensatory effects that occur following MPH self-administration are not dependent on the temporal profile of MPH intake, an effect that is different from previous work with cocaine selfadministration. This highlights that MPH, when taken in larger amounts than prescribed and via any pattern of self-administration, could potentially enhance the abuse and addiction potential of psychostimulant releasers such as AMPH, methamphetamine, and bath salts. Acknowledgements: This work was funded by NIH grants R01 DA024095, R01 DA03016 (SRJ), K99 DA (MJF), T32 DA and F31 DA (ESC).
302 281 References: Ahmed SH, Lin D, Koob GF, Parsons LH. Escalation of cocaine self-administration does not depend on altered cocaine-induced nucleus accumbens dopamine levels. J Neurochem. 86, (2003) Ben-Shahar O, Moscarello JM, Ettenberg A. One hour, but not six hours, of daily access to self-administered cocaine results in elevated levels of the dopamine transporter. Brain Res. 1095, (2006) Berridge CW, Devilbiss DM, Andrzejewski ME, Arnsten AF, Kelley AE, Schmeichel B, Hamilton C, Spencer RC. Methylphenidate preferentially increases catecholamine neurotransmission within the prefrontal cortex at low doses that enhance cognitive function. Biol Psychiatry 60, (2006) Calipari ES, Ferris MJ, Salahpour A, Caron MG, Roberts DC, Jones SR. Methylphenidate amplifies the potency and reinforcing effects of amphetamines by increasing dopamine transporter expression. In press (2013a) Calipari ES, Ferris MJ, Melchior JR, Bermejo K, Salahpour A, Roberts DC, Jones SR. Methylphenidate and cocaine self-administration produce distinct dopamine terminal alterations. Addict Biol. In press (2013b) Calipari ES, Ferris MJ, Zimmer BA, Roberts DCS, Jones SR. Temporal pattern of cocaine intake determines tolerance versus sensitization of cocaine effects at the dopamine transporter. Neuropsychopharm. In press (2013c) Calipari ES, Ferris MJ, Jones SR Extended access cocaine self-administration results in tolerance to the dopamine-elevating and locomotor-stimulating effects of cocaine. J. Neurochem. In press (2013d) Calipari ES, Beveridge TJ, Jones SR, Porrino LJ. Withdrawal from extended access cocaine self-administration results in dysregulated functional activity and altered locomotor activity in rats. Eur. J. Neurosci. In press. (2013e) Chadchankar H, Ihalainen J, Tanila H, Yavich L. Methylphenidate modifies overflow and presynaptic compartmentalization of dopamine via an α-synuclein-dependent mechanism. J Pharmacol Exp Ther. 341, (2012) Cohen A, Koob GF, George O. Robust escalation of nicotine intake with extended access to nicotine self-administration and intermittent periods of abstinence. Neuropsychopharmacology. 37, (2012) Dar DE, Mayo C, Uhl GR. The interaction of methylphenidate and benztropine with the dopamine transporter is different than other substrates and ligands. Biochem Pharmacol. 70, (2005)
303 282 Devilbiss DM, Berridge CW. Cognition-enhancing doses of methylphenidate preferentially increase prefrontal cortex neuronal responsiveness. Biol Psychiatry 64, (2008) Dyck LE, Boulton AA, Jones RS. A comparison of the effects of methylphenidate and amphetamine on the simultaneous release of radiolabelled dopamine and p- or m- tyramine from rat striatal slices. Eur J Pharmacol. 68, (1980) Ferris MJ, Calipari ES, Mateo Y, Melchior JR, Roberts DC, Jones SR. Cocaine selfadministration produces pharmacodynamic tolerance: differential effects on the potency of dopamine transporter blockers, releasers, and methylphenidate. Neuropsychopharmacology. 37, (2012) Ferris MJ, Calipari ES, Melchior JR, Roberts DC, España RA, Jones SR. Paradoxical tolerance to cocaine after initial supersensitivity in drug-use-prone animals. Eur J Neurosci. In press (2013) Ferris MJ, Mateo Y, Roberts DC, Jones SR. Cocaine-insensitive dopamine transporters with intact substrate transport produced by self-administration. Biol Psychiatry. 69, (2011) Flagel SB, Clark JJ, Robinson TE, Mayo L, Czuj A, Willuhn I, Akers CA, Clinton SM, Phillips PE, Akil H. A selective role for dopamine in stimulus-reward learning. Nature 469, 53-7 (2011) Frauger E, Moracchini C, Le Boisselier R, Braunstein D, Thirion X, Micallef J. OPPIDUM surveillance program: 20 years of information on drug abuse in France. Fundam Clin Pharmacol. In press (2013) Griggs R, Weir C, Wayman W, Koeltzow TE. Intermittent methylphenidate during adolescent development produces locomotor hyperactivity and an enhanced response to cocaine compared to continuous treatment in rats. Pharmacol Biochem Behav. 96, (2010) Jones SR, Gainetdinov RR, Wightman RM, Caron MG. Mechanisms of amphetamine action revealed in mice lacking the dopamine transporter. J Neurosci. 18, (1998) Jones SR, Garris PA, Wightman RM. Different effects of cocaine and nomifensine on dopamine uptake in the caudate-putamen and nucleus accumbens. J Pharmacol Exp Ther. 274, (1995) Jones SR, Lee TH, Wightman RM, Ellinwood EH. Effects of intermittent and continuous cocaine administration on dopamine release and uptake regulation in the striatum: in vitro voltammetric assessment. Psychopharmacology (Berl). 126, (1996) Kitamura O, Wee S, Specio SE, Koob GF, Pulvirenti L. Escalation of methamphetamine self-administration in rats: a dose-effect function. Psychopharmacology (Berl). 186, (2006)
304 283 Koob GF, Le Moal M. Drug abuse: hedonic homeostatic dysregulation. Science 278, 52-8 (1997) Liu Y, Morgan D, Roberts DC. Cross-sensitization of the reinforcing effects of cocaine and amphetamine in rats. Psychopharmacology (Berl). 195, (2007) Marusich JA, Beckmann JS, Gipson CD, Bardo MT. Methylphenidate as a reinforcer for rats: contingent delivery and intake escalation. Exp Clin Psychopharmacol. 18, (2010) Mateo Y, Lack CM, Morgan D, Roberts DC, Jones SR. Reduced dopamine terminal function and insensitivity to cocaine following cocaine binge self-administration and deprivation. Neuropsychopharmacology. 30, (2005) Parran TV Jr, Jasinski DR. Intravenous methylphenidate abuse. Prototype for prescription drug abuse. Arch Intern Med. 151, (1991) Ritz MC, Kuhar MJ. Relationship between self-administration of amphetamine and monoamine receptors in brain: comparison with cocaine. J Pharmacol Exp Ther. 248, (1989) Roberts DC, Corcoran ME, Fibiger HC. On the role of ascending catecholaminergic systems in intravenous self-administration of cocaine. Pharmacol Biochem Behav. 6, (1977) Rosen JB, Young AM, Beuthin FC, Louis-Ferdinand RT. Discriminative stimulus properties of amphetamine and other stimulants in lead-exposed and normal rats, Pharmacol, Biochem Behav 24, (1985) Russell V, de Villiers A, Sagvolden T, Lamm M, Taljaard J. Differences between electrically-, ritalin- and D-amphetamine-stimulated release of [3H]dopamine from brain slices suggest impaired vesicular storage of dopamine in an animal model of Attention- Deficit Hyperactivity Disorder. Behav Brain Res. 94, (1998) Salahpour A, Ramsey AJ, Medvedev IO, Kile B, Sotnikova TD, Holmstrand E, Ghisi V, Nicholls PJ, Wong L, Murphy K, Sesack SR, Wightman RM, Gainetdinov RR, Caron MG. Increased amphetamine-induced hyperactivity and reward in mice overexpressing the dopamine transporter. Proc Natl Acad Sci U S A. 105, (2008) Sherman CB, Hudson LD, Pierson DJ. Severe precocious emphysema in intravenous methylphenidate (Ritalin) abusers. Chest 92, (1987) Silverman PB, Ho BT. The discriminative stimulus properties of 2,5-dimethoxy-4- methylamphetamine (DOM): differentiation from amphetamine. Psychopharmacology (Berl) (1980) Sonders MS, Zhu SJ, Zahniser NR, Kavanaugh MP, Amara SG. Multiple ionic conductances of the human dopamine transporter: the actions of dopamine and psychostimulants. J Neurosci. 17, (1997)
305 284 Substance Abuse and Mental Health Services Administration (SAMHSA) (2008) National Survey on Drug Use and Health. Retrieved at Accessed 15 May Teter CJ, McCabe SE, LaGrange K, Cranford JA, Boyd CJ. Illicit use of specific prescription stimulants among college students: prevalence, motives, and routes of administration. Pharmacotherapy 2, (2006) Tilley MR, Gu HH. The effects of methylphenidate on knockin mice with a methylphenidate-resistant dopamine transporter. J Pharmacol Exp Ther. 327, (2008) Volz TJ, Farnsworth SJ, Hanson GR, Fleckenstein AE. Methylphenidate-induced alterations in synaptic vesicle trafficking and activity. Ann N Y Acad Sci. 1139, (2008) Wanat MJ, Willuhn I, Clark JJ, Phillips PE. Phasic dopamine release in appetitive behaviors and drug addiction. Curr Drug Abuse Rev. 2, (2009) Wayment HK, Deutsch H, Schweri MM, Schenk JO. Effects of methylphenidate analogues on phenethylamine substrates for the striatal dopamine transporter: potential as amphetamine antagonists? J Neurochem. 72, (1999) Willuhn I, Wanat MJ, Clark JJ, Phillips PE. Dopamine signaling in the nucleus accumbens of animals self-administering drugs of abuse. Curr Top Behav Neurosci. 3, (2010) Yorgason JT, España RA, Jones SR. Demon voltammetry and analysis software: analysis of cocaine-induced alterations in dopamine signaling using multiple kinetic measures. J Neurosci Methods. 202, (2011) Zimmer BA, Oleson EB, Roberts DC. The motivation to self-administer is increased after a history of spiking brain levels of cocaine. Neuropsychopharmacology. 37, (2012)
306 285 CHAPTER IX DISCUSSION
307 Introduction A new, well-documented trend in the adolescent and young adult population is diverting methylphenidate (MPH) prescriptions for recreational use. Individuals administer MPH at levels 2-10 times the prescribed dose, however, until recently, there has been no systematic evidence in the basic research literature on the neurochemical and behavioral effects of this type of abuse (Teter et al., 2006). This work demonstrates that high-dose MPH self-administration, which models abuse of the compound, leads to sensitized responses of the dopamine system to amphetamines and enhanced amphetamine (AMPH)-seeking in rats (Calipari et al., 2013a, b). These data suggest that human MPH abusers may be more susceptible to abuse of, and addiction to, AMPH and related compounds. To our surprise, a second major discovery was a basic mechanism that may predict vulnerability to abuse AMPH and other dopamine (DA) releasers. Namely, increases in DA transporter (DAT) levels produce increases in AMPH and DA releaser potency (Calipari et al., 2013a). This is important because it suggests that known variability in human dopamine transporter expression, particularly in disorders such as ADHD (Madras et al., 2005), may serve as a marker for vulnerability for substance abuse disorder/addiction. Because both MPH and AMPH are readily available and are used commonly in the clinic, these results may have important implications for treating at risk individuals. A third major discovery is that MPH may represent a new class of psychostimulant compounds. Although considered a DAT blocker, MPH has actions
308 287 similar AMPH, where increases in DAT levels increase MPH potency, a phenomenon that does not extend to traditional blockers (Calipari et al., 2013a). In addition to its acute actions at the DAT, MPH is unique in the compensatory changes that occur after repeated self-administration of the compound (Calipari et al., 2013b). The sensitized releaser and MPH potency that occur following MPH self-administration are different from the alterations that occur after both cocaine and AMPH self-administration (Ferris et al., 2011, 2012, 2013; Calipari et al., 2013c, d, e). Further, while cocaine selfadministration-induced alterations are dependent on the temporal pattern of cocaine intake (Calipari et al., 2013c), the alterations that occur with MPH are independent of self-administration paradigm (Chapter 8). These results, taken together, necessitate a revision of the current two-category model for psychostimulants to allow for classifications beyond purely blocker or releaser.. 2. The Effects of Methylphenidate Self-Administration on the Dopamine System are Different from Other Psychostimulants One of the first major findings of this work, outlined in Chapter 5, is that MPH, a DAT blocker, had dopaminergic effects that were opposite from cocaine selfadministration (Figure 1; Calipari et al., 2013b, c, d, e). We aimed to determine if the effects of MPH self-administration on the DA system would resemble that of blockers or releasers, as we had originally hypothesized that the compensatory effects that occurred following cocaine self-administration were due to the functional properties of cocaine as a DAT blocker (Mateo et al., 2005; Ferris et al., 2011, 2012, 2013). However, the lack of
309 288
310 289 Figure 1. Representative voltammetric dopamine traces highlighting the effects of cocaine, amphetamine (AMPH), and methylphenidate (MPH) self-administration (SA) on cocaine potency. For all panels (not real data), red traces represent typical baseline (pre-drug) release and uptake of dopamine in both naïve and cocaine SA animals. Black traces represent typical dopamine release and uptake following application of cocaine in naïve animals. Naïve animals represent animals with no drug history, but have cocaine applied to brain slices. Uptake is inhibited when cocaine is applied, leading to greater maximal peak-height and slower return to baseline. Blue traces represent inhibition of dopamine uptake following application of cocaine in animals with a history of cocaine (A), AMPH (B), or MPH (C) SA. Note that the blue trace in Panel A (cocaine SA) exhibits a reduced maximal peak-height and reduced rightward-shift in the descending limb of the curve relative to the black trace (naïve). Therefore, cocaine SA produces tolerance to the neurochemical effects of cocaine (A), while AMPH (B) or MPH (C) self-administration does not affect cocaine potency. Reprinted with permission from Ferris et al. Copyright (2013) American Chemical Society.
311 290 similarities between the compensatory effects following MPH and cocaine selfadministration demonstrated that the functional classification was not predictive of the neurochemical outcomes following self-administration. Despite almost identical behavioral profiles, baseline changes in maximal DA uptake rate (V max ) and stimulated DA release following MPH or cocaine selfadministration were found to be opposite. Cocaine self-administration resulted in decreased stimulated DA release, decreased V max, and decreased DAT density, while MPH self-administration resulted in an increase in V max and DAT density as well as an increase in stimulated DA release in the nucleus accumbens core. The effects of MPH and cocaine self-administration on drug potencies were opposite as well. Indeed, MPH self-administration increased the potency of MPH, but not cocaine, and cocaine selfadministration decreased the potency of cocaine, but not MPH, demonstrating that two drugs that result in identical self-administration behavior are capable of causing opposite neurobiological adaptations. Additionally, the changes following MPH selfadministration are not in line with the changes associated with AMPH self-administration (no changes; Ferris et al., 2011), suggesting that MPH is unique and does not fit into either the blocker or releaser class. While MPH and cocaine are both classified as DA uptake blockers, they have different chemical structures, which may account for the disparate neurochemical changes that occur following self-administration. MPH is structurally related to AMPH, while cocaine is a tropane (Uhl et al., 2002; Volkow et al, 2002; Heal et al, 2009; Han & Gu 2006). Additionally, MPH, exhibits binding properties that are distinct from both prototypical blockers and releasers, and these unique interactions with the DAT could, at
312 291 least in part, drive the differential compensatory changes associated with MPH selfadministration as compared to AMPH or cocaine (Dar et al., 2005; Wayment et al., 1999). MPH is not a DAT substrate, as it is not transported into cells (Sonders et al., 1997); however, some studies have shown that MPH has the ability to promote reversetransport of DA through the DAT at very high doses (Sproson et al., 2001; Dyck et al., 1980; Russell et al., 1998; Heal et al., 1996). It is clear that DAT inhibition-induced increases in extracellular DA levels do not explain the distinct neurochemical adaptations MPH or cocaine self-administration, and we hypothesize that drug interactions with specific regions of the DAT may explain the differences observed. 3. Psychostimulant Self-Administration-Induced Changes at the DAT are not Dependent on the Function of the Compound as a Blocker or Releaser One of the questions addressed by this work was whether the structure of the compound, or its function, as a blocker or releaser, was responsible for the compensatory changes that occur at the DAT following self-administration. We conducted a number of studies using cocaine and MPH self-administration and were able to compare the results to previous work with AMPH self-administration (Ferris et al., 2011). Using MPH, AMPH, and cocaine as comparator drugs gives the unique advantage of having a prototypical blocker (cocaine), a prototypical releaser (AMPH), and a compound that is classified as a blocker but is structurally similar to releaser (MPH). The unique structural and functional profile of MPH allows for the determination of how structure and function affect the DAT. We hypothesized that if psychostimulant self-administration-induced
313 292 alterations to the DAT were due to structure, MPH effects would resemble those of AMPH. Alternatively if the changes were due to the function of the compound as a blocker or releaser then MPH-induced alterations would closely resemble those of cocaine. Ferris et al. (2012) demonstrated that cocaine self-administration resulted in tolerance to the ability of cocaine and other DAT blockers, but not releasers, to inhibit DA uptake at the DAT. It was initially hypothesized that the effects were due to the function of cocaine as a blocker, thus all compounds of the same class were affected. However, following cocaine self-administration, MPH potency was unaffected, suggesting that the effects were not purely due to the function of the compound. MPH is structurally similar to releasers, and is an AMPH analog (Uhl et al., 2002; Heal et al., 2009; Han & Gu 2006), thus we hypothesized that the changes following cocaine selfadministration were due to the structure of the compound, but not the function. Using AMPH, a prototypical releaser, Ferris et al., (2011) determined the effects of releaser self-administration on the DA system. Not surprisingly, self-administration of releasers (AMPH self-administration) results in no change in potency for any of the drugs tested (blockers or releasers). This is likely due to the fact that the structures of many DA releaser drugs are closely related to the structure of DA itself. Because of the structural similarities between DA and releaser compounds, any changes in the potency of these compounds for the DAT would likely also result in changes in the affinity of DA for the transporter.
314 293 In order to determine if cocaine and AMPH self-administration-induced changes at the DAT are due to structure or function of the compound we then assessed the consequences of MPH self-administration on the DA system, as discussed in Chapter 6. Because we initially hypothesized that the compensatory changes at the DAT were associated with the function of the compound, we expected MPH-induced changes to resemble the changes associated with cocaine self-administration. However, MPH selfadministration produced a neurochemical profile that did not resemble the adaptations associated with blocker or releaser self-administration. Because MPH interacts with the transporter in a way that is distinct from both blockers and releasers (Dar et al., 2005; Wayment et al., 1999), it is likely that the compensatory changes associated with psychostimulant self-administration are actually due to the interactions of the compound with the DAT, and not their function as a blocker or releaser. Of particular importance, these data confirm that the changes associated with psychostimulant self-administration are not due to the ability of the compounds to elevate DA levels, as all of the drugs do this acutely, and to a similar extent at the doses tested. If the psychostimulant-induced alterations were due to elevations in extracellular DA, the compounds tested here should result in similar changes to the DA system following selfadministration. In contrast, we observed distinct neurochemical adaptations following self-administration of MPH, AMPH or cocaine, demonstrating that the differential DA system alterations following psychostimulant self-administration are likely a result of the compounds interacting with the DAT at different binding sites. Although it is likely that the structure of the compound is the main factor dictating the compensatory changes that occur at the DAT following psychostimulant self-
315 294 administration, there are a number of potential caveats. First, MPH has been shown to have release capabilities at high doses (discussed in chapter 8), thus it is possible that the effects of MPH self-administration, which are divergent from cocaine self-administration, are due to MPH acting as a releaser, and not a blocker. However, this is unlikely because if MPH was acting as a releaser to cause its effects, it would be expected that the changes associated with MPH self-administration would resemble that of AMPH selfadministration. Second, it is possible that the differences between the affinities of these compounds for the DAT, serotonin transporter (SERT), and norepinephrine transporter (NET) could play a role in the differential neurochemical consequences observed following self-administration of each compound. While AMPH and cocaine have high affinity for the DAT, SERT and NET, MPH has low affinity for the SERT as evidenced by studies demonstrating that physiologically relevant doses of MPH do not result in increased serotonin levels or SERT inhibition (John & Jones, 2007; Gatley et al., 1996). It is possible that differential effects on serotonin levels underlie discrepancies in compensatory changes that occur following MPH self-administration as compared to selfadministration of cocaine and AMPH. This could be tested by administering a serotonin reuptake inhibitor, such as fluoxetine, in combination with MPH to determine if modulation of serotonin levels can account for differences between MPH and cocaineinduced alteration of dopaminergic function
316 The Pattern of Intake (Intermittent vs. Continuous) Dictates the Compensatory Changes that Occur at the DAT Following Cocaine, but not MPH Self- Administration. In the literature, there are reports of increased, decreased and unaltered DA uptake rates following cocaine self-administration (Ferris et al., 2011; Ramamoorthy et al., 2010; Mateo et al., 2005; Wilson and Kish, 1996). The discrepancies in previous results may result from differential self-administration paradigms producing different neurobiological effects, emphasizing the need for equating doses as well as injection frequency, timing and duration when comparing the compensatory effects of different drugs. We aimed to determine what was driving MPH and cocaine-induced neurochemical changes to DA system. We have observed both sensitization of effects (following MPH selfadministration; intermittent cocaine self-administration) and tolerance to the effects of drugs (following long-access cocaine self-administration); thus, we focused on exploring how intake and temporal profile of self-administration dictate the compensatory effects of MPH and cocaine. As outlined in Chapter 4, for cocaine self-administration intake is responsible for the development of neurochemical tolerance, while temporal pattern of selfadministration (intermittency) dictates the development of a sensitized response to cocaine (Calipari et al., 2013c). These findings are consistent with previous work using intermittent experimenter-administered intraperitoneal (IP) injections of cocaine, which has documented sensitization of the mesolimbic DA system and corresponding behavior (Kalivas and Duffy, 1990, 1993; Parsons and Justice, 1993; Addy et al., 2010). Conversely, experiments using continuous cocaine administration (via mini-pumps or
317 296 long-access self-administration), like the long-access model within this work, resulted in neurochemical tolerance (Jones et al., 1996; King et al., 1998; Inada et al., 1992; Calipari et al., 2013b, c), highlighting the importance that pattern of administration plays in determining the neurochemical effects elicited by cocaine. Discussed in Chapter 5, in contrast to the changes observed following cocaine self-administration, temporal pattern had no effect on MPH self-administration-induced DA system adaptations (Calipari et al., 2013a, c). Both extended-access and intermittent access MPH self-administration resulted in increased releaser and MPH, but not cocaine potency, as well as elevated maximal rates of uptake and enhanced pre-drug evoked DA release. While previous work with many other compounds has shown tolerance with continuous administration and sensitization with intermittent administration, MPH is unique as MPH self-administration causes sensitized effects of both MPH and releaser compounds when given both continuously and intermittently (Jones et al., 1996; Griggs et al., 2010). The fact that sensitization of MPH and amphetamines occurs independent of the pattern of MPH intake, suggests that MPH may be particularly dangerous, even following limited, sporadic administration. 5. Methylphenidate Abuse May Result in an Increased Abuse Liability for Releaser Psychostimulants Our results demonstrating increased potencies of MPH and releasers following MPH self-administration suggest that there may be an increased abuse liability for these compounds following MPH abuse in the human population. These effects occur
318 297 following both intermittent and extended access self-administration further supporting the idea that MPH is dangerous when abused, and, unlike cocaine, any pattern of selfadministration results in increased potency of psychostimulant releasers. The increase in neurochemical potency is accompanied by an increase in reinforcement behavior (Calipari et al., 2013a), where animals will expend more effort in order to obtain MPH at both high and low doses. Increased reinforcement behavior is thought to model abuse liability and increases in effort on higher order schedules such as progressive ratio or the threshold procedure used here, are thought to signify an increased abuse liability in humans. Although this work has implications for augmented abuse potential of MPH following prior MPH use, the increase in motivation to obtain releaser compounds demonstrates that the effects of MPH extend beyond MPH abuse in isolation. It has been proposed in the literature that MPH does not have significant abuse/addiction potential, and that, accordingly, changes in the motivation to administer MPH do not have clinical relevance (Schubiner, 2005). Although there is some epidemiological data to support these ideas, the reduced addiction rates of MPH as compared to cocaine are likely due to differences in the route of administration, and not due to an intrinsic property of the compound (Parasrampuria et al., 2007; Kollins et al., 1998). The idea that MPH is safe and has limited addiction potential has prevented much of the MPH abuse literature from being deemed clinically relevant. Here we demonstrate with a number of studies that the perceived lack of clinical relevance is particularly misguided based on the effects of MPH self-administration on other psychostimulants. Independent of the effects of MPH self-administration on MPH reinforcement, MPH also increases the
319 298 neurochemical potency of releasers, such as AMPH and methamphetamine (METH). Because AMPHs have relatively high addiction potential, it is possible that MPH abuse could lead to an increased propensity for AMPH and METH abuse/addiction. This is particularly relevant given the co-abuse of MPH with a number of other compounds including nicotine, alcohol, and other stimulants such as AMPH and METH (Sepúlveda et al., 2011) 6. Therapeutic Administration of MPH Does Not Result in the Same Dopamine System Changes as Abuse Relevant Doses Although we have shown that pattern of self-administration does not dictate the dopamine system changes that occur following MPH self-administration, intake plays an obvious role in the effects of MPH administration on the DA system. Here we demonstrate that oral administration of low, therapeutic doses and i.v. administration of high, abuse-relevant doses have completely different neurochemical consequences. The lack of effect of oral MPH administration on the DA system is consistent with published work showing no significant neurochemical changes following long-term therapeutic MPH administration (Gill et al., 2012; Humphreys et al., 2013). These data suggest that although MPH could enhance the reinforcing efficacy of MPH and releasers when taken via non-oral routes, and in larger doses than prescribed, the therapeutic use of MPH, taken as prescribed, may be safe. MPH has differential effects at high and low doses, which is likely due to the fact that low-dose MPH preferentially increases DA in the PFC, while higher doses increase
320 299 DA in both the PFC and nucleus accumbens (Berridge et al., 2006; Devilbiss et al., 2006). The selective increase in the PFC could be involved in the cognitive enhancing effects of MPH, but is likely independent of its reinforcing effects (Devilbiss et al., 2008). The inability of MPH to elevate DA in striatal regions at therapeutic doses could explain the lack of effects of oral MPH administration on DA terminals and psychostimulant potencies measured here. Together these data further highlight the dissociation between the effects of therapeutic and abused doses of MPH. 7. DAT Levels Dictate Releaser, but Not Blocker, Potency This work uses a comprehensive approach to look at the effects of DAT levels on psychostimulant potency in neurochemistry and behavior, and highlights that current theories on DAT levels influencing blocker potency may need revision. The current consensus is that increases in DAT levels lead to a reduction in cocaine potency, and that lower DAT levels results in increased cocaine potency. This theory originates from two main sources: 1) Cell culture work showing that DAT overexpression in cells results in decreased cocaine potency with no change in AMPH potency (Chen & Reith, 2007), and 2) Historically, the ability of cocaine to elevate DA in the NAc shell, where DAT levels are low, is much greater than that of the dorsal striatum, where DAT levels are high. Here we demonstrate both behaviorally and neurochemically, that increasing DAT levels in vivo does not change cocaine potency but does alter AMPH potency. Thus, we are proposing a novel theory by which posits that cell surface expression dictates the potency of releasers, but not blockers.
321 300 First, with respect to discrepancies between the findings demonstrated here and previous cell culture work, one possibility is that the use of endogenous DA in voltammetry may produce different results than [ 3 H]-DA used in cell culture studies (Chen & Reith, 2007). It is possible that [ 3 H]-DA is sequestered into intracellular compartments differently than endogenous DA and that releaser compounds do not have access to this [ 3 H]-DA. Additionally, it should be noted that many cell culture studies have determined the absolute effect of psychostimulants on DA levels, neglecting the influence of baseline rates of uptake in the overall effects of these drugs. This is particularly relevant as much work has suggested that behavioral outputs are dependent on a change relative to baseline, not the overall DA levels in isolation (Cragg 2006; Threlfell & Cragg. 2011; Calipari & Ferris, 2013). For example, when DAT levels are increased, as seen in mice with genetic overexpression of functional DATs, baseline uptake is faster. Thus, even with comparable absolute effects between animals, the change from baseline is greater in an animal with increased baseline uptake. Voltammetry determines the effects of the drug while accounting for baseline uptake rates, and therefore, our voltammetry data is consistent with the behavioral outcome of the current study as well as a number of other studies (Salahpour et al, 2008). Second, the increased potency of cocaine in the shell relative to the caudate has been demonstrated in vivo with microdialysis; however, there are a number of factors that could be influencing the differential cocaine potency across regions. In fact, previous work using voltammetry in freely-moving animals has confirmed that brain regional variation in the potency of cocaine are attributed to differences in D2 autoreceptor
322 301 sensitivity in regulating DA release, and not differences in DAT levels (Aragona et al., 2008). Regardless of possible differences in cocaine potency due to other factors such as D2 receptor function or methodological differences, we demonstrate that the changes observed using voltammetry are predictive of behavioral outcomes in regards to both locomotor activity and reinforcing efficacy. Increases in DAT levels as observed following DAT overexpression or following MPH self-administration result in increased MPH and AMPH, but not cocaine potency. This was consistent with both locomotor activity and reinforcement behavior showing an augmentation of AMPH and MPHinduced behaviors, with no change in the effects of cocaine. It is important to note that, previously, Salahpour et al. (2008) demonstrated that there was no difference in MPHinduced locomotor responses at 20 mg/kg in DAT-tg mice. We also found no change in MPH-induced locomotor activity at the 20 mg/kg dose, however, the dose was on the descending limb of the inverted U shaped dose-response curve dose-response curve, and at lower doses, MPH-induced locomotion was enhanced in DAT-tg mice. 8. Methylphenidate is Unique in the Way that it Interacts with the DAT As discussed previously, MPH is traditional classified as a blocker, as cell culture studies have demonstrated that it is not transported into the cell via the DAT, and thus cannot interact with vesicles and cause active release of DA, as has been observed with releasers (Sonders et al., 1997). Even though MPH is categorized as a blocker, it has been shown to have releasing capabilities at 0.1mM, suggesting that MPH may have
323 302 properties of blocker and releaser compounds (Sproson et al., 2001; Russell et al., 1998; Heal et al., 1996). Although MPH has been suggested to have release capabilities, within this work (discussed in chapters 7 & 8) we show that MPH exhibits the release profile of a blocker, which is characterized by increased DA release with increasing concentrations. This demonstrates that although MPH classes with releasers in regards to its potency effects following MPH and cocaine self-administration, it is not a pure DA releaser, but rather exhibits properties of both classes of psychostimulants. Since MPH is not a substrate, like the prototypical blocker cocaine, yet classes with releasers in its acute effects at the DAT following cocaine and MPH self-administration, MPH may function as an atypical DAT inhibitor, exhibiting pharmacological characteristics similar to AMPH as well as COC via a number of mechanisms. In regards to effects at the DAT, MPH actions seem to resemble those of DA releasers, even though it is traditionally categorized as a DAT blocker. A number of other studies have found similar results in regards to DAT binding as well as following cocaine self-administration where MPH seems to group with releasers (Dar et al., 2005; Ferris et al., 2011, 2012; Wayment et al., 1999). Because MPH has been shown to promote reverse transport of DA at high doses (Russel et al., 1998; Dyck et al., 1980) it is possible that these effects are responsible for its classification as a releaser in these studies. However, as mentioned previously MPH is not transported into the cell (Sonders et al., 1997), therefore its releasing properties must act through different mechanisms than traditional releasers such as AMPH. These release capabilities, independent of being transported into the cell, could be due to a number of factors. 1) MPH binds to the
324 303 DAT in a manner that is distinct from prototypical blockers or releasers (Dar et al., 2005; Wayment et al., 1999), with significant overlap between the binding sites for both cocaine and AMPH. It is possible that, although MPH is not transported into the cell, the interaction with the AMPH binding site on the DAT results in conformational changes in the DAT which promote reverse transport in the same way as AMPH. 2) MPH has been demonstrated to result in the mobilization of vesicles into the releasable pool. It is possible that this effect of MPH is dependent on the DAT, thus when there are increased DAT levels, MPH is better able to induce exocytotic release from vesicles (Volz et al., 2007, 2009; Sandoval et al., 2002; Chadchankar et al., 2012). 9. Clinical Relevance This work has revealed an important, novel concept, whereby increased DAT levels, such as those documented in individuals suffering from ADHD (Spencer et al., 2012), lead to an increase in potency of DA releasers such as AMPH. Many of these individuals, as is the case with ADHD, are treated with stimulant drugs, making it critically important to understand the effects that these drugs have in the case of elevated DAT levels in order to minimize abuse liability, neurotoxicity, and negative side effects. Additionally, models of PTSD (Hoexter et al., 2012), early life stress (Yorgason et al., 2012), and repeated stress (Kohut et al., 2012) all result in similar increases in DAT levels; therefore, many individuals fall into this at risk category, making these findings relevant to a large population that extends far beyond the drug abuse field.
325 304 In regards to drug abuse, in the last SAMHSA survey, 4.8 million people reported abusing MPH, compared to 5.2 million for cocaine. While these reports are already alarming, they rely on the self-reporting of abuse of these compounds and as a result the numbers could be an underestimation for what is actually occurring in the human population. Given the high prevalence of MPH abuse, it is important to determine the neurochemical and behavioral consequences in pre-clinical models. There are numerous reports in humans of co-abuse of MPH with a number of other compounds including AMPH and cocaine (Barrett et al., 2005; Teter et al., 2006; Wilens et al., 2008), and here we show that prior MPH abuse may sensitize releaser reinforcement. This sensitized reinforcement may lead to an increased propensity for abuse of releaser compounds, such as AMPH and methamphetamine. 10. Future Directions Future work should focus on finding the mechanism for differential alterations in DAT levels/uptake rates and stimulant potency caused by MPH and cocaine selfadministration. Functional regulation of the DAT protein occurs by a number of processes including phosphorylation, glycosylation, palmitoylation, and oligomerization of the DAT protein itself, as well as interactions with other presynaptic receptor subtypes such as D2 autoreceptors (Ferris et al., 2013). Discussed below are some potential mechanisms for drug-induced DAT trafficking changes following MPH selfadministration and the DAT-trafficking-independent tolerance caused by cocaine selfadministration.
326 Cocaine Self-Administration Mechanisms N-Terminal Interactions Both the N and C terminals of the DAT are cytoplasmic, and various regions of both of these domains are involved in protein interactions that are capable of modifying the function of the transporter (Giros and Caron, 1993). Of particular importance are the N-terminal and the first transmembrane domain, which is extremely important for the binding of transporter substrates such as DA. A large body of research exists concerning the modulation of the DAT at the N-terminus by phosphorylation and interaction with intracellular signaling molecules (Chen et al., 2010; Giros and Caron, 1993; Guptaroy et al., 2008, 2010). The N-terminus of the DAT contains phosphorylation sites for a number of kinases including protein kinase C (PKC), protein kinase A (PKA), and Ca2+/Calmodulin protein kinase II, and is also important for substrate binding (Giros and Caron, 1993). A number of studies have used human embryonic kidney (HEK) cells stably transfected with various mutated forms of the human DAT (hdat) to study the role that the N terminal threonine residue (Thr62) has on the functionality of the transporter (Guptaroy et al., 2008, 2010). One of the mutants, T62D-hDAT (threonine to aspartic acid), was particularly interesting, in regards to cocaine self-administration effects. In line with the compensatory effects that occur following cocaine selfadministration the T62D mutant had reduced surface expression as well as reduced uptake rates (Guptaroy et al. 2008). Further, the T62D-hDAT also had a reduced potency
327 306 of cocaine to inhibit [ 3 H]-DA uptake compared to hdat; however, their efficacy was unaffected as they were able to completely block DA uptake. This suggests a large role for Thr62 in transporter function and psychostimulant action. It is possible that following cocaine self-administration, modifications to this region of the DAT are responsible for the reduced cocaine effects as well as the other dopaminergic changes that have been observed Dopamine Transporter Complexes: Oligomers and D2 Interactions It has been demonstrated that the DAT forms oligomer complexes with itself and other proteins such as D2 autoreceptors (Chen & Reith 2008; Hadlock et al., 2010). Further, it has been suggested that the oligomerization of DAT proteins could lead to changes in uptake rates, where by the complexes have reduced uptake function than the same number of monomer DAT. It is possible that DAT complex formation reduces the potency of cocaine to inhibit DA uptake. In line with this, it is possible that cocaine selfadministration is resulting in DAT complex formation, an effect that leads to reduced uptake and reduced cocaine potency. This can be easily tested and would be a powerful mechanism to explain the reductions in cocaine potency that occur following cocaine self-administration. Western blot hybridization can be utilized to determine the formation of higher-order complexes; however, it is difficult using western blot to determine what each complex is comprised of. Pull-down assays can be used to assess the changes in the interaction of D2 receptors with the DAT to determine if the complexes contain D2 autoreceptors. Determining if
328 307 these complexes are responsible for the changes associated with cocaine selfadministration and what proteins are within the complex will allow new insights into the neural mechanisms driving the addiction process Methylphenidate Self-Administration Mechanisms Substrate-Induced Rapid Trafficking of DAT Because of the structural similarities of MPH and AMPH, it is possible that MPH is increasing DAT levels via some of the same mechanisms as AMPH. AMPH has been shown to rapidly increase surface expression of DAT (Johnson et al., 2005; Furman et al., 2009). AMPH-induced changes in DAT surface expression happened almost instantaneously with significant increases occurring after only ten seconds. Using reversible biotinylation it was determined that AMPH increases surface expression by promoting insertion of transporters into the membrane, and not by delaying the endocytosis of transporters already present in the membrane (Johnson et al., 2005; Furman et al., 2009). The effect on DAT trafficking in these studies was brief with the increased surface expression lasting only 2.5 minutes. However, these studies were done in cell culture and it is possible that the effects would be more sustained in an in vivo model. Although the mechanism for this effect is unclear, it is possible that MPH is upregulating DAT levels via a similar mechanism, an effect that is a promising avenue for exploration.
329 D2-DAT Interactions Another possible mechanism for the upregulation of DAT function/number following MPH is through D2 autoreceptors. D2 receptors are located presynaptically on DA nerve terminals and function to inhibit DA release and synthesis in order to reduce synaptic DA levels. Additionally, it has been demonstrated that activation of D2/D3 type autoreceptors can cause rapid DAT toward the cell surface an increases in uptake rates (Cass & Gerhard, 1994; Thompson et al., 2001 a, b). The DAT and D2 interact through direct physical interactions at N-terminus and the third intracellular loop of the D2 autoreceptor (Thompson et al., 2001b), which has been hypothesized to facilitate intracellular DAT trafficking to the surface. The disruption of this interaction in vivo results in decreased striatal synaptosomal DA uptake and increased locomotor activity, mimicking DAT-knockout mice (Thompson et al., 2001b), indicating that this interaction is essential for DAT function. Further, voltammetry studies have shown that D2 agonists increase the rate of uptake (V max ) (Meiergerd et al., 1993; Batchelor and Schenk, 1998; Dickinson et al., 1999). V max is often regulated by DAT levels, thus it is likely that the increased V max is due to increases in DAT surface expression. Because D2 activation leads to increases in DAT surface expression and V max, it is possible that MPH-induced activation of D2 autoreceptors is leading to the increasing DAT levels following MPH self-administration. Because MPH is structurally similar to both AMPH and DA it is possible that MPH has binding properties at the D2 receptor. However, it has been demonstrated that at physiologically relevant concentrations that MPH only exhibits limited affinity for D2 receptors (Gatley et al., 1996). Therefore, if D2 receptor activation is the mechanism of the upregulated DAT levels it is likely that it is
330 309 due to MPH-induced elevations in DA levels. However, because cocaine, which increases DA to a similar extent, causes opposite changes this suggests that either elevations in DA does not mediate this effect, or for cocaine there is another competing mechanism causing the decreases in DAT expression observed following cocaine selfadministration. It is possible that MPH is activating some of the signaling cascades originating from the D2 receptor, which leads to these effects. D2 receptors signal via G proteins to a number of downstream effectors, including adenylyl cyclase, ERK1/2, and the PI3k cascade via phosphorylation of Akt (Neve et al., 2004; Brami-Cherrier et al., 2002; Beom et al., 2004; Burgering and Coffer, 1995; Brami-Cherrier et al., 2002; Beaulieu et al., 2005). Recent work has shown that the PI3K pathway (Carvelli et al., 2002) and the mitogen-activated protein kinase pathway (MAPK; Moron et al., 2003) modulate DAT function by altering cell surface expression, suggesting that if MPH was modulating this pathway that these effects could occur. Indeed, Pascoli et al., (2005) showed that MPH increased ERK phosphorylation. Increasing ERK1 and ERK2 phosphorylation affects DAT function by increasing surface DAT expression, which is consistent with MPH increasing DAT levels via this pathway (Parnas & Vaughn, 2008; Morón et al., 2003; Schmitt & Reith, 2010). Therefore it is possible that MPH self-administration results in increased activity of the MAPK pathway containing ERK 1 and ERK 2, which leads to increased DAT expression at the surface. However, conversely, Mines and Jope (2012) show that after eight days of methylphenidate treatment, striatal Akt was dephosphorylated. Although this is consistent with D2 activation, which has been shown to reduce Akt levels (Beaulieu et
331 310 al., 2005, 2008), reductions in Akt levels/phosphorylation would likely lead to reduced surface expression. It is also possible that immediately following or during the MPH selfadministration session that Akt levels/phosphorylation are reduced, and following acute withdrawal the levels increase, increasing DAT levels, in order to compensate for the sustained reduction. Within this work, changes following MPH self-administration were only assessed at one time point 18hrs ( following the commencement of the final session), thus it is unclear as to what changes occur at other points of withdrawal. A time course of DAT changes following MPH self-administration would elucidate this and could determine if this is a potential mechanism Determine Regional Specificity of Effects Although the nucleus accumbens (NAc) core plays an integral role in reinforcement behavior (Roberts et al., 1977), it is possible that other striatal brain regions may mediate different aspects of MPH self-administration, such as reinstatement responding, especially after long periods of exposure. The dorsal striatum has been shown to mediate cocaine seeking following abstinence, whereby, inactivation of the dorsal striatum reduces cocaine seeking, and inactivation of the ventral striatum has no effect (See et al., 2007). In humans, presentation of drug-paired cues to cocaine addicts increases DA activity in the dorsal striatum, which is correlated with the intensity of cueinduced craving, suggesting that the dorsal striatum may play a role in relapse in humans as well as cue-induced reinstatement in rodents (Volkow et al., 2006; Wong et al., 2006). Because of this, it is possible that the ventral striatum is mediating the
332 311 rewarding/reinforcing properties of drugs during and immediately following selfadministration paradigms, such as what was seen in this work with MPH selfadministration. However, following abstinence, it is possible that the effects are driven by changes in the dorsal striatum. This is particularly important for drugs like MPH, which are taken intermittently, as there are repeated abstinence periods that occur between exposures. It will be important to determine the changes that occur during different periods of abstinence and how these neurochemical changes in the ventral versus dorsal stratum differentially mediate reinstatement and reinforcement behaviors. The dorsolateral striatum is thought to control well-established cue-controlled cocaine seeking, and it mediates habit learning, specifically the association of motor outputs with reinforcement, such as an animal responding on a lever for drug (Everitt and Robbins, 2005; Pierce and Vanderschuren, 2010; Ito et al., 2000, 2002; Vanderschuren et al., 2005; Belin and Everitt, 2008; Packard and Knowlton, 2002; Yin and Knowlton, 2006). This region plays a role in drug seeking following repeated self-administration sessions, especially when drug administration becomes habitual and stimulus driven (Everitt and Robbins, 2005; Pierce and Vanderschuren, 2010). Because of the importance of the dorsolateral striatum in drug-seeking and habitual responding, two behaviors that are integral to the addiction process, it will be important to determine the effects of MPH self-administration both neurochemically and behaviorally in this region. This can be done pairing voltammetry with reinstatement studies as well as inactivation studies and studies determining the functional activity of specific brain regions following self-administration, abstinence, and reinstatement in rodents.
333 312 Additionally, it has been shown with cocaine self-administration in monkeys in regards to the DA system, that in the early stages of cocaine self-administration the ventral striatum is most active, while over time the activity shifts to include the dorsal striatum (Porrino et al., 2007; Moore et al., 1998; Letchworth et al., 2001; Nader et al., 2002; Porrino et al., 2004). It is possible that the same changes in drug potency occur in all of the striatal regions following MPH self-administration; however, that the changes in the more dorsal regions of the striatum are what is really driving the reinforcement behavior. Inactivation studies could be done in order to determine the contribution of these regions to threshold responding following MPH self-administration MPH Allows for the Determination of the Relationship Between Tolerance and the Development of Addictive-Like Behaviors. Because MPH and cocaine are virtually identical in their behavioral selfadministration profile in regards to FR1 self-administration (Calipari et al., 2013a, b, c, d, e), but cause sensitization and tolerance, respectively, studies that compare these drugs provide unique insight as to the contribution of tolerance to the drug addiction process. Although tolerance to the euphorigenic and neurochemical effects of cocaine is often seen in cocaine addicts and abusers, it is unclear if this tolerance is a cause or consequence of the addiction process. It is possible that tolerance to the effects of cocaine are a byproduct of repeated cocaine self-administration, however, it is also possible that tolerance is driving many of the behavioral effects that occur during the addiction process, such as increased motivation to administer drug. In animals the
334 313 addiction process has been modeled using a variety of techniques, including measures of reinforcing efficacy such as progressive ratio (PR) paradigms. Although the PR paradigm of self-administration is thought to assess reinforcing efficacy it is possible that it is not assessing solely that. It is currently accepted that neurochemical tolerance to a compound would result in reduced responding on a PR schedule, and decrease breakpoints. Because of this much literature has used increases in PR to suggest that animals are not tolerant to the effects of cocaine following long-access cocaine self-administration (Paterson and Markou, 2003; Wee et al., 2008). However, contrary to this theory, Lack et al., (2008) demonstrate that tolerance to the locomotor activating effects of cocaine occur following escalation on a PR schedule. Therefore, increased breakpoints are associated with tolerance to cocaine s effects behaviorally. Additionally, because locomotor activity is mediated by the DA system, it is likely that these animals are tolerant to the effects of cocaine neurochemically, a phenomenon that has been shown repeatedly following a number of cocaine self-administration paradigms (Hurd et al., 1989; Calipari et al., 2013b, c, d, e; Ferris et al., 2011; Mateo et al., 2005). The threshold procedure of reinforcement (Oleson et al., 2009, 2011, 2012) provides a perfect avenue to study the effects of neurochemical tolerance and sensitization on reinforcement behavior. Although the threshold procedure can still show information about reinforcing efficacy, where the P max is positively correlated with PR, it also allows for the assessment of other factors. For example, while Zimmer et al., (2012) demonstrate that the P max for cocaine is increased following long-access cocaine selfadministration, Oleson et al., (2009) show that intake is only increased at high doses where price is low and is actually decreased at doses where animals have to expend more
335 314 effort to obtain drug, indicating tolerance. Taking advantage of the threshold procedure following MPH and cocaine self-administration may allow for the determination of how tolerance and sensitization change different aspects of drug reinforcement. 11. Conclusions Taken together, this work provides important information, not only about the consequences of MPH and cocaine self-administration on the DA system, but also about how compensatory effects of psychostimulant self-administration occur at the DAT following repeated drug administration. These data have implications for not only the DAT, but also all other transporters in the solute carrier 6 (SLC6; also known as: neurotransmitter-sodium-symporter family or Na(+)/Cl(-)-dependent transporters) family of transporters (Kristensen et al., 2011). These transporters include transporters for serotonin, norepinephrine, GABA and glycine, and it is likely that the compensatory changes that occur at these transporters do so in a similar fashion. Further, these data highlight the importance in continuing work on the neurochemical consequences of high-dose MPH self-administration. The neurochemical changes that occur following MPH self-administration suggest that individuals that abuse MPH may be more susceptible to the abuse and addiction of a number of highly addictive psychostimulants. Given the high prescription rates of MPH, it is readily available, and highly abused, making studies highlighting the consequences of its misuse imperative.
336 315 And finally, this work elucidates some of the factors that influence the compensatory changes that occur at the DAT following cocaine self-administration. Because intake and pattern play a prominent role in determining the neurochemical adaptations that occur, it is important to choose a model that accurately mimics human administration of the compound.
337 316 References: Addy NA, Daberkow DP, Ford JN, Garris PA, Wightman RM. (2010) Sensitization of rapid dopamine signaling in the nucleus accumbens core and shell after repeated cocaine in rats. J Neurophysiol. 104(2): Aragona BJ, Cleaveland NA, Stuber GD, Day JJ, Carelli RM, Wightman RM. (2008) Preferential enhancement of dopamine transmission within the nucleus accumbens shell by cocaine is attributable to a direct increase in phasic dopamine release events. J Neurosci. 28: Batchelor M, Schenk JO. (1998) Protein kinase A activity may kinetically upregulate the striatal transporter for dopamine. J Neurosci. 18(24): Beaulieu JM, Marion S, Rodriguiz RM, Medvedev IO, Sotnikova TD, Ghisi V, Wetsel WC, Lefkowitz RJ, Gainetdinov RR, Caron MG. (2008) A beta-arrestin 2 signaling complex mediates lithium action on behavior. Cell. 132(1): Beaulieu JM, Sotnikova TD, Marion S, Lefkowitz RJ, Gainetdinov RR, Caron MG. (2005) An Akt/beta-arrestin 2/PP2A signaling complex mediates dopaminergic neurotransmission and behavior. Cell. 122(2): Belin D, Everitt BJ. (2008) Cocaine seeking habits depend upon dopamine-dependent serial connectivity linking the ventral with the dorsal striatum. Neuron. 57: Beom S, Cheong D, Torres G, Caron MG, Kim KM. (2004) Comparative studies of molecular mechanisms of dopamine D2 and D3 receptors for the activation of extracellular signal-regulated kinase. J Biol Chem. 279(27): Berridge CW, Devilbiss DM, Andrzejewski ME, Arnsten AF, Kelley AE, Schmeichel B, Hamilton C, Spencer RC. (2006) Methylphenidate preferentially increases catecholamine neurotransmission within the prefrontal cortex at low doses that enhance cognitive function. Biol Psychiatry. 60; Brami-Cherrier K, Valjent E, Garcia M, Pagès C, Hipskind RA, Caboche J. (2002) Dopamine induces a PI3-kinase-independent activation of Akt in striatal neurons: a new route to camp response element-binding protein phosphorylation. J Neurosci. 22(20): Burgering BM, Coffer PJ. (1995) Protein kinase B (c-akt) in phosphatidylinositol-3-oh kinase signal transduction. Nature. 376(6541): Calipari ES, Ferris MJ, Salahpour A, Caron MG, Roberts DCS, Jones SR (2013a) Methylphenidate amplifies the potency and reinforcing effects of amphetamines by increasing dopamine transporter expression Calipari ES, Ferris MJ, Melchior JR, Bermejo K, Salahpour A, Roberts DC et al (2013b). Methylphenidate and cocaine self-administration produce distinct dopamine terminal alterations. Addict Biol (in press).
338 317 Calipari ES, Ferris MJ, Zimmer BA, Roberts DC, Jones SR. (2013c) Temporal Pattern of Cocaine Intake Determines Tolerance vs Sensitization of Cocaine Effects at the Dopamine Transporter. Neuropsychopharmacology. In press. Calipari ES, Beveridge TJ, Jones SR, Porrino LJ (2013d) Withdrawal from extended access cocaine self-administration results in dysregulated functional activity and altered locomotor activity in rats. Eur. J. Neurosci. In press. Calipari ES, Ferris MJ, Jones SR (2013e) Extended access cocaine self-administration results in toleramce to the dopamine-elevating and locomotor-stimulating effects of cocaine. J. Neurochem. In press. Calipari ES, Ferris MJ. (2013) Amphetamine mechanisms and actions at the dopamine terminal revisited. J Neurosci. 33: Carvelli L, Morón JA, Kahlig KM, Ferrer JV, Sen N, Lechleiter JD, Leeb-Lundberg LM, Merrill G, Lafer EM, Ballou LM, Shippenberg TS, Javitch JA, Lin RZ, Galli A. (2002) PI 3-kinase regulation of dopamine uptake. J Neurochem. 81(4): Cass WA, Gerhardt GA. (1994) Direct in vivo evidence that D2 dopamine receptors can modulate dopamine uptake. Neurosci Lett. 176(2):259. Chadchankar H, Ihalainen J, Tanila H, Yavich L. (2012) Methylphenidate modifies overflow and presynaptic compartmentalization of dopamine via an α-synucleindependent mechanism. J Pharmacol Exp Ther. 341(2): Chen N, Reith ME. (2007) Substrates and inhibitors display different sensitivity to expression level of the dopamine transporter in heterologously expressing cells. J Neurochem. 101(2): Chen R, Furman CA, Gnegy ME. (2010) Dopamine transporter trafficking: rapid response on demand. Future Neurol. 5(1):123. Cragg SJ. (2006)Meaningful silences: how dopamine listens to the ACh pause. Trends Neurosci. 29: Dar DE, Mayo C, Uhl GR. (2005) The interaction of methylphenidate and benztropine with the dopamine transporter is different than other substrates and ligands. Biochem Pharmacol. 70, Devilbiss DM, Berridge CW. (2006) Low-dose methylphenidate actions on tonic and phasic locus coeruleus discharge. J Pharmacol Exp Ther. 319; Devilbiss DM, Berridge CW. (2008) Cognition-enhancing doses of methylphenidate preferentially increase prefrontal cortex neuronal responsiveness. Biol Psychiatry. 64; Dickinson SD, Sabeti J, Larson GA, Giardina K, Rubinstein M, Kelly MA, Grandy DK, Low MJ, Gerhardt GA, Zahniser NR. (1999) Dopamine D2 receptor-deficient mice
339 318 exhibit decreased dopamine transporter function but no changes in dopamine release in dorsal striatum. J Neurochem. 72(1): dopamine in the nucleus accumbens. Synapse 5: Dyck LE, Boulton AA, Jones RS. (1980) A comparison of the effects of methylphenidate and amphetamine on the simultaneous release of radiolabelled dopamine and p- or m- tyramine from rat striatal slices. Eur J Pharmacol. 68(1): Everitt BJ, Robbins TW. (2005) Neural systems of reinforcement for drug addiction: from actions to habits to compulsion. Nat Neurosci. 8: Ferris MJ, Calipari ES, Mateo Y, Melchior JR, Roberts DC, Jones SR (2012). Cocaine self-administration produces pharmacodynamic tolerance: differential effects on the potency of dopamine transporter blockers, releasers, and methylphenidate. Neuropsychopharmacology 37: Ferris MJ, Calipari ES, Melchior JR, Roberts DC, España RA, Jones SR. (2013) Paradoxical tolerance to cocaine after initial supersensitivity in drug-use-prone animals. Eur J Neurosci. In press. Ferris MJ, Calipari ES, Yorgason JT, Jones SR. (2013) Examining the complex regulation and drug-induced plasticity of dopamine release and uptake using voltammetry in brain slices. ACS Chem Neurosci. 4(5): Ferris MJ, Mateo Y, Roberts DC, Jones SR (2011) Cocaine-insensitive dopamine transporters with intact substrate transport produced by self-administration. Biol Psychiatry 69(3): Furman CA, Chen R, Guptaroy B, Zhang M, Holz RW, Gnegy M. (2009) Dopamine and amphetamine rapidly increase dopamine transporter trafficking to the surface: live-cell imaging using total internal reflection fluorescence microscopy. J Neurosci. 29(10): Furman CA, Lo CB, Stokes S, Esteban JA, Gnegy ME. (2009) Rab 11 regulates constitutive dopamine transporter trafficking and function in N2A neuroblastoma cells. Neurosci Lett. 463(1): Gatley SJ, Pan D, Chen R, Chaturvedi G, Ding YS. (1996) Affinities of methylphenidate derivatives for dopamine, norepinephrine and serotonin transporters. Life Sci. 58(12):231-9.
340 319 Gill KE, Pierre PJ, Daunais J, Bennett AJ, Martelle S, Gage HD, Swanson JM, Nader MA, Porrino LJ. (2012) Chronic Treatment with Extended Release Methylphenidate Does Not Alter Dopamine Systems or Increase Vulnerability for Cocaine Self- Administration: A Study in Nonhuman Primates. Neuropsychopharmacology. In press. Giros B, Caron MG. (1993) Molecular characterization of the dopamine transporter. Trends Pharmacol Sci. 2:43-9. Griggs R, Weir C, Wayman W, Koeltzow TE. (2010) Intermittent methylphenidate during adolescent development produces locomotor hyperactivity and an enhanced response to cocaine compared to continuous treatment in rats. Pharmacol Biochem Behav. 96(2): Guptaroy B, Fraser R, Desai A, Zhang M, Gnegy ME. (2010) Site directed mutations near transmembrane domain 1 (TM1) alter conformation and function of norepinephrine and dopamine transporters. Mol Pharmacol. 79(3):5ss20-32 Guptaroy B, Zhang M, Bowton E, Binda F, Shi L, Weinstein H, Galli A, Javitch JA, Neubig RR, Gnegy ME. (2009) A juxtamembrane mutation in the N terminus of the dopamine transporter induces preference for an inward-facing conformation. Mol Pharmacol. 75(3): Hadlock GC, Chu PW, Walters ET, Hanson GR, Fleckenstein AE. (2010) Methamphetamine-induced dopamine transporter complex formation and dopaminergic deficits: the role of D2 receptor activation. J Pharmacol Exp Ther.335(1): Han DD, Gu HH (2006) Comparison of the monoamine transporters from human and mouse in their sensitivities to psychostimulant drugs. BMC Pharmacol. 3(6):6. Heal DJ, Cheetham SC, Smith SL (2009) The neuropharmacology of ADHD drugs in vivo: insights on efficacy and safety. Neuropharmacology 57(7-8): Heal DJ, Prow MR, Hearson M, Buckett WR (1996) Efflux of [3H]dopamine from superfused rat striatal slices: predictive value for detecting stimulant drugs of abuse. Br. J. Pharmacol 117:325P. Humphreys KL, Eng T, Lee SS. (2012) Stimulant Medication and Substance Use Outcomes: A Meta-analysis. JAMA Psychiatry. 29:1-9. Hurd YL, Weiss F, Koob GF, And NE, Ungerstedt U. (1989) Cocaine reinforcement and extracellular dopamine overflow in rat nucleus accumbens: an in vivo microdialysis study. Brain Res. 498(1): Inada T, Polk K, Purser C, Hume A, Hoskins B, Ho IK, Rockhold RW. (1992) Behavioral and neurochemical effects of continuous infusion of cocaine in rats. Neuropharmacology. 31(7): Ito R, Dalley JW, Howes SR, Robbins TW, Everitt BJ. (2000) Dissociation in conditioned dopamine release in the nucleus accumbens core and shell in response to cocaine cues and during cocaine-seeking behavior in rats. J Neurosci. 20:
341 320 Ito R, Dalley JW, Robbins TW, Everitt BJ. (2002) Dopamine release in the dorsal striatum during cocaine-seeking behavior under the control of a drug-associated cue. J Neurosci. 22: John CE, Jones SR. (2007) Voltammetric characterization of the effect of monoamine uptake inhibitors and releasers on dopamine and serotonin uptake in mouse caudateputamen and substantia nigra slices. Neuropharmacology. 52(8): Jones SR, Lee TH, Wightman RM, Ellinwood EH. (1996) Effects of intermittent and continuous cocaine administration on dopamine release and uptake regulation in the striatum: in vitro voltammetric assessment. Psychopharmacology (Berl). 126(4): Kalivas PW, Duffy P (1993). Time course of extracellular dopamine and behavioral sensitization to cocaine. I. Dopamine axon terminals. J Neurosci 13: King GR, Xiong Z, Ellinwood EH Jr. (1998) Blockade of the expression of sensitization and tolerance by ondansetron, a 5-HT3 receptor antagonist, administered during withdrawal from intermittent and continuous cocaine. Psychopharmacology (Berl). 135(3): Kollins SH, Rush CR, Pazzaglia PJ, Ali JA. (1998) Comparison of acute behavioral effects of sustained-release and immediate-release methylphenidate. Exp Clin Psychopharmacol. 6(4): Kollins SH. (2003) Comparing the abuse potential of methylphenidate versus other stimulants: a review of available evidence and relevance to the ADHD patient. J Clin Psychiatry. 64 Suppl 11:14-8. Kristensen AS, Andersen J, Jørgensen TN, Sørensen L, Eriksen J, Loland CJ, Strømgaard K, Gether U. (2011) SLC6 neurotransmitter transporters: structure, function, and regulation. Pharmacol Rev. 63(3): Letchworth SR, Nader MA, Smith HR, Friedman DP, Porrino LJ. (2001) Progression of changes in dopamine transporter binding site density as a result of cocaine selfadministration in rhesus monkeys. J Neurosci. 21: Madras BK, Miller GM, Fischman AJ. (2005) The dopamine transporter and attentiondeficit/hyperactivity disorder. Biol Psychiatry. 57, Mateo Y, Lack CM, Morgan D, Roberts DC, Jones SR (2005) Reduced dopamine terminal function and insensitivity to cocaine following cocaine binge self-administration and deprivation. Neuropsychopharm 30(8): Meiergerd SM, Patterson TA, Schenk JO. (1993) D2 receptors may modulate the function of the striatal transporter for dopamine: kinetic evidence from studies in vitro and in vivo. J Neurochem. 61(2): Moore RJ, Vinsant SL, Nader MA, Porrino LJ, Friedman DP. (1998) Effect of cocaine self-administration on dopamine D2 receptors in rhesus monkeys. Synapse. 30:88 96.
342 321 Morón JA, Zakharova I, Ferrer JV, Merrill GA, Hope B, Lafer EM, Lin ZC, Wang JB, Javitch JA, Galli A, Shippenberg TS. (2003) Mitogen-activated protein kinase regulates dopamine transporter surface expression and dopamine transport capacity. J Neurosci. 23(24): Morón, J.A., I. Zakharova, J.V. Ferrer, et al Mitogen-activated protein kinase regulates dopamine transporter surface expression and dopamine transport capacity. J. Neurosci. 23: Nader MA, Daunais JB, Moore T, Nader SH, Moore RJ, Smith HR, Friedman DP, Porrino LJ. (2002) Effects of cocaine self-administration on striatal dopamine systems in rhesus monkeys: initial and chronic exposure. Neuropsychopharmacology. 27: Neve KA, Seamans JK, Trantham-Davidson H. (2004) Dopamine receptor signaling. J Recept Signal Transduct Res. 24(3): Oleson EB, Richardson JM, Roberts DC. (2011) A novel IV cocaine self-administration procedure in rats: differential effects of dopamine, serotonin, and GABA drug pretreatments on cocaine consumption and maximal price paid. Psychopharmacology (Berl). 214(2): Oleson EB, Roberts DC. (2009) Behavioral economic assessment of price and cocaine consumption following self-administration histories that produce escalation of either final ratios or intake. Neuropsychopharmacology. 34(3): Oleson EB, Roberts DC. (2012) Cocaine self-administration in rats: threshold procedures. Methods Mol Biol. 829: Packard MG, Knowlton BJ. (2002) Learning and memory functions of the basal ganglia. Annu Rev Neurosci. 25: Parasrampuria DA, Schoedel KA, Schuller R, Silber SA, Ciccone PE, Gu J, Sellers EM. (2007) Do formulation differences alter abuse liability of methylphenidate? A placebocontrolled, randomized, double-blind, crossover study in recreational drug users. J Clin Psychopharmacol. 27(5): Parnas, M.L. & R.A. Vaughan Molecular structure and composition of dopamine transporters. In Dopamine Transporters: Chemistry, Biology and Pharmacology. M.L.Trudell & S.Izenwasser, Eds.: Wiley. Hoboken, NJ. Parsons LH, Justice JBJr (1993). Serotonin and dopamine sensitization in the nucleus accumbens, ventral tegmental area, and dorsal raphe nucleus following repeated cocaine administration. J Neurochem 61: Paterson NE, Markou A (2003). Increased motivation for self-administered cocaine after escalated cocaine intake. Neuroreport 14: Pierce RC, Vanderschuren LJMJ. Kicking the habit: the neural basis of ingrained behaviors in cocaine addiction. (2010) Neurosci Biobehav Rev. 35:
343 322 Porrino LJ, Lyons D, Smith HR, Daunais JB, Nader MA. (2004) Cocaine selfadministration produces a progressive involvement of limbic, association, and sensorimotor striatal domains. J Neurosci. 24: Porrino LJ, Smith HR, Nader MA, Beveridge TJ. (2007) The effects of cocaine: a shifting target over the course of addiction. Prog Neuropsychopharmacol Biol Psychiatry. 31(8): Ramamoorthy S, Samuvel DJ, Balasubramaniam A, See RE, Jayanthi LD (2010) Altered dopamine transporter function and phosphorylation following chronic cocaine selfadministration and extinction in rats. Biochem Biophys Res Commun. 391(3): Roberts DC, Corcoran ME, Fibiger HC. (1977) On the role of ascending catecholaminergic systems in intravenous self-administration of cocaine. Pharmacol Biochem Behav. 6(6): Rothman RB, Baumann MH. (2003) Monoamine transporters and psychostimulant drugs. Eur J Pharmacol. 479(1-3): Rudnick G, Clark J. (1993) From synapse to vesicle: the reuptake and storage of biogenic amine neurotransmitters. Biochim Biophys Acta. 1144(3): Russell V, de Villiers A, Sagvolden T, Lamm M, Taljaard J (1998) Differences between electrically-, ritalin- and D-amphetamine-stimulated release of [3H]dopamine from brain slices suggest impaired vesicular storage of dopamine in an animal model of Attention- Deficit Hyperactivity Disorder. Behav Brain Res. 94(1): Salahpour A, Ramsey AJ, Medvedev IO, Kile B, Sotnikova TD, Holmstrand E, Ghisi V, Nicholls PJ, Wong L, Murphy K, Sesack SR, Wightman RM, Gainetdinov RR, Caron MG. (2008) Increased amphetamine-induced hyperactivity and reward in mice overexpressing the dopamine transporter. Proc Natl Acad Sci U S A (11): Sandoval V, Riddle EL, Hanson GR, Fleckenstein AE. (2002) Methylphenidate redistributes vesicular monoamine transporter-2: role of dopamine receptors. J Neurosci. 22(19): Schmitt KC, Reith ME. (2010) Regulation of the dopamine transporter: aspects relevant to psychostimulant drugs of abuse. Ann N Y Acad Sci. 1187: Schubiner H. (2005) Substance abuse in patients with attention-deficit hyperactivity disorder : therapeutic implications. CNS Drugs. 19(8): See RE, Elliott JC, Feltenstein MW. (2007) The role of dorsal vs ventral striatal pathways in cocaine-seeking behavior after prolonged abstinence in rats. Psychopharmacology (Berl). 194(3): Sepúlveda DR, Thomas LM, McCabe SE, Cranford JA, Boyd CJ, Teter CJ. (2011) Misuse of prescribed stimulant medication for ADHD and associated patterns of substance use: preliminary analysis among college students. J Pharm Pract. 24(6):
344 323 Sonders MS, Zhu SJ, Zahniser NR, Kavanaugh MP, Amara SG (1997) Multiple ionic conductances of the human dopamine transporter: the actions of dopamine and psychostimulants. J Neurosci 17(3): Sorkina T, Doolen S, Zahniser NR, Sorkin A. (2003) Oligomerization of dopamine transporters visualized in living cells by fluorescence resonance energy transfer microscopy. J Biol Chem 278: Sproson EJ, Chantrey J, Hollis C, Marsden CA, Fonel KC (2001) Effect of repeated methylphenidate administration on presynaptic dopamine and behaviour in young adult rats. J Psychopharmacol 15(2): Teter CJ, McCabe SE, LaGrange K, Cranford JA, Boyd CJ (2006) Illicit use of specific prescription stimulants among college students: prevalence, motives, and routes of administration. Pharmacotherapy 26(10): Thompson TL, Bridges S, Miller C. (2001a) Modulation of dopamine uptake in rat nucleus accumbens: effect of specific dopamine receptor antagonists and sigma ligands. Neurosci Lett. 312(3):169. Thompson TL, Bridges SR, Weirs WJ. (2001b) Alteration of dopamine transport in the striatum and nucleus accumbens of ovariectomized and estrogen-primed rats following N-(p-isothiocyanatophenethyl) spiperone (NIPS) treatment. Brain Res Bull. 54(6):631 Threlfell S, Cragg SJ. (2011) Dopamine signaling in dorsal versus ventral striatum: the dynamic role of cholinergic interneurons. Front Syst Neurosci. 5: 11 Uhl GR, Hall FS, Sora I. (2002) Cocaine, reward, movement and monoamine transporters. Mol Psychiatry. 7(1):21-6. Vanderschuren LJMJ, Di Ciano P, Everitt BJ. (2005) Involvement of the dorsal striatum in cue-controlled cocaine seeking. J Neurosci. 25: Vanderschuren LJMJ, Everitt BJ. (2005) Behavioral and neural mechanisms of compulsive drug seeking. Eur J Pharmacol. 526: Volkow ND, Fowler JS, Wang G, Ding Y, Gatley SJ. (2002) Mechanism of action of methylphenidate: insights from PET imaging studies. J Atten Disord 6 Suppl 1:S Volz TJ, Farnsworth SJ, Hanson GR, Fleckenstein AE. (2009) Methylphenidate-Induced Alterations in Synaptic Vesicle Trafficking and Activity: Functional Consequences and Therapeutic Implications. Ann N Y Acad Sci. 1139: Volz TJ, Farnsworth SJ, King JL, Riddle EL, Hanson GR, Fleckenstein AE. (2007) Methylphenidate administration alters vesicular monoamine transporter-2 function in cytoplasmic and membrane-associated vesicles. J Pharmacol Exp Ther. 323(2):
345 324 Wayment HK, Deutsch H, Schweri MM, Schenk JO. (1999) Effects of methylphenidate analogues on phenethylamine substrates for the striatal dopamine transporter: potential as amphetamine antagonists? J Neurochem. 72, Wee S, Mandyam CD, Lekic DM, Koob GF (2008). Alpha 1-noradrenergic system role in increased motivation for cocaine intake in rats with prolonged access. Eur Neuropsychopharmacol 18: Wilson JM, Kish SJ. (1996) The vesicular monoamine transporter, in contrast to the dopamine transporter, is not altered by chronic cocaine self-administration in the rat. J Neurosci. 16(10): Yin HH, Knowlton BJ. The role of the basal ganglia in habit formation. (2006) Nat Rev Neurosci. 7:
346 325 APPENDIX I CONSERVED DORSAL-VENTRAL GRADIENT OF DOPAMINE RELEASE AND UPTAKE RATE IN MICE, RATS, AND RHESUS MACAQUES Erin S. Calipari, Kimberly N. Huggins, Tiffany A. Mathews, and Sara. R. Jones The following manuscript was published in Neurochemistry International, volme 61(7), pages , 2012, and is reprinted with permission. Stylistic variations are due to the requirements of the journal. Erin S. Calipari performed all rat and mouse experiments, all data analysis, and prepared the manuscript. Dr. Tiffany Mathews performed all monkey voltammetry experiments. Drs. Kimberly N. Huggins and Sara R. Jones acted in an advisory and editorial capacity.
347 326 Abstract Although the vast majority of research on the dopamine system has been performed in rodents, and it is assumed that this work will inform us about the human condition, there have been very few direct comparisons of presynaptic dopamine terminal function across multiple species. Because it is difficult to query rapid sub-second dopamine signaling in humans using voltammetric methods, we chose to compare dopamine signals across multiple striatal subregions in slices from C57BL/6J mice, Sprague-Dawley rats and rhesus macaques. We found a dorsal to ventral gradient of dopamine uptake rates with highest levels in the dorsal striatum and lowest levels in the nucleus accumbens shell, which is conserved across species. In addition to uptake rates, there was also a dorsal to ventral, high to low, gradient in the magnitude of stimulated DA release observed in monkeys, mice, and rats. These data demonstrate that there is considerable functional homology across striatal regions in non-human primates and rodents, lending support to the use of rodents as model systems to study dopamine-related circuitry and disorders that are clinically relevant to the human population.
348 Introduction Because of the ethical limitations of conducting research in humans, model systems must be used to gather information about human disorders. Monkeys and rodents are both commonly used as model systems to study neurobiology, and there is a great deal of homology across various neural networks as well as the actions of drugs across species (Bradberry, 2008; Levey et al., 1993). Currently, monkeys are the closest models of human conditions, but rodents are commonly used to model processes such as neurodegenerative diseases, stress, anxiety and addiction, due to the ability to use more invasive and comprehensive techniques (Chesselet and Richter, 2011;Roberts et al., 2007; Silberman et al., 2010). Although it is obvious that humans and rats differ significantly, it is important to determine the translational capability of neurobiological studies in rodents by determining how well functional parameters in the rat and mouse brain are similar to non-human primates and humans. The dopamine (DA) system is dysregulated in many human disorders, including addiction, schizophrenia, Parkinson s disease, anxiety, attention deficit/hyperactivity disorder (ADHD) and many others (Carlsson, 1988; Jentsch et al., 2000; Goto et al., 2010). Thus, comparing the functionality of dopaminergic regions across species is vital for determining the translational validity of animal models of a multitude of human neurological conditions. There are some cross species comparisons using PET imaging and dopamine transporter (DAT) occupancy in humans, baboons and rats (Ding et al., 1997; Fowler et al., 1998), however, there is limited information on region specific DA kinetic parameters, such as release and uptake (Cragg et al., 2000, 2002). Anatomical distinctions within the striatum are generally similar in rodents and nonhuman primates;
349 328 however, there are some important differences. Striatal organization differs in rodents and primates, most notably with a lack of anatomical separation of caudate and putamen structures in rodents, marked by the internal capsule in primates. In the rodent brain these structures are not anatomically distinct and are referred to as the caudate-putamen (CPu); however the striatum of both primates and rodents contains the nucleus accumbens (Mitchell et al., 1999). The lack of completely homologous structures across species complicates the translation of results of rodent studies to humans; however, additional knowledge of functional homology could lead to better and more accurate translational studies. Although anatomically divergent, inputs into the striatum are largely conserved across species with the two major dopaminergic inputs being the mesolimbic and nigrostrial pathways from the A10 neurons of the ventral tegmental area (VTA) and the A9 neurons of the substantia nigra pars compacta, respectively (Zahm et al., 2011). These projections separate along a ventromedial-dorsolateral axis with ventromedial striatum predominately receiving A10 and dorsolateral striatum receiving A9 projections. Even more important is the functional homology across species with more dorsal regions of the striatum being associated with motor planning and execution, central regions with associative learning and directed attention, and ventral regions with emotional and reward-related processing (Lynd-Balta and Haber, 1994a,b; Reep et al., 2003; Kunzle 1975; Selemon and Goldman-Rakic 1985; Haber and McFarland 1999). Although neuroanatomical homology is convenient for comparisons across species, it is also important to determine functional homology, as changes in structure that do not result in changes in function are not as directly relevant to human behavior/disease processes.
350 329 This study was aimed at assessing functional homology of the DA system by measuring regional DA system parameters in the caudate and nucleus accumbens in three different species, monkeys (Rhesus Macaques), rats (Sprague-Dawley) and mice (C57BL/6J). Using fast-scan cyclic voltammetry, we measured release and uptake parameters for five specific regions in each species: dorsal CPu, medial CPu, ventral CPu, nucleus accumbens core, and nucleus accumbens shell. We found that there is a conserved gradient of both uptake and release with highest levels of both measures occurring in the dorsal caudate of all species, and lowest levels in the nucleus accumbens shell. This conservation of functionality across these species further validates the use of rodent models for translational studies of the DA system. 2. Materials and methods 2.1 Tissue preparation Rats and mice were sacrificed by decapitation and brains were rapidly removed and transferred into ice-cold, pre-oxygenated (95% O 2 /5% CO 2 ) artificial cerebral spinal fluid (acsf) consisting of (in mm): NaCl (126), KCl (2.5), NaH 2 PO 4 (1.2), CaCl 2 (2.4), MgCl 2 (1.2), NaHCO 3 (25), glucose (11), L-ascorbic acid (0.4) and ph was adjusted to 7.4. Rhesus macaques were anesthetized with ketamine (10 mg/kg, i.m.) and maintained at a deep surgical plane of anesthesia with pentobarbital. Transcardial perfusion (90 s) with cold modified Kreb s buffer (ph =7.4) was performed and brains were rapidly removed and cooled in pre-oxygenated acsf (Daunais et al., 2010). For all samples, the tissue was sectioned into 400 μm-thick coronal slices containing the striatum with a vibrating tissue slicer (Leica VT1000S, Vashaw Scientific, Norcross, GA). Brain slices
351 330 were transferred to a submersion recording chamber, perfused at 1 ml/min at 32 C with oxygenated acsf. 2.2 Fast Scan Cyclic Voltammetry After a 30-min equilibration period, a carbon fiber microelectrode ( 150 μm length, 7 μm radius) and a bipolar stimulating electrode were placed in close proximity to each other ( 100 μm) into the dorsal, medial, or ventral striatum, or the nucleus accumbens core or shell. DA was evoked by a single, rectangular, electrical pulse (300 μa, 2 ms) applied every 5 min. Extracellular DA was recorded every 100 ms using fast-scan cyclic voltammetry (Kennedy et al., 1992) by applying a triangular waveform ( 0.4 to +1.2 to 0.4V vs Ag/AgCl, 400 V/s). One recording was obtained per subregion in each animal (rhesus n = 5, mouse n = 3, rat n = 4).Once the extracellular DA response was stable the electrode was moved to the next subregion for recording, where the process was repeated. Immediately following the completion of each experiment, recording electrodes were calibrated by recording their response (in current; na) to 3 μm DA in acsf using a flow-injection system. To determine kinetic parameters, evoked levels of DA were modeled using Michaelis Menten kinetics, as a balance between release and uptake (Wightman et al., 1988). Michaelis Menten modeling of baseline DA signals provides parameters that describe the amount of DA released following stimulation (ie, the peak height of the signal) and the maximal rate of DA uptake (V max ). For baseline modeling, we followed standard voltammetric modeling procedures by setting the apparent K m value to 160 nm for each animal based on well-established research on the affinity of DA for the DAT
352 331 (Wu et al., 2001), whereas baseline V max values were allowed to vary as the maximal rate of DA uptake. All voltammetry data were collected and modeled using Demon Voltammetry and Analysis Software (Yorgason et al., 2011). 2.3 Statistics Baseline voltammetry data were compared across groups using one-way analysis of variance (ANOVA) using Graphpad statistical software (La Jolla, CA, USA). Data obtained from the five striatal areas was subjected to a two-way ANOVA, with species and brain region as the factors. When significant main effects were obtained (p < 0.05), differences between groups at each dose were tested using Bonferroni post hoc tests. Comparisons of release to V max ratios, as well as comparisons between species at specific brain regions were subjected to one-way ANOVA. 3. Results 3.1 Electrically stimulated DA release in the striatum follows a dorsal to ventral gradient. Fast scan cyclic voltammetry was used to assess DA system kinetics in five striatal subregions from C57BL/6J mice, Sprague-Dawley rats, and rhesus macaques (Fig. 1). Electrically stimulated DA release followed a similar pattern across the five measured regions in the three species. A two-way ANOVA revealed a significant difference by region (F 2,45 = 6.906, p < 0.01) in all species (Fig. 2 A, B). Bonferroni post hoc analysis determined that the dorsal caudate had the highest level of DA release and release was reduced in each region along a dorsal to ventral axis, with the nucleus accumbens shell displaying the lowest level of DA release (Fig 2B). In addition, there were significant differences by species (F 4,45 = 5.937, p<0.01), with mouse release
353 332 Figure 1. Location of voltammetric recordings in striatal slices from rat, mouse, and rhesus macaque. Five regions spanning the caudate putamen (CPu) and nucleus accumbens (NAc) were assessed across three species: Dorsolateral CPu (DC), medial CPu (MC), ventromedial CPu (VC), NAc core (core), and NAc shell (shell). (A) Diagram of a rat coronal section from Paxinos and Watson (2007) denoting the location where the recordings were made in each of the five regions, AP 1.32 (B) Diagram of a mouse coronal section adapted from Paxinos and Franklin (2007), AP 1.10 (C) Diagram of coronal section in rhesus macaque, AP AP anterior-posterior coordinates, Put putamen, IC-internal capsule.
 Mean peak release ± SEM evoked at five loci in each species showing an overall gradient of release from dorsal (highest) to ventral (lowest) that is conserved across species. *** p < 0.")
354 333 Figure 2. Dopamine (DA) release from five striatal regions in monkey, rat, and mouse. (A) Representative traces of the five striatal regions in monkey rat and mouse expressed as µm DA over time. (B) Mean peak release ± SEM evoked at five loci in each species showing an overall gradient of release from dorsal (highest) to ventral (lowest) that is conserved across species. *** p < across region; ### p < versus monkey; &&& p < versus rat.
355 334 measurements being significantly higher overall than both monkey and rat (Figure 2 A,B). 3.2 Maximal rate of DA uptake follows a dorsal to ventral gradient This study also assessed the rate of DA uptake following electrical stimulation and revealed a gradient of uptake across the striatal regions. There were significant differences by region (F 2,45 = 32.47, p < 0.001) which were present in all species (Fig 3A, B). Bonferroni post hoc analysis revealed the highest levels of uptake in the dorsal striatum and lower levels in a gradient from more dorsal to more ventral regions with the lowest level of uptake being present in the nucleus accumbens shell (Fig. 3B). In addition to a conserved gradient of uptake across species, there were differences in overall uptake rates between rat, mouse, and monkey (F 4,45 = 42.24, p < 0.001). Post hoc analysis revealed that rat and mouse rates of uptake were significantly higher than the uptake rates recorded for the monkeys (Fig. 3B). 3.3 Ratio of DA release to V max across species The ratio of DA release to V max was used to determine if there were differences in the regulation of presynaptic DA kinetics, across regions or species. There was a significantly lower ratio of release to uptake in rats in the dorsal caudate (F 2,6 = 10.60, p < 0.05) (Fig. 4C) as compared to both monkey and mice. This difference between species was maintained across all regions. In addition, there were no regional differences in the ratio of DA release to V max in any of the species tested, suggesting that this measure is inherent to the type of animal and not regionally variable.
 Representative traces of DA over time normalized for peak height.")
356 335 Figure 3. Maximal dopamine (DA) uptake rate (V max ) as measured by fast scan cyclic voltammetry from dorsal striatum through ventral striatum in each species. (A) Representative traces of DA over time normalized for peak height. (B) V max across five regions (dorsal caudate-putamen (CPu), medial CPu, ventral CPu, nucleus accumbens core, nucleus accumbens shell) showing a gradient of uptake rate in three species. V max was determined using Michaelis-Menten modeling to determine kinetic parameters. Mean peak Vmax (µm/s) ± SEM evoked at the five listed loci. *** p < across region; ### p < versus monkey.

357 336
MOLECULAR BIOLOGY OF DRUG ADDICTION. Sylvane Desrivières, SGDP Centre
 1 MOLECULAR BIOLOGY OF DRUG ADDICTION Sylvane Desrivières, SGDP Centre Reward 2 Humans, as well as other organisms engage in behaviours that are rewarding The pleasurable feelings provide positive reinforcement
1 MOLECULAR BIOLOGY OF DRUG ADDICTION Sylvane Desrivières, SGDP Centre Reward 2 Humans, as well as other organisms engage in behaviours that are rewarding The pleasurable feelings provide positive reinforcement
Methylphenidate amplifies the potency and reinforcing effects of amphetamines by increasing dopamine transporter expression
 Received 1 Aug 213 Accepted 7 Oct 213 Published 5 Nov 213 DOI: 1.138/ncomms372 Methylphenidate amplifies the potency and reinforcing effects of amphetamines by increasing dopamine transporter expression
Received 1 Aug 213 Accepted 7 Oct 213 Published 5 Nov 213 DOI: 1.138/ncomms372 Methylphenidate amplifies the potency and reinforcing effects of amphetamines by increasing dopamine transporter expression
David C.S. Roberts, Ph.D., Advisor Examining Committee: Thomas Martin, Ph.D., Chairman Allyn Howlett, Ph.D. Sara Jones, Ph.D. Wayne Pratt, Ph.D.
 PARSING DISTINCT ASPECTS OF THE ADDICTION PROCESS USING COCAINE SELF-ADMINISTRATION, BEHAVIORAL ECONOMICS, NEUROPHARMACOLOGY AND NEUROCHEMISTRY By Erik B. Oleson A Dissertation Submitted to the Graduate
PARSING DISTINCT ASPECTS OF THE ADDICTION PROCESS USING COCAINE SELF-ADMINISTRATION, BEHAVIORAL ECONOMICS, NEUROPHARMACOLOGY AND NEUROCHEMISTRY By Erik B. Oleson A Dissertation Submitted to the Graduate
The Adolescent Developmental Stage
 The Adolescent Developmental Stage o Physical maturation o Drive for independence o Increased salience of social and peer interactions o Brain development o Inflection in risky behaviors including experimentation
The Adolescent Developmental Stage o Physical maturation o Drive for independence o Increased salience of social and peer interactions o Brain development o Inflection in risky behaviors including experimentation
Neurobiology of Addiction
 Neurobiology of Addiction Tiffany Love, Ph.D. Department of Psychiatry The University of Utah What is Addiction? Addiction is a chronic, relapsing, and treatable brain disorder. Compulsive drug seeking
Neurobiology of Addiction Tiffany Love, Ph.D. Department of Psychiatry The University of Utah What is Addiction? Addiction is a chronic, relapsing, and treatable brain disorder. Compulsive drug seeking
Erin S. Calipari, Mark J. Ferris, Cody A. Siciliano, and Sara R. Jones*
 This is an open access article published under an ACS AuthorChoice License, which permits copying and redistribution of the article or any adaptations for non-commercial purposes. pubs.acs.org/chemneuro
This is an open access article published under an ACS AuthorChoice License, which permits copying and redistribution of the article or any adaptations for non-commercial purposes. pubs.acs.org/chemneuro
Food restriction: enhancing effects on drug reward and striatal cell signaling
 Food restriction: enhancing effects on drug reward and striatal cell signaling K.D. Carr Departments of Psychiatry & Pharmacology NYU School of Medicine Common Neural Substrates for Incentive-Motivating
Food restriction: enhancing effects on drug reward and striatal cell signaling K.D. Carr Departments of Psychiatry & Pharmacology NYU School of Medicine Common Neural Substrates for Incentive-Motivating
Using Neuroimaging to Explore Addiction in Animal Models
 Using Neuroimaging to Explore Addiction in Animal Models NHPs in neuroimaging can be used for Radiotracer development / Occupancy studies Modeling human drug addiction and other brain diseases Radiotracer
Using Neuroimaging to Explore Addiction in Animal Models NHPs in neuroimaging can be used for Radiotracer development / Occupancy studies Modeling human drug addiction and other brain diseases Radiotracer
ELUCIDATING AMPHETAMINE ACTIONS AT THE DOPAMINE TERMINAL: POTENTIAL FOR COCAINE ADDICTION TREATMENT CODY ALEXANDER SICILIANO
 ELUCIDATING AMPHETAMINE ACTIONS AT THE DOPAMINE TERMINAL: POTENTIAL FOR COCAINE ADDICTION TREATMENT BY CODY ALEXANDER SICILIANO A Dissertation Submitted to the Graduate Faculty of WAKE FOREST UNIVERSITY
ELUCIDATING AMPHETAMINE ACTIONS AT THE DOPAMINE TERMINAL: POTENTIAL FOR COCAINE ADDICTION TREATMENT BY CODY ALEXANDER SICILIANO A Dissertation Submitted to the Graduate Faculty of WAKE FOREST UNIVERSITY
Chapter 5: Learning and Behavior Learning How Learning is Studied Ivan Pavlov Edward Thorndike eliciting stimulus emitted
 Chapter 5: Learning and Behavior A. Learning-long lasting changes in the environmental guidance of behavior as a result of experience B. Learning emphasizes the fact that individual environments also play
Chapter 5: Learning and Behavior A. Learning-long lasting changes in the environmental guidance of behavior as a result of experience B. Learning emphasizes the fact that individual environments also play
Serotonin System May Have Potential as a Target for Cocaine Medications
 NIDA - Publications - NIDA Notes - Vol. 21, No. 3 - Research Findings of 4 http://www.drugabuse.gov/nida_notes/nnvol21n3/serotonin.html 9/26/2011 3:45 PM NIDA NEWS NIDA Home > Publications > NIDA Notes
NIDA - Publications - NIDA Notes - Vol. 21, No. 3 - Research Findings of 4 http://www.drugabuse.gov/nida_notes/nnvol21n3/serotonin.html 9/26/2011 3:45 PM NIDA NEWS NIDA Home > Publications > NIDA Notes
Understanding Addiction and Its Impact on the Brain. SDSMA Webinar Matthew Stanley, DO
 Understanding Addiction and Its Impact on the Brain SDSMA Webinar Matthew Stanley, DO Estimated Economic Cost to Society Due to Substance Abuse and Addiction: Illegal drugs: Alcohol: Tobacco: $181 billion/year
Understanding Addiction and Its Impact on the Brain SDSMA Webinar Matthew Stanley, DO Estimated Economic Cost to Society Due to Substance Abuse and Addiction: Illegal drugs: Alcohol: Tobacco: $181 billion/year
590,000 deaths can be attributed to an addictive substance in some way
 Mortality and morbidity attributable to use of addictive substances in the United States. The Association of American Physicians from 1999 60 million tobacco smokers in the U.S. 14 million dependent on
Mortality and morbidity attributable to use of addictive substances in the United States. The Association of American Physicians from 1999 60 million tobacco smokers in the U.S. 14 million dependent on
Brain Imaging studies in substance abuse. Jody Tanabe, MD University of Colorado Denver
 Brain Imaging studies in substance abuse Jody Tanabe, MD University of Colorado Denver NRSC January 28, 2010 Costs: Health, Crime, Productivity Costs in billions of dollars (2002) $400 $350 $400B legal
Brain Imaging studies in substance abuse Jody Tanabe, MD University of Colorado Denver NRSC January 28, 2010 Costs: Health, Crime, Productivity Costs in billions of dollars (2002) $400 $350 $400B legal
Drugs, addiction, and the brain
 Drugs, addiction, and the brain Topics to cover: What is addiction? How is addiction studied in the lab? The neuroscience of addiction. Caffeine Cocaine Marijuana (THC) What are the properties of addiction?
Drugs, addiction, and the brain Topics to cover: What is addiction? How is addiction studied in the lab? The neuroscience of addiction. Caffeine Cocaine Marijuana (THC) What are the properties of addiction?
At a Glance. Background Information. Lesson 3 Drugs Change the Way Neurons Communicate
 Lesson 3 Drugs Change the Way Neurons Communicate Overview Students build upon their understanding of neurotransmission by learning how different drugs of abuse disrupt communication between neurons. Students
Lesson 3 Drugs Change the Way Neurons Communicate Overview Students build upon their understanding of neurotransmission by learning how different drugs of abuse disrupt communication between neurons. Students
The Neurobiology of Addiction
 The Neurobiology of Addiction Jodi Gilman, Ph.D. Center for Addiction Medicine Massachusetts General Hospital Associate Professor, Department of Psychiatry Harvard Medical School What is Addiction? commonly
The Neurobiology of Addiction Jodi Gilman, Ph.D. Center for Addiction Medicine Massachusetts General Hospital Associate Professor, Department of Psychiatry Harvard Medical School What is Addiction? commonly
Differential sensitivity to the acute and sensitizing behavioral effects of methylphenidate as a function of strain in adolescent and young adult rats
 Yetnikoff and Arvanitogiannis Behavioral and Brain Functions 2013, 9:38 SHORT PAPER Open Access Differential sensitivity to the acute and sensitizing behavioral effects of methylphenidate as a function
Yetnikoff and Arvanitogiannis Behavioral and Brain Functions 2013, 9:38 SHORT PAPER Open Access Differential sensitivity to the acute and sensitizing behavioral effects of methylphenidate as a function
Elise Cook. BForensics (Hons) Forensic Biology and Toxicology. BSc Biomedical Science and Molecular Biology
 Forensic Biology and Toxicology. BSc Biomedical Science and Molecular Biology") Acute and chronic toxicity of methamphetamine exposure in cultured neuronal cells Elise Cook BForensics (Hons) Forensic Biology and Toxicology BSc Biomedical Science and Molecular Biology This thesis is
Acute and chronic toxicity of methamphetamine exposure in cultured neuronal cells Elise Cook BForensics (Hons) Forensic Biology and Toxicology BSc Biomedical Science and Molecular Biology This thesis is
General introduction. Chapter 1
 General introduction Chapter 1 General introduction Historical aspects of drug use Religious, medicinal and recreational use of mind-altering substances by humans has a history of thousands of years 1.
General introduction Chapter 1 General introduction Historical aspects of drug use Religious, medicinal and recreational use of mind-altering substances by humans has a history of thousands of years 1.
Psychopharmacology of ADHD. Copyright 2006 Neuroscience Education Institute. All rights reserved.
 Psychopharmacology of ADHD Persistence (Predicted Value) Persistence of ADHD Into Adulthood 90 80 70 60 50 40 30 20 10 0 NA 10 15 20 25 30 Age at Follow-Up Syndromatic Persistence Symptomatic Persistence
Psychopharmacology of ADHD Persistence (Predicted Value) Persistence of ADHD Into Adulthood 90 80 70 60 50 40 30 20 10 0 NA 10 15 20 25 30 Age at Follow-Up Syndromatic Persistence Symptomatic Persistence
JOURNAL OF NEUROCHEMISTRY doi: /jnc.12808
 JOURNAL OF NEUROCHEMISTRY 2014 131 348 355 doi: 10.1111/jnc.12808 *Department of Physiology and Pharmacology, Wake Forest School of Medicine, Winston-Salem, North Carolina, USA Fishberg Department of Neuroscience,
JOURNAL OF NEUROCHEMISTRY 2014 131 348 355 doi: 10.1111/jnc.12808 *Department of Physiology and Pharmacology, Wake Forest School of Medicine, Winston-Salem, North Carolina, USA Fishberg Department of Neuroscience,
MANAGING PAIN IN PATIENTS WITH SUBSTANCE USE DISORDER
 MANAGING PAIN IN PATIENTS WITH SUBSTANCE USE DISORDER Melissa B. Weimer, DO, MCR Chief of Behavioral Health & Addiction Medicine St. Peter s Health Partners Grand Rounds October 11, 2017 Disclosures One
MANAGING PAIN IN PATIENTS WITH SUBSTANCE USE DISORDER Melissa B. Weimer, DO, MCR Chief of Behavioral Health & Addiction Medicine St. Peter s Health Partners Grand Rounds October 11, 2017 Disclosures One
Effects of lesions of the nucleus accumbens core and shell on response-specific Pavlovian i n s t ru mental transfer
 Effects of lesions of the nucleus accumbens core and shell on response-specific Pavlovian i n s t ru mental transfer RN Cardinal, JA Parkinson *, TW Robbins, A Dickinson, BJ Everitt Departments of Experimental
Effects of lesions of the nucleus accumbens core and shell on response-specific Pavlovian i n s t ru mental transfer RN Cardinal, JA Parkinson *, TW Robbins, A Dickinson, BJ Everitt Departments of Experimental
MeCP2 and psychostimulantinduced behavioral adaptations. Anne E. West, M.D., Ph.D. Department of Neurobiology Duke University Medical Center
 MeCP2 and psychostimulantinduced behavioral adaptations Anne E. West, M.D., Ph.D. Department of Neurobiology Duke University Medical Center Psychostimulants and antidepressants slowly change behavior Psychostimulants
MeCP2 and psychostimulantinduced behavioral adaptations Anne E. West, M.D., Ph.D. Department of Neurobiology Duke University Medical Center Psychostimulants and antidepressants slowly change behavior Psychostimulants
Basic definition and Classification of Anhedonia. Preclinical and Clinical assessment of anhedonia.
 Basic definition and Classification of Anhedonia. Preclinical and Clinical assessment of anhedonia. Neurobiological basis and pathways involved in anhedonia. Objective characterization and computational
Basic definition and Classification of Anhedonia. Preclinical and Clinical assessment of anhedonia. Neurobiological basis and pathways involved in anhedonia. Objective characterization and computational
Low Dopamine Receptor Availability May Promote Cocaine Addiction
 of 5 10/10/2011 10:56 AM NIDA NEWS NIDA Home > Publications > NIDA Notes > Vol. 22, No. 3 > Research Findings Low Dopamine Receptor Availability May Promote Cocaine Addiction Research Findings Vol. 22,
of 5 10/10/2011 10:56 AM NIDA NEWS NIDA Home > Publications > NIDA Notes > Vol. 22, No. 3 > Research Findings Low Dopamine Receptor Availability May Promote Cocaine Addiction Research Findings Vol. 22,
Repeated stress exposure causes strain-dependent shifts in the behavioral economics of cocaine in rats
 bs_bs_banneraddiction Biology PRECLINICAL STUDY doi:10.1111/adb.12123 Repeated stress exposure causes strain-dependent shifts in the behavioral economics of cocaine in rats Peter A. Groblewski 1, Chad
bs_bs_banneraddiction Biology PRECLINICAL STUDY doi:10.1111/adb.12123 Repeated stress exposure causes strain-dependent shifts in the behavioral economics of cocaine in rats Peter A. Groblewski 1, Chad
The Biological Perspective. Jørg Mørland Senior researcher, Norwegian Institute of Public Health Professor em of Medicine University of Oslo
 The Biological Perspective Jørg Mørland Senior researcher, Norwegian Institute of Public Health Professor em of Medicine University of Oslo The Biological Perspective What is it? More than «the» one biological
The Biological Perspective Jørg Mørland Senior researcher, Norwegian Institute of Public Health Professor em of Medicine University of Oslo The Biological Perspective What is it? More than «the» one biological
Any use of the contents of this presentation is authorized, on the condition of informing the author about it and quoting the source.
 Any use of the contents of this presentation is authorized, on the condition of informing the author about it and quoting the source. Toute utilisation du contenu de cette présentation est autorisée, à
Any use of the contents of this presentation is authorized, on the condition of informing the author about it and quoting the source. Toute utilisation du contenu de cette présentation est autorisée, à
The Neurobiology of Learning and Memory
 The Neurobiology of Learning and Memory JERRY W. RUDY University of Colorado, Boulder Sinauer Associates, Inc. Publishers Sunderland, Massachusetts 01375 Table of Contents CHAPTER 1 Introduction: Fundamental
The Neurobiology of Learning and Memory JERRY W. RUDY University of Colorado, Boulder Sinauer Associates, Inc. Publishers Sunderland, Massachusetts 01375 Table of Contents CHAPTER 1 Introduction: Fundamental
Qualifying Examination (Part I)
") Department of Pharmacology Qualifying Examination (Part I) December 14 & 15, 2010 Committee Members: Eugenia Gurevich Vsevolod Gurevich Tina Iverson Heidi Hamm Joey Barnett, for retakes Exam Schedule:
Department of Pharmacology Qualifying Examination (Part I) December 14 & 15, 2010 Committee Members: Eugenia Gurevich Vsevolod Gurevich Tina Iverson Heidi Hamm Joey Barnett, for retakes Exam Schedule:
BRAIN MECHANISMS OF REWARD AND ADDICTION
 BRAIN MECHANISMS OF REWARD AND ADDICTION TREVOR.W. ROBBINS Department of Experimental Psychology, University of Cambridge Many drugs of abuse, including stimulants such as amphetamine and cocaine, opiates
BRAIN MECHANISMS OF REWARD AND ADDICTION TREVOR.W. ROBBINS Department of Experimental Psychology, University of Cambridge Many drugs of abuse, including stimulants such as amphetamine and cocaine, opiates
The Neuroscience of Addiction: A mini-review
 The Neuroscience of Addiction: A mini-review Jim Morrill, MD, PhD MGH Charlestown HealthCare Center Massachusetts General Hospital Disclosures Neither I nor my spouse/partner has a relevant financial relationship
The Neuroscience of Addiction: A mini-review Jim Morrill, MD, PhD MGH Charlestown HealthCare Center Massachusetts General Hospital Disclosures Neither I nor my spouse/partner has a relevant financial relationship
The Role of Smoking in Cocaine. Addiction
 The Role of Smoking in Cocaine Addiction Luca Colnaghi Eric Kandel Laboratory Columbia University in the City of New York Department of Neuroscience Index 1- The Brain, memory, metaplasticity 2- Cocaine
The Role of Smoking in Cocaine Addiction Luca Colnaghi Eric Kandel Laboratory Columbia University in the City of New York Department of Neuroscience Index 1- The Brain, memory, metaplasticity 2- Cocaine
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION doi:10.1038/nature12024 entary Figure 1. Distribution of the number of earned cocaine Supplementary Figure 1. Distribution of the number of earned cocaine infusions in Shock-sensitive
SUPPLEMENTARY INFORMATION doi:10.1038/nature12024 entary Figure 1. Distribution of the number of earned cocaine Supplementary Figure 1. Distribution of the number of earned cocaine infusions in Shock-sensitive
Adolescent Prozac Exposure Enhances Sensitivity to Cocaine in Adulthood INTRODUCTION
 INTRODUCTION Epidemiologic reports indicate that mood disorders in children and adolescents are quite common, with up to 70% of depressed children and adolescents experiencing a recurrence within 5 years
INTRODUCTION Epidemiologic reports indicate that mood disorders in children and adolescents are quite common, with up to 70% of depressed children and adolescents experiencing a recurrence within 5 years
Behavioral Neuroscience: Fear thou not. Rony Paz
 Behavioral Neuroscience: Fear thou not Rony Paz Rony.paz@weizmann.ac.il Thoughts What is a reward? Learning is best motivated by threats to survival? Threats are much better reinforcers? Fear is a prime
Behavioral Neuroscience: Fear thou not Rony Paz Rony.paz@weizmann.ac.il Thoughts What is a reward? Learning is best motivated by threats to survival? Threats are much better reinforcers? Fear is a prime
Relationship Between Stress and Substance Use Disorders: Neurobiologic Interface
 Relationship Between Stress and Substance Use Disorders: Neurobiologic Interface Kathleen Brady, M.D., Ph.D. Professor of Psychiatry Associate Dean of Clinical and Translational Research Medical University
Relationship Between Stress and Substance Use Disorders: Neurobiologic Interface Kathleen Brady, M.D., Ph.D. Professor of Psychiatry Associate Dean of Clinical and Translational Research Medical University
A NOVEL MODEL OF TASK-SWITCHING AND DECISION MAKING NICK BENTLEY WAKE FOREST UNIVERSITY GRADUATE SCHOOL OF ARTS AND SCIENCES
 A NOVEL MODEL OF TASK-SWITCHING AND DECISION MAKING BY NICK BENTLEY A Dissertation Submitted to the Graduate Faculty of WAKE FOREST UNIVERSITY GRADUATE SCHOOL OF ARTS AND SCIENCES In Partial Fulfillment
A NOVEL MODEL OF TASK-SWITCHING AND DECISION MAKING BY NICK BENTLEY A Dissertation Submitted to the Graduate Faculty of WAKE FOREST UNIVERSITY GRADUATE SCHOOL OF ARTS AND SCIENCES In Partial Fulfillment
Understanding Addiction: Why Can t Those Affected Just Say No?
 Understanding Addiction: Why Can t Those Affected Just Say No? 1 The Stigma of Addiction There continues to be a stigma surrounding addiction even among health care workers. Consider the negative opinions
Understanding Addiction: Why Can t Those Affected Just Say No? 1 The Stigma of Addiction There continues to be a stigma surrounding addiction even among health care workers. Consider the negative opinions
CHAPTER 2. THE NEUROBIOLOGY OF SUBSTANCE USE, MISUSE, AND ADDICTION
 CHAPTER 2. THE NEUROBIOLOGY OF SUBSTANCE USE, MISUSE, AND ADDICTION Chapter 2 Preview A substantial body of research has accumulated over several decades and transformed our understanding of substance
CHAPTER 2. THE NEUROBIOLOGY OF SUBSTANCE USE, MISUSE, AND ADDICTION Chapter 2 Preview A substantial body of research has accumulated over several decades and transformed our understanding of substance
by popular demand: Addiction II
 by popular demand: Addiction II PSY/NEU338: Animal learning and decision making: Psychological, computational and neural perspectives drug addiction huge and diverse field of research (many different drugs)
by popular demand: Addiction II PSY/NEU338: Animal learning and decision making: Psychological, computational and neural perspectives drug addiction huge and diverse field of research (many different drugs)
Part IV: Slipping Up: Neuroscience Basis of Relapse & Recovery July 31, am 12:30 pm with Break 10-10:15am
 Eighth Edition Congratulations and Thank You All for Attending Part IV: Slipping Up: Neuroscience Basis of Relapse & Recovery July 31, 2015 8 am 12:30 pm with Break 10-10:15am Preventing Recrudescence
Eighth Edition Congratulations and Thank You All for Attending Part IV: Slipping Up: Neuroscience Basis of Relapse & Recovery July 31, 2015 8 am 12:30 pm with Break 10-10:15am Preventing Recrudescence
Neurobiology of Addiction JeanAnne Johnson Talbert, DHA, APRN BC, FNP, CARN AP
 Neurobiology of Addiction JeanAnne Johnson Talbert, DHA, APRN BC, FNP, CARN AP Disclosures This speaker has no conflicts of interest to disclose Objectives Define drug abuse and addiction Identify the
Neurobiology of Addiction JeanAnne Johnson Talbert, DHA, APRN BC, FNP, CARN AP Disclosures This speaker has no conflicts of interest to disclose Objectives Define drug abuse and addiction Identify the
America is a drugged society
 Overview of Drug Abuse Basic Considerations. M. Imad Damaj, Ph.D. Associate Professor Dept. of Pharmacology/Toxicology, Virginia Commonwealth University America is a drugged society 90% of all drugs manufactured
Overview of Drug Abuse Basic Considerations. M. Imad Damaj, Ph.D. Associate Professor Dept. of Pharmacology/Toxicology, Virginia Commonwealth University America is a drugged society 90% of all drugs manufactured
DRUGS AND SOCIETY. Behavioral Medicines and Abusabie Drugs. Arthur P. Leccese, Ph.D. Kenyon College. PRENTICE HALL, Englewood Cliffs, New Jersey 07632
 DRUGS AND SOCIETY Behavioral Medicines and Abusabie Drugs Arthur P. Leccese, Ph.D. Kenyon College PRENTICE HALL, Englewood Cliffs, New Jersey 07632 CONTENTS PREFACE xv PART A PSYCHOPHARMACOLOGY CHAPTER
DRUGS AND SOCIETY Behavioral Medicines and Abusabie Drugs Arthur P. Leccese, Ph.D. Kenyon College PRENTICE HALL, Englewood Cliffs, New Jersey 07632 CONTENTS PREFACE xv PART A PSYCHOPHARMACOLOGY CHAPTER
The Brain, Behavior and Addiction National Family Dialogue January 27, 2010 Presenter: Flo Hilliard, MSH University of Wisconsin-Madison
 The Brain, Behavior and Addiction National Family Dialogue January 27, 2010 Presenter: Flo Hilliard, MSH University of Wisconsin-Madison Attitudes about addiction and recovery throughout history Disease?
The Brain, Behavior and Addiction National Family Dialogue January 27, 2010 Presenter: Flo Hilliard, MSH University of Wisconsin-Madison Attitudes about addiction and recovery throughout history Disease?
BIPN 140 Problem Set 6
 BIPN 140 Problem Set 6 1) Hippocampus is a cortical structure in the medial portion of the temporal lobe (medial temporal lobe in primates. a) What is the main function of the hippocampus? The hippocampus
BIPN 140 Problem Set 6 1) Hippocampus is a cortical structure in the medial portion of the temporal lobe (medial temporal lobe in primates. a) What is the main function of the hippocampus? The hippocampus
 No! No! No! No! With the possible exception of humans Public Health Question Does the compound have the potential to be abused? Public Health Question Does the compound have the potential to be abused?
No! No! No! No! With the possible exception of humans Public Health Question Does the compound have the potential to be abused? Public Health Question Does the compound have the potential to be abused?
Paradoxical reactions to cocaine
 Paradoxical reactions to cocaine Jan 18, 2006. Abstract. Quantitative and conventional microdialysis were used to investigate the effects of constitutive deletion of the prodynorphin gene on basal dopamine
Paradoxical reactions to cocaine Jan 18, 2006. Abstract. Quantitative and conventional microdialysis were used to investigate the effects of constitutive deletion of the prodynorphin gene on basal dopamine
nucleus accumbens septi hier-259 Nucleus+Accumbens birnlex_727
 Nucleus accumbens From Wikipedia, the free encyclopedia Brain: Nucleus accumbens Nucleus accumbens visible in red. Latin NeuroNames MeSH NeuroLex ID nucleus accumbens septi hier-259 Nucleus+Accumbens birnlex_727
Nucleus accumbens From Wikipedia, the free encyclopedia Brain: Nucleus accumbens Nucleus accumbens visible in red. Latin NeuroNames MeSH NeuroLex ID nucleus accumbens septi hier-259 Nucleus+Accumbens birnlex_727
Addiction in the Brain - Latest Research. Gary M. Henschen, MD, LFAPA Chief Behavioral Health Officer Magellan Healthcare, Inc.
 Addiction in the Brain - Latest Research Gary M. Henschen, MD, LFAPA Chief Behavioral Health Officer Magellan Healthcare, Inc. The Neurobiology of Addiction About the speaker Gary M. Henschen, M.D. is
Addiction in the Brain - Latest Research Gary M. Henschen, MD, LFAPA Chief Behavioral Health Officer Magellan Healthcare, Inc. The Neurobiology of Addiction About the speaker Gary M. Henschen, M.D. is
BIPN 140 Problem Set 6
 BIPN 140 Problem Set 6 1) The hippocampus is a cortical structure in the medial portion of the temporal lobe (medial temporal lobe in primates. a) What is the main function of the hippocampus? The hippocampus
BIPN 140 Problem Set 6 1) The hippocampus is a cortical structure in the medial portion of the temporal lobe (medial temporal lobe in primates. a) What is the main function of the hippocampus? The hippocampus
Psychoactive Drugs & The Brain
 Psychoactive Drugs & The Brain Psychoactive Substances & The Brain 1. Psychoactive substances enter the bloodstream via oral administration, inhalation, smoking, or injection 2. Psychoactive substances
Psychoactive Drugs & The Brain Psychoactive Substances & The Brain 1. Psychoactive substances enter the bloodstream via oral administration, inhalation, smoking, or injection 2. Psychoactive substances
Synaptic plasticity and addiction
 Synaptic plasticity and addiction Julie A. Kauer* and Robert C. Malenka Abstract Addiction is caused, in part, by powerful and long-lasting memories of the drug experience. Relapse caused by exposure to
Synaptic plasticity and addiction Julie A. Kauer* and Robert C. Malenka Abstract Addiction is caused, in part, by powerful and long-lasting memories of the drug experience. Relapse caused by exposure to
The Neurobiology of Drug Addiction
 The Neurobiology of Drug Addiction Glen R. Hanson, Ph.D., D.D.S. Director, Utah Addiction Center Professor of Pharmacology and Toxicology, U of U Senior Advisor, NIDA Drug Abuse and Addiction are Among
The Neurobiology of Drug Addiction Glen R. Hanson, Ph.D., D.D.S. Director, Utah Addiction Center Professor of Pharmacology and Toxicology, U of U Senior Advisor, NIDA Drug Abuse and Addiction are Among
If you give any person a prescription of something like Valium and have them take it on
 As always I am happy to do this presentation, which is my favorite topic in addiction medicine. I am an internist, and I have done healthcare for the homeless in Springfield as well as been the medical
As always I am happy to do this presentation, which is my favorite topic in addiction medicine. I am an internist, and I have done healthcare for the homeless in Springfield as well as been the medical
Behavioral Neuroscience: Fear thou not. Rony Paz
 Behavioral Neuroscience: Fear thou not Rony Paz Rony.paz@weizmann.ac.il Thoughts What is a reward? Learning is best motivated by threats to survival Threats are much better reinforcers Fear is a prime
Behavioral Neuroscience: Fear thou not Rony Paz Rony.paz@weizmann.ac.il Thoughts What is a reward? Learning is best motivated by threats to survival Threats are much better reinforcers Fear is a prime
Neurotransmitter Functioning In Major Depressive Disorder
 Neurotransmitter Functioning In Major Depressive Disorder Otsuka Pharmaceutical Development & Commercialization, Inc. 2017 Otsuka Pharmaceutical Development & Commercialization, Inc., Rockville, MD January
Neurotransmitter Functioning In Major Depressive Disorder Otsuka Pharmaceutical Development & Commercialization, Inc. 2017 Otsuka Pharmaceutical Development & Commercialization, Inc., Rockville, MD January
Book 3: Lab Procedures Book 3: Ch. 1: The Hypothesis and Overview
 Book 3: Lab Procedures Book 3: Ch. 1: The Hypothesis and Overview 13 Introduction This experiment will investigate how cocaine acts on dopamine neurons in the brain. Cocaine is a drug of abuse that increases
Book 3: Lab Procedures Book 3: Ch. 1: The Hypothesis and Overview 13 Introduction This experiment will investigate how cocaine acts on dopamine neurons in the brain. Cocaine is a drug of abuse that increases
Supplementary Figure 1
 Supplementary Figure 1 Localization of virus injections. (a) Schematic showing the approximate center of AAV-DIO-ChR2-YFP injection sites in the NAc of Dyn-cre mice (n=8 mice, 16 injections; caudate/putamen,
Supplementary Figure 1 Localization of virus injections. (a) Schematic showing the approximate center of AAV-DIO-ChR2-YFP injection sites in the NAc of Dyn-cre mice (n=8 mice, 16 injections; caudate/putamen,
HISTORY MPH PHARMACOLOGY METHYLPHENIDATE HYDROCHOLORIDE- RITALIN. RITALIN ADDICTION PAOLA ROSCA Head Depart. for the Treatment of Substance Abuse
 METHYLPHENIDATE HYDROCHOLORIDE- RITALIN RITALIN ADDICTION PAOLA ROSCA Head Depart. for the Treatment of Substance Abuse Ministry of Health December 2013 HISTORY Ritalin was synthesized in 1944 by the chemist
METHYLPHENIDATE HYDROCHOLORIDE- RITALIN RITALIN ADDICTION PAOLA ROSCA Head Depart. for the Treatment of Substance Abuse Ministry of Health December 2013 HISTORY Ritalin was synthesized in 1944 by the chemist
Long-Term Effects of Chronic Oral Ritalin Administration on Cognitive and Neural Development in Adolescent Wistar Kyoto Rats
 Brain Sci.,, -; doi:.9/brainsci Article OPEN ACCESS brain sciences ISSN - www.mdpi.com/journal/brainsci/ Long-Term Effects of Chronic Oral Ritalin Administration on Cognitive and Neural Development in
Brain Sci.,, -; doi:.9/brainsci Article OPEN ACCESS brain sciences ISSN - www.mdpi.com/journal/brainsci/ Long-Term Effects of Chronic Oral Ritalin Administration on Cognitive and Neural Development in
What are Substance Use Disorders?
 What are Substance Use Disorders? Sanchit Maruti, MD Michael Goedde, MD University of Vermont Medical Center 1 Disclosures } Drs. Maruti and Goedde receive compensation as consultants to the American Academy
What are Substance Use Disorders? Sanchit Maruti, MD Michael Goedde, MD University of Vermont Medical Center 1 Disclosures } Drs. Maruti and Goedde receive compensation as consultants to the American Academy
- Neurotransmitters Of The Brain -
 - Neurotransmitters Of The Brain - INTRODUCTION Synapsis: a specialized connection between two neurons that permits the transmission of signals in a one-way fashion (presynaptic postsynaptic). Types of
- Neurotransmitters Of The Brain - INTRODUCTION Synapsis: a specialized connection between two neurons that permits the transmission of signals in a one-way fashion (presynaptic postsynaptic). Types of
Na + /Cl - -dependent Neurotransmitter Transporters. Outline. Synaptic Transmission. How did the neurotransmitter cross the membrane?
 Na /Cl - -dependent Neurotransmitter Transporters Stine Meinild Lundby The Neurobiology Group Molecular, Cellular, and Integrative Physiology Section November 26 th, 2007 slide 1 Outline Synaptic neurotransmission
Na /Cl - -dependent Neurotransmitter Transporters Stine Meinild Lundby The Neurobiology Group Molecular, Cellular, and Integrative Physiology Section November 26 th, 2007 slide 1 Outline Synaptic neurotransmission
The Biology of Addiction
 The Biology of Addiction Risk factors for addiction: Biological/Genetic Family history of addiction Being male Having mental illness Exposure to substances in utero * The genes that people are born with
The Biology of Addiction Risk factors for addiction: Biological/Genetic Family history of addiction Being male Having mental illness Exposure to substances in utero * The genes that people are born with
Psychoactive drugs Drugs which affect mental processes. Legal but restricted (by prescription only)
") Psychoactive drugs Drugs which affect mental processes mood, perception, memory, state of consciousness May be illegal: Heroin Cocaine Legal: Alcohol nicotine Legal but restricted (by prescription only)
Psychoactive drugs Drugs which affect mental processes mood, perception, memory, state of consciousness May be illegal: Heroin Cocaine Legal: Alcohol nicotine Legal but restricted (by prescription only)
Supplementary Information
 Supplementary Information The neural correlates of subjective value during intertemporal choice Joseph W. Kable and Paul W. Glimcher a 10 0 b 10 0 10 1 10 1 Discount rate k 10 2 Discount rate k 10 2 10
Supplementary Information The neural correlates of subjective value during intertemporal choice Joseph W. Kable and Paul W. Glimcher a 10 0 b 10 0 10 1 10 1 Discount rate k 10 2 Discount rate k 10 2 10
Behavioral Pharmacology
 Psych 181: Dr. Anagnostaras Lecture 4 Behavioral Pharmacology Behavioral Pharmacology Behavioral Pharmacology The study of the relationship between the physiological actions of drugs and their effects
Psych 181: Dr. Anagnostaras Lecture 4 Behavioral Pharmacology Behavioral Pharmacology Behavioral Pharmacology The study of the relationship between the physiological actions of drugs and their effects
The Neurobiology of Drug Addiction
 National Institute on Drug Abuse (NIDA) The Neurobiology of Drug Addiction Last Updated January 2007 https://www.drugabuse.gov 1 Table of Contents The Neurobiology of Drug Addiction Section I: Introduction
National Institute on Drug Abuse (NIDA) The Neurobiology of Drug Addiction Last Updated January 2007 https://www.drugabuse.gov 1 Table of Contents The Neurobiology of Drug Addiction Section I: Introduction
DOES THE D 3 DOPAMINE RECEPTOR (D 3 r)play A ROLE IN ADDICTION? Isabelle Boileau, PhD
play A ROLE IN ADDICTION? Isabelle Boileau, PhD") DOES THE D 3 DOPAMINE RECEPTOR (D 3 r)play A ROLE IN ADDICTION? Isabelle Boileau, PhD Acknowledgments Human Neurochemical Pathology Lab Stephen J. Kish, PhD Jun Chao Tong, PhD Centre for Movement Disorders
DOES THE D 3 DOPAMINE RECEPTOR (D 3 r)play A ROLE IN ADDICTION? Isabelle Boileau, PhD Acknowledgments Human Neurochemical Pathology Lab Stephen J. Kish, PhD Jun Chao Tong, PhD Centre for Movement Disorders
Dopamine in Ube3a m-/p+ mice. Online Supplemental Material
 Online Supplemental Material S1 Supplemental Figure 1. Schematic of rate-dependent intracranial self-stimulation (ICSS) (A) Mice implanted with monopolar stimulating electrodes to the medial forebrain
Online Supplemental Material S1 Supplemental Figure 1. Schematic of rate-dependent intracranial self-stimulation (ICSS) (A) Mice implanted with monopolar stimulating electrodes to the medial forebrain
NIH Public Access Author Manuscript Biochem Pharmacol. Author manuscript; available in PMC 2009 January 1.
 NIH Public Access Author Manuscript Published in final edited form as: Biochem Pharmacol. 2008 January 1; 75(1): 218 265. Glutamatergic substrates of drug addiction and alcoholism 1 Justin T. Gass and
NIH Public Access Author Manuscript Published in final edited form as: Biochem Pharmacol. 2008 January 1; 75(1): 218 265. Glutamatergic substrates of drug addiction and alcoholism 1 Justin T. Gass and
A comparison of MDMA and d-amphetamine in the drug discrimination paradigm.
 A comparison of MDMA and d-amphetamine in the drug discrimination paradigm. D.N. Harper, A. Crowther, S. Schenk School of Psychology Victoria University of Wellington, New Zealand AIM To compare the subjective
A comparison of MDMA and d-amphetamine in the drug discrimination paradigm. D.N. Harper, A. Crowther, S. Schenk School of Psychology Victoria University of Wellington, New Zealand AIM To compare the subjective
Council on Chemical Abuse Annual Conference November 2, The Science of Addiction: Rewiring the Brain
 Council on Chemical Abuse Annual Conference November 2, 2017 The Science of Addiction: Rewiring the Brain David Reyher, MSW, CAADC Behavioral Health Program Director Alvernia University Defining Addiction
Council on Chemical Abuse Annual Conference November 2, 2017 The Science of Addiction: Rewiring the Brain David Reyher, MSW, CAADC Behavioral Health Program Director Alvernia University Defining Addiction
Mk-801 Administration in Adolescent Male Rats and Cocaine Conditioned Place
 Mk-801 Administration in Adolescent Male Rats and Cocaine Conditioned Place Preference Stephanie Willis, Jonnique Adjmul, Shabaaz Sandhu, Antoniette M. Maldonado-Devincci, Cheryl Kirsten ABSTRACT The present
Mk-801 Administration in Adolescent Male Rats and Cocaine Conditioned Place Preference Stephanie Willis, Jonnique Adjmul, Shabaaz Sandhu, Antoniette M. Maldonado-Devincci, Cheryl Kirsten ABSTRACT The present
Jennifer Lynn Green. Chapel Hill 2012 Approved by: Regina M. Carelli. Linda Dysktra. Todd Thiele
 NUCLEUS ACCUMBENS NEURONS DIFFERENTIALLY ENCODE INFORMATION ABOUT AVERSIVE CUES THAT PREDICT COCAINE AVAILABILITY AND COCAINE SELF-ADMINISTRATION FOLLOWING EXTENDED TASTE-DRUG PAIRINGS. Jennifer Lynn Green
NUCLEUS ACCUMBENS NEURONS DIFFERENTIALLY ENCODE INFORMATION ABOUT AVERSIVE CUES THAT PREDICT COCAINE AVAILABILITY AND COCAINE SELF-ADMINISTRATION FOLLOWING EXTENDED TASTE-DRUG PAIRINGS. Jennifer Lynn Green
The Role of NEUROIMAGING In Diagnostic and Clinical Practice
 The Role of NEUROIMAGING In Diagnostic and Clinical Practice ADHD Schizophrenia Autism Addiction Altzheimer s Disease Nora D. Volkow, M.D. Director National Institute on Drug Abuse National Institutes
The Role of NEUROIMAGING In Diagnostic and Clinical Practice ADHD Schizophrenia Autism Addiction Altzheimer s Disease Nora D. Volkow, M.D. Director National Institute on Drug Abuse National Institutes
Localization of brain reinforcement mechanisms: intracranial self-administration and intracranial place-conditioning studies
 Behavioural Brain Research 101 (1999) 129 152 Review article Localization of brain reinforcement mechanisms: intracranial self-administration and intracranial place-conditioning studies William J. McBride
Behavioural Brain Research 101 (1999) 129 152 Review article Localization of brain reinforcement mechanisms: intracranial self-administration and intracranial place-conditioning studies William J. McBride
FOOD REWARD AND MODULATION BY NUTRITIONAL STATUS, INSULIN AND LEPTIN. Dianne Figlewicz Lattemann, Ph.D.
 FOOD REWARD AND MODULATION BY NUTRITIONAL STATUS, INSULIN AND LEPTIN Dianne Figlewicz Lattemann, Ph.D. ECOR SYMPOSIUM FEBRUARY 23, 2006 PASSION LIVES HERE, TOO Model for regulation of body adiposity Hypothalamic
FOOD REWARD AND MODULATION BY NUTRITIONAL STATUS, INSULIN AND LEPTIN Dianne Figlewicz Lattemann, Ph.D. ECOR SYMPOSIUM FEBRUARY 23, 2006 PASSION LIVES HERE, TOO Model for regulation of body adiposity Hypothalamic
The role of the metabotropic glutamate 5 and. adenosine 2A receptors in. methamphetamine addiction
 The role of the metabotropic glutamate 5 and adenosine 2A receptors in methamphetamine addiction Rose Chesworth (B. Psych, Hons.) Submitted in total fulfillment of the requirements of the degree of Doctor
The role of the metabotropic glutamate 5 and adenosine 2A receptors in methamphetamine addiction Rose Chesworth (B. Psych, Hons.) Submitted in total fulfillment of the requirements of the degree of Doctor
Cogs 107b Systems Neuroscience lec9_ neuromodulators and drugs of abuse principle of the week: functional anatomy
 Cogs 107b Systems Neuroscience www.dnitz.com lec9_02042010 neuromodulators and drugs of abuse principle of the week: functional anatomy Professor Nitz circa 1986 neurotransmitters: mediating information
Cogs 107b Systems Neuroscience www.dnitz.com lec9_02042010 neuromodulators and drugs of abuse principle of the week: functional anatomy Professor Nitz circa 1986 neurotransmitters: mediating information
The future of pharmacological treatment.
 The future of pharmacological treatment. Anne Lingford-Hughes Professor of Addiction Biology, Imperial College. Hon Consultant CNWL NHS Foundation Trust. What substances and when? What Nicotine Alcohol
The future of pharmacological treatment. Anne Lingford-Hughes Professor of Addiction Biology, Imperial College. Hon Consultant CNWL NHS Foundation Trust. What substances and when? What Nicotine Alcohol
Modeling Neurotransmission and Drugs. CHM 108 Lab. Spring Dr. Angela King
 Modeling Neurotransmission and Drugs CHM 18 Lab Spring 28 Dr. Angela King Part A: How does the brain communicate? The human brain is the most complex organ in the body. This three-pound mass of gray and
Modeling Neurotransmission and Drugs CHM 18 Lab Spring 28 Dr. Angela King Part A: How does the brain communicate? The human brain is the most complex organ in the body. This three-pound mass of gray and
Outline of the presentation
 Understanding and Managing Food Addiction Nicole M. Avena, Ph.D. Institute of Human Nutrition Outline of the presentation The problem of obesity Our modern food landscape Some factors contributing to the
Understanding and Managing Food Addiction Nicole M. Avena, Ph.D. Institute of Human Nutrition Outline of the presentation The problem of obesity Our modern food landscape Some factors contributing to the
Drugs, The Brain, and Behavior
 Drugs, The Brain, and Behavior John Nyby Department of Biological Sciences Lehigh University What is a drug? Difficult to define Know it when you see it Neuroactive vs Non-Neuroactive drugs Two major types
Drugs, The Brain, and Behavior John Nyby Department of Biological Sciences Lehigh University What is a drug? Difficult to define Know it when you see it Neuroactive vs Non-Neuroactive drugs Two major types
ADHD & Addictions -What We Know
 ADHD & Addictions -What We Know Dr. David Teplin, Psy.D., C.Psych. 4th CADDAC Annual Conference, Toronto October 14, 2012 1 Introduction Adult ADHD commonly co-exists with several other disorders, including
ADHD & Addictions -What We Know Dr. David Teplin, Psy.D., C.Psych. 4th CADDAC Annual Conference, Toronto October 14, 2012 1 Introduction Adult ADHD commonly co-exists with several other disorders, including
The Neurobiology of Addiction. Angela Haliburda, DO
 The Neurobiology of Addiction Angela Haliburda, DO Currently, 8-10% of those age 12+ years, or 20 million to 22 million Americans are addicted to alcohol or other drugs. Drug overdoses are now the leading
The Neurobiology of Addiction Angela Haliburda, DO Currently, 8-10% of those age 12+ years, or 20 million to 22 million Americans are addicted to alcohol or other drugs. Drug overdoses are now the leading
Classes of Neurotransmitters. Neurotransmitters
 1 Drugs Outline 2 Neurotransmitters Agonists and Antagonists Cocaine & other dopamine agonists Alcohol & its effects / Marijuana & its effects Synthetic & Designer Drugs: Ecstasy 1 Classes of Neurotransmitters
1 Drugs Outline 2 Neurotransmitters Agonists and Antagonists Cocaine & other dopamine agonists Alcohol & its effects / Marijuana & its effects Synthetic & Designer Drugs: Ecstasy 1 Classes of Neurotransmitters
ADHD Medications & How They Work. Gail C. Rodin, Ph.D. January 21, 2008
 ADHD Medications & How They Work Gail C. Rodin, Ph.D. January 21, 2008 Agenda How the (ADHD) Brain Works (or doesn t) Neurons and neurotransmitters NE & DA: the major players in ADHD Channel vs. state
ADHD Medications & How They Work Gail C. Rodin, Ph.D. January 21, 2008 Agenda How the (ADHD) Brain Works (or doesn t) Neurons and neurotransmitters NE & DA: the major players in ADHD Channel vs. state
Adolescent d-amphetamine treatment in a rodent model of ADHD: pro-cognitive effects during adolescence and cocaine abuse risk during adulthood
 Boston University OpenBU Psychological and Brain Sciences http://open.bu.edu BU Open Access Articles 2015-11 Adolescent d-amphetamine treatment in a rodent model of ADHD: pro-cognitive effects during adolescence
Boston University OpenBU Psychological and Brain Sciences http://open.bu.edu BU Open Access Articles 2015-11 Adolescent d-amphetamine treatment in a rodent model of ADHD: pro-cognitive effects during adolescence
Supplemental Information. Dorsal Raphe Dual Serotonin-Glutamate Neurons. Drive Reward by Establishing Excitatory Synapses
 Cell Reports, Volume 26 Supplemental Information Dorsal Raphe Dual Serotonin-Glutamate Neurons Drive Reward by Establishing Excitatory Synapses on VTA Mesoaccumbens Dopamine Neurons Hui-Ling Wang, Shiliang
Cell Reports, Volume 26 Supplemental Information Dorsal Raphe Dual Serotonin-Glutamate Neurons Drive Reward by Establishing Excitatory Synapses on VTA Mesoaccumbens Dopamine Neurons Hui-Ling Wang, Shiliang
SUPPLEMENTARY INFORMATION
 doi: 10.1038/nature06994 A phosphatase cascade by which rewarding stimuli control nucleosomal response A. Stipanovich*, E. Valjent*, M. Matamales*, A. Nishi, J.H. Ahn, M. Maroteaux, J. Bertran-Gonzalez,
doi: 10.1038/nature06994 A phosphatase cascade by which rewarding stimuli control nucleosomal response A. Stipanovich*, E. Valjent*, M. Matamales*, A. Nishi, J.H. Ahn, M. Maroteaux, J. Bertran-Gonzalez,
Understanding Addiction
 Understanding Addiction How Addiction Hijacks the Brain Addiction involves craving for something intensely, loss of control over its use, and continuing involvement with it despite adverse consequences.
Understanding Addiction How Addiction Hijacks the Brain Addiction involves craving for something intensely, loss of control over its use, and continuing involvement with it despite adverse consequences.
Brain Health and Opioid Abuse
 2018 Statewide Tribal Opioid Summit Brain Health and Opioid Abuse Healing From Opiate Addiction Requires Comprehensive Approaches Psychological, Socio-Cultural, and Biological Donald R. Vereen, Jr., M.D.,
2018 Statewide Tribal Opioid Summit Brain Health and Opioid Abuse Healing From Opiate Addiction Requires Comprehensive Approaches Psychological, Socio-Cultural, and Biological Donald R. Vereen, Jr., M.D.,
Dangers Lurking on Social Media for Kids BRADLEY BRIDGES, LMSW
 Dangers Lurking on Social Media for Kids BRADLEY BRIDGES, LMSW Impact of Social Media on Children Impact on brain development Impact on psychological development Why technology is so addictive What to
Dangers Lurking on Social Media for Kids BRADLEY BRIDGES, LMSW Impact of Social Media on Children Impact on brain development Impact on psychological development Why technology is so addictive What to
Subsecond dopamine release promotes cocaine seeking
 Nature 422, 614-618 (2003) Subsecond dopamine release promotes cocaine seeking Paul E. M. Phillips *, Garret D. Stuber, Michael L. A. V. Heien, R. Mark Wightman and Regina M. Carelli * * Department of
Nature 422, 614-618 (2003) Subsecond dopamine release promotes cocaine seeking Paul E. M. Phillips *, Garret D. Stuber, Michael L. A. V. Heien, R. Mark Wightman and Regina M. Carelli * * Department of
Prefrontal dysfunction in drug addiction: Cause or consequence? Christa Nijnens
 Prefrontal dysfunction in drug addiction: Cause or consequence? Master Thesis Christa Nijnens September 16, 2009 University of Utrecht Rudolf Magnus Institute of Neuroscience Department of Neuroscience
Prefrontal dysfunction in drug addiction: Cause or consequence? Master Thesis Christa Nijnens September 16, 2009 University of Utrecht Rudolf Magnus Institute of Neuroscience Department of Neuroscience