SUPPLEMENTARY INFORMATION
|
|
|
- Ashlie Douglas
- 5 years ago
- Views:
Transcription
1 Encoding of conditioned fear in central amygdala inhibitory circuits Stephane Ciocchi 1,*, Cyril Herry 1,,*, François Grenier 1, Steffen B.E. Wolff 1, Johannes J. Letzkus 1, Ioannis Vlachos 3, Ingrid Ehrlich 1,4, Rolf Sprengel 5, Karl Deisseroth 6, Michael Stadler 1, Christian Müller 1 and Andreas Lüthi 1 1 Friedrich Miescher Institute for Biomedical Research, Maulbeerstrasse 66, CH458 Basel, Switzerland. Present address: INSERM U86, Neurocentre Magendie, 146 Rue LéoSaignat, 3377 Bordeaux, France. 3 Bernstein Center for Computational Neuroscience, Freiburg, Germany. 4 Present address: Hertie Institute for Clinical Brain Research, 776 Tübingen, Germany. 5 Department of Molecular Neurobiology, Max Planck Institute for Medical Research, Jahnstrasse 9, 691 Heidelberg, Germany. 6 Department of Bioengineering, Stanford University, Stanford, CA 9435, USA. * These authors contributed equally to this work. Supplementary notes Given that both CEl off and CEl on neurons can form anatomical and functional inhibitory connections in, the question arises why output neurons are disinhibited, rather than inhibited i.e. why the CEl off pathway predominates over the CEl on pathway during sensory stimulation. A possible explanation could be that unitary synaptic connections made by CEl on and CEl off neurons onto output neurons have distinct properties in terms of their strength or their shortterm dynamics. Moreover, based on our data, we cannot exclude that CEl on and CEl off neurons might target distinct subtypes of output neurons. More detailed in vitro electrophysiological and anatomical studies will be required to address these questions. Our results on cell activity are in accord with most studies on the effects of CEA activation on fear behavior. However, the decrease in firing of brainstem projecting cells upon presentation reported by Pascoe and Kapp (1985) 1 seems at odds with our results, which can be at least partly traced to methodological differences. Their neuronal groupings were purely functional, and did not distinguish between the subnuclei in which their neurons were recorded. Furthermore, their analysis was on pooled nonnormalized data, making it hard to appreciate quantitatively the activity that can be ascribed only to brainstem projecting cells. 1. Pascoe, J.P. & Kapp, B.S. Electrophysiological characteristics of amygdaloid central nucleus neurons during Pavlovian fear conditioning in the rabbit. Behav. Brain. Res. 16, (1985). 1
2 Statistical analysis Fig. 1c. Left: Blue laser light (473 nm) was used for bilateral stimulation of ChR expressing neurons (1s periods of light stimulation with interstimulation intervals of 36s). Time bins for freezing determination were 1 s for light stimulation and 36 s for interstimulation intervals. Right: The summary data shows significant lightinduced freezing in AAVChRAtdimer infected animals compared to shamoperated controls. There were significant main effects for infection: F (1,5) = , P <.1; and light stimulation: F (1,5) = , P <.1, two way ANOVA; significant interaction infection x light stimulation: F (1,5) = 5.53, P <.1. Infected animals spent 15.9 ± 6.% of time freezing without light, and 63.1 ± 4.% with light (n = 5, P <.5, posthoc Bonferroni ttest). Freezing in shamoperated controls: without light, 7.6 ± 1.8%; with light: 4.8 ±.4% (n = 1, P >.5 vs. no light, P <.5 vs. infected animals with light). Fig. 1e. Freezing Values for inactivation of the different subnuclei: CEl: 5.9 ± 5.%, n = 8, F (3,) = 6.65, P <.1, one way ANOVA, P <.5, CEl vs. Ctrl and, posthoc Bonferroni ttest. :.9 ± 3.4%, n = 8, P >.5 vs. Ctrl. CEA: 6.8 ± 4.5%, n = 5, P >.5 vs. Ctrl. Control: 6. ± 9.6%, n = 5. Fig. 1f. Freezing Values for inactivation of the different subnuclei: Control: 59.4 ± 8.6%, n = 5. CEl: 4.4 ± 3.7%, n = 14, F (3,31) = 1.719, P <.1, one way ANOVA, P <.1, posthoc Bonferroni ttest. CEA: 7.4 ± 6.4%, n = 7, P <.1. : 54.3 ± 5.5%, n = 9, P >.5. Fig. 1g. Freezing Values for inactivation of the different subnuclei: Control: 65.3 ± 8.%, n = 7. : 39. ± 6.%, n = 9, F (3,) = 5.345, P <.1, one way ANOVA, P <.5, posthoc Bonferroni ttest. CEA: 3.1 ± 1.%, n = 7, P <.1. CEl: 57.7 ± 7.6%, n = 8. Fig. a. CEl on neurons: n = 5 units, 1 mice; zscore, habituation:.79 ±.8; post fear conditioning:.45 ±.39, P <.1, blocks of 4 s. Fig. b. CEl off neurons: n = 41 units, 1 mice; zscore, habituation:.14 ±.31; post fear conditioning: 1.3 ±.3, P <.1, blocks of 4 s. Fig. 3c. Fear conditioninginduced changes in evoked firing in units: n = 15 units, 6 mice; average zscore, 1 ms, habituation:.15 ±.3; post fear conditioning: 3.13 ±.77, P <.1. Fig. 4a. Fear conditioninginduced changes in tonic activity: neurons: before: 9.88 ±.99 Hz; after: 5.98 ±.3 Hz, n = 15, P <.1. CEl on neurons: before: 3.99 ±.7 Hz; after: 3. ±.54 Hz, n = 5, P <.5. CEl off neurons: before: 4.65 ± 1.1 Hz; after: 6.11 ± 1.41 Hz, n = 41, P <.5. Fig. 4d. Correlations between tonic activity and generalization: neurons: freezing ratios for discriminating trial blocks: 1.83 ±.11, n = 4 trial blocks; generalizing trial blocks:.9 ±.11, n = 8, P <.1 vs. discriminating trial blocks; Z score ratios of phasic responses for discriminating trial blocks: 1.79 ±.49, n = 4 trial blocks; generalizing trial blocks:.81 ±.16, n = 8, P <.5 vs. discriminating trial blocks. CEl off neurons: freezing ratios for discriminating trial blocks: 3.48 ±.66, n =
3 19 trial blocks; generalizing trial blocks:.7 ±.9, n = 3, P <.1 vs. discriminating trial blocks; Zscore ratios of phasic responses for discriminating trial blocks: 4.15 ± 1.46, n = 19 trial blocks; generalizing trial blocks: 1.6 ±.35, n = 3, P <.5 vs. discriminating trial blocks. CEl on neurons: freezing ratios for discriminating trial blocks: 3.31 ±.63, n = 1 trial blocks; generalizing trial blocks:.8 ±.11, n = 19, P <.1 vs. discriminating trial blocks; Zscore ratios of phasic responses for discriminating trial blocks:.97 ±., n = 1 trial blocks; generalizing trial blocks: 1.56 ±.3, n = 19, P =. vs. discriminating trial blocks. 3


4 Light on a b 6 Counts Supplementary Figure 1 ChRmediated optical control of neuronal firing in vivo. a, Highmagnification picture illustrating AAVinfected neurons. Scale bar is 5 µm. b, Example raster plot and PSTH illustrating light induced sustained firing of an amygdala neuron recorded in an anaesthetised mouse. Similar lightinduced responses were observed in several neurons (n = 5) in AAVChRAtdimer infected animals. 4
5 a Flinching prob. b Flinching prob. c Freezing (%) 1..5 No drug Muscimol CEA CEl.5 No drug Muscimol US intensity (ma) US intensity (ma) no *** no CEA CEl *** Vocalisation prob. Vocalisation prob No drug Muscimol CEA US intensity (ma) CEl No drug Muscimol US intensity (ma) Supplementary Figure Targeted inactivation of CEA or CEl does not affect US sensitivity and does not lead to permanent damage. a, Inactivation of CEA with muscimol does not affect US sensitivity as measured by the US threshold inducing flinching behavior (left, n = 5, P >.5 vs. control) or vocalization (right, n = 5, P >.5 vs. control). b, Likewise, inactivation of CEl with muscimol does not affect US sensitivity (left: flinching behavior, n = 8, P >.5 vs. control; right: vocalizations, n = 8, P >.5 vs. control). c, Reconditioning of mice 4 hrs after inactivation of CEA or CEl. After washout of muscimol, mice in which CEA or CEl had been inactivated showed normal acquisition and retrieval of conditioned fear responses. Twenty four hours after muscimol application, mice (n = 7 in each group) were reconditioned and tested for fear memory retrieval again 4 hrs later (CEA, 65.3 ± 8.% during exposure vs ± 6.6% before exposure; CEl, 57.7 ± 7.6% during exposure vs. 7.1 ± 4.9% before exposure). **P <
6 Freezing (%) 1 CEACtrl CEA CEl 5 BPY Musc ** ns *** *** ns ns *** ns *** *** *** ** Supplementary Figure 3 Differential role of CEl and in fear expression. Animals were fear conditioned in the absence of BPY or muscimolbpy and tested 4 hrs later. BPY or muscimolbpy was applied before animals were retested on the same day. Bar graphs illustrate freezing levels before and during ex posure in the presence and absence of BPY or muscimol. Consistent with the results obtained in naïve unconditioned animals, we found that inactivation of CEl in conditioned animals resulted in increased baseline (pre) freezing levels. However, when animals with CEl inactivation were exposed to the, they exhibited a further increase in freezing levels. This could be explained by several possibilities. First, it is possible that we did not inactivate the entire CEl, and that conditioned freezing was therefore not completely occluded by unconditioned freezing. Alternatively, our findings are also consistent with the notion that conditioned freezing might be elicited through other pathways (e.g. via disinhibition of output neurons by ITCs or by direct excitatory inputs from BLA or thalamus). Data were analyzed by one way ANOVA followed by posthoc Bonferroni ttest (vs. the no, notreatment group). CEACtrl, n = 7; CEA, n = 7;, n = 9; CEl, n = 8. Error bars indicate mean ± s.e.m. ns: not significant, **P <.1, ***P <
7 a c e Zscore Zscore 1 CEl 1.46 mm CEl on b Freezing (%) d Zscore Zscore no 1 *** CEl on CEl off f CEl off g Zscore i Zscore CEl on CEl off Time (ms) Time (ms) h j Zscore Zscore ms Time (ms) 35 ms Time (ms) Supplementary Figure 4 Characterization of fear conditioninginduced plasticity of evoked firing of CEl units. a, Coronal section of the am ygdala showing the location of the recordings sites of CEl on (green) and CEl off (blue) units. Numbers indicate the anteroposterior coordinates caudal to bregma. b, Summary graph illustrating changes in freezing behavior during fear conditioning. Comparing the first with the last reveals an increase in freezing behavior during conditioning (1 st : 6.3 ± 4.9% of the time spent freezing; 5 th : 6. ± 5.1%). c,d, Increased freezing levels were associated with significant changes in evoked activity in CEl on neurons (n = 15 neurons from 14 mice; note that zscored 1 responses do not reach significance) and CEl off neurons (n = 3 neurons from 17 mice). Note that not all units recorded before and after fear conditioning could be reliably identified during conditioning. e,f, Discrimination between and. Averaged and evoked responses of all CEl on units (n = 3) and CEl off units (n = 3) recorded from a mouse exhibiting behavioral discrimination. g,h, Analysis of response latencies of CEl on units. Normalized population peristimulus time histograms illustrating evoked responses of CEl on neurons (n = 5) before (left panel; onset latency of excitation: 115 ms) and after fear conditioning (right panel; onset latency: 115 ms). i,j, Analysis of response latencies of CEl off units. Normalized population peristimulus time histograms illustrating evoked responses of CEl off neurons (n = 41) before (left panel) and after fear conditioning (right panel; onset latency of inhibition; 354 ms). ***P <




8 a b CEl CEl Injection site c CEl HSVGFP d CEl Injection site HSVGFP Supplementary Figure 5 Retrograde tracing of CEl to projections. a, HSVGFP injection site in revealed by coinjection of fluorescent latex beads. Anteroposterior coordinates caudal to bregma: 1.34 mm. b, HSVGFP injection to reveals retrograde labeling of CEl neurons detected with antigfp immunostaining. c, HSVGFP injection site in CEl revealed by coinjection of fluorescent latex beads. Anteroposterior coordinates caudal to bregma: 1. mm. d, HSVGFP injection to CEl reveals local labeling in CEl as detected with antigfp immunostaining. Very few labeled neurons were observed in, indicating that the CEl to projection is unidirectional. Scale bars: 13 µm. 8


9 a CEl on b CEl on Frequency (Hz) c CEl off d Counts Frequency (Hz) Counts CEl off Supplementary Figure 6 CEl on and CEl off units functionally inhibit output neurons. a, Raster plot illustrating evoked responses of an identified CEl on unit. b, Crosscorrelation analysis reveals a short latency inhibitory interaction between the CEl on unit taken as a reference and a neuron. Dotted lines indicate 95% confidence intervals. Dashed vertical line indicates time of reference spikes from CEl on unit. Because the presynaptic neuron exhibited bursting activity, inhibition of the postsynaptic neuron appears to start before the presynaptic spike. c, Raster plot illustrating evoked responses of an identified CEl off unit. d, Crosscorrelation analysis reveals a short latency inhibitory interaction between the CEl off unit taken as a reference and a neuron. Dotted lines indicate 95% confidence intervals. Dashed vertical line indicates time of reference spikes from CEl off unit. Probabilites for finding inhibitory crosscorrelations between identified CEl units and output neurons were: CEl on pairs: 1/; CEl off pairs: /. These data were obtained from a single mouse. Because of the low n, additional multisite unit recordings or other approaches (paired recordings in acute slices) will be necessary in order to obtain a quantitative understanding of the connectivity between defined subtypes of CEl neurons and ouput neurons. 9

10 a d Zscore 3 1 CEl 1.46 mm Time (ms) b Zscore e Zscore ms Time (ms) 45 ms c Zscore Supplementary Figure 7 Characterization of fear conditioninginduced plasticity of evoked firing of units. a, Coronal section of the amygdala showing the location of the recordings sites of units. Numbers indicate the anteroposterior coordinates caudal to bregma. b, evoked firing of units changes during fear conditioning. Increased freezing levels were associated with significant changes in evoked activity in neurons (n = 7 neurons from 4 mice). Note that not all units recorded before and after fear conditioning could be reliably identified during conditioning. c, Discrimination between and. Averaged and evoked response of a unit recorded from a mouse exhibiting behavioral discrimination. d,e, Analysis of response latencies of units. Normalized population peristimulus time histograms illustrating evoked responses of neurons (n = 15) before (left panel; onset latency of excitation: 115 ms) and after fear conditioning (right panel; onset latencies: 115 ms and 455 ms). 1
11 TEST a BLA Thal. CELon CELoff inhibitory excitatory b HABITUATION / CELoff FEAR CONDITIONING DISCRIMINATION Freezing GENERALIZATION Supplementary Figure 8 Summary of main results. a, Functional connectivity of the CEA microcircuit. The CEl contains two distinct neuronal populations changing responses with fear conditioning: CElon neurons acquire an excitatory response, whereas neurons are inhibited by the. CEloff neurons express PKCδ; REF 1). These two populations are reciprocally connected, and in addition project to. Firing of neurons triggers freezing. In addition to these local inhibitory interactions within CEA, both CElon and neurons also receive shortlatency excitatory input, presumable from thalamus and/or BLA. Based on our data, we cannot exclude that CElon and CEloff neurons might target distinct subtypes of output neurons. b, Changes in activity of CEloff and neurons with discriminative fear conditioning. During habituation, both populations are spontaneously active but showed no response. After conditioning, CEloff neurons acquired an inhibitory response, whereas neurons increased their activity in response to the. Mice were split into a group with discriminating fear responses and one group displaying stimulus generalization. In discriminating animals, baseline firing rates where very similar to habituation (dashed grey line). In contrast, generalization was associated with changes in baseline firing of opposite polarity for the two neuronal populations: CEloff neurons increased tonic firing, whereas cells displayed decreased tonic activity. The tonic changes were of opposite direction to the phasic response in both neuronal populations, and increased the signaltonoise ratio of both and, leading to reduced neuronal discrimination between the two stimuli. 11
12 a b c Freezing (%) Freezing (%) 1 5 r =.43 p< Phasic activity (Zscore) r =.81 p> Tonic activity (Hz) r =.119 p< Phasic activity (Zscore) r =.7 p> Tonic activity (Hz) r =.98 p< Phasic activity (Zscore) r =.39 p> Tonic activity (Hz) Freezing (%) 1 5 r =.17 p> r =.1 p> r =.33 p> Tonic activity (Zscore) Tonic activity (Zscore) Tonic activity (Zscore) Supplementary Figure 9 Correlations of phasic and tonic activity with postconditioning freezing levels: raw data. a, Phasic zscored evoked responses of neurons (left), CEl off neurons (middle), and CEl on neurons (right) are highly correlated with behavioral freezing. Correlations were obtained by plotting the averaged zscored responses for all neurons of a given subtype for each animal over blocks of s. Plots include both and responses. P values indicate significance levels of Pearson s correlation coefficients. Dashed lines indicate 95% confidence intervals. b, Absolute levels of tonic activity do not correlate with freezing behavior. Correlations were obtained as described above. c, Zscored tonic activity does not correlate with freezing behavior. Correlations were obtained as described above. 1
13 Δ Phasic activity (Zscore) 1 5 r =.53 p< Δ Tonic activity (Hz) CEloff r =.17 p< Δ Tonic activity (Hz) 4 CElon r =.4 p> Δ Tonic activity (Hz) Supplementary Figure 1 Changes in tonic activity offset phasic responses. Direct correlation between fear conditioninginduced changes in tonic activity and in evoked phasic activity (zscored) in neurons (left, n = 15), CEl off neurons (middle, n = 41), and CEl on neurons (right, n = 5). For clarity, data were averaged and binned (bins contain equal number of data points). Linear correlations were performed using the nonbinned raw data. P values indicate significance levels of Pearson s correlation coefficients. Dashed lines indicate 95% confidence intervals. 13
14 Freezing (%) 1 5 r =.17 p> r =.1 p> r =.33 p> Tonic activity (Zscore) Tonic activity (Zscore) Tonic activity (Zscore) Supplementary Figure 11 Zscored tonic activity does not correlate with freezing behavior. Zscored tonic activity at test (4 hrs after fear conditioning) of neurons (left), CEl off neurons (middle), and CEl on neurons (right) is poorly correlated with behavioral freezing. Correlations were obtained by plotting the averaged zscored tonic activity levels for all neurons of a given subtype for each animal over blocks of s. For clarity, data were averaged and binned (bins contain equal number of data points). Linear correlations were performed using the nonbinned raw data. P values indicate significance levels of Pearson s correlation coefficients. Dashed lines indicate 95% confidence intervals. 14
15 Frequency (%) Hab. Test CEl off Hab. Test CEl on Hab. Test.5..5 Supplementary Figure 1 Changes in tonic activity during stimulation. Normalized population peristimulus ( ) time histograms of neurons (left), CEl off neurons (middle), and CEl on neurons (right). Fear conditioning was associated with decreased tonic activity in units (n = 15) and increased tonic activity in CEl off units (n = 41). 15
16 Tonic activity (Hz) (gen.) CEl off (gen.) CEl on (disc.) * *** *** * * * * ** ** pre pre pre Hab. Test Hab. Test Hab. Test Supplementary Figure 13 Changes in tonic activity across behavioral sessions. Plots show averaged absolute firing frequencies of, CEl off and CEl on units. Averages were obtained from behavioral trials (blocks of two ) in which changes in tonic activity were observed (e.g. trial blocks in which animals exhibited generalization for recordings, n = 8 trial blocks; CEl off, n = 3 trial blocks; or discrimination CEl on, n = 1 trial blocks). Spontaneous activity levels before onset ( pre ) were measured when animals were first placed in context B. 16

17 a b Absolute values CEloff CElon J3 DB J3 DB J3DB Channels w/o units J3 DB Supplementary Figure 14 Isolation of unit recordings. a, Left: Superimposed waveforms recorded from four different units. Right: Spikes originating from individual units were sorted using 3Dprincipal component analysis. b, Quantitative J3 and Davies Bouldin validity index (DB) statistics calculated for, CEl off and CEl on neurons. Controls values were obtained using two clusters defined from the centered cloud of points from channels in which no units could be detected. High values for the J3 and low values for the DB are indicative of good single unit isolation. 17
18 a b c Probability of freezing Probability of freezing Δ freezing probability Supplementary Figure 15 Time course of freezing behavior during stimulation. a, Freezing probability starts increasing early on during presentation. Freezing probability in relation to presentation in fear conditioned mice. Pooled data from mice with 1 presentations on the day following fear conditioning. Sampling period for freezing was 1 ms. The consisted of a series of 7 pips of 5 ms presented at.9 Hz. Vertical red lines indicate the occurrence of the individual pips composing the. The whole is displayed. b, Freezing probability to the first few pips is displayed, showing that freezing probability increases around 1.3 s after the onset of the. c, Freezing increase following onset can be fitted with an exponential curve rising to a maximum. Baseline freezing was subtracted from total freezing to yield freezing increase linked to presentation. The zero time point was set at 1.3 s following onset, when the freezing starts increasing. 18
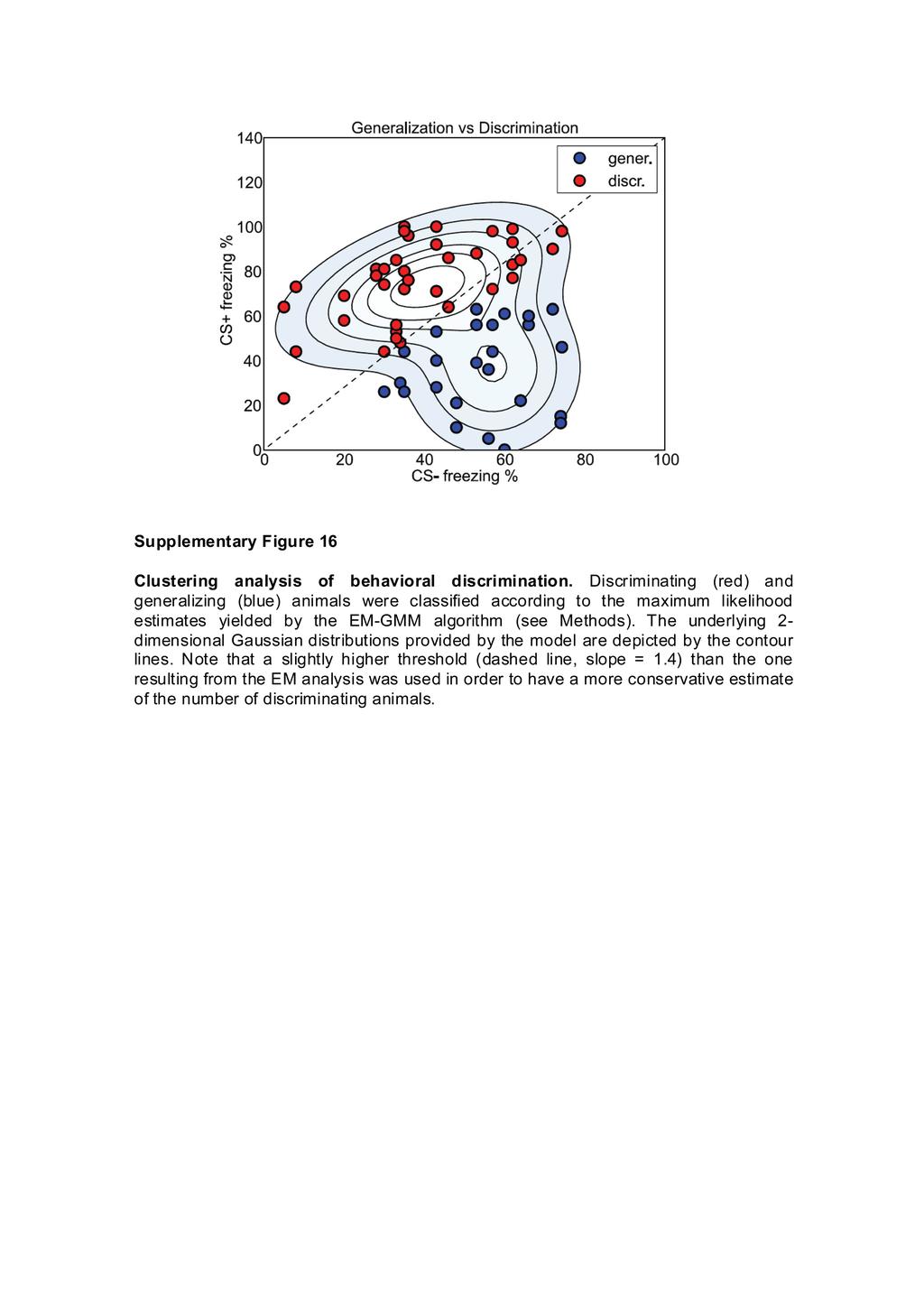

19 19
SUPPLEMENTARY INFORMATION
 doi:1.138/nature9553 Supplementary Table 1. Overlap of neuronal marker and PKC- expression in CEl. Marker/PKC- PKC- Marker Gad65 87.4±4.7 5.3±12.6 CRH 1.2±1. 16.9±15.2 Dyn 1.9±1.2 4.5±2.9 Enk 42.8±7.4
doi:1.138/nature9553 Supplementary Table 1. Overlap of neuronal marker and PKC- expression in CEl. Marker/PKC- PKC- Marker Gad65 87.4±4.7 5.3±12.6 CRH 1.2±1. 16.9±15.2 Dyn 1.9±1.2 4.5±2.9 Enk 42.8±7.4
Encoding of conditioned fear in central amygdala inhibitory circuits
 ARTICLE doi:1.138/nature9559 Encoding of conditioned fear in central amygdala inhibitory circuits Stephane Ciocchi 1 *, Cyril Herry 1 *{, François Grenier 1, Steffen B. E. Wolff 1, Johannes J. Letzkus
ARTICLE doi:1.138/nature9559 Encoding of conditioned fear in central amygdala inhibitory circuits Stephane Ciocchi 1 *, Cyril Herry 1 *{, François Grenier 1, Steffen B. E. Wolff 1, Johannes J. Letzkus
Supplementary Figure 1. Recording sites.
 Supplementary Figure 1 Recording sites. (a, b) Schematic of recording locations for mice used in the variable-reward task (a, n = 5) and the variable-expectation task (b, n = 5). RN, red nucleus. SNc,
Supplementary Figure 1 Recording sites. (a, b) Schematic of recording locations for mice used in the variable-reward task (a, n = 5) and the variable-expectation task (b, n = 5). RN, red nucleus. SNc,
Behavioral generalization
 Supplementary Figure 1 Behavioral generalization. a. Behavioral generalization curves in four Individual sessions. Shown is the conditioned response (CR, mean ± SEM), as a function of absolute (main) or
Supplementary Figure 1 Behavioral generalization. a. Behavioral generalization curves in four Individual sessions. Shown is the conditioned response (CR, mean ± SEM), as a function of absolute (main) or
Nature Neuroscience: doi: /nn Supplementary Figure 1. Diverse anorexigenic signals induce c-fos expression in CEl PKC-δ + neurons
 Supplementary Figure 1 Diverse anorexigenic signals induce c-fos expression in CEl PKC-δ + neurons a-c. Quantification of CEl c-fos expression in mice intraperitoneal injected with anorexigenic drugs (a),
Supplementary Figure 1 Diverse anorexigenic signals induce c-fos expression in CEl PKC-δ + neurons a-c. Quantification of CEl c-fos expression in mice intraperitoneal injected with anorexigenic drugs (a),
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature21682 Supplementary Table 1 Summary of statistical results from analyses. Mean quantity ± s.e.m. Test Days P-value Deg. Error deg. Total deg. χ 2 152 ± 14 active cells per day per mouse
doi:10.1038/nature21682 Supplementary Table 1 Summary of statistical results from analyses. Mean quantity ± s.e.m. Test Days P-value Deg. Error deg. Total deg. χ 2 152 ± 14 active cells per day per mouse
Nature Neuroscience: doi: /nn Supplementary Figure 1. Trial structure for go/no-go behavior
 Supplementary Figure 1 Trial structure for go/no-go behavior a, Overall timeline of experiments. Day 1: A1 mapping, injection of AAV1-SYN-GCAMP6s, cranial window and headpost implantation. Water restriction
Supplementary Figure 1 Trial structure for go/no-go behavior a, Overall timeline of experiments. Day 1: A1 mapping, injection of AAV1-SYN-GCAMP6s, cranial window and headpost implantation. Water restriction
Nature Neuroscience: doi: /nn.4642
 Supplementary Figure 1 Recording sites and example waveform clustering, as well as electrophysiological recordings of auditory CS and shock processing following overtraining. (a) Recording sites in LC
Supplementary Figure 1 Recording sites and example waveform clustering, as well as electrophysiological recordings of auditory CS and shock processing following overtraining. (a) Recording sites in LC
Supplementary figure 1: LII/III GIN-cells show morphological characteristics of MC
 1 2 1 3 Supplementary figure 1: LII/III GIN-cells show morphological characteristics of MC 4 5 6 7 (a) Reconstructions of LII/III GIN-cells with somato-dendritic compartments in orange and axonal arborizations
1 2 1 3 Supplementary figure 1: LII/III GIN-cells show morphological characteristics of MC 4 5 6 7 (a) Reconstructions of LII/III GIN-cells with somato-dendritic compartments in orange and axonal arborizations
Supplementary Figure 1
 Supplementary Figure 1 Miniature microdrive, spike sorting and sleep stage detection. a, A movable recording probe with 8-tetrodes (32-channels). It weighs ~1g. b, A mouse implanted with 8 tetrodes in
Supplementary Figure 1 Miniature microdrive, spike sorting and sleep stage detection. a, A movable recording probe with 8-tetrodes (32-channels). It weighs ~1g. b, A mouse implanted with 8 tetrodes in
Tuning properties of individual circuit components and stimulus-specificity of experience-driven changes.
 Supplementary Figure 1 Tuning properties of individual circuit components and stimulus-specificity of experience-driven changes. (a) Left, circuit schematic with the imaged component (L2/3 excitatory neurons)
Supplementary Figure 1 Tuning properties of individual circuit components and stimulus-specificity of experience-driven changes. (a) Left, circuit schematic with the imaged component (L2/3 excitatory neurons)
Nature Neuroscience: doi: /nn Supplementary Figure 1
 Supplementary Figure 1 Drd1a-Cre driven ChR2 expression in the SCN. (a) Low-magnification image of a representative Drd1a-ChR2 coronal brain section (n = 2) showing endogenous tdtomato fluorescence (magenta).
Supplementary Figure 1 Drd1a-Cre driven ChR2 expression in the SCN. (a) Low-magnification image of a representative Drd1a-ChR2 coronal brain section (n = 2) showing endogenous tdtomato fluorescence (magenta).
Nature Neuroscience: doi: /nn Supplementary Figure 1. ACx plasticity is required for fear conditioning.
 Supplementary Figure 1 ACx plasticity is required for fear conditioning. (a) Freezing time of conditioned and control mice before CS presentation and during CS presentation in a new context. Student s
Supplementary Figure 1 ACx plasticity is required for fear conditioning. (a) Freezing time of conditioned and control mice before CS presentation and during CS presentation in a new context. Student s
Supplementary Figure 1 Information on transgenic mouse models and their recording and optogenetic equipment. (a) 108 (b-c) (d) (e) (f) (g)
 108 (b-c) (d) (e) (f) (g)") Supplementary Figure 1 Information on transgenic mouse models and their recording and optogenetic equipment. (a) In four mice, cre-dependent expression of the hyperpolarizing opsin Arch in pyramidal cells
Supplementary Figure 1 Information on transgenic mouse models and their recording and optogenetic equipment. (a) In four mice, cre-dependent expression of the hyperpolarizing opsin Arch in pyramidal cells
Title: Plasticity of intrinsic excitability in mature granule cells of the dentate gyrus
 Title: Plasticity of intrinsic excitability in mature granule cells of the dentate gyrus Authors: Jeffrey Lopez-Rojas a1, Martin Heine b1 and Michael R. Kreutz ac1 a Research Group Neuroplasticity, b Research
Title: Plasticity of intrinsic excitability in mature granule cells of the dentate gyrus Authors: Jeffrey Lopez-Rojas a1, Martin Heine b1 and Michael R. Kreutz ac1 a Research Group Neuroplasticity, b Research
SUPPLEMENTARY INFORMATION
 doi:1.138/nature1139 a d Whisker angle (deg) Whisking repeatability Control Muscimol.4.3.2.1 -.1 8 4-4 1 2 3 4 Performance (d') Pole 8 4-4 1 2 3 4 5 Time (s) b Mean protraction angle (deg) e Hit rate (p
doi:1.138/nature1139 a d Whisker angle (deg) Whisking repeatability Control Muscimol.4.3.2.1 -.1 8 4-4 1 2 3 4 Performance (d') Pole 8 4-4 1 2 3 4 5 Time (s) b Mean protraction angle (deg) e Hit rate (p
Nature Neuroscience: doi: /nn.4335
 Supplementary Figure 1 Cholinergic neurons projecting to the VTA are concentrated in the caudal mesopontine region. (a) Schematic showing the sites of retrograde tracer injections in the VTA: cholera toxin
Supplementary Figure 1 Cholinergic neurons projecting to the VTA are concentrated in the caudal mesopontine region. (a) Schematic showing the sites of retrograde tracer injections in the VTA: cholera toxin
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION doi:10.1038/nature12024 entary Figure 1. Distribution of the number of earned cocaine Supplementary Figure 1. Distribution of the number of earned cocaine infusions in Shock-sensitive
SUPPLEMENTARY INFORMATION doi:10.1038/nature12024 entary Figure 1. Distribution of the number of earned cocaine Supplementary Figure 1. Distribution of the number of earned cocaine infusions in Shock-sensitive
Supplemental Information. A Visual-Cue-Dependent Memory Circuit. for Place Navigation
 Neuron, Volume 99 Supplemental Information A Visual-Cue-Dependent Memory Circuit for Place Navigation Han Qin, Ling Fu, Bo Hu, Xiang Liao, Jian Lu, Wenjing He, Shanshan Liang, Kuan Zhang, Ruijie Li, Jiwei
Neuron, Volume 99 Supplemental Information A Visual-Cue-Dependent Memory Circuit for Place Navigation Han Qin, Ling Fu, Bo Hu, Xiang Liao, Jian Lu, Wenjing He, Shanshan Liang, Kuan Zhang, Ruijie Li, Jiwei
Supplementary Figure 1
 Supplementary Figure 1 Arcuate ChIEF-tdTomato neurons expressed TH These micrographs show that TH-Cre-ChIEF-tdTomato (magenta), expressed by AAV in a TH-Cre mouse, were immunostained with TH (green) in
Supplementary Figure 1 Arcuate ChIEF-tdTomato neurons expressed TH These micrographs show that TH-Cre-ChIEF-tdTomato (magenta), expressed by AAV in a TH-Cre mouse, were immunostained with TH (green) in
SUPPLEMENTARY INFORMATION
 Supplementary Figure 1. Normal AMPAR-mediated fepsp input-output curve in CA3-Psen cdko mice. Input-output curves, which are plotted initial slopes of the evoked fepsp as function of the amplitude of the
Supplementary Figure 1. Normal AMPAR-mediated fepsp input-output curve in CA3-Psen cdko mice. Input-output curves, which are plotted initial slopes of the evoked fepsp as function of the amplitude of the
Supplementary Material for
 Supplementary Material for Selective neuronal lapses precede human cognitive lapses following sleep deprivation Supplementary Table 1. Data acquisition details Session Patient Brain regions monitored Time
Supplementary Material for Selective neuronal lapses precede human cognitive lapses following sleep deprivation Supplementary Table 1. Data acquisition details Session Patient Brain regions monitored Time
Supplementary Figure 1. Example of an amygdala neuron whose activity reflects value during the visual stimulus interval. This cell responded more
 1 Supplementary Figure 1. Example of an amygdala neuron whose activity reflects value during the visual stimulus interval. This cell responded more strongly when an image was negative than when the same
1 Supplementary Figure 1. Example of an amygdala neuron whose activity reflects value during the visual stimulus interval. This cell responded more strongly when an image was negative than when the same
SUPPLEMENTARY INFORMATION. Supplementary Figure 1
 SUPPLEMENTARY INFORMATION Supplementary Figure 1 The supralinear events evoked in CA3 pyramidal cells fulfill the criteria for NMDA spikes, exhibiting a threshold, sensitivity to NMDAR blockade, and all-or-none
SUPPLEMENTARY INFORMATION Supplementary Figure 1 The supralinear events evoked in CA3 pyramidal cells fulfill the criteria for NMDA spikes, exhibiting a threshold, sensitivity to NMDAR blockade, and all-or-none
Nature Neuroscience: doi: /nn Supplementary Figure 1
 Supplementary Figure 1 Bidirectional optogenetic modulation of the tonic activity of CEA PKCδ + neurons in vitro. a, Top, Cell-attached voltage recording illustrating the blue light-induced increase in
Supplementary Figure 1 Bidirectional optogenetic modulation of the tonic activity of CEA PKCδ + neurons in vitro. a, Top, Cell-attached voltage recording illustrating the blue light-induced increase in
Nature Neuroscience: doi: /nn Supplementary Figure 1. Confirmation that optogenetic inhibition of dopaminergic neurons affects choice
 Supplementary Figure 1 Confirmation that optogenetic inhibition of dopaminergic neurons affects choice (a) Sample behavioral trace as in Figure 1d, but with NpHR stimulation trials depicted as green blocks
Supplementary Figure 1 Confirmation that optogenetic inhibition of dopaminergic neurons affects choice (a) Sample behavioral trace as in Figure 1d, but with NpHR stimulation trials depicted as green blocks
Unique functional properties of somatostatin-expressing GABAergic neurons in mouse barrel cortex
 Supplementary Information Unique functional properties of somatostatin-expressing GABAergic neurons in mouse barrel cortex Luc Gentet, Yves Kremer, Hiroki Taniguchi, Josh Huang, Jochen Staiger and Carl
Supplementary Information Unique functional properties of somatostatin-expressing GABAergic neurons in mouse barrel cortex Luc Gentet, Yves Kremer, Hiroki Taniguchi, Josh Huang, Jochen Staiger and Carl
Supplementary Figure 1
 Supplementary Figure 1 Localization of virus injections. (a) Schematic showing the approximate center of AAV-DIO-ChR2-YFP injection sites in the NAc of Dyn-cre mice (n=8 mice, 16 injections; caudate/putamen,
Supplementary Figure 1 Localization of virus injections. (a) Schematic showing the approximate center of AAV-DIO-ChR2-YFP injection sites in the NAc of Dyn-cre mice (n=8 mice, 16 injections; caudate/putamen,
Supplementary Figure 1
 8w Pia II/III IV V VI PV EYFP EYFP PV EYFP PV d PV EYFP Supplementary Figure a Spike probability x - PV-Cre d Spike probability x - RS RS b e Spike probability Spike probability.6......8..... FS FS c f
8w Pia II/III IV V VI PV EYFP EYFP PV EYFP PV d PV EYFP Supplementary Figure a Spike probability x - PV-Cre d Spike probability x - RS RS b e Spike probability Spike probability.6......8..... FS FS c f
Cellular Neurobiology BIPN140
 Cellular Neurobiology BIPN140 1st Midterm Exam Ready for Pickup By the elevator on the 3 rd Floor of Pacific Hall (waiver) Exam Depot Window at the north entrance to Pacific Hall (no waiver) Mon-Fri, 10:00
Cellular Neurobiology BIPN140 1st Midterm Exam Ready for Pickup By the elevator on the 3 rd Floor of Pacific Hall (waiver) Exam Depot Window at the north entrance to Pacific Hall (no waiver) Mon-Fri, 10:00
Nature Neuroscience: doi: /nn Supplementary Figure 1
 Supplementary Figure 1 Atlas representations of the midcingulate (MCC) region targeted in this study compared against the anterior cingulate (ACC) region commonly reported. Coronal sections are shown on
Supplementary Figure 1 Atlas representations of the midcingulate (MCC) region targeted in this study compared against the anterior cingulate (ACC) region commonly reported. Coronal sections are shown on
Supplementary Materials for
 advances.sciencemag.org/cgi/content/full/3/3/e1600955/dc1 Supplementary Materials for Flexible and stretchable nanowire-coated fibers for optoelectronic probing of spinal cord circuits Chi Lu, Seongjun
advances.sciencemag.org/cgi/content/full/3/3/e1600955/dc1 Supplementary Materials for Flexible and stretchable nanowire-coated fibers for optoelectronic probing of spinal cord circuits Chi Lu, Seongjun
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature10776 Supplementary Information 1: Influence of inhibition among blns on STDP of KC-bLN synapses (simulations and schematics). Unconstrained STDP drives network activity to saturation
doi:10.1038/nature10776 Supplementary Information 1: Influence of inhibition among blns on STDP of KC-bLN synapses (simulations and schematics). Unconstrained STDP drives network activity to saturation
Supplementary Figure 1. GABA depolarizes the majority of immature neurons in the
 Supplementary Figure 1. GABA depolarizes the majority of immature neurons in the upper cortical layers at P3 4 in vivo. (a b) Cell-attached current-clamp recordings illustrate responses to puff-applied
Supplementary Figure 1. GABA depolarizes the majority of immature neurons in the upper cortical layers at P3 4 in vivo. (a b) Cell-attached current-clamp recordings illustrate responses to puff-applied
Bursting dynamics in the brain. Jaeseung Jeong, Department of Biosystems, KAIST
 Bursting dynamics in the brain Jaeseung Jeong, Department of Biosystems, KAIST Tonic and phasic activity A neuron is said to exhibit a tonic activity when it fires a series of single action potentials
Bursting dynamics in the brain Jaeseung Jeong, Department of Biosystems, KAIST Tonic and phasic activity A neuron is said to exhibit a tonic activity when it fires a series of single action potentials
Analysis of in-vivo extracellular recordings. Ryan Morrill Bootcamp 9/10/2014
 Analysis of in-vivo extracellular recordings Ryan Morrill Bootcamp 9/10/2014 Goals for the lecture Be able to: Conceptually understand some of the analysis and jargon encountered in a typical (sensory)
Analysis of in-vivo extracellular recordings Ryan Morrill Bootcamp 9/10/2014 Goals for the lecture Be able to: Conceptually understand some of the analysis and jargon encountered in a typical (sensory)
Electrophysiological and firing properties of neurons: categorizing soloists and choristers in primary visual cortex
 *Manuscript Click here to download Manuscript: Manuscript revised.docx Click here to view linked Referenc Electrophysiological and firing properties of neurons: categorizing soloists and choristers in
*Manuscript Click here to download Manuscript: Manuscript revised.docx Click here to view linked Referenc Electrophysiological and firing properties of neurons: categorizing soloists and choristers in
Supplementary Table I Blood pressure and heart rate measurements pre- and post-stroke
 SUPPLEMENTARY INFORMATION doi:10.1038/nature09511 Supplementary Table I Blood pressure and heart rate measurements pre- and post-stroke Pre Post 7-days Systolic Diastolic BPM Systolic Diastolic BPM Systolic
SUPPLEMENTARY INFORMATION doi:10.1038/nature09511 Supplementary Table I Blood pressure and heart rate measurements pre- and post-stroke Pre Post 7-days Systolic Diastolic BPM Systolic Diastolic BPM Systolic
Nature Neuroscience: doi: /nn Supplementary Figure 1. Behavioral training.
 Supplementary Figure 1 Behavioral training. a, Mazes used for behavioral training. Asterisks indicate reward location. Only some example mazes are shown (for example, right choice and not left choice maze
Supplementary Figure 1 Behavioral training. a, Mazes used for behavioral training. Asterisks indicate reward location. Only some example mazes are shown (for example, right choice and not left choice maze
File name: Supplementary Information Description: Supplementary Figures, Supplementary Table and Supplementary References
 File name: Supplementary Information Description: Supplementary Figures, Supplementary Table and Supplementary References File name: Supplementary Data 1 Description: Summary datasheets showing the spatial
File name: Supplementary Information Description: Supplementary Figures, Supplementary Table and Supplementary References File name: Supplementary Data 1 Description: Summary datasheets showing the spatial
Astrocyte signaling controls spike timing-dependent depression at neocortical synapses
 Supplementary Information Astrocyte signaling controls spike timing-dependent depression at neocortical synapses Rogier Min and Thomas Nevian Department of Physiology, University of Berne, Bern, Switzerland
Supplementary Information Astrocyte signaling controls spike timing-dependent depression at neocortical synapses Rogier Min and Thomas Nevian Department of Physiology, University of Berne, Bern, Switzerland
Cholinergic Activation of M2 Receptors Leads to Context- Dependent Modulation of Feedforward Inhibition in the Visual Thalamus
 Cholinergic Activation of M2 Receptors Leads to Context- Dependent Modulation of Feedforward Inhibition in the Visual Thalamus Miklos Antal., Claudio Acuna-Goycolea., R. Todd Pressler, Dawn M. Blitz, Wade
Cholinergic Activation of M2 Receptors Leads to Context- Dependent Modulation of Feedforward Inhibition in the Visual Thalamus Miklos Antal., Claudio Acuna-Goycolea., R. Todd Pressler, Dawn M. Blitz, Wade
A genetically targeted optical sensor to monitor calcium signals in astrocyte processes
 A genetically targeted optical sensor to monitor calcium signals in astrocyte processes 1 Eiji Shigetomi, 1 Sebastian Kracun, 2 Michael V. Sofroniew & 1,2 *Baljit S. Khakh Ψ 1 Departments of Physiology
A genetically targeted optical sensor to monitor calcium signals in astrocyte processes 1 Eiji Shigetomi, 1 Sebastian Kracun, 2 Michael V. Sofroniew & 1,2 *Baljit S. Khakh Ψ 1 Departments of Physiology
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION doi:10.1038/nature11306 Supplementary Figures Supplementary Figure 1. Basic characterization of GFP+ RGLs in the dentate gyrus of adult nestin-gfp mice. a, Sample confocal images
SUPPLEMENTARY INFORMATION doi:10.1038/nature11306 Supplementary Figures Supplementary Figure 1. Basic characterization of GFP+ RGLs in the dentate gyrus of adult nestin-gfp mice. a, Sample confocal images
The storage and recall of memories in the hippocampo-cortical system. Supplementary material. Edmund T Rolls
 The storage and recall of memories in the hippocampo-cortical system Supplementary material Edmund T Rolls Oxford Centre for Computational Neuroscience, Oxford, England and University of Warwick, Department
The storage and recall of memories in the hippocampo-cortical system Supplementary material Edmund T Rolls Oxford Centre for Computational Neuroscience, Oxford, England and University of Warwick, Department
Supplemental Information. In Vivo Optogenetic Stimulation. of Neocortical Excitatory Neurons. Drives Brain-State-Dependent Inhibition
 Current Biology, Volume 21 Supplemental Information In Vivo Optogenetic Stimulation of Neocortical Excitatory Neurons Drives Brain-State-Dependent Inhibition Celine Mateo, Michael Avermann, Luc J. Gentet,
Current Biology, Volume 21 Supplemental Information In Vivo Optogenetic Stimulation of Neocortical Excitatory Neurons Drives Brain-State-Dependent Inhibition Celine Mateo, Michael Avermann, Luc J. Gentet,
Spiking Inputs to a Winner-take-all Network
 Spiking Inputs to a Winner-take-all Network Matthias Oster and Shih-Chii Liu Institute of Neuroinformatics University of Zurich and ETH Zurich Winterthurerstrasse 9 CH-857 Zurich, Switzerland {mao,shih}@ini.phys.ethz.ch
Spiking Inputs to a Winner-take-all Network Matthias Oster and Shih-Chii Liu Institute of Neuroinformatics University of Zurich and ETH Zurich Winterthurerstrasse 9 CH-857 Zurich, Switzerland {mao,shih}@ini.phys.ethz.ch
Theta sequences are essential for internally generated hippocampal firing fields.
 Theta sequences are essential for internally generated hippocampal firing fields. Yingxue Wang, Sandro Romani, Brian Lustig, Anthony Leonardo, Eva Pastalkova Supplementary Materials Supplementary Modeling
Theta sequences are essential for internally generated hippocampal firing fields. Yingxue Wang, Sandro Romani, Brian Lustig, Anthony Leonardo, Eva Pastalkova Supplementary Materials Supplementary Modeling
Thalamo-Cortical Relationships Ultrastructure of Thalamic Synaptic Glomerulus
 Central Visual Pathways V1/2 NEUR 3001 dvanced Visual Neuroscience The Lateral Geniculate Nucleus () is more than a relay station LP SC Professor Tom Salt UCL Institute of Ophthalmology Retina t.salt@ucl.ac.uk
Central Visual Pathways V1/2 NEUR 3001 dvanced Visual Neuroscience The Lateral Geniculate Nucleus () is more than a relay station LP SC Professor Tom Salt UCL Institute of Ophthalmology Retina t.salt@ucl.ac.uk
Nature Methods: doi: /nmeth Supplementary Figure 1. Activity in turtle dorsal cortex is sparse.
 Supplementary Figure 1 Activity in turtle dorsal cortex is sparse. a. Probability distribution of firing rates across the population (notice log scale) in our data. The range of firing rates is wide but
Supplementary Figure 1 Activity in turtle dorsal cortex is sparse. a. Probability distribution of firing rates across the population (notice log scale) in our data. The range of firing rates is wide but
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature22324 Effects of photoinhibition on licking Photoinhibition of ALM or thalamus caused only small changes in lick early rates, no response rates, and licking latency. ALM photoinhibition
doi:10.1038/nature22324 Effects of photoinhibition on licking Photoinhibition of ALM or thalamus caused only small changes in lick early rates, no response rates, and licking latency. ALM photoinhibition
Sum of Neurally Distinct Stimulus- and Task-Related Components.
 SUPPLEMENTARY MATERIAL for Cardoso et al. 22 The Neuroimaging Signal is a Linear Sum of Neurally Distinct Stimulus- and Task-Related Components. : Appendix: Homogeneous Linear ( Null ) and Modified Linear
SUPPLEMENTARY MATERIAL for Cardoso et al. 22 The Neuroimaging Signal is a Linear Sum of Neurally Distinct Stimulus- and Task-Related Components. : Appendix: Homogeneous Linear ( Null ) and Modified Linear
doi: /nature09554
 SUPPLEMENTARY INFORMATION doi:10.1038/nature09554 Supplementary Figure 1: Optical Tracing with New Photoactivatable GFP Variants Reveals Enhanced Labeling of Neuronal Processes We qualitatively compare
SUPPLEMENTARY INFORMATION doi:10.1038/nature09554 Supplementary Figure 1: Optical Tracing with New Photoactivatable GFP Variants Reveals Enhanced Labeling of Neuronal Processes We qualitatively compare
In Vitro Analog of Operant Conditioning in Aplysia
 The Journal of Neuroscience, March 15, 1999, 19(6):2261 2272 In Vitro Analog of Operant Conditioning in Aplysia. II. Modifications of the Functional Dynamics of an Identified Neuron Contribute to Motor
The Journal of Neuroscience, March 15, 1999, 19(6):2261 2272 In Vitro Analog of Operant Conditioning in Aplysia. II. Modifications of the Functional Dynamics of an Identified Neuron Contribute to Motor
Chapter 4. Activity of human hippocampal and amygdala neurons during retrieval of declarative memories
 131 Chapter 4. Activity of human hippocampal and amygdala neurons during retrieval of declarative memories 4.1 Introduction 4 Episodic memories allow us to remember not only whether we have seen something
131 Chapter 4. Activity of human hippocampal and amygdala neurons during retrieval of declarative memories 4.1 Introduction 4 Episodic memories allow us to remember not only whether we have seen something
Two distinct mechanisms for experiencedependent
 SUPPLEMENTARY INFORMATION Articles https://doi.org/10.1038/s41593-018-0150-0 In the format provided by the authors and unedited. Two distinct mechanisms for experiencedependent homeostasis Michelle C.
SUPPLEMENTARY INFORMATION Articles https://doi.org/10.1038/s41593-018-0150-0 In the format provided by the authors and unedited. Two distinct mechanisms for experiencedependent homeostasis Michelle C.
Supplementary Figure 1. SybII and Ceb are sorted to distinct vesicle populations in astrocytes. Nature Neuroscience: doi: /nn.
 Supplementary Figure 1 SybII and Ceb are sorted to distinct vesicle populations in astrocytes. (a) Exemplary images for cultured astrocytes co-immunolabeled with SybII and Ceb antibodies. SybII accumulates
Supplementary Figure 1 SybII and Ceb are sorted to distinct vesicle populations in astrocytes. (a) Exemplary images for cultured astrocytes co-immunolabeled with SybII and Ceb antibodies. SybII accumulates
Profound Context-Dependent Plasticity of Mitral Cell Responses in Olfactory Bulb
 Profound Context-Dependent Plasticity of Mitral Cell Responses in Olfactory Bulb Wilder Doucette, Diego Restrepo * PLoS BIOLOGY Department of Cell and Developmental Biology, Neuroscience Program and Rocky
Profound Context-Dependent Plasticity of Mitral Cell Responses in Olfactory Bulb Wilder Doucette, Diego Restrepo * PLoS BIOLOGY Department of Cell and Developmental Biology, Neuroscience Program and Rocky
Nature Neuroscience: doi: /nn Supplementary Figure 1. Splenic atrophy and leucopenia caused by T3 SCI.
 Supplementary Figure 1 Splenic atrophy and leucopenia caused by T3 SCI. (a) Gross anatomy of representative spleens from control and T3 SCI mice at 28 days post-injury. (b and c) Hematoxylin and eosin
Supplementary Figure 1 Splenic atrophy and leucopenia caused by T3 SCI. (a) Gross anatomy of representative spleens from control and T3 SCI mice at 28 days post-injury. (b and c) Hematoxylin and eosin
Shadowing and Blocking as Learning Interference Models
 Shadowing and Blocking as Learning Interference Models Espoir Kyubwa Dilip Sunder Raj Department of Bioengineering Department of Neuroscience University of California San Diego University of California
Shadowing and Blocking as Learning Interference Models Espoir Kyubwa Dilip Sunder Raj Department of Bioengineering Department of Neuroscience University of California San Diego University of California
Dep. Control Time (min)
") aa Control Dep. RP 1s 1 mv 2s 1 mv b % potentiation of IPSP 2 15 1 5 Dep. * 1 2 3 4 Time (min) Supplementary Figure 1. Rebound potentiation of IPSPs in PCs. a, IPSPs recorded with a K + gluconate pipette
aa Control Dep. RP 1s 1 mv 2s 1 mv b % potentiation of IPSP 2 15 1 5 Dep. * 1 2 3 4 Time (min) Supplementary Figure 1. Rebound potentiation of IPSPs in PCs. a, IPSPs recorded with a K + gluconate pipette
Spike Sorting and Behavioral analysis software
 Spike Sorting and Behavioral analysis software Ajinkya Kokate Department of Computational Science University of California, San Diego La Jolla, CA 92092 akokate@ucsd.edu December 14, 2012 Abstract In this
Spike Sorting and Behavioral analysis software Ajinkya Kokate Department of Computational Science University of California, San Diego La Jolla, CA 92092 akokate@ucsd.edu December 14, 2012 Abstract In this
Active Control of Spike-Timing Dependent Synaptic Plasticity in an Electrosensory System
 Active Control of Spike-Timing Dependent Synaptic Plasticity in an Electrosensory System Patrick D. Roberts and Curtis C. Bell Neurological Sciences Institute, OHSU 505 N.W. 185 th Avenue, Beaverton, OR
Active Control of Spike-Timing Dependent Synaptic Plasticity in an Electrosensory System Patrick D. Roberts and Curtis C. Bell Neurological Sciences Institute, OHSU 505 N.W. 185 th Avenue, Beaverton, OR
Lecture 1: Neurons. Lecture 2: Coding with spikes. To gain a basic understanding of spike based neural codes
 Lecture : Neurons Lecture 2: Coding with spikes Learning objectives: To gain a basic understanding of spike based neural codes McCulloch Pitts Neuron I w in Σ out Θ Examples: I = ; θ =.5; w=. - in = *.
Lecture : Neurons Lecture 2: Coding with spikes Learning objectives: To gain a basic understanding of spike based neural codes McCulloch Pitts Neuron I w in Σ out Θ Examples: I = ; θ =.5; w=. - in = *.
OPTO 5320 VISION SCIENCE I
 OPTO 5320 VISION SCIENCE I Monocular Sensory Processes of Vision: Color Vision Mechanisms of Color Processing . Neural Mechanisms of Color Processing A. Parallel processing - M- & P- pathways B. Second
OPTO 5320 VISION SCIENCE I Monocular Sensory Processes of Vision: Color Vision Mechanisms of Color Processing . Neural Mechanisms of Color Processing A. Parallel processing - M- & P- pathways B. Second
SUPPLEMENTARY INFORMATION
 doi: 1.138/nature588 SUPPLEMENTARY INFORMATION Supplemental Information Sensory neuron sodium channel Na v 1.8 is essential for pain at cold temperatures Katharina Zimmermann*, Andreas Leffler*, Alexandru
doi: 1.138/nature588 SUPPLEMENTARY INFORMATION Supplemental Information Sensory neuron sodium channel Na v 1.8 is essential for pain at cold temperatures Katharina Zimmermann*, Andreas Leffler*, Alexandru
Social transmission and buffering of synaptic changes after stress
 SUPPLEMENTARY INFORMATION Articles https://doi.org/10.1038/s41593-017-0044-6 In the format provided by the authors and unedited. Social transmission and buffering of synaptic changes after stress Toni-Lee
SUPPLEMENTARY INFORMATION Articles https://doi.org/10.1038/s41593-017-0044-6 In the format provided by the authors and unedited. Social transmission and buffering of synaptic changes after stress Toni-Lee
NSCI 324 Systems Neuroscience
 NSCI 324 Systems Neuroscience Dopamine and Learning Michael Dorris Associate Professor of Physiology & Neuroscience Studies dorrism@biomed.queensu.ca http://brain.phgy.queensu.ca/dorrislab/ NSCI 324 Systems
NSCI 324 Systems Neuroscience Dopamine and Learning Michael Dorris Associate Professor of Physiology & Neuroscience Studies dorrism@biomed.queensu.ca http://brain.phgy.queensu.ca/dorrislab/ NSCI 324 Systems
-51mV 30s 3mV. n=14 n=4 p=0.4. Depolarization (mv) 3
 3") Supplementary Figure 1 a optoβ 2 -AR b ChR2-51mV 30s 3mV -50mV 30s 3mV c 4 n=14 n=4 p=0.4 Depolarization (mv) 3 2 1 0 Both optogenetic actuators, optoβ 2 AR and ChR2, were effective in stimulating astrocytes
Supplementary Figure 1 a optoβ 2 -AR b ChR2-51mV 30s 3mV -50mV 30s 3mV c 4 n=14 n=4 p=0.4 Depolarization (mv) 3 2 1 0 Both optogenetic actuators, optoβ 2 AR and ChR2, were effective in stimulating astrocytes
Increased serotonin transporter expression reduces fear and recruitment of. parvalbumin interneurons of the amygdala
 Increased serotonin transporter expression reduces fear and recruitment of parvalbumin interneurons of the amygdala Marco Bocchio, Giulia Fucsina, Lydia Oikonomidis, Stephen B McHugh, David M Bannerman,
Increased serotonin transporter expression reduces fear and recruitment of parvalbumin interneurons of the amygdala Marco Bocchio, Giulia Fucsina, Lydia Oikonomidis, Stephen B McHugh, David M Bannerman,
SUPPLEMENTARY INFORMATION. Rett Syndrome Mutation MeCP2 T158A Disrupts DNA Binding, Protein Stability and ERP Responses
 SUPPLEMENTARY INFORMATION Rett Syndrome Mutation T158A Disrupts DNA Binding, Protein Stability and ERP Responses Darren Goffin, Megan Allen, Le Zhang, Maria Amorim, I-Ting Judy Wang, Arith-Ruth S. Reyes,
SUPPLEMENTARY INFORMATION Rett Syndrome Mutation T158A Disrupts DNA Binding, Protein Stability and ERP Responses Darren Goffin, Megan Allen, Le Zhang, Maria Amorim, I-Ting Judy Wang, Arith-Ruth S. Reyes,
MECHANISMS INVOLVED IN STORAGE OF FEAR IN LATERAL AMYGDALA A COMPUTATIONAL STUDY
 MECHANISMS INVOLVED IN STORAGE OF FEAR IN LATERAL AMYGDALA A COMPUTATIONAL STUDY A Thesis presented to the Faculty of the Graduate School University of Missouri Columbia In Partial Fulfillment of the Requirement
MECHANISMS INVOLVED IN STORAGE OF FEAR IN LATERAL AMYGDALA A COMPUTATIONAL STUDY A Thesis presented to the Faculty of the Graduate School University of Missouri Columbia In Partial Fulfillment of the Requirement
Supporting Information
 ATP from synaptic terminals and astrocytes regulates NMDA receptors and synaptic plasticity through PSD- 95 multi- protein complex U.Lalo, O.Palygin, A.Verkhratsky, S.G.N. Grant and Y. Pankratov Supporting
ATP from synaptic terminals and astrocytes regulates NMDA receptors and synaptic plasticity through PSD- 95 multi- protein complex U.Lalo, O.Palygin, A.Verkhratsky, S.G.N. Grant and Y. Pankratov Supporting
NIH Public Access Author Manuscript Nat Neurosci. Author manuscript; available in PMC 2010 October 1.
 NIH Public Access Author Manuscript Published in final edited form as: Nat Neurosci. 2010 April ; 13(4): 489 494. doi:10.1038/nn.2499. SYNAPTIC CORRELATES OF FEAR EXTINCTION IN THE AMYGDALA Taiju Amano
NIH Public Access Author Manuscript Published in final edited form as: Nat Neurosci. 2010 April ; 13(4): 489 494. doi:10.1038/nn.2499. SYNAPTIC CORRELATES OF FEAR EXTINCTION IN THE AMYGDALA Taiju Amano
Theme 2: Cellular mechanisms in the Cochlear Nucleus
 Theme 2: Cellular mechanisms in the Cochlear Nucleus The Cochlear Nucleus (CN) presents a unique opportunity for quantitatively studying input-output transformations by neurons because it gives rise to
Theme 2: Cellular mechanisms in the Cochlear Nucleus The Cochlear Nucleus (CN) presents a unique opportunity for quantitatively studying input-output transformations by neurons because it gives rise to
Nature Neuroscience doi: /nn Supplementary Figure 1. Characterization of viral injections.
 Supplementary Figure 1 Characterization of viral injections. (a) Dorsal view of a mouse brain (dashed white outline) after receiving a large, unilateral thalamic injection (~100 nl); demonstrating that
Supplementary Figure 1 Characterization of viral injections. (a) Dorsal view of a mouse brain (dashed white outline) after receiving a large, unilateral thalamic injection (~100 nl); demonstrating that
Microcircuitry coordination of cortical motor information in self-initiation of voluntary movements
 Y. Isomura et al. 1 Microcircuitry coordination of cortical motor information in self-initiation of voluntary movements Yoshikazu Isomura, Rie Harukuni, Takashi Takekawa, Hidenori Aizawa & Tomoki Fukai
Y. Isomura et al. 1 Microcircuitry coordination of cortical motor information in self-initiation of voluntary movements Yoshikazu Isomura, Rie Harukuni, Takashi Takekawa, Hidenori Aizawa & Tomoki Fukai
Supplementary Figure 1. Basic properties of compound EPSPs at
 Supplementary Figure 1. Basic properties of compound EPSPs at hippocampal CA3 CA3 cell synapses. (a) EPSPs were evoked by extracellular stimulation of the recurrent collaterals and pharmacologically isolated
Supplementary Figure 1. Basic properties of compound EPSPs at hippocampal CA3 CA3 cell synapses. (a) EPSPs were evoked by extracellular stimulation of the recurrent collaterals and pharmacologically isolated
Systems Neuroscience November 29, Memory
 Systems Neuroscience November 29, 2016 Memory Gabriela Michel http: www.ini.unizh.ch/~kiper/system_neurosci.html Forms of memory Different types of learning & memory rely on different brain structures
Systems Neuroscience November 29, 2016 Memory Gabriela Michel http: www.ini.unizh.ch/~kiper/system_neurosci.html Forms of memory Different types of learning & memory rely on different brain structures
Short- and long-lasting consequences of in vivo nicotine treatment
 Short- and long-lasting consequences of in vivo nicotine treatment on hippocampal excitability Rachel E. Penton, Michael W. Quick, Robin A. J. Lester Supplementary Figure 1. Histogram showing the maximal
Short- and long-lasting consequences of in vivo nicotine treatment on hippocampal excitability Rachel E. Penton, Michael W. Quick, Robin A. J. Lester Supplementary Figure 1. Histogram showing the maximal
SUPPLEMENTARY INFORMATION
 doi: 10.1038/nature06310 SUPPLEMENTARY INFORMATION www.nature.com/nature 1 www.nature.com/nature 2 www.nature.com/nature 3 Supplementary Figure S1 Spontaneous duration of wake, SWS and REM sleep (expressed
doi: 10.1038/nature06310 SUPPLEMENTARY INFORMATION www.nature.com/nature 1 www.nature.com/nature 2 www.nature.com/nature 3 Supplementary Figure S1 Spontaneous duration of wake, SWS and REM sleep (expressed
The control of spiking by synaptic input in striatal and pallidal neurons
 The control of spiking by synaptic input in striatal and pallidal neurons Dieter Jaeger Department of Biology, Emory University, Atlanta, GA 30322 Key words: Abstract: rat, slice, whole cell, dynamic current
The control of spiking by synaptic input in striatal and pallidal neurons Dieter Jaeger Department of Biology, Emory University, Atlanta, GA 30322 Key words: Abstract: rat, slice, whole cell, dynamic current
Supplementary Figure 1: Validation of labeling specificity of immature OSNs and presynaptic terminals. (A) (B) (C) (D) (E)
 (B) (C) (D) (E)") Supplementary Figure 1: Validation of labeling specificity of immature OSNs and presynaptic terminals. (A) Confocal images of septal olfactory epithelium of an adult Gγ8-sypGFP-tdTom mouse showing colocalization
Supplementary Figure 1: Validation of labeling specificity of immature OSNs and presynaptic terminals. (A) Confocal images of septal olfactory epithelium of an adult Gγ8-sypGFP-tdTom mouse showing colocalization
Hong Jiang 1, Dong Fang 1, Ling-Yu Kong 1, Zi-Run Jin 1, Jie Cai 1, Xue-Jing Kang 1, You Wan 2,3* and Guo-Gang Xing 1,2,3*
 Jiang et al. Molecular Brain 2014, 7:72 RESEARCH Open Access Sensitization of neurons in the central nucleus of the amygdala via the decreased GABAergic inhibition contributes to the development of neuropathic
Jiang et al. Molecular Brain 2014, 7:72 RESEARCH Open Access Sensitization of neurons in the central nucleus of the amygdala via the decreased GABAergic inhibition contributes to the development of neuropathic
An acetylcholine-activated microcircuit drives temporal dynamics of cortical activity
 An acetylcholine-activated microcircuit drives temporal dynamics of cortical activity Naiyan Chen, Hiroki Sugihara, & Mriganka Sur Nature America, nc. All rights reserved. Cholinergic modulation of cortex
An acetylcholine-activated microcircuit drives temporal dynamics of cortical activity Naiyan Chen, Hiroki Sugihara, & Mriganka Sur Nature America, nc. All rights reserved. Cholinergic modulation of cortex
Supralinear increase of recurrent inhibition during sparse activity in the somatosensory cortex
 Supralinear increase of recurrent inhibition during sparse activity in the somatosensory cortex Christoph Kapfer 1,2, Lindsey L Glickfeld 1,3, Bassam V Atallah 1,3 & Massimo Scanziani 1 The balance between
Supralinear increase of recurrent inhibition during sparse activity in the somatosensory cortex Christoph Kapfer 1,2, Lindsey L Glickfeld 1,3, Bassam V Atallah 1,3 & Massimo Scanziani 1 The balance between
Hormonal gain control of a medial preoptic area social reward circuit
 CORRECTION NOTICE Nat. Neurosci. 20, 449 458 (2017) Hormonal gain control of a medial preoptic area social reward circuit Jenna A McHenry, James M Otis, Mark A Rossi, J Elliott Robinson, Oksana Kosyk,
CORRECTION NOTICE Nat. Neurosci. 20, 449 458 (2017) Hormonal gain control of a medial preoptic area social reward circuit Jenna A McHenry, James M Otis, Mark A Rossi, J Elliott Robinson, Oksana Kosyk,
Brief presynaptic bursts evoke synapse-specific retrograde inhibition mediated by endogenous cannabinoids
 Brief presynaptic bursts evoke synapse-specific retrograde inhibition mediated by endogenous cannabinoids Solange P Brown 1 3,Stephan D Brenowitz 1,3 & Wade G Regehr 1 Many types of neurons can release
Brief presynaptic bursts evoke synapse-specific retrograde inhibition mediated by endogenous cannabinoids Solange P Brown 1 3,Stephan D Brenowitz 1,3 & Wade G Regehr 1 Many types of neurons can release
A general error-based spike-timing dependent learning rule for the Neural Engineering Framework
 A general error-based spike-timing dependent learning rule for the Neural Engineering Framework Trevor Bekolay Monday, May 17, 2010 Abstract Previous attempts at integrating spike-timing dependent plasticity
A general error-based spike-timing dependent learning rule for the Neural Engineering Framework Trevor Bekolay Monday, May 17, 2010 Abstract Previous attempts at integrating spike-timing dependent plasticity
Supplementary information for:
 Supplementary information for: Independent hypothalamic circuits for social and predator fear Bianca A. Silva, Camilla Mattucci, Piotr Krzywkowski, Emanuele Murana, Anna Illarionova, Valery Grinevich,
Supplementary information for: Independent hypothalamic circuits for social and predator fear Bianca A. Silva, Camilla Mattucci, Piotr Krzywkowski, Emanuele Murana, Anna Illarionova, Valery Grinevich,
SUPPLEMENTARY INFORMATION
 doi: 1.138/nature6416 Supplementary Notes Spine Ca 2+ signals produced by glutamate uncaging We imaged uncaging-evoked [Ca 2+ ] transients in neurons loaded with a green Ca 2+ - sensitive indicator (G;
doi: 1.138/nature6416 Supplementary Notes Spine Ca 2+ signals produced by glutamate uncaging We imaged uncaging-evoked [Ca 2+ ] transients in neurons loaded with a green Ca 2+ - sensitive indicator (G;
Cellular Bioelectricity
 ELEC ENG 3BB3: Cellular Bioelectricity Notes for Lecture 24 Thursday, March 6, 2014 8. NEURAL ELECTROPHYSIOLOGY We will look at: Structure of the nervous system Sensory transducers and neurons Neural coding
ELEC ENG 3BB3: Cellular Bioelectricity Notes for Lecture 24 Thursday, March 6, 2014 8. NEURAL ELECTROPHYSIOLOGY We will look at: Structure of the nervous system Sensory transducers and neurons Neural coding
Efficient Emulation of Large-Scale Neuronal Networks
 Efficient Emulation of Large-Scale Neuronal Networks BENG/BGGN 260 Neurodynamics University of California, San Diego Week 6 BENG/BGGN 260 Neurodynamics (UCSD) Efficient Emulation of Large-Scale Neuronal
Efficient Emulation of Large-Scale Neuronal Networks BENG/BGGN 260 Neurodynamics University of California, San Diego Week 6 BENG/BGGN 260 Neurodynamics (UCSD) Efficient Emulation of Large-Scale Neuronal
Supplementary Information
 Supplementary Information D-Serine regulates cerebellar LTD and motor coordination through the 2 glutamate receptor Wataru Kakegawa, Yurika Miyoshi, Kenji Hamase, Shinji Matsuda, Keiko Matsuda, Kazuhisa
Supplementary Information D-Serine regulates cerebellar LTD and motor coordination through the 2 glutamate receptor Wataru Kakegawa, Yurika Miyoshi, Kenji Hamase, Shinji Matsuda, Keiko Matsuda, Kazuhisa
Lateralized hippocampal oscillations underlie distinct aspects of human spatial memory and navigation. Jacobs et al.
 Lateralized hippocampal oscillations underlie distinct aspects of human spatial memory and navigation Jacobs et al. Supplementary Information Lateralized hippocampal oscillations underlie distinct aspects
Lateralized hippocampal oscillations underlie distinct aspects of human spatial memory and navigation Jacobs et al. Supplementary Information Lateralized hippocampal oscillations underlie distinct aspects
Basal Ganglia Anatomy, Physiology, and Function. NS201c
 Basal Ganglia Anatomy, Physiology, and Function NS201c Human Basal Ganglia Anatomy Basal Ganglia Circuits: The Classical Model of Direct and Indirect Pathway Function Motor Cortex Premotor Cortex + Glutamate
Basal Ganglia Anatomy, Physiology, and Function NS201c Human Basal Ganglia Anatomy Basal Ganglia Circuits: The Classical Model of Direct and Indirect Pathway Function Motor Cortex Premotor Cortex + Glutamate
Research Paper. Christopher W. Butler, 1 Yvette M. Wilson, 1 Jenny M. Gunnersen, and Mark Murphy
 Research Paper Tracking the fear memory engram: discrete populations of neurons within amygdala, hypothalamus, and lateral septum are specifically activated by auditory fear conditioning Christopher W.
Research Paper Tracking the fear memory engram: discrete populations of neurons within amygdala, hypothalamus, and lateral septum are specifically activated by auditory fear conditioning Christopher W.
G5)H/C8-)72)78)2I-,8/52& ()*+,-./,-0))12-345)6/3/782 9:-8;<;4.= J-3/ J-3/ "#&' "#% "#"% "#%$
H/C8-)72)78)2I-,8/52& ()*+,-./,-0))12-345)6/3/782 9:-8;<;4.= J-3/ J-3/ #&' #% #% #%$") # G5)H/C8-)72)78)2I-,8/52& #% #$ # # &# G5)H/C8-)72)78)2I-,8/52' @5/AB/7CD J-3/ /,?8-6/2@5/AB/7CD #&' #% #$ # # '#E ()*+,-./,-0))12-345)6/3/782 9:-8;;4. @5/AB/7CD J-3/ #' /,?8-6/2@5/AB/7CD #&F #&' #% #$
# G5)H/C8-)72)78)2I-,8/52& #% #$ # # &# G5)H/C8-)72)78)2I-,8/52' @5/AB/7CD J-3/ /,?8-6/2@5/AB/7CD #&' #% #$ # # '#E ()*+,-./,-0))12-345)6/3/782 9:-8;;4. @5/AB/7CD J-3/ #' /,?8-6/2@5/AB/7CD #&F #&' #% #$
Behavioral Neuroscience: Fear thou not. Rony Paz
 Behavioral Neuroscience: Fear thou not Rony Paz Rony.paz@weizmann.ac.il Thoughts What is a reward? Learning is best motivated by threats to survival? Threats are much better reinforcers? Fear is a prime
Behavioral Neuroscience: Fear thou not Rony Paz Rony.paz@weizmann.ac.il Thoughts What is a reward? Learning is best motivated by threats to survival? Threats are much better reinforcers? Fear is a prime
Beyond Vanilla LTP. Spike-timing-dependent-plasticity or STDP
 Beyond Vanilla LTP Spike-timing-dependent-plasticity or STDP Hebbian learning rule asn W MN,aSN MN Δw ij = μ x j (v i - φ) learning threshold under which LTD can occur Stimulation electrode Recording electrode
Beyond Vanilla LTP Spike-timing-dependent-plasticity or STDP Hebbian learning rule asn W MN,aSN MN Δw ij = μ x j (v i - φ) learning threshold under which LTD can occur Stimulation electrode Recording electrode