University of Warwick institutional repository:
|
|
|
- Mariah Walker
- 6 years ago
- Views:
Transcription

1 University of Warwick institutional repository: This paper is made available online in accordance with publisher policies. Please scroll down to view the document itself. Please refer to the repository record for this item and our policy information available from the repository home page for further information. To see the final version of this paper please visit the publisher s website. Access to the published version may require a subscription. Author(s): Ulyana Lalo, Oleg Palygin, Richard Alan North, Alexei Verkhratsky, Yuriy Pankratov Article Title: Age-dependent remodelling of ionotropic signalling in cortical astroglia Year of publication: 2011 Link to published article: Publisher statement: The definitive version is available at wileyonlinelibrary.com
2 Age-dependent remodelling of ionotropic signalling in cortical astroglia Ulyana Lalo 1, Oleg Palygin 2, R. Alan North 3, Alexei Verkhratsky 3,4 & Yuriy Pankratov 2 1 Cell Physiology and Pharmacology, University of Leicester, Leicester LE1 9HN, UK, 2 Department of Biological Sciences, University of Warwick, Coventry, CV4 7AL, UK, 3 Faculty of Life Sciences, The University of Manchester, Manchester M13 9PT, UK and 4 Institute of Experimental Medicine, ASCR, Videnska 1083, Prague 4, Czech Republic Abbreviated title: Synaptic currents in aging astrocytes Address for correspondence: Prof. A. Verkhratsky, Faculty of Life Sciences, The University of Manchester, Smith Building, Oxford Road, Manchester M13 9PT UK Tel: ( ) Alexej.Verkhratsky@manchester.ac.uk OR Dr. Yuriy Pankratov Department of Biological Sciences, University of Warwick, Coventry, CV4 7AL, UK tel: ( ) Y.Pankratov@warwick.ac.uk Keywords: astrocytes; ageing; synaptic transmission; AMPA receptors; NMDA receptors; P2X receptors
3 2 Abstract Cortical astrocytes express fast ionotropic receptors for glutamate and ATP, although their role in neurone-glia communication remains controversial. Stimulation of neuronal afferents in mice neocortex triggers complex glial synaptic currents (GSCs) mediated by NMDA, P2X and AMPA receptors and glutamate transporters. In addition, astrocytes demonstrate spontaneous "miniature" GSCs resulting from quantal release of neurotransmitters. Here we demonstrate that maturation and ageing of the brain of mice (from 1 to 21 months) affects the density of ionotropic receptors in astrocytes and their role in GSCs generation. The AMPA receptor-mediated component is the largest in young animals and progressively declines with age. The P2X and NMDA components of GSC are smallest in young, maximal in adult (3 and 6 months old) and once more decrease in old mice, probably reflecting the remodelling of neuronal-glial circuitry. Our results demonstrate that fast synaptic transmission between neurones and astrocytes in neocortex that may be involved in information processing in neuronal-glial networks undergoes remodelling during brain maturation and ageing.
4 3 Introduction The development and ageing of the brain is associated with remodelling of synaptic contacts and plastic changes in neural networks. The formation, maintenance and modification of these networks depend on the interaction between neurones and neuroglia. It is now universally acknowledged that glial cells express an extensive complement of neurotransmitter receptors that can be activated during synaptic transmission in the CNS (Verkhratsky et al. 1998; Volterra & Meldolesi 2005; Fellin et al. 2006; Haydon & Carmignoto 2006). According to the "tripartite architecture of CNS synapses (Araque et al. 1999; Bezzi et al. 2001; Haydon 2001; Halassa et al. 2007) parts of astroglial membranes are located in close vicinity to neurotransmitter release sites. Ionotropic receptors located in such areas may therefore mediate evoked and spontaneous currents similar to those observed in neurone (Bergles et al. 2000; Lalo et al. 2006). Membrane physiology of maturing and ageing astrocytes is virtually unexplored. All information about active neuronal-glial connections has been derived from the experiments on newborn or young animals, whereas the age-dependent modification of these connections remains unknown. Our previous analysis of cortical astrocytes revealed functional expression of NMDA receptors as well as AMPA and P2X 1/5 receptors; unusually the NMDA receptors showed no Mg 2+ block (Lalo et al. 2006; Lalo et al. 2008). These ionotropic receptors can be activated by release of glutamate and ATP from cortical synaptic terminals (Pankratov et al. 2007) at physiological membrane potentials. Here we report in depth analysis of the age-dependent changes in synaptically activated responses in astroglia which we performed trying to understand how maturation of the brain affects signalling in neuronal-glial circuitry. Materials and Methods Neocortical slice preparation Experiments were performed on somato-sensory cortex isolated from the brains of transgenic mice expressing enhanced green fluorescent protein (EGFP) under the control of the human glial fibrillary acidic protein (GFAP) promoter (line TgN(GFAP-EGFP)GFEC-FKi; see (Nolte et al. 2001; Matthias et al. 2003; Grass et al. 2004; Lalo et al. 2008)). Mice (1-21 months old) were anaesthetized by halothane and then decapitated, in accordance with UK legislation. Slices were prepared using the technique described previously (Lalo et al. 2006). Brains were rapidly dissected and placed in physiological saline containing (in mm): 135 NaCl, 3 KCl, 1 MgCl 2, 2.4 CaCl 2, 26 NaHCO 3, 1 NaH 2 PO 4, 15 glucose, ph 7.4 when gassed with 95% O 2 / 5%CO 2. Cortical slices ( µm thick) were cut at 4 o C and kept at room temperature for 1-2 h prior the cell isolation. Acute isolation of astrocytes Astrocytes were acutely isolated using the modified vibrating ball technique (Pankratov et al. 2002; Lalo et al. 2006). The glass ball (200 µm diameter) was moved slowly some µm above the slice surface, while vibrating at 100 Hz (lateral displacements µm). The composition of external solution for isolated cell experiments was (mm): 135 NaCl; 2.7 KCl; 2.5 CaCl 2 ; 1 MgCl 2 ; 10 HEPES, 1 NaH 2 PO 4, 15 glucose, ph adjusted with NaOH to 7.3. Identification of astrocytes Astrocytes were identified by EGFP fluorescence. For this purpose cells were illuminated at 488 nm and observed at 520 ± 20 nm. Visually identified cells demonstrated electrophysiological signature characteristic of astrocytes (e.g. low input resistance, lack of fast time-dependent conductances, Fig. 1).
5 4 Electrophysiological recordings Whole-cell voltage clamp recordings from isolated astrocytes and astrocytes in situ were made with patch pipettes (5-6 M ) filled with intracellular solution (in mm): 110 KCl, 10 NaCl, 10 HEPES, 5 MgATP, 0.2 EGTA, ph The membrane potential was clamped at -80 mv unless stated otherwise. Currents were monitored using an AxoPatch200B patch-clamp amplifier (Axon Instruments, USA) filtered at 2 khz and digitized at 4 khz. Experiments were controlled by PCI data acquisition board (National Instruments, USA) and WinFluor software (Strathclyde Electrophysiology Software, UK); data were analyzed by self-designed software. Liquid junction potentials were measured with the patch-clamp amplifier; all voltages reported were corrected accordingly. Recordings commenced 10 min after whole-cell access was gained, to ensure equilibration between the pipette solution and the cytosol. Series resistances were 5-12 M and input resistances were MΩ; both varied by less than 20% in the cells accepted for analysis. A modified square-pulse concentration jump method (Lalo et al. 2001; Pankratov et al. 2002) was used for rapid (exchange time ~ 5 ms) applications of solutions containing various agents to single cells. Antagonists of ATP and glutamate receptors (PPADS, D-AP5 and CNQX) were pre-applied for 2 min before application of agonists. In situ recordings Whole cell voltage-clamp recordings were made from identified astrocytes situated in the layer II of somato-sensory cortex. All recordings shown were made at temperature of o C in the presence of 100 µm picrotoxin, membrane potential was kept at -80 mv unless specifically indicated. Axons originating from layer IV-VI neurones were stimulated at Hz with a bipolar coaxial electrode (WPI, USA) placed in the layer IV approximately opposite the site of recording; stimulus duration was 300 µs. The stimulus magnitude was set 3-4 times higher than minimal stimulus adjusted to activate the single-axon response in the layer II pyramidal neurons as described in (Pankratov et al. 2007). At such stimulus magnitude (further referred as moderate, typically μa), neuronal EPSCs were 3-5 times higher than those evoked by minimal stimulation, but still exhibited large amplitude fluctuations with failures in 10-15% of trials. Thus, one could expect that just a few intra-cortical axons and limited number of synapses were activated at chosen stimulus intensity. The currents evoked in the layer II astrocytes by moderate stimulus exhibited large amplitude fluctuations, their average amplitude reached 15-30% of maximal current evoked by saturating stimulus (30-40 μa); no noticeable astroglial response was recorded at stimulus intensity below minimal (i.e. threshold intensity for neuronal EPSCs). Fluorescent Ca 2+ -imaging To monitor the cytoplasmic free Ca 2+ concentraton ([Ca 2+ ] i ), cortical astrocytes were loaded with 100 µm Fura-2 (Ca 2+ probe equimolarly substituting EGTA in the intracellular saline) through the patch-pipette in whole-cell configuration. For fura-2 excitation, cells were alternately illuminated at wavelengths 340 ± 5 nm and 380 ± 5 nm using OptoScan monochromator (Cairn, Faversham, UK). Fluorescent images were recorded using Olympus BX51 microscope equipped with UMPLFL20x/NA0.95 objective and 2x intermediate magnification and Andor ixon885 EMCCD camera; exposure time was 35 ms at 2 x 2 binning. Astrocyte whole-cell currents were recorded simultaneously; experiments were controlled by WinFluor software (Strathclyde Electrophysiology Software, Glasgow, UK). The [Ca 2+ ] i levels were expressed as F 340 /F 380 ratio after background subtraction. To quantify the average amplitude of Ca 2+ -transients in different age groups, the F 340 /F 380 ratios were averaged over the whole cell image over two 1s-long time windows, in the rest (immediately before stimulus) and at the peak of transient; after then the resting F 340 /F 380 value was subtracted from the peak value.
6 5 Pharmacological isolation of different components of glial currents To functionally characterise the components of glial currents activated by synaptic stimulation, cortical slices were treated with the following pharmacological agents: (i) a combination of the antagonists of glutamate transporters (3S)-3-[[3-[[4-(Trifluoromethyl)benzoyl]amino]phenyl]me thoxy]-l-aspartic acid (TFB-TBOA), 1 μm and DL-threo-b-Benzyloxyaspartic acid (DL- TBOA), 30 µm (this mixture, referred to as TBOA-mix, blocks excitatory amino-acid transporters of EAAT1-5 types, some of which (EAAT1/2) are expressed in astroglia (Danbolt 2001; Hu et al. 2003)); (ii) a selective antagonist of AMPA receptors, 6-Cyano-7- nitroquinoxaline-2,3-dione (CNQX), 50 µm; (iii) a selective antagonist of NMDA receptors, D-(- )-2-Amino-5-phosphonopentanoic acid (D-AP5), 30 µm and (iv) the P2X receptor antagonist Pyridoxalphosphate-6-azophenyl-2',4'-disulfonic acid tetrasodium salt (PPADS), 10 µm. In some experiments we also used a selective P2X 1/5 inhibitor NF449 at 10 nm concentration (Rettinger et al. 2005). According to our previous reports (Lalo et al. 2006; Lalo et al. 2008) these concentrations of antagonists effectively inhibited P2X and glutamate receptors expressed in the cortical astrocytes. Data analysis All data are presented as mean SD, the statistical significance of difference between age groups was tested by one-way ANOVA test, unless indicated otherwise. The synaptically-evoked and spontaneous astroglial transmembrane currents were analysed off-line using methods described in (Pankratov & Krishtal 2003; Pankratov et al. 2007). Briefly, preliminary evaluation of the amplitude bi-phasic glial synaptic currents (GSCs) was made by averaging the amplitude of GSC in 1-msec time window at peak (I peak ) and 200 ms after peak (I 200 ). After that, each GSC was fitted with two model curves (e.g. fast and slow components), each model curve had monoexponential rise and decay phases. The decay time constant of fast component was varying in the range of ms, while decay time constant of slow component varied between 400 ms and 2.5 s; such difference in the kinetics ensured unambiguous separation of components. The minimal square root procedure was used to determine the amplitude of the model curves, while the time constants and offset were optimised by the gradient method to minimize the mean square error. For the initial detection of spontaneous events, the inward transmembrane currents of amplitude higher than 2 SD of baseline noise were selected. After then every single spontaneous event was analyzed within the 140 ms time window and its amplitude and kinetics were determined by fitting the model curve with single exponential rise and decay phases. As a rule, mean square error of fit amounted to 5-15% of peak amplitude for the GSCs and 5-20% for the spontaneous currents, depending on the background noise. Whenever error of fit exceeded 25%, spontaneous currents were discarded from further analysis. The amplitude distributions of spontaneous and evoked currents were analyzed with the aid of probability density functions and likelihood maximization techniques, as described in (Stricker et al. 1996; Pankratov & Krishtal 2003); all histograms shown were calculated as density functions. Drugs Receptor antagonists were from Tocris (Bristol, UK). Other salts and chemicals were from Sigma (Dorset, UK) unless specifically indicated. Results Membrane properties of astrocytes in different age groups We performed voltage-clamp recordings from protoplasmic astrocytes in layer II of somatosensory cortex of EFGP/GFAP mice (Nolte et al. 2001) of six different age groups of: 1, 3, 6, 9,
7 12 and months of age. The astrocytes in acutely isolated slices were identified by green fluorescence and by their characteristic electrophysiological signature (Fig. 1A). Ageing did not affect resting potential of astroglial cells (V mem values were ± 1.6 mv, ± 2.4 mv, ± 2.9, ± 1.7 mv, n = 7 for 1,3,6 and 9 months old animals respectively and ± 2.1 and 80.9 ± 2.6 mv, n = 5 for 12 and months old groups). Similarly we have not observed significant changes in membrane current pattern in astrocytes of different ages. Instead we detected a small (~ 20-30%) yet statistically significant decrease in the input resistance of astrocytes in the 3 and 6-months old mice (Fig. 1B). Age-dependent changes in evoked synaptic currents We confirmed our previous observations (Lalo et al. 2006) that stimulation of neuronal afferents originating from layers IV-VI induced complex ion currents in astrocytes (Fig. 2, 3). These currents were directly associated with synaptic transmission because (i) treatment of slices with 1 M TTX completely abolished glial responses (Fig. 2A), (ii) the amplitude of the glial response was stimulus-dependent in a manner similar to the synaptic current evoked in the neighbouring neurones (Fig 2A) and (iii) glial postsynaptic currents could be recorded for a long time at frequencies 0.5 Hz, this demonstrating frequency-dependence similar to neuronal EPSCs (Fig 2B). Therefore, we identified astroglial responses as glial synaptic currents (GSCs); the GSCs were recorded in all age groups. The maximal amplitude of astrocytic response to the synaptic stimulation ranged between 20 and 200 pa depending on the age (see below), the amplitude of GSC evoked by moderate stimulus varied typically between 5 and 60 pa. The GSCs recorded from the astrocytes of old mice (12 and months old) were markedly depressed in the course of repetitive stimulation, whereas amplitude of GSCs in younger animals (1-6 months) was stable at high (1 Hz) stimulation frequency (Fig 2B). The GSCs recorded in astrocytes had complex kinetics, with fast and slow components. Fast component of GSCs had a decay time fast = 27.2 ± 8.8 ms whereas the slow component of GSCs decayed with slow = 1.4 ± 0.8 s (n = 38). The antagonists, applied in appropriate concentrations to block glutamate transporters (TBOA-mix) and astroglial ionotropic receptors (CNQX, D-AP5 and PPADS), inhibited both peak (I peak ) and delayed (I 200ms ) components of GSC (Fig. 3). Effects of antagonists were reversible and the GSC amplitude gradually recovered after washout. This pharmacological profile confirmed our previous findings of the role for AMPA and NMDA glutamate receptors in generation of astroglial synaptic currents (Lalo et al. 2006), and demonstrated the involvement of P2X receptors (Lalo et al. 2008) in shaping synaptically activated astroglial currents. The glial synaptic responses demonstrated age-dependent remodelling. First, the maximal amplitude of synaptic response increased significantly after 1 months attaining highest levels ( pa) at mature adult (3 and 6 months) animals; it declined again at more advanced (9-21 months) age to the lowest level of pa measured in old (18-21 months) group (Fig. 3C). Second, ageing affected the relative weight of fast and slow components of GSCs (Fig. 3C). The relative contribution of the fast component was the lowest at 1 month of age, became predominant in young adults (3 and 6 months), and declined at advanced age (9-21 months). Third, the changes in GSCs kinetics reflected age-dependent changes in their pharmacological profiles, thus indicating changes in the underlying molecular machinery (Fig. 3D). The contribution of P2X receptors to GSCs was higher in younger ages, reaching maximal values of 56 ± 6% (fast component) and 39 ± 4% (slow component) at 3 months (n = 9). In contrast, the contribution of NMDA receptors grew steadily with age, being maximal at 9-21 months for the fast component (53 ± 6%, n = 10) and at 6 months for the slow component (53 ± 7%, n = 9); at the older ages contribution of NMDA component slightly declined. AMPA receptors contributed 6
8 exclusively to the fast component of GSCs, their contribution was maximal at 1 month (18 ± 4%, n = 10) and sharply declined at older ages. The contribution of glutamate transporters to GSCs was minimal in 3 months, but increased again at 9-21 months. Age-dependent changes in spontaneous synaptic currents Similarly to our previous reports (Lalo et al. 2006) we observed spontaneous "miniature" glial synaptic currents (mgscs) in the presence of Na + -channel blocker tetrodotoxin (1 M) and GABA receptor antagonist picrotoxin (100 M) at holding potential -80 mv in astrocytes of all age groups (Fig. 4). Analysis of the spontaneous events revealed two distinct groups of mgscs with different decay kinetics. Fast mgscs had an amplitude in the range of 5-10 pa and a decay time of 10.8 ± 2.9 ms; slow mgscs amplitudes varied between 5 and 25 pa and a decay time was 28.5 ± 11.2 ms (n = 46). Depending on the age of animals, the amplitude ratio of fast to slow mgscs was in the range of , the relative frequency of fast mgscs was 40-60% of the total. The mgscs were insensitive to TBOA-mix but were differentially inhibited by antagonists of glutamate and P2X receptors. Inhibition of the AMPA receptors with 50 µm CNQX did not affect the fast mgscs (Fig. 4) although it somewhat reduced the amplitude of slow mgscs by 23 ± 11% (n = 41, all ages) without affecting their frequency. In the presence of CNQX the time of decay of slow mgscs was slightly increased (13 ± 9%, n = 41). Subsequent (i.e. in the presence of CNQX) inhibition of NMDA receptors by D-AP5 eliminated slow currents in all cells leaving the fast mgscs intact (Fig. 3A, B). In contrast to glutamate receptor antagonists, application of PPADS did not affect the slow mgscs but significantly inhibited fast currents (Fig. 4A, B). On average 10 µm PPADS decreased the amplitude and frequency of fast mgscs by 67 ± 12 and 82 ± 15% (n = 37), respectively. Therefore cortical astrocytes demonstrate two distinct populations of spontaneous currents: fast mgscs mediated by P2X receptors and slow glutamatergic mgscs mediated mainly by NMDA receptors with minor contribution of AMPA receptors. The unitary size of purinergic and glutamatergic mgscs was evaluated through the corresponding peaks in the amplitude histograms. The unitary size of P2X receptor-mediated mgscs varied in an age-dependent manner between 2.8 and 7.5 pa, whereas the unitary size of NMDA receptor-mediated mgscs varied between 3.4 and 10.3 pa (Fig. 4D). Importantly, unitary size of purinergic and glutamatergic mgscs correlated with quantal size of corresponding fraction of fast evoked GSCs at all ages, as discussed below. The differences in the properties of the fast and slow components of GSCs may indicate that they are generated at the different parts of astroglial membrane. The fast component mediated mostly by ionotropic receptors (including AMPA receptors (Lalo et al. 2006)) with low affinity to glutamate) could be generated in areas exposed to the rapid transients of high concentrations of glutamate. These areas of astroglial membranes are located close to the release sites and may even protrude into the synaptic cleft (Verkhratsky & Kirchhoff 2007). In contrast, the slow GSC component has a significant contribution of glutamate transporter-mediated current and therefore may originate from areas exposed to small but long-lasting elevations in concentration of neurotransmitters spilled out of the synaptic cleft. The contribution of both P2X and NMDA receptors to the slow component may be attributed to the high affinity of these receptors to ATP and glutamate and their slow desensitization (Lalo et al. 2006; Lalo et al. 2008). The age-dependent remodelling of astroglial synaptic responses may reflect changes in the release of neurotransmitters and/or changes in the density/efficacy of astroglial receptors. The 7
9 underlying mechanisms can be determined by analysis of the quantal size of GSCs (directly related to the activity of postsynaptic receptors) and by analysis of the mean quantal content (which is indicative for changes in the neurotransmitter release). At the stimulation strength used, both the fast component of evoked GSCs and neuronal synaptic response exhibited similar large fluctuations, which enabled evaluation of the quantal parameters of pharmacologically isolated purinergic and glutamatergic GSCs (Fig. 4C) using quantal basic analysis techniques. The quantal content of evoked purinergic GSCs increased between1 and 3 months and then steadily declined. The quantal content of glutamatergic GSCs was much smaller in young age but exhibited considerable and sustained increase towards the mid-life and then a sharp decrease in the old age (Fig 4C). Quantal size of both P2X and NMDA receptor-mediated GSCs had a similar behaviour with significant increase at 3 and 6 months old animals, which followed by substantial decrease at older ages. Quantal size of the AMPA receptor-mediated GSCs was maximal at 1 month and was steadily decreasing at later ages. These results suggest that the strength of fast ionotropic signalling in cortical astrocytes reaches its peak at the early adulthood and then begins to decline. This was further corroborated by analysis of spontaneous GSCs (Fig. 4D). The frequency of mgscs was higher at younger ages reaching maximum at 3 months and then steadily declined. The unitary size of P2X receptormediated mgscs was increasing until 6 months and decreased significantly afterwards. The unitary size of NMDA receptor-mediated mgscs was maximal at 6 months and decreased at older age. The unitary size of the AMPA receptor-mediated sgscs was maximal in very young animals and declined significantly with ageing. Age-dependent changes in agonist-evoked currents in acutely isolated astrocytes The data described above suggest age-dependent changes in the expression of ionotropic receptors in cortical astrocytes. This was directly confirmed in voltage-clamp experiments on astrocytes acutely isolated from the cortices of animals of different ages. In isolated cells glutamate (100 M) typically triggered biphasic current responses (Fig. 5A). Application of the NMDA receptor antagonist D-AP5 (10 M) selectively reduced the steady-state component but did not affect the fast portion of the response. This latter was selectively inhibited by CNQX (30 µm). The residual sustained component of the glutamate-induced current was completely blocked by the TBOA-mix in all cells tested. Applications of NMDA and ATP (Fig. 5A) triggered current responses which were completely blocked by D-AP5 (10 M) and PPADS (100 M) respectively. Pharmacological properties of the agonist-evoked currents did not change with age and were consistent with properties of astroglial receptors previously described in young animals (Lalo et al. 2006; Lalo et al. 2008). Instead, ageing significantly affected the amplitudes of responses to ATP, NMDA, AMPA and glutamate. We observed a 3-fold and 6-fold increase in the density of P2X and NMDA receptor-mediated currents in isolated astrocytes in the adult age (Fig. 5). Both P2X and NMDA receptor-mediated currents were maximal at 6 months, which was followed by considerable decline at older ages. The density of AMPA-receptor mediated currents was maximal at 1 month and gradually declined with age. These data agree with age-related changes of the astrocytic GSCs measured in situ. The density of glutamate-transporter mediated current (recorded in the presence of CNQX and D-APV) also increased at 3 and even more at 6 months contrasting to the decrease in the transporter-mediated component of slow GSCs in situ (Fig. 5B). This may be explained by much lower density of transporter-mediated currents compared to NMDA- and P2X-currents at all ages (Fig. 5B); hence the relative contribution of transporters to GSCs was smaller at the advanced ages. 8
10 9 Age-dependent changes in ionotropic receptor-mediated astroglial Ca 2+ signals We recorded calcium signals in protoplasmic cortical astrocytes triggered by synaptic stimulation of ionotropic receptors (Fig. 6A). Slices incubation with selective AMPA receptor blocker CNQX (30 µm) did not affect Ca 2+ transients (n = 8, data not shown). In contrast NMDA receptor blocker D-AP5 (30 M) and selective inhibitor of P2X 1/5 receptors NF449 (10 nm) partially inhibited Ca 2+ responses in all 14 astrocytes tested. The relative contribution of NMDA and P2X receptors into shaping Ca 2+ signals changes with age (Fig. 6B), following the general pattern of changes in the GCS and agonist activated-currents. The amplitudes of ionotropicreceptors mediated Ca 2+ signals were maximal at 3 and 6 months and decreased significantly at advanced ages (Fig. 6B). Discussion In this report we demonstrated the age-dependent modification of synaptic transmission onto protoplasmic astrocytes in the neocortical structures. Neuronal activity evokes glial currents generally similar to postsynaptic currents in neurones thus indicating that the astroglial membrane acts as a postsynaptic compartment. Astrocytes receive fast quantal signals, which most likely originate from vesicular release of glutamate and ATP from the presynaptic terminals. In addition to fast currents, astrocytes also possess slow synaptically activated currents mediated by glutamate transporters. Fast GSCs were apparently evoked in areas of astroglial membrane exposed to high concentrations of glutamate since part of this component was mediated by astrocytic AMPA receptors with relatively low glutamate sensitivity (Lalo et al. 2006). These astroglial compartments may surround or even protrude into the synaptic cleft (Volterra & Meldolesi 2005; Fiacco et al. 2009) or may be a direct target of axon collaterals, like synapses on NG-2 glial cells (Paukert & Bergles 2006). Slow GSCs are generated in the areas exposed to small but long-lasting elevations in concentration of neurotransmitters (probably due to the spillover of the neurotransmitter from the synaptic cleft) as suggested by the lack of CNQX sensitivity and significant contribution of high-affinity NMDA receptors and glutamate transporters. The role of ectopic neurotransmitter release in inducing GSC in the cortex remains doubtful. Indeed in our previous experiments in the young mice (Lalo et al. 2006) we found that cyclothiazide (CTZ; a compound that increases the apparent affinity of AMPA receptors (Yamada & Tang 1993; Partin et al. 1994)) increases AMPA-mediated component of glial synaptically activated current by 96%, which is in a good agreement with effect of CTZ on neuronal synaptic currents (Yamada & Tang 1993). Such an increase in astroglial currents also closely resembles a twofold increase in the amplitude of response of the climbing fibre synapses on NG-2 glial cells in cerebellum (Lin et al. 2005). In contrast, currents activated in the Bergmann glia following ectopic release exhibited more than eightfold increase under CTZ due to exposure of glial AMPA receptors to low concentrations of glutamate (Dzubay & Jahr 1999). Additional similarity of cortical GSCs in young mice to synaptically-activated currents in the NG-2 cells (Paukert & Bergles 2006) is their sustainability at repetitive stimulation (we could record GSCs for more min at Hz see Fig 2B) whereas glial currents activated by ectopic release fade rapidly due to pre-synaptic depression and fast depletion of ectopic transmitter pools (Matsui & Jahr 2004; Paukert & Bergles 2006). The GSCs recorded in old animals are much more susceptible to the depression at high stimulation frequency, which may be explained by re-modelling of the neuron-driven input onto astrocytes that may include an increased contribution of ectopic release. Alternatively, the synaptic fatigue may develop faster in old animals leading to the use-dependent decrease in neurotransmitter release.
11 10 Glial synaptic responses were modified through the maturation and ageing of the brain. The agerelated changes in the GSCs amplitude cannot be attributed to changes in the astrocytes input resistance. First, increase in GCS amplitudes recorded in 3- and 6- months was accompanied by decrease in the input resistance (which would decrease the current measured at soma). Second, changes in the input resistance in astrocytes in 3 and 6 months old animals were rather minor. Using the cable theory of whole-cell voltage-clamp recording in situ (Major et al. 1993) it is possible to estimate that 30% change in the input resistance may account for only 3-4% change in the amplitude of current measured at soma in our experimental conditions (the series/input resistance ratio of 8-15). In young animals the synaptic glial responses were relatively small; in the course of CNS maturation they increased several times, mostly because of the increase in the density of NMDA and P2X receptors (Fig. 5). The maximal strength of glial synaptic responses was attained at 6 month. Further increase in age resulted in the decrease in the density of ionotropic receptors and overall diminution of astroglial synaptic currents. Importantly, the considerable decrease in the amplitude of the synaptically-driven currents in old age (Fig. 3C) originates both from decrease in the density of ionotropic receptors, as indicated by the diminution of the quantal size of evoked and spontaneous GSCs, and decline of the release of glutamate and ATP from neurones onto glial cells, manifested by substantial decrease in the quantal content of evoked GSCs and the frequency of spontaneous currents (Fig. 4C,D). Glutamate transporters do not seem to play a major role in astroglial ionotropic signalling, especially at 3 and 6 months, most likely due their lower efficiency in the electric charge transfer per single glutamate molecule bound. The major role for transporters seems to be clearing glutamate from extracellular space and increase in their density at 3 and 6 months most probably reflects the homeostatic response to the increased release of glutamate (as could be postulated based on the analysis of quantal content). We also found that NMDA and P2X receptors provide significant Ca 2+ influx in the astrocytes which is subjected to age-dependent remodelling. Naturally our results offer a wide and open field for speculations, indicating for example an agedependent decrease in synaptic communication in neuronal-glial circuitry that may in turn signify the beginning of age-dependent decline in brain functions; alternatively however, the bell-shaped changes in glial synaptic transmission may reflect maturation of the CNS, pruning of functionally irrelevant synaptic contacts and manifest the attainment of wisdom which generally accompanies advanced age. Acknowledgements Authors research was supported by BBSRC (BB/F to YP) and by Grant Agency of the Czech Republic (GACR 305/08/1381 and GACR 305/08/1384 to AV).
12 References Araque A, Parpura V, Sanzgiri RP, Haydon PG (1999). Tripartite synapses: glia, the unacknowledged partner. Trends Neurosci. 22, Bergles DE, Roberts JD, Somogyi P, Jahr CE (2000). Glutamatergic synapses on oligodendrocyte precursor cells in the hippocampus. Nature. 405, Bezzi P, Domercq M, Vesce S, Volterra A (2001). Neuron-astrocyte cross-talk during synaptic transmission: physiological and neuropathological implications. Prog Brain Res. 132, Danbolt NC (2001). Glutamate uptake. Progr. Neurobiol. 65, Dzubay JA, Jahr CE (1999). The concentration of synaptically released glutamate outside of the climbing fiber- Purkinje cell synaptic cleft. J Neurosci. 19, Fellin T, Pozzan T, Carmignoto G (2006). Purinergic receptors mediate two distinct glutamate release pathways in hippocampal astrocytes. J Biol Chem. 281, Fiacco TA, Agulhon C, McCarthy KD (2009). Sorting out astrocyte physiology from pharmacology. Annu Rev Pharmacol Toxicol. 49, Grass D, Pawlowski PG, Hirrlinger J, Papadopoulos N, Richter DW, Kirchhoff F, Hulsmann S (2004). Diversity of functional astroglial properties in the respiratory network. J Neurosci. 24, Halassa MM, Fellin T, Haydon PG (2007). The tripartite synapse: roles for gliotransmission in health and disease. Trends Mol Med. 13, Haydon PG (2001). GLIA: listening and talking to the synapse. Nat Rev Neurosci. 2, Haydon PG, Carmignoto G (2006). Astrocyte control of synaptic transmission and neurovascular coupling. Physiol Rev. 86, Hu WH, Walters WM, Xia XM, Karmally SA, Bethea JR (2003). Neuronal glutamate transporter EAAT4 is expressed in astrocytes. Glia. 44, Lalo U, Pankratov Y, Kirchhoff F, North RA, Verkhratsky A (2006). NMDA receptors mediate neuron-to-glia signaling in mouse cortical astrocytes. J Neurosci. 26, Lalo U, Pankratov Y, Wichert SP, Rossner MJ, North RA, Kirchhoff F, Verkhratsky A (2008). P2X 1 and P2X 5 subunits form the functional P2X receptor in mouse cortical astrocytes. J Neurosci. 28, Lalo UV, Pankratov YV, Arndts D, Krishtal OA (2001). Omega-conotoxin GVIA potently inhibits the currents mediated by P2X receptors in rat DRG neurons. Brain Res Bull. 54, Lin SC, Huck JH, Roberts JD, Macklin WB, Somogyi P, Bergles DE (2005). Climbing fiber innervation of NG2- expressing glia in the mammalian cerebellum. Neuron. 46, Major G, Evans JD, Jack JJ (1993). Solutions for transients in arbitrarily branching cables: II. Voltage clamp theory. Biophys J. 65, Matsui K, Jahr CE (2004). Differential control of synaptic and ectopic vesicular release of glutamate. J Neurosci. 24, Matthias K, Kirchhoff F, Seifert G, Huttmann K, Matyash M, Kettenmann H, Steinhauser C (2003). Segregated expression of AMPA-type glutamate receptors and glutamate transporters defines distinct astrocyte populations in the mouse hippocampus. J Neurosci. 23, Nolte C, Matyash M, Pivneva T, Schipke CG, Ohlemeyer C, Hanisch UK, Kirchhoff F, Kettenmann H (2001). GFAP promoter-controlled EGFP-expressing transgenic mice: a tool to visualize astrocytes and astrogliosis in living brain tissue. Glia. 33, Pankratov Y, Lalo U, Krishtal O, Verkhratsky A (2002). Ionotropic P2X purinoreceptors mediate synaptic transmission in rat pyramidal neurones of layer II/III of somato-sensory cortex. J Physiol. 542, Pankratov Y, Lalo U, Verkhratsky A, North RA (2007). Quantal release of ATP in mouse cortex. J Gen Physiol. 129, Pankratov YV, Krishtal OA (2003). Distinct quantal features of AMPA and NMDA synaptic currents in hippocampal neurons: implication of glutamate spillover and receptor saturation. Biophys J. 85, Partin KM, Patneau DK, Mayer ML (1994). Cyclothiazide differentially modulates desensitization of alpha-amino- 3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor splice variants. Mol Pharmacol. 46, Paukert M, Bergles DE (2006). Synaptic communication between neurons and NG2+ cells. Curr Opin Neurobiol. 16, Rettinger J, Braun K, Hochmann H, Kassack MU, Ullmann H, Nickel P, Schmalzing G, Lambrecht G (2005). Profiling at recombinant homomeric and heteromeric rat P2X receptors identifies the suramin analogue NF449 as a highly potent P2X1 receptor antagonist. Neuropharmacology. 48, Stricker C, Field AC, Redman SJ (1996). Statistical analysis of amplitude fluctuations in EPSCs evoked in rat CA1 pyramidal neurones in vitro. J Physiol. 490 ( Pt 2), Verkhratsky A, Kirchhoff F (2007). NMDA Receptors in glia. Neuroscientist. 13, Verkhratsky A, Orkand RK, Kettenmann H (1998). Glial calcium: homeostasis and signaling function. Physiol Rev. 78, Volterra A, Meldolesi J (2005). Astrocytes, from brain glue to communication elements: the revolution continues. Nat Rev Neurosci. 6, Yamada KA, Tang CM (1993). Benzothiadiazides inhibit rapid glutamate receptor desensitization and enhance glutamatergic synaptic currents. J Neurosci. 13,
13 Figure legends 12 Figure 1. Identification and basic electrophysiological properties of cortical astrocytes in situ. A. Representative EGFP images (left column) of cortical layer II astrocytes of different age groups and corresponding whole-cell currents (right column) evoked to depolarizing steps from the holding potential of -80 mv; the voltage protocol is shown on the top. B. Age-related changes in the input resistance of cortical astrocytes. Data are presented as mean ± SD for 15 cells in the each age group; decrease in the astrocyte R in measured in 3 and 6 months in comparison to 1 month was statistically significant with P < 0.05 (one-way ANOVA). Figure 2. General functional properties of glial synaptic currents. A. Stimulus dependence and tetrodotoxin (TTX) sensitivity of glial synaptic currents. Left panel shown synaptic current recoded separately in astrocyte and neurone in the same cortical slice at different stimulation amplitudes (1,2,5,10,20,30 and 40 µa). The mean ± SD values of normalised amplitudes of astroglial/neuronal synaptic currents are shown on the graph (right panel); data are shown for 5 astrocytes and 7 neurons from 6-10 week-old mice. Incubation with 1 M of TTX completely abolished both neuronal and astroglial responses. B. Frequency-dependence of glial synaptic currents. The glial responses were reliably evoked for min at stimulation frequency of 0.5 Hz, increasing of stimulation frequency to 1.0 Hz causes only minor decrease in the GSC amplitude in the young (3 and 6 months old) animals but causes considerable decrease the GSCs recorded in the old (12-20 months) mice. Representative current traces are shown on the top. The bottom plot shows changes in the GSCs amplitude quantified for 5 astrocytes for each group; normalized amplitudes of glial currents (mean ± SD for 5 cells) are plotted versus recording time. Each point represents the average amplitude (relative to control) of 10 sequential currents. Figure 3. Synaptic currents in the cortical astrocytes. Transmembrane currents evoked in the astrocytes in neocortical layer II of 3 month old (A) and 9 monthold (B) mice in situ by moderate stimulation of neuronal afferents were recorded at membrane potential of -80 mv in the presence of 100 µm picrotoxin. The glial synaptically-activated currents (GSCs) were inhibited by consecutive application of mixture of 1 µm TFB-TBOA and 30 µm DL-TBOA, 50 µm CNQX, 30 µm D-AP5 and 10 µm PPADS. Amplitudes of GSC were monitored at peak (I peak, grey dots) and 200 after peak (I 200, red dots) when only slow component remained. Each point on the time graphs represents the mean ± SD for five GSCs; examples of GSCs traces and corresponding fast (blue line) and slow (red line) components are shown below. Note the difference in the shape and pharmacological sensitivity of GSC recorded in mice of different age. C. Age-dependent changes in peak amplitude of total synaptic current (black), of the fast (blue) and slow (red) components of GSC evoked by the maximal stimulation; amplitude of fast and slow components were determined by fitting two model curves as shown in (A,B) and described in the Methods. D. Pharmacological profile of fast and slow components of evoked GSCs in mice of different ages. The data, presented as mean ± SD for 6-10 cells for each age group, were calculated from the inhibitory effect of the appropriate agonist. Figure 4. Miniature spontaneous currents (mgscs) in cortical astrocytes are mediated by the P2X, AMPA and NMDA receptors. A. Representative whole-cell recordings from the cortical astrocyte of 6 months-old mouse in control and after consecutive application of 50 µm CNQX, 30 µm D-AP5 and 10 µm PPADS. The right panels represent averaged waveforms of mgscs (25 events). B. Corresponding histograms for amplitude (left column) and decay time (right column) distribution of mgscs. Fast mgscs are indicated by blue lines; slow mgscs are shown in red. Note the fast and slow
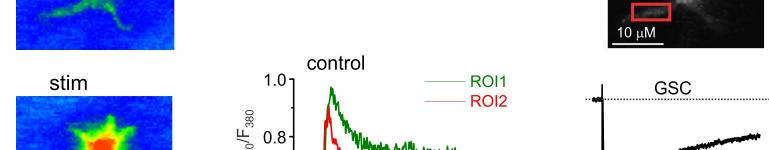
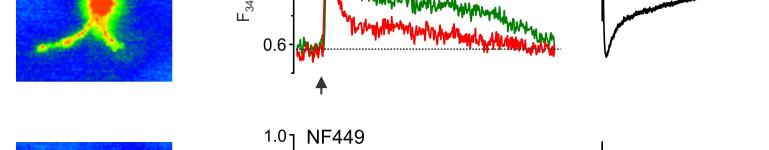
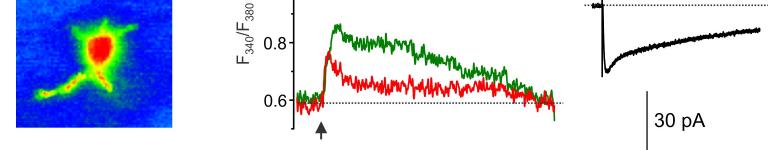
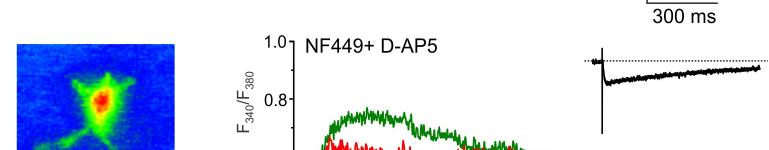
14 mgscs correspond to different peaks in the amplitude histograms. Antagonists of AMPA and NMDA receptors selectively inhibited slow currents while PPADS selectively inhibited fast currents. These results indicate activation of two separate populations of purinergic and glutamatergic mgscs. C. Quantal analysis shows the age-related changes in the mean quantal content and quantal size of pharmacologically isolated P2X, NMDA and AMPA receptor-mediated evoked GSCs. Data are presented for 6-8 cells for each age. Statistical significance of difference between values of quantal content at 1 months and quantal content at 3, 6, and 9 months P<0.01 (ANOVA); statistical significance of difference in the quantal size P<0.02. D. Age-related changes in the frequency and unitary size of spontaneous miniature synaptic currents in the cortical astrocytes. Data presented for 6-11 cells for each age. Note the consistent changes in the quantal size of evoked GSCs and unitary size of mgscs. Statistical significance of difference in the mgscs frequency at 1 month and all other ages P <0.02 for both purinergic and glutamatergic currents; statistical significance of difference in the quantal size at 1, 3 and 6 months P< Figure 5. Ageing affects the density of glutamate transporter and ionotropic receptor-mediated currents in cortical astrocytes. A. Representative whole-cell currents elicited in the acutely isolated astrocytes by application of 100 µm glutamate (left column), 10 µm NMDA (middle column) and 10 µm ATP/αβmeATP at holding potential of -80 mv. Glutamate- and NMDA-evoked currents were inhibited by 10µM D-AP5 and 30µM CNQX; ATP-evoked currents were inhibited by 10 µm PPADS. B. The density of currents mediated by P2X, NMDA and AMPA receptors and glutamate transporters in cortical astrocytes (mean ± SD for 9-12 cells for each age group); statistical significance of difference between average value for 1 month and corresponding values for 3 and 6 months P<0.02 (ANOVA) for all types of currents. Figure 6. Age-dependent changes in synaptically induced ionotropic Ca 2+ astrocytes. signals in protoplasmic A. Cortical layer II astrocyte of 9 months-old mouse was loaded with Fura-2 in situ via patch pipette. Fluorescent images were recorded simultaneously with glial currents evoked by neuronal afferent stimulation in presence of TBOA-mix and CNQX (control) and after consecutive application of 10 nm NF-449 (selective antagonist of P2X receptors) and 30 µm D-AP5. Representative images (pseudocolour, pipette image subtracted) and GSCs (right column) were recorded before (rest) and after stimulation as indicated. Ca 2+ -transients (middle column) are expressed as F 340 /F 380 ratio averaged over the corresponding regions of interest shown in the GFAP image of astrocyte (top right). B. Age-related changes in the astrocytic Ca2+-signalling Left panel, average peak amplitudes of [Ca 2+ ] i increases induced by stimulation of neuronal afferents in cortical astrocytes of different ages. Right panel, average inhibitory effect of antagonists of P2X (NF449) and NMDA (D-AP5) receptors on the amplitudes of [Ca 2+ ] i increases responses in cortical astrocytes. Data are presented as mean ± SD for 3-4 cells for each age group; * P < 0.05, ** P < 0.01 one-way ANOVA compared to 1 month.




15 Fig.1 14
16 15
17 16
18 17
19 18

20 19
Supporting Information
 ATP from synaptic terminals and astrocytes regulates NMDA receptors and synaptic plasticity through PSD- 95 multi- protein complex U.Lalo, O.Palygin, A.Verkhratsky, S.G.N. Grant and Y. Pankratov Supporting
ATP from synaptic terminals and astrocytes regulates NMDA receptors and synaptic plasticity through PSD- 95 multi- protein complex U.Lalo, O.Palygin, A.Verkhratsky, S.G.N. Grant and Y. Pankratov Supporting
Supporting Online Material for
 www.sciencemag.org/cgi/content/full/317/5841/183/dc1 Supporting Online Material for Astrocytes Potentiate Transmitter Release at Single Hippocampal Synapses Gertrudis Perea and Alfonso Araque* *To whom
www.sciencemag.org/cgi/content/full/317/5841/183/dc1 Supporting Online Material for Astrocytes Potentiate Transmitter Release at Single Hippocampal Synapses Gertrudis Perea and Alfonso Araque* *To whom
Is action potential threshold lowest in the axon?
 Supplementary information to: Is action potential threshold lowest in the axon? Maarten H. P. Kole & Greg J. Stuart Supplementary Fig. 1 Analysis of action potential (AP) threshold criteria. (a) Example
Supplementary information to: Is action potential threshold lowest in the axon? Maarten H. P. Kole & Greg J. Stuart Supplementary Fig. 1 Analysis of action potential (AP) threshold criteria. (a) Example
Neuronal-glial chemical transmission mediated by glutamate and ATP Prof. Alexei Verkhratsky
 MD, PhD, DSc Professor of Neurophysiology, The University of Manchester, Faculty of Life Sciences, Manchester, UK 1 Experiments on glial cells in tissue culture demonstrated expression of neurotransmitter
MD, PhD, DSc Professor of Neurophysiology, The University of Manchester, Faculty of Life Sciences, Manchester, UK 1 Experiments on glial cells in tissue culture demonstrated expression of neurotransmitter
Supplementary Information
 Hyperpolarization-activated cation channels inhibit EPSPs by interactions with M-type K + channels Meena S. George, L.F. Abbott, Steven A. Siegelbaum Supplementary Information Part 1: Supplementary Figures
Hyperpolarization-activated cation channels inhibit EPSPs by interactions with M-type K + channels Meena S. George, L.F. Abbott, Steven A. Siegelbaum Supplementary Information Part 1: Supplementary Figures
Neuronal-glial chemical transmission mediated by glutamate and ATP
 1 Experiments on glial cells in tissue culture demonstrated expression of neurotransmitter receptors 2 Neuronal-glial chemical transmission mediated by glutamate and TP lexei Verkhratsky The University
1 Experiments on glial cells in tissue culture demonstrated expression of neurotransmitter receptors 2 Neuronal-glial chemical transmission mediated by glutamate and TP lexei Verkhratsky The University
The mammalian cochlea possesses two classes of afferent neurons and two classes of efferent neurons.
 1 2 The mammalian cochlea possesses two classes of afferent neurons and two classes of efferent neurons. Type I afferents contact single inner hair cells to provide acoustic analysis as we know it. Type
1 2 The mammalian cochlea possesses two classes of afferent neurons and two classes of efferent neurons. Type I afferents contact single inner hair cells to provide acoustic analysis as we know it. Type
SUPPLEMENTARY INFORMATION. Supplementary Figure 1
 SUPPLEMENTARY INFORMATION Supplementary Figure 1 The supralinear events evoked in CA3 pyramidal cells fulfill the criteria for NMDA spikes, exhibiting a threshold, sensitivity to NMDAR blockade, and all-or-none
SUPPLEMENTARY INFORMATION Supplementary Figure 1 The supralinear events evoked in CA3 pyramidal cells fulfill the criteria for NMDA spikes, exhibiting a threshold, sensitivity to NMDAR blockade, and all-or-none
Supporting Online Material for
 www.sciencemag.org/cgi/content/full/312/5779/1533/dc1 Supporting Online Material for Long-Term Potentiation of Neuron-Glia Synapses Mediated by Ca 2+ - Permeable AMPA Receptors Woo-Ping Ge, Xiu-Juan Yang,
www.sciencemag.org/cgi/content/full/312/5779/1533/dc1 Supporting Online Material for Long-Term Potentiation of Neuron-Glia Synapses Mediated by Ca 2+ - Permeable AMPA Receptors Woo-Ping Ge, Xiu-Juan Yang,
SUPPLEMENTARY INFORMATION
 Supplementary Figure 1. Normal AMPAR-mediated fepsp input-output curve in CA3-Psen cdko mice. Input-output curves, which are plotted initial slopes of the evoked fepsp as function of the amplitude of the
Supplementary Figure 1. Normal AMPAR-mediated fepsp input-output curve in CA3-Psen cdko mice. Input-output curves, which are plotted initial slopes of the evoked fepsp as function of the amplitude of the
Neuroscience 201A (2016) - Problems in Synaptic Physiology
 - Problems in Synaptic Physiology") Question 1: The record below in A shows an EPSC recorded from a cerebellar granule cell following stimulation (at the gap in the record) of a mossy fiber input. These responses are, then, evoked by stimulation.
Question 1: The record below in A shows an EPSC recorded from a cerebellar granule cell following stimulation (at the gap in the record) of a mossy fiber input. These responses are, then, evoked by stimulation.
BIPN 140 Problem Set 6
 BIPN 140 Problem Set 6 1) The hippocampus is a cortical structure in the medial portion of the temporal lobe (medial temporal lobe in primates. a) What is the main function of the hippocampus? The hippocampus
BIPN 140 Problem Set 6 1) The hippocampus is a cortical structure in the medial portion of the temporal lobe (medial temporal lobe in primates. a) What is the main function of the hippocampus? The hippocampus
Neurons of the Bed Nucleus of the Stria Terminalis (BNST)
") Neurons of the Bed Nucleus of the Stria Terminalis (BNST) Electrophysiological Properties and Their Response to Serotonin DONALD G. RAINNIE a Harvard Medical School and Department of Psychiatry, Brockton
Neurons of the Bed Nucleus of the Stria Terminalis (BNST) Electrophysiological Properties and Their Response to Serotonin DONALD G. RAINNIE a Harvard Medical School and Department of Psychiatry, Brockton
BIPN 140 Problem Set 6
 BIPN 140 Problem Set 6 1) Hippocampus is a cortical structure in the medial portion of the temporal lobe (medial temporal lobe in primates. a) What is the main function of the hippocampus? The hippocampus
BIPN 140 Problem Set 6 1) Hippocampus is a cortical structure in the medial portion of the temporal lobe (medial temporal lobe in primates. a) What is the main function of the hippocampus? The hippocampus
Ultrastructural Contributions to Desensitization at the Cerebellar Mossy Fiber to Granule Cell Synapse
 Ultrastructural Contributions to Desensitization at the Cerebellar Mossy Fiber to Granule Cell Synapse Matthew A.Xu-Friedman and Wade G. Regehr Department of Neurobiology, Harvard Medical School, Boston,
Ultrastructural Contributions to Desensitization at the Cerebellar Mossy Fiber to Granule Cell Synapse Matthew A.Xu-Friedman and Wade G. Regehr Department of Neurobiology, Harvard Medical School, Boston,
Ube3a is required for experience-dependent maturation of the neocortex
 Ube3a is required for experience-dependent maturation of the neocortex Koji Yashiro, Thorfinn T. Riday, Kathryn H. Condon, Adam C. Roberts, Danilo R. Bernardo, Rohit Prakash, Richard J. Weinberg, Michael
Ube3a is required for experience-dependent maturation of the neocortex Koji Yashiro, Thorfinn T. Riday, Kathryn H. Condon, Adam C. Roberts, Danilo R. Bernardo, Rohit Prakash, Richard J. Weinberg, Michael
Astrocyte signaling controls spike timing-dependent depression at neocortical synapses
 Supplementary Information Astrocyte signaling controls spike timing-dependent depression at neocortical synapses Rogier Min and Thomas Nevian Department of Physiology, University of Berne, Bern, Switzerland
Supplementary Information Astrocyte signaling controls spike timing-dependent depression at neocortical synapses Rogier Min and Thomas Nevian Department of Physiology, University of Berne, Bern, Switzerland
Sample Lab Report 1 from 1. Measuring and Manipulating Passive Membrane Properties
 Sample Lab Report 1 from http://www.bio365l.net 1 Abstract Measuring and Manipulating Passive Membrane Properties Biological membranes exhibit the properties of capacitance and resistance, which allow
Sample Lab Report 1 from http://www.bio365l.net 1 Abstract Measuring and Manipulating Passive Membrane Properties Biological membranes exhibit the properties of capacitance and resistance, which allow
Part 11: Mechanisms of Learning
 Neurophysiology and Information: Theory of Brain Function Christopher Fiorillo BiS 527, Spring 2012 042 350 4326, fiorillo@kaist.ac.kr Part 11: Mechanisms of Learning Reading: Bear, Connors, and Paradiso,
Neurophysiology and Information: Theory of Brain Function Christopher Fiorillo BiS 527, Spring 2012 042 350 4326, fiorillo@kaist.ac.kr Part 11: Mechanisms of Learning Reading: Bear, Connors, and Paradiso,
CELLULAR NEUROPHYSIOLOGY
 CELLULAR NEUROPHYSIOLOGY CONSTANCE HAMMOND 4. SYNAPTIC TRANSMISSION II: GLUTAMATERGIC TRANSMISSION Video 4-1: Observations and glutamate receptor channels Synaptic transmission II 1 Constance Hammond Observation
CELLULAR NEUROPHYSIOLOGY CONSTANCE HAMMOND 4. SYNAPTIC TRANSMISSION II: GLUTAMATERGIC TRANSMISSION Video 4-1: Observations and glutamate receptor channels Synaptic transmission II 1 Constance Hammond Observation
Synaptic Integration
 Synaptic Integration 3 rd January, 2017 Touqeer Ahmed PhD Atta-ur-Rahman School of Applied Biosciences National University of Sciences and Technology Excitatory Synaptic Actions Excitatory Synaptic Action
Synaptic Integration 3 rd January, 2017 Touqeer Ahmed PhD Atta-ur-Rahman School of Applied Biosciences National University of Sciences and Technology Excitatory Synaptic Actions Excitatory Synaptic Action
Chapter 5 subtitles GABAergic synaptic transmission
 CELLULAR NEUROPHYSIOLOGY CONSTANCE HAMMOND Chapter 5 subtitles GABAergic synaptic transmission INTRODUCTION (2:57) In this fifth chapter, you will learn how the binding of the GABA neurotransmitter to
CELLULAR NEUROPHYSIOLOGY CONSTANCE HAMMOND Chapter 5 subtitles GABAergic synaptic transmission INTRODUCTION (2:57) In this fifth chapter, you will learn how the binding of the GABA neurotransmitter to
File name: Supplementary Information Description: Supplementary Figures, Supplementary Table and Supplementary References
 File name: Supplementary Information Description: Supplementary Figures, Supplementary Table and Supplementary References File name: Supplementary Data 1 Description: Summary datasheets showing the spatial
File name: Supplementary Information Description: Supplementary Figures, Supplementary Table and Supplementary References File name: Supplementary Data 1 Description: Summary datasheets showing the spatial
Ligand-Gated Ion Channels
 Ligand-Gated Ion Channels The Other Machines That Make It Possible... Topics I Introduction & Electrochemical Gradients Passive Membrane Properties Action Potentials Voltage-Gated Ion Channels Topics II
Ligand-Gated Ion Channels The Other Machines That Make It Possible... Topics I Introduction & Electrochemical Gradients Passive Membrane Properties Action Potentials Voltage-Gated Ion Channels Topics II
Abstract. 1 Introduction
 Biophysical model of a single synaptic connection: transmission properties are determined by the cooperation of pre- and postsynaptic mechanisms Julia Trommershäuser and Annette Zippelius Institut für
Biophysical model of a single synaptic connection: transmission properties are determined by the cooperation of pre- and postsynaptic mechanisms Julia Trommershäuser and Annette Zippelius Institut für
Synaptic Transmission: Ionic and Metabotropic
 Synaptic Transmission: Ionic and Metabotropic D. Purves et al. Neuroscience (Sinauer Assoc.) Chapters 5, 6, 7. C. Koch. Biophysics of Computation (Oxford) Chapter 4. J.G. Nicholls et al. From Neuron to
Synaptic Transmission: Ionic and Metabotropic D. Purves et al. Neuroscience (Sinauer Assoc.) Chapters 5, 6, 7. C. Koch. Biophysics of Computation (Oxford) Chapter 4. J.G. Nicholls et al. From Neuron to
Supplementary Figure 1. Basic properties of compound EPSPs at
 Supplementary Figure 1. Basic properties of compound EPSPs at hippocampal CA3 CA3 cell synapses. (a) EPSPs were evoked by extracellular stimulation of the recurrent collaterals and pharmacologically isolated
Supplementary Figure 1. Basic properties of compound EPSPs at hippocampal CA3 CA3 cell synapses. (a) EPSPs were evoked by extracellular stimulation of the recurrent collaterals and pharmacologically isolated
MOLECULAR AND CELLULAR NEUROSCIENCE
 MOLECULAR AND CELLULAR NEUROSCIENCE BMP-218 November 4, 2014 DIVISIONS OF THE NERVOUS SYSTEM The nervous system is composed of two primary divisions: 1. CNS - Central Nervous System (Brain + Spinal Cord)
MOLECULAR AND CELLULAR NEUROSCIENCE BMP-218 November 4, 2014 DIVISIONS OF THE NERVOUS SYSTEM The nervous system is composed of two primary divisions: 1. CNS - Central Nervous System (Brain + Spinal Cord)
Problem Set 3 - Answers. -70mV TBOA
 Harvard-MIT Division of Health Sciences and Technology HST.131: Introduction to Neuroscience Course Director: Dr. David Corey HST 131/ Neuro 200 18 September 05 Explanation in text below graphs. Problem
Harvard-MIT Division of Health Sciences and Technology HST.131: Introduction to Neuroscience Course Director: Dr. David Corey HST 131/ Neuro 200 18 September 05 Explanation in text below graphs. Problem
STRUCTURAL ELEMENTS OF THE NERVOUS SYSTEM
 STRUCTURAL ELEMENTS OF THE NERVOUS SYSTEM STRUCTURE AND MAINTENANCE OF NEURONS (a) (b) Dendrites Cell body Initial segment collateral terminals (a) Diagrammatic representation of a neuron. The break in
STRUCTURAL ELEMENTS OF THE NERVOUS SYSTEM STRUCTURE AND MAINTENANCE OF NEURONS (a) (b) Dendrites Cell body Initial segment collateral terminals (a) Diagrammatic representation of a neuron. The break in
Chapter 2: Cellular Mechanisms and Cognition
 Chapter 2: Cellular Mechanisms and Cognition MULTIPLE CHOICE 1. Two principles about neurons were defined by Ramón y Cajal. The principle of connectional specificity states that, whereas the principle
Chapter 2: Cellular Mechanisms and Cognition MULTIPLE CHOICE 1. Two principles about neurons were defined by Ramón y Cajal. The principle of connectional specificity states that, whereas the principle
Chapter 3 subtitles Action potentials
 CELLULAR NEUROPHYSIOLOGY CONSTANCE HAMMOND Chapter 3 subtitles Action potentials Introduction (3:15) This third chapter explains the calcium current triggered by the arrival of the action potential in
CELLULAR NEUROPHYSIOLOGY CONSTANCE HAMMOND Chapter 3 subtitles Action potentials Introduction (3:15) This third chapter explains the calcium current triggered by the arrival of the action potential in
Supplementary Information
 Supplementary Information D-Serine regulates cerebellar LTD and motor coordination through the 2 glutamate receptor Wataru Kakegawa, Yurika Miyoshi, Kenji Hamase, Shinji Matsuda, Keiko Matsuda, Kazuhisa
Supplementary Information D-Serine regulates cerebellar LTD and motor coordination through the 2 glutamate receptor Wataru Kakegawa, Yurika Miyoshi, Kenji Hamase, Shinji Matsuda, Keiko Matsuda, Kazuhisa
When cells are already maximally potentiated LTP is occluded.
 When cells are already maximally potentiated LTP is occluded. Stein, V et al., (2003) J Neurosci, 23:5503-6606. Also found in Rat Barrel Cortex Ehrlich & Malinow (2004) J. Neurosci. 24:916-927 Over-expression
When cells are already maximally potentiated LTP is occluded. Stein, V et al., (2003) J Neurosci, 23:5503-6606. Also found in Rat Barrel Cortex Ehrlich & Malinow (2004) J. Neurosci. 24:916-927 Over-expression
Supplementary Figure 1. SybII and Ceb are sorted to distinct vesicle populations in astrocytes. Nature Neuroscience: doi: /nn.
 Supplementary Figure 1 SybII and Ceb are sorted to distinct vesicle populations in astrocytes. (a) Exemplary images for cultured astrocytes co-immunolabeled with SybII and Ceb antibodies. SybII accumulates
Supplementary Figure 1 SybII and Ceb are sorted to distinct vesicle populations in astrocytes. (a) Exemplary images for cultured astrocytes co-immunolabeled with SybII and Ceb antibodies. SybII accumulates
Ivy/Neurogliaform Interneurons Coordinate Activity in the Neurogenic Niche
 Ivy/Neurogliaform Interneurons Coordinate Activity in the Neurogenic Niche Sean J. Markwardt, Cristina V. Dieni, Jacques I. Wadiche & Linda Overstreet-Wadiche Supplementary Methods. Animals We used hemizygous
Ivy/Neurogliaform Interneurons Coordinate Activity in the Neurogenic Niche Sean J. Markwardt, Cristina V. Dieni, Jacques I. Wadiche & Linda Overstreet-Wadiche Supplementary Methods. Animals We used hemizygous
photometry on the extruded cytoplasm.
 Answers To Midterm 2011 Question 1. a) Isoproterenol. Used to dissect presynaptic and postsynaptic components of sympathetic modulation of neuromuscular junction (Orbelli effect). Specifically activates
Answers To Midterm 2011 Question 1. a) Isoproterenol. Used to dissect presynaptic and postsynaptic components of sympathetic modulation of neuromuscular junction (Orbelli effect). Specifically activates
Supplementary figure 1: LII/III GIN-cells show morphological characteristics of MC
 1 2 1 3 Supplementary figure 1: LII/III GIN-cells show morphological characteristics of MC 4 5 6 7 (a) Reconstructions of LII/III GIN-cells with somato-dendritic compartments in orange and axonal arborizations
1 2 1 3 Supplementary figure 1: LII/III GIN-cells show morphological characteristics of MC 4 5 6 7 (a) Reconstructions of LII/III GIN-cells with somato-dendritic compartments in orange and axonal arborizations
Cellular Neurobiology / BIPN 140
 SECOND MIDTERM EXAMINATION Fall, 2015 GENERAL INSTRUCTIONS 1. Please write your name on ALL 6 pages. 2. Please answer each question IN THE SPACE ALLOTTED. 1) /10 pts 2) /10 pts 3) /15 pts 4) /15 pts 5)
SECOND MIDTERM EXAMINATION Fall, 2015 GENERAL INSTRUCTIONS 1. Please write your name on ALL 6 pages. 2. Please answer each question IN THE SPACE ALLOTTED. 1) /10 pts 2) /10 pts 3) /15 pts 4) /15 pts 5)
Lecture 22: A little Neurobiology
 BIO 5099: Molecular Biology for Computer Scientists (et al) Lecture 22: A little Neurobiology http://compbio.uchsc.edu/hunter/bio5099 Larry.Hunter@uchsc.edu Nervous system development Part of the ectoderm
BIO 5099: Molecular Biology for Computer Scientists (et al) Lecture 22: A little Neurobiology http://compbio.uchsc.edu/hunter/bio5099 Larry.Hunter@uchsc.edu Nervous system development Part of the ectoderm
Chapter 6 subtitles postsynaptic integration
 CELLULAR NEUROPHYSIOLOGY CONSTANCE HAMMOND Chapter 6 subtitles postsynaptic integration INTRODUCTION (1:56) This sixth and final chapter deals with the summation of presynaptic currents. Glutamate and
CELLULAR NEUROPHYSIOLOGY CONSTANCE HAMMOND Chapter 6 subtitles postsynaptic integration INTRODUCTION (1:56) This sixth and final chapter deals with the summation of presynaptic currents. Glutamate and
Nature Methods: doi: /nmeth Supplementary Figure 1. Activity in turtle dorsal cortex is sparse.
 Supplementary Figure 1 Activity in turtle dorsal cortex is sparse. a. Probability distribution of firing rates across the population (notice log scale) in our data. The range of firing rates is wide but
Supplementary Figure 1 Activity in turtle dorsal cortex is sparse. a. Probability distribution of firing rates across the population (notice log scale) in our data. The range of firing rates is wide but
Enhancement of synaptic transmission by cyclic AMP modulation of presynaptic I h channels. Vahri Beaumont and Robert S. Zucker
 Enhancement of synaptic transmission by cyclic AMP modulation of presynaptic I h channels Vahri Beaumont and Robert S. Zucker Background I h channels discovered in 1976 (Noma A. and Irisawa H.) Voltage-gated
Enhancement of synaptic transmission by cyclic AMP modulation of presynaptic I h channels Vahri Beaumont and Robert S. Zucker Background I h channels discovered in 1976 (Noma A. and Irisawa H.) Voltage-gated
Dep. Control Time (min)
") aa Control Dep. RP 1s 1 mv 2s 1 mv b % potentiation of IPSP 2 15 1 5 Dep. * 1 2 3 4 Time (min) Supplementary Figure 1. Rebound potentiation of IPSPs in PCs. a, IPSPs recorded with a K + gluconate pipette
aa Control Dep. RP 1s 1 mv 2s 1 mv b % potentiation of IPSP 2 15 1 5 Dep. * 1 2 3 4 Time (min) Supplementary Figure 1. Rebound potentiation of IPSPs in PCs. a, IPSPs recorded with a K + gluconate pipette
Synaptic communication between neurons and NG2 + cells Martin Paukert and Dwight E Bergles
 Synaptic communication between neurons and NG2 + cells Martin Paukert and Dwight E Bergles Chemical synaptic transmission provides the basis for much of the rapid signaling that occurs within neuronal
Synaptic communication between neurons and NG2 + cells Martin Paukert and Dwight E Bergles Chemical synaptic transmission provides the basis for much of the rapid signaling that occurs within neuronal
BIPN100 F15 Human Physiology 1 Lecture 3. Synaptic Transmission p. 1
 BIPN100 F15 Human Physiology 1 Lecture 3. Synaptic Transmission p. 1 Terms you should know: synapse, neuromuscular junction (NMJ), pre-synaptic, post-synaptic, synaptic cleft, acetylcholine (ACh), acetylcholine
BIPN100 F15 Human Physiology 1 Lecture 3. Synaptic Transmission p. 1 Terms you should know: synapse, neuromuscular junction (NMJ), pre-synaptic, post-synaptic, synaptic cleft, acetylcholine (ACh), acetylcholine
SYNAPTIC COMMUNICATION
 BASICS OF NEUROBIOLOGY SYNAPTIC COMMUNICATION ZSOLT LIPOSITS 1 NERVE ENDINGS II. Interneuronal communication 2 INTERNEURONAL COMMUNICATION I. ELECTRONIC SYNAPSE GAP JUNCTION II. CHEMICAL SYNAPSE SYNAPSES
BASICS OF NEUROBIOLOGY SYNAPTIC COMMUNICATION ZSOLT LIPOSITS 1 NERVE ENDINGS II. Interneuronal communication 2 INTERNEURONAL COMMUNICATION I. ELECTRONIC SYNAPSE GAP JUNCTION II. CHEMICAL SYNAPSE SYNAPSES
Supplementary Figure 1. GABA depolarizes the majority of immature neurons in the
 Supplementary Figure 1. GABA depolarizes the majority of immature neurons in the upper cortical layers at P3 4 in vivo. (a b) Cell-attached current-clamp recordings illustrate responses to puff-applied
Supplementary Figure 1. GABA depolarizes the majority of immature neurons in the upper cortical layers at P3 4 in vivo. (a b) Cell-attached current-clamp recordings illustrate responses to puff-applied
Quantal Release of ATP in Mouse Cortex
 ARTICLE Quantal Release of ATP in Mouse Cortex Yuriy Pankratov, Ulyana Lalo, Alexei Verkhratsky, and R. Alan North Faculty of Life Sciences, University of Manchester, Manchester M13 9PT, UK Transient currents
ARTICLE Quantal Release of ATP in Mouse Cortex Yuriy Pankratov, Ulyana Lalo, Alexei Verkhratsky, and R. Alan North Faculty of Life Sciences, University of Manchester, Manchester M13 9PT, UK Transient currents
Chapter 3 Neurotransmitter release
 NEUROPHYSIOLOGIE CELLULAIRE CONSTANCE HAMMOND Chapter 3 Neurotransmitter release In chapter 3, we proose 3 videos: Observation Calcium Channel, Ca 2+ Unitary and Total Currents Ca 2+ and Neurotransmitter
NEUROPHYSIOLOGIE CELLULAIRE CONSTANCE HAMMOND Chapter 3 Neurotransmitter release In chapter 3, we proose 3 videos: Observation Calcium Channel, Ca 2+ Unitary and Total Currents Ca 2+ and Neurotransmitter
Modeling Depolarization Induced Suppression of Inhibition in Pyramidal Neurons
 Modeling Depolarization Induced Suppression of Inhibition in Pyramidal Neurons Peter Osseward, Uri Magaram Department of Neuroscience University of California, San Diego La Jolla, CA 92092 possewar@ucsd.edu
Modeling Depolarization Induced Suppression of Inhibition in Pyramidal Neurons Peter Osseward, Uri Magaram Department of Neuroscience University of California, San Diego La Jolla, CA 92092 possewar@ucsd.edu
The action potential travels down both branches because each branch is a typical axon with voltage dependent Na + and K+ channels.
 BIO 360 - MIDTERM FALL 2018 This is an open book, open notes exam. PLEASE WRITE YOUR NAME ON EACH SHEET. Read each question carefully and answer as well as you can. Point values are shown at the beginning
BIO 360 - MIDTERM FALL 2018 This is an open book, open notes exam. PLEASE WRITE YOUR NAME ON EACH SHEET. Read each question carefully and answer as well as you can. Point values are shown at the beginning
Human TRPC6 Ion Channel Cell Line
 TECHNICAL DATA SHEET ValiScreen Ion Channel Cell Line Caution: For Laboratory Use. A research product for research purposes only Human TRPC6 Ion Channel Cell Line Product No.: AX-012-C Lot No.: 512-548-A
TECHNICAL DATA SHEET ValiScreen Ion Channel Cell Line Caution: For Laboratory Use. A research product for research purposes only Human TRPC6 Ion Channel Cell Line Product No.: AX-012-C Lot No.: 512-548-A
Changes in Extracellular Ionic Composition q
 Changes in Extracellular Ionic Composition q JL Stringer, Baylor College of Medicine, Houston, TX, United States Ó 2017 Elsevier Inc. All rights reserved. Introduction 1 Background 1 Methods 2 Recent Results
Changes in Extracellular Ionic Composition q JL Stringer, Baylor College of Medicine, Houston, TX, United States Ó 2017 Elsevier Inc. All rights reserved. Introduction 1 Background 1 Methods 2 Recent Results
Chapter 2. The Cellular and Molecular Basis of Cognition Cognitive Neuroscience: The Biology of the Mind, 2 nd Ed.,
 Chapter 2. The Cellular and Molecular Basis of Cognition Cognitive Neuroscience: The Biology of the Mind, 2 nd Ed., M. S. Gazzaniga, R. B. Ivry, and G. R. Mangun, Norton, 2002. Summarized by B.-W. Ku,
Chapter 2. The Cellular and Molecular Basis of Cognition Cognitive Neuroscience: The Biology of the Mind, 2 nd Ed., M. S. Gazzaniga, R. B. Ivry, and G. R. Mangun, Norton, 2002. Summarized by B.-W. Ku,
Nervous System. Master controlling and communicating system of the body. Secrete chemicals called neurotransmitters
 Nervous System Master controlling and communicating system of the body Interacts with the endocrine system to control and coordinate the body s responses to changes in its environment, as well as growth,
Nervous System Master controlling and communicating system of the body Interacts with the endocrine system to control and coordinate the body s responses to changes in its environment, as well as growth,
Supplementary Figure 1) GABAergic enhancement by leptin hyperpolarizes POMC neurons A) Representative recording samples showing the membrane
 GABAergic enhancement by leptin hyperpolarizes POMC neurons A) Representative recording samples showing the membrane") Supplementary Figure 1) GABAergic enhancement by leptin hyperpolarizes POMC neurons A) Representative recording samples showing the membrane potential recorded from POMC neurons following treatment with
Supplementary Figure 1) GABAergic enhancement by leptin hyperpolarizes POMC neurons A) Representative recording samples showing the membrane potential recorded from POMC neurons following treatment with
de Montpellier, Montpellier, France and Ecole Pratique des Hautes Etudes Sorbonne, Paris, France.
 SHORT COMMUNICATION Analysis of Ca 2+ Signaling Mechanisms Our Experience on the Intercellular Communication in Muscle Remodeling S. Filip 1, J. Mokrý 2, O. Forostyak 3, G. Dayanithi 4,5 1 Charles University,
SHORT COMMUNICATION Analysis of Ca 2+ Signaling Mechanisms Our Experience on the Intercellular Communication in Muscle Remodeling S. Filip 1, J. Mokrý 2, O. Forostyak 3, G. Dayanithi 4,5 1 Charles University,
Activity-Dependent Change in AMPA Receptor Properties in Cerebellar Stellate Cells
 The Journal of Neuroscience, May 15, 2002, 22(10):3881 3889 Activity-Dependent Change in AMPA Receptor Properties in Cerebellar Stellate Cells Siqiong June Liu and Stuart G. Cull-Candy Department of Pharmacology,
The Journal of Neuroscience, May 15, 2002, 22(10):3881 3889 Activity-Dependent Change in AMPA Receptor Properties in Cerebellar Stellate Cells Siqiong June Liu and Stuart G. Cull-Candy Department of Pharmacology,
Alterations in Synaptic Strength Preceding Axon Withdrawal
 Alterations in Synaptic Strength Preceding Axon Withdrawal H. Colman, J. Nabekura, J.W. Lichtman presented by Ana Fiallos Synaptic Transmission at the Neuromuscular Junction Motor neurons with cell bodies
Alterations in Synaptic Strength Preceding Axon Withdrawal H. Colman, J. Nabekura, J.W. Lichtman presented by Ana Fiallos Synaptic Transmission at the Neuromuscular Junction Motor neurons with cell bodies
Neuroscience 201A Problem Set #1, 27 September 2016
 Neuroscience 201A Problem Set #1, 27 September 2016 1. The figure above was obtained from a paper on calcium channels expressed by dentate granule cells. The whole-cell Ca 2+ currents in (A) were measured
Neuroscience 201A Problem Set #1, 27 September 2016 1. The figure above was obtained from a paper on calcium channels expressed by dentate granule cells. The whole-cell Ca 2+ currents in (A) were measured
Chapter 4 Neuronal Physiology
 Chapter 4 Neuronal Physiology V edit. Pg. 99-131 VI edit. Pg. 85-113 VII edit. Pg. 87-113 Input Zone Dendrites and Cell body Nucleus Trigger Zone Axon hillock Conducting Zone Axon (may be from 1mm to more
Chapter 4 Neuronal Physiology V edit. Pg. 99-131 VI edit. Pg. 85-113 VII edit. Pg. 87-113 Input Zone Dendrites and Cell body Nucleus Trigger Zone Axon hillock Conducting Zone Axon (may be from 1mm to more
Chapter 11 Introduction to the Nervous System and Nervous Tissue Chapter Outline
 Chapter 11 Introduction to the Nervous System and Nervous Tissue Chapter Outline Module 11.1 Overview of the Nervous System (Figures 11.1-11.3) A. The nervous system controls our perception and experience
Chapter 11 Introduction to the Nervous System and Nervous Tissue Chapter Outline Module 11.1 Overview of the Nervous System (Figures 11.1-11.3) A. The nervous system controls our perception and experience
Chapter 2. The Cellular and Molecular Basis of Cognition
 Chapter 2. The Cellular and Molecular Basis of Cognition Cognitive Neuroscience: The Biology of the Mind, 2 nd Ed., M. S. Gazzaniga,, R. B. Ivry,, and G. R. Mangun,, Norton, 2002. Summarized by B.-W. Ku,
Chapter 2. The Cellular and Molecular Basis of Cognition Cognitive Neuroscience: The Biology of the Mind, 2 nd Ed., M. S. Gazzaniga,, R. B. Ivry,, and G. R. Mangun,, Norton, 2002. Summarized by B.-W. Ku,
Neurobiology: The nerve cell. Principle and task To use a nerve function model to study the following aspects of a nerve cell:
 Principle and task To use a nerve function model to study the following aspects of a nerve cell: INTRACELLULAR POTENTIAL AND ACTION POTENTIAL Comparison between low and high threshold levels Comparison
Principle and task To use a nerve function model to study the following aspects of a nerve cell: INTRACELLULAR POTENTIAL AND ACTION POTENTIAL Comparison between low and high threshold levels Comparison
The Journal of Physiology
 J Physiol 589.9 (2011) pp 2301 2319 2301 Contribution of astrocytic glutamate and GABA uptake to corticostriatal information processing Valérie Goubard, Elodie Fino and Laurent Venance Dynamic and Pathophysiology
J Physiol 589.9 (2011) pp 2301 2319 2301 Contribution of astrocytic glutamate and GABA uptake to corticostriatal information processing Valérie Goubard, Elodie Fino and Laurent Venance Dynamic and Pathophysiology
Neurotransmitter Systems II Receptors. Reading: BCP Chapter 6
 Neurotransmitter Systems II Receptors Reading: BCP Chapter 6 Neurotransmitter Systems Normal function of the human brain requires an orderly set of chemical reactions. Some of the most important chemical
Neurotransmitter Systems II Receptors Reading: BCP Chapter 6 Neurotransmitter Systems Normal function of the human brain requires an orderly set of chemical reactions. Some of the most important chemical
Bursting dynamics in the brain. Jaeseung Jeong, Department of Biosystems, KAIST
 Bursting dynamics in the brain Jaeseung Jeong, Department of Biosystems, KAIST Tonic and phasic activity A neuron is said to exhibit a tonic activity when it fires a series of single action potentials
Bursting dynamics in the brain Jaeseung Jeong, Department of Biosystems, KAIST Tonic and phasic activity A neuron is said to exhibit a tonic activity when it fires a series of single action potentials
Differential Mechanisms of Ca 2 Responses in Glial Cells Evoked by Exogenous and Endogenous Glutamate in Rat Hippocampus
 Differential Mechanisms of Ca 2 Responses in Glial Cells Evoked by Exogenous and Endogenous Glutamate in Rat Hippocampus Isabelle Latour, Christine E. Gee, Richard Robitaille, and Jean-Claude Lacaille*
Differential Mechanisms of Ca 2 Responses in Glial Cells Evoked by Exogenous and Endogenous Glutamate in Rat Hippocampus Isabelle Latour, Christine E. Gee, Richard Robitaille, and Jean-Claude Lacaille*
Short- and long-lasting consequences of in vivo nicotine treatment
 Short- and long-lasting consequences of in vivo nicotine treatment on hippocampal excitability Rachel E. Penton, Michael W. Quick, Robin A. J. Lester Supplementary Figure 1. Histogram showing the maximal
Short- and long-lasting consequences of in vivo nicotine treatment on hippocampal excitability Rachel E. Penton, Michael W. Quick, Robin A. J. Lester Supplementary Figure 1. Histogram showing the maximal
Unique functional properties of somatostatin-expressing GABAergic neurons in mouse barrel cortex
 Supplementary Information Unique functional properties of somatostatin-expressing GABAergic neurons in mouse barrel cortex Luc Gentet, Yves Kremer, Hiroki Taniguchi, Josh Huang, Jochen Staiger and Carl
Supplementary Information Unique functional properties of somatostatin-expressing GABAergic neurons in mouse barrel cortex Luc Gentet, Yves Kremer, Hiroki Taniguchi, Josh Huang, Jochen Staiger and Carl
Prolonged Synaptic Integration in Perirhinal Cortical Neurons
 RAPID COMMUNICATION Prolonged Synaptic Integration in Perirhinal Cortical Neurons JOHN M. BEGGS, 1 JAMES R. MOYER, JR., 1 JOHN P. MCGANN, 2 AND THOMAS H. BROWN 1 3 1 Department of Psychology, 2 Interdepartmental
RAPID COMMUNICATION Prolonged Synaptic Integration in Perirhinal Cortical Neurons JOHN M. BEGGS, 1 JAMES R. MOYER, JR., 1 JOHN P. MCGANN, 2 AND THOMAS H. BROWN 1 3 1 Department of Psychology, 2 Interdepartmental
Decreased Frequency But Not Amplitude of Quantal Synaptic Responses Associated with Expression of Corticostriatal Long-Term Depression
 The Journal of Neuroscience, November 1, 1997, 17(21):8613 8620 Decreased Frequency But Not Amplitude of Quantal Synaptic Responses Associated with Expression of Corticostriatal Long-Term Depression Sukwoo
The Journal of Neuroscience, November 1, 1997, 17(21):8613 8620 Decreased Frequency But Not Amplitude of Quantal Synaptic Responses Associated with Expression of Corticostriatal Long-Term Depression Sukwoo
What Cell Make Up the Brain and Spinal Cord
 What Cell Make Up the Brain and Spinal Cord Jennifer LaVail, Ph.D. (http://anatomy.ucsf.edu/pages/lavaillab/index.html) What kinds of cells are these?" Neuron?" Epithelial cell?" Glial cell?" What makes
What Cell Make Up the Brain and Spinal Cord Jennifer LaVail, Ph.D. (http://anatomy.ucsf.edu/pages/lavaillab/index.html) What kinds of cells are these?" Neuron?" Epithelial cell?" Glial cell?" What makes
Ameen Alsaras. Ameen Alsaras. Mohd.Khatatbeh
 9 Ameen Alsaras Ameen Alsaras Mohd.Khatatbeh Nerve Cells (Neurons) *Remember: The neural cell consists of: 1-Cell body 2-Dendrites 3-Axon which ends as axon terminals. The conduction of impulse through
9 Ameen Alsaras Ameen Alsaras Mohd.Khatatbeh Nerve Cells (Neurons) *Remember: The neural cell consists of: 1-Cell body 2-Dendrites 3-Axon which ends as axon terminals. The conduction of impulse through
How neurons talk. Nivedita Chatterjee Vision Research Foundation, Sankara Nethralaya, Chennai
 How neurons talk Nivedita Chatterjee Vision Research Foundation, Sankara Nethralaya, Chennai The Synaptic Bouton of a chemical synapse Chemical synapse with synaptic cleft The Electrical synapse works
How neurons talk Nivedita Chatterjee Vision Research Foundation, Sankara Nethralaya, Chennai The Synaptic Bouton of a chemical synapse Chemical synapse with synaptic cleft The Electrical synapse works
Electrical Properties of Neurons. Steven McLoon Department of Neuroscience University of Minnesota
 Electrical Properties of Neurons Steven McLoon Department of Neuroscience University of Minnesota 1 Neuronal Communication Neurons communicate with other cells, often over long distances. The electrical
Electrical Properties of Neurons Steven McLoon Department of Neuroscience University of Minnesota 1 Neuronal Communication Neurons communicate with other cells, often over long distances. The electrical
Charlie Taylor, PhD CpTaylor Consulting Chelsea, MI, USA
 Contribution of Calcium Channel α 2 δ Binding Sites to the Pharmacology of Gabapentin and Pregabalin Charlie Taylor, PhD CpTaylor Consulting Chelsea, MI, USA Disclosure Information Charlie Taylor, PhD
Contribution of Calcium Channel α 2 δ Binding Sites to the Pharmacology of Gabapentin and Pregabalin Charlie Taylor, PhD CpTaylor Consulting Chelsea, MI, USA Disclosure Information Charlie Taylor, PhD
Structure of a Neuron:
 Structure of a Neuron: At the dendrite the incoming signals arrive (incoming currents) At the soma current are finally integrated. At the axon hillock action potential are generated if the potential crosses
Structure of a Neuron: At the dendrite the incoming signals arrive (incoming currents) At the soma current are finally integrated. At the axon hillock action potential are generated if the potential crosses
Chapter 12 Nervous Tissue. Copyright 2009 John Wiley & Sons, Inc. 1
 Chapter 12 Nervous Tissue Copyright 2009 John Wiley & Sons, Inc. 1 Terms to Know CNS PNS Afferent division Efferent division Somatic nervous system Autonomic nervous system Sympathetic nervous system Parasympathetic
Chapter 12 Nervous Tissue Copyright 2009 John Wiley & Sons, Inc. 1 Terms to Know CNS PNS Afferent division Efferent division Somatic nervous system Autonomic nervous system Sympathetic nervous system Parasympathetic
The Nervous System. Nervous System Functions 1. gather sensory input 2. integration- process and interpret sensory input 3. cause motor output
 The Nervous System Nervous System Functions 1. gather sensory input 2. integration- process and interpret sensory input 3. cause motor output The Nervous System 2 Parts of the Nervous System 1. central
The Nervous System Nervous System Functions 1. gather sensory input 2. integration- process and interpret sensory input 3. cause motor output The Nervous System 2 Parts of the Nervous System 1. central
NS200: In vitro electrophysiology section September 11th, 2013
 NS200: In vitro electrophysiology section September 11th, 2013 Quynh Anh Nguyen, 4 th Year Nicoll Lab quynhanh.nguyen@ucsf.edu N276 Genentech Hall, Mission Bay Outline Part I: Theory Review of circuit
NS200: In vitro electrophysiology section September 11th, 2013 Quynh Anh Nguyen, 4 th Year Nicoll Lab quynhanh.nguyen@ucsf.edu N276 Genentech Hall, Mission Bay Outline Part I: Theory Review of circuit
Introduction to Electrophysiology
 Introduction to Electrophysiology Dr. Kwangyeol Baek Martinos Center for Biomedical Imaging Massachusetts General Hospital Harvard Medical School 2018-05-31s Contents Principles in Electrophysiology Techniques
Introduction to Electrophysiology Dr. Kwangyeol Baek Martinos Center for Biomedical Imaging Massachusetts General Hospital Harvard Medical School 2018-05-31s Contents Principles in Electrophysiology Techniques
Supralinear increase of recurrent inhibition during sparse activity in the somatosensory cortex
 Supralinear increase of recurrent inhibition during sparse activity in the somatosensory cortex Christoph Kapfer 1,2, Lindsey L Glickfeld 1,3, Bassam V Atallah 1,3 & Massimo Scanziani 1 The balance between
Supralinear increase of recurrent inhibition during sparse activity in the somatosensory cortex Christoph Kapfer 1,2, Lindsey L Glickfeld 1,3, Bassam V Atallah 1,3 & Massimo Scanziani 1 The balance between
QUIZ/TEST REVIEW NOTES SECTION 7 NEUROPHYSIOLOGY [THE SYNAPSE AND PHARMACOLOGY]
![QUIZ/TEST REVIEW NOTES SECTION 7 NEUROPHYSIOLOGY [THE SYNAPSE AND PHARMACOLOGY]](/thumbs/86/94924826.jpg "QUIZ/TEST REVIEW NOTES SECTION 7 NEUROPHYSIOLOGY [THE SYNAPSE AND PHARMACOLOGY]") QUIZ/TEST REVIEW NOTES SECTION 7 NEUROPHYSIOLOGY [THE SYNAPSE AND PHARMACOLOGY] Learning Objectives: Explain how neurons communicate stimulus intensity Explain how action potentials are conducted along
QUIZ/TEST REVIEW NOTES SECTION 7 NEUROPHYSIOLOGY [THE SYNAPSE AND PHARMACOLOGY] Learning Objectives: Explain how neurons communicate stimulus intensity Explain how action potentials are conducted along
EE 791 Lecture 2 Jan 19, 2015
 EE 791 Lecture 2 Jan 19, 2015 Action Potential Conduction And Neural Organization EE 791-Lecture 2 1 Core-conductor model: In the core-conductor model we approximate an axon or a segment of a dendrite
EE 791 Lecture 2 Jan 19, 2015 Action Potential Conduction And Neural Organization EE 791-Lecture 2 1 Core-conductor model: In the core-conductor model we approximate an axon or a segment of a dendrite
Model neurons!!!!synapses!
 Model neurons ynapses uggested reading: Chapter 5.8 in Dayan,. & Abbott, L., Theoretical Neuroscience, MIT ress, 200. Model neurons: ynapse Contents: ynapses ynaptic input into the RC-circuit pike-rate
Model neurons ynapses uggested reading: Chapter 5.8 in Dayan,. & Abbott, L., Theoretical Neuroscience, MIT ress, 200. Model neurons: ynapse Contents: ynapses ynaptic input into the RC-circuit pike-rate
What is Anatomy and Physiology?
 Introduction BI 212 BI 213 BI 211 Ecosystems Organs / organ systems Cells Organelles Communities Tissues Molecules Populations Organisms Campbell et al. Figure 1.4 Introduction What is Anatomy and Physiology?
Introduction BI 212 BI 213 BI 211 Ecosystems Organs / organ systems Cells Organelles Communities Tissues Molecules Populations Organisms Campbell et al. Figure 1.4 Introduction What is Anatomy and Physiology?
Cellular mechanisms of information transfer: neuronal and synaptic plasticity
 Cellular mechanisms of information transfer: neuronal and synaptic plasticity Ivan Pavlov (UCL Institute of Neurology, UK) Anton Chizhov (Ioffe Physical Technical Institute) Pavel Zykin (St.-Petersburg
Cellular mechanisms of information transfer: neuronal and synaptic plasticity Ivan Pavlov (UCL Institute of Neurology, UK) Anton Chizhov (Ioffe Physical Technical Institute) Pavel Zykin (St.-Petersburg
Chapter 11: Nervous System and Nervous Tissue
 Chapter 11: Nervous System and Nervous Tissue I. Functions and divisions of the nervous system A. Sensory input: monitor changes in internal and external environment B. Integrations: make decisions about
Chapter 11: Nervous System and Nervous Tissue I. Functions and divisions of the nervous system A. Sensory input: monitor changes in internal and external environment B. Integrations: make decisions about
Activity Dependent Changes At the Developing Neuromuscular Junction
 Activity Dependent Changes At the Developing Neuromuscular Junction (slides 16, 17 and 18 have been slightly modified for clarity) MCP Lecture 2-3 9.013/7.68 04 Neuromuscular Junction Development 1. Muscle
Activity Dependent Changes At the Developing Neuromuscular Junction (slides 16, 17 and 18 have been slightly modified for clarity) MCP Lecture 2-3 9.013/7.68 04 Neuromuscular Junction Development 1. Muscle
Human Brain and Senses
 Human Brain and Senses Outline for today Levels of analysis Basic structure of neurons How neurons communicate Basic structure of the nervous system Levels of analysis Organism Brain Cell Synapses Membrane
Human Brain and Senses Outline for today Levels of analysis Basic structure of neurons How neurons communicate Basic structure of the nervous system Levels of analysis Organism Brain Cell Synapses Membrane
3) Most of the organelles in a neuron are located in the A) dendritic region. B) axon hillock. C) axon. D) cell body. E) axon terminals.
 Most of the organelles in a neuron are located in the A) dendritic region. B) axon hillock. C) axon. D) cell body. E) axon terminals.") Chapter 48 Neurons, Synapses, and Signaling Multiple-Choice Questions 1) A simple nervous system A) must include chemical senses, mechanoreception, and vision. B) includes a minimum of 12 ganglia. C) has
Chapter 48 Neurons, Synapses, and Signaling Multiple-Choice Questions 1) A simple nervous system A) must include chemical senses, mechanoreception, and vision. B) includes a minimum of 12 ganglia. C) has
Seizure: the clinical manifestation of an abnormal and excessive excitation and synchronization of a population of cortical
 Are There Sharing Mechanisms of Epilepsy, Migraine and Neuropathic Pain? Chin-Wei Huang, MD, PhD Department of Neurology, NCKUH Basic mechanisms underlying seizures and epilepsy Seizure: the clinical manifestation
Are There Sharing Mechanisms of Epilepsy, Migraine and Neuropathic Pain? Chin-Wei Huang, MD, PhD Department of Neurology, NCKUH Basic mechanisms underlying seizures and epilepsy Seizure: the clinical manifestation
Excitatory synaptic transmission in the central nervous system. Mechanisms underlying kainate receptor-mediated disinhibition in the hippocampus
 Mechanisms underlying kainate receptor-mediated disinhibition in the hippocampus M. Frerking*, C. C. H. Petersen*, and R. A. Nicoll* Departments of *Cellular and Molecular Pharmacology and Physiology,
Mechanisms underlying kainate receptor-mediated disinhibition in the hippocampus M. Frerking*, C. C. H. Petersen*, and R. A. Nicoll* Departments of *Cellular and Molecular Pharmacology and Physiology,
BIPN140 Lecture 8: Synaptic Transmission II
 BIPN140 Lecture 8: Synaptic Transmission II 1. Postsynaptic Receptors: Metabotropic & Ionotropic 2. Postsynaptic Responses (Postsynaptic Potentials, PSPs) 3. Neurotransmitters Su (FA16) Chemical Synapse:
BIPN140 Lecture 8: Synaptic Transmission II 1. Postsynaptic Receptors: Metabotropic & Ionotropic 2. Postsynaptic Responses (Postsynaptic Potentials, PSPs) 3. Neurotransmitters Su (FA16) Chemical Synapse:
Quantal Analysis Problems
 Quantal Analysis Problems 1. Imagine you had performed an experiment on a muscle preparation from a Drosophila larva. In this experiment, intracellular recordings were made from an identified muscle fibre,
Quantal Analysis Problems 1. Imagine you had performed an experiment on a muscle preparation from a Drosophila larva. In this experiment, intracellular recordings were made from an identified muscle fibre,
Memory Systems II How Stored: Engram and LTP. Reading: BCP Chapter 25
 Memory Systems II How Stored: Engram and LTP Reading: BCP Chapter 25 Memory Systems Learning is the acquisition of new knowledge or skills. Memory is the retention of learned information. Many different
Memory Systems II How Stored: Engram and LTP Reading: BCP Chapter 25 Memory Systems Learning is the acquisition of new knowledge or skills. Memory is the retention of learned information. Many different
Supporting Online Material for
 www.sciencemag.org/cgi/content/full/331/6017/599/dc1 Supporting Online Material for Action-Potential Modulation During Axonal Conduction Takuya Sasaki, Norio Matsuki, Yuji Ikegaya* *To whom correspondence
www.sciencemag.org/cgi/content/full/331/6017/599/dc1 Supporting Online Material for Action-Potential Modulation During Axonal Conduction Takuya Sasaki, Norio Matsuki, Yuji Ikegaya* *To whom correspondence