Transplant Rabbit Embryos1
|
|
|
- Randell Harmon
- 5 years ago
- Views:
Transcription

1 BIOLOGY OF REPRODUCTION 46, (1992) Effect of Donor Cell Cycle Stage on Chromatin and Spindle Morphology in Nuclear Transplant Rabbit Embryos1 PHILIPPE COLLAS,3 CLARA PINTO-CORREIA, F. ABEL PONCE DE LEON, and JAMES M. ROBL2 Department of Veterinary and Animal Sciences, University of Massachusetts, Amherst, Massachusetts ABSTRACT We investigated the influence of the cell cycle stage of the nuclear donor on prematurely condensed chromatin (PCC) and spindle morphology and on chromosome constitution in rabbit nuclear transplant embryos. The configuration of PCC following nuclear transplantation with Gl, early S, and late 5 phase donor nuclei (Gl, early S, and late S transplants, respectively) was characterized in whole mounts and chromosome spreads. In addition, the influence of the donor cell cycle stage on chromosome constitution in cleavage stage-manipulated embryos was determined. Within 2 h after fusion of the donor blastomere, the recipient oocyte cytoplasm was able to induce formation de novo of a metaphase plate associated with a spindle in G 1, early S, and late 5 transplants. Metaphase chromosomes and spindle were intact in most cases of PCC in Gi transplants. However, these structures displayed minor abnormalities in early S transplants and gross abnormalities in late S transplants, such as incomplete or absent spindle formation and incomplete chromatin condensation. Normal chromosomes were present in GI and early S transplants, whereas chromosome abnormalities were detected in late S transplants. The results indicate that morphology of prematurely condensed Gi and early S chromatin has a minor influence on chromosome constitution of manipulated embryos. That of late S chromatin, however, affects chromosome constitution in embryos and may account for reduced development of nuclear transplant embryos when late S phase donor nuclei are used. INTRODUCTION Transplantation of an interphase nucleus into an enucleated, mature oocvte leads to premature condensation of the donor chromatin, followed by formation and swelling of the nucleus [1]. Premature chromosome condensation (PCC) has been observed in mouse [2-6] and rabbit nuclear transplant embryos [1]. PCC and nuclear swelling have been shown to be beneficial for development of reconstituted rabbit embryos [1]. Furthermore, the position of the donor nucleus in the cell cycle influences the extent of development in vitro of nuclear transplant rabbit embryos [7]. Transfer of donor nuclei in the Gi phase of the cell cycle yields high rates of development to blastocysts; however, development is reduced with donor nuclei in early S phase and is dramatically impaired with those in late S phase. Therefore, as donor nuclei progress in the cell cycle, the extent of development of reconstituted embryos is reduced [7]. Fusion of a somatic cell in interphase with a mitotic cell also results in PCC of the interphase nucleus [8-10]. In somatic cells, PCC is manifested by breakdown of the nuclear envelope and condensation of the interphase chromatin without formation of a spindle [8]. In addition, the morphological changes that occur in the interphase chromatin have been shown to depend on the position of the interphase cell in the division cycle. In particular, PCC in Gi and G2 phases results in elongated chromosomes with single- and double-stranded chromatids, respectively [8-il]. Chromatin in S phase, however, condenses less completely Accepted November 4, Received July 17, This work was supported in part by USDA grant No to J.M.R. Correspondence. tcurrent address: GeivMark. Inc Wakara Way. Salt Lake Cirs-, UT and displays numerous fragments, separated by condensed regions [8-ui]. These studies indicate, therefore, that the cell cycle stage of the interphase nucleus greatly affects the configuration of PCC in mitotic somatic cells. The stage of the cycle of the interphase donor nucleus not only influences the morphology of PCC, it also affects the extent of the genetic contribution of the interphase nucleus to the genome of the hybrid cell [12-15]. Several studies have demonstrated that chromosome fragments, hut not intact chromosomes, resulting from prematurely condensed chromatin, are able to survive in the descendents of heterokaryons [12, 13]. It was suggested that such fragments survive either independently or by integration into chromosomes of the partner nucleus. However, results demonstrating the stability of these integrations have been inconsistent [12, 14, 15]. In addition, it has been reported that the likelihood of contributing genetic markers to the partner nucleus was higher with prematurely condensed Gi or G2 chromosomes than with S phase chromatin [13]. These results support the conclusion that the developmental rate of nuclear transplant embryos is higher with GI than with S phase donor nuclei [7]. The importance of cell cycle stage of the donor nucleus on development of nuclear transplant embryos has already been documented. The present study investigated the effect of donor cell cycle stage on the morphology of PCC and spindle, and on chromosome constitution in nuclear transplant embryos. We characterized the morphology of chromatin that had been prematurely condensed in the Gi, early 5, and late S phases of the cell cycle. The influence of donor cell cycle stage on chromosome constitution in embryos reconstituted in the cleavage stage was also determined. 501
 [16] followed by a single administration of 75 IU hcg (Sigma, St. Louis, MO).")
2 502 COLLAS ET AL. MATERIALS AND METHODS Embryo Micromanipulation Recipient Oocyte and Donor Embrpo Collection Mature mixed-breed female rabbits were superovulated with six injections of 0.3 mg FSH (Burnes Biotech, Omaha, NE) [16] followed by a single administration of 75 IU hcg (Sigma, St. Louis, MO). Ovulated oocytes were recovered from the oviducts h after hcg injection by flushing with 37#{176}C gassed Earles balanced salt solution and 10% fetal calf serum (EBSS/FCS; ph 7.2), and the cumulus was removed with hyaluronidase (Sigma) as described elsewhere [1, 17, 18]. Nonsuperovulated females served as embryo donors and were mated to an intact male. Late 16-cellstage embryos were recovered from the oviducts with 37#{176}C PBS 54 h after mating and were cultured until blastomere synchronization. Embryo Synchronization Donor blastomeres were used in the Gi phase (Gi transplants), early S phase (early S transplants), and late S phase (late S transplants) of the cell cycle. A procedure for synchronizing donor blastomeres in Gi in a reversible manner was previously determined [7]. Briefly, 16-cell-stage embryos were cultured for 10 h in the presence of 0.5 p.g/ ml of the microtuhule inhibitor, colcemid, to arrest them in M phase between the 16- and 32-cell stages. They were then extensively washed in PBS and immediately incubated in 0.1 g/ml of the DNA synthesis inhibitor aphidicolin. Cleavage to the 32-cell stage in aphidicolin was allowed, and embryos remained arrested at the GuS transition until used for nuclear transfer [7]. Blastomeres were isolated in the presence of aphidicolin as described previously [1, 17, 18]. Synchronization of 32-cell-stage embryos in early S entailed culturing 16-cell-stage embryos in 0.5 g/ml colcemid for 10 h. After a thorough wash in PBS, they were allowed to cleave in culture without aphidicolin, and blastomeres were used within 1.5 h after mitosis. Thirty-two-cell-stage embryos used in late S were cultured without any inhibitor for 16 h after recovery from the oviducts at the 16-cell stage. Early and late S donor blastomeres were isolated. Colcemid and aphidicolin (Sigma) were reconstituted at 2 mg/mi in PBS and dimethylsulfoxide (DMSO), respectively, aliquoted, and stored at -10#{176}C, Embryo Culture Embryos were cultured in vitro in vitreous humor (VH) from the eyes of female rabbits as reported elsewhere [1, 7; P. Collas, R.T. Duby, and J.M. Robl, unpublished results]. Briefly, VH was aspirated aseptically from the eyes of killed rabbits and centrifuged at X g for 2 mm to pellet cellular debris. The supernatant was filtered (0.2 pan) in microdrops overlaid with equilibrated paraffin oil. Culture dishes (Falcon No. 1007, \WR Scientific, Boston, MA) were incubated at 39#{176}C in an atmosphere of 5% CO2 in air for 12 h prior to use [1]. Nuclear transfers were performed in gassed EBSS/FCS with 7.5 pg/ml cytochalasin B (Sigma) as previously described [1, 18]. With Gi donor nuclei, however, micromanipulation was carried out in the presence of 0.1 g/ml aphidicolin, and embryos were cultured with aphidicolin until fusion and activation as described previously [7]. Blastomere Fusion and Oocyte Activation Fusion and activation were performed electrically [1, 17, 18]. Manipulated embryos were placed between two electrodes 0.5 mm apart, overlaid with 0.30 M mannitol containing 100 p.m CaCl2 and MgC12 [16] at 30#{176}C. Six direct current pulses of 2.2 ky/cm for 60 p.sec, each pulse 30 mm apart, starting 18 h post-hcg, were given using a BTX Electro Cell Manipulator 200 (BTX, Inc., San Diego, CA); and embryos were placed in culture between each pulse. For Gi transplants, fused embryos were cultured with 0.1 p.g/ ml aphidicolin for 30 mm after the first pulse, to prevent DNA synthesis prior to chromosome condensation [1]. Embryos allowed to develop in vitro were incubated in 7.5 p.g/ml cytochalasin B for 1 h after the last stimulation [16]. In all cases, time zero was considered to be the time of cell fusion, which occurs about 15 mm after the first pulse. Spnchronization Controls A control was established to determine whether colcemid or aphidicolin treatments had an effect on the morphology of PCC in Gi transplants. Donor embryos were synchronized in Gi without colcemid, using two different procedures. (1) Isolated late 16-cell-stage blastomeres were cultured in vitro, and cleavage to the 32-cell-stage was monitored every 15 mm starting 60 h after mating. Elongated (cleaving) blastomeres were manipulated in the presence of 0.1 p.g/ml aphidicolin until complete fusion to the recipient oocyte. (2) To test the effect of aphidicolin, a portion of cleaving 16-cell-stage blastomeres were slowed in development by cooling at 4#{176}C in PBS for 2-3 h, until micromanipulation and fusion as previously described [7]. The maximum interval between cold treatment and complete blastomere fusion was 30 mm in this experiment. Chromatin and microtubule immunolocalization and chromosome analyses were performed as described below. The two synchronization procedures were directly compared, and two replicates were carried out. Localization of Chromatin and Microtubules Chromatin and microtubules were detected by immunofluorescence in whole mounts of nuclear transplant embryos. Immunofluorescence was performed with mature oocytes 18 h post-hcg and with Gi, early 5, and late S transplants 2 h after nuclear transplantation, once PCC had occurred [1]. Oocytes and embryos were permeabilized for 1 h at 37#{176}C in microtubule-stabilization buffer [19] with 2.5%
 diluted 1:100 in PBS containing 0.1% Tween-20 (PBS-T).")
3 CHROMOSOME CONDENSATION IN NUCLEAR TRANSPLANT EMBRYOS 503 Triton X-100 and fixed for 30 mm in 2% paraformaldehyde. After six washes in PBS and a 10-mm treatment with 0.1 M glycmne, embryos were incubated overnight at 4#{176}C with antia tubulin monoclonal antibody (Sigma) diluted 1:100 in PBS containing 0.1% Tween-20 (PBS-T). As a control for specificity of the anti-a tubulin, monoclonal antibody oocytes and embryos were treated with the same concentration of affinity-purified mouse IgG (Sigma). After several washes in PBS-T, oocytes and embryos were again incubated overnight at 4#{176}C in fluorescein isothiocyanate (FITC)- conjugated affinity-purified goat anti-mouse IgG (Sigma) diluted 1:20 in PBS-T. After an overnight wash at 4#{176}C with PBS-T, specimens were mounted in glycerol containing 0.5 p.g/ml Hoechst (Sigma). Coverslips were sealed to glass slides with nail polish, and slides were stored in the dark at 4#{176}C. Three replicates were performed with each twe of donor nucleus. In all cases, statistical comparisons were made by means of chi-square analysis [20]. Chromosome Spreads Chromosome spreads were prepared at the same time as whole mounts in each replicate. Eighteen-hour post-hcg mature oocytes and G 1, early 5, and late S transplants 2 h after nuclear transplantation were used; and spreads were prepared on acid-washed coverslips according to Matsuda and Tobari [21]. After alcohol dehydration, coverslips were allowed to air-dry and were mounted in anti-fade solution containing 2.3% of 1,4-diazabicyclo(2,2,2)octane (DAPCO, Sigma) [22; unpublished results] and 0.2 p.g/ml propidium iodide (Sigma). As for whole mounts, coverslips were sealed to a glass slide and stored at 4#{176}C in the dark. Three replicates were performed with each type of nuclear donor. Chromosome spreads were also prepared from nonmanipulated in vivo-fertilized 16-cell-stage embryos and Gi (8-16-cell stage), early S (8-16-cell stage), and late S (8- cell stage) transplants (n = 3, 2, and 4, respectively). Embryos were arrested in metaphase (M phase) with 0.5 p.g/ ml colcemid and 10 p.g/ml ethidium bromide (Sigma) for 2-5 h. The zona pellucida and the mucin coat, when present, were removed [18], and embryos were processed for chromosome spreads as described. In all cases, comparisons were made by means of chi-square analysis. Microscopy Whole mounts and chromosome spreads were examined at 100 x under a Zeiss Axioskop microscope equipped with epifluorescence. Hoechst, propidium iodide, and FITC signals were detected using 365/400, 546/590, and 450/ 520 nm excitation/barrier filter combinations, respectively. An SIT 66 video camera (DAGE MTI, Michigan City, IN) fixed to the microscope was used to capture images that were digitized using the Image 1 AT software (Universal Imaging, Media, PA). Images were stored in 44-mb disk cartridges, and photomicrographs were prepared with a video printer (Sony, model UP 5000, Park Ridge, NJ). RESULTS Chromatin and Microtubule Organization in Whole Mounts Two Hours after Nuclear Transfer Unfertilized oocytes. Mature oocytes 18 h post-hcg showed, as expected, an M phase plate (12/12 oocytes, 100%; Fig. Ia) at the equator of the meiotic spindle (Fig. ub). The morphology of this spindle differed from that of a typical meiotic barrel-shaped spindle, such as that in oocytes 14 h post-hcg. This difference in spindle morphology may be due to aging of the oocyte [C. Pinto-Correia, P. Collas, and J.M. RobI, unpublished results]. The spindle was consistently located in the cortex of the oocyte, parallel to the plasma membrane. Gi transplants. PCC of donor nuclei in the Gi phase was manifested by the formation of an M phase plate in 16/19 embryos (84%; Fig. 2, a and c). In some instances (44%, 7/16), the plate was perfectly regular (Fig. 2a); however, in most cases (9/16, 56%), it showed a few isolated chromosomes or, possibly, chromosome fragments in its vicinity (Fig. 2c). Chromosomes were consistently aligned at the equator of spindles. These spindles were morphologically similar to those of 18-h post-hcg oocytes (Fig. ib) and to mitotic spindles (Fig. le). The spindles were located in the oocyte cortex, either parallel or perpendicular to the plasma membrane. In 3/19 embryos (i6%), PCC were scattered in the cytoplasm with no visible microtubules (Fig. 2e, f). Early S transplants. PCC in the early S phase was manifested by the formation of M phase plates in 10/11 embryos (91%; Fig. 2, i, k, and m). This proportion did not differ from that in Gi transplants (p > 0.1). One or two M phase plates were formed (7/ui embryos, 64%, and 3/11 embryos, 36%, respectively), and each was associated with a spindle (Fig. 2, j, 1, and n). Interestingly, when two spindles were formed, their width and length were half those of single spindles. Single metaphase plates were less organized than those of Gi transplants, and all single plates were disrupted. In addition, spindles surrounding these plates showed microtubules radiating from the poles and from the sides of the spindle into the cytoplasm (Fig. 2, and 1). One embryo had no spindle and contained only dispersed chromatin (not shown). Late S transplants. PCC in the late S phase was characterized in 7/13 embryos (54%) by the formation of a very irregular, large, and thick M phase plate, and 85% of these plates (6/7) were surrounded by isolated chromosomes and/ or chromosome fragments. The M phase plates were located at the equator of a spindle with microtubules radiating from the poles and/or from the sides. This configuration was very similar to that found in early S transplants. The morphology of these spindles was similar to that of early S transplants. In 3/i3 embryos (23%), two distinct masses of chromatmn were located outside a small spindle,
 and by indirect immunofluorescence with anti-tubulin antibody (b, e).")
 showed unorganized chromatin with corresponding half spindles (Fig.")
.")
.")
. Therefore, morphology of PCC in Gi transplants was not caused by colcemid or aphidicolin treatments themselves.")

4 504 COLLAS ET AL. FIG. 1. Meiotic apparatus in metaphase II rabbit oocyte 18 h post-hcg (a-c) and mitotic apparatus in pronuclear embryo id-f) (whole mounts). Oocytes and embryos were double-stained with the DNA-specific Hoechst (a, d) and by indirect immunofluorescence with anti-tubulin antibody (b, e). The M phase plate occupies the equator the meiotic spindle (arrows). (c, f) lgg control. Bar, 10 gm. adjacent to the poles (Fig. 2, q and r). In addition, 2/13 embryos (15%) showed unorganized chromatin with corresponding half spindles (Fig. 2, s and t) and, in one embryo, three M phase plates were formed at the equators of three small spindles (Fig. 2, u and v). Despite the discrepancies in configuration, the overall frequency of spindle and plate formation in late S transplants was not different from that of Gi and early S transplants (8/13 embryos,p > 0.1). Controls. Omission of colcemid and aphidicolin for blastomere synchronization did not alter the morphology of PCC in G 1 transplants. Synchronization without colcemid but with aphidicolmn resulted in embryos with PCC organized in an M phase plate at the equator of an intact spindle (7/7 embryos; Fig. 3, a and b). Similarly, when blastomeres were synchronized in Gi without colcemid or aphidicolin (cold treatment), M phase plates were formed at the equators of spindles (5/6 embryos). One embryo was already in anaphase (Fig. 3, d and e). Therefore, morphology of PCC in Gi transplants was not caused by colcemid or aphidicolin treatments themselves. Rather, it reflected the stage of the cell cycle of the donor blastomere at the time of fusion. In addition, we examined whether the structures observed in manipulated embryos were due to the introduction of the exogenous nucleus and not to fusion or activation stimuli themselves. An anucleate cytoplasmic bleb was fused to an enucleated oocyte. As expected, no DNA label was detected (4/4 embryos; Fig. 3g). Furthermore, there was no detectable microtubule organization, and the pool of unpolymerized tubulin was scattered in the entire oocyte cytoplasm (Fig. 3h). This confirms, therefore, that M phase structures and spindles form only in the presence of the exogenous nucleus. Chromosome Spreads from Embryos Tu o Hours after Nuclear Transplantation. Unfertilized oocvtes. Chromosome spreads of nonmanipulated 18-h post-hcg oocytes showed haploid sets of 22 chromosomes, each with 2 distinct chromatids (17/17 oocytes, 100%; Fig. 4a). Gi transplants. PCC in GI revealed two types of configuration. In 8/12 transplants (67%), chromosomes were formed but were not fully individualized (Fig. 4b). The remaining embryos (33%) displayed individualized chromosomes (Fig. 4c) together with unorganized chromatmn (Fig. 4c). In both cases, some chromosomes seemed paired (Fig. 4, b and c). In addition, these chromosomes were shorter and less defined than typical mitotic chromosomes (see Fig. 5a) and displayed a meiotic configuration, similar to that of oocyte chromosomes (Fig. 4a). Early S transplants. PCC in the early S phase resulted in the formation of chromosomes that were not individualized (3/5 embryos, 60%; Fig. 4d). Alternatively, no chromosomes were formed, and only patches of unorganized chromatmn were present (Fig. 4e). Late S transplants. PCC in late S phase was unorganized and never formed chromosomes. Rather, the chromatin remained in 1, 2 or 4 patches (5/1 0, 4/10, and 1/10 transplants, respectively; Fig. 4, f and g).



.")

.")


.")

; (c, dl")

;")


 metaphase plate and")
;")


")




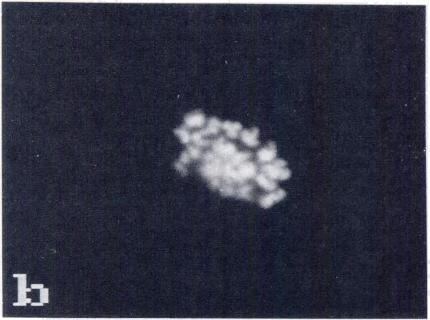
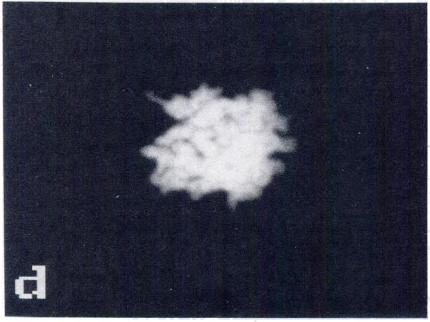
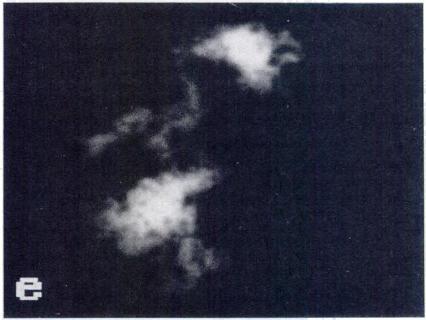
5 CHROMOSOME CONDENSATION IN NUCLEAR TRANSPLANT EMBRYOS 505 FIG. 2. PCC and newly formed spindle after transfer of nuclei in Gi, early S and late S phases (whole mounts). DNA was stained with Hoechst (Gi: a, c, e, g; early S: i, k, m, o; late S: q, s, u, w). Microtubules were stained with anti-tubulin antibody (Gi: b, d, f; early S: i. I, n; late S: r, t, v). Control embryos (h, p. x). Gi transplants: (a, b) metaphase plate formed at the equator of a spindle (arrows); (c, dl metaphase plate with isolated chromosomes or, possibly, chromosome fragments (arrowheads); (e, f) chromosomes dispersed in the oocyte without a spindle; (g, h) lgg controls. Early S transplants: (i, i) metaphase plate and spindle with radiating microtubules (arrows); (k, I) disrupted metaphase plate, and spindle with radiating microtubules (arrows); (m, n) two small metaphase plates formed at the equator of two distinct spindles; (0, p) IgG controls. Late S transplants: (q, r) two sets of chromatin at the poles of a small spindle (arrows); is, t) unorganized chromatin within an uneven number of half spindles; (u, v) three small metaphase plates formed at the equator of three spindles; (w, x) lgg controls. Bar, 10 p.m.
 or cold treatment id-f).")
 Whole mounts with chromosomes already in anaphase along a normal spindle; if) smear with not fully individualized chromosomes.")
. Bar, 10 p.m. Synchronization controls.")
.")
 revealed normal chromosomes (Fig.")


 indicated that chromosomes were normal (7/7 M phase spreads; 100%; Fig. Sb). FIG. 4. Chromosome spreads from a mature oocyte, and PCC in Gi, early S, and late S transplants.")
6 506 COLIAS ET AL. FIG. 3. PCC and spindle in synchronization and anucleate control manipulated embryos. Prior to nuclear transfer, non-colcemid-treated donor biastomeres were synchronized in Gi with aphidicolin (a-c) or cold treatment id-f). (a, b) Whole mounts showing a metaphase plate at the equator of an intact spindle; (c) smear with individualized chromosomes. (d, e) Whole mounts with chromosomes already in anaphase along a normal spindle; if) smear with not fully individualized chromosomes. (g, h) When an anucleate cytoplasmic fragment was transferred into a nenucleated oocyte, no chromatin was present (g), and only the unpolymerized pool of tubulin was detected without any signs of spindle formation (h). Bar, 10 p.m. Synchronization controls. Synchronization in the Gi phase without colcemid but with aphidicolin resulted in spreads with individualized chromosomes (5/5 embryos; Fig. 4c). Synchronization in the Gi phase by cold treatment resulted in spreads with chromosomes not fully individualized (5/5 embryos; Fig. 4f). These results indicate that use of colcemid and aphidicolin for blastomere synchronization did not affect chromosome configuration in Gi transplants. Early S transplants. Two 8-16-cell stage embryos were processed. Fourteen M phase spreads were obtained; 12 (86%) revealed normal chromosomes (Fig. 5c), and 2 blastomeres contained a total of 7 chromosomes and chromosome fragments (2/14, 14%, Fig. 5d). Chromosome morphology did not differ from that of G1 transplants or nonmanipulated embryos. Chromatmn fragments were structures irregular in shape and size and smaller than in- Chromosome Spreads of Cleavage Stage Nuclear Transplant Embryos GI transplants. Three manipulated embryos were successfully prepared at the 8-16-cell stage, and 7 M phase spreads were obtained. Chromosome counts (n = 44) and morphological comparisons with chromosome spreads from nonmanipulated control embryos (Fig. Sa) indicated that chromosomes were normal (7/7 M phase spreads; 100%; Fig. Sb). FIG. 4. Chromosome spreads from a mature oocyte, and PCC in Gi, early S, and late S transplants. (a) Haploid set of 22 bivalent chromosomes in a metaphase II rabbit oocyte. Note the short and condensed morphology, characteristic of meiotic chromosomes (compare with mitotic chromosomes in Fig. 5a). In Gi transplants, the donor chromatin (b) forms plates with chromosomes not fully individualized or (c) condenses into chromosomes with two chromatids (arrowheads), chromosomes with one chromatid (short arrow), and random clusters (long arrow). (dl In early S transplants, the chromatin is either organized into aggregated chromosomes, or (e) displays an unorganized conformation. (f, g) In late S transplants, chromosomes are never formed, and the chromatin condenses into one or two clusters. Bar, 10 p.m.




7 CHROMOSOME CONDENSATION IN NUCLEAR TRANSPLANT EMBRYOS 507

8 508 COLLAS ET AL. tact chromosomes. In addition, one blastomere contained 2 interphase micronuclei (Fig. Sd). Micronuclei consisted of round entities smaller than an intact nucleus. They probably resulted from the formation of a nuclear envelope around a single chromosome or small cluster of chromosomes [23]. Therefore, although each embryo contained cells with normal chromosomes, some defects also occurred in other blastomeres. Late S transplants. Four 8-cell-stage embryos were examined, and 20 M phase spreads were obtained. In 15 of these spreads (75%), chromosomes tended to be larger than those of GI transplants, early S transplants, and nonmanipulated embryos (Fig. Se). Width and length of chromosomes 7 and 18 were measured in one blastomere of nonmanipulated 8-cell- and 64-cell-stage embryos, as well as of Gi (8-16-cell), early S (8-16-cell), and late S (8-cell) transplants (Table 1). Chromosomes 7 and 18 were chosen because they were easily identifiable owing to their morphology. Since only 2 chromosomes were measured in each type of embryo, the data indicate only a trend, and statistical comparisons were not made. However, this phenomenon was consistently observed in these 15 chromosome spreads. The fact that chromosome size was similar in 8- and 64-cell-stage controls indicates that the increase in chromosome size in late S transplants was not due to a difference in embryo cell stage at the time chromosome spreads were made; rather, it reveals a peculiarity of late S transplants. The increase in width/length (W/L) ratio in late S transplants indicates that an increase in size is due more to an increase in chromosome width than in length. In addition, 4 chromosome spreads revealed extremely damaged chromatin (4/20; 20%; Fig. 5h). It is likely that these cells were already degenerated when embryos were fixed. Furthermore, one spread displayed paired chromosomes, as in diplo-chromosomes (1/20, 5%; Fig. 5g). In addition to the 20 M phase spreads, over 300 interphase micronuclei were detected in a total of 8-10 cells (Fig. Sf). In contrast, micronuclei were not detected in nonmanipulated embryos cultured in vitro. A characteristic of late S transplants, therefore, was that each embryo contained blastomeres with large chromosomes, and a high proportion of cells with abnormalities. DISCUSSION In this study, we investigated the effect of donor cell cycle stage on morphology of PCC and spindle and on chromosome structure in nuclear transplant embryos. The results indicate that after nuclear transfer, the recipient oocyte cytoplasm was typically able to induce the formation de novo of an M phase plate associated with a spindle. M phase plates and spindles were intact in most cases of PCC in GI transplants. However, their integrity was lost in most early S transplants, and gross abnormalities were detected in late S transplants. Furthermore, normal chromosomes were obtained in Gi and most of early S transplants, whereas a large proportion of chromosome abnormalities was present in late S transplants. M Phase Plate and Spindle in Nuclear Transplant Embryos Two Hours after Nuclear Transfer Interactions between donor chromatin and recipient oocyte microtubules may determine spindle and M phase plate conformation. Formation of a spindle in Gi, early 5, and, to some extent, late S transplants suggests that initiation of microtubule nucleation in the oocyte is the result of a signal common to donor blastomeres in G1 and S phases. Nevertheless, a normal spindle can be associated with an intact or an abnormal M phase plate (Gi transplants). Alternatively, an abnormal spindle is always associated with an abnormal M phase plate (early and late S transplants). These observations suggest that morphology of the new spindle influences integrity of the M phase plate. It is not excluded, however, that the condensed chromatin also sends specific instructions to oocyte microtubules; these instructions may differ with the cell cycle stage at which the chromatmn is condensed. Thus, it is possible that only Gi donor chromatin sends correct instructions for spindle assembly, whereas altered information from chromatin condensed in the S phase may result in abnormal or incomplete spindle formation. Alternatively, final spindle assembly may depend simply on the physical conformation of the condensed chromatin. It is suggested, therefore, that a first set of instructions, nonspecific for the stage of the donor blastomere in the cell cycle, triggers the nucleation of microtubules. Then, physical configuration of the condensed chromatin or a signal specific for the cell cycle stage of the condensed chromatmn may determine the extent of final spindle assembly. The comparison between M phase and spindle morphology in whole mounts and chromatin configuration in chromosome spreads 2 h after nuclear transfer supports this hypothesis. It is likely that M phase spreads showing chromosomes in Gi and early S transplants (Fig. 4, b and d) may correspond to the M phase plates found in whole mounts (Fig. 2, a, c, i, and k). Furthermore, in both Gi and early S transplants, isolated chromosomes or chromosome fragments present in whole mounts (Fig. 2, FIG. 5. Chromosome spreads in cleavage stage G1, early S, and late S transplants. DNA was double-stained with Hoechst and propidium iodide. (a) M phase chromosomes from a nonmanipulated embryo. (b) Chromosomes from a Gi transplant; no morphological differences with (a) were detected. (c) Within early S transplants, some blastomeres show normal chromosomes, and (d) others reveal abnormalities, such as a micronucleus (arrow) coexisting with chromosomes (arrowheads), or, possibly, chromosome fragments. (e-h) Late S phase transplants containing (e) M phase chromosomes, larger than in controls, Gi transplants, or early S transplants; if) a micronucleus (arrow) next to a normal interphase nucleus; (g) paired chromosomes; and (h) abnormal chromatin. The proportion of abnormalities within embryos was increased between early S and late S transplants. Bar, 10 p.m.








9 CHROMOSOME CONDENSATION IN NUCLEAR TRANSPLANT EMBRYOS 509
 Width Length W/L WxL Width Length W/L WxL Ctrl 8c 1.93 2.38 0.81 4.59 2.68 2.38 1.13 6.39 Ctrl 64c 2.07 2.67 0.76 5.53 2.68 2.22 1.21 5.95 G1 trans 2.05 2.57 0.82 5.27 2.43 2.00 1.25 4.")
10 510 COLIAS ET AL. TABLE 1. Chromosome size in cleavage stage Gi, early S (ES), and late S (LS) transplants. Embryo Typeb Big submetacentric (Chrom. 71 Big acroc (Chrom. entric 18) Width Length W/L WxL Width Length W/L WxL Ctrl 8c Ctrl 64c G1 trans ES trans LS trans Chromosomes 7 and 18 were chosen because they were easily identifiable due to their morphology. Width and length, p.m; W x L, width x length product, p.m2; W/L, width/length ratio. 6Ctrl 8c and Mc: nonmanipulated 8-cell- and 64-cell-stage embryos, respectively; one embryo was examined in each group. c, i, and k) were not found in chromosome spreads (Fig. 4, h, and d). It is possible that the were lost during chromosome spread preparation. In addition, absence of M phase plate formation revealed in whole mounts ma\ he manifested in chromosome spreads with non-uniformly condensed chromatin (Fig. 4, c, e, f, and g). Also, the multiple M phase plates in earl and late S transplants (Fig. 2, m and u) ma have been manifested h multiple chromatin patches in chromosome spreads (Fig. 4, e and g). These obseryations suggest that chromosomes, as opposed to non-uniform chroniatin, may he present in the M phase plates of GI and early S transplants. However, defects in spindle formation may he related to the non-chromosome structures observed in M phase spreads. l remature Condensation of Gi and S Chronzatin in Nuclear Transplant Embryos Gi chromatin condenses into chromosomes in nuclear transplants; however, the pattern of PCC differs between somatic cell h brids and nuclear transplants. First, PCC was found not to he accompanied by formation of a spindle in somatic cell hybrids [8]. This may reflect a difference in recipient cell type, as discussed below. Second, in contrast to findings in somatic cell hybrids [8, 11], not all chromosomes were fully individualized in nuclear transplants. Because presumably the nuclear envelope breaks down after blastomere fusion, remnants of nuclear matrix not completelv extracted at permeahilization ma have held the chromosomes together. Third, chromosomes in GI transplants were extremely short and condensed and showed a meiotic configuration. Ln contrast, prematurely condensed Gi chromosomes in somatic hybrids have been found to he abnormally elongated when compared to control meiotic chromosomes [8, 11]. Again, a difference in recipient cell wpe may he responsible for this phenomenon (see below). Fourth, a proportion of chromosomes in Gi transplants revealed a hivalent structure, similar to that of metaphase II chromosomes. The fact that Gi chromosomes are normally nionoyalent suggests that some chromosome pairing occurred. Whether pairing occurred between homologous chromosomes, however, was not investigated. PCC in S phase also displays morphological differences in somatic cell hybrids and nuclear transplants. As with Gi cells, no evidence of spindle formation has been reported after FCC in S phase in somatic hybrids [8]. Furthermore, PCC in S phase results in extensive chromatmn fragmentation in somatic hybrids [8, 1 1 ]. Fragmentation was not observed in rabbit nuclear transplants; however, as in somatic cell hybrids [8, 1 1 ], chromosomes were never formed with late S chromatin. The reason for condensation into a nonchromosome structure may be similar in both studies. Replication forks in S phase DNA create a discontinuity and, presumably, weak points in the chromatin due to absence of base pairing. This condition may be responsihle for nonuniform condensation, s ielding the structures observed. As with PCC in somatic cells, our findings support the conclusion that the ability of the donor nucleus to participate fulls iii nietaphase plate formation in the oocvte decreases as donor blastomeres progress in the cell cycle. Several factors may account for differences in the morphology of FCC in somatic cells and nuclear transplants. Differentiated cells have previously been used in somatic cell fusion experiments [6-9], whereas embryonic blastomeres were used in our study. Therefore, the differentiated state of the interphase nucleus may have affected FCC. In a separate experiment, however, differentiated granulosa cell nuclei fused to enucleated oocvtes also condensed into a metaphase plate within a spindle [unpublished observation]. This finding, therefore, rules out the above hpothesis. Another factor that may influence the spindle and the chromatin in nuclear transplants is the absence of the meiotic spindle in recipient oocytes. Since the metaphase plate no longer has to he maintained, the tubulin pool is entirely available for the establishment of a new spindle. Further studies indicate, however, that metaphase plate and spindle form de novo even in non-enucleated oocvtes [unpublished observation]. Thus, it is unlikely that absence of the meiotic spindle per se allowed spindle formation in nuclear transplants. The most significant factor affecting FCC is probably the pe of M phase recipient cell used. Mitotic cells were employed in previous cell fusions [8-11], whereas a meiotic cell (oocyte) was used for nuclear transfer. It is possible that meiotic-specific factors present in the oocte ma\ be responsible for PCC into a metaphase plate, rneioticlike configuration of chromosomes, and chromosome pairing in nuclear transplants. These factors max or may not be associated with a maturation-promoting factor. Mature oocytes have also been used as recipient cells in mouse thymocvte nuclear transplants [2-6]; however, absence of control of the cell cle stage of donor thvmocvtes makes comparisons difficult. The data suggest, therefore, that the mature oocvte is a privileged environment for condensanon of the chromatin into a metaphase plate and a spindle.

11 CHROMOSOME CONDENSATION IN NUCLEAR TRANSPLANT EMBRYOS 511 Chromosome Constitution in Cleavage Stage Nuclear Transplant Embryos The morphology of the metaphase plate upon PCC in nuclear transplants does not, to some extent, affect chromosome structure and number and early embryo development In particular, formation of an intact metaphase plate upon FCC is compatible with normal chromosome complement (Gi transplants). Similarly, a less defined or disrupted metaphase plate inside an abnormal spindle does not alter subsequent chromosome structure in Gi and early S transplants. Furthermore, Gi and early S transplants developed to blastocysts at high and similar rates in a separate study (71% and 59%, respectively; p > 0.1) [7]. These results suggest that formation of a metaphase plate and a spindle upon PCC is compatible with normal preimplantation development of manipulated embryos. These indications do not imply, however, that all embryos with intact chromosomes develop to blastocysts. In addition, a small proportion of Gi and early S transplants do not activate or develop beyond 2 or 3 cleavages [7]. The data suggest that these embryos may be those in which the chromatin condenses into a non-uniform mesh with no spindle. Furthermore, FCC of late S chromatin is also compatible with subsequent formation of chromosomes in cleavage stage embryos. These chromosomes, however, are larger than those of Gi and early S transplants. Also, a low percentage of these embryos develop to blastocysts (15% [7]). FCC in late 5, therefore, is detrimental, and blastomeres containing abnormally thick chromosomes are likely to be nonviable. Reasons for this defect remain to be determined; they may entail reduced chromosome compaction at M phase. These observations indicate that the ability of the donor nucleus to direct development in vitro of nuclear transplant embryos does not depend solely on the morphology of FCC and, therefore, on the cell cycle stage of the donor nucleus. Other defects that may not be related to PCC morphology may arise during embryo development. Absence of cell division immediately after FCC in nuclear transplants may account for the normal chromosome structure in cleavage stage embryos. Completion of mitosis after FCC in somatic cell hybrids has been shown to be responsible for the deleterious effect of PCC in such systems [12-15]. In nuclear transplant embryos, however, the condensed chromatin decondenses into a pronucleus upon activation of the oocyte [1]. It is possible, therefore, that the chromatin may be reorganized in the interphase pronucleus. However, this does not exclude the possibility that chromosomes or chromosome fragments may be lost during this process. Alternatively, chromosome abnormalities that would not be detected in chromosome spreads may arise. These defects may, however, remain compatible with development of nuclear transplant embryos in vitro. ratorv. ACKNOWLEDGMENT The authors are grateful to John Balise for his technical expertise in the laho. REFERENCES 1. Collas P. RobI JM. Relationship between nuclear remodeling and development in nuclear transplant rabbit embryos. Biol Reprod 1991; 45: Czolowska R, Modlinski JA, Tarkowski AK. Behaviour of thymoevte nuclei in nonactivated and activated mouse oocvtes. J Cell Sci ]984; 69: Balakier H, Masui Y. Chromosome condensation activity in the cytoplasm of anucleate and nucleate fragments of mouse oocvte. Dev Biol 1986; 113: Czolowska R, Waksmundzka M, Kubiak JZ, Tarkowski AK. Chromosome conden. sation activity in ovulated metaphase II mouse oocvtes assayed by fusion with interphase blastomeres. J Cell Sci 1986; 84: Szollosi D, Czolowska R, Soltynska M5, Tarkowski AK. Remodelling of thymocvte nuclei in activated mouse oocytes: an ultrastructural study. Europ J Cell Biol 1986; 42: Szollosi D, Czolowska R, Szollosi MS, Tarkoki AK. Remodeling of mouse thymocyte nuclei depends on the time of their transfer into activated, homologous oocvtes.j Cell Sci 1988; 91: Collas P. Balise B RobI JM. Influence of donor cell cycle stage on development of nuclear transplant rabbit embryos. Biol Reprod 1992; 46: Johnson RI, Rao PN. Mammalian cell fusion: induction of premature chromosome condensation in interphase nuclei. Nature 1970; 226: Johnson RT, Rao PN, Hughes HD. Mammalian cell fusion. III. A HeLa cell inducer of premature chromosome condensation active in cells from a variety of animal species. J Cell Physiol 1970; 76: Rao PN, Johnson RI. Mammalian cell fusion: studies on the regulation of DNA synthesis and mitosis. Nature 1970; 225: Johnson RT, Mullinger AM. The induction of DNA synthesis in the chick red cell nucleus in heterokarvons during the first cell cycle after fusion with HeLa cells. Cell Sci 1975; 18: Schwartz AG, Cook PR, Harris H. Correction of a genetic defect in a mammalian cell. Nature New Biol 1971; 230: Boyd YL, Harris H. Correction of genetic defects in mammalian cells by the input of small amounts of foreign genetic material. J Cell Sci 1973; 13: Kao FT. Identification of chick chromosomes in cell hybrids formed between chick ei-vthrocvtes and adenine requiring mutants of Chinese hamster cells. Proc Nat Acad Sci USA 1973; 70: Klinger u, Shin SI. Modulation of the activity of an avian gene transferred into a mammalian cell by cell fusion. Proc Nat Acad Sci USA 1974; 71: Kenelly JJ, Foote RH. Superovulatorv response of pre- and post.puberal rabbits to commercially available gonadotropins. J Reprod Fertil 1965; 9: Stice SL, RobI JM. Nuclear reprogramming in nuclear transplant rabbit embryos. Biol Reprod 1988; 39: Collas F, RobI JM. Factors affecting the efficiency of nuclear transplantation in the rabbit embro. Biol Reprod 1990; 43: Schatten GC, Simerly C, Schatten H. Microtubule configuration during fertilization. mitosis, and early development in the mouse and the requirement for egg microtubule-mediated motility during mammalian fertilization. Proc Nat Acad Sci USA 1985; 82: , Snedecor GW, Cochran WG. Sampleing from the binomial distribution. In: Statistical Methods, 6th ed. Ames, IA: Iowa State University Press; 1980: Matsuda Y, Tobari I. Chromosomal analysis in mouse eggs fertilized in vitro with sperm exposed to ultraviolet light (UV) and methyl and ethyl methanesulfonate (MMS and EMS). Muta Res 1988; 198: Lichter P. Tang CC. Call K, Hermanson G, Evans GA, Housman D, Ward DC. High resolution mapping of human chromosome 11 by in situ hybridization with cos mid clones. Science 1991; 247: McNeill CA, Brown RL. Genetic manipulation by means of microcell-mediated transfer of normal human chromosomes into recipient mouse cells. Proc Nat Acad Sci USA 1980; 77:
Cell cycle co-ordination in embryo cloning by nuclear transfer
 Reviews of Reproduction (1996) 1, 40 46 Cell cycle co-ordination in embryo cloning by nuclear transfer Keith H. S. Campbell, Pasqualino Loi*, Pedro J. Otaegui and Ian Wilmut Roslin Institute (Edinburgh),
Reviews of Reproduction (1996) 1, 40 46 Cell cycle co-ordination in embryo cloning by nuclear transfer Keith H. S. Campbell, Pasqualino Loi*, Pedro J. Otaegui and Ian Wilmut Roslin Institute (Edinburgh),
CELL CYCLE INTRODUCTION PART I ANIMAL CELL CYCLE INTERPHASE
 CELL CYCLE INTRODUCTION The nuclei in cells of eukaryotic organisms contain chromosomes with clusters of genes, discrete units of hereditary information consisting of double-stranded DNA. Structural proteins
CELL CYCLE INTRODUCTION The nuclei in cells of eukaryotic organisms contain chromosomes with clusters of genes, discrete units of hereditary information consisting of double-stranded DNA. Structural proteins
Chapter 8: Cellular Reproduction
 Chapter 8: Cellular Reproduction 1. The Cell Cycle 2. Mitosis 3. Meiosis 2 Types of Cell Division 2n 1n Mitosis: occurs in somatic cells (almost all cells of the body) generates cells identical to original
Chapter 8: Cellular Reproduction 1. The Cell Cycle 2. Mitosis 3. Meiosis 2 Types of Cell Division 2n 1n Mitosis: occurs in somatic cells (almost all cells of the body) generates cells identical to original
Preimplantation genetic diagnosis: polar body and embryo biopsy
 Human Reproduction, Vol. 15, (Suppl. 4), pp. 69-75, 2000 Preimplantation genetic diagnosis: polar body and embryo biopsy Luca Gianaroli SISMER, Via Mazzini 12, 40138 Bologna, Italy Scientific Director
Human Reproduction, Vol. 15, (Suppl. 4), pp. 69-75, 2000 Preimplantation genetic diagnosis: polar body and embryo biopsy Luca Gianaroli SISMER, Via Mazzini 12, 40138 Bologna, Italy Scientific Director
The Cell Cycle CHAPTER 12
 The Cell Cycle CHAPTER 12 The Key Roles of Cell Division cell division = reproduction of cells All cells come from pre-exisiting cells Omnis cellula e cellula Unicellular organisms division of 1 cell reproduces
The Cell Cycle CHAPTER 12 The Key Roles of Cell Division cell division = reproduction of cells All cells come from pre-exisiting cells Omnis cellula e cellula Unicellular organisms division of 1 cell reproduces
The Cellular Basis of Reproduction and Inheritance
 Chapter 8 The Cellular Basis of Reproduction and Inheritance PowerPoint Lectures for! Biology: Concepts and Connections, Fifth Edition! Campbell, Reece, Taylor, and Simon Lectures by Chris Romero Objective:
Chapter 8 The Cellular Basis of Reproduction and Inheritance PowerPoint Lectures for! Biology: Concepts and Connections, Fifth Edition! Campbell, Reece, Taylor, and Simon Lectures by Chris Romero Objective:
Bacterial cell. Origin of replication. Septum
 Bacterial cell Bacterial chromosome: Double-stranded DNA Origin of replication Septum 1 2 3 Chromosome Rosettes of Chromatin Loops Scaffold protein Chromatin Loop Solenoid Scaffold protein Chromatin loop
Bacterial cell Bacterial chromosome: Double-stranded DNA Origin of replication Septum 1 2 3 Chromosome Rosettes of Chromatin Loops Scaffold protein Chromatin Loop Solenoid Scaffold protein Chromatin loop
Ploidy and Human Cell Types. Cell Cycle and Mitosis. DNA and Chromosomes. Where It All Began 11/19/2014. Chapter 12 Pg
 Ploidy and Human Cell Types Cell Cycle and Mitosis Chapter 12 Pg. 228 245 Cell Types Somatic cells (body cells) have 46 chromosomes, which is the diploid chromosome number. A diploid cell is a cell with
Ploidy and Human Cell Types Cell Cycle and Mitosis Chapter 12 Pg. 228 245 Cell Types Somatic cells (body cells) have 46 chromosomes, which is the diploid chromosome number. A diploid cell is a cell with
Mitosis THE CELL CYCLE. In unicellular organisms, division of one cell reproduces the entire organism Multicellular organisms use cell division for..
 Mitosis THE CELL CYCLE In unicellular organisms, division of one cell reproduces the entire organism Multicellular organisms use cell division for.. Development from a fertilized cell Growth Repair Cell
Mitosis THE CELL CYCLE In unicellular organisms, division of one cell reproduces the entire organism Multicellular organisms use cell division for.. Development from a fertilized cell Growth Repair Cell
Chapter 2. Mitosis and Meiosis
 Chapter 2. Mitosis and Meiosis Chromosome Theory of Heredity What structures within cells correspond to genes? The development of genetics took a major step forward by accepting the notion that the genes
Chapter 2. Mitosis and Meiosis Chromosome Theory of Heredity What structures within cells correspond to genes? The development of genetics took a major step forward by accepting the notion that the genes
Origin of replication. Septum
 Bacterial cell Bacterial chromosome: Double-stranded DNA Origin of replication Septum 1 2 3 Chromosome Rosettes of Chromatin Loops Chromatin Loop Solenoid Scaffold protein Scaffold protein Chromatin loop
Bacterial cell Bacterial chromosome: Double-stranded DNA Origin of replication Septum 1 2 3 Chromosome Rosettes of Chromatin Loops Chromatin Loop Solenoid Scaffold protein Scaffold protein Chromatin loop
Chapter 10. Cell Cycle - Mitosis
 Chapter 10 Cell Cycle - Mitosis WHAT CELL REPRODUCTION ACCOMPLISHES Cell division plays important roles in the lives of organisms. Cell division replaces damaged or lost cells permits growth allows for
Chapter 10 Cell Cycle - Mitosis WHAT CELL REPRODUCTION ACCOMPLISHES Cell division plays important roles in the lives of organisms. Cell division replaces damaged or lost cells permits growth allows for
Unit 4: Cell Division Guided Notes
 Unit 4: Cell Division Guided Notes 1 Chromosomes are structures that contain material When Eukaryotes are not dividing, DNA and Proteins are in a mass called: When the cell divides, it condenses and becomes
Unit 4: Cell Division Guided Notes 1 Chromosomes are structures that contain material When Eukaryotes are not dividing, DNA and Proteins are in a mass called: When the cell divides, it condenses and becomes
Cell Division. The Process of Cell Division Section Section 10.2: The Process of Cell Division 12/8/2010
 The Process of Cell Division Section 10.2 Biology B Section 10.2: The Process of Cell Division The student will investigate and understand common mechanisms of inheritance and protein synthesis. Key concepts
The Process of Cell Division Section 10.2 Biology B Section 10.2: The Process of Cell Division The student will investigate and understand common mechanisms of inheritance and protein synthesis. Key concepts
Genetics and Cellular Function
 Genetics and Cellular Function DNA replication and the cell cycle Mitosis Mitosis Mitosis: division of cells that results in daughter cells with the same the genetic information that the original cell
Genetics and Cellular Function DNA replication and the cell cycle Mitosis Mitosis Mitosis: division of cells that results in daughter cells with the same the genetic information that the original cell
Cellular Reproduction, Part 1: Mitosis Lecture 10 Fall 2008
 Cell Theory 1 Cellular Reproduction, Part 1: Mitosis Lecture 10 Fall 2008 Cell theory: All organisms are made of cells All cells arise from preexisting cells How do new cells arise? Cell division the reproduction
Cell Theory 1 Cellular Reproduction, Part 1: Mitosis Lecture 10 Fall 2008 Cell theory: All organisms are made of cells All cells arise from preexisting cells How do new cells arise? Cell division the reproduction
Biology 4361 Developmental Biology. October 11, Multiple choice (one point each)
") Biology 4361 Developmental Biology Exam 1 October 11, 2005 Name: ID#: Multiple choice (one point each) 1. Sertoli cells a. surround spermatocytes b. are the structural components of the seminiferous tubules
Biology 4361 Developmental Biology Exam 1 October 11, 2005 Name: ID#: Multiple choice (one point each) 1. Sertoli cells a. surround spermatocytes b. are the structural components of the seminiferous tubules
klp-18 (RNAi) Control. supplementary information. starting strain: AV335 [emb-27(g48); GFP::histone; GFP::tubulin] bleach
![klp-18 (RNAi) Control. supplementary information. starting strain: AV335 [emb-27(g48); GFP::histone; GFP::tubulin] bleach](/thumbs/91/104639484.jpg "klp-18 (RNAi) Control. supplementary information. starting strain: AV335 [emb-27(g48); GFP::histone; GFP::tubulin] bleach") DOI: 10.1038/ncb1891 A. starting strain: AV335 [emb-27(g48); GFP::histone; GFP::tubulin] bleach embryos let hatch overnight transfer to RNAi plates; incubate 5 days at 15 C RNAi food L1 worms adult worms
DOI: 10.1038/ncb1891 A. starting strain: AV335 [emb-27(g48); GFP::histone; GFP::tubulin] bleach embryos let hatch overnight transfer to RNAi plates; incubate 5 days at 15 C RNAi food L1 worms adult worms
Molecular Cell Biology - Problem Drill 22: The Mechanics of Cell Division
 Molecular Cell Biology - Problem Drill 22: The Mechanics of Cell Division Question No. 1 of 10 1. Which of the following statements about mitosis is correct? Question #1 (A) Mitosis involves the dividing
Molecular Cell Biology - Problem Drill 22: The Mechanics of Cell Division Question No. 1 of 10 1. Which of the following statements about mitosis is correct? Question #1 (A) Mitosis involves the dividing
基醫所. The Cell Cycle. Chi-Wu Chiang, Ph.D. IMM, NCKU
 基醫所 The Cell Cycle Chi-Wu Chiang, Ph.D. IMM, NCKU 1 1 Introduction to cell cycle and cell cycle checkpoints 2 2 Cell cycle A cell reproduces by performing an orderly sequence of events in which it duplicates
基醫所 The Cell Cycle Chi-Wu Chiang, Ph.D. IMM, NCKU 1 1 Introduction to cell cycle and cell cycle checkpoints 2 2 Cell cycle A cell reproduces by performing an orderly sequence of events in which it duplicates
Chapter 10 Cell Growth and Division
 Chapter 10 Cell Growth and Division 10 1 Cell Growth 2 Limits to Cell Growth The larger a cell becomes, the more demands the cell places on its DNA. In addition, the cell has more trouble moving enough
Chapter 10 Cell Growth and Division 10 1 Cell Growth 2 Limits to Cell Growth The larger a cell becomes, the more demands the cell places on its DNA. In addition, the cell has more trouble moving enough
Name: Date: Block: 10-2 Cell Division Worksheet
 10-2 Cell Division Worksheet W hat do you think would happen if a cell were simple to split into two, without any advance preparation? Would each daughter cell have everything it needed to survive? Because
10-2 Cell Division Worksheet W hat do you think would happen if a cell were simple to split into two, without any advance preparation? Would each daughter cell have everything it needed to survive? Because
Lesson 1. Quiz (short) Cell cycle Chromosomes Mitosis phases
 Cell cycle Chromosomes Mitosis phases") Lesson 1 Quiz (short) Cell cycle Chromosomes Mitosis phases 2 Cell division is needed for Growth (Mitosis) Repair (Mitosis) Reproduction (Meiosis) 3 Mitosis consists of 4 phases (division of the nuclear
Lesson 1 Quiz (short) Cell cycle Chromosomes Mitosis phases 2 Cell division is needed for Growth (Mitosis) Repair (Mitosis) Reproduction (Meiosis) 3 Mitosis consists of 4 phases (division of the nuclear
Chromosomes and Cell Cycle
 Chromosomes and Cell Cycle Cell Basics There are trillions of cells in your body Cells are microscopic Cells have DNA inside a structure called the nucleus The nucleus is enclosed by a structure called
Chromosomes and Cell Cycle Cell Basics There are trillions of cells in your body Cells are microscopic Cells have DNA inside a structure called the nucleus The nucleus is enclosed by a structure called
Biology is the only subject in which multiplication is the same thing as division
 Biology is the only subject in which multiplication is the same thing as division 2007-2008 The Cell Cycle: Cell Growth, Cell Division Ch. 10 Where it all began You started as a cell smaller than a period
Biology is the only subject in which multiplication is the same thing as division 2007-2008 The Cell Cycle: Cell Growth, Cell Division Ch. 10 Where it all began You started as a cell smaller than a period
Unit 4 Student Notes Cell Cycle
 Name Date Unit 4 Student Notes Cell Cycle B-2.6 Summarize the characteristics of the cell cycle: interphase (called G1, S, G2); the phases of mitosis (called prophase, metaphase, anaphase, and telophase);
Name Date Unit 4 Student Notes Cell Cycle B-2.6 Summarize the characteristics of the cell cycle: interphase (called G1, S, G2); the phases of mitosis (called prophase, metaphase, anaphase, and telophase);
Cell cycle and apoptosis
 Cell cycle and apoptosis Cell cycle Definition Stages and steps Cell cycle Interphase (G1/G0, S, and G2) Mitosis (prophase, metaphase, anaphase, telophase, karyokinesis, cytokinesis) Control checkpoints
Cell cycle and apoptosis Cell cycle Definition Stages and steps Cell cycle Interphase (G1/G0, S, and G2) Mitosis (prophase, metaphase, anaphase, telophase, karyokinesis, cytokinesis) Control checkpoints
BIOLOGY LTF DIAGNOSTIC TEST CELL CYCLE & MITOSIS
 Biology Multiple Choice 016044 BIOLOGY LTF DIAGNOSTIC TEST CELL CYCLE & MITOSIS TEST CODE: 016044 Directions: Each of the questions or incomplete statements below is followed by five suggested answers
Biology Multiple Choice 016044 BIOLOGY LTF DIAGNOSTIC TEST CELL CYCLE & MITOSIS TEST CODE: 016044 Directions: Each of the questions or incomplete statements below is followed by five suggested answers
Unduplicated. Chromosomes. Telophase
 10-2 Cell Division The Cell Cycle Interphase Mitosis Prophase Cytokinesis G 1 S G 2 Chromatin in Parent Nucleus & Daughter Cells Chromatin Daughter Nuclei Telophase Mitotic Anaphase Metaphase Use what
10-2 Cell Division The Cell Cycle Interphase Mitosis Prophase Cytokinesis G 1 S G 2 Chromatin in Parent Nucleus & Daughter Cells Chromatin Daughter Nuclei Telophase Mitotic Anaphase Metaphase Use what
A comparison of the effects of estrus cow. nuclear maturation of bovine oocytes
 A comparison of the effects of estrus cow serum and fetal calf serum on in vitro nuclear maturation of bovine oocytes J Spiropoulos, SE Long University of Bristol, School of Veterinary Science, Department
A comparison of the effects of estrus cow serum and fetal calf serum on in vitro nuclear maturation of bovine oocytes J Spiropoulos, SE Long University of Bristol, School of Veterinary Science, Department
Biology 4361 Developmental Biology Exam 1 ID#: October 11, 2005
 Biology 4361 Developmental Biology Name: Key Exam 1 ID#: October 11, 2005 Multiple choice (one point each) 1. Primordial germ cells a. are immortal b. produce polar bodies c. are haploid d. are somatic
Biology 4361 Developmental Biology Name: Key Exam 1 ID#: October 11, 2005 Multiple choice (one point each) 1. Primordial germ cells a. are immortal b. produce polar bodies c. are haploid d. are somatic
Organisms that reproduce Sexually are made up of two different types of cells.
 MEIOSIS Organisms that reproduce Sexually are made up of two different types of cells. 1. Somatic Cells are body cells and contain the normal number of chromosomes.called the Diploid number (the symbol
MEIOSIS Organisms that reproduce Sexually are made up of two different types of cells. 1. Somatic Cells are body cells and contain the normal number of chromosomes.called the Diploid number (the symbol
The Process of Cell Division
 Lesson Overview 10.2 The Process of Cell Division THINK ABOUT IT What role does cell division play in your life? Does cell division stop when you are finished growing? Chromosomes What is the role of chromosomes
Lesson Overview 10.2 The Process of Cell Division THINK ABOUT IT What role does cell division play in your life? Does cell division stop when you are finished growing? Chromosomes What is the role of chromosomes
Creating Identical Body Cells
 Creating Identical Body Cells 5.A Students will describe the stages of the cell cycle, including DNA replication and mitosis, and the importance of the cell cycle to the growth of organisms 5.D Students
Creating Identical Body Cells 5.A Students will describe the stages of the cell cycle, including DNA replication and mitosis, and the importance of the cell cycle to the growth of organisms 5.D Students
10-2 Cell Division. Chromosomes
 Cell Division In eukaryotes, cell division occurs in two major stages. The first stage, division of the cell nucleus, is called mitosis. The second stage, division of the cell cytoplasm, is called cytokinesis.
Cell Division In eukaryotes, cell division occurs in two major stages. The first stage, division of the cell nucleus, is called mitosis. The second stage, division of the cell cytoplasm, is called cytokinesis.
Cell Division. Chromosome structure. Made of chromatin (mix of DNA and protein) Only visible during cell division
 Only visible during cell division") Chromosome structure Made of chromatin (mix of DNA and protein) Only visible during cell division Chromosome structure The DNA in a cell is packed into an elaborate, multilevel system of coiling and folding.
Chromosome structure Made of chromatin (mix of DNA and protein) Only visible during cell division Chromosome structure The DNA in a cell is packed into an elaborate, multilevel system of coiling and folding.
CELL CYCLE INTRODUCTION PART I ANIMAL CELL CYCLE INTERPHASE EVOLUTION/HEREDITY UNIT. Activity #3
 AP BIOLOGY EVOLUTION/HEREDITY UNIT Unit 1 Part 3 Chapter 12 Activity #3 INTRODUCTION CELL CYCLE NAME DATE PERIOD The nuclei in cells of eukaryotic organisms contain chromosomes with clusters of genes,
AP BIOLOGY EVOLUTION/HEREDITY UNIT Unit 1 Part 3 Chapter 12 Activity #3 INTRODUCTION CELL CYCLE NAME DATE PERIOD The nuclei in cells of eukaryotic organisms contain chromosomes with clusters of genes,
The Cell Cycle and How Cells Divide
 The Cell Cycle and How Cells Divide 1 Phases of the Cell Cycle The cell cycle consists of Interphase normal cell activity The mitotic phase cell divsion INTERPHASE Growth G 1 (DNA synthesis) Growth G 2
The Cell Cycle and How Cells Divide 1 Phases of the Cell Cycle The cell cycle consists of Interphase normal cell activity The mitotic phase cell divsion INTERPHASE Growth G 1 (DNA synthesis) Growth G 2
Cellular Reproduction, Part 2: Meiosis Lecture 10 Fall 2008
 Mitosis & 1 Cellular Reproduction, Part 2: Lecture 10 Fall 2008 Mitosis Form of cell division that leads to identical daughter cells with the full complement of DNA Occurs in somatic cells Cells of body
Mitosis & 1 Cellular Reproduction, Part 2: Lecture 10 Fall 2008 Mitosis Form of cell division that leads to identical daughter cells with the full complement of DNA Occurs in somatic cells Cells of body
Why do cells divide? Cells divide in order to make more cells they multiply in order to create a larger surface to volume ratio!!!
 Why do cells divide? Cells divide in order to make more cells they multiply in order to create a larger surface to volume ratio!!! Chromosomes Are made of chromatin: a mass of genetic material composed
Why do cells divide? Cells divide in order to make more cells they multiply in order to create a larger surface to volume ratio!!! Chromosomes Are made of chromatin: a mass of genetic material composed
Biology is the only subject in which multiplication is the same thing as division
 Biology is the only subject in which multiplication is the same thing as division The Cell Cycle: Cell Growth, Cell Division 2007-2008 2007-2008 Getting from there to here Going from egg to baby. the original
Biology is the only subject in which multiplication is the same thing as division The Cell Cycle: Cell Growth, Cell Division 2007-2008 2007-2008 Getting from there to here Going from egg to baby. the original
Chapter 12. living /non-living? growth repair renew. Reproduction. Reproduction. living /non-living. fertilized egg (zygote) next chapter
 next chapter") Chapter 12 How cells divide Reproduction living /non-living? growth repair renew based on cell division first mitosis - distributes identical sets of chromosomes cell cycle (life) Cell Division in Bacteria
Chapter 12 How cells divide Reproduction living /non-living? growth repair renew based on cell division first mitosis - distributes identical sets of chromosomes cell cycle (life) Cell Division in Bacteria
Change in frequency of radiation induced micronuclei during interphase of four-cell mouse embryos in vitro
 Radiat Environ Biophys (1986) 25:195-199 Radiation and Environmental Biophysics Springer-Verlag 1986 Change in frequency of radiation induced micronuclei during interphase of four-cell mouse embryos in
Radiat Environ Biophys (1986) 25:195-199 Radiation and Environmental Biophysics Springer-Verlag 1986 Change in frequency of radiation induced micronuclei during interphase of four-cell mouse embryos in
Cell Division and Mitosis
 Chromatin-Uncoiled DNA during interphase Cell Division and Mitosis Chromosomes-Tightly coiled DNA Chromatid-One half of a duplicated chromosome. Each is identical and called sister chromatids Centromere-The
Chromatin-Uncoiled DNA during interphase Cell Division and Mitosis Chromosomes-Tightly coiled DNA Chromatid-One half of a duplicated chromosome. Each is identical and called sister chromatids Centromere-The
Outline Interphase Mitotic Stage Cell Cycle Control Apoptosis Mitosis Mitosis in Animal Cells Cytokinesis Cancer Prokaryotic Cell Division
 The Cell Cycle and Cellular Reproduction Chapter 9 Outline Interphase Mitotic Stage Cell Cycle Control Apoptosis Mitosis Mitosis in Animal Cells Cytokinesis Cancer Prokaryotic Cell Division 1 2 Interphase
The Cell Cycle and Cellular Reproduction Chapter 9 Outline Interphase Mitotic Stage Cell Cycle Control Apoptosis Mitosis Mitosis in Animal Cells Cytokinesis Cancer Prokaryotic Cell Division 1 2 Interphase
Regulators of Cell Cycle Progression
 Regulators of Cell Cycle Progression Studies of Cdk s and cyclins in genetically modified mice reveal a high level of plasticity, allowing different cyclins and Cdk s to compensate for the loss of one
Regulators of Cell Cycle Progression Studies of Cdk s and cyclins in genetically modified mice reveal a high level of plasticity, allowing different cyclins and Cdk s to compensate for the loss of one
SUPPORTING ONLINE MATERIAL
 SUPPORTING ONLINE MATERIAL SUPPORTING ONLINE TEXT Efficiency of SCNT Alive fetuses at mid-gestation The rate of viable (beating heart) embryos at day 12.5-14.5 dpc was assessed after sacrifice of foster
SUPPORTING ONLINE MATERIAL SUPPORTING ONLINE TEXT Efficiency of SCNT Alive fetuses at mid-gestation The rate of viable (beating heart) embryos at day 12.5-14.5 dpc was assessed after sacrifice of foster
The Cell Cycle. Dr. SARRAY Sameh, Ph.D
 The Cell Cycle Dr. SARRAY Sameh, Ph.D Overview When an organism requires additional cells (either for growth or replacement of lost cells), new cells are produced by cell division (mitosis) Somatic cells
The Cell Cycle Dr. SARRAY Sameh, Ph.D Overview When an organism requires additional cells (either for growth or replacement of lost cells), new cells are produced by cell division (mitosis) Somatic cells
Why do cells reproduce?
 Outline Cell Reproduction 1. Overview of Cell Reproduction 2. Cell Reproduction in Prokaryotes 3. Cell Reproduction in Eukaryotes 1. Chromosomes 2. Cell Cycle 3. Mitosis and Cytokinesis Examples of Cell
Outline Cell Reproduction 1. Overview of Cell Reproduction 2. Cell Reproduction in Prokaryotes 3. Cell Reproduction in Eukaryotes 1. Chromosomes 2. Cell Cycle 3. Mitosis and Cytokinesis Examples of Cell
Mitosis. AND Cell DiVISION
 Mitosis AND Cell DiVISION Cell Division Characteristic of living things: ability to reproduce their own kind. Cell division purpose: When unicellular organisms such as amoeba divide to form offspring reproduction
Mitosis AND Cell DiVISION Cell Division Characteristic of living things: ability to reproduce their own kind. Cell division purpose: When unicellular organisms such as amoeba divide to form offspring reproduction
Biology is the only subject in which multiplication is the same thing as division
 Biology is the only subject in which multiplication is the same thing as division 2007-2008 The Cell Cycle: Cell Growth, Cell Division 2007-2008 Getting from there to here Going from egg to baby. the original
Biology is the only subject in which multiplication is the same thing as division 2007-2008 The Cell Cycle: Cell Growth, Cell Division 2007-2008 Getting from there to here Going from egg to baby. the original
F ertilizability of Rabbit Ova after Removal of the Corona Radiata
 F ertilizability of Rabbit Ova after Removal of the Corona Radiata M. C. CHANG, Ph.D., and J. M. BEDFORD, M.R.C.V.S." FRESHLY ovulated rabbit ova are surrounded by a mass of follicular cells in a mucous
F ertilizability of Rabbit Ova after Removal of the Corona Radiata M. C. CHANG, Ph.D., and J. M. BEDFORD, M.R.C.V.S." FRESHLY ovulated rabbit ova are surrounded by a mass of follicular cells in a mucous
NOTES- CHAPTER 6 CHROMOSOMES AND CELL REPRODUCTION
 NOTES- CHAPTER 6 CHROMOSOMES AND CELL REPRODUCTION Section I Chromosomes Formation of New Cells by Cell Division New cells are formed when old cells divide. 1. Cell division is the same as cell reproduction.
NOTES- CHAPTER 6 CHROMOSOMES AND CELL REPRODUCTION Section I Chromosomes Formation of New Cells by Cell Division New cells are formed when old cells divide. 1. Cell division is the same as cell reproduction.
BIOLOGY 4/6/2015. Cell Cycle - Mitosis. Outline. Overview: The Key Roles of Cell Division. identical daughter cells. I. Overview II.
 2 Cell Cycle - Mitosis CAMPBELL BIOLOGY TENTH EDITION Reece Urry Cain Wasserman Minorsky Jackson Outline I. Overview II. Mitotic Phase I. Prophase II. III. Telophase IV. Cytokinesis III. Binary fission
2 Cell Cycle - Mitosis CAMPBELL BIOLOGY TENTH EDITION Reece Urry Cain Wasserman Minorsky Jackson Outline I. Overview II. Mitotic Phase I. Prophase II. III. Telophase IV. Cytokinesis III. Binary fission
Abstract. Introduction. Materials and methods
 RBMOnline - Vol 10. No 2. 2005 199-204 Reproductive BioMedicine Online; www.rbmonline.com/article/1592 on web 15 December 2004 Article Cytogenetic analysis of human somatic cell haploidization Dr Vasiliy
RBMOnline - Vol 10. No 2. 2005 199-204 Reproductive BioMedicine Online; www.rbmonline.com/article/1592 on web 15 December 2004 Article Cytogenetic analysis of human somatic cell haploidization Dr Vasiliy
Mitosis: cell division that forms identical daughter cells with the same number of chromosomes as the parent cell (duplicate and divide)
") Mitosis: cell division that forms identical daughter cells with the same number of chromosomes as the parent cell (duplicate and divide) Meiosis: cell division that forms daughter cells with half the number
Mitosis: cell division that forms identical daughter cells with the same number of chromosomes as the parent cell (duplicate and divide) Meiosis: cell division that forms daughter cells with half the number
Name Date Class. Interphase. (1) The. grows. DNA is duplicated.
 The. grows. DNA is duplicated.") Concept Mapping The Cell Cycle Complete the cycle map about the cell cycle. These terms may be used more than once: cell, cytoplasm, metaphase, nuclear membrane, nucleoli, poles. (1) The Interphase grows.
Concept Mapping The Cell Cycle Complete the cycle map about the cell cycle. These terms may be used more than once: cell, cytoplasm, metaphase, nuclear membrane, nucleoli, poles. (1) The Interphase grows.
Cell plate Carcinogen Oncogenes. Haploid cell Diploid cell Chromosome. Telophase Keywords Mitosis
 Cell Division Cell plate Carcinogen Oncogenes Haploid cell Diploid cell Chromosome Gene Interphase Anaphase Metaphase Telophase Keywords Mitosis Meiosis Centromere Cleavage furrow Middle lamella What is
Cell Division Cell plate Carcinogen Oncogenes Haploid cell Diploid cell Chromosome Gene Interphase Anaphase Metaphase Telophase Keywords Mitosis Meiosis Centromere Cleavage furrow Middle lamella What is
Chapter 12. The Cell Cycle
 Chapter 12 The Cell Cycle The Key Roles of Cell Division The ability of organisms to produce more of their own kind is the one characteristic that best distinguishes living things from nonliving things.
Chapter 12 The Cell Cycle The Key Roles of Cell Division The ability of organisms to produce more of their own kind is the one characteristic that best distinguishes living things from nonliving things.
10-2 Cell Division mitosis. cytokinesis. Chromosomes chromosomes Slide 1 of 38
 In eukaryotes, cell division occurs in two major stages. The first stage, division of the cell nucleus, is called mitosis. The second stage, division of the cell cytoplasm, is called cytokinesis. Chromosomes
In eukaryotes, cell division occurs in two major stages. The first stage, division of the cell nucleus, is called mitosis. The second stage, division of the cell cytoplasm, is called cytokinesis. Chromosomes
Cell Division. Learning Objectives: Introduction. Revised Fall 2018
 Revised Fall 2018 Cell Division Learning Objectives: 1. Define cell cycle and the ordered sequence of events in the cell cycle (Interphase and The divisional phase or M phase) 2. Explain the stages in
Revised Fall 2018 Cell Division Learning Objectives: 1. Define cell cycle and the ordered sequence of events in the cell cycle (Interphase and The divisional phase or M phase) 2. Explain the stages in
BIOLOGY. Cell Cycle - Mitosis. Outline. Overview: The Key Roles of Cell Division. identical daughter cells. I. Overview II.
 2 Cell Cycle - Mitosis CAMPBELL BIOLOGY TENTH EDITION Reece Urry Cain Wasserman Minorsky Jackson Outline I. Overview II. Mitotic Phase I. Prophase II. III. Telophase IV. Cytokinesis III. Binary fission
2 Cell Cycle - Mitosis CAMPBELL BIOLOGY TENTH EDITION Reece Urry Cain Wasserman Minorsky Jackson Outline I. Overview II. Mitotic Phase I. Prophase II. III. Telophase IV. Cytokinesis III. Binary fission
Cell Cycle and Cell Division
 122 Cell Cycle and Cell Division 1. Meiosis I is reductional division. Meiosis II is equational division due to [1988] (a) pairing of homologous chromosomes (b) crossing over (c) separation of chromatids
122 Cell Cycle and Cell Division 1. Meiosis I is reductional division. Meiosis II is equational division due to [1988] (a) pairing of homologous chromosomes (b) crossing over (c) separation of chromatids
General Embryology. School of Medicine Department of Anatomy and Histology School of medicine The University of Jordan
 General Embryology 2019 School of Medicine Department of Anatomy and Histology School of medicine The University of Jordan https://www.facebook.com/dramjad-shatarat What is embryology? Is the science that
General Embryology 2019 School of Medicine Department of Anatomy and Histology School of medicine The University of Jordan https://www.facebook.com/dramjad-shatarat What is embryology? Is the science that
8.4 The cell cycle multiplies cells. 8.4 The cell cycle multiplies cells
 8.4 The cell cycle multiplies cells! Cell division is a highly orchestrated process! The cell cycle is an ordered sequence of events that extends from the time a cell is first formed from a dividing parent
8.4 The cell cycle multiplies cells! Cell division is a highly orchestrated process! The cell cycle is an ordered sequence of events that extends from the time a cell is first formed from a dividing parent
Biology is the only subject in which multiplication is the same thing as division
 Biology is the only subject in which multiplication is the same thing as division 2007-2008 The Cell Cycle: Cell Growth, Cell Division 2007-2008 Where it all began You started as a cell smaller than a
Biology is the only subject in which multiplication is the same thing as division 2007-2008 The Cell Cycle: Cell Growth, Cell Division 2007-2008 Where it all began You started as a cell smaller than a
U3.2.3: Eukaryotic chromosomes are linear DNA molecules associated with histone proteins. (Oxford Biology Course Companion page 151).
.") Cell Division Study Guide U3.2.3: Eukaryotic chromosomes are linear DNA molecules associated with histone proteins. (Oxford Biology Course Companion page 151). 1. Describe the structure of eukaryotic DNA
Cell Division Study Guide U3.2.3: Eukaryotic chromosomes are linear DNA molecules associated with histone proteins. (Oxford Biology Course Companion page 151). 1. Describe the structure of eukaryotic DNA
APGRU4L1 Chap 12 Extra Reading Cell Cycle and Mitosis
 APGRU4L1 Chap 12 Extra Reading Cell Cycle and Mitosis Dr. Ramesh Biology is the only subject in which multiplication is the same thing as division 2007-2008 The Cell Cycle: Cell Growth, Cell Division 2007-2008
APGRU4L1 Chap 12 Extra Reading Cell Cycle and Mitosis Dr. Ramesh Biology is the only subject in which multiplication is the same thing as division 2007-2008 The Cell Cycle: Cell Growth, Cell Division 2007-2008
Cell Cycle and Mitosis
 Cell Cycle and Mitosis Name Period A# THE CELL CYCLE The cell cycle, or cell-division cycle, is the series of events that take place in a eukaryotic cell between its formation and the moment it replicates
Cell Cycle and Mitosis Name Period A# THE CELL CYCLE The cell cycle, or cell-division cycle, is the series of events that take place in a eukaryotic cell between its formation and the moment it replicates
LECTURE PRESENTATIONS
 LECTURE PRESENTATIONS For CAMPBELL BIOLOGY, NINTH EDITION Jane B. Reece, Lisa A. Urry, Michael L. Cain, Steven A. Wasserman, Peter V. Minorsky, Robert B. Jackson Chapter 12 The Cell Cycle Lectures by Erin
LECTURE PRESENTATIONS For CAMPBELL BIOLOGY, NINTH EDITION Jane B. Reece, Lisa A. Urry, Michael L. Cain, Steven A. Wasserman, Peter V. Minorsky, Robert B. Jackson Chapter 12 The Cell Cycle Lectures by Erin
Biology is the only subject in which multiplication is the same thing as division
 Biology is the only subject in which multiplication is the same thing as division 2007-2008 The Cell Cycle: Cell Growth, Cell Division 2007-2008 Where it all began You started as a cell smaller than a
Biology is the only subject in which multiplication is the same thing as division 2007-2008 The Cell Cycle: Cell Growth, Cell Division 2007-2008 Where it all began You started as a cell smaller than a
Cell Cycle/Mitosis -Notes-
 Cell Cycle/Mitosis -Notes- LIMITS TO CELL GROWTH The a cell becomes, the more demands the cell places on DNA. Additionally, the cell has more trouble moving enough and wastes across the cell membrane.
Cell Cycle/Mitosis -Notes- LIMITS TO CELL GROWTH The a cell becomes, the more demands the cell places on DNA. Additionally, the cell has more trouble moving enough and wastes across the cell membrane.
Prentice Hall Biology Slide 1 of 38
 Prentice Hall Biology 1 of 38 2 of 38 In eukaryotes, cell division occurs in two major stages. The first stage, division of the cell nucleus, is called mitosis. The second stage, division of the cell cytoplasm,
Prentice Hall Biology 1 of 38 2 of 38 In eukaryotes, cell division occurs in two major stages. The first stage, division of the cell nucleus, is called mitosis. The second stage, division of the cell cytoplasm,
Biology is the only subject in which multiplication is the same thing as division
 The Cell Cycle Biology is the only subject in which multiplication is the same thing as division Why do cells divide? For reproduction asexual reproduction For growth one-celled organisms from fertilized
The Cell Cycle Biology is the only subject in which multiplication is the same thing as division Why do cells divide? For reproduction asexual reproduction For growth one-celled organisms from fertilized
Mitosis and Cytokinesis
 B-2.6 Summarize the characteristics of the cell cycle: interphase (called G1, S, G2); the phases of mitosis (called prophase, metaphase, anaphase, and telophase); and plant and animal cytokinesis. The
B-2.6 Summarize the characteristics of the cell cycle: interphase (called G1, S, G2); the phases of mitosis (called prophase, metaphase, anaphase, and telophase); and plant and animal cytokinesis. The
Cell Division Questions. Mitosis and Meiosis
 Cell Division Questions Mitosis and Meiosis 1 10 Do not write outside the box 5 Figure 3 shows a pair of chromosomes at the start of meiosis. The letters represent alleles. Figure 3 E E e e F F f f 5 (a)
Cell Division Questions Mitosis and Meiosis 1 10 Do not write outside the box 5 Figure 3 shows a pair of chromosomes at the start of meiosis. The letters represent alleles. Figure 3 E E e e F F f f 5 (a)
Omnis cellula e cellula
 Chapter 12 The Cell Cycle Omnis cellula e cellula 1855- Rudolf Virchow German scientist all cells arise from a previous cell Every cell from a cell In order for this to be true, cells must have the ability
Chapter 12 The Cell Cycle Omnis cellula e cellula 1855- Rudolf Virchow German scientist all cells arise from a previous cell Every cell from a cell In order for this to be true, cells must have the ability
The Cell Cycle. Chapter 12. Biology Eighth Edition Neil Campbell and Jane Reece. PowerPoint Lecture Presentations for
 Chapter 12 The Cell Cycle PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions from Joan Sharp Copyright
Chapter 12 The Cell Cycle PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions from Joan Sharp Copyright
10-2 Cell Division. Slide 1 of 38. End Show. Copyright Pearson Prentice Hall
 1 of 38 Cell Division In eukaryotes, cell division occurs in two major stages. The first stage, division of the cell nucleus, is called mitosis. The second stage, division of the cell cytoplasm, is called
1 of 38 Cell Division In eukaryotes, cell division occurs in two major stages. The first stage, division of the cell nucleus, is called mitosis. The second stage, division of the cell cytoplasm, is called
The Cell Cycle. Chapter 12. PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece
 Chapter 12 The Cell Cycle PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions from Joan Sharp Overview:
Chapter 12 The Cell Cycle PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions from Joan Sharp Overview:
Mitosis & Meiosis. Diploid cells- (2n)- a cell that has 2 of each chromosome - 1 from mom, 1 from dad = 1 pair
- a cell that has 2 of each chromosome - 1 from mom, 1 from dad = 1 pair") Mitosis & Meiosis Diploid cells- (2n)- a cell that has 2 of each chromosome - 1 from mom, 1 from dad = 1 pair The pair is called homologous chromosomes The homologous chromosomes contain the same gene
Mitosis & Meiosis Diploid cells- (2n)- a cell that has 2 of each chromosome - 1 from mom, 1 from dad = 1 pair The pair is called homologous chromosomes The homologous chromosomes contain the same gene
The Cell Cycle. Packet #9. Thursday, August 20, 2015
 1 The Cell Cycle Packet #9 2 Introduction Cell Cycle An ordered sequence of events in the life of a dividing eukaryotic cell and is a cellular asexual reproduction. The contents of the parent s cell nucleus
1 The Cell Cycle Packet #9 2 Introduction Cell Cycle An ordered sequence of events in the life of a dividing eukaryotic cell and is a cellular asexual reproduction. The contents of the parent s cell nucleus
(i) List these events in the correct order, starting with D.... (1)... (1)... (1)
 List these events in the correct order, starting with D.... (1)... (1)... (1)") Q1. (a) Boxes A to E show some of the events of the cell cycle. A Chromatids seperate B Nuclear envelopes disappears C Cytoplasm divides D Chromosomes condense and become visible E Chromosomes on the equator
Q1. (a) Boxes A to E show some of the events of the cell cycle. A Chromatids seperate B Nuclear envelopes disappears C Cytoplasm divides D Chromosomes condense and become visible E Chromosomes on the equator
Biology 4A Laboratory MITOSIS Asexual Reproduction OBJECTIVE
 Biology 4A Laboratory MITOSIS Asexual Reproduction OBJECTIVE To study the cell cycle and understand how, when and why cells divide. To study and identify the major stages of cell division. To relate the
Biology 4A Laboratory MITOSIS Asexual Reproduction OBJECTIVE To study the cell cycle and understand how, when and why cells divide. To study and identify the major stages of cell division. To relate the
BIOLOGY 111. CHAPTER 9: The Links in Life s Chain Genetics and Cell Division
 BIOLOGY 111 CHAPTER 9: The Links in Life s Chain Genetics and Cell Division The Links in Life s Chain: Genetics and Cell Division 9.1 An Introduction to Genetics 9.2 An Introduction to Cell Division 9.3
BIOLOGY 111 CHAPTER 9: The Links in Life s Chain Genetics and Cell Division The Links in Life s Chain: Genetics and Cell Division 9.1 An Introduction to Genetics 9.2 An Introduction to Cell Division 9.3
Stages of Mitosis. Introduction
 Name: Due: Stages of Mitosis Introduction Mitosis, also called karyokinesis, is division of the nucleus and its chromosomes. It is followed by division of the cytoplasm known as cytokinesis. Both mitosis
Name: Due: Stages of Mitosis Introduction Mitosis, also called karyokinesis, is division of the nucleus and its chromosomes. It is followed by division of the cytoplasm known as cytokinesis. Both mitosis
meiosis asexual reproduction CHAPTER 9 & 10 The Cell Cycle, Meiosis & Sexual Life Cycles Sexual reproduction mitosis
 meiosis asexual reproduction CHAPTER 9 & 10 The Cell Cycle, Meiosis & Sexual Sexual reproduction Life Cycles mitosis Chromosomes Consists of a long DNA molecule (represents thousands of genes) Also consists
meiosis asexual reproduction CHAPTER 9 & 10 The Cell Cycle, Meiosis & Sexual Sexual reproduction Life Cycles mitosis Chromosomes Consists of a long DNA molecule (represents thousands of genes) Also consists
Cell Division (Mitosis)
") Cell Division (Mitosis) Chromosomes The essential part of a chromosome is a single very long strand of DNA. This DNA contains all the genetic information for creating and running the organism. Each chromosome
Cell Division (Mitosis) Chromosomes The essential part of a chromosome is a single very long strand of DNA. This DNA contains all the genetic information for creating and running the organism. Each chromosome
The Cell Cycle. Chapter 12. Biology. Edited by Shawn Lester. Eighth Edition Neil Campbell and Jane Reece. PowerPoint Lecture Presentations for
 Chapter 12 The Cell Cycle Edited by Shawn Lester PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions
Chapter 12 The Cell Cycle Edited by Shawn Lester PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions
(FITC) or rhodamine blue isothiocyanate (RBITC) for use in mixed egg-transfer experiments. Both FITC and RBITC bind to the zona pellucida
 or rhodamine blue isothiocyanate (RBITC) for use in mixed egg-transfer experiments. Both FITC and RBITC bind to the zona pellucida") THE LABELLING OF LIVING RABBIT OVA WITH FLUORESCENT DYES J. W. OVERSTREET Department of Anatomy and International Institute for the Study of Human Reproduction, Columbia University, College of Physicians
THE LABELLING OF LIVING RABBIT OVA WITH FLUORESCENT DYES J. W. OVERSTREET Department of Anatomy and International Institute for the Study of Human Reproduction, Columbia University, College of Physicians
Cell Cycle. Cell Cycle the cell s life cycle that extends from one division to the next G1 phase, the first gap phase. S phase, synthesis phase
 Cell Cycle the cell s life cycle that extends from one division to the next G1 phase, the first gap phase Cell Cycle interval between cell division and DNA replication accumulates materials needed to replicate
Cell Cycle the cell s life cycle that extends from one division to the next G1 phase, the first gap phase Cell Cycle interval between cell division and DNA replication accumulates materials needed to replicate
Cell Cycle, Mitosis, and Microtubules. LS1A Final Exam Review Friday 1/12/07. Processes occurring during cell cycle
 Cell Cycle, Mitosis, and Microtubules LS1A Final Exam Review Friday 1/12/07 Processes occurring during cell cycle Replicate chromosomes Segregate chromosomes Cell divides Cell grows Cell Growth 1 The standard
Cell Cycle, Mitosis, and Microtubules LS1A Final Exam Review Friday 1/12/07 Processes occurring during cell cycle Replicate chromosomes Segregate chromosomes Cell divides Cell grows Cell Growth 1 The standard
The Cell Cycle. Chapter 12. Biology Eighth Edition Neil Campbell and Jane Reece. PowerPoint Lecture Presentations for
 Chapter 12 The Cell Cycle PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions from Joan Sharp 1
Chapter 12 The Cell Cycle PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions from Joan Sharp 1
Mitosis Notes AP Biology Mrs. Laux
 I. Cell Cycle-includes interphase and mitosis (IPPMAT) A. Interphase 1. accounts for 90% of the cycle 2. cell grows and copies its chromosomes in preparation for cell division 3. produces proteins and
I. Cell Cycle-includes interphase and mitosis (IPPMAT) A. Interphase 1. accounts for 90% of the cycle 2. cell grows and copies its chromosomes in preparation for cell division 3. produces proteins and
KEY CONCEPT Cells have distinct phases of growth, reproduction, and normal functions.
 5.1 The Cell Cycle KEY CONCEPT Cells have distinct phases of growth, reproduction, and normal functions. Objective: Cells have distinct phases of growth, reproduction and normal functions. APK: Why do
5.1 The Cell Cycle KEY CONCEPT Cells have distinct phases of growth, reproduction, and normal functions. Objective: Cells have distinct phases of growth, reproduction and normal functions. APK: Why do
The Cell Life Cycle. S DNA replication, INTERPHASE. G 2 Protein. G 1 Normal THE CELL CYCLE. Indefinite period. synthesis. of histones.
 Mitosis & Meiosis The Cell Life Cycle INTERPHASE G 1 Normal cell functions plus cell growth, duplication of organelles, protein synthesis S DNA replication, synthesis of histones THE CELL CYCLE M G 2 Protein
Mitosis & Meiosis The Cell Life Cycle INTERPHASE G 1 Normal cell functions plus cell growth, duplication of organelles, protein synthesis S DNA replication, synthesis of histones THE CELL CYCLE M G 2 Protein
Mitosis and Cellular Division. EQ: How do the cells in our body divide?
 Mitosis and Cellular Division EQ: How do the cells in our body divide? Cell division is the process by which cellular material is divided between two new daughter cells. 1 Mother Cell 2 Daughter cells.
Mitosis and Cellular Division EQ: How do the cells in our body divide? Cell division is the process by which cellular material is divided between two new daughter cells. 1 Mother Cell 2 Daughter cells.
LECTURE PRESENTATIONS
 LECTURE PRESENTATIONS For CAMPBELL BIOLOGY, NINTH EDITION Jane B. Reece, Lisa A. Urry, Michael L. Cain, Steven A. Wasserman, Peter V. Minorsky, Robert B. Jackson Chapter 12 The Cell Cycle Lectures by Erin
LECTURE PRESENTATIONS For CAMPBELL BIOLOGY, NINTH EDITION Jane B. Reece, Lisa A. Urry, Michael L. Cain, Steven A. Wasserman, Peter V. Minorsky, Robert B. Jackson Chapter 12 The Cell Cycle Lectures by Erin
MITOSIS AND THE CELL CYCLE PowerPoint Notes
 1 Name: Date: MITOSIS AND THE CELL CYCLE PowerPoint Notes THE FUNCTIONS OF CELL DIVISION 1. Cell division is vital for all. living organisms This is the only process that can create. new cells 2. Cell
1 Name: Date: MITOSIS AND THE CELL CYCLE PowerPoint Notes THE FUNCTIONS OF CELL DIVISION 1. Cell division is vital for all. living organisms This is the only process that can create. new cells 2. Cell