ATTENUATION OF CIRCADIAN DYSFUNCTION IMPROVES SLEEP, MOOD AND NEUROPSYCHOMETRIC PERFORMANCE
|
|
|
- Hannah Green
- 5 years ago
- Views:
Transcription
1 ATTENUATION OF CIRCADIAN DYSFUNCTION IMPROVES SLEEP, MOOD AND NEUROPSYCHOMETRIC PERFORMANCE BY SHADAB ATAUR RAHMAN A THESIS SUBMITTED IN CONFORMITY WITH THE REQUIREMENTS FOR THE DEGREE OF DOCTOR OF PHILOSOPHY GRADUATE DEPARTMENT OF PHYSIOLOGY UNIVERSITY OF TORONTO COPYRIGHT BY SHADAB ATAUR RAHMAN 2010
2 ATTENUATION OF CIRCADIAN DYSFUNCTION IMPROVES SLEEP, MOOD AND NEUROPSYCHOMETRIC PERFORMANCE Shadab Ataur Rahman, Doctor of Philosophy, 2010 Graduate Department of Physiology, University of Toronto, Toronto, Ontario, Canada ABSTRACT Mood and cognition, along with numerous other physiological processes, are under circadian regulation. The synthesis and secretion rhythm of the pineal hormone melatonin is under the direct regulation of the central circadian pacemaker and the secretion rhythm of melatonin can be used to assess circadian alterations. In this thesis, it was demonstrated that low levels of endogenous nocturnal melatonin was associated with subsyndromal depression and alterations in sleep architecture. Studies in individuals with endogenous circadian rhythm disorder, with and without comorbid depressive symptoms, revealed that individuals with depressive symptoms had a greater phase delay in melatonin profiles as compared to individuals without depressive symptoms. Furthermore, in the same study, exogenous melatonin administered to induce phase advances significantly improved depression scores and sleep initiation. In addition to endogenous circadian disruption, circadian rhythms can also be disrupted by repeated atypical alterations in environmental time cues. In mammals, light is the strongest environmental cue that can modulate circadian rhythms. Recent studies suggest that circadian response to photic stimuli is preferentially sensitive to short wavelengths in the range of nm. Using an animal model it was demonstrated that filtering a 10 nm bandwidth between nm from polychromatic white light prevents nocturnal light exposure induced disruptions in melatonin and corticosterone secretion as well as central and peripheral clock gene expression. These findings ii
3 were further investigated in humans and revealed that filtering short wavelengths below 480 nm attenuates 12 h nocturnal light exposure induced suppression of melatonin secretion, increased cortisol secretion and disrupted peripheral clock gene expression. Furthermore, attenuation of these changes was associated with improvements in mood, alertness and vigilance at a time close to the endogenous circadian wake drive. However, filtering short wavelengths below 460 nm or reducing the optical transmission by up to 30% below 480 nm did not attenuate the disruptive effects of nocturnal light exposure on physiological and behavioural variables. Overall, the results presented in this thesis support the role of circadian dysfunction in neuropsychometric impairment and presents evidence supporting spectral modulation as a promising approach to attenuate light-mediated chronodisruption. iii
4 ACKNOWLEDGMENTS ALLAHU NURU SAMAWATI WALARDDI DIXITQUE DEUS FIAT LUX ET FACTA EST LUX To: ALLAH, Rasul, my Gaus, my Huzur My parents, sister, wife, family & friends Dr. Casper Dr. Brown All my teachers Alex, Domenica, Dr. Tone-A, Prem, Shawn, Shakib Bhai Thank you iv
5 TABLE OF CONTENTS ACKNOWLEDGMENTS... iv TABLE OF CONTENTS... v LIST OF TABLES... x LIST OF FIGURES... xi ABBREVIATIONS... xiii 1 LITERATURE REVIEW OVERVIEW A BRIEF CHRONOLOGY OF CHRONOBIOLOGY CIRCADIAN RHYTHM QUALIFICATION GENERAL PRINCIPLES OF CIRCADIAN RHYTHM BIOLOGY A linear model of circadian synchronization Core parameters of circadian rhythms Models of entrainment The circadian system is built on multiple circadian oscillators THE OSCILLATORY MECHANISM Molecular oscillatory loops The master clock: the suprachiasmatic nucleus Organization of the SCN CIRCADIAN RESPONSE TO LIGHT (PHOTIC) STIMULI Evolution of circadian photic response Mammalian circadian phototransduction begins at the retina Dedicated circadian photoreceptor Resetting of the central clock by light v
6 1.6.5 Unique spectral sensitivity of circadian photic response EFFECTS OF LIGHT EXPOSURE ON CIRCADIAN ENDOCRINE PATHWAYS Classical circadian endocrine pathways Melatonin as an endogenous neuroendocrine time cue Melatonin as an endogenous circadian phase marker EFFECTS OF LIGHT EXPOSURE ON CIRCADIAN BEHAVIORAL PATHWAYS Sleep Alertness Mood The role of circadian rhythms in mood disorders Categorizing depression CIRCADIAN RHYTHM DYSFUNCTION Endogenous circadian dysfunction and circadian rhythm sleep disorders Prevalence of circadian rhythm sleep disorders Shift work and exogenous circadian dysfunction Health effects of shift work Shift work countermeasures THESIS HYPOTHESIS AND OBJECTIVES ALTERED SLEEP ARCHITECTURE AND HIGHER INCIDENCE OF SUBSYNDROMAL DEPRESSION IN LOW ENDOGENOUS MELATONIN SECRETORS ABSTRACT INTRODUCTION METHODS Sample population Study design vi
7 2.3.3 Objective measures: two night polysomnography Subjective measures Circadian rhythm parameters Psychiatric parameters Data analysis RESULTS Melatonin secretion profiles in low endogenous melatonin secretors and controls Sleep architecture in low endogenous melatonin secretors and controls Subjective Neuropsychometrics in Low Endogenous Melatonin Secretors and Controls DISCUSSION ANTIDEPRESSANT ACTION OF MELATONIN IN THE TREATMENT OF DELAYED SLEEP PHASE SYNDROME ABSTRACT INTRODUCTION MATERIALS AND METHODS Study population Study design Sleep physiology measures Psychometric measures Circadian profile measures Data analysis RESULTS Melatonin treatment on depression in DSPS Melatonin treatment on sleep architecture in DSPS Circadian profile of DSPS patients with and without comorbid depression vii
8 3.5 DISCUSSION SELECTIVELY FILTERING SHORT WAVELENGTHS ATTENUATES THE DISRUPTIVE EFFECTS OF NOCTURNAL LIGHT ON ENDOCRINE AND MOLECULAR CIRCADIAN PHASE MARKERS IN RATS ABSTRACT INTRODUCTION MATERIALS AND METHODS Animals Light exposure protocol Sample collection Hormone assays Quantitative real time RT-PCR Cell maintenance, treatments, and cell proliferation assay Western blot analysis Statistics RESULTS Normalization of Circadian Endocrine Phase Markers under Nocturnal Light Exposure ACTH-Independent Modulation of Glucocorticoid Secretion in Response to Nocturnal Light Normalization of central and peripheral clock gene expression patterns Differential wavelength-dependent normalization of endocrine circadian phase markers and clock gene expression Cortisol secretion modulation by melatonin in human adrenocortical cells DISCUSSION ATTENUATING THE DIRECT EFFECTS OF LIGHT EXPOSURE ON CIRCADIAN PHASE MARKERS BY FILTERING SHORT WAVELENGTHS ABSTRACT INTRODUCTION viii
9 5.3 METHODS Study population Light exposure conditions and overall study design Sample collection Hormone analysis Neuropsychometrics Gene expression analysis by quantitative real time RT-PCR Statistical analysis RESULTS Filtering short wavelengths below 480 nm attenuates direct endocrine disruption induced by nocturnal light exposure Filtering short wavelengths below 480 nm attenuates direct peripheral clock gene expression alterations induced by nocturnal light exposure Filtering short wavelengths below 480 nm attenuates direct neuropsychometric impairment induced by nocturnal light exposure DISCUSSION OVERALL DISCUSSION REFERENCES COPYRIGHT ACKNOWLEDGEMENTS ix
10 LIST OF TABLES Table 2.1: Sleep physiology parameters of low endogenous melatonin secretors and the normal secretors Table 3.1: Sleep polysomnographic variables during the course of a 9-week randomized, doubleblind placebo-controlled crossover study in patients with Delayed Sleep Phase Syndrome (DSPS) and comorbid depression Table 3.2: Sleep polysomnographic variables during the course of a 9-week randomized, doubleblind, placebo-controlled crossover study in non-depressed patients with DSPS x
11 LIST OF FIGURES Figure 1.1: Linear model of entrainment Figure 1.2: Fundamental circadian rhythm parameters Figure 1.3: Hierarchical organization of the biological clock Figure 1.4: The mammalian circadian molecular clock Figure 1.5: A schematic representation of the SCN and its efferent and afferent pathways Figure 1.6: A schematic representation of the mammalian retina and photic signal transduction pathway Figure 2.1: Melatonin secretion profile of low endogenous melatonin secretors (LEMS) and normal secretors (controls) Figure 2.2: Psychiatric profile of low endogenous melatonin secretors (LEMS) and normal secretors (controls) Figure 3.1: Ameliorating depression in delayed sleep phase patients with exogenous melatonin treatment Figure 3.2: Altered melatonin secretion profile in DSPS patients with comorbid depression compared to DSPS patients with no depression Figure 4.1: Normalization of melatonin and glucocorticoid secretion independent of ACTH Figure 4.2: Normalization of Per2 and Bmal1 gene expression in the hypothalamus and adrenal gland Figure 4.3: Wavelength-dependent differential normalization of endocrine circadian phase markers and tissue-specific clock gene expression xi
12 Figure 4.4: Inhibition of camp-mediated cortisol secretion from NCI-H295R human adrenocortical cells by melatonin at physiological levels Figure 4.5: Proposed model of differential SCN activation leading to different thresholds in circadian response elicited by polychromatic white light Figure 5.1: Normalization of melatonin and glucocorticoid secretion and clock gene expression under nocturnal light exposure Figure 5.2: Effects of filtering short wavelengths on neuropsychometric performance under nocturnal light exposure Supplemental Figures Supplemental Figure 5.1: Spectral characteristics of the light sources before and after filtering Supplemental Figure 5.2: Normalization of circadian endocrine and clock gene expression rhythms facilitates early morning neuropsychometric performance under nocturnal light exposure xii
13 ABBREVIATIONS µg Microgram µl Microlitre µm Micromolar µw Microwatt 5-HT Serotonin AA-NAT Serotonin N-acetyl transferase AAS Ascending arousal system AC Adenylyl cyclase ACTH Adrenocorticotropic hormone AI Arousal index AMPA α-amino-3-hydroxyl-5-methyl-4-isoxazole-propionate amt6s 6-sulphatoxymelatonin ANOVA Analysis of variance ASPS Advanced sleep phase syndrome bhlh Basic helix loop helix Bmal1/2 Brain and muscle ARNT (aryl hydrocarbon receptor nuclear translocator)-like protein 1/2 camp Cyclic adenosine monophosphate CBT Core body temperature CCG Clock controlled gene cdna Complementary deoxyribonucleic acid Cebpb CCAAT/enhancer-binding protein beta CES-D Center for epidemiologic studies depression scale CK1 ε Casein kinase 1ε Clock Circadian locomotor output cycles kaput CNS Central nervous system CR Constant routine CRE camp response element CREB camp response element binding protein CREM camp responsive element modulator CRH Corticotropin-releasing hormone CRSD Circadian rhythm sleep disorder Cry Cryptochrome CSNK1D/E Casein kinase 1 δ/ε CYP11A1/11B2/21 Cytochrome P450 11A1/11B2/21 Dbp D site of albumin promoter (albumin D-box) binding protein Dec1/2 Deleted in esophageal cancer 1/2 DLMO Dim light melatonin onset DMH Dorsomedial hypothalamus xiii
14 DNase1 Deoxyribonuclease I DSM Diagnostic and statistical manual of mental disorders DSPS Delayed sleep phase syndrome DVT Digit vigilance test E4BP4 E4 promoter binding-protein 4 EEG Electroencephalogram ELISA Enzyme-linked immunosorbent assay EMG Electromyogram EOG Electrooculogram ER Estrogen receptor ESS Epworth sleepiness scale E-type Evening type FD Force desynchrony FEO Food entrainable oscillator Fig. Figure fmri Functional magnetic resonance imaging g Gravitational force GABA γ-aminobutyric acid GAPDH Glyceraldehyde 3-phosphate dehydrogenase GCL Ganglion cell layer GHT Geniculohypothalamic tract Glu Glutamate GnRH Gonadotropin-releasing hormone GPCR G-protein coupled receptor GRP Gastrin releasing peptide GSK 3β Glycogen synthase kinase 3β h Hour HAT Histone acetyl transferase HDL High density lipoprotein HDRS-17 Hamilton depression rating scale-17 HEK293 Human embryonic kidney 293 HEPES (4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid HPA Hypothalamo-pituitary-adrenal axis ICSD International classification of sleep disorders IGL Intergeniculate leaflet IML Intermediolateral nucleus INL Inner nuclear layer IPL Inner plexiform layer iprgc intrinsically photosensitive RGC ISWD Irregular sleep wake disorder LD Light-dark LEMS Low endogenous melatonin secretors xiv
15 LH Lateral hypothalamus m metre MAPK Mitogen activated protein kinase MD Major depression MDE Major depressive episode MEL Melatonin min Minute MinD Minor depression MPO Medial preoptic area mrna Messenger ribonucleic acid MT1/2 Melatonin receptor 1/2 M-type Morning type NA Noradrenaline nm nanometer nm nanomolar NMDA N-methyl-D-aspartic acid NO Nitric Oxide NPY Neuropeptide Y NREM Non-rapid eye movement sleep ONL Outer nuclear layer OPL Outer plexiform layer PAC1 Procaspase activating compound 1 PACAP Pituitary adenylate cyclase-activating peptide PAS Per (Period) -ARNT (Aryl hydrocarbon receptor nuclear translocator) - Sim (Single-minded) PCR Polymerase chain reaction Per Period PET Positron Emission Tomography pg Picogram PIH Prolactin-inhibiting hormone PKA/G/C Protein kinase A/G/C PRC Phase response curve PSG Polysomnography PT Pars tuberalis PVN Paraventricular nucleus QR2 Quinone reductase 2 qrt-pcr Quantitative reverse transcriptase polymerase chain reaction RAR Retinoic acid receptor RBD Recurrent brief depression REM Rapid eye movement sleep RGC Retinal ganglion cell RHT Retinohypothalamic tract xv
16 RIPA ROR RPE RRE RT-PCR RZR SCG SCN SD SDS-PAGE SE SEM SOL spvz SSS SWA SWS TAS-20 THAT TIM TSH TST Ub UV VAS VIP VLPO VMH VP VPAC WASO ZAS Radioimmunoassay precipitation assay RAR-related orphan receptor Retinal pigment epithelium ROR/RZR response element Reverse transcriptase polymerase chain reaction Retinoic Z receptor Superior cervical ganglion Suprachiasmatic nucleus Subsyndromal depression Sodium dodecyl sulfate polyacrylamide gel electrophoresis Sleep efficiency Standard error of mean Sleep onset latency Sub-paraventricular zone Stanford sleepiness scale Slow wave activity Slow wave sleep Toronto alexythimia scale-20 Toronto hospital alertness test Timeless Thyroid-stimulating hormone (Thyrotropin) Total sleep time Ubiquitin Ultra violet Visual analog scale Vasoactive intestinal peptide Ventrolateral preoptic nucleus Ventromedial hypothalamus Vasopressin Vasoactive intestinal peptide receptor Wakefulness after sleep onset Zung anxiety scale xvi
17 1 1 LITERATURE REVIEW 1.1 OVERVIEW Various aspects of human physiology and behavior exhibit near-24-hour rhythms defined as circadian rhythms. Oscillations in individual physiological processes can be regulated by tissue specific oscillators, however the synchronized oscillation of multiple processes is coordinated by a central pacemaker located in the suprachiasmatic nuclei (SCN) of the anterior hypothalamus 1-6. While the SCN can maintain near 24-h synchronicity in various physiological processes even in the absence of external environmental influences, the SCN can also respond to external environmental changes such as oscillations in the light-dark cycle and can reset the timing of physiological rhythms so that they occur at the most appropriate time of the day. However, improper time-keeping function by the SCN, either due to endogenous dysfunction of the SCN itself or aberrant alterations in exogenous environmental time cues can disrupt the temporal organization of individual physiological rhythms and lead to overall homeostatic disorganization 7,8. While the circadian disorder delayed sleep phase syndrome (DSPS) is likely mediated by an endogenous dysfunction of the pacemaker itself 9-12, shift work is a common cause for circadian disruption that is exogenously mediated due to repeated aberrant alterations of the light-dark cycle 13. The sleep-wake cycle, mood and cognition are under strong circadian regulation and show marked diurnal variation 14,15. Circadian rhythm disruption due to DSPS or shift work is associated with sleep disruption and significant impairment of mood and cognition 13,16. The studies presented in this thesis demonstrate that a significant reduction in nocturnal melatonin secretion, an endogenous circadian time cue, is associated with sleep and mood impairment. Furthermore, evidence will be presented supporting the use of exogenous melatonin to improve mood and sleep in individuals with delayed sleep phase syndrome, an endogenous
18 2 circadian disorder. In addition, a novel approach to minimize exogenously mediated circadian rhythm disruption will be presented. It will be demonstrated that spectral modulation of light, the strongest environmental time cue, attenuates nocturnal light exposure-induced alterations in endocrine, molecular and behavioral variables. 1.2 A BRIEF CHRONOLOGY OF CHRONOBIOLOGY In 1729, French astronomer Jean Jacques d Ortous de Mairan demonstrated that the upright movement of leaves at night and the lowering of the leaves during the day continued over several days even when the Mimosa pudica plant was maintained in constant darkness. This simple experiment suggested that the timely movement of the leaves was controlled by an endogenous clock instead of the movements being passive changes to sunlight 17. Another interesting experiment by French researcher Michel Siffre demonstrated that endogenous cycles may not follow an exact 24 h period. Siffre lived in an underground cave in 1962 for 60 days and again in 1972 for 205 days. However, at the end of the first experiment, he believed that only 43 days had passed instead of 60 days, and at the end of the second experiment, he believed that only 175 days had passed instead of 205 days 18,19. While Siffre had counted the number of days spent as the completion of each of his own rest-activity cycles, his consistent reports of a shorter number of days spent in the caves as compared to the number of days counted using the geophysical 24 h light-dark (LD) cycle suggests that endogenous cycles may be longer when not synchronized to the 24 h LD cycle 18,19. Biological oscillations are very common and cycle with different durations ranging from a few milliseconds in the case of neuronal responses to multiple years such as those found in animal population dynamics Nathaniel Kleitman defined a biological rhythm as a regularly recurring quantitative change in some particular variable biological process, irrespective of whether or not it takes place in a cell, tissue, structure, organism or population 23. Chronobiology
19 3 is the field of science that studies cyclical events in living organisms and their adaptation to solar and lunar related rhythms 24. Studies conducted in the 1950s on 24-h rhythmicity in fruit flies by Colin Pittendrigh and in humans and other animals by Jurgen Aschoff can be considered as the foundation for Chronobiology and circadian rhythm research. Work during the early 1970s formed the groundwork for our understanding of the genetic basis of circadian rhythms and of the neuroanatomical pathways regulating mammalian timekeeping mechanisms. The first biological clock mutants were generated using chemical mutagenesis in the fruitfly Drosophila melanogaster and the first clock gene Period (Per) was identified in The very next year, Stephan and Zucker and Moore and Eichler individually demonstrated the anatomical location of the endogenous clock in the rat to be in the suprachiasmatic nuclei (SCN) 26,27. The discovery of the circadian tau mutation in the Syrian hamster by Ralph and Menaker in 1988 introduced the use of mutagenesis and molecular biology to study biological rhythms in mammals and defined the SCN as a primary site in the mammalian brain for the generation of biological rhythmicity 28,29. Since then, several animal mutants have been generated and a whole host of seminal experiments using various experimental models have advanced our understanding of chronobiology and its role in maintaining organism homeostasis and disease pathogenesis. 1.3 CIRCADIAN RHYTHM QUALIFICATION Endogenous daily rhythms that follow a near 24 h period are called circadian rhythms originating from the Latin words circa and dies translating to approximately and day respectively. Circadian rhythms in all organisms share defining properties which include (i) a rhythm with a periodicity of about 24 h, even in the absence of an environmental cue (ii) the ability of the clock to be entrained in a time-dependent manner by environmental stimuli and (iii) compensation of period length for a range of changes in temperature 30. Almost all physiological
20 4 processes ranging from gene expression to overt behavior demonstrate circadian rhythms 2. Even complex physiological processes such as childbirth show circadian rhythms 31,32. Circadian rhythms are driven by the endogenous clock, a self-sustained oscillator, and their oscillatory amplitude (oscillatory strength) is not diminished over time in the absence of environmental cues. On the other hand, some biological rhythms, that also oscillate every 24 h, lose their oscillations once the environmental cues are removed and cannot be defined as circadian rhythms GENERAL PRINCIPLES OF CIRCADIAN RHYTHM BIOLOGY A linear model of circadian synchronization In its simplest form the circadian time keeping pathway can be depicted as a linear pathway composed of three components (Fig. 1.1) 33,34. The input component, composed of specialized receptors, receives and transduces environmental signals to the central component which is composed of pacemakers and oscillators. The central component generates rhythmicity autonomously, measures endogenous time and regulates temporal organization of physiological rhythms. The pacemakers respond to environmental cues and drive rhythmic outputs either directly or through other oscillators, whereas oscillators can drive rhythmic outputs but they cannot respond directly to environmental cues and depend on the pacemakers for their own synchronization 1,30. Efferent to the central component are the output pathways leading to effector systems that express overt physiological rhythms. However, biological systems are rarely as straightforward as models predict and the circadian system is not an exception. Numerous feedback cycles between the input/output pathways and the central time-keeping mechanisms exist that can modulate the sensitivity of the afferent and efferent pathways, also known as circadian gating, adding another level of complexity in the design of the overall system.
21 5 Figure 1.1: Linear model of entrainment. The circadian time keeping mechanism can be conceptually viewed as a linear pathway with three major components that enable entrainment of biological processes to environmental cues. Receptors detect changes in external cues which are then relayed through input signal transduction pathways to the central component composed of pacemakers and oscillators. Following integration of afferent signals, temporal cues are transduced to peripheral clocks through output signal transduction pathways. There can be numerous feedback operations between relay centers within the input and output signal transduction pathways as well as between the central pacemaker, receptors and relay centers modulating sensitivity of the pathway in a time specific manner (circadian gating) (Adapted from Roenneberg et al., ).
22 6 Nonetheless, the simplified linear model has powerful predictive capacity and is useful to convey the fundamental circadian mechanisms Core parameters of circadian rhythms The temporal synchronization of endogenous biological processes between each other as well as with external environmental cycles is critical to the health and survival of diverse organisms, from bacteria to humans. Entrainment is the process by which endogenous circadian rhythms maintain a stable and predictable phase relationship with external environmental cycles, such as the 24 h LD cycle 33. This also ensures that individual endogenous rhythms are synchronized to each other. Some of the key parameters of circadian rhythms are illustrated in Fig The amplitude of the cycle is the maximal deviation between the peak and trough of the circadian cycle and is often a measure of the strength of the cycle. A reduction in the strength or amplitude of the cycle often suggests a loss of circadian control over the particular biological process and likely of the overall circadian regulatory system which can lead to desynchronization or loss of entrainment between multiple physiological processes 35,36. The period of the rhythm is the time required to complete one cycle. In the absence of environmental time cues the period of the endogenous clock approximates 24 h (slightly more or less depending on the species) and is defined as a free running rhythm. Under entrained conditions, circadian rhythms are set to a period that matches exactly the period of the environmental cycle, usually 24 h. The phase of a circadian cycle is its relative position in reference to either another endogenous rhythm or another external environmental cycle. While the onset of activity can serve as an endogenous reference point, dawn or dusk can serve as external environmental reference points to measure the phase of a given circadian rhythm. The time difference between two rhythms is defined as the phase angle difference.
23 7 Figure 1.2: Fundamental circadian rhythm parameters. The amplitude of the cycle is the maximal deviation between the peak and trough of the circadian cycle (a). The period of the rhythm is the time required to complete one cycle. Under light-dark entrained conditions the endogenous period matches the period of the external time cue cycle, however under constant darkness the rhythms following the innate period of the endogenous clock, defined as the free running period (a). The phase of a circadian cycle is its relative position in reference to either another endogenous rhythm or external environmental cycle (a). Light exposure at night can phase shift endogenous rhythms in a time dependent manner (b).
24 8 Alterations in the timing of endogenous or exogenous cues induce changes in the phase of endogenous cycles and the magnitude of this change is defined as the phase shift. For example, exposure to light at night will shift the phase of the sleep-wake cycle relative to the 24 h clock. Furthermore, the direction of the phase shift depends on the timing of the stimulus. Since free running circadian periods deviate from the geo-physical 24 h LD cycle, it is helpful to give endogenous circadian rhythms their own time frame as they do not always correspond to the geo-physical LD cycle or night and day when under constant conditions. Under constant conditions, the half of the endogenous cycle that coincides with daytime when the animal is under an LD cycle is called subjective day and the other half is called subjective night. Exposure to light in the early part of the subjective night will cause a phase delay, that is, delay the onset of sleep and wake in the next cycle. Conversely, exposure to light in the late part of the subjective night or early morning will cause a phase advance, that is, advance the onset of sleep and wake in the next cycle (Fig. 1.2). Since entrainment allows an endogenous oscillator to adopt a stable and reproducible phase relationship with the environmental time cue through inducing reproducible degrees of phase shifts, the relationship of the phase shifts in the organism to the circadian timing of the stimulus can be generated as the phase response curve (PRC) 37. Depending on the strength or amplitude of the stimuli the resulting phase shifts and PRC can be classified as either strong (Type 0) phase resetting stimuli or weak (Type 1) phase resetting stimuli Models of entrainment The model of discrete pulses of light inducing phase shifts defines the nonparametric model of entrainment that is most widely accepted 39. The nonparametric entrainment principle can be summarized as daily corrections to the free-running period of the circadian system for the difference between the endogenous period and 24 h 39,40. The model of nonparametric
25 9 entrainment stems for the PRCs developed by Pittendrigh, Hastings and Rawson 39. However, in nature, exposure to time cues is seldom discrete or phasic and instead is usually continuous as in the case of light exposure in diurnal animals. While the nonparametric entrainment model works well for nocturnal animals who may be exposed to light strong enough to induce phase shifts in a phasic manner at dawn and dusk, diurnal animals are exposed to light in a tonic or continuous manner. Entrainment to continuous or tonic time cue stimuli is explained by the parametric model of entrainment 39. The parametric entrainment principle can be summarized as the continuous response of the circadian system to the intensity of light. Instead of phase shifts, the endogenous period itself changes by continuously accelerating or decelerating (angular velocity) as a function of the ambient light intensity 40,41. Thus, the nonparametric model of entrainment allows organisms to synchronize to environmental cues primarily by means of phase shifts in their endogenous cycle, whereas the parametric model induces continuous changes primarily in the period of the endogenous cycle. In addition to phase and period alterations, amplitude modulation has also been proposed as a means of entrainment 35,36. Though not widely studied or incorporated into models of entrainment, amplitude alterations in circadian rhythms may play an important role in pathological states as observed with circadian rhythm sleep disorders 42. However, the phase response model for nonparametric entrainment has been most widely accepted among chronobiologists, perhaps for its greater predictive capacity as a model, and has formed the basis for most of the work done in behavioral, physiological and theoretical chronobiology 39. In addition to clock mediated entrainment, environmental perturbations can directly exert potent changes in physiological variables that are otherwise regulated by circadian rhythms. For example melatonin suppression and inhibition of locomotor activity induced by nocturnal light exposure are direct effects that mask or hide clock controlled mechanisms. Since the effect is
26 10 observed in physiological parameters that are under strong circadian control, it may suggest that the observed changes are due to alterations in the circadian clock even though the response is often earlier than associated alterations in the clock mechanism. Nonetheless, the direct effects of light are likely mediated by the same neural and molecular pathways of the circadian clock, and may serve to fine tune the circadian entrainment mechanism 43, The circadian system is built on multiple circadian oscillators The circadian time-keeping mechanism starts at the level of individual cells. Cells in the kidney, liver, pancreas, heart, skeletal muscles and brain all contain the same set of core clock genes that are transcribed and translated rhythmically forming a 24 h oscillatory loop. These genes also regulate the expression of other cell/tissue specific genes (clock controlled genes) and this generates rhythmic tissue specific function However, in mammals, the SCN is the master pacemaker as SCN ablation abolishes rhythmic behavioral activity and disrupts the coordinated expression of clock genes in different organ systems 52,53. Time cues are relayed to peripheral organs from the SCN either through direct neural connections or by using hormones such as melatonin 54,55 and glucocorticoids 56. Such a hierarchical multioscillatory system allows for precise phase control and stability of the wide range of physiological systems that are under circadian regulation (Fig. 1.3). 1.5 THE OSCILLATORY MECHANISM Molecular oscillatory loops The set of core clock genes generate and maintain circadian rhythms under entrained and free running conditions (Fig. 1.4). In mammals, basic-helix-loop-helix Per-ARNT-Sim (bhlh- PAS) domain-containing transcription factors Clock (circadian locomotor output cycles kaput) and Bmal1 (brain and muscle ARNTL-like protein 1) act as positive regulators that drive the clock forward. Three Period proteins (Per1, Per2, and Per3) and two Cryptochrome proteins
27 11 Figure 1.3: Hierarchical organization of the biological clock. The mammalian suprachiasmatic nuclei (SCN) respond to changes in environmental time cues via dedicated receptor pathways in the case of photic cues and synchronize the timing of peripheral clocks by relaying temporal cues either using direct neuronal connections (solid arrows) or endocrine (gray spheres) and humoral (red spheres) factors. In the absence of the SCN connection peripheral oscillator rhythms are desynchronized and often damp out within a few cycles. Therefore, the peripheral clocks are slave to one endogenous circadian time set by the master clock (Adapted from Herzog and Tosini, ).
28 12 Figure 1.4: The mammalian circadian molecular clock. The circadian clock mechanism involves transcription-translation feedback loops comprised of a primary negative feedback loop involving the genes, Clock, Bmal1, Period 1(Per1), Per2, Cryptochrome 1(Cry1) and Cry 2. Clock and Bmal1 transcription factors activate transcription of the Per and Cry genes. The resulting Per and Cry proteins heterodimerize, translocate to the nucleus and interact with the Clock-Bmal1 complex to inhibit their own transcription. The negative control is removed by controlled degradation of the Per-Cry repressor complex and this activates a new cycle of transcription. The secondary autoregulatory feedback loop is composed of Rev-ERBα, which is also transcribed by Clock-Bmal1. Rev-ERBα feeds back to repress Bmal1 transcription and competes with a retinoic acid-related orphan receptor (ROR) to bind ROR response elements (RREs) in the Bmal1 promoter. In addition to the transcriptional activators and repressors, post-translational modification and degradation of circadian clock proteins are crucial steps for determining circadian periodicity. Key kinases for Per (and Cry) phosphorylation are casein kinase 1 delta (CSNK1D) and epsilon (CSNK1E). One of the roles for phosphorylation of clock proteins is to target them for polyubiquitylation and degradation by the 26S proteosomal pathway. (CCG, clock-controlled genes; CREB, camp response element-binding; E-box, CACGTG/T consensus sequence; MAPK, mitogen-activated protein kinase; Ub, ubiquitin).
29 13
30 14 (Cry1 and Cry2) operate as negative regulators that repress the clock 57,58. During the subjective day, Clock-Bmal1 heterodimers bind to E-box (CACGTG) enhancer elements on Per and Cry transcription promoter sites to activate their transcription 2. Following their transcription, Per and Cry proteins are translated in the cytoplasm and transported to the nucleus where they inhibit Clock-Bmal1-dependent transcriptional activation, resulting in a decrease in their own transcripts during the subjective dark period 59,60. As Per and Cry concentrations increase, regulated degradation of Per and Cry proteins leads to a restart of the activation and inhibition cycle of E-box mediated gene transcription, allowing the circadian oscillations of mrna and protein levels of both Per and Cry 1,2. This forms the major transcription-translation feedback loop. In addition to the major loop, a minor loop adds finer control to the molecular timekeeping mechanism. The clock protein Timeless (Tim) plays a role in the negative feedback arm of the clock oscillation mechanism by interacting with PER 61. In addition, bhlh transcription factors Dec1 (deleted in esophageal cancer 1) and Dec2 (deleted in esophageal cancer 2), which are transcribed by Clock-Bmal1, inhibit Clock-Bmal1 mediated transcription thereby acting as additional negative regulators in the feedback loop 62,63. Furthermore, the transcription of Bmal1 is regulated negatively and positively by the orphan nuclear receptors Rev-Erbα, and RORα respectively, in a competitive manner 64. This regulation results in circadian oscillation of Bmal1 expression in antiphase with the rhythm of Per expression. Furthermore, basic-leucine-zipper transcription factors Dbp (D site of albumin promoter (albumin D-box) binding protein) and E4BP4 (E4 promoter binding protein 4) activate Per1 and Per2 gene expression respectively and facilitate circadian clock repression 65,66. More recently, the Bmal1 paralog Bmal2 has also been shown to play a positive role in clock mediated
31 15 transcription by interacting with Clock 67. Moreover, Per2 homodimers have been shown to positively regulate Bmal1 transcription 58. Clock proteins also undergo extensive post-translational modifications including phosphorylation and ubiquitination 68. Casein kinase Iδ/ε (CKIδ/ε), MAPK (mitogen-activated protein kinase) and GSK 3β (glycogen synthase kinase-3β) phosphorylate several clock proteins including Per1, Per2 and Bmal1 to modulate their stability and/or function 69,70. Interactions between PER and CRY proteins regulate their nucleocytoplasmic localization and ubiquitinationmediated degradation 68. Similarly, the interaction between Clock and Bmal1 controls their nucleocytoplasmic localization, phosphorylation and degradation 71. Post-translational modifications play a major role in maintaining the 24 h periodicity of the molecular clock. While usual transcription-translation feedback loops generally operate on a timescale of several hours, reversible phosphorylation mediated regulation of important processes such as nuclear entry, formation of protein complexes and protein degradation, adds a length of delay in the system that keeps the period at near 24 h The master clock: the suprachiasmatic nucleus While clock genes are present in almost all cells and allow each tissue system to oscillate with its own phase, the SCN ensure proper temporal coordination between individual tissue specific oscillators 72. The central role of the SCN as the master pacemaker is demonstrated by SCN lesion studies. The homozygous tau mutant hamster has a characteristic period of about 20 h and when the SCN from these animals are transplanted into arrhythmic SCN-lesioned wildtype mice it restores a 20 h period 28,29. Moreover, transplanting wild type hamster SCN into SCN-lesioned tau mutant hamsters restores a wild type period (24 h) in tau mutant hamsters that originally had a 20 h period 29. These transplantation studies demonstrate that the circadian period is an intrinsic property of the SCN. Furthermore, transplantation of SCN grafts from wild-type
32 16 mice into Cry1/Cry2 double knockouts, which are congenitally arrhythmic, restores behavioural rhythmicity, demonstrating that the SCN is able to control rhythmic behaviour independent of any other putative master clocks Organization of the SCN Morphologically, the SCN can be divided into the dorsomedial shell and the ventrolateral core, based on differential neuropeptide content 74 (Fig. 1.5). In the rat, most of the neurons in the shell synthesize vasopressin (VP), whereas neurons in the core synthesize vasoactive intestinal peptide (VIP), peptide histidine isoleucine (PHI) and/or gastrin releasing peptide (GRP). A smaller proportion of somatostatin-producing neurons are found in between these two cell populations. In addition, γ amino-butyric acid (GABA) is present in most SCN neurons 74. Neurons in the core receive glutamatergic (Glu) input from the retina via the retinohypothalamic tract (RHT), neuropeptide Y (NPY) input via the geniculohypothalamic tract (GHT) originating from the intergenicular leaflet (IGL), and serotonergic (5-HT) input from the raphe nuclei. Neurons in the dmscn shell receive non-photic input from the cortex, basal forebrain and hypothalamus 75. The core is densely innervated by afferent fibers, and the shell contains more efferent fibers than the core. Most of the latter project to other hypothalamic areas regulating neuroendocrine and autonomic functions 76,77. Such morphological and topographical differences support the possibility of the two parts playing differential roles in the generating and regulating circadian rhythms. 1.6 CIRCADIAN RESPONSE TO LIGHT (PHOTIC) STIMULI Evolution of circadian photic response Nearly all organisms, from the simplest algae to mammals, use sunlight to adjust their period of activity to optimize survival and light is the strongest time cue in mammals including
33 17 Figure 1.5: A schematic representation of the SCN and its efferent and afferent pathways. The SCN are located in the anterior hypothalamus above the optic chiasm and receive afferent signals from the cortex, limbic and visceral pathways (a). Though direct SCN innervations are limited to the subparaventricular zone (spvz) and the paraventricular nucleus (PVN), the SCN pathways regulate coherent timing of diverse physiological functions, including endocrine rhythms, sleep-wake cycle, feeding behavior, and thermoregulation. The SCN output to the spvz is relayed to the medial preoptic region (MPO) to control circadian rhythms of body temperature; indirect projections through the dorsomedial nucleus of the hypothalamus (DMH) to the lateral hypothalamus (LH) controls behavioural rhythms of feeding; similarly indirect pathway to the ventrolateral preoptic nucleus (VLPO)via the DMH may regulate sleep (b). The SCN can be morphologically and functionally divided into the core region, which receives photic stimuli through direct projections from intrinsically photoresponsive retinal ganglion cells via the retinohypothalamic tract (RHT), and the shell region which mostly makes efferent projections to hypothalamic homeostatic centers (c) (Adapted from Takahashi et al., ).
34 18
35 19 humans. The evolutionary pressure for coupling the circadian clock to light has been so strong that even in a simple organism such as the zebrafish, light can directly affect clock mechanisms in organs or muscle tissue 79. In lower order animals such as fruit flies and zebrafish light is able to penetrate all tissues and almost all parts of the body have pacemaker cells, with a higher concentration in the retina 80. In contrast, light-receptive cells in birds, are present in the retina and pineal gland 81 while in mammals the light signal can only reach the central nervous system (CNS) via the retina 82. In mammals, pacemaker cells are only present in the retina and the SCN. Light was not always considered to be the dominant circadian time cue in humans. Work in the 1960s and 1970s suggested that social non-photic cues were most important in humans 83. Food and exercise have been established as potent time cues in animals and similar to light and melatonin, an exercise PRC has also been generated When food is restricted to certain times of the day animals display increased activity prior to feeding time, which persists in the absence of food on consecutive days. This suggests that the anticipatory response is generated endogenously. Based on such observations it has been proposed that there exists an independent food entrainable oscillator (FEO) 86. Such anticipatory behaviors occur even in SCN lesioned animals 87 suggesting that the FEO is independent of the SCN. The exact neural location of the FEO has not yet been identified and the role of canonical clock genes is debatable at best, and results from recent studies taken together suggest that the FEO is a complex of central and peripheral oscillators driven by metabolic oscillations 86,87. Nevertheless, light remains to be the dominant circadian time cue in mammals, especially humans. Light exposure can induce strong (type 0) resetting of the human circadian pacemaker affecting both oscillatory amplitude and phase 6. The photic response of the circadian pacemaker depends on the timing, intensity, duration and history of light exposure 6, Light pulses can also induce singularity behavior
36 20 which is a loss of circadian amplitude (strength) resulting from desynchronization of SCN activity Mammalian circadian phototransduction begins at the retina Visible light is electromagnetic radiation that activates retinal photoreceptors and generates vision. Visible light is a portion of the larger electromagnetic spectrum and falls within the nm wavelength range 93. The different wavelengths correspond to different colors ranging from violet ( nm) and blue ( nm), which constitute the short wavelength portion of visible light, to orange ( nm) and red ( nm), which constitute the longer wavelength portion of the visible spectrum 94. In mammals all photoreceptors are present in the eye and therefore the retina is of fundamental importance for entrainment to the LD cycle. The retina is made up of five major classes of neurons (photoreceptors, horizontal cells, bipolar cells, amacrine cells, and ganglion cells) and glia (Fig. 1.6). The structural organization of the retina comprises three nuclear layers which include the outer nuclear layer (ONL), inner nuclear layer (INL), and ganglion cell layer (GCL). The retina is apposed to the pigment epithelium (RPE) that lines the back of the eye. Photoreceptors, bipolar and horizontal cells make synaptic connections with each other in the outer plexiform layer (OPL), whereas the bipolar, amacrine and ganglion cells make contact in the inner plexiform layer (IPL). In most of the retina, light passes through layers of nerve cells (GCL and INL) and their processes (IPL and OPL) before reaching the photoreceptors 97. Following photoreception, signals are transduced vertically from photoreceptors to bipolar cells and to ganglion cells. In addition, signals are transduced laterally mediated by horizontal cells in OPL and amacrine cells in the IPL 96,97. Traditionally, vertebrate photoreceptors in the eye were thought to be of only two classes: the rods and cones. Normally there are more rods than cones and rods are also more sensitive than cones. Therefore, rods are
37 21 Figure 1.6: A schematic representation of the mammalian retina and photic signal transduction pathway. The retina contains five major classes of neurons (photoreceptors, horizontal cells, bipolar cells, amacrine cells, and ganglion cells) arranged into three nuclear layers: outer nuclear layer (rods (R) and cones(c)) (ONL), inner nuclear layer (bipolar (B) and amacrine (A) cells) (INL) and ganglion cell layer ((G)ganglion cells) (GCL). The retina is apposed to the pigment epithelium (RPE) that lines the back of the eye. Photoreceptors, bipolar and horizontal cells make synapses with each other in the outer plexiform layer (OPL). The bipolar, amacrine and ganglion cells make contact in the inner plexiform layer (IPL). In most of the retina, light passes through layers of nerve cells (GCL and INL) and their processes (IPL and OPL) before reaching the photoreceptors. After photoreception, information flows vertically from photoreceptors to bipolar cells to ganglion cells, and laterally mediated by horizontal cells in OPL and amacrine cells in the IPL. Ganglion cells form the major conduit for signal transduction. Only a small subset of ganglion cells express melanopsin making them intrinsically photosensitive (iprgc). Following photic stimuli exposure, the signal is transduced to the suprachiasmatic nuclei (SCN) via the retinohypothalamic tract using PACAP and glutamate as neurotransmitters which induces phosphorylation of camp response element binding protein (CREB) and leads to Period gene expression and associated phase shifts (Adapted from Hankins et al., ).
38 22 used for detecting light at low intensities (scotopic vision) whereas cones are used for detecting light at higher intensities (photopic vision). There are three classes of cones (short, mid, and long wavelength) that differentially respond to the various wavelengths incident on the retina and thereby facilitate colour vision. However, more recently, a third class of photoreceptor was identified which may play a minimal role in vision, but acts as the principal circadian photoreceptor 99, Dedicated circadian photoreceptor In mammals, photic resetting is abolished by bilateral enucleation suggesting the circadian photoreceptor is localized solely within the eye. Importantly, visually blind mice lacking both rod and cone photoreceptors, have normal photic circadian resetting, suggesting that circadian photic response is not dependent on the traditional rods and cones but likely uses a dedicated photoreceptor 101,102. Recent studies strongly point to melanopsin, a novel opsin-like protein as the photopigment in the circadian photoreceptor 102. Melanopsin was originally identified as a photopigment expressed in Xenopus skin melanophores 103, and later melanopsin expression was found in a subset of retinal ganglion cells (RGCs) but not in rods and cones 104,105. The melanopsin-containing RGCs extend axons directly to the SCN and this projection forms the RHT 106. While ganglion cells are common in the retina, only 1-2% of RGCs express melanopsin and are therefore intrinsically photosensitive (iprgc). Despite the limited expression of melanopsin in RGCs, the iprgcs form a diffuse photosensitive layer that covers almost the entire retina 104,106. In melanopsin knockout mice, the intrinsic photosensitivity of the RGCs is eliminated, supporting the role of melanopsin as the photopigment that confers photosensitivity to RGCs 107. However, the melanopsin knockout mice are still able to entrain to light-dark cycles, although photic resetting is greatly attenuated in these animals 108. This highlights a possible
39 23 contribution of rod and cone photoreceptors in the circadian phototransduction. Mice lacking both the functional rod-cone system and melanopsin exhibit complete loss of the photic resetting indicating that the visual rod-cone photoreceptors and non-visual melanopsin expressing RGCs together accounts for all circadian photoreceptors Resetting of the central clock by light Photic signals are transduced to the SCN via the RHT using glutamate and PACAP, which are colocalized in the RHT 1 (Fig. 1.6). Glutamate and PACAP activate several intracellular signaling cascades in the core SCN neurons including nitric oxide, protein kinase A, protein kinase G and MAP kinase pathways, all of which converge on the transcriptional activator camp response element binding protein (CREB) 110. Activated CREB binds to campresponse elements (CREs) at the promoter sites of the clock genes Per1, Per2 and Dec1 63,111. Light-induced CREB activation and the subsequent de novo synthesis of the corresponding clock proteins result in prolonged suppression of Clock-Bmal1 activity by Per, Cry and Dec protein complexes. Light-induced suppression of Clock-Bmal1 transcriptional drive allows entrainment of the molecular clock to the LD cycle 112. Interestingly, Per1 mutant mice have attenuated phase advances in response to early morning light exposure with no impairment of phase delays in response to early night light exposure, whereas, Per2 mutant mice demonstrate the reversed trend 113. This suggests that mammalian Per isoforms may play critical and differential roles in regulating light mediated circadian phase shifts instead of merely adding robustness to the system through redundancy Unique spectral sensitivity of circadian photic response To determine the differential sensitivity of biological responses to optical wavelengths, an action spectrum can be generated. Action spectra can be either polychromatic or monochromatic (analytic) 114. Polychromatic action spectra are developed by using broader
40 24 optical wavelength ranges (bandwidth), often greater than nm, whereas monochromatic action spectra are generated using narrow bandwidth (between 5-10 nm) light 114. Early circadian action spectrum studies using polychromatic stimuli examined the effects of light exposure on pineal melatonin synthesis, core body temperature (CBT) phase shifts and photoperiodic responses These studies strongly suggest that the spectral region between 450 nm and 550 nm provides the strongest stimulation of circadian and neuroendocrine responses in mammals with minimal responsiveness to the long wavelength end of the spectrum above 550 nm. Although, at very high intensities, long wavelength light in the red portion of the spectrum can acutely suppress melatonin, as well as phase shift or entrain circadian rhythms in both rodents and humans 118. At the very low end of the spectrum, near-ultraviolet radiation (UV- A: nm) has little to no effect on rodent circadian photic response. In adult humans the ocular lens does not transmit UV radiation to the retina and therefore has minimal effects on circadian responses 119. While polychromatic action spectra are useful, they are limited in determining peak sensitivities of a biological response as a function of optical wavelengths, thus analytic (monochromatic) action spectra must be used. By using narrower bandwidths the peak sensitivity of a biological response to one wavelength can be determined. Monochromatic action spectra studies confirmed the short wavelength sensitivity of circadian responses determined by polychromatic action spectra studies and also determined the maximal photosensitivity range to be between 459 to 480 nm, corresponding to blue light EFFECTS OF LIGHT EXPOSURE ON CIRCADIAN ENDOCRINE PATHWAYS Classical circadian endocrine pathways Several endocrine rhythms have been shown to have robust circadian rhythmicity, including, but not limited to, adrenocorticotropic hormone (ACTH), gonadotropin releasing
41 25 hormone (GnRH), corticotropin releasing hormone (CRH), thyrotropin (TSH), leptin and testosterone The two endocrine pathways that have been extensively studied for their circadian regulation are those of melatonin and glucocorticoid secretion. Both pathways are under the control of the SCN and are often used as endocrine markers of circadian function. Alterations in their secretion amounts (amplitude) or timing (phase) can be used as markers for evaluating changes in the endogenous circadian pacemaker induced by environmental changes. Light exposure induces alterations in both endocrine pathways in rodents and humans making them effective markers to evaluate light induced alterations in circadian function Hypothalamo-Pituitary-Adrenal axis Glucocorticoids are essential for the maintenance of homeostasis and enable organisms to respond to and manage stress. While cortisol is the principal glucocorticoid in humans, corticosterone is the principal glucocorticoid in rats. Both are synthesized in the adrenal cortex and their secretion is governed by the hypothalamus and pituitary 130. In response to stress (disruptions to the homeostatic balance of the organism), the hypothalamus releases CRH and VP from parvocellular neurons projecting from the paraventricular nucleus (PVN) to the median eminence. These neurohormones are released into the hypothalamic-hypophyseal portal vessel targeting the anterior pituitary where they act synergistically to trigger the release of ACTH from the corticotrope cells into the systemic circulation. In turn, ACTH acts on the adrenal cortex to initiate the synthesis of cortisol or corticosterone, which are released immediately into the systemic circulation by diffusion 124,130. Hypothalamic CRH and VP, pituitary ACTH and adrenal glucocorticoids comprise the hypothalamic-pituitary-adrenal (HPA) axis. However, comparing ACTH and corticosterone responses under various experimental conditions reveals that the circadian release of corticosterone does not depend solely on the release of ACTH, contrary to the stress induced release of corticosterone 131. Tracing studies have
42 26 revealed a multisynaptic pathway from the SCN to the adrenal gland, which passes via preautonomic PVN neurons to the intermediolateral column (IML) of the spinal cord. The preautonomic PVN neurons contact sympathetic preganglionic neurons in the IML, which then innervate the adrenal gland through the splanchnic nerve 132. Interestingly, this pathway does not overlap with the SCN connections to the PVN via the spvz and DMH described above, indicating a separation of the autonomic from the HPA axis targeted pathways of SCN control at the level of the PVN 132. This autonomic neural pathway has been shown to be critical for transmitting photic stimuli to the adrenal gland and mediating changes in glucocorticoid release independent of the HPA axis 132,133. In addition, this pathway modulates the adrenal sensitivity to ACTH and likely adds another dimension of control over regulating circadian rhythms of corticosterone release directly by the master circadian clock 134. Furthermore, in the rat, the current model proposed for circadian regulation of corticosterone release suggests that the increased release of endogenous VP from the SCN during the first part of the light period causes corticosterone levels to remain at low basal levels during the initial part of the sleep period 135. Subsequently, the concomitant arrest of VP release and the increased release of an additional SCN transmitter (not yet identified) that can stimulate corticosterone release during the second part of the light period results in the daily corticosterone surge just before awakening 124,135. After the onset of the dark period, the release of the putative stimulatory SCN transmitter decreases and corticosterone levels slowly decline. However, the circadian peak in glucocorticoid release is locked to the activity phase, that is, it occurs in the early morning in diurnal animals (humans) and in the early night in nocturnal animals (rats) 124. Interestingly, the inhibitory role of VP in the nocturnal rat is reversed in the diurnal rodent Arvicanthis ansorgei 136. Furthermore, the timing of clock gene expression, neural activity and transmitter release by the SCN are similar in both species, whereas the effect on the
43 27 brain targets is different 136. Therefore, it has been postulated that the SCN projections likely contact different interneurons in the spvz and/or DMH areas, inhibitory GABAergic neurons in the case of the rat and excitatory glutamatergic neurons in A. ansorgei, which then exert their species specific effects on PVN neurons. Light exposure has differential effects on glucocorticoid release in a manner which is sensitive to both time-of-exposure and duration-of-exposure. In rats, a rapid 5 min nocturnal light exposure induces a fast repression of corticosterone release when given at the beginning of the night, but not later in the night or at the beginning of the day 132. However, a longer 1 h nocturnal light exposure given at the beginning of the night significantly increases corticosterone levels 133. Importantly, in both cases, the changes in corticosterone are not accompanied by changes in ACTH and SCN ablation abolishes the light dependent glucocorticoid release demonstrating that the photic response must be mediated independent of the HPA axis 132,133. Furthermore, light exposure increases adrenal nerve activity, but only if the SCN is intact, and adrenal denervation blocks the light induced corticosterone release 133. In addition, the signal to the adrenal cortex likely involves epinephrine release by the adrenal medulla after the light pulse to induce glucocorticoid release 133,137. Several human studies have demonstrated that early morning light exposure can significantly increase cortisol secretion above levels normally associated with arousal However, short duration (1 h) light exposure or even moderately long duration (6.5 h) light exposure in the early evening does not increase cortisol secretion 139,141. Taken together, the reports suggest that light exposure around the usual time of cortisol secretion onset, it may increase glucocorticoid secretion. While no action spectra studies exist for the spectral sensitivity of glucocorticoid secretion to nocturnal light exposure either in humans or rats, one study did compare glucocorticoid secretion after exposure to discrete 6.5 h monochromatic 460 nm and
44 nm pulses and found no significant differences 141. However, based on the SCN mediated control of glucocorticoid release from the adrenal cortex in response to light exposure, and the short wavelength sensitivity of SCN photic responses, it may be that glucocorticoid secretion also has short wavelength sensitivity to photic stimuli Hypothalamo-Pineal axis Melatonin is synthesized in the pinealocytes in the pineal gland and is secreted during the subjective night, only under darkness, in both nocturnal and diurnal animals 142. In mammals, the pineal is not directly light sensitive instead a polysynaptic pathway transmits photic stimuli from the retina to the pineal 143. Photic signals are transduced from iprgcs via the RHT to the SCN. The signal is further relayed from the SCN to the autonomic division of the PVN connecting it to the superior cervical ganglion (SCG) and then to the IML column of the upper thoracic cord. From the IML noradrenergic postganglionic fibers innervate the pineal gland and norepinepherine released from these fibers induces melatonin synthesis by the pinealocytes 142. The effects of light on melatonin have been extensively studied. Light exerts two distinct effects on melatonin production. First, light exposure at night affects both melatonin biosynthetic and catabolic pathways. Nocturnal light exposure rapidly decreases the activity of the rate limiting enzyme in melatonin biosynthesis, serotonin N-acetyltransferase (AA-NAT), decreases circulating melatonin levels and decreases the levels of the major metabolite 6- sulfatoxymelatonin (amt6s) 142,144,145. This suppressive effect can be caused by full-spectrum white light, monochromatic visible light, as well as with near ultraviolet radiation (UV-A) exposure, although the UV-A exposure mediated suppression is species specific. Melatonin suppression is maximal by short wavelength light peaking around 480 nm, which corresponds to the maximal activation range of iprgcs 114,120,121. The amount of light required to suppress melatonin production during the night is also species specific, generally the murine system being
45 29 much more sensitive than in humans 146. The second effect of nocturnal light exposure is altered timing of melatonin secretion. While light exposure late in the subjective day and early in the subjective night delays the phase of the melatonin circadian rhythm, exposure during the second half of the subjective night produces a phase advance of the rhythm 147,148. While there is some evidence that supports the role of VP in regulating melatonin release from the pineal gland, similar to glucocorticoid regulation by the SCN, most studies now support the role of the inhibitory neurotransmitter GABA in mediating melatonin secretion regulation by the SCN. Infusion of muscimol, a GABA agonist, in the DMH during the onset of darkness prevents the nocturnal increase in plasma melatonin levels, whereas infusions later in the night cause an immediate inhibition of melatonin release 149. Nocturnal light exposure causes the release of GABA in the vicinity of the PVN and inhibits those PVN neurons that are in control of the final part of the pineal-activating pathway 150. Infusion of the GABA antagonist bicuculline to the PVN in the middle of the subjective day evokes pineal melatonin release to levels equivalent to those at night 151. However, the excitatory neurotransmitter glutamate also plays a critical role in maintaining melatonin levels. Blockade of the glutamatergic input from the SCN to the PVN in the middle of the night significantly decreases melatonin levels 151. Taken together, the proposed model suggests that the SCN uses a combination of daytime GABA-ergic inhibitory and nocturnal glutamatergic stimulatory signals toward the PVN-pineal pathway to control the daily rhythm of melatonin synthesis 150,151. However, there are additional players that have not yet been identified, since blockade of GABAergic signaling in the PVN at dawn does not prevent the early morning decline of melatonin completely Melatonin as an endogenous neuroendocrine time cue Melatonin production has been demonstrated in several tissues however, the circulating melatonin is derived mainly from the pineal organ 142,152. Melatonin is an indoleamine, with low
46 30 molecular weight (mol. wt ), it is lipophilic and is rapidly carried by blood and cerebrospinal fluid to all tissues of the organism. Its production is tightly restricted to the dark phase of the LD cycle 153. The duration of the nocturnal melatonin peak reflects the duration of the photoperiod, that is, the short days that define winter result in long duration of the melatonin peak. As night length changes over the annual cycle, the alteration in melatonin profile induces profound changes in the reproductive state of seasonally breeding animals 142. Melatonin levels follow a robust circadian rhythm and in both nocturnal and diurnal animals melatonin secretion occurs under darkness during the subjective dark phase. While this corresponds to habitual sleep times in humans, in rats, which are nocturnal animals, melatonin secretion corresponds to a time of increased activity. This suggests that melatonin may act as a neuroendocrine signal that relays temporal cues rather than acting as a sleep promoting agent. Although pinealectomy has minimal effect on circadian activity rhythms in mammals, pinealectomized rodents entrain to a reversed photoperiod regimen more quickly than intact animals, suggesting that melatonin adds resilience to perturbations in the circadian system 154,155. In addition, pinealectomy decreases the amplitude of the firing rate rhythm in the SCN, providing additional support to the role of melatonin as a stabilizer of circadian output strength 156. Daily melatonin administration can entrain activity rhythms in free running rats 157. Importantly, melatonin administration cannot entrain free running rhythms in SCN ablated rats 157. The SCN is a major site of melatonin binding in the rodent brain and the SCN also exhibits circadian variation in the density of melatonin binding sites 158,159. Even subcutaneous melatonin injection alters the metabolic activity of rat SCN in a time dependent manner 160. Single unit activity in rat and hamster SCN brain slices is acutely suppressed when melatonin is applied near the transition to subjective night 161,162. Taken together, these studies strongly suggest that melatonin can entrain circadian rhythms, and that it acts at the level of the SCN.
47 31 In humans, melatonin administration during the subjective day induces earlier sleep onset and generates longer sleep duration Daily melatonin ingestion can entrain free running circadian rhythms in blind individuals and pharmacological suppression of nocturnal melatonin secretion increases total wake time and concomitantly decreases both Non-Rapid Eye Movement (NREM) Sleep and Rapid Eye Movement (REM) Sleep 170. The circadian rhythm of plasma melatonin also has a temporal association with circadian rhythms observed in cortical EEG activity during sleep in humans 171, suggesting a direct influence of melatonin on sleepwake regulation. Similar to light, a physiological dose of orally administered melatonin shifts circadian rhythms in humans according to a PRC. However, the melatonin PRC is about 12 h out of phase (opposite phase) to that of the light PRC 148. Melatonin delays circadian rhythms when administered in the morning and advances them when administered in the afternoon or early evening 172. Based on the melatonin PRC, exogenous melatonin administration can be timed to induce phase alterations in individuals with underlying circadian disorders 148,173,174. Melatonin acts through two high affinity G protein-coupled membrane bound receptor isoforms MT1 and MT2 and may also act on a nuclear receptor from the retinoic acid orphan receptors family, RZR (retinoic Z receptor) and ROR (RAR-related orphan receptor) 142,175. In addition, a third isoform of the melatonin receptor, MT3, has been identified and characterized as the enzyme quinone reductase 2 (QR2) 176 however, the physiological role and significance of melatonin acting upon QR2 remains elusive 177. Melatonin receptors are widely expressed both centrally and peripherally 178,179. In humans, melatonin membrane-bound receptors are found in the SCN and pars tuberalis (PT), cerebellum, brain blood vessels, kidney and also prostate 142. Melatonin regulates different second messenger cascades positively or negatively in a tissue specific manner and regulates a wide range of physiological processes ranging from cell cycle
48 32 regulation to insulin secretion 175,180,181. However, certain effects of melatonin on particular genes have significant bearing on its role as a circadian rhythm modulator. One of the best studied effects of melatonin on modulating second messenger pathways is the inhibition of adenylyl cyclase (AC) which inhibits camp (cyclic adenosine monophosphate) generation, and inhibition of protein kinase A (PKA) activity, ultimately inhibiting CREB mediated gene transcription 175. In ovine PT cells, which are thought to mediate some of the melatonin regulated seasonal changes, melatonin blocks the forskolin-stimulated increase in the levels of immediate early genes c-fos and junb, both key factors in regulating neuronal activity in the mammalian master clock as well as photic resetting 182,183. Furthermore, Per1 is campresponsive and its expression is repressed by the melatonin-induced inhibition of camp production 184. Furthermore, AC inhibition by melatonin over a period of several hours sensitizes AC to subsequent stimulation by activators of the enzyme 185. This AC hypersensitization allows Per1 to be transcribed at higher levels when melatonin is removed from the system; however, in the absence of melatonin, this AC hypersensitization is lost and Per1 transcription amplitude is blunted suggesting the importance of melatonin in regulating circadian oscillatory strength even at the molecular level 186. Importantly, melatonin also inhibits the pituitary PACAP induced CREB phosphorylation in the SCN via MT1/MT2 receptors 187,188. As discussed, PACAP is critical in transducing photic signals to the SCN and such a function of melatonin suggests an integral role for melatonin to regulate circadian rhythms, even at the level of input signal transduction pathways Melatonin as an endogenous circadian phase marker It is not possible to directly measure SCN activity in humans due to the level of invasiveness involved. Instead, pacemaker activity in humans is assessed indirectly by studying physiological variables directly controlled by the SCN. In principle, any physiological variable
49 33 that is regulated by the SCN can be used to measure endogenous circadian phase; however, masking effects that can directly modulate the variables independent of circadian pacemaker alterations must be negated. While numerous physiological processes are regulated by the SCN, the critical constraint of removing masking effects greatly limits the choices of possible biological parameters that can be used to measure circadian phase. The sleep-wake cycle itself is a rough indicator of circadian phase, but it can be easily masked by homeostatic sleep need and prior wakefulness. Wake up time provides a relatively accurate estimate of circadian phase in subjects allowed to sleep on an ad libitum schedule, but under 24 h entrained conditions 189,190. Another important necessity for such a marker of circadian phase is the possibility of frequent serial sampling. Without enough data points the resolution of the marker to detect alterations in circadian phase is greatly diminished. In the past, one of the most commonly used circadian phase markers was the CBT rhythm but similar to sleep, the circadian signal from the CBT rhythm can be easily masked by activity, food intake and by sleep. CBT serves well when all possible sources of externally mediated perturbations of the circadian clock are held constant 191. While this technique is very useful and provides sufficient resolution for use in research, implementation in clinical protocols is difficult. Since melatonin secretion is also under the direct control of the circadian pacemaker and is less sensitive to environmental/behavioural perturbations it is becoming increasingly more popular as a circadian phase marker Another major advantage of using melatonin is that it is readily measurable in saliva and hence provides a convenient, non-invasive method for sampling. Furthermore, salivary melatonin level, although lower than in plasma, is reliably detectable, and strongly correlated 195. Its adaptation has also been facilitated by the commercial availability of immunoassays with sufficient sensitivity and specificity. It is also possible to
50 34 measure the major metabolite of melatonin, amt6s, in urine using commercially available immunoassays 192. Different parameters of the melatonin secretion rhythm such as time of peak value, time of onset and offset, mid-point of the rhythm curve, can all be used to determine circadian phase 196. However, most studies have used the dramatic elevation in melatonin levels in the evening compared to low baseline daytime levels for determining circadian phase. Since, melatonin secretion is suppressed by light exposure (a masking effect), samples need to be obtained under darkness or dim light (less than 5 Lux) conditions, and thus the procedure is often termed the dim light melatonin onset test and the reference point on the rhythm curve is referred to as dim light melatonin onset (DLMO). It has also been reported that posture and drugs (such as beta-blockers, NSAIDS, and caffeine) may influence melatonin levels and thus may mask, to some degree, the melatonin rhythm; however, such confounders can be controlled for with greater ease than those affecting the sleep-wake cycle and CBT There is high level of correlation between CBT (using constant routine conditions) and DLMO 5,201. Even repeated measures of circadian phase on three occasions, spaced five days apart, in the same individuals demonstrated lowest standard deviation using melatonin as a marker compared to CBT and cortisol 202. These findings provide strong support for the melatonin profile as the most stable and, therefore, presumably the most accurate, currently available marker for circadian phase. In a clinical evaluation, DLMO test showed high sensitivity and specificity in phase typing individuals with circadian disorders compared to individuals who manifested similar sleep pathology without any underlying circadian misalignment 203. In addition to such endogenous biological markers, behavioral markers can also be used to assess global patterns of sleep-wake activity such as sleep diaries and actigraphy. While such methods are useful for longitudinal studies where daily serial sampling, even from saliva, may be
51 35 demanding both on the study participants and researchers, there are no standardized widely accepted sleep diaries and wearing actigraphs regularly for long durations can lead to low compliance. 1.8 EFFECTS OF LIGHT EXPOSURE ON CIRCADIAN BEHAVIORAL PATHWAYS Sleep Sleep is more than a passive rest phase; it is a dynamic process when certain brain regions show the same (or increased) activity as during wakefulness 204,205. According to the flipflop model of sleep regulation, the neurobiological maintenance of sleep and wakefulness is controlled by two mutually inhibiting neural networks, the ascending arousal system (AAS), which maintains wakefulness, and the ventrolateral preoptic nucleus (VLPO), which promotes sleep 206,207. Monoaminergic nuclei, including the locus coeruleus, the serotonergic dorsal and median raphe nuclei and the histaminergic tuberomammilary neurons constitute the AAS and promotes wakefulness by direct excitatory effects on the cortex and inhibit sleep promoting neurons of the VLPO. During sleep, the VLPO inhibits monoaminergic-mediated arousal regions through GABAergic and galaninergic projections. Orexin (hypocretin) containing neurons in the lateral hypothalamus play an important stabilizing role in regulating sleep-wake transitions by reinforcing the arousal system However, the precise mechanism of sleep-wake transition regulation is yet to be determined. It may be that with progressively increased durations of wakefulness, sleep-promoting (somnogeneic) substances accumulate that enhance the activity of sleep-promoting cells and reduce the activity of wake-promoting neurons. The range of such somnogenic substances includes neuropeptides, cytokines and hormones and amongst these, adenosine and adenosinergic signalling pathway are currently the most promising candidates 207,208.
52 36 The active neural process of sleep can be measured non-invasively by polysomnography (PSG) which breaks down sleep into different stages based on neural activity. PSG includes three electrophysiologic measures; the electroencephalogram (EEG) measuring brain activity, electromyogram (EMG) measuring muscle tone, and electrooculogram (EOG) 209 measuring motion of eyes. These measures are used to divide sleep into two broad categories of REM and non-rem sleep. REM periods occur in cycles of approximately 90 minutes throughout the night, with more REM toward the end of the major sleep episode. While REM sleep distribution shows a significant ultradian pattern, several studies have shown that the temporal distribution of REM to be shifted to the latter part of the night is under circadian regulation Consequently, circadian misalignment affects REM sleep and depressed individuals who often have underlying circadian misalignment as well often show alterations or reversal in REM sleep distribution compared to healthy individuals, that is, in depressed individuals more REM occurs in the early part of the sleep episode instead of the later part 213,214. Non-REM sleep is further subdivided into Stages 1, 2, 3, and 4. Stages 3 and 4, also called delta or slow-wave sleep (SWS), represent the deepest sleep in humans 209,210,212. The intensity of SWS is measured as Slow Wave Activity (SWA) which also serves as a measure of sleep depth. The amount of SWA increases in recovery sleep following sleep deprivation. There are several aspects of sleep, including the continuity, timing, and patterning of different stages of sleep that influence sleep quality 215,216. According to the two-process model of sleep, the timing of sleep is regulated by two dynamic and independent processes, namely the circadian drive for wakefulness (Process C) and the homeostatic sleep drive or the need for sleep (Process S) 8. Process C is driven by endogenous pacemaker whereas Process S is driven by the homeostatic need for sleep based on prior wakefulness. While the two processes may be independent, they heavily interact with significant commonalities at the molecular level 217.
53 37 During the day Process C opposes Process S to promote wakefulness and counter the exponentially growing pressure of Process S due to increasing durations of wakefulness. Optimal sleep occurs when maximal sleep pressure is synchronized to the decline of Process C 8,54. However, circadian alterations, as can be caused by nocturnal light exposure, can change the timing of Process C and concurrently imposed time restrictions due to social opportunities can easily force individuals to sleep at a time when the two processes are not properly synchronized for promoting highest sleep quality. As commonly observed in shift workers, even after a night of sleep deprivation workers find it difficult to initiate and maintain sleep in the morning hours because the circadian drive for wakefulness is high at that time even though the need for sleep is very high Sleep consolidation, that is regulating the sleep period to one part of the day and activity to another instead of fragmenting sleep across different times of the day, is perhaps one of the most significant contributions of the circadian system to sleep physiology. Light exposure can modulate the sleep-wake cycle as well as sleep architecture. Bright light in the morning shortens sleep duration 226,227 and advances circadian rhythms without any effects on non-rem sleep homeostasis Light exposure in the evening increases sleep latency to stage 2 and changes the temporal distribution of SWS such that EEG delta power is lower during the first and higher during the fourth sleep cycle 228,229. Furthermore, REM latency is decreased the night following morning light exposure compared with evening bright light exposure 230. Taken together, the studies suggest that light exposure primarily affects the temporal regulation of sleep, most likely by modulating underlying circadian regulatory processes. Furthermore, direct iprgc projections to the VLPO, suggests that sleep architecture may be differentially modulated by optical wavelengths 231,232. This is supported by the findings that when individuals are exposed to 550 nm green light or to 460 nm blue light for 2 h prior to habitual sleep time, blue light but not green light exposure modestly reduces SWA in the first but
54 38 significantly increases SWA during the third sleep cycle in parietal and occipital brain regions 233. Moreover, blue light significantly shortens REM sleep duration during these two sleep cycles 233. Thus, the effects of light on the dynamics of SWA and REM sleep durations are also likely blue shifted similar to other circadian outputs such as melatonin secretion and locomotor activity Alertness Cognitive performance is often impaired in individuals with disrupted circadian rhythms 15, Attention shows clear circadian variation, usually being the highest during day time and progressively worsening during the night 15. Memory performance also follows similar circadian variation, being worse at night compared to day time 15, Even higher order aspects of cognition including executive functions, inhibition and processing of the rapid temporal characteristics embedded in both speech and non-speech signals show circadian rhythms in performance 15. Alertness is a psychosomatic state that has often been considered to be the inverse of sleepiness and not surprisingly then has frequently been measured using subjective sleepiness scales and even objective measures of sleep such as EEG changes 244,245. Subjective sleepiness can be described as one s perceived experience of the propensity to fall asleep and similarly objective measures of sleepiness measure the same homeostatic propensity to fall asleep 245. On the other hand, alertness can be described as the capacity to respond correctly to internal or external stimuli and hence has often been equated to task performance and has also been measured as such 245. Using such a construct of alertness it is understandable then that alertness can be more heavily influenced by neurophysiological substrates governing motivation, affect and cognitive capacities as compared to sleepiness. Both circadian rhythms and the sleep homeostatic processes impact alertness such that the circadian process represents the wake-
55 39 promoting drive to balance the accumulating homeostatic drive for sleep during wakefulness 244,246. Light exposure can strongly modulate alertness in a time-dependent manner. Light stimulates the ascending arousal system and eventually the cortex to enhance alertness and cognition 206. Several studies using different light intensities and light exposure durations have shown that exposure to bright nocturnal light increases alertness levels as compared to being exposed to dim light Most studies associate this increased alertness at night by light exposure with the suppression of melatonin. Administration of supraphysiologic doses of melatonin can increase sleepiness and induce changes in the waking EEG opposite to those induced by bright light, and the effect of light on the EEG and sleepiness can be counteracted by exogenous melatonin 250. It has been hypothesized that melatonin elicits these effects by attenuating SCN-dependent mechanisms responsible for promoting and maintaining cortical and behavioral arousal at particular times in the circadian cycle 165. However, light exposure can enhance alertness even during daytime hours, when circulating melatonin levels are very low, suggesting that extra-scn extra-pineal pathways can also mediate the effects of light on increasing alertness 251,252. Positron Emission Tomography (PET) results on the effects of polychromatic light exposure during the biological night demonstrated that alerting responses are not confined to hypothalamic areas in the vicinity of the SCN but extend to modulation of a large-scale network of cortical areas involved in regulating attention, particularly a large-scale occipito-parietal attention network, including the right intraparietal sulcus 253. These findings are supported by recent functional magnetic resonance imaging (fmri) studies that demonstrate that short exposure to bright light during subjective daytime transiently attenuates sleepiness developed in constant darkness and corresponds with enhanced responses in the posterior thalamus, including
56 40 the pulvinar nucleus, which has been implicated in visual pattern discrimination, visual attention and alertness regulation 251,254. Light exposure also increases activity in a large scale cortical network, involving parietal, temporal, occipital and insular areas, all involved in regulating attention 251. Several studies have examined the relationship between the optical wavelengths and its alerting response using monochromatic light exposures for different durations at night 141,251,255,256. All studies report higher efficacy of short wavelength light (470 nm and lower) over longer wavelengths (550 nm and greater) in improving alertness Mood Similar to alertness, subjective measures of mood are lowest near the minimum of the CBT cycle, under constant routine (CR), which corresponds to roughly the middle of the major nocturnal sleep episode in entrained individuals 257,258. Under a CR protocol, individuals remain awake under constant (unchanging) environmental conditions. This allows studying biological rhythms independent of the affects of behavioral cycles such as the sleep-wake cycle. However, remaining awake for long durations (greater than 24 h) during the CR protocol can impair mood due to sleep deprivation 259. To circumvent this issue, the forced desynchrony (FD) protocol can be used which manipulates the inability of the circadian pacemaker to entrain to periods significantly longer than 24 h. Under FD, individuals maintain h days with approximately 9 h of sleep and 19 h of wakefulness. This long period is outside of the range of circadian entrainment resulting in desynchrony between the circadian timing system (which free runs) and the imposed sleep-wake cycle. Subjective mood assessed at different circadian phases using the forced desynchronization protocol also demonstrates a significant variation in mood similar to variations observed under CR but circumvents the confounding impact of sleep deprivation 14. Furthermore, this approach revealed that the temporal alignment between the sleep-wake cycle
57 41 and the endogenous circadian rhythms affects mood even in healthy subjects. When sleep is displaced the phase relationship between the sleep-wake cycle and the endogenous circadian pacemaker changes and may alter mood while awake 14. This phase misalignment is common in shiftworkers as well as in individuals with endogenously mediated circadian rhythm sleep disorders. In depressed individuals, one of the prominent symptoms is a diurnal variation in mood and is typically experienced as mood being worse during the day and progressively better in the evening, converse to what is observed in healthy individuals It is suggested that this shift in the diurnal mood variation is mediated by a weakened circadian function 260. Furthermore, some of the major neurotransmitters implicated in mood regulation such as norepinephrine, dopamine and serotonin (5-HT), all exhibit circadian rhythms in their levels, release, and synthesis-related enzyme expression 263. The expression and activity of several of the receptors that bind these neurotransmitters also follow circadian rhythms, suggesting that these entire systems are under circadian control and likely accounts for an additional means of mood impairment associated with circadian desynchrony The role of circadian rhythms in mood disorders Mood can be defined as a long-term background affective state 264 with numerous contributing physiological processes ranging from hormones to social interaction. Circadian rhythms also regulate many of the biological substrates underlying mood regulation including neurotransmitters, the timing of social interaction and even cognitive functions that modulate perception of social interaction 15, Sleep also plays a critical role in modulating mood and is often disturbed in individuals with mood disorders. Both the homeostatic and circadian components of sleep are affected in depression and sleep disturbance constitutes one of the major diagnostic symptoms of depression. As mentioned previously, one of the commonly observed alterations in sleep associated with depression is a reduction in REM sleep onset latency (the
58 42 time required to reach REM episode) and a reversal of REM occurrence; that is, more REM occurs earlier in sleep than during the later portion as observed in healthy individuals. According to the internal coincidence hypothesis, an abnormal relationship between the circadian system and the sleep-wake cycle occurs in which an abnormal phase-angle between an advanced/delayed circadian pacemaker and the sleep-wake cycle promotes depression 265, Categorizing depression According to the Diagnostic and Statistical Manual of Mental Disorders (IV) 273 mood disorders are categorized as Depressive Disorders (unipolar depression), Bipolar Disorders, and two disorders based on etiology (Mood Disorder Due to a General Medical Condition and Substance-Induced Mood Disorder). The differentiation between conditions is based on the presence or absence of characteristic symptoms and diagnosis is based on meeting specific criteria. For example, to be diagnosed with major depression (MD) a patient must present with five or more of nine listed symptoms which must be present for a duration of two weeks or more and at least one of the symptoms must be depressed (low) mood or loss of interest/pleasure (anhedonia). The remaining seven symptoms include significant weight alterations, increased or decreased sleep durations, psychomotor agitation or retardation, fatigue or low energy, low self esteem, reduced capacity to concentrate and suicidal ideation. However, patients can often present with some of these symptoms with significant impairment of well being but do not qualify for MD, in other words they are sub-threshold or fall below the minimum standards for being diagnosed with MD. Therefore, two sub-threshold categories of depression were added to the diagnostic module. When a patient presents with more than two but less than five of these symptoms for at least two weeks duration it is classified as Minor Depression (MinD) and when patients present with all the symptoms to qualify for major depression but do not meet the minimum two week
59 43 criteria and the episode cycles or repeats they are diagnosed with recurrent brief depression (RBD). Even with these additions, approximately two-thirds to three-fourths of patients with sub-threshold depression present with depressive symptoms, causing significant psychosocial impairment, but which still do not satisfy any DSM-IV diagnosis and often do not complain of anhedonia and depressed mood, the necessary criterion for all DMS-IV categories of depression 274,275. This remaining group is labeled as subsyndromal symptomatic depression or subsyndromal depression (SD) and defined as greater than, or equal to, two symptoms of depression of the same quality as in MD, but which symptoms excluded depressed mood and anhedonia. The symptoms must be present for more than 2 weeks and be associated with social dysfunction 274. Further, MD, MinD, and dysthymia must be excluded. However, it must be noted that even with the additional group of SD, individuals that have one symptom of depression, including depressed mood or anhedonia, or those who have more than two symptoms of depression, occurring less than 2 weeks, are not included in any of the depressive disorder categories. These patients are broadly categorized as having sub-threshold depressive symptoms. The most commonly reported symptoms of subsyndromal depression include insomnia, fatigue and diminished ability to concentrate similar to MD and MinD supporting the role of sleep and circadian disturbance as a significant commonality between various forms of depression 274. The 1-month point prevalence of SD in the general population is 3.9% as compared to 1.5% for MinD and 2.3% for MD 275. Results from the Epidemiologic Catchment Area study established that the 1-year prevalence of SD in the general population is 8.4% versus 3.5% for minor depression 274. The combined 1-year prevalence of minor depression and SD equals 11.8%, which is more than the 1-year prevalence of all DSM-III mood disorders combined (9.5%) 274. This finding established the existence of a high prevalence of subthreshold forms of depression in the general population.
60 CIRCADIAN RHYTHM DYSFUNCTION The entrainment process is a dynamic interaction between endogenous biological timekeeping mechanisms and environmental cues. However, if either the endogenous mechanism is faulty or the environmental cues are outside the range of entrainment then it is possible to induce circadian rhythm dysfunction. While the former process may be defined as endogenous circadian dysfunction the latter can be defined as exogenous circadian dysfunction. Circadian dysfunction can be either circadian misalignment and/or desynchronization. Circadian misalignment can be defined as the condition when individual physiological rhythms maintain stable periods and relative phases; however, the phase angle difference is abnormal. For example, the phase angle difference between the onset of melatonin secretion and mid-sleep can be advanced or delayed in SAD patients compared to healthy individuals 276,277. The altered or misaligned phase angle can then adversely affect physiology such as mood or neuropsychometric impairment. Circadian desynchronization can be defined as the condition when oscillators follow their individual phases instead of being entrained to one phase set by the central pacemaker. This may be induced by repeated phase shifts as different oscillators have differential rates of entrainment and while some shift to the new phase of entrainment set by central pacemaker rapidly others lag behind. In the case of shift work, phase alterations can be more rapid than the time required by the slowerentraining rhythms to synchronize, thereby inducing circadian desynchronization. It is important to note that both misalignment and desynchronization are relative, that is, an individual rhythm can be misaligned or desynchronized from another endogenous rhythm or an external rhythm such as the 24 h LD cycle. Therefore, there can be internal and/or external circadian misalignment or desynchronization and in the case of shift work both forms are present.
61 Endogenous circadian dysfunction and circadian rhythm sleep disorders Circadian rhythm sleep disorders (CRSDs) arise from a desynchronized biological clock and often exhibit psychiatric comorbidities including psychological and functional difficulties such as personality disorders and depression 278. CRSDs constitute a misalignment between the patient s sleep pattern and that, which is desired or regarded as the societal norm. It is important to note that CRSDs may not be simply extreme versions of circadian M-type or E-type chronotypes. People with M-type or E-type chronotype prefer morning or evening shifted activity patterns and their underlying circadian rhythms may also demonstrate similar phase of entrainment; however they are flexible to change their routines with sufficient education and time 278,279. However, CRSD patients appear to be unable to change their clock by means of motivation or education. While treatment with chronobiotic agents can be beneficial in many cases, there is also a high incidence of recurrence once treatment is stopped 280. This suggests that CRSDs are a manifestation of an underlying dysfunction of the circadian system. CRSDs often present with alterations in the phase of not only the major sleep-wake cycle relative to external clock time, but also in several other underlying circadian parameters such as CBT and melatonin 281,282. The phase alteration may add to the difficulty faced by CRSD patients to switch their rest-activity rhythms with education and motivation. Furthermore, studies suggest that the relative phase between two or more endogenous rhythms such as the CBT to sleep-wake or melatonin secretion to sleep-wake cycles are also different between healthy controls and CRSD patients, supporting the role of underlying circadian desynchronization that promotes CRSDs 281. Delayed Sleep Phase Syndrome (DSPS) is the most prevalent CRSD both in general and clinical populations 283,284. DSPS is the most common sleep-wake scheduling disorder observed and accounts for 10% of chronic insomnia patients 11. DSPS is characterized by the inability to fall asleep and arise at conventional times with sleep onset times well past midnight and sleep
62 46 offset times in the early afternoon 11,283. When individuals with DSPS are made to conform to earlier retiring times it leads to marked sleep onset insomnia due to an abnormal entrainment of the endogenous biological clock located in the SCN 285. The etiology of DSPS include abnormalities of the circadian timing system, such as long intrinsic period or a weak phase advance portion of the light PRC 9,286. In addition to the hallmark delays of sleep onset and offset, patients also manifest delayed timing of melatonin secretion patterns and CBT minimum It has also been demonstrated that there is increased suppression of melatonin to light exposure in DSPS patients versus controls, suggesting that the hypersensitivity to evening light may be a precipitating or maintaining factor for the phase delay in DSPS 287. Another line of experimental investigation suggests deficits in sleep-wake homeostasis in the etiology of DSPS. When DSPS patients and healthy controls are studied under CR protocol followed by an ultra-short sleep-wake cycle, DSPS patients show reduced recovery sleep duration compared to healthy controls following sleep deprivation 288. This suggests that DSPS patients may have poor ability to compensate for lost sleep and, thus, have difficulty falling asleep even when they wake early or have shortened sleep. Difference between DSPS patients and healthy controls on several PSG sleep variables, including SWS has been reported 289, further suggesting involvement of the homeostatic system. These findings suggest that DSPS may involve abnormalities of the circadian timing system and to some extent the sleep homeostatic process. In addition to DSPS, other forms of endogenous circadian rhythm misalignment include familial Advanced Sleep Phase Syndrome (ASPS) and non-24 h sleep-wake syndrome. The timing of both sleep and endogenous circadian rhythms are advanced in ASPS individuals 290,291. However, the internal phase relationships between sleep and the circadian rhythms of melatonin and CBT appear normal in these patients. Sleep architecture also appeared normal, suggesting
63 47 that the primary change in these patients is not associated with the sleep homeostatic facet of sleep-wake regulation but rather in the circadian aspect. A third form of endogenous circadian rhythm disorder is the non-24h sleep-wake syndrome which is characterized by a chronic, regular pattern of daily delays in sleep onset and wake times such that the sleep period eventually comes in and out of phase with local night time hours for sleep 280,292. The sleep-wake pattern resembles that of normal subjects free-running in temporal isolation studies. Non-24 h sleepwake syndrome is hypothesized to result from a failure of the biological clock to entrain normally to the environmental LD cycle and is more common in blind individuals. Similar to non-24 h sleep-wake syndrome another variant of endogenous circadian rhythm disorder includes is Irregular Sleep Wake Disorder (ISWD), that is an individual with an undefined sleepwake cycle. Unlike non-24 h sleep wake syndrome which is characterized by a non-entrained, continuously shifting sleep-wake cycle, in ISWD, sleep is fragmented or broken into a series of naps that occur throughout a 24-hour period 42. Patients complain of chronic insomnia, excessive sleepiness or both 42. A low-amplitude or irregular circadian rhythm of sleep-wake pattern may be seen in association with neurological disorders such as dementia and Alzheimer s disease. However, both ASPS and non-24 h sleep-wake syndrome are rare in the general population Prevalence of circadian rhythm sleep disorders There are very few community based epidemiological studies of CRSDs, hence the exact prevalence is not known 42. From the limited few that were conducted the prevalence of CRSD was 0.13% of adults in Japan, 0.17% of adults in Norway and 7.3% of adolescents in the American population 278,293. The proportion of patients who are diagnosed with a CRSD in current sleep disorders medicine clinics is limited compared to other diagnostic categories 283,294. However amongst CRSD patients, DSPS is the most common CRSD diagnosis (83%), followed by free-running disorder (12%). ASPS and ISWD are rarely diagnosed, accounting for less than
64 48 2% of the CRSD patients. On the other hand, the number of patients presenting to a clinic may be quite different from the prevalence in the population. Patients with CRSDs may not recognize that their problem has a physiological basis, or may not know that medical help is available. However, criterion based clinical diagnosis of CRSD necessitates that CRSD associated sleep disturbance causes clinically significant distress or impairment in social, occupational, or other important areas of functioning 295. It is important to note that many individuals may not meet this strict criterion, but still have underlying circadian misalignment. This may be potentiated by the individual differences in being able to tolerate circadian phase misalignment, and without recognizing or addressing the underlying pathologic condition of circadian misalignment, overall well-being may ultimately worsen Shift work and exogenous circadian dysfunction The major difficulty with nocturnal work is functioning at a time when the wakefulness drive is low and sleep pressure is high and trying to sleep when the circadian wakefulness drive is high and opposing sleep 218,220,299. This results in decreased alertness, increased errors and accidents and poor daytime sleep 241, In addition, the repeated irregular light exposure schedules in shift work alternate too quickly for the endogenous biological clock to be able to stably adapt to any one schedule and since different physiological rhythms change their phase and period at different rates , repeated alterations caused by shiftwork induces desynchronization between different endogenous circadian rhythms as well as between circadian rhythms and the external LD cycle 265,307. This desynchronization has profound effects on mood and cognition and can adversely impact performance and the safety of the workers and other individuals directly or indirectly associated with them 263,265,308. Controlled lab studies simulating shift work have demonstrated that altering the light dark cycle can change the rhythms of melatonin, cortisol and peripheral clock gene expression in
65 49 humans demonstrating that under shift work conditions light may still be the principal circadian time cue altering the phase and periodicity of the system 224. Furthermore, altering the light-dark cycle to simulate shift work conditions also results in systematic increases in postprandial glucose, insulin, mean arterial pressure, systematic decreases in leptin and sleep efficiency 309. These deleterious changes may contribute to the myriad of health disorders associated with shift work Health effects of shift work Depression is common in the work force; 3.7% of the Canadian work force experienced a major depressive episode (MDE) in ,311. Similar to the general population, the occurrence of depression in women in the workforce is more than in men (5.1% vs. 2.6% respectively) 311. However, the prevalence of MDE in regular evening and night shift workers is almost doubled (5.6%) compared to those working a regular day schedule (3.5%) 311. Interestingly, the differential prevalence of MDE between men and women is also lost in shiftwork, suggesting that it affects both genders equally. Furthermore, chronobiological observations have also raised concern that shiftworkers may be at an increased risk of developing mood disorders. Circadian desynchronization due to shiftwork include symptoms of psychophysiological disturbances of well-being, resembling symptoms of affective disorders, such as irritability, fatigue and poor sleep, apathy, poor appetite, and psychosomatic complaints 312,313. In addition, changes in mood have been shown to be associated with irregularity of sleep patterns and with sleep deprivation 272, Several studies indicate that imposed time shifts may lead to the development of clinical depression in vulnerable individuals. An increase in depressive symptoms has been reported after experiencing a phase-delay time shift 318. Several studies suggest shiftwork may predispose certain individuals to affective disorders. Nurses have increased depression, anxiety and irritability during night shifts compared to day shifts 319,320. In retired workers, cases of
66 50 depression are more frequent in shiftworkers compared to day workers and in current and former shiftworkers there is a trend of increasing prevalence of major depressive disorder with increasing years of shiftwork 313,321. Shift workers have lower cognitive performance than individuals who never worked shifts. Memory performance decreases with increasing shift-work duration, but importantly, the cognitive performance of a shift worker who has stopped shiftwork for more than 4 years increases, suggesting it is possible to reverse these effects 322. Studies on health care providers, including nurses and physicians, demonstrate that working nights can increase task completion times and error rates as their shift progresses Subjects also rate themselves to be in significantly worse mood, and have more unclear thinking when working the night versus day shift 323. In a population-based study involving more than 27,000 individuals, obesity, high triglycerides and low levels of high density lipoprotein (HDL) cholesterol clustered more often in shift workers than day workers 328,329. Another study examining 5,517 individuals over 12 years demonstrated that shift workers have a relative risk of 1.31 for developing all forms of circulatory diseases 330. A prospective study of over 14,000 men in Japan demonstrated that shift workers have a 3-fold higher risk of developing prostate cancer 331. In addition, female shift workers have a 50% greater risk of developing breast cancer than non-shift workers 332. Moreover, with both prostate and breast cancer, the relative risk increased significantly with longer durations of shift work. Another major health concern in shift workers is poor sleep 218. EEG studies demonstrate a significant 2-4 h reduction in total sleep time in shift workers compared to total sleep times in day workers 218,333. Most of the sleep loss involves stage 2 and REM sleep, whereas SWS is unaffected 219. The subjective aspects of sleep seem little affected, apart from the reports of
67 51 premature awakenings and of not getting enough sleep 219. Rotating shift workers face greater sleep challenges than permanent shift workers. This is because permanent shift workers have an inverted but constant sleep schedule. On the other hand, the more prevalent rotating type shift work offers no opportunity to be entrained to any one schedule Shift work countermeasures With more than 14 million shiftworkers in North America and the emerging evidence supporting a strong association between shift work and major health disorders including various forms of cancer, affective disorders, obesity and cardiovascular diseases 220,328,331,332,334,335, various countermeasures have been tested to improve circadian rhythmicity in shift workers. The most common approach to counteract the circadian disruption caused by shiftwork has been to alter the circadian phase of subjects by regulating the time of exposure to bright light 221, In combination to appropriately timed bright light exposure, exogenous melatonin treatment has also been used to induce phase shifts according the melatonin PRC. Bright light and melatonin can be used separately or together. In the latter case, the time of melatonin administration is separated by 12 hours from bright light treatment due to their opposite phase shifting effects. The primary objective of most of these countermeasures has been to shift circadian rhythms so that the period of wakefulness regulated by Process C falls during the nocturnal work period, thereby improving task performance, reducing accidents and errors and improving daytime. However, a complete inversion of circadian rhythms is seldom reached, even across successive night shifts and after years of night work experience 340. One of the principal practical difficulties with maintaining an artificially altered internal circadian phase of subjects is exposure to bright light at times when individuals are to remain in darkness. While darkness can be strictly controlled under laboratory conditions this approach may often face practical challenges.
68 THESIS HYPOTHESIS AND OBJECTIVES The synchronization of endogenous rhythms between each other and relative to the external environment is regulated by the master circadian clock. Endogenous melatonin biosynthesis and secretion is directly regulated by the SCN making melatonin rhythm a highly reliable circadian phase marker for studying phase alterations of the central pacemaker. Furthermore, exogenous melatonin application can act directly on the SCN and alter the timing of circadian rhythms, suggesting that it plays a critical role in transducing temporal cues back to the master clock as well as to peripheral organs and thereby regulating the temporal order of physiological processes. Mood and neuropsychometric performance (measures of subjective alertness, sleepiness and fatigue) are under circadian regulation and are often impaired due to circadian dysfunction. My overall hypothesis was that correcting endogenous and exogenously-mediated circadian dysfunction will improve mood and neuropsychometric performance. The data chapters included in this thesis addressed aspects of this hypothesis as follows: In the first objective presented in this thesis (Chapter 2), the effects of low endogenous melatonin on mood and sleep are examined. In the second study (Chapter 3), the effects of exogenous melatonin treatment on mood and sleep in individuals with endogenous circadian disorder (DSPS) was investigated. In the third study (Chapter 4), the efficacy of spectral modulation in attenuating nocturnal light exposure-induced alteration in circadian phase markers was explored using a rodent model. In the final objective presented in this thesis (Chapter 5), the effects of attenuating nocturnal light exposure-induced alteration in endocrine and molecular circadian phase markers on neuropsychometrics was examined in humans.
69 53 2 ALTERED SLEEP ARCHITECTURE AND HIGHER INCIDENCE OF SUBSYNDROMAL DEPRESSION IN LOW ENDOGENOUS MELATONIN SECRETORS Shadab Ataur Rahman 1, Shai Marcu 2,3, Leonid Kayumov 2, Colin Michael Shapiro 1,2,3,4 * 1 Sleep Research Laboratory, University Health Network, Toronto, Ontario; Canada 2 Dept. of Psychiatry, University of Toronto, Toronto, Ontario; Canada 3 Youthdale Child and Adolescent Sleep Centre, Toronto, Ontario; Canada 4 Dept. of Ophthalmology, University of Toronto, Toronto, Ontario; Canada This work was published in the journal European Archives of Psychiatry and Clinical Neuroscience 2009 Dec 18. [Epub ahead of print; DOI: /s ] NOTES: SAR completed data collection, analysis, interpretation and manuscript preparation. SM assisted with data collection. LK and CMS were involved with study design and interpretation. CMS conducted all clinical psychiatric assessment and final manuscript edification.
70 ABSTRACT Melatonin secretion is synchronized to the sleep/wake cycle and has been suggested to have somnogenic properties. Sleep/wake cycle disruption and alterations in the secretary pattern of melatonin is present in various psychiatric disorders. The objective of this study was to investigate the sleep architecture and the presence of depression in individuals with low endogenous melatonin levels. The study included 16 participants (mean age 30.3 ± 14.9 years). The first night of testing included psychiatric evaluation followed by melatonin secretion profile evaluation by Dim Light Melatonin Onset test and then standard montage polysomnographic testing. On the second night, only polysomnographic testing was carried out with an imposed sleep period of 8 h. Low endogenous melatonin secretors (LEMS) showed no discernible peaks in melatonin secretion compared to normal secretors (controls). LEMS demonstrated significant alterations in rapid eye movement sleep but not in non-rapid eye movement sleep along with poor sleep initiation and quality compared to controls. 55.6% of the low melatonin secretors group presented with subsyndromal depression. Melatonin has significant bearing on sleep architecture and a lack of melatonin may desynchronize endogenous rhythms allowing subsyndromal depression to manifest.
71 INTRODUCTION Melatonin (5-methoxy N-acetyltryptamine) is a neurohormone secreted primarily by the pineal gland under the control of the master circadian clock, the suprachiasmatic nucleus (SCN) 341. In humans, melatonin secretion is closely synchronized with the habitual hours of sleep, increasing soon after the onset of darkness, peaking in the middle of the night (between 0200 and 0400 hours), and then gradually returning to baseline levels by the end of the night 342,343. One of the principal functions of melatonin is to relay temporal queues to various organs, including the SCN itself 162,342. Several studies suggest a role for melatonin influencing the timing of sleep onset and sleep duration 54,314,341. In humans, melatonin administration during the subjective daytime promotes sleep by inducing earlier sleep onset and generating longer sleep duration Daily melatonin ingestion can entrain free running circadian rhythms in blind individuals and pharmacological suppression of nocturnal melatonin secretion increases total wake time and concomitantly decreases both non-rapid eye movement (NREM) sleep and rapid eye movement (REM) sleep 170. The circadian rhythm of plasma melatonin also has a temporal association with circadian rhythms observed in cortical EEG activity during sleep in humans 171, suggesting a direct influence of melatonin on sleep wake regulation. However, the effectiveness of melatonin in improving sleep depends on the nature of the sleep disorder and the timing of melatonin administration. In the elderly population, who show a decrease in melatonin secretion and often exhibit marked sleep wake cycle alterations, only certain melatonin replacement protocols tend to improve sleep 55, Based on whether the sleep disorder is primary or secondary in nature, exogenous melatonin may or may not be beneficial 347,348. This suggests that melatonin is not directly somnogenic but instead it plays an intricate role in the timing and synchronization of the sleep wake cycle.
72 56 Melatonin can regulate the timing of various physiological processes other than the sleep wake cycle 341,349,350. Interestingly, melatonin concentrations are altered in patients with major depression, additionally many other circadian rhythms measured in depressive patients are abnormal Endogenously depressed patients have also been recognized to have clear circadian rhythm abnormalities, consisting mainly in amplitude reduction 354. Sleep wake cycle disorders are often associated with depression, suggesting a role for circadian rhythm misalignment in generating psychosomatic and mood disorders 265,317,351,355. Since melatonin can regulate the timing of circadian rhythms, it is likely that alterations in melatonin secretion can lead to mood disorders or depression. Our working hypothesis for this study was that endogenous melatonin functions as synchronizer of circadian rhythms and a reduction in circulating levels of melatonin will lead to desynchronization of circadian processes leading to endogenous depression. The primary objective of this study was to examine if endogenous melatonin levels modulate sleep architecture and to investigate the possible presence of psychiatric comorbidity among this unique population of low endogenous melatonin secretors (LEMS) compared to normal melatonin secretors (control). 2.3 METHODS Sample population A total of 16 individuals (6 females), mean age of 30.3 ± 14.9 years, participated in the study at a single sleep clinic. The group of low melatonin secretors (n = 9) were selected from individuals who had previously been examined for sleep disorders at the same sleep clinic and had demonstrated low endogenous melatonin secretion tested by a Dim Light Melatonin Onset (DLMO) test. The original cohort consisted of 27 individuals who were referred for circadian rhythm disorders. Within that cohort, individuals were diagnosed with sleep apnea, periodic limb
73 57 movement syndrome and alexithymia. Moreover, several individuals were on regular medication including antidepressants that can alter the secretion of melatonin. Therefore, only individuals who were not on any medication and did not have any other sleep disorders were randomly called upon for participation in the study out of which nine individuals were available for the study. The control group (n = 7) consisted of healthy individuals recruited from a cohort of individuals who had previously been referred to the clinic for the assessment of circadian rhythm disorders based on subjective complaints of an inability to initiate sleep. The individuals who were selected for this study were diagnosed with no sleep disorders and assessed to have poor sleep hygiene which was addressed through subsequent follow-up visits to the clinic prior to this study. None of the individuals were on any regular medication including antidepressants that can alter the secretion of melatonin. All participants maintained sleep diaries for 2 weeks prior to starting the study. The mean bedtime and rising time according to the sleep logs were 0153 h (range h) and 0748 h (range h) for the low melatonin secretors, respectively. The mean bedtime and rising time according to the sleep logs were 2317 h (range h) and 0823 h (range h) for the controls, respectively. Before further testing, all major psychopathologies including schizophrenia were ruled out for both groups by conducting a mini evaluation based on the Diagnostic and Statistical Manual of Mental Disorders (DSM-IV) 273. All differential diagnoses resulting from sleep restriction or secondary sleep disorders were also excluded using The International Classification of Sleep Disorders 295. All assessments were conducted by a psychiatrist trained and experienced in sleep medicine. The study protocol was approved by the Research Human Ethics Committee of the University Health Network and all patients signed consent forms prior to being assessed.
74 Study design During their initial visit with the psychiatrist/sleep specialists, the participants were asked to complete a battery of questionnaires which included measures of subjective levels of sleepiness, self-esteem, depression and anxiety. All clinical interviews and questionnaires were completed during the day of this initial visit. After their initial assessment, the patients underwent a two night sleep study comprised of self-report questionnaires, saliva sample collection for melatonin assay and a PSG which included electroencephalography, electrooculography, electromyography and respiratory monitoring. The first night involved completion of pre-sleep questionnaires, followed by saliva sample collection and PSG. Saliva samples were acquired hourly from 1900 until 0300 h. The patients then retired to sleep and their next morning waking time was left to them to choose. An hour after waking up, they completed a series of post-sleep questionnaires. The second night sleep study involved completion of the same pre-sleep questionnaires as the previous night and all patients retired to bed at a fixed time of 2300 h and were awoken the next morning at a fixed time of 0700 h, thus creating an imposed sleep period. They again completed the same series of post-sleep questionnaires. No saliva samples were collected on the second night. Sleep physiology parameters Objective measures: two night polysomnography A standard montage including electroencephalography, electrooculography, electromyography and respiratory monitoring (oxygen saturation, nasal airflow, and breathing effort) was used. The polysomnographs were scored by a single-blinded scorer according to standardized criteria 356. The sleep parameters included sleep onset latency (SOL), sleep efficiency (SE%), REM latency, slow wave sleep (SWS%), REM% and arousal index (AI). SOL was defined as the time required to reach the first 30-s epoch of stage 2 sleep. Arousals were
75 59 defined as an abrupt shift in EEG frequency, which may include theta, alpha and/or frequencies greater than 16 Hz but not spindles. Ten seconds of continuous sleep must precede the arousal. The arousal must last 3 s and it must be accompanied by an increase in chin EMG if it occurs during REM sleep 357, Subjective measures Patients completed questionnaires pre- and post-sleep study on both nights of testing. The questionnaires were used as a subjective measure of fatigue, sleepiness, alexithymia (emotional connectedness) and alertness. The questionnaires were the Fatigue Severity Scale (FSS) 359, Stanford Sleepiness Scale (SSS) 360, Toronto Alexithymia Scale (TAS-20) 361, ZOGIM-A (questionnaire name is not abbreviated) 362, and the Toronto Hospital Alertness Scale (THAT) 362. The THAT is a rating scale of alertness that assesses a range of activities such as ability to concentrate, think of new ideas, or focusing on the task at hand. The ZOGIM-A describes a range of activities that might impact on alertness (sleep loss, exercise, caffeine, etc.) and inquires about how a person may function differently if he or she was more alert 362. The post-sleep study questionnaires completed in the morning after the sleep study were the FSS, SSS, ZOGIM-A, and the THAT. All questionnaires were completed an hour before retiring to bed and within an hour after waking up. In addition, subjective sleepiness was evaluated using the Epworth Sleepiness Scale (ESS) 363, Circadian rhythm parameters Sample collection for melatonin was carried out as previously described 365,366. Briefly, individuals were seated in a dark room from 1900 to 0300 h and saliva specimens were collected using the Sali-Saver (ALPCO, American Laboratory Products Company, New Hampshire, USA). Samples from each hour were refrigerated until all samples were collected at the end of the night. Sample collection was carried out under dim red lighting (<5 lux). Saliva specimens
76 60 were analyzed immediately after collection of the last sample. Salivary melatonin was determined by Direct Saliva Melatonin ELISA kit from Buhlman Laboratories (Allschwil, Switzerland). Saliva specimens from a given subject were run with the same assay kit; all kits used in this study were from the same batch. Assay functional sensitivity was 1.3 pg/ml, the maximum intra-assay and inter-assay coefficients of variability were 6.5% (n = 12) and 11.3% (n = 12), respectively (in the range of concentrations of melatonin between 1 and 81 pg/ml). The DLMO, defined as the point at which the melatonin secretion begins, was used as a marker of the circadian phase. DLMO was defined as the time of the sampling of the first salivary sample to show an elevated melatonin measurement that remained elevated in the subsequent sampling time. To be considered elevated the melatonin concentration had to be 20% greater than the 1900 h baseline value and greater than a minimum value of 4 pg/ml 367, Psychiatric parameters The parameters evaluated were depression, anxiety, self-esteem and subjective perception of sleepiness. The scales used were previously validated questionnaires, namely, the Centre for Epidemiological Studies Depression Scale (CES-D) 369, the Rosenberg Scale (RS) 370, the Zung Anxiety Scale (ZAS) 371 for each of the measures, respectively. Coupled with the self-report questionnaires, the final diagnosis of subsyndromal depression was made by psychiatrists who were blinded to melatonin secretion type using the definition provided by Judd et al., greater than, or equal to, two symptoms of depression of the same quality as in MD, but which symptoms excluded depressed mood and anhedonia. The symptoms must be present for more than 2 weeks and be associated with social dysfunction 274, Data analysis Results are expressed as mean ± SEM. All questionnaire data were analyzed with the non-parametric Mann Whitney test with a Bonferroni s alpha adjustment of for each
77 61 psychiatric questionnaire and 0.01 for each sleep questionnaire. Objective sleep variables and circadian parameters were compared between the two groups with Student s t test with Bonferroni correction for repeated t tests for multiple variables. Statistical significance was set at P = First and second night data were analyzed separately to negate a possible first night effect RESULTS Melatonin secretion profiles in low endogenous melatonin secretors and controls Low endogenous melatonin secretors were defined as subjects with no discernible DLMO point throughout the duration of testing. Based on the melatonin secretion profile, subjects were dichotomized into groups: normal melatonin secretors (controls) (n = 7; 3 females) and low endogenous secretors (n = 9; 2 females). The mean age of the controls was 31.7 years (range years) and the mean age of the LEMS was 27 years (range years). The mean amount of melatonin secreted at DLMO by the control group was 23.1 ± 9.2 pg/ml with a night time peak of 59.9 ± 10.7 pg/ml. In contrast, the low secretors had no clearly discernible peak or inflection in the rise of melatonin with the highest concentrations reaching only 7.8 ± 2.8 pg/ml. Melatonin levels were significantly higher in controls than in LEMS starting from 2100 h (DLMO time) until the end of the testing period (0300 h) (Fig. 2.1) Sleep architecture in low endogenous melatonin secretors and controls None of the sleep architecture parameters on the first night of testing were significantly different between LEMS and normal secretors (controls) (Table 2.1). While normal secretors retired to bed at 0331 ± 0112 h, LEMS retired at 0339 ± 0021 h. The mean wake up time for LEMS was 0908 ± 0227 h and for normal secretors was 1054 ± 0124 h. The second night of
78 62 Figure 2.1: Melatonin secretion profile of low endogenous melatonin secretors (LEMS) and normal secretors (controls). Saliva samples were collected hourly from 1900 to 0300 hours and batch processed by ELISA. Data shown are mean ± SEM. Circadian parameters were compared between the two groups with Student s t test with Bonferroni correction for repeated t tests for multiple variables (significance at **P < between LEMS and controls).
79 63 Table 2.1: Sleep physiology parameters of low endogenous melatonin secretors (LEMS; n = 9) and the normal secretors (controls; n = 7). Sleep Parameter Sleep Onset Latency Sleep Efficiency% Slow Wave Sleep% REM LATENCY FIRST NIGHT SECOND NIGHT LEMS CONTROLS LEMS CONTROLS 10.9 ± ± ± 7.4** 12.5 ± ± ± ± 2.8** 89.3 ± ± ± ± ± ± ± ± 3.3 ** 76.4 ± 7.2 REM% 17.8 ± ± ± 3.3** 23.6 ± 2.7 Arousal Index 8.3 ± ± ± ± 2.1 Subjects underwent full montage polysomnography for two nights. The second night of testing had an imposed sleep period from 2300 to 0700 hours. Data shown are mean ± SEM. Objective sleep variables were compared between the two groups with Student s t test with Bonferroni correction for repeated t tests for multiple variables (significance at **P < between LEMS and controls).
80 64 sleep studies with imposed sleep period revealed significant differences in four parameters between LEMS and controls (Table 2.1). SOL was significantly higher in LEMS (35.2 ± 7.4 min) than in controls (12.5 ± 4.2 min); SE was significantly lower in LEMS (68.9 ± 2.8%) than in controls (89.3 ± 2.1%); REM latency was significantly higher in LEMS (193.0 ± 40.5 min) than in controls (76.4 ± 7.2 min) and REM% was significantly less in LEMS (13.1 ± 3.3%) than in controls (23.6 ± 2.7%) Subjective Neuropsychometrics in Low Endogenous Melatonin Secretors and Controls No significant differences were noted in the pre- and post-psg questionnaires between the groups on either the first or the second night (data not shown). ESS scores were significantly higher in LEMS (10.6 ± 1.5) than in the controls (5.2 ± 1.2) (Fig. 2.2a). While LEMS and controls scored comparably on the RS and ZAS, LEMS scored significantly higher on the CES- D (17.8 ± 3.0) than controls (7.4 ± 1.4) (Fig. 2.2a). The clinical evaluation revealed a higher prevalence of subsyndromal depression in LEMS than in controls, only 14.3% controls (n = 1) were diagnosed with subsyndromal depression whereas 55.6% of LEMS (n = 5) were diagnosed with subsyndromal depression by psychiatrists (Fig. 2.2b). 2.5 DISCUSSION Our study is the first to demonstrate that a natural reduction in endogenous melatonin secretion is associated with altered sleep architecture and sleep phase duration. A salient feature of our study design was to be able to compare the natural sleep pattern with that of an imposed sleep period which can be helpful in diagnosing circadian rhythm sleep disorders (CRSDs), sleep onset insomnia and sleep/wake cycle phase alterations 285. While SOL levels were comparable in the controls between the first and second night of PSG testing, the LEMS had higher SOL between the first and second night suggesting that low levels of melatonin may promote sleep
81 65 Figure 2.2: Psychiatric profile of low endogenous melatonin secretors (LEMS) and normal secretors (controls). LEMS demonstrated significantly increased daytime sleepiness compared to controls with no significant differences in self-esteem and anxiety levels (a). LEMS demonstrated elevated depression levels and higher incidence of subsyndromal depression (b). Data shown are mean ± SEM. All questionnaire data were analyzed with the nonparametric Mann Whitney test with a Bonferroni s alpha adjustment (significance at **P < between LEMS and controls).
82 66 onset insomnia and alterations in sleep-wake cycle. Furthermore, lower SE in LEMS compared to controls demonstrates that a reduction in endogenous melatonin levels leads to poor sleep quality. A routine example of poor sleep maintenance and increased sleep fragmentation is observed in daytime sleep in shift workers who also have lower melatonin levels during the main sleep episode 218,219,374. Our study also demonstrates elevated ESS levels in LEMS suggesting increased daytime sleepiness in LEMS that may be directly induced by poor sleep quality. Individuals from both groups were awake until 0300 h the first night due to DLMO testing and this may have caused a phase delay influencing sleep homeostatic aspects the second night. However, we did not find any significant increase in the SOL of the controls between the first and second night (with imposed sleep) of testing which would be indicative of a phase delay in the major sleep episode. Furthermore, there was no significant difference between the first and second night SWS% in controls suggesting that the participants had adequate sleep the first night and were not sleep deprived. It is possible that the control group can entrain their circadian rhythms faster than low melatonin secretors. Exposure to light after awakening entrained the normal secretors but not the low secretors. However, the differential ability in entrainment of circadian rhythms between low melatonin secretors and normal secretors requires further investigation. We also observed significant alterations in REM latency and the amount of REM sleep (REM%) in LEMS compared to normal melatonin secretors without affecting SWS. According to the two-process model of sleep, SOL is modulated by the circadian propensity for sleep (Process C) and REM sleep has also a circadian component whereas SWS is driven by the homeostatic propensity for sleep (Process S) 272. This suggests that melatonin is required for the finer alignment of the sleep wake cycle especially those parameters that have a circadian
83 67 component. A significant delay in sleep onset and REM sleep onset strongly suggests a shift in the circadian timing of the major sleep episode in low melatonin secretors. Alterations in REM sleep is also associated with depression; however, these alterations are commonly seen as an increase in REM sleep and decrease in REM latency 375,376. This difference in REM sleep between the individuals with subsyndromal depression as in our study and major depression as in other studies suggests that the sleep architecture alterations observed with major depression has different underlying neurobiological mechanisms than those inducing sleep architecture alterations in subsyndromal depression. It is possible that low melatonin secretors also have disrupted serotonin (5-HT) secretion We hypothesize that the low melatonin secretors in our study may have high higher than normal central 5-HT availability. Moreover, it may be that 5-HT levels do not differ during the day between the low melatonin producers and healthy individuals. However, nocturnal 5-HT levels are higher in low melatonin secretors than in healthy individuals due to the reduced serotonin to melatonin conversion in the low melatonin producers 380. This excess in pineal 5-HT may then play an additive role in total central 5-HT availability and adversely affect 5-HT signaling. REM sleep expression is promoted in part by cholinergic neurons 381,382 and inhibited by serotonergic and noradrenergic neurons in the dorsal pontine tegmentum 383. Fluoxetine a selective serotonin reuptake inhibitor decreases REM sleep and increases REM latency in both healthy and depressed subjects 384 suggesting that increased serotonergic signaling may induce the changes in REM sleep observed in our study. The increased SOL and REM latency suggest that there may be a delay in the sleep cycle compared to other endogenous rhythms. According to the phase advance theory of affective disorders 7,272, the oscillator regulating endogenous rhythms has a phase that is advanced compared to the oscillator regulating sleep. This causes the patient to sleep at a time when the
84 68 other rhythms are at an inappropriate phase for sleep initiation or maintenance, promoting desynchronization and depression. However, further investigation is required to determine the phase angle difference between endogenous rhythms such as body temperature and cortisol secretion and the sleep cycle in LEMS to confirm the phase advance hypothesis of affective disorders in our patient population. Prior studies have demonstrated that exogenous melatonin administration even in healthy individuals increases the amount of REM sleep obtained 385,386, suggesting that a lack of melatonin can cause a reduction in REM sleep as observed in our study. In addition, the reduction in melatonin secretion may induce circadian desynchrony promoting the development of subsyndromal depression. Furthermore, steady internal and external phase relationships appear to be crucial for a stable and euthymic mood state (i.e. the timing between core body rhythms such as cortisol and temperature as well as the timing of sleep with respect to the day night cycle) 265,353,387. Any misalignment may bring with it the propensity for mood fluctuation, particularly in vulnerable individuals such as low melatonin secretors who have marked alterations in their sleep wake rhythm. It has been postulated that alterations in circadian phase markers such as low melatonin secretion can be a biological marker for susceptibility to endogenous depression 388,389. Both the amplitude and rhythm of melatonin secretion can be altered in patients suffering from unipolar depression as well as in patients suffering from bipolar affective disorders 390. Abnormal levels and patterns of melatonin secretion have been observed in depressed patients in some , but not all studies 394. The circadian rhythm of individual components such as cortisol and melatonin rhythm, core body temperature rhythm, may have different controls and alteration in any one control is likely to have significant impacts on other circadian controls as well. Therefore, inconsistent findings may also arise from the multifactorial nature of circadian mechanisms and heterogeneity of symptoms in mood disorders. For example, a reduction in amplitude of 24-h
85 69 cortisol levels is apparent in non-psychotic depressed patients, but not in patients with the psychotic subtype of depression 395. In addition, a growing number of studies indicate that genetic vulnerability moderates the nature of circadian disturbances in mood disorders 396,397. Delayed sleep phase syndrome (DSPS) is a CRSD, caused by a desynchronized central biological clock 289. DSPS patients show emotional features such as low self-esteem, nervousness and diminished control of emotional expression 16. Our results suggest that similar to DSPS, low endogenous melatonin levels may promote alterations in the timing of the major sleep episode. However, the low melatonin secretors in our study cannot be categorized as DSPS patients based on the lack of characteristic delayed retiring times when comparing retiring times between weekdays (imposed sleeping period) and weekends (free to choose sleeping period) 398,399, furthermore, due to the lack of a clearly discernable DLMO which is observed in DSPS patients 400. In this present study, we used self-reporting questionnaires and psychiatric interviews to evaluate depression, anxiety and self-esteem. The presence of subsyndromal depression and high RS and CES-D scores suggests a similar pattern of depression and low self-esteem arising from underlying desynchronization of the circadian system due to a lack of melatonin and its chronobiotic properties. Our study is the first to show that there is a very high prevalence of subsyndromal depression in LEMS. The prevalence of subsyndromal depression in the general population is 8.4% and females more frequently present with subsyndromal depression 372. In our clinical study, 14.3% of normal melatonin secretors (n = 1) were diagnosed with subsyndromal depression whereas in low melatonin secretors 55.6% (n = 5) were diagnosed with subsyndromal depression. Even though the number of participants in each group were low, making it a limiting factor in the present study, the high prevalence of subsyndromal depression in low melatonin
86 70 secretors is alarming because of a high possibility of this condition leading to major depression. Individuals with minor or subsyndromal depression have a 5.5-fold higher risk for major depression after 1 year compared to individuals who are not depressed 401. In addition, the age at which this population starts to exhibit such symptoms is also concerning. The mean age of low melatonin secretors was early 30s, much earlier than the observed decline in endogenous melatonin levels in elderly individuals. Similar problems of sleep/wake cycle desynchronization and subsyndromal depression with very fast progression into major depression are evident in the elderly population 401. Though we did not look into specific causes for this early onset of melatonin decline, a hyperactive hypothalamic pituitary adrenal (HPA) axis may play a role since elevated cortisol levels can reduce melatonin secretion 402. Another possibility is genetic polymorphisms and loss of function mutations in the genes coding the necessary enzymes that catalyze the reactions in the melatonin biosynthetic pathway. To our knowledge, our study is the first to examine sleep physiology in endogenously low melatonin secretors and it demonstrates markedly altered sleep architecture in low melatonin secretors with significant sleep wake cycle disruption. Moreover, an alarmingly high prevalence of subsyndromal depression was identified in low melatonin secretors. While previous studies have documented a correlation between circadian rhythm disruption and depression, our findings suggest a potential direct role of melatonin with depression which warrants further investigation.
87 71 3 ANTIDEPRESSANT ACTION OF MELATONIN IN THE TREATMENT OF DELAYED SLEEP PHASE SYNDROME Shadab A. Rahman 1, Leonid Kayumov 1,2 and *Colin M. Shapiro 1,2,3 1 Sleep Research Laboratory, University Health Network, Toronto, Ontario; 2 Dept. of Psychiatry, University of Toronto, Toronto, Ontario; 3 Dept. of Ophthalmology, University of Toronto, Toronto, Ontario This work was published in the journal Sleep Medicine 2009 Dec 29. [Epub ahead of print; DOI: /j.sleep ] NOTES: SAR completed data collection, analysis, interpretation and manuscript preparation. SAR, LK and CMS were involved with study design and interpretation. CMS conducted all clinical psychiatric assessment and final manuscript edification.
88 ABSTRACT Depression is a common problem in patients with Delayed Sleep Phase Syndrome (DSPS). This study used a randomized, double-blind, crossover, placebo-controlled approach to test the hypothesis that exogenous melatonin (5 mg) can attenuate depressive symptomatology in DSPS patients. Twenty patients with an established diagnosis of DSPS were dichotomized into DSPS with depressive symptoms (Group I; n = 8) and without depressive symptoms (Group II; n = 12) based on structured clinical interviews and a score greater than 17 on Center for Epidemiologic Studies Depression Scale (CES-D). Both groups received melatonin and placebo treatment for 4 weeks with a 1-week washout period in between. Participants underwent a clinical interview and psychometric evaluation to assess depression, and overnight polysomnographic sleep studies were carried out at baseline and at the end of melatonin and placebo treatments. Furthermore, melatonin secretion rhythm as a circadian phase marker was assessed by measuring urinary 6-sulphatoxymelatonin levels. Melatonin treatment significantly reduced depression scores in the depressed patients as measured by the CES-D and Hamilton Depression Rating Scale 17. Melatonin treatment improved sleep continuity in both groups compared to placebo and baseline conditions. Group I individuals showed marked alterations in melatonin rhythms compared to Group II individuals. Exogenous melatonin treatment may be an effective treatment modality for individuals with circadian rhythm sleep disorders and associated comorbid depressive symptomatology.
89 INTRODUCTION Depression is often associated with circadian rhythm abnormalities, and many diverse rhythms such as hormone secretion, neurotransmitter secretion and synthesis and behavioural rhythms can be disrupted in depressed patients, suggesting that such disturbances are not unique to a specific rhythm, but instead involve the central circadian pacemaker which regulates the various rhythms 403. One rhythm that is often disrupted in depression is the sleep-wake cycle, a disruption that, in turn, might lead to other rhythm disturbances 317,351. Delayed Sleep Phase Syndrome (DSPS) falls under the group of intrinsic Circadian Rhythm Sleep Disorders (CRSD), amongst which DSPS is most common both in general and clinical populations 283,284. DSPS is characterized by the inability to fall asleep and to awake at conventional times. Sleep onset is usually well past midnight, and sleep offset time is typically past noon 9,11,283. When individuals with DSPS are made to conform to earlier retiring times this leads to marked sleep onset insomnia 9,203,285,289. DSPS often exhibits psychiatric comorbidities including psychological and functional difficulties such as personality disorders and depression 11,278,404. DSPS patients may show marked nervousness and lack of control of emotional expression 16. These characteristics may worsen social withdrawal, causing a loss of social cues in synchronizing their circadian rhythm and a further exacerbation of the circadian problem. The neurohormone melatonin regulates the timing of the central circadian pacemaker located in the Suprachiasmatic Nuclei (SCN) 162,405,406. Significant alterations in melatonin secretion in depressed patients during the acute phase of illness have been documented 353,354,391,392,407. Stemming from this observation, several strategies have been utilized to manipulate circadian rhythm to alleviate depression 353, Exogenous melatonin can modulate the timing of the major sleep-wake episode and have beneficial effects on mood 265,353. The aim of this study was to investigate the role of exogenous melatonin treatment as a
90 74 chronobiotic in ameliorating depressive symptomatology in DSPS patients. In addition, we evaluated urinary sulphatoxymelatonin (amt6s) as a marker of circadian phase and the effects of melatonin on sleep in DSPS patients. 3.3 MATERIALS AND METHODS Study population Thirteen males aged 35.6 ± 14.0 years and seven females aged 30.8 ± 12.4 years with an established diagnosis of DSPS participated in the study. The following exclusion criteria were applied: shift work, presence of other sleep disorders, age under 16 years, alcohol or drug abuse, current use of psychotropic medications or any other form of medication affecting melatonin secretion, active behavioural treatment, and severe psychiatric and neurological disorders. Prior to inclusion in the study participants were reevaluated based on the International Classification of Sleep Disorders 295, 2-week sleep logs and a clinical interview by a psychiatrist/sleep specialist. The mean bedtime and rising times according to the sleep logs were 0247 h (range h) and 1045 h (range h) respectively. The study protocol was approved by the Research Human Ethics Committee of the University Health Network, and all patients signed consent forms prior to being assessed Study design The trial was randomized, double-blind, crossover, placebo-controlled and was conducted over nine consecutive weeks during one season in order to control for seasonal variations in endogenous melatonin levels. Since all patient enrolment and data collection was completed before 2004, the study was not registered as a clinical trial. All patients were divided into two groups based on the presence or absence of comorbid depressive features as judged by a structured clinical assessment, including HDRS and CES-D scale 369. Patients diagnosed who scored 17 or greater on the CES-D scale were assigned to the group of DSPS patients with
91 75 comorbid depressive symptoms. Following the clinical interview and psychometric evaluation to assess depression, participants spent two consecutive nights at the sleep clinic for overnight polysomnographic (PSG) sleep studies. Sleep studies were conducted on two consecutive nights to negate a possible confounding effect of the first night 373 on sleep data. This set of psychometric and sleep studies comprised the baseline evaluation. During the baseline sleep studies, participants chose their own retiring and wake up times to be consistent with their normal routines. After the second night of sleep studies, patients were randomly assigned to either the placebo group or the melatonin treatment group. Each participant took 5 mg/day of either melatonin or placebo between 1900 and 2100 h (Day 1). Both melatonin and placebo capsules were supplied by Penn Pharmaceutical Limited, UK. Treatment continued for 4 weeks followed by a 1-week washout period prior to treatment crossover. Two-night sleep studies were again conducted on the last two nights of melatonin and placebo treatment (Day 27 and 28; Day 62 and 63) Sleep physiology measures The sleep studies after the baseline evaluation had an imposed sleep period from midnight to 08:00 h. The imposed sleep period was used only during the two nights of sleep studies, once while on melatonin and once again during placebo treatment. During the rest of the trial participants were allowed to adhere to their habitual bedtime. Evening exposure to light was not controlled while they were at home during the duration of the study. PSG results obtained on the first night on each occasion were not included in the analysis due to a possible first night effect 373. A standard montage including electroencephalography, electrooculography, electromyography and respiratory monitoring (oxygen saturation, nasal airflow, and breathing effort) was used. The polysomnographs were scored by a single blinded scorer according to standardized criteria 356. The sleep parameters included sleep continuity (Sleep Onset Latency
92 76 (SOL), Total Sleep Time (TST), Sleep Efficiency (SE), and Wakefulness After Sleep Onset (WASO); Arousal Index (AI), sleep architecture (% of each sleep stage; Slow Wave Sleep% in the first and second sleep cycles), and REM sleep details (REM latency; Number of REM episodes) were analyzed Psychometric measures During the initial clinical interview participants were dichotomized based on the presence (n = 8) or absence (n = 12) of comorbid depressive features and a CES-D score of 17 or greater. This categorization, however, did not affect the order of treatment (melatonin/placebo) received. Both HDRS and CES-D scales 369 were used; the former was completed by the psychiatrist, the latter by patients. Clinical interviews and psychometric evaluations were conducted before the start of the study (baseline) and on the second night of each treatment period (placebo or melatonin) corresponding to the sleep study sessions and the end of each treatment phase Circadian profile measures Urine samples for amt6s measurements were collected during the 24 h period at 2100, 0300, 0900 and 1500 h. Urine was collected during placebo treatment for all participants. During the dark period samples were collected under dim light conditions (safe red light less than 5 Lux). Immunoassay for urinary amt6s was carried out per manufacturer s instructions (CIDtech Research Inc., Ont., Canada) Data analysis One-way analysis of variance (ANOVA) was used to determine statistical significance of sleep variables, subjective scores and 6-sulphatoxymelatonin (amt6s) levels between baseline, melatonin and placebo treatments. Significance based on the ANOVA (p < 0.05) was followed by Tukey post-hoc paired comparisons to determine statistically different groups (baseline,
93 77 melatonin, placebo treatment) using the Statistical Package for the Social Sciences. Results in the text are expressed as mean ± Standard Error of Mean (SEM). 3.4 RESULTS Melatonin treatment on depression in DSPS Group I (DSPS patients with comorbid depressive symptoms; n = 8) included three women and five men (mean age: 31.5 ± 7.2 years). Group II (DSPS patients without comorbid depressive symptoms n = 12) included four women and eight men (mean age: 36.2 ± 15.7 years). The mean scores on the CES-D and HDRS-17 were significantly higher in Group I (33.5 ± 9.4 and 13.2 ± 3.6) than in Group II (13.3 ± 3.6 and 5.6 ± 2.4). While at baseline both scales indicated depression in Group I, after melatonin treatment there was a significant decrease in HDRS-17 (Fig. 3.1a) and CES-D (Fig. 3.1b) scores, whereas placebo treatment had no effect on either score (Fig. 3.1a and b). Changes on the depression scales in Group II were not significant in any limb of the trial (Fig. 3.1c and d) Melatonin treatment on sleep architecture in DSPS Sleep variables in DSPS patients with comorbid depression (Group I) during the course of the double-blind, placebo-controlled study are presented in Table 3.1. In Group I mean lights out time at baseline was at 0220 h (range: h). The mean sleep offset time on this occasion was at 1040 h (range: h). On melatonin treatment, the sleep onset latency was significantly reduced compared to both placebo and baseline measures. There were no significant differences in the other sleep continuity variables (sleep efficiency, WASO, arousal index, and alpha scores) other than TST at baseline, melatonin and placebo treatments. Sleep architecture (percentage of each sleep stage and REM sleep parameters) was also similar for all three conditions of the trial. Interestingly, at baseline a reversal of SWS from the first to the second sleep cycle was noted. SWS distribution on melatonin and placebo was normal.
94 78 Figure 3.1: Ameliorating depression in delayed sleep phase patients with exogenous melatonin treatment. Patients were treated with 5 mg exogenous melatonin or placebo, each treatment lasting 4 weeks. Depression was rated using the Center for Epidemiologic Studies Depression Scale (CES-D) and the Hamilton Depression Rating Scale (HDRS-17). DSPS patients with comorbid depression had a significant decrease in their HDRS-17 (a) and CES-D (b) depression ratings while on melatonin treatment but not on placebo treatment. DSPS patients with no comorbid depression did not show any significant alterations in their HDRS-17 (c) or CES-D (d) depression ratings on melatonin or placebo treatment. Bars represent mean ± SEM. Bars with different letters are statistically different from one another as determined by Analysis Of Variance (ANOVA) followed by Tukey post-hoc paired comparisons analysis. Significance was set to p < 0.05.
95 79
96 80 Table 3.1: Sleep polysomnographic variables during the course of a 9-week randomized, double-blind placebo-controlled crossover study in patients with Delayed Sleep Phase Syndrome (DSPS) and comorbid depression (n = 8). Sleep variables Baseline Melatonin Placebo F p Sleep continuity Sleep onset latency (SOL): minutes 31.6±9.4 a 15.3±5.4 b 61.4±15.3 c Total sleep time (TST): minutes 487.7±19.1 a 436.0±17.8 a 383.7±19.7 b Sleep efficiency 92.9± ± ± n/s Wakefulness after sleep onset (WASO) 36.5± ± ± n/s Arousal index 15.4± ± ± n/s Alpha score 2.9± ± ± n/s Sleep architecture %stage 1 2.8± ± ± n/s %stage ± ± ± n/s %stage 3 7.2± ± ± n/s %stage ± ± ± n/s %REM 20.8± ± ± n/s SWS in the 1 st cycle 23.0± ± ± n/s SWS in the 2 nd cycle 47.6± ± ± n/s REM sleep REM latency: minutes 83.1± ± ± n/s Number of REM episodes 4.4± ± ± n/s p-values <0.05 for Tukey post-hoc analysis. Different letters signify statistical difference.
97 81 Polysomnographic findings in DSPS patients without symptoms of depression (Group II) are presented in Table 3.2. In Group II mean lights out time at baseline was at 0125 h (range: h). The mean sleep offset time at baseline in Group II was at 1020 h (range: h). Similar to Group I, melatonin induced a significant advance in sleep onset latency compared to placebo and baseline conditions. TST was also significantly reduced on placebo treatment compared to melatonin treatment as observed with Group I. All other sleep continuity and architecture parameters were similar on baseline, melatonin treatment and placebo in Group II. Comparison between sleep architecture parameters in Group I and Group II did not reveal any statistically significant differences Circadian profile of DSPS patients with and without comorbid depression DSPS patients with marked depressive symptoms showed an abnormal circadian pattern of endogenous melatonin secretion on placebo treatment (Fig. 3.2a). Their peak excretion of a major melatonin metabolite, 6-sulphatoxymelatonin (amt6s), did not occur at night as would normally be expected, but was observed in the morning at 0800 h (Fig. 3.2a). The DSPS patients without comorbid depression had the usual nocturnal rise in melatonin production (Fig. 3.2b). 3.5 DISCUSSION Numerous studies have linked disruptions in the sleep-wake cycle and fundamental circadian dysfunctions to affective illnesses 16,265,403, ; however, to the best of our knowledge, this study is the first to demonstrate that exogenous melatonin treatment in individuals with endogenous circadian rhythm disruption significantly ameliorates depressive symptomatology. In our sample of 20 DSPS patients 8 individuals had comorbid depressive symptomatology, and even though our sampling may have been biased due to patient recruitment
98 82 Table 3.2: Sleep polysomnographic variables during the course of a 9-week randomized, double-blind, placebo-controlled crossover study in non-depressed patients with DSPS (n = 12). Sleep variables Baseline Melatonin Placebo F p Sleep continuity Sleep onset latency (SOL) (min.) 38.6±14.5 a 21.2±5.6 b 58.9±8.9 c Total sleep time (TST) (min.) 513.7±15.4 a 418.3±8.9 a 362.0±9.8 c Sleep efficiency (%) 88.6± ± ± n/s Wakefulness after sleep onset (WASO) 46.3± ± ± n/s (min.) Arousal index 6.4± ± ± n/s Alpha score 2.0± ± ± n/s Sleep architecture %stage 1 3.5± ± ± n/s %stage ± ± ± n/s %stage 3 5.3± ± ± n/s %stage 4 7.3± ± ± n/s %REM 19.9± ± ± n/s SWS in the 1 st cycle (min.) 41.3± ± ± n/s SWS in the 2 nd cycle (min.) 16.9± ± ± n/s REM sleep REM latency (min.) 83.7± ± ± n/s Number of REM episodes 4.4± ± ± n/s p-values <0.05 for Tukey post-hoc analysis. Different letters signify statistical difference.
99 83 Figure 3.2: Altered melatonin secretion profile in DSPS patients with comorbid depression compared to DSPS patients with no depression. Urinary 6-sulphatoxymelatonin (amt6s) levels were measured four times over a 24-h period while patients were on placebo treatment. DSPS patients with comorbid depression had marked alterations in their melatonin secretion profile (a) compared to DSPS patients with no depression (b). Bars represent mean ± SEM.
100 84 from a sleep and neuropsychiatry clinic (leading to increased prevalence rates), several studies have demonstrated a high prevalence of depression in DSPS patients 11,16,411,418. The circadian phase of several hormonal rhythms is delayed in DSPS patients compared to healthy controls 419. A functional disturbance in the timing of these rhythms with the timing of sleep-wake rhythm has also been demonstrated 419. Therefore, DSPS patients may have double desynchronization: one that is internal and exists between different endogenous rhythms, such as melatonin secretion and sleep-wake cycle, and at another level, an external desynchronization that exists between endogenous circadian rhythms with the external light dark cycle and social clock times. Winter depression, or Seasonal Affective Disorder, is also associated with internal circadian misalignment between those rhythms related to the sleep-wake cycle and those that are more tightly coupled to the endogenous circadian pacemaker 276,420. At baseline the DSPS patients with depressive symptoms displayed a SWS reversal in the first and second sleep cycles which is a characteristic finding in depression 259,284,421,422. Increased arousal index was another distinguishing feature of sleep in DSPS patients with and without depressive symptoms. Interestingly, the common polysomnographic markers for depression such as short REM sleep latency and increased duration of REM sleep 7,317,376,417 were not observed in DSPS patients with depressive features compared to non-depressed DSPS patients. Sleep studies carried out during melatonin and placebo treatment were under imposed sleep periods ( h), whereas, under baseline conditions, patients were allowed to choose their own retiring and wake up times. Under an imposed sleep period the patients had increased SOL and increased difficulty in sleep initiation, resulting in decreased TST. With melatonin treatment, however, even under imposed sleep periods SOL and difficulties with sleep initiation were decreased, resulting in TST that was not significantly different from levels observed with a non-imposed sleep period. Sleep efficiency showed a corresponding non-
101 85 significant decrease likely resulting from the decreased TST under an imposed sleep period with placebo treatment. Although our study showed no major impact of melatonin on sleep architecture in DSPS patients, further studies are warranted to investigate the effect of melatonin on more precise and intrinsic mechanisms involved such as REM density in different cycles and REM sleep and SWS distribution. One of the principal differences between DSPS patients with and without depressive symptoms was observed in their melatonin excretion patterns. DSPS patients with comorbid depressive symptomatology did show a marked delay in their melatonin rhythm compared to non-depressed DSPS patients. The profile of endogenous melatonin secretion is a reliable circadian phase marker 423 which suggests that DSPS patients with depressive symptoms have a greater degree of circadian rhythm disturbance. Altered rhythm of melatonin secretion in DSPS has been reported, and it has also been suggested that not only the delay of the circadian clock, but also a functional disturbance of the sleep-wake mechanism underlies DSPS 9,203,424,425. Furthermore, our study lends support for melatonin rhythms serving as a marker for depression. Though our current sample size was limited, the marked alterations in melatonin rhythms observed in this study warrant further investigation of the association between the extent of circadian phase misalignment and depression symptom severity. Further studies with greater resolution in phase angle determination are warranted to correlate the degree of circadian misalignment with symptom severity in depressed DSPS patients. Based on our findings we can conclude that there is a strong association between circadian rhythm disorder and depression and that melatonin treatment ameliorates depressive symptomatology. Whether exogenous melatonin treatment corrected the underlying circadian misalignment in the present study cannot be concluded directly from the results of this study; however, several studies have demonstrated that exogenous melatonin treatment can modulate
102 86 the phase of the endogenous pacemaker, and, more importantly, administration of exogenous melatonin during the early evening induces phase advances 148,172,173,193. It is likely then that melatonin treatment phase advances the delayed circadian rhythms in DSPS patients and may mediate its beneficial effects on ameliorating depressive symptomatology in DSPS patients through this chronobiotic mechanism. A steady internal and external phase relationship appears to be crucial for a stable and euthymic mood state (i.e., the timing between core body rhythms such as cortisol and temperature as well as the timing of sleep with respect to the day night cycle) 265,353,387. Any misalignment may bring with it the propensity for mood fluctuation.
103 87 4 SELECTIVELY FILTERING SHORT WAVELENGTHS ATTENUATES THE DISRUPTIVE EFFECTS OF NOCTURNAL LIGHT ON ENDOCRINE AND MOLECULAR CIRCADIAN PHASE MARKERS IN RATS Shadab A Rahman 1,2, Alexandra Kollara 1, Theodore J Brown 1,2, Robert F Casper 1,2 1 Fran and Lawrence Bloomberg Department of Obstetrics and Gynecology, and Samuel Lunenfeld Research Institute, Mount Sinai Hospital, Toronto, Ontario, Canada, M5G 1X5; 2 Department of Physiology, University of Toronto, Toronto, Ontario, Canada, M5S 3G5. This work was published in the journal Endocrinology 2008, Dec; 149(12): [Epub 2008 Aug 7] NOTES: SAR completed data collection, analysis, interpretation and manuscript preparation. AK assisted with study design and training SAR in molecular biology techniques. SAR, TJ and RFC were involved with study design and interpretation.
104 ABSTRACT Various physiological processes exhibit a circadian rhythm synchronized to the geophysical light/dark cycle. Our study using a rat model demonstrated that exposure to light at night suppressed the expected nocturnal rise in melatonin, increased plasma corticosterone, and disrupted core clock gene expression in the hypothalamus and the adrenal gland. These effects were prevented by filtration of a 10-nm bandwidth of light between 470 and 480 nm, whereas filtration of light between 452 and 462 nm prevented the rise of corticosterone without restoring normal melatonin secretion or hypothalamic clock gene expression. This is the first demonstration of a wavelength dependency of glucocorticoid secretion and clock gene expression. Our results in an animal model suggest that filtering a narrow bandwidth of light from nocturnal lighting may efficiently attenuate overall disruption of circadian endocrine rhythms and clock gene expression in the hypothalamus and adrenal gland. Because a narrow bandwidth of light is filtered, the color distribution of the illumination source is not altered, and this may be of practical importance for potential future studies in shift workers.
105 INTRODUCTION Numerous physiological processes including the sleep/wake cycle, alertness, mood, hormone secretion, body temperature, feeding/drinking, and locomotor activity exhibit a circadian rhythm that promotes organism homeostasis 426. The rhythms originate from the master biological clock located in the suprachiasmatic nuclei (SCN) 427. Environmental time cues such as the geophysical light/dark cycle can act on the SCN to reset endogenous phase and periodicity of these rhythms enabling synchronization and entrainment of the rhythms to a 24-h period 427. Nocturnal light exposure and associated changes in entrainment cues can reset the SCN and disrupt almost all of these essential physiological rhythms, subsequently altering organism homeostasis 2,334,428,429. Nocturnal light exposure in shift workers, and presumably subsequent disruption of circadian rhythms, has been linked to pathological conditions such as breast 332,430 and colorectal cancer 335, cardiovascular disease, depression and anxiety disorders 334,428, obesity, and type 2 diabetes 340,431. More than 11 million North Americans, employed in various industries, are currently involved in shift work and the associated morbidity and mortality has become a major health concern 334. Several strategies have been tested for correcting circadian rhythm disruption induced by nocturnal light exposure. Many of the adverse health effects thought to be due to nocturnal light exposure have been associated with melatonin suppression 146,341,343. Thus, most of these approaches primarily focused on modulating melatonin secretion, such as alteration of the melatonin onset and peak time by using bright light exposure to correspond to daytime sleep in night shift workers 221,432. However, these strategies have proven difficult to implement in rotating shifts, the most common type of shift work 433. A complete inversion of the circadian rhythm is seldom reached, even across successive night shifts and after years of night work experience 340. Furthermore, clinical and animal studies demonstrate significant alteration in gene
106 90 expression rhythms and other endocrine rhythms besides melatonin under nocturnal light exposure that may all contribute to pathogenesis associated with shift work 224,434,435. Recent human and animal studies demonstrate a major role of clock genes in mediating various health disorders ranging from obesity to seasonal affective disorders 397,436,437. Furthermore, elevated cortisol levels have been linked to cardiovascular diseases 438,439, depression, and metabolic disorders 440,441. All of these health disorders are also associated with night shift work 334,442,443, suggesting a possible role of clock genes and glucocorticoids and disruptions in their circadian rhythms in mediating the deleterious effects of night-shift work. In humans and rodents, circadian rhythm responses to light starts at the retina, and recent studies demonstrate that intrinsically photosensitive melanopsin expressing retinal ganglion cells (RGCs) play a principal role in mediating these responses 444,445. The traditional rods and cones, which are the principal photoreceptors associated with vision, can modulate RGC response but only in a duration, irradiance level, and wavelength-dependent manner 107,435. The peak activation of intrinsically photosensitive RGCs occurs in the range of the maximal absorption spectrum of melanopsin ( nm) 48,445. RGCs convey light signals directly to the SCN via the retinohypothalamic tract 444,446 eliciting light-induced circadian alterations by modulating SCN activity 102. Pineal melatonin secretion is regulated by the SCN, making it a reliable circadian phase marker, and its secretion is acutely suppressed by nocturnal light exposure 172,341. The adrenal hormone cortisol also shows a marked circadian rhythm under the control of the SCN and nocturnal light exposure increases cortisol secretion in a time-of-day-dependent manner in both humans and rodents 133,139. Interestingly, nocturnal light-induced suppression of melatonin secretion is wavelength dependent, with short wavelengths (between 420 and 520 nm) having a pronounced effect and maximal suppression occurring in the 446 to 477 nm range 120,121 which is
107 91 coincident with the RGC absorption spectrum. This wavelength dependency is also observed in light-mediated phase shifts in locomotor activity 109, providing further support for RGC activity regulating light-mediated circadian alterations. However, such a wavelength dependency has not been demonstrated for glucocorticoid secretion in humans or rodents. In the current study, we hypothesized that filtering short wavelengths in the range of maximal RGC activity may effectively attenuate circadian rhythm disruptions. Whereas we previously demonstrated that blocking all wavelengths less than 530 nm is effective in normalizing melatonin secretion in humans 365,366, the objective of the current study was to determine whether blocking a narrow bandwidth of wavelengths in the maximal activation range of RGCs would be effective in normalizing melatonin secretion as well as other circadian phase markers such as glucocorticoid secretion and clock gene expression under nocturnal light exposure. Melatonin and corticosterone secretion are established circadian phase markers and are routinely used to examine alterations in the phase and periodicity of endogenous rhythms 423,447, MATERIALS AND METHODS Animals Ten-week-old male Sprague Dawley rats were obtained from Charles River Laboratories (Montréal, Canada). We used male animals to avoid possible confounding effects of estrus cyclicity. Before all experiments the rats were entrained for at least 2 wk in a 12-h light, 12-h dark cycle (0800 h lights on; 2000 h lights off, with 450 Lux light intensity at animal eye level during the light phase; all times are presented as clock time, i.e. the time of day, to distinguish from circadian time and Zeitgeber time commonly used in circadian physiology). All animals were allowed ad libitum feeding on standard rat chow and were handled as per approved protocol by the Samuel Lunenfeld Research Institute Animal Care Committee.
108 Light exposure protocol Light exposure experiments were carried out in three phases. Each experimental phase was repeated at least twice. The first phase determined the baseline (control) hormone levels and clock gene expression rhythms (n = 32). The second phase of the experiment was to determine the hormone levels and clock gene expression rhythms in animals exposed to 12 h light, 12 h light (continuous lighting) cycles (n = 32). The third phase of the experiment determined the melatonin and corticosterone secretion profile in rats exposed to 12 h light, 12 h light cycles with optically filtered lighting (n = 40). Total number of animals used in the study in all three phases was 104. To maintain light intensity and exposure at a constant level, a custom-designed optically sealed wooden box was used. The box did not allow external lighting to enter or internal light to escape, thus preventing alterations in light intensity within the holding conditions. The box was 24 x 24 x 14 in. and held two regulation-size rat cages. The rat cages were also modified such that more than 80% of each plastic side was replaced with galvanized steel mesh wire. This modification was to ensure that light could enter freely and not be refracted or filtered by the original plastic sides and still ensure that the animals were kept under regulation holding conditions. The dimensions and bedding used were the same as during entrainment. Each modified cage housed two non-breeding male animals. Animals were housed in pairs from the beginning of the entrainment period, and the same pairs were transferred to their modified holding cages to negate stress caused by exposure to a new animal. Animals were transferred from their colony room to the box at 2000 h. To ensure equal treatment, control animals were also maintained in the modified box but with no light exposure. During light exposure experiments (second and third phase), the animals were exposed to light from 2000 to 0800 h in addition to regular light exposure between 0800 and 2000 h. Light
109 93 was fed uniformly throughout the box using six evenly spaced FSPT-50 fiber optic bundles, each containing 50 random sorted end-emitting fiber optic cables (Magic Lite, Mississauga, Ontario, Canada) running from a fiber optic FS-7L illuminator (Magic Lite) fitted with a 183-W tungsten halogen MR-16 bulb (3000 K color temperature) generating 481 Lux photopic light intensity of full-spectrum polychromatic white light at animal eye level. During the third phase, light was filtered at the source of the light input for the optical fibers using holographic notch filters (Kaiser Optical Systems, Ann Arbor, MI) designed to block approximately 10-nm wavelength bandwidths. The two filters used selectively blocked wavelengths between 452 and 462 and 470 and 480 nm. When stacked together, the combined filters were able to block the range of approximately nm with the exception of an average of 28% transmission between 462 and 470 nm. Total scotopic illuminosity and spectral irradiance of unfiltered lighting was 481 Lux; mw/m 2, after nm filtration, 432 Lux; mw/m 2, after nm filtration, 424 Lux; mw/m 2 and after combined filtration, 387 Lux; mw/m 2. Spectral irradiance and illuminosity were measured using a calibrated Ocean Optics S2000 spectroradiometer (Dunedin, FL) Sample collection Starting from 2000 until 0800 h, animals (n = 8) were killed every 4 h for blood and tissue sample collection under darkness using a safe red light lamp (<5 Lux illuminosity) and with the aid of night vision equipment. The animals were anesthetized with isoflurane gas (5% induction; 3% maintenance), and 5 ml of blood were aspirated by cardiac puncture into tuberculin syringes (20 gauge, 1-in. needles) and stored on ice at 4 C in prechilled heparin coated tubes (BD Biosciences, Ontario, Canada) until all samples were collected. Immediately after blood collection, animals were euthanized by decapitation. The whole brain and the left adrenal
110 94 gland were harvested. The hypothalamus was dissected from the whole brain. All tissue was snap frozen in liquid nitrogen and stored at 80 C. After collection of all blood samples, the plasma was extracted by centrifuging the samples at 1000 x g for 15 min and then stored at 80 C Hormone assays All hormones were assayed in plasma. All frozen plasma samples were defrosted on ice and centrifuged again at 1000 x g for 15 min at 4 C before assaying. Melatonin was extracted from plasma before assaying by ELISA (Alpco Diagnostics, Salem, NH). ELISAs were also used for ACTH (Alpco Diagnostics), corticosterone (Cayman Chemical, Ann Arbor, MI), and cortisol (Cayman Chemical) as per the manufacturer s instructions Quantitative real time RT-PCR PCR primers used [forward (F) 5' to 3', reverse (R) 5' to 3'] were as follows: Per2 (NM_ ), F: GAC GGG TCG AGC AAA GGA, R: GGG AAA AGT CCA CAT ATC CAT TCA; Bmal1 (NM_ ), F: TCT ATC CGA TGA CGA ACT GAA ACA, R: CCC TCG GTC ACA TCC TAC GA; and GAPDH (XM_221254), F: AAC CTG CCA AGT ATG ATG ACA TCA, R: TGT TGA AGT CAC AGG AGA CAA CCT. Whole hypothalami and adrenal glands were homogenized, and total RNA was extracted with Trizol (Invitrogen Life Technologies, Ontario, Canada) according to the manufacturer s instructions. RNA absorbance was measured at 260 nm by µquant microplate spectrophotometer (Bio-Tek Instruments, Winooski, VT) to quantify RNA amounts, and samples were treated with DNaseI (Ambion, Ontario, Canada) according to the manufacturer s instructions. Total RNA (1 µg) was reverse transcribed using 200 ng random hexamers, 200 U SuperScript II reverse transcriptase, 10 mm dithiothreitol, and 1x first-strand buffer (Invitrogen) at 25 C for 10 min and 48 C for 30 min in a reaction volume of 20 µl (PTC-200 Peltier thermal cycler; Bio-Rad, Ontario, Canada). Real-time quantitative RT-PCR was performed for total RNA using SYBR
111 95 Green (Applied Biosystems, Ontario, Canada) and detected using the Applied Biosystems 7900 HT thermal cycler with 25 µl SYBR Green master mix (Applied Biosystems), 50 ng cdna, and primers in a total volume of 50 µl. Quantitative RT-PCR conditions were as follows: 95 C for 10 min, and then 40 cycles of 95 C for 15 sec and 60 C for 15 sec. Expression was quantified, using glyceraldehyde-3-phosphate dehydrogenase as a reference, by the comparative cycle threshold method, as per the manufacturer s instructions (Applied Biosystems). Dissociation curves, gel analysis, and sequencing of certain PCR products confirmed gene-specific product amplification Cell maintenance, treatments, and cell proliferation assay NCI-H295R, a human pluripotent adrenocortical carcinoma cell line was obtained from American Type Culture Collection (Manassas, VA) and grown as monolayer cultures in DMEM/Ham s F-12 nutrient 1:1 mix (Invitrogen) without phenol red and supplemented with 2.5% FBS-serum (BD Biosciences), 1% insulin/transferrin/selenium + premix (BD Biosciences), 0.1% penicillin/streptomycin, and 0.25% Fungizone (250 µg amphotericin B and 205 µg sodium deoxycholate per milliliter) (Invitrogen) at 37 C in a humidified CO 2 incubator. Cells were then plated to a density of 5 x 10 5 cells/well in six-well plates and serum starved for 24 h before treatment. Culture medium was replaced with 1 ml fresh serum-free medium with the addition of 1 µm forskolin (Sigma Aldrich, Ontario, Canada), 1 nm epinephrine (Sigma Aldrich), or 1 nm ACTH (Amphastar Pharmaceuticals, Rancho Cucamonga, CA) in the presence or absence of 1 nm melatonin (Sigma Aldrich). Cells were also treated with a combination of ACTH and epinephrine in the presence or absence of melatonin. All treatment compounds were dissolved in ethanol and diluted with culture medium before addition to cell cultures. Vehicle treatment consisted of an equivalent amount of ethanol diluted with culture medium. Treatments were carried out for 24 h, after which medium was collected for cortisol measurement by ELISA. Immediately after collection, 1 ml fresh serum-free medium was placed into each well with 150
112 96 µl of 5 mg/ml methylthiazolyldiphenyl-tetrazolium bromide (Sigma Aldrich) for cell proliferation assessment. Cells were incubated for 4 h at 37 C in a humidified CO 2 incubator after which the medium was aspirated and the metabolized formazan crystals were dissolved in dimethylsulfoxide for 5 min. Absorbance (relative OD) was measured at 570 nm with a µ-quant spectrophotometer (Bio-Tek Instruments) Western blot analysis Whole-cell lysates were extracted from cells suspended in radioimmunoprecipitation assay buffer (50 mm HEPES, 150 mm NaCl, 1% Triton X-100, 0.1% sodium dodecyl sulfate, 1% sodium deoxycholate) supplemented with Complete protease inhibitor cocktail (Roche, Ontario, Canada). Protein was quantitated by a BCA protein assay (Pierce, Rockford, IL) following the manufacturer s recommended protocol. Total protein lysates (20 µg) were loaded and resolved by 10% SDS-PAGE, transferred to nitrocellulose membranes, and blotted for 2 h at room temperature with affinity-purified goat polyclonal anti-mt1 or anti-mt2 antibodies (both at 1:500 dilution; both from Santa Cruz Biotechnology, Santa Cruz, CA). Immunoreactive proteins were visualized with horseradish peroxidase-labeled secondary donkey antigoat antisera (1:2500) (Santa Cruz Biotechnology) for 1 h and detected by enhanced chemiluminescence (Amersham Pharmacia Biotech, Ontario, Canada) Statistics Data obtained in the hormone assays and cell proliferation assays were expressed as the mean ± SEM and were subjected to ANOVA followed by Student-Newman-Keuls multiple comparison test (P < 0.05). Data obtained in the gene expression assays were expressed as mean relative fold change in gene expression ±95% confidence interval. Mean relative fold change values between two points were considered statistically different when the two confidence intervals did not intersect.
113 RESULTS Normalization of Circadian Endocrine Phase Markers under Nocturnal Light Exposure Exposure to unfiltered nocturnal lighting (481 Lux; mw/m 2 ) significantly suppressed the night time increase in melatonin secretion observed in the control group and maintained melatonin secretion at basal levels (peak melatonin value in light exposed animals was 63.1 ± 2.8 pg/ml; in control animals ± 9.1 pg/ml; P < 0.001). Filtering short wavelengths between 450 and 480 nm with the combined notch filters (387 Lux; mw/m 2 ) restored melatonin secretion, generating a peak that corresponded to that observed in the control group (peak melatonin value in filtered light exposed animals 99.0 ± 6.9 pg/ml; in control animals, ± 9.1 pg/ml; P > 0.05) (Fig. 4.1A). Nocturnal light exposure resulted in a significant increase in plasma corticosterone levels at 2400, 0400, and 0800 h compared with levels measured in control animals (P < 0.001) (Fig. 4.1B). Filtering light wavelengths between 450 and 480 nm effectively normalized corticosterone secretion and inhibited the increase in corticosterone secretion observed with unfiltered light exposure (P > 0.05) (Fig. 4.1B) ACTH-Independent Modulation of Glucocorticoid Secretion in Response to Nocturnal Light To examine whether the ACTH pathway is involved in light-induced changes in corticosterone levels, we measured plasma ACTH levels under unfiltered and filtered light exposure and compared them with levels measured in control animals kept in darkness. ACTH
114 98 Figure 4.1: Normalization of melatonin and glucocorticoid secretion independent of ACTH. Normalization of nocturnal light exposure induced disruption of melatonin (A) and corticosterone (B) secretion in male Sprague Dawley rats by filtering short wavelengths ( nm). Nocturnal light-induced disruption of corticosterone secretion was not associated with changes in ACTH levels (C). Animals were exposed to bright polychromatic white light, filtered bright polychromatic white light, or no light (control). Starting from 2000 to 0800 h, every 4 h (n = 8 per time point), blood was collected for measuring hormone levels by ELISA. Data represent the mean ± SEM. Statistical differences between groups were analyzed by one-way ANOVA followed by Student-Newman-Keuls multiple comparison test. Significance was set to P < **, Significant difference of P < 0.001; compared with levels obtained with light exposure.
115 99
116 100 levels were not significantly different between any of these three conditions at any time point throughout the 12 h of testing (P > 0.05) (Fig. 4.1C) Normalization of central and peripheral clock gene expression patterns Our results demonstrate that exposure to unfiltered nocturnal light disrupts clock gene expression. Per2 expression was significantly increased compared with controls at 2400, 0400, and 0800 h in the hypothalamus (Fig. 4.2A) and at 0400 and 0800 h in the adrenal gland (Fig. 4.2C). Bmal1 expression was increased in the hypothalamus only at 0400 h, 4 h after significantly increased Per2 expression (Fig. 4.2B). Bmal1 expression in the adrenal gland was significantly increased compared with controls at 0800 h, again 4 h after significantly increased Per2 expression (Fig. 4.2D). Filtering short wavelengths from nocturnal light effectively normalized clock gene expression both in the hypothalamus (Fig. 4.2, A and B) and the adrenal gland (Fig. 4.2, C and D) Differential wavelength-dependent normalization of endocrine circadian phase markers and clock gene expression We studied the effectiveness of filtering narrower ranges of short wavelengths in preserving physiologic and endocrine rhythms. We subsequently tested each of the notch filters alone to selectively block a 10 nm range of wavelengths, resulting in maximum transmission at all other wavelengths (Fig. 4.3, A-E). Spectral irradiance distributions (Fig. 4.3, A and B) and color chart comparisons (Fig. 4.3, C-E) demonstrate that the notch filters effectively block a selective 10 nm bandwidth and allow maximal transmission at all other wavelengths, resulting in minimal color distortion. We examined the effects of each notch filter individually on melatonin and corticosterone secretion profiles and on clock gene expression under unfiltered or filtered nocturnal lighting conditions compared with control conditions. Melatonin secretion was normalized by the 470 to
117 101 Figure 4.2: Normalization of Per2 and Bmal1 gene expression in the hypothalamus and adrenal gland. Normalization of nocturnal light exposure induced disruption of Per2 (A) and Bmal1 (B) gene expression in the hypothalamus and Per2 (C) and Bmal1 (D) gene expression in the adrenal gland in male Sprague Dawley rats by filtering short wavelengths ( nm). Animals were exposed to bright polychromatic white light, filtered bright polychromatic white light, or no light (control). Starting from 2000 to 0800 h, every 4 h (n = 8 per time point), the hypothalamus and left adrenal gland were collected and clock gene expression was measured by quantitative realtime RT-PCR. Data represent mean relative fold change in gene expression ±95% confidence interval. **, Statistically significant difference compared with levels obtained with light exposure.
118 102 Figure 4.3: Wavelength-dependent differential normalization of endocrine circadian phase markers and tissue-specific clock gene expression. Spectral irradiance distribution of the different lighting conditions between 380 and 780 nm at 1nm intervals (A) and between 440 and 520 nm (RGC activation range) at 1-nm intervals (B). Color charts obtained with nm filtration (C), no filtration (D), and nm filtration (E), showing minimal color distortion after filtering. Normalization of nocturnal light exposure induced disruption of melatonin (F) and corticosterone (G) secretion and Per2 clock gene expression in the hypothalamus (H) and adrenal gland (I) in male Sprague Dawley rats by filtering short wavelengths using the two individual notch filters. Melatonin secretion was normalized using only the 470 to 480 nm filter and not the 452 to 462 nm filter, whereas corticosterone secretion was normalized by both filters. Per2 gene expression was normalized in both the hypothalamus and adrenal gland using only the 470 to 480 nm filter, whereas the 452 to 462 nm filter was effective in normalizing Per2 expression only in the adrenal gland. Animals were exposed to bright polychromatic white light, filtered bright polychromatic white light, or no light (control). Starting from 2000 to 0800 h, every 4 h (n = 4 per time point), blood was collected for measuring hormone levels by ELISA. Data represent the mean ± SEM. Statistical differences between groups were analyzed by ANOVA followed by Student-Newman-Keuls multiple comparison test. Significance was set to P < **, Significant difference compared with levels obtained with light exposure;, significant difference in gene expression levels between 452 and 462 nm and 470 and 480 nm filtered light exposure).
119 103
120 nm filter (432 Lux; mw/m 2 ) but not the 452- to 462-nm filter (424 Lux; mw/m 2 ) (Fig. 4.3F), whereas corticosterone secretion was normalized by either filter (Fig. 4.3G). Whereas the 470 to 480 nm filter normalized clock gene expression in the hypothalamus (Fig. 4.3H) and the adrenal gland (Fig. 4.3I), the 452 to 462 nm filter normalized clock gene expression only in the adrenal gland (Fig. 4.3I). The 452 to 462 nm filter reduced the nocturnal light induced increase in hypothalamic Per2 expression, although the levels were still significantly elevated compared with control levels until 2400 h (Fig. 4.3H), the time of peak melatonin secretion Cortisol secretion modulation by melatonin in human adrenocortical cells NCI-H295R, a human adrenocortical carcinoma cell line, was used to examine whether melatonin treatment can modulate cortisol secretion. The presence of both melatonin receptors, MT1 and MT2, was determined in this cell line by Western blot analysis (Fig. 4.4A). NCI- H295R cells were treated with forskolin, ACTH, or epinephrine in the presence or absence of melatonin. Melatonin treatment did not affect basal cortisol secretion. Forskolin, epinephrine, and ACTH treatments significantly increased cortisol secretion from control levels by 63.9, 17.9, and 27.0%, respectively, whereas melatonin treatment partially but significantly attenuated the induced increase in cortisol secretion by 13.0, 21.6, and 12.7%, respectively (Fig. 4.4B). 4.5 DISCUSSION Both melatonin and glucocorticoid secretion demonstrate robust circadian rhythmicity. The phase and periodicity in their secretion rhythms is directly regulated by the SCN 127,449, making them reliable circadian phase markers. A disruption or change in the timing of the endogenous circadian pacemaker is directly evinced by alterations in the rhythmicity of these phase markers 202. Therefore, light exposure mediated RGC activity may in turn modulate glucocorticoid secretion by causing photic resetting of the endogenous circadian pacemaker and
121 105 Figure 4.4: Inhibition of camp-mediated cortisol secretion from NCI-H295R human adrenocortical cells by melatonin at physiological levels. The presence of both the melatonin receptors, Mt1 and Mt2, was determined in the NCI-H295R cell line by Western blot analysis (A). Furthermore, the presence of both melatonin receptors was determined in the rat adrenal gland (A). NCI-H295R cells were treated with 1 µm forskolin, 1 nm epinephrine, or 1 nm ACTH in the presence or absence of 1 nm melatonin (B). Cells were also treated with melatonin alone or equivalent amount of ethanol (vehicle control). Culture medium was collected 24 h later and assayed for cortisol by ELISA. Bars, mean ± SEM of three independent cell cultures. Bars with different letters are statistically different from one another as determined by ANOVA followed by Student-Newman-Keuls multiple comparison test. Significance was set to P < 0.05.
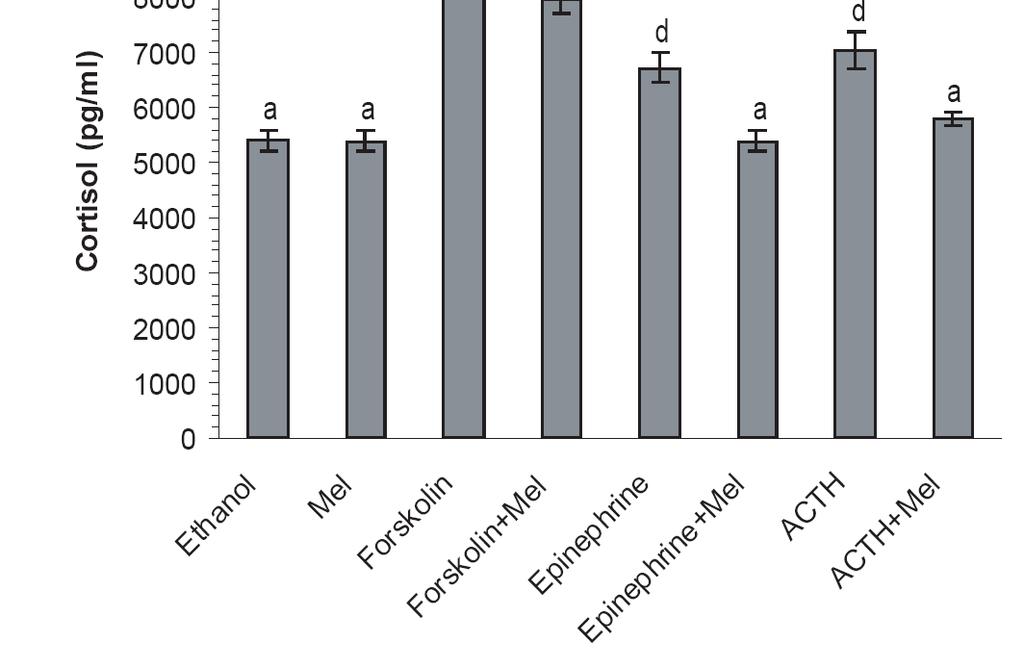
122 106
Sleep-Wake Cycle I Brain Rhythms. Reading: BCP Chapter 19
 Sleep-Wake Cycle I Brain Rhythms Reading: BCP Chapter 19 Brain Rhythms and Sleep Earth has a rhythmic environment. For example, day and night cycle back and forth, tides ebb and flow and temperature varies
Sleep-Wake Cycle I Brain Rhythms Reading: BCP Chapter 19 Brain Rhythms and Sleep Earth has a rhythmic environment. For example, day and night cycle back and forth, tides ebb and flow and temperature varies
Biological Clocks. Lu Chen, Ph.D. MCB, UC Berkeley. What is biological clock?
 Biological Clocks Lu Chen, Ph.D. MCB, UC Berkeley 1 What is biological clock? All eukaryotes and some prokaryotes display changes in gene activity, biochemistry, physiology, and behavior that wax and wane
Biological Clocks Lu Chen, Ph.D. MCB, UC Berkeley 1 What is biological clock? All eukaryotes and some prokaryotes display changes in gene activity, biochemistry, physiology, and behavior that wax and wane
Biological rhythms. Types of biological rhythms
 Biological rhythms Types of biological rhythms 2/33 what do we call rhythm in a living organism? physiological events occurring at approximately regular times internally controlled rhythms: breathing,
Biological rhythms Types of biological rhythms 2/33 what do we call rhythm in a living organism? physiological events occurring at approximately regular times internally controlled rhythms: breathing,
Biological Clocks. Lu Chen, Ph.D. MCB, UC Berkeley. Why Does Melatonin Now Outsell Vitamin C??
 Biological Clocks Lu Chen, Ph.D. MCB, UC Berkeley 1 Why Does Melatonin Now Outsell Vitamin C?? Wake / sleep complaints are extremely prevalent. Much melatonin is consumed in an attempt to overcome the
Biological Clocks Lu Chen, Ph.D. MCB, UC Berkeley 1 Why Does Melatonin Now Outsell Vitamin C?? Wake / sleep complaints are extremely prevalent. Much melatonin is consumed in an attempt to overcome the
PHYSIOLOGY AND MAINTENANCE Vol. V - Biological Rhythms - Tarja Porkka-Heiskanen, Jarmo T. Laitinen
 BIOLOGICAL RHYTHMS Tarja Porkka-Heiskanen, Institute of Biomedicine, University of Helsinki, Finland Jarmo T. Laitinen Department of Physiology, University of Kuopio, Finland Keywords: Light, melatonin,
BIOLOGICAL RHYTHMS Tarja Porkka-Heiskanen, Institute of Biomedicine, University of Helsinki, Finland Jarmo T. Laitinen Department of Physiology, University of Kuopio, Finland Keywords: Light, melatonin,
CIRCADIAN SIGNALING NETWORKS
 Transcription Regulation And Gene Expression in Eukaryotes Cycle G2 (lecture 13709) FS 2014 P. Matthias and RG Clerc Roger G. Clerc 07.05.2014 CIRCADIAN SIGNALING NETWORKS Master pacemaker SCN «Slave clocks»
Transcription Regulation And Gene Expression in Eukaryotes Cycle G2 (lecture 13709) FS 2014 P. Matthias and RG Clerc Roger G. Clerc 07.05.2014 CIRCADIAN SIGNALING NETWORKS Master pacemaker SCN «Slave clocks»
Transcription Regulation And Gene Expression in Eukaryotes (Cycle G2 # )
") Transcription Regulation And Gene Expression in Eukaryotes (Cycle G2 #13709-01) CIRCADIAN SIGNALING NETWORKS RG. Clerc May 19. 2010 www.fmi.ch/training/teaching Circadian rythms : most physiological processes
Transcription Regulation And Gene Expression in Eukaryotes (Cycle G2 #13709-01) CIRCADIAN SIGNALING NETWORKS RG. Clerc May 19. 2010 www.fmi.ch/training/teaching Circadian rythms : most physiological processes
Hypothalamus. Small, central, & essential.
 Hypothalamus Small, central, & essential. Summary: You can t live without a hypothalamus. Located at the junction between the brain stem and the forebrain Medial hypothalamus: interface between the brain
Hypothalamus Small, central, & essential. Summary: You can t live without a hypothalamus. Located at the junction between the brain stem and the forebrain Medial hypothalamus: interface between the brain
Circadian Rhythm Disturbances: What Happens When Your Biological Clock Is In The Wrong Time Zone
 Circadian Rhythm Disturbances: What Happens When Your Biological Clock Is In The Wrong Time Zone Steven A. Thau MD Chief, Pulmonary, Sleep Department. Phelps Hospital, Northwell Health Internal Clock Examples
Circadian Rhythm Disturbances: What Happens When Your Biological Clock Is In The Wrong Time Zone Steven A. Thau MD Chief, Pulmonary, Sleep Department. Phelps Hospital, Northwell Health Internal Clock Examples
Neurobiology of Circadian Rhythms
 ARC-IBRO ISN Joined Neuroscience School Behavioural Bioassays in Neuroscience: Brain and Behavior From Invertabrates To Small Mammals 4-14 December 2014 ICIPE, Nairobi KENYA Neurobiology of Circadian Rhythms
ARC-IBRO ISN Joined Neuroscience School Behavioural Bioassays in Neuroscience: Brain and Behavior From Invertabrates To Small Mammals 4-14 December 2014 ICIPE, Nairobi KENYA Neurobiology of Circadian Rhythms
Circadian photoreception in humans: More than meets the eye
 DAYLIGHTING (4.430) MIT Architecture Circadian photoreception in humans: More than meets the eye Steven W. Lockley, Ph.D. Division of Sleep Medicine, Brigham and Women s Hospital, Boston, MA Division of
DAYLIGHTING (4.430) MIT Architecture Circadian photoreception in humans: More than meets the eye Steven W. Lockley, Ph.D. Division of Sleep Medicine, Brigham and Women s Hospital, Boston, MA Division of
Sleep, Dreaming and Circadian Rhythms
 Sleep, Dreaming and Circadian Rhythms People typically sleep about 8 hours per day, and spend 16 hours awake. Most people sleep over 175,000 hours in their lifetime. The vast amount of time spent sleeping
Sleep, Dreaming and Circadian Rhythms People typically sleep about 8 hours per day, and spend 16 hours awake. Most people sleep over 175,000 hours in their lifetime. The vast amount of time spent sleeping
Circadian rhythm and Sleep. Radwan Banimustafa MD
 Circadian rhythm and Sleep Radwan Banimustafa MD Homeostasis Maintenance of equilibrium by active regulation of internal states: Cardiovascular function (blood pressure, heart rate) Body temperature Food
Circadian rhythm and Sleep Radwan Banimustafa MD Homeostasis Maintenance of equilibrium by active regulation of internal states: Cardiovascular function (blood pressure, heart rate) Body temperature Food
Biological Rhythms. Today s lecture
 Biological Rhythms (a review of general endocrinology) 35 Neuroendocrine control: homeostatic responses and biological rhythms. A role for anticipation or feed-forward mechanisms or scheduled events. Biological
Biological Rhythms (a review of general endocrinology) 35 Neuroendocrine control: homeostatic responses and biological rhythms. A role for anticipation or feed-forward mechanisms or scheduled events. Biological
LESSON 4.5 WORKBOOK How do circuits regulate their output?
 DEFINITIONS OF TERMS Homeostasis tendency to relatively stable equilibrium. Feed-forward inhibition control mechanism whereby the output of one pathway inhibits the activity of another pathway. Negative
DEFINITIONS OF TERMS Homeostasis tendency to relatively stable equilibrium. Feed-forward inhibition control mechanism whereby the output of one pathway inhibits the activity of another pathway. Negative
The Success of Decomposition
 11/21/11 Mechanism and Levels of Organization: Recomposing and Situating Circadian Clocks The Success of Decomposition Moving beyond per, researchers in the 1990s and early 2000s identified many clock
11/21/11 Mechanism and Levels of Organization: Recomposing and Situating Circadian Clocks The Success of Decomposition Moving beyond per, researchers in the 1990s and early 2000s identified many clock
Neurons and Hormones 1. How do animals perform the right behaviors at the right time? In the right context?
 Neurons and Hormones 1 How do animals perform the right behaviors at the right time? In the right context? Active at night only What if conflicting signals? Magnetic cues are always present But migrate
Neurons and Hormones 1 How do animals perform the right behaviors at the right time? In the right context? Active at night only What if conflicting signals? Magnetic cues are always present But migrate
Neuroscience - Problem Drill 13: The Eye and Visual Processing
 Neuroscience - Problem Drill 13: The Eye and Visual Processing Question No. 1 of 10 needed, (3) Pick the answer, and (4) Review the core concept tutorial as needed. 1. Which of the following statements
Neuroscience - Problem Drill 13: The Eye and Visual Processing Question No. 1 of 10 needed, (3) Pick the answer, and (4) Review the core concept tutorial as needed. 1. Which of the following statements
Index. sleep.theclinics.com. Note: Page numbers of article titles are in boldface type.
 Note: Page numbers of article titles are in boldface type. A Accidents, at work, effect of shift work disorder on, 263 264 Acetylcholine, in circadian rhythms, 100 105 Acrophase, definition of, 301 Actigraphy,
Note: Page numbers of article titles are in boldface type. A Accidents, at work, effect of shift work disorder on, 263 264 Acetylcholine, in circadian rhythms, 100 105 Acrophase, definition of, 301 Actigraphy,
Transcription Regulation And Gene Expression in Eukaryotes FS 2016 Graduate Course G2
 Transcription Regulation And Gene Expression in Eukaryotes FS 2016 Graduate Course G2 P. Matthias and RG Clerc Pharmazentrum Hörsaal 2 16h15-18h00 CIRCADIAN SIGNALING NETWORKS Master pacemaker SCN «slave
Transcription Regulation And Gene Expression in Eukaryotes FS 2016 Graduate Course G2 P. Matthias and RG Clerc Pharmazentrum Hörsaal 2 16h15-18h00 CIRCADIAN SIGNALING NETWORKS Master pacemaker SCN «slave
Altered Entrainment to the Day/Night Cycle Attenuates the Daily Rise in Circulating Corticosterone in the Mouse
 University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Papers in Veterinary and Biomedical Science Veterinary and Biomedical Sciences, Department of 2014 Altered Entrainment to
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Papers in Veterinary and Biomedical Science Veterinary and Biomedical Sciences, Department of 2014 Altered Entrainment to
Clicker Question. The Need to Decompose. Mechanism and Reduction: Decomposing Circadian Clocks
 Mechanism and Reduction: Decomposing Circadian Clocks Clicker Question On the Deductive-Nomological (DN) model of reduction, which of the following does not figure in providing the explanation (i.e., is
Mechanism and Reduction: Decomposing Circadian Clocks Clicker Question On the Deductive-Nomological (DN) model of reduction, which of the following does not figure in providing the explanation (i.e., is
Rhythms in the brain
 NeuroScience Lecture 5 Rhythms in the brain Ana-Maria Zagrean MD, PhD Assoc. Prof., Division of Physiology & Fundamental Neuroscience Carol Davila University of Medicine and Pharmacy, Bucharest DEVELOPMENTAL
NeuroScience Lecture 5 Rhythms in the brain Ana-Maria Zagrean MD, PhD Assoc. Prof., Division of Physiology & Fundamental Neuroscience Carol Davila University of Medicine and Pharmacy, Bucharest DEVELOPMENTAL
Review Article Circadian Rhythm Disturbances in Mood Disorders: Insights into the Role of the Suprachiasmatic Nucleus
 Hindawi Neural Plasticity Volume 2017, Article ID 1504507, 28 pages https://doi.org/10.1155/2017/1504507 Review Article Circadian Rhythm Disturbances in Mood Disorders: Insights into the Role of the Suprachiasmatic
Hindawi Neural Plasticity Volume 2017, Article ID 1504507, 28 pages https://doi.org/10.1155/2017/1504507 Review Article Circadian Rhythm Disturbances in Mood Disorders: Insights into the Role of the Suprachiasmatic
Light as a central modulator of circadian rhythms, sleep and affect
 Light as a central modulator of circadian rhythms, sleep and affect Tara A. LeGates 1,2, Diego C. Fernandez 1 and Samer Hattar 1,3 Abstract Light has profoundly influenced the evolution of life on earth.
Light as a central modulator of circadian rhythms, sleep and affect Tara A. LeGates 1,2, Diego C. Fernandez 1 and Samer Hattar 1,3 Abstract Light has profoundly influenced the evolution of life on earth.
Pediatrics Grand Rounds 16 July University of Texas Health Science Center at San Antonio
 Light Pollution and Melatonin Suppression: Implications for Pediatrics Russel J. Reiter University of Texas Health Science Center San Antonio, Texas reiter@uthscsa.edu Evolution has dictated biological
Light Pollution and Melatonin Suppression: Implications for Pediatrics Russel J. Reiter University of Texas Health Science Center San Antonio, Texas reiter@uthscsa.edu Evolution has dictated biological
Light and the circadian clock : effects on human health
 Light and the circadian clock : effects on human health Claude Gronfier, Ph.D. Département de Chronobiologie Inserm U846, Institut Cellule Souche et Cerveau Lyon, France Colloque Eclairage et Santé Association
Light and the circadian clock : effects on human health Claude Gronfier, Ph.D. Département de Chronobiologie Inserm U846, Institut Cellule Souche et Cerveau Lyon, France Colloque Eclairage et Santé Association
Make sure you remember the Key Concepts
 A2 Psychology Term 1 Module 4 Physiological Psychology Biological Rhythms, Sleep and Dreaming Area of Study: Biological Rhythms. Lesson 7 Getting you Thinking pg 403 Make sure you remember the Key Concepts
A2 Psychology Term 1 Module 4 Physiological Psychology Biological Rhythms, Sleep and Dreaming Area of Study: Biological Rhythms. Lesson 7 Getting you Thinking pg 403 Make sure you remember the Key Concepts
Monday, 7 th of July 2008 ( ) University of Buea MED30. (GENERAL ENDOCRINOLOGY) Exam ( )
 University of Buea MED30. (GENERAL ENDOCRINOLOGY) Exam ( )") .. Monday, 7 th of July 2008 (8 30-11. 30 ) Faculty of Health Sciences University of Buea MED30 304 Programme in Medicine (GENERAL ENDOCRINOLOGY) Exam (2007-2008).. Multiple Choice Identify the letter
.. Monday, 7 th of July 2008 (8 30-11. 30 ) Faculty of Health Sciences University of Buea MED30 304 Programme in Medicine (GENERAL ENDOCRINOLOGY) Exam (2007-2008).. Multiple Choice Identify the letter
Targeting of the attenuated diphtheria toxin (adta) into the melanopsin locus. a,
 into the melanopsin locus. a,") doi: 1.138/nature6829 a DTA HSV- TK PGK-Neo Targeting construct b kb.85.65 L WT adta/+ adta/ adta Melanopsin (Opn 4) Genomic Locus 1 kb.4 Supplementary Figure 1: Targeting of the attenuated diphtheria
doi: 1.138/nature6829 a DTA HSV- TK PGK-Neo Targeting construct b kb.85.65 L WT adta/+ adta/ adta Melanopsin (Opn 4) Genomic Locus 1 kb.4 Supplementary Figure 1: Targeting of the attenuated diphtheria
Introduction to Physiological Psychology
 Introduction to Physiological Psychology Vision ksweeney@cogsci.ucsd.edu cogsci.ucsd.edu/~ksweeney/psy260.html This class n Sensation vs. Perception n How light is translated into what we see n Structure
Introduction to Physiological Psychology Vision ksweeney@cogsci.ucsd.edu cogsci.ucsd.edu/~ksweeney/psy260.html This class n Sensation vs. Perception n How light is translated into what we see n Structure
Endocrine Glands: Hormone-secreting organs are called endocrine glands
 University of Jordan Department of Physiology and Biochemistry Nursing students, Academic year 2017/2018. ******************************************************************* Ref: Principles of Anatomy
University of Jordan Department of Physiology and Biochemistry Nursing students, Academic year 2017/2018. ******************************************************************* Ref: Principles of Anatomy
Neuro-Physiology Kamal Mohammed Lecturer Of Physiology LECTURE NO (-) Hypothalamus. Faculty Of Medicine Dept.Of Physiology
 Hypothalamus. Faculty Of Medicine Dept.Of Physiology") LECTURE NO (-) Neuro-Physiology Kamal Mohammed Lecturer Of Physiology Hypothalamus Faculty Of Medicine Dept.Of Physiology Hypothalamus Less than 1% of the brain mass Many connect the hypothalamus to the
LECTURE NO (-) Neuro-Physiology Kamal Mohammed Lecturer Of Physiology Hypothalamus Faculty Of Medicine Dept.Of Physiology Hypothalamus Less than 1% of the brain mass Many connect the hypothalamus to the
REVIEWS. Linking neural activity and molecular oscillations in the SCN
 Linking neural activity and molecular oscillations in the SCN Christopher S. Colwell Abstract Neurons in the suprachiasmatic nucleus (SCN) function as part of a central timing circuit that drives daily
Linking neural activity and molecular oscillations in the SCN Christopher S. Colwell Abstract Neurons in the suprachiasmatic nucleus (SCN) function as part of a central timing circuit that drives daily
Physiology Unit 2 CONSCIOUSNESS, THE BRAIN AND BEHAVIOR
 Physiology Unit 2 CONSCIOUSNESS, THE BRAIN AND BEHAVIOR In Physiology Today What the Brain Does The nervous system determines states of consciousness and produces complex behaviors Any given neuron may
Physiology Unit 2 CONSCIOUSNESS, THE BRAIN AND BEHAVIOR In Physiology Today What the Brain Does The nervous system determines states of consciousness and produces complex behaviors Any given neuron may
4/23/2018. Endocrine System: Overview. Endocrine System: Overview
 Endocrine System: Overview With nervous system, coordinates and integrates activity of body cells Influences metabolic activities via hormones transported in blood Response slower but longer lasting than
Endocrine System: Overview With nervous system, coordinates and integrates activity of body cells Influences metabolic activities via hormones transported in blood Response slower but longer lasting than
Physiology Unit 2 CONSCIOUSNESS, THE BRAIN AND BEHAVIOR
 Physiology Unit 2 CONSCIOUSNESS, THE BRAIN AND BEHAVIOR What the Brain Does The nervous system determines states of consciousness and produces complex behaviors Any given neuron may have as many as 200,000
Physiology Unit 2 CONSCIOUSNESS, THE BRAIN AND BEHAVIOR What the Brain Does The nervous system determines states of consciousness and produces complex behaviors Any given neuron may have as many as 200,000
BIOLOGY - CLUTCH CH.45 - ENDOCRINE SYSTEM.
 !! www.clutchprep.com Chemical signals allow cells to communicate with each other Pheromones chemical signals released to the environment to communicate with other organisms Autocrine signaling self-signaling,
!! www.clutchprep.com Chemical signals allow cells to communicate with each other Pheromones chemical signals released to the environment to communicate with other organisms Autocrine signaling self-signaling,
Biological Rhythms, Sleep, and Dreaming. Elaine M. Hull
 Biological Rhythms, Sleep, and Dreaming Elaine M. Hull Rhythms of Waking and Sleeping Animals generate 24 hour cycles of wakefulness and sleep. Some animals generate endogenous circannual rhythms (yearly
Biological Rhythms, Sleep, and Dreaming Elaine M. Hull Rhythms of Waking and Sleeping Animals generate 24 hour cycles of wakefulness and sleep. Some animals generate endogenous circannual rhythms (yearly
The Endocrine System. I. Overview of the Endocrine System. II. Three Families of Hormones. III. Hormone Receptors. IV. Classes of Hormone Receptor
 The Endocrine System I. Overview of the Endocrine System A. Regulates long term metabolic processes B. Releases hormones from endocrine cells 1. Hormones are chemicals 2. Alter metabolism of cells 3. Release
The Endocrine System I. Overview of the Endocrine System A. Regulates long term metabolic processes B. Releases hormones from endocrine cells 1. Hormones are chemicals 2. Alter metabolism of cells 3. Release
Neuroanatomy, Text and Atlas (J. H. Martin), 3 rd Edition Chapter 7, The Visual System, pp ,
, 3 rd Edition Chapter 7, The Visual System, pp ,") Normal CNS, Special Senses, Head and Neck TOPIC: FACULTY: LECTURE: READING: RETINA and CENTRAL VISUAL PATHWAYS P. Hitchcock, Ph.D. Department Cell and Developmental Biology Kellogg Eye Center Friday, 20
Normal CNS, Special Senses, Head and Neck TOPIC: FACULTY: LECTURE: READING: RETINA and CENTRAL VISUAL PATHWAYS P. Hitchcock, Ph.D. Department Cell and Developmental Biology Kellogg Eye Center Friday, 20
The Science of Wellness: Why Your Doctor Continues to Insist You Sleep and Exercise to be Well. Nicole Rausch, DO
 The Science of Wellness: Why Your Doctor Continues to Insist You Sleep and Exercise to be Well Nicole Rausch, DO Sleep Cycle O Spend 1/3 of our time in sleep O Two types of Sleep O Non-rapid eye movement
The Science of Wellness: Why Your Doctor Continues to Insist You Sleep and Exercise to be Well Nicole Rausch, DO Sleep Cycle O Spend 1/3 of our time in sleep O Two types of Sleep O Non-rapid eye movement
Lecture 8. Arousal & Sleep. Cogs17 * UCSD
 Lecture 8 Arousal & Sleep Cogs17 * UCSD Arousal in the Brain Stimulated by sensory input Initiated, maintained endogenously Basal Forebrain Delivers ACh throughout cortex Arousal in the Brain Lateral Hypothalamus
Lecture 8 Arousal & Sleep Cogs17 * UCSD Arousal in the Brain Stimulated by sensory input Initiated, maintained endogenously Basal Forebrain Delivers ACh throughout cortex Arousal in the Brain Lateral Hypothalamus
The Functional and Clinical Significance of the 24-Hour Rhythm of Circulating Glucocorticoids
 REVIEW The Functional and Clinical Significance of the 24-Hour Rhythm of Circulating Glucocorticoids Henrik Oster,* Etienne Challet,* Volker Ott, Emanuela Arvat, E. Ronald de Kloet, Derk-Jan Dijk, Stafford
REVIEW The Functional and Clinical Significance of the 24-Hour Rhythm of Circulating Glucocorticoids Henrik Oster,* Etienne Challet,* Volker Ott, Emanuela Arvat, E. Ronald de Kloet, Derk-Jan Dijk, Stafford
NEURAL MECHANISMS OF SLEEP (p.1) (Rev. 3/21/07)
 (Rev. 3/21/07)") NEURAL MECHANISMS OF SLEEP (p.1) (Rev. 3/21/07) 1. Revisitation of Bremer s 1936 Isolated Brain Studies Transected the brain: a. Cut between the medulla and the spinal cord ( encephale isole ) Note: recall
NEURAL MECHANISMS OF SLEEP (p.1) (Rev. 3/21/07) 1. Revisitation of Bremer s 1936 Isolated Brain Studies Transected the brain: a. Cut between the medulla and the spinal cord ( encephale isole ) Note: recall
NROSCI/BIOSC 1070 and MSNBIO 2070 September 11, 2017 Control Mechanisms 2: Endocrine Control
 NROSCI/BIOSC 1070 and MSNBIO 2070 September 11, 2017 Control Mechanisms 2: Endocrine Control Hormones are chemical messengers that are secreted into the blood by endocrine cells or specialized neurons.
NROSCI/BIOSC 1070 and MSNBIO 2070 September 11, 2017 Control Mechanisms 2: Endocrine Control Hormones are chemical messengers that are secreted into the blood by endocrine cells or specialized neurons.
EEG Sleep Circadian rhythms Learning Objectives: 121, 122
 EEG Sleep Circadian rhythms Learning Objectives: 121, 122 Zoltán Lelkes Electroencenphalography Hans Berger pen time amplifier electrodes 1 The waves of the EEG gamma > 30 Hz beta: 13-30 Hz Mental activity:
EEG Sleep Circadian rhythms Learning Objectives: 121, 122 Zoltán Lelkes Electroencenphalography Hans Berger pen time amplifier electrodes 1 The waves of the EEG gamma > 30 Hz beta: 13-30 Hz Mental activity:
Dr Alex Bartle. Medical Director Sleep Well Clinic Christchurch
 Dr Alex Bartle Medical Director Sleep Well Clinic Christchurch 11:00-11:55 WS #113: Circadian Sleep Disorders 12:05-13:00 WS #125: Circadian Sleep Disorders (Repeated) Overview The Structure of Sleep
Dr Alex Bartle Medical Director Sleep Well Clinic Christchurch 11:00-11:55 WS #113: Circadian Sleep Disorders 12:05-13:00 WS #125: Circadian Sleep Disorders (Repeated) Overview The Structure of Sleep
SLEEP DISORDERS IN HUNTINGTON S DISEASE. Gary L. Dunbar, Ph.D.
 SLEEP DISORDERS IN HUNTINGTON S DISEASE Gary L. Dunbar, Ph.D. Executive Director, Field Neurosciences Institute Co-Director, Program in Neuroscience Central Michigan University Pre-Talk Test 1. Which type
SLEEP DISORDERS IN HUNTINGTON S DISEASE Gary L. Dunbar, Ph.D. Executive Director, Field Neurosciences Institute Co-Director, Program in Neuroscience Central Michigan University Pre-Talk Test 1. Which type
A CLOCKWORK WEB: CIRCADIAN TIMING IN BRAIN AND PERIPHERY, IN HEALTH AND DISEASE
 A CLOCKWORK WEB: CIRCADIAN TIMING IN BRAIN AND PERIPHERY, IN HEALTH AND DISEASE Michael H. Hastings, Akhilesh B. Reddy and Elizabeth S. Maywood The hypothalamic suprachiasmatic nuclei (SCN) are our principal
A CLOCKWORK WEB: CIRCADIAN TIMING IN BRAIN AND PERIPHERY, IN HEALTH AND DISEASE Michael H. Hastings, Akhilesh B. Reddy and Elizabeth S. Maywood The hypothalamic suprachiasmatic nuclei (SCN) are our principal
Physiology Unit 2 SENSORY PHYSIOLOGY
 Physiology Unit 2 SENSORY PHYSIOLOGY In Physiology Today Sensory System Sensory information Conscious sensations Unconscious sensations Sensory processing Transferring stimulus energy into a graded potential
Physiology Unit 2 SENSORY PHYSIOLOGY In Physiology Today Sensory System Sensory information Conscious sensations Unconscious sensations Sensory processing Transferring stimulus energy into a graded potential
Ch45: Endocrine System
 Ch45: Endocrine System Endocrine System Homeostasis is the tendency to maintain a stable internal environment. Function = coordinate and control the body with hormones to maintain homeostasis Works with
Ch45: Endocrine System Endocrine System Homeostasis is the tendency to maintain a stable internal environment. Function = coordinate and control the body with hormones to maintain homeostasis Works with
PSYCH 260 Exam 2. March 2, Answer the questions using the Scantron form. Name:
 PSYCH 260 Exam 2 March 2, 2017 Answer the questions using the Scantron form. Name: 1 1 Main Please put in their proper order the steps that lead to synaptic communication between neurons. Begin with the
PSYCH 260 Exam 2 March 2, 2017 Answer the questions using the Scantron form. Name: 1 1 Main Please put in their proper order the steps that lead to synaptic communication between neurons. Begin with the
The Nobel Assembly at Karolinska Institutet has today decided to award. the 2017 Nobel Prize in Physiology or Medicine. jointly to
 The Nobel Assembly at Karolinska Institutet has today decided to award the 2017 Nobel Prize in Physiology or Medicine jointly to Jeffrey C. Hall, Michael Rosbash and Michael W. Young for their discoveries
The Nobel Assembly at Karolinska Institutet has today decided to award the 2017 Nobel Prize in Physiology or Medicine jointly to Jeffrey C. Hall, Michael Rosbash and Michael W. Young for their discoveries
Carlson (7e) PowerPoint Lecture Outline Chapter 9: Sleep and Biological Rhythms
 PowerPoint Lecture Outline Chapter 9: Sleep and Biological Rhythms") Carlson (7e) PowerPoint Lecture Outline Chapter 9: Sleep and Biological Rhythms This multimedia product and its contents are protected under copyright law. The following are prohibited by law: any public
Carlson (7e) PowerPoint Lecture Outline Chapter 9: Sleep and Biological Rhythms This multimedia product and its contents are protected under copyright law. The following are prohibited by law: any public
Beyond our eyes: the non-visual impact of light
 Beyond our eyes: the non-visual impact of light Christian Cajochen, PhD Centre for Chronobiology Psychiatric Hospital of the University of Basel, Switzerland Workshop Intelligent Efficient Human Centric
Beyond our eyes: the non-visual impact of light Christian Cajochen, PhD Centre for Chronobiology Psychiatric Hospital of the University of Basel, Switzerland Workshop Intelligent Efficient Human Centric
Functions of hypothalamus
 The Hypothalamus Functions of hypothalamus Endocrine function Caloric balance Osmolarity balance Thermal regulation Autonomic balance Sleep Affective behavior Memory Somatic movements Anatomy of
The Hypothalamus Functions of hypothalamus Endocrine function Caloric balance Osmolarity balance Thermal regulation Autonomic balance Sleep Affective behavior Memory Somatic movements Anatomy of
Art labeling Activity: Figure 16.1
 ANP 1105D Winter 2013 Assignment 6 part I: The Endocrine Sy... Assignment 6 part I: The Endocrine System, Chapter 16 Due: 11:59pm on Monday, March 4, 2013 Note: To understand how points are awarded, read
ANP 1105D Winter 2013 Assignment 6 part I: The Endocrine Sy... Assignment 6 part I: The Endocrine System, Chapter 16 Due: 11:59pm on Monday, March 4, 2013 Note: To understand how points are awarded, read
Nonvisual effects of light. Prof. Grega Bizjak, PhD Laboratory of Lighting and Photometry Faculty of Electrical Engineering University of Ljubljana
 Nonvisual effects of light Prof. Grega Bizjak, PhD Laboratory of Lighting and Photometry Faculty of Electrical Engineering University of Ljubljana Visual and nonvisual effects of light MIND VISION HEALTH
Nonvisual effects of light Prof. Grega Bizjak, PhD Laboratory of Lighting and Photometry Faculty of Electrical Engineering University of Ljubljana Visual and nonvisual effects of light MIND VISION HEALTH
Endocrine Notes Mrs. Laux AP Biology I. Endocrine System consists of endocrine glands (ductless), cells, tissues secrete hormones
, cells, tissues secrete hormones") I. Endocrine System consists of endocrine glands (ductless), cells, tissues secrete hormones regulates metabolism, fluid balance, growth, reproduction A. Hormones 1. chemical signals-cell to cell communication
I. Endocrine System consists of endocrine glands (ductless), cells, tissues secrete hormones regulates metabolism, fluid balance, growth, reproduction A. Hormones 1. chemical signals-cell to cell communication
Light Therapy in Sleep Disorders 2014
 Light Therapy in Sleep Disorders 2014 Andrew G Veale Light and Health Effects through the eye Effects through the skin Positive effects Mood Vitality Alertess Light on skin Vitamin D production light therapy
Light Therapy in Sleep Disorders 2014 Andrew G Veale Light and Health Effects through the eye Effects through the skin Positive effects Mood Vitality Alertess Light on skin Vitamin D production light therapy
Chapter 6 Communication, Integration, and Homeostasis
 Chapter 6 Communication, Integration, and Homeostasis About This Chapter Cell-to-cell communication Signal pathways Novel signal molecules Modulation of signal pathways Homeostatic reflex pathways Cell-to-Cell
Chapter 6 Communication, Integration, and Homeostasis About This Chapter Cell-to-cell communication Signal pathways Novel signal molecules Modulation of signal pathways Homeostatic reflex pathways Cell-to-Cell
Physiology of the Mammalian Circadian System Alan M. Rosenwasser Fred W. Turek
 29 Physiology of the Mammalian Circadian System Alan M. Rosenwasser Fred W. Turek ABSTRACT Our understanding of the physiology of the mammalian circadian system has increased enormously in just the past
29 Physiology of the Mammalian Circadian System Alan M. Rosenwasser Fred W. Turek ABSTRACT Our understanding of the physiology of the mammalian circadian system has increased enormously in just the past
GENERAL CHARACTERISTICS OF THE ENDOCRINE SYSTEM FIGURE 17.1
 GENERAL CHARACTERISTICS OF THE ENDOCRINE SYSTEM FIGURE 17.1 1. The endocrine system consists of glands that secrete chemical signals, called hormones, into the blood. In addition, other organs and cells
GENERAL CHARACTERISTICS OF THE ENDOCRINE SYSTEM FIGURE 17.1 1. The endocrine system consists of glands that secrete chemical signals, called hormones, into the blood. In addition, other organs and cells
Hypothalamic Control of Posterior Pituitary
 Hypothalamic Control of Posterior Pituitary Hypothalamus neuron cell bodies produce ADH: supraoptic nuclei Oxytocin: paraventricular nuclei Transported along the hypothalamohypophyseal tract Stored in
Hypothalamic Control of Posterior Pituitary Hypothalamus neuron cell bodies produce ADH: supraoptic nuclei Oxytocin: paraventricular nuclei Transported along the hypothalamohypophyseal tract Stored in
Foundations. 1. Introduction 2. Gross Anatomy of the Eye 3. Simple Anatomy of the Retina
 Foundations 2. Gross Anatomy of the Eye 3. Simple Anatomy of the Retina Overview Central and peripheral retina compared Muller Glial Cells Foveal Structure Macula Lutea Blood supply to the retina Degenerative
Foundations 2. Gross Anatomy of the Eye 3. Simple Anatomy of the Retina Overview Central and peripheral retina compared Muller Glial Cells Foveal Structure Macula Lutea Blood supply to the retina Degenerative
Marieb, E.N. (2004), Human Anatomy & Physiology, Pearson Education, Inc, San Francisco, CA, USA, p459
, Human Anatomy & Physiology, Pearson Education, Inc, San Francisco, CA, USA, p459") Neuroscience Tutorial: Sleep and Dreams Discuss the following diagram with the students: Marieb, E.N. (2004), Human Anatomy & Physiology, Pearson Education, Inc, San Francisco, CA, USA, p459 1 Purves,
Neuroscience Tutorial: Sleep and Dreams Discuss the following diagram with the students: Marieb, E.N. (2004), Human Anatomy & Physiology, Pearson Education, Inc, San Francisco, CA, USA, p459 1 Purves,
Cephalization. Nervous Systems Chapter 49 11/10/2013. Nervous systems consist of circuits of neurons and supporting cells
 Nervous Systems Chapter 49 Cephalization Nervous systems consist of circuits of neurons and supporting cells Nervous system organization usually correlates with lifestyle Organization of the vertebrate
Nervous Systems Chapter 49 Cephalization Nervous systems consist of circuits of neurons and supporting cells Nervous system organization usually correlates with lifestyle Organization of the vertebrate
Hormones and the Endocrine System Chapter 45. Intercellular communication. Paracrine and Autocrine Signaling. Signaling by local regulators 11/26/2017
 Hormones and the Endocrine System Chapter 45 Intercellular communication Endocrine signaling Local regulators Paracrine and autocrine signaling Neuron signaling Synaptic and neuroendocrine signaling Paracrine
Hormones and the Endocrine System Chapter 45 Intercellular communication Endocrine signaling Local regulators Paracrine and autocrine signaling Neuron signaling Synaptic and neuroendocrine signaling Paracrine
Chronobiology Biological rhythm Ultradian Infradian Circadian Circatidal Circalunar Circannual Endogenous Free-running Pacemaker Target tissue
 Chronobiology Biological rhythm Ultradian Infradian Circadian Circatidal Circalunar Circannual Endogenous Free-running Pacemaker Target tissue Overt rhythm Nocturnal Diurnal LL DD LD 12:12 Study of biological
Chronobiology Biological rhythm Ultradian Infradian Circadian Circatidal Circalunar Circannual Endogenous Free-running Pacemaker Target tissue Overt rhythm Nocturnal Diurnal LL DD LD 12:12 Study of biological
Chapter 11 - Endocrine System
 Chapter 11 - Endocrine System 11.1 Introduction A. The endocrine system is made up of the cells, tissues, and organs that secrete hormones into body fluids. B. The body has two kinds of glands, exocrine
Chapter 11 - Endocrine System 11.1 Introduction A. The endocrine system is made up of the cells, tissues, and organs that secrete hormones into body fluids. B. The body has two kinds of glands, exocrine
The Visual System. Retinal Anatomy Dr. Casagrande February 2, Phone: Office: T2302 MCN
 The Visual System Retinal Anatomy Dr. Casagrande February 2, 2004 Phone: 343-4538 Email: vivien.casagrande@mcmail.vanderbilt.edu Office: T2302 MCN Reading assignments and Good Web Sites Chapter 2 in Tovée,
The Visual System Retinal Anatomy Dr. Casagrande February 2, 2004 Phone: 343-4538 Email: vivien.casagrande@mcmail.vanderbilt.edu Office: T2302 MCN Reading assignments and Good Web Sites Chapter 2 in Tovée,
Light, body clocks and sleep - SE13
 Light, body clocks and sleep - SE13 Friday 4th January 2013 10:15-11:00 Location: Palmer, 109 Russell G. Foster FRS Professor of Circadian Neuroscience Head, Nuffield Laboratory of Ophthalmology Fellow,
Light, body clocks and sleep - SE13 Friday 4th January 2013 10:15-11:00 Location: Palmer, 109 Russell G. Foster FRS Professor of Circadian Neuroscience Head, Nuffield Laboratory of Ophthalmology Fellow,
Modeling Rhythms on Differents Levels: Cells, Tissues, and Organisms
 Modeling Rhythms on Differents Levels: Cells, Tissues, and Organisms Hanspeter Herzel Institute for Theoretical Biology (ITB) Charité and Humboldt University Berlin Molecular Chronobiology SCN-neuron nucleus
Modeling Rhythms on Differents Levels: Cells, Tissues, and Organisms Hanspeter Herzel Institute for Theoretical Biology (ITB) Charité and Humboldt University Berlin Molecular Chronobiology SCN-neuron nucleus
Homeostasis. Endocrine System Nervous System
 Homeostasis Endocrine System Nervous System 2004-2005 Regulation Why are hormones needed? chemical messages from one body part to another communication needed to coordinate whole body homeostasis & regulation
Homeostasis Endocrine System Nervous System 2004-2005 Regulation Why are hormones needed? chemical messages from one body part to another communication needed to coordinate whole body homeostasis & regulation
Circadian Rhythms in Physiology and Behavior. The Persistence of Memory, Salvador Dali, 1931
 Circadian Rhythms in Physiology and Behavior The Persistence of Memory, Salvador Dali, 1931 Homeostasis and Rhythms? Homeostasis (Bernard, 1878): All the vital mechanisms, however varied they may be, have
Circadian Rhythms in Physiology and Behavior The Persistence of Memory, Salvador Dali, 1931 Homeostasis and Rhythms? Homeostasis (Bernard, 1878): All the vital mechanisms, however varied they may be, have
Overview. Surviving shift work. What is the circadian rhythm? Components of a Generic Biological Timing System 31/10/2017
 Overview Surviving shift work Dr Claire M. Ellender Respiratory and Sleep Physician Princess Alexandra Hospital Conflicts nil relevant Circadian rhythm Impacts of shift work on health Case example Circadian
Overview Surviving shift work Dr Claire M. Ellender Respiratory and Sleep Physician Princess Alexandra Hospital Conflicts nil relevant Circadian rhythm Impacts of shift work on health Case example Circadian
Endocrine secretion cells secrete substances into the extracellular fluid
 Animal Hormones Concept 30.1 Hormones Are Chemical Messengers Endocrine secretion cells secrete substances into the extracellular fluid Exocrine secretion cells secrete substances into a duct or a body
Animal Hormones Concept 30.1 Hormones Are Chemical Messengers Endocrine secretion cells secrete substances into the extracellular fluid Exocrine secretion cells secrete substances into a duct or a body
Supplementary Figure 1 Validation of Per2 deletion in neuronal cells in N Per2 -/- mice. (a) Western blot from liver extracts of mice held under ad
 Western blot from liver extracts of mice held under ad") Supplementary Figure 1 Validation of Per2 deletion in neuronal cells in N Per2 -/- mice. (a) Western blot from liver extracts of mice held under ad libitum conditions detecting PER2 protein in brain and
Supplementary Figure 1 Validation of Per2 deletion in neuronal cells in N Per2 -/- mice. (a) Western blot from liver extracts of mice held under ad libitum conditions detecting PER2 protein in brain and
Physiology of Sleep. Dr Nervana
 Physiology of Sleep Dr Nervana Objectives: 1. Explain the difference between sleep and coma. 2. Define NREM (non-rapid eye movement, SWS) and REM (rapid eye movement) sleep. 3. Describe how NREM and REM
Physiology of Sleep Dr Nervana Objectives: 1. Explain the difference between sleep and coma. 2. Define NREM (non-rapid eye movement, SWS) and REM (rapid eye movement) sleep. 3. Describe how NREM and REM
Chapter 20. Endocrine System Chemical signals coordinate body functions Chemical signals coordinate body functions. !
 26.1 Chemical signals coordinate body functions Chapter 20 Endocrine System! Hormones Chemical signals Secreted by endocrine glands Usually carried in the blood Cause specific changes in target cells Secretory
26.1 Chemical signals coordinate body functions Chapter 20 Endocrine System! Hormones Chemical signals Secreted by endocrine glands Usually carried in the blood Cause specific changes in target cells Secretory
Overview of the Biology of Sleep and Circadian Rhythms
 Overview of the Biology of Sleep and Circadian Rhythms Daniel J. Buysse, MD UPMC Professor of Sleep Medicine Professor of Psychiatry and Clinical and Translational Science University of Pittsburgh School
Overview of the Biology of Sleep and Circadian Rhythms Daniel J. Buysse, MD UPMC Professor of Sleep Medicine Professor of Psychiatry and Clinical and Translational Science University of Pittsburgh School
Developmental regulation of melanopsincontaining retinal ganglion cells and its effects on circadian function
 Oregon Health & Science University OHSU Digital Commons Scholar Archive October 2008 Developmental regulation of melanopsincontaining retinal ganglion cells and its effects on circadian function Linda
Oregon Health & Science University OHSU Digital Commons Scholar Archive October 2008 Developmental regulation of melanopsincontaining retinal ganglion cells and its effects on circadian function Linda
The Endocrine System. PowerPoint Lecture Presentations prepared by Jason LaPres. Lone Star College North Harris
 18 The Endocrine System PowerPoint Lecture Presentations prepared by Jason LaPres Lone Star College North Harris NOTE: Presentations extensively modified for use in MCB 244 & 246 at the University of Illinois
18 The Endocrine System PowerPoint Lecture Presentations prepared by Jason LaPres Lone Star College North Harris NOTE: Presentations extensively modified for use in MCB 244 & 246 at the University of Illinois
Chapter 26. Hormones and the Endocrine System. Lecture by Edward J. Zalisko
 Chapter 26 Hormones and the Endocrine System PowerPoint Lectures for Biology: Concepts & Connections, Sixth Edition Campbell, Reece, Taylor, Simon, and Dickey Copyright 2009 Pearson Education, Inc. Lecture
Chapter 26 Hormones and the Endocrine System PowerPoint Lectures for Biology: Concepts & Connections, Sixth Edition Campbell, Reece, Taylor, Simon, and Dickey Copyright 2009 Pearson Education, Inc. Lecture
Manipulating the sleep-wake cycle and circadian rhythms to improve clinical management of major depression
 See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/236073666 Manipulating the sleep-wake cycle and circadian rhythms to improve clinical management
See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/236073666 Manipulating the sleep-wake cycle and circadian rhythms to improve clinical management
Chapter 11. Endocrine System
 Chapter 11 Endocrine System 1 Introduction A. The endocrine system is made up of the cells, tissues, and organs that secrete hormones into body fluids. B. Hormones diffuse into the bloodstream to act target
Chapter 11 Endocrine System 1 Introduction A. The endocrine system is made up of the cells, tissues, and organs that secrete hormones into body fluids. B. Hormones diffuse into the bloodstream to act target
This brief animation illustrates the EEG patterns of the different stages of sleep, including NREM and REM sleep.
 Brain wave frequency and amplitude This brief animation illustrates the EEG patterns of the different stages of sleep, including NREM and REM sleep. http://www.youtube.com/watch?v=u WYwMnMMEoU&feature=related
Brain wave frequency and amplitude This brief animation illustrates the EEG patterns of the different stages of sleep, including NREM and REM sleep. http://www.youtube.com/watch?v=u WYwMnMMEoU&feature=related
THE VISUAL WORLD! Visual (Electromagnetic) Stimulus
 Stimulus") THE VISUAL WORLD! Visual (Electromagnetic) Stimulus Perceived color of light is determined by 3 characteristics (properties of electromagnetic energy): 1. Hue: the spectrum (wavelength) of light (color)
THE VISUAL WORLD! Visual (Electromagnetic) Stimulus Perceived color of light is determined by 3 characteristics (properties of electromagnetic energy): 1. Hue: the spectrum (wavelength) of light (color)
Parallel pathways in the retina
 Retinal origins of parallel pathways in the primate visual system Wednesday, September 23, 2015 Sherry, 2002 1 Parallel pathways in the retina Several different images of the outside world are sent simultaneously
Retinal origins of parallel pathways in the primate visual system Wednesday, September 23, 2015 Sherry, 2002 1 Parallel pathways in the retina Several different images of the outside world are sent simultaneously
Dark and light adaptation: a job that is accomplished mainly in the retina
 Dark and light adaptation: a job that is accomplished mainly in the retina Dark adaptation: recovery in darkness (of sensitivity) and photoreceptor pigment. Light adaptation: The ability of the visual
Dark and light adaptation: a job that is accomplished mainly in the retina Dark adaptation: recovery in darkness (of sensitivity) and photoreceptor pigment. Light adaptation: The ability of the visual
Endocrine Effects of Circadian Disruption
 I PH78CH01-Nelson ARI 10 July 2015 13:37 R E V I E W S Review in Advance first posted online on July 22, 2015. (Changes may still occur before final publication online and in print.) E N C A D V A N Endocrine
I PH78CH01-Nelson ARI 10 July 2015 13:37 R E V I E W S Review in Advance first posted online on July 22, 2015. (Changes may still occur before final publication online and in print.) E N C A D V A N Endocrine
Advance in circadian rhythm genetics in mammals
 16 2 2004 4 Chinese Bulletin of Life Sciences Vol. 16, No. 2 Apr., 2004 1004-0374 (2004) 02-0104-05 1 100101 2 434025 9 24, Q41 A Advance in circadian rhythm genetics in mammals XU Zu-Yuan 1,2 (1 Beijing
16 2 2004 4 Chinese Bulletin of Life Sciences Vol. 16, No. 2 Apr., 2004 1004-0374 (2004) 02-0104-05 1 100101 2 434025 9 24, Q41 A Advance in circadian rhythm genetics in mammals XU Zu-Yuan 1,2 (1 Beijing
CP116. Mixed-Color LED Lighting with Circadian Benefits
 Mixed-Color LED Lighting with Circadian Benefits Y. J. Tam, V. Kalavally, J. Parkkinen Department of Electrical and Computer Systems Engineering, School of Engineering, Monash University, Jalan Lagoon
Mixed-Color LED Lighting with Circadian Benefits Y. J. Tam, V. Kalavally, J. Parkkinen Department of Electrical and Computer Systems Engineering, School of Engineering, Monash University, Jalan Lagoon
NOTES: ENDOCRINE SYSTEM (CH 9)
") NOTES: ENDOCRINE SYSTEM (CH 9) Endocrine System *The endocrine system consists of a range of glands and tissues throughout the body Functions of the Endocrine System: 1) Maintain balance within body (homeostasis)
NOTES: ENDOCRINE SYSTEM (CH 9) Endocrine System *The endocrine system consists of a range of glands and tissues throughout the body Functions of the Endocrine System: 1) Maintain balance within body (homeostasis)
Rodney Heller LC, CLEP Senior Lighting Designer & Lighting Evangelist Energy Performance Lighting Cottage Grove, WI
 Rodney Heller LC, CLEP Senior Lighting Designer & Lighting Evangelist Energy Performance Lighting Cottage Grove, WI Not promoting any product or manufacturer Based on existing body of research & my experience
Rodney Heller LC, CLEP Senior Lighting Designer & Lighting Evangelist Energy Performance Lighting Cottage Grove, WI Not promoting any product or manufacturer Based on existing body of research & my experience
Mohammad Tarek. Wahab Al-tekreeti Tamer Barakat. Faisal Mohammad
 15 Mohammad Tarek Wahab Al-tekreeti Tamer Barakat Faisal Mohammad Things to remember Types of synapse: Neuron types and neurotransmitters When it happens between an axon and dendrites it is called axodendritic
15 Mohammad Tarek Wahab Al-tekreeti Tamer Barakat Faisal Mohammad Things to remember Types of synapse: Neuron types and neurotransmitters When it happens between an axon and dendrites it is called axodendritic
Psychology - Problem Drill 05: Endocrine System & Influence on Behavior
 Psychology - Problem Drill 05: Endocrine System & Influence on Behavior No. 1 of 10 1. Which of the following statements is FALSE regarding the interaction between the nervous an endocrine systems? (A)
Psychology - Problem Drill 05: Endocrine System & Influence on Behavior No. 1 of 10 1. Which of the following statements is FALSE regarding the interaction between the nervous an endocrine systems? (A)
Testosterone and other male hormones seem to be related to aggressive behavior in some species
 Testosterone and Male Aggression Testosterone and other male hormones seem to be related to aggressive behavior in some species In the fish species Oreochromis mossambicus, elevated levels have been found
Testosterone and Male Aggression Testosterone and other male hormones seem to be related to aggressive behavior in some species In the fish species Oreochromis mossambicus, elevated levels have been found
Sleep Wake Cycle in Depression
 Sleep Wake Cycle in Depression Constantin R. Soldatos Professor of Psychiatry & Founder of Sleep Study Center Eginition Hospital University of Athens Lecture in Suzdal School, 20/04/2013 SLEEP WAKE CYCLE
Sleep Wake Cycle in Depression Constantin R. Soldatos Professor of Psychiatry & Founder of Sleep Study Center Eginition Hospital University of Athens Lecture in Suzdal School, 20/04/2013 SLEEP WAKE CYCLE