Large-Scale Network Coupling with the Fusiform Cortex Facilitates Future Social Motivation
|
|
|
- Tobias Doyle
- 6 years ago
- Views:
Transcription

1 This Accepted Manuscript has not been copyedited and formatted. The final version may differ from this version. Research Article: New Research Cognition and Behavior Large-Scale Network Coupling with the Fusiform Cortex Facilitates Future Social Motivation Network Coupling Facilitates Social Motivation Amanda V. Utevsky 1,2, David V. Smith 3, Jacob S. Young 4 and Scott A. Huettel 1,2 1 Center for Cognitive Neuroscience, Duke University, Durham, NC Department of Psychology and Neuroscience, Duke University, Durham, NC Department of Psychology, Temple University, Philadelphia, PA Pritzker School of Medicine, University of Chicago, Chicago, IL DOI: /ENEURO Received: 15 March 2017 Revised: 20 September 2017 Accepted: 26 September 2017 Published: 3 October 2017 Author Contributions: AVU, DVS, JSY, and SAH designed research. AVU, DVS, JSY performed research. AVU, DVS, JYS analyzed data. AVU, DVS, SAH wrote the paper. Funding: HHS NIH National Institute of Mental Health (NIMH) F31-MH Funding: HHS NIH National Institute of Mental Health (NIMH) F32-MH Funding: HHS National Institutes of Health (NIH) NIMH RC Funding: Duke Institute for Brain Sciences, Duke University (DIBS) Conflict of Interest: Authors report no conflict of interest. This study was funded by a grant from the National Institutes of Health (NIMH RC ), an Incubator Award from the Duke Institute for Brain Sciences (SAH), and by NIMH National Research Service Awards F31- MH (AVU) and F32-MH (DVS). Correspondence should be addressed to Address for correspondence: Scott A. Huettel, Box 90999, Duke University, Durham, NC 27708, USA. scott.huettel@duke.edu phone: (919) , fax: (919) Cite as: eneuro 2017; /ENEURO Alerts: Sign up at eneuro.org/alerts to receive customized alerts when the fully formatted version of this article is published. Accepted manuscripts are peer-reviewed but have not been through the copyediting, formatting, or proofreading process. Copyright 2017 Utevsky et al. This is an open-access article distributed under the terms of the Creative Commons Attribution 4.0 International license, which permits unrestricted use, distribution and reproduction in any medium provided that the original work is properly attributed.
2 Large-Scale Network Coupling with the Fusiform Cortex Facilitates Future Social Motivation Abbreviated Title: Network Coupling Facilitates Social Motivation Amanda V. Utevsky 1,2, David V. Smith 3, Jacob S. Young 4, Scott A. Huettel 1,2 * 1 Center for Cognitive Neuroscience, Duke University, Durham, NC, Department of Psychology and Neuroscience, Duke University, Durham, NC, Department of Psychology, Temple University, Philadelphia, PA, Pritzker School of Medicine, University of Chicago, Chicago, IL, AVU, DVS, JSY, and SAH designed research. AVU, DVS, JSY performed research. AVU, DVS, JYS analyzed data. AVU, DVS, SAH wrote the paper. *Address for correspondence: Scott A. Huettel Box 90999, Duke University Durham, NC 27708, USA scott.huettel@duke.edu phone: (919) fax: (919) Number of Figures: 4 Number of Tables: 2 Number of words for Abstract: 233 Number of words for Significance Statement: 116 Number of words for Introduction: 637 Number of words for Discussion: 1759 Acknowledgments: This study was funded by a grant from the National Institutes of Health (NIMH RC ), an Incubator Award from the Duke Institute for Brain Sciences (SAH), and by NIMH National Research Service Awards F31-MH (AVU) and F32-MH (DVS). The authors thank Christopher Coutlee and Cary Politzer for their assistance in task design and data collection. Conflict of Interest: Authors report no conflict of interest. Funding sources: National Institutes of Health (NIMH RC ), the Duke Institute for Brain Sciences (SAH), NIMH National Research Service Awards F31-MH105137, F32- MH (DVS). Page 1 of 38
3 Abstract Large-scale functional networks as identified through the coordinated activity of spatially distributed brain regions have become central objects of study in neuroscience because of their contributions to many processing domains. Yet, it remains unclear how these domaingeneral networks interact with focal brain regions to coordinate thought and action. Here, we investigated how the default-mode network (DMN) and executive control network (ECN) two networks associated with goal-directed behavior shape task performance through their coupling with other cortical regions several seconds in advance of behavior. We measured these networks connectivity during an adaptation of the monetary incentive delay response-time task in which human participants viewed social and non-social images (i.e., pictures of faces and landscapes, respectively) while brain activity was measured using fmri. We found that participants displayed slower response times (RTs) subsequent to social trials relative to nonsocial trials. To examine the neural mechanisms driving this subsequent-rt effect, we integrated independent components analysis (ICA) and a networkbased psychophysiological interaction (nppi) analysis; this allowed us to investigate taskrelated changes in network coupling that preceded the observed trial-to-trial variation in RT. Strikingly, when subjects viewed social rewards, an area of the fusiform gyrus consistent with the functionally-defined fusiform face area (FFA) exhibited increased coupling with the ECN (relative to the DMN) and the relative magnitude of coupling tracked the slowing of RT on the following trial. These results demonstrate how large-scale, domain-general networks can interact with focal, domain-specific cortical regions to orchestrate subsequent behavior. Page 2 of 38
4 Keywords: effective connectivity, executive control, default mode network, faces, goal- directed behavior Page 3 of 38
5 Significance A diverse set of behaviors from normal to pathological has been linked to the responses of large-scale functional networks. Yet, it remains unclear how these domain-general networks shape subsequent processing in cortical regions with domain-specific function. Here, we examine how two such networks the default-mode network (DMN) and executive control network (ECN) connect functionally with other cortical regions to alter performance in an incentive-compatible task. We found that differential coupling between a prototypical face processing region and DMN and ECN tracked subsequent improvements in performance to social stimuli. Our approach allowed us to examine direct coupling with functional networks to future behavior, providing a significant step forward in understanding how large-scale networks coordinate thought and action Page 4 of 38
6 Introduction Since the discovery of functionally correlated brain regions (Biswal et al., 1995), large-scale functional networks have been considered fundamental features of brain activity (Beckmann et al., 2005; Behrens and Sporns, 2012; Gusnard and Raichle, 2001). These networks are highly reliable across large samples of participants (Biswal et al., 2010; Smith et al., 2013) and are thought to reflect intrinsic properties of brain organization. Many of these networks reflect sensory and perceptual processes instantiated within visual or auditory regions (Smith et al., 2009). Still others contribute broadly to cognitive processing across stimulus and task domains, including the default-mode (DMN), executive control (ECN), and frontoparietal (FPN) networks (Dosenbach et al., 2007; Smith et al., 2009). These functional networks have been linked to various aspects of behavior including demographic variables (Cowdrey et al., 2012; Filippini et al., 2009), traits (Kucyi et al., 2014; Shannon et al., 2011), and cognitive states (De Havas et al., 2012; Smith et al., 2009). Yet, for effective behavior in a particular task, these large-scale domain-general networks must alter ongoing task-specific processing in focal brain regions. To claim that such interactions (i.e., between large-scale networks and specific brain regions) are critical for behavior change, several conditions should be met. First, a large-scale network should be identifiable during task performance independently of other concurrent activation; that is, networks should be able to be extracted regardless of other processes occurring in focal brain regions that might contribute to that task (i.e., without relying on seed-based analyses; Cole et al., 2010; Smith et al., 2014b). Second, the coupling between a given network and a given focal brain region should systematically vary across task conditions according to the relative engagement of the task (Friston et al., 1997; O Reilly et al., 2012). Third, those Page 5 of 38
7 changes in coupling (e.g., effective connectivity (Friston, 2011)) should predict the characteristics of subsequent behavior, to provide evidence that the coupling contributes to effective task performance. If these conditions are met, there would be strong evidence that coupling between a functional network and a focal brain region contributes to a specific cognitive process. In the current study, we examined effective connectivity between large-scale networks and focal brain regions while subjects played a response-time game to receive social and nonsocial rewards in a modified monetary incentive delay (MID) task (Knutson et al., 2000). We have previously used this task to produce meaningful variability in reaction time behavior that reflects relative motivation (Clithero et al., 2011). We focused on the DMN and ECN since both have been linked to task performance, engagement, and other markers of executive function and preparatory behavior. Behaviorally, we found that participants exhibited slower response times (RTs) subsequent to social relative to nonsocial trials, reflecting a change in motivation according to social stimulus type. We investigated whether coupling with DMN and ECN contributed to this subsequent-rt effect by combining independent components (ICA) and psychophysiological interactions (PPI) analyses. This network PPI (nppi) pipeline allowed us to examine how DMN and ECN contribute to other cortical regions to shape subsequent motivated behavior up to several seconds later. Strikingly, we found that DMN and ECN differentially coupled with a region in the fusiform gyrus (FG) when subjects viewed the social rewards, and that changes in this coupling tracked the effect of stimulus type on subsequent RT. This region of the FG is consistent with the functionally-defined fusiform face area (FFA) a region classically associated with face processing (Kanwisher et al., 1997; McCarthy et al., 1997). Page 6 of 38
8 These findings highlight functional network interactions that guide subsequent changes in reaction time behavior. While the cognitive mechanisms underlying the behavioral effect of previous stimulus type on subsequent reaction time require future research, we propose that this effect may be driven by increased attentional interference of social rewards relative to nonsocial rewards or a subsequent change in task motivation. Collectively, these results indicate that functional networks associated with goal-directed behavior can interact with focal brain regions to support future motivated behavior. 151 Materials and Methods Participants A group of 50 self-reported heterosexual males completed the task (mean age: 23.8 years, range 18-32); this sample size was established prior to data collection. All participants were screened prior to data collection to rule out prior or current psychiatric or neurological illness. We excluded four participants because of data quality issues (see Preprocessing), and we excluded five participants because of a malfunctioning button box in the scanner. These exclusions left a final sample of 41 participants (mean age: 24.1 years, range: 18-32). All participants gave written informed consent as part of a protocol approved by the Institutional Review Board of Duke University Medical Center Stimuli and Tasks The experiment consisted of four components: 1) a training session outside the scanner; 2) a modified monetary incentive delay task (MID; Knutson et al., 2000, 2001); 3) a passive viewing task with results previously published (Young et al., 2015); 4) two Page 7 of 38
9 additional post-scan rating tasks. All participants received variable cash payment depending on their performance. In advance of the neuroimaging session, participants memorized the associations of four fractal images with different potential rewards (see an example in Figure 1). On the day of the scan, subjects were tested on the fractal images associations to ensure they remembered the meaning of each image; if participants identified every association correctly on the first attempt (n = 31), they received an extra $5 in addition to their study compensation. All participants successfully learned the image associations before entering the scanner. Each trial of the modified MID task began with a fractal image presented as a cue for 1500 ms (Figure 1). The different cues indicated the different potential benefits of a fast response on that trial: [1] reducing the delay until the presentation of a face image of a variable attractiveness rating (Social Delay; SD), [2] increasing the attractiveness of a face image presented after a variable duration (Social Attractiveness; SA), [3] reducing the delay until the presentation of a landscape image of a variable attractiveness rating (Nonsocial Delay; ND), [4] increasing the attractiveness of a landscape image presented after a variable duration (Nonsocial Attractiveness; NA). Following the cue, a fixation cross appeared for a variable duration (interstimulus interval; ms), followed by a target (a white triangle) that appeared on the screen for 750 ms. Participants then responded by pressing a button box with their right index finger before the target disappeared. A feedback shape then appeared for 750 ms; that shape was colored green if the participant s response was sufficiently fast (a win), and was colored blue if the response was too slow (a loss). After the feedback image disappeared, Page 8 of 38
10 participants waited a delay period that varied according to the cue and whether the subject won or lost that trial. Lastly, the reward image (either a face or a landscape) was shown for 2000 ms, and there was an intertrial interval (ITI) that was jittered between 1500 and 4500 ms (with the 2000 ms reward image duration, the minimum and maximum time delays between reward onset and subsequent cue onset was 3500 and 6500 ms, respectively). Each run of the modified MID task consisted of 36 trials, nine images of each SD, SA, ND, and NA in randomized order. Across all runs included in the analyses, the four types of transitions (e.g., social followed by social, social followed by nonsocial, etc.) each occurred on approximately 25±1% of trials, reflecting effective randomization of stimulus types across trials. Social images were photographs of young adult women and were cropped to show only the face (images were drawn from Smith et al. 2010). Nonsocial images were photographs of landscapes, and did not contain any body or facial features. All images were resized to uniform dimensions. Attractiveness (high/medium/low) of images was determined by ratings by an independent group. Additionally, performance thresholds were determined independently for each subject by an algorithm that adapted the task difficulty so that accuracy would be approximately 60% (observed hit rate = 60.7%). After completing the four runs of the modified MID task, participants completed an unrelated passive-viewing task described elsewhere (Young et al., 2015). Following this second scanner task, participants completed two tasks outside the scanner. In the first task, participants rated the attractiveness of each image they viewed in the modified MID task on a scale of 1 to 7. Each participant s highest-rated face and landscape images were then used in a second task involving forced choices between a landscape image and a face image presented simultaneously. Page 9 of 38
11 All tasks were programmed and displayed using the Psychophysics Toolbox (Version 3; Brainard, 1997) for MATLAB (Mathworks Inc.) Behavioral Analysis Behavioral data were analyzed using MATLAB and IBM SPSS Statistics 20. A response time (RT) difference was calculated for each participant by subtracting their mean RT on the trials subsequent to nonsocial trials from their mean RT on trials subsequent to social trials. Consequently, positive RT differences indicate shorter RTs subsequent to nonsocial trials (relative to social trials), whereas negative differences indicate shorter RTs subsequent to social trials (relative to nonsocial trials). To control for factors other than prior stimulus category such as attractiveness of the prior trial s reward image, the prior trial s RT, and the subsequent trial s stimulus type we incorporated these factors into a general linear model (GLM) for each subject. The GLM included the following as regressors: (1) stimulus type on previous (t-1) trial, (2) RT on previous trial, (3) attractiveness rating of the reward image of the previous trial, and (4) stimulus type on current trial t. This analysis indicates the specific influence of each regressor on the current trial s RT, and allowed us to examine the unique influence of the previous trial s stimulus category (social or nonsocial) on current RT. In our analyses, positive stimulus type beta weights reflect slower RTs on or following social trials, whereas negative stimulus type betas weights reflect faster RTs on or following social trials Image Acquisition Neuroimaging data were collected using a General Electric MR Tesla scanner equipped with an 8-channel parallel imaging system. We used a T 2*-weighted spiral-in Page 10 of 38
12 sensitivity encoding sequence (SENSE factor = 2), with slices parallel to the axial plane connecting the anterior and posterior commissures (repetition time (TR): 1580 ms; echo time (TE): 30 ms; matrix: 64 x 64; field of view (FOV): 243 mm; voxel size: 3.8 x 3.8 x 3.8 mm; 37 axial slices; flip angle: 70 degrees). The first eight volumes of each run were removed to allow for magnetic stabilization. We additionally acquired whole-brain highresolution anatomical scans (T1-weighted FSPGR sequence; TR: 7.58 ms; TE: 2.93 ms; matrix: 256 x 256; FOV: 256 mm; voxel size: 1 x 1 x 1 mm; 206 axial slices; flip angle: 12 degrees) to allow for coregistration and normalization Preprocessing Our preprocessing used tools from the FMRIB Software Library package (FSL Version 4.1.8; Smith et al., 2004; Woolrich et al., 2009). We corrected for head motion by realigning the time series to the middle volume (Jenkinson and Smith, 2001), and then removed non-brain material using a brain extraction tool (Smith, 2002). We then corrected intravolume slice-timing differences using Fourier-space phase shifting to align to the middle slice (Sladky et al., 2011). After spatially smoothing the image using a 5-mm full-width-half-maximum isotropic Gaussian kernel, we applied a high-pass temporal filter with a 100-second cutoff, and we normalized each 4-dimensional dataset to the grand-mean intensity using a single multiplicative factor. Lastly, we spatially normalized the functional data to the Montreal Neurological Institute (MNI) Template avg152 T1- weighted template (3 mm isotropic resolution) using a 12-parameter affine transformation implemented in FLIRT (Jenkinson and Smith, 2001). As part of our preprocessing and quality control, we additionally examined three partially correlated measures of quality assurance: signal-to-fluctuation-noise ratio (SFNR; Page 11 of 38
13 Friedman and Glover, 2006), volume-to-volume head motion, and number of motion spikes within the time series (motion spikes were identified by evaluating the root-mean-squareerror of each volume relative to the middle time point). Measures on each metric were considered outliers if they exceeded the 75 th percentile plus the value of 150% of the interquartile range (i.e., a standard boxplot threshold); runs that were identified as outliers were excluded from further analyses. Additionally, any participant who had fewer than two good runs (out of four total runs) was excluded from further analyses. These criteria eliminated four participants Neuroimaging Analysis To best meet the conditions for identifying task-specific interactions between largescale networks and focal brain regions (see Introduction), our neuroimaging analyses proceeded in two phases, each described in a separate section below. First, we used independent components analysis (ICA; Beckmann & Smith, 2004) and spatial regression (Filippini et al., 2009) to identify the large-scale neural networks of interests (DMN and ECN) and to examine the networks levels of activation over the course of each run. Second, we used generalized network psychophysiological interaction (nppi) models (adapted from McLaren et al., 2012) to identify brain regions whose coupling with the ECN and DMN changed as a function of the effect of stimulus type on subsequent RT. Importantly, this ICAbased nppi approach follows the logic of ROI-based PPI analyses, with the critical difference of examining connectivity with data-driven large-scale neural networks instead of a specific seed region. Critically, we note that this nppi pipeline allowed us to test the three necessary conditions for inferences that functional networks alter task-specific processing in focal brain regions. Page 12 of 38
14 Identifying Large-Scale Functional Networks We used FSL s Multivariate Exploratory Linear Decomposition into Independent Components (MELODIC) Version 3.10 to identify large-scale functional networks in the neuroimaging data (Beckmann and Smith, 2004). MELODIC ICA was implemented using temporal concatenation, which looks for common spatial patterns of components across participants data without assuming a specific or common time course across all participants; we note that previous research has also used temporal concatenation-based ICA on task-based fmri data (Utevsky et al., 2014; Young et al., 2015). The preprocessed data were whitened and projected into a 25-dimensional subspace (Ray et al., 2013; Utevsky et al., 2014). The whitened data were decomposed into sets of vectors describing the temporal and spatial signal variation, using a fixed-point iteration technique to optimize non-gaussian spatial source distribution (Hyvärinen, 1999). The estimated component maps were then thresholded by dividing the maps by the standard deviation of the residual noise, then fitting a Gaussian-Gamma mixture model to the histogram of the normalized intensity values (Beckmann and Smith, 2004). This first step provided a data-driven means to identify functional networks present during task performance; this allowed us to meet the first condition for identifying task-specific interactions between large-scale networks and specific brain regions. All unthresholded spatial maps from the ICA were then submitted to a spatial regression (part of FSL s dual regression analysis) to estimate the time courses of each network (Filippini et al., 2009; Leech et al., 2011). In this analysis, spatial maps are regressed onto each participant s functional data, resulting in a matrix of T (time points) x C Page 13 of 38
15 (components) beta coefficients that characterize each subject s time courses for each network Characterizing Reward-Related Network Connectivity and Activation We assessed task-dependent network coupling using a generalized nppi model. The generalized PPI, in contrast to a standard PPI, computes a separate PPI term for each task condition. This approach has been shown to yield more accurate estimates of how connectivity varies as a function of psychological context (McLaren et al., 2012). The generalized nppi analysis was carried out using FMRI Expert Analysis Tool (FEAT) Version The run-level model included six task regressors: social cue (duration = s), nonsocial cue (duration = s), hits (duration = 0.75 s), misses (duration = 0.75 s), social reward outcome (duration = 2 s), and nonsocial reward outcome (duration = 2 s). We additionally included time courses of both the DMN and ECN that were produced by the spatial regression. Because of the minimum ITI of 1500 ms and the potential confound of the cue presentation occurring between the outcome and subsequent target phases of the task, we examined whether there was any collinearity between the outcomes and cues across the runs included in our analysis. For each run included in our analyses, we calculated the correlation values between the face and land outcome regressors and the face and land cue regressors and then averaged these correlation values across all runs. The mean correlation values between face or land cues and face or land outcomes ranged from r = to r = (minimum r = -0.28; maximum r = -0.10), indicating that our run-level analyses were able to isolate and model cues and outcomes independently, with minimal collinearity between regressors. Page 14 of 38
16 For our network interaction analysis, nppi regressors were formed by multiplying the DMN and ECN time courses (zeroed to the mean of the time course), separately, by the social outcome and nonsocial outcome regressors (zeroed to the minimum value of the task time course) (McLaren et al., 2012); this yielded four nppi regressors: (1) DMN*social, (2) DMN*nonsocial, (3) ECN*social, (4) ECN*nonsocial. To control for motion in the scanner, we additionally included motion spikes and motion parameters as regressors. Lastly, to control for the influence of other networks and potential artifacts on our generalized nppi, we included the time courses of the remaining 23 components from the ICA. The nppi analysis allowed us to examine task-specific coupling between ECN and DMN with other regions in the brain, fulfilling the second condition for identifying the task-specific interactions of interest. Subject-level analyses for the generalized nppi were run using FEAT and implementing FMRIB s Local Analysis of Mixed Effects (FLAME 1), and examined activation across runs within each participant. Group-level analyses included each subject s demeaned betas from the prior stimulus-category regressor (see Behavioral Analysis); this allowed us to examine whether network coupling predicts the characteristics of subsequent RT, and fulfill the third condition for identifying task-specific interactions of interest. The group-level analysis additionally included the main effect of group and three motion-related parameters (SFNR, volume-to-volume head motion, and number of motion spikes within the time series). All resulting z-statistic images were thresholded using a cluster-forming threshold of 2.3 and a corrected cluster-significance threshold of p < Although this threshold combined with FSL s FLAME 1 protects against false positives, we note that all of our results also survived permutation-based testing (Eklund et al., 2016). In these supplemental tests, Page 15 of 38
17 statistical significance was assessed in a nonparametric fashion via FSL s randomise; this tool uses Monte Carlo permutation-based testing with 10,000 permutations and = 0.05, corrected for multiple comparisons across the whole brain (Nichols and Holmes, 2002; Winkler et al., 2014). Brain images and activations are displayed using MRIcroGL ( Rorden et al., 2007). All coordinates are reported in Montreal Neurological Institute (MNI) space. 354 Results Previous stimulus category influences current behavior Social rewards, such as images of individuals or interactions with others, provide a useful tool for examining the effects of both social context and non-consumable rewards on motivated behavior. However, relatively little is known about the effects of social and nonsocial rewards on future motivated behavior, and how the brain orchestrates future motivated action. To examine the effect of social rewards on subsequent motivated actions, we calculated a subsequent-rt effect by subtracting RTs following nonsocial trials from RTs following social trials. When averaging across value and delay trials, we found an overall effect of previous trial stimulus type on current trial RT: participants exhibited increased (slower) RTs subsequent to social trials (M = s, SD = s) compared to subsequent to nonsocial trials (M = s, SD = s; t (40) = 2.63, p = 0.01, d = 0.41; Figure 2A). This pattern replicated when examining hit trials only (social: M = s, SD = s; Page 16 of 38
18 nonsocial: M = s, SD = s; t (40) = 2.63, p = 0.01, d = 0.41). Thus, participants were slower after performing a social trial compared to a nonsocial trial. We next ran a general linear model (GLM) to control for properties of the previous and current trials (see Behavioral Analysis). Across participants, current stimulus category was the strongest predictor of current RT (t (40) = -4.31, p <.0001, d = 0.67), reflecting that participants were faster to respond during a social trial than during a nonsocial trial. The next strongest predictor was the previous trial s stimulus category (t (40) = 2.46, p = 0.01, d = 0.38), reflecting that participants were slower to respond following a social trial than following a nonsocial trial. Neither previous trial s RT nor previous trial s reward attractiveness rating significantly predicted the current trial s RT (Figure 2B; Table 1). We note that the effect of previous stimulus type on current RT was still significant even after accounting for the large effect of current stimulus type, reflecting a distinct role of previous stimulus type unaccounted for by the other variables included in the GLM. These results indicate that of the measured properties of the previous trial, prior stimulus type had the strongest effect on current RT Network coupling with fusiform gyrus tracks effect of stimulus category on subsequent behavior Our behavioral results indicated that current-trial stimulus type influences motivated behavior on the subsequent trial (which occurred seven seconds or more later), such that participants were slower to respond following social trials compared to following nonsocial trials. However, it remains unclear how interactions in the brain during this previous stimulus outcome affect the subsequent RT. To investigate this, we examined how effective Page 17 of 38
19 connectivity with the ECN a network implicated in cognitive control and goal-directed behavior and the DMN a network linked with task engagement orchestrate this change in future motivated behavior. We predicted that this subsequent-rt effect would be guided, in part, by changes in the coupling between these large-scale functional networks and domainspecific brain regions when viewing social images compared to when viewing nonsocial images. In particular, we predicted that the subsequent-rt effect would be driven by changes in the relative coupling of the ECN and the DMN during the reward outcome phase of the previous trial. Our analysis pipeline allowed us to meet the criteria to claim that interactions between large-scale networks and specific brain regions are critical for behavior changes (see Introduction). After running the ICA, we identified the DMN and ECN maps by running spatial correlations between the unthresholded maps from our ICA and the DMN and ECN maps from Smith and colleagues (2009). From our 25 components, we selected the maps that best matched the DMN (R = 0.776; other components: R mean = , R min = , R max = 0.124) and ECN (R = 0.64; other components: R mean = 0.019, R min = , R max = 0.296) maps from Smith and colleagues (2009) (see Table 2). For ease of visualization, thresholded maps (Z > 4) are shown in Figure 3. Following network identification, we ran a nppi analysis that used participantspecific DMN and ECN time courses (estimated by the spatial regression analysis) as the physiological regressors and presentation of social and nonsocial images as the psychological regressors. This nppi identified regions that are influenced by the ECN and DMN in a task-dependent manner. We then tested whether these influences on cortex predicted the effect of prior stimulus type on current RT. Our nppi analysis indicated that Page 18 of 38
20 effective connectivity between the fusiform gyrus (FG; peak: x = 38, y = -64, z = -20, p < , voxel extent = 383) and the ECN increased (compared to FG-DMN connectivity) when participants viewed social rewards (Figure 4A) and that the magnitude of this increase tracked the slowing of RT on the subsequent trial (Figure 4B). Strikingly, this peak voxel is consistent with the often functionally-defined FFA (Kanwisher and Yovel, 2006; McCarthy et al., 1997). Identification of the terms associated with this peak voxel using the meta-analytical tool Neurosynth ( (Yarkoni et al., 2011) yielded faces, FFA, and fusiform face within its top four associations. These results suggest that functional networks associated with goal-directed and preparatory behavior can interact with focal brain regions to support task-relevant behavior. To ensure that this connectivity was associated with the previous trial s outcome rather than the current trial s cue, we ran an additional nppi to examine whole-brain connectivity during social cues, nonsocial cues, and nonsocial > social cues. Notably, there were no regions exhibiting changes in connectivity with the ECN relative to the DMN in any of the three contrasts tested. To further investigate this, we used the FG region showing changes in ECN-DMN connectivity during the previous outcome phase as a mask during the current cue phase, and examined whether the ECN-DMN connectivity from this region of interest (ROI) tracked the effect of prior stimulus on current RT. Across the three contrasts tested (social, nonsocial, nonsocial > social), there was no correlation between the ROI connectivity estimates and the effect of previous stimulus type on current RT (social: r = , p = 0.45; nonsocial: r = 0.01, p = 0.94; nonsocial > social: r = 0.23, p = 0.15). Collectively, these results support the claim that changes in subsequent RT are associated with Page 19 of 38
21 differential network connectivity with the FG during the previous outcome phase, and cannot be attributed to other aspects of task performance. No regions showed increased coupling with ECN (relative to DMN) during nonsocial rewards that tracked the effect on subsequent RT, nor did any regions show increased coupling with DMN (relative to ECN) during social or nonsocial rewards that tracked the effect on subsequent RT. Additionally, there were no regions that exhibited changes in coupling with the DMN or ECN when comparing social and nonsocial rewards (e.g., DMNsocial > DMN-nonsocial; ECN-social > ECN-nonsocial; and the inverse contrasts) that tracked the effect on subsequent RT. Lastly, we ran a whole-brain GLM using the RT betas as a covariate to examine whether any regions activation tracked the effect of stimulus type on subsequent RT; we found that no regions tracked this effect using this traditional GLM. Thus, coupling between large-scale networks and the FG that tracked the effect on subsequent RT was only observed during the viewing of social rewards Discussion Recent neuroscience research has highlighted the relevance of large-scale functional networks to various aspects of behavior (Anticevic et al., 2010; Eichele et al., 2008; Kelly et al., 2008; Rosen et al., 2015). While many of these studies have linked network activation and connectivity to behavior, the contribution of these networks to motivated behaviors via focal cortical regions has been relatively understudied. For example, although previous work has found correlations between DMN and working memory (Piccoli et al., 2015; Sambataro et al., 2010) or sustained attention (Bonnelle et al., 2011; Gui et al., 2015), understanding how these distributed functional networks influence other cortical regions to influence Page 20 of 38
22 behavior has remained a significant challenge. Here, we found that participants were slower in a response time task after having performed a trial for a social reward relative to a nonsocial reward (after accounting for the influence of the current trial cue type), reflecting a change in motivated behavior according to previous social stimulus type (Clithero et al., 2011). We then examined the neural mechanisms underlying this effect using nppi analysis an adaptation of generalized PPI analysis (i.e., effective connectivity) that examines effective connectivity with large-scale functional networks. This analysis pipeline allowed us to identify changes in coupling between large-scale networks and focal brain regions that predicted subsequent motivated behavior. Our results demonstrate that two goal-relevant networks, the DMN and ECN, interact with FG in a manner that predicts trial-to-trial adjustments in response time. Our analysis on our modified MID task identified that participants exhibit slower reaction times on trials subsequent to social outcomes, relative to trials subsequent to nonsocial outcomes. These results are consistent with a difference in motivational processes due to the social nature of the previous reward outcome, analogous to effects on motivation observed in similar tasks with nonsocial stimuli (Clithero et al., 2011). There are at least two potential explanations for this subsequent reaction time effect. The first hypothesis is that social images are more distracting than nonsocial images, and participants may still covertly attend to a prior social reward more so than they do a prior nonsocial reward. Previous research indicates that social images interfere with visual attention to a greater degree than nonsocial images (Ebitz et al., 2013); thus, it is possible that this differential interference may affect subsequent trials differently, as well. A second and related potential explanation is that participants decreased motivation can be attributed to the satisfaction of receiving a Page 21 of 38
23 motivating reward on the previous trial, much akin to results seen in satisfaction of search (SOS) research. In SOS research, participants performing visual search tasks tend to discontinue their search after finding an initial item, and either miss or display slower reaction times for subsequent items (Berbaum et al., 1990; Fleck et al., 2010). As applied to our MID task, a more motivating reward outcome (social images) may hinder the performance on the subsequent trial. Future research will be needed to explore the cognitive mechanisms driving the observed subsequent-rt effect and to determine whether it is specific to social versus nonsocial stimuli, or may generalize to items that are more motivating versus less motivating. Our findings expand on recent research examining the relevance of DMN and ECN activation to behavior, potentially in task-relevant contexts. Specifically, we found that increased ECN (relative to DMN) coupling with the FG is associated with enhanced subsequent task performance. These results are consistent with previous studies demonstrating different relationships between the DMN and ECN with task behavior: while ECN activation is often associated with heightened task performance and behavior (Dosenbach et al., 2007; Seeley et al., 2007), DMN activation is frequently linked to decrements in behavior and engagement in both humans (Eichele et al., 2008; Weissman et al., 2006) and non-human primates (Hayden et al., 2009, 2010; Heilbronner and Platt, 2013). Importantly, however, our findings extend these previous results by demonstrating direct coupling of these networks with a prototypical social-perception processing region in a taskdependent manner. Our results additionally highlight that the opposing effects of DMN and ECN are not limited to concurrent behavior, but also affect subsequent behavior with effects observed seven or more seconds later. These findings support the idea that large-scale Page 22 of 38
24 networks interact with lower-level perceptual regions to contribute to motivational processes (Clithero et al., 2011) and shape later behavior. Unlike previous studies examining large-scale networks (Brewer et al., 2011; Ossandon et al., 2011; Seeley et al., 2007; Utevsky et al., 2014), our experiment demonstrates network coupling that directly shapes subsequent response time. While prior studies report associations between functional connectivity and behavior, estimates of functional connectivity solely report on correlations in activation (not coupling) between regions, which can be the result of various phenomena (Friston, 2011). Specifically, reported changes in functional connectivity can arise from numerous causes, including: changes in connectivity with another region, changes in observation noise (or signal-to-noise ratios), and changes in the degree of neuronal fluctuations (for a review, see Friston, 2011). Thus, changes in functional connectivity may not reflect changes in coupling between cortical regions. In contrast to functional connectivity, we implemented an adaptation of a traditional PPI analysis to measure effective connectivity between functional networks and other cortical regions. Psychophysiological interaction analyses measure whether a psychological context (e.g., outcome stimulus category) influences how one brain region or network (the seed ) contributes to another (the target ) by examining whether an interaction between the psychological context and the seed is identified in the target (Smith et al., 2016). Thus, a PPI analysis can eschew the potential confounds of functional connectivity analyses and reflects a change in neural coupling (e.g., effective connectivity). We note that our novel approach of generalized nppi applying generalized PPI analyses to large-scale networks allows us to examine task-dependent contributions between networks and other cortical Page 23 of 38
25 regions (Friston et al., 1997; Friston, 2011). In this way, our study extends prior work by demonstrating specific task-dependent coupling of the DMN/ECN. Although recent meta-analytic work has demonstrated that PPI produces consistent and specific patterns of connectivity (Smith et al., 2016; Smith and Delgado, 2017), it is important to note that PPI results can be interpreted in two ways (Friston et al., 1997). First, our effects could reflect a context-specific modulation of effective connectivity. In this case, face presentations modulate the degree to which the DMN and ECN contribute to FG. Our results focus on the difference between DMN and ECN contributions to FG which seem to facilitate social motivation (Clithero et al., 2011). Alternatively, our effects could reflect a modulation of stimulus-specific responses. In this case, the DMN and ECN influence how FG responds to the presentation of the face; under this interpretation, our results suggest that the degree to which DMN enhances face responses in FG is greater than that of ECN. In either interpretation, the resulting effect on FG connectivity predicts behavior. A better understanding of the mechanisms and causal relationship underlying our results may be facilitated by other analytical approaches, such as dynamic causal modeling (DCM) (Friston et al., 2003; Friston, 2011), although relatively less is understood regarding how the biophysical models implemented in DCM apply to distributed functional networks (Buxton et al., 1998; Friston et al., 2003). Additionally, other methodological approaches, such as transcranial magnetic stimulation (Fox et al., 2012; Luber and Lisanby, 2014; Mueller et al., 2014) or transcranial current stimulation (Hampstead et al., 2014; Keeser et al., 2011) may also better inform the specific interactions between FG and the ECN and DMN. One potential caveat to note is that our task design included a cue to indicate the current trial s stimulus type in between the previous trial s outcome phase and the current trial s Page 24 of 38
26 response phase. Because the current trial s stimulus type is the strongest predictor of the current trial s RT, there may be concerns over whether the neural coupling we observe can be attributed to the cue, rather than the previous trial s outcome. This concern is ameliorated through both the randomization of trial types (i.e., the identity of the outcome on the previous trial is uncorrelated with the trial type on the subsequent trial), as well as the run-level models implemented in our PPI analysis that included separate regressors for both the outcome phase (social and nonsocial outcomes, separately) and the cue phase (social and nonsocial cues, separately). Including these separate regressors for each phase allowed us to distinguish effects of the cue and outcome phases on connectivity. This concern is additionally mitigated by the analyses examining connectivity during the cue phase. A nppi showed no changes in ECN-DMN connectivity during social cues, nonsocial cues, or social > nonsocial cues when looking across the whole brain. A supplementary ROI analysis using the FG region indicated that FG connectivity with the ECN (relative to the DMN) during these three contrasts did not track the effect of the previous trial stimulus type on subsequent RT (as illustrated by the lack of significant correlations between the connectivity estimates and the effect of previous stimulus type on subsequent RT). However, future research may benefit from a modified task design in which there are no informative stimuli presented between a reward image and subsequent response. As an additional caveat, we note that our paradigm and results leave room for interpretive challenges. Because participants exhibited faster response times to view social images compared to nonsocial images, we cannot discern whether our nppi results from the social-stimuli condition are due to the stimulus type itself (i.e. face images) or would generalize to other highly motivating stimuli. While prior work linking FG to face processing Page 25 of 38
27 supports the interpretation that our results are indeed due to that specific stimulus type (Engell and McCarthy, 2014; Kanwisher et al., 1997; Kanwisher and Yovel, 2006; McCarthy et al., 1997), future studies could compare within-domain images of varying attractiveness to other categories of motivating stimuli (e.g., money). Such an analysis would speak to whether these behavioral and neural results are unique to face image rewards, or are due to participant differences in subjective value (Bos et al., 2013; Smith et al., 2014a). Our results demonstrate task-dependent contributions between the DMN and ECN and the FG that shape subsequent motivated behavior. These large-scale networks are known to be disrupted in a variety of psychopathologies marked by impairments in attention and reward processing, including autism spectrum disorder (Assaf et al., 2010; Young et al., 2015), obsessive compulsive disorder (Stern et al., 2011, 2012), and major depressive disorder (Grimm et al., 2011; Sheline et al., 2009), and so an improved understanding of how they influence moment-to-moment behavior could have clinical relevance and advance models of pathophysiology (Cuthbert and Insel, 2013; Insel et al., 2010). Thus, this study marks a significant step toward better understanding and treatment of disorders characterized by impaired social and reward processing. Page 26 of 38
28 References Anticevic, A., Repovs, G., Shulman, G.L., Barch, D.M., When less is more: TPJ and default network deactivation during encoding predicts working memory performance. NeuroImage 49, doi: /j.neuroimage Assaf, M., Jagannathan, K., Calhoun, V.D., Miller, L., Stevens, M.C., Sahl, R., O Boyle, J.G., Schultz, R.T., Pearlson, G.D., Abnormal functional connectivity of default mode sub-networks in autism spectrum disorder patients. NeuroImage 53, doi: /j.neuroimage Beckmann, C.F., DeLuca, M., Devlin, J.T., Smith, S.M., Investigations into resting-state connectivity using independent component analysis. Philos. Trans. R. Soc. Lond. 360, doi: /rstb Beckmann, C.F., Smith, S.M., Probabilistic independent component analysis for functional magnetic resonance imaging. IEEE Trans. Med. Imaging 23, doi: /tmi Behrens, T.E., Sporns, O., Human connectomics. Curr. Opin. Neurobiol. 22, doi: /j.conb Berbaum, K.S., Franken, E.A., Dorfman, D.D., Rooholamini, S.A., Kathol, M.H., Barloon, T.J., Behlke, F.M., Sato, Y., Lu, C.H., el-khoury, G.Y., Satisfaction of search in diagnostic radiology. Invest. Radiol. 25, Biswal, B., Zerrin Yetkin, F., Haughton, V.M., Hyde, J.S., Functional connectivity in the motor cortex of resting human brain using echo-planar mri. Magn. Reson. Med. 34, doi: /mrm Biswal, B.B., Mennes, M., Zuo, X.-N., Gohel, S., Kelly, C., Smith, S.M., Beckmann, C.F., Adelstein, J.S., Buckner, R.L., Colcombe, S., Dogonowski, A.-M., Ernst, M., Fair, D., Hampson, M., Hoptman, M.J., Hyde, J.S., Kiviniemi, V.J., Kötter, R., Li, S.-J., Lin, C.-P., Lowe, M.J., Mackay, C., Madden, D.J., Madsen, K.H., Margulies, D.S., Mayberg, H.S., McMahon, K., Monk, C.S., Mostofsky, S.H., Nagel, B.J., Pekar, J.J., Peltier, S.J., Petersen, S.E., Riedl, V., Rombouts, S.A.R.B., Rypma, B., Schlaggar, B.L., Schmidt, S., Seidler, R.D., Siegle, G.J., Sorg, C., Teng, G.-J., Veijola, J., Villringer, A., Walter, M., Wang, L., Weng, X.-C., Whitfield-Gabrieli, S., Williamson, P., Windischberger, C., Zang, Y.-F., Zhang, H.-Y., Castellanos, F.X., Milham, M.P., Toward discovery science of human brain function. Proc. Natl. Acad. Sci. 107, doi: /pnas Bonnelle, V., Leech, R., Kinnunen, K.M., Ham, T.E., Beckmann, C.F., Boissezon, X.D., Greenwood, R.J., Sharp, D.J., Default Mode Network Connectivity Predicts Sustained Attention Deficits after Traumatic Brain Injury. J. Neurosci. 31, doi: /jneurosci Page 27 of 38
29 Bos, W. van den, Talwar, A., McClure, S.M., Neural Correlates of Reinforcement Learning and Social Preferences in Competitive Bidding. J. Neurosci. 33, doi: /jneurosci Brainard, D.H., The Psychophysics Toolbox. Spat. Vis. 10, Brewer, J.A., Worhunsky, P.D., Gray, J.R., Tang, Y.-Y., Weber, J., Kober, H., Meditation experience is associated with differences in default mode network activity and connectivity. Proc. Natl. Acad. Sci. 108, doi: /pnas Buxton, R.B., Wong, E.C., Frank, L.R., Dynamics of blood flow and oxygenation changes during brain activation: the balloon model. Magn. Reson. Med. 39, Clithero, J.A., Reeck, C., Carter, R.M., Smith, D.V., Huettel, S.A., Nucleus accumbens mediates relative motivation for rewards in the absence of choice. Front. Hum. Neurosci. 5, 87. doi: /fnhum Cole, D.M., Smith, S.M., Beckmann, C.F., Advances and pitfalls in the analysis and interpretation of resting-state FMRI data. Front. Syst. Neurosci. 4, 8. doi: /fnsys Cowdrey, F.A., Filippini, N., Park, R.J., Smith, S.M., McCabe, C., Increased resting state functional connectivity in the default mode network in recovered anorexia nervosa. Hum. Brain Mapp. 0. doi: /hbm Cuthbert, B.N., Insel, T.R., Toward the future of psychiatric diagnosis: the seven pillars of RDoC. BMC Med. 11, 126. doi: / De Havas, J.A., Parimal, S., Soon, C.S., Chee, M.W.L., Sleep deprivation reduces default mode network connectivity and anti-correlation during rest and task performance. NeuroImage 59, doi: /j.neuroimage Dosenbach, N.U.F., Fair, D.A., Miezin, F.M., Cohen, A.L., Wenger, K.K., Dosenbach, R. a T., Fox, M.D., Snyder, A.Z., Vincent, J.L., Raichle, M.E., Schlaggar, B.L., Petersen, S.E., Distinct brain networks for adaptive and stable task control in humans. Proc. Natl. Acad. Sci. 104, doi: /pnas Ebitz, R.B., Watson, K.K., Platt, M.L., Oxytocin blunts social vigilance in the rhesus macaque. Proc. Natl. Acad. Sci. 110, doi: /pnas Eichele, T., Debener, S., Calhoun, V.D., Specht, K., Engel, A.K., Hugdahl, K., von Cramon, D.Y., Ullsperger, M., Prediction of human errors by maladaptive changes in event-related brain networks. Proc. Natl. Acad. Sci. 105, doi: /pnas Eklund, A., Nichols, T.E., Knutsson, H., Cluster failure: Why fmri inferences for spatial extent have inflated false-positive rates. Proc. Natl. Acad. Sci. 113, doi: /pnas Page 28 of 38
30 Engell, A.D., McCarthy, G., Face, eye, and body selective responses in fusiform gyrus and adjacent cortex: an intracranial EEG study. Front. Hum. Neurosci. 8. doi: /fnhum Filippini, N., MacIntosh, B.J., Hough, M.G., Goodwin, G.M., Frisoni, G.B., Smith, S.M., Matthews, P.M., Beckmann, C.F., Mackay, C.E., Distinct patterns of brain activity in young carriers of the APOE-ϵ4 allele. Proc. Natl. Acad. Sci. 106, doi: /pnas Fleck, M.S., Samei, E., Mitroff, S.R., Generalized satisfaction of search : adverse influences on dual-target search accuracy. J. Exp. Psychol. 16, doi: /a Fox, M.D., Halko, M.A., Eldaief, M.C., Pascual-Leone, A., Measuring and manipulating brain connectivity with resting state functional connectivity magnetic resonance imaging (fcmri) and transcranial magnetic stimulation (TMS). NeuroImage 62, doi: /j.neuroimage Friedman, L., Glover, G.H., Reducing interscanner variability of activation in a multicenter fmri study: controlling for signal-to-fluctuation-noise-ratio (SFNR) differences. NeuroImage 33, doi: /j.neuroimage Friston, K.J., Functional and Effective Connectivity: A Review. Brain Connect. 1, doi: /brain Friston, K.J., Buechel, C., Fink, G.R., Morris, J., Rolls, E., Dolan, R.J., Psychophysiological and Modulatory Interactions in Neuroimaging. NeuroImage 6, doi: /nimg Friston, K.J., Harrison, L., Penny, W., Dynamic causal modelling. NeuroImage 19, doi: /s (03) Grimm, S., Ernst, J., Boesiger, P., Schuepbach, D., Boeker, H., Northoff, G., Reduced negative BOLD responses in the default-mode network and increased self-focus in depression. World J. Biol. Psychiatry 12, doi: / Gui, D., Xu, S., Zhu, S., Fang, Z., Spaeth, A.M., Xin, Y., Feng, T., Rao, H., Resting spontaneous activity in the default mode network predicts performance decline during prolonged attention workload. NeuroImage. doi: /j.neuroimage Gusnard, D.A., Raichle, M.E., Searching for a baseline: Functional imaging and the resting human brain. Nat. Rev. Neurosci. 2, doi: / Hampstead, B.M., Brown, G.S., Hartley, J.F., Transcranial Direct Current Stimulation Modulates Activation and Effective Connectivity During Spatial Navigation. Brain Stimulat. 7, doi: /j.brs Page 29 of 38
31 Hayden, B.Y., Smith, D.V., Platt, M.L., Cognitive control signals in posterior cingulate cortex. Front. Hum. Neurosci. 4, 223. doi: /fnhum Hayden, B.Y., Smith, D.V., Platt, M.L., Electrophysiological correlates of default-mode processing in macaque posterior cingulate cortex. Proc. Natl. Acad. Sci. 106, doi: /pnas Heilbronner, S.R., Platt, M.L., Causal Evidence of Performance Monitoring by Neurons in Posterior Cingulate Cortex during Learning. Neuron 80, doi: /j.neuron Hyvärinen, A., Fast and robust fixed-point algorithms for independent component analysis. IEEE Trans. Neural Netw. 10, doi: / Insel, T., Cuthbert, B., Garvey, M., Heinssen, R., Pine, D.S., Quinn, K., Sanislow, C., Wang, P., Research domain criteria (RDoC): Toward a new classification framework for research on mental disorders. Am. J. Psychiatry 167, doi: /appi.ajp Jenkinson, M., Smith, S., A global optimisation method for robust affine registration of brain images. Med. Image Anal. 5, Kanwisher, N., McDermott, J., Chun, M.M., The fusiform face area: A module in human extrastriate cortex specialized for face perception. J. Neurosci. 17, Kanwisher, N., Yovel, G., The fusiform face area: a cortical region specialized for the perception of faces. Philos. Trans. R. Soc. B Biol. Sci. 361, doi: /rstb Keeser, D., Meindl, T., Bor, J., Palm, U., Pogarell, O., Mulert, C., Brunelin, J., Moller, H.-J., Reiser, M., Padberg, F., Prefrontal Transcranial Direct Current Stimulation Changes Connectivity of Resting-State Networks during fmri. J. Neurosci. 31, doi: /jneurosci Kelly, A., Uddin, L.Q., Biswal, B.B., Castellanos, F.X., Milham, M.P., Competition between functional brain networks mediates behavioral variability. NeuroImage 39, doi: /j.neuroimage Knutson, B., Adams, C.M., Fong, G.W., Hommer, D., Anticipation of Increasing Monetary Reward Selectively Recruits Nucleus Accumbens. J. Neurosci. 21, RC159-RC159. Knutson, B., Westdorp, A., Kaiser, E., Hommer, D., FMRI visualization of brain activity during a monetary incentive delay task. NeuroImage 12, doi: /nimg Kucyi, A., Moayedi, M., Weissman-Fogel, I., Goldberg, M.B., Freeman, B.V., Tenenbaum, H.C., Davis, K.D., Enhanced Medial Prefrontal-Default Mode Network Functional Page 30 of 38
32 Connectivity in Chronic Pain and Its Association with Pain Rumination. J. Neurosci. 34, doi: /jneurosci Leech, R., Kamourieh, S., Beckmann, C.F., Sharp, D.J., Fractionating the default mode network: distinct contributions of the ventral and dorsal posterior cingulate cortex to cognitive control. J. Neurosci. 31, doi: /jneurosci Luber, B., Lisanby, S.H., Enhancement of human cognitive performance using transcranial magnetic stimulation (TMS). NeuroImage 85, Part 3, doi: /j.neuroimage McCarthy, G., Puce, A., Gore, J.C., Allison, T., Face-Specific Processing in the Human Fusiform Gyrus. J. Cogn. Neurosci. 9, doi: /jocn McLaren, D.G., Ries, M.L., Xu, G., Johnson, S.C., A generalized form of context-dependent psychophysiological interactions (gppi): A comparison to standard approaches. NeuroImage 61, doi: /j.neuroimage Mueller, J.K., Grigsby, E.M., Prevosto, V., Petraglia Iii, F.W., Rao, H., Deng, Z.-D., Peterchev, A.V., Sommer, M.A., Egner, T., Platt, M.L., Grill, W.M., Simultaneous transcranial magnetic stimulation and single-neuron recording in alert non-human primates. Nat. Neurosci. advance online publication. doi: /nn.3751 Nichols, T.E., Holmes, A.P., Nonparametric permutation tests for functional neuroimaging: a primer with examples. Hum. Brain Mapp. 15, O Reilly, J.X., Woolrich, M.W., Behrens, T.E., Smith, S.M., Johansen-Berg, H., Tools of the trade: psychophysiological interactions and functional connectivity. Soc. Cogn. Affect. Neurosci. 7, doi: /scan/nss055 Ossandon, T., Jerbi, K., Vidal, J.R., Bayle, D.J., Henaff, M.-A., Jung, J., Minotti, L., Bertrand, O., Kahane, P., Lachaux, J.-P., Transient Suppression of Broadband Gamma Power in the Default-Mode Network Is Correlated with Task Complexity and Subject Performance. J. Neurosci. 31, doi: /jneurosci Piccoli, T., Valente, G., Linden, D.E.J., Re, M., Esposito, F., Sack, A.T., Salle, F.D., The Default Mode Network and the Working Memory Network Are Not Anti-Correlated during All Phases of a Working Memory Task. PLoS ONE 10, e doi: /journal.pone Ray, K.L., McKay, D.R., Fox, P.M., Riedel, M.C., Uecker, A.M., Beckmann, C.F., Smith, S.M., Fox, P.T., Laird, A.R., ICA model order selection of task co-activation networks. Front. Neurosci. 7. doi: /fnins Rorden, C., Karnath, H.-O., Bonilha, L., Improving Lesion-Symptom Mapping. J. Cogn. Neurosci. 19, doi: /jocn Page 31 of 38
33 Rosen, M.L., Stern, C.E., Michalka, S.W., Devaney, K.J., Somers, D.C., Cognitive Control Network Contributions to Memory-Guided Visual Attention. Cereb. Cortex bhv028. doi: /cercor/bhv028 Sambataro, F., Murty, V.P., Callicott, J.H., Tan, H.-Y., Das, S., Weinberger, D.R., Mattay, V.S., Age-related alterations in default mode network: Impact on working memory performance. Neurobiol. Aging 31, doi: /j.neurobiolaging Seeley, W.W., Menon, V., Schatzberg, A.F., Keller, J., Glover, G.H., Kenna, H., Reiss, A.L., Greicius, M.D., Dissociable intrinsic connectivity networks for salience processing and executive control. J. Neurosci. 27, doi: /jneurosci Shannon, B.J., Raichle, M.E., Snyder, A.Z., Fair, D.A., Mills, K.L., Zhang, D., Bache, K., Calhoun, V.D., Nigg, J.T., Nagel, B.J., Stevens, A.A., Kiehl, K.A., Premotor functional connectivity predicts impulsivity in juvenile offenders. Proc. Natl. Acad. Sci. U. S. A. 108, doi: /pnas Sheline, Y.I., Barch, D.M., Price, J.L., Rundle, M.M., Vaishnavi, S.N., Snyder, A.Z., Mintun, M.A., Wang, S., Coalson, R.S., Raichle, M.E., The default mode network and selfreferential processes in depression. Proc. Natl. Acad. Sci. 106, doi: /pnas Sladky, R., Friston, K.J., Tröstl, J., Cunnington, R., Moser, E., Windischberger, C., Slicetiming effects and their correction in functional MRI. NeuroImage 58, doi: /j.neuroimage Smith, D.V., Clithero, J.A., Boltuck, S.E., Huettel, S.A., 2014a. Functional connectivity with ventromedial prefrontal cortex reflects subjective value for social rewards. Soc. Cogn. Affect. Neurosci. 9, doi: /scan/nsu005 Smith, D.V., Delgado, M.R., Meta-analysis of psychophysiological interactions: Revisiting cluster-level thresholding and sample sizes. Hum. Brain Mapp. n/a-n/a. doi: /hbm Smith, D.V., Gseir, M., Speer, M.E., Delgado, M.R., Toward a cumulative science of functional integration: A meta-analysis of psychophysiological interactions. Hum. Brain Mapp. Smith, D.V., Hayden, B.Y., Truong, T.-K., Song, A.W., Platt, M.L., Huettel, S.A., Distinct value signals in anterior and posterior ventromedial prefrontal cortex. J. Neurosci. 30, doi: /jneurosci Smith, D.V., Utevsky, A.V., Bland, A.R., Clement, N., Clithero, J.A., Harsch, A.E.W., McKell Carter, R., Huettel, S.A., 2014b. Characterizing individual differences in functional connectivity using dual-regression and seed-based approaches. NeuroImage 95, doi: /j.neuroimage Page 32 of 38
34 Smith, S.M., Fast robust automated brain extraction. Hum. Brain Mapp. 17, doi: /hbm Smith, S.M., Beckmann, C.F., Andersson, J., Auerbach, E.J., Bijsterbosch, J., Douaud, G., Duff, E., Feinberg, D.A., Griffanti, L., Harms, M.P., Kelly, M., Laumann, T., Miller, K.L., Moeller, S., Petersen, S., Power, J., Salimi-Khorshidi, G., Snyder, A.Z., Vu, A.T., Woolrich, M.W., Xu, J., Yacoub, E., Uğurbil, K., Van Essen, D.C., Glasser, M.F., Resting-state fmri in the Human Connectome Project. NeuroImage, Mapping the Connectome 80, doi: /j.neuroimage Smith, S.M., Fox, P.T., Miller, K.L., Glahn, D.C., Fox, P.M., Mackay, C.E., Filippini, N., Watkins, K.E., Toro, R., Laird, A.R., Beckmann, C.F., Correspondence of the brain s functional architecture during activation and rest. Proc. Natl. Acad. Sci. 106, doi: /pnas Smith, S.M., Jenkinson, M., Woolrich, M.W., Beckmann, C.F., Behrens, T.E., Johansen-Berg, H., Bannister, P.R., De Luca, M., Drobnjak, I., Flitney, D.E., Niazy, R.K., Saunders, J., Vickers, J., Zhang, Y., De Stefano, N., Brady, J.M., Matthews, P.M., Advances in functional and structural MR image analysis and implementation as FSL. NeuroImage 23 Suppl 1, S doi: /j.neuroimage Stern, E.R., Fitzgerald, K.D., Welsh, R.C., Abelson, J.L., Taylor, S.F., Resting-State Functional Connectivity between Fronto-Parietal and Default Mode Networks in Obsessive-Compulsive Disorder. PLoS ONE 7, e doi: /journal.pone Stern, E.R., Welsh, R.C., Fitzgerald, K.D., Gehring, W.J., Lister, J.J., Himle, J.A., Abelson, J.L., Taylor, S.F., Hyperactive Error Responses and Altered Connectivity in Ventromedial and Frontoinsular Cortices in Obsessive-Compulsive Disorder. Biol. Psychiatry, Genes and Anxiety 69, doi: /j.biopsych Utevsky, A.V., Smith, D.V., Huettel, S.A., Precuneus is a functional core of the defaultmode network. J. Neurosci. 34, doi: /jneurosci Weissman, D.H., Roberts, K.C., Visscher, K.M., Woldorff, M.G., The neural bases of momentary lapses in attention. Nat. Neurosci. 9, doi: /nn1727 Winkler, A.M., Ridgway, G.R., Webster, M.A., Smith, S.M., Nichols, T.E., Permutation inference for the general linear model. NeuroImage 92, doi: /j.neuroimage Woolrich, M.W., Jbabdi, S., Patenaude, B., Chappell, M., Makni, S., Behrens, T.E., Beckmann, C.F., Jenkinson, M., Smith, S.M., Bayesian analysis of neuroimaging data in FSL. NeuroImage 45, S doi: /j.neuroimage Yarkoni, T., Poldrack, R.A., Nichols, T.E., Van Essen, D.C., Wager, T.D., Large-scale automated synthesis of human functional neuroimaging data. Nat. Methods 8, doi: /nmeth.1635 Page 33 of 38
35 Young, J.S., Smith, D.V., Coutlee, C.G., Huettel, S.A., Synchrony between sensory and cognitive networks is associated with subclinical variation in autistic traits. Front. Hum. Neurosci doi: /fnhum Page 34 of 38
36 841 Figure Captions Figure 1: Task Design Fifty heterosexual men performed a modified MID task which examined timed responses to view social and nonsocial images as rewards (faces and landscapes, respectively). Each trial of the task began with an abstract image presented as a cue for 1500 ms indicating the potential reward for that trial. Following the cue, a fixation cross appeared for a variable duration (interstimulus interval; ms), followed by a target (a white triangle) that appeared on the screen for 750 ms; participants responded by pressing a button box with their right index finger before the target disappeared. After the target, a feedback image (a colored triangle) appeared for 750 ms; the feedback image was colored green if the participants response was sufficiently fast on the trial (a win), and was blue if the response was too slow (a loss). After the feedback image disappeared, the participants waited a delay period that varied according to the cue and whether the subject won or lost that trial. Lastly, the reward image (either a face or a landscape) was shown for 2000 ms. Consent was provided for the use of the image in this figure Figure 2: Response Times are Slower Following Face Trials Compared to Landscape Trials (A) Distribution of response time (RT) differences according to previous trial s stimulus type, calculated by subtracting RTs following nonsocial trials from RTs following social trials. Response times on trials following social rewards were greater than those on trials following nonsocial rewards, indicating an effect of previous reward stimulus type on subsequent behavior. (B) Average beta weights (with S.E.M. plotted) across subjects from a behavioral Page 35 of 38
37 regression predicting current RT. We regressed current trial RT on a model including the following regressors: stimulus category (social or nonsocial) on previous trial, RT on previous trial, attractiveness rating of the previous trial s reward, and stimulus category on current trial. Of these four regressors, current stimulus category most strongly predicted current RT; the negative beta weight indicates that participants are faster to respond during a social trial compared to during a nonsocial trial. The next most predictive regressor was previous trial s stimulus category; the positive beta weight indicates that participants were slower to respond following social trials compared to following nonsocial trials. Neither previous trial s RT nor previous trial s reward attractiveness significantly predicted the current trial s RT Figure 3: Networks Identified from Independent Components Analysis Using independent components analysis across our task runs, we identified large-scale functional networks. This analysis produced 25 components. Our analyses focused on the components that best matched default-mode network (highest-correlating component illustrated on top) and the executive control network (highest-correlating component illustrated on bottom) from Smith and colleagues (2009). For visualization purposes, maps are thresholded at Z > Figure 4: Fusiform Gyrus Connectivity Tracks Effect of Stimulus Type on Subsequent Behavior (A) A network PPI indicated that an area in the fusiform gyrus (FG) exhibits heightened effective connectivity with ECN (compared to DMN) during social reward outcomes; thresholded at p < We note that this result also held with permutation-based testing Page 36 of 38
38 (Eklund et al., 2016). (B) Parameters estimates extracted from the FG connectivity track the effect of stimulus type on subsequent behavior (stimulus-type beta weights estimated from our behavioral GLM; see Behavioral Analysis): as FG-DMN connectivity increases relative to FG-ECN connectivity, RTs are further slowed following social trials compared to following nonsocial trials. 890 Page 37 of 38
39 891 Table Captions Table 1. Behavioral Regression Results To examine the effects of other trial characteristics on current trial RT, we regressed current trial RT on a model including: stimulus category (social or nonsocial) on previous trial, RT on previous trial, attractiveness rating of the previous trial s reward, and stimulus category on current trial. Our analysis indicated that current stimulus type had the strongest effect on current RT; however, of all the characteristics from the prior trial, only prior stimulus type had a significant effect on current RT. Regressor Parameter Estimate (s.e.m) t-stat p-value Current trial (t) stimulus type (0.013) < Prior trial (t-1) stimulus type (0.013) Prior trial (t-1) attractiveness (0.011) Prior trial (t-1) response time < (0.013) Table 2. Spatial Correspondence with Canonical Networks To identify the DMN and ECN, we ran spatial correlations between canonical neural networks (Smith et al., 2009) and the 25 components from our ICA. The highest-correlating ICA component numbers for each network map and the correlation values are listed. Canonical Network Independent Component No. Spatial Correlation (r) Visual 1 IC Visual 2 IC Visual 3 IC Default-Mode IC Cerebellar IC Sensorimotor IC Auditory IC Executive Control IC R Frontoparietal IC L Frontoparietal IC Page 38 of 38

40 Outcome Cue ISI Target Feedback Delay ms ITI 1500ms ms 750ms 750ms ms ms 2000ms
 0.02 0-0.02-0.04-0.06 ** p < 0.")
41 A 8 Number of Participants Faster following face trials Faster following landscape trials Face-Land RT Difference (s) B 0.04 * Previous trial attractiveness Previous trial stimulus type Previous trial reaction time Current trial stimulus type Parameter Estimates (β) ** p < * p < 0.05 **

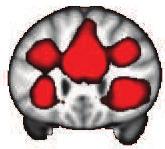
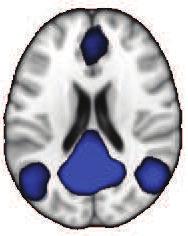
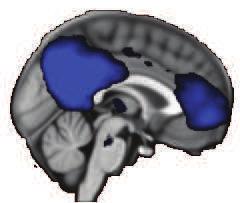
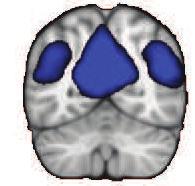
42 Z = 21 Y = -61 X = 0 Default-Mode Network Z = 12 Y = 27 X = 0 Executive Control Network
 β) Faster after social trials Slower after social trials")
43 A B Increased FG-ECN coupling Increased FG-DMN coupling Network Coupling with FG (DMN - ECN PPI β) Z = -17 Y = -65 X = Change in Behavior (Face-Scene (Current Stimulus RT Effect β) β) Faster after social trials Slower after social trials
Supplementary Information Methods Subjects The study was comprised of 84 chronic pain patients with either chronic back pain (CBP) or osteoarthritis
 or osteoarthritis") Supplementary Information Methods Subjects The study was comprised of 84 chronic pain patients with either chronic back pain (CBP) or osteoarthritis (OA). All subjects provided informed consent to procedures
Supplementary Information Methods Subjects The study was comprised of 84 chronic pain patients with either chronic back pain (CBP) or osteoarthritis (OA). All subjects provided informed consent to procedures
Investigations in Resting State Connectivity. Overview
 Investigations in Resting State Connectivity Scott FMRI Laboratory Overview Introduction Functional connectivity explorations Dynamic change (motor fatigue) Neurological change (Asperger s Disorder, depression)
Investigations in Resting State Connectivity Scott FMRI Laboratory Overview Introduction Functional connectivity explorations Dynamic change (motor fatigue) Neurological change (Asperger s Disorder, depression)
Classification and Statistical Analysis of Auditory FMRI Data Using Linear Discriminative Analysis and Quadratic Discriminative Analysis
 International Journal of Innovative Research in Computer Science & Technology (IJIRCST) ISSN: 2347-5552, Volume-2, Issue-6, November-2014 Classification and Statistical Analysis of Auditory FMRI Data Using
International Journal of Innovative Research in Computer Science & Technology (IJIRCST) ISSN: 2347-5552, Volume-2, Issue-6, November-2014 Classification and Statistical Analysis of Auditory FMRI Data Using
Supplementary information Detailed Materials and Methods
 Supplementary information Detailed Materials and Methods Subjects The experiment included twelve subjects: ten sighted subjects and two blind. Five of the ten sighted subjects were expert users of a visual-to-auditory
Supplementary information Detailed Materials and Methods Subjects The experiment included twelve subjects: ten sighted subjects and two blind. Five of the ten sighted subjects were expert users of a visual-to-auditory
Group-Wise FMRI Activation Detection on Corresponding Cortical Landmarks
 Group-Wise FMRI Activation Detection on Corresponding Cortical Landmarks Jinglei Lv 1,2, Dajiang Zhu 2, Xintao Hu 1, Xin Zhang 1,2, Tuo Zhang 1,2, Junwei Han 1, Lei Guo 1,2, and Tianming Liu 2 1 School
Group-Wise FMRI Activation Detection on Corresponding Cortical Landmarks Jinglei Lv 1,2, Dajiang Zhu 2, Xintao Hu 1, Xin Zhang 1,2, Tuo Zhang 1,2, Junwei Han 1, Lei Guo 1,2, and Tianming Liu 2 1 School
SUPPLEMENT: DYNAMIC FUNCTIONAL CONNECTIVITY IN DEPRESSION. Supplemental Information. Dynamic Resting-State Functional Connectivity in Major Depression
 Supplemental Information Dynamic Resting-State Functional Connectivity in Major Depression Roselinde H. Kaiser, Ph.D., Susan Whitfield-Gabrieli, Ph.D., Daniel G. Dillon, Ph.D., Franziska Goer, B.S., Miranda
Supplemental Information Dynamic Resting-State Functional Connectivity in Major Depression Roselinde H. Kaiser, Ph.D., Susan Whitfield-Gabrieli, Ph.D., Daniel G. Dillon, Ph.D., Franziska Goer, B.S., Miranda
Functional connectivity in fmri
 Functional connectivity in fmri Cyril Pernet, PhD Language and Categorization Laboratory, Brain Research Imaging Centre, University of Edinburgh Studying networks fmri can be used for studying both, functional
Functional connectivity in fmri Cyril Pernet, PhD Language and Categorization Laboratory, Brain Research Imaging Centre, University of Edinburgh Studying networks fmri can be used for studying both, functional
Supplementary Online Content
 Supplementary Online Content Green SA, Hernandez L, Tottenham N, Krasileva K, Bookheimer SY, Dapretto M. The neurobiology of sensory overresponsivity in youth with autism spectrum disorders. Published
Supplementary Online Content Green SA, Hernandez L, Tottenham N, Krasileva K, Bookheimer SY, Dapretto M. The neurobiology of sensory overresponsivity in youth with autism spectrum disorders. Published
Procedia - Social and Behavioral Sciences 159 ( 2014 ) WCPCG 2014
 WCPCG 2014") Available online at www.sciencedirect.com ScienceDirect Procedia - Social and Behavioral Sciences 159 ( 2014 ) 743 748 WCPCG 2014 Differences in Visuospatial Cognition Performance and Regional Brain Activation
Available online at www.sciencedirect.com ScienceDirect Procedia - Social and Behavioral Sciences 159 ( 2014 ) 743 748 WCPCG 2014 Differences in Visuospatial Cognition Performance and Regional Brain Activation
Supporting Online Material for
 www.sciencemag.org/cgi/content/full/324/5927/646/dc1 Supporting Online Material for Self-Control in Decision-Making Involves Modulation of the vmpfc Valuation System Todd A. Hare,* Colin F. Camerer, Antonio
www.sciencemag.org/cgi/content/full/324/5927/646/dc1 Supporting Online Material for Self-Control in Decision-Making Involves Modulation of the vmpfc Valuation System Todd A. Hare,* Colin F. Camerer, Antonio
WHAT DOES THE BRAIN TELL US ABOUT TRUST AND DISTRUST? EVIDENCE FROM A FUNCTIONAL NEUROIMAGING STUDY 1
 SPECIAL ISSUE WHAT DOES THE BRAIN TE US ABOUT AND DIS? EVIDENCE FROM A FUNCTIONAL NEUROIMAGING STUDY 1 By: Angelika Dimoka Fox School of Business Temple University 1801 Liacouras Walk Philadelphia, PA
SPECIAL ISSUE WHAT DOES THE BRAIN TE US ABOUT AND DIS? EVIDENCE FROM A FUNCTIONAL NEUROIMAGING STUDY 1 By: Angelika Dimoka Fox School of Business Temple University 1801 Liacouras Walk Philadelphia, PA
Twelve right-handed subjects between the ages of 22 and 30 were recruited from the
 Supplementary Methods Materials & Methods Subjects Twelve right-handed subjects between the ages of 22 and 30 were recruited from the Dartmouth community. All subjects were native speakers of English,
Supplementary Methods Materials & Methods Subjects Twelve right-handed subjects between the ages of 22 and 30 were recruited from the Dartmouth community. All subjects were native speakers of English,
Supporting online material. Materials and Methods. We scanned participants in two groups of 12 each. Group 1 was composed largely of
 Placebo effects in fmri Supporting online material 1 Supporting online material Materials and Methods Study 1 Procedure and behavioral data We scanned participants in two groups of 12 each. Group 1 was
Placebo effects in fmri Supporting online material 1 Supporting online material Materials and Methods Study 1 Procedure and behavioral data We scanned participants in two groups of 12 each. Group 1 was
Distinct Value Signals in Anterior and Posterior Ventromedial Prefrontal Cortex
 Supplementary Information Distinct Value Signals in Anterior and Posterior Ventromedial Prefrontal Cortex David V. Smith 1-3, Benjamin Y. Hayden 1,4, Trong-Kha Truong 2,5, Allen W. Song 2,5, Michael L.
Supplementary Information Distinct Value Signals in Anterior and Posterior Ventromedial Prefrontal Cortex David V. Smith 1-3, Benjamin Y. Hayden 1,4, Trong-Kha Truong 2,5, Allen W. Song 2,5, Michael L.
Temporal preprocessing of fmri data
 Temporal preprocessing of fmri data Blaise Frederick, Ph.D., Yunjie Tong, Ph.D. McLean Hospital Brain Imaging Center Scope This talk will summarize the sources and characteristics of unwanted temporal
Temporal preprocessing of fmri data Blaise Frederick, Ph.D., Yunjie Tong, Ph.D. McLean Hospital Brain Imaging Center Scope This talk will summarize the sources and characteristics of unwanted temporal
Supporting Information. Demonstration of effort-discounting in dlpfc
 Supporting Information Demonstration of effort-discounting in dlpfc In the fmri study on effort discounting by Botvinick, Huffstettler, and McGuire [1], described in detail in the original publication,
Supporting Information Demonstration of effort-discounting in dlpfc In the fmri study on effort discounting by Botvinick, Huffstettler, and McGuire [1], described in detail in the original publication,
Resistance to forgetting associated with hippocampus-mediated. reactivation during new learning
 Resistance to Forgetting 1 Resistance to forgetting associated with hippocampus-mediated reactivation during new learning Brice A. Kuhl, Arpeet T. Shah, Sarah DuBrow, & Anthony D. Wagner Resistance to
Resistance to Forgetting 1 Resistance to forgetting associated with hippocampus-mediated reactivation during new learning Brice A. Kuhl, Arpeet T. Shah, Sarah DuBrow, & Anthony D. Wagner Resistance to
Temporal preprocessing of fmri data
 Temporal preprocessing of fmri data Blaise Frederick, Ph.D. McLean Hospital Brain Imaging Center Scope fmri data contains temporal noise and acquisition artifacts that complicate the interpretation of
Temporal preprocessing of fmri data Blaise Frederick, Ph.D. McLean Hospital Brain Imaging Center Scope fmri data contains temporal noise and acquisition artifacts that complicate the interpretation of
Face-specific resting functional connectivity between the fusiform gyrus and posterior superior temporal sulcus
 HUMAN NEUROSCIENCE Original Research Article published: 24 September 2010 doi: 10.3389/fnhum.2010.00176 Face-specific resting functional connectivity between the fusiform gyrus and posterior superior temporal
HUMAN NEUROSCIENCE Original Research Article published: 24 September 2010 doi: 10.3389/fnhum.2010.00176 Face-specific resting functional connectivity between the fusiform gyrus and posterior superior temporal
Functional Elements and Networks in fmri
 Functional Elements and Networks in fmri Jarkko Ylipaavalniemi 1, Eerika Savia 1,2, Ricardo Vigário 1 and Samuel Kaski 1,2 1- Helsinki University of Technology - Adaptive Informatics Research Centre 2-
Functional Elements and Networks in fmri Jarkko Ylipaavalniemi 1, Eerika Savia 1,2, Ricardo Vigário 1 and Samuel Kaski 1,2 1- Helsinki University of Technology - Adaptive Informatics Research Centre 2-
Neuroimaging. BIE601 Advanced Biological Engineering Dr. Boonserm Kaewkamnerdpong Biological Engineering Program, KMUTT. Human Brain Mapping
 11/8/2013 Neuroimaging N i i BIE601 Advanced Biological Engineering Dr. Boonserm Kaewkamnerdpong Biological Engineering Program, KMUTT 2 Human Brain Mapping H Human m n brain br in m mapping ppin can nb
11/8/2013 Neuroimaging N i i BIE601 Advanced Biological Engineering Dr. Boonserm Kaewkamnerdpong Biological Engineering Program, KMUTT 2 Human Brain Mapping H Human m n brain br in m mapping ppin can nb
Functional MRI Mapping Cognition
 Outline Functional MRI Mapping Cognition Michael A. Yassa, B.A. Division of Psychiatric Neuro-imaging Psychiatry and Behavioral Sciences Johns Hopkins School of Medicine Why fmri? fmri - How it works Research
Outline Functional MRI Mapping Cognition Michael A. Yassa, B.A. Division of Psychiatric Neuro-imaging Psychiatry and Behavioral Sciences Johns Hopkins School of Medicine Why fmri? fmri - How it works Research
Network-based pattern recognition models for neuroimaging
 Network-based pattern recognition models for neuroimaging Maria J. Rosa Centre for Neuroimaging Sciences, Institute of Psychiatry King s College London, UK Outline Introduction Pattern recognition Network-based
Network-based pattern recognition models for neuroimaging Maria J. Rosa Centre for Neuroimaging Sciences, Institute of Psychiatry King s College London, UK Outline Introduction Pattern recognition Network-based
Inverse problems in functional brain imaging Identification of the hemodynamic response in fmri
 Inverse problems in functional brain imaging Identification of the hemodynamic response in fmri Ph. Ciuciu1,2 philippe.ciuciu@cea.fr 1: CEA/NeuroSpin/LNAO May 7, 2010 www.lnao.fr 2: IFR49 GDR -ISIS Spring
Inverse problems in functional brain imaging Identification of the hemodynamic response in fmri Ph. Ciuciu1,2 philippe.ciuciu@cea.fr 1: CEA/NeuroSpin/LNAO May 7, 2010 www.lnao.fr 2: IFR49 GDR -ISIS Spring
Personal Space Regulation by the Human Amygdala. California Institute of Technology
 Personal Space Regulation by the Human Amygdala Daniel P. Kennedy 1, Jan Gläscher 1, J. Michael Tyszka 2 & Ralph Adolphs 1,2 1 Division of Humanities and Social Sciences 2 Division of Biology California
Personal Space Regulation by the Human Amygdala Daniel P. Kennedy 1, Jan Gläscher 1, J. Michael Tyszka 2 & Ralph Adolphs 1,2 1 Division of Humanities and Social Sciences 2 Division of Biology California
Experimental Design. Outline. Outline. A very simple experiment. Activation for movement versus rest
 Experimental Design Kate Watkins Department of Experimental Psychology University of Oxford With thanks to: Heidi Johansen-Berg Joe Devlin Outline Choices for experimental paradigm Subtraction / hierarchical
Experimental Design Kate Watkins Department of Experimental Psychology University of Oxford With thanks to: Heidi Johansen-Berg Joe Devlin Outline Choices for experimental paradigm Subtraction / hierarchical
Supplementary materials for: Executive control processes underlying multi- item working memory
 Supplementary materials for: Executive control processes underlying multi- item working memory Antonio H. Lara & Jonathan D. Wallis Supplementary Figure 1 Supplementary Figure 1. Behavioral measures of
Supplementary materials for: Executive control processes underlying multi- item working memory Antonio H. Lara & Jonathan D. Wallis Supplementary Figure 1 Supplementary Figure 1. Behavioral measures of
Number of studies with resting and state and fmri in their title/abstract
 430 Chapter 11 seed voxel A voxel chosen as a starting point for a connectivity analysis. Figure 11.13 Studies of spontaneous BOLD fluctuations in the resting state have become an increasingly important
430 Chapter 11 seed voxel A voxel chosen as a starting point for a connectivity analysis. Figure 11.13 Studies of spontaneous BOLD fluctuations in the resting state have become an increasingly important
FMRI Data Analysis. Introduction. Pre-Processing
 FMRI Data Analysis Introduction The experiment used an event-related design to investigate auditory and visual processing of various types of emotional stimuli. During the presentation of each stimuli
FMRI Data Analysis Introduction The experiment used an event-related design to investigate auditory and visual processing of various types of emotional stimuli. During the presentation of each stimuli
Changes in Default Mode Network as Automaticity Develops in a Categorization Task
 Purdue University Purdue e-pubs Open Access Theses Theses and Dissertations January 2015 Changes in Default Mode Network as Automaticity Develops in a Categorization Task Farzin Shamloo Purdue University
Purdue University Purdue e-pubs Open Access Theses Theses and Dissertations January 2015 Changes in Default Mode Network as Automaticity Develops in a Categorization Task Farzin Shamloo Purdue University
BRAIN STATE CHANGE DETECTION VIA FIBER-CENTERED FUNCTIONAL CONNECTIVITY ANALYSIS
 BRAIN STATE CHANGE DETECTION VIA FIBER-CENTERED FUNCTIONAL CONNECTIVITY ANALYSIS Chulwoo Lim 1, Xiang Li 1, Kaiming Li 1, 2, Lei Guo 2, Tianming Liu 1 1 Department of Computer Science and Bioimaging Research
BRAIN STATE CHANGE DETECTION VIA FIBER-CENTERED FUNCTIONAL CONNECTIVITY ANALYSIS Chulwoo Lim 1, Xiang Li 1, Kaiming Li 1, 2, Lei Guo 2, Tianming Liu 1 1 Department of Computer Science and Bioimaging Research
Investigations in Resting State Connectivity. Overview. Functional connectivity. Scott Peltier. FMRI Laboratory University of Michigan
 Investigations in Resting State Connectivity Scott FMRI Laboratory Overview Introduction Functional connectivity explorations Dynamic change (motor fatigue) Neurological change (Asperger s Disorder, depression)
Investigations in Resting State Connectivity Scott FMRI Laboratory Overview Introduction Functional connectivity explorations Dynamic change (motor fatigue) Neurological change (Asperger s Disorder, depression)
Prediction of Successful Memory Encoding from fmri Data
 Prediction of Successful Memory Encoding from fmri Data S.K. Balci 1, M.R. Sabuncu 1, J. Yoo 2, S.S. Ghosh 3, S. Whitfield-Gabrieli 2, J.D.E. Gabrieli 2 and P. Golland 1 1 CSAIL, MIT, Cambridge, MA, USA
Prediction of Successful Memory Encoding from fmri Data S.K. Balci 1, M.R. Sabuncu 1, J. Yoo 2, S.S. Ghosh 3, S. Whitfield-Gabrieli 2, J.D.E. Gabrieli 2 and P. Golland 1 1 CSAIL, MIT, Cambridge, MA, USA
FINAL PROGRESS REPORT
 (1) Foreword (optional) (2) Table of Contents (if report is more than 10 pages) (3) List of Appendixes, Illustrations and Tables (if applicable) (4) Statement of the problem studied FINAL PROGRESS REPORT
(1) Foreword (optional) (2) Table of Contents (if report is more than 10 pages) (3) List of Appendixes, Illustrations and Tables (if applicable) (4) Statement of the problem studied FINAL PROGRESS REPORT
smokers) aged 37.3 ± 7.4 yrs (mean ± sd) and a group of twelve, age matched, healthy
 aged 37.3 ± 7.4 yrs (mean ± sd) and a group of twelve, age matched, healthy") Methods Participants We examined a group of twelve male pathological gamblers (ten strictly right handed, all smokers) aged 37.3 ± 7.4 yrs (mean ± sd) and a group of twelve, age matched, healthy males,
Methods Participants We examined a group of twelve male pathological gamblers (ten strictly right handed, all smokers) aged 37.3 ± 7.4 yrs (mean ± sd) and a group of twelve, age matched, healthy males,
Functional topography of a distributed neural system for spatial and nonspatial information maintenance in working memory
 Neuropsychologia 41 (2003) 341 356 Functional topography of a distributed neural system for spatial and nonspatial information maintenance in working memory Joseph B. Sala a,, Pia Rämä a,c,d, Susan M.
Neuropsychologia 41 (2003) 341 356 Functional topography of a distributed neural system for spatial and nonspatial information maintenance in working memory Joseph B. Sala a,, Pia Rämä a,c,d, Susan M.
A possible mechanism for impaired joint attention in autism
 A possible mechanism for impaired joint attention in autism Justin H G Williams Morven McWhirr Gordon D Waiter Cambridge Sept 10 th 2010 Joint attention in autism Declarative and receptive aspects initiating
A possible mechanism for impaired joint attention in autism Justin H G Williams Morven McWhirr Gordon D Waiter Cambridge Sept 10 th 2010 Joint attention in autism Declarative and receptive aspects initiating
Comparing event-related and epoch analysis in blocked design fmri
 Available online at www.sciencedirect.com R NeuroImage 18 (2003) 806 810 www.elsevier.com/locate/ynimg Technical Note Comparing event-related and epoch analysis in blocked design fmri Andrea Mechelli,
Available online at www.sciencedirect.com R NeuroImage 18 (2003) 806 810 www.elsevier.com/locate/ynimg Technical Note Comparing event-related and epoch analysis in blocked design fmri Andrea Mechelli,
Experimental design for Cognitive fmri
 Experimental design for Cognitive fmri Alexa Morcom Edinburgh SPM course 2017 Thanks to Rik Henson, Thomas Wolbers, Jody Culham, and the SPM authors for slides Overview Categorical designs Factorial designs
Experimental design for Cognitive fmri Alexa Morcom Edinburgh SPM course 2017 Thanks to Rik Henson, Thomas Wolbers, Jody Culham, and the SPM authors for slides Overview Categorical designs Factorial designs
Brain Imaging Investigation of the Impairing Effect of Emotion on Cognition
 Brain Imaging Investigation of the Impairing Effect of Emotion on Cognition Gloria Wong 1,2, Sanda Dolcos 1,3, Ekaterina Denkova 1, Rajendra A. Morey 4,5, Lihong Wang 4,5, Nicholas Coupland 1, Gregory
Brain Imaging Investigation of the Impairing Effect of Emotion on Cognition Gloria Wong 1,2, Sanda Dolcos 1,3, Ekaterina Denkova 1, Rajendra A. Morey 4,5, Lihong Wang 4,5, Nicholas Coupland 1, Gregory
Effects Of Attention And Perceptual Uncertainty On Cerebellar Activity During Visual Motion Perception
 Effects Of Attention And Perceptual Uncertainty On Cerebellar Activity During Visual Motion Perception Oliver Baumann & Jason Mattingley Queensland Brain Institute The University of Queensland The Queensland
Effects Of Attention And Perceptual Uncertainty On Cerebellar Activity During Visual Motion Perception Oliver Baumann & Jason Mattingley Queensland Brain Institute The University of Queensland The Queensland
Visual Selection and Attention
 Visual Selection and Attention Retrieve Information Select what to observe No time to focus on every object Overt Selections Performed by eye movements Covert Selections Performed by visual attention 2
Visual Selection and Attention Retrieve Information Select what to observe No time to focus on every object Overt Selections Performed by eye movements Covert Selections Performed by visual attention 2
Functional Magnetic Resonance Imaging with Arterial Spin Labeling: Techniques and Potential Clinical and Research Applications
 pissn 2384-1095 eissn 2384-1109 imri 2017;21:91-96 https://doi.org/10.13104/imri.2017.21.2.91 Functional Magnetic Resonance Imaging with Arterial Spin Labeling: Techniques and Potential Clinical and Research
pissn 2384-1095 eissn 2384-1109 imri 2017;21:91-96 https://doi.org/10.13104/imri.2017.21.2.91 Functional Magnetic Resonance Imaging with Arterial Spin Labeling: Techniques and Potential Clinical and Research
Supporting Information
 Revisiting default mode network function in major depression: evidence for disrupted subsystem connectivity Fabio Sambataro 1,*, Nadine Wolf 2, Maria Pennuto 3, Nenad Vasic 4, Robert Christian Wolf 5,*
Revisiting default mode network function in major depression: evidence for disrupted subsystem connectivity Fabio Sambataro 1,*, Nadine Wolf 2, Maria Pennuto 3, Nenad Vasic 4, Robert Christian Wolf 5,*
Fusing Simultaneous EEG-fMRI by Linking Multivariate Classifiers
 Fusing Simultaneous EEG-fMRI by Linking Multivariate Classifiers Bryan R. Conroy 1, Jordan Muraskin 1, and Paul Sajda 1 Dept. of Biomedical Engineering, Columbia University, Columbia NY, USA Abstract.
Fusing Simultaneous EEG-fMRI by Linking Multivariate Classifiers Bryan R. Conroy 1, Jordan Muraskin 1, and Paul Sajda 1 Dept. of Biomedical Engineering, Columbia University, Columbia NY, USA Abstract.
Neuroimaging vs. other methods
 BASIC LOGIC OF NEUROIMAGING fmri (functional magnetic resonance imaging) Bottom line on how it works: Adapts MRI to register the magnetic properties of oxygenated and deoxygenated hemoglobin, allowing
BASIC LOGIC OF NEUROIMAGING fmri (functional magnetic resonance imaging) Bottom line on how it works: Adapts MRI to register the magnetic properties of oxygenated and deoxygenated hemoglobin, allowing
Training-Induced Functional Activation Changes in Dual-Task Processing: An fmri Study
 Cerebral Cortex January 2007;17:192-204 doi:10.1093/cercor/bhj137 Advance Access publication February 8, 2006 Training-Induced Functional Activation Changes in Dual-Task Processing: An fmri Study Kirk
Cerebral Cortex January 2007;17:192-204 doi:10.1093/cercor/bhj137 Advance Access publication February 8, 2006 Training-Induced Functional Activation Changes in Dual-Task Processing: An fmri Study Kirk
Activated Fibers: Fiber-centered Activation Detection in Task-based FMRI
 Activated Fibers: Fiber-centered Activation Detection in Task-based FMRI Jinglei Lv 1, Lei Guo 1, Kaiming Li 1,2, Xintao Hu 1, Dajiang Zhu 2, Junwei Han 1, Tianming Liu 2 1 School of Automation, Northwestern
Activated Fibers: Fiber-centered Activation Detection in Task-based FMRI Jinglei Lv 1, Lei Guo 1, Kaiming Li 1,2, Xintao Hu 1, Dajiang Zhu 2, Junwei Han 1, Tianming Liu 2 1 School of Automation, Northwestern
The role of cognitive effort in subjective reward devaluation and risky decision-making
 The role of cognitive effort in subjective reward devaluation and risky decision-making Matthew A J Apps 1,2, Laura Grima 2, Sanjay Manohar 2, Masud Husain 1,2 1 Nuffield Department of Clinical Neuroscience,
The role of cognitive effort in subjective reward devaluation and risky decision-making Matthew A J Apps 1,2, Laura Grima 2, Sanjay Manohar 2, Masud Husain 1,2 1 Nuffield Department of Clinical Neuroscience,
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and
 This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution
Data-driven Structured Noise Removal (FIX)
") Hamburg, June 8, 2014 Educational Course The Art and Pitfalls of fmri Preprocessing Data-driven Structured Noise Removal (FIX) Ludovica Griffanti! FMRIB Centre, University of Oxford, Oxford, United Kingdom
Hamburg, June 8, 2014 Educational Course The Art and Pitfalls of fmri Preprocessing Data-driven Structured Noise Removal (FIX) Ludovica Griffanti! FMRIB Centre, University of Oxford, Oxford, United Kingdom
Supporting Online Material for
 www.sciencemag.org/cgi/content/full/317/5835/215/dc1 Supporting Online Material for Prefrontal Regions Orchestrate Suppression of Emotional Memories via a Two- Phase Process Brendan E. Depue,* Tim Curran,
www.sciencemag.org/cgi/content/full/317/5835/215/dc1 Supporting Online Material for Prefrontal Regions Orchestrate Suppression of Emotional Memories via a Two- Phase Process Brendan E. Depue,* Tim Curran,
Supplemental Material
 1 Supplemental Material Golomb, J.D, and Kanwisher, N. (2012). Higher-level visual cortex represents retinotopic, not spatiotopic, object location. Cerebral Cortex. Contents: - Supplemental Figures S1-S3
1 Supplemental Material Golomb, J.D, and Kanwisher, N. (2012). Higher-level visual cortex represents retinotopic, not spatiotopic, object location. Cerebral Cortex. Contents: - Supplemental Figures S1-S3
The Neural Correlates of Moral Decision-Making in Psychopathy
 University of Pennsylvania ScholarlyCommons Neuroethics Publications Center for Neuroscience & Society 1-1-2009 The Neural Correlates of Moral Decision-Making in Psychopathy Andrea L. Glenn University
University of Pennsylvania ScholarlyCommons Neuroethics Publications Center for Neuroscience & Society 1-1-2009 The Neural Correlates of Moral Decision-Making in Psychopathy Andrea L. Glenn University
Functional Connectivity with Distinct Neural Networks Tracks Fluctuations in Gain/Loss Framing Susceptibility
 r Human Brain Mapping 00:00 00 (2015) r Functional Connectivity with Distinct Neural Networks Tracks Fluctuations in Gain/Loss Framing Susceptibility David V. Smith, 1 * Kamila E. Sip, 1,2 and Mauricio
r Human Brain Mapping 00:00 00 (2015) r Functional Connectivity with Distinct Neural Networks Tracks Fluctuations in Gain/Loss Framing Susceptibility David V. Smith, 1 * Kamila E. Sip, 1,2 and Mauricio
Behavioural Brain Research
 Behavioural Brain Research 197 (2009) 186 197 Contents lists available at ScienceDirect Behavioural Brain Research j o u r n a l h o m e p a g e : www.elsevier.com/locate/bbr Research report Top-down attentional
Behavioural Brain Research 197 (2009) 186 197 Contents lists available at ScienceDirect Behavioural Brain Research j o u r n a l h o m e p a g e : www.elsevier.com/locate/bbr Research report Top-down attentional
Category: Life sciences Name: Seul Lee SUNetID: seul
 Independent Component Analysis (ICA) of functional MRI (fmri) data Category: Life sciences Name: Seul Lee SUNetID: seul 05809185 Introduction Functional Magnetic Resonance Imaging (fmri) is an MRI method
Independent Component Analysis (ICA) of functional MRI (fmri) data Category: Life sciences Name: Seul Lee SUNetID: seul 05809185 Introduction Functional Magnetic Resonance Imaging (fmri) is an MRI method
SUPPLEMENTARY INFORMATION. Predicting visual stimuli based on activity in auditory cortices
 SUPPLEMENTARY INFORMATION Predicting visual stimuli based on activity in auditory cortices Kaspar Meyer, Jonas T. Kaplan, Ryan Essex, Cecelia Webber, Hanna Damasio & Antonio Damasio Brain and Creativity
SUPPLEMENTARY INFORMATION Predicting visual stimuli based on activity in auditory cortices Kaspar Meyer, Jonas T. Kaplan, Ryan Essex, Cecelia Webber, Hanna Damasio & Antonio Damasio Brain and Creativity
DATA MANAGEMENT & TYPES OF ANALYSES OFTEN USED. Dennis L. Molfese University of Nebraska - Lincoln
 DATA MANAGEMENT & TYPES OF ANALYSES OFTEN USED Dennis L. Molfese University of Nebraska - Lincoln 1 DATA MANAGEMENT Backups Storage Identification Analyses 2 Data Analysis Pre-processing Statistical Analysis
DATA MANAGEMENT & TYPES OF ANALYSES OFTEN USED Dennis L. Molfese University of Nebraska - Lincoln 1 DATA MANAGEMENT Backups Storage Identification Analyses 2 Data Analysis Pre-processing Statistical Analysis
Reporting Checklist for Nature Neuroscience
 Corresponding Author: Manuscript Number: Manuscript Type: Simon Musall NNA47695 Article Reporting Checklist for Nature Neuroscience # Main Figures: 6 # Supplementary Figures: 14 # Supplementary Tables:
Corresponding Author: Manuscript Number: Manuscript Type: Simon Musall NNA47695 Article Reporting Checklist for Nature Neuroscience # Main Figures: 6 # Supplementary Figures: 14 # Supplementary Tables:
Statistical parametric mapping
 350 PRACTICAL NEUROLOGY HOW TO UNDERSTAND IT Statistical parametric mapping Geraint Rees Wellcome Senior Clinical Fellow,Institute of Cognitive Neuroscience & Institute of Neurology, University College
350 PRACTICAL NEUROLOGY HOW TO UNDERSTAND IT Statistical parametric mapping Geraint Rees Wellcome Senior Clinical Fellow,Institute of Cognitive Neuroscience & Institute of Neurology, University College
Supplementary Online Content
 Supplementary Online Content Redlich R, Opel N, Grotegerd D, et al. Prediction of individual response to electroconvulsive therapy via machine learning on structural magnetic resonance imaging data. JAMA
Supplementary Online Content Redlich R, Opel N, Grotegerd D, et al. Prediction of individual response to electroconvulsive therapy via machine learning on structural magnetic resonance imaging data. JAMA
Overview. Connectivity detection
 Overview Introduction Functional connectivity explorations Dynamic change Effect of motor fatigue Neurological change Effect of Asperger s Disorder, Depression Consciousness change Effect of anesthesia
Overview Introduction Functional connectivity explorations Dynamic change Effect of motor fatigue Neurological change Effect of Asperger s Disorder, Depression Consciousness change Effect of anesthesia
SUPPLEMENTARY INFORMATION. Table 1 Patient characteristics Preoperative. language testing
 Categorical Speech Representation in the Human Superior Temporal Gyrus Edward F. Chang, Jochem W. Rieger, Keith D. Johnson, Mitchel S. Berger, Nicholas M. Barbaro, Robert T. Knight SUPPLEMENTARY INFORMATION
Categorical Speech Representation in the Human Superior Temporal Gyrus Edward F. Chang, Jochem W. Rieger, Keith D. Johnson, Mitchel S. Berger, Nicholas M. Barbaro, Robert T. Knight SUPPLEMENTARY INFORMATION
Introduction to Computational Neuroscience
 Introduction to Computational Neuroscience Lecture 5: Data analysis II Lesson Title 1 Introduction 2 Structure and Function of the NS 3 Windows to the Brain 4 Data analysis 5 Data analysis II 6 Single
Introduction to Computational Neuroscience Lecture 5: Data analysis II Lesson Title 1 Introduction 2 Structure and Function of the NS 3 Windows to the Brain 4 Data analysis 5 Data analysis II 6 Single
HUMAN SOCIAL INTERACTION RESEARCH PROPOSAL C8CSNR
 HUMAN SOCIAL INTERACTION RESEARCH PROPOSAL C8CSNR Applicants Principal Investigator Student ID 4039921 Collaborators Name(s) Institution(s) Title of project: Neural basis of verbal and non-verbal false
HUMAN SOCIAL INTERACTION RESEARCH PROPOSAL C8CSNR Applicants Principal Investigator Student ID 4039921 Collaborators Name(s) Institution(s) Title of project: Neural basis of verbal and non-verbal false
Sum of Neurally Distinct Stimulus- and Task-Related Components.
 SUPPLEMENTARY MATERIAL for Cardoso et al. 22 The Neuroimaging Signal is a Linear Sum of Neurally Distinct Stimulus- and Task-Related Components. : Appendix: Homogeneous Linear ( Null ) and Modified Linear
SUPPLEMENTARY MATERIAL for Cardoso et al. 22 The Neuroimaging Signal is a Linear Sum of Neurally Distinct Stimulus- and Task-Related Components. : Appendix: Homogeneous Linear ( Null ) and Modified Linear
FREQUENCY DOMAIN HYBRID INDEPENDENT COMPONENT ANALYSIS OF FUNCTIONAL MAGNETIC RESONANCE IMAGING DATA
 FREQUENCY DOMAIN HYBRID INDEPENDENT COMPONENT ANALYSIS OF FUNCTIONAL MAGNETIC RESONANCE IMAGING DATA J.D. Carew, V.M. Haughton, C.H. Moritz, B.P. Rogers, E.V. Nordheim, and M.E. Meyerand Departments of
FREQUENCY DOMAIN HYBRID INDEPENDENT COMPONENT ANALYSIS OF FUNCTIONAL MAGNETIC RESONANCE IMAGING DATA J.D. Carew, V.M. Haughton, C.H. Moritz, B.P. Rogers, E.V. Nordheim, and M.E. Meyerand Departments of
An Introduction to Translational Neuroscience Approaches to Investigating Autism
 An Introduction to Translational Neuroscience Approaches to Investigating Autism Gabriel S. Dichter, PhD Departments of Psychiatry & Psychology, UNC-Chapel Hill Carolina Institute for Developmental Disabilities
An Introduction to Translational Neuroscience Approaches to Investigating Autism Gabriel S. Dichter, PhD Departments of Psychiatry & Psychology, UNC-Chapel Hill Carolina Institute for Developmental Disabilities
Supplementary Information on TMS/hd-EEG recordings: acquisition and preprocessing
 Supplementary Information on TMS/hd-EEG recordings: acquisition and preprocessing Stability of the coil position was assured by using a software aiming device allowing the stimulation only when the deviation
Supplementary Information on TMS/hd-EEG recordings: acquisition and preprocessing Stability of the coil position was assured by using a software aiming device allowing the stimulation only when the deviation
Supplemental Data. Inclusion/exclusion criteria for major depressive disorder group and healthy control group
 1 Supplemental Data Inclusion/exclusion criteria for major depressive disorder group and healthy control group Additional inclusion criteria for the major depressive disorder group were: age of onset of
1 Supplemental Data Inclusion/exclusion criteria for major depressive disorder group and healthy control group Additional inclusion criteria for the major depressive disorder group were: age of onset of
QUANTIFYING CEREBRAL CONTRIBUTIONS TO PAIN 1
 QUANTIFYING CEREBRAL CONTRIBUTIONS TO PAIN 1 Supplementary Figure 1. Overview of the SIIPS1 development. The development of the SIIPS1 consisted of individual- and group-level analysis steps. 1) Individual-person
QUANTIFYING CEREBRAL CONTRIBUTIONS TO PAIN 1 Supplementary Figure 1. Overview of the SIIPS1 development. The development of the SIIPS1 consisted of individual- and group-level analysis steps. 1) Individual-person
Attention Response Functions: Characterizing Brain Areas Using fmri Activation during Parametric Variations of Attentional Load
 Attention Response Functions: Characterizing Brain Areas Using fmri Activation during Parametric Variations of Attentional Load Intro Examine attention response functions Compare an attention-demanding
Attention Response Functions: Characterizing Brain Areas Using fmri Activation during Parametric Variations of Attentional Load Intro Examine attention response functions Compare an attention-demanding
5/2/2013. Real-time fmri: Methods and applications
 Real-time fmri: Methods and applications Frank Scharnowski Non-invasive imaging technique PET Magnetic Resonance Imaging (MRI) NIRS EEG MEG Functional MRI Structural MRI Perfusion MRI Diffusion MRI MR
Real-time fmri: Methods and applications Frank Scharnowski Non-invasive imaging technique PET Magnetic Resonance Imaging (MRI) NIRS EEG MEG Functional MRI Structural MRI Perfusion MRI Diffusion MRI MR
International Journal of Innovative Research in Advanced Engineering (IJIRAE) Volume 1 Issue 10 (November 2014)
 Volume 1 Issue 10 (November 2014)") Technique for Suppression Random and Physiological Noise Components in Functional Magnetic Resonance Imaging Data Mawia Ahmed Hassan* Biomedical Engineering Department, Sudan University of Science & Technology,
Technique for Suppression Random and Physiological Noise Components in Functional Magnetic Resonance Imaging Data Mawia Ahmed Hassan* Biomedical Engineering Department, Sudan University of Science & Technology,
Identification of Neuroimaging Biomarkers
 Identification of Neuroimaging Biomarkers Dan Goodwin, Tom Bleymaier, Shipra Bhal Advisor: Dr. Amit Etkin M.D./PhD, Stanford Psychiatry Department Abstract We present a supervised learning approach to
Identification of Neuroimaging Biomarkers Dan Goodwin, Tom Bleymaier, Shipra Bhal Advisor: Dr. Amit Etkin M.D./PhD, Stanford Psychiatry Department Abstract We present a supervised learning approach to
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature11239 Introduction The first Supplementary Figure shows additional regions of fmri activation evoked by the task. The second, sixth, and eighth shows an alternative way of analyzing reaction
doi:10.1038/nature11239 Introduction The first Supplementary Figure shows additional regions of fmri activation evoked by the task. The second, sixth, and eighth shows an alternative way of analyzing reaction
Combining tdcs and fmri. OHMB Teaching Course, Hamburg June 8, Andrea Antal
 Andrea Antal Department of Clinical Neurophysiology Georg-August University Goettingen Combining tdcs and fmri OHMB Teaching Course, Hamburg June 8, 2014 Classical Biomarkers for measuring human neuroplasticity
Andrea Antal Department of Clinical Neurophysiology Georg-August University Goettingen Combining tdcs and fmri OHMB Teaching Course, Hamburg June 8, 2014 Classical Biomarkers for measuring human neuroplasticity
Experimental design. Experimental design. Experimental design. Guido van Wingen Department of Psychiatry Academic Medical Center
 Experimental design Guido van Wingen Department of Psychiatry Academic Medical Center guidovanwingen@gmail.com Experimental design Experimental design Image time-series Spatial filter Design matrix Statistical
Experimental design Guido van Wingen Department of Psychiatry Academic Medical Center guidovanwingen@gmail.com Experimental design Experimental design Image time-series Spatial filter Design matrix Statistical
Supplementary Information
 Supplementary Information The neural correlates of subjective value during intertemporal choice Joseph W. Kable and Paul W. Glimcher a 10 0 b 10 0 10 1 10 1 Discount rate k 10 2 Discount rate k 10 2 10
Supplementary Information The neural correlates of subjective value during intertemporal choice Joseph W. Kable and Paul W. Glimcher a 10 0 b 10 0 10 1 10 1 Discount rate k 10 2 Discount rate k 10 2 10
Selective bias in temporal bisection task by number exposition
 Selective bias in temporal bisection task by number exposition Carmelo M. Vicario¹ ¹ Dipartimento di Psicologia, Università Roma la Sapienza, via dei Marsi 78, Roma, Italy Key words: number- time- spatial
Selective bias in temporal bisection task by number exposition Carmelo M. Vicario¹ ¹ Dipartimento di Psicologia, Università Roma la Sapienza, via dei Marsi 78, Roma, Italy Key words: number- time- spatial
CS/NEUR125 Brains, Minds, and Machines. Due: Friday, April 14
 CS/NEUR125 Brains, Minds, and Machines Assignment 5: Neural mechanisms of object-based attention Due: Friday, April 14 This Assignment is a guided reading of the 2014 paper, Neural Mechanisms of Object-Based
CS/NEUR125 Brains, Minds, and Machines Assignment 5: Neural mechanisms of object-based attention Due: Friday, April 14 This Assignment is a guided reading of the 2014 paper, Neural Mechanisms of Object-Based
Voxel-based Lesion-Symptom Mapping. Céline R. Gillebert
 Voxel-based Lesion-Symptom Mapping Céline R. Gillebert Paul Broca (1861) Mr. Tan no productive speech single repetitive syllable tan Broca s area: speech production Broca s aphasia: problems with fluency,
Voxel-based Lesion-Symptom Mapping Céline R. Gillebert Paul Broca (1861) Mr. Tan no productive speech single repetitive syllable tan Broca s area: speech production Broca s aphasia: problems with fluency,
Source Memory is not Properly Differentiated from Object Memory in Early Schizophrenia An fmri study using virtual reality
 Source Memory is not Properly Differentiated from Object Memory in Early Schizophrenia An fmri study using virtual reality Colin S. Hawco, PhD Lisa Buchy Michael Bodnar, Sarah Izadi, Jennifer Dell Elce,
Source Memory is not Properly Differentiated from Object Memory in Early Schizophrenia An fmri study using virtual reality Colin S. Hawco, PhD Lisa Buchy Michael Bodnar, Sarah Izadi, Jennifer Dell Elce,
Experimental Design I
 Experimental Design I Topics What questions can we ask (intelligently) in fmri Basic assumptions in isolating cognitive processes and comparing conditions General design strategies A few really cool experiments
Experimental Design I Topics What questions can we ask (intelligently) in fmri Basic assumptions in isolating cognitive processes and comparing conditions General design strategies A few really cool experiments
The Neural Correlates of Subjective Utility of Monetary Outcome and Probability Weight in Economic and in Motor Decision under Risk
 8822 The Journal of Neuroscience, June 15, 2011 31(24):8822 8831 Behavioral/Systems/Cognitive The Neural Correlates of Subjective Utility of Monetary Outcome and Probability Weight in Economic and in Motor
8822 The Journal of Neuroscience, June 15, 2011 31(24):8822 8831 Behavioral/Systems/Cognitive The Neural Correlates of Subjective Utility of Monetary Outcome and Probability Weight in Economic and in Motor
An introduction to functional MRI
 An introduction to functional MRI Bianca de Haan and Chris Rorden Introduction...1 1 The basics of MRI...1 1.1 The brain in a magnetic field...1 1.2 Application of the radiofrequency pulse...3 1.3 Relaxation...4
An introduction to functional MRI Bianca de Haan and Chris Rorden Introduction...1 1 The basics of MRI...1 1.1 The brain in a magnetic field...1 1.2 Application of the radiofrequency pulse...3 1.3 Relaxation...4
SUPPLEMENTARY METHODS. Subjects and Confederates. We investigated a total of 32 healthy adult volunteers, 16
 SUPPLEMENTARY METHODS Subjects and Confederates. We investigated a total of 32 healthy adult volunteers, 16 women and 16 men. One female had to be excluded from brain data analyses because of strong movement
SUPPLEMENTARY METHODS Subjects and Confederates. We investigated a total of 32 healthy adult volunteers, 16 women and 16 men. One female had to be excluded from brain data analyses because of strong movement
Spatiotemporal clustering of synchronized bursting events in neuronal networks
 Spatiotemporal clustering of synchronized bursting events in neuronal networks Uri Barkan a David Horn a,1 a School of Physics and Astronomy, Tel Aviv University, Tel Aviv 69978, Israel Abstract in vitro
Spatiotemporal clustering of synchronized bursting events in neuronal networks Uri Barkan a David Horn a,1 a School of Physics and Astronomy, Tel Aviv University, Tel Aviv 69978, Israel Abstract in vitro
Virtual Reality Testing of Multi-Modal Integration in Schizophrenic Patients
 Virtual Reality Testing of Multi-Modal Integration in Schizophrenic Patients Anna SORKIN¹, Avi PELED 2, Daphna WEINSHALL¹ 1 Interdisciplinary Center for Neural Computation, Hebrew University of Jerusalem,
Virtual Reality Testing of Multi-Modal Integration in Schizophrenic Patients Anna SORKIN¹, Avi PELED 2, Daphna WEINSHALL¹ 1 Interdisciplinary Center for Neural Computation, Hebrew University of Jerusalem,
NIH Public Access Author Manuscript Neuroimage. Author manuscript; available in PMC 2011 February 1.
 NIH Public Access Author Manuscript Published in final edited form as: Neuroimage. 2010 February 1; 49(3): 2163 2177. doi:10.1016/j.neuroimage.2009.10.080. Reliable Intrinsic Connectivity Networks: Test-Retest
NIH Public Access Author Manuscript Published in final edited form as: Neuroimage. 2010 February 1; 49(3): 2163 2177. doi:10.1016/j.neuroimage.2009.10.080. Reliable Intrinsic Connectivity Networks: Test-Retest
What do you think of the following research? I m interested in whether a low glycemic index diet gives better control of diabetes than a high
 What do you think of the following research? I m interested in whether a low glycemic index diet gives better control of diabetes than a high glycemic index diet. So I randomly assign 100 people with type
What do you think of the following research? I m interested in whether a low glycemic index diet gives better control of diabetes than a high glycemic index diet. So I randomly assign 100 people with type
Data Visualization for MRI
 Data Visualization for MRI Cyril Pernet Centre for Clinical Brain Sciences (CCBS) Neuroimaging Sciences Edinburgh Biennial SPM course 2019 @CyrilRPernet What to visualize? Brain and tissues / Brains and
Data Visualization for MRI Cyril Pernet Centre for Clinical Brain Sciences (CCBS) Neuroimaging Sciences Edinburgh Biennial SPM course 2019 @CyrilRPernet What to visualize? Brain and tissues / Brains and
Supplementary Online Content
 Supplementary Online Content Gregg NM, Kim AE, Gurol ME, et al. Incidental cerebral microbleeds and cerebral blood flow in elderly individuals. JAMA Neurol. Published online July 13, 2015. doi:10.1001/jamaneurol.2015.1359.
Supplementary Online Content Gregg NM, Kim AE, Gurol ME, et al. Incidental cerebral microbleeds and cerebral blood flow in elderly individuals. JAMA Neurol. Published online July 13, 2015. doi:10.1001/jamaneurol.2015.1359.
Tools of the trade: psychophysiological interactions and functional connectivity
 doi:10.1093/scan/nss055 Tools of the trade: psychophysiological interactions and functional connectivity SCAN (2012) 7, 604^609 Jill X. O Reilly, 1 Mark W. Woolrich, 1,2 Timothy E.J. Behrens, 1,3 Stephen
doi:10.1093/scan/nss055 Tools of the trade: psychophysiological interactions and functional connectivity SCAN (2012) 7, 604^609 Jill X. O Reilly, 1 Mark W. Woolrich, 1,2 Timothy E.J. Behrens, 1,3 Stephen
The effects of single-trial averaging upon the spatial extent of fmri activation
 BRAIN IMAGING NEUROREPORT The effects of single-trial averaging upon the spatial extent of fmri activation Scott A. Huettel,CA and Gregory McCarthy, Brain Imaging and Analysis Center, Duke University Medical
BRAIN IMAGING NEUROREPORT The effects of single-trial averaging upon the spatial extent of fmri activation Scott A. Huettel,CA and Gregory McCarthy, Brain Imaging and Analysis Center, Duke University Medical
Supplementary Online Content
 1 Supplementary Online Content Miller CH, Hamilton JP, Sacchet MD, Gotlib IH. Meta-analysis of functional neuroimaging of major depressive disorder in youth. JAMA Psychiatry. Published online September
1 Supplementary Online Content Miller CH, Hamilton JP, Sacchet MD, Gotlib IH. Meta-analysis of functional neuroimaging of major depressive disorder in youth. JAMA Psychiatry. Published online September
Reporting Checklist for Nature Neuroscience
 Corresponding Author: Manuscript Number: Manuscript Type: Alex Pouget NN-A46249B Article Reporting Checklist for Nature Neuroscience # Main Figures: 7 # Supplementary Figures: 3 # Supplementary Tables:
Corresponding Author: Manuscript Number: Manuscript Type: Alex Pouget NN-A46249B Article Reporting Checklist for Nature Neuroscience # Main Figures: 7 # Supplementary Figures: 3 # Supplementary Tables:
Distinguishing informational from value-related encoding of rewarding and punishing outcomes in the human brain
 European Journal of Neuroscience, Vol. 39, pp. 2014 2026, 2014 doi:10.1111/ejn.12625 Distinguishing informational from value-related encoding of rewarding and punishing outcomes in the human brain Ryan
European Journal of Neuroscience, Vol. 39, pp. 2014 2026, 2014 doi:10.1111/ejn.12625 Distinguishing informational from value-related encoding of rewarding and punishing outcomes in the human brain Ryan