Characterization of drug-resistant influenza A(H1N1) and A(H3N2) variants selected in vitro with laninamivir.
|
|
|
- Zoe Sherman
- 5 years ago
- Views:
Transcription
1 AAC Accepts, published online ahead of print on 23 June 2014 Antimicrob. Agents Chemother. doi: /aac Copyright 2014, American Society for Microbiology. All Rights Reserved. 1 2 Characterization of drug-resistant influenza A(H1N1) and A(H3N2) variants selected in vitro with laninamivir Mélanie Samson 1, Yacine Abed 1, François-Marc Desrochers 1, Stephanie Hamilton 2, Angela Luttick 2, Simon P. Tucker 2, Melinda J. Pryor 2 and Guy Boivin 1#. 1 Research Center in Infectious Diseases of the CHU de Québec-CHUL and Laval University, Québec City, Québec, Canada. 2 Biota Scientific Management, 10/585 Blackburn Road, Notting Hill, Australia. Running title: Laninamivir-resistant influenza A viruses. Abstract word count: 249 Text word count: 3702 Inserts: 4 tables and 3 figures #Corresponding author: Guy Boivin, MD CHUL, room RC blvd Laurier, Sainte-Foy, Québec, Canada G1V 4G2 Tel: (418) Fax: (418) Guy.Boivin@crchul.ulaval.ca 1
2 ABSTRACT Neuraminidase inhibitors (NAIs) play a major role for managing influenza infections. The widespread oseltamivir resistance among seasonal A(H1N1) viruses and community outbreaks of oseltamivir-resistant A(H1N1)pdm09 strains highlight the need for additional anti-influenza agents. Laninamivir is a novel long-lasting NAI that has demonstrated in vitro activity against influenza A and B viruses and its prodrug (laninamivir octanoate) is in phase II clinical trials in US and other countries. Currently, little information is available on the mechanisms of resistance to laninamivir. In this study, we first performed neuraminidase (NA) inhibition assays to determine the activity of laninamivir against a set of influenza A viruses containing NA mutations conferring resistance to one or many other NAIs. We also generated drug-resistant A(H1N1) and A(H3N2) viruses under in vitro laninamivir pressure. Laninamivir demonstrated a profile of susceptibility that was similar to that of zanamivir. More specifically, it retained activity against oseltamivir-resistant H275Y and N295S A(H1N1) mutantsand the E119V A(H3N2) variant. In vitro, laninamivir pressure selected the E119A NA substitution in the A/Solomon Islands/3/2006 A(H1N1) background whereas E119K and G147E NA changes along with a K133E HA substitution were selected in the A/Quebec/144147/2009 A(H1N1)pdm09 strain. In the A/Brisbane/10/2007 A(H3N2) background, a large NA deletion accompanied with S138A/P194L HA substitutions were selected. This H3N2 variant had altered receptor-binding properties and was highly resistant to laninamivir in plaque reduction assays. Overall, we confirmed the similarity between zanamivir and laninamivir susceptibility profiles and demonstrated that both NA and HA changes can contribute to laninamivir resistance in vitro. Keywords: Influenza, antivirals, neuraminidase inhibitors, laninamivir, resistance. 2
3 INTRODUCTION Influenza is a highly transmissible viral infection associated with serious public health and economic problems. Each year, various strains of influenza viruses circulate throughout the world and cause significant morbidity in the general population as well as substantial mortality among high-risk individuals. Among the 18 hemagglutinin (HA) and 11 neuraminidase (NA) subtypes, influenza A(H1N1) and A(H3N2) strains have been predominantly associated with epidemics during the last century. In addition to immunization programs, antiviral agents constitute an important means in the management of seasonal influenza and play a major role as a first-line defense in the case of pandemics. Two neuraminidase inhibitors (NAIs) are currently licensed worldwide for treatment and prophylaxis of influenza infections: oseltamivir phosphate (Tamiflu; Hoffmann-La Roche) and zanamivir (Relenza; GlaxoSmithKline). Oseltamivir is the most widely used and stockpiled NAI due to its good bioavailability as an oral preparation (1-3). However, recent emergence of oseltamivir-resistant variants, especially among A(H1N1) viruses (4-6), is a matter of great concern. Oseltamivir has a large hydrophobic side chain requiring a conformational rearrangement in the viral NA that is essential to accommodate the drug. Any mutations that affect this rearrangement may reduce the binding affinity of oseltamivir leading to viral resistance (7, 8). Notably, the H275Y mutation (N1 numbering), that is most commonly associated with oseltamivir resistance, has been shown to inhibit such rearrangement (9). In addition, influenza viruses of the N1 subtype containing the H275Y mutation, including A(H1N1) and A(H5N1) strains, are also resistant to peramivir, a cyclopentane NAI whose intravenous formulation is approved in a 3
4 few countries (10). Fortunately, most oseltamivir-resistant strains exhibit susceptibility to zanamivir, a drug that is administered by an inhaler device (Diskhaler ). On the other hand, zanamivir must be administered twice daily over 5 consecutive days for optimal benefits (11). Therefore, there is a need to develop new inhibitors that possess long term half-life and that are also effective against oseltamivir-resistant influenza viruses. Laninamivir (R ) is a long-acting NAI which demonstrated a broad range of activity against influenza A (N1 to N9) and influenza B strains (12, 13). Laninamivir octanoate (CS- 8958), the octanoyl prodrug of laninamivir, is commercially available in Japan under the name of Inavir (Daiichi Sankyo). As for zanamivir, laninamivir octanoate is formulated as a dry powder that has to be administered with a specific inhaler device. Importantly, a single nasal administration is associated with prolonged retention in the lungs, which confers a longacting anti-neuraminidase effect (12) comparable to oseltamivir twice daily for 5 days (14, 15). Additionally, laninamivir was shown to be active against oseltamivir-resistant A(H5N1) and A(H1N1) variants containing the H275Y NA mutation (16). For these reasons, laninamivir octanoate offers advantages over both oseltamivir and zanamivir. To date, no clinical cases of laninamivir-resistant influenza strains have been reported. Moreover, there are currently no data on the in vitro selection of viral mutations conferring resistance to laninamivir. Nevertheless, as for other NAIs, the emergence of laninamivir resistance should be considered. The objectives of the present study were, firstly, to evaluate the activity of laninamivir against a collection of NAI-resistant seasonal A(H1N1), A(H3N2) and 2009 pandemic A(H1N1)pdm09 viruses. Secondly, we aimed to generate and 4
5 98 99 characterize laninamivir-resistant influenza A(H1N1) and A(H3N2) variants following in vitro passaging under laninamivir pressure MATERIALS AND METHODS Cells culture. ST6Gal1 Madin-Darby canine kidney cells, overexpressing the α2,6 sialic acid receptors (MDCK α2,6; kindly provided by Y. Kawaoka from the University of Wisconsin, Madison, WI) and human embryonic kidney 293T cells (ATCC) were maintained in Dulbecco s modified Eagle s medium (DMEM) (Invitrogen, Carlsbad, CA), supplemented with 10% fetal bovine serum (Invitrogen, Carlsbad, CA). Madin-Darby canine kidney (MDCK) cells were sourced from the European Collection of Cell Cultures (ECACC, Wiltshire, United Kingdom). These cells were maintained to generate cell bank stocks in Minimal Essential Medium without L-glutamine (Invitrogen, Carlsbad, CA) supplemented with 10% Fetal Bovine Serum (Invitrogen, Carlsbad, CA). Drug susceptibility testing. A selection of seasonal A(H1N1), A(H3N2) and A(H1N1)pdm09 viruses harboring NAIresistant NA mutations (Table 1) were used for assessing susceptibility to laninamivir (R ) (Biota Scientific Managament, Notting Hill, Australia), oseltamivir carboxylate (Hoffmann-La Roche, Basel, Switzerland), zanamivir (GlaxoSmithKline, Stevenage, United Kingdom) and peramivir (BioCryst, Birmingham, United States) by NA inhibition assays as previously described (17) with minor modifications. Briefly, viruses were standardized to a NA activity level 10-fold higher than that of the background, as measured by the production 5
6 of a fluorescent product from the 2 -(4-methylumbelliferyl)-α-D-N-acetylneuraminic acid (MUNANA; Sigma, St-Louis, MO) substrate. Drug susceptibility profiles were determined by the extent of NA inhibition after incubation with 3-fold serial dilutions of NAIs at final concentrations ranging between 0 and 10,800 nm. The 50% inhibitory concentrations (IC 50 s) were determined from the dose-response curve. Some drug-selected viruses were also assessed for their susceptibility to laninamivir and oseltamivir by using a plaque reduction assay. In Method 1 (A/Quebec/144147/2009 and A/Brisbane/10/2007 influenza viruses), confluent monolayers of MDCK α2,6 cells were grown in 12-well dishes and infected with a dilution of virus required to obtain plaques per well. After an hour incubation step at 37 C with 5% CO 2, unbound viruses were removed and cells were overlaid with 0.8% agarose-containing maintenance medium in the presence of TPCK-treated trypsin and different concentrations of NAIs. Three days later, the mean viral plaque areas were determined from a minimum of 15 plaques using the ImageJ software (version 1.41, developed by Wayne Rasband of the National Institutes of Health). The IC 50 value was defined as the drug concentration resulting in the reduction of viral plaque areas by 50%. In method 2 (A/Auckland/3/2009 and A/Solomon Islands/3/2006 influenza viruses), confluent monolayers of MDCK cells were grown in six-well plates and infected with a dilution of virus required to obtain plaques per well. After an hour incubation step at 37 C with 5% CO 2, unbound viruses were removed and cells were overlaid with 0.5% seakem agarose-containing maintenance medium in the presence of TPCK-treated trypsin and 6
7 different concentrations of NAIs. Three days later, plaque diameter measurements were determined using Adobe Photoshop software CS3 (version 10.01,). The IC 50 value was defined as the drug concentration that results in a 50% reduction in influenza induced plaque diameter. Selection of drug-resistant viruses in vitro. Initial passages of A(H1N1)pdm09 [A/Quebec/144147/2009], which is an A/California/7/2009-like isolate, and A(H3N2) [A/Brisbane/10/2007-like] influenza viruses were performed by infecting MDCK α2,6 cells at a multiplicity of infection (MOI) of 0.01 in presence of 50 nm of laninamivir. On the other hand, passages of A(H1N1)pdm09 [A/Auckland/3/2009], another A/California/7/2009-like isolate, and A(H1N1) [A/Solomon Islands/3/2006-like] influenza viruses were performed by infecting MDCK cells at a MOI of 0.05 in presence of 48 nm and 8 nm of laninamivir, respectively. The drug concentration was then slowly increased with some passages being performed at the same level to increase viral replication. Control viruses were also passaged concomitantly in the absence of drug. At passages 3, 6, 9 and 12 for A/Quebec/144147/2009; 1-5 for A/Auckland/3/2009; 1-11 for A/Solomon Islands/3/2006-like and 3, 6 and 9 for A/Brisbane/10/2007-like viruses, viral RNA was isolated from supernatants of infected cell cultures and reverse-transcribed before PCR amplification of the entire HA and NA genes. PCR products were purified and sequenced using an Applied Biosystems 3730xl DNA Analyzer (Life Technologies Corporation, Carlsbad, CA). Generation of recombinant viruses and proteins. 7
8 The pllb plasmid containing the A/Quebec/144147/2009 NA gene was used for the introduction of 3 single substitutions (N1 numbering: E119A, E119K, G147E) using appropriate primers and the QuikChange site-directed mutagenesis kit (Stratagene, La Jolla, USA). Similarly, the plasmid containing the HA gene was used for the introduction of 3 single substitutions (H3 numbering: K133E, K156E, D225G). We used the plasmid pllb- NA E119K to introduce the second substitution G147E and the plasmid pllb-ha K156E to introduce the second substitution D225G. The 8 plasmids were co-transfected into 293T cells by using the Lipofectamine 2000 reagent (Invitrogen, Carlsbad, CA) as previously described (18). Supernatants were collected at 72 h post-transfection and used to inoculate MDCK α2,6 cells. The resulting recombinant wild-type (WT) as well as mutant viruses were subsequently sequenced and titrated by plaque assays in MDCK α2,6 cells. The pcaggs-pa, -PB1, -PB2 and -NP expression plasmids and the pllb plasmid containing the WT or variant (E119A, E119K, G147E and E119K/G147E) NA were used to co-transfect 293T cells in order to express recombinant NA proteins (19). Forty-eight h after transfection, the cells were briefly treated with trypsin-edta and neutralized by the addition of serum followed by centrifugation at 3,000 RPM for 5 min. After washing twice with PBS, the cells were resuspended in PBS containing 3.5 mm CaCl2 and used in an NA assay using the MUNANA substrate. Hemagglutination and hemagglutination-elution assays. Serial two-fold dilutions of viruses were prepared in 50 µl of PBS in round-bottomed 96-well plates to which 50 µl of 0.7% suspension of erythrocytes (from guinea pigs and turkeys 8
9 expressing predominantly α2,6 and α2,3 receptors, respectively) were added. Plates were incubated at 4 C for 1 h and HA titers, given in hemagglutinating units (HAU), were determined. HA-elution assays were performed as described elsewhere (20). Briefly, 4 HAU of viruses were pre-incubated for 30 min at room temperature with 1 µm of NAIs. Guinea pig or turkey erythrocytes were then added and the plates were incubated at 4 C for 1 h to allow agglutination to occur. The plates were then incubated at 37 C to allow virus elution. Elution was followed by the appearance of pelleted erythrocytes. One well contained the virus in the absence of drug as the elution control. The plates were monitored at 15, 30, 45, 60, 90, 120 and 240 min to determine the rate at which elution occurred. Statistical analyses Amounts of NA activity of recombinant proteins were compared to those of the WT by the use of unpaired two-tailed t tests. RESULTS Laninamivir susceptibility profiles of influenza viruses harboring mutations of resistance to other NAIs. The IC 50 values of laninamivir against various NAI-resistant influenza A variants as determined by NA inhibition assays are summarized in Table 1. All viruses that were susceptible to zanamivir also had a susceptible phenotype to laninamivir, including oseltamivir-resistant A(H1N1) variants containing H275Y and N295S substitutions as well as the A(H3N2) variant with the E119V change. Influenza A(H1N1)pdm09 variants containing the E119V/G and Q136K substitutions, which conferred resistance to zanamivir, exhibited 9
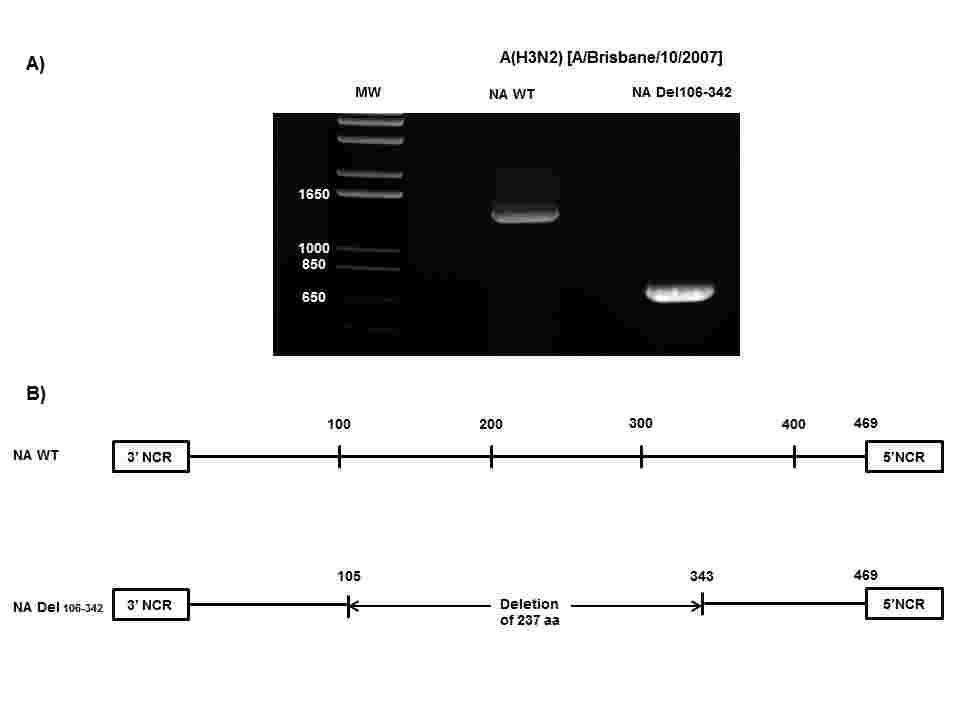
10 reduced or highly reduced inhibition to laninamivir. Of note, a multidrug resistance phenotype to laninamivir, zanamivir, peramivir and oseltamivir was observed for the E119V A(H1N1)pdm09 recombinant variant Selection of laninamivir-resistant variants in vitro. Pandemic influenza A(H1N1)pdm09 [A/Quebec/144147/2009 and A/Auckland/3/2009] strains as well as seasonal A(H1N1) [A/Solomon Islands/3/2006-like] and A(H3N2) [A/Brisbane/10/2007-like] viruses were passaged under laninamivir pressure. Sequence changes in the NA and HA proteins were analyzed at different passages (Table 2). Sequence analysis of assay 1 with recombinant A/Quebec/144147/2009 virus passaged in presence of laninamivir revealed two HA substitutions (V135A and G158E), but the latter was also found in control-passaged virus. In assay 2, two NA substitutions (E119K and G147E) and one HA change (K133E) were detected. The E119K NA substitution was also selected in the A/Auckland/3/2009 virus along with K156E and D225G HA substitutions after 5 passages under laninamivir pressure whereas an A/Solomon Islands/3/2006 variant containing an E119A substitution in the NA protein emerged after 11 passages in presence of laninamivir. The passage of the A/Brisbane/10/2007-like (H3N2) virus under laninamivir pressure resulted in the emergence of a large deletion (237 aa) in the NA protein (Fig. 1) which was observed after 6 passages with no changes being detected in the NA protein of the controlpassaged virus. On the other hand, the S138A and P194L HA changes were concomitantly found in the laninamivir-passaged virus whereas the S138A and N144K HA substitutions were found in the control-passaged virus. 10
11 Characterization of laninamivir-resistant influenza mutants selected in vitro. Herein, we further characterized the A(H1N1)pdm09 [A/Quebec/144147/2009 and A/Auckland/3/2009], A(H1N1) [A/Solomon Islands/3/2006-like] and the A(H3N2) [A/Brisbane/10/2007-like] influenza variants selected with laninamivir. In plaque reduction assays, these variants exhibited a reduced or highly reduced susceptibility phenotype to laninamivir and oseltamivir by using criteria analogous to those reported for NA inhibition assays (Table 3 and Fig. 2). We further evaluated the laninamivir-passaged A(H3N2) virus (NA deletion, S138A and P194L HA changes), the control-passaged A(H3N2) virus (S138A and N144K HA substitutions), and the initial WT virus in a hemagglutination-elution (HAE) assay using either turkey or guinea pig red blood cells (RBCs) (data not shown). The HAE assay was done in the absence and in the presence of laninamivir, zanamivir or oseltamivir. No difference in elution time was observed between the control-passaged virus and the initial virus in absence or in presence of NAIs; however, elution time was faster with turkey RBCs than with guinea pig RBCs equivalent to 30 and 90 min, respectively. On the other hand, the laninamivir-passaged virus containing the NA deletion and the two HA substitutions S138A / P194L did not agglutinate the two types of RBCs. Generation and characterization of laninamivir-resistant recombinant A(H1N1)pdm09 viruses and proteins. A recombinant A(H1N1)pdm09 [A/Quebec/144147/2009] E119A NA variant was successfully generated by reverse genetics. Susceptibility profiles of the WT and E119A recombinants viruses to laninamivir, zanamivir, oseltamivir and peramivir as determined by NA inhibition assays, are summarized in Table 4. The E119A NA substitution conferred 11
12 reduced susceptibility to laninamivir, zanamivir, oseltamivir and peramivir with 82-, 90-, 17- and 12- fold increases in IC 50 values, respectively, compared with those of the WT virus. A recombinant A(H1N1)pdm09 [A/Quebec/144147/2009] harboring E119K NA and K133E HA substitutions was also generated by reverse genetics. Despite multiple attempts, we were unable to generate the NA E119K or E119K / G147E viruses without the HA change. Consequently, we generated recombinant NA proteins expressed in 293T cells and assessed their NA enzymatic activity. As shown in Figure 3, the E119A, E119K and E119K / G147E variants were associated with a significant reduction of total NA activity, with relative total NA activities of 36.5 % (P < 0.001), 1.0 % (P < 0.001) and 1.1 % (P < 0.001), respectively, compared to the WT protein. Of note, we cannot distinguish between decreased activity or expression based on the current assay. The G147E substitution alone did not significantly affect the relative total NA activity (104 %) or susceptibility to laninamivir (7-fold decrease in IC 50 value compared to WT). DISCUSSION NAIs are expected to play a major role for the control of seasonal and eventual pandemic influenza infections. However, the emergence and spread of NAI-resistant variants is a serious concern. The identification of amino acid substitutions conferring resistance to NAIs from in vitro studies may help to understand mechanisms of resistance and to predict clinical cases of resistance to this class of antivirals. In fact, the well-known NA changes conferring resistance to oseltamivir in humans, including the H1N1 H275Y variant and the H3N2 E119V and R292K mutants were previously predicted by in vitro studies (21-23). In the 12
13 present study, we used an in vitro approach to investigate mechanisms of resistance to laninamivir, a novel NAI By testing several A(H1N1)pdm09 as well as seasonal A(H1N1) and A(H3N2) variants, previously found to be resistant to at least one NAI, we demonstrated a similar pattern of susceptibility between laninamivir and zanamivir. More specifically, laninamivir was shown to be active against oseltamivir-resistant H1N1-H275Y as reported (24) and H3N2-E119V variants. Therefore, laninamivir could constitute an antiviral option for the treatment of severe oseltamivir-resistant cases. By contrast, the recombinant A(H1N1)pdm09 viruses containing E119A/G/V framework NA substitutions exhibited reduced or highly reduced susceptibility to zanamivir and laninamivir in our NA inhibition assays. Accordingly, passaging of the seasonal A(H1N1) [A/Solomon/3/2006] as well as the two A(H1N1)pdm09 [A/Quebec/144147/2009 and A/Auckland/3/2009] strains under laninamivir pressure induced an amino acid substitution at position 119 (E119A/K) of the viral NA. In previous reports, E119A/G/D NA changes were described in A(H3N2), A(H1N9), A(H5N1) and B variants that were selected through in vitro passages in presence of zanamivir(25). The structural similarity between laninamivir and zanamivir confers to these two NAIs a common binding process with the enzyme (26). The latter involves the 4-guanidino group which interacts with the carboxylate of the E119 side chain. The loss of this interaction in the E119A/K/G/V A(H1N1) variants described in this study may result in reduced binding to both zanamivir and laninamivir(27). Of note, as peramivir also contains a guanidine group, the zanamivir/laninamivir-resistant viruses also demonstrated resistance to peramivir (Tables 1 and 4). In contrast to the three above NAIs, oseltamivir contains an acetamide group instead 13
14 of guanidine at that position which explains the lower IC 50 values for the 119 A(H1N1) mutants In a previous work, we demonstrated that a.a. substitutions at residue 119 in recombinant A(H1N1)pdm09 viruses (i.e. E119A/G/V) were associated with altered NA activity resulting in reduced viral titers in replicative capacity experiments (18). Moreover, in experimentallyinfected mice, the recombinant A/Québec/144747/2009 E119G variant was associated with a significant reduction of lung viral titers as compared to the WT and a reversion of this variant to the WT genotype was observed in the ferret model (28). In this study, the E119K change was shown to cause an even greater reduction in NA activity and/or cell surface expression, which may explain why we were unable to perform NA inhibition assays and to generate the NA variant without HA mutation by reverse genetics. Thus, we believe that the E119K substitution would alter the viral fitness of influenza A(H1N1)pdm09 viruses although this hypothesis remains to be confirmed. The Q136K mutation is another NA change that was shown to confer resistance to laninamivir in our NA inhibition assays. This NA substitution was previously described in A(H3N2) and A(H1N1) cultured isolates (29, 30) although the clinical relevance of this substitution remains unclear as the Q136K change could only be detected in infected culture cells in some cases whereas it was absent in clinical samples. Nonetheless, a recent report has described the emergence of this mutation under in vitro zanamivir pressure in A(H1N1)pdm09 viruses(31). The Q136 residue is located at the periphery of the NA binding site and the Q136K change was suggested to alter interactions with D151 and R156, thereby 14
15 disrupting hydrogen links between D151 and zanamivir(30, 32, 33). In one of the laninamivir-resistant A(H1N1)pdm09 variants described in our study, the E119K substitution was accompanied by an additional G147E NA change. However, the characterization of recombinant A(H1N1)pdm09 proteins in this study revealed no significant impact of the G147E change on the susceptibility to laninamivir and NA activity (Figure 3) when compared to the recombinant WT protein. Following passages of the A(H3N2) [A/Brisbane/10/2007-like] virus under laninamivir pressure, we observed a large deletion in the NA protein. Of interest, almost the whole enzymatic head domain of the NA protein (residues 106 to 342) including the active site was missing. Consequently, this A(H3N2) variant had undetectable NA enzymatic activity. We and others previously observed a similar deletion in the seasonal A(H1N1) [A/WSN/33] virus exposed to zanamivir selective pressure (34, 35). In two of these cases, the NA deletion was accompanied by HA substitutions, S138A / P194L for A(H3N2) and A200T for A(H1N1). The hemagglutination elution (HAE) assay showed that our A(H3N2) mutant containing the NA deletion and the two HA substitutions did not agglutinate two RBC species. It is plausible that these HA mutations reduce affinity or alter receptors specificity resulting in a virus that is less dependent on the NA and allowing the replication of the NA-deficient viruses. HA mutations were observed in all A(H1N1)pdm09 and A(H3N2) viruses passaged under laninamivir pressure. In the first series of passages of A(H1N1)pdm09 [A/Quebec/144147/2009] virus under laninamivir pressure, we did not observe any NA 15
16 substitution but detected two HA substitutions. We further cultured this variant in the presence of a high laninamivir concentration (100 µm) and obtained good cytopathic effects. One of the HA mutations (V135A) is located on the right edge of the receptor binding site (RBS) that comprises the residues (36). We also detected a HA substitution preceding the selection of the NA changes in the second series of passages of A(H1N1)pdm09 [A/Quebec/144147/2009] virus. This HA substitution at position 133 appeared after 3 passages under laninamivir pressure and was conserved throughout the selection process even after emergence of NA substitutions at positions 119 and 147, which occurred at passages 6 and 9, respectively. The residue 133 is located near the right edge of the RBS. The experiment with a different A(H1N1)pdm09 virus [A/Auckland/3/2009] in MDCK cells, also resulted in the selection of the E119K NA variant along with two HA substitutions, K156E and D225G. Of note, the selected K156E substitution is located near the RBS and it has been shown that the D225G change switches receptor binding specificity from α2,6 linkage binding to dual receptor binding (37). Similarly, the A(H3N2) [A/Brisbane/10/2007-like] virus containing an HA substitution at position 194 appeared after 3 passages under laninamivir pressure concomitantly with the NA deletion. The 194 residue is located in the RBS, which probably explains the growth of the variant in cell culture, despite a NA deletion impairing its enzymatic activity. Interestingly, we observed that control-passaged viruses, i.e. not submitted to drug pressure, acquired some HA mutations during viral propagation in MDCK cells overexpressing the α2,6 sialic acid receptors. The A(H1N1)pdm09 [A/Quebec/144147/2009] virus acquired the G158E and K122N HA substitutions during passages in assays 1 and 2, respectively, while 16
17 A(H3N2) [A/Brisbane/10/2007-like] acquired two HA substitutions, S138A and N144K. It has been previously reported that HA substitutions G158E and K122N could improve the growth of A(H1N1)pdm09 virus in MDCK cells and eggs and that the G158E change drastically reduced viral antigenicity (38). We selected another substitution of interest in the A(H3N2) virus antigenic site A, the N144K HA substitution. This amino acid change is present along with others (E62K, K158N, K173Q and N189K) in a drifted strain that emerged in 2009 (39, 40). In summary, our results demonstrate that the drug resistance pattern of laninamivir and zanamivir appears similar and that both NA and HA changes emerge during the in vitro selection of influenza A(H1N1)pdm09 and A(H3N2) variants with reduced susceptibility to laninamivir. Furthermore, the selection of HA mutations in or near the RBS in all A(H1N1)pdm09 and A(H3N2) viruses passaged under laninamivir pressure emphasizes the importance of HA binding affinity on the replicative capacity of laninamivir-resistant viruses. Overall, our results highlight the importance of the HA/NA balance in the resistance phenotype to NAIs such as laninamivir as well as the need to develop drugs with new viral targets. ACKNOWLEDGMENTS This study was supported by the Canadian Institutes of Health Research (grants and to G.B.), Biota Scientific Management and by the public Health Agency of Canada/Canadian Institutes of Health Research Influenza Research Network (PCIRN) 17
18 (postdoctoral training award to MS). G.B. is the holder of the Canada research chair on influenza and other respiratory viruses REFERENCES 1. Davies BE Pharmacokinetics of oseltamivir: an oral antiviral for the treatment and prophylaxis of influenza in diverse populations. J Antimicrob Chemother. 65 (Suppl. 2):ii5-ii Dutkowski R, Thakrar B, Froehlich E, Suter P, Oo C, Ward P Safety and pharmacology of oseltamivir in clinical use. Drug Saf. 26: He G, Massarella J, Ward P Clinical pharmacokinetics of the prodrug oseltamivir and its active metabolite Ro Clin Pharmacokinet. 37: Dharan NJ, Gubareva LV, Meyer JJ, Okomo-Adhiambo M, McClinton RC, Marshall SA, St George K, Epperson S, Brammer L, Klimov AI, Bresee JS, Fry AM Infections with oseltamivir-resistant influenza A(H1N1) virus in the United States. JAMA. 301: Hauge SH, Dudman S, Borgen K, Lackenby A, Hungnes O Oseltamivirresistant influenza viruses A (H1N1), Norway, Emerg Infect Dis. 15: Meijer A, Lackenby A, Hungnes O, Lina B, van-der-werf S, Schweiger B, Opp M, Paget J, van-de-kassteele J, Hay A, Zambon M Oseltamivir-resistant influenza virus A (H1N1), Europe season. Emerg Infect Dis. 15: Collins PJ, Haire LF, Lin YP, Liu J, Russell RJ, Walker PA, Skehel JJ, Martin SR, Hay AJ, Gamblin SJ Crystal structures of oseltamivir-resistant influenza virus neuraminidase mutants. Nature. 453: Malaisree M, Rungrotmongkol T, Decha P, Intharathep P, Aruksakunwong O, Hannongbua S Understanding of known drug-target interactions in the catalytic pocket of neuraminidase subtype N1. Proteins. 71: Wang MZ, Tai CY, Mendel DB Mechanism by which mutations at his274 alter sensitivity of influenza a virus n1 neuraminidase to oseltamivir carboxylate and zanamivir. Antimicrob Agents Chemother. 46: Kohno S, Kida H, Mizuguchi M, Hirotsu N, Ishida T, Kadota J, Shimada J Intravenous peramivir for treatment of influenza A and B virus infection in high-risk patients. Antimicrob Agents Chemother. 55: FDA Relenza (zanamivir) Information.: forpatientsandproviders/ucm htm. 12. Kubo S, Tomozawa T, Kakuta M, Tokumitsu A, Yamashita M Laninamivir prodrug CS-8958, a long-acting neuraminidase inhibitor, shows superior antiinfluenza virus activity after a single administration. Antimicrob Agents Chemother. 54:
19 Yamashita M, Tomozawa T, Kakuta M, Tokumitsu A, Nasu H, Kubo S CS-8958, a prodrug of the new neuraminidase inhibitor R , shows long-acting anti-influenza virus activity. Antimicrob Agents Chemother. 53: Sugaya N, Ohashi Y Long-acting neuraminidase inhibitor laninamivir octanoate (CS-8958) versus oseltamivir as treatment for children with influenza virus infection. Antimicrob Agents Chemother. 54: Watanabe A, Chang SC, Kim MJ, Chu DW, Ohashi Y Long-acting neuraminidase inhibitor laninamivir octanoate versus oseltamivir for treatment of influenza: A double-blind, randomized, noninferiority clinical trial. Clin Infect Dis. 51: Yamashita M Laninamivir and its prodrug, CS-8958: long-acting neuraminidase inhibitors for the treatment of influenza. Antivir Chem Chemother. 21: Potier M, Mameli L, Bélisle M, Dallaire L, Melançon S Fluorometric assay of neuraminidase with a sodium (4-methylumbelliferyl-α-D-N-acetylneuraminate) substrate. Anal Biochem. 94: Pizzorno A, Bouhy X, Abed Y, Boivin G Generation and characterization of recombinant pandemic influenza A(H1N1) viruses resistant to neuraminidase inhibitors. J Infect Dis. 203: Abed Y, Goyette N, Boivin G A reverse genetics study of resistance to neuraminidase inhibitors in an influenza A/H1N1 virus. Antivir Ther. 9: McKimm-Breschkin JL, Blick TJ, Sahasrabudhe A, Tiong T, Marshall D, Hart GJ, Bethell RC, Penn CR Generation and characterization of variants of NWS/G70C influenza virus after in vitro passage in 4-amino-Neu5Ac2en and 4- guanidino-neu5ac2en. Antimicrob Agents Chemother. 40: Gubareva LV Molecular mechanisms of influenza virus resistance to neuraminidase inhibitors. Virus Res. 103: McKimm-Breschkin JL Resistance of influenza viruses to neuraminidase inhibitors-a review. Antiviral Res. 47: Tai CY, Escarpe PA, Sidwell RW, Williams MA, Lew W, Wu H, Kim CU, Mendel DB Characterization of human influenza virus variants selected in vitro in the presence of the neuraminidase inhibitor GS Antimicrob Agents Chemother. 42: Kiso M, Shinya K, Shimojima M, Takano R, Takahashi K, Katsura H, Kakugawa S, Le MT, Yamashita M, Furuta Y, Ozawa M, Kawaoka Y Characterization of oseltamivir-resistant 2009 H1N1 pandemic influenza A viruses. PLoS pathogens 6:e Samson M, Pizzorno A, Abed Y, Boivin G Influenza virus resistance to neuraminidase inhibitors. Antivir Res. 98: Vavricka CJ, Li Q, Wu Y, Qi J, Wang M, Liu Y, Gao F, Liu J, Feng E, He J, Wang J, Liu H, Jiang H, Gao GF Structural and functional analysis of laninamivir and its octanoate prodrug reveals group specific mechanisms for influenza NA inhibition. PLoS Pathog. 7:e Smith BJ, McKimm-Breshkin JL, McDonald M, Fernley RT, Varghese JN, Colman PM Structural studies of the resistance of influenza virus neuramindase to inhibitors. J Med Chem. 45:
20 Pizzorno A, Abed Y, Rhéaume C, Bouhy X, Boivin G Evaluation of recombinant 2009 pandemic influenza A (H1N1) viruses harboring zanamivir resistance mutations in mice and ferrets. Antimicrob Agents Chemother. 57: Dapat C, Suzuki Y, Saito R, Kyaw Y, Myint YY, Lin N, Oo HN, Oo KY, Win N, Naito M, Hasegawa G, Dapat IC, Zaraket H, Baranovich T, Nishikawa M, Saito T, Suzuki H Rare influenza A (H3N2) variants with reduced sensitivity to antiviral drugs. Emerg Infect Dis. 16: Hurt AC, Holien JK, Parker M, Kelso A, Barr IG Zanamivir-resistant influenza viruses with a novel neuraminidase mutation. J Virol. 83: Kaminski MM, Ohnemus A, Staeheli P, Rubbenstroth D Pandemic 2009 H1N1 influenza A virus carrying a Q136K mutation in the neuraminidase gene is resistant to zanamivir but exhibits reduced fitness in the guinea pig transmission model. J Virol. 87: Han N, Liu X, Mu Y Exploring the mechanism of zanamivir resistance in a neuraminidase mutant: a molecular dynamics study. PLoS One 7:e Russell RJ, Haire LF, Stevens DJ, Collins PJ, Lin YP, Blackburn GM, Hay AJ, Gamblin SJ, Skehel JJ The structure of H5N1 avian influenza neuraminidase suggests new opportunities for drug design. Nature. 443: Nedyalkova MS, Hayden FG, Webster RG, Gubareva LV Accumulation of defective neuraminidase (NA) genes by influenza A viruses in the presence of NA inhibitors as a marker of reduced dependence on NA. The Journal of infectious diseases 185: Baz M, Abed Y, Boivin G Characterization of drug-resistant recombinant influenza A/H1N1 viruses selected in vitro with peramivir and zanamivir. Antiviral Res. 74: Nobusawa E, Aoyama T, Kato H, Suzuki Y, Tateno Y, Nakajima K Comparison of complete amino acid sequences and receptor-binding properties among 13 serotypes of hemagglutinins of influenza A viruses. Virology. 182: Zhang W, Shi Y, Qi J, Gao F, Li Q, Fan Z, Yan J, Gao GF Molecular basis of the receptor binding specificity switch of the hemagglutinins from both the 1918 and 2009 pandemic influenza A viruses by a D225G substitution. J Virol. 87: Chen Z, Wang W, Zhou H, Suguitan AL, Shambaugh C, Kim L, Zhao J, Kemble G, Jin H Generation of live attenuated novel influenza virus A/California/7/09 (H1N1)vaccines with high yield in embryonated chicken eggs. J Virol. 84: Ann J, Papenburg J, Bouhy X, Rhéaume C, Hamelin MÈ, Boivin G Molecular and antigenic evolution of human influenza A/H3N2 viruses in Quebec, Canada, J Clin Virol. 53: Yang JR, Lin CH, Chen CJ, Liu JL, Huang YP, Kuo CY, Yao CY, Hsu LC, Lo J, Ho Y, Wu HS, Liu MT A new antigenic variant of human influenza A (H3N2) virus isolated fromairport and community surveillance in Taiwan in early Virus Res. 151: Pizzorno A, Abed Y, Bouhy X, Beaulieu E, Mallett C, Russell R, Boivin G Impact of mutations at residue I223 of the neuraminidase protein on the resistance 20
21 profile, replication level, and virulence of the 2009 pandemic influenza virus. Antimicrob Agents Chemother. 56: Abed Y, Baz M, Boivin G Impact of neuraminidase mutations conferring influenza resistance to neuraminidase inhibitors in the N1 and N2 genetic backgrounds. Antivir Ther. 11: Baz M, Abed Y, Simon P, Hamelin ME, Boivin G Effect of the neuraminidase mutation H274Y conferring resistance to oseltamivir on the replicative capacity and virulence of old and recent human influenza A(H1N1) viruses. J Infect Dis. 201: Baz M, Abed Y, McDonald J, Boivin G Characterization of multidrugresistant influenza A/H3N2 viruses shed during 1 year by an immunocompromised child. Clin Infect Dis. 43: Abed Y, Baz M, Boivin G A novel neuraminidase deletion mutation conferring resistance to oseltamivir in clinical influenza A/H3N2 virus. J Infect Dis. 199: Downloaded from on October 9, 2018 by guest 21
22 Table 1. Laninamivir susceptibility profiles of influenza A(H1N1)pdm09, A(H1N1) and A(H3N2) viruses harboring NA substitutions mediating resistance to other neuraminidase inhibitors. Influenza strains A(H1N1)pdm09 [A/Quebec/144147/2009] [Recombinant] A(H1N1) [A/WSN/33] [Recombinant] A(H1N1) [A/Brisbane/59/07] [Recombinant] A(H3N2) [Clinical isolates] NA IC 50 in nm (fold increase) b Phenotype in NA inhibition assays Mutations a Laninamivir Zanamivir Laninamivir Zanamivir Oseltamivir Peramivir References WT 1.41 ± 0.01 (1) 0.15±0.01 (1) S S S S (18) ± (18, 28) E119G ± 7.6 (832) HRI HRI S RI (108) E119V ± 8.80 (49) 85.7 ± 10.6 (571) RI HRI RI RI (18) I223R 5.91 ± 2.20 (4) 1.10 ± 0.1 (7) S S RI RI (41) I223R-H275Y 8.45 ± 3.06 (6) 2.32 ± 0.1 (16) S RI HRI HRI (41) I223V 1.71 ± 0.01 (1) 0.35 ± 0.02 (2) S S S S (18, 41) I223V-H275Y 2.01 ± 0.11 (1) 0.32 ± 0.01 (2) S S HRI HRI (18, 41) Q136K ± (45) ± 15.8 (749) RI HRI S ND (28) D199G 2.46 ± 0.80 (2) 0.90 ± 0.06 (6) S S RI S (18) H275Y 4.22 ± 0.11 (3) 0.14 ± 0.01 (0.9) S S HRI HRI (18) N295S 3.70 ± 0.33 (3) 0.49 ± 0.02 (3) S S HRI RI (18) WT 0.66 ± 0.01 (1) 0.56 ± 0.07 (1) S S S S (19) H275Y 1.35 ± 0.02 (2) 0.62 ± 0.04 (1) S S HRI RI (19) N295S 1.28 ± 0.10 (2) 3.17 ± 0.3 (5) S S HRI S (42) WT 4.50 ± 0.03 (1) 0.25 ± 0 (1) S S S S (43) H275Y 3.32 ± 0.17 (1) 0.5 ± 0.15 (2) S S HRI HRI (43) WT 4.61 ± 2.00 (1) 3.6 ± 0.5 (1) S S S S (44) E119V ± 8.50 (5) 4.7 ± 0.4 (1) S S HRI S (44) E119V-I223V 8.59 ± 0.95 (2) 5.29 ± 2.43 (2) S S HRI S (44) Del ± 1.87 (2) 10.1 ± 2.4 (2.8) S S HRI S (45) S, susceptibility or normal inhibition (<10-fold increase in IC 50 over WT); RI, reduced inhibition ( fold increase in IC 50 over WT); HRI, highly reduced inhibition ( 100-fold increase in IC 50 over WT); ND, not determined. a Residues are numbered according to N1 numbering for H1N1 variants and N2 numbering for H3N2 variants. b Values are means ±standard deviations from three independent experiments. 22
23 Table 2. Sequence changes in the neuraminidase and hemagglutinin genes of influenza viruses selected with laninamivir. Influenza strains A(H1N1)pdm09 [A/Quebec/144147/2009] Assay 1 A(H1N1)pdm09 [A/Quebec/144147/2009] Assay 2 A(H1N1)pdm09 [A/Auckland/3/2009] A(H1N1) [A/Solomon Islands/3/2006] A(H3N2) [A/Brisbane/10/2007] a Based on N1 numbering for H1N1 variants and N2 numbering for H3N2 variants. b Based on H3 numbering as in Nobusawa et al. (36). Passage number Laninamivir (µm) NA amino acid substitution a HA amino acid substitution b 0 (Control) No mutation G158E 1.00 No mutation G158E Mixed population 135V : 135A 0 (Control) No mutation G158E 4.00 No mutation V135A / G158E 0 (Control) No mutation G158E No mutation V135A / G158E 0 (Control) No mutation G158E No mutation V135A / G158E 0 (Control) No mutation No mutation 1.00 No mutation K133E 0 (Control) No mutation K122N 3.00 Mixed population 119E : 119K K133E 0 (Control) No mutation K122N Mixed population 119E : 119K Mixed population 147G : 147E K133E 0 (Control) No mutation K122N E119K / G147E K133E 0 (Control) No mutation K136E 0.77 E119K K156E / D225G 0 (Control) No mutation No mutation 8.20 E119A No mutation 0 (Control) No mutation 0.22 No mutation 0 (Control) No mutation 1.80 Del Mixed population 138S : 138A Mixed population 144N : 144K Mixed population 138S : 138A Mixed population 138S : 138A Mixed population 144N : 144K S138A Mixed population 194P : 194L 0 (Control) No mutation S138A / N144K 2.00 Del S138A / P194L 23
24 Table 3. NAIs susceptibility profiles of A(H3N2) and A(H1N1)pdm09 viruses selected with laninamivir as assessed by plaque reduction assay. Influenza strains Passage number Amino acid changes a IC 50 in nm (fold increase) b Laninamivir Oseltamivir A(H1N1)pdm09 [A/Quebec/144147/2009] Assay 1 A(H1N1)pdm09 [A/Quebec/144147/2009] Assay 2 A(H3N2) [A/Brisbane/10/2007] A(H1N1)pdm09 [A/Auckland/3/2009] A(H1N1) [A/Solomon Islands/3/2006] a NA numbering based on N1 numbering for H1N1 variants and N2 numbering for H3N2 variants and HA numbering based on H3 numbering as in Nobusawa et al. (36). b Values are means ± standard deviations from two independent experiments. 0 WT 5.89 ± 0.84 (1.0) 7.55 ± 0.37 (1.0) 12 Control HA G158E 19.37± 2.41 (3.3) 4.65 ± 0.32 (0.6) 12 HA V135A / G158E ± (183.3) ± (26.1) 3 HA K133E 7.62 ± 1.01 (1.3) 8.43 ± 0.08 (1.1) 12 Control 12 HA K122N 4.90 ± 0.11 (0.8) 4.03 ± 0.17 (0.5) NA E119K / G147E HA K133E 100,000 ( 10,000) 100,000 ( 10,000) 0 WT 3.11 ± 0.83 (1.0) 1.87 ± 0.04 (1.0) 9 Control 9 HA S138A / N144K 3.71 ± 0.15 (1.2) 3.12 ± 0.13 (1.7) NA Del HA S138A / P194L 100,000 ( 10,000) 100,000 ( 10,000) 0 WT ± ± Control 5 HA K136E ± ± NA E119K HA K156E / D225G >100,000 (>394) >100,000 (>708) 0 WT ± ± Control WT ± ± NA E119 A ± (325) ± (31) 24
25 Table 4. Susceptibility profiles to neuraminidase inhibitors of a recombinant A(H1N1)pdm09 virus harboring the E119A neuraminidase substitution Influenza strains Mutations a IC 50 in nm (fold increase) b A(H1N1)pdm09 [A/Québec/144147/09] WT NA E119A Laninamivir Zanamivir Oseltamivir Peramivir 1.41 ± 0.01 (1) ± 9.50 (82) 1.53 ± 0.02 (1) ± 8.8 (90) 3.20 ± 0.01 (1) ± 1.21 (17) 0.61± 0.00 (1) 7.22 ± 0.77 (12) a Based on N1 numbering system. b Values are means ± standard deviations from three independent experiments. 25
26 Figures legends Figure 1. A(H3N2) [A/Brisbane/10/2007] laninamivir-selected variant with a major deletion in the region encompassing the active site of the neuraminidase (NA) protein. (A) PCR products of the NA gene from the wild-type (WT) strain and the NA variant were separated on a 1% electrophoresis gel in presence of a 1 Kb molecular weight (MW) marker. (B) Schematic representation of the full-length (469 aa) and the deleted (232 aa) NA proteins. Figure 2. Plaque size of the A(H1N1)pdm09 [A/Quebec/144147/2009] and A(H3N2) [A/Brisbane/10/2007] laninamivir-selected variants. Viruses selected with laninamivir were propagated in 12-well plates containing MDCK α2,6 cells under 0.8% agarose. Plaques were visualized after 48 h of incubation at 37 C in the absence or presence of laninamivir. Figure 3. Activity of recombinant A(H1N1)pdm09 [A/Quebec/144147/2009] neuraminidase proteins. 293T cells were transfected with pcaggs-pa, -PB1, -PB2, and -NP plasmids in addition to plasmids expressing the WT or varianta/quebec/144147/2009 neuraminidase (NA)proteins. At 48 h post-transfection, NA activity was measured by using the fluorogenic substrate (MUNANA). Percent NA activities were determined in triplicate experiments ± standard deviations. *P< compared to the WT NA activity. 26
27

28
29
Detection and management of antiviral resistance for influenza viruses
 DOI:10.1111/irv.12176 www.influenzajournal.com Review Article Detection and management of antiviral resistance for influenza viruses Guy Boivin CHUQ-CHUL and Laval University, Quebec, QC, Canada. Correspondence:
DOI:10.1111/irv.12176 www.influenzajournal.com Review Article Detection and management of antiviral resistance for influenza viruses Guy Boivin CHUQ-CHUL and Laval University, Quebec, QC, Canada. Correspondence:
Flu, Avian Flu and emerging aspects (H1N1 resistance)
") EU-CIS Seminar New trends in Infectious Diseases 26 28 November 2008 / Lyon, France Flu, Avian Flu and emerging aspects (H1N1 resistance) Pr. Florence MORFIN FRE 3011 Université Lyon 1 - CNRS Laboratory
EU-CIS Seminar New trends in Infectious Diseases 26 28 November 2008 / Lyon, France Flu, Avian Flu and emerging aspects (H1N1 resistance) Pr. Florence MORFIN FRE 3011 Université Lyon 1 - CNRS Laboratory
Emergence of Multidrug-Resistant Influenza A(H1N1)pdm09 Virus Variants in an Immunocompromised Child Treated With Oseltamivir and Zanamivir
pdm09 Virus Variants in an Immunocompromised Child Treated With Oseltamivir and Zanamivir") BRIEF REPORT Emergence of Multidrug-Resistant Influenza A(H1N1)pdm09 Virus Variants in an Immunocompromised Child Treated With Oseltamivir and Zanamivir Daisuke Tamura, 1,a Roberta L. DeBiasi, 2,4,5,a
BRIEF REPORT Emergence of Multidrug-Resistant Influenza A(H1N1)pdm09 Virus Variants in an Immunocompromised Child Treated With Oseltamivir and Zanamivir Daisuke Tamura, 1,a Roberta L. DeBiasi, 2,4,5,a
Antiviral-resistant B viruses with a novel mutation detected by antiviral resistance surveillance in Japan
 6thMeeting of NICs in the WPR and SEAR 29-31 May, 2012 Hanoi VN Antiviral-resistant B viruses with a novel mutation detected by antiviral resistance surveillance in Japan Masaki Imai WHO Collaborating
6thMeeting of NICs in the WPR and SEAR 29-31 May, 2012 Hanoi VN Antiviral-resistant B viruses with a novel mutation detected by antiviral resistance surveillance in Japan Masaki Imai WHO Collaborating
TREATING MEXICAN (NOVEL) INFLUENZA IN AN IMMUNOCOMPROMISED PATIENT
 INFLUENZA IN AN IMMUNOCOMPROMISED PATIENT") TREATING MEXICAN (NOVEL) INFLUENZA IN AN IMMUNOCOMPROMISED PATIENT Zanamivir (carbons coloured grey) No conformational change required to allow binding Sialic acid Natural Substrate (carbons coloured
TREATING MEXICAN (NOVEL) INFLUENZA IN AN IMMUNOCOMPROMISED PATIENT Zanamivir (carbons coloured grey) No conformational change required to allow binding Sialic acid Natural Substrate (carbons coloured
Review. The antiviral resistance of influenza virus. Vanessa Escuret 1,2, Olivier Ferraris 2 & Bruno Lina 1,2
 Review The antiviral resistance of influenza virus The 2009 pandemic confirmed the increasing role antiviral treatment plays in influenza disease management in severe cases. In the 1960s adamantane derivatives
Review The antiviral resistance of influenza virus The 2009 pandemic confirmed the increasing role antiviral treatment plays in influenza disease management in severe cases. In the 1960s adamantane derivatives
Influenza A Virus Hemagglutinin Antibody Escape Promotes Neuraminidase Antigenic Variation and Drug Resistance
 Influenza A Virus Hemagglutinin Antibody Escape Promotes Neuraminidase Antigenic Variation and Drug Resistance Scott E. Hensley, Suman R. Das, James S. Gibbs, Adam L. Bailey, Loren M. Schmidt, Jack R.
Influenza A Virus Hemagglutinin Antibody Escape Promotes Neuraminidase Antigenic Variation and Drug Resistance Scott E. Hensley, Suman R. Das, James S. Gibbs, Adam L. Bailey, Loren M. Schmidt, Jack R.
Review Influenza virus susceptibility and resistance to oseltamivir
 Review Influenza virus susceptibility and resistance to oseltamivir Fred Y Aoki 1, Guy Boivin 2 and Noel Roberts 3 * Antiviral Therapy 12:603 616 1 Department of Medical Microbiology, University of Manitoba,
Review Influenza virus susceptibility and resistance to oseltamivir Fred Y Aoki 1, Guy Boivin 2 and Noel Roberts 3 * Antiviral Therapy 12:603 616 1 Department of Medical Microbiology, University of Manitoba,
TITLE: Influenza A (H7N9) virus evolution: Which genetic mutations are antigenically important?
 virus evolution: Which genetic mutations are antigenically important?") TITLE: Influenza A (H7N9) virus evolution: Which genetic mutations are antigenically important? AUTHORS: Joshua G. Petrie 1, Adam S. Lauring 2,3 AFFILIATIONS: 1 Department of Epidemiology, University of
TITLE: Influenza A (H7N9) virus evolution: Which genetic mutations are antigenically important? AUTHORS: Joshua G. Petrie 1, Adam S. Lauring 2,3 AFFILIATIONS: 1 Department of Epidemiology, University of
Michael M. Kaminski, Annette Ohnemus, Peter Staeheli *, and Dennis Rubbenstroth *
 JVI Accepts, published online ahead of print on 28 November 2012 J. Virol. doi:10.1128/jvi.02507-12 Copyright 2012, American Society for Microbiology. All Rights Reserved. Zanamivir resistance in 2009
JVI Accepts, published online ahead of print on 28 November 2012 J. Virol. doi:10.1128/jvi.02507-12 Copyright 2012, American Society for Microbiology. All Rights Reserved. Zanamivir resistance in 2009
J.A.L. Ives a, *, J.A. Carr a, D.B. Mendel b, C.Y. Tai b, R. Lambkin c,1, L. Kelly c, J.S. Oxford c, F.G. Hayden d, N.A. Roberts a. 1.
 Antiviral Research 55 (2002) 307 317 www.elsevier.com/locate/antiviral The H274Y mutation in the influenza A/H1N1 neuraminidase active site following oseltamivir phosphate treatment leave virus severely
Antiviral Research 55 (2002) 307 317 www.elsevier.com/locate/antiviral The H274Y mutation in the influenza A/H1N1 neuraminidase active site following oseltamivir phosphate treatment leave virus severely
Influenza Virus Genotypes Circulating In Central Greece During And Vaccine Strain Match
 ISPUB.COM The Internet Journal of Microbiology Volume 13 Number 1 Influenza Virus Genotypes Circulating In Central Greece During 2012-2014 And Vaccine Strain Match E Plakokefalos, A Vontas, Z Florou, G
ISPUB.COM The Internet Journal of Microbiology Volume 13 Number 1 Influenza Virus Genotypes Circulating In Central Greece During 2012-2014 And Vaccine Strain Match E Plakokefalos, A Vontas, Z Florou, G
Antiviral Therapy 2015; 20:49 55 (doi: /IMP2798)
") Antiviral Therapy 2015; 20:49 55 (doi: 10.3851/IMP2798) Original article Comparison between virus shedding and fever duration after treating children with pandemic A H1N1/09 and children with A H3N2 with
Antiviral Therapy 2015; 20:49 55 (doi: 10.3851/IMP2798) Original article Comparison between virus shedding and fever duration after treating children with pandemic A H1N1/09 and children with A H3N2 with
Recent H3N2 influenza virus clinical isolates rapidly acquire hemagglutinin or neuraminidase mutations when propagated for antigenic analyses
 JVI Accepts, published online ahead of print on 2 July 2014 J. Virol. doi:10.1128/jvi.01077-14 Copyright 2014, American Society for Microbiology. All Rights Reserved. 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
JVI Accepts, published online ahead of print on 2 July 2014 J. Virol. doi:10.1128/jvi.01077-14 Copyright 2014, American Society for Microbiology. All Rights Reserved. 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
Emergence and Fixing of Antiviral Resistance in Influenza A Via Recombination and Hitch Hiking. Henry L Niman
 Emergence and Fixing of Antiviral Resistance in Influenza A Via Recombination and Hitch Hiking Henry L Niman Recombinomics, Inc, Pittsburgh, Pennsylvania USA Department of Influenza Recombination Recombinomics,
Emergence and Fixing of Antiviral Resistance in Influenza A Via Recombination and Hitch Hiking Henry L Niman Recombinomics, Inc, Pittsburgh, Pennsylvania USA Department of Influenza Recombination Recombinomics,
Influenza surveillance and pandemic preparedness - a global challenge Anne Kelso
 Influenza surveillance and pandemic preparedness - a global challenge Anne Kelso WHO Collaborating Centre for Reference and Research on Influenza Melbourne, Australia Three global health challenges 243
Influenza surveillance and pandemic preparedness - a global challenge Anne Kelso WHO Collaborating Centre for Reference and Research on Influenza Melbourne, Australia Three global health challenges 243
Supplementary Figure 1 Weight and body temperature of ferrets inoculated with
 Supplementary Figure 1 Weight and body temperature of ferrets inoculated with A/Anhui/1/2013 (H7N9) influenza virus. (a) Body temperature and (b) weight change of ferrets after intranasal inoculation with
Supplementary Figure 1 Weight and body temperature of ferrets inoculated with A/Anhui/1/2013 (H7N9) influenza virus. (a) Body temperature and (b) weight change of ferrets after intranasal inoculation with
This product was developed by the Victorian Infectious Diseases Reference Laboratory (VIDRL) in its capacity as a WHO Collaborating Centre for
 in its capacity as a WHO Collaborating Centre for") This product was developed by the Victorian Infectious Diseases Reference Laboratory (VIDRL) in its capacity as a WHO Collaborating Centre for Reference and Research on Influenza, with material provided
This product was developed by the Victorian Infectious Diseases Reference Laboratory (VIDRL) in its capacity as a WHO Collaborating Centre for Reference and Research on Influenza, with material provided
The A(H7N9) influenza outbreak in China
 influenza outbreak in China") Viruses in May, Katoomba, 9 11 May 2013 The A(H7N9) influenza outbreak in China Anne Kelso Director WHO Collaborating Centre for Reference and Research on Influenza Melbourne Influenza in the 21 st century:
Viruses in May, Katoomba, 9 11 May 2013 The A(H7N9) influenza outbreak in China Anne Kelso Director WHO Collaborating Centre for Reference and Research on Influenza Melbourne Influenza in the 21 st century:
Reagents for the Typing of Human Influenza Isolates 2011
 Reagents for the Typing of Human Influenza Isolates 2011 This product was developed by the Victorian Infectious Diseases Reference Laboratory (VIDRL) in its capacity as a WHO Collaborating Centre for Reference
Reagents for the Typing of Human Influenza Isolates 2011 This product was developed by the Victorian Infectious Diseases Reference Laboratory (VIDRL) in its capacity as a WHO Collaborating Centre for Reference
Pandemic Influenza influenza epidemic: realization of a worst-case scenario
 Pandemic Influenza October 9, 2006 1918 influenza epidemic: realization of a worst-case scenario First case: Albert Mitchell, Camp Funston, KS, March 11, 1918 Up to 20% of all humans infected 20-50 million
Pandemic Influenza October 9, 2006 1918 influenza epidemic: realization of a worst-case scenario First case: Albert Mitchell, Camp Funston, KS, March 11, 1918 Up to 20% of all humans infected 20-50 million
ph1n1 H3N2: A Novel Influenza Virus Reassortment
 ph1n1 H3N2: A Novel Influenza Virus Reassortment Jonathan Gubbay Medical Microbiologist Public Health Laboratory Public Health Ontario June 16, 2011 ph1n1 H3N2 Reassortment: Talk Overview Explain strain
ph1n1 H3N2: A Novel Influenza Virus Reassortment Jonathan Gubbay Medical Microbiologist Public Health Laboratory Public Health Ontario June 16, 2011 ph1n1 H3N2 Reassortment: Talk Overview Explain strain
Influenza antiviral susceptibility: methods and challenges to detect resistant virus
 SARINet Pan-American Health Organization - PAHO Influenza antiviral susceptibility: methods and challenges to detect resistant virus Brazilian experience Paola Cristina Resende, PhD Researcher National
SARINet Pan-American Health Organization - PAHO Influenza antiviral susceptibility: methods and challenges to detect resistant virus Brazilian experience Paola Cristina Resende, PhD Researcher National
NEXT GENERATION SEQUENCING OPENS NEW VIEWS ON VIRUS EVOLUTION AND EPIDEMIOLOGY. 16th International WAVLD symposium, 10th OIE Seminar
 NEXT GENERATION SEQUENCING OPENS NEW VIEWS ON VIRUS EVOLUTION AND EPIDEMIOLOGY S. Van Borm, I. Monne, D. King and T. Rosseel 16th International WAVLD symposium, 10th OIE Seminar 07.06.2013 Viral livestock
NEXT GENERATION SEQUENCING OPENS NEW VIEWS ON VIRUS EVOLUTION AND EPIDEMIOLOGY S. Van Borm, I. Monne, D. King and T. Rosseel 16th International WAVLD symposium, 10th OIE Seminar 07.06.2013 Viral livestock
Reassortment and Mutations Associated with Emergence and Spread of Oseltamivir-Resistant Seasonal Influenza A/H1N1 Viruses in
 Reassortment and Mutations Associated with Emergence and Spread of Oseltamivir-Resistant Seasonal Influenza A/H1N1 Viruses in 2005 2009 Ji-Rong Yang 1, Yu-Cheng Lin 1, Yuan-Pin Huang 1, Chun-Hui Su 1,JeLo
Reassortment and Mutations Associated with Emergence and Spread of Oseltamivir-Resistant Seasonal Influenza A/H1N1 Viruses in 2005 2009 Ji-Rong Yang 1, Yu-Cheng Lin 1, Yuan-Pin Huang 1, Chun-Hui Su 1,JeLo
Blocking Interhost Transmission of Influenza Virus by Vaccination in the Guinea Pig Model
 JOURNAL OF VIROLOGY, Apr. 2009, p. 2803 2818 Vol. 83, No. 7 0022-538X/09/$08.00 0 doi:10.1128/jvi.02424-08 Copyright 2009, American Society for Microbiology. All Rights Reserved. Blocking Interhost Transmission
JOURNAL OF VIROLOGY, Apr. 2009, p. 2803 2818 Vol. 83, No. 7 0022-538X/09/$08.00 0 doi:10.1128/jvi.02424-08 Copyright 2009, American Society for Microbiology. All Rights Reserved. Blocking Interhost Transmission
A study of oseltamivir-resistant influenza viruses in Thailand,
 A study of oseltamivir-resistant influenza viruses in Thailand, 2008-2010 Malinee Chittaganpitch*, Sunthareeya Waicharoen*, Jiranan Warachit De silva*, Krongkaew Supawat*, Sirima Pattamadilok*, Wattana
A study of oseltamivir-resistant influenza viruses in Thailand, 2008-2010 Malinee Chittaganpitch*, Sunthareeya Waicharoen*, Jiranan Warachit De silva*, Krongkaew Supawat*, Sirima Pattamadilok*, Wattana
ECDC TECHNICAL REPORT
 TECHNICAL REPORT External quality assessment scheme for antiviral susceptibility detection in influenza viruses for the Community Network of Reference Laboratories for Human Influenza in Europe 2010/2011
TECHNICAL REPORT External quality assessment scheme for antiviral susceptibility detection in influenza viruses for the Community Network of Reference Laboratories for Human Influenza in Europe 2010/2011
on September 26, 2018 by guest
 AAC Accepts, published online ahead of print on 18 January 2013 Antimicrob. Agents Chemother. doi:10.1128/aac.02145-12 Copyright 2013, American Society for Microbiology. All Rights Reserved. 1 2 Title:
AAC Accepts, published online ahead of print on 18 January 2013 Antimicrob. Agents Chemother. doi:10.1128/aac.02145-12 Copyright 2013, American Society for Microbiology. All Rights Reserved. 1 2 Title:
Chang Gung University co-commissioned final report. Research Ttitle: Antiviral mechanism study for 254 UVC robot system
 co-commissioned final report Research Ttitle: Antiviral mechanism study for 254 UVC robot system Project/Research Number:: Execution duration: 2014.10.16-2015.01.16 Principle Investigator: Professor Shin-Ru
co-commissioned final report Research Ttitle: Antiviral mechanism study for 254 UVC robot system Project/Research Number:: Execution duration: 2014.10.16-2015.01.16 Principle Investigator: Professor Shin-Ru
In Vitro Generation of Neuraminidase Inhibitor Resistance in A(H5N1) Influenza Viruses
 Influenza Viruses") ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, Oct. 2009, p. 4433 4440 Vol. 53, No. 10 0066-4804/09/$08.00 0 doi:10.1128/aac.00334-09 Copyright 2009, American Society for Microbiology. All Rights Reserved. In
ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, Oct. 2009, p. 4433 4440 Vol. 53, No. 10 0066-4804/09/$08.00 0 doi:10.1128/aac.00334-09 Copyright 2009, American Society for Microbiology. All Rights Reserved. In
Antiviral Research xxx (2005) xxx xxx. Short communication
 xxx xxx. Short communication") Antiviral Research xxx (2005) xxx xxx 3 4 5 6 7 8 Short communication Sensitivity of influenza viruses to zanamivir and oseltamivir: A study performed on viruses circulating in France prior to the introduction
Antiviral Research xxx (2005) xxx xxx 3 4 5 6 7 8 Short communication Sensitivity of influenza viruses to zanamivir and oseltamivir: A study performed on viruses circulating in France prior to the introduction
Supporting Information
 Supporting Information Yen et al. 10.1073/pnas.1111000108 SI Materials and Methods Cells. Madin Darby canine kidney (MDCK) cells and human embryonic kidney 293T cells were obtained from the American Type
Supporting Information Yen et al. 10.1073/pnas.1111000108 SI Materials and Methods Cells. Madin Darby canine kidney (MDCK) cells and human embryonic kidney 293T cells were obtained from the American Type
REAGENTS FOR THE TYPING OF HUMAN INFLUENZA ISOLATES 2017
 REAGENTS FOR THE TYPING OF HUMAN INFLUENZA ISOLATES 2017 This product was developed by the Victorian Infectious Diseases Reference Laboratory (VIDRL) in its capacity as a WHO Collaborating Centre for Reference
REAGENTS FOR THE TYPING OF HUMAN INFLUENZA ISOLATES 2017 This product was developed by the Victorian Infectious Diseases Reference Laboratory (VIDRL) in its capacity as a WHO Collaborating Centre for Reference
Anti-influenza (M2 and NA) Drugs
 Drugs") Anti-influenza (M2 and NA) Drugs Alan J. Hay MRC National Institute for Medical Research, London Summer School on Influenza Siena 1-5 August 2011 Antiviral drugs licensed for use against influenza M2 (Flu
Anti-influenza (M2 and NA) Drugs Alan J. Hay MRC National Institute for Medical Research, London Summer School on Influenza Siena 1-5 August 2011 Antiviral drugs licensed for use against influenza M2 (Flu
Application of Reverse Genetics to Influenza Vaccine Development
 NIAID Application of Reverse Genetics to Influenza Vaccine Development Kanta Subbarao Laboratory of Infectious Diseases NIAID, NIH Licensed Vaccines for Influenza Principle: Induction of a protective
NIAID Application of Reverse Genetics to Influenza Vaccine Development Kanta Subbarao Laboratory of Infectious Diseases NIAID, NIH Licensed Vaccines for Influenza Principle: Induction of a protective
Influenza virus.
 INFLUENZA VIRUS Adapté en partie des exposés de la Chaire Franqui 2003 "Antiviral drugs and Discoveries in Medicine" Prof. E. De Clercq, KU-Leuven http://www.md.ucl.ac.be/chaire-francqui/ Influenza virus
INFLUENZA VIRUS Adapté en partie des exposés de la Chaire Franqui 2003 "Antiviral drugs and Discoveries in Medicine" Prof. E. De Clercq, KU-Leuven http://www.md.ucl.ac.be/chaire-francqui/ Influenza virus
Practical guidance for national influenza centres establishing or implementing neuraminidase inhibitor susceptibility surveillance
 Practical guidance for national influenza centres establishing or implementing neuraminidase inhibitor susceptibility surveillance WHO Library Cataloguing-in-Publication Data Practical guidance for national
Practical guidance for national influenza centres establishing or implementing neuraminidase inhibitor susceptibility surveillance WHO Library Cataloguing-in-Publication Data Practical guidance for national
Planning for Respiratory Pandemics. Michael G. Ison, MD MS. Viruses in May
 Planning for Respiratory Pandemics Michael G. Ison, MD MS Divisions of Infectious Diseases & Organ Transplantation Transplant Infectious Diseases Service Northwestern University Comprehensive Transplant
Planning for Respiratory Pandemics Michael G. Ison, MD MS Divisions of Infectious Diseases & Organ Transplantation Transplant Infectious Diseases Service Northwestern University Comprehensive Transplant
AGLOBAL EMERGENCE AND
 BRIEF REPORT Morbidity and Mortality Associated With Nosocomial Transmission of - Resistant Influenza A(H1N1) Virus Jairo Gooskens, MD Marcel Jonges, MSc Eric C. J. Claas, PhD Adam Meijer, PhD Peterhans
BRIEF REPORT Morbidity and Mortality Associated With Nosocomial Transmission of - Resistant Influenza A(H1N1) Virus Jairo Gooskens, MD Marcel Jonges, MSc Eric C. J. Claas, PhD Adam Meijer, PhD Peterhans
Amino acid changes in hemagglutinin contribute to the replication of. oseltamivir-resistant H1N1 influenza viruses
 JVI Accepts, published online ahead of print on 19 October 2011 J. Virol. doi:10.1128/jvi.06085-11 Copyright 2011, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.
JVI Accepts, published online ahead of print on 19 October 2011 J. Virol. doi:10.1128/jvi.06085-11 Copyright 2011, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.
Supplementary Figure 1. FACS analysis of cells infected with TY93/H5N1 GFP-627E,
 Supplementary Figure 1. FACS analysis of cells infected with TY93/H5N1 GFP-627E, TY93/H5N1 GFP-627K, or the TY93/H5N1 PB2(588-759) virus library. To establish our GFP- FACS screening platform, we compared
Supplementary Figure 1. FACS analysis of cells infected with TY93/H5N1 GFP-627E, TY93/H5N1 GFP-627K, or the TY93/H5N1 PB2(588-759) virus library. To establish our GFP- FACS screening platform, we compared
Supporting Information
 Supporting Information Valkenburg et al. 10.1073/pnas.1403684111 SI Materials and Methods ELISA and Microneutralization. Sera were treated with Receptor Destroying Enzyme II (RDE II, Accurate) before ELISA
Supporting Information Valkenburg et al. 10.1073/pnas.1403684111 SI Materials and Methods ELISA and Microneutralization. Sera were treated with Receptor Destroying Enzyme II (RDE II, Accurate) before ELISA
Ralf Wagner Paul-Ehrlich-Institut
 www.pei.de Other Assays for the Detection of Neuraminidase (NA)-Specific Antibodies Ralf Wagner Paul-Ehrlich-Institut Overview to presented assays Assay principle based on: Chemical substrates: Protein
www.pei.de Other Assays for the Detection of Neuraminidase (NA)-Specific Antibodies Ralf Wagner Paul-Ehrlich-Institut Overview to presented assays Assay principle based on: Chemical substrates: Protein
INFLUENZA VACCINATION AND MANAGEMENT SUMMARY
 INFLUENZA VACCINATION AND MANAGEMENT SUMMARY Morbidity and mortality related to influenza occur at a higher rate in people over 65 and those with underlying chronic medical conditions. Annual influenza
INFLUENZA VACCINATION AND MANAGEMENT SUMMARY Morbidity and mortality related to influenza occur at a higher rate in people over 65 and those with underlying chronic medical conditions. Annual influenza
ADCC Assay Protocol Vikram Srivastava 1, Zheng Yang 1, Ivan Fan Ngai Hung 2, Jianqing Xu 3, Bojian Zheng 3 and Mei- Yun Zhang 3*
 ADCC Assay Protocol Vikram Srivastava 1, Zheng Yang 1, Ivan Fan Ngai Hung 2, Jianqing Xu 3, Bojian Zheng 3 and Mei- Yun Zhang 3* 1 Department of Microbiology, Li Ka Shing Faculty of Medicine, University
ADCC Assay Protocol Vikram Srivastava 1, Zheng Yang 1, Ivan Fan Ngai Hung 2, Jianqing Xu 3, Bojian Zheng 3 and Mei- Yun Zhang 3* 1 Department of Microbiology, Li Ka Shing Faculty of Medicine, University
Influenza or flu is a
 Clinical and Research Area Infectious Diseases Influenza Virus Types A and B Influenza or flu is a respiratory illness that is caused by influenza viruses. Influenza viruses type A and type B cause seasonal
Clinical and Research Area Infectious Diseases Influenza Virus Types A and B Influenza or flu is a respiratory illness that is caused by influenza viruses. Influenza viruses type A and type B cause seasonal
1918 Influenza; Influenza A, H1N1. Basic agent information. Section I- Infectious Agent. Section II- Dissemination
 1918 Influenza; Influenza A, H1N1 Basic agent information Section I- Infectious Agent Risk Group: - RG3 Synonym or Cross reference: - Spanish Flu - 1918 Flu - El Grippe Characteristics: - SELECT AGENT
1918 Influenza; Influenza A, H1N1 Basic agent information Section I- Infectious Agent Risk Group: - RG3 Synonym or Cross reference: - Spanish Flu - 1918 Flu - El Grippe Characteristics: - SELECT AGENT
External quality assessment for influenza antiviral susceptibility for the European Reference Laboratory Network for human influenza
 TECHNICAL REPORT External quality assessment for influenza antiviral susceptibility for the European Reference Laboratory Network for human influenza 2013 www.ecdc.europa.eu ECDC TECHNICAL REPORT External
TECHNICAL REPORT External quality assessment for influenza antiviral susceptibility for the European Reference Laboratory Network for human influenza 2013 www.ecdc.europa.eu ECDC TECHNICAL REPORT External
10 3 pfu A B 2004/ FluA. Viral Shedding. Viral Shedding. Frank Houston Family Study 2005/
 56 pp.109-1162006 6. 1 2 1 2 10 3 pfu A B 2004/2005 1500 1700 120 1000 1500 420 50 Viral Shedding Frank 1975 1980 Houston Family Study 12 4 A 110-8645 2 23 16 TEL 03-3833-8381 FAX 03-3831-9488 E-mail mitamura@eijuhp.com
56 pp.109-1162006 6. 1 2 1 2 10 3 pfu A B 2004/2005 1500 1700 120 1000 1500 420 50 Viral Shedding Frank 1975 1980 Houston Family Study 12 4 A 110-8645 2 23 16 TEL 03-3833-8381 FAX 03-3831-9488 E-mail mitamura@eijuhp.com
6.2 Pandemic influenza
 6.2 Pandemic influenza See Background Paper 6.2 (BP6_2Pandemic.pdf) Background and developments since 2004 The WHO estimates that annual influenza epidemics account for about 3 to 5 million cases of severe
6.2 Pandemic influenza See Background Paper 6.2 (BP6_2Pandemic.pdf) Background and developments since 2004 The WHO estimates that annual influenza epidemics account for about 3 to 5 million cases of severe
Relative activity (%) SC35M
 SC35M") a 125 Bat (H17N) b 125 A/WSN (H1N1) Relative activity (%) 0 75 50 25 Relative activity (%) 0 75 50 25 0 Pos. Neg. PA PB1 Pos. Neg. NP PA PB1 PB2 0 Pos. Neg. NP PA PB1 PB2 SC35M Bat Supplementary Figure
a 125 Bat (H17N) b 125 A/WSN (H1N1) Relative activity (%) 0 75 50 25 Relative activity (%) 0 75 50 25 0 Pos. Neg. PA PB1 Pos. Neg. NP PA PB1 PB2 0 Pos. Neg. NP PA PB1 PB2 SC35M Bat Supplementary Figure
Medicinal Chemistry Feature Molecule
 Current Medicinal Chemistry, 2000, 7, 000-000 663 Medicinal Chemistry Feature Molecule Discovery and Development of GS 4104 (oseltamivir): An rally Active Influenza Neuraminidase Inhibitor Willard Lew*,
Current Medicinal Chemistry, 2000, 7, 000-000 663 Medicinal Chemistry Feature Molecule Discovery and Development of GS 4104 (oseltamivir): An rally Active Influenza Neuraminidase Inhibitor Willard Lew*,
Original Article Development and Sequence Analysis of a Cold-Adapted Strain of Influenza A/New Caledonia/20/1999(H1N1) Virus
 Virus") Iranian Journal of Virology 2011;5(4): 6-10 2011, Iranian Society for Virology Original Article Development and Sequence Analysis of a Cold-Adapted Strain of Influenza A/New Caledonia/20/1999(H1N1) Virus
Iranian Journal of Virology 2011;5(4): 6-10 2011, Iranian Society for Virology Original Article Development and Sequence Analysis of a Cold-Adapted Strain of Influenza A/New Caledonia/20/1999(H1N1) Virus
Report on the Deliberation Results
 Report on the Deliberation Results August 3, 2010 Evaluation and Licensing Division, Pharmaceutical and Food Safety Bureau Ministry of Health, Labour and Welfare [Brand name] Inavir Dry Powder Inhaler
Report on the Deliberation Results August 3, 2010 Evaluation and Licensing Division, Pharmaceutical and Food Safety Bureau Ministry of Health, Labour and Welfare [Brand name] Inavir Dry Powder Inhaler
In the Name of God. Talat Mokhtari-Azad Director of National Influenza Center
 In the Name of God Overview of influenza laboratory diagnostic technology: advantages and disadvantages of each test available Talat Mokhtari-Azad Director of National Influenza Center Tehran- Iran 1 1)
In the Name of God Overview of influenza laboratory diagnostic technology: advantages and disadvantages of each test available Talat Mokhtari-Azad Director of National Influenza Center Tehran- Iran 1 1)
An Exploration to Determine if Fab Molecules are Efficacious in Neutralizing Influenza H1 and H3 Subtypes. Nick Poulton June September 2012
 An Exploration to Determine if Fab Molecules are Efficacious in Neutralizing Influenza H1 and H3 Subtypes Nick Poulton June 2012- September 2012 Epidemiology of Influenza Infection Causes between 250,000
An Exploration to Determine if Fab Molecules are Efficacious in Neutralizing Influenza H1 and H3 Subtypes Nick Poulton June 2012- September 2012 Epidemiology of Influenza Infection Causes between 250,000
Emergence of distinct avian-like influenza A H1N1 viruses in pigs in Ireland and their reassortment with cocirculating H3N2 viruses
 International Congress Series 1263 (2004) 209 213 Emergence of distinct avian-like influenza A H1N1 viruses in pigs in Ireland and their reassortment with cocirculating H3N2 viruses Y.P. Lin a, *, M. Bennett
International Congress Series 1263 (2004) 209 213 Emergence of distinct avian-like influenza A H1N1 viruses in pigs in Ireland and their reassortment with cocirculating H3N2 viruses Y.P. Lin a, *, M. Bennett
Characteristics of oseltamivir-resistant influenza A (H1N1) pdm09 virus during the influenza season in Mainland China
 pdm09 virus during the influenza season in Mainland China") Huang et al. Virology Journal (2015) 12:96 DOI 10.1186/s12985-015-0317-1 RESEARCH Open Access Characteristics of oseltamivir-resistant influenza A (H1N1) pdm09 virus during the 2013 2014 influenza season
Huang et al. Virology Journal (2015) 12:96 DOI 10.1186/s12985-015-0317-1 RESEARCH Open Access Characteristics of oseltamivir-resistant influenza A (H1N1) pdm09 virus during the 2013 2014 influenza season
Preparing for the Fall Flu Season. Jonathan Gubbay Medical Microbiologist Public Health Laboratory OAHPP
 Preparing for the Fall Flu Season Laboratory Perspective Jonathan Gubbay Medical Microbiologist Public Health Laboratory OAHPP September 21, 2009 Objectives 1. Review the emergence of Novel Influenza A
Preparing for the Fall Flu Season Laboratory Perspective Jonathan Gubbay Medical Microbiologist Public Health Laboratory OAHPP September 21, 2009 Objectives 1. Review the emergence of Novel Influenza A
What is influenza virus? 13,000 base RNA genome: 1/ the size of the human genome
 What is influenza virus? 13,000 base RNA genome: 1/246153 the size of the human genome CDC Principles of Virology, 4e Neumann et al. Nature. 2009. Influenza virus is one of the most deadly viral pathogens
What is influenza virus? 13,000 base RNA genome: 1/246153 the size of the human genome CDC Principles of Virology, 4e Neumann et al. Nature. 2009. Influenza virus is one of the most deadly viral pathogens
INFLUENZA VIRUS. Influenza virus
 IFLUEZA VIRUS Adapté en partie des exposés de la Chaire Franqui 2003 "Antiviral drugs and Discoveries in Medicine" Prof. E. De Clercq, KU-Leuven http://www.md.ucl.ac.be/chaire-francqui/ Influenza virus
IFLUEZA VIRUS Adapté en partie des exposés de la Chaire Franqui 2003 "Antiviral drugs and Discoveries in Medicine" Prof. E. De Clercq, KU-Leuven http://www.md.ucl.ac.be/chaire-francqui/ Influenza virus
Review Report. Inavir Dry Powder Inhaler 20 mg
 Review Report June 27, 2016 Pharmaceuticals and Medical Devices Agency The following are the results of a review of the following pharmaceutical product submitted for marketing approval conducted by the
Review Report June 27, 2016 Pharmaceuticals and Medical Devices Agency The following are the results of a review of the following pharmaceutical product submitted for marketing approval conducted by the
Persistent Infection of MDCK Cells by Influenza C Virus: Initiation and Characterization
 J. gen. Virol. (199), 70, 341-345. Printed in Great Britain 341 Key words: influenza C virus/interferon/persistent infection Persistent Infection of MDCK Cells by Influenza C Virus: Initiation and Characterization
J. gen. Virol. (199), 70, 341-345. Printed in Great Britain 341 Key words: influenza C virus/interferon/persistent infection Persistent Infection of MDCK Cells by Influenza C Virus: Initiation and Characterization
Temperature-Sensitive Mutants Isolated from Hamster and
 JOURNAL OF VIROLOGY, Nov. 1975, p. 1332-1336 Copyright i 1975 American Society for Microbiology Vol. 16, No. 5 Printed in U.S.A. Temperature-Sensitive Mutants Isolated from Hamster and Canine Cell Lines
JOURNAL OF VIROLOGY, Nov. 1975, p. 1332-1336 Copyright i 1975 American Society for Microbiology Vol. 16, No. 5 Printed in U.S.A. Temperature-Sensitive Mutants Isolated from Hamster and Canine Cell Lines
Spread of Influenza A(H1N1) oseltamivir-resistant viruses in Africa in 2008 confirmed by multiple introductions in Senegal
 oseltamivir-resistant viruses in Africa in 2008 confirmed by multiple introductions in Senegal") Dia et al. BMC Infectious Diseases 2013, 13:106 RESEARCH ARTICLE Open Access Spread of Influenza A(H1N1) oseltamivir-resistant viruses in Africa in 2008 confirmed by multiple introductions in Senegal Ndongo
Dia et al. BMC Infectious Diseases 2013, 13:106 RESEARCH ARTICLE Open Access Spread of Influenza A(H1N1) oseltamivir-resistant viruses in Africa in 2008 confirmed by multiple introductions in Senegal Ndongo
Public health relevant virological features of Influenza A(H7N9) causing human infection in China
 causing human infection in China") Public health relevant virological features of Influenza A(H7N9) causing human infection in China Address requests about publications of the WHO Regional Office for Europe to: Publications WHO Regional
Public health relevant virological features of Influenza A(H7N9) causing human infection in China Address requests about publications of the WHO Regional Office for Europe to: Publications WHO Regional
Recommended laboratory tests to identify influenza A/H5 virus in specimens from patients with an influenza-like illness
 World Health Organization Recommended laboratory tests to identify influenza A/H5 virus in specimens from patients with an influenza-like illness General information Highly pathogenic avian influenza (HPAI)
World Health Organization Recommended laboratory tests to identify influenza A/H5 virus in specimens from patients with an influenza-like illness General information Highly pathogenic avian influenza (HPAI)
Cristina Cassetti, Ph.D.
 NIAID Extramural Research Update: Recombinant Influenza Viruses and Biosafety Cristina Cassetti, Ph.D. Influenza Program Officer Division of Microbiology and Infectious Diseases NIAID Influenza virus DMID
NIAID Extramural Research Update: Recombinant Influenza Viruses and Biosafety Cristina Cassetti, Ph.D. Influenza Program Officer Division of Microbiology and Infectious Diseases NIAID Influenza virus DMID
INFLUENZA EVOLUTION: Challenges for diagnosis
 INFLUENZA EVOLUTION: Challenges for diagnosis Jairo A. Méndez-Rico Influenza Team PAHO/WHO, Washington, DC Overview Every year, influenza infects up to one in five people around the world, and causes up
INFLUENZA EVOLUTION: Challenges for diagnosis Jairo A. Méndez-Rico Influenza Team PAHO/WHO, Washington, DC Overview Every year, influenza infects up to one in five people around the world, and causes up
Prophylaxis and Treatment for Influenza among the Elderly
 Respiratory Diseases in the Elderly Prophylaxis and Treatment for Influenza among the Elderly JMAJ 45(6): 245 250, 2002 Hajime GOTO Professor, The First Department of Internal Medicine, Kyorin University
Respiratory Diseases in the Elderly Prophylaxis and Treatment for Influenza among the Elderly JMAJ 45(6): 245 250, 2002 Hajime GOTO Professor, The First Department of Internal Medicine, Kyorin University
Update on influenza monitoring and vaccine development
 Update on influenza monitoring and vaccine development Annette Fox WHO Collaborating Centre for Reference and Research on Influenza at The Peter Doherty Institute for Infection and Immunity 1 Outline Why
Update on influenza monitoring and vaccine development Annette Fox WHO Collaborating Centre for Reference and Research on Influenza at The Peter Doherty Institute for Infection and Immunity 1 Outline Why
Report on virological surveillance of influenza activity in the Romania 2014/15 season
 National Institute for Research Cantacuzino National Influenza Centre, Laboratory for Respiratory Viruses Spl.Independentei 103, sector 5, 050096 Bucuresti Romania Report on virological surveillance of
National Institute for Research Cantacuzino National Influenza Centre, Laboratory for Respiratory Viruses Spl.Independentei 103, sector 5, 050096 Bucuresti Romania Report on virological surveillance of
C~ 6 D~~J ~ old ~ s rw~ o, - _ - 2 DEPARTMENT OF HEALTH AND HUMAN SERVICES. Food and Drug Administration. 21 CFR Part 530
 DEPARTMENT OF HEALTH AND HUMAN SERVICES Food and Drug Administration 21 CFR Part 530, s rw~ o, - _ - 2 [Docket No. 2006N-0106] New Animal Drugs ; Adamantane and Neuraminidase Inhibitor Anti-influenza Drugs
DEPARTMENT OF HEALTH AND HUMAN SERVICES Food and Drug Administration 21 CFR Part 530, s rw~ o, - _ - 2 [Docket No. 2006N-0106] New Animal Drugs ; Adamantane and Neuraminidase Inhibitor Anti-influenza Drugs
Development of safe and immunogenic reassortant viruses with 5:3 genotype for live attenuated influenza vaccine
 Development of safe and immunogenic reassortant viruses with 5:3 genotype for live attenuated influenza vaccine Irina Isakova-Sivak, PhD Institute of Experimental Medicine, Saint Petersburg, Russia The
Development of safe and immunogenic reassortant viruses with 5:3 genotype for live attenuated influenza vaccine Irina Isakova-Sivak, PhD Institute of Experimental Medicine, Saint Petersburg, Russia The
2009 H1N1 Influenza ( Swine Flu ) Hemagglutinin ELISA kit
 Hemagglutinin ELISA kit") 2009 H1N1 Influenza ( Swine Flu ) Hemagglutinin ELISA kit Catalog Number : SEK001 To achieve the best assay results, this manual must be read carefully before using this product and the assay is run as
2009 H1N1 Influenza ( Swine Flu ) Hemagglutinin ELISA kit Catalog Number : SEK001 To achieve the best assay results, this manual must be read carefully before using this product and the assay is run as
Influenza: The past, the present, the (future) pandemic
 pandemic") Influenza: The past, the present, the (future) pandemic Kristin Butler, MLS (ASCP) cm Department of Clinical Laboratory Sciences Louisiana Health Sciences Center - Shreveport Fall 2017 Objectives 1) Detail
Influenza: The past, the present, the (future) pandemic Kristin Butler, MLS (ASCP) cm Department of Clinical Laboratory Sciences Louisiana Health Sciences Center - Shreveport Fall 2017 Objectives 1) Detail
Characterization of two distinct neuraminidases from avian-origin human-infecting H7N9 influenza viruses
 ORIGINAL ARTICLE Cell Research (2013) 23:1347-1355. 2013 IBCB, SIBS, CAS All rights reserved 1001-0602/13 www.nature.com/cr npg Characterization of two distinct neuraminidases from avian-origin human-infecting
ORIGINAL ARTICLE Cell Research (2013) 23:1347-1355. 2013 IBCB, SIBS, CAS All rights reserved 1001-0602/13 www.nature.com/cr npg Characterization of two distinct neuraminidases from avian-origin human-infecting
Supplemental Information Dose Response Parameters for Gain of Function Pathogens
 Supplemental Information Dose Response Parameters for Gain of Function Pathogens Infection Dose-Response To quantify the likelihood of an individual or animal becoming infected from exposure to virus,
Supplemental Information Dose Response Parameters for Gain of Function Pathogens Infection Dose-Response To quantify the likelihood of an individual or animal becoming infected from exposure to virus,
Active and Passive Immunization for Avian Influenza Virus Infections
 NIAID Active and Passive Immunization for Avian Influenza Virus Infections Kanta Subbarao, MD, MPH Laboratory of Infectious Diseases NIAID, NIH Immortalizing H5 HA-Specific Memory B Cells Collection of
NIAID Active and Passive Immunization for Avian Influenza Virus Infections Kanta Subbarao, MD, MPH Laboratory of Infectious Diseases NIAID, NIH Immortalizing H5 HA-Specific Memory B Cells Collection of
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION doi:10.1038/nature10831 1. Pathological analyses of H5 avian-human reassortant viruses. The rgca04 infection in the lungs of ferrets mainly caused severe bronchopneumonia with
SUPPLEMENTARY INFORMATION doi:10.1038/nature10831 1. Pathological analyses of H5 avian-human reassortant viruses. The rgca04 infection in the lungs of ferrets mainly caused severe bronchopneumonia with
Anti-Influenza Agents Quantity Limit Program Summary
 Anti-Influenza Agents Quantity Limit Program Summary FDA APPROVED INDICATIONS DOSAGE 1,2 Agent Indication Dosage & Administration Relenza Treatment of influenza in Treatment of influenza: (zanamivir) patients
Anti-Influenza Agents Quantity Limit Program Summary FDA APPROVED INDICATIONS DOSAGE 1,2 Agent Indication Dosage & Administration Relenza Treatment of influenza in Treatment of influenza: (zanamivir) patients
Influenza A Virus Transmission Bottlenecks Are Defined by Infection Route and Recipient Host
 Cell Host & Microbe, Volume 16 Supplemental Information Influenza A Virus Transmission Bottlenecks Are Defined by Infection Route and Recipient Host Andrew Varble, Randy A. Albrecht, Simone Backes, Marshall
Cell Host & Microbe, Volume 16 Supplemental Information Influenza A Virus Transmission Bottlenecks Are Defined by Infection Route and Recipient Host Andrew Varble, Randy A. Albrecht, Simone Backes, Marshall
Alberta Health. Seasonal Influenza in Alberta. 2012/2013 Season. Surveillance and Assessment Branch. November Government of Alberta 1
 Alberta Health Seasonal Influenza in Alberta 2012/2013 Season Surveillance and Assessment Branch November 2013 2013 Government of Alberta 1 For more information contact: Surveillance and Assessment Branch
Alberta Health Seasonal Influenza in Alberta 2012/2013 Season Surveillance and Assessment Branch November 2013 2013 Government of Alberta 1 For more information contact: Surveillance and Assessment Branch
Diagnosing and managing
 www.bpac.org.nz keyword: influenza influenza Diagnosing and managing Key reviewers: Associate Professor Mark Thomas, Infectious Disease Specialist, School of Medical Sciences, University of Auckland Dr
www.bpac.org.nz keyword: influenza influenza Diagnosing and managing Key reviewers: Associate Professor Mark Thomas, Infectious Disease Specialist, School of Medical Sciences, University of Auckland Dr
Tamiflu. Tamiflu (oseltamivir) Description
 Description") Federal Employee Program 1310 G Street, N.W. Washington, D.C. 20005 202.942.1000 Fax 202.942.1125 5.01.19 Subject: Tamiflu Page: 1 of 5 Last Review Date: March 18, 2016 Tamiflu Description Tamiflu (oseltamivir)
Federal Employee Program 1310 G Street, N.W. Washington, D.C. 20005 202.942.1000 Fax 202.942.1125 5.01.19 Subject: Tamiflu Page: 1 of 5 Last Review Date: March 18, 2016 Tamiflu Description Tamiflu (oseltamivir)
Identification of Microbes Lecture: 12
 Diagnostic Microbiology Identification of Microbes Lecture: 12 Electron Microscopy 106 virus particles per ml required for visualization, 50,000-60,000 magnification normally used. Viruses may be detected
Diagnostic Microbiology Identification of Microbes Lecture: 12 Electron Microscopy 106 virus particles per ml required for visualization, 50,000-60,000 magnification normally used. Viruses may be detected
Reverse genetic platform for inactivated and live-attenuated influenza vaccine
 EXPERIMENTAL and MOLECULAR MEDICINE, Vol. 42, No. 2, 116-121, February 2010 Reverse genetic platform for inactivated and live-attenuated influenza vaccine Eun-Ju Jung, Kwang-Hee Lee and Baik Lin Seong
EXPERIMENTAL and MOLECULAR MEDICINE, Vol. 42, No. 2, 116-121, February 2010 Reverse genetic platform for inactivated and live-attenuated influenza vaccine Eun-Ju Jung, Kwang-Hee Lee and Baik Lin Seong
Avian Influenza: Armageddon or Hype? Bryan E. Bledsoe, DO, FACEP The George Washington University Medical Center
 Avian Influenza: Armageddon or Hype? Bryan E. Bledsoe, DO, FACEP The George Washington University Medical Center Definitions: Epidemic The occurrence of cases of an illness in a community or region which
Avian Influenza: Armageddon or Hype? Bryan E. Bledsoe, DO, FACEP The George Washington University Medical Center Definitions: Epidemic The occurrence of cases of an illness in a community or region which
A Novel Duplex Real-Time Reverse-Transcription PCR Assay for the Detection of Influenza A and the Novel Influenza A(H1N1) Strain
 Strain") Viruses 2009, 1, 1204-1208; doi:10.3390/v1031204 OPEN ACCESS viruses ISSN 1999-4915 www.mdpi.com/journal/viruses Article A Novel Duplex Real-Time Reverse-Transcription PCR Assay for the Detection of Influenza
Viruses 2009, 1, 1204-1208; doi:10.3390/v1031204 OPEN ACCESS viruses ISSN 1999-4915 www.mdpi.com/journal/viruses Article A Novel Duplex Real-Time Reverse-Transcription PCR Assay for the Detection of Influenza
Influenza A H1N1 (Swine Flu 2009) Hemagglutinin / HA ELISA Pair Set
 Hemagglutinin / HA ELISA Pair Set") Influenza A H1N1 (Swine Flu 2009) Hemagglutinin / HA ELISA Pair Set Catalog Number : SEK001 To achieve the best assay results, this manual must be read carefully before using this product and the assay
Influenza A H1N1 (Swine Flu 2009) Hemagglutinin / HA ELISA Pair Set Catalog Number : SEK001 To achieve the best assay results, this manual must be read carefully before using this product and the assay
INFLUENZA VIRUS. INFLUENZA VIRUS CDC WEBSITE
 INFLUENZA VIRUS INFLUENZA VIRUS CDC WEBSITE http://www.cdc.gov/ncidod/diseases/flu/fluinfo.htm 1 THE IMPACT OF INFLUENZA Deaths: PANDEMICS 1918-19 S p a n is h flu 5 0 0,0 0 0 U S 2 0,0 0 0,0 0 0 w o rld
INFLUENZA VIRUS INFLUENZA VIRUS CDC WEBSITE http://www.cdc.gov/ncidod/diseases/flu/fluinfo.htm 1 THE IMPACT OF INFLUENZA Deaths: PANDEMICS 1918-19 S p a n is h flu 5 0 0,0 0 0 U S 2 0,0 0 0,0 0 0 w o rld
Influenza viruses. Virion. Genome. Genes and proteins. Viruses and hosts. Diseases. Distinctive characteristics
 Influenza viruses Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Virion Enveloped particles, quasi-spherical or filamentous Diameter 80-120 nm Envelope is derived
Influenza viruses Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Virion Enveloped particles, quasi-spherical or filamentous Diameter 80-120 nm Envelope is derived
Supplementary Figure 1. SC35M polymerase activity in the presence of Bat or SC35M NP encoded from the phw2000 rescue plasmid.
 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 Supplementary Figure 1. SC35M polymerase activity in the presence of Bat or SC35M NP encoded from the phw2000 rescue plasmid. HEK293T
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 Supplementary Figure 1. SC35M polymerase activity in the presence of Bat or SC35M NP encoded from the phw2000 rescue plasmid. HEK293T
Evolution of influenza
 Evolution of influenza Today: 1. Global health impact of flu - why should we care? 2. - what are the components of the virus and how do they change? 3. Where does influenza come from? - are there animal
Evolution of influenza Today: 1. Global health impact of flu - why should we care? 2. - what are the components of the virus and how do they change? 3. Where does influenza come from? - are there animal
Influenza: Seasonal, Avian, and Otherwise
 Influenza: Seasonal, Avian, and Otherwise Lisa Winston, MD University of California, San Francisco San Francisco General Hospital Influenza biology Antiviral medications Seasonal influenza Vaccination
Influenza: Seasonal, Avian, and Otherwise Lisa Winston, MD University of California, San Francisco San Francisco General Hospital Influenza biology Antiviral medications Seasonal influenza Vaccination
External quality assessment scheme for influenza antiviral susceptibility for the European Reference Laboratory Network for Human Influenza
 TECHNICAL REPORT External quality assessment scheme for influenza antiviral susceptibility for the European Reference Laboratory Network for Human Influenza 2015 www.ecdc.europa.eu ECDC TECHNICAL REPORT
TECHNICAL REPORT External quality assessment scheme for influenza antiviral susceptibility for the European Reference Laboratory Network for Human Influenza 2015 www.ecdc.europa.eu ECDC TECHNICAL REPORT
Understanding mortality from pandemic and seasonal influenza
 Understanding mortality from pandemic and seasonal influenza Jonathan A. McCullers Associate Member Department of Infectious Diseases St. Jude Children s Research Hospital H1 H2 H3 H4 H5 H6 H7 H8 H9 H10
Understanding mortality from pandemic and seasonal influenza Jonathan A. McCullers Associate Member Department of Infectious Diseases St. Jude Children s Research Hospital H1 H2 H3 H4 H5 H6 H7 H8 H9 H10
Guidelines for clinical management of severe influenza infection. Aeron Hurt
 Guidelines for clinical management of severe influenza infection Aeron Hurt Current WHO guidelines WHO guidance documents for the clinical management of influenza virus infection were published in 2007
Guidelines for clinical management of severe influenza infection Aeron Hurt Current WHO guidelines WHO guidance documents for the clinical management of influenza virus infection were published in 2007
Cluster-Randomized Controlled Trial of a Brief Theory-Based Avian Influenza Prevention Program for Poultry Workers in Taiwan
 Cluster-Randomized Controlled Trial of a Brief Theory-Based Avian Influenza Prevention Program for Poultry Workers in Taiwan Jiun-Hau Huang 1 2 +, Yen-Yu Miao 1 and Pei-Chun Kuo 1 1 Institute of Health
Cluster-Randomized Controlled Trial of a Brief Theory-Based Avian Influenza Prevention Program for Poultry Workers in Taiwan Jiun-Hau Huang 1 2 +, Yen-Yu Miao 1 and Pei-Chun Kuo 1 1 Institute of Health