Specific and overlapping sphingosine-1-phosphate receptor functions in human. synoviocytes: impact of TNF-alpha
|
|
|
- Wilfrid Goodman
- 6 years ago
- Views:
Transcription
1 Specific and overlapping sphingosine-1-phosphate receptor functions in human synoviocytes: impact of TNF-alpha Chenqi Zhao, Maria J. Fernandes, Mélanie Turgeon, Sabrina Tancrède, John Di Battista, Patrice E. Poubelle, and Sylvain G. Bourgoin Centre de Recherche en Rhumatologie et Immunologie, Centre de Recherche du CHUQ- CHUL, Départements d Anatomie-Physiologie et Médecine, Faculté de Médecine, Université Laval, Québec, Canada, G1V 4G2; and Département de Rhumatologie et Immunologie, Centre Universitaire McGill, Montréal, Québec, Canada, H3A 1A1 Abbreviated title: The role of S1P in synoviocyte function Contact information for correspondence author: Sylvain G. Bourgoin, PhD Centre de Recherche en Rhumatologie et Immunologie, Local T1-49 Centre de Recherche du CHUQ-CHUL 2705 Boul. Laurier, Québec (QC), Canada, G1V 4G2 Phone: (418) , ext Fax: (418) Electronic address: sylvain.bourgoin@crchul.ulaval.ca Abbreviations: COX-2, cyclooxygenase-2; EDG, endothelial differentiation gene; ELISA, enzyme-linked immunosorbent assay; FLS, fibroblast-like synoviocyte; IL, interleukin; IP- 10, interferon-γ-inducible protein 10; MAPK, mitogen-activated protein kinase; MCP-1,
2 monocyte chemotactic protein-1; PGE 2, prostaglandin E 2 ; RA, rheumatoid arthritis; RANTES, regulated on activation normal T-cell expressed and secreted; RT-PCR, reverse transcriptase-polymerase chain reaction; S1P, sphingosine-1-phosphate; SphK, sphingosine kinase; TNF-α, tumor necrosis factor-α. 1
3 Abstract Sphingosine-1-phosphate (S1P), via interaction with its G protein-coupled receptors, regulates various physiological and pathological responses. The present study investigated the role of S1P/S1P receptor signaling in several functional responses of human fibroblastlike synoviocytes (FLS) that may contribute to the pathogenesis of rheumatoid arthritis (RA). We report that FLS express the S1P 1, S1P 2 and S1P 3 receptors. Moreover, exogenously applied S1P induces FLS 1) migration, 2) secretion of inflammatory cytokine/chemokine, and 3) protection from apoptosis. Using specific S1P receptor agonists/antagonists, we determined that S1P stimulates FLS migration through S1P 1 and S1P 3, induces cytokine/chemokine secretion through S1P 2 and S1P 3, whereas protects from cell apoptosis via S1P 1. The S1P-mediated cell motility and cytokine/chemokine secretion seem to be regulated by the p38 mitogen-activated protein kinase (MAPK), p42/44 MAPK and Rho kinase signal transduction pathways. Interestingly, treatment of FLS with TNF-α increases S1P 3 expression and correlates with the enhancement of S1P-induced cytokine/chemokine production. Our data suggest that S1P 1, S1P 2, and S1P 3 play essential roles in the pathogenesis of RA by modulating FLS migration, cytokine/chemokine production, and cell survival. Moreover, the cytokine-rich environment of the inflamed synovium may synergize with S1P signaling to exacerbate the clinical manifestations of this autoimmune disease. Supplementary key words: inflammation G protein-coupled receptor Interleukin-8 cell migration apoptosis mitogen-activated protein kinase Rho kinase 2
4 Introduction Rheumatoid arthritis (RA) is a chronic inflammatory disorder that primarily affects diarthroidal joints leading to their progressive destruction. The disease is characterized by hyperplasia of fibroblast-like synoviocytes (FLS) and a massive infiltration of inflammatory cells in the affected joint. FLS comprise the synovial lining, a thin membrane in direct contact with cartilage and bone. In RA, FLS play a key role in the pathogenesis (reviewed in (1)). FLS increase in number, invade adjacent tissues and produce pro-inflammatory cytokines, chemokines, and matrix metalloproteinases that promote inflammation and joint destruction (2). Proinflammatory cytokines, such as TNF-α and IL-1β, appear to play a key role in the stimulation of FLS towards this aggressive phenotype. There is mounting evidence, however, that the activation of FLS can be maintained in the absence of inflammatory cytokines (3), suggesting the contribution of other pro-inflammatory molecules to the pathogenesis of RA. Sphingosine-1-phosphate (S1P) is a biologically active sphingolipid that transmits potent signals through five G protein-coupled EDG (endothelial differentiation gene) receptors, namely S1P 1 /EDG-1, S1P 2 /EDG-5, S1P 3 /EDG-3, S1P 4 /EDG-6 and S1P 5 /EDG-8 (4). Among the five S1P receptors, S1P 1, S1P 2, and S1P 3 are widely expressed in various tissues, whereas the expression of S1P 4 and S1P 5 is mainly confined to cells of the immune and nervous systems, respectively (5). S1P receptors activate a variety of heterologous signaling pathways through coupling with multiple G proteins (G q, G i/o, and G 12/13 ) that regulate both physiological (cell growth, differentiation, migration, and survival) and pathological processes (angiogenesis, cancer, and autoimmunity) (reviewed in (6)). Activated platelets are the main source of S1P (7). However, other cell types, such as neutrophils and mononuclear cells, also produce S1P (8, 9). S1P synthesis and secretion are stimulated by inflammatory mediators such as TNF-α (10). Virtually all cells that participate 3
5 in immune responses express multiple S1P receptors (reviewed in (11)). While S1P enhances the expression of inflammation-related genes (12), S1P signaling through the S1P 1 receptor also controls T cell migration and tissue distribution as well as the initiation of early events in the differentiation of T cells into effector states (13). Several lines of evidence strongly suggest that S1P contributes to the pathogenesis of RA. S1P is produced by neutrophils, the most abundant cell type in synovial fluids of RA patients. Also, TNF-α, a cytokine that plays a key role in the pathogenesis of RA, as well as other inflammatory mediators induce the synthesis and secretion of S1P. High levels of S1P have, indeed, been reported in the synovial fluid of patients with RA (14). Not only is the synthesis and secretion of S1P induced by proinflammatory mediators but, in turn, S1P also triggers the expression of inflammatory genes such as cyclooxygenase-2 (COX-2), and a super-production of prostaglandin E 2 (PGE 2 ) by FLS (14). Moreover, the expression of sphingosine kinase (SphK), the enzyme that generates S1P by phosphorylation of sphingosine (15), is significantly increased in the cells derived from RA B lymphoblastoid cells (16). These findings from in vitro experiments ought to be at least partly representative of the in vivo physiological function of S1P in the RA synovium. Since FLS activation is partly cytokine-independent and S1P is known to induce gene expression in FLS, we sought to fully characterize the role of the S1P/S1P receptor signaling pathway in FLS function. The objective of the present study was thus to determine whether the S1P/S1P receptor signaling pathway is involved in RA. Herein, we report that FLS express the S1P 1, S1P 2 and S1P 3 receptors. We also provide evidence that these S1P receptors triggered specific and overlapping functional responses. S1P 1 was essential for the survival of FLS and S1P 1 and S1P 3 stimulated FLS migration. In contrast, the activation of S1P 2 and S1P 3 enhanced cytokine/chemokine secretion. In addition, S1P 3 expression by FLS was enhanced by TNF-α and S1P-mediated chemokine secretion was super-induced in TNF- 4
6 α-primed FLS. The results suggest that in addition to S1P 1, S1P 2 and S1P 3 may also play important roles in the pathogenesis of RA. 5
7 Materials and Methods Reagents. S1P was purchased from Biomol (Plymouth Meeting, PA, USA). The specific S1P 1/3 receptor antagonist VPC23019 was obtained from Avanti Polar Lipids Inc (Alabaster, AL, USA). The S1P 1 agonist SEW2871, the S1P 2 antagonist JTE-013, and the S1P 3 antagonist CAY10444, were from Cayman Chemical (Ann Arbor, MI, USA). TNF-α was from PeproTech Inc (Rocky Hill, NJ, USA). Human IL-8 and IL-6 ELISA (Enzyme- Linked Immunosorbent Assay) kits were purchased from BioSource International Inc (Camarillo, CA, USA). The apoptosis assay kit (Annexin V-FITC) was from BD Pharmingen (Oakville, ON, Canada). The cell proliferation assay kit (CellTiter 96 Aqueous One Solution Cell Proliferation) was purchased from Promega (Medison, WI, USA). Sodium nitroferricyanide (III) dihydrate (sodium nitroprusside, SNP), and SYBR Green JumpStart TM Ready Mix kit were obtained from Sigma (St. Louis, MO, USA). TRIzol Reagent was from Invitrogen (Burlington, ON, Canada). Inhibitors of p42/44 mitogen-activated protein kinase (MAPK) PD98059, of p38 MAPK SB203580, and of Rho kinase Y27632 were purchased from Calbiochem (San Diego, CA, USA). S1P 1 and S1P 3 antibodies were from Cayman Chemical (Ann Arbor, MI, USA). Antibodies to total and phosphorylated forms of p42/44 MAPK, of p38mapk, and of JNK were purchased from Cell Signaling Technology (Beverly, MA, USA). The human cytokine/chemokine LINCOplex kit was purchased from Linco Research (St. Charles, MI, USA). Cell culture reagents were purchased from Wisent Inc (St-Bruno, QC, Canada). Cell culture. Human primary FLS were obtained from RA patients, who were diagnosed according to the criteria developed by the American College of Rheumatology (ACR) (17) and were undergoing joint surgery on the knee or hip. Cells were maintained under standard conditions (37 C and 5% CO 2 ) and grown in DMEM supplemented with 10% fetal 6
8 bovine serum (FBS), penicillin (100 IU), and streptomycin (100 µm). Cells were used at passages 5 to 15. Cell treatment. Semi-confluent cells were starved with serum-free medium for 24 h before treatment. At the moment of cell treatment, the culture medium was replaced with fresh serum-free medium containing various concentrations of the tested compounds as indicated in details below. Semi-quantitative reverse transcription (RT)-PCR and real-time PCR analyses. Total cellular RNA was extracted using TRIzol reagent according to the instructions from the manufacturer. RNA (0.5-1 µg) was reverse-transcribed using random priming and Superscript II Reverse Transcriptase system (Invitrogen, Burlington, ON, Canada). Oligonucleotides used as primers were designed to amplify specific cdna sequences. Primer sequences and PCR conditions are as follows: S1P 1 (778-bp product), sense, 5 -GGA- AGG-GAG-TAT-GTT-TGT-GGC-3, antisense, 5 -TGA-CGT-TTC-CAG-AAG-ACA-TA- 3 ; S1P 2 (425-bp product), sense, 5 -AGT-GGC-CAT-TGC-CAA-GGT-CAA-G-3, antisense, 5 -TAG-TGG-GCT-TTG-TAG-AGG-A-3 ; S1P 3 (466-bp product), sense, 5 - AGG-GAG-GGC-AGT-ATG-TTC-G-3, antisense, 5 -GCC-ACA-TCA-ATG-AGG-AAG- AGG-AT-3 ; S1P 4, (672-bp product), sense, 5 - ATC-ACG-CTG-AGT-GAC-CTG-CTC-A- 3, antisense, 5 -TGC-GGA-AGG-AGT-AGA-TGA-3. S1P 5, (658-bp product), sense, 5 - CTA-CTG-TCG-GGG-CCG-CTC-AC-3, antisense, 5 -CGG-TTG-GTG-AAC-GTG-TAG- ATG-A-3 ; IL-8 (562-bp product), sense, 5 -TGG-GTG-CAG-AGG-GTT-GTG-3, antisense, 5 -CAG-ACT-AGG-GTT-GCC-AGA-TTT-3. To ensure linear cdna amplification, different amplifying cycles were tried. The experiments revealed linear amplification within 35 cycles. A total of 35 PCR cycles were run at 94 C (denaturation, 1 min), 63 C for S1P 1-3, 61 C for S1P 4-5 and IL-8 (annealing, 1 min), and 72 C (extension, 1 min). The ribosomal protein RPLP0 mrna was used as an internal PCR control. RPLP0 7
9 (248-bp product) primer sequences are as follows: sense, 5 -GTT-GTA-GAT-GCT-GCC- ATT-G-3, antisense, 5 -CCA-TGT-GAA-GTC-ACT-GTG-C-3. The PCR products were subjected to electrophoresis on a 0.8% agarose gel and visualized by ethidium bromide staining. Densitometry analysis was used for band quantification using the software Alphamage The results were expressed as a ratio of the band intensity relative to the corresponding RPLP0 band obtained by amplification of the same template cdna. Semiquantitative real-time PCR was also conducted to examine the mrna expression of S1P 1-3 receptors. In real-time PCR experiments, we used the same primers as for RT-PCR to amplify S1P 1-3. The thermal cycling conditions were as follows: 95 C (initial denaturation, 3 min) followed by 40 cycles of 95 C (denaturation, 15 sec), 63 C for S1P 1-3 (annealing, 20 sec), and 72 C (extension, 20 sec). Western blot analyses. To monitor the activation of p42/44 MAPK, p38 MAPK, and JNK, cells were exposed to S1P (5 µm) for various times (5-30 min). Where indicated, cells were pre-treated with PD98059 (25 µm), SB (10 µm) and Y27632 (10 µm) for 30 min prior to S1P stimulation (5 µm, 15 min). Cells were lysed in boiling sample buffer [50 mm Tris/HCL (ph 6.8), 10% (v/v) glycerol, 50 mm DTT, 4% (v/v) SDS] and boiled for 7-10 min. Equal amounts of protein (50 µg) were separated by 10% SDS-polyacrylamide gel electrophoresis. For analysis of S1P 1 /S1P 3 receptor expression, cell membrane fractions were prepared as described previously (18). Briefly, cells were treated with accutase and harvested in icecold KCl Hepes relaxation buffer (50 mm Hepes, 100 mm KCl, 5 mm NaCl, 1 mm MgCl 2, 0.5 mm EGTA, 10 µg/ml aprotinin, 10 µg/ml leupeptin, 2 mm PMSF and 1 mm sodium orthovanadate, adjusted to ph 7.2), sonicated 20 sec and the samples were centrifuged for 7 min at 1000 x g. Unbroken cells and nuclei were discarded and the supernatants were ultracentrifuged at 180,000 x g for 45 min in a Beckman TL-100 ultracentrifuge. Membrane 8
10 pellets were washed once and re-suspended in a small volume of solubilization buffer containing 0.25 M Na 2 HPO 4, 0.3 M NaCl, 2.5% sodium dodecyl sulfate (SDS), 10 µg/ml aprotinin, 10 µg/ml leupeptin and 2 mm PMSF, and samples were assayed for protein content with Pierce Coomassie Brilliant Blue Protein Assay reagent. To determine whether TNF-α can regulate the expression of S1P 3, cells were cultured for 24 h in the presence of TNF-α (100 ng/ml). Protein samples (150 µg) were separated by gradient (7.5-20%) SDSpolyacrylamide gel electrophoresis. Proteins were later transferred from polyacrylamide gel to methanol-soaked Immobilon PVDF membranes (Millipore corporation, Bedford, MA, USA). Primary antibody incubation was performed overnight at 4 C. The membranes were then washed three times and incubated with appropriate horseradish peroxidase-conjugated secondary antibodies at room temperature for 1 h. Membranes were washed three times and antibody-antigen complexes were revealed using ECL according to the manufacturer s instructions (Perkin Elmer Life Sciences, Boston, MA, USA). The ECL signal was visualized by developing the signal on a film. Wound closing assay. Cells were plated at a concentration of cells/ml in 12- well plates. After routine starvation, a plastic pipette tip (200 µl) was drawn across the center of the well to produce a clean wound area. Free cells were removed and the medium was replaced with serum-free medium containing S1P, selective agonists/antagonists of S1P receptors, and the specific inhibitors of p44/42 MAPK, p38 MAPK and Rho kinase at indicated concentrations. Immediately following scratch wounding (0 h) and after incubation for 24 h, the wound closing process was photographed with an inverted microscope (Nikon TE300). The cells that had migrated into the wound area were examined and monitored with MetaMorph software. 9
11 ELISA and Luminex 100 multiplex array assay. Cells plated at a concentration of cells/ml in 24-well plates were stimulated with S1P or the S1P 1 agonist SEW2871 at indicated concentrations or times. Where indicated cells were pre-treated with the S1P receptor antagonists or the inhibitors of p42/44 MAPK, p38 MAPK, and Rho kinase for 30 min prior to stimulation with S1P. To evaluate the effect of TNF-α on S1P-mediated cytokine secretion, cells were pre-treated with TNF-α for 2 h, 8 h, and 24 h and washed extensively prior to stimulation with S1P (5 µm, 24 h). Cell culture supernatants were collected and stored at -80 C until the cytokine ELISAs were performed. IL-8 and IL-6 in all samples were monitored in duplicate, according to the manufacturer s protocol. Optical densities were determined using a SoftMaxPro40 plate reader at 450 nm. The results were compared with a standard curve that was generated using known concentrations (pg/ml) of IL-8 and IL-6. The results were expressed in pg/ml. For the Luminex 100 multiplex array assay, cells were stimulated with S1P (5 µm, 24 h) with or without pre-treatment of TNF-α (100 ng/ml, 8 h) and cell supernatants were collected for multiple cytokine and chemokine analyses. The multiplex bead-based sandwich immunoassay kit was used to measure the levels of IL-1α, IL-1β, IL-8, IL-15, Eotaxin, Granulocyte-monocyte colony-stimulating factor (GM-CSF), interferon-γ-inducible protein 10 (IP-10), monocyte chemotactic protein-1 (MCP-1), macrophage inflammatory protein 1α (MIP-1α), MIP-1β, and regulated on activation normal T cells expressed and secreted (RANTES). Apoptosis assay and proliferation assay. For the apoptosis assay, cells were stimulated with 5 µm S1P or µm SEW2871 for 8 h prior to stimulation with 1 mm SNP for an additional 16 h. Cells were then harvested and apoptosis was evaluated, using Annexin V-FITC labeling according to the manufacturer s instructions. Cells were then 10
12 analyzed by flow cytometry. For the proliferation assay, cells were cultured with or without S1P (1 nm-5 µm) in triplicate in 96-well microplates at concentration of 2 X 10 5 /ml in DMEM without serum. After 96 h, cell viability was assessed by measuring the conversion of the CellTiter 96 Aqueous One Solution Reagent (3-(4,5-dimethythizol-2-yl)-5-(3- carboxymethoxyphenyl)-2-(4-sulfophenyl)-2h-tetrazolium) to a water-soluble formazan product that absorbs light at 490 nm and is proportional to the number of live cells. Statistical analysis. Unless otherwise stated, experiments were performed in triplicate and results presented are expressed as mean values ± SE. In Table 1 and Figure 10C results are expressed as mean values ± SD. Statistical significance of the difference between treated and untreated samples was determined by t-test (two tailed P value). For the dose response and time course studies, statistical significance between untreated samples and samples treated with S1P, S1P 1 receptor agonist, TNF-α, and kinase inhibitors (dose response experiments) and between samples treated at 0 h with those treated at indicated time points (time course study) was determined by ANOVA, Dunnett s Multiple Comparison Test. Calculations were made with the Prism 4.0 software. P values less than 0.05 were considered statistically significant. 11
13 Results S1P 1, S1P 2, and S1P 3 receptor expression in human FLS. As the biological activity of S1P is via interaction with specific cell surface receptors, we examined whether human FLS express all the known receptors for S1P. The expression of S1P 1, S1P 2, and S1P 3 receptor transcripts in FLS was investigated by using both semi-quantitative RT-PCR (Figure 1A) and real-time PCR (Figure 1B). Oligonucleotide primers designed for S1P 4 and S1P 5 did not yield a detectable signal in human FLS but amplified the expected 672-bp (S1P 4 ) and 658-bp (S1P 5 ) bands in human monocytes that were used as positive controls (data not shown). To confirm the expression of these receptors at the protein level, Western blot analysis was performed using purified membrane fractions of FLS. The only antibodies available for Western blot analysis are those for the S1P 1 and S1P 3 receptors. As a positive control, cdnas coding for the HA-tagged S1P 1 and S1P 3 receptors were expressed in CHO cells. As shown in Figure 10C, an expression of S1P 3 (47 kd) was detected in FLS. S1P 1 expression, however, was lower than the threshold for detection by the S1P 1 antibody (data not shown). Induction of human FLS migration by S1P via S1P 1 and/or S1P 3 receptors. S1P is known to induce cell migration. We therefore investigated whether S1P could enhance the migration of FLS using a wound-closing assay. As shown in Figure 2, cell migration was stimulated by S1P at concentration of 1 µm or 5 µm, by 1.7±0.3 fold (p<0.05) and 3.2±0.2 fold (p<0.01) above the non-treated control, respectively. Since FLS express the S1P 1, S1P 2, and S1P 3 receptors, the contribution of each of these receptors to S1P-mediated cell motility was then examined using selective S1P receptor agonists/antagonists. The S1P 1 specific agonist SEW2871 stimulated FLS migration in a concentration-dependent manner (Figure 3A). SEW2871 at 1 µm, 5 µm, and 10 µm 12
14 increased cell motility by 1.8±0.01 fold, 3.3±0.3 fold (p<0.01), and 3.5±0.4 fold (p<0.01) when compared to non-stimulated FLS, respectively. The specific S1P 1/3 antagonist VPC23019 and the S1P 3 antagonist CAY10444 decreased S1P-induced cell motility by 90.8±9.3% (p<0.01) and 84.4±4.7% (p<0.01), respectively (Figure 3B). In contrast, the S1P 2 antagonist JTE-013 had no significant effect on cell motility induced by S1P (p=0.74, Figure 3B). It should be noted that the S1P receptor agonists/antagonists used throughout our study did not affect cell viability, as measured using Annexin V/PI staining (data not shown). S1P mediated production of IL-8 and IL-6 through S1P 2 and S1P 3 in human FLS. Although S1P was recently shown to induce the expression of inflammatory genes in FLS such as COX-2 (14), the release of pro-inflammatory chemokines in response to stimulation of FLS with S1P has not yet been investigated. We chose to investigate IL-8 and IL-6 because IL-8 is one of the most abundant chemokines in RA synovial tissues and both IL-8 and IL-6 are potent chemoattractants that are involved in the recruitment of inflammatory cells into the synovium (19). As shown in Figure 4, a significant release of IL- 8 (Figure 4A) was detected with 2-5 µm S1P (p<0.01 for both 2 µm and 5 µm S1P versus non-treated FLS). S1P-mediated IL-8 (Figure 4C) secretion increased in a linear manner up to 48 h, the last time point tested. S1P also stimulated IL-6 secretion in a concentration- and time-dependent manner (Figure 4B and 4D). The contribution of each S1P receptor in S1P-mediated IL-8 and IL-6 production was then evaluated using specific S1P receptor agonists/antagonists. Interestingly, the S1P 1 agonist SEW2871 had no significant stimulatory effect on IL-8 and IL-6 secretion by FLS (data not shown). In contrast, the S1P 1/3 antagonist, VPC23019, the S1P 2 antagonist JTE-013, and the S1P 3 antagonist CAY10444 significantly decreased S1P-induced IL-8 and IL-6 secretion (Figure 5). S1P-induced IL-8 secretion was inhibited by 58±9% with VPC23019 (p<0.01), 85±3% with JTE-013 (p<0.01), and 82±6% with CAY10444 (p<0.01) (Figure 5A). 13
15 VPC23019, JTE-013 and CAY10444 also inhibited S1P-induced IL-6 secretion by 76±0.6% (p<0.01), 69±3% (p<0.05), and 71±2% (p<0.01), respectively (Figure 5B). Taken together the data suggest a role for S1P 2 and S1P 3 but not S1P 1, in S1P-mediated cytokine/chemokine synthesis. Protective effect of S1P on SNP-mediated FLS apoptosis. Abnormal proliferation and resistance to apoptosis is believed to contribute to both chronic inflammation and hyperplasia of the lining layer (20). Therefore, the next series of experiments was designed to examine the regulation of FLS proliferation and survival by S1P. Under routine serumstarvation conditions, no increase in cell proliferation was observed following stimulation with S1P (1 nm-5 µm) for up to 96 h (data not shown). In contrast, FLS incubated with 5 µm S1P for 8 h reduced FLS apoptosis, induced by the NO donor sodium nitroprusside (SNP), by 28±0.7% (p<0.001, Figure 6A). NO was chosen as a pro-apoptotic agent due to its overproduction in RA joint and its ability to induce apoptosis of FLS in vitro (21, 22). Whereas the protective effect of S1P on SNP-mediated FLS apoptosis was not blocked by a pre-treatment of the cells with the S1P 2 antagonist JTE-013 and the S1P 3 antagonist CAY10444 (data not shown), the S1P 1 agonist SEW2871 significantly reduced SNPmediated FLS cell apoptosis in a concentration-dependent manner (Figure 6B). SEW2871 at 0.3 µm, 1 µm and 3 µm reduced SNP-mediated cell apoptosis by 14.8±6.7 (p<0.05), 47.6±2.9% (p<0.01) and 61.3±7.3% (p<0.01), respectively. The data suggest that S1P 1 receptor protects FLS from SNP-induced apoptosis. Analysis of downstream signaling pathways coupled to S1P functional responses. Activation of S1P receptors induces a range of downstream signaling cascades. These include mitogen-activated protein kinase activation and activation of small GTPases Rho, Rac and Ras, to name a few (23). The pathways and the functional responses are dependent on several variables that include receptor usage, receptor expression level and cell type. We therefore 14
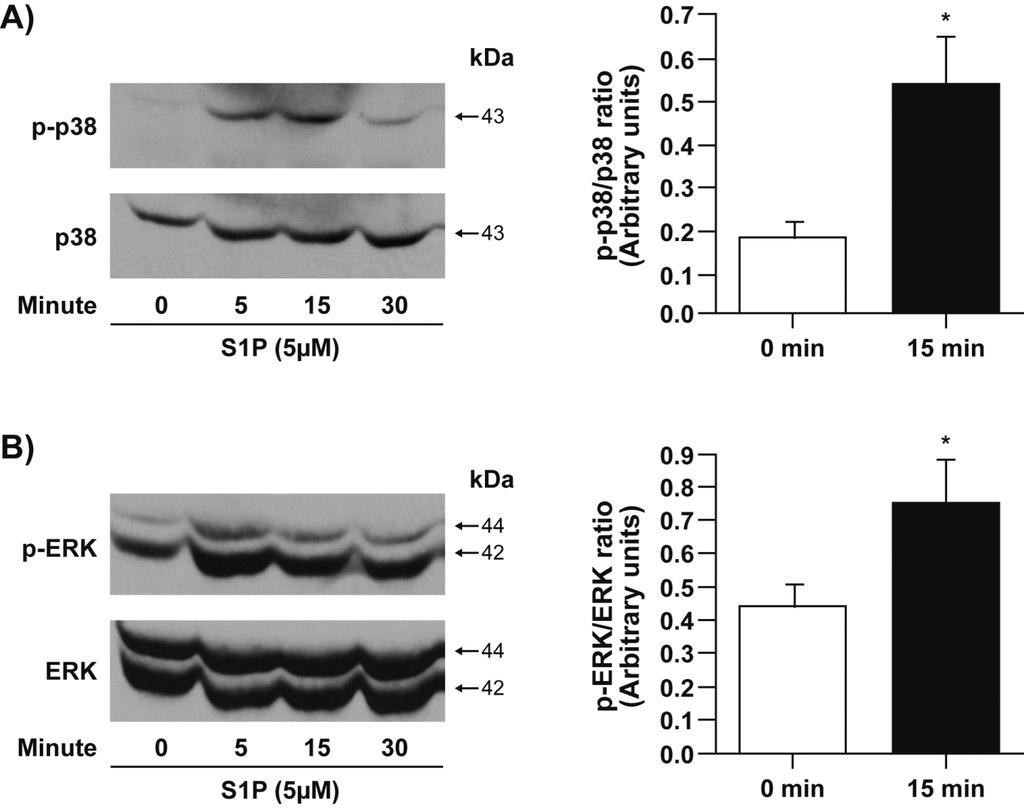
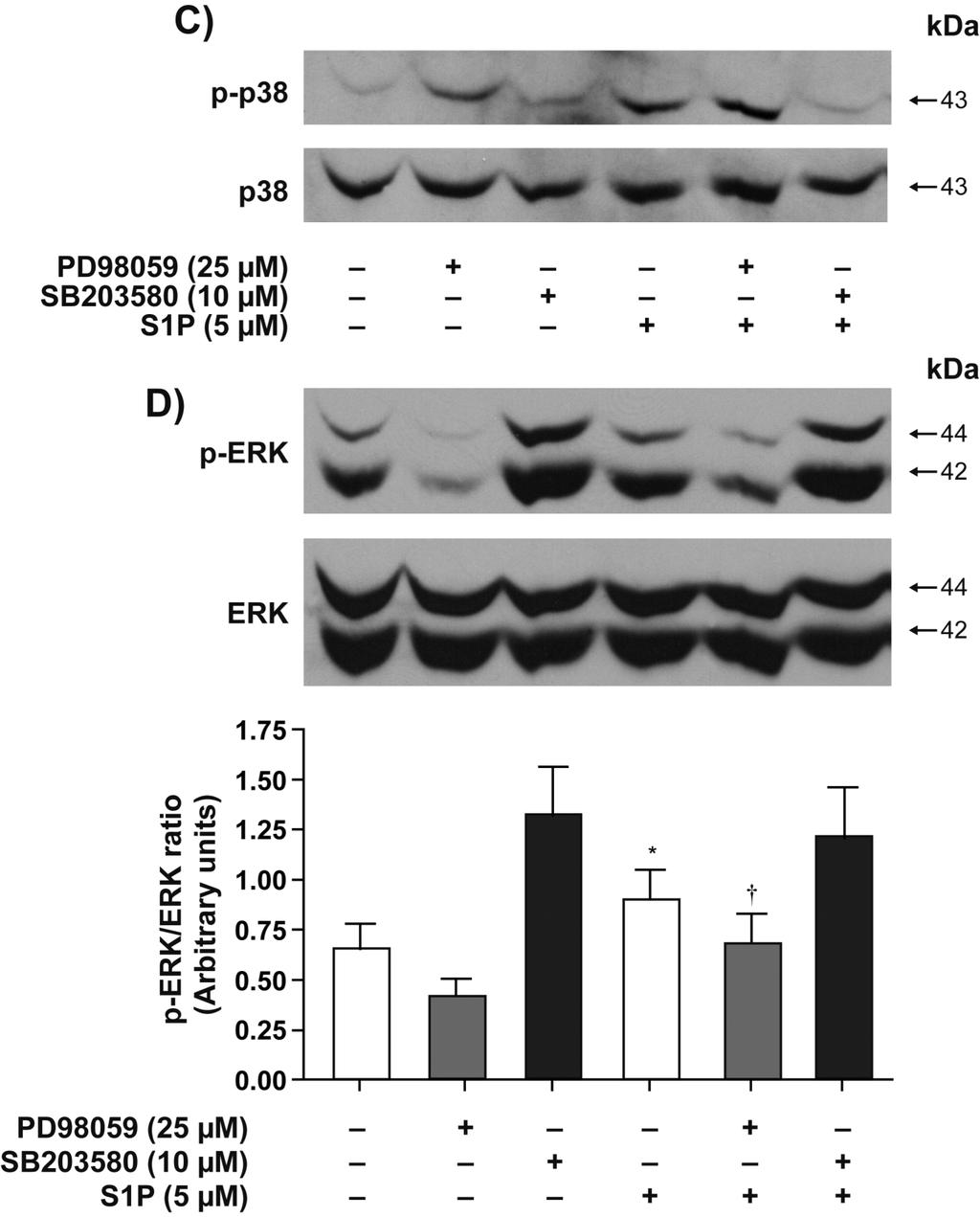
16 explored the involvement of MAPK and Rho kinase pathways in S1P-mediated functional responses. We first assessed the activation state of p42/44 MAPK, p38 MAPK and JNK. S1P enhanced the phosphorylation of p38 and p42/44 MAPK (Figure 7). Phosphorylation of p38 (Figure 7A) and p42/44 MAPK (Figure 7B) peaked at 5 min and 15 min, respectively. No phosphorylation of JNK was detected following stimulation with S1P for up to 30 min (data not shown). As expected, SB blocked S1P-induced phosphorylation of p38 MAPK (Figure 7C) but not that of p42/44 MAPK (Figure 7D). In unstimulated RA synovial cells from RA patients, p42/44 MAPK was phosphorylated (24, 25). PD98059 similarly attenuated basal and S1P-induced phosphorylation of p42/44 MAPK (Figure 7D, p<0.05 for S1P versus S1P+PD98059) but had no significant effect on S1P-induced phosphorylation of p38 MAPK (Figure 7C). Interestingly, the Rho kinase inhibitor Y27632 significantly diminished S1Pinduced activation of p38 MAPK (Figure 7E) but had no effect on S1P-mediated activation of p42/44 MAPK (Figure 7F). The data suggests that p38 MAPK may act, at least partly, downstream of Rho/Rho kinase to promote S1P-induced IL-8 secretion and cell motility. The following experiments addressed the involvement of p38 and p42/44 MAPK and Rho kinase in S1P-induced cell motility. As shown in Figure 8, all three inhibitors attenuated S1P-induced cell migration. PD98059 at 25 µm decreased S1P-induced cell motility by 53.1±2.6% (p<0.01) (Figure 8A). SB at 3 µm and 10 µm decreased S1P-mediated cell migration by 35.5±4.3% (p<0.01) and 79.6±1.6% (p<0.01), respectively (Figure 8B). Y27632 at 1 µm and 10 µm also reduced S1P-mediated cell migration by 61.8±2.4% (p<0.01) and 71.6±6.6% (p<0.01), respectively (Figure 8C). Regarding cytokine production, the inhibitor of p42/44 MAPK PD98059 had marginal inhibitory effect on S1P-induced IL-8 (Figure 9A) and IL-6 secretion (Figure 9D). On the one hand, 3 µm and 10 µm SB reduced S1P-mediated IL-8 secretion by 38.7±4.0% (p<0.01, Figure 9B) and 40.2±3.6% (p<0.01, Figure 9B) and that of IL-6 by 15
17 38.1±6.9% (p<0.01, Figure 9E) and 42±4.1% (p<0.01, Figure 9E), respectively. On the other hand, Y27632 reduced S1P-mediated IL-8 and IL-6 secretion in a dose-dependent manner. At a higher concentration (10 µm), Y27632 reduced S1P-mediated IL-8 and IL-6 secretion by 96.9±0.2% (p<0.01, Figure 9C) and 83.6±1.7% (p<0.01, Figure 9F), respectively. It is worthy to note that the MAPK and Rho kinase inhibitors used throughout our study did not affect cell viability, as measured using Annexin V/PI staining (data not shown). Regulation of S1P 3 receptor expression by TNF-α. Given that TNF-α is a key inflammatory molecule in RA (26), we analyzed the expression profile of S1P receptors in response to TNF-α by RT-PCR, real-time PCR and Western blot analyses. Treatment of FLS with TNF-α up-regulated the expression of S1P 3 mrna (Figure 10A and 10B) in a concentration dependent manner. A 3.1±0.5 (p<0.01) fold increase in S1P 3 expression was observed with 100 ng/ml of TNF-α, the highest concentration tested. Under the same conditions, S1P 1 and S1P 2 expression was not altered by TNF-α (data not shown). Upregulation by TNF-α of S1P 3 expression at the protein level (1.8±0.5 fold, n=2) was confirmed by Western blot (Figure 10C). Effect of TNF-α on S1P-induced cytokine/chemokine secretion in human FLS. The next series of experiments was designed to examine the effect of a proinflammatory environment, established by a pre-treatment of FLS with TNF-α, on S1P-induced secretion of cytokine/chemokine that are relevant to RA pathogenesis (27). Starved FLS were treated with S1P (5 µm) for 24 h with or without a pre-treatment of TNF-α (100 ng/ml) for 8 h. Cell culture supernatants were analyzed using a Luminex 100 array assay (Table 1). Among the 11 cytokine/chemokine tested, IL-1α, IL-1β, IL-15, Eotaxin, GM-CSF, MIP-1α, and MIP-1β were not secreted in response to S1P, TNF-α, or their combination. In contrast, significant amounts of IL-8, MCP-1, and RANTES were secreted following stimulation with S1P and, 16
18 most importantly, S1P-mediated cytokine synthesis was strongly enhanced in TNF-α-primed FLS. The release of IL-8, MCP-1, and RANTES was increased 5.0±0.3 (p<0.01), 1.7±0.2 (p<0.05), and 66±6.4 (p<0.01) fold, respectively, in TNF-α-primed and S1P stimulated samples as compared to unprimed cells stimulated with S1P. Moreover, S1P or TNF-α priming did not stimulate the secretion of IP-10, but their combination resulted in a significant secretion of this chemokine. As shown in Figure 11A, after priming FLS with TNF-α for 2 h, 8 h and 24 h, S1Pinduced IL-8 secretion was super-induced. To determine the relevance of the S1P receptor(s) to this process, FLS were treated with the S1P 1/3 receptor antagonist, VPC VPC23019 had no effect on the release of IL-8 by TNF-α-primed FLS, but almost completely inhibited the enhanced secretion of cytokine induced by S1P in these cells (Figure 11B). S1P-mediated super-production of IL-8 in TNF-α-primed cells was also completely abolished by a selective S1P 3 antagonist, CAY10444 (Figure 11C). The data suggest that TNF-α enhances S1P receptor (S1P 3 ) expression and S1P 3 -dependent responses in human FLS. 17
19 Discussion S1P has been previously implicated in RA due to the expression of the S1P 1 and S1P 3 receptors in the synovium of RA patients (14). Moreover, S1P was shown to potentiate cytokine-induced COX-2 expression and PGE 2 production (14). S1P, however, mediates a wide variety of cellular responses including cell migration, apoptosis, cell proliferation and differentiation. The effect of S1P on these cellular processes in FLS has not been investigated. Herein, we report that S1P stimulates FLS migration, and cytokine/chemokine synthesis, and that S1P protects FLS from apoptosis. The S1P receptors mediating these effects were also identified. Moreover, we provide direct evidence for the modulation of S1Pinduced cytokine secretion by TNF-α. Our expression analysis also revealed that FLS not only express the S1P 1 and S1P 3 receptors but also express S1P 2. The observation that S1P induces FLS migration is significant in the context of RA since one of the hallmarks of RA is synovial hyperplasia. This pro-migratory effect of S1P was mimicked by the S1P 1 specific agonist SEW2871 but abolished by the S1P 1/3 antagonist VPC23019 or the S1P 3 selective antagonist CAY The results suggest a prevailing role for S1P 1 and S1P 3 receptors in this process. These observations corroborate with previous reports that identified a role for S1P 1 and S1P 3 in the migratory response toward S1P in other cell types (28-30). Since FLS express S1P 2 and this receptor is known to decrease growth factor-mediated cell migration (31), we hypothesized that blocking this receptor would increase cell migration. Our results reveal, however, that blocking the S1P 2 receptor with the selective antagonist JTE-013 had no significant effect on S1P-induced FLS migration. These data suggest that the activation of S1P 2 by S1P does not act as a negative regulator of S1Pinduced migration in human FLS. Whether S1P 2 can counteract growth factor-, CC or CXC chemokine-mediated migration of FLS remains to be elucidated. 18
20 A key feature of RA is the infiltration of immune cells such as neutrophils, monocytes and T lymphocytes into the joints due to the large amount of CC and CXC chemokines, (32-34) produced by activated cells of the synovial lining. In particular, IL-8 exhibits selective chemotactic activity for neutrophils, whereas MCP-1, MIP-1α, -1β and RANTES primarily attract monocytes (19). To gain insight into additional molecular mechanisms by which S1P may contribute to RA pathogenesis, we profiled the secretion of inflammatory CC and CXC chemokines by FLS stimulated with S1P. FLS do not secret detectable levels of cytokines or chemokines, except for low amounts of MCP-1 in the resting state. Upon treatment with S1P, the secretion of IL-8, IL-6, MCP-1 and RANTES was strongly induced, thereby suggesting that S1P can contribute to and/or amplify the secretion of chemokines by cells of the inflamed synovium. Since immune cells express a wide repertoire of chemokine receptors, including IL-8, MCP-1, SDF-1α, IP-10 and RANTES (35), our results suggest that S1Pmediated cytokine secretion may contribute to the recruitment and retention of inflammatory cells in RA. Chemokines such as MCP-1, SDF-1α, IP-10 and RANTES enhance the migration and proliferation of FLS and upregulate matrix metalloproteinase production by FLS (27), indicating a direct role for FLS in the destructive phase of RA beyond the regulation of immune cell trafficking. Studies using S1P receptor agonists/antagonists revealed that S1P 2 and S1P 3 are likely involved in S1P-mediated cytokine/chemokine secretion in FLS. The ability of the S1P 1 agonist SEW2871 to induce cell migration but not cytokine/chemokine secretion implies that S1P-driven FLS migration is independent of and not secondary to synthesis of CC or CXC chemokines (27). In contrast to the induction of FLS migration and cytokine/chemokine secretion by S1P, this bioactive lipid had no effect on FLS proliferation (data not shown). Our observations differ from those of Kitano et al. who reported that S1P induces FLS 19
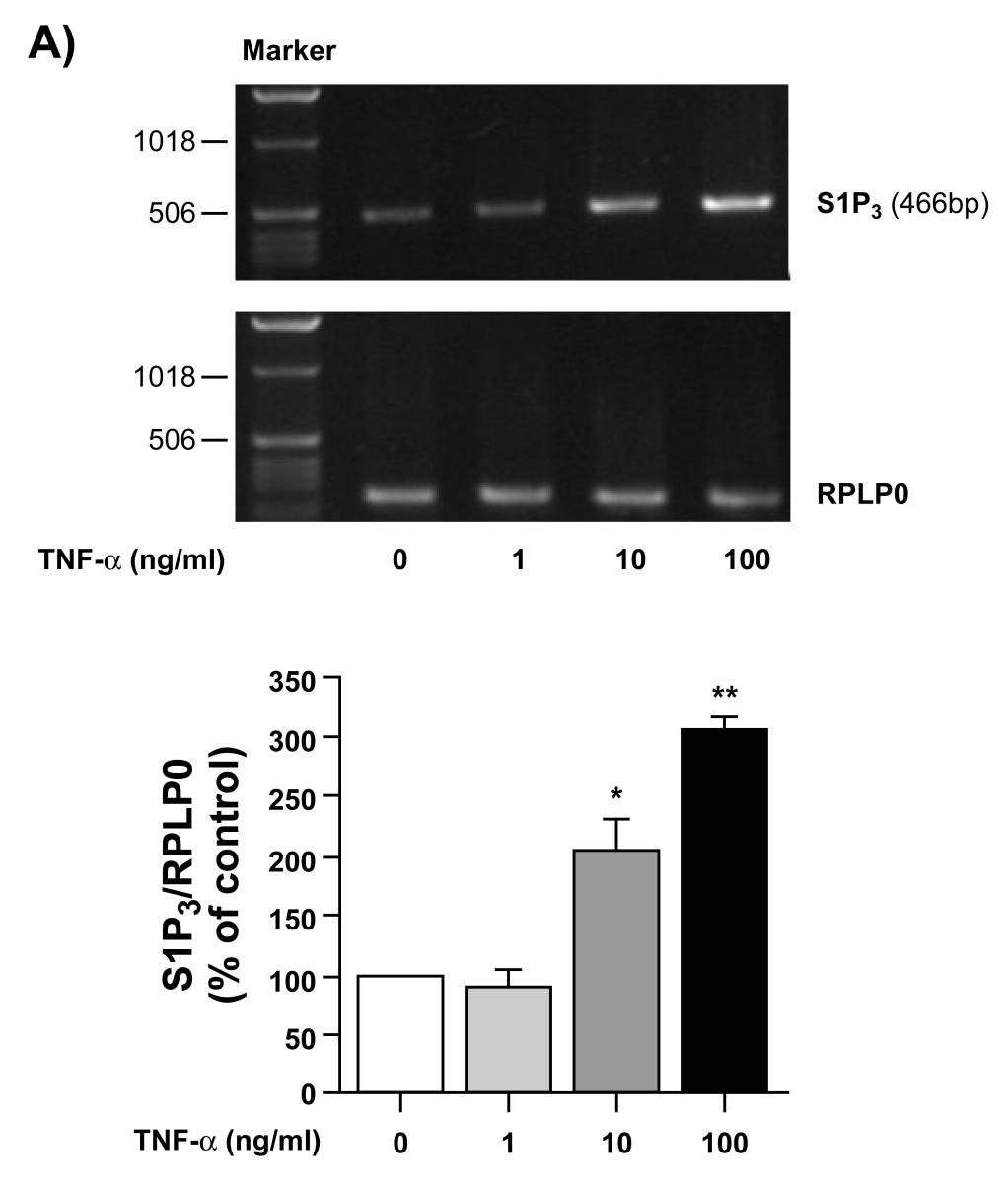
21 proliferation (14). The discrepancy between these two observations may be partly explained by the different experimental conditions. Kitano et al. performed their experiments in the presence of serum as opposed to our serum-starved culture of FLS during the proliferation assay. Since proliferation is scarce in RA synovium (36), dysregulation of apoptosis has been proposed to explain synovial hyperplasia (37). We also provide direct evidence for the inhibition of FLS apoptosis by S1P. Indeed, S1P appears capable of increasing cell survival and inhibiting apoptosis of various other cell types (38-40), including B lymphoblastoid cells derived from patients with RA (16). TNF-α plays a key role in the pathogenesis of RA. Khoa et al. reported that TNF-α can cross-talk with the G protein-coupled receptor adenosine A 2A receptor (A 2A R) (41). Since S1P receptors are coupled to G proteins, we sought to determine whether the cross-talk between TNF-α and G protein-coupled receptors is a general phenomenon. We provide direct evidence for the significant enhancement of S1P-induced cytokine/chemokine secretion, including IL-6, IL-8, MCP-1, and RANTES, in FLS primed with TNF-α. Moreover, S1P and TNF-α alone were not able to stimulate IP-10 secretion. Their combination, however, resulted in an impressive induction of IP-10 production, suggestive of cross-talk between TNF-α and S1P receptors. These observations corroborate with the findings of Kitano et al. who reported that S1P and TNF-α signaling pathways synergize (14). The increase in S1Pinduced cytokine secretion by TNF-α can be partly explained by the induction of S1P 3 expression by TNF-α. Although a large number of genes up-regulated by TNF-α have been identified in FLS (26), this is the first report of the regulation of S1P 3 receptor expression by TNF-α. This raises the possibility of a causal relationship between enhanced expression of S1P 3 receptor and production of chemokine by S1P following priming of FLS with TNF-α. 20
22 Indeed, the S1P 1/3 receptor antagonist VPC23019 and the selective S1P 3 antagonist CAY10444 did not reduce TNF-α-induced cytokine secretion but totally blocked that induced by S1P in TNF-α-primed FLS. Together, these observations suggest that the proinflammatory environment potentiates some of the functional outcomes of the S1P/S1P receptor signaling pathway. To identify the signaling molecules involved in the functional responses of FLS to S1P, the activation of the classical signaling proteins involved in G protein-coupled receptor signaling was investigated. We suggest that the main pathway that regulates S1P-induced IL- 8 and IL-6 secretion is Rho/Rho kinase related, while p38 MAPK is also involved in this process. On the other hand, p42/44 MAPK, p38 MAPK and Rho kinase but not JNK are all involved in S1P-induced FLS motility. The results suggest that the coupling of S1P receptors to various heterotrimeric G proteins and consequently distinct downstream signaling pathways lead to downstream pathological phenomena, such as FLS invasion into cartilage and bone and recruitment of immune cells into the inflamed synovium. We previously reported that a lysophospholipid that is structurally related to S1P, lysophosphatidic acid (LPA), can also stimulate FLS cytokine secretion and migration by interacting with its cognate receptors (42). The functional responses induced by LPA, however, differ from those induced by S1P in that LPA-induced cytokine synthesis is strongly dependent on p42/44 MAPK activity whereas signal transduction through p42/44 MAPK is dispensable for LPAmediated FLS migration. As reported previously (24, 25), p42/44 MAPK is slightly phosphorylated in synovial cells from RA patients, even in the absence of stimulation. Basal levels of p42/44 MAPK may result from autocrine activation by unknown stimulatory factor(s) released by synoviocytes in culture. In summary, the present study provides evidence that S1P/S1P receptor signaling may contribute to RA pathogenesis by stimulating FLS migration, cytokine/chemokine secretion 21
23 and inhibiting apoptosis. Though S1P receptors exert receptor subtype-specific responses, we also demonstrated a redundancy of function of the three S1P receptors expressed by FLS. Furthermore, our data suggest that up-regulation of S1P 3 receptor expression and enhanced S1P-induced cytokine secretion by TNF-α-primed FLS may amplify the inflammatory process in RA. Moreover, our results are suggestive of a role for S1P/S1P receptor signaling in the development and progression of RA, and that S1P 3 may represent a critical player in the events that take place in the RA synovium. Thus, subtype-specific antagonists for S1P receptors could be novel therapeutic modalities for limiting inflammation in the destructive phase of RA. 22
24 Acknowledgements We would like to thank the Bioimaging Platform of the Centre de Recherche en Infectiologie, Centre de Recherche du CHUQ-CHUL, for Luminex analysis. This project is supported by research grants from Canadian Institutes of Health Research and the Arthritis Society of Canada (S.G.B.) and the Canadian Arthritis Network Post-Doctoral Fellowship Award (C.Z.). 23
25 References 1. Pap, T., U. Muller-Ladner, R.E. Gay, and S. Gay Fibroblast biology. Role of synovial fibroblasts in the pathogenesis of rheumatoid arthritis. Arthritis Res 2: Feldmann, M., F.M. Brennan, and R.N. Maini Rheumatoid arthritis. Cell 85: Muller-Ladner, U., J. Kriegsmann, B.N. Franklin, S. Matsumoto, T. Geiler, R.E. Gay, and S. Gay Synovial fibroblasts of patients with rheumatoid arthritis attach to and invade normal human cartilage when engrafted into SCID mice. Am J Pathol 149: Kluk, M.J., and T. Hla Signaling of sphingosine-1-phosphate via the S1P/EDG-family of G-protein-coupled receptors. Biochim Biophys Acta 1582: Rosenfeldt, H.M., J.P. Hobson, S. Milstien, and S. Spiegel The sphingosine-1- phosphate receptor EDG-1 is essential for platelet-derived growth factor-induced cell motility. Biochem Soc Trans 29: Hla, T Physiological and pathological actions of sphingosine 1-phosphate. Semin Cell Dev Biol 15: Yatomi, Y., F. Ruan, S. Hakomori, and Y. Igarashi Sphingosine-1-phosphate: a platelet-activating sphingolipid released from agonist-stimulated human platelets. Blood 86: Spiegel, S., and S. Milstien Sphingosine 1-phosphate, a key cell signaling molecule. J Biol Chem 277: Yang, L., Y. Yatomi, Y. Miura, K. Satoh, and Y. Ozaki Metabolism and functional effects of sphingolipids in blood cells. Br J Haematol 107:
26 10. Xia, P., L. Wang, J.R. Gamble, and M.A. Vadas Activation of sphingosine kinase by tumor necrosis factor-alpha inhibits apoptosis in human endothelial cells. J Biol Chem 274: Lin, D.A., and J.A. Boyce Lysophospholipids as mediators of immunity. Adv Immunol 89: Lin, C.I., C.N. Chen, P.W. Lin, and H. Lee Sphingosine 1-phosphate regulates inflammation-related genes in human endothelial cells through S1P1 and S1P3. Biochem Biophys Res Commun 355: Dorsam, G., M.H. Graeler, C. Seroogy, Y. Kong, J.K. Voice, and E.J. Goetzl Transduction of multiple effects of sphingosine 1-phosphate (S1P) on T cell functions by the S1P1 G protein-coupled receptor. J Immunol 171: Kitano, M., T. Hla, M. Sekiguchi, Y. Kawahito, R. Yoshimura, K. Miyazawa, T. Iwasaki, H. Sano, J.D. Saba, and Y.Y. Tam Sphingosine 1- phosphate/sphingosine 1-phosphate receptor 1 signaling in rheumatoid synovium: regulation of synovial proliferation and inflammatory gene expression. Arthritis Rheum 54: Spiegel, S., and S. Milstien Sphingosine-1-phosphate: an enigmatic signalling lipid. Nat Rev Mol Cell Biol 4: Pi, X., S.Y. Tan, M. Hayes, L. Xiao, J.A. Shayman, S. Ling, and J. Holoshitz Sphingosine kinase 1-mediated inhibition of Fas death signaling in rheumatoid arthritis B lymphoblastoid cells. Arthritis Rheum 54: Faour, W.H., A. Mancini, Q.W. He, and J.A. Di Battista T-cell-derived interleukin-17 regulates the level and stability of cyclooxygenase-2 (COX-2) mrna through restricted activation of the p38 mitogen-activated protein kinase cascade: role 25
27 of distal sequences in the 3'-untranslated region of COX-2 mrna. J Biol Chem 278: Marcil, J., D. Harbour, M.G. Houle, P.H. Naccache, and S. Bourgoin Monosodium urate-crystal-stimulated phospholipase D in human neutrophils. Biochem J 337 (Pt 2): Koch, A.E Chemokines and their receptors in rheumatoid arthritis: future targets? Arthritis Rheum 52: Ospelt, C., M. Neidhart, R.E. Gay, and S. Gay Synovial activation in rheumatoid arthritis. Front Biosci 9: Aupperle, K.R., D.L. Boyle, M. Hendrix, E.A. Seftor, N.J. Zvaifler, M. Barbosa, and G.S. Firestein Regulation of synoviocyte proliferation, apoptosis, and invasion by the p53 tumor suppressor gene. Am J Pathol 152: Borderie, D., P. Hilliquin, A. Hernvann, H. Lemarechal, C.J. Menkes, and O.G. Ekindjian Apoptosis induced by nitric oxide is associated with nuclear p53 protein expression in cultured osteoarthritic synoviocytes. Osteoarthritis Cartilage 7: Anliker, B., and J. Chun Cell surface receptors in lysophospholipid signaling. Semin Cell Dev Biol 15: Ha, J.E., Y.E. Choi, J. Jang, C.H. Yoon, H.Y. Kim, and Y.S. Bae FLIP and MAPK play crucial roles in the MLN51-mediated hyperproliferation of fibroblast-like synoviocytes in the pathogenesis of rheumatoid arthritis. Febs J 25. Harigai, M., M. Hara, M. Kawamoto, Y. Kawaguchi, T. Sugiura, M. Tanaka, M. Nakagawa, H. Ichida, K. Takagi, S. Higami-Ohsako, K. Shimada, and N. Kamatani Amplification of the synovial inflammatory response through activation of mitogen-activated protein kinases and nuclear factor kappab using ligation of CD40 26
28 on CD14+ synovial cells from patients with rheumatoid arthritis. Arthritis Rheum 50: Taberner, M., K.F. Scott, L. Weininger, C.R. Mackay, and M.S. Rolph Overlapping gene expression profiles in rheumatoid fibroblast-like synoviocytes induced by the proinflammatory cytokines interleukin-1 beta and tumor necrosis factor. Inflamm Res 54: Garcia-Vicuna, R., M.V. Gomez-Gaviro, M.J. Dominguez-Luis, M.K. Pec, I. Gonzalez-Alvaro, J.M. Alvaro-Gracia, and F. Diaz-Gonzalez CC and CXC chemokine receptors mediate migration, proliferation, and matrix metalloproteinase production by fibroblast-like synoviocytes from rheumatoid arthritis patients. Arthritis Rheum 50: Okamoto, H., N. Takuwa, T. Yokomizo, N. Sugimoto, S. Sakurada, H. Shigematsu, and Y. Takuwa Inhibitory regulation of Rac activation, membrane ruffling, and cell migration by the G protein-coupled sphingosine-1-phosphate receptor EDG5 but not EDG1 or EDG3. Mol Cell Biol 20: Balthasar, S., J. Samulin, H. Ahlgren, N. Bergelin, M. Lundqvist, E.C. Toescu, M.C. Eggo, and K. Tornquist Sphingosine 1-phosphate receptor expression profile and regulation of migration in human thyroid cancer cells. Biochem J 398: Ishii, I., N. Fukushima, X. Ye, and J. Chun Lysophospholipid receptors: signaling and biology. Annu Rev Biochem 73: Goparaju, S.K., P.S. Jolly, K.R. Watterson, M. Bektas, S. Alvarez, S. Sarkar, L. Mel, I. Ishii, J. Chun, S. Milstien, and S. Spiegel The S1P2 receptor negatively regulates platelet-derived growth factor-induced motility and proliferation. Mol Cell Biol 25:
29 32. Lin, F., C.M. Nguyen, S.J. Wang, W. Saadi, S.P. Gross, and N.L. Jeon Effective neutrophil chemotaxis is strongly influenced by mean IL-8 concentration. Biochem Biophys Res Commun 319: Patel, D.D., J.P. Zachariah, and L.P. Whichard CXCR3 and CCR5 ligands in rheumatoid arthritis synovium. Clin Immunol 98: Arend, W.P., and J.M. Dayer Cytokines and cytokine inhibitors or antagonists in rheumatoid arthritis. Arthritis Rheum 33: Chen, X., J.J. Oppenheim, and O.M. Howard Chemokines and chemokine receptors as novel therapeutic targets in rheumatoid arthritis (RA): inhibitory effects of traditional Chinese medicinal components. Cell Mol Immunol 1: Lalor, P.A., P.I. Mapp, P.A. Hall, and P.A. Revell Proliferative activity of cells in the synovium as demonstrated by a monoclonal antibody, Ki67. Rheumatol Int 7: Baier, A., I. Meineckel, S. Gay, and T. Pap Apoptosis in rheumatoid arthritis. Curr Opin Rheumatol 15: Donati, C., F. Cencetti, P. Nincheri, C. Bernacchioni, S. Brunelli, E. Clementi, G. Cossu, and P. Bruni Sphingosine 1-phosphate mediates proliferation and survival of mesoangioblasts. Stem Cells 25: Goetzl, E.J., Y. Kong, and B. Mei Lysophosphatidic acid and sphingosine 1- phosphate protection of T cells from apoptosis in association with suppression of Bax. J Immunol 162: Kwon, Y.G., J.K. Min, K.M. Kim, D.J. Lee, T.R. Billiar, and Y.M. Kim Sphingosine 1-phosphate protects human umbilical vein endothelial cells from serumdeprived apoptosis by nitric oxide production. J Biol Chem 276:
30 41. Khoa, N.D., M. Postow, J. Danielsson, and B.N. Cronstein Tumor necrosis factor-alpha prevents desensitization of Galphas-coupled receptors by regulating GRK2 association with the plasma membrane. Mol Pharmacol 69: Zhao, C., M.J. Fernandes, G.D. Prestwich, M. Turgeon, J. Di Battista, T. Clair, P.E. Poubelle, and S.G. Bourgoin Regulation of lysophosphatidic Acid receptor expression and function in human synoviocytes: implications for rheumatoid arthritis? Mol Pharmacol 73:
31 Legends for Figures Fig. 1. Expression of S1P 1, S1P 2, and S1P 3 mrna in human FLS. Semi-quantitative RT- PCR (A) and real-time PCR (B) analyses of S1P 1, S1P 2, and S1P 3 mrna expression in FLS. Total RNA from resting state FLS was extracted for RT-PCR or real-time PCR. The reaction performed without oligonucleotide primers was used as a negative control and RPLP0 was used as an internal control. Data shown are representative of three separate experiments. Fig. 2. Stimulation of human FLS motility by S1P. A clean wound area was made on a monolayer of FLS. After removing free cells, the wound was allowed to close for 24 h in serum-free medium containing S1P (0.1-5 µm). The wound closing process was photographed at 0 h and at 24 h (upper panels). The data shown are representative of three separate experiments. The migration index corresponds to the percentage of cells that migrated in the presence of S1P over that of non-treated cells (lower panel). For statistical comparative analyses we compared cells treated with and without S1P. Data shown are means ± SE of three independent experiments. (*p < 0.05; **p < 0.01) Fig. 3. Effect of selective S1P receptor agonists/antagonists on FLS motility. (A) Effect of S1P 1 agonist SEW2871 on cell motility. After scratching the monolayer and removing free cells, the wound was allowed to close for 24 h in serum-free medium containing SEW2871 (1-10 µm). The wound closing process was photographed at 0 h and at 24 h (upper panels). The data shown are representative of three separate experiments. Migrated cell numbers were expressed as percentage of non-treated cells (lower panel). For statistical comparative analyses we compared cells treated with and without SEW2871. (B) Effect of the S1P 1/3 antagonist VPC23019, S1P 2 antagonist JTE-013, and S1P 3 antagonist CAY10444 on cell motility. The wound was allowed to close for 24 h in serum-free medium containing S1P (5 30
32 µm) in the presence or absence of VPC23019 (5 µm), JTE-013 (5 µm), and CAY10444 (5 µm). Migrated cell numbers were expressed as percentage of non-treated cells. For statistical comparative analyses, samples treated with S1P were compared to those treated with S1P+VPC23019/JTE-013/CAY Data shown are means ± SE of three independent experiments. (**p < 0.01) Fig. 4. S1P-mediated IL-8 and IL-6 secretion. (A, B) Dose-response effect of S1P on IL-8/IL- 6 secretion. FLS were treated with the indicated concentrations of S1P for 24 h before collecting supernatants for cytokine quantification. (C, D) Kinetics of IL-8/IL-6 secretion. Cells were incubated with S1P (5 µm) for the indicated times and the cell culture supernatants were collected for the ELISA assay. Experiments were repeated three times and the results are displayed as mean value ± SE. For statistical analyses samples incubated with diluents were compared to those treated with S1P at indicated concentrations (A, B), or times (C, D). ((*p < 0.05; **p < 0.01) Fig. 5. Effect of selective S1P receptor antagonists on S1P-induced IL-8 and IL-6 secretion. Cells were incubated with or without 5 µm VPC23019, JTE-013, and CAY10444 for 30 min prior to stimulation with S1P (5 µm) for 24 h. Cell culture supernatants were collected and IL-8 (A) and IL-6 (B) were quantified by ELISA. The results are displayed as mean value ± SE (n=3). For statistical comparative analyses we compared the samples stimulated with S1P to those treated with S1P+VPC23019/JTE-013/CAY (*p < 0.05; **p < 0.01) Fig. 6. Protection of FLS from apoptosis by S1P and the S1P 1 agonist SEW2871. Human FLS were pre-treated for 8 h with 5 µm S1P (A) or the indicated concentrations of the S1P 1 receptor agonist SEW2871 (B) prior to the addition of 1mM SNP for 16 h. Apoptotic cells 31
33 were analyzed by Annexin V-propidium iodide (PI) labeling. Total apoptotic cells represent the cells that were Annexin V+ /PI+ and Annexin V+/PI-. Early apoptotic cells correspond to the percentage of cells that were Annexin V+ /PI-. The results are displayed as mean value ± SE (n=3). For statistical comparative analyses we compared samples incubated with SNP to those treated with SNP+S1P/SEW2871. (*p < 0.05; **p < 0.01) Fig. 7. S1P-induced activation of p42/44mapk and p38 MAPK. (A) Effect of S1P on the activation of p38 MAPK. Cells were stimulated with S1P (5 µm) for up to 30 min, and cell lysates were analyzed by Western blot analysis. Blot shown is a representative example from three independent experiments with similar results. Bands corresponding to phosphorylated p38 (15 min time point) were quantified densitometrically, and were normalized with respect to the total amounts of p38 (n=4). Data are shown as the ratio of arbitrary units for the phosphorylated p38 to total p38. (B) Effect of S1P on the activation of p42/44 MAPK. Cells were stimulated with S1P (5 µm) for up to 30 min, and cell lysates were analyzed by Western blot analysis. Blot shown is a representative example from three independent experiments with similar results. Bands corresponding to phosphorylated p42/44 (15 min time point) were quantified densitometrically, and were normalized with respect to the total amounts of p42/44 (n=4). Data are shown as the ratio of arbitrary units for the phosphorylated p42/44 to total p42/44. (C-F) Effect of p42/44 MAPK, p38 MAPK, and Rho kinase inhibitors on S1P-induced activation of p42/44 MAPK and p38 MAPK. Cells were pretreated with 25 µm PD98059 (C, D), 10 µm SB (C, D) and 10 µm Y27632 (E, F) for 30 min prior to stimulation with 5 µm S1P for 15 min. Cell lysates were analyzed by Western blot analysis. Data shown are representative of three separate experiments. In panel D, bands corresponding to phosphorylated p42/44 were quantified densitometrically, and were normalized with respect to the total amounts of p42/44. For statistical comparative analyses we compared samples of 32
34 non-treated to those treated with S1P (*p < 0.05), and samples treated with S1P to those treated with S1P+PD98059 ( p < 0.05). Fig. 8. MAPKs and Rho kinase regulation on S1P-induced FLS migration. After scratching the monolayer and removing free cells, the wound was allowed to close for 24 h in serumfree medium containing 5 µm S1P, with or without PD98059 (A), SB (B), or Y27632 (C), at the indicated concentrations. Migrated cell numbers were expressed as percentage of non-treated cells. Data shown are means ± SE of three independent experiments. For statistical comparative analyses the samples treated with S1P and S1P+inhibitors were compared. (**p < 0.01) Fig. 9. Role of p42/44 MAPK, p38 MAPK and Rho kinase in S1P-induced cytokine secretion. Cells were treated with S1P (5 µm) for 24 h, in the presence or absence of PD98059 (A, D), SB (B, E), or Y27632 (C, F), at the indicated concentrations. Secreted IL-8 (A-C) and IL-6 (D-F) were quantified by ELISA. Data shown are means ± SE of three independent experiments. For statistical comparative analyses the samples incubated with S1P and S1P+inhibitors were compared. (*p < 0.05; **p < 0.01) Fig. 10. Regulation of S1P 3 receptor expression by TNF-α. (A) Concentration-dependent response of S1P 3 mrna expression to TNF-α (RT-PCR). Cells were treated with TNF-α (2 h, at indicated concentrations) prior to RNA extraction and RT-PCR analyses. Results are presented as a representative agarose gel (upper panels) and/or as ratios (means ± SE, n=3) of S1P 3 and RPLP0 (lower panels). (B) Real-time PCR analysis of S1P 3 mrna expression upon TNF-α treatment. Cells were incubated with TNF-α (100 ng/ml) for 2 h prior to RNA extraction and real time PCR analyses. (C) Western blot analysis of S1P 3 protein expression 33
35 upon TNF-α treatment. Cells were incubated with or without TNF-α (100 ng/ml) for 24 h. Cell lysates were subjected to % gradient SDS/PAGE and samples were probed with antibodies to S1P 3, HA-tag and flotillin. Blot shown is a representative example from two independent experiments with similar results (upper panel). Bands corresponding to S1P 3 were quantified densitometrically, and their levels were normalized with respect to flotillin. Data are shown as the ratio of arbitrary units for S1P 3 to flotillin and as means ± SD (n=2, lower panel). For statistical comparative analysis we compared cells of non-treated (NT) to those treated with TNF-α. (*p < 0.05; **p < 0.01) Fig. 11. Super-induction of S1P-induced IL-8 secretion by cell priming with TNF-α. (A) Kinetics of TNF-α pre-treatment on S1P-induced IL-8 secretion. Cells were pre-treated with TNF-α (100 ng/ml) for different time lengths as indicated prior to stimulation with 5 µm S1P for 24 h. Cell culture supernatants were harvested for IL-8 measurement. Results are presented as means ± SE (n=3). For statistical comparative analyses we compared the samples incubated with S1P to those treated with S1P+TNF-α. (**p < 0.01; *** p < 0.001) (B, C) Effect of S1P 1/3 antagonist VPC23019 and S1P 3 antagonist CAY10444 on S1P-induced super production of IL-8 after TNF-α priming. Cells were stimulated with TNF-α (100 ng/ml) for 8 h before stimulation with S1P (5 µm) for another 24 h in the presence/absence of 5 µm of VPC23019 and CAY The results are presented as means ± SE (n=3). For statistical comparative analyses we compared the samples treated with TNFα+S1P to those incubated with TNF-α+S1P+VPC23019/CAY (*p < 0.05; *** p < 0.001) 34
36 Table 1. Effect of TNF-α priming on S1P-induced cytokine/chemokine secretion. Cells were pre-treated with TNF-α (100 ng/ml) for 8 h prior to stimulation with 5 µm S1P for 24 h as described in Materials and Methods. Cell culture supernatants were harvested to monitor cytokine/chemokine secretion using Luminex. Results are presented as means ± SD (n=2). For statistical comparative analyses we compared non-treated cells to S1P/TNF-α/TNFα+S1P-treated cells, respectively. (*p < 0.05; ** p < 0.01) Table 2. S1P receptor agonists/antagonists. 35
37 36

38 Figure 1 37

39 Figure 2 38

40 Figure 3 39
41 Figure 4 40
42 Figure 5 41

43 Figure 6 42
44 43
45 44

46 Figure 7 45

47 Figure 8 46

48 Figure 9 47
49 48
Potential Role of Sphingosine 1-Phosphate in the. Pathogenesis of Rheumatoid Arthritis
 Potential Role of Sphingosine 1-Phosphate in the Pathogenesis of Rheumatoid Arthritis COMMENTARY for Zhao, C., Fernandes, M.J., Turgeon, M., Tancrede, S., Di Battista, J., Poubelle, P.E. and Bourgoin,
Potential Role of Sphingosine 1-Phosphate in the Pathogenesis of Rheumatoid Arthritis COMMENTARY for Zhao, C., Fernandes, M.J., Turgeon, M., Tancrede, S., Di Battista, J., Poubelle, P.E. and Bourgoin,
CD31 5'-AGA GAC GGT CTT GTC GCA GT-3' 5 ' -TAC TGG GCT TCG AGA GCA GT-3'
 Table S1. The primer sets used for real-time RT-PCR analysis. Gene Forward Reverse VEGF PDGFB TGF-β MCP-1 5'-GTT GCA GCA TGA ATC TGA GG-3' 5'-GGA GAC TCT TCG AGG AGC ACT T-3' 5'-GAA TCA GGC ATC GAG AGA
Table S1. The primer sets used for real-time RT-PCR analysis. Gene Forward Reverse VEGF PDGFB TGF-β MCP-1 5'-GTT GCA GCA TGA ATC TGA GG-3' 5'-GGA GAC TCT TCG AGG AGC ACT T-3' 5'-GAA TCA GGC ATC GAG AGA
Abbreviations: P- paraffin-embedded section; C, cryosection; Bio-SA, biotin-streptavidin-conjugated fluorescein amplification.
 Supplementary Table 1. Sequence of primers for real time PCR. Gene Forward primer Reverse primer S25 5 -GTG GTC CAC ACT ACT CTC TGA GTT TC-3 5 - GAC TTT CCG GCA TCC TTC TTC-3 Mafa cds 5 -CTT CAG CAA GGA
Supplementary Table 1. Sequence of primers for real time PCR. Gene Forward primer Reverse primer S25 5 -GTG GTC CAC ACT ACT CTC TGA GTT TC-3 5 - GAC TTT CCG GCA TCC TTC TTC-3 Mafa cds 5 -CTT CAG CAA GGA
Supplementary Materials
 Supplementary Materials 1 Supplementary Table 1. List of primers used for quantitative PCR analysis. Gene name Gene symbol Accession IDs Sequence range Product Primer sequences size (bp) β-actin Actb gi
Supplementary Materials 1 Supplementary Table 1. List of primers used for quantitative PCR analysis. Gene name Gene symbol Accession IDs Sequence range Product Primer sequences size (bp) β-actin Actb gi
c Tuj1(-) apoptotic live 1 DIV 2 DIV 1 DIV 2 DIV Tuj1(+) Tuj1/GFP/DAPI Tuj1 DAPI GFP
 apoptotic live 1 DIV 2 DIV 1 DIV 2 DIV Tuj1(+) Tuj1/GFP/DAPI Tuj1 DAPI GFP") Supplementary Figure 1 Establishment of the gain- and loss-of-function experiments and cell survival assays. a Relative expression of mature mir-484 30 20 10 0 **** **** NCP mir- 484P NCP mir- 484P b Relative
Supplementary Figure 1 Establishment of the gain- and loss-of-function experiments and cell survival assays. a Relative expression of mature mir-484 30 20 10 0 **** **** NCP mir- 484P NCP mir- 484P b Relative
Figure S1. Analysis of genomic and cdna sequences of the targeted regions in WT-KI and
 Figure S1. Analysis of genomic and sequences of the targeted regions in and indicated mutant KI cells, with WT and corresponding mutant sequences underlined. (A) cells; (B) K21E-KI cells; (C) D33A-KI cells;
Figure S1. Analysis of genomic and sequences of the targeted regions in and indicated mutant KI cells, with WT and corresponding mutant sequences underlined. (A) cells; (B) K21E-KI cells; (C) D33A-KI cells;
Supplementary Table 3. 3 UTR primer sequences. Primer sequences used to amplify and clone the 3 UTR of each indicated gene are listed.
 Supplemental Figure 1. DLKI-DIO3 mirna/mrna complementarity. Complementarity between the indicated DLK1-DIO3 cluster mirnas and the UTR of SOX2, SOX9, HIF1A, ZEB1, ZEB2, STAT3 and CDH1with mirsvr and PhastCons
Supplemental Figure 1. DLKI-DIO3 mirna/mrna complementarity. Complementarity between the indicated DLK1-DIO3 cluster mirnas and the UTR of SOX2, SOX9, HIF1A, ZEB1, ZEB2, STAT3 and CDH1with mirsvr and PhastCons
Supplementary Document
 Supplementary Document 1. Supplementary Table legends 2. Supplementary Figure legends 3. Supplementary Tables 4. Supplementary Figures 5. Supplementary References 1. Supplementary Table legends Suppl.
Supplementary Document 1. Supplementary Table legends 2. Supplementary Figure legends 3. Supplementary Tables 4. Supplementary Figures 5. Supplementary References 1. Supplementary Table legends Suppl.
Supplementary Appendix
 Supplementary Appendix This appendix has been provided by the authors to give readers additional information about their work. Supplement to: Sherman SI, Wirth LJ, Droz J-P, et al. Motesanib diphosphate
Supplementary Appendix This appendix has been provided by the authors to give readers additional information about their work. Supplement to: Sherman SI, Wirth LJ, Droz J-P, et al. Motesanib diphosphate
a) Primary cultures derived from the pancreas of an 11-week-old Pdx1-Cre; K-MADM-p53
 Primary cultures derived from the pancreas of an 11-week-old Pdx1-Cre; K-MADM-p53") 1 2 3 4 5 6 7 8 9 10 Supplementary Figure 1. Induction of p53 LOH by MADM. a) Primary cultures derived from the pancreas of an 11-week-old Pdx1-Cre; K-MADM-p53 mouse revealed increased p53 KO/KO (green,
1 2 3 4 5 6 7 8 9 10 Supplementary Figure 1. Induction of p53 LOH by MADM. a) Primary cultures derived from the pancreas of an 11-week-old Pdx1-Cre; K-MADM-p53 mouse revealed increased p53 KO/KO (green,
Toluidin-Staining of mast cells Ear tissue was fixed with Carnoy (60% ethanol, 30% chloroform, 10% acetic acid) overnight at 4 C, afterwards
 overnight at 4 C, afterwards") Toluidin-Staining of mast cells Ear tissue was fixed with Carnoy (60% ethanol, 30% chloroform, 10% acetic acid) overnight at 4 C, afterwards incubated in 100 % ethanol overnight at 4 C and embedded in
Toluidin-Staining of mast cells Ear tissue was fixed with Carnoy (60% ethanol, 30% chloroform, 10% acetic acid) overnight at 4 C, afterwards incubated in 100 % ethanol overnight at 4 C and embedded in
Supplementary Figure 1 MicroRNA expression in human synovial fibroblasts from different locations. MicroRNA, which were identified by RNAseq as most
 Supplementary Figure 1 MicroRNA expression in human synovial fibroblasts from different locations. MicroRNA, which were identified by RNAseq as most differentially expressed between human synovial fibroblasts
Supplementary Figure 1 MicroRNA expression in human synovial fibroblasts from different locations. MicroRNA, which were identified by RNAseq as most differentially expressed between human synovial fibroblasts
Supplementary Figure 1 a
 Supplementary Figure a Normalized expression/tbp (A.U.).6... Trip-br transcripts Trans Trans Trans b..5. Trip-br Ctrl LPS Normalized expression/tbp (A.U.) c Trip-br transcripts. adipocytes.... Trans Trans
Supplementary Figure a Normalized expression/tbp (A.U.).6... Trip-br transcripts Trans Trans Trans b..5. Trip-br Ctrl LPS Normalized expression/tbp (A.U.) c Trip-br transcripts. adipocytes.... Trans Trans
Regulation of Lysophosphatidic Acid Receptor Expression and Function in Human Synoviocytes: Implications for Rheumatoid Arthritis?
 0026-895X/08/7302-587 600 MOLECULAR PHARMACOLOGY Vol. 73, No. 2 U.S. Government work not protected by U.S. copyright 38216/3299899 Mol Pharmacol 73:587 600, 2008 Printed in U.S.A. Regulation of Lysophosphatidic
0026-895X/08/7302-587 600 MOLECULAR PHARMACOLOGY Vol. 73, No. 2 U.S. Government work not protected by U.S. copyright 38216/3299899 Mol Pharmacol 73:587 600, 2008 Printed in U.S.A. Regulation of Lysophosphatidic
Plasmids Western blot analysis and immunostaining Flow Cytometry Cell surface biotinylation RNA isolation and cdna synthesis
 Plasmids psuper-retro-s100a10 shrna1 was constructed by cloning the dsdna oligo 5 -GAT CCC CGT GGG CTT CCA GAG CTT CTT TCA AGA GAA GAA GCT CTG GAA GCC CAC TTT TTA-3 and 5 -AGC TTA AAA AGT GGG CTT CCA GAG
Plasmids psuper-retro-s100a10 shrna1 was constructed by cloning the dsdna oligo 5 -GAT CCC CGT GGG CTT CCA GAG CTT CTT TCA AGA GAA GAA GCT CTG GAA GCC CAC TTT TTA-3 and 5 -AGC TTA AAA AGT GGG CTT CCA GAG
BHP 2-7 and Nthy-ori 3-1 cells were grown in RPMI1640 medium (Hyclone) supplemented with 10% fetal bovine serum (Gibco), 2mM L-glutamine, and 100 U/mL
 supplemented with 10% fetal bovine serum (Gibco), 2mM L-glutamine, and 100 U/mL") 1 2 3 4 Materials and Methods Cell culture BHP 2-7 and Nthy-ori 3-1 cells were grown in RPMI1640 medium (Hyclone) 5 supplemented with 10% fetal bovine serum (Gibco), 2mM L-glutamine, and 100 U/mL 6 penicillin-streptomycin.
1 2 3 4 Materials and Methods Cell culture BHP 2-7 and Nthy-ori 3-1 cells were grown in RPMI1640 medium (Hyclone) 5 supplemented with 10% fetal bovine serum (Gibco), 2mM L-glutamine, and 100 U/mL 6 penicillin-streptomycin.
Supplemental Data. Shin et al. Plant Cell. (2012) /tpc YFP N
 /tpc YFP N") MYC YFP N PIF5 YFP C N-TIC TIC Supplemental Data. Shin et al. Plant Cell. ()..5/tpc..95 Supplemental Figure. TIC interacts with MYC in the nucleus. Bimolecular fluorescence complementation assay using
MYC YFP N PIF5 YFP C N-TIC TIC Supplemental Data. Shin et al. Plant Cell. ()..5/tpc..95 Supplemental Figure. TIC interacts with MYC in the nucleus. Bimolecular fluorescence complementation assay using
Supplementary data Supplementary Figure 1 Supplementary Figure 2
 Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
Nature Structural & Molecular Biology: doi: /nsmb Supplementary Figure 1
 Supplementary Figure 1 U1 inhibition causes a shift of RNA-seq reads from exons to introns. (a) Evidence for the high purity of 4-shU-labeled RNAs used for RNA-seq. HeLa cells transfected with control
Supplementary Figure 1 U1 inhibition causes a shift of RNA-seq reads from exons to introns. (a) Evidence for the high purity of 4-shU-labeled RNAs used for RNA-seq. HeLa cells transfected with control
Supplementary Figure 1. ROS induces rapid Sod1 nuclear localization in a dosagedependent manner. WT yeast cells (SZy1051) were treated with 4NQO at
 were treated with 4NQO at") Supplementary Figure 1. ROS induces rapid Sod1 nuclear localization in a dosagedependent manner. WT yeast cells (SZy1051) were treated with 4NQO at different concentrations for 30 min and analyzed for
Supplementary Figure 1. ROS induces rapid Sod1 nuclear localization in a dosagedependent manner. WT yeast cells (SZy1051) were treated with 4NQO at different concentrations for 30 min and analyzed for
A smart acid nanosystem for ultrasensitive. live cell mrna imaging by the target-triggered intracellular self-assembly
 Electronic Supplementary Material (ESI) for Chemical Science. This journal is The Royal Society of Chemistry 2017 A smart ZnO@polydopamine-nucleic acid nanosystem for ultrasensitive live cell mrna imaging
Electronic Supplementary Material (ESI) for Chemical Science. This journal is The Royal Society of Chemistry 2017 A smart ZnO@polydopamine-nucleic acid nanosystem for ultrasensitive live cell mrna imaging
Supplementary Table 2. Conserved regulatory elements in the promoters of CD36.
 Supplementary Table 1. RT-qPCR primers for CD3, PPARg and CEBP. Assay Forward Primer Reverse Primer 1A CAT TTG TGG CCT TGT GCT CTT TGA TGA GTC ACA GAA AGA ATC AAT TC 1B AGG AAA TGA ACT GAT GAG TCA CAG
Supplementary Table 1. RT-qPCR primers for CD3, PPARg and CEBP. Assay Forward Primer Reverse Primer 1A CAT TTG TGG CCT TGT GCT CTT TGA TGA GTC ACA GAA AGA ATC AAT TC 1B AGG AAA TGA ACT GAT GAG TCA CAG
Supplementary Figure 1
 Metastatic melanoma Primary melanoma Healthy human skin Supplementary Figure 1 CD22 IgG4 Supplementary Figure 1: Immunohisochemical analysis of CD22+ (left) and IgG4 (right), cells (shown in red and indicated
Metastatic melanoma Primary melanoma Healthy human skin Supplementary Figure 1 CD22 IgG4 Supplementary Figure 1: Immunohisochemical analysis of CD22+ (left) and IgG4 (right), cells (shown in red and indicated
Formylpeptide receptor2 contributes to colon epithelial homeostasis, inflammation, and tumorigenesis
 Supplementary Data Formylpeptide receptor2 contributes to colon epithelial homeostasis, inflammation, and tumorigenesis Keqiang Chen, Mingyong Liu, Ying Liu, Teizo Yoshimura, Wei Shen, Yingying Le, Scott
Supplementary Data Formylpeptide receptor2 contributes to colon epithelial homeostasis, inflammation, and tumorigenesis Keqiang Chen, Mingyong Liu, Ying Liu, Teizo Yoshimura, Wei Shen, Yingying Le, Scott
Supplementary Figures
 Supplementary Figures Supplementary Figure 1. H3F3B expression in lung cancer. a. Comparison of H3F3B expression in relapsed and non-relapsed lung cancer patients. b. Prognosis of two groups of lung cancer
Supplementary Figures Supplementary Figure 1. H3F3B expression in lung cancer. a. Comparison of H3F3B expression in relapsed and non-relapsed lung cancer patients. b. Prognosis of two groups of lung cancer
Supplemental Figures: Supplemental Figure 1
 Supplemental Figures: Supplemental Figure 1 Suppl. Figure 1. BM-DC infection with H. pylori does not induce cytotoxicity and treatment of BM-DCs with H. pylori sonicate, but not heat-inactivated bacteria,
Supplemental Figures: Supplemental Figure 1 Suppl. Figure 1. BM-DC infection with H. pylori does not induce cytotoxicity and treatment of BM-DCs with H. pylori sonicate, but not heat-inactivated bacteria,
Phylogenetic analysis of human and chicken importins. Only five of six importins were studied because
 Supplementary Figure S1 Phylogenetic analysis of human and chicken importins. Only five of six importins were studied because importin-α6 was shown to be testis-specific. Human and chicken importin protein
Supplementary Figure S1 Phylogenetic analysis of human and chicken importins. Only five of six importins were studied because importin-α6 was shown to be testis-specific. Human and chicken importin protein
SUPPLEMENTARY INFORMATION
 doi: 10.1038/nature05883 SUPPLEMENTARY INFORMATION Supplemental Figure 1 Prostaglandin agonists and antagonists alter runx1/cmyb expression. a-e, Embryos were exposed to (b) PGE2 and (c) PGI2 (20μM) and
doi: 10.1038/nature05883 SUPPLEMENTARY INFORMATION Supplemental Figure 1 Prostaglandin agonists and antagonists alter runx1/cmyb expression. a-e, Embryos were exposed to (b) PGE2 and (c) PGI2 (20μM) and
Expression of Selected Inflammatory Cytokine Genes in Bladder Biopsies
 Borneo Journal of Resource Science and Technology (2013) 3(2): 15-20 Expression of Selected Inflammatory Cytokine Genes in Bladder Biopsies EDMUND UI-HANG SIM *1, NUR DIANA ANUAR 2, TENG-AIK ONG 3, GUAN-
Borneo Journal of Resource Science and Technology (2013) 3(2): 15-20 Expression of Selected Inflammatory Cytokine Genes in Bladder Biopsies EDMUND UI-HANG SIM *1, NUR DIANA ANUAR 2, TENG-AIK ONG 3, GUAN-
Nature Immunology: doi: /ni.3836
 Supplementary Figure 1 Recombinant LIGHT-VTP induces pericyte contractility and endothelial cell activation. (a) Western blot showing purification steps for full length murine LIGHT-VTP (CGKRK) protein:
Supplementary Figure 1 Recombinant LIGHT-VTP induces pericyte contractility and endothelial cell activation. (a) Western blot showing purification steps for full length murine LIGHT-VTP (CGKRK) protein:
Table S1. Oligonucleotides used for the in-house RT-PCR assays targeting the M, H7 or N9. Assay (s) Target Name Sequence (5 3 ) Comments
 Target Name Sequence (5 3 ) Comments") SUPPLEMENTAL INFORMATION 2 3 Table S. Oligonucleotides used for the in-house RT-PCR assays targeting the M, H7 or N9 genes. Assay (s) Target Name Sequence (5 3 ) Comments CDC M InfA Forward (NS), CDC M
SUPPLEMENTAL INFORMATION 2 3 Table S. Oligonucleotides used for the in-house RT-PCR assays targeting the M, H7 or N9 genes. Assay (s) Target Name Sequence (5 3 ) Comments CDC M InfA Forward (NS), CDC M
HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation
 SUPPLEMENTARY INFORMATION Materials and Methods Human cell lines and culture conditions HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation in exon 20 of BRCA1
SUPPLEMENTARY INFORMATION Materials and Methods Human cell lines and culture conditions HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation in exon 20 of BRCA1
Electrolytes. Summary: (This area will include a brief description of what the protocol is used for and why someone would need to use it.
 Electrolytes Version: 1 Edited by: Jason Kim (note that the following list should be linked to the appropriate location.) Summary Reagents and Materials Protocol Reagent Preparation Reagent 1 Reagent 2
Electrolytes Version: 1 Edited by: Jason Kim (note that the following list should be linked to the appropriate location.) Summary Reagents and Materials Protocol Reagent Preparation Reagent 1 Reagent 2
Supplementary Figure 1
 Supplementary Figure 1 Supplementary Figure 1: Cryopreservation alters CD62L expression by CD4 T cells. Freshly isolated (left) or cryopreserved PBMCs (right) were stained with the mix of antibodies described
Supplementary Figure 1 Supplementary Figure 1: Cryopreservation alters CD62L expression by CD4 T cells. Freshly isolated (left) or cryopreserved PBMCs (right) were stained with the mix of antibodies described
Introduction: 年 Fas signal-mediated apoptosis. PI3K/Akt
 Fas-ligand (CD95-L; Fas-L) Fas (CD95) Fas (apoptosis) 年 了 不 度 Fas Fas-L 力 不 Fas/Fas-L T IL-10Fas/Fas-L 不 年 Fas signal-mediated apoptosis 度降 不 不 力 U-118, HeLa, A549, Huh-7 MCF-7, HepG2. PI3K/Akt FasPI3K/Akt
Fas-ligand (CD95-L; Fas-L) Fas (CD95) Fas (apoptosis) 年 了 不 度 Fas Fas-L 力 不 Fas/Fas-L T IL-10Fas/Fas-L 不 年 Fas signal-mediated apoptosis 度降 不 不 力 U-118, HeLa, A549, Huh-7 MCF-7, HepG2. PI3K/Akt FasPI3K/Akt
Supplementary Figure 1
 Supplementary Figure 1 3 3 3 1 1 Bregma -1.6mm 3 : Bregma Ref) Http://www.mbl.org/atlas165/atlas165_start.html Bregma -.18mm Supplementary Figure 1 Schematic representation of the utilized brain slice
Supplementary Figure 1 3 3 3 1 1 Bregma -1.6mm 3 : Bregma Ref) Http://www.mbl.org/atlas165/atlas165_start.html Bregma -.18mm Supplementary Figure 1 Schematic representation of the utilized brain slice
Anti-inflammatory properties of SM04690, a small molecule inhibitor of the Wnt pathway as a potential treatment for knee osteoarthritis
 Anti-inflammatory properties of SM04690, a small molecule inhibitor of the Wnt pathway as a potential treatment for knee osteoarthritis V. Deshmukh 1, T. Seo 1, C. Swearingen 1, Y. Yazici 1 1 Samumed,
Anti-inflammatory properties of SM04690, a small molecule inhibitor of the Wnt pathway as a potential treatment for knee osteoarthritis V. Deshmukh 1, T. Seo 1, C. Swearingen 1, Y. Yazici 1 1 Samumed,
Citation for published version (APA): Oosterveer, M. H. (2009). Control of metabolic flux by nutrient sensors Groningen: s.n.
: Oosterveer, M. H. (2009). Control of metabolic flux by nutrient sensors Groningen: s.n.") University of Groningen Control of metabolic flux by nutrient sensors Oosterveer, Maaike IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite from it.
University of Groningen Control of metabolic flux by nutrient sensors Oosterveer, Maaike IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite from it.
Islet viability assay and Glucose Stimulated Insulin Secretion assay RT-PCR and Western Blot
 Islet viability assay and Glucose Stimulated Insulin Secretion assay Islet cell viability was determined by colorimetric (3-(4,5-dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide assay using CellTiter
Islet viability assay and Glucose Stimulated Insulin Secretion assay Islet cell viability was determined by colorimetric (3-(4,5-dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide assay using CellTiter
p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO
 Supplementary Information p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO Yuri Shibata, Masaaki Oyama, Hiroko Kozuka-Hata, Xiao Han, Yuetsu Tanaka,
Supplementary Information p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO Yuri Shibata, Masaaki Oyama, Hiroko Kozuka-Hata, Xiao Han, Yuetsu Tanaka,
Description of Supplementary Files. File Name: Supplementary Information Description: Supplementary Figures and Supplementary Tables
 Description of Supplementary Files File Name: Supplementary Information Description: Supplementary Figures and Supplementary Tables Supplementary Figure 1: (A), HCT116 IDH1-WT and IDH1-R132H cells were
Description of Supplementary Files File Name: Supplementary Information Description: Supplementary Figures and Supplementary Tables Supplementary Figure 1: (A), HCT116 IDH1-WT and IDH1-R132H cells were
Cytokines modulate the functional activities of individual cells and tissues both under normal and pathologic conditions Interleukins,
 Cytokines http://highered.mcgraw-hill.com/sites/0072507470/student_view0/chapter22/animation the_immune_response.html Cytokines modulate the functional activities of individual cells and tissues both under
Cytokines http://highered.mcgraw-hill.com/sites/0072507470/student_view0/chapter22/animation the_immune_response.html Cytokines modulate the functional activities of individual cells and tissues both under
Culture Density (OD600) 0.1. Culture Density (OD600) Culture Density (OD600) Culture Density (OD600) Culture Density (OD600)
 0.1. Culture Density (OD600) Culture Density (OD600) Culture Density (OD600) Culture Density (OD600)") A. B. C. D. E. PA JSRI JSRI 2 PA DSAM DSAM 2 DSAM 3 PA LNAP LNAP 2 LNAP 3 PAO Fcor Fcor 2 Fcor 3 PAO Wtho Wtho 2 Wtho 3 Wtho 4 DTSB Low Iron 2 4 6 8 2 4 6 8 2 22 DTSB Low Iron 2 4 6 8 2 4 6 8 2 22 DTSB
A. B. C. D. E. PA JSRI JSRI 2 PA DSAM DSAM 2 DSAM 3 PA LNAP LNAP 2 LNAP 3 PAO Fcor Fcor 2 Fcor 3 PAO Wtho Wtho 2 Wtho 3 Wtho 4 DTSB Low Iron 2 4 6 8 2 4 6 8 2 22 DTSB Low Iron 2 4 6 8 2 4 6 8 2 22 DTSB
MAF Shalaby Prof. Rheumatology Al Azhar University, Cairo, Egypt.
 MAF Shalaby Prof. Rheumatology Al Azhar University, Cairo, Egypt. AUTOIMMUNE DISEASE RA SLE VASCULITIS RELAPSING POLYCHONDRITIS SS DM/PM SJOGREN S SYNDROME RHEUMATOID ARTHRITIS Classically immune mediated
MAF Shalaby Prof. Rheumatology Al Azhar University, Cairo, Egypt. AUTOIMMUNE DISEASE RA SLE VASCULITIS RELAPSING POLYCHONDRITIS SS DM/PM SJOGREN S SYNDROME RHEUMATOID ARTHRITIS Classically immune mediated
Supplementary Materials and Methods
 DD2 suppresses tumorigenicity of ovarian cancer cells by limiting cancer stem cell population Chunhua Han et al. Supplementary Materials and Methods Analysis of publicly available datasets: To analyze
DD2 suppresses tumorigenicity of ovarian cancer cells by limiting cancer stem cell population Chunhua Han et al. Supplementary Materials and Methods Analysis of publicly available datasets: To analyze
SUPPLEMENTARY DATA. Supplementary Table 1. Primer sequences for qrt-pcr
 Supplementary Table 1. Primer sequences for qrt-pcr Gene PRDM16 UCP1 PGC1α Dio2 Elovl3 Cidea Cox8b PPARγ AP2 mttfam CyCs Nampt NRF1 16s-rRNA Hexokinase 2, intron 9 β-actin Primer Sequences 5'-CCA CCA GCG
Supplementary Table 1. Primer sequences for qrt-pcr Gene PRDM16 UCP1 PGC1α Dio2 Elovl3 Cidea Cox8b PPARγ AP2 mttfam CyCs Nampt NRF1 16s-rRNA Hexokinase 2, intron 9 β-actin Primer Sequences 5'-CCA CCA GCG
Supplemental Information. Cancer-Associated Fibroblasts Neutralize. the Anti-tumor Effect of CSF1 Receptor Blockade
 Cancer Cell, Volume 32 Supplemental Information Cancer-Associated Fibroblasts Neutralize the Anti-tumor Effect of CSF1 Receptor Blockade by Inducing PMN-MDSC Infiltration of Tumors Vinit Kumar, Laxminarasimha
Cancer Cell, Volume 32 Supplemental Information Cancer-Associated Fibroblasts Neutralize the Anti-tumor Effect of CSF1 Receptor Blockade by Inducing PMN-MDSC Infiltration of Tumors Vinit Kumar, Laxminarasimha
Astaxanthin prevents and reverses diet-induced insulin resistance and. steatohepatitis in mice: A comparison with vitamin E
 Supplementary Information Astaxanthin prevents and reverses diet-induced insulin resistance and steatohepatitis in mice: A comparison with vitamin E Yinhua Ni, 1,2 Mayumi Nagashimada, 1 Fen Zhuge, 1 Lili
Supplementary Information Astaxanthin prevents and reverses diet-induced insulin resistance and steatohepatitis in mice: A comparison with vitamin E Yinhua Ni, 1,2 Mayumi Nagashimada, 1 Fen Zhuge, 1 Lili
Beta Thalassemia Case Study Introduction to Bioinformatics
 Beta Thalassemia Case Study Sami Khuri Department of Computer Science San José State University San José, California, USA sami.khuri@sjsu.edu www.cs.sjsu.edu/faculty/khuri Outline v Hemoglobin v Alpha
Beta Thalassemia Case Study Sami Khuri Department of Computer Science San José State University San José, California, USA sami.khuri@sjsu.edu www.cs.sjsu.edu/faculty/khuri Outline v Hemoglobin v Alpha
Supplementary information
 Supplementary information Unique polypharmacology nuclear receptor modulator blocks inflammatory signaling pathways Mi Ra Chang 1, Anthony Ciesla 1, Timothy S. Strutzenberg 1, Scott J. Novick 1, Yuanjun
Supplementary information Unique polypharmacology nuclear receptor modulator blocks inflammatory signaling pathways Mi Ra Chang 1, Anthony Ciesla 1, Timothy S. Strutzenberg 1, Scott J. Novick 1, Yuanjun
Cross-talk between mineralocorticoid and angiotensin II signaling for cardiac
 ONLINE SUPPLEMENT TO Crosstalk between mineralocorticoid and angiotensin II signaling for cardiac remodeling An Di ZHANG,,3, Aurelie NGUYEN DINH CAT*,,3, Christelle SOUKASEUM *,,3, Brigitte ESCOUBET, 4,
ONLINE SUPPLEMENT TO Crosstalk between mineralocorticoid and angiotensin II signaling for cardiac remodeling An Di ZHANG,,3, Aurelie NGUYEN DINH CAT*,,3, Christelle SOUKASEUM *,,3, Brigitte ESCOUBET, 4,
Immunological Aspect of Ozone in Rheumatic Diseases
 Immunological Aspect of Ozone in Rheumatic Diseases Prof. Dr. med. Z. Fahmy Chief Consulting Rheumatologist Augusta Clinic for Rheumatic Diseases And Rehabilitation Bad Kreuznach Germany Rheumatoid arthritis
Immunological Aspect of Ozone in Rheumatic Diseases Prof. Dr. med. Z. Fahmy Chief Consulting Rheumatologist Augusta Clinic for Rheumatic Diseases And Rehabilitation Bad Kreuznach Germany Rheumatoid arthritis
CIRCRESAHA/2004/098145/R1 - ONLINE 1. Validation by Semi-quantitative Real-Time Reverse Transcription PCR
 CIRCRESAHA/2004/098145/R1 - ONLINE 1 Expanded Materials and Methods Validation by Semi-quantitative Real-Time Reverse Transcription PCR Expression patterns of 13 genes (Online Table 2), selected with respect
CIRCRESAHA/2004/098145/R1 - ONLINE 1 Expanded Materials and Methods Validation by Semi-quantitative Real-Time Reverse Transcription PCR Expression patterns of 13 genes (Online Table 2), selected with respect
Supplemental Information. Th17 Lymphocytes Induce Neuronal. Cell Death in a Human ipsc-based. Model of Parkinson's Disease
 Cell Stem Cell, Volume 23 Supplemental Information Th17 Lymphocytes Induce Neuronal Cell Death in a Human ipsc-based Model of Parkinson's Disease Annika Sommer, Franz Maxreiter, Florian Krach, Tanja Fadler,
Cell Stem Cell, Volume 23 Supplemental Information Th17 Lymphocytes Induce Neuronal Cell Death in a Human ipsc-based Model of Parkinson's Disease Annika Sommer, Franz Maxreiter, Florian Krach, Tanja Fadler,
SUPPLEMENTARY RESULTS
 SUPPLEMENTARY RESULTS Supplementary Table 1. hfpr1- Flpln-CHO hfpr2-flpln-cho pec 50 E max (%) Log( /K A) Log( /K A) N pec 50 E max (%) Log( /K A) Log( /K A) n ERK1/2 phosphorylation fmlp 9.0±0.6 80±7
SUPPLEMENTARY RESULTS Supplementary Table 1. hfpr1- Flpln-CHO hfpr2-flpln-cho pec 50 E max (%) Log( /K A) Log( /K A) N pec 50 E max (%) Log( /K A) Log( /K A) n ERK1/2 phosphorylation fmlp 9.0±0.6 80±7
Doctoral Degree Program in Marine Biotechnology, College of Marine Sciences, Doctoral Degree Program in Marine Biotechnology, Academia Sinica, Taipei,
 Cyclooxygenase 2 facilitates dengue virus replication and serves as a potential target for developing antiviral agents Chun-Kuang Lin 1,2, Chin-Kai Tseng 3,4, Yu-Hsuan Wu 3,4, Chih-Chuang Liaw 1,5, Chun-
Cyclooxygenase 2 facilitates dengue virus replication and serves as a potential target for developing antiviral agents Chun-Kuang Lin 1,2, Chin-Kai Tseng 3,4, Yu-Hsuan Wu 3,4, Chih-Chuang Liaw 1,5, Chun-
BIOLOGY 621 Identification of the Snorks
 Name: Date: Block: BIOLOGY 621 Identification of the Snorks INTRODUCTION: In this simulation activity, you will examine the DNA sequence of a fictitious organism - the Snork. Snorks were discovered on
Name: Date: Block: BIOLOGY 621 Identification of the Snorks INTRODUCTION: In this simulation activity, you will examine the DNA sequence of a fictitious organism - the Snork. Snorks were discovered on
Assay Kit for Measurement of Proteoglycan. (Sulfated Glycosaminoglycan Quantification Kit)
") Assay Kit for Measurement of Proteoglycan. (Sulfated Glycosaminoglycan Quantification Kit) Cat. No. 280560-N INTRODUCTION Glycosaminoglycans (GAGs) are a major component of the extracellular matrix (ECM)
Assay Kit for Measurement of Proteoglycan. (Sulfated Glycosaminoglycan Quantification Kit) Cat. No. 280560-N INTRODUCTION Glycosaminoglycans (GAGs) are a major component of the extracellular matrix (ECM)
ice-cold 70% ethanol with gentle vortexing, incubated at -20 C for 4 hours, and washed with PBS.
 Cell cycle analysis For cell cycle analysis, single cell suspensions of E12.5 fetal liver cells were suspended in 4 ml ice-cold 7% ethanol with gentle vortexing, incubated at -2 C for 4 hours, and washed
Cell cycle analysis For cell cycle analysis, single cell suspensions of E12.5 fetal liver cells were suspended in 4 ml ice-cold 7% ethanol with gentle vortexing, incubated at -2 C for 4 hours, and washed
Suppl Video: Tumor cells (green) and monocytes (white) are seeded on a confluent endothelial
 and monocytes (white) are seeded on a confluent endothelial") Supplementary Information Häuselmann et al. Monocyte induction of E-selectin-mediated endothelial activation releases VE-cadherin junctions to promote tumor cell extravasation in the metastasis cascade
Supplementary Information Häuselmann et al. Monocyte induction of E-selectin-mediated endothelial activation releases VE-cadherin junctions to promote tumor cell extravasation in the metastasis cascade
SUPPORTING INFORMATION
 SUPPORTING INFORMATION Biology is different in small volumes: endogenous signals shape phenotype of primary hepatocytes cultured in microfluidic channels Amranul Haque, Pantea Gheibi, Yandong Gao, Elena
SUPPORTING INFORMATION Biology is different in small volumes: endogenous signals shape phenotype of primary hepatocytes cultured in microfluidic channels Amranul Haque, Pantea Gheibi, Yandong Gao, Elena
Table S1. Primers used to quantitatively amplify the human mirnas precursors and indicated genes
 Table S1. Primers used to quantitatively amplify the human mirnas precursors and indicated genes Forward primer (5 3 ) Rervese primer (5 3 ) U6 CTCGCTTCGGCAGCACA AACGCTTCACGAATTTGCGT 5S TACGGCCATACCACCCTGAA
Table S1. Primers used to quantitatively amplify the human mirnas precursors and indicated genes Forward primer (5 3 ) Rervese primer (5 3 ) U6 CTCGCTTCGGCAGCACA AACGCTTCACGAATTTGCGT 5S TACGGCCATACCACCCTGAA
Beta Thalassemia Sami Khuri Department of Computer Science San José State University Spring 2015
 Bioinformatics in Medical Product Development SMPD 287 Three Beta Thalassemia Sami Khuri Department of Computer Science San José State University Hemoglobin Outline Anatomy of a gene Hemoglobinopathies
Bioinformatics in Medical Product Development SMPD 287 Three Beta Thalassemia Sami Khuri Department of Computer Science San José State University Hemoglobin Outline Anatomy of a gene Hemoglobinopathies
SUPPLEMENTAL MATERIAL. Supplementary Methods
 SUPPLEMENTAL MATERIAL Supplementary Methods Culture of cardiomyocytes, fibroblasts and cardiac microvascular endothelial cells The isolation and culturing of neonatal rat ventricular cardiomyocytes was
SUPPLEMENTAL MATERIAL Supplementary Methods Culture of cardiomyocytes, fibroblasts and cardiac microvascular endothelial cells The isolation and culturing of neonatal rat ventricular cardiomyocytes was
Title Page. Regulation of Lysophosphatidic Acid Receptor Expression and Function in Human. Synoviocytes: Implications for Rheumatoid Arthritis?
 Molecular Pharmacology This article Fast has not Forward. been copyedited Published and formatted. on The November final version 15, may 2007 differ as from doi:10.1124/mol.107.038216 this version. Title
Molecular Pharmacology This article Fast has not Forward. been copyedited Published and formatted. on The November final version 15, may 2007 differ as from doi:10.1124/mol.107.038216 this version. Title
well for 2 h at rt. Each dot represents an individual mouse and bar is the mean ±
 Supplementary data: Control DC Blimp-1 ko DC 8 6 4 2-2 IL-1β p=.5 medium 8 6 4 2 IL-2 Medium p=.16 8 6 4 2 IL-6 medium p=.3 5 4 3 2 1-1 medium IL-1 n.s. 25 2 15 1 5 IL-12(p7) p=.15 5 IFNγ p=.65 4 3 2 1
Supplementary data: Control DC Blimp-1 ko DC 8 6 4 2-2 IL-1β p=.5 medium 8 6 4 2 IL-2 Medium p=.16 8 6 4 2 IL-6 medium p=.3 5 4 3 2 1-1 medium IL-1 n.s. 25 2 15 1 5 IL-12(p7) p=.15 5 IFNγ p=.65 4 3 2 1
Protocol for Gene Transfection & Western Blotting
 The schedule and the manual of basic techniques for cell culture Advanced Protocol for Gene Transfection & Western Blotting Schedule Day 1 26/07/2008 Transfection Day 3 28/07/2008 Cell lysis Immunoprecipitation
The schedule and the manual of basic techniques for cell culture Advanced Protocol for Gene Transfection & Western Blotting Schedule Day 1 26/07/2008 Transfection Day 3 28/07/2008 Cell lysis Immunoprecipitation
INTRODUCTION. Induction of Monocyte Chemoattractant Protein-1 (MCP-1) Expression by Angiotensin II (AngII) in the Pancreatic Islets and Beta Cells
 Expression by Angiotensin II (AngII) in the Pancreatic Islets and Beta Cells") Induction of Monocyte Chemoattractant Protein-1 (MCP-1) Expression by Angiotensin II (AngII) in the Pancreatic Islets and Beta Cells Galina Chipitsyna, Qiaoke Gong, Chance F. Gray et al. Endocrinology,
Induction of Monocyte Chemoattractant Protein-1 (MCP-1) Expression by Angiotensin II (AngII) in the Pancreatic Islets and Beta Cells Galina Chipitsyna, Qiaoke Gong, Chance F. Gray et al. Endocrinology,
Basis of Immunology and
 Basis of Immunology and Immunophysiopathology of Infectious Diseases Jointly organized by Institut Pasteur in Ho Chi Minh City and Institut Pasteur with kind support from ANRS & Université Pierre et Marie
Basis of Immunology and Immunophysiopathology of Infectious Diseases Jointly organized by Institut Pasteur in Ho Chi Minh City and Institut Pasteur with kind support from ANRS & Université Pierre et Marie
Supplementary Figure 1a
 Supplementary Figure 1a Hours: E-cadherin TGF-β On TGF-β Off 0 12 24 36 48 24 48 72 Vimentin βactin Fig. S1a. Treatment of AML12 cells with TGF-β induces EMT. Treatment of AML12 cells with TGF-β results
Supplementary Figure 1a Hours: E-cadherin TGF-β On TGF-β Off 0 12 24 36 48 24 48 72 Vimentin βactin Fig. S1a. Treatment of AML12 cells with TGF-β induces EMT. Treatment of AML12 cells with TGF-β results
B cells: a fundamental role in the pathogenesis of rheumatoid arthritis?
 Rheumatology 2005;44(Suppl. 2):ii3 ii7 B cells: a fundamental role in the pathogenesis of rheumatoid arthritis? doi:10.1093/rheumatology/keh616 The role of T cells in the pathogenesis of RA is well established,
Rheumatology 2005;44(Suppl. 2):ii3 ii7 B cells: a fundamental role in the pathogenesis of rheumatoid arthritis? doi:10.1093/rheumatology/keh616 The role of T cells in the pathogenesis of RA is well established,
Supplementary Information
 Supplementary Information Supplementary Figure 1. CD4 + T cell activation and lack of apoptosis after crosslinking with anti-cd3 + anti-cd28 + anti-cd160. (a) Flow cytometry of anti-cd160 (5D.10A11) binding
Supplementary Information Supplementary Figure 1. CD4 + T cell activation and lack of apoptosis after crosslinking with anti-cd3 + anti-cd28 + anti-cd160. (a) Flow cytometry of anti-cd160 (5D.10A11) binding
colorimetric sandwich ELISA kit datasheet
 colorimetric sandwich ELISA kit datasheet For the quantitative detection of human TNF-alpha in serum, plasma and cell culture supernatants. general information Catalogue Number Product Name Species cross-reactivity
colorimetric sandwich ELISA kit datasheet For the quantitative detection of human TNF-alpha in serum, plasma and cell culture supernatants. general information Catalogue Number Product Name Species cross-reactivity
Figure 1. Effects of FGF21 on adipose tissue. (A) Representative histological. findings of epididymal adipose tissue (B) mrna expression of
 Representative histological. findings of epididymal adipose tissue (B) mrna expression of") SUPPLEMENTAL MATERIAL EN-12-2276 Figure 1. Effects of FGF21 on adipose tissue. (A) Representative histological findings of epididymal adipose tissue (B) mrna expression of adipocytokines in adipose tissue.
SUPPLEMENTAL MATERIAL EN-12-2276 Figure 1. Effects of FGF21 on adipose tissue. (A) Representative histological findings of epididymal adipose tissue (B) mrna expression of adipocytokines in adipose tissue.
CHAPTER 4 RESULTS. showed that all three replicates had similar growth trends (Figure 4.1) (p<0.05; p=0.0000)
 (p<0.05; p=0.0000)") CHAPTER 4 RESULTS 4.1 Growth Characterization of C. vulgaris 4.1.1 Optical Density Growth study of Chlorella vulgaris based on optical density at 620 nm (OD 620 ) showed that all three replicates had similar
CHAPTER 4 RESULTS 4.1 Growth Characterization of C. vulgaris 4.1.1 Optical Density Growth study of Chlorella vulgaris based on optical density at 620 nm (OD 620 ) showed that all three replicates had similar
General Laboratory methods Plasma analysis: Gene Expression Analysis: Immunoblot analysis: Immunohistochemistry:
 General Laboratory methods Plasma analysis: Plasma insulin (Mercodia, Sweden), leptin (duoset, R&D Systems Europe, Abingdon, United Kingdom), IL-6, TNFα and adiponectin levels (Quantikine kits, R&D Systems
General Laboratory methods Plasma analysis: Plasma insulin (Mercodia, Sweden), leptin (duoset, R&D Systems Europe, Abingdon, United Kingdom), IL-6, TNFα and adiponectin levels (Quantikine kits, R&D Systems
- 1 - Cell types Monocytes THP-1 cells Macrophages. LPS Treatment time (Hour) IL-6 level (pg/ml)
 IL-6 level (pg/ml)") Supplementary Table ST1: The dynamic effect of LPS on IL-6 production in monocytes and THP-1 cells after GdA treatment. Monocytes, THP-1 cells and macrophages (5x10 5 ) were incubated with 10 μg/ml of
Supplementary Table ST1: The dynamic effect of LPS on IL-6 production in monocytes and THP-1 cells after GdA treatment. Monocytes, THP-1 cells and macrophages (5x10 5 ) were incubated with 10 μg/ml of
Supplementary Information. Bamboo shoot fiber prevents obesity in mice by. modulating the gut microbiota
 Supplementary Information Bamboo shoot fiber prevents obesity in mice by modulating the gut microbiota Xiufen Li 1,2, Juan Guo 1, Kailong Ji 1,2, and Ping Zhang 1,* 1 Key Laboratory of Tropical Plant Resources
Supplementary Information Bamboo shoot fiber prevents obesity in mice by modulating the gut microbiota Xiufen Li 1,2, Juan Guo 1, Kailong Ji 1,2, and Ping Zhang 1,* 1 Key Laboratory of Tropical Plant Resources
Supplementary Materials and Methods
 Supplementary Materials and Methods Immunoblotting Immunoblot analysis was performed as described previously (1). Due to high-molecular weight of MUC4 (~ 950 kda) and MUC1 (~ 250 kda) proteins, electrophoresis
Supplementary Materials and Methods Immunoblotting Immunoblot analysis was performed as described previously (1). Due to high-molecular weight of MUC4 (~ 950 kda) and MUC1 (~ 250 kda) proteins, electrophoresis
The Schedule and the Manual of Basic Techniques for Cell Culture
 The Schedule and the Manual of Basic Techniques for Cell Culture 1 Materials Calcium Phosphate Transfection Kit: Invitrogen Cat.No.K2780-01 Falcon tube (Cat No.35-2054:12 x 75 mm, 5 ml tube) Cell: 293
The Schedule and the Manual of Basic Techniques for Cell Culture 1 Materials Calcium Phosphate Transfection Kit: Invitrogen Cat.No.K2780-01 Falcon tube (Cat No.35-2054:12 x 75 mm, 5 ml tube) Cell: 293
Supplementary Data Table of Contents:
 Supplementary Data Table of Contents: - Supplementary Methods - Supplementary Figures S1(A-B) - Supplementary Figures S2 (A-B) - Supplementary Figures S3 - Supplementary Figures S4(A-B) - Supplementary
Supplementary Data Table of Contents: - Supplementary Methods - Supplementary Figures S1(A-B) - Supplementary Figures S2 (A-B) - Supplementary Figures S3 - Supplementary Figures S4(A-B) - Supplementary
The synergistic effect of homocysteine and lipopolysaccharide on the differentiation and conversion of raw264.7 macrophages
 Gao et al. Journal of Inflammation 2014, 11:13 RESEARCH Open Access The synergistic effect of homocysteine and lipopolysaccharide on the differentiation and conversion of raw264.7 macrophages Shanshan
Gao et al. Journal of Inflammation 2014, 11:13 RESEARCH Open Access The synergistic effect of homocysteine and lipopolysaccharide on the differentiation and conversion of raw264.7 macrophages Shanshan
CYTOKINE RECEPTORS AND SIGNAL TRANSDUCTION
 CYTOKINE RECEPTORS AND SIGNAL TRANSDUCTION What is Cytokine? Secreted popypeptide (protein) involved in cell-to-cell signaling. Acts in paracrine or autocrine fashion through specific cellular receptors.
CYTOKINE RECEPTORS AND SIGNAL TRANSDUCTION What is Cytokine? Secreted popypeptide (protein) involved in cell-to-cell signaling. Acts in paracrine or autocrine fashion through specific cellular receptors.
Cytokines, adhesion molecules and apoptosis markers. A comprehensive product line for human and veterinary ELISAs
 Cytokines, adhesion molecules and apoptosis markers A comprehensive product line for human and veterinary ELISAs IBL International s cytokine product line... is extremely comprehensive. The assays are
Cytokines, adhesion molecules and apoptosis markers A comprehensive product line for human and veterinary ELISAs IBL International s cytokine product line... is extremely comprehensive. The assays are
Procaspase-3. Cleaved caspase-3. actin. Cytochrome C (10 M) Z-VAD-fmk. Procaspase-3. Cleaved caspase-3. actin. Z-VAD-fmk
 Z-VAD-fmk. Procaspase-3. Cleaved caspase-3. actin. Z-VAD-fmk") A HeLa actin - + + - - + Cytochrome C (1 M) Z-VAD-fmk PMN - + + - - + actin Cytochrome C (1 M) Z-VAD-fmk Figure S1. (A) Pan-caspase inhibitor z-vad-fmk inhibits cytochrome c- mediated procaspase-3 cleavage.
A HeLa actin - + + - - + Cytochrome C (1 M) Z-VAD-fmk PMN - + + - - + actin Cytochrome C (1 M) Z-VAD-fmk Figure S1. (A) Pan-caspase inhibitor z-vad-fmk inhibits cytochrome c- mediated procaspase-3 cleavage.
Targeting tumour associated macrophages in anti-cancer therapies. Annamaria Gal Seminar Series on Drug Discovery Budapest 5 January 2018
 Targeting tumour associated macrophages in anti-cancer therapies Annamaria Gal Seminar Series on Drug Discovery Budapest 5 January 2018 Macrophages: Professional phagocytes of the myeloid lineage APC,
Targeting tumour associated macrophages in anti-cancer therapies Annamaria Gal Seminar Series on Drug Discovery Budapest 5 January 2018 Macrophages: Professional phagocytes of the myeloid lineage APC,
The Annexin V Apoptosis Assay
 The Annexin V Apoptosis Assay Development of the Annexin V Apoptosis Assay: 1990 Andree at al. found that a protein, Vascular Anticoagulant α, bound to phospholipid bilayers in a calcium dependent manner.
The Annexin V Apoptosis Assay Development of the Annexin V Apoptosis Assay: 1990 Andree at al. found that a protein, Vascular Anticoagulant α, bound to phospholipid bilayers in a calcium dependent manner.
Human Urokinase / PLAU / UPA ELISA Pair Set
 Human Urokinase / PLAU / UPA ELISA Pair Set Catalog Number : SEK10815 To achieve the best assay results, this manual must be read carefully before using this product and the assay is run as summarized
Human Urokinase / PLAU / UPA ELISA Pair Set Catalog Number : SEK10815 To achieve the best assay results, this manual must be read carefully before using this product and the assay is run as summarized
Supporting Information
 Supporting Information Identification of Novel ROS Inducers: Quinone Derivatives Tethered to Long Hydrocarbon Chains Yeonsun Hong,, Sandip Sengupta,, Wooyoung Hur, *, Taebo Sim,, * KU-KIST Graduate School
Supporting Information Identification of Novel ROS Inducers: Quinone Derivatives Tethered to Long Hydrocarbon Chains Yeonsun Hong,, Sandip Sengupta,, Wooyoung Hur, *, Taebo Sim,, * KU-KIST Graduate School
Nori Rabbit IL-2 ELISA Kit DataSheet
 Nori Rabbit IL-2 ELISA Kit DataSheet IL-2 is an interleukin, a type of cytokine immune system signaling molecule. IL-2 is a T cell stimulatory cytokine best known for inducing T cell proliferation, the
Nori Rabbit IL-2 ELISA Kit DataSheet IL-2 is an interleukin, a type of cytokine immune system signaling molecule. IL-2 is a T cell stimulatory cytokine best known for inducing T cell proliferation, the
Supplementary Fig. 1. Identification of acetylation of K68 of SOD2
 Supplementary Fig. 1. Identification of acetylation of K68 of SOD2 A B H. sapiens 54 KHHAAYVNNLNVTEEKYQEALAK 75 M. musculus 54 KHHAAYVNNLNATEEKYHEALAK 75 X. laevis 55 KHHATYVNNLNITEEKYAEALAK 77 D. rerio
Supplementary Fig. 1. Identification of acetylation of K68 of SOD2 A B H. sapiens 54 KHHAAYVNNLNVTEEKYQEALAK 75 M. musculus 54 KHHAAYVNNLNATEEKYHEALAK 75 X. laevis 55 KHHATYVNNLNITEEKYAEALAK 77 D. rerio
HTRF MEASUREMENT OF CYTOKINE RELEASE FROM FRESH BLOOD SAMPLES
 HTRF MEASUREMENT OF CYTOKINE RELEASE FROM FRESH BLOOD SAMPLES APPLICATION NOTE ABSTRACT Cisbio offers a comprehensive line of HTRF cytokine assays to investigate the functional response of immune cells.
HTRF MEASUREMENT OF CYTOKINE RELEASE FROM FRESH BLOOD SAMPLES APPLICATION NOTE ABSTRACT Cisbio offers a comprehensive line of HTRF cytokine assays to investigate the functional response of immune cells.
Focus Application. Compound-Induced Cytotoxicity
 xcelligence System Real-Time Cell Analyzer Focus Application Compound-Induced Cytotoxicity Featured Study: Using the Time Resolving Function of the xcelligence System to Optimize Endpoint Viability and
xcelligence System Real-Time Cell Analyzer Focus Application Compound-Induced Cytotoxicity Featured Study: Using the Time Resolving Function of the xcelligence System to Optimize Endpoint Viability and
TNF-alpha ELISA. For Research Use Only. Not For Use In Diagnostic Procedures.
 TNF-alpha ELISA For the quantitative determination of TNF-alpha in serum, plasma, buffered solution or cell culture medium. For Research Use Only. Not For Use In Diagnostic Procedures. Catalog Number:
TNF-alpha ELISA For the quantitative determination of TNF-alpha in serum, plasma, buffered solution or cell culture medium. For Research Use Only. Not For Use In Diagnostic Procedures. Catalog Number:
(14): DOI: /j.issn ORCID: ( )
: DOI: /j.issn ORCID: ( )") 20 14 2016 04 01 Chinese Journal of Tissue Engineering Research April 1, 2016 Vol.20, No.14 ( 473000). [J]. 2016 20(14):2033-2039. DOI: 10.3969/j.issn.2095-4344.2016.14.008 ORCID: 0000-0002-8672-7109()
20 14 2016 04 01 Chinese Journal of Tissue Engineering Research April 1, 2016 Vol.20, No.14 ( 473000). [J]. 2016 20(14):2033-2039. DOI: 10.3969/j.issn.2095-4344.2016.14.008 ORCID: 0000-0002-8672-7109()
B-cell. Astrocyte SCI SCI. T-cell
 RF #2015 P-01 PI: Azizul Haque, PhD Grant Title: Targeting Enolase in Spinal Cord Injury 12-month Technical Progress Report Progress Report (First Six Months): Enolase is one of the most abundantly expressed
RF #2015 P-01 PI: Azizul Haque, PhD Grant Title: Targeting Enolase in Spinal Cord Injury 12-month Technical Progress Report Progress Report (First Six Months): Enolase is one of the most abundantly expressed
Annals of Oncology Advance Access published January 10, 2005
 Annals of Oncology Advance Access published January 10, 2005 Original article Annals of Oncology doi:10.1093/annonc/mdi077 Expression of survivin and bax/bcl-2 in peroxisome proliferator activated receptor-g
Annals of Oncology Advance Access published January 10, 2005 Original article Annals of Oncology doi:10.1093/annonc/mdi077 Expression of survivin and bax/bcl-2 in peroxisome proliferator activated receptor-g
Supplementary material: Materials and suppliers
 Supplementary material: Materials and suppliers Electrophoresis consumables including tris-glycine, acrylamide, SDS buffer and Coomassie Brilliant Blue G-2 dye (CBB) were purchased from Ameresco (Solon,
Supplementary material: Materials and suppliers Electrophoresis consumables including tris-glycine, acrylamide, SDS buffer and Coomassie Brilliant Blue G-2 dye (CBB) were purchased from Ameresco (Solon,
HCV Persistence and Immune Evasion in the Absence of Memory T Cell Help.
 SOM Text HCV Persistence and Immune Evasion in the Absence of Memory T Cell Help. Arash Grakoui 1, Naglaa H. Shoukry 2, David J. Woollard 2, Jin-Hwan Han 1, Holly L. Hanson 1, John Ghrayeb 3, Krishna K.
SOM Text HCV Persistence and Immune Evasion in the Absence of Memory T Cell Help. Arash Grakoui 1, Naglaa H. Shoukry 2, David J. Woollard 2, Jin-Hwan Han 1, Holly L. Hanson 1, John Ghrayeb 3, Krishna K.
Department of Pharmaceutical Sciences, School of Pharmacy, Northeastern University, Boston, MA 02115, USA 2
 Pancreatic Cancer Cell Exosome-Mediated Macrophage Reprogramming and the Role of MicroRNAs 155 and 125b2 Transfection using Nanoparticle Delivery Systems Mei-Ju Su 1, Hibah Aldawsari 2, and Mansoor Amiji
Pancreatic Cancer Cell Exosome-Mediated Macrophage Reprogramming and the Role of MicroRNAs 155 and 125b2 Transfection using Nanoparticle Delivery Systems Mei-Ju Su 1, Hibah Aldawsari 2, and Mansoor Amiji