Accepted Manuscript. Increased expression of cutaneous α 1 -adrenoceptors after chronic constriction injury in rats
|
|
|
- Maude Stanley
- 5 years ago
- Views:
Transcription
01351-5 10.")
1 Accepted Manuscript Increased expression of cutaneous α 1 -adrenoceptors after chronic constriction injury in rats Eleanor S. Drummond, Linda F. Dawson, Philip M. Finch, Gary J. Bennett, Peter D. Drummond PII: DOI: S (13) /j.jpain Reference: YJPAI 2863 To appear in: Journal of Pain Received Date: 6 August 2013 Revised Date: 2 October 2013 Accepted Date: 2 October 2013 Please cite this article as: Drummond ES, Dawson LF, Finch PM, Bennett GJ, Drummond PD, Increased expression of cutaneous α 1 -adrenoceptors after chronic constriction injury in rats, Journal of Pain (2013), doi: /j.jpain This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
2 1 Increased expression of cutaneous α 1 -adrenoceptors after chronic constriction injury in rats Eleanor S. Drummond, a Linda F. Dawson, a Philip M. Finch, a Gary J. Bennett, b Peter D. Drummond a a Centre for Research on Chronic Pain and Inflammatory Diseases, Murdoch University, Perth, Western Australia; b Department of Anesthesia, Faculty of Dentistry, and The Alan Edwards Centre for Research on Pain, McGill University, Montréal, QC, Canada. Address for correspondence: Professor Peter Drummond, School of Psychology and Exercise Science, Murdoch University, 6150 Western Australia. Ph: Fax: P.Drummond@murdoch.edu.au
3 2 Abstract Alpha 1 -adrenoceptor expression on nociceptors may play an important role in sympathetic-sensory coupling in certain neuropathic pain syndromes. The aim of this study was to determine whether α 1 - adrenoceptor expression was up-regulated on surviving peptidergic, non-peptidergic and myelinated nerve fiber populations in the skin after chronic constriction injury of the sciatic nerve in rats. Seven days after surgery, α 1 -adrenoceptor expression was up-regulated in the epidermis and on dermal nerve fibres in plantar skin ipsilateral to the injury, but not around blood vessels. This α 1 - adrenoceptor up-regulation in the plantar skin was observed on all nerve fiber populations examined. However, α 1 -adrenoceptor expression was unaltered in dorsal hind paw skin after the injury. The increased expression of α 1 -adrenoceptors on cutaneous nociceptors in plantar skin after chronic constriction injury suggests that this may be a site of sensory-sympathetic coupling that increases sensitivity to adrenergic agonists after nerve injury. In addition, activation of up-regulated α 1 -adrenoceptors in the epidermis might cause release of factors that stimulate nociceptive signalling. Perspective: Our findings indicate that peripheral nerve injury provokes up-regulation of α 1 - adrenoceptors on surviving nociceptive afferents and epidermal cells in the skin. This might contribute to sympathetically maintained pain in conditions such as complex regional pain syndrome, painful diabetic neuropathy and post-herpetic neuralgia. Keywords: α 1 -adrenoceptors; nociceptors; neuropathic pain; chronic constriction injury; keratinocytes; sensory-sympathetic coupling
4 3 Introduction Activation of the sympathetic nervous system with consequent release of norepinephrine does not normally activate nociceptors. However, after injury, sensory-sympathetic coupling in the skin may 1, 10, become a source of pain by activating α 1 -adrenoceptors(α 1 -ARs) on nociceptive afferent fibers 12, 28, 43. After nerve injury, messenger RNA for the α 1B -AR subtype increases in the dorsal root ganglia 29, 50, but whether this results in an up-regulation of α 1 -ARs on nociceptors in the skin is unknown. Chronic constriction injury (CCI) of the rat sciatic nerve triggers signs of pain and hyperalgesia as early as 24 hours post-injury 3, 5, 9, 18, together with neurodegeneration of sensory nerve fibers in the injured sciatic nerve and the skin. The injured nerve fibers regenerate gradually in the weeks following injury, resulting in a de novo, spatially intimate association of sprouting sympathetic efferent fibers and sensory afferent fibers in the upper dermis 25-27, 37, 51. Chemical and surgical sympathectomy generally attenuates allodynia and thermal hyperalgesia after CCI 7, 34 (but see 18 ), suggesting that certain features of the neuropathic pain associated with CCI may be sympathetically maintained. Therefore, the aim of this study was to determine whether α 1 -AR expression was upregulated on surviving peptidergic, non-peptidergic and myelinated nerve fiber populations in the skin after CCI. As α 1 -AR expression in the epidermis and on blood vessels could indirectly influence nociceptor signalling, α 1 -AR expression changes on these skin structures were also examined. Chronic constriction injury Methods The experiments conformed to the ethical guidelines of the International Association for the Study of Pain, the National Institutes of Health (USA), and the Canadian Institutes of Health Research. All protocols were approved by the Animal Care Committee of the Faculty of Medicine, McGill University, in accordance with the regulations of the Canadian Council on Animal Care, and by the Animal Ethics Committee of Murdoch University. All animals used in this study were male Sprague
5 4 Dawley rats weighing g. Six rats underwent unilateral CCI of the common sciatic nerve 5 and three rats were examined as naive, un-operated age-matched controls. Histological processing and immunohistochemsitry Seven days after surgery rats were anesthetised with a lethal dose of sodium pentobarbital (100 mg/kg, IP) and perfused transcardially with a vascular rinse (0.1M phosphate buffered saline containing 0.05% sodium bicarbonate and 0.1% sodium nitrite) followed by freshly prepared 4% paraformaldehyde in 0.1M phosphate buffered saline (PBS), ph 7.4. The hind paws were severed and post-fixed overnight, after which a sample of glabrous, plantar skin was excised from the wide part of the plantar hind paw that lies distal to the calcaneus and proximal to the digital tori and another sample of skin was taken from the dorsum of the hind paw proximal to the toes. The samples were cryoprotected in 30% sucrose at 4 C overnight and then embedded in Optimal Cutting Temperature compound, frozen on dry ice, and stored at -80 C. 10 µm thick cross-sections were cut using a cryostat and collected onto silane coated slides (Hurst Scientific). Cryosections were stained with the following combinations of antibodies (details shown in Table 1): α 1 -AR/calcitonin gene related peptide (CGRP)/pan-neuronal marker (TUJ1) to examine α 1 -AR expression on peptidergic afferents; α 1 -AR/isolectin B4 (IB4)/TUJ1 to examine α 1 -AR expression on non-peptidergic afferents; α 1 -AR/neurofilament 200 (NF200)/TUJ1 to examine α 1 -AR expression on myelinated fibers; and α 1 -AR/smooth muscle actin (SMA) to examine α 1 -AR expression on blood vessels. For immunohistochemistry, sections initially were washed in 0.1M PBS (3x10 min) and then incubated with 0.2% Triton X-100 for 7.5 min at room temperature. Sections were washed with PBS (3x5 min) and blocked for 2 hrs in 10% donkey serum in PBS at room temperature. Sections were incubated for 48 hrs at 4 C with primary antibodies using the concentrations shown in Table 1, diluted in blocking solution. Sections were washed with PBS (3x15 min) and then incubated with the
6 5 appropriate secondary antibodies diluted in 5% donkey serum (Sigma) in PBS for 4 hrs at room temperature. Sections were washed with PBS (3x15 min) and cover-slipped with Prolong Gold antifade mounting media. The peptide sequence recognized by the α 1 -AR antibody used in this study is unique to the α 1 -AR, a G-protein-coupled receptor (National Center for Biotechnology Information, National Institutes of Health, BLAST program, In an examination of the specificity of the α 1 -AR antibody, the pattern of staining on blood vessels, nerves and epidermal cells in rat skin tissue resembled the staining pattern produced by BODIPY FL-prazosin, a fluorescent α 1 -AR antagonist. 6 In addition, staining was eliminated following pre-adsorption of the anti-sera with an α 1 -AR-specific peptide. 6 In the present study, no staining was observed on negative control sections that had the primary antibodies omitted. Two consecutive sections per sample were stained with a combination of antibodies, and immunohistochemistry for each antibody combination was performed in one run to ensure staining consistency. Quantification of Immunohistochemistry α 1 -AR staining intensity was quantified in the epidermis, dermal blood vessels, nerve fibers in the upper dermis, and nerve fibers in large dermal nerve bundles by a blinded investigator. The epidermis was identified by morphology, blood vessels were identified by SMA staining and nerve fibers in the dermis and nerve bundles were identified using the pan-neuronal marker TUJ1. α 1 -AR staining intensity in the epidermis and nerve fibers was quantified in 7 sections per sample and averaged. Since many blood vessels were present in each skin section, α 1 -AR staining intensity in small blood vessels in the papillary dermis proximal to the epidermis was quantified as the average intensity in the smooth muscle surrounding all blood vessels in two representative images taken from one section per sample. One additional image from each α 1 -AR/SMA stained section was
7 6 collected from the deeper reticular dermis and used to quantify α 1 -AR staining intensity in the smooth muscle surrounding large blood vessels. Images of immunostained skin sections were collected using a Nikon A1 confocal microscope. For quantification of α 1 -AR intensity, two 200X magnification confocal image stacks were collected per section; one image stack contained the papillary dermis, epidermis and dermal nerve fibers, and the other image stack was of the large dermal nerve bundles in the reticular dermis. Two 200X images of the papillary dermis and one from the reticular dermis were collected from sections stained with α 1 - AR and SMA for quantification of α 1 -AR expression on blood vessels. All confocal stacks consisted of consecutive images collected with an optical section thickness of 2 µm, and the maximum intensity projection overlay of the resulting stack was used for quantification. Special care was taken to ensure that there was no bleed-through between channels using the imaging settings chosen, and imaging settings were identical for all sections in each staining run. Quantification of immunohistochemistry staining intensity was performed using ImageJ software (available from The proportion of each nerve bundle occupied by nerve fibers was calculated in 4-7 nerve bundles per sample. Nerve bundles were identified using a combination of morphology and TUJ1 staining. The perimeter of each nerve bundle was manually traced around and the area was measured. An intensity threshold was applied to all TUJ1 images, which ensured that only immuno-positive TUJ1 (TUJ1 + ) pixels were included in analysis, and the proportion of TUJ1 + pixels in each nerve bundle was determined. α 1 -AR staining intensity was measured as the average pixel intensity in each area of interest.tuj1 staining was used to identify the location of nerve fibers, which was then used as a mask on the corresponding α 1 -AR stained image, ensuring that α 1 -AR expression was only examined in those pixels also positive for TUJ1 staining. Similarly, quantification of the average α 1 -AR intensity on blood vessels was performed by creating a mask from SMA immunostaining and using that to define the
8 7 location of blood vessels in the corresponding α 1 -AR image. Quantification of α 1 -ARs in the epidermis was performed by manually drawing around the keratinocyte layer and measuring the average α 1 -AR intensity in the defined region. α 1 -AR staining intensity was also examined in specific primary afferent neuron subpopulations: peptidergic CGRP + neurons, non-peptidergic IB4 + neurons and myelinated NF200 + neurons. For this quantification, a co-localisation analysis was performed between each of these specific neuronal markers and TUJ1 using the Co-localization Finder plugin to create a mask of pixels that identified each individual neuronal population. This mask was applied to the corresponding α 1 -AR image and α 1 -AR staining intensity was then quantified in these specific neuronal populations. The proportion of nerve fibers belonging to each of these populations was quantified by expressing the number of pixels co-localized for TUJ1 and CGRP, IB4 or NF200 as a percentage of the total number of pixels labelled for TUJ1. Statistical approach To combine data for neural markers across multiple immunohistochemistry runs, α 1 -AR + pixel intensity within each run was expressed as a Z-score for each region of interest (nerve fibers in the papillary dermis and nerve bundles in the reticular dermis). Mean scores were then compared between the injured and contralateral limb with Wilcoxon s Signed Ranks test, and between experimental and control animals with the Mann-Whitney U test. A similar approach was used to investigate differences in α 1 -AR + pixel intensity in the epidermis and blood vessel walls, and in peptidergic, non-peptidergic and myelinated neuron populations throughout the papillary and reticular dermis. Results are reported as the mean ± standard error, and the criterion of statistical significance was p<0.05.
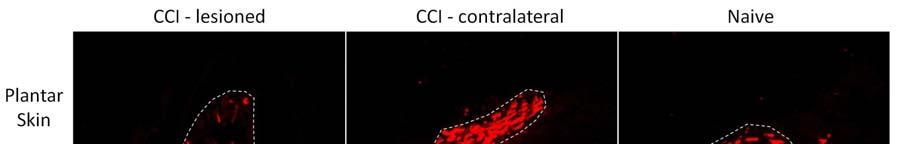
9 8 Results Neurodegeneration after CCI The proportion of pixels within nerve bundles labelled by the pan-neuronal marker TUJ1 was used to index the extent of neurodegeneration. There was a trend for this index to be lower in skin ipsilateral than contralateral to CCI and in comparison to skin from naive animals both in plantar and hairy skin (Table 2), but this trend did not achieve statistical significance. The surviving dermal TUJ1 + fibers were then separated into peptidergic, non-peptidergic and myelinated neuron populations, and the proportions of these populations were compared across experimental groups to determine whether any of these populations were particularly vulnerable to CCI. There was a profound degeneration of NF200 + fibers in plantar skin ipsilateral to CCI (Figure 1); the percentage of NF200 + pixels co-labelled for TUJ1 decreased to 6±2% of the total number of TUJ1 + pixels, which was lower than in skin contralateral to CCI (24±5%, p<0.05) and in naive animals (28± 6%, p<0.05). This degeneration was specific to NF200 + fibers as the proportions of CGRP + and IB4 + pixels remained unchanged after CCI (Table 3). Interestingly, the degeneration of myelinated fibers was observed in plantar skin but not in hairy skin on the dorsal paw (Figure 1 and Table 3). α 1 -AR expression in skin of naïve rats The strongest α 1 -AR expression was observed in the epidermis. α 1 -AR expression was also observed around blood vessels and in nerve fibers in the dermis and in large dermal nerve bundles. The strong α 1 -AR expression throughout the epidermis precluded observation of α 1 -ARs on intra-epidermal nerve fibers. The pattern of α 1 -AR staining was consistent with a previous study that examined α 1 -AR staining in skin from uninjured rats 6. α 1 -AR up-regulation in plantar skin after nerve injury
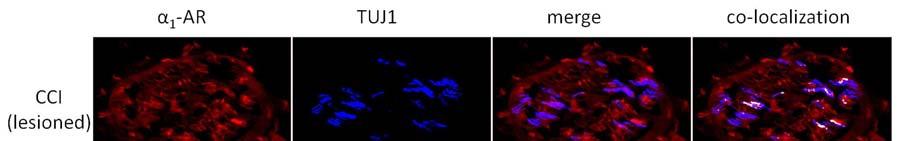
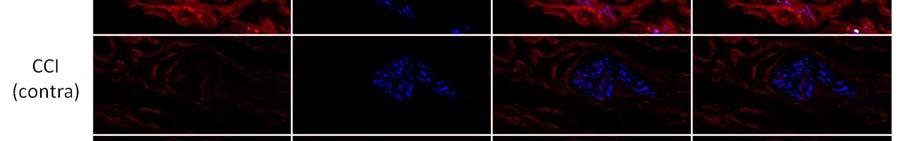
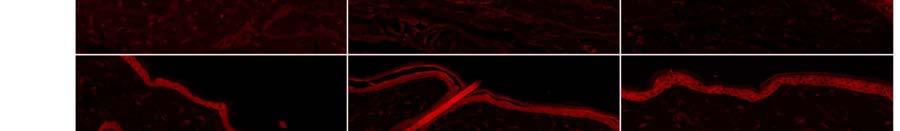
10 9 α 1 -AR expression was up-regulated after CCI in all regions examined in ipsilateral plantar skin except around blood vessels. CCI resulted in significantly increased α 1 -AR expression on nerve fibers labelled with TUJ1 (TUJ1 + ) and in nerve bundles in the deep dermis in comparison to skin contralateral to CCI (p<0.05) and to skin from naive animals (p<0.05)(figure 2, Figure 3A). α 1 -AR expression was also significantly higher in TUJ1 + nerve fibers in the papillary dermis in skin ipsilateral to CCI than in contralateral skin (p<0.05)(figure 3B). α 1 -AR expression was then examined in individual primary afferent subpopulations. In those nerve fibers that remained after injury, α 1 -AR expression was significantly higher on CGRP + fibers in skin ipsilateral than contralateral to CCI (p<0.05)(figure 4A). α 1 -AR expression was also significantly higher in IB4 + fibers in skin ipsilateral than contralateral to CCI (p<0.05) and in comparison to skin from naive animals (p<0.05)(figure 4B). Similarly, in the few remaining plantar dermal NF200 + fibers, α 1 -AR expression was higher ipsilateral than contralateral to CCI (p<0.05)(figure 4C). α 1 -AR expression was significantly higher in the epidermis ipsilateral than contralateral to CCI (p<0.05) and in comparison to skin from naive animals (p<0.01)(figure 3C, Figure 5). There was no difference in α 1 -AR expression in the epidermis from skin contralateral to CCI compared to skin from naive animals. There were also no significant differences in the average α 1 -AR staining intensity in small blood vessels in the papillary dermis or large blood vessels in the reticular dermis after CCI in comparison to skin contralateral to CCI or to skin from naive animals (Figure 6A). α 1 -AR expression in hairy skin α 1 -AR expression was not altered in the epidermis, dermal blood vessels, nerve fibers in the dermis or nerve fibers in dermal nerve bundles in hairy skin after CCI in comparison either to skin from the contralateral paw or from naive rats (Figure 3D-F and Figure 6B). As immunohistochemistry of hairy skin was performed at the same time as plantar skin, and α 1 -AR staining intensity was consistent
11 10 between groups in all structures examined in hairy skin, these results suggest that α 1 -AR expression was not up-regulated in hairy skin at 7 days post-cci. Discussion α 1 -AR expression was increased on cutaneous nerve fibers and in the epidermis after CCI. We have previously shown that nociceptors express α 1 -ARs basally 6. Others have shown that surviving nerve fibers become more sensitive to α 1 -AR agonists after nerve injury 2, 33 and that administration of α 1 - AR antagonists reduces thermal and mechanical hyperalgesia in animal models of neuropathic pain 14, 19, 20, 22, 49. The increased expression of α 1 -ARs on cutaneous nociceptors after CCI suggests that this may be a site of sensory-sympathetic coupling, and could provide a mechanism underlying this hypersensitivity to adrenergic agonists after nerve injury. It has been hypothesised that direct activation of α 1 -ARs on non-peptidergic nociceptors expressing the P2X3 receptor enhances the firing rate to painful stimuli by activating protein kinase C 29, 31. Considering that we also found α 1 -AR up-regulation on non-peptidergic nociceptors after CCI, this could provide a potential direct link between increased α 1 -AR expression on nociceptors and neuropathic pain. α 1 -AR expression was up-regulated on nerve fibers co-labelled with CGRP + or IB4 +, and was also upregulated in nerve bundles on fibers co-labelled with NF200, a marker of myelinated nociceptive and non-nociceptive neurons. These nerve fibers may carry different types of pain information; CGRP + nociceptors respond to noxious heat and are partly responsible for hypersensitivity to heat after nerve injury 17, 30, whereas non-peptidergic nociceptors respond both to noxious heat and mechanical stimuli and contribute to heat hypersensitivity and mechanical allodynia after nerve injury 16, 41, 42, 45, 46. Similarly, decreases in the firing threshold of myelinated nociceptors may increase the intensity of sharp, pricking pain after nerve injury 8. Therefore, if α 1 -AR expression increases the excitability of these nociceptors, α 1 -AR up-regulation could potentially contribute to symptoms of neuropathic pain.
12 11 α 1 -AR expression may also influence pain signalling indirectly by stimulating the release of secondary mediators that act on nociceptors. α 1 -AR expression was increased throughout the epidermis in plantar skin affected by CCI. The normal epidermis contains nerve fibers, Langerhans cells, melanocytes, and keratinocytes. Melanocytes and Langerhans cells are relatively rare and are confined almost exclusively to the basal (germinative) layer, whereas keratinocytes are present in all the vital layers. Keratinocytes can influence nerve signalling by releasing various factors that are capable of activating and sensitizing nerve fibers after injury including nerve growth factor (NGF), pro-inflammatory cytokines, CGRP and ATP 15, 23, 24, 38, 40, 53. Given the up-regulation of α 1 -AR expression in the CCI-affected plantar epidermis, it could be hypothesised that activation of these receptors might cause release of factors from keratinocytes and/or other epidermal cells that are capable of stimulating nociceptive signalling, resulting in sensitization of these nociceptive nerve fibers and contributing to neuropathic pain. The release of NGF could be of particular importance as it might not only sensitize nociceptive afferents but could also promote the growth of re-innervating cutaneous sympathetic nerve fibers into the upper dermis after CCI; these fibers grow in close proximity to nociceptors and are a possible source of the norepinephrine needed to activate α 1 -ARs 51. Keratinocytes and melanocytes also have the capacity to synthesize catecholamines 13. Thus, one could speculate that autocrine stimulation of α 1 - or β 2 -ARs 23 on these epidermal cells triggers a cascade of inflammatory mediators that augment neuropathic pain. α 1 -AR up-regulation was observed in plantar skin but not in hairy skin on the dorsal paw. This was an unexpected finding as both types of skin are innervated by branches of the sciatic nerve, and TUJ1 staining suggested a trend for neurodegeneration in both regions. Interestingly, in a previous study, neurons in hairy skin regenerated faster than neurons in plantar skin after sciatic nerve injury 39, suggesting that there may be some inherent difference between these regions that results in different responses to injury. Our finding that NF200 + fibers were profoundly degenerated in plantar, but not hairy, skin supports this hypothesis. Extensive degeneration of myelinated fibers after CCI
13 12 has been observed previously 4, 32, 37. However, this is the first study to compare the degeneration in plantar and hairy skin. Why there was a difference between the two types of skin in the response to nerve injury is not yet understood, but one major difference is that the plantar skin is completely innervated by branches of the sciatic nerve whereas the medial section of the dorsal hind paw is innervated by the saphenous nerve 47. One could speculate that the presence of uninjured saphenous nerve fibers in the hairy skin could provide a neuroprotective effect, at least during the first week after injury (the only time point examined in this study). In addition, it would be interesting to determine whether a functional process (e.g., triggered by weight-bearing on the plantar surface of the paw 11 ) influenced the response to CCI. α 1 -AR expression on large and small blood vessels was unaffected by CCI. This is interesting as blood vessels appear to have heightened sensitivity to α 1 -AR agonists after injury 21, 49, 52. This has resulted in the hypothesis that α 1 -AR expression may be increased on blood vessels in cases of sympathetically-maintained pain and that α 1 -AR activation causes pain by inducing vasoconstriction 48. Our results suggest that increased vascular sensitivity to α 1 -AR agonists after injury is not due to increased vascular expression of α 1 -ARs, at least in the CCI model, but instead may be a direct consequence of denervation of blood vessels and consequent loss of neuronal norepinephrine transporters 44. This could explain both the lack of α 1 -AR up-regulation on blood vessels and increased sensitivity to α 1 -AR agonists observed after CCI. One limitation of this study was that α 1 -AR expression was examined at only one time point, 7 days post-injury. This time point was chosen because previous studies have consistently reported the presence of mechanical and thermal hyperalgesia at this early post-injury time 5, 18, 35, 36. However, it would be interesting in future studies to determine whether α 1 -AR up-regulation is present at later time points. In addition, it would be useful to include sham operated animals in future studies to investigate possible nonspecific effects of surgery.
14 13 In conclusion, the up-regulation of α 1 -ARs both in the epidermis and on nerve fibers in skin affected by CCI provides insights into the mechanism of involvement of the sympathetic nervous system in neuropathic pain. The increased expression of α 1 -ARs after injury suggests that epidermal cells and nociceptive nerve fibers may become more sensitive to epinephrine and norepinephrine released as a result of sympathetic neural or adrenal gland excitation, or perhaps even synthesized locally within the epidermis. This could either directly activate α 1 -ARs expressed on nociceptors or indirectly excite nociceptors by activation of epidermal cells and consequent release of factors that act on nociceptive nerve fibers. Acknowledgements and disclosures This work was supported by grants from the National Health and Medical Research Council of Australia (grant numbers APP , ); the Australian and New Zealand College of Anaesthetists (grant number 12/024); and unrestricted grants from Pfizer, Medtronic Australasia and St Jude Medical. None of the authors has a conflict of interest with the contents of this paper.
15 14 References 1. Ali Z, Raja SN, Wesselmann U, Fuchs PN, Meyer RA, Campbell JN. Intradermal injection of norepinephrine evokes pain in patients with sympathetically maintained pain. Pain. 88: , Ali Z, Ringkamp M, Hartke TV, Chien HF, Flavahan NA, Campbell JN, Meyer RA. Uninjured C- fiber nociceptors develop spontaneous activity and alpha-adrenergic sensitivity following L6 spinal nerve ligation in monkey. J Neurophysiol. 81: , Attal N, Jazat F, Kayser V, Guilbaud G. Further evidence for 'pain-related' behaviours in a model of unilateral peripheral mononeuropathy. Pain. 41: , Basbaum AI, Gautron M, Jazat F, Mayes M, Guilbaud G. The spectrum of fiber loss in a model of neuropathic pain in the rat: an electron microscopic study. Pain. 47: , Bennett GJ, Xie YK. A peripheral mononeuropathy in rat that produces disorders of pain sensation like those seen in man. Pain. 33:87-107, Dawson LF, Phillips JK, Finch PM, Inglis JJ, Drummond PD. Expression of alpha1- adrenoceptors on peripheral nociceptive neurons. Neuroscience. 175: , Desmeules JA, Kayser V, Weil-Fuggaza J, Bertrand A, Guilbaud G. Influence of the sympathetic nervous system in the development of abnormal pain-related behaviours in a rat model of neuropathic pain. Neuroscience. 67: , Djouhri L, Fang X, Koutsikou S, Lawson SN. Partial nerve injury induces electrophysiological changes in conducting (uninjured) nociceptive and nonnociceptive DRG neurons: Possible relationships to aspects of peripheral neuropathic pain and paresthesias. Pain. 153: , Dowdall T, Robinson I, Meert TF. Comparison of five different rat models of peripheral nerve injury. Pharmacol Biochem Behav. 80:93-108, Drummond PD, Skipworth S, Finch PM. alpha 1-adrenoceptors in normal and hyperalgesic human skin. Clin Sci (Lond). 91:73-77, Eliasson P, Andersson T, Aspenberg P. Influence of a single loading episode on gene expression in healing rat Achilles tendons. J Appl Physiol. 112: , Gibbs GF, Drummond PD, Finch PM, Phillips JK. Unravelling the pathophysiology of complex regional pain syndrome: focus on sympathetically maintained pain. Clin Exp Pharmacol Physiol. 35: , Grando SA, Pittelkow MR, Schallreuter KU. Adrenergic and cholinergic control in the biology of epidermis: physiological and clinical significance. The Journal of investigative dermatology. 126: , Hord AH, Denson DD, Stowe B, Haygood RM. alpha-1 and alpha-2 Adrenergic antagonists relieve thermal hyperalgesia in experimental mononeuropathy from chronic constriction injury. Anesth Analg. 92: , Hou Q, Barr T, Gee L, Vickers J, Wymer J, Borsani E, Rodella L, Getsios S, Burdo T, Eisenberg E, Guha U, Lavker R, Kessler J, Chittur S, Fiorino D, Rice F, Albrecht P. Keratinocyte expression of calcitonin gene-related peptide beta: implications for neuropathic and inflammatory pain mechanisms. Pain. 152: , Hsieh YL, Chiang H, Lue JH, Hsieh ST. P2X3-mediated peripheral sensitization of neuropathic pain in resiniferatoxin-induced neuropathy. Exp Neurol. 235: , Hsieh YL, Lin CL, Chiang H, Fu YS, Lue JH, Hsieh ST. Role of Peptidergic Nerve Terminals in the Skin: Reversal of Thermal Sensation by Calcitonin Gene-Related Peptide in TRPV1-Depleted Neuropathy. PLoS One. 7:e50805, Kim KJ, Yoon YW, Chung JM. Comparison of three rodent neuropathic pain models. Exp Brain Res. 113: , 1997
16 Kim SK, Min BI, Kim JH, Hwang BG, Yoo GY, Park DS, Na HS. Effects of alpha1- and alpha2- adrenoreceptor antagonists on cold allodynia in a rat tail model of neuropathic pain. Brain Res. 1039: , Kim SK, Min BI, Kim JH, Hwang BG, Yoo GY, Park DS, Na HS. Individual differences in the sensitivity of cold allodynia to phentolamine in neuropathic rats. Eur J Pharmacol. 523:64-66, Kurvers H, Daemen M, Slaaf D, Stassen F, van den Wildenberg F, Kitslaar P, de Mey J. Partial peripheral neuropathy and denervation induced adrenoceptor supersensitivity. Functional studies in an experimental model. Acta Orthop Belg. 64:64-70, Lee DH, Liu X, Kim HT, Chung K, Chung JM. Receptor subtype mediating the adrenergic sensitivity of pain behavior and ectopic discharges in neuropathic Lewis rats. J Neurophysiol. 81: , Li W, Shi X, Wang L, Guo T, Wei T, Cheng K, Rice KC, Kingery WS, Clark JD. Epidermal adrenergic signaling contributes to inflammation and pain sensitization in a rat model of complex regional pain syndrome. Pain. 154: , Li WW, Guo TZ, Li XQ, Kingery WS, Clark JD. Fracture induces keratinocyte activation, proliferation, and expression of pro-nociceptive inflammatory mediators. Pain. 151: , Lindenlaub T, Sommer C. Epidermal innervation density after partial sciatic nerve lesion and pain-related behavior in the rat. Acta Neuropathol. 104: , Lindenlaub T, Teuteberg P, Hartung T, Sommer C. Effects of neutralizing antibodies to TNFalpha on pain-related behavior and nerve regeneration in mice with chronic constriction injury. Brain Res. 866:15-22, Ma W, Bisby MA. Calcitonin gene-related peptide, substance P and protein gene product 9.5 immunoreactive axonal fibers in the rat footpad skin following partial sciatic nerve injuries. J Neurocytol. 29: , Mailis-Gagnon A, Bennett GJ. Abnormal contralateral pain responses from an intradermal injection of phenylephrine in a subset of patients with complex regional pain syndrome (CRPS). Pain. 111: , Maruo K, Yamamoto H, Yamamoto S, Nagata T, Fujikawa H, Kanno T, Yaguchi T, Maruo S, Yoshiya S, Nishizaki T. Modulation of P2X receptors via adrenergic pathways in rat dorsal root ganglion neurons after sciatic nerve injury. Pain. 120: , McCoy ES, Taylor-Blake B, Street SE, Pribisko AL, Zheng J, Zylka MJ. Peptidergic CGRPalpha Primary Sensory Neurons Encode Heat and Itch and Tonically Suppress Sensitivity to Cold. Neuron. 78: , Meisner JG, Waldron JB, Sawynok J. Alpha1-adrenergic receptors augment P2X3 receptormediated nociceptive responses in the uninjured state. J Pain. 8: , Munger BL, Bennett GJ, Kajander KC. An experimental painful peripheral neuropathy due to nerve constriction. I. Axonal pathology in the sciatic nerve. Exp Neurol. 118: , Nam TS, Yeon DS, Leem JW, Paik KS. Adrenergic sensitivity of uninjured C-fiber nociceptors in neuropathic rats. Yonsei Med J. 41: , Neil A, Attal N, Guilbaud G. Effects of guanethidine on sensitization to natural stimuli and self-mutilating behaviour in rats with a peripheral neuropathy. Brain Res. 565: , Obata K, Yamanaka H, Dai Y, Mizushima T, Fukuoka T, Tokunaga A, Noguchi K. Differential activation of MAPK in injured and uninjured DRG neurons following chronic constriction injury of the sciatic nerve in rats. Eur J Neurosci. 20: , Okamoto K, Martin DP, Schmelzer JD, Mitsui Y, Low PA. Pro- and anti-inflammatory cytokine gene expression in rat sciatic nerve chronic constriction injury model of neuropathic pain. Exp Neurol. 169: , 2001
17 Peleshok JC, Ribeiro-da-Silva A. Delayed reinnervation by nonpeptidergic nociceptive afferents of the glabrous skin of the rat hindpaw in a neuropathic pain model. J Comp Neurol. 519:49-63, Peleshok JC, Ribeiro-da-Silva A. Neurotrophic factor changes in the rat thick skin following chronic constriction injury of the sciatic nerve. Mol Pain. 8:1, Povlsen B, Hildebrand C, Stankovic N. Functional projection of sensory lateral plantar and superficial peroneal nerve axons to glabrous and hairy skin of the rat hindfoot after sciatic nerve lesions. Exp Neurol. 128: , Roggenkamp D, Falkner S, Stab F, Petersen M, Schmelz M, Neufang G. Atopic keratinocytes induce increased neurite outgrowth in a coculture model of porcine dorsal root ganglia neurons and human skin cells. The Journal of investigative dermatology. 132: , Tarpley JW, Kohler MG, Martin WJ. The behavioral and neuroanatomical effects of IB4- saporin treatment in rat models of nociceptive and neuropathic pain. Brain Res. 1029:65-76, Taylor AM, Osikowicz M, Ribeiro-da-Silva A. Consequences of the ablation of nonpeptidergic afferents in an animal model of trigeminal neuropathic pain. Pain. 153: , Torebjork E, Wahren L, Wallin G, Hallin R, Koltzenburg M. Noradrenaline-evoked pain in neuralgia. Pain. 63:11-20, Tripovic D, Pianova S, McLachlan EM, Brock JA. Transient supersensitivity to alphaadrenoceptor agonists, and distinct hyper-reactivity to vasopressin and angiotensin II after denervation of rat tail artery. Br J Pharmacol. 159: , Vilceanu D, Honore P, Hogan QH, Stucky CL. Spinal nerve ligation in mouse upregulates TRPV1 heat function in injured IB4-positive nociceptors. J Pain. 11: , Vulchanova L, Olson TH, Stone LS, Riedl MS, Elde R, Honda CN. Cytotoxic targeting of isolectin IB4-binding sensory neurons. Neuroscience. 108: , Wall JT, Cusick CG. Cutaneous responsiveness in primary somatosensory (S-I) hindpaw cortex before and after partial hindpaw deafferentation in adult rats. J Neurosci. 4: , Xanthos DN, Bennett GJ, Coderre TJ. Norepinephrine-induced nociception and vasoconstrictor hypersensitivity in rats with chronic post-ischemia pain. Pain. 137: , Xanthos DN, Coderre TJ. Sympathetic vasoconstrictor antagonism and vasodilatation relieve mechanical allodynia in rats with chronic postischemia pain. J Pain. 9: , Xie J, Ho Lee Y, Wang C, Mo Chung J, Chung K. Differential expression of alpha1- adrenoceptor subtype mrnas in the dorsal root ganglion after spinal nerve ligation. Brain Res Mol Brain Res. 93: , Yen LD, Bennett GJ, Ribeiro-da-Silva A. Sympathetic sprouting and changes in nociceptive sensory innervation in the glabrous skin of the rat hind paw following partial peripheral nerve injury. J Comp Neurol. 495: , Yoshimura T, Ito A, Saito SY, Takeda M, Kuriyama H, Ishikawa T. Calcitonin ameliorates enhanced arterial contractility after chronic constriction injury of the sciatic nerve in rats. Fundam Clin Pharmacol. 26: , Zhao P, Barr TP, Hou Q, Dib-Hajj SD, Black JA, Albrecht PJ, Petersen K, Eisenberg E, Wymer JP, Rice FL, Waxman SG. Voltage-gated sodium channel expression in rat and human epidermal keratinocytes: evidence for a role in pain. Pain. 139:90-105, 2008
18 17 Figure Legends Figure 1. Representative images of NF200 + fibers in nerve bundles in the reticular dermis of plantar and hairy skin. Dashed lines identify the perimeter of nerve bundles. Scale bar = 50 µm Figure 2. Representative images of α 1 -AR expression in nerve bundles in the reticular dermis of plantar skin. α 1 -AR expression was increased in nerve bundles in skin affected by CCI (lesioned) in comparison to skin on the contralateral limb to CCI (contra) and from naive animals. α 1 -AR immunoreactivity co-localised with the pan-neuronal marker TUJ1. Co-localized pixels are shown in white in the co-localized panel of images. Scale bar = 50 µm Figure 3. α 1 -AR immunoreactivity (expressed as Z-scores ± S.E.) in dermal TUJ + nerve fibers and on epidermal cells on the injured and contralateral sides after chronic constriction injury (N = 6) and in naïve animals (N = 3). A-C: In plantar skin, α 1 -AR immunoreactivity was greater on the injured than contralateral side in nerve fibers and on epidermal cells (* p<0.05), and was greater on the injured side than in uninjured naïve animals (#p<0.05). D-F: In the dorsal paw, α 1 -AR immunoreactivity was similar in injured and naïve animals. Figure 4. α 1 -AR immunoreactivity (expressed as Z-scores ± S.E.) in TUJ + nerve fibers in plantar skin co-labelled with CGRP, IB4 or NF200 after chronic constriction injury (N = 6) and in naïve animals (N = 3). A: α 1 -AR expression on fibers co-labelled with CGRP was greater on the injured than contralateral side (* p<0.05). B: α 1 -AR expression on fibers co-labelled with IB4 was greater on the injured than contralateral side (* p<0.05), and was greater on the injured side than in uninjured naïve animals (#p<0.05). C: Trends were similar in nerve fibers co-labelled with NF200. Figure 5. Representative images of α 1 -AR expression in the epidermis. α 1 -AR immunoreactivity was increased after CCI in plantar skin, but not hairy skin in the dorsal hind paw, in comparison to skin
19 18 contralateral to CCI and from naive animals. HF: indicates location of hair follicles in hairy skin. Scale bar = 100 µm Figure 6. Representative image of α 1 -AR expression in a dermal blood vessel stained with smooth muscle actin (SMA) (scale bar = 40 µm). α 1 -AR immunoreactivity (expressed as Z-scores ± S.E.) on cells co-labelled with SMA was similar on the injured and contralateral sides after chronic constriction injury (N = 6) and in naïve animals (N = 3) both (A) in plantar and (B) hairy skin.
20 Table 1: Primary and secondary antibodies Antibody Dilution Product code and Source anti α 1 -AR, rabbit polyclonal 1:200 A270, Sigma-Aldrich anti BIII-tubulin (TUJ1), mouse monoclonal 1:800 MMS-435P, Covance anti CGRP, goat polyclonal 1: , AbD Serotec IB4, FITC conjugate 1:250 L2895, Sigma-Aldrich anti NF200, chicken polyclonal 1:4000 Jackson ImmunoResearch anti-sma, mouse monoclonal 1:4000 A2547, Sigma-Aldrich anti-chicken Cy2 1:600 Jackson ImmunoResearch anti-goat 488 1:600 Jackson ImmunoResearch anti-rabbit 549 1:1200 Jackson ImmunoResearch anti-mouse 647 1:1000 Jackson ImmunoResearch
21 Table 2: Proportion of TUJ1 + pixels within nerve bundles after CCI Mean ± S.E. (% of total area) Naïve (N = 3) CCI ipsilateral (N = 6) CCI contralateral (N = 6) Plantar skin 21±4 11±3 17±5 Dorsal skin 32±6 17±3 30±6
22 Table 3: Neural markers expressed as a proportion of dermal TUJ1 + pixels Mean ± S.E. (% of TUJ1 + pixels) Naïve CCI ipsilateral (N = 3) (N = 6) Plantar skin CCI contralateral (N = 6) CGRP 35±6 37±8 40±5 IB4 35±1 34±5 37±7 NF200 28±6 6±2 * 24±5 Dorsal skin CGRP 38±19 45±13 40±8 IB4 38±5 36±8 51±11 NF200 23±3 23±5 24±5 * p<0.05 compared with CCI contralateral and naïve animals


23

24
25
26

27

28
MURDOCH RESEARCH REPOSITORY.
 MURDOCH RESEARCH REPOSITORY http://researchrepository.murdoch.edu.au This is the author's final version of the work, as accepted for publication following peer review but without the publisher's layout
MURDOCH RESEARCH REPOSITORY http://researchrepository.murdoch.edu.au This is the author's final version of the work, as accepted for publication following peer review but without the publisher's layout
Accepted Manuscript. S (13) Reference: PAIN To appear in: PAIN
 Reference: PAIN To appear in: PAIN") Accepted Manuscript Up-regulation of α 1 -adrenoceptors on cutaneous nerve fibres after partial sciatic nerve ligation and in complex regional pain syndrome type II Peter D. Drummond, Eleanor S. Drummond,
Accepted Manuscript Up-regulation of α 1 -adrenoceptors on cutaneous nerve fibres after partial sciatic nerve ligation and in complex regional pain syndrome type II Peter D. Drummond, Eleanor S. Drummond,
A role for uninjured afferents in neuropathic pain
 Acta Physiologica Sinica, October 25, 2008, 60 (5): 605-609 http://www.actaps.com.cn 605 Review A role for uninjured afferents in neuropathic pain Richard A. Meyer 1,2,3,*, Matthias Ringkamp 1 Departments
Acta Physiologica Sinica, October 25, 2008, 60 (5): 605-609 http://www.actaps.com.cn 605 Review A role for uninjured afferents in neuropathic pain Richard A. Meyer 1,2,3,*, Matthias Ringkamp 1 Departments
Mechanical sensitization of cutaneous sensory fibers in the spared nerve injury mouse model
 Smith et al. Molecular Pain 2013, 9:61 MOLECULAR PAIN SHORT REPORT Open Access Mechanical sensitization of cutaneous sensory fibers in the spared nerve injury mouse model Amanda K Smith, Crystal L O Hara
Smith et al. Molecular Pain 2013, 9:61 MOLECULAR PAIN SHORT REPORT Open Access Mechanical sensitization of cutaneous sensory fibers in the spared nerve injury mouse model Amanda K Smith, Crystal L O Hara
Enhanced formalin nociceptive responses following L5 nerve ligation in the rat reveals neuropathy-induced inflammatory hyperalgesia
 University of Kentucky From the SelectedWorks of Renee R. Donahue 2001 Enhanced formalin nociceptive responses following L5 nerve ligation in the rat reveals neuropathy-induced inflammatory hyperalgesia
University of Kentucky From the SelectedWorks of Renee R. Donahue 2001 Enhanced formalin nociceptive responses following L5 nerve ligation in the rat reveals neuropathy-induced inflammatory hyperalgesia
Animal Model of Trigeminal Neuralgia Induced by Chronic Constriction Injury Applied to the Ophthalmic Nerve in the Rat
 Showa Univ. J. Med. Sci. 9(2), 8995, December 1997 Original Animal Model of Trigeminal Neuralgia Induced by Chronic Constriction Injury Applied to the Ophthalmic Nerve in the Rat Kazuo HANAKAWA, Takao
Showa Univ. J. Med. Sci. 9(2), 8995, December 1997 Original Animal Model of Trigeminal Neuralgia Induced by Chronic Constriction Injury Applied to the Ophthalmic Nerve in the Rat Kazuo HANAKAWA, Takao
Sensory coding and somatosensory system
 Sensory coding and somatosensory system Sensation and perception Perception is the internal construction of sensation. Perception depends on the individual experience. Three common steps in all senses
Sensory coding and somatosensory system Sensation and perception Perception is the internal construction of sensation. Perception depends on the individual experience. Three common steps in all senses
NIH Public Access Author Manuscript J Neuropathic Pain Symptom Palliation. Author manuscript; available in PMC 2007 March 26.
 NIH Public Access Author Manuscript Published in final edited form as: J Neuropathic Pain Symptom Palliation. 2005 ; 1(1): 19 23. Sympathetic Fiber Sprouting in Chronically Compressed Dorsal Root Ganglia
NIH Public Access Author Manuscript Published in final edited form as: J Neuropathic Pain Symptom Palliation. 2005 ; 1(1): 19 23. Sympathetic Fiber Sprouting in Chronically Compressed Dorsal Root Ganglia
RESEARCH REPOSITORY.
 RESEARCH REPOSITORY This is the author s final version of the work, as accepted for publication following peer review but without the publisher s layout or pagination. The definitive version is available
RESEARCH REPOSITORY This is the author s final version of the work, as accepted for publication following peer review but without the publisher s layout or pagination. The definitive version is available
Sympathetic fibre sprouting in the skin contributes to pain related behaviour in spared nerve injury and cuff models of neuropathic pain
 DOI 10.1186/s12990-015-0062-x RESEARCH Open Access Sympathetic fibre sprouting in the skin contributes to pain related behaviour in spared nerve injury and cuff models of neuropathic pain Francisney P.
DOI 10.1186/s12990-015-0062-x RESEARCH Open Access Sympathetic fibre sprouting in the skin contributes to pain related behaviour in spared nerve injury and cuff models of neuropathic pain Francisney P.
Original Research Article Up-Regulation of Cutaneous α 1 -Adrenoceptors in Complex Regional Pain Syndrome Type I
 bs_bs_banner Pain Medicine 2014; 15: 1945 1956 Wiley Periodicals, Inc. NEUROPATHIC PAIN SECTION Original Research Article Up-Regulation of Cutaneous α 1 -Adrenoceptors in Complex Regional Pain Syndrome
bs_bs_banner Pain Medicine 2014; 15: 1945 1956 Wiley Periodicals, Inc. NEUROPATHIC PAIN SECTION Original Research Article Up-Regulation of Cutaneous α 1 -Adrenoceptors in Complex Regional Pain Syndrome
P2X3 receptors are present on sensory afferent neurons. Nociceptive Responses in the Uninjured State
 The Journal of Pain, Vol 8, No 7 (July), 2007: pp 556-562 Available online at www.sciencedirect.com 1 -Adrenergic Receptors Augment P2X 3 Receptor Mediated Nociceptive Responses in the Uninjured State
The Journal of Pain, Vol 8, No 7 (July), 2007: pp 556-562 Available online at www.sciencedirect.com 1 -Adrenergic Receptors Augment P2X 3 Receptor Mediated Nociceptive Responses in the Uninjured State
Introduction to some interesting research questions: Molecular biology of the primary afferent nociceptor
 Introduction to some interesting research questions: Molecular biology of the primary afferent nociceptor NOCICEPTORS ARE NOT IDENTICAL PEPTIDE SubP/CGRP Trk A NON-PEPTIDE IB4 P2X 3 c-ret Snider and McMahon
Introduction to some interesting research questions: Molecular biology of the primary afferent nociceptor NOCICEPTORS ARE NOT IDENTICAL PEPTIDE SubP/CGRP Trk A NON-PEPTIDE IB4 P2X 3 c-ret Snider and McMahon
Diabetic Complications Consortium
 Diabetic Complications Consortium Application Title: Cathepsin S inhibition and diabetic neuropathy Principal Investigator: Nigel A Calcutt 1. Project Accomplishments: We investigated the efficacy of cathepsin
Diabetic Complications Consortium Application Title: Cathepsin S inhibition and diabetic neuropathy Principal Investigator: Nigel A Calcutt 1. Project Accomplishments: We investigated the efficacy of cathepsin
Update on the Neurophysiology of Pain Transmission and Modulation: Focus on the NMDA-Receptor
 S2 Journal of Pain and Symptom Management Vol. 19 No. 1(Suppl.) January 2000 Proceedings Supplement NMDA-Receptor Antagonists: Evolving Role in Analgesia Update on the Neurophysiology of Pain Transmission
S2 Journal of Pain and Symptom Management Vol. 19 No. 1(Suppl.) January 2000 Proceedings Supplement NMDA-Receptor Antagonists: Evolving Role in Analgesia Update on the Neurophysiology of Pain Transmission
Repeated intra-articular injections of acidic saline produce long-lasting joint pain and. widespread hyperalgesia
 Repeated intra-articular injections of acidic saline produce long-lasting joint pain and widespread hyperalgesia N. Sugimura 1, M. Ikeuchi 1 *, M. Izumi 1, T. Kawano 2, K. Aso 1, T. Kato 1, T. Ushida 3,
Repeated intra-articular injections of acidic saline produce long-lasting joint pain and widespread hyperalgesia N. Sugimura 1, M. Ikeuchi 1 *, M. Izumi 1, T. Kawano 2, K. Aso 1, T. Kato 1, T. Ushida 3,
The biochemical origin of pain: The origin of all pain is inflammation and the inflammatory response: Inflammatory profile of pain syndromes
 The biochemical origin of pain: The origin of all pain is inflammation and the inflammatory response: Inflammatory profile of pain syndromes 1 Medical Hypothesis 2007, Vol. 69, pp. 1169 1178 Sota Omoigui
The biochemical origin of pain: The origin of all pain is inflammation and the inflammatory response: Inflammatory profile of pain syndromes 1 Medical Hypothesis 2007, Vol. 69, pp. 1169 1178 Sota Omoigui
A subpopulation of nociceptors specifically linked to itch
 Supplementary Information A subpopulation of nociceptors specifically linked to itch Liang Han 1, Chao Ma 3,4, Qin Liu 1,2, Hao-Jui Weng 1,2, Yiyuan Cui 5, Zongxiang Tang 1,2, Yushin Kim 1, Hong Nie 4,
Supplementary Information A subpopulation of nociceptors specifically linked to itch Liang Han 1, Chao Ma 3,4, Qin Liu 1,2, Hao-Jui Weng 1,2, Yiyuan Cui 5, Zongxiang Tang 1,2, Yushin Kim 1, Hong Nie 4,
MURDOCH RESEARCH REPOSITORY.
 MURDOCH RESEARCH REPOSITORY http://researchrepository.murdoch.edu.au This is the author's final version of the work, as accepted for publication following peer review but without the publisher's layout
MURDOCH RESEARCH REPOSITORY http://researchrepository.murdoch.edu.au This is the author's final version of the work, as accepted for publication following peer review but without the publisher's layout
A Review of Neuropathic Pain: From Diagnostic Tests to Mechanisms
 DOI 10.1007/s40122-017-0085-2 REVIEW A Review of Neuropathic Pain: From Diagnostic Tests to Mechanisms Andrea Truini Received: September 19, 2017 Ó The Author(s) 2017. This article is an open access publication
DOI 10.1007/s40122-017-0085-2 REVIEW A Review of Neuropathic Pain: From Diagnostic Tests to Mechanisms Andrea Truini Received: September 19, 2017 Ó The Author(s) 2017. This article is an open access publication
Cancer-induced bone pain
 Cancer-induced bone pain Common Prevalent in particular cancers: breast (73%), prostate (68%), thyroid (42%), lung (36%), renal (35%), colon (5%) Correlates with an increased morbidity Reduced performance
Cancer-induced bone pain Common Prevalent in particular cancers: breast (73%), prostate (68%), thyroid (42%), lung (36%), renal (35%), colon (5%) Correlates with an increased morbidity Reduced performance
David S. Warner, M.D., Editor. Testing the Link between Sympathetic Efferent and Sensory Afferent Fibers in Neuropathic Pain
 David S. Warner, M.D., Editor Testing the Link between Sympathetic Efferent and Sensory Afferent Fibers in Neuropathic Pain Srinivasa N. Raja, M.D.,* Rolf-Detlef Treede, M.D., Ph.D. Systemic -adrenergic
David S. Warner, M.D., Editor Testing the Link between Sympathetic Efferent and Sensory Afferent Fibers in Neuropathic Pain Srinivasa N. Raja, M.D.,* Rolf-Detlef Treede, M.D., Ph.D. Systemic -adrenergic
The Egyptian Journal of Hospital Medicine (January 2018) Vol. 70 (12), Page
 Vol. 70 (12), Page") The Egyptian Journal of Hospital Medicine (January 2018) Vol. 70 (12), Page 2172-2177 Blockage of HCN Channels with ZD7288 Attenuates Mechanical Hypersensitivity in Rats Model of Diabetic Neuropathy Hussain
The Egyptian Journal of Hospital Medicine (January 2018) Vol. 70 (12), Page 2172-2177 Blockage of HCN Channels with ZD7288 Attenuates Mechanical Hypersensitivity in Rats Model of Diabetic Neuropathy Hussain
Introduction ORIGINAL ARTICLE. Seiji Ohtori Kazuhisa Takahashi Hideshige Moriya
 Eur Spine J (2003) 12 :211 215 DOI 10.1007/s00586-002-0506-7 ORIGINAL ARTICLE Seiji Ohtori Kazuhisa Takahashi Hideshige Moriya Calcitonin gene-related peptide immunoreactive DRG neurons innervating the
Eur Spine J (2003) 12 :211 215 DOI 10.1007/s00586-002-0506-7 ORIGINAL ARTICLE Seiji Ohtori Kazuhisa Takahashi Hideshige Moriya Calcitonin gene-related peptide immunoreactive DRG neurons innervating the
Somatic Sensation (MCB160 Lecture by Mu-ming Poo, Friday March 9, 2007)
") Somatic Sensation (MCB160 Lecture by Mu-ming Poo, Friday March 9, 2007) Introduction Adrian s work on sensory coding Spinal cord and dorsal root ganglia Four somatic sense modalities Touch Mechanoreceptors
Somatic Sensation (MCB160 Lecture by Mu-ming Poo, Friday March 9, 2007) Introduction Adrian s work on sensory coding Spinal cord and dorsal root ganglia Four somatic sense modalities Touch Mechanoreceptors
Seizure: the clinical manifestation of an abnormal and excessive excitation and synchronization of a population of cortical
 Are There Sharing Mechanisms of Epilepsy, Migraine and Neuropathic Pain? Chin-Wei Huang, MD, PhD Department of Neurology, NCKUH Basic mechanisms underlying seizures and epilepsy Seizure: the clinical manifestation
Are There Sharing Mechanisms of Epilepsy, Migraine and Neuropathic Pain? Chin-Wei Huang, MD, PhD Department of Neurology, NCKUH Basic mechanisms underlying seizures and epilepsy Seizure: the clinical manifestation
effect on the upregulation of these cell surface markers. The mean peak fluorescence intensity
 SUPPLEMENTARY FIGURE 1 Supplementary Figure 1 ASIC1 disruption or blockade does not effect in vitro and in vivo antigen-presenting cell activation. (a) Flow cytometric analysis of cell surface molecules
SUPPLEMENTARY FIGURE 1 Supplementary Figure 1 ASIC1 disruption or blockade does not effect in vitro and in vivo antigen-presenting cell activation. (a) Flow cytometric analysis of cell surface molecules
Nature Neuroscience: doi: /nn Supplementary Figure 1
 Supplementary Figure 1 Atlas representations of the midcingulate (MCC) region targeted in this study compared against the anterior cingulate (ACC) region commonly reported. Coronal sections are shown on
Supplementary Figure 1 Atlas representations of the midcingulate (MCC) region targeted in this study compared against the anterior cingulate (ACC) region commonly reported. Coronal sections are shown on
Somatosensory System. Steven McLoon Department of Neuroscience University of Minnesota
 Somatosensory System Steven McLoon Department of Neuroscience University of Minnesota 1 Course News Dr. Riedl s review session this week: Tuesday (Oct 10) 4-5pm in MCB 3-146B 2 Sensory Systems Sensory
Somatosensory System Steven McLoon Department of Neuroscience University of Minnesota 1 Course News Dr. Riedl s review session this week: Tuesday (Oct 10) 4-5pm in MCB 3-146B 2 Sensory Systems Sensory
Nerve Demyelination Increases Metabotropic Glutamate Receptor Subtype 5 Expression in Peripheral Painful Mononeuropathy
 Int. J. Mol. Sci. 2015, 16, 4642-4665; doi:10.3390/ijms16034642 Article OPEN ACCESS International Journal of Molecular Sciences ISSN 1422-0067 www.mdpi.com/journal/ijms Nerve Demyelination Increases Metabotropic
Int. J. Mol. Sci. 2015, 16, 4642-4665; doi:10.3390/ijms16034642 Article OPEN ACCESS International Journal of Molecular Sciences ISSN 1422-0067 www.mdpi.com/journal/ijms Nerve Demyelination Increases Metabotropic
Pathophysiology of Pain
 Pathophysiology of Pain Wound Inflammatory response Chemical mediators Activity in Pain Path PAIN http://neuroscience.uth.tmc.edu/s2/chapter08.html Chris Cohan, Ph.D. Dept. of Pathology/Anat Sci University
Pathophysiology of Pain Wound Inflammatory response Chemical mediators Activity in Pain Path PAIN http://neuroscience.uth.tmc.edu/s2/chapter08.html Chris Cohan, Ph.D. Dept. of Pathology/Anat Sci University
211MDS Pain theories
 211MDS Pain theories Definition In 1986, the International Association for the Study of Pain (IASP) defined pain as a sensory and emotional experience associated with real or potential injuries, or described
211MDS Pain theories Definition In 1986, the International Association for the Study of Pain (IASP) defined pain as a sensory and emotional experience associated with real or potential injuries, or described
What is on the Horizon in Drug Therapy for OAB?
 What is on the Horizon in Drug Therapy for OAB? K-E Andersson, MD, PhD Wake Forest Institute for Regenerative Medicine Wake Forest University School of Medicine Winston Salem, North Carolina Disclosures
What is on the Horizon in Drug Therapy for OAB? K-E Andersson, MD, PhD Wake Forest Institute for Regenerative Medicine Wake Forest University School of Medicine Winston Salem, North Carolina Disclosures
Somatosensory modalities!
 Somatosensory modalities! The somatosensory system codes five major sensory modalities:! 1. Discriminative touch! 2. Proprioception (body position and motion)! 3. Nociception (pain and itch)! 4. Temperature!
Somatosensory modalities! The somatosensory system codes five major sensory modalities:! 1. Discriminative touch! 2. Proprioception (body position and motion)! 3. Nociception (pain and itch)! 4. Temperature!
Bi/CNS/NB 150: Neuroscience. November 11, 2015 SOMATOSENSORY SYSTEM. Ralph Adolphs
 Bi/CNS/NB 150: Neuroscience November 11, 2015 SOMATOSENSORY SYSTEM Ralph Adolphs 1 Menu for today Touch -peripheral -central -plasticity Pain 2 Sherrington (1948): senses classified as --teloreceptive
Bi/CNS/NB 150: Neuroscience November 11, 2015 SOMATOSENSORY SYSTEM Ralph Adolphs 1 Menu for today Touch -peripheral -central -plasticity Pain 2 Sherrington (1948): senses classified as --teloreceptive
Norepinephrine-induced nociception and vasoconstrictor hypersensitivity in rats with chronic post-ischemia pain
 Pain 137 (2008) 640 651 www.elsevier.com/locate/pain Norepinephrine-induced nociception and vasoconstrictor hypersensitivity in rats with chronic post-ischemia pain Dimitris N. Xanthos a,e, Gary J. Bennett
Pain 137 (2008) 640 651 www.elsevier.com/locate/pain Norepinephrine-induced nociception and vasoconstrictor hypersensitivity in rats with chronic post-ischemia pain Dimitris N. Xanthos a,e, Gary J. Bennett
Chapter 13 PNS and reflex activity
 Chapter 13 PNS and reflex activity I. Peripheral nervous system A. PNS links CNS to the body B. Sensory: the afferent division C. Motor: the efferent division D. Ganglia: collections of cell bodies in
Chapter 13 PNS and reflex activity I. Peripheral nervous system A. PNS links CNS to the body B. Sensory: the afferent division C. Motor: the efferent division D. Ganglia: collections of cell bodies in
CHAPTER 10 THE SOMATOSENSORY SYSTEM
 CHAPTER 10 THE SOMATOSENSORY SYSTEM 10.1. SOMATOSENSORY MODALITIES "Somatosensory" is really a catch-all term to designate senses other than vision, hearing, balance, taste and smell. Receptors that could
CHAPTER 10 THE SOMATOSENSORY SYSTEM 10.1. SOMATOSENSORY MODALITIES "Somatosensory" is really a catch-all term to designate senses other than vision, hearing, balance, taste and smell. Receptors that could
Supplementary Figure 1
 Supplementary Figure 1 The average sigmoid parametric curves of capillary dilation time courses and average time to 50% peak capillary diameter dilation computed from individual capillary responses averaged
Supplementary Figure 1 The average sigmoid parametric curves of capillary dilation time courses and average time to 50% peak capillary diameter dilation computed from individual capillary responses averaged
Published online October 10, 2016
 Published online October 10, 2016 reviewed by Jim C. Eisenach, Wake Forest University; Ronald Lindsay, Zebra Biologics; Remi Quirion, McGill University; and Tony L. Yaksh, University of California, San
Published online October 10, 2016 reviewed by Jim C. Eisenach, Wake Forest University; Ronald Lindsay, Zebra Biologics; Remi Quirion, McGill University; and Tony L. Yaksh, University of California, San
Analgesia Mediated by the TRPM8 Cold Receptor in Chronic Neuropathic Pain
 Current Biology 16, 1591 1605, August 22, 2006 ª2006 Elsevier Ltd All rights reserved DOI 10.1016/j.cub.2006.07.061 Analgesia Mediated by the TRPM8 Cold Receptor in Chronic Neuropathic Pain Article Clare
Current Biology 16, 1591 1605, August 22, 2006 ª2006 Elsevier Ltd All rights reserved DOI 10.1016/j.cub.2006.07.061 Analgesia Mediated by the TRPM8 Cold Receptor in Chronic Neuropathic Pain Article Clare
Pain teaching. Muhammad Laklouk
 Pain teaching Muhammad Laklouk Definition Pain An unpleasant sensory and emotional experience associated with actual or potential tissue damage or described in terms of such damage. Sensory (discriminatiory)
Pain teaching Muhammad Laklouk Definition Pain An unpleasant sensory and emotional experience associated with actual or potential tissue damage or described in terms of such damage. Sensory (discriminatiory)
Potential for delta opioid receptor agonists as analgesics in chronic pain therapy
 Potential for delta opioid receptor agonists as analgesics in chronic pain therapy David Kendall & Bengt von Mentzer; PharmNovo AB/UK Alex Conibear & Eamonn Kelly, University of Bristol Junaid Asghar,
Potential for delta opioid receptor agonists as analgesics in chronic pain therapy David Kendall & Bengt von Mentzer; PharmNovo AB/UK Alex Conibear & Eamonn Kelly, University of Bristol Junaid Asghar,
Supplemental Information. Otic Mesenchyme Cells Regulate. Spiral Ganglion Axon Fasciculation. through a Pou3f4/EphA4 Signaling Pathway
 Neuron, Volume 73 Supplemental Information Otic Mesenchyme Cells Regulate Spiral Ganglion Axon Fasciculation through a Pou3f4/EphA4 Signaling Pathway Thomas M. Coate, Steven Raft, Xiumei Zhao, Aimee K.
Neuron, Volume 73 Supplemental Information Otic Mesenchyme Cells Regulate Spiral Ganglion Axon Fasciculation through a Pou3f4/EphA4 Signaling Pathway Thomas M. Coate, Steven Raft, Xiumei Zhao, Aimee K.
Ch 9. The Autonomic Nervous System
 Ch 9 The Autonomic Nervous System SLOs Review the organization of the ANS Describe how neural regulation of smooth and cardiac muscles differs from that of skeletal muscles Describe the structure and innervation
Ch 9 The Autonomic Nervous System SLOs Review the organization of the ANS Describe how neural regulation of smooth and cardiac muscles differs from that of skeletal muscles Describe the structure and innervation
MYOFASCIAL PAIN. Dr. Janet Travell ( ) credited with bringing MTrPs to the attention of healthcare providers.
 credited with bringing MTrPs to the attention of healthcare providers.") Myofascial Trigger Points background info Laurie Edge-Hughes BScPT, MAnimSt (Animal Physio), CAFCI, CCRT History lesson Dr. Janet Travell (1901 1997) credited with bringing MTrPs to the attention of healthcare
Myofascial Trigger Points background info Laurie Edge-Hughes BScPT, MAnimSt (Animal Physio), CAFCI, CCRT History lesson Dr. Janet Travell (1901 1997) credited with bringing MTrPs to the attention of healthcare
What is Pain? An unpleasant sensory and emotional experience associated with actual or potential tissue damage. Pain is always subjective
 Pain & Acupuncture What is Pain? An unpleasant sensory and emotional experience associated with actual or potential tissue damage. NOCICEPTION( the neural processes of encoding and processing noxious stimuli.)
Pain & Acupuncture What is Pain? An unpleasant sensory and emotional experience associated with actual or potential tissue damage. NOCICEPTION( the neural processes of encoding and processing noxious stimuli.)
Product Datasheet. Vanilloid R1/TRPV1 Antibody NB Unit Size: 0.05 ml. Aliquot and store at -20C or -80C. Avoid freeze-thaw cycles.
 Product Datasheet Vanilloid R1/TRPV1 Antibody NB100-1617 Unit Size: 0.05 ml Aliquot and store at -20C or -80C. Avoid freeze-thaw cycles. Publications: 15 Protocols, Publications, Related Products, Reviews,
Product Datasheet Vanilloid R1/TRPV1 Antibody NB100-1617 Unit Size: 0.05 ml Aliquot and store at -20C or -80C. Avoid freeze-thaw cycles. Publications: 15 Protocols, Publications, Related Products, Reviews,
Expression of the transient receptor potential channels TRPV1, TRPA1 and TRPM8 in mouse trigeminal primary afferent neurons innervating the dura
 Washington University School of Medicine Digital Commons@Becker Open Access Publications 212 Expression of the transient receptor potential channels TRPV1, TRPA1 and TRPM8 in mouse trigeminal primary afferent
Washington University School of Medicine Digital Commons@Becker Open Access Publications 212 Expression of the transient receptor potential channels TRPV1, TRPA1 and TRPM8 in mouse trigeminal primary afferent
Proceedings of the World Small Animal Veterinary Association Sydney, Australia 2007
 Proceedings of the World Small Animal Sydney, Australia 2007 Hosted by: Next WSAVA Congress REDUCING THE PAIN FACTOR AN UPDATE ON PERI-OPERATIVE ANALGESIA Sandra Forysth, BVSc DipACVA Institute of Veterinary,
Proceedings of the World Small Animal Sydney, Australia 2007 Hosted by: Next WSAVA Congress REDUCING THE PAIN FACTOR AN UPDATE ON PERI-OPERATIVE ANALGESIA Sandra Forysth, BVSc DipACVA Institute of Veterinary,
Pharmacology of Pain Transmission and Modulation
 Pharmacology of Pain Transmission and Modulation 2 Jürg Schliessbach and Konrad Maurer Nociceptive Nerve Fibers Pain is transmitted to the central nervous system via thinly myelinated Aδ and unmyelinated
Pharmacology of Pain Transmission and Modulation 2 Jürg Schliessbach and Konrad Maurer Nociceptive Nerve Fibers Pain is transmitted to the central nervous system via thinly myelinated Aδ and unmyelinated
Neuropathic pain, pain matrix dysfunction, and pain syndromes
 Neuropathic pain, pain matrix dysfunction, and pain syndromes MSTN121 - Neurophysiology Session 3 Department of Myotherapy Session objectives Describe the mechanism of nociceptive chronic pain. Define
Neuropathic pain, pain matrix dysfunction, and pain syndromes MSTN121 - Neurophysiology Session 3 Department of Myotherapy Session objectives Describe the mechanism of nociceptive chronic pain. Define
EDUCATION M.D., Peking Union Medical College, Beijing, China, 1999 B.S., Beijing University, College of Life Science, Beijing, China, 1994
 CHAO MA, M.D. Yale University School of Medicine, Department of Anesthesiology, 333 Cedar Street, TMP3, New Haven, CT 06510, USA. Phone: 203-785-3522 (O), 203-606-7959 (C), Fax: 203-737-1528, Email: chao.ma@yale.edu
CHAO MA, M.D. Yale University School of Medicine, Department of Anesthesiology, 333 Cedar Street, TMP3, New Haven, CT 06510, USA. Phone: 203-785-3522 (O), 203-606-7959 (C), Fax: 203-737-1528, Email: chao.ma@yale.edu
EFFECTS OF Pueraria mirifica ON NEUROPATHIC PAIN IN RATS
 EFFECTS OF Pueraria mirifica ON NEUROPATHIC PAIN IN RATS Kamolchanok Tanchotikul 1, Quankamon Dejativongse Na Ayudhya 1*, Orawan Piyaboon 1 and Supin Chompoopong 2 1 Department of Biology, Mahidol Wittayanusorn
EFFECTS OF Pueraria mirifica ON NEUROPATHIC PAIN IN RATS Kamolchanok Tanchotikul 1, Quankamon Dejativongse Na Ayudhya 1*, Orawan Piyaboon 1 and Supin Chompoopong 2 1 Department of Biology, Mahidol Wittayanusorn
NURSE-UP INTRODUCTION TO THE NERVOUS SYSTEM
 NURSE-UP INTRODUCTION TO THE NERVOUS SYSTEM FUNCTIONS OF THE NERVOUS SYSTEM Body s primary communication and control system. Integrates and regulates body function Collects information specialized nervous
NURSE-UP INTRODUCTION TO THE NERVOUS SYSTEM FUNCTIONS OF THE NERVOUS SYSTEM Body s primary communication and control system. Integrates and regulates body function Collects information specialized nervous
What is pain?: An unpleasant sensation. What is an unpleasant sensation?: Pain. - Aristotle.
 What is pain?: An unpleasant sensation. What is an unpleasant sensation?: Pain. - Aristotle. Nociception The detection of tissue damage or impending tissue damage, but There can be tissue damage without
What is pain?: An unpleasant sensation. What is an unpleasant sensation?: Pain. - Aristotle. Nociception The detection of tissue damage or impending tissue damage, but There can be tissue damage without
Special Issue on Pain and Itch
 Special Issue on Pain and Itch Title: Recent Progress in Understanding the Mechanisms of Pain and Itch Guest Editor of the Special Issue: Ru-Rong Ji, PhD Chronic pain is a major health problem world-wide.
Special Issue on Pain and Itch Title: Recent Progress in Understanding the Mechanisms of Pain and Itch Guest Editor of the Special Issue: Ru-Rong Ji, PhD Chronic pain is a major health problem world-wide.
Centrally Mediated Anti-Hyperalgesic and Anti-Allodynic Effect of Tolperisone in Spared Nerve Injury Model of Neuropathic Pain
 Centrally Mediated Anti-Hyperalgesic and Anti-Allodynic Effect of Tolperisone in Spared Nerve Injury Model of Neuropathic Pain Keyur S Patel *, Punam D Sachdeva Department of Pharmacology, A.R.College
Centrally Mediated Anti-Hyperalgesic and Anti-Allodynic Effect of Tolperisone in Spared Nerve Injury Model of Neuropathic Pain Keyur S Patel *, Punam D Sachdeva Department of Pharmacology, A.R.College
Accepted Manuscript. Stephanie Kao, BA, Alexi Kiss, MD, Tatiana Efimova, PhD, Adam Friedman, MD
 Accepted Manuscript An ex-vivo evaluation of cytotoxicity and melanocyte viability after A-101 hydrogen peroxide topical solution 40% or cryosurgery treatment in Seborrheic Keratosis lesions Stephanie
Accepted Manuscript An ex-vivo evaluation of cytotoxicity and melanocyte viability after A-101 hydrogen peroxide topical solution 40% or cryosurgery treatment in Seborrheic Keratosis lesions Stephanie
Receptor Subtype Mediating the Adrenergic Sensitivity of Pain Behavior and Ectopic Discharges in Neuropathic Lewis Rats
 Receptor Subtype Mediating the Adrenergic Sensitivity of Pain Behavior and Ectopic Discharges in Neuropathic Lewis Rats DOO HYUN LEE, 1 XIANZENG LIU, 1 HYUN TAEK KIM, 1 KYUNGSOON CHUNG, 1,2 AND JIN MO
Receptor Subtype Mediating the Adrenergic Sensitivity of Pain Behavior and Ectopic Discharges in Neuropathic Lewis Rats DOO HYUN LEE, 1 XIANZENG LIU, 1 HYUN TAEK KIM, 1 KYUNGSOON CHUNG, 1,2 AND JIN MO
Effects of decompression on neuropathic pain behaviors and skin reinnervation in chronic constriction injury
 Experimental Neurology 204 (2007) 574 582 www.elsevier.com/locate/yexnr Effects of decompression on neuropathic pain behaviors and skin reinnervation in chronic constriction injury To-Jung Tseng a, Chih-Cheng
Experimental Neurology 204 (2007) 574 582 www.elsevier.com/locate/yexnr Effects of decompression on neuropathic pain behaviors and skin reinnervation in chronic constriction injury To-Jung Tseng a, Chih-Cheng
Degeneration of Myelinated Efferent Fibers Induces Spontaneous Activity in Uninjured C-Fiber Afferents
 The Journal of Neuroscience, September 1, 2002, 22(17):7746 7753 Degeneration of Myelinated Efferent Fibers Induces Spontaneous Activity in Uninjured C-Fiber Afferents Gang Wu, 1 Matthias Ringkamp, 1 Beth
The Journal of Neuroscience, September 1, 2002, 22(17):7746 7753 Degeneration of Myelinated Efferent Fibers Induces Spontaneous Activity in Uninjured C-Fiber Afferents Gang Wu, 1 Matthias Ringkamp, 1 Beth
Risk Factors That Predispose Patients to Orofacial Pain
 Risk Factors That Predispose Patients to Orofacial Pain Paul Durham, PhD Distinguished Professor of Cell Biology Director Center for Biomedical & Life Sciences Missouri State University Cluster Headache
Risk Factors That Predispose Patients to Orofacial Pain Paul Durham, PhD Distinguished Professor of Cell Biology Director Center for Biomedical & Life Sciences Missouri State University Cluster Headache
Pain Mechanisms. Prof Michael G Irwin MD, FRCA, FANZCA FHKAM Head Department of Anaesthesiology University of Hong Kong. The Somatosensory System
 ain Mechanisms rof Michael G Irwin MD, FRCA, FANZCA FHKAM Head Department of Anaesthesiology University of Hong Kong The Somatosensory System Frontal cortex Descending pathway eriaqueductal gray matter
ain Mechanisms rof Michael G Irwin MD, FRCA, FANZCA FHKAM Head Department of Anaesthesiology University of Hong Kong The Somatosensory System Frontal cortex Descending pathway eriaqueductal gray matter
Peripheral nerve injury alters excitatory synaptic transmission in lamina II of the rat dorsal horn
 J Physiol (2003), 548.1, pp. 131 138 DOI: 10.1113/jphysiol.2002.036186 The Physiological Society 2003 www.jphysiol.org Peripheral nerve injury alters excitatory synaptic transmission in lamina II of the
J Physiol (2003), 548.1, pp. 131 138 DOI: 10.1113/jphysiol.2002.036186 The Physiological Society 2003 www.jphysiol.org Peripheral nerve injury alters excitatory synaptic transmission in lamina II of the
Autonomic Nervous System
 ANS..??? Autonomic Nervous System Nervous system CNS PNS Autonomic Somatic Symp Parasymp Enteric SOMATIC AUTONOMIC Organ supplied Skeletal muscle Other organs Distal most synapse Nerve fibre Peripheral
ANS..??? Autonomic Nervous System Nervous system CNS PNS Autonomic Somatic Symp Parasymp Enteric SOMATIC AUTONOMIC Organ supplied Skeletal muscle Other organs Distal most synapse Nerve fibre Peripheral
Effect of Propecia on the Hair Follicle in Male Androgenetic Alopecia: A Confocal Laser Scanning Microscopy and Video Imaging Study
 Effect of Propecia on the Hair Follicle in Male Androgenetic Alopecia: A Confocal Laser Scanning Microscopy and Video Imaging Study Investigators: Department of Dermatology, University of Minnesota, Minneapolis,
Effect of Propecia on the Hair Follicle in Male Androgenetic Alopecia: A Confocal Laser Scanning Microscopy and Video Imaging Study Investigators: Department of Dermatology, University of Minnesota, Minneapolis,
Module H NERVOUS SYSTEM
 Module H NERVOUS SYSTEM Topic from General functions of the nervous system Organization of the nervous system from both anatomical & functional perspectives Gross & microscopic anatomy of nervous tissue
Module H NERVOUS SYSTEM Topic from General functions of the nervous system Organization of the nervous system from both anatomical & functional perspectives Gross & microscopic anatomy of nervous tissue
Venipuncture-induced neuropathic pain: the clinical syndrome, with comparisons to experimental nerve injury models
 Pain 94 (2001) 225 229 Topical review Venipuncture-induced neuropathic pain: the clinical syndrome, with comparisons to experimental nerve injury models Steven H. Horowitz* Division of Neurology, University
Pain 94 (2001) 225 229 Topical review Venipuncture-induced neuropathic pain: the clinical syndrome, with comparisons to experimental nerve injury models Steven H. Horowitz* Division of Neurology, University
Downregulation of angiotensin type 1 receptor and nuclear factor-κb. by sirtuin 1 contributes to renoprotection in unilateral ureteral
 Supplementary Information Downregulation of angiotensin type 1 receptor and nuclear factor-κb by sirtuin 1 contributes to renoprotection in unilateral ureteral obstruction Shao-Yu Yang 1,2, Shuei-Liong
Supplementary Information Downregulation of angiotensin type 1 receptor and nuclear factor-κb by sirtuin 1 contributes to renoprotection in unilateral ureteral obstruction Shao-Yu Yang 1,2, Shuei-Liong
What Does the Mechanism of Spinal Cord Stimulation Tell Us about Complex Regional Pain Syndrome?pme_
 Pain Medicine 2010; 11: 1278 1283 Wiley Periodicals, Inc. What Does the Mechanism of Spinal Cord Stimulation Tell Us about Complex Regional Pain Syndrome?pme_915 1278..1283 Joshua P. Prager, MD, MS Center
Pain Medicine 2010; 11: 1278 1283 Wiley Periodicals, Inc. What Does the Mechanism of Spinal Cord Stimulation Tell Us about Complex Regional Pain Syndrome?pme_915 1278..1283 Joshua P. Prager, MD, MS Center
Changes in Hind Paw Epidermal Thickness, Peripheral Nerve Distribution and Mechanical Sensitivity After Immobilization in Rats
 Physiol. Res. 61: 643-647, 2012 SHORT COMMUNICATION Changes in Hind Paw Epidermal Thickness, Peripheral Nerve Distribution and Mechanical Sensitivity After Immobilization in Rats J. NAKANO 1, Y. SEKINO
Physiol. Res. 61: 643-647, 2012 SHORT COMMUNICATION Changes in Hind Paw Epidermal Thickness, Peripheral Nerve Distribution and Mechanical Sensitivity After Immobilization in Rats J. NAKANO 1, Y. SEKINO
Accepted Manuscript. Alternative splicing in heart surgery: lost in translation?
 Accepted Manuscript Alternative splicing in heart surgery: lost in translation? Tyson A. Fricke, MBBS, BMedSci, Michael ZL. Zhu, MD, Igor E. Konstantinov, MD, PhD, FRACS PII: S0022-5223(18)32871-X DOI:
Accepted Manuscript Alternative splicing in heart surgery: lost in translation? Tyson A. Fricke, MBBS, BMedSci, Michael ZL. Zhu, MD, Igor E. Konstantinov, MD, PhD, FRACS PII: S0022-5223(18)32871-X DOI:
Laiche Djouhri 1, Xin Fang 2, Stella Koutsikou, Sally N. Lawson PAIN Ò 153 (2012) article info. abstract. 1.
 article info. abstract. 1.") PAIN Ò 13 (212) 1824 1836 www.elsevier.com/locate/pain Partial nerve injury induces electrophysiological changes in conducting (uninjured) nociceptive and nonnociceptive DRG neurons: Possible relationships
PAIN Ò 13 (212) 1824 1836 www.elsevier.com/locate/pain Partial nerve injury induces electrophysiological changes in conducting (uninjured) nociceptive and nonnociceptive DRG neurons: Possible relationships
Inflammation-Induced Airway Hypersensitivity: From Ion Channels to Patients
 Inflammation-Induced Airway Hypersensitivity: From Ion Channels to Patients Lu-Yuan Lee, Ph.D. Airway Sensory Neurobiology Laboratory Department of Physiology University of Kentucky Medical Center BACKGROUND
Inflammation-Induced Airway Hypersensitivity: From Ion Channels to Patients Lu-Yuan Lee, Ph.D. Airway Sensory Neurobiology Laboratory Department of Physiology University of Kentucky Medical Center BACKGROUND
PAIN MANAGEMENT in the CANINE PATIENT
 PAIN MANAGEMENT in the CANINE PATIENT Laurie Edge-Hughes, BScPT, MAnimSt (Animal Physio), CAFCI, CCRT Part 1: Laurie Edge-Hughes, BScPT, MAnimSt (Animal Physio), CAFCI, CCRT 1 Pain is the most common reason
PAIN MANAGEMENT in the CANINE PATIENT Laurie Edge-Hughes, BScPT, MAnimSt (Animal Physio), CAFCI, CCRT Part 1: Laurie Edge-Hughes, BScPT, MAnimSt (Animal Physio), CAFCI, CCRT 1 Pain is the most common reason
Lesson 33. Objectives: References: Chapter 16: Reading for Next Lesson: Chapter 16:
 Lesson 33 Lesson Outline: Nervous System Structure and Function Neuronal Tissue Supporting Cells Neurons Nerves Functional Classification of Neuronal Tissue Organization of the Nervous System Peripheral
Lesson 33 Lesson Outline: Nervous System Structure and Function Neuronal Tissue Supporting Cells Neurons Nerves Functional Classification of Neuronal Tissue Organization of the Nervous System Peripheral
Lecture 17, 28 Oct 2003 Chapter 12, Circulation (con t) Vertebrate Physiology ECOL 437 University of Arizona Fall 2003
 Vertebrate Physiology ECOL 437 University of Arizona Fall 2003") 1 Lecture 17, 28 Oct 2003 Chapter 12, Circulation (con t) Vertebrate Physiology ECOL 437 University of Arizona Fall 2003 instr: Kevin Bonine t.a.: Bret Pasch Vertebrate Physiology 437 2 1. Circulation
1 Lecture 17, 28 Oct 2003 Chapter 12, Circulation (con t) Vertebrate Physiology ECOL 437 University of Arizona Fall 2003 instr: Kevin Bonine t.a.: Bret Pasch Vertebrate Physiology 437 2 1. Circulation
Neuropsychiatry Block
 Neuropsychiatry Block Physiology of the Autonomic Nervous System By Laiche Djouhri, PhD Dept. of Physiology Email: ldjouhri@ksu.edu.sa Ext:71044 References The Autonomic Nervous System and the Adrenal
Neuropsychiatry Block Physiology of the Autonomic Nervous System By Laiche Djouhri, PhD Dept. of Physiology Email: ldjouhri@ksu.edu.sa Ext:71044 References The Autonomic Nervous System and the Adrenal
Lujain Hamdan. Ayman Musleh & Yahya Salem. Mohammed khatatbeh
 12 Lujain Hamdan Ayman Musleh & Yahya Salem Mohammed khatatbeh the last lecture, we have studied the differences between the two divisions of the ANS: sympathetic and parasympathetic pathways which work
12 Lujain Hamdan Ayman Musleh & Yahya Salem Mohammed khatatbeh the last lecture, we have studied the differences between the two divisions of the ANS: sympathetic and parasympathetic pathways which work
Pain Pathways. Dr Sameer Gupta Consultant in Anaesthesia and Pain Management, NGH
 Pain Pathways Dr Sameer Gupta Consultant in Anaesthesia and Pain Management, NGH Objective To give you a simplistic and basic concepts of pain pathways to help understand the complex issue of pain Pain
Pain Pathways Dr Sameer Gupta Consultant in Anaesthesia and Pain Management, NGH Objective To give you a simplistic and basic concepts of pain pathways to help understand the complex issue of pain Pain
ANAT2010. Concepts of Neuroanatomy (II) S2 2018
 S2 2018") ANAT2010 Concepts of Neuroanatomy (II) S2 2018 Table of Contents Lecture 13: Pain and perception... 3 Lecture 14: Sensory systems and visual pathways... 11 Lecture 15: Techniques in Neuroanatomy I in vivo
ANAT2010 Concepts of Neuroanatomy (II) S2 2018 Table of Contents Lecture 13: Pain and perception... 3 Lecture 14: Sensory systems and visual pathways... 11 Lecture 15: Techniques in Neuroanatomy I in vivo
Dissecting out mechanisms responsible for peripheral neuropathic pain: Implications for diagnosis and therapy
 Life Sciences 74 (2004) 2605 2610 www.elsevier.com/locate/lifescie Dissecting out mechanisms responsible for peripheral neuropathic pain: Implications for diagnosis and therapy Clifford J. Woolf* Neural
Life Sciences 74 (2004) 2605 2610 www.elsevier.com/locate/lifescie Dissecting out mechanisms responsible for peripheral neuropathic pain: Implications for diagnosis and therapy Clifford J. Woolf* Neural
Title proteinase-activated receptor 2 ago. Published by Elsevier Ireland Ltd.
 NAOSITE: Nagasaki University's Ac Title Author(s) Olopatadine hydrochloride inhibits proteinase-activated receptor 2 ago Yoshizaki, Ayumi; Sato, Shinichi Citation Journal of dermatological science, Issue
NAOSITE: Nagasaki University's Ac Title Author(s) Olopatadine hydrochloride inhibits proteinase-activated receptor 2 ago Yoshizaki, Ayumi; Sato, Shinichi Citation Journal of dermatological science, Issue
Peripheral Nervous System
 Peripheral Nervous System 1 Sensory Receptors Sensory Receptors and Sensation Respond to changes (stimuli) in the environment Generate graded potentials that can trigger an action potential that is carried
Peripheral Nervous System 1 Sensory Receptors Sensory Receptors and Sensation Respond to changes (stimuli) in the environment Generate graded potentials that can trigger an action potential that is carried
NIH Public Access Author Manuscript Ann Neurol. Author manuscript; available in PMC 2009 October 19.
 NIH Public Access Author Manuscript Published in final edited form as: Ann Neurol. 2009 March ; 65(3): 348 351. doi:10.1002/ana.21601. Sympathetic Block with Botulinum Toxin to Treat Complex Regional Pain
NIH Public Access Author Manuscript Published in final edited form as: Ann Neurol. 2009 March ; 65(3): 348 351. doi:10.1002/ana.21601. Sympathetic Block with Botulinum Toxin to Treat Complex Regional Pain
Supplementary Table 1. List of primers used in this study
 Supplementary Table 1. List of primers used in this study Gene Forward primer Reverse primer Rat Met 5 -aggtcgcttcatgcaggt-3 5 -tccggagacacaggatgg-3 Rat Runx1 5 -cctccttgaaccactccact-3 5 -ctggatctgcctggcatc-3
Supplementary Table 1. List of primers used in this study Gene Forward primer Reverse primer Rat Met 5 -aggtcgcttcatgcaggt-3 5 -tccggagacacaggatgg-3 Rat Runx1 5 -cctccttgaaccactccact-3 5 -ctggatctgcctggcatc-3
Somatic Sensory System I. Background
 Somatic Sensory System I. Background A. Differences between somatic senses and other senses 1. Receptors are distributed throughout the body as opposed to being concentrated at small, specialized locations
Somatic Sensory System I. Background A. Differences between somatic senses and other senses 1. Receptors are distributed throughout the body as opposed to being concentrated at small, specialized locations
Chapter 7. Objectives
 Chapter 7 The Nervous System: Structure and Control of Movement Objectives Discuss the general organization of the nervous system Describe the structure & function of a nerve Draw and label the pathways
Chapter 7 The Nervous System: Structure and Control of Movement Objectives Discuss the general organization of the nervous system Describe the structure & function of a nerve Draw and label the pathways
The Conundrum Of Sensitization When Recording From Nociceptors
 University of New England DUNE: DigitalUNE Biomedical Sciences Faculty Publications Biomedical Sciences Faculty Works 5-15-2010 The Conundrum Of Sensitization When Recording From Nociceptors Geoffrey M.
University of New England DUNE: DigitalUNE Biomedical Sciences Faculty Publications Biomedical Sciences Faculty Works 5-15-2010 The Conundrum Of Sensitization When Recording From Nociceptors Geoffrey M.
Biomechanics of Pain: Dynamics of the Neuromatrix
 Biomechanics of Pain: Dynamics of the Neuromatrix Partap S. Khalsa, D.C., Ph.D. Department of Biomedical Engineering The Neuromatrix From: Melzack R (1999) Pain Suppl 6:S121-6. NIOSH STAR Symposium May
Biomechanics of Pain: Dynamics of the Neuromatrix Partap S. Khalsa, D.C., Ph.D. Department of Biomedical Engineering The Neuromatrix From: Melzack R (1999) Pain Suppl 6:S121-6. NIOSH STAR Symposium May
The Autonomic Nervous System Outline of class lecture for Physiology
 The Autonomic Nervous System Outline of class lecture for Physiology 1 After studying the endocrine system you should be able to: 1. Describe the organization of the nervous system. 2. Compare and contrast
The Autonomic Nervous System Outline of class lecture for Physiology 1 After studying the endocrine system you should be able to: 1. Describe the organization of the nervous system. 2. Compare and contrast
Role of Peptidergic Nerve Terminals in the Skin: Reversal of Thermal Sensation by Calcitonin Gene-Related Peptide in TRPV1-Depleted Neuropathy
 Role of Peptidergic Nerve Terminals in the Skin: Reversal of Thermal Sensation by Calcitonin Gene-Related Peptide in TRPV1-Depleted Neuropathy Yu-Lin Hsieh 1,2., Chih-Lung Lin 3,4., Hao Chiang 2, Yaw-Syan
Role of Peptidergic Nerve Terminals in the Skin: Reversal of Thermal Sensation by Calcitonin Gene-Related Peptide in TRPV1-Depleted Neuropathy Yu-Lin Hsieh 1,2., Chih-Lung Lin 3,4., Hao Chiang 2, Yaw-Syan
2401 : Anatomy/Physiology
 Dr. Chris Doumen Week 11 2401 : Anatomy/Physiology Autonomic Nervous System TextBook Readings Pages 533 through 552 Make use of the figures in your textbook ; a picture is worth a thousand words! Work
Dr. Chris Doumen Week 11 2401 : Anatomy/Physiology Autonomic Nervous System TextBook Readings Pages 533 through 552 Make use of the figures in your textbook ; a picture is worth a thousand words! Work
Integrated Cardiopulmonary Pharmacology Third Edition
 Integrated Cardiopulmonary Pharmacology Third Edition Chapter 3 Pharmacology of the Autonomic Nervous System Multimedia Directory Slide 19 Slide 37 Slide 38 Slide 39 Slide 40 Slide 41 Slide 42 Slide 43
Integrated Cardiopulmonary Pharmacology Third Edition Chapter 3 Pharmacology of the Autonomic Nervous System Multimedia Directory Slide 19 Slide 37 Slide 38 Slide 39 Slide 40 Slide 41 Slide 42 Slide 43
Brian Kahan, D.O. FAAPMR, DABPM, DAOCRM, FIPP Center for Pain Medicine and Physiatric Rehabilitation 2002 Medical Parkway Suite 150 Annapolis, MD
 Brian Kahan, D.O. FAAPMR, DABPM, DAOCRM, FIPP Center for Pain Medicine and Physiatric Rehabilitation 2002 Medical Parkway Suite 150 Annapolis, MD 1630 Main Street Suite 215 Chester, MD 410-571-9000 www.4-no-pain.com
Brian Kahan, D.O. FAAPMR, DABPM, DAOCRM, FIPP Center for Pain Medicine and Physiatric Rehabilitation 2002 Medical Parkway Suite 150 Annapolis, MD 1630 Main Street Suite 215 Chester, MD 410-571-9000 www.4-no-pain.com
Antigen-induced arthritis in rats is associated with increased growth-associated protein 43 positive intraepidermal nerve fibres remote from the joint
 Jochmann et al. Arthritis Research & Therapy (215) 17:299 DOI 1.1186/s1375-15-818-8 RESEARCH ARTICLE Open Access Antigen-induced arthritis in rats is associated with increased growth-associated protein
Jochmann et al. Arthritis Research & Therapy (215) 17:299 DOI 1.1186/s1375-15-818-8 RESEARCH ARTICLE Open Access Antigen-induced arthritis in rats is associated with increased growth-associated protein
NORLAND AVENUE PHARMACY PRESCRIPTION COMPOUNDING FOR PAIN MANAGEMENT
 NOVEMBER 2011 NORLAND AVENUE PHARMACY PRESCRIPTION COMPOUNDING N ORLANDA VENUEP HARMACY. COM We customize individual prescriptions for the specific needs of our patients. INSIDE THIS ISSUE: Sciatic Pain
NOVEMBER 2011 NORLAND AVENUE PHARMACY PRESCRIPTION COMPOUNDING N ORLANDA VENUEP HARMACY. COM We customize individual prescriptions for the specific needs of our patients. INSIDE THIS ISSUE: Sciatic Pain