2 Tissue Amino Acid Metabolism. Lecture #17-19 Lecturer: Alexander Koval
|
|
|
- Annabelle Caldwell
- 5 years ago
- Views:
Transcription


1 2 Tissue Amino Acid Metabolism Lecture #17-19 Lecturer: Alexander Koval
2 Introduction to Amino Acid Metabolism Amino Acid Catabolism Non Essential Amino Acid Biosynthesis Plants and microorganisms can synthesize all 20 amino acids required for protein synthesis whereas vertebrates can only synthesize about half this number Koval, 2007 (C) 2
3 Inborn Errors of Metabolism Can Disrupt Amino Acid Degradation Errors in amino acid metabolism served as sources of some of the first insights into the correlation between pathology and biochemistry. Although there are many hereditary errors of amino acid metabolism, phenylketonuria is the best known. This condition is the result of the accumulation of high levels of phenylalanine in the body fluids. By unknown mechanisms, this accumulation results in mental retardation unless the afflicted are placed on low phenylalanine diets immediately after birth Koval, 2007 (C) 3
4 Glutamine/Glutamate and Asparagine/Aspartate Catabolism Glutaminase is an important kidney tubule enzyme involved in converting glutamine (from liver and from other tissue) to glutamate and NH 3+, with the NH 3+ being excreted in the urine. Glutaminase activity is present in many other tissues as well, although its activity is not nearly as prominent as in the kidney. The glutamate produced from glutamine is converted to a-ketoglutarate, making glutamine a glucogenic amino acid. Asparaginase is also widely distributed within the body, where it converts asparagine into ammonia and aspartate. Aspartate transaminates to oxaloacetate, which follows the gluconeogenic pathway to glucose. Glutamate and aspartate are important in collecting and eliminating amino nitrogen via glutamine synthetase and the urea cycle, respectively. The catabolic path of the carbon skeletons involves simple 1-step aminotransferase reactions that directly produce net quantities of a TCA cycle intermediate. The glutamate dehydrogenase reaction operating in the direction of a-ketoglutarate production provides a second avenue leading from glutamate to gluconeogenesis Koval, 2007 (C) 4
5 Alanine Catabolism Alanine is also important in intertissue nitrogen transport as part of the glucose-alanine cycle. Alanine's catabolic pathway involves a simple aminotransferase reaction that directly produces pyruvate. Generally pyruvate produced by this pathway will result in the formation of oxaloacetate, although when the energy charge of a cell is low the pyruvate will be oxidized to C 2 and H 2 via the PDH complex and the TCA cycle. This makes alanine a glucogenic amino acid Koval, 2007 (C) 5


6 Ala Conversion to Urea Koval, 2007 (C) 6
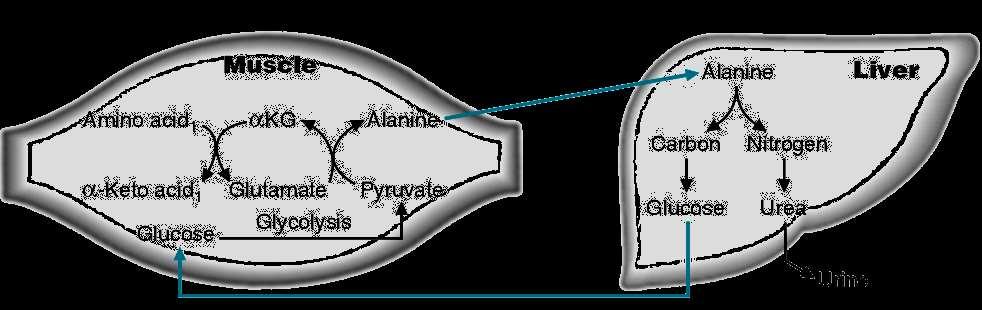
7 Glucose-Alanine Cycle Koval, 2007 (C) 7
8 Arginine, rnithine and Proline Catabolism The catabolism of arginine begins within the context of the urea cycle. Proline catabolism is a reversal of its synthesis process. Thus arginine, ornithine and proline, are glucogenic Koval, 2007 (C) 8


9 Arginine Metabolism ARG is hydrolyzed to urea and ornithine by arginase. rnithine, in excess of urea cycle needs, is transaminated to form glutamate semialdehyde. Glutamate semialdehyde can serve as the precursor for proline biosynthesis as described above or it can be converted to glutamate Koval, 2007 (C) 9

10 Prolin Metabolism The glutamate semialdehyde generated from ornithine and proline catabolism is oxidized to glutamate by an ATPindependent glutamate semialdehyde dehydrogenase. The glutamate can then be converted to a-ketoglutarate in a transamination reaction Koval, 2007 (C) 10
11 Serine Catabolism Serine can be converted to glycine and then glycine can be oxidized to C 2 and NH 3, with the production of two equivalents of N 5,N 10 - methylene-thf Koval, 2007 (C) 11

12 Serine Catabolism: 3-Phosphoglycerate and Pyruvate Formation Marks, 2000 Serine can be catabolized back to the glycolytic intermediate, 3- phosphoglycerate, by a pathway that is essentially a reversal of serine biosynthesis. However, the enzymes are different. Serine can also be converted to pyruvate through a deamination reaction catalyzed by serine/threonine dehydratase Koval, 2007 (C) 12

13 Threonine Catabolism There are at least 3 pathways for threonine catabolism. ne involves a pathway initiated by threonine dehydrogenase yielding a-amino-bketobutyrate. The a-amino-b-ketobutyrate is either converted to acetyl-coa and glycine or spontaneously degrades to aminoacetone which is converted to pyruvate. The second pathway involves serine/threonine dehydratase yielding a- ketobutyrate which is further catabolized to propionyl-coa and finally the TCA cycle intermediate, succinyl-coa. The third pathway utilizes threonine aldolase. The products of this reaction are both ketogenic (acetyl-coa) and glucogenic (pyruvate) Koval, 2007 (C) 13
14 Glycine Catabolism Glycine can be converted to serine by serine hydroxymethyltransferase, serine can be converted back to 3- phosphoglycerate or to pyruvate by serine/threonine dehydratase. the main glycine catabolic pathway leads to the production of C 2, NH 4, and one equivalent of N 5,N 10 -methylene-thf by the mitochondrial glycine cleavage complex Koval, 2007 (C) 14
15 Glycine xidation in Mitochondria Glycine produced from serine or from the diet can also be oxidized by glycine cleavage complex, GCC, to yield a second equivalent of N 5,N 10 -methylenetetrahydrofolate as well as ammonia and C 2. Glycine is involved in many anabolic reactions other than protein synthesis including the synthesis of purine nucleotides, heme, glutathione, creatine and serine. H 2 N Glycine H H 4 folate C 2 + NH NADH +H + N 5, N 10 -CH 2 -H 4 folate Koval, 2007 (C) 15

16 Glycine, Serine & Threonine Metabolism Marks, Koval, 2007 (C) 16

17 Structure of Glycine Cleavage Complex Koval, 2007 (C) 17
18 Cysteine Catabolism There are several pathways for cysteine catabolism. The simplest, but least important pathway is catalyzed by a liver desulfurase and produces hydrogen sulfide, (H 2 S) and pyruvate. The more important catabolic pathway is via a cytochrome-p 450 -coupled enzyme, cysteine dioxygenase that oxidizes the cysteine sulfhydryl to sulfinate, producing the intermediate cysteinesulfinate Koval, 2007 (C) 18

 and H 2 2.")
19 Cysteine Metabolism-1 Cysteinesulfinate can serve as a biosynthetic intermediate undergoing decarboxylation and oxidation to produce taurine. Catabolism of cysteinesulfinate proceeds through transamination to b-sulfinylpyruvate which is in undergoes desulfuration yielding bisulfite, (HS 3- ) and the glucogenic product, pyruvate. The enzyme sulfite oxidase uses 2 and H 2 to convert HS 3 - to sulfate, (S 4- ) and H 2 2. The resultant sulfate is used as a precursor for the formation of 3'-phosphoadenosine- 5'-phosphosulfate (PAPS) Koval, 2007 (C) 19
 20")
20 Formation of 3'-phosphoadenosine-5'- phosphosulfate (PAPS) PAPS is used for the transfer of sulfate to biological molecules such as the sugars of the glycosphingolipids Koval, 2007 (C) 20
21 ther Cysteine Catabolism Pathways ther than protein, the most important product of cysteine metabolism is the bile salt precursor taurine, which is used to form the bile acid conjugates taurocholate and taurochenodeoxycholate Koval, 2007 (C) 21
, to produce thiosulfate, (S 2 3 2- ) and pyruvate.")
22 Cysteine Metabolism-2 The enzyme cystathionase can also transfer the sulfur from one cysteine to another generating thiocysteine and pyruvate. Transamination of cysteine yields b-mercaptopyruvate which then reacts with sulfite, (S 3 2- ), to produce thiosulfate, (S ) and pyruvate. Both thiocysteine and thiosulfate can be used by the enzyme rhodanese to incorporate sulfur into cyanide, (CN - ), thereby detoxifying the cyanide to thiocyanate Koval, 2007 (C) 22
23 Methionine Catabolism The principal fates of the essential amino acid methionine are incorporation into polypeptide chains, and use in the production of a-ketobutyrate and cysteine via SAM as described above. The transulfuration reactions that produce cysteine from homocysteine and serine also produce a-ketobutyrate, the latter being converted to succinyl-coa. Regulation of the methionine metabolic pathway is based on the availability of methionine and cysteine. If both amino acids are present in adequate quantities, SAM accumulates and is a positive effector on cystathionine synthase, encouraging the production of cysteine and a-ketobutyrate (both of which are glucogenic). However, if methionine is scarce, SAM will form only in small quantities, thus limiting cystathionine synthase activity. Under these conditions accumulated homocysteine is remethylated to methionine, using N 5 -methyl-thf and other compounds as methyl donors Koval, 2007 (C) 23
 methyltransferase reactions and is conveted to S- adenosylhomocysteine (SAH). (3) adenosylhomocysteinase converts SAH to homocysteine.")
24 Methionine Cycle Methionine is converted to S-adenosylmethionine (SAM) by (1) methionine adenosyltransferase. SAM serves as a methyldonor in many (2) methyltransferase reactions and is conveted to S- adenosylhomocysteine (SAH). (3) adenosylhomocysteinase converts SAH to homocysteine. methionine can be regenerated from homocysteine via (4) methionine synthase Koval, 2007 (C) 24
25 Valine, Leucine and Isoleucine Catabolism This group of essential amino acids are identified as the branched-chain amino acids, BCAAs. Because this arrangement of carbon atoms cannot be made by humans, these amino acids are an essential element in the diet. The catabolism of all three compounds initiates in muscle and yields NADH and FADH 2 which can be utilized for ATP generation. The catabolism of all three of these amino acids uses the same enzymes in the first two steps Koval, 2007 (C) 25


26 BCAA Catabolism-1 The first step in each case is a transamination using a single BCAA aminotransferase, with a-ketoglutarate as amine acceptor. As a result, three different a- keto acids are produced and are oxidized using a common branchedchain a-keto acid dehydrogenase, yielding the three different CoA derivatives. Subsequently the metabolic pathways diverge, producing many intermediates Koval, 2007 (C) 26
27 BCAA Catabolism (2) The principal product from valine is propionylcoa, the glucogenic precursor of succinyl-coa. Isoleucine catabolism terminates with production of acetylcoa and propionylcoa; thus isoleucine is both glucogenic and ketogenic. Leucine gives rise to acetylcoa and acetoacetylcoa, and is thus classified as strictly ketogenic Koval, 2007 (C) 27
28 Maple Syrup Urine Disease, MSUD There are a number of genetic diseases associated with faulty catabolism of the BCAAs. The most common defect is in the branched-chain a- keto acid dehydrogenase. all three a-keto acids accumulate and are excreted in the urine. Maple syrup urine disease because of the characteristic odor of the urine in afflicted individuals. Mental retardation in these cases is extensive. The life of afflicted individuals is short and development is abnormal. The main neurological problems are due to poor formation of myelin in the CNS Koval, 2007 (C) 28
29 Phenylalanine and Tyrosine Catabolism 1. Incorporated into polypeptide chains 2. Production of tyrosine from phenylalanine. Phe and Tyr degradation conversion to fumarate and acetoacetate: both glucogenic and ketogenic AA. 3. Biosynthesis of physiologically important metabolites e.g. Dopamine Norepinephrine Epinephrine Koval, 2007 (C) 29
30 Aromatic Amino Acid Catabolism type II tyrosinemia neonatal tyrosinemia alkaptonuria type I tyrosinemia, or tyrosinosis Blocks in pathology 1. type II tyrosinemia; 2. neonatal tyrosinemia; 3. alkaptonuria; 4. type I tyrosinemia, or tyrosinosis Murray et al., Koval, 2007 (C) 30
31 Defective Aromatic AA Catabolism Deficiency of tyrosine aminotransferase (TAT) leads to: Hypertyrosinemia Urinary excretion of tyrosine and the catabolic intermediates The neurological symptoms are similar for both deficiencies. Hypertyrosinemia leads to painful corneal eruptions and photophobia. Alkaptonuria: defective homogentisic acid oxidase. Homogentisic acid accumulation is relatively innocuous, causing urine to darken on exposure to air, but no lifethreatening effects accompany the disease Koval, 2007 (C) 31
32 Lysine Catabolism H 2 - a-ketoglutarate - - N - H 2 N H 2 N H 2 N NADH + H + H Lysine Lysine catabolism is unusual in the way NAD + that the e-amino group is transferred to a- ketoglutarate and into the general nitrogen pool. The reaction is a transamination in which the e-amino group is transferred to the a- - keto carbon of a-ketoglutarate forming the metabolite, saccharopine. This transamination reactions does not employ pyridoxal phosphate as a cofactor Koval, 2007 (C) 32 HN - H 2 N H saccharopine dehydrogenase (NAD+, L-lysine-forming) EC H L-Saccharopine N 6 -(L-1,3-dicarboxypropyl)-L-lysine
33 Lysine Catabolism (cont d) NADH + H + NAD N H 2 N H 2 H - saccharopine HN H 2 N dehydrogenase (NAD+, L-glutamateforming) EC H L-Saccharopine N 6 -(L-1,3-dicarboxypropyl)-L-lysine - H Glutamate NH 2 - H 2 N - L-2-Aminoadipate semialdehyde Saccharopine is immediately hydrolyzed by the enzyme a- aminoadipic semialdehyde synthase; and the amino nitrogen remains with the a-carbon of a- ketoglutarate, producing glutamate and a-aminoadipic semialdehyde. Because this transamination reaction is not reversible, lysine is an essential amino acid Koval, 2007 (C) 33
34 Lysine Catabolism (final of pathway) - H 2 N - L-2-Aminoadipate semialdehyde Glutaryl-CoA S NAD + NADH + H + H 2 L-aminoadipatesemialdehyde dehydrogenase EC CoA C 2 [] - - NH 2 L-2-Aminoadipate 2-aminoadipate transaminase EC HSCoA 2-Ketoadipate - a-kg Glu - The ultimate endproduct of lysine catabolism is acetoacetyl-coa. This one catabolizes by b-oxidation pathway to 2 acetyl-coa C 2 and H 2 are produced in TCA. Acetoacetyl-CoA 2 Acetyl-CoA C 2 + H Koval, 2007 (C) 34
35 Pathology of Lysine Catabolism Genetic deficiencies in the enzyme a-aminoadipic semialdehyde synthase have been observed in individuals who excrete large quantities of urinary lysine and some saccharopine. The lysinemia and associated lysinuria are benign. ther serious disorders associated with lysine metabolism are due to failure of the transport system for lysine and the other dibasic amino acids across the intestinal wall. Lysine is essential for protein synthesis; a deficiencies of its transport into the body can cause seriously diminished levels of protein synthesis. Probably more significant however, is the fact that arginine is transported on the same dibasic amino acid carrier, and resulting arginine deficiencies limit the quantity of ornithine available for the urea cycle. The result is severe hyperammonemia after a meal rich in protein. The addition of citrulline to the diet prevents the hyperammonemia. Koval, 2007 (C) 35
36 Lysine is Important for Carnitine Synthesis Lysine is also important as a precursor for the synthesis of carnitine, required for the transport of fatty acids into the mitochondria for oxidation. Free lysine does not serve as the precursor for this reaction, rather the modified lysine found in certain proteins. Some proteins modify lysine to trimethyllysine using SAM as the methyl donor to transfer methyl groups to the e-amino of the lysine side chain. Hydrolysis of proteins containing trimethyllysine provide the substrate for the subsequent conversion to carnitine Koval, 2007 (C) 36
37 Histidine Catabolism - N HN NH 2 L-Histidine a-ketoglutarate - NH 4 + histidase EC H HN N Urocanate L-Glutamate Koval, 2007 (C) H 2 urocanase EC N 5 -Formimino H NH 4 folate 2 glutamate formiminotransferase EC N N - 4-Imidazolone-5-propionate H 4 folate HN H 2 - N H imidazolonepropionase EC H N-Formiminoglutamate Histidine catabolism begins with release of the a-amino group catalyzed by histidase. The deaminated product, urocanate, is not the usual a-keto acid. The end product of histidine catabolism is glutamate (histidine is glucogenic amino acids).
38 Histidine is a Source of N 5 -formiminothf H 2 N HN N HN N NH HN HN H N 5 -formimidoyl tetrahydrofolate Another key feature of histidine catabolism is that it serves as a source of ring nitrogen to combine with tetrahydrofolate (THF), producing the 1-carbon THF intermediate known as N 5 - formiminothf (or N 5 -formimidoyl tetrahydrofolate). The latter reaction is one of two routes to N 5 -formiminothf Koval, 2007 (C) 38 -
39 Decarboxylation of Histidine N HN C 2 N HN NH 2 Histidine Histamine H Histidine decarboxylase NH 2 Decarboxylation of histidine in the intestine by bacteria gives rise to histamine. Similarly, histamine arises in many tissues by the decarboxylation of histidine, which in excess causes constriction or dilation of various blood vessels. The general symptoms are those of asthma and various allergic reactions Koval, 2007 (C) 39
40 Histidine Catabolism Pathology The principal genetic deficiency associated with histidine metabolism is absence or deficiency of the first enzyme of the pathway, histidase. The resultant histidinemia is relatively benign. The disease, which is of relatively high incidence (1 in 10,000), is most easily detected by the absence of urocanate from skin and sweat, where it is normally found in relative abundance Koval, 2007 (C) 40
41 Tryptophan N H Tryptophan NH 2 H Serotonin Melatonin Niacin The main function of tryptophan is as a building block in protein synthesis. Tryptophan is a precursor for serotonin (a neurotransmitter), melatonin (a neurohormone), and niacin. The functional group of trytophan is indole. Tryptophan has been implicated as a possible cause of schizophrenia in people who cannot metabolize it properly. When improperly metabolized it creates a waste product in the brain which is toxic and causes hallucinations and delusions. Tryptophan has also been indicated as an aid for schizophrenic patients Koval, 2007 (C) 41
42 Tryptophan Catabolism: Kynurenic Acid Formation (1) tryptophan 2,3-dioxygenase EC H N-Formylkynurenine arylformamidase EC N H NH NH 2 2 Fe 2 + H NH 2 Tryptophan H 2 NH 2 NH 2 H H Formate Kynurenine A number of important side reactions occur during the catabolism of tryptophan on the pathway to acetoacetate. The first enzyme of the catabolic pathway is an iron tryptophan 2,3- dioxygenase that opens the indole ring. The latter enzyme is highly inducible, its concentration rising almost 10-fold on a diet high in tryptophan. Kynurenine is the first key branch point intermediate in the pathway Koval, 2007 (C) 42
43 Kynurenic Acid Formation (2) NH 2 NH 2 Kynurenine undergoes deamniation in a standard transamination reaction yielding kynurenic acid. Kynurenic acid and metabolites have been shown to act as antiexcitotoxics and anticonvulsives. H Kynurenine a-kg Glu kynurenineoxoglutarate transaminase EC H 2 NH Koval, 2007 (C) 43 N H spontaneously Kynurenic acid H CH
44 Tryptophan Catabolism: Kynurenine Degradation H H NH 2 - NH 2 NH 2 H NH 2 NH 2 Kynurenine kynurenine 2 + NADPH 3-monooxygenase H 2 + NADP + EC CH kynureninase EC H2 3-Hydroxykynurenine CH H 3 C CH NH 2 Alanine 3-Hydroxyanthranilate Kynurenic acid A second side branch reaction produces anthranilic acid plus alanine. Another equivalent of alanine is produced further along the main catabolic pathway, and it is the production of these alanine residues that allows tryptophan to be classified among the glucogenic and ketogenic amino acids Koval, 2007 (C) 44
45 Further Tryptophan Catabolism The second important branch point converts kynurenine into 2-amino-3-carboxymuconic semialdehyde, which has two fates. The main flow of carbon to glutarate. Important side reaction in liver transamination, nicotinic acid small amount of NAD + and NADP + Also Trp roles as AA in protein biosynthesis, precursor of serotonin and melatonin Koval, 2007 (C) 45
46 Non Essential Amino Acid Biosynthesis Glutamate and Aspartate Alanine and the Glucose-Alanine Cycle Cysteine Tyrosine rnithine and Proline Serine Glycine Aspartate/Asparagine and Glutamate/Glutamine Koval, 2007 (C) 46
47 Glutamate and Aspartate - a Ketoglutarate - - NH 2 L-Glutamate - NH 4 + H 2 NAD(P)H + H + NAD(P) + The glutamate dehydrogenase Glutamate and aspartate are synthesized from their widely distributed a-keto acid precursors by simple 1-step transamination reactions. The former catalyzed by glutamate dehydrogenase and the latter by aspartate aminotransferase, Koval, 2007 (C) AST. 47
48 Transaminase reactions - NH 2 H Pyruvate Alanine Glu or Asp a-ketoglutarate or oxaloacetate Transamination is the most widespread form of nitrogen transfer Koval, 2007 (C) 48
49 Asparaginase H 2 N NH 2 H 2 NH 3 NH 2 H H L-Asparagine L-Aspartate - Aspartate is also derived from asparagine through the action of asparaginase Koval, 2007 (C) 49
50 Alanine and the Glucose-Alanine Cycle Aside from its role in protein synthesis, alanine is second only to glutamine in prominence as a circulating amino acid. In this capacity it serves a unique role in the transfer of nitrogen from peripheral tissue to the liver. Alanine is transferred to the circulation by many tissues, but mainly by muscle, in which alanine is formed from pyruvate at a rate proportional to intracellular pyruvate levels Koval, 2007 (C) 50
51 Glucose-Alanine Cycle Liver accumulates plasma alanine, reverses the transamination that occurs in muscle, and proportionately increases urea production. The pyruvate is either oxidized or converted to glucose via gluconeogenesis. When alanine transfer from muscle to liver is coupled with glucose transport from liver back to muscle, the process is known as the glucose-alanine cycle. The key feature of the cycle is that in 1 molecule, alanine, peripheral tissue exports pyruvate and ammonia (which are potentially rate-limiting for metabolism) to the liver, where the carbon skeleton is recycled and most nitrogen eliminated Koval, 2007 (C) 51
52 2 Main Pathways of Muscle Alanine Production There are 2 main pathways to production of muscle alanine: directly from protein degradation, and via the transamination of pyruvate by glutamate-pyruvate aminotransferase (also called alanine transaminase, ALT). glutamate + pyruvate a-kg + alanine Koval, 2007 (C) 52
53 Cysteine metabolism H H 2 N SH cysteine Synthesis Cysteine derives its carbon and nitrogen from serine. The essential amino acid methionine supplies the sulfur. Degradation Forms pyruvate. Its sulfur is converted to H 2 S Koval, 2007 (C) 53
. 10.")
54 Cysteine Biosynthesis The sulfur for cysteine synthesis comes from the essential amino acid methionine. A condensation of ATP and methionine catalyzed by methionine adenosyltransferase yields S-adenosylmethionine (SAM or AdoMet) Koval, 2007 (C) 54
55 Transmethylation Reactions SAM serves as a precurosor for numerous methyl transfer reactions (e.g. the conversion of norepinephrine to epinenephrine). The result of methyl transfer is the conversion of SAM to S-adenosylhomocysteine. S-adenosylhomocysteine is then cleaved by adenosylhomocyteinase to yield homocysteine and adenosine. Homocysteine can be converted back to methionine by methionine synthase, a reaction that occurs under methionine-sparing conditions and requires N5-methyl-tetrahydrofolate as methyl donor Koval, 2007 (C) 55


56 Utilization of Methionine in the Synthesis f Cysteine Koval, 2007 (C) 56
57 Transmethylation Reactions (cont d) Transmethylation reactions employing SAM are extremely important, but in this case the role of S-adenosylmethionine in transmethylation is secondary to the production of homocysteine (essentially a byproduct of transmethylase activity). In the production of SAM all phosphates of an ATP are lost: one as Pi and two as PPi. It is adenosine which is transferred to methionine and not AMP Koval, 2007 (C) 57
58 Cysteine Formation In cysteine synthesis, homocysteine condenses with serine to produce cystathionine, which is subsequently cleaved by cystathionase to produce cysteine and a- ketobutyrate. The sum of the latter two reactions is known as trans-sulfuration. Cysteine is used for protein synthesis and other body needs, while the a-ketobutyrate is decarboxylated and converted to propionyl- CoA Koval, 2007 (C) 58

59 Glutathione Cysteine readily oxidizes in air to form the disulfide cystine. cells contain little free cystine because glutathione reverses the formation of cystine by a non-enzymatic reduction reaction Koval, 2007 (C) 59
60 Homocystinuria The 2 key enzymes of this pathway, cystathionine synthase and cystathionase (cystathionine lyase), both use pyridoxal phosphate as a cofactor, and both are under regulatory control. Cystathionase is under negative allosteric control by cysteine, as well, cysteine inhibits the expression of the cystathionine synthase gene Koval, 2007 (C) 60
61 Homocystinyria (cont d) Genetic defects are known for both the synthase and the lyase. Missing or impaired cystathionine synthase leads to homocystinuria and is often associated with mental retardation, although the complete syndrome is multifaceted and many individuals with this disease are mentally normal. Some instances of genetic homocystinuria respond favorably to pyridoxine therapy, suggesting that in these cases the defect in cystathionine synthase is a decreased affinity for the cofactor. Missing or impaired cystathionase leads to excretion of cystathionine in the urine but does not have any other untoward effects. Rare cases are known in which cystathionase is defective and operates at a low level. This genetic disease leads to methioninuria with no other consequences Koval, 2007 (C) 61
62 Tyrosine Biosynthesis H NH 2 tyrosine H Tyrosine is produced in cells by hydroxylating the essential amino acid phenylalanine. This relationship is much like that between cysteine and methionine. Half of the phenylalanine required goes into the production of tyrosine; if the diet is rich in tyrosine itself, the requirements for phenylalanine are reduced by about 50% Koval, 2007 (C) 62


63 Biosynthesis of tyrosine from phenylalanine Koval, 2007 (C) 63
64 Phenylalanine Hydroxylase Functions H 2 N HN H N H N N H Tetrahydrobiopterin H Phenylalanine hydroxylase is a mixed-function oxygenase: one atom of oxygen is incorporated into water and the other into the hydroxyl of tyrosine. The reductant is the tetrahydrofolate-related cofactor tetrahydrobiopterin, which is maintained in the reduced state by the NADHdependent enzyme dihydropteridine reductase (DHPR) Koval, 2007 (C) 64
65 Disorders of phenylalanine metabolism Missing or deficient phenylalanine hydroxylase results in hyperphenylalaninemia. Hyperphenylalaninemia is defined as a plasma phenylalanine concentration greater than 2 mg/dl (120 mm). The most widely recognized hyperphenylalaninemia (and most severe) is the genetic disease known as phenlyketonuria (PKU). Patients suffering from PKU have plasma phenylalanine levels >1000mM, whereas the non-pku hyperphenylalaninemias exhibit levels of plasma phenylalanine <1000mM Koval, 2007 (C) 65
66 Untreated PKU Untreated PKU leads to severe mental retardation. The mental retardation is caused by the accumulation of phenylalanine, which becomes a major donor of amino groups in aminotransferase activity and depletes neural tissue of a-ketoglutarate. This absence of a-ketoglutarate in the brain shuts down the TCA cycle and the associated production of aerobic energy, which is essential to normal brain development Koval, 2007 (C) 66
67 PKU Diagnostics The product of phenylalanine transamination, phenylpyruvic acid, is reduced to phenylacetate and phenyllactate, and all 3 compounds appear in the urine. The presence of phenylacetate in the urine imparts a "mousy" odor. If the problem is diagnosed early, the addition of tyrosine and restriction of phenylalanine from the diet can minimize the extent of mental retardation Koval, 2007 (C) 67
68 Tetrahydrobiopterin and the Effects of Missing or Defective DHPR Because of the requirement for tetrahydrobiopterin in the function of phenylalanine hydroxylase, deficiencies in DHPR can manifest with hyperphenylalaninemia. Tetrahydrobiopterin is a cofactor in several other reactions (e.g. in the synthesis of the neurotransmitters as well as nitric oxide), the effects of missing or defective DHPR cause even more severe neurological difficulties than those usually associated with PKU caused by deficient phenylalanine hydroxylase activity Koval, 2007 (C) 68
69 rnithine and Proline Biosynthesis Glutamate is the precursor of both proline and ornithine, with glutamate semialdehyde being a branch point intermediate leading to one or the other of these 2 products. While ornithine is not one of the 20 amino acids used in protein synthesis, it plays a significant role as the acceptor of carbamoyl phosphate in the urea cycle Koval, 2007 (C) 69
70 rnithine and Proline Biosynthesis (cont d) NH 3 glutamate - NADH NH 3 - L-glutamate- -semialdehyde Glu a-kg H 3 N NH 3 rnithine - NH 2 - NADH H 2 NH - L-Proline 2 -Pyrrolidine- 5-carboxylate Koval, 2007 (C) 70
71 H 2 N ornithine NH 2 H H 2 N SAM NH 2 putrescine SAM 5'-methylthioadenosine H 2 N N H NH 2 5'-methylthioadenosine C 2 DC spermidine H 2 N N H H N NH2 spermine rnithine serves an additional important role as the precursor for the synthesis of the polyamines. The production of ornithine from glutamate is important when dietary arginine, the other principal source of ornithine, is limited Koval, 2007 (C) 71
72 The Fate of Glutamate Semialdehyde The fate of glutamate semialdehyde depends on prevailing cellular conditions. rnithine production occurs from the semialdehyde via a simple glutamate-dependent transamination, producing ornithine. At high arginine concentrations the ornithine contributed from the urea cycle plus that from glutamate semialdehyde inhibit the aminotransferase reaction. Semialdehyde is accumulated. The semialdehyde cyclizes spontaneously to 2 - pyrrolidine-5-carboxylate which is then reduced to proline by an NADPH-dependent reductase Koval, 2007 (C) 72
73 Serine Biosynthesis H P H D-3-Phosphoglycerate NH 2 H H L-serine NADH P i H 2 P H NH 2 Phosphohydroxypyruvate a-aa P H a-ka Phospho-L-serine The main pathway to serine starts with the glycolytic intermediate 3- phosphoglycerate. An NADH-linked dehydrogenase converts 3- phosphoglycerate into a keto acid, 3-phosphopyruvate, suitable for subsequent transamination. Aminotransferase activity with glutamate as a donor produces 3-phosphoserine, which is converted to serine by phosphoserine phosphatase Koval, 2007 (C) 73
74 Glycine Biosynthesis NH 2 H H L-Serine H4 folate Methylene H4 folate NH 2 H Glycine The main pathway to glycine is a 1-step reaction catalyzed by serine hydroxymethyl-transferase. This reaction involves the transfer of the hydroxymethyl group from serine to the cofactor tetrahydrofolate (THF), producing glycine and N 5,N 10 - methylene-thf Koval, 2007 (C) 74
75 Glycine Formation from Choline H N + Choline [2H] Choline oxidase N + Betaine aldehyde NAD + NADH + H + Betaine aldehyde dehydrogenase - Betaine N + H 2 N H Glycine [CH 2 ] formaldehyde Sarcosine oxidase H NH Sarcosine [CH 2 ] formaldehyde Dimethylglycine oxidase H [CH 3 ] N Dimethylglycine Koval, 2007 (C) 75
76 Aspartate/Asparagine and Glutamate/Glutamine Biosynthesis Glutamate is synthesized by the reductive amination of a-ketoglutarate catalyzed by glutamate dehydrogenase; a-kg + NH 3 + NADPH+H + Glu + NADP + +H 2 it is thus a nitrogen-fixing reaction. In addition, glutamate arises by aminotransferase reactions, with the amino nitrogen being donated by a number of different amino acids. Thus, glutamate is a general collector of amino nitrogen Koval, 2007 (C) 76
77 H Aspartate/Asparagine and Glutamate/Glutamine Biosynthesis (cont d) NH 3 H 2 H 2 N H NH 2 L-Aspartate NH 2 L-Asparagine H Aspartate is formed in a transamintion reaction catalyzed by aspartate transaminase, AST. This reaction uses the aspartate a- keto acid analog, oxaloacetate, and glutamate as the amino donor. Aspartate can also be formed by deamination of asparagine catalyzed by asparaginase Koval, 2007 (C) 77
78 Aspartate/Asparagine and Glutamate/Glutamine Biosynthesis (cont d) Asparagine synthetase and glutamine synthetase, catalyze the production of asparagine and glutamine from their respective a-amino acids. Glutamine is produced from glutamate by the direct incorporation of ammonia; and this can be considered another nitrogen fixing reaction. Asparagine, however, is formed by an amidotransferase reaction Koval, 2007 (C) 78
79 Selected Biomolecules Derived from Amino Acids Koval, 2007 (C) 79

")

80 Koval, 2007 (C) 80
Biochemistry: A Short Course
 Tymoczko Berg Stryer Biochemistry: A Short Course Second Edition CHAPTER 30 Amino Acid Degradation and the Urea Cycle 2013 W. H. Freeman and Company Chapter 30 Outline Amino acids are obtained from the
Tymoczko Berg Stryer Biochemistry: A Short Course Second Edition CHAPTER 30 Amino Acid Degradation and the Urea Cycle 2013 W. H. Freeman and Company Chapter 30 Outline Amino acids are obtained from the
Amino acid metabolism I
 Amino acid metabolism I Jana Novotná Department of the Medical Chemistry and Clinical Biochemistry The 2nd Faculty of Medicine, Charles Univ. Metabolic relationship of amino acids DIETARY PROTEINS GLYCOLYSIS
Amino acid metabolism I Jana Novotná Department of the Medical Chemistry and Clinical Biochemistry The 2nd Faculty of Medicine, Charles Univ. Metabolic relationship of amino acids DIETARY PROTEINS GLYCOLYSIS
Amino Acid Oxidation and the Urea Cycle
 Amino Acid Oxidation and the Urea Cycle Amino Acids: Final class of biomolecules whose oxidation contributes significantly to the generation of energy Undergo oxidation in three metabolic circumstances
Amino Acid Oxidation and the Urea Cycle Amino Acids: Final class of biomolecules whose oxidation contributes significantly to the generation of energy Undergo oxidation in three metabolic circumstances
Biochemistry: A Short Course
 Tymoczko Berg Stryer Biochemistry: A Short Course Second Edition CHAPTER 30 Amino Acid Degradation and the Urea Cycle 2013 W. H. Freeman and Company In the cytosol of a cell amino groups from amino acids
Tymoczko Berg Stryer Biochemistry: A Short Course Second Edition CHAPTER 30 Amino Acid Degradation and the Urea Cycle 2013 W. H. Freeman and Company In the cytosol of a cell amino groups from amino acids
Catabolism of Carbon skeletons of Amino acids. Amino acid metabolism
 Catabolism of Carbon skeletons of Amino acids Amino acid metabolism Carbon skeleton Carbon Skeleton a carbon skeleton is the internal structure of organic molecules. Carbon Arrangements The arrangement
Catabolism of Carbon skeletons of Amino acids Amino acid metabolism Carbon skeleton Carbon Skeleton a carbon skeleton is the internal structure of organic molecules. Carbon Arrangements The arrangement
number Done by Corrected by Doctor Dr.Diala
 number 32 Done by Mousa Salah Corrected by Bahaa Najjar Doctor Dr.Diala 1 P a g e In the last lecture we talked about the common processes between all amino acids which are: transamination, deamination,
number 32 Done by Mousa Salah Corrected by Bahaa Najjar Doctor Dr.Diala 1 P a g e In the last lecture we talked about the common processes between all amino acids which are: transamination, deamination,
Lecture 10 - Protein Turnover and Amino Acid Catabolism
 Lecture 10 - Protein Turnover and Amino Acid Catabolism Chem 454: Regulatory Mechanisms in Biochemistry University of Wisconsin-Eau Claire 1 Introduction 2 Proteins are degraded into amino acids. Protein
Lecture 10 - Protein Turnover and Amino Acid Catabolism Chem 454: Regulatory Mechanisms in Biochemistry University of Wisconsin-Eau Claire 1 Introduction 2 Proteins are degraded into amino acids. Protein
The diagram below summarizes the conversion of the twenty standard amino acids. Copyright Mark Brandt, Ph.D. 23
 Amino acid breakdown Amino acids comprise one of the three major energy sources for animals. They are an especially important energy source for carnivorous animals, and for all animals during early starvation
Amino acid breakdown Amino acids comprise one of the three major energy sources for animals. They are an especially important energy source for carnivorous animals, and for all animals during early starvation
PROTEIN METABOLISM: SPECIFIC WAYS OF AMINO ACIDS CATABOLISM AND SYNTHESIS
 PROTEIN METABOLISM: SPECIFIC WAYS OF AMINO ACIDS CATABOLISM AND SYNTHESIS SPECIFIC WAYS OF AMINO ACID CATABOLISM After removing of amino group the carbon skeletons of amino acids are transformed into metabolic
PROTEIN METABOLISM: SPECIFIC WAYS OF AMINO ACIDS CATABOLISM AND SYNTHESIS SPECIFIC WAYS OF AMINO ACID CATABOLISM After removing of amino group the carbon skeletons of amino acids are transformed into metabolic
Nitrogen Metabolism. Overview
 Nitrogen Metabolism Pratt and Cornely Chapter 18 Overview Nitrogen assimilation Amino acid biosynthesis Nonessential aa Essential aa Nucleotide biosynthesis Amino Acid Catabolism Urea Cycle Juicy Steak
Nitrogen Metabolism Pratt and Cornely Chapter 18 Overview Nitrogen assimilation Amino acid biosynthesis Nonessential aa Essential aa Nucleotide biosynthesis Amino Acid Catabolism Urea Cycle Juicy Steak
AMINO ACID METABOLISM
 AMINO ACID METABOLISM Synthesis of Urea in Liver The series of reactions that form urea is known as the Urea Cycle or the Krebs-Henseleit Cycle. The urea cycle operates only to eliminate excess nitrogen.
AMINO ACID METABOLISM Synthesis of Urea in Liver The series of reactions that form urea is known as the Urea Cycle or the Krebs-Henseleit Cycle. The urea cycle operates only to eliminate excess nitrogen.
Amino acid metabolism
 Amino acid metabolism The important reaction commonly employed in the breakdown of an amino acid is always the removal of its -amino group. The product ammonia is excreted after conversion to urea or other
Amino acid metabolism The important reaction commonly employed in the breakdown of an amino acid is always the removal of its -amino group. The product ammonia is excreted after conversion to urea or other
Nitrogen Metabolism. Overview
 Nitrogen Metabolism Pratt and Cornely Chapter 18 Overview Nitrogen assimilation Amino acid biosynthesis Nonessential aa Essential aa Nucleotide biosynthesis Amino Acid Catabolism Urea Cycle Juicy Steak
Nitrogen Metabolism Pratt and Cornely Chapter 18 Overview Nitrogen assimilation Amino acid biosynthesis Nonessential aa Essential aa Nucleotide biosynthesis Amino Acid Catabolism Urea Cycle Juicy Steak
AMINO ACID METABOLISM. Sri Widia A Jusman Dept. of Biochemistry & Molecular Biology FMUI
 AMINO ACID METABOLISM Sri Widia A Jusman Dept. of Biochemistry & Molecular Biology FMUI Amino acids derived from dietary protein absorbed from intestine through blood taken up by tissues used for biosynthesis
AMINO ACID METABOLISM Sri Widia A Jusman Dept. of Biochemistry & Molecular Biology FMUI Amino acids derived from dietary protein absorbed from intestine through blood taken up by tissues used for biosynthesis
Nitrogen Metabolism. Pratt and Cornely Chapter 18
 Nitrogen Metabolism Pratt and Cornely Chapter 18 Overview Nitrogen assimilation Amino acid biosynthesis Nonessential aa Essential aa Nucleotide biosynthesis Amino Acid Catabolism Urea Cycle Juicy Steak
Nitrogen Metabolism Pratt and Cornely Chapter 18 Overview Nitrogen assimilation Amino acid biosynthesis Nonessential aa Essential aa Nucleotide biosynthesis Amino Acid Catabolism Urea Cycle Juicy Steak
Amino acid metabolism II
 Amino acid metabolism II Fates of amino acid carbon skeleton degradation to common intermediates pyruvate, intermediates of citric acid cycle, acetyl-coa Glucogenic AA precursors of glucose - degradation
Amino acid metabolism II Fates of amino acid carbon skeleton degradation to common intermediates pyruvate, intermediates of citric acid cycle, acetyl-coa Glucogenic AA precursors of glucose - degradation
Metabolism of amino acids. Vladimíra Kvasnicová
 Metabolism of amino acids Vladimíra Kvasnicová Classification of proteinogenic AAs -metabolic point of view 1) biosynthesis in a human body nonessential (are synthesized) essential (must be present in
Metabolism of amino acids Vladimíra Kvasnicová Classification of proteinogenic AAs -metabolic point of view 1) biosynthesis in a human body nonessential (are synthesized) essential (must be present in
Amino Acid Metabolism: Amino Acid Degradation & Synthesis
 Amino Acid Metabolism: Amino Acid Degradation & Synthesis Dr. Diala Abu-Hassan, DDS, PhD Medical students-first semester All images are taken from Lippincott s Biochemistry textbook except where noted
Amino Acid Metabolism: Amino Acid Degradation & Synthesis Dr. Diala Abu-Hassan, DDS, PhD Medical students-first semester All images are taken from Lippincott s Biochemistry textbook except where noted
Midterm 2. Low: 14 Mean: 61.3 High: 98. Standard Deviation: 17.7
 Midterm 2 Low: 14 Mean: 61.3 High: 98 Standard Deviation: 17.7 Lecture 17 Amino Acid Metabolism Review of Urea Cycle N and S assimilation Last cofactors: THF and SAM Synthesis of few amino acids Dietary
Midterm 2 Low: 14 Mean: 61.3 High: 98 Standard Deviation: 17.7 Lecture 17 Amino Acid Metabolism Review of Urea Cycle N and S assimilation Last cofactors: THF and SAM Synthesis of few amino acids Dietary
Amino acid Catabolism
 Enzymatic digestion of dietary proteins in gastrointestinal-tract. Amino acid Catabolism Amino acids: 1. There are 20 different amino acid, they are monomeric constituents of proteins 2. They act as precursors
Enzymatic digestion of dietary proteins in gastrointestinal-tract. Amino acid Catabolism Amino acids: 1. There are 20 different amino acid, they are monomeric constituents of proteins 2. They act as precursors
AMINO ACID METABOLISM
 AMINO ACID METABOLISM PHL-285 Biochemistry-2 Mahmoud N. Nagi, Ph.D. Professor of Biochemistry Overview of amino acid metabolism. Classification of amino acids. Biosynthesis of nonessential amino acids.
AMINO ACID METABOLISM PHL-285 Biochemistry-2 Mahmoud N. Nagi, Ph.D. Professor of Biochemistry Overview of amino acid metabolism. Classification of amino acids. Biosynthesis of nonessential amino acids.
Fate of Dietary Protein
 Fate of Dietary Protein Dietary protein Stomach: l, pepsin Denatured and partially hydrolyzed protein (large polypeptides) small intestine: proteases Amino acids and dipeptides intestinal lining: proteases
Fate of Dietary Protein Dietary protein Stomach: l, pepsin Denatured and partially hydrolyzed protein (large polypeptides) small intestine: proteases Amino acids and dipeptides intestinal lining: proteases
Amino Acid Metabolism
 Amino Acid Metabolism Fate of Dietary Protein Dietary protein Stomach: l, pepsin Denatured and partially hydrolyzed protein (large polypeptides) small intestine: proteases Amino acids and dipeptides intestinal
Amino Acid Metabolism Fate of Dietary Protein Dietary protein Stomach: l, pepsin Denatured and partially hydrolyzed protein (large polypeptides) small intestine: proteases Amino acids and dipeptides intestinal
Lecture 17: Nitrogen metabolism 1. Urea cycle detoxification of NH 3 2. Amino acid degradation
 Lecture 17: Nitrogen metabolism 1. Urea cycle detoxification of NH 3 2. Amino acid degradation Reference material Biochemistry 4 th edition, Mathews, Van Holde, Appling, Anthony Cahill. Pearson ISBN:978
Lecture 17: Nitrogen metabolism 1. Urea cycle detoxification of NH 3 2. Amino acid degradation Reference material Biochemistry 4 th edition, Mathews, Van Holde, Appling, Anthony Cahill. Pearson ISBN:978
SYNTHESIS OF NON-ESSENTIAL AMINO ACIDS [LIPPINCOTT S ] Deeba S. Jairajpuri
![SYNTHESIS OF NON-ESSENTIAL AMINO ACIDS [LIPPINCOTT S ] Deeba S. Jairajpuri](/thumbs/80/80689583.jpg "SYNTHESIS OF NON-ESSENTIAL AMINO ACIDS [LIPPINCOTT S ] Deeba S. Jairajpuri") SYNTHESIS OF NON-ESSENTIAL AMINO ACIDS [LIPPINCOTT S 267-274] Deeba S. Jairajpuri TYPES OF AMINO ACIDS Amino acids that cannot be synthesized by the human body and are required to be taken in the diet
SYNTHESIS OF NON-ESSENTIAL AMINO ACIDS [LIPPINCOTT S 267-274] Deeba S. Jairajpuri TYPES OF AMINO ACIDS Amino acids that cannot be synthesized by the human body and are required to be taken in the diet
Part III => METABOLISM and ENERGY. 3.5 Protein Catabolism 3.5a Protein Degradation 3.5b Amino Acid Breakdown 3.5c Urea Cycle
 Part III => METABOLISM and ENERGY 3.5 Protein Catabolism 3.5a Protein Degradation 3.5b Amino Acid Breakdown 3.5c Urea Cycle Section 3.5a: Protein Degradation Synopsis 3.5a - Dietary proteins are degraded
Part III => METABOLISM and ENERGY 3.5 Protein Catabolism 3.5a Protein Degradation 3.5b Amino Acid Breakdown 3.5c Urea Cycle Section 3.5a: Protein Degradation Synopsis 3.5a - Dietary proteins are degraded
1 Digestion and absorption. Lecture #14 Lecturer: PhD Alexander N. Koval
 1 Digestion and absorption Lecture #14 Lecturer: PhD Alexander N. Koval Presentation of Protein 12/22/2016 A. Koval (C), 2016 2 Lectures plan 12/22/2016 A. Koval (C), 2016 3 Overview of Protein Metabolism
1 Digestion and absorption Lecture #14 Lecturer: PhD Alexander N. Koval Presentation of Protein 12/22/2016 A. Koval (C), 2016 2 Lectures plan 12/22/2016 A. Koval (C), 2016 3 Overview of Protein Metabolism
Amino Acid Metabolism
 Amino Acid Metabolism Last Week Most of the Animal Kingdom = Lazy - Most higher organisms in the animal kindom don t bother to make all of the amino acids. - Instead, we eat things that make the essential
Amino Acid Metabolism Last Week Most of the Animal Kingdom = Lazy - Most higher organisms in the animal kindom don t bother to make all of the amino acids. - Instead, we eat things that make the essential
Midterm 2 Results. Standard Deviation:
 Midterm 2 Results High: Low: Mean: Standard Deviation: 97.5% 16% 58% 16.3 Lecture 17 Amino Acid Metabolism Urea Cycle N and S assimilation Last cofactors: THF and SAM Dietary (Exogenous) Proteins Hydrolyzed
Midterm 2 Results High: Low: Mean: Standard Deviation: 97.5% 16% 58% 16.3 Lecture 17 Amino Acid Metabolism Urea Cycle N and S assimilation Last cofactors: THF and SAM Dietary (Exogenous) Proteins Hydrolyzed
Amino acid metabolism I
 Amino acid metabolism I Jana Novotná, Bruno Sopko Department of the Medical Chemistry and Clinical Biochemistry The 2nd Faculty of Medicine, Charles Univ. Metabolic relationship of amino acids Body proteins
Amino acid metabolism I Jana Novotná, Bruno Sopko Department of the Medical Chemistry and Clinical Biochemistry The 2nd Faculty of Medicine, Charles Univ. Metabolic relationship of amino acids Body proteins
Welcome to Class 14! Class 14: Outline and Objectives. Overview of amino acid catabolism! Introductory Biochemistry!
 Welcome to Class 14 Introductory Biochemistry Class 14: Outline and Objectives Amino Acid Catabolism Fates of amino groups transamination urea cycle Fates of carbon skeletons important cofactors metabolic
Welcome to Class 14 Introductory Biochemistry Class 14: Outline and Objectives Amino Acid Catabolism Fates of amino groups transamination urea cycle Fates of carbon skeletons important cofactors metabolic
NITROGEN METABOLISM An Overview
 1 University of Papua New Guinea School of Medicine and Health Sciences Division of Basic Medical Sciences Discipline of Biochemistry and Molecular Biology PBL Seminar & Health Sciences NITROGEN METABOLISM
1 University of Papua New Guinea School of Medicine and Health Sciences Division of Basic Medical Sciences Discipline of Biochemistry and Molecular Biology PBL Seminar & Health Sciences NITROGEN METABOLISM
Lecture 11 - Biosynthesis of Amino Acids
 Lecture 11 - Biosynthesis of Amino Acids Chem 454: Regulatory Mechanisms in Biochemistry University of Wisconsin-Eau Claire 1 Introduction Biosynthetic pathways for amino acids, nucleotides and lipids
Lecture 11 - Biosynthesis of Amino Acids Chem 454: Regulatory Mechanisms in Biochemistry University of Wisconsin-Eau Claire 1 Introduction Biosynthetic pathways for amino acids, nucleotides and lipids
Amino Acid Metabolism (Nitrogen Metabolism) Dec Dr. Robert Lyons
 Dec Dr. Robert Lyons") 1 Amino Acid Metabolism (itrogen Metabolism) Dec 9-11 008 Dr. Robert Lyons See: http://seqcore.brcf.med.umich.edu/mcb500 for supplementary (non-required) course materials. Medical relevance of amino acid
1 Amino Acid Metabolism (itrogen Metabolism) Dec 9-11 008 Dr. Robert Lyons See: http://seqcore.brcf.med.umich.edu/mcb500 for supplementary (non-required) course materials. Medical relevance of amino acid
Chapter 26. Outline. Nitrogen. Nitrogen and Amino Acid Metabolism. BCH 4054 Spring 2001 Chapter 26 Lecture Notes. Slide 1. Slide 2
 BCH 4054 Spring 2001 Chapter 26 Lecture Notes 1 Chapter 26 Nitrogen and Amino Acid Metabolism 2 utline No time to cover entire chapter, therefore concentrate on a few focal points Assimilation of inorganic
BCH 4054 Spring 2001 Chapter 26 Lecture Notes 1 Chapter 26 Nitrogen and Amino Acid Metabolism 2 utline No time to cover entire chapter, therefore concentrate on a few focal points Assimilation of inorganic
18 Amino Acid Oxidation and Production of Urea W. H. Freeman and Company
 18 Amino Acid Oxidation and Production of Urea 2013 W. H. Freeman and Company 1 Last Class of Biomolecules For Energy 1. Production of acetyl-coa. Glucose. To pyruvate via glycolysis. To acetyl-coa by
18 Amino Acid Oxidation and Production of Urea 2013 W. H. Freeman and Company 1 Last Class of Biomolecules For Energy 1. Production of acetyl-coa. Glucose. To pyruvate via glycolysis. To acetyl-coa by
Amino Acid Metabolism
 Amino Acid Metabolism The continuous degradation and synthesis of cellular proteins occur in all forms of life. Each day humans turn over 1 2% of their total body protein, principally muscle protein. Approximately
Amino Acid Metabolism The continuous degradation and synthesis of cellular proteins occur in all forms of life. Each day humans turn over 1 2% of their total body protein, principally muscle protein. Approximately
Amino Acid Catabolism
 Amino Acid atabolism 3-1 Lec #8 To date we have considered the catabolism of carbohydrates and lipids with the object of generating energy in the form of ATP. Both give rise to AcoA which is fed through
Amino Acid atabolism 3-1 Lec #8 To date we have considered the catabolism of carbohydrates and lipids with the object of generating energy in the form of ATP. Both give rise to AcoA which is fed through
BIOCHEMISTRY Protein Metabolism
 BIOCHEMISTRY Protein Metabolism BIOB111 CHEMISTRY & BIOCHEMISTRY Session 25 Session Plan Digestion & Absorption of Proteins Amino Acid Utilization Amino Acid Degradation Transamination Oxidative Deamination
BIOCHEMISTRY Protein Metabolism BIOB111 CHEMISTRY & BIOCHEMISTRY Session 25 Session Plan Digestion & Absorption of Proteins Amino Acid Utilization Amino Acid Degradation Transamination Oxidative Deamination
AMINOACID METABOLISM FATE OF AMINOACIDS & UREA CYCLE
 AMINOACID METABOLISM FATE OF AMINOACIDS & UREA CYCLE SOURCE & FATE OF AA The aminoacids obtained from DIETARY SOURCE or BODY PROTEIN TURNOVER are utilized for protein biosynthesis and the production of
AMINOACID METABOLISM FATE OF AMINOACIDS & UREA CYCLE SOURCE & FATE OF AA The aminoacids obtained from DIETARY SOURCE or BODY PROTEIN TURNOVER are utilized for protein biosynthesis and the production of
Jana Novotná, Bruno Sopko. Department of the Medical Chemistry and Clinical Biochemistry The 2nd Faculty of Medicine, Charles Univ.
 Amino acid metabolism II. Urea cycle Jana Novotná, Bruno Sopko Department of the Medical Chemistry and Clinical Biochemistry The 2nd Faculty of Medicine, Charles Univ. Nitrogen balance Tissue proteins
Amino acid metabolism II. Urea cycle Jana Novotná, Bruno Sopko Department of the Medical Chemistry and Clinical Biochemistry The 2nd Faculty of Medicine, Charles Univ. Nitrogen balance Tissue proteins
Lecture: Amino Acid catabolism: Nitrogen-The Urea cycle
 BIOC 423: Introductory Biochemistry Biochemistry Education Department of Biochemistry & Molecular Biology University of New Mexico Lecture: Amino Acid catabolism: Nitrogen-The Urea cycle OBJECTIVES Describe
BIOC 423: Introductory Biochemistry Biochemistry Education Department of Biochemistry & Molecular Biology University of New Mexico Lecture: Amino Acid catabolism: Nitrogen-The Urea cycle OBJECTIVES Describe
Urea is the major end product of nitrogen catabolism in humans One nitrogen free NH3 other nitrogen aspartate. carbon oxygen CO2 liver,
 Urea is the major end product of nitrogen catabolism in humans Urea is the major disposal form of amino groups derived from amino acids, and accounts about 90% percent of the nitrogencontaining components
Urea is the major end product of nitrogen catabolism in humans Urea is the major disposal form of amino groups derived from amino acids, and accounts about 90% percent of the nitrogencontaining components
Dental Students Biochemistry Exam V Questions ( Note: In all cases, the only correct answer is the best answer)
") Dental Students Biochemistry Exam V Questions - 2006 ( Note: In all cases, the only correct answer is the best answer) 1. Essential fatty acids are: A. precursors of biotin B. precursors of tyrosine C.
Dental Students Biochemistry Exam V Questions - 2006 ( Note: In all cases, the only correct answer is the best answer) 1. Essential fatty acids are: A. precursors of biotin B. precursors of tyrosine C.
Conversion of amino acids الفريق الطبي األكاديمي
 الفريق الطبي األكاديمي لكية الطب البرشي البلقاء التطبيقية / املركز 2022 2016/ -the ones with * are the essential amino acids- - Histidine decarboxylation -> histamine Page 1 - Glutamine is a nitrogen carrier
الفريق الطبي األكاديمي لكية الطب البرشي البلقاء التطبيقية / املركز 2022 2016/ -the ones with * are the essential amino acids- - Histidine decarboxylation -> histamine Page 1 - Glutamine is a nitrogen carrier
Amino Acid Metabolism Parts I-III
 M1 - Biochemistry Amino Acid Metabolism Parts I-III Dr. Diegelmann 10/2, 10/3, 10/7 1 Biochemical Pathways; If you know all of this you can get an A 2 OBJECTIVES After studying the material presented in
M1 - Biochemistry Amino Acid Metabolism Parts I-III Dr. Diegelmann 10/2, 10/3, 10/7 1 Biochemical Pathways; If you know all of this you can get an A 2 OBJECTIVES After studying the material presented in
NITROGEN METABOLISM: An Overview
 NITROGEN METABOLISM: An Overview University of PNG School of Medicine and Health Sciences Division of Basic Medical Sciences Discipline of Biochemistry & Molecular Biology VJ Temple 1 How are nitrogen-containing
NITROGEN METABOLISM: An Overview University of PNG School of Medicine and Health Sciences Division of Basic Medical Sciences Discipline of Biochemistry & Molecular Biology VJ Temple 1 How are nitrogen-containing
Nitrogen Metabolism. Dr. Kevin Ahern
 Nitrogen Metabolism Dr. Kevin Ahern Nitrogen Metabolism Nitrogen Forms in the Body Nitrogen Balance Critical Body Must Make and Break Down Amino Acids Nitrogen Also Needed for Synthesis of Nucleotides
Nitrogen Metabolism Dr. Kevin Ahern Nitrogen Metabolism Nitrogen Forms in the Body Nitrogen Balance Critical Body Must Make and Break Down Amino Acids Nitrogen Also Needed for Synthesis of Nucleotides
number Done by Corrected by Doctor
 number 33 Done by Omar Sami Corrected by Waseem abu obeida Doctor Diala Too late for second guessing, too late to go back to sleep 1 P age Just try defying gravity This sheet will be divided to two parts,
number 33 Done by Omar Sami Corrected by Waseem abu obeida Doctor Diala Too late for second guessing, too late to go back to sleep 1 P age Just try defying gravity This sheet will be divided to two parts,
Metabolism of amino acids I. Josef Fontana
 Metabolism of amino acids I Josef Fontana EC Overview of the lecture Introduction to protein and amino acids metabolism Metabolic pathways of amino acids Transamination Conversion glutamate - glutamine
Metabolism of amino acids I Josef Fontana EC Overview of the lecture Introduction to protein and amino acids metabolism Metabolic pathways of amino acids Transamination Conversion glutamate - glutamine
Biochemistry 2 Recita0on Amino Acid Metabolism
 Biochemistry 2 Recita0on Amino Acid Metabolism 04-20- 2015 Glutamine and Glutamate as key entry points for NH 4 + Amino acid catabolism Glutamine synthetase enables toxic NH 4 + to combine with glutamate
Biochemistry 2 Recita0on Amino Acid Metabolism 04-20- 2015 Glutamine and Glutamate as key entry points for NH 4 + Amino acid catabolism Glutamine synthetase enables toxic NH 4 + to combine with glutamate
Biochemistry: A Short Course
 Tymoczko Berg Stryer Biochemistry: A Short Course Second Edition CHAPTER 31 Amino Acid Synthesis 2013 W. H. Freeman and Company Chapter 31 Outline Although the atmosphere is approximately 80% nitrogen,
Tymoczko Berg Stryer Biochemistry: A Short Course Second Edition CHAPTER 31 Amino Acid Synthesis 2013 W. H. Freeman and Company Chapter 31 Outline Although the atmosphere is approximately 80% nitrogen,
AMINO ACIDS NON-ESSENTIAL ESSENTIAL
 Edith Frederika Introduction A major component of food is PROTEIN The protein ingested as part of our diet are not the same protein required by the body Only 40 to 50 gr of protein is required by a normal
Edith Frederika Introduction A major component of food is PROTEIN The protein ingested as part of our diet are not the same protein required by the body Only 40 to 50 gr of protein is required by a normal
Amino acid oxidation and the production of urea
 Seminar 10 Urea cycle Amino acid oxidation and the production of urea Oxidation Waste or Reuse Ammonia has to be eliminated ammonia originates in the catabolism of amino acids that are primarily produced
Seminar 10 Urea cycle Amino acid oxidation and the production of urea Oxidation Waste or Reuse Ammonia has to be eliminated ammonia originates in the catabolism of amino acids that are primarily produced
METABOLISM OF AMINO ACIDS
 Dr. M. Sasvari METABOLISM OF AMINO ACIDS 2. The fate of the carbon sceleton 3 N + C R Active C 1 intermediers The folate derivatives structure s Folate (F) - vitamin Folate, 2 F, 4 F Dihydrofolate ( 2
Dr. M. Sasvari METABOLISM OF AMINO ACIDS 2. The fate of the carbon sceleton 3 N + C R Active C 1 intermediers The folate derivatives structure s Folate (F) - vitamin Folate, 2 F, 4 F Dihydrofolate ( 2
CH395G FINAL (3 rd ) EXAM Kitto/Hackert - Fall 2003
 EXAM Kitto/Hackert - Fall 2003") CH395G FINAL (3 rd ) EXAM Kitto/Hackert - Fall 2003 1. A cell in an active, catabolic state has a. a high (ATP/ADP) and a high (NADH/NAD + ) ratio b. a high (ATP/ADP) and a low (NADH/NAD + ) ratio c. a
CH395G FINAL (3 rd ) EXAM Kitto/Hackert - Fall 2003 1. A cell in an active, catabolic state has a. a high (ATP/ADP) and a high (NADH/NAD + ) ratio b. a high (ATP/ADP) and a low (NADH/NAD + ) ratio c. a
endopeptidases aminopeptidases carboxypeptidases hydrolyzes a peptide bond somewhere in the middle of the polypeptide
 1 Amino Acid Metabolism: The primary purpose for s in the body is to provide the building blocks for proteins R other s. owever, if there is no protein synthesis occurring, the s can be broken down (i.e.
1 Amino Acid Metabolism: The primary purpose for s in the body is to provide the building blocks for proteins R other s. owever, if there is no protein synthesis occurring, the s can be broken down (i.e.
E.coli Core Model: Metabolic Core
 1 E.coli Core Model: Metabolic Core 2 LEARNING OBJECTIVES Each student should be able to: Describe the glycolysis pathway in the core model. Describe the TCA cycle in the core model. Explain gluconeogenesis.
1 E.coli Core Model: Metabolic Core 2 LEARNING OBJECTIVES Each student should be able to: Describe the glycolysis pathway in the core model. Describe the TCA cycle in the core model. Explain gluconeogenesis.
UNIT 2 Amino acids and Proteins
 UNIT 2 Amino acids and Proteins Significance of Proteins 1. Keep the cells and tissues growing, renewing and mending 2. Take part in some kinds of important physiological activities 3. Oxidation and supply
UNIT 2 Amino acids and Proteins Significance of Proteins 1. Keep the cells and tissues growing, renewing and mending 2. Take part in some kinds of important physiological activities 3. Oxidation and supply
M1 - Renal, Fall 2007
 University of Michigan Deep Blue deepblue.lib.umich.edu 2007-09 M1 - Renal, Fall 2007 Lyons, R.; Burney, R. Lyons, R., Burney, R. (2008, August 07). Renal. Retrieved from Open.Michigan - Educational Resources
University of Michigan Deep Blue deepblue.lib.umich.edu 2007-09 M1 - Renal, Fall 2007 Lyons, R.; Burney, R. Lyons, R., Burney, R. (2008, August 07). Renal. Retrieved from Open.Michigan - Educational Resources
VITAMIN B6 History B
 VITAMIN B 6 History Gyorgy demonstrated a characteristic type of dermatitis in rats produced by a factor different from known vitamins 1937 : He showed that the factor was present in yeast & liver and
VITAMIN B 6 History Gyorgy demonstrated a characteristic type of dermatitis in rats produced by a factor different from known vitamins 1937 : He showed that the factor was present in yeast & liver and
Biomolecules: amino acids
 Biomolecules: amino acids Amino acids Amino acids are the building blocks of proteins They are also part of hormones, neurotransmitters and metabolic intermediates There are 20 different amino acids in
Biomolecules: amino acids Amino acids Amino acids are the building blocks of proteins They are also part of hormones, neurotransmitters and metabolic intermediates There are 20 different amino acids in
Integration Of Metabolism
 Integration Of Metabolism Metabolism Consist of Highly Interconnected Pathways The basic strategy of catabolic metabolism is to form ATP, NADPH, and building blocks for biosyntheses. 1. ATP is the universal
Integration Of Metabolism Metabolism Consist of Highly Interconnected Pathways The basic strategy of catabolic metabolism is to form ATP, NADPH, and building blocks for biosyntheses. 1. ATP is the universal
Dynamics of Protein And Amino Acid Metabolism
 Dynamics of Protein And Amino Acid Metabolism Dietary Proteins Digestion to Amino Acids Transport in Blood to Cells Protein Synthesis Functional Proteins Amino Acids Metabolites Protein Degradation In
Dynamics of Protein And Amino Acid Metabolism Dietary Proteins Digestion to Amino Acids Transport in Blood to Cells Protein Synthesis Functional Proteins Amino Acids Metabolites Protein Degradation In
Lynne A. Wolfe, MS, ACNP, PNP, BC Department of Genetics Yale School of Medicine
 Lynne A. Wolfe, MS, ACNP, PNP, BC Department of Genetics Yale School of Medicine Harvey Levy, MD Mark Korson, MD Piero Rinaldo, MD, PhD Larry Sweetman, PhD K. Michael Gibson, PhD Charlie Roe, MD Jerry
Lynne A. Wolfe, MS, ACNP, PNP, BC Department of Genetics Yale School of Medicine Harvey Levy, MD Mark Korson, MD Piero Rinaldo, MD, PhD Larry Sweetman, PhD K. Michael Gibson, PhD Charlie Roe, MD Jerry
CITRIC ACID CYCLE ERT106 BIOCHEMISTRY SEM /19 BY: MOHAMAD FAHRURRAZI TOMPANG
 CITRIC ACID CYCLE ERT106 BIOCHEMISTRY SEM 1 2018/19 BY: MOHAMAD FAHRURRAZI TOMPANG Chapter Outline (19-1) The central role of the citric acid cycle in metabolism (19-2) The overall pathway of the citric
CITRIC ACID CYCLE ERT106 BIOCHEMISTRY SEM 1 2018/19 BY: MOHAMAD FAHRURRAZI TOMPANG Chapter Outline (19-1) The central role of the citric acid cycle in metabolism (19-2) The overall pathway of the citric
SCBC203 Amino Acid Metabolism
 Breakdown of proteins Route I: Dietary protein breakdown SCBC203 Amino Acid Metabolism Dr Sarawut Jitrapakdee Professor of Biochemistry Department of Biochemistry Faculty of Science Mahidol University
Breakdown of proteins Route I: Dietary protein breakdown SCBC203 Amino Acid Metabolism Dr Sarawut Jitrapakdee Professor of Biochemistry Department of Biochemistry Faculty of Science Mahidol University
Classification of amino acids: -
 Page 1 of 8 P roteinogenic amino acids, also known as standard, normal or primary amino acids are 20 amino acids that are incorporated in proteins and that are coded in the standard genetic code (subunit
Page 1 of 8 P roteinogenic amino acids, also known as standard, normal or primary amino acids are 20 amino acids that are incorporated in proteins and that are coded in the standard genetic code (subunit
Glycolysis. Cellular Respiration
 Glucose is the preferred carbohydrate of cells. In solution, it can change from a linear chain to a ring. Energy is stored in the bonds of the carbohydrates. Breaking these bonds releases that energy.
Glucose is the preferred carbohydrate of cells. In solution, it can change from a linear chain to a ring. Energy is stored in the bonds of the carbohydrates. Breaking these bonds releases that energy.
Citric Acid Cycle: Central Role in Catabolism. Entry of Pyruvate into the TCA cycle
 Citric Acid Cycle: Central Role in Catabolism Stage II of catabolism involves the conversion of carbohydrates, fats and aminoacids into acetylcoa In aerobic organisms, citric acid cycle makes up the final
Citric Acid Cycle: Central Role in Catabolism Stage II of catabolism involves the conversion of carbohydrates, fats and aminoacids into acetylcoa In aerobic organisms, citric acid cycle makes up the final
Prof Dr Mohammad Ibrahim Prof of Medical Biochemistry
 Amino Acids Metabolism ١ i Metabolism ٢ NH2 Structure It is α-amino acetic acid Nutrional Value It is non-essential amino acid Metabolic Fate It is glucogenic g amino acid ٣ Biosynthesis 1. From 2 and
Amino Acids Metabolism ١ i Metabolism ٢ NH2 Structure It is α-amino acetic acid Nutrional Value It is non-essential amino acid Metabolic Fate It is glucogenic g amino acid ٣ Biosynthesis 1. From 2 and
Metabolism of proteins and amino acids
 BIOQUÍMICA E BIOLOGIA CELULAR António Ascensão, José Magalhães Metabolism of proteins and amino acids Faculdade de Desporto, Universidade do Porto, 1º Ciclo, 1º Ano 202_2013 Humans degradation of ingested
BIOQUÍMICA E BIOLOGIA CELULAR António Ascensão, José Magalhães Metabolism of proteins and amino acids Faculdade de Desporto, Universidade do Porto, 1º Ciclo, 1º Ano 202_2013 Humans degradation of ingested
Lipid and Amino Acid Metabolism
 CHEM 3331 Fundamentals of Biochemistry Chapter 14 Lipid and Amino Acid Metabolism Organic and Biochemistry for Today Spencer L. Seager / Michael R. Slabaugh Mr. Kevin A. Boudreaux Angelo State University
CHEM 3331 Fundamentals of Biochemistry Chapter 14 Lipid and Amino Acid Metabolism Organic and Biochemistry for Today Spencer L. Seager / Michael R. Slabaugh Mr. Kevin A. Boudreaux Angelo State University
CELLULAR METABOLISM. Metabolic pathways can be linear, branched, cyclic or spiral
 CHM333 LECTURE 24 & 25: 3/27 29/13 SPRING 2013 Professor Christine Hrycyna CELLULAR METABOLISM What is metabolism? - How cells acquire, transform, store and use energy - Study reactions in a cell and how
CHM333 LECTURE 24 & 25: 3/27 29/13 SPRING 2013 Professor Christine Hrycyna CELLULAR METABOLISM What is metabolism? - How cells acquire, transform, store and use energy - Study reactions in a cell and how
Chapter 24 Lecture Outline
 Chapter 24 Lecture Outline Carbohydrate Lipid and Protein! Metabolism! In the catabolism of carbohydrates, glycolysis converts glucose into pyruvate, which is then metabolized into acetyl CoA. Prepared
Chapter 24 Lecture Outline Carbohydrate Lipid and Protein! Metabolism! In the catabolism of carbohydrates, glycolysis converts glucose into pyruvate, which is then metabolized into acetyl CoA. Prepared
Reactions and amino acids structure & properties
 Lecture 2: Reactions and amino acids structure & properties Dr. Sameh Sarray Hlaoui Common Functional Groups Common Biochemical Reactions AH + B A + BH Oxidation-Reduction A-H + B-OH + energy ª A-B + H
Lecture 2: Reactions and amino acids structure & properties Dr. Sameh Sarray Hlaoui Common Functional Groups Common Biochemical Reactions AH + B A + BH Oxidation-Reduction A-H + B-OH + energy ª A-B + H
TCA CYCLE (Citric Acid Cycle)
") TCA CYCLE (Citric Acid Cycle) TCA CYCLE The Citric Acid Cycle is also known as: Kreb s cycle Sir Hans Krebs Nobel prize, 1953 TCA (tricarboxylic acid) cycle The citric acid cycle requires aerobic conditions!!!!
TCA CYCLE (Citric Acid Cycle) TCA CYCLE The Citric Acid Cycle is also known as: Kreb s cycle Sir Hans Krebs Nobel prize, 1953 TCA (tricarboxylic acid) cycle The citric acid cycle requires aerobic conditions!!!!
number Done by Corrected by Doctor Nayef Karadsheh
 number 13 Done by Asma Karameh Corrected by Saad hayek Doctor Nayef Karadsheh Gluconeogenesis This lecture covers gluconeogenesis with aspects of: 1) Introduction to glucose distribution through tissues.
number 13 Done by Asma Karameh Corrected by Saad hayek Doctor Nayef Karadsheh Gluconeogenesis This lecture covers gluconeogenesis with aspects of: 1) Introduction to glucose distribution through tissues.
Amino acids. Dr. Mamoun Ahram Summer semester,
 Amino acids Dr. Mamoun Ahram Summer semester, 2017-2018 Resources This lecture Campbell and Farrell s Biochemistry, Chapters 3 (pp.66-76) General structure (Chiral carbon) The amino acids that occur in
Amino acids Dr. Mamoun Ahram Summer semester, 2017-2018 Resources This lecture Campbell and Farrell s Biochemistry, Chapters 3 (pp.66-76) General structure (Chiral carbon) The amino acids that occur in
Marah Bitar. Faisal Nimri ... Nafeth Abu Tarboosh
 8 Marah Bitar Faisal Nimri... Nafeth Abu Tarboosh Summary of the 8 steps of citric acid cycle Step 1. Acetyl CoA joins with a four-carbon molecule, oxaloacetate, releasing the CoA group and forming a six-carbon
8 Marah Bitar Faisal Nimri... Nafeth Abu Tarboosh Summary of the 8 steps of citric acid cycle Step 1. Acetyl CoA joins with a four-carbon molecule, oxaloacetate, releasing the CoA group and forming a six-carbon
Integration of Metabolism
 Integration of Metabolism Metabolism is a continuous process. Thousands of reactions occur simultaneously in order to maintain homeostasis. It ensures a supply of fuel, to tissues at all times, in fed
Integration of Metabolism Metabolism is a continuous process. Thousands of reactions occur simultaneously in order to maintain homeostasis. It ensures a supply of fuel, to tissues at all times, in fed
Test de aminoácidos. Orina.
 SPECIMEN VALIDITY per creatinine INTERVAL 2.5 th 16 th 50 th 84 th 97.5 th Creatinine 57 mg/dl 15-120 Glutamine/Glutamate 4.3 5-160 Ammonia Level (NH 4 ) 42800 μm/g 18000-100000 Specimen Validity Index
SPECIMEN VALIDITY per creatinine INTERVAL 2.5 th 16 th 50 th 84 th 97.5 th Creatinine 57 mg/dl 15-120 Glutamine/Glutamate 4.3 5-160 Ammonia Level (NH 4 ) 42800 μm/g 18000-100000 Specimen Validity Index
Find this material useful? You can help our team to keep this site up and bring you even more content consider donating via the link on our site.
 Find this material useful? You can help our team to keep this site up and bring you even more content consider donating via the link on our site. Still having trouble understanding the material? Check
Find this material useful? You can help our team to keep this site up and bring you even more content consider donating via the link on our site. Still having trouble understanding the material? Check
1-To know what is protein 2-To identify Types of protein 3- To Know amino acids 4- To be differentiate between essential and nonessential amino acids
 Amino acids 1-To know what is protein 2-To identify Types of protein 3- To Know amino acids 4- To be differentiate between essential and nonessential amino acids 5-To understand amino acids synthesis Amino
Amino acids 1-To know what is protein 2-To identify Types of protein 3- To Know amino acids 4- To be differentiate between essential and nonessential amino acids 5-To understand amino acids synthesis Amino
Citric acid cycle and respiratory chain. Pavla Balínová
 Citric acid cycle and respiratory chain Pavla Balínová Mitochondria Structure of mitochondria: Outer membrane Inner membrane (folded) Matrix space (mtdna, ribosomes, enzymes of CAC, β-oxidation of FA,
Citric acid cycle and respiratory chain Pavla Balínová Mitochondria Structure of mitochondria: Outer membrane Inner membrane (folded) Matrix space (mtdna, ribosomes, enzymes of CAC, β-oxidation of FA,
Integrative Metabolism: Significance
 Integrative Metabolism: Significance Energy Containing Nutrients Carbohydrates Fats Proteins Catabolism Energy Depleted End Products H 2 O NH 3 ADP + Pi NAD + NADP + FAD + Pi NADH+H + NADPH+H + FADH2 Cell
Integrative Metabolism: Significance Energy Containing Nutrients Carbohydrates Fats Proteins Catabolism Energy Depleted End Products H 2 O NH 3 ADP + Pi NAD + NADP + FAD + Pi NADH+H + NADPH+H + FADH2 Cell
SIMPLE BASIC METABOLISM
 SIMPLE BASIC METABOLISM When we eat food such as a tuna fish sandwich, the polysaccharides, lipids, and proteins are digested to smaller molecules that are absorbed into the cells of our body. As these
SIMPLE BASIC METABOLISM When we eat food such as a tuna fish sandwich, the polysaccharides, lipids, and proteins are digested to smaller molecules that are absorbed into the cells of our body. As these
Lecture 16. Finish lipid metabolism (Triglycerides, Isoprenoids/Steroids, Glyoxylate cycle) Amino acid metabolism (Urea cycle) Google Man III
 Amino acid metabolism (Urea cycle) Google Man III") Lecture 16 Finish lipid metabolism (Triglycerides, Isoprenoids/Steroids, Glyoxylate cycle) Amino acid metabolism (Urea cycle) Google Man III The Powertrain of Human Metabolism (verview) CARBHYDRATES PRTEINS
Lecture 16 Finish lipid metabolism (Triglycerides, Isoprenoids/Steroids, Glyoxylate cycle) Amino acid metabolism (Urea cycle) Google Man III The Powertrain of Human Metabolism (verview) CARBHYDRATES PRTEINS
2. When a muscle depletes its supply of ATP, the next molecule used as an energy source is: a) pyruvate b) muscle glycogen c) blood glucose d) GTP
 pyruvate b) muscle glycogen c) blood glucose d) GTP") Chapter 34: Carbohydrate Metabolism Multiple Choice 1. The synthesis of glycogen from glucose is known as: a) glycogenolysis b) gluconeogenesis c) glycogenesis d) the Embden-Myerhof pathway 2. When a muscle
Chapter 34: Carbohydrate Metabolism Multiple Choice 1. The synthesis of glycogen from glucose is known as: a) glycogenolysis b) gluconeogenesis c) glycogenesis d) the Embden-Myerhof pathway 2. When a muscle
BY: RASAQ NURUDEEN OLAJIDE
 BY: RASAQ NURUDEEN OLAJIDE LECTURE CONTENT INTRODUCTION CITRIC ACID CYCLE (T.C.A) PRODUCTION OF ACETYL CoA REACTIONS OF THE CITIRC ACID CYCLE THE AMPHIBOLIC NATURE OF THE T.C.A CYCLE THE GLYOXYLATE CYCLE
BY: RASAQ NURUDEEN OLAJIDE LECTURE CONTENT INTRODUCTION CITRIC ACID CYCLE (T.C.A) PRODUCTION OF ACETYL CoA REACTIONS OF THE CITIRC ACID CYCLE THE AMPHIBOLIC NATURE OF THE T.C.A CYCLE THE GLYOXYLATE CYCLE
Chemistry 3503 Final exam April 17, Student s name:
 Chemistry 3503 Final exam April 17, 2008 Student s name: THIS EXAM IS FOR STUDENTS IN D. CRAIG S SECTION. IF YOU ARE IN M. EZE S SECTION THIS EXAM IS NOT FOR YOU. Part I /40 Part II Question 1 /4 Question
Chemistry 3503 Final exam April 17, 2008 Student s name: THIS EXAM IS FOR STUDENTS IN D. CRAIG S SECTION. IF YOU ARE IN M. EZE S SECTION THIS EXAM IS NOT FOR YOU. Part I /40 Part II Question 1 /4 Question
Metabolism of Nucleotides
 Metabolism of Nucleotides Outline Nucleotide degradation Components of Nucleobases Purine and pyrimidine biosynthesis Hyperuricemia Sources Nucleotide degradation The nucleotides are among the most complex
Metabolism of Nucleotides Outline Nucleotide degradation Components of Nucleobases Purine and pyrimidine biosynthesis Hyperuricemia Sources Nucleotide degradation The nucleotides are among the most complex
Oxidation of Long Chain Fatty Acids
 Oxidation of Long Chain Fatty Acids Dr NC Bird Oxidation of long chain fatty acids is the primary source of energy supply in man and animals. Hibernating animals utilise fat stores to maintain body heat,
Oxidation of Long Chain Fatty Acids Dr NC Bird Oxidation of long chain fatty acids is the primary source of energy supply in man and animals. Hibernating animals utilise fat stores to maintain body heat,
Module No. # 01 Lecture No. # 19 TCA Cycle
 Biochemical Engineering Prof. Dr. Rintu Banerjee Department of Agricultural and Food Engineering Asst. Prof. Dr. Saikat Chakraborty Department of Chemical Engineering Indian Institute of Technology, Kharagpur
Biochemical Engineering Prof. Dr. Rintu Banerjee Department of Agricultural and Food Engineering Asst. Prof. Dr. Saikat Chakraborty Department of Chemical Engineering Indian Institute of Technology, Kharagpur
استاذ الكيمياءالحيوية
 قسم الكيمياء الحيوية د.دولت على سالمه استاذ الكيمياءالحيوية ٢٠١٥-٢٠١٤ الرمز الكودي : ٥١٢ المحاضرة األولى ١ Content : Definition of proteins Definition of amino acids Definition of peptide bond General
قسم الكيمياء الحيوية د.دولت على سالمه استاذ الكيمياءالحيوية ٢٠١٥-٢٠١٤ الرمز الكودي : ٥١٢ المحاضرة األولى ١ Content : Definition of proteins Definition of amino acids Definition of peptide bond General
MBB 115:511 and 694:407 Final Exam Niederman/Deis
 MBB 115:511 and 694:407 Final Exam Niederman/Deis Tue. Dec. 19, 2006 Name Row Letter Seat Number This exam consists of two parts. Part I is multiple choice. Each of these 25 questions is worth two points.
MBB 115:511 and 694:407 Final Exam Niederman/Deis Tue. Dec. 19, 2006 Name Row Letter Seat Number This exam consists of two parts. Part I is multiple choice. Each of these 25 questions is worth two points.
Discussion of Prism modules and predicted interactions (Fig. 4)
") SUPPLEMENTARY NOTES Discussion of Prism modules and predicted interactions (Fig. 4) a. Interactions of the TCA-cycle, respiratory chain, and ATP synthetase with the amino acid biosynthesis modules. Given
SUPPLEMENTARY NOTES Discussion of Prism modules and predicted interactions (Fig. 4) a. Interactions of the TCA-cycle, respiratory chain, and ATP synthetase with the amino acid biosynthesis modules. Given
Biological oxidation II. The Cytric acid cycle
 Biological oxidation II The Cytric acid cycle Outline The Cytric acid cycle (TCA tricarboxylic acid) Central role of Acetyl-CoA Regulation of the TCA cycle Anaplerotic reactions The Glyoxylate cycle Localization
Biological oxidation II The Cytric acid cycle Outline The Cytric acid cycle (TCA tricarboxylic acid) Central role of Acetyl-CoA Regulation of the TCA cycle Anaplerotic reactions The Glyoxylate cycle Localization
PROTEIN METABOLISM: NITROGEN CYCLE; DIGESTION OF PROTEINS. Red meat is an important dietary source of protein nitrogen
 PROTEIN METABOLISM: NITROGEN CYCLE; DIGESTION OF PROTEINS Red meat is an important dietary source of protein nitrogen The Nitrogen Cycle and Nitrogen Fixation Nitrogen is needed for amino acids, nucleotides,
PROTEIN METABOLISM: NITROGEN CYCLE; DIGESTION OF PROTEINS Red meat is an important dietary source of protein nitrogen The Nitrogen Cycle and Nitrogen Fixation Nitrogen is needed for amino acids, nucleotides,
Respiration. Respiration. Respiration. How Cells Harvest Energy. Chapter 7
 How Cells Harvest Energy Chapter 7 Organisms can be classified based on how they obtain energy: autotrophs: are able to produce their own organic molecules through photosynthesis heterotrophs: live on
How Cells Harvest Energy Chapter 7 Organisms can be classified based on how they obtain energy: autotrophs: are able to produce their own organic molecules through photosynthesis heterotrophs: live on