The S2 subunit of infectious bronchitis virus Beaudette is a
|
|
|
- Reynold Randall
- 5 years ago
- Views:
Transcription
1 JVI Accepted Manuscript Posted Online 18 July 2018 J. Virol. doi: /jvi Copyright 2018 Bickerton et al. This is an open-access article distributed under the terms of the Creative Commons Attribution 4.0 International license. 1 2 The S2 subunit of infectious bronchitis virus Beaudette is a determinant of cellular tropism Erica Bickerton #, Helena J. Maier, Phoebe Stevenson-Leggett, Maria Armesto and Paul Britton The Pirbright Institute, Surrey, United Kingdom. Running Head: S2 subunit determines IBV tropism # Address correspondence to Erica Bickerton, erica.bickerton@pirbright.ac.uk. 1
2 Abstract The spike glycoprotein (S) of the avian gammacoronavirus, infectious bronchitis virus (IBV), is comprised of two subunits, S1 and S2, and has a role in virulence in vivo as well as 17 being responsible for cellular tropism in vitro. We have previously demonstrated that replacement of the S glycoprotein ectodomain from the avirulent Beaudette strain of IBV with the corresponding region from the virulent M41-CK strain resulted in a recombinant virus, BeauR-M41(S), with the in vitro cell tropism of M41-CK. The IBV Beaudette strain is able to replicate in both primary chick kidney cells and Vero cells, whereas the IBV M41-CK strain replicates in primary cells only. In order to investigate the region of the IBV S responsible for growth in Vero cells we have generated a series of recombinant IBVs expressing chimaeric S glycoproteins, consisting of regions from the Beaudette and M41-CK S gene sequences, within the genomic background of Beaudette. The S2, but not the S1, subunit of the Beaudette S was found to confer the ability to grow in Vero cells. Various combinations of Beaudette-specific amino acids were introduced into the S2 subunit of M41 to determine the minimum requirement to confer tropism for growth in Vero cells. The ability for IBV to grow and produce infectious progeny virus in Vero cells was subsequently narrowed down to just three amino acids surrounding the S2ʹ cleavage site. Conversely, swapping the three Beaudette-associated amino acids with corresponding ones from M41 was sufficient to abolish Beaudette growth in Vero cells. Importance Infectious Bronchitis remains a major problem in the global poultry industry, despite the existence of many different vaccines. IBV vaccines, both live attenuated and inactivated, are currently grown on embryonated hen s eggs, a cumbersome and expensive process due to the fact that most IBV strains do not grow in cultured cells. The reverse genetics system for IBV 2
3 creates the opportunity for generating rationally designed and more effective vaccines. The observation that IBV Beaudette has the additional tropism for growth on Vero cells also invokes the possibility of generating IBV vaccines produced from cultured cells rather than the use of embryonated eggs. The regions of the IBV Beaudette S glycoprotein involved in the determination of extended cellular tropism have been identified in this study, which will enable the rational design of a future generation of IBV vaccines that may be grown on Vero cells. Introduction The avian coronavirus infectious bronchitis virus (IBV), is a gammacoronavirus in the order Nidovirales (1), and the aetiological agent of the disease infectious bronchitis (IB) that affects domestic fowl (2-5). IBV replicates primarily in the respiratory tract (6, 7), causing a highly contagious respiratory disease characterised by nasal discharge, snicking, rales and tracheal ciliostasis in chickens (8), but also in many other epithelial surfaces including enteric surfaces (9), oviducts and kidneys (10-12). Coronaviruses are enveloped viruses with an unsegmented, single-stranded positivesense RNA genome of 26 32kb, which is capped and polyadenylated that replicates in the cell cytoplasm (13-15). The genomic RNA associates with the nucleoprotein (N) forming helical nucleocapsids that are enclosed within lipid envelopes containing the spike (S) glycoprotein, membrane (M) protein and small envelope (E) protein. The coronavirus S glycoprotein is a highly-glycosylated type I membrane glycoprotein that is synthesised as a single polypeptide chain of about 180 kda that oligomerises into homotrimers in the endoplasmic reticulum and is processed in the Golgi apparatus of the host cell (16-18) observable as 20 nm structures projecting from the virion surface by electron microscopy. Cryo-electron microscopy has revealed that the S protein forms a clove-shaped trimer of S1 3
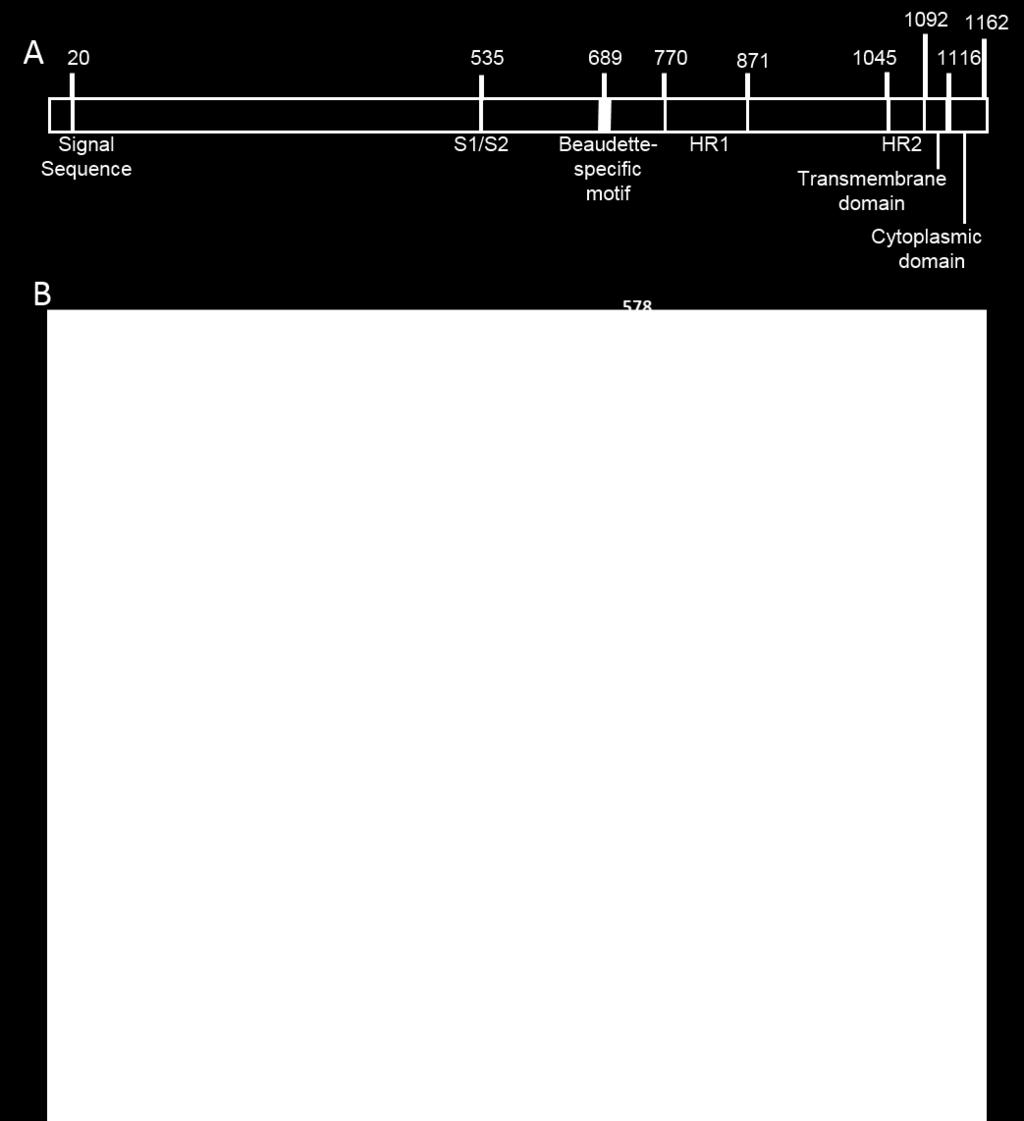
4 subunits linked to a stalk of trimeric S2 subunits (19-21). The S protein is responsible for binding to the target cell receptor and fusion of the viral and cellular membranes, fulfilling a major role in the infection of susceptible cells (22). Coronavirus S glycoproteins consist of four domains; an N-terminal signal sequence that is cleaved during synthesis; the ectodomain, which is present on the outside of the virus particle; the transmembrane region responsible for anchoring the S protein into the lipid bilayer of the virus particle; and the cysteine-rich cytoplasmic tail at the C-terminus. The S glycoprotein of some coronaviruses including IBV, mouse hepatitis virus (MHV) and human coronavirus OC43 (HCoV-OC43), are cleaved during biosynthesis into two subunits, S1 and S2, by a furin-like protease in the Golgi apparatus, which remain non-covalently linked (23). The IBV S glycoprotein (1162 amino acids) is composed of two subunits, S1 (535 amino acids, 90-kDa) comprising the N-terminal half of the S protein and S2 (627 amino acids, 84- kda) comprising the C-terminal half of the S protein. As with other coronaviruses the S2 subunit contains the transmembrane and C-terminal cytoplasmic tail domains and the S1 subunit contains the receptor binding domain (RBD) of the S protein (24-27). Proteolytic activation and binding of the S1 subunit to the host cell receptor induce conformational changes in the spike (28, 29) (30), leading to virus-cell fusion and release of the nucleocapsid into the cytoplasm (31). The ectodomain region of the S2 subunit contains a fusion peptide (32-39) and two heptad repeat regions involved in oligomerisation of the S protein (40) and is required for entry into susceptible cells (41-43). The spike interacts with the M protein on pre-golgi membranes forming multimeric complexes (44) and is incorporated into forming virions (45). The coronavirus RBD is an independently folded region of the S glycoprotein responsible for interacting with the virus receptor on susceptible host cells. Although all coronavirus RBDs are found within the S1 subunit, they are virus-specific and vary in 4
5 position throughout the S1 domain (46-48). The location of the RBD of IBV has been mapped within the N-terminal 253 amino acids of the S1 subunit (49). The host-cell receptors for several coronaviruses have been identified; several of the alphacoronaviruses, porcine transmissible gastroenteritis virus (TGEV), type II feline coronavirus (FCoV), canine coronavirus (CCoV) and human coronavirus-229e (HCoV-229E) use aminopeptidase N as host cell receptor (50-52). In contrast, betacoronaviruses have been found to utilise a variety of different receptors. The principal receptor for MHV is murine carcinoembryonic antigenrelated cell adhesion molecule (CEACAM; (53)); the virus also attaches to O-acetylated sialic acid, which is mediated by an extra structural protein, the haemagglutinin-esterase (HE) protein found in the virus membrane of some coronaviruses in this genus, rather than the S glycoprotein (54). Severe acute respiratory syndrome coronavirus (SARS-CoV) has been shown to use angiotensin-converting enzyme 2 (ACE2) as a receptor (55) along with CD209L (56) and is also thought to use the C-type lectins DC-SIGN and L-SIGN as alternative receptors independent of ACE2 (57). The receptor for the Middle East respiratory syndrome coronavirus (MERS-CoV) has been identified as dipeptidyl peptidase 4, also known as CD26 (58). The protein receptor for IBV has not yet been elucidated, although two strains of IBV, M41 and Beaudette-US, have been shown to use α2,3-linked sialic acid as an attachment factor (59, 60), and for subsequent infection of the tracheal epithelium (61, 62). The interaction between IBV and sialic acid has been confirmed by Madu et al. (63), who also identified a heparan sulfate (HS) binding site between amino acid residues of the S2 subunit of the Beaudette S glycoprotein and showed that HS may be involved as a cofactor in Beaudette virus entry into host cells. Yamada and Liu identified an additional furin cleavage motif within the putative HS binding site identified by Madu et al. (64), with a role in viral entry and syncytium formation in vitro. Cleavage was mapped to arginine residue 690. Susceptibility of cultured cells to infection with Beaudette was subsequently found to 5
6 correlate with cellular furin expression levels (65). This additional proteolytic cleavage site within the S2 subunit, named S2ʹ, has since been identified in SARS-CoV, MERS-CoV and HCoV-229E, and found to mediate membrane fusion and viral infectivity (66-69). Coronaviruses generally exhibit restricted cell and tissue tropism, which is dependent upon the S glycoprotein of individual coronavirus strains (70, 71). However, some strains of IBV, following adaptation, are able to replicate in primary chicken cells such as chick kidney (CK) cells. Interestingly, the Beaudette strain, after several hundred passages in embryonated eggs, was also discovered to have an extended host range with the ability to replicate in a mammalian cell line, Vero cells (72) and to a limited extent in baby hamster kidney (BHK- 21) cell lines (73). We have previously demonstrated that the cellular tropism of IBV is determined by the S glycoprotein; replacement of the ectodomain of the IBV Beaudette S glycoprotein with the corresponding region from the pathogenic IBV M41-CK resulted in a ribv, BeauR-M41(S), which had the tissue tropism associated with M41-CK (74). The present study aims to elucidate the specific regions of the IBV Beaudette S glycoprotein involved in the determination of extended cellular tropism thus enabling the rational design and generation of IBV vaccines that may be grown on Vero cells. Materials and Methods Cells and Viruses The IBV strains used were: (1) M41-CK derived from the pathogenic M41 strain (75), after multiple passages in CK cells (8, 76, 77). This isolate is able to produce infectious virus in CK but not Vero cells; (2) Beau-R (78), a molecular clone of CK cell-adapted Beaudette (79), Beaudette-CK (80). Beau-R is able to produce infectious virus on both CK and Vero cells; and (3) BeauR-M41(S) in which the ectodomain of the Beau-R S gene was replaced 6
7 with the corresponding sequence of the M41-CK S glycoprotein (74). This ribv based on the genome of Beaudette, but with the S glycoprotein ectodomain from M41-CK, has the same tropism as M41-CK and is therefore able to produce infectious virus on CK but not Vero cells. IBV strains were propagated and titrated in CK cells as described previously (81-83). Growth curves and confocal microscopy of IBV infection were carried out using CK and Vero cells (74). Vaccinia viruses (VV) were propagated in Vero cells and large stocks for DNA extraction were prepared in BHK-21 cells as described previously (84, 85). Although Beaudette and M41 viruses belong to the same serogroup, Massachusetts, it is important to note that the two viruses are not genetically related, in the sense that one was derived from the other. Comparison of their nucleotide and amino acid sequences show many differences between the two viruses. Unfortunately, there are instances in the literature where it is implied that Beaudette was derived from M41 (63, 86), leading to the incorrect conclusion that mutations arising in M41 may have led to the ability of Beaudette to produce infectious progeny in Vero cells. This may have arisen from IBV Beaudette being referred to as IBV-42 (sometimes abbreviated to M42) and M41 as IBV-41, respectively (72, 87, 88). IBV Beaudette was isolated in the 1930s (79) and M41 in the 1940s (75). IBV Beaudette is able to grow and produce infectious virus in CK cells and can be also be adapted to Vero cells by repeat passage, as observed by syncytia formation (72). Beau-R, the molecular clone of Beaudette CK, does not produce syncytia when grown on Vero cells (78), however, repeated passage of Beau-R on Vero cells resulted in adaption, as previously observed for Beaudette CK, for syncytia formation on Vero cells (Bickerton PhD Thesis). Construction of chimaeric IBV S genes The S1 and S2 subunits of Beau-R and M41-CK (GenBank Accession numbers AJ and X04722 respectively) were amplified by PCR from two plasmids, pgpt- 7
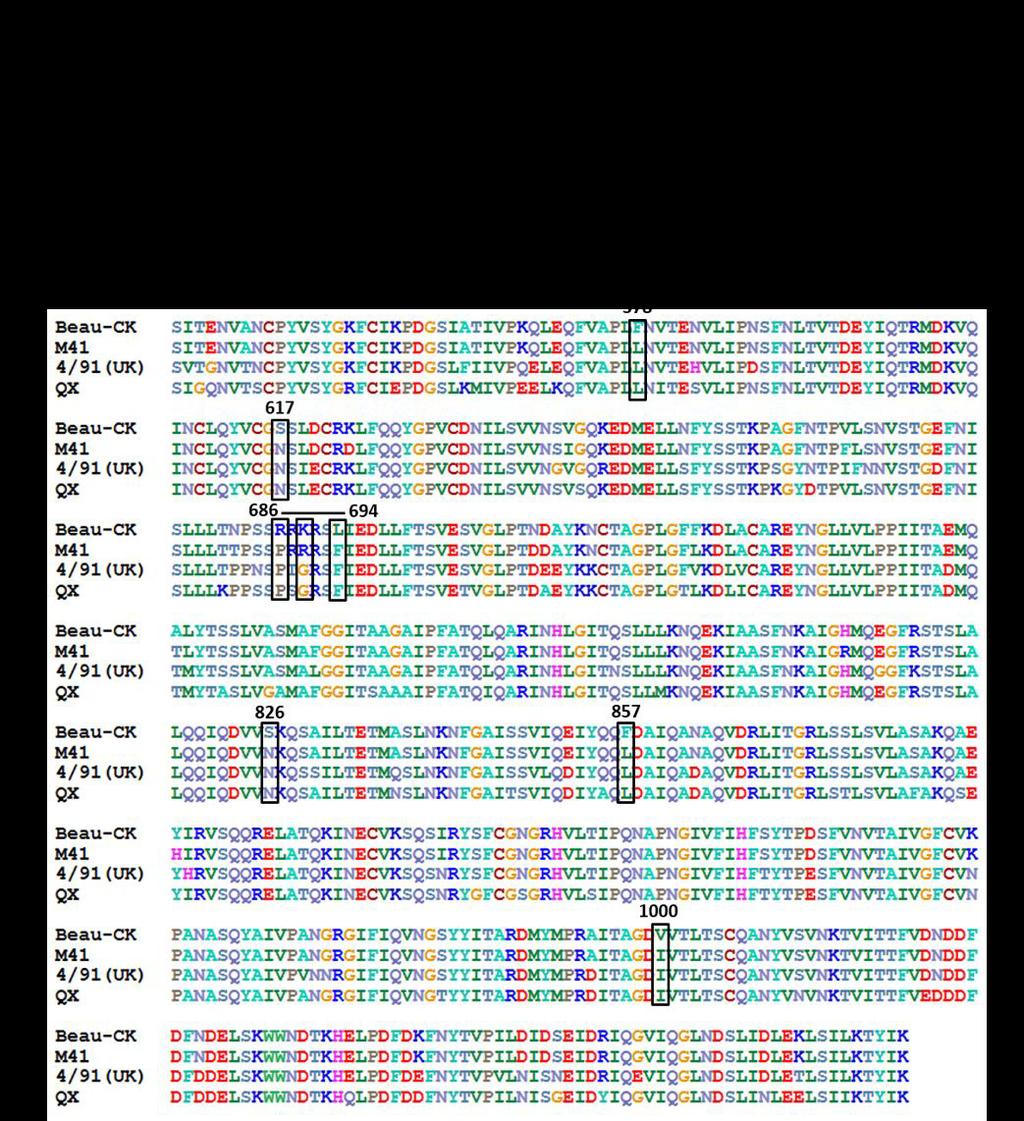
8 M41S and pgpt-ibv-stui-bamhi (84), using primers located within the replicase gene (replicase forward GTGTACCGCATAACATGCGA ), across the S1/S2 junction (S1/S2 forward ATCACTAATGGAACACGTCGTTTTAGACGTTCTATTACTG ) and S1/S2 reverse ( CAGTAATAGAACGTCTAAAACGACGTGTTCCATTAGTAT ) and within gene 3 (BG-51 reverse CTCGTTAACAATAACTGC ). Overlapping PCR was used to combine the subunits, forming the chimaeric S genes, which were then inserted into NsiI and BspEI digested pgpt-ibv-stui-bamhi to create the plasmids pgpt-beaur-s1-m41-s2 and pgpt-m41-s1-beaur-s2. The IBV replicase gene and gene 3 sequences, 5 and 3 to the S gene sequences, respectively, were derived from Beau-R. Sequences corresponding to 715 bp cdnas of the Beaudette and M41 S2 regions were synthesised and cloned into pgpt-neb193 (84) by GeneArt, Thermo Fisher Scientific. pgpt-beaur-s-mm, based on the Beau-R S glycoprotein in which the Beaudette-specific motif (BSM), 686 SRRKRSLIE 694, located within the S2 subunit was replaced with the corresponding sequence, M41 motif (MM), 686 SPRRRSFIE 694 from M41-CK. pgpt-m41-s- BSM, contained a cdna based on the M41-CK S glycoprotein in which the MM sequence, 686SPRRRSFIE 694, was replaced with the BSM 686 SRRKRSLIE 694 motif. Plasmids pgpt- M41-S2-L 578 F-N 617 S and pgpt-m41-s2-n 826 S-L 857 F-I 1000 V, were based on M41-CK S cdna sequences of 747 bp and 1162 bp, respectively, but with nucleotides representing Beaudette-specific amino acids L 578 F and N 617 S in pgpt-m41-s2-l 578 F-N 617 S, and N 826 S, L 857 F and I 1000 V in pgpt-m41-s2-n 826 S-L 857 F-I 1000 V. PCR products derived from the plasmids, the S gene sequence and adjoining regions of the Beaudette genome were sequenced to authenticate the modified sequences. 8
9 Generation of recombinant vaccinia viruses containing modified IBV cdna and recovery of infectious IBV The IBV cdnas contained within plasmids pgpt-beaur-s1-m41-s2, pgpt-m41-s1- BeauR-S2, pgpt-m41-s-bsm, pgpt-beaur-s-mm, pgpt-m41-s2-l 578 F-N 617 S and pgpt-m41-s2-n 826 S-L 857 F-I 1000 V were introduced into the Beau-R cdna within the genome of four different recombinant vaccinia viruses (rvvs) by homologous recombination using the transient dominant selection system (TDS) as previously described (84, 89). The rvvs used were: (1) VV-BeauR-ΔS, containing the full-length IBV Beau-R cdna genome minus the S gene (84) used with pgpt-beaur-s1-m41-s2 and pgpt-m41-s1-beaur-s2; (2) vnoti/ibv FL, containing the full-length IBV Beau-R cdna genome (78) used with pgpt-beaur-s-mm; (3) vnoti/ibv FL -M41S, containing the full-length IBV Beau-R cdna genome with the S gene from M41 (84) used with pgpt-m41-s-bsm to generate (4) VV- M41-S-BSM, containing the full-length IBV Beau-R cdna genome but with the S gene from M41 containing the BSM, which was subsequently used with pgpt-m41-s2-l 578 F-N 617 S and pgpt-m41-s2-n 826 S-L 857 F-I 1000 V. All recombinant IBVs were rescued from plaque-purified clones of rvvs containing the IBV cdna genome. The rvvs were sequenced to verify the correct S gene was present prior to rescue of ribvs, which were passaged three times in CK cells prior to experimental use (84, 89). Assessment of ribv growth in CK and Vero cells Confluent monolayers of CK and Vero cells in six-well plates were infected with ribv and observed for cytopathic effect (CPE) formation by bright-field microscopy using a Leica DM IRB inverted light microscope. CK and Vero cells were grown on 13 mm coverslips in 24-well plates to approximately 50% confluency and infected with 150 µl of ribv at 1x10 5 pfu/ml in duplicate. Infected cells were fixed 24 h post-infection with 4% paraformaldehyde 9
10 in phosphate buffered saline (PBS) and permeabilised using 0.5% Triton-X100. IBV infected cells were identified by incubation with a 1:400 dilution of mouse anti-dsrna J2 IgG2a monoclonal antibody (English and Scientific Consulting Bt.) and detected with Alexa Fluor conjugated goat anti-mouse IgG antibody (Invitrogen) diluted 1:200. Nuclei were stained with 4',6-diamidino-2-phenylindole (DAPI, Invitrogen) Immunolabelled cells were imaged using a Leica TCS SP5 DM6000 confocal microscope using the Leica Microsystems LAS AF software. Growth kinetics of ribv growth in CK and Vero cells Confluent monolayers of Vero and CK cells in 6-well plates were infected with ribv in triplicate at 0.5, 0.05 or multiplicity of infection (MOI) for 1 h at 37 C in 5% CO 2, washed twice with PBS and 3 ml of fresh 1 x BES medium added. Cell supernatants were harvested at 1, 12, 24, 48 and 72 h post-infection and assayed for progeny virus by plaque assay using CK cells. Sequence analysis The plasmid-derived and IBV-derived RT-PCR and PCR products were sequenced using a variety of oligonucleotides, derived from the Beau-CK sequence (90). Assembly of the sequences was performed using Gap4 of the Staden Sequence Software Programs (91). IBV nucleotide and amino acid positions are based on Beau-CK sequence (GenBank accession number AJ311317). Western blot analysis of ribv spike glycoprotein proteolytic cleavage CK cells were mock infected or infected with the indicated viruses and at 24 hours post infection, cells were scraped into the culture medium and pelleted. Cell pellets were washed 10
11 once with cold PBS and then lysed in RIPA buffer (150 mm NaCl, 50 mm Tris (ph 7.4), 1% Igepal, 0.25% sodium deoxycholate, 1 mm sodium orthovanadate and 1x protease inhibitor cocktail) on ice for 20 minutes. Cell lysates were mixed with 4x SB (Biorad) to give 1x final concentration and denatured at 95 oc for 3 minutes. Proteins were separated by SDS-PAGE and transferred onto nitrocellulose membrane. After blocking overnight in 5% milk in PBS-T, the membrane was labelled with anti-s2 (26.1, Thermo Fisher) and anti-beta actin (Abcam) followed by IRDye 680 and 800 labelled secondary antibodies (Li-Cor). Blots were imaged using an Odyssey CLx (Li-Cor). Results Generation of ribvs expressing S genes with chimaeric subunits. Recombinant IBVs, expressing chimaeric S proteins composed of either the S1 subunit derived from M41-CK and the S2 subunit from Beaudette CK, BeauR-M41(S1), or the S1 subunit from Beaudette CK and the S2 subunit from M41-CK, BeauR-M41(S2), were produced (Figure 1) to determine which subunit of the S glycoprotein was responsible for the extended tropism of Beaudette CK for Vero cells. Sequence analysis following rescue of the ribvs identified the presence of three nucleotide substitutions in BeauR-M41(S1) P 3 -CK at positions , GTT to AAC, corresponding to two adjacent codons, amino acids L 624 -F 625, resulting in one amino acid substitution, F 625 T, within the Beaudette S2 subunit (Table 1). One nucleotide substitution, C T, was identified in BeauR-M41(S2) P 3 -CK, resulting in amino acid substitution A 452 T within the Beaudette S1 subunit. These substitutions could have arisen during rescue or passage of the virus on CK cells as they were not present in the parental rvv. Expression of chimaeric S genes does not alter growth on primary chicken cells. 11
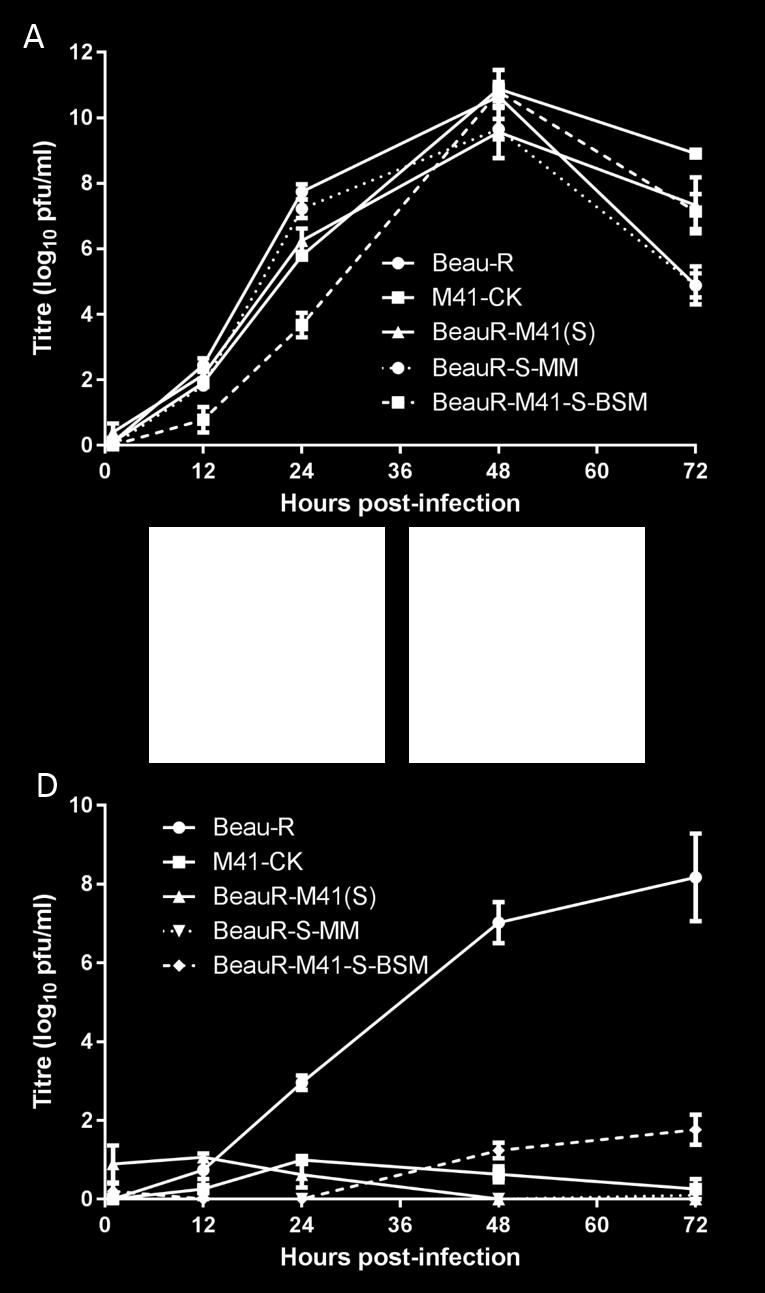
12 Analysis of the growth kinetics of the ribvs, BeauR-M41(S1) P 3 -CK and BeauR- M41(S2) P 3 -CK, in primary chicken kidney (CK) cells identified peak titres equivalent to those of parental viruses, Beau-R, M41-CK and BeauR-M41(S), at 48h post-infection. Interestingly, the growth of the ribvs was initially slower over the first 24h (Figure 2A). Immunofluorescence analysis of IBV infected CK cells showed that the ribvs were able to infect and spread to neighbouring cells, as observed for the parental viruses (data not shown). The S2 subunit of Beaudette is responsible for Vero cell tropism. Immunofluorescence analysis showed that Vero cells could be infected with Beau-R or BeauR-M41(S1) P 3 -CK (Figure 2D and G). However, only a small number of individual Vero cells were infected with M41-CK or BeauR-M41(S) (Figure 2E and F). Confirming previous results that infectious progeny virus does not infect neighbouring cells and there is no virus spread (74); indicating that either progeny virus is not released from infected Vero cells or is not infectious. Although ribv BeauR-M41(S2) P 3 -CK formed infectious centres on CK cells, the ribv had a similar phenotype on Vero cells as observed with M41-CK and BeauR-M41(S) (Figure 2H). Analysis of the growth kinetics of the ribvs on Vero cells confirmed the immunofluorescence studies. BeauR-M41(S1) P 3 -CK demonstrated a similar growth to Beau-R in Vero cells, albeit with a lower peak titre (Figure 2B). M41-CK, BeauR-M41(S) and BeauR-M41(S2) P 3 -CK did not reach titres greater than 1.5 log 10 pfu/ml in Vero cells. Our results show that BeauR-M41(S1) P 3 -CK and Beau-R produce infectious progeny in both CK and Vero cells, whereas BeauR-M41(S2) P 3 -CK resembled the tropism of M41-CK and 285 BeauR-M41(S) for growth in Vero cells. Overall, replacement of the Beaudette S1 subunit with the corresponding S1 sequence from M41-CK did not affect the ability of the ribv to replicate and produce progeny virus in Vero cells. In contrast, replacement of the Beaudette 12
13 S2 subunit with the M41 S2 sequence resulted in loss of the ability of Beau-R to grow in Vero cells. The ability of Beau-R and BeauR-M41(S1) P3-CK to grow on Vero cells resides within the Beaudette S2 subunit and not within the S1 subunit known to contain the RBD Adaptation of ribvs to growth on Vero cells. The P 3 -CK ribvs were passaged three times on Vero cells to determine whether virus replication could be maintained by passage on this mammalian cell line. Passage of BeauR- M41(S1) resulted in CPE from 24 h post-infection, whereas passage of BeauR-M41(S2) did not result in an observable CPE. Additional passages of ribv BeauR-M41(S1) resulted in syncytia formation from P 5., indicating that the virus, as previously observed for Beaudette, had become adapted for growth on Vero cells (data not shown). Analysis of the growth kinetics of BeauR-M41(S1) P 3 -CK passaged on Vero cells showed an increase in titre following passage, to within one log 10 of the titres observed for Beau-R. Overall the growth of the P 7 -Vero isolate matched growth kinetics observed for Beau-R (Figure 3). Sequence analysis of the S gene from BeauR-M41(S1) P 7 -Vero identified one nucleotide difference, located within the S2 subunit, from the parental viruses, Beau-R and BeauR- M41(S). The substitution occurred at nucleotide position C T, resulting in an amino acid change of T 1004 I (Table 1). This change may contribute to the further adaptation of BeauR- M41(S1) for growth in Vero cells and may be involved in syncytium formation. Generation of ribvs with modified S2 subunit Beaudette-specific motifs. Once it was established that the S2 subunit is responsible for the extended tropism of Beaudette in Vero cells, the amino acid sequence of the Beaudette S2 was compared to those in other IBV strains to identify potential amino acids, unique to the Beaudette S2, which may 13
14 313 play a role in the tropism for growth in Vero cells. The positions of the amino acid 314 differences between Beaudette, M41 and two pathogenic field strains of IBV, QX and 4/91, 315 are shown in Figure 1B. A Beaudette-specific motif (BSM), 686 SRRKRSLIE 694, was identified in the Beaudette S2 sequence surrounding the S2ʹ cleavage site at arginine residue 690, which was not found in the S glycoprotein of any other IBV strain; the Beaudettespecific amino acids within the BSM are underlined. In order to determine whether this motif plays a role in the Vero tropism of IBV Beaudette, two full length IBV cdnas were generated; BeauR-S-MM that has the Beau-R genomic background (78) in which the BSM in the S2 subunit was replaced with nucleotides encoding the corresponding M41 motif sequence, 686SPRRRSFIE 694 (Figure 1B), and BeauR-M41-S-BSM which was based on BeauR-M41(S) (74), in which the MM in the M41 S2 subunit was replaced with the BSM (Figure 1B). Following rescue and growth in CK cells sequence analysis identified the presence of one nucleotide substitution in BeauR-M41-S-BSM P 3 -CK, A C, resulting in an amino acid substitution from D 620 A in the S2 subunit (Table 1), which could have arisen during rescue or passage of the virus on CK cells as the substitution was not present in the rvv. No mutations were identified in the BeauR-S-MM P 3 -CK S gene. Analysis of the growth kinetics BeauR-S-MM P 3 -CK and BeauR-M41-S-BSM P 3 -CK in CK cells showed they displayed peak titres equivalent to those of parental viruses, Beau-R, M41-CK and BeauR-M41(S), at 48h post-infection, although growth of BeauR-M41-S-BSM was slower over the first 24h (Figure 4A). Immunofluorescence analysis of infected CK cells confirmed both viruses were able to infect neighbouring cells (data not shown). The Beaudette-specific motif in the S2 subunit is sufficient to confer the tropism for growth in Vero cells. 14
15 Immunofluorescence analysis of infected Vero cells infected with BeauR-M41-S-BSM P 3 -CK (Figure 4B), demonstrated that the introduction of the BSM into the M41 S glycoprotein resulted in the ability of a ribv, expressing the modified M41 S glycoprotein, to grow on Vero cells. Conversely, ribv BeauR-S-MM P 3 -CK, in which only the BSM in the Beaudette S2 subunit was replaced with the corresponding MM sequence from M41, lost its ability to grow in Vero cells (Figure 4C), with a phenotype observed for M41 and BeauR- M41(S). This observation was confirmed by analysis of the growth kinetics of the ribvs in Vero cells (Figure 4D). Production of progeny BeauR-S-MM P 3 -CK virus from Vero cells was undetectable. Although BeauR-M41-S-BSM P 3 -CK produced progeny virus in Vero cells the growth was considerably lower than those observed for Beau-R. Progeny BeauR-M41-S- BSM P 3 -CK virus was only observed from 24 h post-infection and reached a peak titre at 72 h post-infection that was around 6 log 10 lower than those of Beau-R. These results showed that loss of the BSM abolished the ability of Beau-R to grow on Vero cells and that the accruement of the BSM resulted in the ability of a ribv expressing the M41 S to have a tropism for growth on Vero cells, albeit to a lesser extent than replacement with the entire Beaudette S2. Overall, it appeared that either a complete Beaudette S2 subunit or other additional Beaudette S2-specific amino acids may be required for the growth characteristics observed with the complete Beaudette S2. The P 3 -CK viruses, BeauR-M41-S-BSM and BeauR-S-MM, were passaged on Vero cells to determine whether virus replication could be maintained. BeauR-S-MM isolates were unable to maintain replication on Vero cells for three passages as assessed by RT-PCR. The isolates caused no CPE, and no infected cells were observed on either CK or Vero cells (data not shown). The ribv BeauR-M41-S-BSM isolates were successfully passaged on Vero cells and the growth kinetics of one of the BeauR-M41-S-BSM P 7 -Vero passaged isolates 15
16 analysed. The virus had a very similar growth pattern within one log 10 of the growth of Beau-R on Vero cells (Figure 5). No additional mutations were identified in the BeauR- M41-S-BSM P 7 -Vero S gene, other than the nucleotide substitution identified at P 3 -CK; A C, resulting in an amino acid substitution from D 620 A in the S2 subunit. The genetic stability of BeauR-M41-S-BSM observed over seven passages on Vero cells indicates that the spike glycoprotein is not under pressure to mutate in these conditions. Generation of ribvs with additional Beaudette-specific amino acid modifications within the M41-CK S2 subunit. Once it was established that the BSM within the S2 subunit is able to confer the extended tropism of IBV in cell culture, other Beaudette-specific amino acids, identified in the Beaudette S2 subunit in comparison to other IBV sequences (Figure 1B), were introduced into the M41-CK S2 subunit in addition to the BSM. In order to determine the specific amino acids and minimum number of amino acid changes required to fully confer tropism, full length IBV cdnas were generated with various combinations of the Beaudette-specific amino acids in the S2 subunit of M41 based on the BeauR-M41-S-BSM genomic background; L 578 F and N 617 S were introduced by pgpt-m41-s2-l 578 F-N 617 S, and N 826 S, L 857 F and I 1000 V were introduced by pgpt-m41-s2-n 826 S-L 857 F-I 1000 V. Following homologous recombination into the IBV cdna BeauR-M41-S-BSM in vaccinia virus a series of cdnas containing a variety of the Beaudette specific amino acids in the M41 S2 sequence were generated by random homologous recombination using the two pgpt plasmids. These full length IBV cdnas were used to rescue the corresponding ribvs; BeauR-M41-S-BSM-N 617 S, BeauR-M41-S-BSM-L 578 F-N 617 S, BeauR-M41-S-BSM-I 1000 V, BeauR-M41-S-BSM-L 857 F-I 1000 V, BeauR-M41-S-BSM-N 826 S-L 857 F-I 1000 V and BeauR-M41- S-BSM-L 578 F-N 617 S-N 826 S-L 857 F-I 1000 V. 16
17 Sequence analysis of the P 3 -CK ribvs showed the presence of one nucleotide substitution in BeauR-M41-S-BSM-L 578 F-N 617 S, A C, which resulted in an amino acid substitution from E 493 D, and a mixed population at C T/C in BeauR-M41-S-BSM-N 826 S- L 857 F-I 1000 V resulting in amino acid substitution H 43 Y (Table 1), which could have arisen during rescue or passage of the virus on CK cells as the substitutions were not present in the rvv. Analysis of the growth kinetics of the ribvs with the modified M41 S2 subunit on CK cells showed that most of the viruses, apart from BeauR-M41-S-BSM-L 578 F-N 617 S-N 826 S- L 857 F-I 1000 V, had growth patterns that were representative of ribv BeauR-M41(S) in that they produced less progeny virus than Beau-R in the first 24 h post-infection but reached or exceeded the titre of Beau-R over the next 48 h post-infection (Figure 6A). Interestingly, ribv BeauR-M41-S-BSM-L 578 F-N 617 S-N 826 S-L 857 F-I 1000 V had a similar growth pattern as Beau-R in that it had a peak titre at 24 post-infection, which decreased over the 48 hours post-infection, the peak titre was about 1.2 log 10 less than Beau-R at 24 hours post-infection. All the ribvs had a similar growth pattern in CK cells by immunofluorescence analysis of the infected cells (data not shown). Assessment of tropism Analysis of the growth kinetics of the P 3 -CK ribvs on Vero cells showed that they all grew with similar patterns to Beau-R, but with lower peak titres and lower titres throughout infection, whereas BeauR-M41(S) expressing the donor S protein did not grow in Vero cells (Figure 6B). Interestingly, BeauR-M41-S-BSM-L 578 F-N 617 S-N 826 S-L 857 F-I 1000 V P 3 -CK, which contains the most Beaudette-specific amino acids inserted into the M41 S2 subunit, grew the most similar to Beau-R, indicating that only eight Beaudette-derived amino acids are required to confer ability to grow on Vero cells with similar growth kinetics and 17
18 phenotype as Beau-R in Vero cells. The other ribvs with modified M41-CK S2 subunits replicated to a similar titre as BeauR-M41-S-BSM P 3 -CK, reaching a peak titre at 72 h postinfection. Immunofluorescence analysis of infected Vero cells showed that the P 3 -CK ribvs containing the additional Beaudette-specific modifications within the M41-CK S2 subunit formed infectious centres on Vero cells (Figure 6C H). Only small foci of infected Vero cells at 24 hours post-infection were generated by some of these ribvs during the initial passage on Vero cells (Figure 6E and G). This is reflected in the delayed growth during the first 24 hours post-infection as shown in Figure 6B. As shown above passage of BeauR-M41-S-BSM P 3 -CK on Vero cells improved the growth of this virus in Vero cells, therefore the ribvs with the additional Beaudette-derived amino acids in the M41 S2 subunit were passaged on Vero cells and the P 7 -Vero isolates reexamined for growth on Vero cells. Analysis of the growth kinetics of the P 7 -Vero isolates on Vero cells clearly demonstrated that all the ribvs now replicated with similar patterns and peak titres equivalent to those observed for Beau-R (Figure 7). Interestingly, three of the P 7 - Vero passaged ribvs, BeauR-M41-S-BSM-N 617 S, BeauR-M41-S-BSM-L 578 F-N 617 S, and BeauR-M41-S-BSM-N 826 S-L 857 F-I 1000 V, had higher titres of progeny virus, almost 2 log 10 higher to 24 hours post-infection, than Beau-R, at 72 hours post-infection the peak titres were similar to Beau-R. Sequence analysis of the S genes from the P 7 -Vero isolates identified several nucleotide substitutions that may be involved in further conferment of tropism for Vero cells (Table 1); BeauR-M41-S-BSM-L 857 F-I 1000 V P 7 -Vero C T results in an amino acid substitution P 630 S, BeauR-M41-S-BSM-L 578 F-N 617 S-N 826 S-L 857 F-I 1000 V P 7 -Vero G A results in an amino acid substitution G 73 S and BeauR-M41-S-BSM-N 617 S, L 578 F-N 617 S and N 826 S-L 857 F-I 1000 V P 7 -Vero were all found to have a common substitution, A C, resulting in an amino acid 18
19 substitution Q 866 H in the M41 S2 subunit. BeauR-M41-S-BSM-N 826 S-L 857 F-I 1000 V P 7 -Vero had several other substitutions; G A results in V 448 I, C A results in P 659 T and a mixed population of C T corresponding to the first engineered amino acid modification in the M41 S2 subunit L 578 F. The ribv appears to be losing the L 857 F modification resulting in with a mixed population at C T. Interestingly, BeauR-M41-S-BSM-I 1000 V P 7 -Vero was found to gain the Beaudette-specific amino acid, L 578 F, which was introduced into the M41 S2 subunit in some of the ribvs. The ribvs containing the Beaudette-specific motif are proteolytically cleaved at the S2ʹ cleavage site As the Beaudette-specific motif surrounds the S2ʹ cleavage site, susceptibility of ribvs containing the BSM to proteolytic cleavage was investigated. CK cells were infected with BeauR-M41-S-BSM P 3 -CK, BeauR-S-MM P 3 -CK or parent viruses Beau-R, BeauR-M41(S) or M41-CK for 24 hours then lysed and proteolytic cleavage of the spike glycoprotein was analysed by Western blotting with anti-s2 antibody (Figure 8). A clear S2ʹ cleavage product was observed in the Beau-R sample, but this was absent in the BeauR-M41(S), M41-CK and BeauR-S-MM samples, which do not contain the BSM and are unable to grow in Vero cells. A faint band corresponding to the S2ʹ cleavage product was also observed in the BeauR-M41- S-BSM sample, indicating that proteolytic cleavage at the S2ʹ site within the BSM may play a role in extending the host tropism of Beau-R. The faint band generated by BeauR-M41-S- BSM in comparison with that produced by Beau-R may indicate that proteolytic processing is not as efficient in the recombinant spike glycoprotein. This could be due to subtle differences in the tertiary or quaternary structures of the BeauR-M41-S-BSM spike glycoprotein resulting in stearic hindrance of the protease acting at the S2ʹ site. The size of the S2ʹ cleavage product is smaller than expected when analysed by western blotting. It is possible 19
20 that the band we observe at around 30 kda may be a breakdown product of the S2ʹ cleavage product, or the size of the S2ʹ cleavage product may appear to be smaller than expected due to distortion of protein coils in the gel. Nevertheless, it is important to note that this band is only observed in samples infected with ribv containing the Beaudette-specific motif surrounding the S2ʹ cleavage site, indicating that this is where cleavage occurs, so its production may be involved in the extended tropism of Beau-R. Discussion Infectious bronchitis remains a major problem in the global poultry industry, despite the existence of many different vaccines. IBV vaccines, both live attenuated and inactivated, are currently grown in embryonated hen s eggs, due to the fact that most IBV strains do not grow in cultured cells. Production of IBV vaccines in embryonated eggs is expensive and cumbersome with each egg only producing a small volume of allantoic fluid from which the virus is isolated. Furthermore, the supply of embryonated eggs is not guaranteed to be reliable, which may seriously affect production of required vaccines. As demand for seasonal and pandemic influenza vaccines rises, the supply of embryonated eggs for the production of other vaccines may be reduced. Another concern about the use of embryonated eggs is the possible presence of adventitious viruses, which may compromise the vaccine stocks or cause pathology in the vaccinated chickens. In light of the numerous disadvantages of egg-based vaccine production, the ability to produce live vaccines in vitro would be beneficial to the vaccine industry as well as the poultry industry. Vaccine production in vitro is faster, more efficient, and able to produce larger volumes of vaccine than in ovo. This is particularly important given the very competitive vaccine price points in the poultry sector. This approach would also reduce the 20
21 number of embryonated hen s eggs utilised, an important consideration under the principles of the 3Rs. Coronavirus S glycoproteins have recently been shown to undergo large-scale conformational changes upon fusion with the host cell membrane (30). We have demonstrated that it is possible to generate ribv expressing chimaeric S glycoproteins comprised of S1 and S2 subunits from two different strains of IBV that are able to maintain productive infections in vitro and generate peak titres similar to the parent viruses. This indicates that the chimaeric spikes are still able to undergo the conformational changes required during entry to host cells. It is possible that inefficient conformational changes leading to a reduction in rate of viral entry to host cells may be responsible for the lag in viral growth kinetics for some of the ribvs with modified spike glycoproteins at early times in the course of infection. These may be corrected by the additional mutations accrued during adaptation of the ribvs to Vero cells as the growth kinetics of all ribvs increased upon serial passage. Although live attenuated and inactivated vaccines are universally used in the control of IBV, they do not offer cross-protection between the different circulating serotypes of IBV. The advent of a reverse genetics system for IBV (78, 84, 85) creates the opportunity for generating rationally designed and more effective vaccines. The prospect of swapping spike genes from emerging strains of IBV into an attenuated backbone such as Beau-R are promising (8, 92). The observation that IBV Beaudette, a highly-attenuated laboratory strain, has the additional tropism for growth in Vero cells invokes the further possibility of generating IBV vaccines produced from cultured cells rather than the use of embryonated eggs. Vero cells were first isolated in 1962 from kidney epithelial cells extracted from an African green monkey. They have already been validated for virus growth and diagnostic purposes and are 21
22 licensed for use in human vaccine manufacture. Vero cells are currently used in the production of polio and rabies vaccines (93, 94) and several influenza virus vaccines have been developed for growth on Vero cells (95, 96). Vero cells have been extensively tested for tumorigenic properties, can be grown in suspension or flat bed and it is possible to achieve consistent virus yields. Beaudette is itself too attenuated to be an efficacious vaccine (8, 97) however, we have identified the specific region in the Beaudette genome, within the spike glycoprotein, responsible for its ability to grow in Vero cells. Whilst the S1 subunit of IBV contains the receptor binding domain and is responsible for binding to host cells, it was determined that infectivity for Vero cells is conferred by the Beaudette S2 subunit, in particular the Beaudette-specific motif 686 SRRKRSLIE 694 surrounding the S2ʹ cleavage site. Although there is a clear indication of the involvement of the Beaudette-specific motif in the ability of Beau- R to replicate on Vero cells, the addition of other Beaudette-specific amino acids within the S2 subunit or mutations introduced by serial passage in Vero cells were able to further increase the growth kinetics of the ribvs on Vero cells. These additional mutations may serve to optimise the structure of the recombinant S glycoprotein. Interestingly, the Beaudette-specific motif doesn t appear to confer ability to grow on BHK-21 cells in the same way (data not shown) indicating that there may be other regions of the S glycoprotein responsible for the extended host range of Beau-R in mammalian cells other than Vero cells. The Beaudette-specific motif has two putative mechanisms to permit infectivity for Vero cells, either by facilitating binding to additional host attachment factors (63) or as an additional protease cleavage site (64). It is possible that both activities may play a role in the extended tropism of Beaudette in vitro. Results of this study suggest that IBV Beaudette employs a different mechanism of entry into Vero cells than into CK cells. Western blot analysis demonstrates that ribvs containing the Beaudette-specific motif are susceptible to 22
23 cleavage at the S2ʹ site whereas the ribvs containing the equivalent sequence from M41 are not cleaved at the S2ʹ site. Efficiency of this cleavage appears to be lower in the ribvs containing the BSM than in Beau-R however, which may be due to differences in tertiary or quaternary structure between the spike glycoproteins resulting in the S2ʹ site being less accessible to the protease. The S2ʹ cleavage product of 472 amino acids equates to around 52 kda if you assume a linear relationship between molecular weight and size of the protein. The S2 is glycosylated and palmitoylated, which alters the size and can distort the protein coils that form when denatured using SDS. We consistently see a band of around 30 kda in Beau-R infected samples. It is possible that this band may be a breakdown product of the S2ʹ cleavage product, or the size of the S2ʹ cleavage product may appear to be smaller than expected due to distortion of protein coils in the gel. Nonetheless, this study contributes to the mounting evidence from other coronaviruses that cleavage at the S2ʹ site is involved in entry, and this region plays a role in fusion activation (64, 66, 67, 69). Inclusion of this Beaudette-specific region of the spike glycoprotein in recombinant viruses confers the ability to grow in Vero cells to incompetent strains such as BeauR- M41(S). We have previously demonstrated that this recombinant IBV, expressing the M41 spike glycoprotein in the genetic background of Beaudette, is capable of inducing protective immunity against challenge with virulent M41 (8). These findings are significant as they allow the development of vaccine strains expressing different spike glycoproteins that may be propagated in cell culture, saving costs and reducing the number of eggs required for vaccine production. Acknowledgements The authors wish to acknowledge the help and support provided by the Microbiological 561 Services department at The Pirbright Institute, Compton Laboratory. This work was 23
24 supported by Biotechnology and Biological Sciences Research Council (BBSRC) grant BBS/E/I/ and strategic funding to The Pirbright Institute References 1. Carstens EB Ratification vote on taxonomic proposals to the International Committee on Taxonomy of Viruses (2009). Archives of Virology 155: Cavanagh D Coronaviruses in poultry and other birds. Avian Pathology 34: Cavanagh D, Gelb J, Jr Infectious Bronchitis, p In Saif YM (ed), Diseases of Poultry, 12th ed. Blackwell Publishing, Iowa. 4. Jones RC Viral respiratory diseases (ILT, ampv infections, IB): are they ever under control? British Poultry Science 51: de Wit JJ, Cook JKA, van der Heijden HMJF Infectious bronchitis virus variants: a review of the history, current situation and control measures. Avian Pathology 40: Cavanagh D, Naqi S Infectious bronchitis, p In Saif YM, Barnes HJ, Glisson JR, Fadly AM, McDougald LR, Swayne DE (ed), Diseases of Poultry, 11 ed. Iowa State University Press, Ames, Iowa. 7. Britton P, Cavanagh D Avian coronavirus diseases and infectious bronchitis vaccine development, p In Thiel V (ed), Coronaviruses: Molecular and Cellular Biology. Caister Academic Press, Norfolk, UK. 8. Hodgson T, Casais R, Dove B, Britton P, Cavanagh D Recombinant infectious bronchitis coronavirus Beaudette with the spike protein gene of the pathogenic M41 strain remains attenuated but induces protective immunity. Journal of Virology 78: Ambali AG, Jones RC Early pathogenesis in chicks of infection with an enterotropic strain of infectious bronchitis virus. Avian Diseases 34: Cavanagh D Coronavirus avian infectious bronchitis virus. Veterinary Research 38: Lambrechts C, Pensaert M, Ducatelle R Challenge experiments to evaluate cross-protection induced at the trachea and kidney level by vaccine strains and Belgian nephropathogenic isolates of avian infectious bronchitis virus. Avian Pathology 22: Ziegler AF, Ladman BS, Dunn PA, Schneider A, Davison S, Miller PG, Lu H, Weinstock D, Salem M, Eckroade RJ, Gelb J Nephropathogenic infectious bronchitis in Pennsylvania chickens Avian Diseases 46: Britton P, Cavanagh D Nidovirus genome organization and expression mechanisms, p In Perlman S, Gallagher T, Snijder EJ (ed), Nidoviruses. ASM Press, Washington, DC. 14. de Vries AAF, Horzinek MC, Rottier PJM, de Groot RJ The genome organisation of the Nidovirales: Similarities and differences between Arteri-, Toroand Coronaviruses. Seminars in Virology 8: Lai MM, Cavanagh D The molecular biology of coronaviruses. Advances in Virus Research 48: Delmas B, Laude H Assembly of coronavirus spike protein into trimers and its role in epitope expression. Journal of Virology 64:
25 Vennema H, Rottier PJ, Heijnen L, Godeke GJ, Horzinek MC, Spaan WJ Biosynthesis and function of the coronavirus spike protein. Advances in Experimental Medicine and Biology 276: Bosch BJ, van der Zee R, de Haan CA, Rottier PJ The coronavirus spike protein is a class I virus fusion protein: structural and functional characterization of the fusion core complex. Journal of Virology 77: Walls AC, Tortorici MA, Bosch BJ, Frenz B, Rottier PJ, DiMaio F, Rey FA, Veesler D Cryo-electron microscopy structure of a coronavirus spike glycoprotein trimer. Nature 531: Kirchdoerfer RN, Cottrell CA, Wang N, Pallesen J, Yassine HM, Turner HL, Corbett KS, Graham BS, McLellan JS, Ward AB Pre-fusion structure of a human coronavirus spike protein. Nature 531: Shang J, Zheng Y, Yang Y, Liu C, Geng Q, Luo C, Zhang W, Li F, Cryo- EM structure of infectious bronchitis coronavirus spike protein reveals structural and functional evolution of coronavirus spike proteins. PLoS Pathog 14:e Gallagher TM, Buchmeier MJ Coronavirus spike proteins in viral entry and pathogenesis. Virology 279: De Haan CA, Stadler K, Godeke GJ, Bosch BJ, Rottier PJ Cleavage inhibition of the murine coronavirus spike protein by a furin-like enzyme affects cellcell but not virus-cell fusion. Journal of Virology 78: Sturman LS, Ricard CS, Holmes KV Proteolytic cleavage of the E2 glycoprotein of murine coronavirus: activation of cell-fusing activity of virions by trypsin and separation of two different 90K cleavage fragments. Journal of Virology 56: Spaan W, Cavanagh D, Horzinek MC Coronaviruses: structure and genome expression. Journal of General Virology 69: Koch G, Hartog L, Kant A, van Roozelaar DJ Antigenic domains of the peplomer protein of avian infectious bronchitis virus: correlation with biological function. Journal of General Virology 71: Schultze B, Cavanagh D, Herrler G Neuraminidase treatment of avian infectious bronchitis coronavirus reveals a hemagglutinating activity that is dependent on sialic acid-containing receptors on erythrocytes. Virology 189: Zelus BD, Schickli JH, Blau DM, Weiss SR, Holmes KV Conformational changes in the spike glycoprotein of murine coronavirus are induced at 37 degrees C either by soluble murine CEACAM1 receptors or by ph 8. Journal of Virology 77: Matsuyama S, Taguchi F Two-step conformational changes in a coronavirus envelope glycoprotein mediated by receptor binding and proteolysis. Journal of Virology 83: Walls AC, Tortorici MA, Snijder J, Xiong X, Bosch BJ, Rey FA, Veesler D Tectonic conformational changes of a coronavirus spike glycoprotein promote membrane fusion. Proc Natl Acad Sci U S A 114: Risco C, Anton IM, Enjuanes L, Carrascosa JL The transmissible gastroenteritis coronavirus contains a spherical core shell consisting of M and N proteins. Journal of Virology 70: Luo ZL, Weiss SR Roles in cell-to-cell fusion of two conserved hydrophobic regions in the murine coronavirus spike protein. Virology 244: Xu Y, Lou Z, Liu Y, Pang H, Tien P, Gao GF, Rao Z Crystal structure of severe acute respiratory syndrome coronavirus spike protein fusion core. J Biol Chem 279:
26 Lu L, Liu Q, Zhu Y, Chan K, Qin L, Li Y, Wang Q, Chan JF, du L, Yu F, Ma C, Ye S, Yuen K, Zhang R, Jiang S Structure-based discovery of Middle East respiratory syndrome coronavirus fusion inhibitor. Nature Communications Zheng Q, deng Y, Liu J, Van der Hoek L, Berkhout B, Lu M Core structure of S2 from the human coronavirus NL63 spike glycoprotein.. Biochemistry 45: Xu Y, Liu Y, Lou Z, Qin L, Li X, Bai Z, Pang H, Tien P, Gao GF, Rao Z Structural basis for coronavirus-mediated membrane fusion. Crystal structure of mouse hepatitis virus spike protein fusion core. J Biol Chem 279: Duquerroy S, Vigouroux A, Rottier PJ, Rey FA, Jan Bosch B Central ions and lateral asparagine/glutamine zippers stabilize the post-fusion hairpin conformation of the SARS coronavirus spike glycoprotein. Virology 335: Gao J, Lu G, Qi J, Li Y, Wu Y, Deng Y, Geng H, Li H, Wang Q, Xiao H, Tan W, Yan J, Gao GF Structure of the fusion core and inhibition of fusion by a heptad repeat peptide derived from the S protein of Middle East respiratory syndrome coronavirus. J Virol 87: Supekar VM, Bruckmann C, Ingallinella P, Bianchi E, Pessi A, Carfi A Structure of a proteolytically resistant core from the severe acute respiratory syndrome coronavirus S2 fusion protein. Proc Natl Acad Sci U S A 101: de Groot RJ, Lujtjes W, Horzinek MC, van der Zeijst BAM, Spaan WJ, Lenstra JA Evidence for a coiled-coil structure in the spike proteins of coronaviruses. Journal of Molecular Biology 196: Tripet B, Howard MW, Jobling M, Holmes RK, Holmes KV, Hodges RS Structural characterization of the SARS-coronavirus spike S fusion protein core. Journal of Biological Chemistry 279: Guo Y, Tisoncik J, McReynolds S, Farzan M, Prabhakar BS, Gallagher T, Rong L, Caffrey M Identification of a new region of SARS-CoV S protein critical for viral entry. Journal of Molecular Biology 394: Shulla A, Gallagher T Role of spike protein endodomains in regulating coronavirus entry. Journal of Biological Chemistry 284: Opstelten DJ, Raamsman MJ, Wolfs K, Horzinek MC, Rottier PJ Envelope glycoprotein interactions in coronavirus assembly. Journal of Cell Biology 131: de Haan CA, Smeets M, Vernooij F, Vennema H, Rottier PJ Mapping of the coronavirus membrane protein domains involved in interaction with the spike protein. Journal of Virology 73: Kubo H, Yamada YK, Taguchi F Localization of neutralizing epitopes and the receptor-binding site within the amino-terminal 330 amino acids of the murine coronavirus spike protein. J Virol 68: Bonavia A, Zelus BD, Wentworth DE, Talbot PJ, Holmes KV Identification of a receptor-binding domain of the spike glycoprotein of human coronavirus HCoV- 229E. Journal of Virology 77: Babcock GJ, Esshaki DJ, Thomas WD, Jr., Ambrosino DM Amino acids 270 to 510 of the severe acute respiratory syndrome coronavirus spike protein are required for interaction with receptor. J Virol 78: Promkuntod N, van Eijndhoven RE, de Vrieze G, Grone A, Verheije MH Mapping of the receptor-binding domain and amino acids critical for attachment in the spike protein of avian coronavirus infectious bronchitis virus. Virology 448:
27 Delmas B, Gelfi J, L'Haridon R, Vogel LK, Sj"str"m H, Nor n O, Laude H Aminopeptidase N is a major receptor for the enteropathogenic coronavirus TGEV. Nature 357: Yeager CL, Ashmun RA, Williams RK, Cardellichio CB, Shapiro LH, Look AT, Holmes KV Human aminopeptidase N is a receptor for human coronavirus 229E. Nature 357: Tresnan DB, Levis R, Holmes KV Feline aminopeptidase N serves as a receptor for feline, canine, porcine, and human coronaviruses in serogroup I. Journal of Virology 70: Williams RK, Jiang G-S, Holmes KV Receptor for mouse hepatitis virus is a member of the carcinoembryonic antigen family of glycoproteins. Proceedings of the National Academy of Science USA 88: Langereis MA, van Vliet AL, Boot W, de Groot RJ Attachment of mouse hepatitis virus to O-acetylated sialic acid is mediated by hemagglutinin-esterase and not by the spike protein. Journal of Virology 84: Li W, Moore MJ, Vasilieva N, Sui J, Wong SK, Berne MA, Somasundaran M, Sullivan JL, Luzuriaga K, Greenough TC, Choe H, Farzan M Angiotensinconverting enzyme 2 is a functional receptor for the SARS coronavirus. Nature 426: Jeffers SA, Tusell SM, Gillim-Ross L, Hemmila EM, Achenbach JE, Babcock GJ, Thomas WD, Jr., Thackray LB, Young MD, Mason RJ, Ambrosino DM, Wentworth DE, Demartini JC, Holmes KV CD209L (L-SIGN) is a receptor for severe acute respiratory syndrome coronavirus. Proceedings of the National Academy of Science USA 101: Han DP, Lohani M, Cho MW Specific asparagine-linked glycosylation sites are critical for DC-SIGN- and L-SIGN-mediated severe acute respiratory syndrome coronavirus entry. Journal of Virology 81: Raj VS, Mou H, Smits SL, Dekkers DH, Muller MA, Dijkman R, Muth D, Demmers JA, Zaki A, Fouchier RA, Thiel V, Drosten C, Rottier PJ, Osterhaus AD, Bosch BJ, Haagmans BL Dipeptidyl peptidase 4 is a functional receptor for the emerging human coronavirus-emc. Nature 495: Winter C, Schwegmann-Wessels C, Cavanagh D, Neumann U, Herrler G Sialic acid is a receptor determinant for infection of cells by avian Infectious bronchitis virus. Journal of General Virology 87: Wickramasinghe IN, de Vries RP, Grone A, de Haan CA, Verheije MH Binding of avian coronavirus spike proteins to host factors reflects virus tropism and pathogenicity. Journal of Virology 85: Winter C, Herrler G, Neumann U Infection of the tracheal epithelium by infectious bronchitis virus is sialic acid dependent. Microbes and Infection 10: Abd El Rahman S, El-Kenawy AA, Neumann U, Herrler G, Winter C Comparative analysis of the sialic acid binding activity and the tropism for the respiratory epithelium of four different strains of avian infectious bronchitis virus. Avian Pathology 38: Madu IG, Chu VC, Lee H, Regan AD, Bauman BE, Whittaker GR Heparan sulfate is a selective attachment factor for the avian coronavirus infectious bronchitis virus Beaudette. Avian Diseases 51: Yamada Y, Liu DX Proteolytic activation of the spike protein at a novel RRRR/S motif is implicated in furin-dependent entry, syncytium formation, and 27
28 infectivity of coronavirus infectious bronchitis virus in cultured cells. Journal of Virology 83: Tay F, Huang M, Wang L, Yamada Y, Liu DX Characterization of cellular furin content as a potential factor determining the susceptibility of cultured human and animal cells to coronavirus infectious bronchitis virus infection. Virology 433: Belouzard S, Chu VC, Whittaker GR Activation of the SARS coronavirus spike protein via sequential proteolytic cleavage at two distinct sites. Proc Natl Acad Sci U S A 106: Millet JK, Whittaker GR Host cell entry of Middle East respiratory syndrome coronavirus after two-step, furin-mediated activation of the spike protein. Proc Natl Acad Sci U S A 111: Millet JK, Goldstein M, Labitt RN, Hsu H, Daniel S, Whittaker GR A camel-derived MERS-CoV with a variant spike protein cleavage site and distinct fusion activation properties. Emerging Microbes & Infections Bonnin A, Danneels A, Dubuisson J, Goffard A, Belouzard S HCoV-229E spike protein fusion activation by trypsin-like serine proteases is mediated by proteolytic processing in the S2' region. Journal of General Virology doi: /jgv Sanchez CM, Izeta A, Sanchez-Morgado JM, Alonso S, Sola I, Balasch M, Plana- Duran J, Enjuanes L Targeted recombination demonstrates that the spike gene of transmissible gastroenteritis coronavirus is a determinant of its enteric tropism and virulence. Journal of Virology 73: Tekes G, Hofmann-Lehmann R, Bank-Wolf B, Maier R, Thiel HJ, Thiel V Chimeric feline coronaviruses that encode type II spike protein on type I genetic background display accelerated viral growth and altered receptor usage. Journal of Virology 84: Cunningham CH, Spring MP, Nazerian K Replication of Avian Infectious Bronchitis Virus in African Green Monkey Kidney Cell Line VERO. Journal of General Virology 16: Otsuki K, Noro K, Yamamoto H, Tsubokura M Studies on avian infectious bronchitis virus (IBV). II. Propagation of IBV in several cultured cells. Archives of Virology 60: Casais R, Dove B, Cavanagh D, Britton P Recombinant avian infectious bronchitis virus expressing a heterologous spike gene demonstrates that the spike protein is a determinant of cell tropism. Journal of Virology 77: Van Roekel H, Clarke MK, Bullis KL, Olesiuk OM, Sperling FG Infectious Bronchitis. American Journal of Veterinary Research 12: Cook JKA, Darbyshire JH, Peters RW The use of chicken tracheal organ cultures for the isolation and assay of avian infectious bronchitis virus. Archives of Virology 50: Darbyshire JH, Rowell JG, Cook JKA, Peters RW Taxonomic studies on strains of avian infectious bronchitis virus using neutralisation tests in tracheal organ cultures. Archives of Virology 61: Casais R, Thiel V, Siddell SG, Cavanagh D, Britton P Reverse genetics system for the avian coronavirus infectious bronchitis virus. Journal of Virology 75: Beaudette FR, Hudson CB Cultivation of the virus of infectious bronchitis. Journal of the American Veterinary Medical Association 90:
29 Cavanagh D, Davis PJ, Pappin DJC, Binns MM, Boursnell MEG, Brown TDK Coronavirus IBV: partial amino terminal sequencing of spike polypeptide S2 identifies the sequence Arg-Arg-Phe-Arg-Arg at the cleavage site of the spike precursor propolypeptide of IBV strains Beaudette and M41. Virus Research 4: Pénzes Z, Tibbles K, Shaw K, Britton P, Brown TDK, Cavanagh D Characterization of a replicating and packaged defective RNA of avian coronavirus infectious bronchitis virus. Virology 203: Pénzes Z, Wroe C, Brown TD, Britton P, Cavanagh D Replication and packaging of coronavirus infectious bronchitis virus defective RNAs lacking a long open reading frame. Journal of Virology 70: Stirrups K, Shaw K, Evans S, Dalton K, Cavanagh D, Britton P Leader switching occurs during the rescue of defective RNAs by heterologous strains of the coronavirus infectious bronchitis virus. Journal of General Virology 81: Britton P, Evans S, Dove B, Davies M, Casais R, Cavanagh D Generation of a recombinant avian coronavirus infectious bronchitis virus using transient dominant selection. Journal of Virological Methods 123: Armesto M, Casais R, Cavanagh D, Britton P Transient dominant selection for the modification and generation of recombinant infectious bronchitis coronaviruses. Methods in Molecular Biology 454: Wickramasinghe IN, van Beurden SJ, Weerts EA, Verheije MH The avian coronavirus spike protein. Virus Res 194: Cunningham CH Avian Infectious Bronchitis. Advances in Veterinary Science 14: Cunningham CH Immunity to Avian Infectious Bronchitis, p In Perkins FT, Regamey RH (ed), Immunity to Infections of the Respiratory System in Man and Animals, vol 28. Karger, London. 89. Armesto M, Bentley K, Bickerton E, Keep S, Britton P Coronavirus reverse genetics, p In Bridgen A (ed), Reverse Genetics of RNA Viruses: Applications and Perspectives. Wiley-Blackwell Press, United Kingdom. 90. Boursnell MEG, Brown TDK, Foulds IJ, Green PF, Tomley FM, Binns MM Completion of the sequence of the genome of the coronavirus avian infectious bronchitis virus. Journal of General Virology 68: Bonfield JK, Smith KF, Staden R A new DNA sequence assembly program. Nucleic Acids Research 23: Armesto M, Evans S, Cavanagh D, Abu-Median AB, Keep S, Britton P A recombinant avian infectious bronchitis virus expressing a heterologous spike gene belonging to the 4/91 serotype. PLoS ONE 6:e Frazatti-Gallina NM, Mourao-Fuches RM, Paoli RL, Silva MLN, Miyaki C, Valentini EJG, Raw I, Higashi HG Vero-cell rabies vaccine produced using serum-free medium. Vaccine 23: Montagnon BJ, Fanget B, Nicolas AJ The large-scale cultivation of VERO cells in micro-carrier culture for virus vaccine production. Preliminary results for killed poliovirus vaccine. Dev Biol Stand 47: Govorkova EA, Murti G, Meignier B, De Taisne C, Webster RG African Green Monkey Kidney (Vero) Cells Provide an Alternative Host Cell System for Influenza A and B Viruses. Journal of Virology 70: Kistner O, Barrett PN, Mundt W, Reiter M, Schober-Bendixen S, Dorner F Development of a mammalian cell (Vero) derived candidate influenza virus vaccine. Vaccine 16:
30 Geilhausen HE, Ligon FB, Lukert PD The pathogenesis of virulent and avirulent avian infectious bronchitis virus. Archiv für die Gesamte Virusforschung 40: Table 1. Mutations identified in the S genes of ribvs. Mutation Virus Passage nt postion nt change aa position aa change P 3 -CKC GTT - AAC 625 LF - LT BeauR-M41(S1) GTT - AAC 625 LF - LT P 7 -Vero C - T 1004 T - I BeauR-M41(S2) P 3 -CKC G - A 452 A - T BeauR-S-MM P 3 -CKC A - C 620 D - A BeauR-M41-S-BSM P 7 -Vero A - C 620 D - A BeauR-M41-S-BSM-N 617 S P 7 -Vero A - C 866 Q - H P 3 -CKC A - C 493 E - D BeauR-M41-S-BSM-L 578 F A - C 493 E - D N 617 S P 7 -Vero A - C 866 Q - H BeauR-M41-S-BSM-I 1000 V P 7 -Vero C - T 578 L - F BeauR-M41-S-BSM-L 857 F- P 7 -Vero C - T 630 P - S 30
31 I 1000 V P 3 -CKC C - mixed T/C 43 H - Y G - A 448 V - I BeauR-M41-S-BSM-N 826 S- L 857 F-I 1000 V P 7 -Vero C - T 578 L - F C - mixed A/C 659 P - T 866 BeauR-M41-S-BSM-L 578 F- N 617 S-N 826 S-L 857 F-I 1000 V C - mixed T/C 857 L - F A - mixed T/A 866 Q - H P 7 -Vero G - A 73 G - S Downloaded from on December 22, 2018 by guest 31
32 Figure 1. Schematic diagram of the IBV S glycoprotein and identification of Beaudette- specific amino acids in the S2 subunit. A. The S1 domain contains the receptor binding domain and the cleaved signal sequence. The S2 domain contains two heptad repeats, HR1 870 and 2, a transmembrane domain and a cytoplasmic domain. Beau-R also contains a Beaudette-specific sequence not found in M41. B. The ectodomain of the S2 subunit of IBV strains Beau-CK (Accession number AJ311317), M41 (Accession number X04722), 4/91(UK) (Accession number JN192154) and QX L1148 (Accession number KY933090) is shown. There are 19 amino acid differences between the S2 subunits of Beau-R and M41- CK. The Beaudette-specific motif is labelled with a horizontal line. A black box surrounds each amino acid change made in the spike glycoproteins of the recombinant viruses generated in this study and the amino acid positions are shown above. Figure 2. Growth characteristics of ribvs BeauR-M41(S1) and BeauR-M41(S2) P 3 -CK on CK and Vero cells. A. Chick kidney cells and B. Vero cells in 6-well plates were infected with MOI 0.5 Beau-R, M41-CK, BeauR-M41(S), BeauR-M41(S1) P 3 -CK and BeauR-M41(S2) P 3 -CK. Supernatant was harvested at 1, 12, 24, 48 and 72 hours postinfection and titrated on CK cells. Three replicates were performed and the averages taken. Error bars indicate standard error of the mean. Vero cells were infected with C. mock, D. Beau-R, E. M41-CK, F. BeauR-M41(S), G. BeauR-M41(S1) P 3 -CK, H. BeauR-M41(S2) P 3 - CK. Infected cells were fixed 24 hours post-infection and immunolabelled with mouse antidsrna, secondary antibody AlexaFluor 488 goat anti-mouse (green, Invitrogen). Nuclei were labelled with DAPI (blue). The scale bars represent 50 µm. Figure 3. Adaptation of ribv BeauR-M41(S1) to Vero cells. Vero cells in 6-well plates were infected with MOI 0.5 Beau-R and BeauR-M41(S1) P 7 -Vero. Supernatant was 32
33 harvested at 1, 12, 24, 48 and 72 hours post-infection and titrated on CK cells. Three replicates were performed and the averages taken. Error bars indicate standard error of the mean, some error bars are too small to be observed Figure 4. Growth characteristics of ribvs BeauR-S-MM and BeauR-M41-S-BSM P 3 - CK on CK and Vero cells. A. Chick kidney cells and D. Vero cells in 6-well plates were infected with MOI 0.5 Beau-R, M41-CK, BeauR-M41(S), BeauR-S-MM P 3 -CK and BeauR- M41-S-BSM P 3 -CK. Supernatant was harvested at 1, 12, 24, 48 and 72 hours post-infection and titrated on CK cells. Three replicates were performed and the averages taken. Error bars indicate standard error of the mean. Vero cells were infected with B. BeauR-M41-S-BSM P 3 - CK and C. BeauR-S-MM P 3 -CK. Infected cells were fixed 24 hours post-infection and immunolabelled with mouse anti-dsrna, secondary antibody AlexaFluor 488 goat antimouse (green, Invitrogen). Nuclei were labelled with DAPI (blue). The scale bars represent 50 µm. Figure 5. Adaptation of ribv BeauR-M41-S-BSM to Vero cells. Vero cells in 6-well plates were infected with MOI 0.5 Beau-R and BeauR-M41-S-BSM P 7 -Vero. Supernatant was harvested at 1, 12, 24, 48, 72 and 96 hours post-infection and titrated on CK cells. Three replicates were performed and the averages taken. Error bars indicate standard error of the mean, some error bars are too small to be observed. Figure 6. Growth characteristics of ribvs with modified S2 subunits on CK and Vero cells. A. Chick kidney cells and B. Vero cells in 6-well plates were infected with MOI 0.05 Beau-R, BeauR-M41(S), BeauR-M41-S-BSM P 3 -CK, BeauR-M41-S-BSM-N 617 S P 3 -CK, 33
34 BeauR-M41-S-BSM-L 578 F-N 617 S P 3 -CK, BeauR-M41-S-BSM-I 1000 V P 3 -CK, BeauR-M41-S- BSM-L 857 F-I 1000 V P 3 -CK, BeauR-M41-S-BSM-N 826 S-L 857 F-I 1000 V P 3 -CK and BeauR-M41- S-BSM-L 578 F-N 617 S-N 826 S-L 857 F-I 1000 V P 3 -CK. Supernatant was harvested at 1, 12, 24, 48 and 72 hours post-infection and titrated on CK cells. Three replicates were performed and the averages taken. Error bars indicate standard error of the mean. Vero cells were infected with C. BeauR-M41-S-BSM-N 617 S P 3 -CK,, D. BeauR-M41-S-BSM-L 578 F-N 617 S P 3 -CK, E. BeauR-M41-S-BSM-I 1000 V P 3 -CK, F. BeauR-M41-S-BSM-N 826 S-L 857 F-I 1000 V P 3 -CK, G. BeauR-M41-S-BSM-N 826 S-L 857 F-I 1000 V P 3 -CK and H. BeauR-M41-S-BSM-L 578 F-N 617 S- N 826 S-L 857 F-I 1000 V P 3 -CK. Infected cells were fixed 24 hours post-infection and immunolabelled with mouse anti-dsrna, secondary antibody AlexaFluor 488 goat antimouse (green, Invitrogen). Nuclei were labelled with DAPI (blue). The scale bars represent 50 µm. Figure 7. Adaptation of ribvs with modified S2 subunits to Vero cells. Vero cells in 6- well plates were infected with MOI Beau-R, BeauR-M41-S-BSM P 7 -Vero, BeauR- M41-S-BSM-N 617 S P 7 -Vero, BeauR-M41-S-BSM-L 578 F-N 617 S P 7 -Vero, BeauR-M41-S- BSM-I 1000 V P 7 -Vero, BeauR-M41-S-BSM-L 857 F-I 1000 V P 7 -Vero, BeauR-M41-S-BSM-N 826 S- L 857 F-I 1000 V P 7 -Vero and BeauR-M41-S-BSM-L 578 F-N 617 S-N 826 S-L 857 F-I 1000 V P 7 -Vero. Supernatant was harvested at 1, 12, 24, 48 and 72 hours post-infection and titrated on CK cells. Three replicates were performed and the averages taken. Error bars indicate standard error of the mean. Figure 8. Proteolytic cleavage of ribvs at the S2ʹ site is dependent on the Beaudettespecific motif. Chicken kidney cells in six-well plates were 1. Mock infected or infected with 2. Beau-R, 3. M41-CK, 4. BeauR-M41(S), 5 and 6. BeauR-S-MM P 3 -CK, 7 and 8. BeauR- 34
35 M41-S-BSM P 3 -CK. Cells were lysed 24 hours post-infection and spike glycoproteins were analysed by Western blotting with anti-s2 antibody. Actin was detected with anti-beta actin antibody. Two replicates of BeauR-S-MM P 3 -CK and BeauR-M41-S-BSM P 3 -CK were used. Bands corresponding to uncleaved spike, S2 subunit and S2ʹ cleavage product are labelled. The S2ʹ cleavage product was detected in lanes 2, 7 and 8 only. 35
36
37
38

39
40
41


42

43
Coronaviruses. Virion. Genome. Genes and proteins. Viruses and hosts. Diseases. Distinctive characteristics
 Coronaviruses Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Virion Spherical enveloped particles studded with clubbed spikes Diameter 120-160 nm Coiled helical
Coronaviruses Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Virion Spherical enveloped particles studded with clubbed spikes Diameter 120-160 nm Coiled helical
Objective 3. Develop new and improved diagnostic tools, vaccines, and novel management approaches
 Objective 3. Develop new and improved diagnostic tools, vaccines, and novel management approaches Development of novel nanoparticle-base vaccines for infectious bronchitis PI, Mazhar I. Khan; CoPI, Peter
Objective 3. Develop new and improved diagnostic tools, vaccines, and novel management approaches Development of novel nanoparticle-base vaccines for infectious bronchitis PI, Mazhar I. Khan; CoPI, Peter
Viruses 2013, 5, ; doi: /v OPEN ACCESS viruses ISSN
 Viruses 2013, 5, 1924-1933; doi:10.3390/v5081924 OPEN ACCESS viruses ISSN 1999-4915 www.mdpi.com/journal/viruses Article Sialic Acid Binding Properties of Soluble Coronavirus Spike (S1) Proteins: Differences
Viruses 2013, 5, 1924-1933; doi:10.3390/v5081924 OPEN ACCESS viruses ISSN 1999-4915 www.mdpi.com/journal/viruses Article Sialic Acid Binding Properties of Soluble Coronavirus Spike (S1) Proteins: Differences
Part I. Content: History of Viruses. General properties of viruses. Viral structure. Viral classifications. Virus-like agents.
 Viruses Part I Content: History of Viruses. General properties of viruses. Viral structure. Viral classifications. Virus-like agents. History Through the 1800s, many scientists discovered that something
Viruses Part I Content: History of Viruses. General properties of viruses. Viral structure. Viral classifications. Virus-like agents. History Through the 1800s, many scientists discovered that something
PROCEEDINGS OF THE FIFTII INTERNATIONAL SYMPOSIUM ON CORONAVIRUSES. Held September 13-18, 1992, in Chantilly France
 PROCEEDINGS OF THE FIFTII INTERNATIONAL SYMPOSIUM ON CORONAVIRUSES Held September 13-18, 1992, in Chantilly France INDEX Adenovirus expression, ofprcv S protein, 469 oftgev S gene fragments, 455 A59 virus,
PROCEEDINGS OF THE FIFTII INTERNATIONAL SYMPOSIUM ON CORONAVIRUSES Held September 13-18, 1992, in Chantilly France INDEX Adenovirus expression, ofprcv S protein, 469 oftgev S gene fragments, 455 A59 virus,
The humoral immune responses to IBV proteins.
 The humoral immune responses to IBV proteins. E. Dan Heller and Rosa Meir The Hebrew University of Jerusalem, Israel COST FA1207 meeting WG2 + WG3, Budapest, Jan. 2015 1 IBV encodes four major structural
The humoral immune responses to IBV proteins. E. Dan Heller and Rosa Meir The Hebrew University of Jerusalem, Israel COST FA1207 meeting WG2 + WG3, Budapest, Jan. 2015 1 IBV encodes four major structural
Influenza viruses. Virion. Genome. Genes and proteins. Viruses and hosts. Diseases. Distinctive characteristics
 Influenza viruses Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Virion Enveloped particles, quasi-spherical or filamentous Diameter 80-120 nm Envelope is derived
Influenza viruses Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Virion Enveloped particles, quasi-spherical or filamentous Diameter 80-120 nm Envelope is derived
Coronaviruses cause acute, mild upper respiratory infection (common cold).
.") Coronaviruses David A. J. Tyrrell Steven H. Myint GENERAL CONCEPTS Clinical Presentation Coronaviruses cause acute, mild upper respiratory infection (common cold). Structure Spherical or pleomorphic enveloped
Coronaviruses David A. J. Tyrrell Steven H. Myint GENERAL CONCEPTS Clinical Presentation Coronaviruses cause acute, mild upper respiratory infection (common cold). Structure Spherical or pleomorphic enveloped
Recombinant Infectious Bronchitis Viruses expressing chimaeric spike glycoproteins induce
 JVI Accepted Manuscript Posted Online 12 September 2018 J. Virol. doi:10.1128/jvi.01473-18 Copyright 2018 Ellis et al. This is an open-access article distributed under the terms of the Creative Commons
JVI Accepted Manuscript Posted Online 12 September 2018 J. Virol. doi:10.1128/jvi.01473-18 Copyright 2018 Ellis et al. This is an open-access article distributed under the terms of the Creative Commons
Sequencing-Based Identification of a Novel Coronavirus in Ferrets with Epizootic Catarrhal Enteritis and Development of Molecular Diagnostic Tests
 Sequencing-Based Identification of a Novel Coronavirus in Ferrets with Epizootic Catarrhal Enteritis and Development of Molecular Diagnostic Tests A. Wise, Matti Kiupel,, C. Isenhour, R. Maes Coronaviruses
Sequencing-Based Identification of a Novel Coronavirus in Ferrets with Epizootic Catarrhal Enteritis and Development of Molecular Diagnostic Tests A. Wise, Matti Kiupel,, C. Isenhour, R. Maes Coronaviruses
Severe Acute Respiratory Syndrome (SARS) Coronavirus
 Coronavirus") Severe Acute Respiratory Syndrome (SARS) Coronavirus Coronaviruses Coronaviruses are single stranded enveloped RNA viruses that have a helical geometry. Coronaviruses are the largest of RNA viruses with
Severe Acute Respiratory Syndrome (SARS) Coronavirus Coronaviruses Coronaviruses are single stranded enveloped RNA viruses that have a helical geometry. Coronaviruses are the largest of RNA viruses with
Shin-Hee Kim, Yongqi Yan, and Siba K. Samal*
 JOURNAL OF VIROLOGY, Oct. 2009, p. 10250 10255 Vol. 83, No. 19 0022-538X/09/$08.00 0 doi:10.1128/jvi.01038-09 Copyright 2009, American Society for Microbiology. All Rights Reserved. Role of the Cytoplasmic
JOURNAL OF VIROLOGY, Oct. 2009, p. 10250 10255 Vol. 83, No. 19 0022-538X/09/$08.00 0 doi:10.1128/jvi.01038-09 Copyright 2009, American Society for Microbiology. All Rights Reserved. Role of the Cytoplasmic
Department of Veterinary Pathology, University of Liverpool, Leahurst Campus, Neston, South Wirral, CH64 7TE, UNITED KINGDOM
 IMMUNE RESPONSES IN CHICKS AFTER SINGLE OR DUAL VACCINATION WITH LIVE INFECTIOUS BRONCHITIS MASSACHUSETTS AND VARIANT VACCINES: SOME PRELIMINARY FINDINGS GANAPATHY 1 K, ROTHWELL 2 L, LEMIERE 3 S, KAISER
IMMUNE RESPONSES IN CHICKS AFTER SINGLE OR DUAL VACCINATION WITH LIVE INFECTIOUS BRONCHITIS MASSACHUSETTS AND VARIANT VACCINES: SOME PRELIMINARY FINDINGS GANAPATHY 1 K, ROTHWELL 2 L, LEMIERE 3 S, KAISER
TSH Receptor Monoclonal Antibody (49) Catalog Number MA3-218 Product data sheet
 Catalog Number MA3-218 Product data sheet") Website: thermofisher.com Customer Service (US): 1 800 955 6288 ext. 1 Technical Support (US): 1 800 955 6288 ext. 441 TSH Receptor Monoclonal Antibody (49) Catalog Number MA3-218 Product data sheet Details
Website: thermofisher.com Customer Service (US): 1 800 955 6288 ext. 1 Technical Support (US): 1 800 955 6288 ext. 441 TSH Receptor Monoclonal Antibody (49) Catalog Number MA3-218 Product data sheet Details
Institute for Animal Health, Division of Molecular Biology, Compton Laboratory, Compton, Newbury, Berkshire, United Kingdom
 JOURNAL OF VIROLOGY, Dec. 2004, p. 13804 13811 Vol. 78, No. 24 0022-538X/04/$08.00 0 DOI: 10.1128/JVI.78.24.13804 13811.2004 Copyright 2004, American Society for Microbiology. All Rights Reserved. Recombinant
JOURNAL OF VIROLOGY, Dec. 2004, p. 13804 13811 Vol. 78, No. 24 0022-538X/04/$08.00 0 DOI: 10.1128/JVI.78.24.13804 13811.2004 Copyright 2004, American Society for Microbiology. All Rights Reserved. Recombinant
LEC 2, Medical biology, Theory, prepared by Dr. AYAT ALI
 General Characteristics, Structure and Taxonomy of Viruses Viruses A virus is non-cellular organisms made up of genetic material and protein that can invade living cells. They are considered both a living
General Characteristics, Structure and Taxonomy of Viruses Viruses A virus is non-cellular organisms made up of genetic material and protein that can invade living cells. They are considered both a living
Chapter 6- An Introduction to Viruses*
 Chapter 6- An Introduction to Viruses* *Lecture notes are to be used as a study guide only and do not represent the comprehensive information you will need to know for the exams. 6.1 Overview of Viruses
Chapter 6- An Introduction to Viruses* *Lecture notes are to be used as a study guide only and do not represent the comprehensive information you will need to know for the exams. 6.1 Overview of Viruses
Lecture 2: Virology. I. Background
 Lecture 2: Virology I. Background A. Properties 1. Simple biological systems a. Aggregates of nucleic acids and protein 2. Non-living a. Cannot reproduce or carry out metabolic activities outside of a
Lecture 2: Virology I. Background A. Properties 1. Simple biological systems a. Aggregates of nucleic acids and protein 2. Non-living a. Cannot reproduce or carry out metabolic activities outside of a
Influenza or flu is a
 Clinical and Research Area Infectious Diseases Influenza Virus Types A and B Influenza or flu is a respiratory illness that is caused by influenza viruses. Influenza viruses type A and type B cause seasonal
Clinical and Research Area Infectious Diseases Influenza Virus Types A and B Influenza or flu is a respiratory illness that is caused by influenza viruses. Influenza viruses type A and type B cause seasonal
History electron microscopes
 Viruses History Through the 1800s, many scientists discovered that something smaller than bacteria could cause disease and they called it virion (Latin word- poison) In the 1930s, after the invention of
Viruses History Through the 1800s, many scientists discovered that something smaller than bacteria could cause disease and they called it virion (Latin word- poison) In the 1930s, after the invention of
TITLE: Influenza A (H7N9) virus evolution: Which genetic mutations are antigenically important?
 virus evolution: Which genetic mutations are antigenically important?") TITLE: Influenza A (H7N9) virus evolution: Which genetic mutations are antigenically important? AUTHORS: Joshua G. Petrie 1, Adam S. Lauring 2,3 AFFILIATIONS: 1 Department of Epidemiology, University of
TITLE: Influenza A (H7N9) virus evolution: Which genetic mutations are antigenically important? AUTHORS: Joshua G. Petrie 1, Adam S. Lauring 2,3 AFFILIATIONS: 1 Department of Epidemiology, University of
Temperature-Sensitive Mutants Isolated from Hamster and
 JOURNAL OF VIROLOGY, Nov. 1975, p. 1332-1336 Copyright i 1975 American Society for Microbiology Vol. 16, No. 5 Printed in U.S.A. Temperature-Sensitive Mutants Isolated from Hamster and Canine Cell Lines
JOURNAL OF VIROLOGY, Nov. 1975, p. 1332-1336 Copyright i 1975 American Society for Microbiology Vol. 16, No. 5 Printed in U.S.A. Temperature-Sensitive Mutants Isolated from Hamster and Canine Cell Lines
Patricia Fitzgerald-Bocarsly
 FLU Patricia Fitzgerald-Bocarsly October 23, 2008 Orthomyxoviruses Orthomyxo virus (ortho = true or correct ) Negative-sense RNA virus (complementary to mrna) Five different genera Influenza A, B, C Thogotovirus
FLU Patricia Fitzgerald-Bocarsly October 23, 2008 Orthomyxoviruses Orthomyxo virus (ortho = true or correct ) Negative-sense RNA virus (complementary to mrna) Five different genera Influenza A, B, C Thogotovirus
Worldwide perspective on Infectious Bronchitis. Ruth Bouwstra, DVM, PhD Turkey February 2017
 Worldwide perspective on Infectious Bronchitis Ruth Bouwstra, DVM, PhD Turkey February 2017 Infectious bronchitis virus Corona Virus, a ssrna virus - Relatively high rate of mutations (0,0012 subst per
Worldwide perspective on Infectious Bronchitis Ruth Bouwstra, DVM, PhD Turkey February 2017 Infectious bronchitis virus Corona Virus, a ssrna virus - Relatively high rate of mutations (0,0012 subst per
Immunogenicity of Avian Influenza H7N9 Virus in Birds
 Immunogenicity of Avian Influenza H7N9 Virus in Birds Identification of Viral Epitopes Recognized by the Immune System Following Vaccination and Challenge Darrell R. Kapczynski US DEPARTMENT OF AGRICULTURE,
Immunogenicity of Avian Influenza H7N9 Virus in Birds Identification of Viral Epitopes Recognized by the Immune System Following Vaccination and Challenge Darrell R. Kapczynski US DEPARTMENT OF AGRICULTURE,
Identification of Microbes Lecture: 12
 Diagnostic Microbiology Identification of Microbes Lecture: 12 Electron Microscopy 106 virus particles per ml required for visualization, 50,000-60,000 magnification normally used. Viruses may be detected
Diagnostic Microbiology Identification of Microbes Lecture: 12 Electron Microscopy 106 virus particles per ml required for visualization, 50,000-60,000 magnification normally used. Viruses may be detected
Original Article Identification of Different Serotypes of Infectious Bronchitis Viruses in Allantoic Fluid Samples with Single and Multiplex RT- PCR
 Iranian Journal of Virology 2009;3(2): 24-29 2009, Iranian Society for Virology Original Article Identification of Different Serotypes of Infectious Bronchitis Viruses in Allantoic Fluid Samples with Single
Iranian Journal of Virology 2009;3(2): 24-29 2009, Iranian Society for Virology Original Article Identification of Different Serotypes of Infectious Bronchitis Viruses in Allantoic Fluid Samples with Single
Original Article S1 gene sequence analysis of infectious bronchitis virus vaccinal strains (H120 & H52) and their embryo-passaged derivatives
 and their embryo-passaged derivatives") Archives of Razi Institute, Vol. 71, No. 2 (2016) 87-96 Copyright 2016 by Razi Vaccine & Serum Research Institute Original Article S1 gene sequence analysis of infectious bronchitis virus vaccinal strains
Archives of Razi Institute, Vol. 71, No. 2 (2016) 87-96 Copyright 2016 by Razi Vaccine & Serum Research Institute Original Article S1 gene sequence analysis of infectious bronchitis virus vaccinal strains
Identification of Mutation(s) in. Associated with Neutralization Resistance. Miah Blomquist
 in. Associated with Neutralization Resistance. Miah Blomquist") Identification of Mutation(s) in the HIV 1 gp41 Subunit Associated with Neutralization Resistance Miah Blomquist What is HIV 1? HIV-1 is an epidemic that affects over 34 million people worldwide. HIV-1
Identification of Mutation(s) in the HIV 1 gp41 Subunit Associated with Neutralization Resistance Miah Blomquist What is HIV 1? HIV-1 is an epidemic that affects over 34 million people worldwide. HIV-1
Animal hosts Natural host Laboratory animals Rabbits Mice Rats Hamsters Newborn or suckling rodents Animal models for viral pathogenesis 4 Growth of v
 Principles of Virology Department of Molecular Genetics & Microbiology Univ ersity of Florida, Gainesv ille, FL 1 Outline Virus cultivation Assay of viruses Virus genetics 2 Virus isolation Evidence of
Principles of Virology Department of Molecular Genetics & Microbiology Univ ersity of Florida, Gainesv ille, FL 1 Outline Virus cultivation Assay of viruses Virus genetics 2 Virus isolation Evidence of
بسم هللا الرحمن الرحيم
 - 1 - - - 1 P a g e بسم هللا الرحمن الرحيم This sheet was made from record section 1 all information are included - Introduction Our respiratory tract is divided anatomically to upper (URT),middle and
- 1 - - - 1 P a g e بسم هللا الرحمن الرحيم This sheet was made from record section 1 all information are included - Introduction Our respiratory tract is divided anatomically to upper (URT),middle and
Viral vaccines. Lec. 3 أ.د.فائزة عبد هللا مخلص
 Lec. 3 أ.د.فائزة عبد هللا مخلص Viral vaccines 0bjectives 1-Define active immunity. 2-Describe the methods used for the preparation of attenuated live & killed virus vaccines. 3- Comparison of Characteristics
Lec. 3 أ.د.فائزة عبد هللا مخلص Viral vaccines 0bjectives 1-Define active immunity. 2-Describe the methods used for the preparation of attenuated live & killed virus vaccines. 3- Comparison of Characteristics
Investigation of the genetic differences between bovine herpesvirus type 1 variants and vaccine strains
 Investigation of the genetic differences between bovine herpesvirus type 1 variants and vaccine strains Name: Claire Ostertag-Hill Mentor: Dr. Ling Jin Bovine herpesvirus Bovine herpesvirus-1 (BHV-1) Pathogen
Investigation of the genetic differences between bovine herpesvirus type 1 variants and vaccine strains Name: Claire Ostertag-Hill Mentor: Dr. Ling Jin Bovine herpesvirus Bovine herpesvirus-1 (BHV-1) Pathogen
Supplementary Figure 1 Weight and body temperature of ferrets inoculated with
 Supplementary Figure 1 Weight and body temperature of ferrets inoculated with A/Anhui/1/2013 (H7N9) influenza virus. (a) Body temperature and (b) weight change of ferrets after intranasal inoculation with
Supplementary Figure 1 Weight and body temperature of ferrets inoculated with A/Anhui/1/2013 (H7N9) influenza virus. (a) Body temperature and (b) weight change of ferrets after intranasal inoculation with
Influenza A H1N1 HA ELISA Pair Set
 Influenza A H1N1 HA ELISA Pair Set for H1N1 ( A/Puerto Rico/8/1934 ) HA Catalog Number : SEK11684 To achieve the best assay results, this manual must be read carefully before using this product and the
Influenza A H1N1 HA ELISA Pair Set for H1N1 ( A/Puerto Rico/8/1934 ) HA Catalog Number : SEK11684 To achieve the best assay results, this manual must be read carefully before using this product and the
287 Current Topics in Microbiology and Immunology
 287 Current Topics in Microbiology and Immunology Editors R.W. Compans, Atlanta/Georgia M.D. Cooper, Birmingham/Alabama T. Honjo, Kyoto H. Koprowski, Philadelphia/Pennsylvania F. Melchers, Basel M.B.A.
287 Current Topics in Microbiology and Immunology Editors R.W. Compans, Atlanta/Georgia M.D. Cooper, Birmingham/Alabama T. Honjo, Kyoto H. Koprowski, Philadelphia/Pennsylvania F. Melchers, Basel M.B.A.
Reoviruses. Virion. Genome. Genes and proteins. Viruses and hosts. Diseases. Distinctive characteristics
 Reoviruses Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Virion Naked icosahedral capsid (T=13), diameter 60-85 nm Capsid consists of two or three concentric protein
Reoviruses Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Virion Naked icosahedral capsid (T=13), diameter 60-85 nm Capsid consists of two or three concentric protein
SARS Ten Years Later: Lessons for Science and Safety Mark R. Denison M.D.
 SARS Ten Years Later: Lessons for Science and Safety Mark R. Denison M.D. Vanderbilt University School of Medicine SARS Research: You Must be Batty! Mark R. Denison M.D. Vanderbilt University School of
SARS Ten Years Later: Lessons for Science and Safety Mark R. Denison M.D. Vanderbilt University School of Medicine SARS Research: You Must be Batty! Mark R. Denison M.D. Vanderbilt University School of
numbe r Done by Corrected by Doctor
 numbe r 5 Done by Mustafa Khader Corrected by Mahdi Sharawi Doctor Ashraf Khasawneh Viral Replication Mechanisms: (Protein Synthesis) 1. Monocistronic Method: All human cells practice the monocistronic
numbe r 5 Done by Mustafa Khader Corrected by Mahdi Sharawi Doctor Ashraf Khasawneh Viral Replication Mechanisms: (Protein Synthesis) 1. Monocistronic Method: All human cells practice the monocistronic
Materials and Methods , The two-hybrid principle.
 The enzymatic activity of an unknown protein which cleaves the phosphodiester bond between the tyrosine residue of a viral protein and the 5 terminus of the picornavirus RNA Introduction Every day there
The enzymatic activity of an unknown protein which cleaves the phosphodiester bond between the tyrosine residue of a viral protein and the 5 terminus of the picornavirus RNA Introduction Every day there
Coronavirus Pathogenesis and the Emerging Pathogen Severe Acute Respiratory Syndrome Coronavirus
 MICROBIOLOGY AND MOLECULAR BIOLOGY REVIEWS, Dec. 2005, p. 635 664 Vol. 69, No. 4 1092-2172/05/$08.00 0 doi:10.1128/mmbr.69.4.635 664.2005 Copyright 2005, American Society for Microbiology. All Rights Reserved.
MICROBIOLOGY AND MOLECULAR BIOLOGY REVIEWS, Dec. 2005, p. 635 664 Vol. 69, No. 4 1092-2172/05/$08.00 0 doi:10.1128/mmbr.69.4.635 664.2005 Copyright 2005, American Society for Microbiology. All Rights Reserved.
Influenza A H1N1 (Swine Flu 2009) Hemagglutinin / HA ELISA Pair Set
 Hemagglutinin / HA ELISA Pair Set") Influenza A H1N1 (Swine Flu 2009) Hemagglutinin / HA ELISA Pair Set Catalog Number : SEK001 To achieve the best assay results, this manual must be read carefully before using this product and the assay
Influenza A H1N1 (Swine Flu 2009) Hemagglutinin / HA ELISA Pair Set Catalog Number : SEK001 To achieve the best assay results, this manual must be read carefully before using this product and the assay
Viruse associated gastrointestinal infection
 Viruse associated gastrointestinal infection Dr. Hala Al Daghistani Rotaviruses Rotaviruses are a major cause of diarrheal illness in human (infants), and young animals, including calves and piglets. Infections
Viruse associated gastrointestinal infection Dr. Hala Al Daghistani Rotaviruses Rotaviruses are a major cause of diarrheal illness in human (infants), and young animals, including calves and piglets. Infections
Chapter 19: The Genetics of Viruses and Bacteria
 Chapter 19: The Genetics of Viruses and Bacteria What is Microbiology? Microbiology is the science that studies microorganisms = living things that are too small to be seen with the naked eye Microorganisms
Chapter 19: The Genetics of Viruses and Bacteria What is Microbiology? Microbiology is the science that studies microorganisms = living things that are too small to be seen with the naked eye Microorganisms
The chemical fate of biological pollutants in treatment processes
 The chemical fate of biological pollutants in treatment processes http://pubs.rsc.org/en/content/articlehtml/2011/sm/c1sm060 92a http://www.epa.gov/nerlcwww/polio.htm Source: Centers for Disease Control
The chemical fate of biological pollutants in treatment processes http://pubs.rsc.org/en/content/articlehtml/2011/sm/c1sm060 92a http://www.epa.gov/nerlcwww/polio.htm Source: Centers for Disease Control
The N-terminal Region of the Porcine Epidemic Diarrhea Virus Spike Protein is Important for the Receptor Binding
 Korean J. Microbiol. Biotechnol. Vol. 39, No. 2, 140 145 (2011) The N-terminal Region of the Porcine Epidemic Diarrhea Virus Spike Protein is Important for the Receptor Binding Lee, Dong-Kyu 1, Se-Yeoun
Korean J. Microbiol. Biotechnol. Vol. 39, No. 2, 140 145 (2011) The N-terminal Region of the Porcine Epidemic Diarrhea Virus Spike Protein is Important for the Receptor Binding Lee, Dong-Kyu 1, Se-Yeoun
Polyomaviridae. Spring
 Polyomaviridae Spring 2002 331 Antibody Prevalence for BK & JC Viruses Spring 2002 332 Polyoma Viruses General characteristics Papovaviridae: PA - papilloma; PO - polyoma; VA - vacuolating agent a. 45nm
Polyomaviridae Spring 2002 331 Antibody Prevalence for BK & JC Viruses Spring 2002 332 Polyoma Viruses General characteristics Papovaviridae: PA - papilloma; PO - polyoma; VA - vacuolating agent a. 45nm
The Zombies of the Scientific Community Viruses and Other Acellular Infectious Agents. Acellular Agents
 viruses protein and nucleic acid viroids RNA virusoids RNA prions proteins The Zombies of the Scientific Community Viruses and Other Acellular Infectious Agents Acellular Agents Viruses major cause of
viruses protein and nucleic acid viroids RNA virusoids RNA prions proteins The Zombies of the Scientific Community Viruses and Other Acellular Infectious Agents Acellular Agents Viruses major cause of
Supplemental Information Dose Response Parameters for Gain of Function Pathogens
 Supplemental Information Dose Response Parameters for Gain of Function Pathogens Infection Dose-Response To quantify the likelihood of an individual or animal becoming infected from exposure to virus,
Supplemental Information Dose Response Parameters for Gain of Function Pathogens Infection Dose-Response To quantify the likelihood of an individual or animal becoming infected from exposure to virus,
Chair of Medical Biology, Microbiology, Virology, and Immunology STRUCTURE, CLASSIFICATION AND PHYSIOLOGY OF VIRUSES
 Chair of Medical Biology, Microbiology, Virology, and Immunology STRUCTURE, CLASSIFICATION AND PHYSIOLOGY OF VIRUSES Viruses are small obligate intracellular parasites, which by definition contain either
Chair of Medical Biology, Microbiology, Virology, and Immunology STRUCTURE, CLASSIFICATION AND PHYSIOLOGY OF VIRUSES Viruses are small obligate intracellular parasites, which by definition contain either
An outbreak of a respiratory infection of multi-agents occurred in poultry flocks in Tripoli, Libya
 An outbreak of a respiratory infection of multi-agents occurred in poultry flocks in Tripoli, Libya S.O. Al-GARIB*, A. A. ASHEG, A. KAMMON and M.A. HAMID Department of Poultry Disease, Faculty of Veterinary
An outbreak of a respiratory infection of multi-agents occurred in poultry flocks in Tripoli, Libya S.O. Al-GARIB*, A. A. ASHEG, A. KAMMON and M.A. HAMID Department of Poultry Disease, Faculty of Veterinary
Supplementary materials
 Supplementary materials Chemical library from ChemBridge 50,240 structurally diverse small molecule compounds dissolved in DMSO Hits Controls: No virus added μ Primary screening at 20 g/ml of compounds
Supplementary materials Chemical library from ChemBridge 50,240 structurally diverse small molecule compounds dissolved in DMSO Hits Controls: No virus added μ Primary screening at 20 g/ml of compounds
Supporting Information
 Supporting Information Valkenburg et al. 10.1073/pnas.1403684111 SI Materials and Methods ELISA and Microneutralization. Sera were treated with Receptor Destroying Enzyme II (RDE II, Accurate) before ELISA
Supporting Information Valkenburg et al. 10.1073/pnas.1403684111 SI Materials and Methods ELISA and Microneutralization. Sera were treated with Receptor Destroying Enzyme II (RDE II, Accurate) before ELISA
Cristina Cassetti, Ph.D.
 NIAID Extramural Research Update: Recombinant Influenza Viruses and Biosafety Cristina Cassetti, Ph.D. Influenza Program Officer Division of Microbiology and Infectious Diseases NIAID Influenza virus DMID
NIAID Extramural Research Update: Recombinant Influenza Viruses and Biosafety Cristina Cassetti, Ph.D. Influenza Program Officer Division of Microbiology and Infectious Diseases NIAID Influenza virus DMID
HIV-1 Virus-like Particle Budding Assay Nathan H Vande Burgt, Luis J Cocka * and Paul Bates
 HIV-1 Virus-like Particle Budding Assay Nathan H Vande Burgt, Luis J Cocka * and Paul Bates Department of Microbiology, Perelman School of Medicine at the University of Pennsylvania, Philadelphia, USA
HIV-1 Virus-like Particle Budding Assay Nathan H Vande Burgt, Luis J Cocka * and Paul Bates Department of Microbiology, Perelman School of Medicine at the University of Pennsylvania, Philadelphia, USA
Ralf Wagner Paul-Ehrlich-Institut
 www.pei.de Other Assays for the Detection of Neuraminidase (NA)-Specific Antibodies Ralf Wagner Paul-Ehrlich-Institut Overview to presented assays Assay principle based on: Chemical substrates: Protein
www.pei.de Other Assays for the Detection of Neuraminidase (NA)-Specific Antibodies Ralf Wagner Paul-Ehrlich-Institut Overview to presented assays Assay principle based on: Chemical substrates: Protein
Evolution of influenza
 Evolution of influenza Today: 1. Global health impact of flu - why should we care? 2. - what are the components of the virus and how do they change? 3. Where does influenza come from? - are there animal
Evolution of influenza Today: 1. Global health impact of flu - why should we care? 2. - what are the components of the virus and how do they change? 3. Where does influenza come from? - are there animal
Update August Questions & Answers on Middle East Respiratory Syndrome Coronavirus (MERS CoV)
") Update August 2014 - Questions & Answers on Middle East Respiratory Syndrome Coronavirus (MERS CoV) What is MERS-CoV? MERS-CoV is a coronavirus (CoV) which causes Middle East Respiratory Syndrome (MERS),
Update August 2014 - Questions & Answers on Middle East Respiratory Syndrome Coronavirus (MERS CoV) What is MERS-CoV? MERS-CoV is a coronavirus (CoV) which causes Middle East Respiratory Syndrome (MERS),
Suggestions to prevent / control Respiratory Disease Complex in poultry
 Suggestions to prevent / control Respiratory Disease Complex in poultry Dr. J. L. Vegad Adviser Phoenix Group 201/15, Gorakhpur, Jabalpur - 482001 Introduction Today, respiratory disease complex has emerged
Suggestions to prevent / control Respiratory Disease Complex in poultry Dr. J. L. Vegad Adviser Phoenix Group 201/15, Gorakhpur, Jabalpur - 482001 Introduction Today, respiratory disease complex has emerged
Lab 3: Pathogenesis of Virus Infections & Pattern 450 MIC PRACTICAL PART SECTION (30397) MIC AMAL ALGHAMDI 1
 MIC AMAL ALGHAMDI 1") Lab 3: Pathogenesis of Virus Infections & Pattern 450 MIC PRACTICAL PART SECTION (30397) 2018 450 MIC AMAL ALGHAMDI 1 Learning Outcomes The pathogenesis of viral infection The viral disease pattern Specific
Lab 3: Pathogenesis of Virus Infections & Pattern 450 MIC PRACTICAL PART SECTION (30397) 2018 450 MIC AMAL ALGHAMDI 1 Learning Outcomes The pathogenesis of viral infection The viral disease pattern Specific
Structural vs. nonstructural proteins
 Why would you want to study proteins associated with viruses or virus infection? Receptors Mechanism of uncoating How is gene expression carried out, exclusively by viral enzymes? Gene expression phases?
Why would you want to study proteins associated with viruses or virus infection? Receptors Mechanism of uncoating How is gene expression carried out, exclusively by viral enzymes? Gene expression phases?
2009 H1N1 Influenza ( Swine Flu ) Hemagglutinin ELISA kit
 Hemagglutinin ELISA kit") 2009 H1N1 Influenza ( Swine Flu ) Hemagglutinin ELISA kit Catalog Number : SEK001 To achieve the best assay results, this manual must be read carefully before using this product and the assay is run as
2009 H1N1 Influenza ( Swine Flu ) Hemagglutinin ELISA kit Catalog Number : SEK001 To achieve the best assay results, this manual must be read carefully before using this product and the assay is run as
(;[rowth Charaeteristies of Influenza Virus Type C in Avian Hosts
 Archives of Virology 58, 349--353 (1978) Archives of Virology by Springer-Verlag 1978 (;[rowth Charaeteristies of Influena Virus Type C in Avian Hosts Brief Report By M ~R A~N D. AUSTIn, A. S. MONTO, and
Archives of Virology 58, 349--353 (1978) Archives of Virology by Springer-Verlag 1978 (;[rowth Charaeteristies of Influena Virus Type C in Avian Hosts Brief Report By M ~R A~N D. AUSTIn, A. S. MONTO, and
7.012 Problem Set 6 Solutions
 Name Section 7.012 Problem Set 6 Solutions Question 1 The viral family Orthomyxoviridae contains the influenza A, B and C viruses. These viruses have a (-)ss RNA genome surrounded by a capsid composed
Name Section 7.012 Problem Set 6 Solutions Question 1 The viral family Orthomyxoviridae contains the influenza A, B and C viruses. These viruses have a (-)ss RNA genome surrounded by a capsid composed
Introduction. In the past 15 years, several technological advancements have open new perspectives and applications in the field of vaccinology.
 Introduction In the past 15 years, several technological advancements have open new perspectives and applications in the field of vaccinology. - Genomics: fasten antigen discovery for complex pathogens
Introduction In the past 15 years, several technological advancements have open new perspectives and applications in the field of vaccinology. - Genomics: fasten antigen discovery for complex pathogens
Study of rabies virus by Differential Scanning Calorimetry (DSC)
") Study of rabies virus by Differential Scanning Calorimetry (DSC) Frédéric Gréco Analytical Research & Developpement Biochemical & Biophysical Characterization 26 September 2016 1 The rabies virus (RABV)
Study of rabies virus by Differential Scanning Calorimetry (DSC) Frédéric Gréco Analytical Research & Developpement Biochemical & Biophysical Characterization 26 September 2016 1 The rabies virus (RABV)
Gene Vaccine Dr. Sina Soleimani
 Gene Vaccine Dr. Sina Soleimani Human Viral Vaccines Quality Control Laboratory (HVVQC) Titles 1. A short Introduction of Vaccine History 2. First Lineage of Vaccines 3. Second Lineage of Vaccines 3. New
Gene Vaccine Dr. Sina Soleimani Human Viral Vaccines Quality Control Laboratory (HVVQC) Titles 1. A short Introduction of Vaccine History 2. First Lineage of Vaccines 3. Second Lineage of Vaccines 3. New
Chapter 13 Viruses, Viroids, and Prions. Biology 1009 Microbiology Johnson-Summer 2003
 Chapter 13 Viruses, Viroids, and Prions Biology 1009 Microbiology Johnson-Summer 2003 Viruses Virology-study of viruses Characteristics: acellular obligate intracellular parasites no ribosomes or means
Chapter 13 Viruses, Viroids, and Prions Biology 1009 Microbiology Johnson-Summer 2003 Viruses Virology-study of viruses Characteristics: acellular obligate intracellular parasites no ribosomes or means
G. W. WOOD J. C. MUSKETT and D. H. THORNTON MAFF, Central Veterinary Laboratory, New Haw, Weybridge, Surrey, U.K.
 J. Comp. Path. 1986 vol. 96 OBSERVATIONS ON THE ABILITY OF AVIAN REOVIRUS VACCINMATION OF HENS TO PROTECT THEIR PROGENY AGAINST THE EFFECTS OF CHALLENGE WITH HOMOLOGOUS AND HETEROLOGOUS STRAINS By G. W.
J. Comp. Path. 1986 vol. 96 OBSERVATIONS ON THE ABILITY OF AVIAN REOVIRUS VACCINMATION OF HENS TO PROTECT THEIR PROGENY AGAINST THE EFFECTS OF CHALLENGE WITH HOMOLOGOUS AND HETEROLOGOUS STRAINS By G. W.
Orthomyxoviridae and Paramyxoviridae. Lecture in Microbiology for medical and dental medical students
 Orthomyxoviridae and Paramyxoviridae Lecture in Microbiology for medical and dental medical students Orthomyxoviridae and Paramyxoviridae are ss RNA containng viruses Insert Table 25.1 RNA viruses 2 SIZE
Orthomyxoviridae and Paramyxoviridae Lecture in Microbiology for medical and dental medical students Orthomyxoviridae and Paramyxoviridae are ss RNA containng viruses Insert Table 25.1 RNA viruses 2 SIZE
Dr. Ahmed K. Ali Attachment and entry of viruses into cells
 Lec. 6 Dr. Ahmed K. Ali Attachment and entry of viruses into cells The aim of a virus is to replicate itself, and in order to achieve this aim it needs to enter a host cell, make copies of itself and
Lec. 6 Dr. Ahmed K. Ali Attachment and entry of viruses into cells The aim of a virus is to replicate itself, and in order to achieve this aim it needs to enter a host cell, make copies of itself and
MOLECULAR DIVERSITY, EVOLUTIONARY TRENDS, AND MUTATION RATES IN AVIAN CORONAVIRUS INFECTIOUS BRONCHITIS VIRUS ENID THANNA MCKINLEY
 MOLECULAR DIVERSITY, EVOLUTIONARY TRENDS, AND MUTATION RATES IN AVIAN CORONAVIRUS INFECTIOUS BRONCHITIS VIRUS by ENID THANNA MCKINLEY (Under the Direction of Mark W. Jackwood) ABSTRACT Live attenuated
MOLECULAR DIVERSITY, EVOLUTIONARY TRENDS, AND MUTATION RATES IN AVIAN CORONAVIRUS INFECTIOUS BRONCHITIS VIRUS by ENID THANNA MCKINLEY (Under the Direction of Mark W. Jackwood) ABSTRACT Live attenuated
636.09: : :614.91:636.5
 636.09:616.233-002:578.834:614.91:636.5.,..,. : ( ), ( ),,,.,.,,,, [1].,,.. ,,,.,,, [4].,,. :,.. ( ),,,., 7 45. 30-60-., 105 [2,3].,, . ( ) 7. 1930, A. Schalk i M.Hawn 1931., 1933. L. Buchelli i C. Brandly.
636.09:616.233-002:578.834:614.91:636.5.,..,. : ( ), ( ),,,.,.,,,, [1].,,.. ,,,.,,, [4].,,. :,.. ( ),,,., 7 45. 30-60-., 105 [2,3].,, . ( ) 7. 1930, A. Schalk i M.Hawn 1931., 1933. L. Buchelli i C. Brandly.
VIROLOGY. Engineering Viral Genomes: Retrovirus Vectors
 VIROLOGY Engineering Viral Genomes: Retrovirus Vectors Viral vectors Retrovirus replicative cycle Most mammalian retroviruses use trna PRO, trna Lys3, trna Lys1,2 The partially unfolded trna is annealed
VIROLOGY Engineering Viral Genomes: Retrovirus Vectors Viral vectors Retrovirus replicative cycle Most mammalian retroviruses use trna PRO, trna Lys3, trna Lys1,2 The partially unfolded trna is annealed
Practice Problems 8. a) What do we define as a beneficial or advantageous mutation to the virus? Why?
 What do we define as a beneficial or advantageous mutation to the virus? Why?") Life Sciences 1a Practice Problems 8 1. You have two strains of HIV one is a wild type strain of HIV and the second has acquired a mutation in the gene encoding the protease. This mutation has a dual effect
Life Sciences 1a Practice Problems 8 1. You have two strains of HIV one is a wild type strain of HIV and the second has acquired a mutation in the gene encoding the protease. This mutation has a dual effect
The Infectious Cycle. Lecture 2 Biology W3310/4310 Virology Spring You know my methods, Watson --SIR ARTHUR CONAN DOYLE
 The Infectious Cycle Lecture 2 Biology W3310/4310 Virology Spring 2016 You know my methods, Watson --SIR ARTHUR CONAN DOYLE The Infectious Cycle Virologists divide the infectious cycle into steps to facilitate
The Infectious Cycle Lecture 2 Biology W3310/4310 Virology Spring 2016 You know my methods, Watson --SIR ARTHUR CONAN DOYLE The Infectious Cycle Virologists divide the infectious cycle into steps to facilitate
Lecture 11. Immunology and disease: parasite antigenic diversity
 Lecture 11 Immunology and disease: parasite antigenic diversity RNAi interference video and tutorial (you are responsible for this material, so check it out.) http://www.pbs.org/wgbh/nova/sciencenow/3210/02.html
Lecture 11 Immunology and disease: parasite antigenic diversity RNAi interference video and tutorial (you are responsible for this material, so check it out.) http://www.pbs.org/wgbh/nova/sciencenow/3210/02.html
PARAMYXOVIRUS FAMILY properties of attachment protein
 PARAMYXOVIRUS FAMILY properties of attachment protein GENUS GLYCOPROTEINS TYPICAL MEMBERS Paramyxovirus genus Rubulavirus Genus HN, F HN, F HPIV1, HPIV3 HPIV2, HPIV4 mumps virus Morbillivirus genus Pneumovirus
PARAMYXOVIRUS FAMILY properties of attachment protein GENUS GLYCOPROTEINS TYPICAL MEMBERS Paramyxovirus genus Rubulavirus Genus HN, F HN, F HPIV1, HPIV3 HPIV2, HPIV4 mumps virus Morbillivirus genus Pneumovirus
Application of Reverse Genetics to Influenza Vaccine Development
 NIAID Application of Reverse Genetics to Influenza Vaccine Development Kanta Subbarao Laboratory of Infectious Diseases NIAID, NIH Licensed Vaccines for Influenza Principle: Induction of a protective
NIAID Application of Reverse Genetics to Influenza Vaccine Development Kanta Subbarao Laboratory of Infectious Diseases NIAID, NIH Licensed Vaccines for Influenza Principle: Induction of a protective
Overview: Chapter 19 Viruses: A Borrowed Life
 Overview: Chapter 19 Viruses: A Borrowed Life Viruses called bacteriophages can infect and set in motion a genetic takeover of bacteria, such as Escherichia coli Viruses lead a kind of borrowed life between
Overview: Chapter 19 Viruses: A Borrowed Life Viruses called bacteriophages can infect and set in motion a genetic takeover of bacteria, such as Escherichia coli Viruses lead a kind of borrowed life between
Chapter 5. Virus isolation and identification of measles and rubella in cell culture
 Chapter 5. Virus isolation and identification of measles and rubella in cell culture In this chapter: 5.1. Recommended cell line for measles and rubella virus isolation 5.2. Propagation of Vero/hSLAM cells
Chapter 5. Virus isolation and identification of measles and rubella in cell culture In this chapter: 5.1. Recommended cell line for measles and rubella virus isolation 5.2. Propagation of Vero/hSLAM cells
LESSON 4.4 WORKBOOK. How viruses make us sick: Viral Replication
 DEFINITIONS OF TERMS Eukaryotic: Non-bacterial cell type (bacteria are prokaryotes).. LESSON 4.4 WORKBOOK How viruses make us sick: Viral Replication This lesson extends the principles we learned in Unit
DEFINITIONS OF TERMS Eukaryotic: Non-bacterial cell type (bacteria are prokaryotes).. LESSON 4.4 WORKBOOK How viruses make us sick: Viral Replication This lesson extends the principles we learned in Unit
CHAPTER 4 RESULTS. showed that all three replicates had similar growth trends (Figure 4.1) (p<0.05; p=0.0000)
 (p<0.05; p=0.0000)") CHAPTER 4 RESULTS 4.1 Growth Characterization of C. vulgaris 4.1.1 Optical Density Growth study of Chlorella vulgaris based on optical density at 620 nm (OD 620 ) showed that all three replicates had similar
CHAPTER 4 RESULTS 4.1 Growth Characterization of C. vulgaris 4.1.1 Optical Density Growth study of Chlorella vulgaris based on optical density at 620 nm (OD 620 ) showed that all three replicates had similar
Multiplication of the V4 strain of Newcastle disease virus in Madin Derby bovine kidney cells
 Onderstepoort Journal of Veterinary Research, 69:201-206 (2002) Multiplication of the V4 strain of Newcastle disease virus in Madin Derby bovine kidney cells M. SAHLE1*, W.G. BURGESS2 and A. KIDANEMARIAM1
Onderstepoort Journal of Veterinary Research, 69:201-206 (2002) Multiplication of the V4 strain of Newcastle disease virus in Madin Derby bovine kidney cells M. SAHLE1*, W.G. BURGESS2 and A. KIDANEMARIAM1
Chapter 5. Virus isolation and identification of measles and rubella in cell culture
 Chapter 5. Virus isolation and identification of measles and rubella in cell culture In this chapter: 5.1 Recommended cell line for measles and rubella virus isolation 5.2 Propagation of Vero/hSLAM cells
Chapter 5. Virus isolation and identification of measles and rubella in cell culture In this chapter: 5.1 Recommended cell line for measles and rubella virus isolation 5.2 Propagation of Vero/hSLAM cells
Nucleic acid: singled stranded, double stranded, RNA, or DNA, linear or circular. Capsid: protein coat that is most of the mass of the virus.
 Viruses General Characteristics of Viruses 1. Depending on view may be regarded as exceptionally complex aggregates of nonliving chemicals or as exceptionally simple living microbes. 2. Contain a single
Viruses General Characteristics of Viruses 1. Depending on view may be regarded as exceptionally complex aggregates of nonliving chemicals or as exceptionally simple living microbes. 2. Contain a single
7.012 Quiz 3 Answers
 MIT Biology Department 7.012: Introductory Biology - Fall 2004 Instructors: Professor Eric Lander, Professor Robert A. Weinberg, Dr. Claudette Gardel Friday 11/12/04 7.012 Quiz 3 Answers A > 85 B 72-84
MIT Biology Department 7.012: Introductory Biology - Fall 2004 Instructors: Professor Eric Lander, Professor Robert A. Weinberg, Dr. Claudette Gardel Friday 11/12/04 7.012 Quiz 3 Answers A > 85 B 72-84
Chapter 18. Viral Genetics. AP Biology
 Chapter 18. Viral Genetics 2003-2004 1 A sense of size Comparing eukaryote bacterium virus 2 What is a virus? Is it alive? DNA or RNA enclosed in a protein coat Viruses are not cells Extremely tiny electron
Chapter 18. Viral Genetics 2003-2004 1 A sense of size Comparing eukaryote bacterium virus 2 What is a virus? Is it alive? DNA or RNA enclosed in a protein coat Viruses are not cells Extremely tiny electron
Comparison of lentiviruses pseudotyped with S proteins from coronaviruses and cell tropisms of porcine coronaviruses
 VIROLOGICA SINICA 2016, 31 (1): 49 56 DOI: 10.1007/s12250-015-3690-4 RESEARCH ARTICLE Comparison of lentiviruses pseudotyped with S proteins from coronaviruses and cell tropisms of porcine coronaviruses
VIROLOGICA SINICA 2016, 31 (1): 49 56 DOI: 10.1007/s12250-015-3690-4 RESEARCH ARTICLE Comparison of lentiviruses pseudotyped with S proteins from coronaviruses and cell tropisms of porcine coronaviruses
Characterization of an Avian Infectious Bronchitis Virus Isolated in China from Chickens with Nephritis
 J. Vet. Med. B 51, 147 152 (2004) Ó 2004 Blackwell Verlag, Berlin ISSN 0931 1793 Institute of Preventive Veterinary Medicine, Zhejiang University, Hangzhou, China Characterization of an Avian Infectious
J. Vet. Med. B 51, 147 152 (2004) Ó 2004 Blackwell Verlag, Berlin ISSN 0931 1793 Institute of Preventive Veterinary Medicine, Zhejiang University, Hangzhou, China Characterization of an Avian Infectious
Some living things are made of ONE cell, and are called. Other organisms are composed of many cells, and are called. (SEE PAGE 6)
") Section: 1.1 Question of the Day: Name: Review of Old Information: N/A New Information: We tend to only think of animals as living. However, there is a great diversity of organisms that we consider living
Section: 1.1 Question of the Day: Name: Review of Old Information: N/A New Information: We tend to only think of animals as living. However, there is a great diversity of organisms that we consider living
Supplementary Material
 Supplementary Material Nuclear import of purified HIV-1 Integrase. Integrase remains associated to the RTC throughout the infection process until provirus integration occurs and is therefore one likely
Supplementary Material Nuclear import of purified HIV-1 Integrase. Integrase remains associated to the RTC throughout the infection process until provirus integration occurs and is therefore one likely
AP Biology. Viral diseases Polio. Chapter 18. Smallpox. Influenza: 1918 epidemic. Emerging viruses. A sense of size
 Hepatitis Viral diseases Polio Chapter 18. Measles Viral Genetics Influenza: 1918 epidemic 30-40 million deaths world-wide Chicken pox Smallpox Eradicated in 1976 vaccinations ceased in 1980 at risk population?
Hepatitis Viral diseases Polio Chapter 18. Measles Viral Genetics Influenza: 1918 epidemic 30-40 million deaths world-wide Chicken pox Smallpox Eradicated in 1976 vaccinations ceased in 1980 at risk population?
The Coronavirus Spike Protein:
 General Introduction The Coronavirus Spike Protein: Mechanisms of Membrane Fusion and Virion Incorporation Berend Jan Bosch 1 General Introduction Printing by Wöhrmann Print Service, Zutphen Financial
General Introduction The Coronavirus Spike Protein: Mechanisms of Membrane Fusion and Virion Incorporation Berend Jan Bosch 1 General Introduction Printing by Wöhrmann Print Service, Zutphen Financial
Malik Sallam. Ola AL-juneidi. Ammar Ramadan. 0 P a g e
 1 Malik Sallam Ola AL-juneidi Ammar Ramadan 0 P a g e Today's lecture will be about viral upper respiratory tract infections. Those include: common cold, sinusitis, otitis, etc. Infections in the upper
1 Malik Sallam Ola AL-juneidi Ammar Ramadan 0 P a g e Today's lecture will be about viral upper respiratory tract infections. Those include: common cold, sinusitis, otitis, etc. Infections in the upper
Biotechnology-Based Vaccines. Dr. Aws Alshamsan Department of Pharmaceutics Office: AA87 Tel:
 Biotechnology-Based Vaccines Dr. Aws Alshamsan Department of Pharmaceutics Office: AA87 Tel: 4677363 aalshamsan@ksu.edu.sa Objectives of this lecture By the end of this lecture you will be able to: 1.
Biotechnology-Based Vaccines Dr. Aws Alshamsan Department of Pharmaceutics Office: AA87 Tel: 4677363 aalshamsan@ksu.edu.sa Objectives of this lecture By the end of this lecture you will be able to: 1.
HIV-1 p24 ELISA Pair Set Cat#: orb54951 (ELISA Manual)
") HIV-1 p24 ELISA Pair Set Cat#: orb54951 (ELISA Manual) BACKGROUND Human Immunodeficiency Virus ( HIV ) can be divided into two major types, HIV type 1 (HIV-1) and HIV type 2 (HIV-2). HIV-1 is related to
HIV-1 p24 ELISA Pair Set Cat#: orb54951 (ELISA Manual) BACKGROUND Human Immunodeficiency Virus ( HIV ) can be divided into two major types, HIV type 1 (HIV-1) and HIV type 2 (HIV-2). HIV-1 is related to
Variation in the HindlII Restriction Fragments of DNA from the Chinese Tian Tan Strain of Vaccinia Virus
 J. gen. irol. (1985), 66, 1819-1823. Printed in Great Britain 1819 Key words: vaccinia virus~vaccine~restriction Jragrnent variation ariation in the Hindl Restriction Fragments of DNA from the Chinese
J. gen. irol. (1985), 66, 1819-1823. Printed in Great Britain 1819 Key words: vaccinia virus~vaccine~restriction Jragrnent variation ariation in the Hindl Restriction Fragments of DNA from the Chinese
Figure S1. Schematic presentation of genomic replication of idsiv after transfection and infection. After transfection of idsiv plasmid DNA into 293T
 Figure S1. Schematic presentation of genomic replication of idsiv after transfection and infection. After transfection of idsiv plasmid DNA into 293T cells, the RNA genomes with all modifications are generated
Figure S1. Schematic presentation of genomic replication of idsiv after transfection and infection. After transfection of idsiv plasmid DNA into 293T cells, the RNA genomes with all modifications are generated
Fine Mapping of a cis-acting Sequence Element in Yellow Fever Virus RNA That Is Required for RNA Replication and Cyclization
 JOURNAL OF VIROLOGY, Feb. 2003, p. 2265 2270 Vol. 77, No. 3 0022-538X/03/$08.00 0 DOI: 10.1128/JVI.77.3.2265 2270.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved. Fine Mapping
JOURNAL OF VIROLOGY, Feb. 2003, p. 2265 2270 Vol. 77, No. 3 0022-538X/03/$08.00 0 DOI: 10.1128/JVI.77.3.2265 2270.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved. Fine Mapping