Ejaculate Investment in a Promiscuous Rodent, Peromyscus maniculatus: Effects of Population Density and Social Role
|
|
|
- Alisha Webster
- 5 years ago
- Views:
Transcription
1 Wilfrid Laurier University Scholars Laurier Biology Faculty Publications Biology 2006 Ejaculate Investment in a Promiscuous Rodent, Peromyscus maniculatus: Effects of Population Density and Social Role Tristan A.F. Long Wilfrid Laurier University, tlong@wlu.ca Robert Montgomerie Queen's University -- Kingston Follow this and additional works at: Recommended Citation Long, Tristan A.F. and Montgomerie, Robert, "Ejaculate Investment in a Promiscuous Rodent, Peromyscus maniculatus: Effects of Population Density and Social Role" (2006). Biology Faculty Publications This Article is brought to you for free and open access by the Biology at Scholars Laurier. It has been accepted for inclusion in Biology Faculty Publications by an authorized administrator of Scholars Laurier. For more information, please contact scholarscommons@wlu.ca.
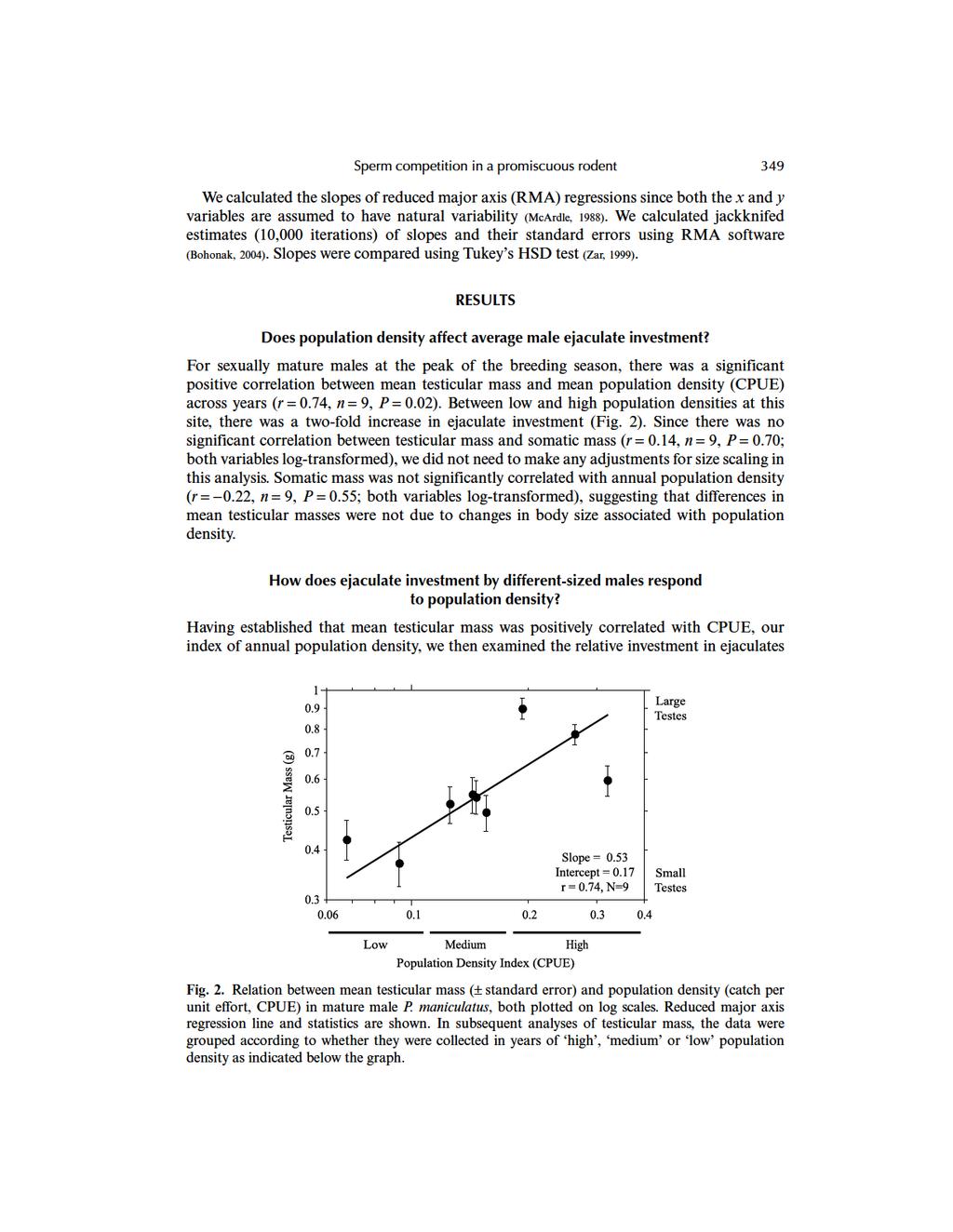
2 Evolutionary Ecology Research, 2006, 8: Ejaculate investment in a promiscuous rodent, Peromyscus maniculatus: effects of population density and social role Tristan A.F. Long* and Robert Montgomerie Department of Biology, Queen s University, Kingston, Ontario K7L 3N6, Canada ABSTRACT Questions: How does average male investment in ejaculates vary with changing population density (and thus with the risk of sperm competition) in a promiscuous species? Do individual male investment strategies vary with population density? Data studied: Total testicular mass, somatic mass and annual population density for wild-caught male deer mice, Peromyscus maniculatus, collected by snap-trapping over a 23-year period in Algonquin Provincial Park, Ontario, Canada. Search methods: We analysed the relation between mean testicular mass and mouse population densities across years. To investigate individual investment patterns, we compared the relation between total testicular mass and somatic mass among males for years differing in population density. Conclusions: Average investment in the testes was positively correlated with annual population density. An individual s investment in testes depended on both the abundance of rival males and on relative body size, a trait associated with social rank. Keywords: ejaculate investment, mating system, Peromyscus maniculatus, population density, sexual selection, sperm competition, testicular mass. INTRODUCTION Sperm competition is the post-copulatory competition between the ejaculates of rival males for the fertilization of a female s ova (Parker, 1970). This widespread phenomenon has influenced the evolution of a broad diversity of sexual behaviours, as well as anatomical and physiological traits (for reviews, see Smith, 1984; Birkhead and Møller, 1992, 1998). Sperm competition theory (Parker, 1990, 1998) predicts that when females mate with more than one male, and their ejaculates overlap temporally in the female s reproductive tract, selection will favour those male traits that improve the ability to secure paternity. Many interspecific comparative studies (e.g. Harcourt et al., 1981; Kenagy and Trombulak, 1986; Briskie, 1993; Hosken, 1997; Byrne et al., 2002) have found that relative testis size is frequently positively correlated with the potential for sperm * Author to whom all correspondence should be addressed. longt@biology.queensu.ca Consult the copyright statement on the inside front cover for non-commercial copying policies Tristan A.F. Long
3 346 Long and Montgomerie competition, supporting the hypothesis that there is an adaptive basis for variation in ejaculate investment. In many species, the opportunity for promiscuity (and thus sperm competition) is closely related to the distribution of sexually mature individuals, which in turn is influenced by the dispersion of resources in the environment (Emlen and Oring, 1977; Davies and Lundberg, 1984; Ostfeld, 1990). Thus, spatial or temporal variation in ecological conditions may shape the probability [or risk (Parker, 1998)] that a population s males experience sperm competition (Birkhead and Møller, 1992), and affect their reproductive tactics. For instance, a high local density of breeding adults can increase the opportunity for promiscuity due to a greater likelihood of interactions between the sexes (Emlen and Oring, 1977; Reynolds, 1996). This prediction has been supported by field studies that have found that the frequency of polyandry or extra-pair copulations and fertilizations is positively related to the local population density (Davies and Lundberg, 1984; Westneat and Sherman, 1997). As a result, the risk of sperm competition experienced by males may vary temporally, and selection should favour the evolution of males who can respond to cues in the social environment and adopt reproductive tactics that maximize fitness (Gage and Barnard, 1996). Most attempts to investigate male responses to variation in the risk of sperm competition have been laboratory-based, and have involved experimentally placing males into social situations that differ in the probability of sperm competition (e.g. Gage and Baker, 1991; Gage and Barnard, 1996; Wedell and Cook, 1999; Hosken and Ward, 2001). Few studies have examined male responses to varying social situations in free-living populations, where individuals experience natural levels of variation in the risk of sperm competition (but see Ribble and Millar, 1992; Stockley and Purvis, 1993; Pitcher and Stutchbury, 1998). The deer mouse, Peromyscus maniculatus (Rodentia; Muridae), is a good model species in which to study whether the response of males to changing socio-sexual conditions is adaptive with respect to the risk of sperm competition. Deer mice have a polygynous or promiscuous mating system (Wolff, 1989; Ribble and Millar, 1996) and lack many of the characteristic traits of monogamy [e.g. sexual monomorphism, small litter size, slow maturation, extensive paternal behaviour (Dewsbury, 1981a)]. Moreover, several ecological and social factors probably alter the actual intensity of their promiscuity (Wolff, 1989). In deer mice, the activity patterns of mature males are closely tied to those of mature females (Metzgar, 1979), and males will maintain home ranges, at least in part, to obtain access to females in reproductive condition (Wolff, 1985; Ostfeld, 1990). The ability to access these females will often depend on the local population density, which normally varies from 1 to 25 individuals per hectare (Banfield, 1974). At high densities, socially dominant males are able to monopolize the home ranges of several females (Mihok, 1979; Wolff and Cicirello, 1990), simultaneously displacing subordinate males [who become transient, and have limited exposure to mature females (Mihok, 1979)]. When population densities are low, neighbouring females no longer occupy contiguous home ranges (Wolff, 1985), and males generally interact with fewer females (Mihok, 1979). Thus, in free-living populations of deer mice, the density of breeding individuals has the potential to dramatically influence the amount of intrasexual competition, and thus the risk of sperm competition, that is faced by males. Studies of wild deer mouse populations have also provided indirect evidence that the local risk of sperm competition can be quite variable. Genetic parentage analyses have shown that between 10% and 43% of litters are sired by multiple males (Birdsall and Nash, 1973; Merrit and Wu, 1975; Ribble and Millar, 1996), and estimates of testis size also vary dramatically between populations (Kenagy and Trombulak, 1986; Ribble and Millar, 1992). Unfortunately, none of these
4 Sperm competition in a promiscuous rodent 347 studies report breeding population densities, so it is impossible to determine whether differences in the risk of sperm competition associated with population density contributed to any of this variation. Evolutionarily stable strategy (ESS) modelling of sperm competition was used by Parker (1990) to predict how a male s investment in his ejaculate should facultatively change in response to the overall risk of sperm competition. In this model, males should increase their expenditure on sperm in direct proportion to the risk of sperm competition to prevent loss of paternity to rival males. We tested this prediction using data collected from adult males in a free-living population of P. maniculatus whose breeding season population density fluctuated almost five-fold over a 23-year period (Fig. 1), and thus has potentially experienced much variation in sperm competition risk. METHODS Study area and field methods We used data obtained from a P. maniculatus population located in the vicinity of Lake Sasajewun, Algonquin Provincial Park, Ontario, Canada (48 30 N, W) that was studied as part of a larger investigation of population dynamics in small mammals (Fryxell et al., 1998). During May September from 1965 to 1987, deer mice were collected during bimonthly trapping sessions that usually spanned three to four consecutive nights. Trapping took place along trap-lines that consisted of stations located at 10-m intervals in stands of hardwood forest dominated by sugar maple, Acer saccharum. At each station, either one or two snap-traps (Woodstream Museum Special or Victor Four-Ways traps) were deployed. Traps were baited with a mixture of rolled oats and peanut butter, and set before sunset. The following morning, traps were checked and dead mice were taken to the laboratory for measurement. Male mice were weighed to the nearest 0.1 g, dissected, and the length and width of the left testis measured to the nearest millimetre. As snap-traps collected both mature and immature mice, it was necessary to differentiate between them, as only the former are relevant for this study of sperm competition. Sexually mature mice typically have testes more than 7 mm long (Jameson, 1953; Gram et al., 1982), so we analysed data only from males that met this criterion. To ensure that we studied only males in full breeding condition, we restricted our analyses to mice collected on days of the year 165 through 215, corresponding to the peak breeding period of May and early June in this (J.B. Falls, personal communication) and other (Beer and MacLeod, 1966; Sadleir, 1974) deer mouse populations. Additionally, we used data only from years where at least 10 sexually mature males had been captured during the peak breeding period, to ensure that the calculated mean values would be reasonably accurate. In total, 124 mice from nine different years were used in the analyses (Fig. 1). Estimates of ejaculate investment Using the testis length (cm) and width (cm) of mature males, we employed Møller s (1991) corrected formula to calculate the combined mass (g) of both testes as [ g π (radius of testis width) 2 radius of testis length], and used this measure (hereafter called testicular mass ) as an index of energetic investment in ejaculates. It is generally accepted that sperm production capacity is correlated with the size of the testes, since the number of

5
6
7 350 Long and Montgomerie by males within years at different annual mean population densities. To do this, we categorized years as having a high, medium or low mouse population density, such that each category contained a roughly equal number of years (see Fig. 2), and then performed an analysis of covariance on testicular mass, with somatic mass and population density category as predictor variables. This analysis revealed a significant interaction between the two predictors (Table 1). Thus, the relation between testicular mass and somatic mass varied significantly between population density categories. We then examined the relation between testicular mass and somatic mass for groups of individual adult males that were collected in years with different population densities (Fig. 3). Consistent with the results of analysis of covariance (Table 1), the regression between testicular mass and somatic mass varied with population density category. In addition, relative testicular mass, expressed as a percentage of total body mass, was significantly larger at high density (3.96%) than at low density (1.95%; t-test, t 72 = 6.4, P < ; Table 2). Moreover, at high densities, there was a negative correlation between testicular mass and soma mass (Fig. 3a), while at low densities there was a strong positive correlation (Fig. 3c). Males collected at medium population densities exhibited an intermediate, positive correlation between their testicular and somatic masses (Fig. 3b). The slopes of the RMA regressions from all three population density categories were significantly different from 0 (all t-statistics > 4.99, all P < 0.05, randomization tests). Furthermore, the slopes of both the high and low, but not the medium, population density regressions were significantly different from isometry (i.e. slope = 1), but in opposite directions. DISCUSSION Does population density affect average male ejaculate investment? In the P. maniculatus population that we studied, the average mass of a male s testes during the peak of the breeding season was positively correlated with the local population density (Fig. 2). This is consistent with Parker s (1990) prediction that males should invest more resources in their ejaculates as the risk of sperm competition rises. Since sperm production is often a direct function of the size of the testes (e.g. Hoditscheck and Best, 1983), building larger sperm-producing organs should enhance a male s ability to transfer large numbers of sperm during copulation, and ultimately secure more paternity following post-copulatory competition between ejaculates (Parker, 1998). In a comparative study of testis size in northern populations of P. maniculatus, Ribble and Millar (1992) found considerable variation in both body and testis length, some of which was attributed to differences in the length of the breeding season from site to site. They Table 1. Results of analysis of covariance comparing the testicular mass of mature P. maniculatus males collected at high, medium and low population densities (F 5,123 = 18.8, P < 0.001, R 2 = 0.44) Source of variation SS d.f. F P Somatic mass (g) , Population density category , <0.001 Somatic mass population density category ,
8
9 352 Long and Montgomerie As a result, males would face a greater risk of sperm competition in populations located in warmer regions compared with those in colder regions (Ribble and Millar, 1992). The results of our study are consistent with this hypothesis that intraspecific variation in testicular mass is associated with differences in the likelihood of sperm competition. When population density was high (increased risk of sperm competition), males had testes that were, on average, twice as large as those of males trapped at relatively low density. An alternative interpretation of the observed pattern of testes investment is that males are responding to some factor other than risk of sperm competition, such as food availability that covaries with population density. Thus, males might produce large testes only if there is ample food available. Although we do not have data on annual food availability, the non-significant negative relation between somatic mass and population density suggests that, if anything, mice were slightly larger in low-density years. Additionally, our interpretation has focused mainly on potential gains in sperm production associated with larger testicular mass, while there may be other testicular products, such as testosterone, whose production is also correlated with organ size (e.g. Schulte-Hostedde et al., 2003; but see Joyce et al., 1993, for an exception). Although we cannot address these potential covariates with the data available, the relationship between testis size, hormone levels and sperm production at different population densities presents an interesting subject for future research. How does ejaculate investment by different-sized males respond to population density? In this study, we make the assumption that male social dominance rank in P. maniculatus is positively correlated with somatic mass. Dominance ranks in male deer mice are rapidly established through antagonistic encounters, and remain stable over time (Dewsbury, 1981b). In laboratory-reared deer mice, heavier males are typically dominant over lighter males in intrasexual aggressive interactions, and dominant males are approached more frequently by females, and complete more mounts, intromissions and ejaculations with females than do subdominant males (Dewsbury, 1979). Ultimately, dominant males are able to secure privileged, though not exclusive, access to females (Dewsbury, 1981b, p. 890). Based on these observations, it appears that somatic mass is a reasonable index of social dominance, and can be used to study whether ejaculate investment strategies of males of different social rank vary with population density. The results of our analyses on the relation between ejaculate investment and sperm competition risk (Fig. 3) are in accord with Parker s (1990) ESS predictions for a loaded raffle fertilization system, wherein some males (in this case, large, dominant ones) are favoured over others (i.e. small males) with respect to sperm competition. The negative correlation between testicular mass and somatic mass among individuals at high population density (Fig. 3a) indicates that smaller males invest relatively more in their ejaculates than do larger (presumably higher-ranked) males. When population densities were low, testicular mass scaled positively with somatic mass (Figs. 3b and 3c). The differences in the relation between investment in ejaculates by males of different sizes collected in years with different population densities presumably reflects how male reproductive tactics are influenced by relative access to females. In the wild, subordinate males typically have little contact with mature females at high population density (Mihok, 1979), and are predicted to invest more in ejaculates (Parker, 1990) to maximize the impact of their limited mating opportunities. Conversely, at low population density when access to females is more evenly distributed across
10 Sperm competition in a promiscuous rodent 353 males, dominance rank becomes less important, and subordinate males are not predicted to invest heavily in their ejaculates (Parker, 1990). When females are distantly spaced, the only way in which a male may enhance his individual reproductive success is through long, extra-territorial excursions to find other potential mates (Mihok, 1979; Wolff and Cicirello, 1990). It is notable that under low-density conditions, there was a positive allometric relationship between total testicular mass and somatic mass in our population (Fig. 3c), suggesting that larger males invested relatively more heavily in their ejaculates so that they could capitalize on these encounters with breeding females. Finally, if the sperm competition raffle had been fair rather than loaded (Parker, 1990), there should have been no difference in the relative ejaculate investment made by males of all sizes at different population densities. The results of our analyses do not support that hypothesis. A previous interspecific analysis of the correlation between testis and soma sizes in mammals (Stockley and Purvis, 1993) found that small males in continuous-breeding species (i.e. those with high risk of sperm competition) had disproportionately larger testes than did seasonally breeding species (with lower risk of sperm competition), where their testicular and somatic masses were positively correlated. Our results are consistent with these findings, as we observed that the relative investment in ejaculates by males of different sizes varied with the local risk of sperm competition (Fig. 3). Since the production of ejaculates is a non-trivial expense for P. maniculatus males (Dewsbury, 1982), we would expect that the costs of increased investment in the testes would be offset by the gains made in sperm competition contests. Dewsbury (1979, 1981b) observed that large (dominant) males often achieved a greater number of ejaculations with females than did small (subordinate) males. Despite this apparent difference in reproductive success, dominant males did not father a greater proportion of their mate s litters than did the subordinate males in some experiments (Table 2 in Dewsbury, 1979). Our findings suggest an explanation based in sperm competition theory: smaller males, with larger testes, are able to offset their copulatory disadvantage through a greater numerical representation of sperm in the female s reproductive tract. Based on the regression for the high population density category (Fig. 3a), a small male (17.0 g) would have had testes that were 2.4 times larger than a male that is much heavier (21.0 g). If the daily sperm production rate per gram of parenchyma for P. maniculatus is similar to that of Rattus norvegicus (see Møller, 1989), the smaller male s testes will produce more sperm per day than those of the larger male. Under high-density social conditions, this additional sperm production capacity may allow small males to compete effectively with larger, dominant males for paternity. Our research highlights some important issues that should be considered when undertaking future intra- and interspecific studies of sperm competition. It is common practice in interspecific studies to use a mean value of testis mass for each species (e.g. Harcourt et al., 1981; Kenagy and Trombulak, 1986). As we have shown, the ejaculate investment by P. maniculatus males depends on both population density and body size and cannot be readily characterized by the species mean value. If the calculation of mean testis mass is based on solitary studies, or on those with limited sampling, this could obviously lead to inaccurate estimates, and incorrect conclusions. Similarly, intraspecific studies may misconstrue the risk of sperm competition within a population. It was only through the extended temporal scale of this study that we were able to show that risk from sperm competition is not constant from year to year. Care should be used when combining data obtained from individuals collected from different populations and times.
11 354 Long and Montgomerie ACKNOWLEDGEMENTS We thank Dr J. Bruce Falls and E. Ann Falls who generously provided us with access to their long-term P. maniculatus data set; the many field biologists who trapped deer mice over the years; the staff of the Wildlife Research Station, Algonquin Provincial Park, for their support; and L. Nagel for helpful comments on the manuscript. T.A.F.L. was supported by an NSERC PGS-B scholarship and R.M. by a Killam Research Fellowship from the Canada Council. This research was funded by a Discovery Grant to R.M. from the Natural Sciences and Engineering Research Council of Canada. REFERENCES Amann, R.P Sperm production rates. In The Testis, Vol. 1 (A.D. Johnson, W.R. Gomes and N.L. Vandermark, eds.), pp New York: Academic Press. Banfield, A The Mammals of Canada. Toronto, ON: University of Toronto Press. Beer, J.R. and MacLeod, C.F Seasonal change in the prairie deer mouse. Am. Midl. Nat., 76: Birdsall, D.H. and Nash, D Occurrence of successful multiple insemination of females in natural populations of deer mice (Peromyscus maniculatus). Evolution, 27: Birkhead, T.R. and Møller, A.P Sperm Competition in Birds: Evolutionary Causes and Consequences. Toronto, ON: Academic Press. Birkhead, T.R. and Møller, A.P Sperm Competition and Sexual Selection. Toronto, ON: Academic Press. Bohonak, A.J Software for reduced major axis regression v San Diego, CA: San Diego State University (available at: Briskie, J.V Anatomical adaptations to sperm competition in Smith s longspurs and other polygynandrous passerines. Auk, 110: Byrne, P.G., Roberts, J.D. and Simmons, L.W Sperm competition selects for increased testes mass in Australian frogs. J. Evol. Biol., 15: Davies, N.B. and Lundberg, A Food distribution and a mating system in the Dunnock, Prunella modularis. J. Anim. Ecol., 53: Dewsbury, D.A Copulatory behavior of deer mice (Peromyscus maniculatus): II. A study of some factors regulating the fine structure of behavior. J. Comp. Physiol. Psychol., 93: Dewsbury, D.A. 1981a. An exercise in the prediction of monogamy in the field from laboratory data on 42 species of muroid rodents. Biologist, 63: Dewsbury, D.A. 1981b. Social dominance, copulatory behavior, and differential reproduction in deer mice (Peromyscus maniculatus). J. Comp. Physiol. Psychol., 95: Dewsbury, D.A Ejaculate cost and male choice. Am. Nat., 119: Emlen, S.T. and Oring L.W Ecology, sexual selection, and the evolution of mating systems. Science, 197: Fryxell, J.M., Falls, J.B., Falls, E.A. and Brooks, R.J Long-term dynamics of small-mammal populations in Ontario. Ecology, 79: Gage, A.R. and Barnard, C.J Male crickets increase sperm number in relation to competition and female size. Behav. Ecol. Sociobiol., 38: Gage, M.J.G. and Baker, R.R Ejaculate size varies with socio-sexual situation in an insect. Ecol. Entomol., 16: Gram, W.D., Heath, H.W., Wichman, H.A. and Lynch, G.R Geographic variation in Peromyscus leucopus: short-day induced reproductive regression and spontaneous recrudescence. Biol. Reprod., 27: Harcourt, A.H., Purvis, A. and Liles, L Sperm competition: mating system, not breeding season, affects testes size in Primates. Funct. Ecol., 9:
12 Sperm competition in a promiscuous rodent 355 Hoditscheck, B. and Best, T.L Reproductive biology of Ord s kangaroo rat (Dipodomys ordii). J. Mammal., 64: Hosken, D.J Sperm competition in bats. Proc. R. Soc. Lond. B, 264: Hosken, D.J. and Ward, P.I Experimental evidence for testis size evolution via sperm competition. Ecol. Lett., 4: Jameson, E.W., Jr Reproduction of deer mice (Peromyscus maniculatus and P. boylei) in the Sierra Nevada, California. J. Mammal., 34: Joyce, K.L., Porcelli, J. and Cooke, P.S Neonatal goitrogen treatment increases adult testis size and sperm production in the mouse. J. Androl., 14: Kenagy, G.J. and Trombulak, S.C Size and function of mammalian testes in relation to body size. J. Mammal., 67: Lunstra, D.D., Wise, T.H. and Ford, J.J Sertoli cells in the boar testis: changes during development and compensatory hypertrophy after hemicastration at different ages. Biol. Reprod., 86: McArdle, B.H The structural relationship: regression in biology. Can. J. Zool., 66: Merritt, R.B. and Wu, B.J On the quantification of promiscuity (or Promyscus maniculatus?). Evolution, 29: Metzgar, L.H Dispersion pattern in a Peromyscus population. J. Mammal., 60: Mihok, S Behaviour and demography of subartic Clethrionomys gapperi and Peromyscus maniculatus. Can. J. Zool., 57: Møller, A.P Ejaculate quality, testes size and sperm production in mammals. Funct. Ecol., 3: Møller, A.P Sperm competition, sperm depletion, paternal care and relative testis size in birds. Am. Nat., 137: Møller, A.P. and Birkhead, T.R Copulation behaviour in mammals: evidence that sperm competition is widespread. Biol. J. Linn. Soc., 38: Ostfeld, R.S The ecology of territoriality in small mammals. Trends Ecol. Evol., 5: Parker, G.A Sperm competition and its evolutionary consequences in the insects. Biol. Rev., 45: Parker, G.A Sperm competition games: raffles and roles. Proc. R. Soc. Lond. B, 242: Parker, G.A Sperm competition and the evolution of ejaculates: towards a theory base. In Sperm Competition and Sexual Selection (T.R. Birkhead and A.P. Møller, eds.), pp Toronto, ON: Academic Press. Peters, R.H The Ecological Implications of Body Size. New York: Cambridge University Press. Pitcher, T.E. and Stutchbury, J.M Latitudinal variation in testis size in six species of North American songbirds. Can. J. Zool., 76: Reynolds, J.D Animal breeding systems. Trends Ecol. Evol., 11: Ribble, D.O. and Millar, J.S Intraspecific variation in testes size among northern populations of Peromyscus. Funct. Ecol., 6: Ribble, D.O. and Millar, J.S The mating system of northern populations of Peromyscus maniculatus as revealed by radiotelemetry and DNA fingerprinting. Écoscience, 3: Sadleir, R.M.F.S The ecology of the deer mouse Peromyscus maniculatus in a coastal coniferous forest. II. Reproduction. Can. J. Zool., 52: SAS Institute Inc JMP Statistics and Graphics Guide v. 5. Cary, NC: SAS Institute Inc. Schulte-Hostedde, A.I., Millar, J.S. and Hickling, G.J Intraspecific variation in testis size of small mammals: implications for muscle mass. Can. J. Zool., 81: Smith, R.L Sperm Competition and the Evolution of Animal Mating Systems. Toronto, ON: Academic Press. Stockley, P. and Purvis, A Sperm competition in mammals: a comparative study of male roles and relative investment in sperm production. Funct. Ecol., 7:
13 356 Long and Montgomerie Wedell, N. and Cook, P.A Butterflies tailor their ejaculate in response to sperm competition risk and intensity. Proc. R. Soc. Lond. B, 266: Westneat, D.F. and Sherman, P.W Density and extra-pair fertilizations in birds: a comparative analysis. Behav. Ecol. Sociobiol., 41: Wolff, J.O The effect of density, food, and interspecific interference on home range size in Peromyscus leucopus and Peromyscus maniculatus. Can. J. Zool., 63: Wolff, J.O Social behaviour. In Advances in the Study of Peromyscus (G. Kirkland and J. Layne, eds.), pp Lubbock, TX: Texas Tech University Press. Wolff, J.O. and Cicirello, D.M Mobility versus territoriality: alternate reproductive strategies in white-footed mice. Anim. Behav., 39: Zar, J.H Biostatistical Analysis, 4th edn. Upper Saddle River, NJ: Prentice-Hall.
Genetic Patterns of Paternity and Testes Size in Mammals
 Genetic Patterns of Paternity and Testes Size in Mammals Carl D. Soulsbury* School of Biological Science, University of Bristol, Bristol, United Kingdom Abstract Background: Testes size is used as a proxy
Genetic Patterns of Paternity and Testes Size in Mammals Carl D. Soulsbury* School of Biological Science, University of Bristol, Bristol, United Kingdom Abstract Background: Testes size is used as a proxy
Sperm competition and the evolution of testes size in birds
 doi:10.1111/j.1420-9101.2004.00874.x Sperm competition and the evolution of testes size in birds T. E. PITCHER,*P.O.DUNN &L.A.WHITTINGHAM *Department of Zoology, University of Toronto, Toronto, Ontario,
doi:10.1111/j.1420-9101.2004.00874.x Sperm competition and the evolution of testes size in birds T. E. PITCHER,*P.O.DUNN &L.A.WHITTINGHAM *Department of Zoology, University of Toronto, Toronto, Ontario,
Sexual Selection. Male and female. Lukas Schärer. Evolutionary Biology Zoological Institute University of Basel
 Sexual Selection Lukas Schärer! Evolutionary Biology Zoological Institute University of Basel 1 8.11.2017 Advanced-level Evolutionary Biology Male and female 2 Summary: Sexual Selection defining sexual
Sexual Selection Lukas Schärer! Evolutionary Biology Zoological Institute University of Basel 1 8.11.2017 Advanced-level Evolutionary Biology Male and female 2 Summary: Sexual Selection defining sexual
Anti-predator behavior: Group defense - Many eyes - Selfish herd - Dilution effects - Tonic immobility. Dispersal:
 Note: These are just the topics that have been covered since Exam 2. The exams are cumulative, so all the material that we have covered in this course can be included in the Final. However, there will
Note: These are just the topics that have been covered since Exam 2. The exams are cumulative, so all the material that we have covered in this course can be included in the Final. However, there will
Sexual Selection and the Evolution of Brain Size in Primates
 Sexual Selection and the Evolution of Brain Size in Primates Michael A. Schillaci* Department of Anthropology, University of Toronto, Toronto, Ontario, Canada Reproductive competition among males has long
Sexual Selection and the Evolution of Brain Size in Primates Michael A. Schillaci* Department of Anthropology, University of Toronto, Toronto, Ontario, Canada Reproductive competition among males has long
MECHANISMS OF SPERM COMPETITION: TESTING THE FAIR RAFFLE
 Evolution, 8(8), 04, pp. 1846 181 MECHANISMS OF SPERM COMPETITION: TESTING THE FAIR RAFFLE BRYAN D. NEFF 1,2 AND LINDI M. WAHL 3,4 1 Department of Biology, University of Western Ontario, London, Ontario
Evolution, 8(8), 04, pp. 1846 181 MECHANISMS OF SPERM COMPETITION: TESTING THE FAIR RAFFLE BRYAN D. NEFF 1,2 AND LINDI M. WAHL 3,4 1 Department of Biology, University of Western Ontario, London, Ontario
The Operational Sex Ratio, the Potential Reproductive Rate, and the Opportunity for Sexual Selection Stephen M. Shuster
 The Operational Sex Ratio, the Potential Reproductive Rate, and the Opportunity for Sexual Selection Stephen M. Shuster Stephen M. Shuster Northern Arizona University Objectives. Definitions of OSR/PRR.
The Operational Sex Ratio, the Potential Reproductive Rate, and the Opportunity for Sexual Selection Stephen M. Shuster Stephen M. Shuster Northern Arizona University Objectives. Definitions of OSR/PRR.
Effect of Fluorescent Powder Marking of Females on Mate Choice by Male White-Footed Mice (Peromyscus leucopus)
") Effect of Fluorescent Powder Marking of Females on Mate Choice by Male White-Footed Mice (Peromyscus leucopus) By: MATINA C. KALCOUNIS-RÜPPELL, AMANDA PATRICK, JOHN S. MILLAR Kalcounis-Rüppell, M. C.,
Effect of Fluorescent Powder Marking of Females on Mate Choice by Male White-Footed Mice (Peromyscus leucopus) By: MATINA C. KALCOUNIS-RÜPPELL, AMANDA PATRICK, JOHN S. MILLAR Kalcounis-Rüppell, M. C.,
FILM SESSIONS. Film session I - Behaviour of Communication (September 30)
") FILM SESSIONS Film session I - Behaviour of Communication (September 30) 1. Talking to Strangers (60 min.) 1 This film surveys some of the general communication signals used by animals from diverse taxa,
FILM SESSIONS Film session I - Behaviour of Communication (September 30) 1. Talking to Strangers (60 min.) 1 This film surveys some of the general communication signals used by animals from diverse taxa,
A Closer Look at the Breeding Season of the Kangaroo Rat Dipodomys heermanni
 A Closer Look at the Breeding Season of the Kangaroo Rat Dipodomys heermanni Brianna R. Lavelle Senior Project for the College of Science and Math Biological Sciences Department, California Polytechnic
A Closer Look at the Breeding Season of the Kangaroo Rat Dipodomys heermanni Brianna R. Lavelle Senior Project for the College of Science and Math Biological Sciences Department, California Polytechnic
Authors: Abstract: Keywords: Introduction
 Light or Dark Preference of Peromyscus leucopus in a Y-maze Choice Experiment Authors: Eleanor Shore, Jimmy Lovett *, and Dr. Karen McBee Abstract: Peromyscus leucopus, or white-footed mouse, is a small
Light or Dark Preference of Peromyscus leucopus in a Y-maze Choice Experiment Authors: Eleanor Shore, Jimmy Lovett *, and Dr. Karen McBee Abstract: Peromyscus leucopus, or white-footed mouse, is a small
Absence Makes the Adaptations Grow Fonder: Proportion of Time Apart from Partner, Male Sexual Psychology, and Sperm Competition in Humans
 Sperm competition in humans p. 1 Running Head: SPERM COMPETITION IN HUMANS [in press, Journal of Comparative Psychology, November 2006] Absence Makes the Adaptations Grow Fonder: Proportion of Time Apart
Sperm competition in humans p. 1 Running Head: SPERM COMPETITION IN HUMANS [in press, Journal of Comparative Psychology, November 2006] Absence Makes the Adaptations Grow Fonder: Proportion of Time Apart
Plasticity in reproductive effort of male dung flies. (Scatophaga stercoraria) as a response to larval density. P. STOCKLEY and N. J.
 as a response to larval density. P. STOCKLEY and N. J.") Functional Ecology 2001 Plasticity in reproductive effort of male dung flies Blackwell Science, Ltd (Scatophaga stercoraria) as a response to larval density P. STOCKLEY and N. J. SEAL Population Biology
Functional Ecology 2001 Plasticity in reproductive effort of male dung flies Blackwell Science, Ltd (Scatophaga stercoraria) as a response to larval density P. STOCKLEY and N. J. SEAL Population Biology
ESRM 350 Reproduction and Mating Systems
 ESRM 350 Reproduction and Mating Systems Autumn 2013 Life s but a walking shadow, a poor player That struts and frets his hour upon the stage And then is heard no more. - Macbeth, William Shakespeare Reproduction
ESRM 350 Reproduction and Mating Systems Autumn 2013 Life s but a walking shadow, a poor player That struts and frets his hour upon the stage And then is heard no more. - Macbeth, William Shakespeare Reproduction
The Relationship between Objective Sperm Competition Risk and Men s Copulatory Interest is. Moderated by Partner s Time Spent with Other Men
 Running head: SPERM COMPETITION 1 [in press, Human Nature, February 2013] The Relationship between Objective Sperm Competition Risk and Men s Copulatory Interest is Moderated by Partner s Time Spent with
Running head: SPERM COMPETITION 1 [in press, Human Nature, February 2013] The Relationship between Objective Sperm Competition Risk and Men s Copulatory Interest is Moderated by Partner s Time Spent with
Structure and allometry of genitalia in males and females of a social African ground squirrel with high polygynandry
 Structure and allometry of genitalia in males and females of a social African ground squirrel with high polygynandry M. B. Manjerovic 1, A. A. Kinahan 2,3, J. M. Waterman 1,2, N. C. Bennett 2 & P. W. Bateman
Structure and allometry of genitalia in males and females of a social African ground squirrel with high polygynandry M. B. Manjerovic 1, A. A. Kinahan 2,3, J. M. Waterman 1,2, N. C. Bennett 2 & P. W. Bateman
Sperm competition in mammals
 Human Fertility, June 2004, 7(2): 91 97 Sperm competition in mammals P. STOCKLEY Department of Clinical Veterinary Science, Faculty of Veterinary Science, University of Liverpool, Leahurst, Neston, UK
Human Fertility, June 2004, 7(2): 91 97 Sperm competition in mammals P. STOCKLEY Department of Clinical Veterinary Science, Faculty of Veterinary Science, University of Liverpool, Leahurst, Neston, UK
Do male fallow deer (Dama dama L) guard female before and after mating.
 guard female before and after mating.") Research Article Animal Science Journal, 3(1): 01-05, 2012 Available online at www.isisn.org ISISnet Publishers Print ISSN: 2220-959 Online ISSN: 2220-9557 Do male fallow deer (Dama dama L) guard female
Research Article Animal Science Journal, 3(1): 01-05, 2012 Available online at www.isisn.org ISISnet Publishers Print ISSN: 2220-959 Online ISSN: 2220-9557 Do male fallow deer (Dama dama L) guard female
Male parental effort and paternity in a variable mating system
 Anim. Behav., 1998, 55, 69 64 Male parental effort and paternity in a variable mating system LINDA A. WHITTINGHAM & PETER O. DUNN Evolutionary Ecology Group, Division of Botany and Zoology, Australian
Anim. Behav., 1998, 55, 69 64 Male parental effort and paternity in a variable mating system LINDA A. WHITTINGHAM & PETER O. DUNN Evolutionary Ecology Group, Division of Botany and Zoology, Australian
Co-evolution of nuptial gift and female multiple mating resulting in diverse breeding systems
 Evolutionary Ecology Research, 2001, 3: 75 89 Co-evolution of nuptial gift and female multiple mating resulting in diverse breeding systems Michio Kondoh* Centre for Ecological Research, Kyoto University,
Evolutionary Ecology Research, 2001, 3: 75 89 Co-evolution of nuptial gift and female multiple mating resulting in diverse breeding systems Michio Kondoh* Centre for Ecological Research, Kyoto University,
Polygyny in animals From Wikipedia, the free encyclopedia
 Page 1 of 6 Polygyny in animals From Wikipedia, the free encyclopedia Polygyny (/pəˈlɪdʒɪniː/; from Neo-Greek πολυγυνία from πολύ- poly- "many", and γυνή gyne "woman" or "wife") [1] is a mating system
Page 1 of 6 Polygyny in animals From Wikipedia, the free encyclopedia Polygyny (/pəˈlɪdʒɪniː/; from Neo-Greek πολυγυνία from πολύ- poly- "many", and γυνή gyne "woman" or "wife") [1] is a mating system
The reproductive choices of eavesdropping female black-capped chickadees, Poecile atricapillus
 Naturwissenschaften (2003) 90:577-582 DOI 10.1007/s00114-003-0479-3 SHORT COMMUNICATION Daniel J. Mennill Peter T. Boag Laurene M. Ratcliffe The reproductive choices of eavesdropping female black-capped
Naturwissenschaften (2003) 90:577-582 DOI 10.1007/s00114-003-0479-3 SHORT COMMUNICATION Daniel J. Mennill Peter T. Boag Laurene M. Ratcliffe The reproductive choices of eavesdropping female black-capped
Test Bank. Chapter 2. Abrams, Sexuality and Its Disorders SAGE Publishing, 2017
 Test Bank Chapter 2 1. Is the male female dichotomy an appropriate mode of defining gender? Why, or why not? Ans: The male female dichotomy is strongly embodied in most of cultures, religions, and even
Test Bank Chapter 2 1. Is the male female dichotomy an appropriate mode of defining gender? Why, or why not? Ans: The male female dichotomy is strongly embodied in most of cultures, religions, and even
Sperm characteristics associated with different male reproductive tactics in bluegills (Lepomis macrochirus)
") Behav Ecol Sociobiol (2000) 49:31 37 Springer-Verlag 2000 ORIGINAL ARTICLE Brenda Leach Robert Montgomerie Sperm characteristics associated with different male reproductive tactics in bluegills (Lepomis
Behav Ecol Sociobiol (2000) 49:31 37 Springer-Verlag 2000 ORIGINAL ARTICLE Brenda Leach Robert Montgomerie Sperm characteristics associated with different male reproductive tactics in bluegills (Lepomis
No evidence of sperm conjugate formation in an Australian mouse bearing sperm with three hooks
 No evidence of sperm conjugate formation in an Australian mouse bearing sperm with three hooks Renee C. Firman, Blair Bentley, Faye Bowman, Fernando Garcıa-Solıs Marchant, Jahmila Parthenay, Jessica Sawyer,
No evidence of sperm conjugate formation in an Australian mouse bearing sperm with three hooks Renee C. Firman, Blair Bentley, Faye Bowman, Fernando Garcıa-Solıs Marchant, Jahmila Parthenay, Jessica Sawyer,
Sperm competition and the evolution of testes size
 Functional Ecology 2008, 22, 655 662 doi: 10.1111/j.1365-2435.2008.01409.x Sperm competition and the evolution of testes size Blackwell Publishing Ltd in terrestrial mammalian carnivores G. Iossa*, C.
Functional Ecology 2008, 22, 655 662 doi: 10.1111/j.1365-2435.2008.01409.x Sperm competition and the evolution of testes size Blackwell Publishing Ltd in terrestrial mammalian carnivores G. Iossa*, C.
Sperm competition and its evolutionary consequences in humans
 6 Sperm competition and its evolutionary consequences in humans AARON T. GOETZ AND TODD K. SHACKELFORD Florida Atlantic University Identifying sperm competition Sexual selection is the mechanism that favors
6 Sperm competition and its evolutionary consequences in humans AARON T. GOETZ AND TODD K. SHACKELFORD Florida Atlantic University Identifying sperm competition Sexual selection is the mechanism that favors
Sperm competition and brain size evolution in mammals
 doi:10.1111/j.1420-9101.2009.01837.x Sperm competition and brain size evolution in mammals J.-F. LEMAÎTRE*, S. A. RAMM*, R. A. BARTON & P. STOCKLEY* *Mammalian Behaviour & Evolution Group, Faculty of Veterinary
doi:10.1111/j.1420-9101.2009.01837.x Sperm competition and brain size evolution in mammals J.-F. LEMAÎTRE*, S. A. RAMM*, R. A. BARTON & P. STOCKLEY* *Mammalian Behaviour & Evolution Group, Faculty of Veterinary
Male monarch butterfly spermatophore mass and mating strategies
 Anita. Behav., 1988, 36, 1384-1388 Male monarch butterfly spermatophore mass and mating strategies KAREN S. OBERHAUSER Department of Ecology and Behavioral Biology, 109 Zoology, University of Minnesota,
Anita. Behav., 1988, 36, 1384-1388 Male monarch butterfly spermatophore mass and mating strategies KAREN S. OBERHAUSER Department of Ecology and Behavioral Biology, 109 Zoology, University of Minnesota,
Biology 352, Spring 2018 Exam Number KEY Second midterm exam Part 1 (short answer worth 21 % of grade)
") Part 1 (short answer worth 21 % of grade) 1-1) Match the following terms used in behavioral ecology with the appropriate topics of behavioral study listed to the right (you may apply more than one topic
Part 1 (short answer worth 21 % of grade) 1-1) Match the following terms used in behavioral ecology with the appropriate topics of behavioral study listed to the right (you may apply more than one topic
Assessment of sperm competition by European bitterling, Rhodeus sericeus
 Behav Ecol Sociobiol (2003) 53:206 213 DOI 10.1007/s00265-002-0576-x ORIGINAL ARTICLE Carl Smith Martin Reichard Pavel Jurajda Assessment of sperm competition by European bitterling, Rhodeus sericeus Received:
Behav Ecol Sociobiol (2003) 53:206 213 DOI 10.1007/s00265-002-0576-x ORIGINAL ARTICLE Carl Smith Martin Reichard Pavel Jurajda Assessment of sperm competition by European bitterling, Rhodeus sericeus Received:
Sperm competition in birds
 Reviews of Reproduction (1998) 3, 123 129 Sperm competition in birds Tim R. Birkhead Department of Animal and Plant Sciences, The University, Sheffield S10 2TN, UK Sperm competition in birds occurs when
Reviews of Reproduction (1998) 3, 123 129 Sperm competition in birds Tim R. Birkhead Department of Animal and Plant Sciences, The University, Sheffield S10 2TN, UK Sperm competition in birds occurs when
Experimental and natural changes in the peacock's (Pavo cristatus} train can affect mating success
 Behav Ecol Sociobiol (1994) 35:213-217 Springer-Verlag 1994 Marion Petrie - Tim Halliday Experimental and natural changes in the peacock's (Pavo cristatus} train can affect mating success Received: 18
Behav Ecol Sociobiol (1994) 35:213-217 Springer-Verlag 1994 Marion Petrie - Tim Halliday Experimental and natural changes in the peacock's (Pavo cristatus} train can affect mating success Received: 18
Shrimp adjust their sex ratio to fluctuating age distributions
 Evolutionary Ecology Research, 2002, 4: 239 246 Shrimp adjust their sex ratio to fluctuating age distributions Eric L. Charnov 1,2 and Robert W. Hannah 3 1 Department of Biology, The University of New
Evolutionary Ecology Research, 2002, 4: 239 246 Shrimp adjust their sex ratio to fluctuating age distributions Eric L. Charnov 1,2 and Robert W. Hannah 3 1 Department of Biology, The University of New
Sex ratio bias in the dung beetle Onthophagus taurus: adaptive allocation or sex-specific offspring mortality?
 Evol Ecol (2011) 25:363 372 DOI 10.1007/s10682-010-9423-0 RESEARCH ARTICLE Sex ratio bias in the dung beetle Onthophagus taurus: adaptive allocation or sex-specific offspring mortality? Clarissa M. House
Evol Ecol (2011) 25:363 372 DOI 10.1007/s10682-010-9423-0 RESEARCH ARTICLE Sex ratio bias in the dung beetle Onthophagus taurus: adaptive allocation or sex-specific offspring mortality? Clarissa M. House
Types of Mating Systems
 The Human Mating System Note: this is the stereotypical EP view (e.g., Buss) Types of Mating Systems Random Monogamy Polygyny: variance in male reproductive success > variance in female reproductive success
The Human Mating System Note: this is the stereotypical EP view (e.g., Buss) Types of Mating Systems Random Monogamy Polygyny: variance in male reproductive success > variance in female reproductive success
Genetic evidence of promiscuity in Peromyscus leucopus
 Behav Ecol Sociobiol (1991) 28:171-178 Behavioral Ecology and Sociobiology Springer-Verlag 199~ Genetic evidence of promiscuity in Peromyscus leucopus Xuhua Xia* and John S. Millar Department of Zoology,
Behav Ecol Sociobiol (1991) 28:171-178 Behavioral Ecology and Sociobiology Springer-Verlag 199~ Genetic evidence of promiscuity in Peromyscus leucopus Xuhua Xia* and John S. Millar Department of Zoology,
Each copy of any part of a JSTOR transmission must contain the same copyright notice that appears on the screen or printed page of such transmission.
 Bat Predation and Sexual Advertisement in a Neotropical Anuran Author(s): Michael J. Ryan, Merlin D. Tuttle, A. Stanley Rand Source: The American Naturalist, Vol. 119, No. 1 (Jan., 1982), pp. 136-139 Published
Bat Predation and Sexual Advertisement in a Neotropical Anuran Author(s): Michael J. Ryan, Merlin D. Tuttle, A. Stanley Rand Source: The American Naturalist, Vol. 119, No. 1 (Jan., 1982), pp. 136-139 Published
Parasitism of mating and non-mating males and body mass of Enallagma hageni
 Parasitism of mating and non-mating males and body mass of Enallagma hageni Sarah T Kaminsky, Akinyemi Oni-Orisan, Stephen Pruett-Jones, Harvey Blankespoor Email: Sarah T Kaminsky - kaminss@umich.edu;
Parasitism of mating and non-mating males and body mass of Enallagma hageni Sarah T Kaminsky, Akinyemi Oni-Orisan, Stephen Pruett-Jones, Harvey Blankespoor Email: Sarah T Kaminsky - kaminss@umich.edu;
This question is taken directly from the list of second test study questions (#6) it should not be a surprise...
 it should not be a surprise...") 1. Female choice of males that have exaggerated characters has been explained by female choice of males that have better genes. Explain how female choice of higher quality males might lead to exaggerated
1. Female choice of males that have exaggerated characters has been explained by female choice of males that have better genes. Explain how female choice of higher quality males might lead to exaggerated
SEX RATIO MANIPULATION BY THE PARASITOID WASP MUSCIDIFURAX RAPTOR IN RESPONSE TO HOST SIZE
 This is an electronic version of an article published as Seidl, S.E. and B.H. King. 1993. Sex ratio response to host size in the parasitoid wasp Muscidifurax raptor. Evolution 47:1876-1882. SEX RATIO MANIPULATION
This is an electronic version of an article published as Seidl, S.E. and B.H. King. 1993. Sex ratio response to host size in the parasitoid wasp Muscidifurax raptor. Evolution 47:1876-1882. SEX RATIO MANIPULATION
Male reproductive allocation in fireflies (Photinus spp.)
") 126(1): 74 80. r 2007, The Authors Journal compilation r 2007, The American Microscopical Society, Inc. DOI: 10.1111/j.1744-7410.2007.00078.x Male reproductive allocation in fireflies (Photinus spp.) Kristian
126(1): 74 80. r 2007, The Authors Journal compilation r 2007, The American Microscopical Society, Inc. DOI: 10.1111/j.1744-7410.2007.00078.x Male reproductive allocation in fireflies (Photinus spp.) Kristian
Some observations. Some traits are difficult to view as adaptations, because they appear to provide a disadvantage to the organism
 Some traits are difficult to view as adaptations, because they appear to provide a disadvantage to the organism Darwin asked: Can natural selection explain these differences? Structural traits: cumbersome
Some traits are difficult to view as adaptations, because they appear to provide a disadvantage to the organism Darwin asked: Can natural selection explain these differences? Structural traits: cumbersome
Environmental Potential for Polygamy. Polygamy Favored. Do females always benefit from being mates of polygynous males? Resource Defense Polygyny
 1 Polygamy Favored Biparental care not mandatory Food super-abundant Resource abundance varies spatially & temporally Environmental Potential for Polygamy Distribution of resources or mates Spatial & temporal
1 Polygamy Favored Biparental care not mandatory Food super-abundant Resource abundance varies spatially & temporally Environmental Potential for Polygamy Distribution of resources or mates Spatial & temporal
Intraspecific variation in testis size of small mammals: implications for muscle mass
 591 Intraspecific variation in testis size of small mammals: implications for muscle mass Albrecht I. Schulte-Hostedde, John S. Millar, and Graham J. Hickling Abstract: Intraspecific variation in testis
591 Intraspecific variation in testis size of small mammals: implications for muscle mass Albrecht I. Schulte-Hostedde, John S. Millar, and Graham J. Hickling Abstract: Intraspecific variation in testis
1. (6 pts) a. Can all characteristics of organisms be explained by natural selection? Explain your answer in a sentence (3 pts)
 a. Can all characteristics of organisms be explained by natural selection? Explain your answer in a sentence (3 pts)") Zoology 357 - Evolutionary Ecology - First Exam 1. (6 pts) a. Can all characteristics of organisms be explained by natural selection? Explain your answer in a sentence (3 pts) b. Name two non-selectionist
Zoology 357 - Evolutionary Ecology - First Exam 1. (6 pts) a. Can all characteristics of organisms be explained by natural selection? Explain your answer in a sentence (3 pts) b. Name two non-selectionist
Sperm competition occurs when gametes from more than
 Behavioral Ecology Vol. 13 No. 5: 705 712 A state-based model of sperm allocation in a group-breeding salamander W. Edwin Harris and Jeffrey R. Lucas Department of Biological Sciences, 1392 Lilly Hall,
Behavioral Ecology Vol. 13 No. 5: 705 712 A state-based model of sperm allocation in a group-breeding salamander W. Edwin Harris and Jeffrey R. Lucas Department of Biological Sciences, 1392 Lilly Hall,
Sex Comb Variation in Four Drosophila Species from North India
 International Journal of Biotechnology and Bioengineering Research. ISSN 2231-1238, Volume 4, Number 4 (2013), pp. 329-334 Research India Publications http://www.ripublication.com/ ijbbr.htm Sex Comb Variation
International Journal of Biotechnology and Bioengineering Research. ISSN 2231-1238, Volume 4, Number 4 (2013), pp. 329-334 Research India Publications http://www.ripublication.com/ ijbbr.htm Sex Comb Variation
Robert E. Taylor Memorial Symposium Applied Reproductive Strategies in Beef Cattle Fort Collins, CO December 2-3, 2
 Robert E. Taylor Memorial Symposium Applied Reproductive Strategies in Beef Cattle Fort Collins, CO December 2-3, 2 2008 Natural Service Mating with Bulls - - Management Considerations - - Roger W. Ellis
Robert E. Taylor Memorial Symposium Applied Reproductive Strategies in Beef Cattle Fort Collins, CO December 2-3, 2 2008 Natural Service Mating with Bulls - - Management Considerations - - Roger W. Ellis
Sexual selection and the evolution of sex differences
 Sexual selection and the evolution of sex differences Males and females have the same genes. Why do the sexes often look and act so differently? Why is the male often insanely ornamented? (Or simply insane?)
Sexual selection and the evolution of sex differences Males and females have the same genes. Why do the sexes often look and act so differently? Why is the male often insanely ornamented? (Or simply insane?)
EVOLUTION OF EJACULATES: PATTERNS OF PHENOTYPIC AND GENOTYPIC VARIATION AND CONDITION DEPENDENCE IN SPERM COMPETITION TRAITS
 Evolution, 56(8), 2002, pp. 1622 1631 EVOLUTION OF EJACULATES: PATTERNS OF PHENOTYPIC AND GENOTYPIC VARIATION AND CONDITION DEPENDENCE IN SPERM COMPETITION TRAITS LEIGH W. SIMMONS 1 AND JANNE S. KOTIAHO
Evolution, 56(8), 2002, pp. 1622 1631 EVOLUTION OF EJACULATES: PATTERNS OF PHENOTYPIC AND GENOTYPIC VARIATION AND CONDITION DEPENDENCE IN SPERM COMPETITION TRAITS LEIGH W. SIMMONS 1 AND JANNE S. KOTIAHO
Intersexual Competition
 (in press, October 2012). In P. Whelan & A. Bolin (Eds.), Encyclopedia of Human Sexuality. Hoboken, NJ: Wiley Blackwell. Intersexual Competition Valerie G. Starratt, Nova Southeastern University, valerie.starratt@nova.edu
(in press, October 2012). In P. Whelan & A. Bolin (Eds.), Encyclopedia of Human Sexuality. Hoboken, NJ: Wiley Blackwell. Intersexual Competition Valerie G. Starratt, Nova Southeastern University, valerie.starratt@nova.edu
3/26/ Sexual dimorphism is differences between males and females of a species. 2. Anisogamy. 1. Fecundity
 Sexual Dimorphism 1. Sexual dimorphism is differences between males and females of a species. 2. Anisogamy A. sexual reproduction involving the fusion of two dissimilar gametes;individuals producing the
Sexual Dimorphism 1. Sexual dimorphism is differences between males and females of a species. 2. Anisogamy A. sexual reproduction involving the fusion of two dissimilar gametes;individuals producing the
The behavioral ecology of animal reproduction
 The behavioral ecology of animal reproduction I. Introduction to Behavioral Ecology Behavioral ecologists test hypotheses about the adaptive function of a behavior Behavioral Ecology Behavioral ecologists
The behavioral ecology of animal reproduction I. Introduction to Behavioral Ecology Behavioral ecologists test hypotheses about the adaptive function of a behavior Behavioral Ecology Behavioral ecologists
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at
 Experimental and Natural Changes in the Peacock's (Pavo cristatus) Train Can Affect Mating Success Author(s): Marion Petrie and Tim Halliday Source: Behavioral Ecology and Sociobiology, Vol. 35, No. 3
Experimental and Natural Changes in the Peacock's (Pavo cristatus) Train Can Affect Mating Success Author(s): Marion Petrie and Tim Halliday Source: Behavioral Ecology and Sociobiology, Vol. 35, No. 3
Threshold dimorphism in ejaculate characteristics in the squid Loligo bleekeri
 Vol. 345: 141 146, 07 doi:.3354/meps06971 MARINE ECOLOGY PROGRESS SERIES Mar Ecol Prog Ser Published September 13 Threshold dimorphism in ejaculate characteristics in the squid Loligo bleekeri Yoko Iwata*,
Vol. 345: 141 146, 07 doi:.3354/meps06971 MARINE ECOLOGY PROGRESS SERIES Mar Ecol Prog Ser Published September 13 Threshold dimorphism in ejaculate characteristics in the squid Loligo bleekeri Yoko Iwata*,
Territorial Contests. Chapter 22: Territorial Signals. Games again. Territories. Ownership asymmetry
 Territorial Contests Chapter 22: Territorial Signals Ownership asymmetry New twist: what significance? Previous models have only considered asymmetries in resource value and cost of conflict (fighting
Territorial Contests Chapter 22: Territorial Signals Ownership asymmetry New twist: what significance? Previous models have only considered asymmetries in resource value and cost of conflict (fighting
Types of behaviors that are elicited in response to simple stimuli
 Lecture 19: Animal Behavior I. Background A. Animal behavior reflects and arises from biological properties 1. Exhibited behavior defends on the physiological systems and processes unique to a given organism
Lecture 19: Animal Behavior I. Background A. Animal behavior reflects and arises from biological properties 1. Exhibited behavior defends on the physiological systems and processes unique to a given organism
Supporting Online Material for
 www.sciencemag.org/cgi/content/full/science.1185550/dc1 Supporting Online Material for Cryptic Sex-Ratio Bias Provides Indirect Genetic Benefits Despite Sexual Conflict Robert M. Cox* and Ryan Calsbeek
www.sciencemag.org/cgi/content/full/science.1185550/dc1 Supporting Online Material for Cryptic Sex-Ratio Bias Provides Indirect Genetic Benefits Despite Sexual Conflict Robert M. Cox* and Ryan Calsbeek
Learning Objectives (Davies et al. 2012, Table 9.3)
") Behavioral Ecology of Vertebrates Unit 9. Mating Systems Module 4 Reproduction j-packard@tamu.edu Previously, we have examined how the physical and social environments have shaped behavioral strategies.
Behavioral Ecology of Vertebrates Unit 9. Mating Systems Module 4 Reproduction j-packard@tamu.edu Previously, we have examined how the physical and social environments have shaped behavioral strategies.
Female mating preferences are widely recognized as being
 Quantitative genetic correlation between trait and preference supports a sexually selected sperm process Leigh W. Simmons* and Janne S. Kotiaho *Centre for Evolutionary Biology, School of Animal Biology
Quantitative genetic correlation between trait and preference supports a sexually selected sperm process Leigh W. Simmons* and Janne S. Kotiaho *Centre for Evolutionary Biology, School of Animal Biology
Female mating preferences for colourful males in a population of guppies subject to high predation
 Journal of Fish Biology (2004) 65, 1154 1159 doi:10.1111/j.1095-8649.2004.00502.x, available online at http://www.blackwell-synergy.com Female mating preferences for colourful males in a population of
Journal of Fish Biology (2004) 65, 1154 1159 doi:10.1111/j.1095-8649.2004.00502.x, available online at http://www.blackwell-synergy.com Female mating preferences for colourful males in a population of
Evolutionary trade-off between weapons and testes
 Evolutionary trade-off between weapons and testes Leigh W. Simmons, and Douglas J. Emlen PNAS published online Oct 19, 2006; doi:10.1073/pnas.0603474103 This information is current as of December 2006.
Evolutionary trade-off between weapons and testes Leigh W. Simmons, and Douglas J. Emlen PNAS published online Oct 19, 2006; doi:10.1073/pnas.0603474103 This information is current as of December 2006.
Variation in sperm size within and between ejaculates. in a cockroach. W. E. HARRIS*, A. J. MOORE and P. J. MOORE
 Functional Ecology 2007 Variation in sperm size within and between ejaculates Blackwell Publishing Ltd in a cockroach W. E. HARRIS*, A. J. MOORE and P. J. MOORE *Faculty of Life Sciences, The University
Functional Ecology 2007 Variation in sperm size within and between ejaculates Blackwell Publishing Ltd in a cockroach W. E. HARRIS*, A. J. MOORE and P. J. MOORE *Faculty of Life Sciences, The University
Seasonal Variation in Mate Choice of Photinus ignitus Fireflies
 Ethology 111, 89 100 (2005) Ó 2005 Blackwell Verlag, Berlin ISSN 0179 1613 Seasonal Variation in Mate Choice of Photinus ignitus Fireflies Christopher K. Cratsley* & Sara M. Lewis *Department of Biology,
Ethology 111, 89 100 (2005) Ó 2005 Blackwell Verlag, Berlin ISSN 0179 1613 Seasonal Variation in Mate Choice of Photinus ignitus Fireflies Christopher K. Cratsley* & Sara M. Lewis *Department of Biology,
Ever since Parker (1970) recognized the importance of
 recognized the importance of") Behavioral Ecology Vol. 12 No. 6: 726 731 Sperm size of African cichlids in relation to sperm competition Sigal Balshine, a Brenda J. Leach, b Francis Neat, a Noam Y. Werner, c and Robert Montgomerie b
Behavioral Ecology Vol. 12 No. 6: 726 731 Sperm size of African cichlids in relation to sperm competition Sigal Balshine, a Brenda J. Leach, b Francis Neat, a Noam Y. Werner, c and Robert Montgomerie b
Rival Male Relatedness Does Not Affect Ejaculate Allocation as Predicted by Sperm Competition Theory
 Rival Male Relatedness Does Not Affect Ejaculate Allocation as Predicted by Sperm Competition Theory Melissa L. Thomas*, Leigh W. Simmons Centre for Evolutionary Biology, School of Animal Biology, The
Rival Male Relatedness Does Not Affect Ejaculate Allocation as Predicted by Sperm Competition Theory Melissa L. Thomas*, Leigh W. Simmons Centre for Evolutionary Biology, School of Animal Biology, The
MATERIALS AND METHODS
 48 M. J. RYAN amount used to produce the signal. However, because of the difficulty in determining energy expenditures associated with acoustic signalling, this comparison has only been made with insects
48 M. J. RYAN amount used to produce the signal. However, because of the difficulty in determining energy expenditures associated with acoustic signalling, this comparison has only been made with insects
OBSERVATION OF TOOTH SCARS ON THE HEAD OF MALE SPERM WHALE, AS AN INDICATION OF INTRA-SEXUAL FIGHTINGS
 OBSERVATION OF TOOTH SCARS ON THE HEAD OF MALE SPERM WHALE, AS AN INDICATION OF INTRA-SEXUAL FIGHTINGS HIDEHIRO KATO Whales Research Institute, Tokyo ABSTRACT In order to examine the relation between intra-sexual
OBSERVATION OF TOOTH SCARS ON THE HEAD OF MALE SPERM WHALE, AS AN INDICATION OF INTRA-SEXUAL FIGHTINGS HIDEHIRO KATO Whales Research Institute, Tokyo ABSTRACT In order to examine the relation between intra-sexual
Adaptation and Optimality Theory
 Adaptation and Optimality Theory Prisoner s Dilemma game: An optimality model What is the optimal strategy when you are playing with different people of unknown reputation? Defect: We call this the evolutionarily
Adaptation and Optimality Theory Prisoner s Dilemma game: An optimality model What is the optimal strategy when you are playing with different people of unknown reputation? Defect: We call this the evolutionarily
A Comparative Study of Mating Strategies in Two Species of Philanthus (Hymenoptera: Sphecidae)
") University of Wyoming National Park Service Research Center Annual Report Volume 4 4th Annual Report, 1980 Article 10 1-1-1980 A Comparative Study of Mating Strategies in Two Species of Philanthus (Hymenoptera:
University of Wyoming National Park Service Research Center Annual Report Volume 4 4th Annual Report, 1980 Article 10 1-1-1980 A Comparative Study of Mating Strategies in Two Species of Philanthus (Hymenoptera:
Superior sperm competitors sire higher-quality young
 FirstCite e-publishing Received 24 February 2003 Accepted 8 May 2003 Published online Superior sperm competitors sire higher-quality young D. J. Hosken 1*, T. W. J. Garner 1, T. Tregenza 2, N. Wedell 2
FirstCite e-publishing Received 24 February 2003 Accepted 8 May 2003 Published online Superior sperm competitors sire higher-quality young D. J. Hosken 1*, T. W. J. Garner 1, T. Tregenza 2, N. Wedell 2
Cooperative Breeding in Groups of Synchronously Mating Females and Evolution of Large Testes to Avoid Sperm Depletion in African Striped Mice 1
 BIOLOGY OF REPRODUCTION 81, 111 117 (2009) Published online before print 4 March 2009. DOI 10.1095/biolreprod.108.075838 Cooperative Breeding in Groups of Synchronously Mating Females and Evolution of
BIOLOGY OF REPRODUCTION 81, 111 117 (2009) Published online before print 4 March 2009. DOI 10.1095/biolreprod.108.075838 Cooperative Breeding in Groups of Synchronously Mating Females and Evolution of
PHOTOPERIODIC CONTROL OF THE TESTICULAR CYCLE IN THE EASTERN RED-WINGED BLACKBIRD, (AGELAIUS PHOENICEUS PHOENICEUS)
") University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Bird Control Seminars Proceedings Wildlife Damage Management, Internet Center for September 1970 PHOTOPERIODIC CONTROL OF
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Bird Control Seminars Proceedings Wildlife Damage Management, Internet Center for September 1970 PHOTOPERIODIC CONTROL OF
Introduction to Biological Anthropology: Notes 15 Primate sociality: Predators and living in groups Copyright Bruce Owen 2010 Most haplorrine, and
 Introduction to Biological Anthropology: Notes 15 Primate sociality: Predators and living in groups Copyright Bruce Owen 2010 Most haplorrine, and many strepsirrhine, primates are social: they live in
Introduction to Biological Anthropology: Notes 15 Primate sociality: Predators and living in groups Copyright Bruce Owen 2010 Most haplorrine, and many strepsirrhine, primates are social: they live in
Cost/benefit approach
 Cost/benefit approach Care FEMALE Abandon MALE Care F: wp 2 WP 1 M: wp 2 WP 1 Abandon F: wp 1 WP 0 M: wp 1 (1+p M ) WP 0 (1+p M ) P 0,1,2 = probability eggs survive given 0, 1, or 2 parents W, w = eggs
Cost/benefit approach Care FEMALE Abandon MALE Care F: wp 2 WP 1 M: wp 2 WP 1 Abandon F: wp 1 WP 0 M: wp 1 (1+p M ) WP 0 (1+p M ) P 0,1,2 = probability eggs survive given 0, 1, or 2 parents W, w = eggs
Territorial defense by males is a conspicuous component
 Behavioral Ecology Vol. 12 No. 5: 626 632 Territoriality and male reproductive success in arctic ground squirrels Eileen A. Lacey and John R. Wieczorek Museum of Zoology and Department of Biology, The
Behavioral Ecology Vol. 12 No. 5: 626 632 Territoriality and male reproductive success in arctic ground squirrels Eileen A. Lacey and John R. Wieczorek Museum of Zoology and Department of Biology, The
Husband s Reaction to His Wife s Sexual Rejection Is Predicted by the Time She Spends With Her Male Friends but Not Her Male Coworkers
 Original Article Husband s Reaction to His Wife s Sexual Rejection Is Predicted by the Time She Spends With Her Male Friends but Not Her Male Coworkers Evolutionary Psychology April-June 2017: 1 5 ª The
Original Article Husband s Reaction to His Wife s Sexual Rejection Is Predicted by the Time She Spends With Her Male Friends but Not Her Male Coworkers Evolutionary Psychology April-June 2017: 1 5 ª The
Animal Behaviour 78 (2009) Contents lists available at ScienceDirect. Animal Behaviour. journal homepage:
 Contents lists available at ScienceDirect. Animal Behaviour. journal homepage:") Animal Behaviour 78 (2009) 685 690 Contents lists available at ScienceDirect Animal Behaviour journal homepage: www.elsevier.com/locate/yanbe Male house mice do not adjust sperm allocation in response
Animal Behaviour 78 (2009) 685 690 Contents lists available at ScienceDirect Animal Behaviour journal homepage: www.elsevier.com/locate/yanbe Male house mice do not adjust sperm allocation in response
Introduction. Chapter 1
 Chapter 1 Introduction I spent my earliest days as a professional biologist happily tramping through the forests of southern Canada recording the survival, reproduction, and movement patterns of tiny mice
Chapter 1 Introduction I spent my earliest days as a professional biologist happily tramping through the forests of southern Canada recording the survival, reproduction, and movement patterns of tiny mice
POST-ALLOGROOMING REDUCTIONS IN SELF-DIRECTED BEHAVIOUR ARE AFFECTED BY ROLE AND STATUS IN THE GREEN WOODHOOPOE
 2 3 4 5 6 POST-ALLOGROOMING REDUCTIONS IN SELF-DIRECTED BEHAVIOUR ARE AFFECTED BY ROLE AND STATUS IN THE GREEN WOODHOOPOE Andrew N. Radford Electronic Supplementary Material 8 9 0 2 3 4 5 6 8 9 Study Species
2 3 4 5 6 POST-ALLOGROOMING REDUCTIONS IN SELF-DIRECTED BEHAVIOUR ARE AFFECTED BY ROLE AND STATUS IN THE GREEN WOODHOOPOE Andrew N. Radford Electronic Supplementary Material 8 9 0 2 3 4 5 6 8 9 Study Species
Evidence for directional testes asymmetry in Hyla gongshanensis jindongensis
 Acta Herpetologica 12(1): 89-93, 2017 DOI: 10.13128/Acta_Herpetol-17902 Evidence for directional testes asymmetry in Hyla gongshanensis jindongensis Qing Gui Wu 1, Wen Bo Liao 2, * 1 Ecological Security
Acta Herpetologica 12(1): 89-93, 2017 DOI: 10.13128/Acta_Herpetol-17902 Evidence for directional testes asymmetry in Hyla gongshanensis jindongensis Qing Gui Wu 1, Wen Bo Liao 2, * 1 Ecological Security
When boys want to be girls: effects of mating system and dispersal on parent offspring sex ratio conflict
 Evolutionary Ecology Research, 2006, 8: 103 113 When boys want to be girls: effects of mating system and dispersal on parent offspring sex ratio conflict Ido Pen* Theoretical Biology Group, Centre for
Evolutionary Ecology Research, 2006, 8: 103 113 When boys want to be girls: effects of mating system and dispersal on parent offspring sex ratio conflict Ido Pen* Theoretical Biology Group, Centre for
Centrobolus dubius (Schubart, 1966) Monomorphism
 Monomorphism") International Journal of Research Studies in Zoology Volume 4, Issue 3, 2018, PP 17-21 ISSN No. 2454-941X DOI: http://dx.doi.org/10.20431/2454-941x.0403003 www.arcjournals.org Mark Cooper School of Animal,
International Journal of Research Studies in Zoology Volume 4, Issue 3, 2018, PP 17-21 ISSN No. 2454-941X DOI: http://dx.doi.org/10.20431/2454-941x.0403003 www.arcjournals.org Mark Cooper School of Animal,
Evolution of Mating Systems. Chapter 8
 Evolution of Mating Systems Chapter 8 Mating Systems-Chapter 8 1 Monogamy 2 Polyandry 3 Polygyny And the many combinations within! Why should a male be monogamous? 1 extension of guarding, little chance
Evolution of Mating Systems Chapter 8 Mating Systems-Chapter 8 1 Monogamy 2 Polyandry 3 Polygyny And the many combinations within! Why should a male be monogamous? 1 extension of guarding, little chance
Good resources. Why are females choosy? Good resources. Resources from males can tip the scales of relative investment
 Why are females choosy? Good resources direct benefits Good genes indirect benefits Sexy son hypothesis (Fisher) Handicap hypothesis (Zahavi) Good genes for sons, daughters Good resources courtship feeding
Why are females choosy? Good resources direct benefits Good genes indirect benefits Sexy son hypothesis (Fisher) Handicap hypothesis (Zahavi) Good genes for sons, daughters Good resources courtship feeding
Good resources. Why are females choosy? Good resources. Good resources direct benefits Good genes indirect benefits
 Why are females choosy? Good resources direct benefits Good genes indirect benefits Sexy son hypothesis (Fisher) Handicap hypothesis (Zahavi) Good genes for sons, daughters courtship feeding Good resources
Why are females choosy? Good resources direct benefits Good genes indirect benefits Sexy son hypothesis (Fisher) Handicap hypothesis (Zahavi) Good genes for sons, daughters courtship feeding Good resources
Sexual size dimorphism in garter snakes (Thamnophis sirtalis), water snakes. (Nerodia sipedon) and black ratsnakes (Elaphe obsoleta) Karen Elgee
, water snakes. (Nerodia sipedon) and black ratsnakes (Elaphe obsoleta) Karen Elgee") Sexual size dimorphism in garter snakes (Thamnophis sirtalis), water snakes (Nerodia sipedon) and black ratsnakes (Elaphe obsoleta) By Karen Elgee Thesis submitted to the Department of Biology in partial
Sexual size dimorphism in garter snakes (Thamnophis sirtalis), water snakes (Nerodia sipedon) and black ratsnakes (Elaphe obsoleta) By Karen Elgee Thesis submitted to the Department of Biology in partial
Time allowed: 2 hours Answer ALL questions in Section A, ALL PARTS of the question in Section B and ONE question from Section C.
 UNIVERSITY OF EAST ANGLIA School of Biological Sciences Main Series UG Examination 2014-2015 BEHAVIOURAL ECOLOGY BIO-5010B Time allowed: 2 hours Answer ALL questions in Section A, ALL PARTS of the question
UNIVERSITY OF EAST ANGLIA School of Biological Sciences Main Series UG Examination 2014-2015 BEHAVIOURAL ECOLOGY BIO-5010B Time allowed: 2 hours Answer ALL questions in Section A, ALL PARTS of the question
Psychological and Physiological Adaptations to Sperm Competition in Humans
 Review of General Psychology Copyright 2005 by the Educational Publishing Foundation 2005, Vol. 9, No. 3, 228 248 1089-2680/05/$12.00 DOI: 10.1037/1089-2680.9.3.228 Psychological and Physiological Adaptations
Review of General Psychology Copyright 2005 by the Educational Publishing Foundation 2005, Vol. 9, No. 3, 228 248 1089-2680/05/$12.00 DOI: 10.1037/1089-2680.9.3.228 Psychological and Physiological Adaptations
The effect of breeding density and male quality on paternity-assurance behaviours in the house sparrow, Passer domesticus
 DOI 10.1007/s10164-010-0217-1 ARTICLE The effect of breeding density and male quality on paternity-assurance behaviours in the house sparrow, Passer domesticus Herbert Hoi Hans Tost Matteo Griggio Received:
DOI 10.1007/s10164-010-0217-1 ARTICLE The effect of breeding density and male quality on paternity-assurance behaviours in the house sparrow, Passer domesticus Herbert Hoi Hans Tost Matteo Griggio Received:
Mate preferences by male guppies (Poecilia reticulata) in relation to the risk of sperm competition
 in relation to the risk of sperm competition") Behav Ecol Sociobiol (2004) 55:266 271 DOI 10.1007/s00265-003-0710-4 ORIGINAL ARTICLE Laura D. Dosen Robert Montgomerie Mate preferences by male guppies (Poecilia reticulata) in relation to the risk of
Behav Ecol Sociobiol (2004) 55:266 271 DOI 10.1007/s00265-003-0710-4 ORIGINAL ARTICLE Laura D. Dosen Robert Montgomerie Mate preferences by male guppies (Poecilia reticulata) in relation to the risk of
Sperm competition roles and ejaculate investment in a promiscuous mammal
 doi: 10.1111/j.1420-9101.2012.02511.x Sperm competition roles and ejaculate investment in a promiscuous mammal J.-F. LEMAÎTRE 1,S.A.RAMM 2,J.L.HURST&P.STOCKLEY Mammalian Behaviour & Evolution Group, Institute
doi: 10.1111/j.1420-9101.2012.02511.x Sperm competition roles and ejaculate investment in a promiscuous mammal J.-F. LEMAÎTRE 1,S.A.RAMM 2,J.L.HURST&P.STOCKLEY Mammalian Behaviour & Evolution Group, Institute
Effect of act on fitness of recipient
 Kin selection (inclusive fitness) and social behavior These worker ants are sterile, and they differ greatly from their queens and males. Darwin was troubled by social-insect workers, because they have
Kin selection (inclusive fitness) and social behavior These worker ants are sterile, and they differ greatly from their queens and males. Darwin was troubled by social-insect workers, because they have
Rapid evolution towards equal sex ratios in a system with heterogamety
 Evolutionary Ecology Research, 1999, 1: 277 283 Rapid evolution towards equal sex ratios in a system with heterogamety Mark W. Blows, 1 * David Berrigan 2,3 and George W. Gilchrist 3 1 Department of Zoology,
Evolutionary Ecology Research, 1999, 1: 277 283 Rapid evolution towards equal sex ratios in a system with heterogamety Mark W. Blows, 1 * David Berrigan 2,3 and George W. Gilchrist 3 1 Department of Zoology,
An Introduction to Behavioural Ecology
 An Introduction to Behavioural Ecology J.R. Krebs FRS Royal Society Research Professor at the Edward Grey Institute of Field Ornithology Department of Zoology University of Oxford and Fellow of Pembroke
An Introduction to Behavioural Ecology J.R. Krebs FRS Royal Society Research Professor at the Edward Grey Institute of Field Ornithology Department of Zoology University of Oxford and Fellow of Pembroke
Optimal skew models focusing on the degree to which
 Reproductive skew in the polygynandrous acorn woodpecker Joseph Haydock and Walter D. Koenig Department of Biology, Gonzaga University, 502 East Boone Avenue, Spokane, WA 99258; and Hastings Natural History
Reproductive skew in the polygynandrous acorn woodpecker Joseph Haydock and Walter D. Koenig Department of Biology, Gonzaga University, 502 East Boone Avenue, Spokane, WA 99258; and Hastings Natural History
White Rose Research Online URL for this paper:
 This is a repository copy of Sperm mobility: mechanisms of fertilizing efficiency, genetic variation and phenotypic relationship with male status in the domestic fowl, Gallus gallus domesticus. White Rose
This is a repository copy of Sperm mobility: mechanisms of fertilizing efficiency, genetic variation and phenotypic relationship with male status in the domestic fowl, Gallus gallus domesticus. White Rose
Supplemental Information. Guarding Males Protect Females. from Predation in a Wild Insect. Rolando Rodríguez-Muñoz, Amanda Bretman, and Tom Tregenza
 Current Biology, Volume 21 Supplemental Information Guarding Males Protect Females from Predation in a Wild Insect Rolando Rodríguez-Muñoz, Amanda Bretman, and Tom Tregenza Figure S1. Spatial Use of Burrow
Current Biology, Volume 21 Supplemental Information Guarding Males Protect Females from Predation in a Wild Insect Rolando Rodríguez-Muñoz, Amanda Bretman, and Tom Tregenza Figure S1. Spatial Use of Burrow
Sexual Selection and Mating Systems. Outline
 Sexual Selection and Mating Systems Stephen M. Shuster BIO 666: Animal Behavior Fall 29 Northern Arizona University Outline.Darwin s contributions 2.Existing emphases in plant and animal mating systems
Sexual Selection and Mating Systems Stephen M. Shuster BIO 666: Animal Behavior Fall 29 Northern Arizona University Outline.Darwin s contributions 2.Existing emphases in plant and animal mating systems