City, University of London Institutional Repository
|
|
|
- Baldric McKinney
- 5 years ago
- Views:
Transcription

1 City Research Online City, University of London Institutional Repository Citation: Merchant, H. and Yarrow, K. (2016). How the motor system both encodes and influences our sense of time. Current Opinion in Behavioral Sciences, 8, pp doi: /j.cobeha This is the accepted version of the paper. This version of the publication may differ from the final published version. Permanent repository link: Link to published version: Copyright and reuse: City Research Online aims to make research outputs of City, University of London available to a wider audience. Copyright and Moral Rights remain with the author(s) and/or copyright holders. URLs from City Research Online may be freely distributed and linked to. City Research Online: publications@city.ac.uk
2 1 This is an author-final version. The final (press-formatted) version of this article will be published by Elsevier in Current Opinion in Behavioral Sciences (DOI: /j.cobeha ) This manuscript is made available under the CC-BY-NC-ND 4.0 license. How the motor system both encodes and influences our sense of time Hugo Merchant a,*&, Kielan Yarrow b*& a. Instituto de Neurobiología, UNAM, campus Juriquilla b. Department of Psychology, City University London *Corresponding authors: Dr. Hugo Merchant, Phone: Dr. Kielan Yarrow, Rhind Building, City University London, Northampton Square, London EC1V 0HB, UK Phone: & Equal contribution
3 2 Abstract Several lines of evidence suggest that motoric brain structures may form the core amodal component of a neural network supporting a wide range of timed behaviours. Here, we review recent findings which elucidate the neural computations that occur within motor regions, and in particular the supplementary motor area, in order to support precisely timed actions. Although motor activity may help us represent time, it is also clear that action both enriches and complicates the interpretation of sensory inputs. Hence, in the second half of this review, we also consider the latest findings regarding the perceptual distortions that our actions can impose upon the subjective timeline.
4 3 Highlights The core timing mechanism resides within the motor system Interval tuning in SMA is a dynamic signal encoding time in different contexts The prediction of sensorimotor events is transmitted from motor to sensory areas Actions lead to highly selective distortions of perceived time Actions distort the subjective timeline before, during and after a movement
5 4 Introduction Precise timing is an implicit component of all skilled actions, and often also an explicit requirement, as in musical production and dancing. It is possible to view such motor timing as a specific process generated by a dedicated timer. However, it is now clear that brain regions traditionally viewed as motoric form part of a core timing network which interacts dynamically with sensory regions to support a range of timing needs. Perception, meanwhile, is not merely a passive process; it is also an active one operating in tandem with the motor system. This implies that, just as for other aspects of perception, we should take account of our actions when interpreting the temporal meaning of sensory inputs. In this paper we provide a selective review of recent work in these two areas. First, we consider the neural mechanisms that support timed behaviours, with a particular focus on the role of the supplementary motor area (SMA), a key node in the corticothalamic-basal ganglia timing circuit (CTBGc). Second, we consider the ways in which temporal perception is affected by our ongoing actions. Neural mechanisms for timed behaviours Speech comprehension and articulation, as well as music appreciation and execution, depend on intricate loops linking the perception and execution of timed intervals. Hence, the timing mechanism engaged in these complex activities is multimodal and needs to extract temporal information from incoming stimuli and develop predictive signals about upcoming sensory and motor events. This scheme distinguishes time sensation (bottom-up) and time prediction (topdown). There is a vast functional-imaging literature that indicates that timing is based on a partially overlapping neural network that has two elements. The first is a core-timing network that resides in the motor system and includes the CTBGc, which emits predictive signals to the rest of the brain independently of the timing context. The second is a set of areas that are selectively engaged depending on the specific behavioural requirements of a task [1]. Thus, on one hand SMA and the basal ganglia of the CTBGc are activated during time production and perception tasks using both visual and auditory stimuli with various interval structures [2,3]. On the other hand, specific sensory and association areas are involved in interval timing, depending of the exact temporal paradigm [3,4]. Two basic features of interval timing are a linear increase in temporal variability as a function of interval duration, a phenomenon called the scalar property, and a systematic bias in estimated durations towards the mean of the input intervals. Using a Bayesian observer model it is
6 5 possible to capture both features when the temporal context changes, creating a testable framework regarding how the core timing network represents the prior distribution of intervals and measures time in a scalar fashion [5]. Indeed, a scalar representation of time has been observed recently in the primate SMA [6**], as well as in the rodent striatum [7]. Recent neurophysiological experiments have confirmed that the SMA is a crucial element in the core timing mechanism. The neural activity from this area in macaques was recorded during a synchronization-continuation task (SCT), where the tapping behaviour of monkeys was synchronized to a metronome of isochronous stimuli, followed by a period where animals internally temporalized their movements [8]. Monkeys use an explicit timing strategy to perform this task, tightly controlling the duration of the pauses (or dwell times) between movements, then making stereotyped pushing movements, across each produced interval in the rhythmic sequence [9]. Interestingly, SMA activity during the SCT indicates that the time-keeping mechanism is governed by different layers of neural clocks (Figure 1). At the bottom of the hierarchy is ramping activity that shows an instantaneous increase or decrease in response magnitude. This peaks at the time of the anticipated response, suggesting its involvement in the temporal construction of motor intentions and actions [10,11]. In fact, a tight interaction between the ramping cells computing elapsed time since the previous tap and a different set of cells encoding time remaining to the next tap generates a coordinated cycle of activation during the SCT that ends with the triggering of each tap [12]. On top of such climbing activity, there are SMA cells that are tuned not only to the duration of the produced intervals but also to the serial-order of elements within each reproduced sequence in the SCT. This suggests that the SMA uses a cell population signal that works rather like the notes of a musical score, representing both the duration and the rank order of the produced intervals in the sequence [13**]. Most importantly, a subgroup of these interval-tuned neurons showed similar preferred intervals across both auditory and visual modalities and during tasks involving the production of one or multiple intervals. This observation supports the notion that SMA is part of the core timing mechanism that uses interval tuning as an abstract signal across timing behaviours and modalities [13**]. The multiplexing of duration and serial order is a dynamic process, where discrete neural ensembles encode these parameters through small-time windows, so that the pattern of neural activity changes dramatically within each interval [6**]. Thus, the rapid activation of small groups of cells provides a strong synaptic drive to the next ensemble producing a neural avalanche across an interval [6**], which is repeated for every serial-order element of the SCT sequence [14*]. A recent
7 6 study has shown that the activity of duration/serial-order tuned cells in SMA can be statistically linked to either the sensory or motor events of the SCT [14*]. Moreover, a substantial group of sensory-aligned cells predict the appearance of the next stimulus in the sequence instead of simply responding to the previous stimulus. Therefore, the activity of this neural subpopulation can act as a top-down predictive signal to create the indispensable sensory-motor coupling that permits an animal to execute the SCT. Such predictive capabilities of the motor system seem well designed to sharpen perception in rhythmic contexts. When attention is allocated to auditory or visual events in a rhythmic sequence, delta oscillations of primary visual and auditory cortices of monkeys are entrained (i.e. phase-locked) to the attended modality [15]. Notably, however, the time-dependent processing of input stimuli can be further enhanced by using top-down signals coordinated by the motor system (Figure 1). A recent psychophysical study provides evidence for the existence of predictive signals associated with a rhythmic motor behaviour that are fed back to the sensory areas through a corollary discharge to enhance processing of incoming auditory signals at a particular cyclic phase [16**]. It seems that the motor system routinely generates strong dynamic signals in order to internally represent time, predict sensory events, and drive behaviour [17]. A possible mechanism for such active information distribution is through small pieces of spike-encoded data embedded in slow brain oscillations that can travel long distances [18]. Indeed, beta oscillations may be the channel for these top-down signals during rhythm perception and entrainment. The exact nature of the cell activity inserted into the beta oscillations is still unknown, but they have been associated with: (1) both the stimulusdriven and the endogenous interpretation of the beat in early auditory areas [19]; (2) the prediction of the onset of the next temporal event [20]; (3) the dynamical coupling between the motor and auditory areas [20], and (4) internally driven timing behaviour [21-23]. Such interactions between top-down information and perceptual processes provide a natural segue to our second area of focus: The tendency for action to bias temporal perception. Time perception in the context of action Actions induce distortions of the subjective timeline. Numerous distortions have been reported, affecting the events (and thus the intervals they define) occurring before, during, and after action (summarised in Figure 2). Here, we provide a brief update on classic findings. In the chronostasis illusion, a new percept that is revealed by an action is experienced as longer than a control percept. For example, the target of a saccadic eye movement has a subjectively
8 7 expanded duration, which may explain the commonly experienced stopped-clock effect (i.e. when we glance at a clock just after the second hand has advanced, and are surprised when it fails to advance again as quickly as we expect; [24]). A similar effect emerges when we reach to touch a vibrating stimulus [25]. Research conducted in the noughties suggested that saccadic chronostasis reflects an active mechanism compensating for sensory degradation during a saccade. The post-saccadic image may be being antedated, to stitch up the temporal gap induced by saccadic suppression and masking [24,26-29]. An active mechanism is implied because chronostasis emerges over and above any time dilation induced in passive control conditions, which simulate the visual effects of a saccade. A recent paper, however, included a very realistic passive control condition, and found temporal distortion similar to that obtained using real saccades (albeit in a very small sample of observers; [30*]). As we will see, action-induced timing illusions are often challenged on the basis that the fundamental cause is some correlate of action, rather than the action per se. For saccadic chronostasis, this issue bears further scrutiny. A similar effect, intentional binding, arises when an action causes a punctate event (such as a beep) to occur after a short delay [31]. Typically, the time of both the action and the beep are evaluated against a rapidly rotating clock in separate blocks. Judgements are made in a condition when the action causes the beep, but also in control conditions when each occurs alone. Generally, the time of the action is perceived relatively later, and the beep relatively earlier, when the action causes the beep; hence they are bound. The logical implication, that the interval from action to beep is perceptually compressed, has been verified directly via interval judgements (e.g. [32]). How central is action to this illusion? The effect is eliminated (even reversed) when the hand movement is triggered by magnetic brain stimulation, removing the sense of agency [30]. However, recent findings suggest that causality (rather than agency or intentionality) may be the critical factor inducing binding effects. For example, a binding-like effect can emerge between the action of an intentionless machine and a consequent flash [33]. It is important to keep in mind that this emphasis on causality stands against a backdrop of several studies emphasizing the importance of intentionality in this illusion [34]. Interestingly, while the focus of the effect has always been the interval between the action and the resulting sensory event, one recent study suggests that a similar temporal binding can occur between an imperative stimulus and an action, i.e. for the interval prior to action [35*]. The relative contributions of intentionality and causality to this new illusion are yet to be fully explored.
9 8 Shortly after intentional binding was described, a possible alternative account emerged in the form of motor-sensory temporal recalibration [36]. Here, an action causes an event, but only after a short lag (in baseline conditions the event occurs almost immediately). Participants adapt to these temporal relationships over multiple trials. On test trials, various timings are used and the time of the sensory consequence is judged relative to the time of the action. Compared to baseline, lag adaptation recalibrates the perceived temporal relationship, so that delayed events are now more likely to be judged synchronous with (or even preceding) the action. Now recall intentional binding. Because that effect is measured in a situation that might be expected to generate recalibration, it could be recast as a consequence of adaptation. However, at least one recent study suggests that these are actually independent effects [37]. Regardless of whether motor-sensory temporal recalibration explains intentional binding, it is interesting in its own right. Recent work shows that recalibration emerges when correlated events consistently lead (as well as lag) tapping actions ([38]; c.f. [35*]) and can also be induced by delayedfeedback tracking tasks [39]. Transfer of recalibration occurs when the stimulus changes ([40,41]; but see [42]). This, along with the details of the transfer observed between hand and foot tapping [43] and the (partial) capacity to adapt the two hands independently [44] suggests that recalibration mainly induces an effector-specific change in the perceived time of actions. The studies described so far focus mainly on the interval after the action. What happens before and during an action? The answers may depend upon which aspect of the subjective timeline we assess. For example, Morrone and colleagues briefly flashed two lines before or during a saccadic eye movement, and found a compression of the interval between them [45,46]. The effect was most pronounced when both lines appeared just before the saccade, and could even result in subjective order reversals, but was not found for auditory stimuli. However, as with other illusions considered here, there are some questions about whether these effects are fundamentally motoric in nature, or arise from correlated changes in visual sensitivity [47]. A recent paper has now found analogous compression for manual actions [48*]. Specifically, an interval defined by two brief tactile stimuli was subjectively compressed immediately prior to and during a reach when the taps were applied to the reaching hand, but not when they were applied to the non-reaching hand. This effect also occurred during static (isometric) force production, but not in a passive control experiment when stimulus intensity was adjusted to approximate the reduced percept occurring during action (as a result of tactile sensory suppression). This manual illusion is intriguing, but also puzzling, because it appears to be at odds with another recent report [49*]. Here, participants lifted either their index or middle fingers. When a
10 9 vibrotactile stimulus was applied to the active finger, its perceived duration increased, but this did not occur for a stimulus applied to the static finger. This result is opposite to that described previously, despite a similar setup. This increase was also observed when visually judging the duration of the same action on a computer screen. It occurred regardless of exact viewing perspective and spatial alignment. It is difficult to know what procedural differences gave rise to these apparently opposite effects, but we might speculate that dilated (filled) intervals generate simultaneous contrast effects affecting the empty intervals that they subsume. At first glance, a similar contradiction seems to emerge when the interval just prior to an action is considered. As already described, when assessed using two taps to the active hand, compression occurs just before movement [48*]. This is broadly in line with the bias to perceive a visual imperative stimulus (a change in the colour of a fixation dot) as later (assessed via comparison with a rotating clock) when it triggers an action [35*]. However, another recent report found that time (assessed visually) is expanded during the preparatory period preceding action [50]. In this case, however, the period in question was earlier, coming prior to the imperative stimulus triggering movement. Hence expansion of that interval is broadly consistent with a delayed perception of the imperative stimulus, and thus contraction of the interval from imperative stimulus to action onset. Conclusions In this brief review, we have highlighted how time is represented within core motor areas and translated into meaningful predictions and actions. Balancing the roles of core and dedicated timing systems in order to provide a more complete mechanistic explanation of their interactions and interdependencies will provide a key challenge for the future. We have also provided a concise update regarding the many ways in which time can be distorted by our actions. Researchers should focus on nailing down the functional relevance of these curious effects, in order to provide a concrete contribution to our understanding of active temporal perception.
11 10 Acknowledgements We thank Raul Paulín and Luis Prado for their technical assistance. Hugo Merchant was supported by the grants CONACYT: and PAPIIT: IN
12 11 References [1] Merchant H, Harrington DL, Meck WH: Neural basis of the perception and estimation of time. Annu Rev Neurosci 2013, 36: [2] Macar F, Coull J, Vidal F: The supplementary motor area in motor and perceptual time processing: fmri studies. Cogn Process 2006, 7(2): [3] Harrington DL, Castillo GN, Fong CH, Reed JD: Neural underpinnings of distortions in the experience of time across senses. Front Integr Neurosci 2011, 5:32. [4] Jantzen K, Steinberg F, Kelso J: Functional MRI reveals the existence of modality and coordination-dependent timing networks. Neuroimage 2005, 25(4): [5] Jazayeri M, Shadlen MN: Temporal context calibrates interval timing. Nat Neurosci 2010, 13(8): [6] ** Crowe DA, Zarco W, Bartolo R, Merchant H: Dynamic representation of the temporal and sequential structure of rhythmic movements in the primate medial premotor cortex. J Neurosci 2014, 34(36): This paper describes a neural correlate of the scalar representation of time, as well as the properties of the dynamic mechanism governing the flux of encoding information across small SMA cell populations that represent the temporal and sequential structure of a rhythmic tapping task. [7] Mello GB, Soares S, Paton JJ: A scalable population code for time in the striatum. Curr Biol 2015, 25(9): [8] Zarco W, Merchant H, Prado L, Mendez JC: Subsecond timing in primates: comparison of interval production between human subjects and rhesus monkeys. J Neurophysiol 2009, 102(6): [9] Donnet S, Bartolo R, Fernandes JM, Cunha JP, Prado L, Merchant H: Monkeys time their pauses of movement and not their movement-kinematics during a synchronization-continuation rhythmic task. J Neurophysiol 2014, 111(10): [10] Renoult L, Roux S, Riehle A: Time is a rubberband: neuronal activity in monkey motor cortex in relation to time estimation. Eur J Neurosci 2006, 23(11): [11] Knudsen EB, Powers ME, Moxon KA: Dissociating movement from movement timing in the rat primary motor cortex. J Neurosci 2014, 34(47): [12] Merchant H, Zarco W, Perez O, Prado L, Bartolo R: Measuring time with different neural chronometers during a synchronization-continuation task. Proc Natl Acad Sci U S A 2011, 108(49): [13] ** Merchant H, Perez O, Zarco W, Gamez J: Interval tuning in the primate medial premotor cortex as a general timing mechanism. J Neurosci 2013, 33(21):
13 12 This study shows that neurons in SMA are tuned to both the intervals and serial order elements of the SCT, and demonstrates that a subgroup of interval-tuned neurons show similar preferred intervals across both auditory and visual modalities and during tasks involving the production of one or multiple intervals, supporting the notion of a core timing mechanism. [14] * Merchant H, Pérez O, Bartolo R, Méndez JC, Mendoza G, Gámez J, Yc K, Prado L: Sensorimotor neural dynamics during isochronous tapping in the medial premotor cortex of the macaque. Eur J Neurosci 2015, 41(5): This paper is the first to show that the activity of duration/serial order tuned cells in SMA can be statistically linked to either the sensory or motor events of the SCT, and that a substantial group of sensory aligned cells predict the appearance of the next stimulus in the sequence. [15] Lakatos P, Karmos G, Mehta AD, Ulbert I, Schroeder CE: Entrainment of neuronal oscillations as a mechanism of attentional selection. Science 2008, 320(5872): [16] ** Morillon B, Schroeder CE, Wyart V: Motor contributions to the temporal precision of auditory attention. Nature communications 2014, 5:5255. This exciting paper shows that the predictive signals derived from rhythmic motor behaviour are send back to sensory areas through a corollary discharge to enhance processing of incoming auditory signals. [17] Laje R, Buonomano DV: Robust timing and motor patterns by taming chaos in recurrent neural networks. Nat Neurosci 2013, 16(7): [18] Buzsáki G, Draguhn A: Neuronal oscillations in cortical networks. Science 2004, 304(5679): [19] Iversen JR, Repp BH, Patel AD: Top-down control of rhythm perception modulates early auditory responses. Ann N Y Acad Sci 2009, 1169(1): [20] Fujioka T, Trainor LJ, Large EW, Ross B: Internalized timing of isochronous sounds is represented in neuromagnetic beta oscillations. J Neurosci 2012, 32(5): [21] Bartolo R, Prado L, Merchant H: Information processing in the primate basal ganglia during sensory-guided and internally driven rhythmic tapping. J Neurosci 2014, 34(11): [22] Kononowicz TW, van Rijn H: Single trial beta oscillations index time estimation. Neuropsychologia 2015, 75: [23] Bartolo R, Merchant H: Beta Oscillations are Linked to the Initiation of Sensory-Cued Movement Sequences and the Internal Guidance of Regular Tapping in the Monkey. J Neurosci 2015, 35(11): [24] Yarrow K, Haggard P, Heal R, Brown P, Rothwell JC: Illusory perceptions of space and time preserve cross-saccadic perceptual continuity. Nature 2001, 414(6861): [25] Yarrow K, Rothwell JCE: Manual chronostasis: Tactile perception precedes physical contact. Curr Biol 2003, 13:
14 13 [26] Yarrow K, Whiteley L, Haggard P, Rothwell JC: Biases in the perceived timing of perisaccadic perceptual and motor events. Percept Psychophys 2006, 68(7): [27] Yarrow K, Johnson H, Haggard P, Rothwell JC: Consistent chronostasis effects across saccade categories imply a subcortical efferent trigger. J Cogn Neurosci 2004, 16(5): [28] Yarrow K, Haggard P, Rothwell JC: Action, arousal, and subjective time. Conscious Cogn 2004, 13(2): [29] Yarrow K: Temporal dilation: the chronostasis illusion and spatial attention. In Attention and time. Edited by Nobre AC, Coull JT.. Oxford University Press; 2010: [30] * Knöll J, Morrone MC, Bremmer F: Spatio-temporal topography of saccadic overestimation of time. Vision Res 2013, 83: This study revisits the saccadic chronostasis illusion using several methodological innovations. It shows spatial as well as temporal dependencies for the effect, and casts doubt on the active/motoric interpretation developed in the original series of papers. [31] Haggard P, Clark S, Kalogeras J: Voluntary action and conscious awareness. Nat Neurosci 2002, 5(4): [32] Moore JW, Wegner DM, Haggard P: Modulating the sense of agency with external cues. Conscious Cogn 2009, 18(4): [33] Buehner MJ: Understanding the past, predicting the future: causation, not intentional action, is the root of temporal binding. Psychol Sci 2012, 23(12): [34] Moore JW, Obhi SS: Intentional binding and the sense of agency: a review. Conscious Cogn 2012, 21(1): [35] * Yabe Y, Goodale MA: Time flies when we intend to act: temporal distortion in a go/no-go task. J Neurosci 2015, 35(12): Here, an interesting new component of the intentional binding illusion is revealed by focussing on the imperative stimulus that precedes the action. [36] Stetson C, Cui X, Montague PR, Eagleman DM: Motor-sensory recalibration leads to an illusory reversal of action and sensation. Neuron 2006, 51(5): [37] Cravo AM, Haddad H, Claessens PM, Baldo MV: Bias and learning in temporal binding: Intervals between actions and outcomes are compressed by prior bias. Conscious Cogn 2013, 22(4): [38] Rohde M, Ernst MO: To lead and to lag forward and backward recalibration of perceived visuo-motor simultaneity. Front Psychol 2012, 3:599. [39] Rohde M, van Dam LC, Ernst MO: Predictability is necessary for closed-loop visual feedback delay adaptation. J Vis 2014, 14(3):4. [40] Heron J, Hanson JV, Whitaker D: Effect before cause: supramodal recalibration of sensorimotor timing. PLoS ONE 2009, 4(11):e7681.
15 14 [41] Sugano Y, Keetels M, Vroomen J: Adaptation to motor-visual and motor-auditory temporal lags transfer across modalities. Exp Brain Res 2010, 201(3): [42] Sugano Y, Keetels M, Vroomen J: The Build-Up and Transfer of Sensorimotor Temporal Recalibration Measured via a Synchronization Task. Front Psychol 2012, 3:246. [43] Yarrow K, Sverdrup-Stueland I, Roseboom W, Arnold DH: Sensorimotor Temporal Recalibration Within and Across Limbs. J Exp Psychol Hum Percept Perform 2013, 39: [44] Sugano Y, Keetels M, Vroomen J: Concurrent sensorimotor temporal recalibration to different lags for the left and right hand. Front Psychol 2014, 5:140. [45] Morrone MC, Ross J, Burr D: Saccadic eye movements cause compression of time as well as space. Nat Neurosci 2005, 8(7): [46] Binda P, Cicchini GM, Burr DC, Morrone MC: Spatiotemporal distortions of visual perception at the time of saccades. J Neurosci 2009, 29(42): [47] Terao M, Watanabe J, Yagi A, Nishida S: Reduction of stimulus visibility compresses apparent time intervals. Nat Neurosci 2008, 11(5): [48] * Tomassini A, Gori M, Baud-Bovy G, Sandini G, Morrone MC: Motor commands induce time compression for tactile stimuli. J Neurosci 2014, 34(27): In this paper, a temporal compression between visual events occurring before or during a saccade is generalised to manual actions by applying tactile taps around the time of movement onset. [49] * Press C, Berlot E, Bird G, Ivry R, Cook R: Moving time: The influence of action on duration perception. J Exp Psychol: Gen 2014, 143(5):1787. These authors demonstrate selective temporal dilation for both tactile and visual intervals with close spatial correspondence to a current action. [50] Hagura N, Kanai R, Orgs G, Haggard P: Ready steady slow: action preparation slows the subjective passage of time. Proc Biol Sci 2012, 279(1746):
16 15 Figure Legends Legend to Figure 1. A flux diagram for time processing using auditory stimuli, ranging from the extraction of temporal features (time sensation) in early sensory areas, to time perception, time production and time prediction (green characters) in the motor system. In turn, time prediction is fed back as a topdown signal to enhance the processing of incoming stimuli in sensory areas. The colored squares on the right highlight the tentative neural signals involved at each processing level of time quantification [6,7,10-22]. Legend to Figure 2 Schematic overview illustrating the contextual effects of action upon perceived time. Temporal distortions are placed along a timeline, beginning with the imperative stimulus that instructs action and ending with the consequences of that action. Widened clocks indicate subjectively expanded intervals, contracted clocks represent subjectively compressed intervals, and arrow shapes indicate subjective shifts of the events demarcating different intervals. Matched shading is used to indicate the event biases that belong with different interval biases. A. Visually defined intervals dilate during action preparation [50]. B. Visual events are perceived later when they trigger actions [35]. C. Vibrotactile and visual filled intervals dilate when they overlap action [49]. D. Empty intervals between pairs of visual/tactile markers are compressed before and during saccades/manual actions respectively [45,48]. E. Tactile (visual) information revealed at the end of a manual action (saccade) is temporally dilated (and antedated) [24-26]. F. Actions and their delayed consequences seem compressed in time [31] and adapting to delayed feedback induces a recalibration of the perceived timing between action and its sensory correlates [36].



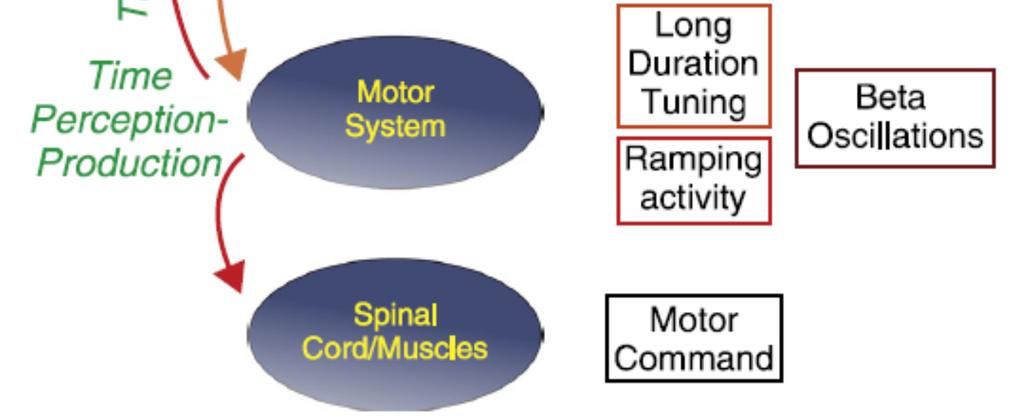
17 16 Figure 1
18 17 Figure 2 A. Preparatory dilation B. Imperative binding F. Intentional binding & motor-sensory C. Action dilation recalibration Time D. Action compression Action E. Chronostasis A Get ready! Go! Immediate feedback Delayed feedback
Selective bias in temporal bisection task by number exposition
 Selective bias in temporal bisection task by number exposition Carmelo M. Vicario¹ ¹ Dipartimento di Psicologia, Università Roma la Sapienza, via dei Marsi 78, Roma, Italy Key words: number- time- spatial
Selective bias in temporal bisection task by number exposition Carmelo M. Vicario¹ ¹ Dipartimento di Psicologia, Università Roma la Sapienza, via dei Marsi 78, Roma, Italy Key words: number- time- spatial
Is Clock Slowing General or Specific?
 Temporal Binding and Internal Clocks: Is Clock Slowing General or Specific? Richard Fereday (FeredayR@cardiff.ac.uk), Marc J. Buehner (BuehnerM@cardiff.ac.uk) Cardiff University, School of Psychology,
Temporal Binding and Internal Clocks: Is Clock Slowing General or Specific? Richard Fereday (FeredayR@cardiff.ac.uk), Marc J. Buehner (BuehnerM@cardiff.ac.uk) Cardiff University, School of Psychology,
Computational Explorations in Cognitive Neuroscience Chapter 7: Large-Scale Brain Area Functional Organization
 Computational Explorations in Cognitive Neuroscience Chapter 7: Large-Scale Brain Area Functional Organization 1 7.1 Overview This chapter aims to provide a framework for modeling cognitive phenomena based
Computational Explorations in Cognitive Neuroscience Chapter 7: Large-Scale Brain Area Functional Organization 1 7.1 Overview This chapter aims to provide a framework for modeling cognitive phenomena based
Oscillatory Neural Network for Image Segmentation with Biased Competition for Attention
 Oscillatory Neural Network for Image Segmentation with Biased Competition for Attention Tapani Raiko and Harri Valpola School of Science and Technology Aalto University (formerly Helsinki University of
Oscillatory Neural Network for Image Segmentation with Biased Competition for Attention Tapani Raiko and Harri Valpola School of Science and Technology Aalto University (formerly Helsinki University of
Evaluating the roles of the basal ganglia and the cerebellum in time perception
 Sundeep Teki Evaluating the roles of the basal ganglia and the cerebellum in time perception Auditory Cognition Group Wellcome Trust Centre for Neuroimaging SENSORY CORTEX SMA/PRE-SMA HIPPOCAMPUS BASAL
Sundeep Teki Evaluating the roles of the basal ganglia and the cerebellum in time perception Auditory Cognition Group Wellcome Trust Centre for Neuroimaging SENSORY CORTEX SMA/PRE-SMA HIPPOCAMPUS BASAL
A model of parallel time estimation
 A model of parallel time estimation Hedderik van Rijn 1 and Niels Taatgen 1,2 1 Department of Artificial Intelligence, University of Groningen Grote Kruisstraat 2/1, 9712 TS Groningen 2 Department of Psychology,
A model of parallel time estimation Hedderik van Rijn 1 and Niels Taatgen 1,2 1 Department of Artificial Intelligence, University of Groningen Grote Kruisstraat 2/1, 9712 TS Groningen 2 Department of Psychology,
How do individuals with congenital blindness form a conscious representation of a world they have never seen? brain. deprived of sight?
 How do individuals with congenital blindness form a conscious representation of a world they have never seen? What happens to visual-devoted brain structure in individuals who are born deprived of sight?
How do individuals with congenital blindness form a conscious representation of a world they have never seen? What happens to visual-devoted brain structure in individuals who are born deprived of sight?
On the nature of Rhythm, Time & Memory. Sundeep Teki Auditory Group Wellcome Trust Centre for Neuroimaging University College London
 On the nature of Rhythm, Time & Memory Sundeep Teki Auditory Group Wellcome Trust Centre for Neuroimaging University College London Timing substrates Timing mechanisms Rhythm and Timing Unified timing
On the nature of Rhythm, Time & Memory Sundeep Teki Auditory Group Wellcome Trust Centre for Neuroimaging University College London Timing substrates Timing mechanisms Rhythm and Timing Unified timing
The role of amplitude, phase, and rhythmicity of neural oscillations in top-down control of cognition
 The role of amplitude, phase, and rhythmicity of neural oscillations in top-down control of cognition Chair: Jason Samaha, University of Wisconsin-Madison Co-Chair: Ali Mazaheri, University of Birmingham
The role of amplitude, phase, and rhythmicity of neural oscillations in top-down control of cognition Chair: Jason Samaha, University of Wisconsin-Madison Co-Chair: Ali Mazaheri, University of Birmingham
Recalibration of temporal order perception by exposure to audio-visual asynchrony Vroomen, Jean; Keetels, Mirjam; de Gelder, Bea; Bertelson, P.
 Tilburg University Recalibration of temporal order perception by exposure to audio-visual asynchrony Vroomen, Jean; Keetels, Mirjam; de Gelder, Bea; Bertelson, P. Published in: Cognitive Brain Research
Tilburg University Recalibration of temporal order perception by exposure to audio-visual asynchrony Vroomen, Jean; Keetels, Mirjam; de Gelder, Bea; Bertelson, P. Published in: Cognitive Brain Research
2012 Course: The Statistician Brain: the Bayesian Revolution in Cognitive Sciences
 2012 Course: The Statistician Brain: the Bayesian Revolution in Cognitive Sciences Stanislas Dehaene Chair of Experimental Cognitive Psychology Lecture n 5 Bayesian Decision-Making Lecture material translated
2012 Course: The Statistician Brain: the Bayesian Revolution in Cognitive Sciences Stanislas Dehaene Chair of Experimental Cognitive Psychology Lecture n 5 Bayesian Decision-Making Lecture material translated
Introduction to the Special Issue on Multimodality of Early Sensory Processing: Early Visual Maps Flexibly Encode Multimodal Space
 BRILL Multisensory Research 28 (2015) 249 252 brill.com/msr Introduction to the Special Issue on Multimodality of Early Sensory Processing: Early Visual Maps Flexibly Encode Multimodal Space Roberto Arrighi1,
BRILL Multisensory Research 28 (2015) 249 252 brill.com/msr Introduction to the Special Issue on Multimodality of Early Sensory Processing: Early Visual Maps Flexibly Encode Multimodal Space Roberto Arrighi1,
Theoretical Neuroscience: The Binding Problem Jan Scholz, , University of Osnabrück
 The Binding Problem This lecture is based on following articles: Adina L. Roskies: The Binding Problem; Neuron 1999 24: 7 Charles M. Gray: The Temporal Correlation Hypothesis of Visual Feature Integration:
The Binding Problem This lecture is based on following articles: Adina L. Roskies: The Binding Problem; Neuron 1999 24: 7 Charles M. Gray: The Temporal Correlation Hypothesis of Visual Feature Integration:
A Multimodal Paradigm for Investigating the Perisaccadic Temporal Inversion Effect in Vision
 A Multimodal Paradigm for Investigating the Perisaccadic Temporal Inversion Effect in Vision Leo G. Trottier (leo@cogsci.ucsd.edu) Virginia R. de Sa (desa@cogsci.ucsd.edu) Department of Cognitive Science,
A Multimodal Paradigm for Investigating the Perisaccadic Temporal Inversion Effect in Vision Leo G. Trottier (leo@cogsci.ucsd.edu) Virginia R. de Sa (desa@cogsci.ucsd.edu) Department of Cognitive Science,
Spectral fingerprints of large-scale neuronal interactions
 Nature Reviews Neuroscience AOP, published online 11 January 212; doi:1.138/nrn3137 REVIEWS Spectral fingerprints of large-scale neuronal interactions Markus Siegel 1 *, Tobias H. Donner 2 * and Andreas
Nature Reviews Neuroscience AOP, published online 11 January 212; doi:1.138/nrn3137 REVIEWS Spectral fingerprints of large-scale neuronal interactions Markus Siegel 1 *, Tobias H. Donner 2 * and Andreas
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature14066 Supplementary discussion Gradual accumulation of evidence for or against different choices has been implicated in many types of decision-making, including value-based decisions
doi:10.1038/nature14066 Supplementary discussion Gradual accumulation of evidence for or against different choices has been implicated in many types of decision-making, including value-based decisions
CS/NEUR125 Brains, Minds, and Machines. Due: Friday, April 14
 CS/NEUR125 Brains, Minds, and Machines Assignment 5: Neural mechanisms of object-based attention Due: Friday, April 14 This Assignment is a guided reading of the 2014 paper, Neural Mechanisms of Object-Based
CS/NEUR125 Brains, Minds, and Machines Assignment 5: Neural mechanisms of object-based attention Due: Friday, April 14 This Assignment is a guided reading of the 2014 paper, Neural Mechanisms of Object-Based
Introduction to Computational Neuroscience
 Introduction to Computational Neuroscience Lecture 11: Attention & Decision making Lesson Title 1 Introduction 2 Structure and Function of the NS 3 Windows to the Brain 4 Data analysis 5 Data analysis
Introduction to Computational Neuroscience Lecture 11: Attention & Decision making Lesson Title 1 Introduction 2 Structure and Function of the NS 3 Windows to the Brain 4 Data analysis 5 Data analysis
5th Mini-Symposium on Cognition, Decision-making and Social Function: In Memory of Kang Cheng
 5th Mini-Symposium on Cognition, Decision-making and Social Function: In Memory of Kang Cheng 13:30-13:35 Opening 13:30 17:30 13:35-14:00 Metacognition in Value-based Decision-making Dr. Xiaohong Wan (Beijing
5th Mini-Symposium on Cognition, Decision-making and Social Function: In Memory of Kang Cheng 13:30-13:35 Opening 13:30 17:30 13:35-14:00 Metacognition in Value-based Decision-making Dr. Xiaohong Wan (Beijing
Agency, Subjective Time, and Other Minds
 Journal of Experimental Psychology: Human Perception and Performance 2007, Vol. 33, No. 6, 1261 1268 Copyright 2007 by the American Psychological Association 0096-1523/07/$12.00 DOI: 10.1037/0096-1523.33.6.1261
Journal of Experimental Psychology: Human Perception and Performance 2007, Vol. 33, No. 6, 1261 1268 Copyright 2007 by the American Psychological Association 0096-1523/07/$12.00 DOI: 10.1037/0096-1523.33.6.1261
Motor Systems I Cortex. Reading: BCP Chapter 14
 Motor Systems I Cortex Reading: BCP Chapter 14 Principles of Sensorimotor Function Hierarchical Organization association cortex at the highest level, muscles at the lowest signals flow between levels over
Motor Systems I Cortex Reading: BCP Chapter 14 Principles of Sensorimotor Function Hierarchical Organization association cortex at the highest level, muscles at the lowest signals flow between levels over
Key questions about attention
 Key questions about attention How does attention affect behavioral performance? Can attention affect the appearance of things? How does spatial and feature-based attention affect neuronal responses in
Key questions about attention How does attention affect behavioral performance? Can attention affect the appearance of things? How does spatial and feature-based attention affect neuronal responses in
Choosing the Greater of Two Goods: Neural Currencies for Valuation and Decision Making
 Choosing the Greater of Two Goods: Neural Currencies for Valuation and Decision Making Leo P. Surgre, Gres S. Corrado and William T. Newsome Presenter: He Crane Huang 04/20/2010 Outline Studies on neural
Choosing the Greater of Two Goods: Neural Currencies for Valuation and Decision Making Leo P. Surgre, Gres S. Corrado and William T. Newsome Presenter: He Crane Huang 04/20/2010 Outline Studies on neural
Neurobiology of Hearing (Salamanca, 2012) Auditory Cortex (2) Prof. Xiaoqin Wang
 Auditory Cortex (2) Prof. Xiaoqin Wang") Neurobiology of Hearing (Salamanca, 2012) Auditory Cortex (2) Prof. Xiaoqin Wang Laboratory of Auditory Neurophysiology Department of Biomedical Engineering Johns Hopkins University web1.johnshopkins.edu/xwang
Neurobiology of Hearing (Salamanca, 2012) Auditory Cortex (2) Prof. Xiaoqin Wang Laboratory of Auditory Neurophysiology Department of Biomedical Engineering Johns Hopkins University web1.johnshopkins.edu/xwang
Adaptation to motor-visual and motor-auditory temporal lags transfer across modalities
 Exp Brain Res (2010) 201:393 399 DOI 10.1007/s00221-009-2047-3 RESEARCH ARTICLE Adaptation to motor-visual and motor-auditory temporal lags transfer across modalities Yoshimori Sugano Mirjam Keetels Jean
Exp Brain Res (2010) 201:393 399 DOI 10.1007/s00221-009-2047-3 RESEARCH ARTICLE Adaptation to motor-visual and motor-auditory temporal lags transfer across modalities Yoshimori Sugano Mirjam Keetels Jean
Supplementary materials for: Executive control processes underlying multi- item working memory
 Supplementary materials for: Executive control processes underlying multi- item working memory Antonio H. Lara & Jonathan D. Wallis Supplementary Figure 1 Supplementary Figure 1. Behavioral measures of
Supplementary materials for: Executive control processes underlying multi- item working memory Antonio H. Lara & Jonathan D. Wallis Supplementary Figure 1 Supplementary Figure 1. Behavioral measures of
Supplementary Motor Area exerts Proactive and Reactive Control of Arm Movements
 Supplementary Material Supplementary Motor Area exerts Proactive and Reactive Control of Arm Movements Xiaomo Chen, Katherine Wilson Scangos 2 and Veit Stuphorn,2 Department of Psychological and Brain
Supplementary Material Supplementary Motor Area exerts Proactive and Reactive Control of Arm Movements Xiaomo Chen, Katherine Wilson Scangos 2 and Veit Stuphorn,2 Department of Psychological and Brain
Cognitive Modelling Themes in Neural Computation. Tom Hartley
 Cognitive Modelling Themes in Neural Computation Tom Hartley t.hartley@psychology.york.ac.uk Typical Model Neuron x i w ij x j =f(σw ij x j ) w jk x k McCulloch & Pitts (1943), Rosenblatt (1957) Net input:
Cognitive Modelling Themes in Neural Computation Tom Hartley t.hartley@psychology.york.ac.uk Typical Model Neuron x i w ij x j =f(σw ij x j ) w jk x k McCulloch & Pitts (1943), Rosenblatt (1957) Net input:
The Integration of Features in Visual Awareness : The Binding Problem. By Andrew Laguna, S.J.
 The Integration of Features in Visual Awareness : The Binding Problem By Andrew Laguna, S.J. Outline I. Introduction II. The Visual System III. What is the Binding Problem? IV. Possible Theoretical Solutions
The Integration of Features in Visual Awareness : The Binding Problem By Andrew Laguna, S.J. Outline I. Introduction II. The Visual System III. What is the Binding Problem? IV. Possible Theoretical Solutions
The Nonhuman Primate as Model System for Mechanistic Studies of Glutamate System Function and Dysfunction
 The Nonhuman Primate as Model System for Mechanistic Studies of Glutamate System Function and Dysfunction FORUM ON NEUROSCIENCE AND NERVOUS SYSTEM DISORDERS Board on Health Sciences Policy Glutamate-related
The Nonhuman Primate as Model System for Mechanistic Studies of Glutamate System Function and Dysfunction FORUM ON NEUROSCIENCE AND NERVOUS SYSTEM DISORDERS Board on Health Sciences Policy Glutamate-related
Coordination in Sensory Integration
 15 Coordination in Sensory Integration Jochen Triesch, Constantin Rothkopf, and Thomas Weisswange Abstract Effective perception requires the integration of many noisy and ambiguous sensory signals across
15 Coordination in Sensory Integration Jochen Triesch, Constantin Rothkopf, and Thomas Weisswange Abstract Effective perception requires the integration of many noisy and ambiguous sensory signals across
Cortical Control of Movement
 Strick Lecture 2 March 24, 2006 Page 1 Cortical Control of Movement Four parts of this lecture: I) Anatomical Framework, II) Physiological Framework, III) Primary Motor Cortex Function and IV) Premotor
Strick Lecture 2 March 24, 2006 Page 1 Cortical Control of Movement Four parts of this lecture: I) Anatomical Framework, II) Physiological Framework, III) Primary Motor Cortex Function and IV) Premotor
Memory, Attention, and Decision-Making
 Memory, Attention, and Decision-Making A Unifying Computational Neuroscience Approach Edmund T. Rolls University of Oxford Department of Experimental Psychology Oxford England OXFORD UNIVERSITY PRESS Contents
Memory, Attention, and Decision-Making A Unifying Computational Neuroscience Approach Edmund T. Rolls University of Oxford Department of Experimental Psychology Oxford England OXFORD UNIVERSITY PRESS Contents
Object Substitution Masking: When does Mask Preview work?
 Object Substitution Masking: When does Mask Preview work? Stephen W. H. Lim (psylwhs@nus.edu.sg) Department of Psychology, National University of Singapore, Block AS6, 11 Law Link, Singapore 117570 Chua
Object Substitution Masking: When does Mask Preview work? Stephen W. H. Lim (psylwhs@nus.edu.sg) Department of Psychology, National University of Singapore, Block AS6, 11 Law Link, Singapore 117570 Chua
Timing and the cerebellum (and the VOR) Neurophysiology of systems 2010
 Neurophysiology of systems 2010") Timing and the cerebellum (and the VOR) Neurophysiology of systems 2010 Asymmetry in learning in the reverse direction Full recovery from UP using DOWN: initial return to naïve values within 10 minutes,
Timing and the cerebellum (and the VOR) Neurophysiology of systems 2010 Asymmetry in learning in the reverse direction Full recovery from UP using DOWN: initial return to naïve values within 10 minutes,
Who is causing what? The sense of agency is relational and efferent-triggered
 Cognition 107 (2008) 693 704 www.elsevier.com/locate/cognit Brief article Who is causing what? The sense of agency is relational and efferent-triggered Kai Engbert a, *, Andreas Wohlschläger a, Patrick
Cognition 107 (2008) 693 704 www.elsevier.com/locate/cognit Brief article Who is causing what? The sense of agency is relational and efferent-triggered Kai Engbert a, *, Andreas Wohlschläger a, Patrick
Carl Wernicke s Contribution to Theories of Conceptual Representation in the Cerebral Cortex. Nicole Gage and Gregory Hickok Irvine, California
 Carl Wernicke s Contribution to Theories of Conceptual Representation in the Cerebral Cortex Nicole Gage and Gregory Hickok Irvine, California Acknowledgments Christian Sekirnjak, Ph.D. San Diego, CA Heidi
Carl Wernicke s Contribution to Theories of Conceptual Representation in the Cerebral Cortex Nicole Gage and Gregory Hickok Irvine, California Acknowledgments Christian Sekirnjak, Ph.D. San Diego, CA Heidi
Motor Functions of Cerebral Cortex
 Motor Functions of Cerebral Cortex I: To list the functions of different cortical laminae II: To describe the four motor areas of the cerebral cortex. III: To discuss the functions and dysfunctions of
Motor Functions of Cerebral Cortex I: To list the functions of different cortical laminae II: To describe the four motor areas of the cerebral cortex. III: To discuss the functions and dysfunctions of
Introduction to Computational Neuroscience
 Introduction to Computational Neuroscience Lecture 5: Data analysis II Lesson Title 1 Introduction 2 Structure and Function of the NS 3 Windows to the Brain 4 Data analysis 5 Data analysis II 6 Single
Introduction to Computational Neuroscience Lecture 5: Data analysis II Lesson Title 1 Introduction 2 Structure and Function of the NS 3 Windows to the Brain 4 Data analysis 5 Data analysis II 6 Single
 Peripheral facial paralysis (right side). The patient is asked to close her eyes and to retract their mouth (From Heimer) Hemiplegia of the left side. Note the characteristic position of the arm with
Peripheral facial paralysis (right side). The patient is asked to close her eyes and to retract their mouth (From Heimer) Hemiplegia of the left side. Note the characteristic position of the arm with
Visual Object Recognition Computational Models and Neurophysiological Mechanisms Neurobiology 130/230. Harvard College/GSAS 78454
 Visual Object Recognition Computational Models and Neurophysiological Mechanisms Neurobiology 130/230. Harvard College/GSAS 78454 Web site: http://tinyurl.com/visionclass (Class notes, readings, etc) Location:
Visual Object Recognition Computational Models and Neurophysiological Mechanisms Neurobiology 130/230. Harvard College/GSAS 78454 Web site: http://tinyurl.com/visionclass (Class notes, readings, etc) Location:
Beat perception in 3D: A comparative analysis between sight, sound, and touch
 Beat perception in 3D: A comparative analysis between sight, sound, and touch Heather Khey Beldman (Dr. Jessica A. Grahn) The Brain and Mind Institute, Western University Physiology 4980E Honors Thesis
Beat perception in 3D: A comparative analysis between sight, sound, and touch Heather Khey Beldman (Dr. Jessica A. Grahn) The Brain and Mind Institute, Western University Physiology 4980E Honors Thesis
Chapter 5. Summary and Conclusions! 131
 ! Chapter 5 Summary and Conclusions! 131 Chapter 5!!!! Summary of the main findings The present thesis investigated the sensory representation of natural sounds in the human auditory cortex. Specifically,
! Chapter 5 Summary and Conclusions! 131 Chapter 5!!!! Summary of the main findings The present thesis investigated the sensory representation of natural sounds in the human auditory cortex. Specifically,
Morton-Style Factorial Coding of Color in Primary Visual Cortex
 Morton-Style Factorial Coding of Color in Primary Visual Cortex Javier R. Movellan Institute for Neural Computation University of California San Diego La Jolla, CA 92093-0515 movellan@inc.ucsd.edu Thomas
Morton-Style Factorial Coding of Color in Primary Visual Cortex Javier R. Movellan Institute for Neural Computation University of California San Diego La Jolla, CA 92093-0515 movellan@inc.ucsd.edu Thomas
Frank Tong. Department of Psychology Green Hall Princeton University Princeton, NJ 08544
 Frank Tong Department of Psychology Green Hall Princeton University Princeton, NJ 08544 Office: Room 3-N-2B Telephone: 609-258-2652 Fax: 609-258-1113 Email: ftong@princeton.edu Graduate School Applicants
Frank Tong Department of Psychology Green Hall Princeton University Princeton, NJ 08544 Office: Room 3-N-2B Telephone: 609-258-2652 Fax: 609-258-1113 Email: ftong@princeton.edu Graduate School Applicants
Entrainment of neuronal oscillations as a mechanism of attentional selection: intracranial human recordings
 Entrainment of neuronal oscillations as a mechanism of attentional selection: intracranial human recordings J. Besle, P. Lakatos, C.A. Schevon, R.R. Goodman, G.M. McKhann, A. Mehta, R.G. Emerson, C.E.
Entrainment of neuronal oscillations as a mechanism of attentional selection: intracranial human recordings J. Besle, P. Lakatos, C.A. Schevon, R.R. Goodman, G.M. McKhann, A. Mehta, R.G. Emerson, C.E.
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and
 This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution
ANTICIPATING DYNAMIC LOADS IN HANDLING OBJECTS.
 ANTICIPATING DYNAMIC LOADS IN HANDLING OBJECTS. Alan M. Wing Sensory Motor Neuroscience Centre, The University of Birmingham, Edgbaston, Birmingham, B15 2TT, UK. a.m.wing@bham.ac.uk J. Randall Flanagan
ANTICIPATING DYNAMIC LOADS IN HANDLING OBJECTS. Alan M. Wing Sensory Motor Neuroscience Centre, The University of Birmingham, Edgbaston, Birmingham, B15 2TT, UK. a.m.wing@bham.ac.uk J. Randall Flanagan
Intentional Binding and the Sense of Agency: A review
 Intentional Binding and the Sense of Agency: A review James W. Moore 1,2 & Sukhvinder S. Obhi 3,4 1. Department of Psychology, Goldsmiths, University of London, London, UK 2. Brain Mapping Unit, Department
Intentional Binding and the Sense of Agency: A review James W. Moore 1,2 & Sukhvinder S. Obhi 3,4 1. Department of Psychology, Goldsmiths, University of London, London, UK 2. Brain Mapping Unit, Department
Sleep-Wake Cycle I Brain Rhythms. Reading: BCP Chapter 19
 Sleep-Wake Cycle I Brain Rhythms Reading: BCP Chapter 19 Brain Rhythms and Sleep Earth has a rhythmic environment. For example, day and night cycle back and forth, tides ebb and flow and temperature varies
Sleep-Wake Cycle I Brain Rhythms Reading: BCP Chapter 19 Brain Rhythms and Sleep Earth has a rhythmic environment. For example, day and night cycle back and forth, tides ebb and flow and temperature varies
Extracting the beat: An experience-dependent complex integration of multisensory information involving multiple levels of the nervous system
 Extracting the beat: An experience-dependent complex integration of multisensory information involving multiple levels of the nervous system LAUREL J. TRAINOR [1] McMaster University ANDREA UNRAU McMaster
Extracting the beat: An experience-dependent complex integration of multisensory information involving multiple levels of the nervous system LAUREL J. TRAINOR [1] McMaster University ANDREA UNRAU McMaster
Selective Attention. Modes of Control. Domains of Selection
 The New Yorker (2/7/5) Selective Attention Perception and awareness are necessarily selective (cell phone while driving): attention gates access to awareness Selective attention is deployed via two modes
The New Yorker (2/7/5) Selective Attention Perception and awareness are necessarily selective (cell phone while driving): attention gates access to awareness Selective attention is deployed via two modes
Consciousness as representation formation from a neural Darwinian perspective *
 Consciousness as representation formation from a neural Darwinian perspective * Anna Kocsis, mag.phil. Institute of Philosophy Zagreb, Croatia Vjeran Kerić, mag.phil. Department of Psychology and Cognitive
Consciousness as representation formation from a neural Darwinian perspective * Anna Kocsis, mag.phil. Institute of Philosophy Zagreb, Croatia Vjeran Kerić, mag.phil. Department of Psychology and Cognitive
Rules of apparent motion: The shortest-path constraint: objects will take the shortest path between flashed positions.
 Rules of apparent motion: The shortest-path constraint: objects will take the shortest path between flashed positions. The box interrupts the apparent motion. The box interrupts the apparent motion.
Rules of apparent motion: The shortest-path constraint: objects will take the shortest path between flashed positions. The box interrupts the apparent motion. The box interrupts the apparent motion.
Lateral view of human brain! Cortical processing of touch!
 Lateral view of human brain! Cortical processing of touch! How do we perceive objects held in the hand?! Touch receptors deconstruct objects to detect local features! Information is transmitted in parallel
Lateral view of human brain! Cortical processing of touch! How do we perceive objects held in the hand?! Touch receptors deconstruct objects to detect local features! Information is transmitted in parallel
Degree of freedom problem
 KINE 4500 Neural Control of Movement Lecture #1:Introduction to the Neural Control of Movement Neural control of movement Kinesiology: study of movement Here we re looking at the control system, and what
KINE 4500 Neural Control of Movement Lecture #1:Introduction to the Neural Control of Movement Neural control of movement Kinesiology: study of movement Here we re looking at the control system, and what
Mirror neurons. Romana Umrianova
 Mirror neurons Romana Umrianova The functional role of the parieto-frontal mirror circuit: interpretations and misinterpretations Giacomo Rizzolatti and Corrado Sinigaglia Mechanism that unifies action
Mirror neurons Romana Umrianova The functional role of the parieto-frontal mirror circuit: interpretations and misinterpretations Giacomo Rizzolatti and Corrado Sinigaglia Mechanism that unifies action
Rhythm and Rate: Perception and Physiology HST November Jennifer Melcher
 Rhythm and Rate: Perception and Physiology HST 722 - November 27 Jennifer Melcher Forward suppression of unit activity in auditory cortex Brosch and Schreiner (1997) J Neurophysiol 77: 923-943. Forward
Rhythm and Rate: Perception and Physiology HST 722 - November 27 Jennifer Melcher Forward suppression of unit activity in auditory cortex Brosch and Schreiner (1997) J Neurophysiol 77: 923-943. Forward
KINE 4500 Neural Control of Movement. Lecture #1:Introduction to the Neural Control of Movement. Neural control of movement
 KINE 4500 Neural Control of Movement Lecture #1:Introduction to the Neural Control of Movement Neural control of movement Kinesiology: study of movement Here we re looking at the control system, and what
KINE 4500 Neural Control of Movement Lecture #1:Introduction to the Neural Control of Movement Neural control of movement Kinesiology: study of movement Here we re looking at the control system, and what
Ch 5. Perception and Encoding
 Ch 5. Perception and Encoding Cognitive Neuroscience: The Biology of the Mind, 2 nd Ed., M. S. Gazzaniga, R. B. Ivry, and G. R. Mangun, Norton, 2002. Summarized by Y.-J. Park, M.-H. Kim, and B.-T. Zhang
Ch 5. Perception and Encoding Cognitive Neuroscience: The Biology of the Mind, 2 nd Ed., M. S. Gazzaniga, R. B. Ivry, and G. R. Mangun, Norton, 2002. Summarized by Y.-J. Park, M.-H. Kim, and B.-T. Zhang
Making Things Happen: Simple Motor Control
 Making Things Happen: Simple Motor Control How Your Brain Works - Week 10 Prof. Jan Schnupp wschnupp@cityu.edu.hk HowYourBrainWorks.net The Story So Far In the first few lectures we introduced you to some
Making Things Happen: Simple Motor Control How Your Brain Works - Week 10 Prof. Jan Schnupp wschnupp@cityu.edu.hk HowYourBrainWorks.net The Story So Far In the first few lectures we introduced you to some
Visual Context Dan O Shea Prof. Fei Fei Li, COS 598B
 Visual Context Dan O Shea Prof. Fei Fei Li, COS 598B Cortical Analysis of Visual Context Moshe Bar, Elissa Aminoff. 2003. Neuron, Volume 38, Issue 2, Pages 347 358. Visual objects in context Moshe Bar.
Visual Context Dan O Shea Prof. Fei Fei Li, COS 598B Cortical Analysis of Visual Context Moshe Bar, Elissa Aminoff. 2003. Neuron, Volume 38, Issue 2, Pages 347 358. Visual objects in context Moshe Bar.
Dr. Mark Ashton Smith, Department of Psychology, Bilkent University
 UMAN CONSCIOUSNESS some leads based on findings in neuropsychology Dr. Mark Ashton Smith, Department of Psychology, Bilkent University nattentional Blindness Simons and Levin, 1998 Not Detected Detected
UMAN CONSCIOUSNESS some leads based on findings in neuropsychology Dr. Mark Ashton Smith, Department of Psychology, Bilkent University nattentional Blindness Simons and Levin, 1998 Not Detected Detected
Manipulating the Experienced Onset of Intention after Action Execution
 Manipulating the Experienced Onset of Intention after Action Execution Hakwan C. Lau 1,2, Robert D. Rogers 2, and Richard E. Passingham 1,2 Abstract & Using transcranial magnetic stimulation (TMS), we
Manipulating the Experienced Onset of Intention after Action Execution Hakwan C. Lau 1,2, Robert D. Rogers 2, and Richard E. Passingham 1,2 Abstract & Using transcranial magnetic stimulation (TMS), we
Introduction. Chapter The Perceptual Process
 Chapter 1 Introduction Most of us take for granted our ability to perceive the external world. However, this is no simple deed at all. Imagine being given a task of designing a machine that can perceive,
Chapter 1 Introduction Most of us take for granted our ability to perceive the external world. However, this is no simple deed at all. Imagine being given a task of designing a machine that can perceive,
Attention Response Functions: Characterizing Brain Areas Using fmri Activation during Parametric Variations of Attentional Load
 Attention Response Functions: Characterizing Brain Areas Using fmri Activation during Parametric Variations of Attentional Load Intro Examine attention response functions Compare an attention-demanding
Attention Response Functions: Characterizing Brain Areas Using fmri Activation during Parametric Variations of Attentional Load Intro Examine attention response functions Compare an attention-demanding
On the Time of Peripheral Sensations and Voluntary Motor Actions. Text
 On the Time of Peripheral Sensations and Voluntary Motor Actions DOUGLAS M. SNYDER ABSTRACT Libet's notions of backwards referral for peripheral sensations and unconscious cerebral initiative accompanying
On the Time of Peripheral Sensations and Voluntary Motor Actions DOUGLAS M. SNYDER ABSTRACT Libet's notions of backwards referral for peripheral sensations and unconscious cerebral initiative accompanying
The influence of visual motion on fast reaching movements to a stationary object
 Supplemental materials for: The influence of visual motion on fast reaching movements to a stationary object David Whitney*, David A. Westwood, & Melvyn A. Goodale* *Group on Action and Perception, The
Supplemental materials for: The influence of visual motion on fast reaching movements to a stationary object David Whitney*, David A. Westwood, & Melvyn A. Goodale* *Group on Action and Perception, The
Why is our capacity of working memory so large?
 LETTER Why is our capacity of working memory so large? Thomas P. Trappenberg Faculty of Computer Science, Dalhousie University 6050 University Avenue, Halifax, Nova Scotia B3H 1W5, Canada E-mail: tt@cs.dal.ca
LETTER Why is our capacity of working memory so large? Thomas P. Trappenberg Faculty of Computer Science, Dalhousie University 6050 University Avenue, Halifax, Nova Scotia B3H 1W5, Canada E-mail: tt@cs.dal.ca
Introduction to Computational Neuroscience
 Introduction to Computational Neuroscience Lecture 7: Network models Lesson Title 1 Introduction 2 Structure and Function of the NS 3 Windows to the Brain 4 Data analysis 5 Data analysis II 6 Single neuron
Introduction to Computational Neuroscience Lecture 7: Network models Lesson Title 1 Introduction 2 Structure and Function of the NS 3 Windows to the Brain 4 Data analysis 5 Data analysis II 6 Single neuron
LEAH KRUBITZER RESEARCH GROUP LAB PUBLICATIONS WHAT WE DO LINKS CONTACTS
 LEAH KRUBITZER RESEARCH GROUP LAB PUBLICATIONS WHAT WE DO LINKS CONTACTS WHAT WE DO Present studies and future directions Our laboratory is currently involved in two major areas of research. The first
LEAH KRUBITZER RESEARCH GROUP LAB PUBLICATIONS WHAT WE DO LINKS CONTACTS WHAT WE DO Present studies and future directions Our laboratory is currently involved in two major areas of research. The first
Ch 5. Perception and Encoding
 Ch 5. Perception and Encoding Cognitive Neuroscience: The Biology of the Mind, 2 nd Ed., M. S. Gazzaniga,, R. B. Ivry,, and G. R. Mangun,, Norton, 2002. Summarized by Y.-J. Park, M.-H. Kim, and B.-T. Zhang
Ch 5. Perception and Encoding Cognitive Neuroscience: The Biology of the Mind, 2 nd Ed., M. S. Gazzaniga,, R. B. Ivry,, and G. R. Mangun,, Norton, 2002. Summarized by Y.-J. Park, M.-H. Kim, and B.-T. Zhang
Physiology of Tactile Sensation
 Physiology of Tactile Sensation Objectives: 1. Describe the general structural features of tactile sensory receptors how are first order nerve fibers specialized to receive tactile stimuli? 2. Understand
Physiology of Tactile Sensation Objectives: 1. Describe the general structural features of tactile sensory receptors how are first order nerve fibers specialized to receive tactile stimuli? 2. Understand
Structure and Function of the Auditory and Vestibular Systems (Fall 2014) Auditory Cortex (3) Prof. Xiaoqin Wang
 Auditory Cortex (3) Prof. Xiaoqin Wang") 580.626 Structure and Function of the Auditory and Vestibular Systems (Fall 2014) Auditory Cortex (3) Prof. Xiaoqin Wang Laboratory of Auditory Neurophysiology Department of Biomedical Engineering Johns
580.626 Structure and Function of the Auditory and Vestibular Systems (Fall 2014) Auditory Cortex (3) Prof. Xiaoqin Wang Laboratory of Auditory Neurophysiology Department of Biomedical Engineering Johns
Visual Processing (contd.) Pattern recognition. Proximity the tendency to group pieces that are close together into one object.
 Pattern recognition. Proximity the tendency to group pieces that are close together into one object.") Objectives of today s lecture From your prior reading and the lecture, be able to: explain the gestalt laws of perceptual organization list the visual variables and explain how they relate to perceptual
Objectives of today s lecture From your prior reading and the lecture, be able to: explain the gestalt laws of perceptual organization list the visual variables and explain how they relate to perceptual
Identify these objects
 Pattern Recognition The Amazing Flexibility of Human PR. What is PR and What Problems does it Solve? Three Heuristic Distinctions for Understanding PR. Top-down vs. Bottom-up Processing. Semantic Priming.
Pattern Recognition The Amazing Flexibility of Human PR. What is PR and What Problems does it Solve? Three Heuristic Distinctions for Understanding PR. Top-down vs. Bottom-up Processing. Semantic Priming.
Carnegie Mellon University Annual Progress Report: 2011 Formula Grant
 Carnegie Mellon University Annual Progress Report: 2011 Formula Grant Reporting Period January 1, 2012 June 30, 2012 Formula Grant Overview The Carnegie Mellon University received $943,032 in formula funds
Carnegie Mellon University Annual Progress Report: 2011 Formula Grant Reporting Period January 1, 2012 June 30, 2012 Formula Grant Overview The Carnegie Mellon University received $943,032 in formula funds
Synfire chains with conductance-based neurons: internal timing and coordination with timed input
 Neurocomputing 5 (5) 9 5 www.elsevier.com/locate/neucom Synfire chains with conductance-based neurons: internal timing and coordination with timed input Friedrich T. Sommer a,, Thomas Wennekers b a Redwood
Neurocomputing 5 (5) 9 5 www.elsevier.com/locate/neucom Synfire chains with conductance-based neurons: internal timing and coordination with timed input Friedrich T. Sommer a,, Thomas Wennekers b a Redwood
Chapter 6. Attention. Attention
 Chapter 6 Attention Attention William James, in 1890, wrote Everyone knows what attention is. Attention is the taking possession of the mind, in clear and vivid form, of one out of what seem several simultaneously
Chapter 6 Attention Attention William James, in 1890, wrote Everyone knows what attention is. Attention is the taking possession of the mind, in clear and vivid form, of one out of what seem several simultaneously
Importance of Deficits
 Importance of Deficits In complex systems the parts are often so integrated that they cannot be detected in normal operation Need to break the system to discover the components not just physical components
Importance of Deficits In complex systems the parts are often so integrated that they cannot be detected in normal operation Need to break the system to discover the components not just physical components
Visual Selection and Attention
 Visual Selection and Attention Retrieve Information Select what to observe No time to focus on every object Overt Selections Performed by eye movements Covert Selections Performed by visual attention 2
Visual Selection and Attention Retrieve Information Select what to observe No time to focus on every object Overt Selections Performed by eye movements Covert Selections Performed by visual attention 2
Hippocampal mechanisms of memory and cognition. Matthew Wilson Departments of Brain and Cognitive Sciences and Biology MIT
 Hippocampal mechanisms of memory and cognition Matthew Wilson Departments of Brain and Cognitive Sciences and Biology MIT 1 Courtesy of Elsevier, Inc., http://www.sciencedirect.com. Used with permission.
Hippocampal mechanisms of memory and cognition Matthew Wilson Departments of Brain and Cognitive Sciences and Biology MIT 1 Courtesy of Elsevier, Inc., http://www.sciencedirect.com. Used with permission.
Auditory Scene Analysis
 1 Auditory Scene Analysis Albert S. Bregman Department of Psychology McGill University 1205 Docteur Penfield Avenue Montreal, QC Canada H3A 1B1 E-mail: bregman@hebb.psych.mcgill.ca To appear in N.J. Smelzer
1 Auditory Scene Analysis Albert S. Bregman Department of Psychology McGill University 1205 Docteur Penfield Avenue Montreal, QC Canada H3A 1B1 E-mail: bregman@hebb.psych.mcgill.ca To appear in N.J. Smelzer
THE BRAIN HABIT BRIDGING THE CONSCIOUS AND UNCONSCIOUS MIND
 THE BRAIN HABIT BRIDGING THE CONSCIOUS AND UNCONSCIOUS MIND Mary ET Boyle, Ph. D. Department of Cognitive Science UCSD How did I get here? What did I do? Start driving home after work Aware when you left
THE BRAIN HABIT BRIDGING THE CONSCIOUS AND UNCONSCIOUS MIND Mary ET Boyle, Ph. D. Department of Cognitive Science UCSD How did I get here? What did I do? Start driving home after work Aware when you left
Cognitive Neuroscience Section 4
 Perceptual categorization Cognitive Neuroscience Section 4 Perception, attention, and memory are all interrelated. From the perspective of memory, perception is seen as memory updating by new sensory experience.
Perceptual categorization Cognitive Neuroscience Section 4 Perception, attention, and memory are all interrelated. From the perspective of memory, perception is seen as memory updating by new sensory experience.
University of Padova, Padova, Italy b University College London, London, UK PLEASE SCROLL DOWN FOR ARTICLE
 This article was downloaded by: [Agrillo, Christian] On: 15 March 2010 Access details: Access Details: [subscription number 919880202] Publisher Psychology Press Informa Ltd Registered in England and Wales
This article was downloaded by: [Agrillo, Christian] On: 15 March 2010 Access details: Access Details: [subscription number 919880202] Publisher Psychology Press Informa Ltd Registered in England and Wales
Understanding Users. - cognitive processes. Unit 3
 Understanding Users - cognitive processes Unit 3 Why do we need to understand users? Interacting with technology involves a number of cognitive processes We need to take into account Characteristic & limitations
Understanding Users - cognitive processes Unit 3 Why do we need to understand users? Interacting with technology involves a number of cognitive processes We need to take into account Characteristic & limitations
Sperling conducted experiments on An experiment was conducted by Sperling in the field of visual sensory memory.
 Levels of category Basic Level Category: Subordinate Category: Superordinate Category: Stages of development of Piaget 1. Sensorimotor stage 0-2 2. Preoperational stage 2-7 3. Concrete operational stage
Levels of category Basic Level Category: Subordinate Category: Superordinate Category: Stages of development of Piaget 1. Sensorimotor stage 0-2 2. Preoperational stage 2-7 3. Concrete operational stage
Thesis. Attentional modulation of frequency specific distortions of event time. Utrecht University
 Utrecht University Master Psychology, Toegepaste Cognitieve Psychologie. Thesis Attentional modulation of frequency specific distortions of event time Pascal Fruneaux, 3269264 26-06- 2012 Supervisors:
Utrecht University Master Psychology, Toegepaste Cognitieve Psychologie. Thesis Attentional modulation of frequency specific distortions of event time Pascal Fruneaux, 3269264 26-06- 2012 Supervisors:
Unconscious determinants of free decisions in the human brain
 Unconscious determinants of free decisions in the human brain Chun Siong Soon 1,2, Marcel Brass 1,3, Hans-Jochen Heinze 4 & John-Dylan Haynes* 1,2 1 Max Planck Institute for Human Cognitive and Brain Sciences,
Unconscious determinants of free decisions in the human brain Chun Siong Soon 1,2, Marcel Brass 1,3, Hans-Jochen Heinze 4 & John-Dylan Haynes* 1,2 1 Max Planck Institute for Human Cognitive and Brain Sciences,
Do We Have a Common Mechanism for Measuring Time in the Hundreds of Millisecond Range? Evidence From Multiple-Interval Timing Tasks
 J Neurophysiol 99: 939 949, 28. First published December 19, 27; doi:1.1152/jn.1225.27. Do We Have a Common Mechanism for Measuring Time in the Hundreds of Millisecond Range? Evidence From Multiple-Interval
J Neurophysiol 99: 939 949, 28. First published December 19, 27; doi:1.1152/jn.1225.27. Do We Have a Common Mechanism for Measuring Time in the Hundreds of Millisecond Range? Evidence From Multiple-Interval
Optical Illusions and Human Visual System: Can we reveal more? CIS Micro-Grant Executive Report and Summary of Results
 Optical Illusions and Human Visual System: Can we reveal more? CIS Micro-Grant 2011 Executive Report and Summary of Results Prepared By: Principal Investigator: Siddharth Khullar 1, Ph.D. Candidate (sxk4792@rit.edu)
Optical Illusions and Human Visual System: Can we reveal more? CIS Micro-Grant 2011 Executive Report and Summary of Results Prepared By: Principal Investigator: Siddharth Khullar 1, Ph.D. Candidate (sxk4792@rit.edu)
Distortions of Subjective Time Perception Within and Across Senses
 Distortions of Subjective Time Perception Within and Across Senses Virginie van Wassenhove 1 *, Dean V. Buonomano 2,3, Shinsuke Shimojo 1, Ladan Shams 2 1 Division of Biology, California Institute of Technology,
Distortions of Subjective Time Perception Within and Across Senses Virginie van Wassenhove 1 *, Dean V. Buonomano 2,3, Shinsuke Shimojo 1, Ladan Shams 2 1 Division of Biology, California Institute of Technology,
The role of phase synchronization in memory processes
 The role of phase synchronization in memory processes Juergen Fell and Nikolai Axmacher Abstract In recent years, studies ranging from single-unit recordings in animals to electroencephalography and magnetoencephalography
The role of phase synchronization in memory processes Juergen Fell and Nikolai Axmacher Abstract In recent years, studies ranging from single-unit recordings in animals to electroencephalography and magnetoencephalography
Flexible Retinotopy: Motion-Dependent Position Coding in the Visual Cortex
 Flexible Retinotopy: Motion-Dependent Position Coding in the Visual Cortex David Whitney,* 1 Herbert C. Goltz, 2 Christopher G. Thomas, 1 Joseph S. Gati, 2 Ravi S. Menon, 2 Melvyn A. Goodale 1 1 The Department
Flexible Retinotopy: Motion-Dependent Position Coding in the Visual Cortex David Whitney,* 1 Herbert C. Goltz, 2 Christopher G. Thomas, 1 Joseph S. Gati, 2 Ravi S. Menon, 2 Melvyn A. Goodale 1 1 The Department
Introductory Motor Learning and Development Lab
 Introductory Motor Learning and Development Lab Laboratory Equipment & Test Procedures. Motor learning and control historically has built its discipline through laboratory research. This has led to the
Introductory Motor Learning and Development Lab Laboratory Equipment & Test Procedures. Motor learning and control historically has built its discipline through laboratory research. This has led to the
Long-range dependence in human movements: detection, interpretation, and modeling perspectives
 Long-range dependence in human movements: detection, interpretation, and modeling perspectives Didier Delignières E.A. 2991 Motor Efficiency and Deficiency University Montpellier I, France Telephone: +33
Long-range dependence in human movements: detection, interpretation, and modeling perspectives Didier Delignières E.A. 2991 Motor Efficiency and Deficiency University Montpellier I, France Telephone: +33
Dynamics of Color Category Formation and Boundaries
 Dynamics of Color Category Formation and Boundaries Stephanie Huette* Department of Psychology, University of Memphis, Memphis, TN Definition Dynamics of color boundaries is broadly the area that characterizes
Dynamics of Color Category Formation and Boundaries Stephanie Huette* Department of Psychology, University of Memphis, Memphis, TN Definition Dynamics of color boundaries is broadly the area that characterizes
Sparse Coding in Sparse Winner Networks
 Sparse Coding in Sparse Winner Networks Janusz A. Starzyk 1, Yinyin Liu 1, David Vogel 2 1 School of Electrical Engineering & Computer Science Ohio University, Athens, OH 45701 {starzyk, yliu}@bobcat.ent.ohiou.edu
Sparse Coding in Sparse Winner Networks Janusz A. Starzyk 1, Yinyin Liu 1, David Vogel 2 1 School of Electrical Engineering & Computer Science Ohio University, Athens, OH 45701 {starzyk, yliu}@bobcat.ent.ohiou.edu
Neural Coding. Computing and the Brain. How Is Information Coded in Networks of Spiking Neurons?
 Neural Coding Computing and the Brain How Is Information Coded in Networks of Spiking Neurons? Coding in spike (AP) sequences from individual neurons Coding in activity of a population of neurons Spring
Neural Coding Computing and the Brain How Is Information Coded in Networks of Spiking Neurons? Coding in spike (AP) sequences from individual neurons Coding in activity of a population of neurons Spring