An inquiry of mediating events in the solution of the matching-to-sample problem.
|
|
|
- Sherman Dalton
- 5 years ago
- Views:
Transcription
1 University of Massachusetts Amherst Amherst Masters Theses February An inquiry of mediating events in the solution of the matching-to-sample problem. Richard Paul Mack University of Massachusetts Amherst Follow this and additional works at: Mack, Richard Paul, "An inquiry of mediating events in the solution of the matching-to-sample problem." (1971). Masters Theses February Retrieved from This thesis is brought to you for free and open access by ScholarWorks@UMass Amherst. It has been accepted for inclusion in Masters Theses February 2014 by an authorized administrator of ScholarWorks@UMass Amherst. For more information, please contact scholarworks@library.umass.edu.


2

3 AN INQUIRY OF MEDIATING EVENTS IN THE SOLUTION OF THE MATCHING-TO- SAMPLE PROBLEM A Thesis Presented by Richard Paul Mack Submitted to the Graduate School of the University of Massachusetts in partial fulfillment of the requirements for the degree of MASTER OF SCIENCE July, 1971 Major Subject: Psycho J jgy

 mead of the i Department t n a (Member")
4 AN INQUIRY OF MEDIATING EVENTS THE SOLUTION OF THE MATCHING -TO- SAMPLE PROBLEM A Thesis by Richard Paul Mack Approved as to style and content by: (ChairtrjJan of the Committee) mead of the i Department t n a (Member July, 1971


5 iii TABLE OF CONTENTS Page LIST OF TABLES iv LIST OF FIGURES v LIST OF APPENDICES vi INTRODUCTION 1 METHOD 16 RESULTS 22 DISCUSSION 26 REFERENCES 50 I
6 IV LIST OF TABLES Page 1. Analysis of variance of savings score for transfer 40 1

7 V LIST OF FIGURES Page 1. Percentage correct matching in acquisition transfer, FR 1, and R-G testing Latencies on ST responding ; Percentage correct in delay testing 43


8 vi LIST OF APPENDICES I Percentage correct on probe and regular trials Page II Analysis of variance of delay testing 45 III F-values of contrasts for delay testing 46 IV Distribution of errors on first session of transfer. 47 V Percentage correct matching in acguisition, transfer, FR 1, and R-G testing for individual subjects. 48 VI Percentage correct matching in delay for individual subjects 49
 only the differential number of responses could be employed.")
9 ABSTRACT Pigeons were trained on a zero-delay matching- tosample task under one of three conditions; (1) only the hue oi the standard stimulus (ST) could be used as the discriminative stimulus, (2) either the hue or differential number of responses on the ST could be used, (3) only the differential number of responses could be employed. Following acquisition training all subjects were given delay testing and then transferred to the acquisition procedure but using new hues. The results demonstrated the groups with the hue on the ST had superior performance in acquisition, delay, and transfer, compared with the group that had only differential responses. In addition, the group which could use either the hue or differential responding as the discriminative stimulus appeared to employ the hue dimension. However, it was found that by having more than one response required on the ST, performance under acquisition, delay, and transfer procedures was much superior to previous research. The results were discussed in terms of a modification of the coding hypothesis (Camming, Berryraan and Cohen, 1965).

 induced by these requirements provides the basis for correct matching behavior.")
10 INTRODUCTION The matching-to-sample (MTS) task, as pointed out by Ginsberg (1957), is a "special case" of a conditional discrimination problem. It is "special" in that the conditional stimulus is identical to the stimulus the subject is to choose. Thus, in the MTS task, the subject can use either the physical properties of the sample (ST) stimulus directly, or employ some mediating event as the discriminative stimulus. The present study is an attempt to isolate the functional discriminative stimulus in the MTS task. The study varied the response requirements to the STs, and hypothesized the information (i.e. resnonse produced cues or stimuli) induced by these requirements provides the basis for correct matching behavior. Before proceeding to the proposed method of testing this hypothesis a description of the MTS task will be given, along with a brief review of the theories and data leading to this research. The matching- to-samole task. In the MTS task S_ is presented with three stimuli on each trial. The center stimulus, referred to as the sample or standard stimulus (ST), is identical to one of the other two stimuli lateral to it. These lateral stimuli are called comparison stimuli (COs). The typical trial begins by the presentation of the ST. Usually, a response to the ST is required for the presentation of the COs. 1
 where many different stimuli are used.")

11 2 After the COs are presented,s is then reinforced for a response to the CO which matches the ST. It should be noted that a stimulus population of only two or three stimulus values are usually used across all trials in contrast to learning sets (Harlow, 1949) where many different stimuli are used. The particular stimulus value used as the ST and the position of this value as a CO varies randomly over trials Two variations of this paradigm are nonmatching and amatching (Ginsberg, 1957). Nonmatching differs from matching in that S^ is reinforced for choosing the CO which is different from the ST. With amatching the STs are different from the COs. For example, if the ST is green, the correct CO is yellow, if the ST is red, the correct CO is blue. All three of these procedures; matching, nonmatching, and amatching are conditional discrimination tasks. Theories and review. Yerkes and Nissen (19 39) hypothesized that a crude "symbolic process" was used in solving the MTS task. They describe the process as: a differential and usually implicit response established by previous training, whose "meaning", as exemplified by its positive or negative valence, is extremely labile. The sign response is- differential in the sen:- and to the extent it varies for different stimuli... lability of the consequence of the sign response is manifest by the readiness with which approach and avoidance may replace one another, (pp» ) 1


12 3 Nissen, Blum and Blum (1948) suggested three mechanisms that could be operating independently or conjunctively in the MTS task. One is the "abstract generalization mechanism," and appears quite similar to the "symbolic process" hypothesis. This mechanism assumes the approach response is not conditioned to the physical characteristics of the stimulus but to a secondary neural response of "similarity." Furthermore, there is an avoidance response conditioned to difference. The previous two hypotheses suggest the physical characteristics of the stimuli to-be-matched are of secondary importance, while some mediating event is of primary importance Unfortunately, they offer inadequate or no hypothetical constructs through which the mediating event functions. The words "symbolic" and "abstract" imply the Ss may be operating on a "higher" level of learning or using a conceptual scheme. One such scheme could be the "principle of sameness" (Weinstein, 1941); that is, Ss would choose the CO which is the "same" as the ST. If such a principle were operating, one -would expect positive transfer of matching to a new set of stimuli. Most transfer studies in MTS involve initial training tc a particular set cf stimuli to a level of competent matching, followed by trials with a new stimulus (or stimuli). It is then noted whether performance is lower than expected by chance (negative transfer), chance, or above chance (positive transfer). Another measure would be a savings score. Most data from pigeons fail to demon-
 which would be indicative of using a conceptual scheme.")
13 strate enough transfer to warrant the use of a conceptual scheme by Ss. Ginsberg (1957) initially trained Ss on red and green stimuli and then switched to blue and yellow. In contrast to most data, one S_ appeared to demonstrate complete positive transfer (i.e., a score of zero) which would be indicative of using a conceptual scheme. However, the data of this S_ is questionable in that a correction procedure was used. The S could have had numerous errors which were not taken into account in the transfer functions. The other two Ss took 360 and 680 less trials to learn the transfer task than the original task. Although there is positive transfer, it doesn't appear significantly strong enough as would be required by conceptual scheme interpretation. A nonspecific transfer (i.e., familiarity with the apparatus and the MTS task) can easily account: for this faster acquisition during transfer. In another transfer study, Gumming and Berryman (1961) trained Ss on a MTS task involving three hues: blue, red, and green. After 22 sessions a yellow light was substituted ^ Kr* vo,,^ Dorfnrn^nro nn n pw hue droooed to chance while matching of other hues remained unimpaired. Again, the data suggest the lack of a conceptual scheme. In a nonmatching task S would be required to learn the "principle of difference" in contrast to the "principle of
 did a simultaneous nonmatching task, (In simultaneous tasks the ST and COS are all present when the choice response is made.")
14 5 sameness" for Hatching. Intuitively there should be no difference between these two principles in terms of the steepness of the acquisition function and amount of transfer. Berryman, Crowning, Cohen and Johnson (1965) did a simultaneous nonmatching task, (In simultaneous tasks the ST and COS are all present when the choice response is made. ) in which Ss were initially trained on red, green, and blue stimuli with yellow being substituted for blue during transfer. First, they compared the acquisition functions of this task with a previous matching study (Cumming and Berryman, 1961) and found the matching function to be much steeper. Again there is no apparent reason why these functions should differ if Ss were using conceptual schemes. Secondly, they foiled to find the amount of transfer predicted by the "principle of difference." In addition to the "abstract generalization mechanism," Nissen, Blum and Blum (1948) suggested a psychologically simpler mechanism; "the perceptual mechanism." In matching, _S would solve the task by learning that responses to the large, instead of the small, portion of the stimulus display is correct. Of course the opposite would be true for nonmatching. It is evident that this mochanism is not capable of handling delayed matching (DMTS) tasks (the COs are presented some time after the ST is removed, thus the choice response is done without all the stimuli), in that it re-

15 6 quires the ST and COs to be simultaneously present when the choice response is made. One may argue that the "perception mechanism" is being used in the simultaneous matching task and some other mechanism in delayed matching. That is, assuming an organism attempts to maximize its efficiency in dealing with the environment the "perceptual mechanism" fulfills this requirement in a simultaneous procedure. On the other hand, a DMTS task would require some mediating event in that the information provided by the ST must span the temporal gap and be available during the choice response. Berryman, Cumming, and Nevin (1963) found suggestions of a mediating event in a simultaneous MTS task in which there was positive transfer from the simultaneous procedure to the delayed procedure. If Ss were not solving the simultaneous task by employing a mediator, zero or negative transfer would be exoected upon initiation of DMTS. On the other hand, as demonstrated by Blough (1959) and Sacks (1971), Ss given extensive training on zero delay matching (ST goes off and COs come on simultaneously) dropped to chance performance and failed to recover after additional training when switched to delayed MTS. The "perception" hypothesis would also predict that increased exposure or attention to the stimulus display would induce better performance. Berryman, Cumming, and Nevin x

16 7 (1963) required an FR5 on the ST in order to light the COs in an attempt to increase the time Ss would be attending to the stimulus display. Results showed that longer exposure to the ST did not result in better performance compared to Ss that made only one response to the ST. Cumming, Berryman, and Cohen (1965) and Berryman, Cumming, Cohen and Johnson (1965) have made use of the "coding hypothesis" (Lawrence, 1963) in interpreting both matching and nonmatching tasks. The hypothesis first assumes there is no direct relation between the physical properties of the ST and the response to the correct CO, but there is always an intervening, or mediating, event. This event is an assumed implicit response, in which initially there is a trial and error period of coding responses until one is encountered that brings consistent reinforcement. The response produces a "code item" which serves as a representation of the original input. The "code item" of this implicit response then serves as the S D for- the correct matching behavior. Cumming, Berryman, and Cohen (1965) describe it in the following way learns to make an appropriate coding response (rx) in the presence of a particular standard stimulus (ST ). In the presence of rx, the two comparison stimuli (CO r and CO, ) are presented. The chain ST x - rx/co"^ - R is reinforced, while ST X - rx/co X - R X is extinguished. Within x y y
 y x Sacks (1971) found evidence for the coding hypothesis in a zero delay MTS task. Initial training was on red and green stimuli followed by a transfer to blue and yellow.")
17 8 the same situation, coding responses to ST are also being acquired, with ST. - ry/coy - Ry reinforced, and ST - ry^co - R extinguished. (p. 437) y x Sacks (1971) found evidence for the coding hypothesis in a zero delay MTS task. Initial training was on red and green stimuli followed by a transfer to blue and yellow. Acquisition to a criteria of 90% matching was reached sooner for the latter stimuli; but only to a level which suggests practice effects. After this, Ss were switched back to red and green stimuli. Results showed that Ss were around the 90% matching criteria. Next Ss were given sessions involving all four stimuli in which stimuli never before used together were presented on the same trial (e.g., green and yellow). Initial performances for the Ss were around 70-30%, and criteria was generally reached in several sessions. This data suggests that Ss were learning something other than particular responses to certain stimulus arrangements of the hues on the stimulus display panel. A coding hypothesis would predict that once a stimulus is coded it can easily be used within the context of other uncoded or coded items, as was demonstrated in this study. Tne Cumming and Berryman (1961) study mentioned earlier also supported the coding position in which there was no effect upon green and red matching when a yellow stimulus was substituted for blue.


18 9 Cumming, Berryman, and Cohen (1365), again substituting yellow for blue, examined three types of trials: (1) old stimuli as both ST and COs ; (2) yellow appears only as a CO; (3) yellow is both an ST and a CO. Upon transfer, there was no effect upon the red and green accuracy on trials like number 1 above. When one CO was yellow, but the ST had not been yellow, two Ss performed at the same accuracy as they had with blue, while one _S improved. On the third type of trial, performance dropped from 92% accuracy to 28%. For one of the Ss at least, it appeared that the yellow ST took on the code for red. That is, on trials when the ST was yellow and the COs were yellow and red, S_ consistently chose the red CO. More support for the coding hypothesis was found in a transfer study (Berryman, Cumming, Cohen and Johnson, 1965) using a simultaneous nonmatching task. In addition, the study provides evidence for the rule Ss may be using in solving the task. S_s were trained on red, green, and blue stimuli with yellow being substituted for blue during transfer. During transfer the S rule would predict no change in performance because 3_ is just to avoid the ST informs,s to select red or blue, but neither are present. The results supported the S J rule. The experimenters claim the slower acquisition rate of nonmatching as compared to Matching (Cumming and 3erryman, 1961) is consistent with the coding hypothesis. In the nonmatching task, S_s have a
 is very similar to the coding hypothesis, but is more specific in that it provided a hypothetical construct through which the mediator may be functioning.")
 was presented than when the other ST (steady light) was displayed.")
19 10 more difficult code to acquire (i.e., S red or green) while matching has only one hue per code (i.e., S D red). An hypothesis by Blough (1959) is very similar to the coding hypothesis, but is more specific in that it provided a hypothetical construct through which the mediator may be functioning. Blough noticed that pigeons in a DMTS task developed chains of behavior between the offset of the ST and onset of the COs. Furthermore, some Ss would display a chain of responses distinctly different from the other chain when one ST (flickering light) was presented than when the other ST (steady light) was displayed. That is, _Ss developed response chains that were specific to the ST presented. Even more significant is that the Ss which demonstrated stimulus specific chains had greater matching accuracy for long delays than Ss that failed to acquire these stimulus specific chains. Blough believed the chains provided a mechanism by. which information or feedback could bridge the temporal gap and be available during presentation of the COs. Although Blough has been the only one to report such distinct evert response chains, Gumming, Berryman and Cohen (1965) mentioned that Ss often develop discriminably different ways of responding to ST huej. With the exception of the requirement of a response to the ST tor the presentation of the COs (by definition of the task) it has been shown that responses to the ST are not necessary for MTS
 that required ST responses for presentation of the COs.")

20 11 (Yerkes and Nissen, 1939). However, they do have a definite effect upon performance. Eckerman, Lanson, and Curaming (1968) used two procedures to test for effects of ST responding. The first compared a simultaneous MTS task in which no response was required to the ST with a zero-delay task (Gumming and Berryman, 1961) that required ST responses for presentation of the COs. Asymtotic performance was not reached until the 45th session in the "no ST required study/' while the "ST response required" procedure reached asymtote by the tenth session. The better performance occurred even though zerodelay MTS is typically considered a more difficult task than simultaneous MTS. The second precedure used a sequence of changes in the ST response requirements. The requirements were as follows: (1) NSTR - no ST response required; (2) PRO 10 SEC - in which responses to the ST during the IT1 delayed the onset of the next trial and the NSTR was still operating; (3) STR - a recovery stage in which ST responses were required; (4) CANCEL (C) - any response to the ST during the trial cancelled zhe trial by extinguishing Lights and having a 25 second ITI; (5) back to an STR contingency. The results differed between the three S_s. One S_ dropped in accuracy fro 100% to 15% matching wh^n ST responses were weakened (i.e., institution of NSTR, CRO, and C). The second

21 12 S_ maintained high performance during the NSTR but dropped to 50% matching when the DRO or C conditions were introduced. The last never dropped to chance performance perhaps because a "chain of pecking around the center key at the time of its onset was noted " It appears, at least for pigeons, that ST responses are not necessary for matching, but they do speed up acquisition and maintain more accurate performance that if the responses were absent. Since it was a simultaneous MTS task it raises the question of the tenability of the "perception mechanism" (Nissen et al., 1948). In all conditions, S^s had equal opportunity to view the stimulus complex (i.e., all three and any other visual stimuli). Yet, performance differed by the requirement and nonrequirernent of a response to the ST. One may argue the ST response requirement induced greater "attention" to it, resulting in better performance. However, as mentioned earlier, Berryman, Cumming and Nevin (1963) required an FR5 on the ST in order to light the COs. Results showed that longer exposure to the ST did not result in better performance compared to Sis that made only one response the ST. How do responses to the ST facilitate performance? How can they make a normally more difficult task (i.e., delayed MTS) be acquired faster than an easier task (i.e., simultaneous matching)? Do these responses serve as discriminative stimuli


22 13 for the choice response, and if so, what is the nature of the stimuli? Some studies outside the NTS literature suggest that responses early in a response sequence can provide information for subsequent responding. Responses as Discriminati ve Stimuli. Schulte ( using rats, attempted to determine if the first component (R^) of a response sequence could exert control on the second component (R^ ) * was a barpressing response in which + R.j required a particular force to depress the bar, and required a different force. ' was consistently followed by R2 and reinforcement, while was followed by nonreinf orcemen t. cons is ted of approaching and nos ing open two doors for reinforcement* The dependent measure was the speed (reciprocal of latency) of k 0 * It was assumed that the discrimination could be solved only by the stimuli arising from the differential consequences of R. and R.^~. Results showed that Ss trained with Rj* and R^~ acquired the discrimination faster than control Ss. Furthermore, they displayed a generalization gradient that covaried with the differences between cue 1C1 C6S Utii. xiiiiiy cm^.. Three experiments have provided additional evidence suggesting that responses (or their stimulus consequences) serve as discriminative stimuli. Pliskoff and Goldiamond (1966) had pigeons respond on a red key at PR's 95-5 to FR's If S responded on the larger schedule, the was to peck the red I

23 14 key to receive reinforcement. If the FR was small, S was to respond to the left. FR was a control. Results showed that S's responses became more frequent the smaller the difference between the mixed FR's. After initial training on 1-sec delay between the red light terminating and the white lights being presented, Ss were tested over various delays. However, in this study there is a confounding between the number of responses in the FR and the amount cf time needed to complete the FR; either of these could be the effective S. Rilling (1967) isolated these variables and found the percentage of correct responses to varv with the number of responses rather than the length of time to complete these schedules. Rilling (1968) in another study established a spatial discrimination between FR's On probe trials a TO was introduced after the twenty-sixth response on the FR 50 and after the first response on the FR 25. TO durations were 0.5, 1, 2, 5, 10, and 30 seconds. As predicted the TO after the twenty-sixth response impaired discrimination in that Ss would perform the response conditional upon the FR lb schedule. Furthermore, the probability cf ir.ccrrccc responding increased with longer TO's. A review of the MTS literature suggests a mediating event may be used in solution to the task, but provides little evidence as to the nature of the mediator. In light of Biough's (1959) differential response chains, the effect


24 15 of the requirement and nonrequirement of ST response (Eckerman, Lanson, and Cumming, 1968), and the informative responding literature, it is possible to hypothesize that the mediator consists of response-produced cues arising from differential responding to the ST 1 s This study attempts to evaluate the differences between conditional hue discrimination tasks in a between-groups design. The first group is an MTS group in which the ST hues are the conditional stimuli. The second group is like the first, except a certain number of responses (i.e. FR) to the ST, specific to the hues, is required to terminate the ST and present the COs. The last group is like the second except that the ST is always a white light; thus, the conditional stimuli are response produced cues. Although the first group may have differential responding to the hues (hen differential response-produced cues), it is assumed that the second group has a greater probability of developing differential response-produced cues due to the response requirements to the ST. The last group tests whether responsenrnrlnc.pd cues are sufficient to solve the task. After acquisition of the task, delay, transfer, and probe techniques are employed in an attempt to map some of the characteristics of the mediating process.

 modified so that three keys were on the stimulus")
25 METHOD Subj ects Four male and 12 female White Carneaux pigeons obtained from the Palmetto Pigeon Plant of Sumter, South Carolina were given preliminary training. One male and 3 females were then randomly assigned to each group. Subjects ( Ss ) were maintained at 75-80% of their free feeding weight and were housed in individual cages with water continuously available. The reinforcement mixture and home cage feed was Purina Pigeon Grains Apparatus The apparatus was a Lehigh Vally Electronics pigeon chamber (#1519) modified so that three keys were on the stimulus display panel. The three keys, illuminated by IEE projectors, were horizontally located 5.5 cm apart and 21 cm above the floor. The food magazine was directly beneath the center key and 15 cm above the floor. A light inside the food magazine was illuminated whenever the feeder was operated. A small house-light was mounted on the top of the stimulus display panel and arranged so that it would not shine directly on the keys. White noise was provided via a speaker located in the lower left corner of the stimulus display panel. A fan on the opposite side of the chamber from the keys provided ventilation. Viewing of Ss was provided by a 16 x 18 cm
 they were habituated to the apparatus with the house-light and white noise on, bat neither the key lights nor the feeder were")
26 17 one-way window located on the door of the chamber. Programming was by relay control equipment located in an adjacent room. Procedure Preliminary training. While Ss were being accustomed to the food deprivation schedule (13 days) they were habituated to the apparatus with the house-light and white noise on, bat neither the key lights nor the feeder were operative. During magazine training the birds were given access to the grain hopper for 5 sec on a VI 1 min schedule. If after 5 days Ss had not begun to eat, they were given longer sessions and longer presentations of the grain hopper. Shaping consisted of training S^ to peck the lighted (white) key for reinforcement. If js did not immediately acquire the key peck response, it was established through successive approximation procedures. A specified sequence of response requirements (i.e., FRs ) were followed, in which Ss were first required to peck the center key once for reinforcement, then increasing to 8, 12, and lastly 16 responses in order to be reinforced. Next, S^ had to peck the illuminated side key upon which only one response was required for reinforcement throughout training. Forty reinforcements of 2 sec grain access were given on each key for two consecutive days. One of the 4 male and 3 of the 12 female Ss were randomly excluded from the oool of Ss. The remaining Ss were then randomly
 followed typical zero-delay MTS procedures, with the exception that,s was required to peck the center key the sample or standard stimulus (ST) 16 times for the")
27 10 assigned to one of three groups, with the stipulation that 1 male js would be in each group. Acquisition. The first group. Matching- to-sample-only (MTS-Only) followed typical zero-delay MTS procedures, with the exception that,s was required to peck the center key the sample or standard stimulus (ST) 16 times for the presentation of the side keys comparison stimuli (COs). Each trial began by the ST being illuminated with either red or green hues. Whenever S^ completed the 16 responses (FR 16) on the ST, it darkened and the COs were illuminated. One CO was green while the other was red. The animal then made its choice response to the CO. If correct, js was given 2 sec access to the grain magazine. If the incorrect CO was chosen the houselight and keys were turned off, leaving _S in the darkened cage for a 10 sec timeout (TO). A noncorrection procedure was used throughout and no intertrial interval employed. The hue of the ST was green on 50% of the trials and red on the other 50% with the order random throughout the session. The position of the hue as a CO was random, occurring an equal number ot times on each CO. Each daily session consisted of a tape with 120 trials. Five tapes with randomly determined trial sequences were used throughout the experiment. Ss were run under the acquisition procedure until a criterion of 35% correct matching for 5 consecutive days was reached, and then were switched to the delay procedure.

28 19 Latencies of the time to complete ST responding were recorded for the MTS-Only group. The FR 16 was divided into several segments: time to make the first response; time to complete the responses 2-6, 7-11, and The duration of each interval was recorded on each trial in tenths of a second. A second group, Differential Fixed-Ratio Matching-to- Sample ( DFR-MTS ), was exactly like the MTS-Only group except for the following requirements. An FR 27 was required on the ST when it was red in order to light the COs. When the ST was green an FR 5 was required. After reaching criterion, in this case 3 consecutive days of above 85% correct matching, Ss were given the probe procedure (see below). The third group, Differential Fixed-Ratio Only (DFR-Only) always had the center key illuminated white. Randomly programmed on this key were FR's 27 and 5. When the programmed FR was completed, the center key went out and the red and green COs were presented. When the FR had been 2 7, Ss were reinforced for choosing the red CO, and the green when the FR 5 had just been completed. Two of the Ss (# s s 1 and 5) were inadvertently run at only 3 days of 85% or above correct matching, while the other 2 Ss were run for the full five davs 4
.")
29 20 Probe Testing. Probe testing involved only the DFR-MTS group. Ss were given 12 probe trials in addition to the 120 regular trials in the same session for 2 consecutive days. The probe trials consisted of reversing the FR of the ST hue used in the regular trials (i.e., FR 27 on a green ST and FR 5 on a red ST). Randomly distributed throughout the session, 3 trials on each of the four possible stimulus patterns (RRG, GRR, RGG and GGR) were given each day. Two different tapes were used on the 2 testing days. On probe trials all choice responses were reinforced. The purpose of the probe trials was to test whether Ss were using the hue or information from differential responding in performing correct MTS behavior. Delay Testing. The day after Ss reached criterion (or finished probe testing in the case of the DFR-MTS group) they underwent delay testing. The delay interval was the time between the termination of the ST hue and presentation of CO hues. Ss were tested at.25,.50, 1, 2, 4, 8 and 16 sec delays, in that order. Each S^ received one session in which ail 120 trials were of the same delay value. If performance dropped below 85% correct matching for the session. S_ was returned to the zero-delay (acquisition) procedure unril it attained 85% accuracy again, then the next delay value was administered.
 and yellow (with FR 5) for the red and green hues on the ST under the acguisition procedure of the DFR-MTS group.")
30 21 Transfe r. After reinstatement of the baseline (i.e., 35% correct matching) for 1 session following delay testing, each S was tested for transfer. This procedure simply involved the substitution of blue (with FR 27) and yellow (with FR 5) for the red and green hues on the ST under the acguisition procedure of the DFR-MTS group. The MTS-Only group also had the hues changed, but maintained the FR 16 on the ST. The DFR-Only group retained the white center key. Of course, the COs in all groups were given the new hues. Ml J3s were run to a criterion of 85% correct matching for 3 consecutive days. FR 1 Testing. The MTS-Only and the DFR-MTS groups were given a single session in which only 1 response to the ST was required on the blue and yellow hues. Following this, the baseline was reestablished. Red-Green Retest. On the last day, all 3s were returned to a single session of matching under the original acquisition procedure (i.e., using red and green hues).


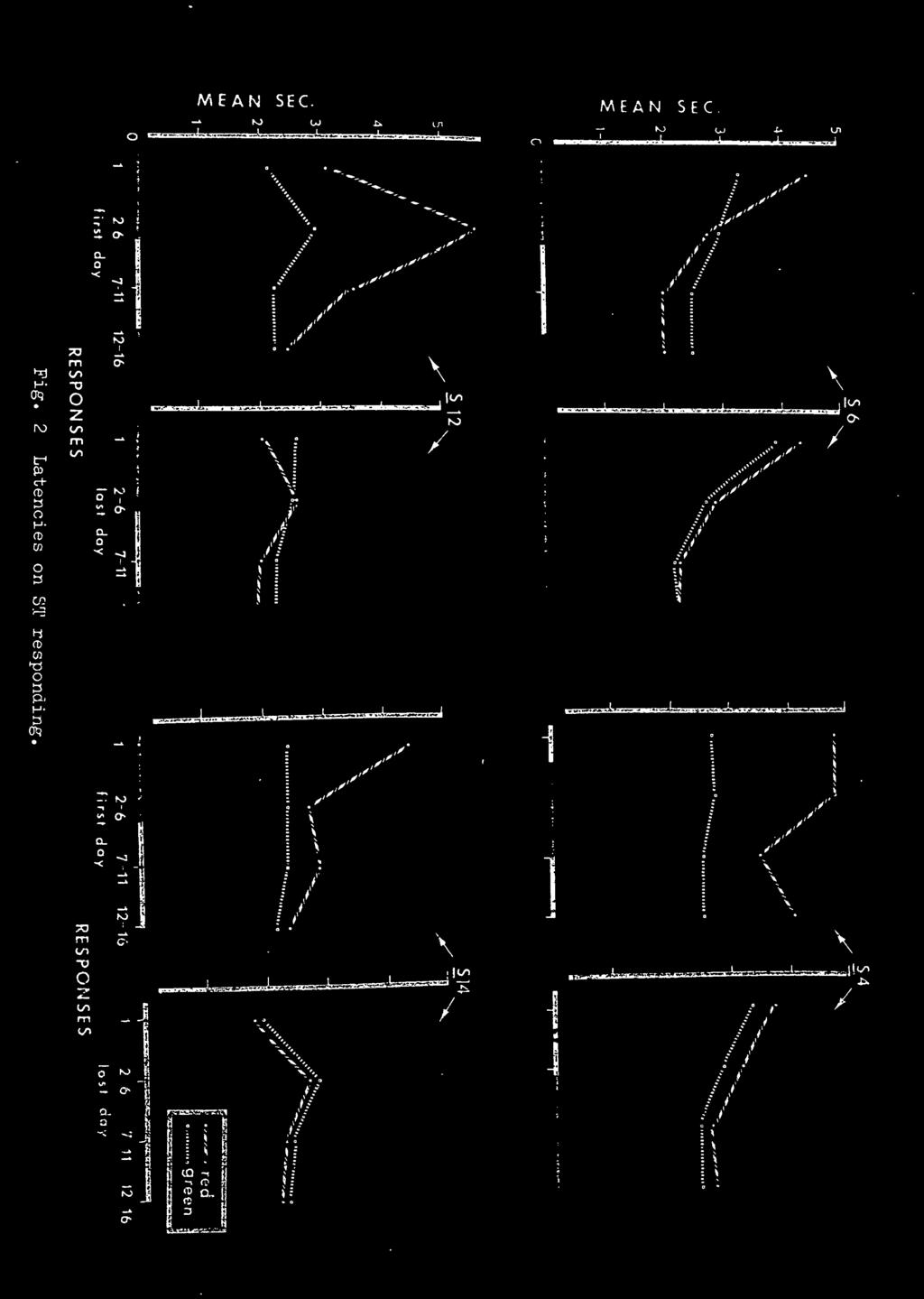
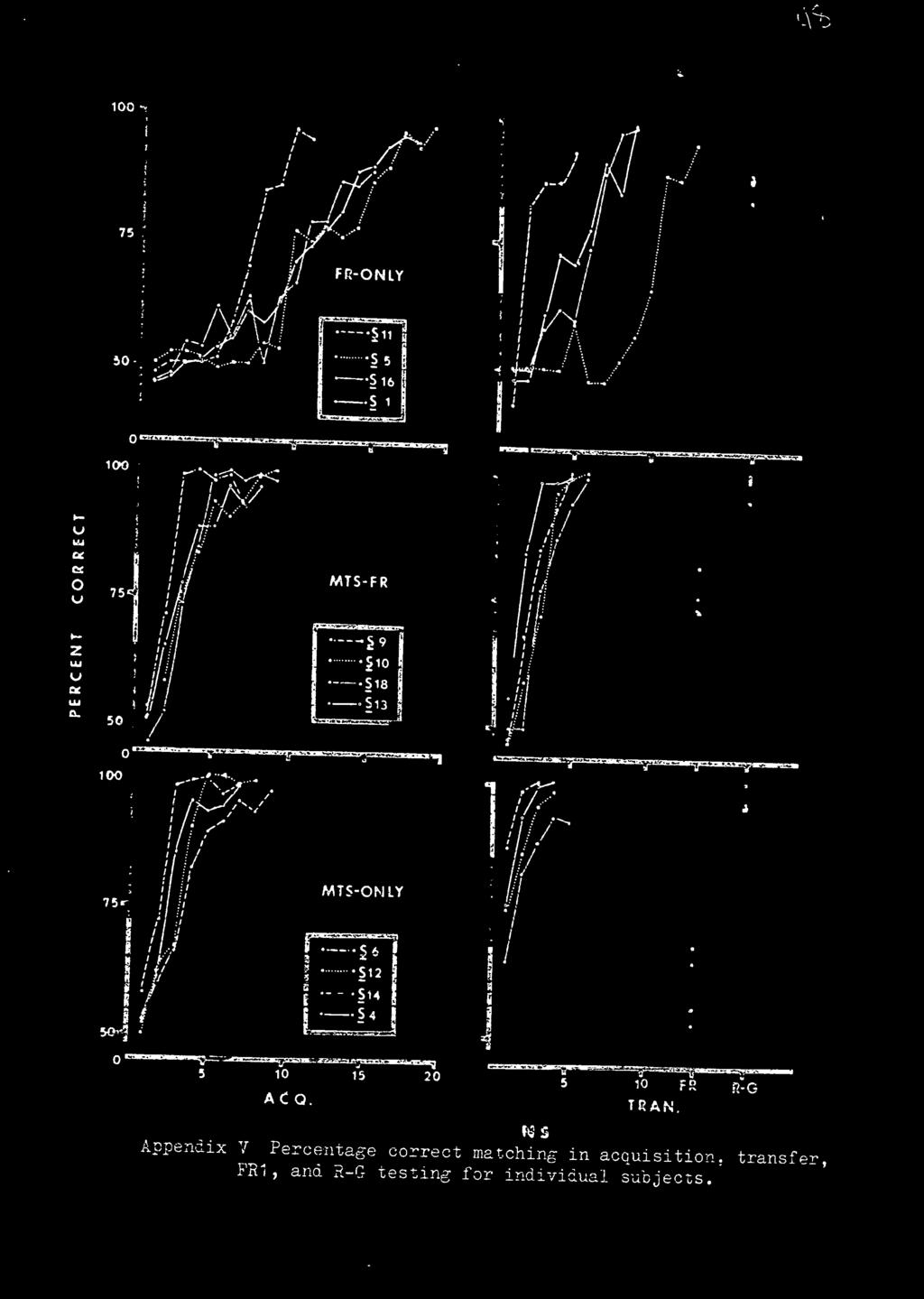
31 RESULTS The presentation and discussion of the results is based on group data in that they are representative of the individual subjects. Since Ss did not all reach criterion in the same number of sessions, the percentage correct of the last session was used in the computation of the completion of the group curves. Individual subject data are presented in the appendices. Acquisition. The acquisition curves are shown in Figure la. The MTS-Only and DFR-MTS groups are virtually identical, and differed by no more than 4 percentage points on any day. Both of these groups were above 85% correct matching by the 4th session and both groups reached criterion by an average of 7.75 sessions. All the Ss of these two groups reached criterion within two days of one another. The DFR-Only group also had greater day to day variability as shown by the peaks and troughs in the curve. All Ss in this group reached criterion within 6 days of one another. Insert Figure 1 about here In the MTS-Only group it was expected that if Ss were using response-produced cues as the source of stimulation to solve the task, they would develop distinctly different ways of responding to the ST hues. The results were the opposite

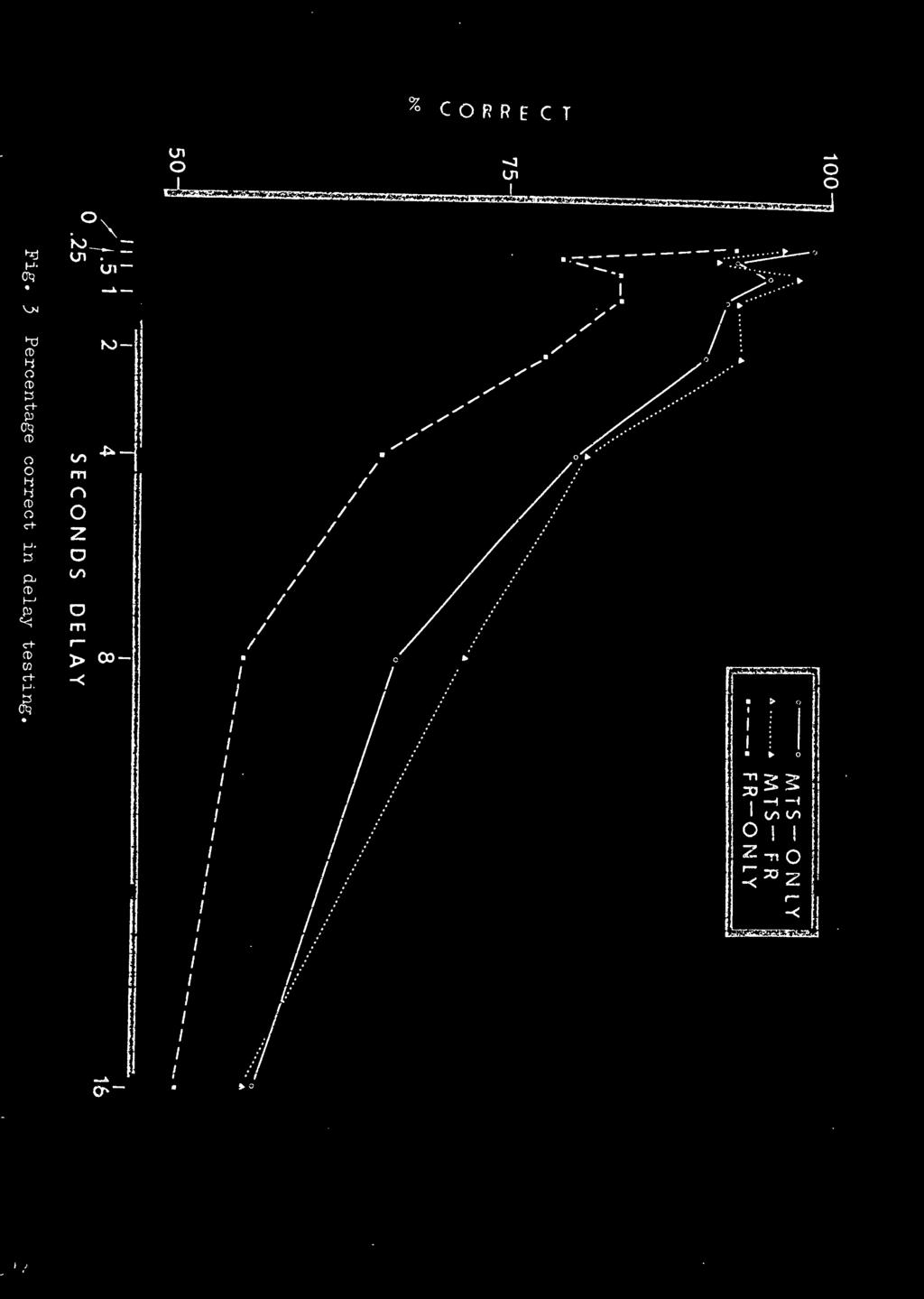
32 23 Insert Figure 2 about here of what was expected (see Figure 2). The differences in mean seconds to complete the response interval for the different hues became less rather than greater from the first day of acquisition to the last day. Probe Testing. The data for the probe and regular trials (on probe testing days) are given in Appendix I. Each cell of the matrices contains the percentage correct for each stimulus pattern and S^. The percentages on probe trials are based upon an N of 6 per cell, while regular trials are based upon an N of 60. As can be seen there is a small disruption of performance on the probe trials, but the percentage correct is still well above chance. A t-test of matched pairs resulted in a significant difference between probe and regular trials (t = 3.75, df = 15, p.001). The group data, also in Appendix I, show the probe trials to be consistently lower in percentage correct than the regular trials. Delay Testing. The curves showing the percentage correct over delay testing are presented in Figure 3. Three main Insert Figure 3 about here features of this data should be noted. First, all groups had a lower percentage correct matching at the.25 sec
 revealed a significant groups' main effect (F = 14.")
33 24 delay value than at the.50 sec delay value. Secondly, the DFR-Only group was consistently lower in correct matching than the other two groups. Last, the MTS-Only and the DFR-MTS groups differed by no more than 6 percentage points at any delay value. An analysis of variance (see Appendix II) revealed a significant groups' main effect (F = 14.76; 6,54 df) at the.001 level of significance. Scheffe's multiple comparison method (Myers, 1966; pp ) demonstrated that all comparisons with the DFR-Only group were significant at the.05 level except at the 1 sec and 16 sec delay values (see Appendix III). Transfer. Figure lb reveals that each group had a transfer function distinctly different from the other groups. The MTS-Only group had 76% correct matching on the first day with all subjects reaching criterion by the 5th day. The DFR-MTS group was performing at chance in the first transfer session, but improved rapidly. All Ss reached criterion by the 6th day. The DFR-Only group, again having the poorest performance, took over twice as many trials to reach criterion as the other two groups. The number of errors to 85% matching was computed for both acquisition and. transfer, and a savings score (i.e., difference between the number of errors in acquisition and transfer) was computed. An analysis of variance (see Table 1) yielded a significant difference (F = 19.2; 2,9 df) between groups at the.001 level.


34 25 Insert Table 1 about here PR 1 Testing. Only the MTS-Only and DFR-MTS groups were given FR 1 testing. Figure lb shows both groups performed well above chance, with the DFR-MTS group being superior. However, there was a noticeable decrement from asymtotic performance. Red-Green Retest. Again looking at Figure lb, all groups were above 85% correct. The MTS-Only and DFR-MTS groups were virtually identical and superior to the DFR-Only group.

 the high degree of similarity between these two groups in rate of acquisition; and 2) the decrease in the differences of the latencies")
35 DISCUSSION Original Hypotheses. it was hypothesized that Ss were solving the UTS task by response-pro* K*d cues arising from differential responding to different ST hues. Thus, the FR 5 and PR 2 7 requirements in the DFR-MTS group was expected to facilitate the differential responding resulting in faster acquisition. The data suggest, however, that the MTS-Only and DFR-MTS groups both used information provided by the ST hue rather than information from differential responding to the ST to solve the task. Two facts from the acquisition data lead to this conclusion: 1) the high degree of similarity between these two groups in rate of acquisition; and 2) the decrease in the differences of the latencies to complete the FR on the ST in the MTS-Only group. The high degree of similarity between the MTS-Only and DFR-MTS groups suggests that although Ss in the DFR-MTS group were provided a response dimension with which they could solve the task, they apparently chose the hue dimension. As demonstrated by the DFR-Only group, the response dimension provided by the different FRs is sufficient to solve the task; however, it appears more difficult to use. Furthermore, there does not appear to be any additivity between the response and hue dimensions for the DFR-MTS group. That is, the addition of an FR 2 7 requirement on the red ST and an FR 5 requirement on a green ST failed to make the discriminative stimuli more distinct from each other, which would result in faster

36 27 acquisition. The decrease in differences in latencies to the STs from the first to last days of acquisition argues directly against the development of differential responding (hence differential response produced cues) to the ST. In fact, on the first day of acquisition the latencies to the red and green STs were distinctly different, but by the last day of acquisition they were nearly identical. The latency results are the opposite of what one would expect if they were entertaining the notion that differential response-produced cues were the basis for solution of the task. However, the possibility still exists that a response dimension other than the latency on the ST may be used. Nevertheless, the high degree of similarity between the MTS-Only and DFR-MTS groups strongly suggests they are using the same dimension in solving the task; it is presently believed to be the hue dimension. Although the addition of differential fixed-ratios to the hue of the DFR-MTS group did not affect the acquisition rate, the results of probe trials suggest that induced differential responding was not completely ignored. A significant disruption of matching occurred on these trials, although performance remained well above chance. These results clearly demonstrate that choice responses were not based solely upon the DFR cues; otherwise the probe trials would have had zero percent, or nearly so, correct matching. Perhaps the disruption

37 20 that did occur was due to generalization decrement from regular trials to probe trials. The virtually identical delay functions of the MTS-Only and DPR-MTS groups support the contention that these groups are using the same source of information in solving the task, and that this information is probably the ST hue. If the DFR-MTS group were using information from the differential FKs we would expect its performance to be comparable to the DFR-Only group. Neither of these appear to be the case, in that the DFR-Only delay function was consistently lower than the other two, and the DFR-MTS and MTS-Only groups had quite similar delay functions. The differences of the delay functions between the groups that had a hue on the ST and the DFR-Only group tells something about the information used by these groups in solving the task. First, the information (response-produced cues) used by the DFR-Only group is presumably of shorter duration, or at least more difficult to use during delay testing than the information provided by the hue on the ST. Secondly, this loss, or difficulty of use, of information in the DFR-Only group is a relatively constant amount over all delay values. Furthermore, the percentage correct matching in delay of the DFR-Only group is more similar to Rilling'S (1968) results which used FRs as discriminative stimuli, than the MTS-Only and DFR-MTS groups which apparently used the hues.

38 29 The increase in matching accuracy from.25 to.50 sec delay values was initially puzzling. However, informal observations of Ss while making their choice responses during zero-delay procedure revealed that Ss looked at one of the COs first. If it was the correct one, the bird pecked it, while if it were incorrect, the other CO would be immediately and presumably indiscriminately pecked. It is possible that with the.25 sec delay the would look at one CO during the delay, but since it did not "match" the sample, since it was dark, they would immediately respond to the other key which was lit by the time 5^ get there. With longer delays, however, would be exposed to both COs as dark. Presumably S_ would then wait until both COs were illuminated before making their comparisons and choice responses An initial comparison of the MTS-Only and DFR-MTS transfer functions seem to show a break in the consistency of similar results between these two groups. However, the chance performance of the DFR-MTS group on the first day may have been due to a color preference factor. All S_s of the DFR-MTS group formed a yellow color pre;.eieike, *niie u^nc tne Only Sis formed any kind of preference The yellow preference may be due to the pairing of the yellow with the shorter PR, making it preferable over the longer PR on the blue ST. In fact, 3 of the 4 Ss in the DFR-MTS group during initial ac-

39 30 quisition had preferences for the green stimulus which was paired with the FR 5. It is believed that the reason for the different transfer functions between MTS-Only and the DFR-MTS groups is due to the depression of correct matching on the first day caused by color preferences and not differences in the learning processes In order to see if the high initial performance (i.e., positive transfer) of the MTS-Only group was immediate or due to rapid learning within the session, the distribution of errors throughout the session were examined (see Appendix IV)* The errors were distributed relatively evenly throughout the session with only a small decrease in the number of errors during the second half of the session. At present it is unclear whether this positive transfer is specific, such as in a "conceptual scheme," as suggested by the "abstract generalization mechanism" (Nissen, Blum and Blum, 1948) and the "symbolic process" interpretation (Yerkes and Nissen, 1938), or due to a general practice effect. Comparing the DFR-Only transfer function with the other 2 groups, poorer performance is observed. Again, as in the original acquisition and delay data, ue find that response produced cue information is more difficult to utilize than information from the hue on an ST, as shown by the slower rate of acquisition.

40 31 As expected, the MTS-Only and DFR-MTS groups both showed a decrement in performance during FR 1 testing; however, the MTS-Only group had the largest disruption. A possible reason for the difference between these groups is the difference in the amount of generalization decrement. As mentioned earlier, although these grcups solved the task by using hues, the response dimension was not completely ignored. Possibly the MTS- Only group experienced a greater generalization decrement when given FR 1 testing because it was accustomed to consistently responding 16 times to the ST, while the DFR-MTS group was used to responding a variable number of times (i.e., 5 or 27). Furthermore, the FR 5 is relatively close to the FR 1, making this new situation seem less novel for the DFR-MTS group. It would also be expected that Ss in the DFR-MTS group would perform better when the correct CO was a green hue (i.e., FR 5 and FR 1 are relatively similar). The results showed that the DFR-MTS group had 209 correct responses to the green CO and only 165 correct responses to the red, while the MTS-Only group had 148 and 142 correct response respectively. In addition to the differences found between groups in the present study, there are also large differences in the results between present and previous research using the MTS paradigm. The differences have suggested a model which will be presented in the following section.
 obtained; but comparable to previous simultaneous matching results (e.g., Gumming and Berryman, 1961).")
41 32 A Modified Coding Model. The results of the MTS-Only and DFR-MTS groups of the present study differ from previous research results in 3 main ways. First, there was faster acquisition with zero-delay procedures than, for example, Cumming _et aj^. (1965) obtained; but comparable to previous simultaneous matching results (e.g., Gumming and Berryman, 1961). Secondly, delay matching performance was better than either Berryman et al. 's (1963) or Sack's (1971) results. Last, the present study demonstrated superior positive transfer to the matching of new hues than previously reported (e.g.,, Sacks, 1971). The most evident orocedural difference between the Drevious and present research is an PR requirement greater than 1 on the ST. Only two previous studies have employed FRs on the ST. Berryman et aj_. (1963) had an FR 5 required on the ST in a simultaneous procedure during acquisition, and then underwent delay testing at 0, 1, 2, 4, 10 and 24 sec delay values with the FR requirement still present. Comparing their results with an earlier simultaneous matching study, Cumming and Berryman { ; concluded that the dddi Lional exposure Lo the ST induce^ by the FR requirement ^id not facilitate acquisition of simultaneous matching, However, comparing Sack's (1971) delay data, which was at chance performance at 1 sec delay, with Cumming and Berryman 1 s data, it appears

42 33 that FR did have an effect upon delay matching accuracy in that chance performance' was not reached until the 4 sec delay value. Two explanations are possible. First, perhaps the simultaneous matching task is "easy" enough for the Ss that the FR has no effect. More specifically, the information provided by the ST is present during the choice response, thus the FR adds nothing to the information provided by the ST. Secondly, there may be a "ceiling effect" in which acquisition occurs so rapidly in the simultaneous task that the FR-effect fails to show up in the data. Thus, assuming delay matching to be more difficult, due to the absence of the ST during the choice response, than simultaneous matching, and comparing the present and Cumming and Berryman's data with Sacks', it appears that increased responding to the ST yields superior performance with the more difficult delayed matching task. This is further supported by acquisition data. The MTS-Only and DFR-MTS Ss of the present study had much steeper acquisition runctions than those of Sacks or Gumming _et al. (1965), although all studies employed zero-delay procedures. The second study using an FR on the ST i Gumming et al., 1965) also failed to find any facilitation due to instituting the FR requirement. Although this was a zero-delay task (hence FR-effect should presumably have occurred) the FR was not instituted until the 13th day of transfer training.

43 34 As pointed out by the authors, this previous training with FR 1 could have disrupted the chain of "peck center once, then peck side. The question arises as to how this FR-effect occurs; that is, how does an FR requirement on the ST facilitate acquisition, improve delayed matching performance, and increase the amount of positive transfer. It is proposed that the FR requirement induces _S to attend to the ST and its stimulus properties more than a single response does. Hence, S_ is exposed to it longer making it a more "significant" or salient part of its environment. This increased exposure presumably has two main results: 1) a faster "attentional learning" in which _S learns to attend to the relevant dimension (i.e., hue) faster than if only an FR 1 were required, and 2) a strengthening of the mediational, or coding response beyond that ordinarily occurring after a single response to the ST. It should be mentioned that the original notion of a laarned mediator (a development of differential responding leading to differential response-produced cues) has been abandoned. It is now assumed the mediator is a naturally elicited response (with stimulus properties) of the onanism to the relevant cue dimension that j3 uses to solve the task. More specifically, the mediator is an unconditioned response to the hue of the ST, and is presumably central and neural in nature. This

44 35 neural response, with its stimulus properties, is the functional cue used as the discriminative stimulus in making the choice response. Thus, in the present model; _S learns to attend to the relevant dimension, and to make a certain response in the presence of a particular mediating cue. The modified coding aspect of the model will now be discussed. It is considered "modified" in that it differs from the forms presented by Lawrence (1963) and Cumming et al. (1965). The most significant change, as discussed earlier, is that the coding response (r ) is not learned, but an unconditioned response to the stimulus properties of the ST. Additional modifications should become evident in the following example. CO > r /R rf green green green ST red ' L red - CO \ * r,/r, rf red red red Time As shown above, a red sample stimulus (ST ) -A- ^w- \ is presented at the beginning of the trial. This red ST elicits an unconditioned mediating neural response (r red ). Subsequently, the COs are presented which also elicit unconditioned responses respective to their color (i.e., CO g r "^^ > green
 in that it elicits the r which is the red same mediator that the red ST elicits.")
45 36 and CO r --^ >r re(j ). ;S then acquires, through the reinforcement contingencies, the choice response to the red CO (in this example) in that it elicits the r which is the red same mediator that the red ST elicits. Thus, the mediating responses which serve as the discriminative cues inform the j3 of the correct response. An attempt will now be made to show how the FR-effect interacts with the modified coding hypothesis, and how it apparently accounts for much of the present data. There are two ways of looking at the acquisition functions: how soon, in terms of sessions, performance begins to rise above chance; and the slope of these functions. It is assumed the first tells us about how soon S_ attends to the relevant dimension and the latter about the learning of the correct response. The acquisition function of the MTS-Only group suggests that the hue dimension was attended to almost immediately. Although the DFR-MTS group could have solved the task on either the hue or the response dimension, it appears the hue dimension was used by the Ss. Two facts lead to this conclusion; 1) as already mentioned, the high similarity of these two g^cups in acquisition, delay, and transfer testing, and 2) the saliency of color and visual cues for pigeons (Reynolds, 1961). This latter fact, along with the possibility that DFR-Only Ss may have to learn to attend to two dimensions

46 37 may account for their slower rise from chance performance. Perhaps this requirement of having to attend to two dimensions can be seen more clearly in the following schema. C green ^ - n ^ ^reen^green ~* rf ST 2T > r 2?r ' ' C0_~ ± ' red > r_. "red' J /R... ix red J rf That is, Si must learn to attend to the FR mediating dimension as well as the color mediating dimension. Next looking at the groups with the hue on the ST, we find very steep acquisition functions. As mentioned earlier these functions are comparable to simultaneous acquisition functions with FR 1 on the center key. It is being proposed that the larger FR requirement on the vst, through increased exposure to the hue dimension, increases the strength of the unconditioned mediating response. This increased strength allows the information provided by the ST to persist until the CDs are present; thus making it more similar to the simultaneous task in which the ST information is always present when the CO choice is made. In the case of the DFR-Only group, two possible reasons will be suggested for its less steep acquisition function. The first is a function of the animal itself. As stated earlier, color and visual stimuli are salient cues for pigeons. Thus, the sensory and central


47 38 nervous apparatus of the is more developed and efficient with the use of visual stimuli than stimuli produced by differential responding. This less developed apparatus of response-produced cues makes the handling of information more difficult, resulting in slower acquisition. Another reason may be that it is more difficult to match a mediating response from a response-produced cue dimension with a mediator from a hue dimension than if both were from the same dimension. The argument for the FR-effect during delay testing on the groups with the hue on the ST follows directly from the argument given for the acquisition results. That is, the increased responding on the ST increases the s trength of the unconditioned mediation response. This allows the information to persist longer into the delay interval. Thus, there is a higher probability of having a correct choice response because the information is present when the COs are presented. The delay data presented in Figure 2 show the FR-effect was operating; chat is, a higher percentage correct matching over longer o^lay intervals. Furthermore, the notion that pigeons are better at handling visual information than that orovided bv resoonse-oroduced curs is evidenced by the fact that the DFR-Only group has consistently lower performance.
.")

48 39 However, due to the procedure employed the possibility exists that the performance at the higher delay values may be confound with experience at earlier values. Figure lb shows the transfer functions (the reasons for the differences between these groups has already been discussed). Due to the high initial performance of the MTS-Only group and the rapid rise from chance performance of the other two groups, we find strong evidence that Ss are already attending to the relevant dimensions. In addition, the DFR-MTS group which has a choice of two dimensions, apparently employs the one previously used. Thus, the attention stage is practically eliminated. The rapid improvement thereafter is apparently due to the s trong mediators provided through the FR on the ST. Again, the DFR-Only group provides evidence that response-produced cue information is more difficult to utilize than that provided by hues. In conclusion, it appears that Ss use the hue dimension when given a choice between response and hue dimensions. In addition, the hue dimension appears easier to utilize, and that is believed to be a function of the animal itself. A model has been proposed in which the mediator is assumed to be an unconditioned response to the stimuli employed, rather than a learned response. Perhaps the most significant aspect of the present research was the uncovering of the FR-effect, in which performance in the MTS task was greatly improved.

49 40 TABLE 1 ANALYSIS OF VARIANCE OF SAVINGS SCORE FOR TRANSFER Source of Degrees of Sum of Mean F-Ratio Variance Freedom Squares Squares Total ,802 Groups 2 121, , Error 9 28,489 3,


50 U1 O %CO R RE C T O O p. CD o CD P IU V J o 4 ^ o > O M <> V\ o 0) o 1 c+ CD CO O & c+ a in m o P3 O o H* CO H- c+ H- O -» c+ ^r^ui CO Hj (D 4 > z ^3 o

51 MEAN SEC MEAN SEC

52 % CORRECT


53 APPENDIX I PERCENTAGE "CORRECT" ON PROBE AND REGULAR TRIALS Stimulus Pattern Subject Number Group RRG FR probe RRG 71 regular RGG FR probe RGG regular GRR FR5 probe GRR ' regular GRR FR probe GRR regular i

54 45 APPENDIX II ANALYSIS OF VARIANCE OF DELAY TESTING Source of Degrees of Freedom Sum of k ' v_i U X- *w Mean S rru srpq F-Ratio Total Between Subj ects 11 3, 89 7 Group 2 2,986 1, * Error Within Subj ects 12 16, 234 Delay Value 6 14,626 2, ** Interaction of Group and Delay Error 54 1, * p.005 ** * o

55 46 APPENDIX III F-VALUES OF CONTRASTS FOR BELAY TESTING Contrast Delay Value 12 in Seconds MTS MTS-FR MTS-FR FR MTS FR


56 47 APPENDIX IV DISTRIBUTION OF ERRORS ON FIRST SESSION OF TRANSFER Subj ect Number Fifteen Trial Segments 1st 2nd 3rd 4th Second Half

57 100



58 FR-ONLY \ MTS-ONLY.25^ 8 <= r DELAY Appendix VI Percentage correct matching in for individual subjects.

59 50 REFERENCES Berryman, R., Cumming, W.W., and Nevin, J. A. Acquisition of delayed matching in the pigeon. Journal of the Experimental Analysis of Behavior. 1963, 6_, Berryman, R., Cumming, W.W., Cohen, L.R., and Johnson, D.F. Acquisition and transfer of simultaneous oddity. Psycho - logical Reports, 1965, Blough, D.S. Delayed matching in the pigeon. J ournal of t he Experimental Analysis of Behavior. 1959, 2,, Cumming, W.W. and Berryman, R. Some data on matching behavior in the pigeon. Journal of the Experimental Analysis of Behavior, 4, , Cumming, W.W., Berryman, R. and Cohen, L.R. Acquisition and transfer of zero-delay matching. Psychological Reports, 1965, 12, Eckerman, E.A., Lanson, R.N. and Cumming, W.W. Acquisition and maintenance of matching without a required observing response. Journal of the Experimental Analysis of Behavio r, 1968, 11., Ginsberg, N. Matching in pigeons. Journal of Comparative and Physiological Psychology, 1943, 3, Lawrence, D.H. The nature of a stimulus-: some relations between learning and perception. In S, Koch (Ed.), Psychology: A Study of a Sc ience. Vol. 5. New York: McGraw-Hill, 1963,

60 / 51 Nissen, H.W., Blu, J.S., and Blum, R.A. Analysis of Matching behavior in chimpanzees. Journal of Comparative and Physiological Psychology. 1948, 4_1, Piiskoff, S.S. and Goldiamond, I. Some discriminative properties of fixed-ratio performance in the pigeon. Journal of the Experimental Analysis of 3ehavior. 1966, 9_, 1-9. Reynolds, G.S. Attention in the pigeon. Journal of the Experimental Analysis of Behavior. 1961, 4_, Rilling, M.E. Number of responses as a stimulus in fixedinterval and fixed-ratio schedules. Journal of Comparative and Physiological Psychology, 196 7, 6_3, Rilling, M.E. Effects of timeout on a discrimination between fixed-ratio schedules. Journal of the Experimental Analysis of Behavior, 1968, JL1, Sacks, R. The coding hypothesis and zero-delay matching-tosarnple in the pigeon. Unpublished master's thesis, University of Massachusetts, Schulte, V.G. The discrimination of the stimulus consequences of responses differing in work requirement. Unpublished Doctoral Dissertation. University of Kentucky Weinstein, 3. Matching- to-sample by rhesus monkeys and by children. Journal of Comparative Psychology, 1941, 31.,
61 52 Yerkes, R.M. and Nissen, H.W. Prelinguis tice sign behavior in chimpanzee. Science, 1939, 89,
62



63
Within-event learning contributes to value transfer in simultaneous instrumental discriminations by pigeons
 Animal Learning & Behavior 1999, 27 (2), 206-210 Within-event learning contributes to value transfer in simultaneous instrumental discriminations by pigeons BRIGETTE R. DORRANCE and THOMAS R. ZENTALL University
Animal Learning & Behavior 1999, 27 (2), 206-210 Within-event learning contributes to value transfer in simultaneous instrumental discriminations by pigeons BRIGETTE R. DORRANCE and THOMAS R. ZENTALL University
ON THE EFFECTS OF EXTENDED SAMPLE-OBSERVING RESPONSE REQUIREMENTS ON ADJUSTED DELAY IN A TITRATING DELAY MATCHING-TO-SAMPLE PROCEDURE WITH PIGEONS
 ON THE EFFECTS OF EXTENDED SAMPLE-OBSERVING RESPONSE REQUIREMENTS ON ADJUSTED DELAY IN A TITRATING DELAY MATCHING-TO-SAMPLE PROCEDURE WITH PIGEONS Brian D. Kangas, B.A. Thesis Prepared for the Degree of
ON THE EFFECTS OF EXTENDED SAMPLE-OBSERVING RESPONSE REQUIREMENTS ON ADJUSTED DELAY IN A TITRATING DELAY MATCHING-TO-SAMPLE PROCEDURE WITH PIGEONS Brian D. Kangas, B.A. Thesis Prepared for the Degree of
Value transfer in a simultaneous discrimination by pigeons: The value of the S + is not specific to the simultaneous discrimination context
 Animal Learning & Behavior 1998, 26 (3), 257 263 Value transfer in a simultaneous discrimination by pigeons: The value of the S + is not specific to the simultaneous discrimination context BRIGETTE R.
Animal Learning & Behavior 1998, 26 (3), 257 263 Value transfer in a simultaneous discrimination by pigeons: The value of the S + is not specific to the simultaneous discrimination context BRIGETTE R.
CAROL 0. ECKERMAN UNIVERSITY OF NORTH CAROLINA. in which stimulus control developed was studied; of subjects differing in the probability value
 JOURNAL OF THE EXPERIMENTAL ANALYSIS OF BEHAVIOR 1969, 12, 551-559 NUMBER 4 (JULY) PROBABILITY OF REINFORCEMENT AND THE DEVELOPMENT OF STIMULUS CONTROL' CAROL 0. ECKERMAN UNIVERSITY OF NORTH CAROLINA Pigeons
JOURNAL OF THE EXPERIMENTAL ANALYSIS OF BEHAVIOR 1969, 12, 551-559 NUMBER 4 (JULY) PROBABILITY OF REINFORCEMENT AND THE DEVELOPMENT OF STIMULUS CONTROL' CAROL 0. ECKERMAN UNIVERSITY OF NORTH CAROLINA Pigeons
Attention shifts during matching-to-sample performance in pigeons
 Animal Learning & Behavior 1975, Vol. 3 (2), 85-89 Attention shifts during matching-to-sample performance in pigeons CHARLES R. LEITH and WILLIAM S. MAKI, JR. University ofcalifornia, Berkeley, California
Animal Learning & Behavior 1975, Vol. 3 (2), 85-89 Attention shifts during matching-to-sample performance in pigeons CHARLES R. LEITH and WILLIAM S. MAKI, JR. University ofcalifornia, Berkeley, California
Pigeons transfer between conditional discriminations with differential outcomes in the absence of differential-sample-responding cues
 Animal Learning & Behavior 1995, 23 (3), 273-279 Pigeons transfer between conditional discriminations with differential outcomes in the absence of differential-sample-responding cues LOU M. SHERBURNE and
Animal Learning & Behavior 1995, 23 (3), 273-279 Pigeons transfer between conditional discriminations with differential outcomes in the absence of differential-sample-responding cues LOU M. SHERBURNE and
KEY PECKING IN PIGEONS PRODUCED BY PAIRING KEYLIGHT WITH INACCESSIBLE GRAIN'
 JOURNAL OF THE EXPERIMENTAL ANALYSIS OF BEHAVIOR 1975, 23, 199-206 NUMBER 2 (march) KEY PECKING IN PIGEONS PRODUCED BY PAIRING KEYLIGHT WITH INACCESSIBLE GRAIN' THOMAS R. ZENTALL AND DAVID E. HOGAN UNIVERSITY
JOURNAL OF THE EXPERIMENTAL ANALYSIS OF BEHAVIOR 1975, 23, 199-206 NUMBER 2 (march) KEY PECKING IN PIGEONS PRODUCED BY PAIRING KEYLIGHT WITH INACCESSIBLE GRAIN' THOMAS R. ZENTALL AND DAVID E. HOGAN UNIVERSITY
Value Transfer in a Simultaneous Discrimination Appears to Result From Within-Event Pavlovian Conditioning
 Journal of Experimental Psychology: Animal Behavior Processes 1996, Vol. 22. No. 1, 68-75 Copyright 1996 by the American Psychological Association. Inc. 0097-7403/96/53.00 Value Transfer in a Simultaneous
Journal of Experimental Psychology: Animal Behavior Processes 1996, Vol. 22. No. 1, 68-75 Copyright 1996 by the American Psychological Association. Inc. 0097-7403/96/53.00 Value Transfer in a Simultaneous
Timing in pigeons: The choose-short effect may result from pigeons confusion between delay and intertrial intervals
 Psychonomic Bulletin & Review 1998, 5 (3), 516-522 Timing in pigeons: The choose-short effect may result from pigeons confusion between delay and intertrial intervals LOU M. SHERBURNE Wabash College, Crawfordsville,
Psychonomic Bulletin & Review 1998, 5 (3), 516-522 Timing in pigeons: The choose-short effect may result from pigeons confusion between delay and intertrial intervals LOU M. SHERBURNE Wabash College, Crawfordsville,
Animal memory: The contribution of generalization decrement to delayed conditional discrimination retention functions
 Learning & Behavior 2009, 37 (4), 299-304 doi:10.3758/lb.37.4.299 Animal memory: The contribution of generalization decrement to delayed conditional discrimination retention functions REBECCA RAYBURN-REEVES
Learning & Behavior 2009, 37 (4), 299-304 doi:10.3758/lb.37.4.299 Animal memory: The contribution of generalization decrement to delayed conditional discrimination retention functions REBECCA RAYBURN-REEVES
Transitive inference in pigeons: Control for differential value transfer
 Psychonomic Bulletin & Review 1997, 4 (1), 113-117 Transitive inference in pigeons: Control for differential value transfer JANICE E. WEAVER University of Kentucky, Lexington, Kentucky JANICE N. STEIRN
Psychonomic Bulletin & Review 1997, 4 (1), 113-117 Transitive inference in pigeons: Control for differential value transfer JANICE E. WEAVER University of Kentucky, Lexington, Kentucky JANICE N. STEIRN
Stimulus control of foodcup approach following fixed ratio reinforcement*
 Animal Learning & Behavior 1974, Vol. 2,No. 2, 148-152 Stimulus control of foodcup approach following fixed ratio reinforcement* RICHARD B. DAY and JOHN R. PLATT McMaster University, Hamilton, Ontario,
Animal Learning & Behavior 1974, Vol. 2,No. 2, 148-152 Stimulus control of foodcup approach following fixed ratio reinforcement* RICHARD B. DAY and JOHN R. PLATT McMaster University, Hamilton, Ontario,
The coding hypothesis and zero-delay matching-tosample
 University of Massachusetts Amherst ScholarWorks@UMass Amherst Masters Theses 1911 - February 2014 1971 The coding hypothesis and zero-delay matching-tosample in the pigeon. Robert A. Sacks University
University of Massachusetts Amherst ScholarWorks@UMass Amherst Masters Theses 1911 - February 2014 1971 The coding hypothesis and zero-delay matching-tosample in the pigeon. Robert A. Sacks University
Coding of hedonic and nonhedonic samples by pigeons in many-to-one delayed matching
 Animal Learning & Behavior 1995, 23 (2), 189 196 Coding of hedonic and nonhedonic samples by pigeons in many-to-one delayed matching THOMAS R. ZENTALL and LOU M. SHERBURNE University of Kentucky, Lexington,
Animal Learning & Behavior 1995, 23 (2), 189 196 Coding of hedonic and nonhedonic samples by pigeons in many-to-one delayed matching THOMAS R. ZENTALL and LOU M. SHERBURNE University of Kentucky, Lexington,
Some Parameters of the Second-Order Conditioning of Fear in Rats
 University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Papers in Behavior and Biological Sciences Papers in the Biological Sciences 1969 Some Parameters of the Second-Order Conditioning
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Papers in Behavior and Biological Sciences Papers in the Biological Sciences 1969 Some Parameters of the Second-Order Conditioning
PROBABILITY OF SHOCK IN THE PRESENCE AND ABSENCE OF CS IN FEAR CONDITIONING 1
 Journal of Comparative and Physiological Psychology 1968, Vol. 66, No. I, 1-5 PROBABILITY OF SHOCK IN THE PRESENCE AND ABSENCE OF CS IN FEAR CONDITIONING 1 ROBERT A. RESCORLA Yale University 2 experiments
Journal of Comparative and Physiological Psychology 1968, Vol. 66, No. I, 1-5 PROBABILITY OF SHOCK IN THE PRESENCE AND ABSENCE OF CS IN FEAR CONDITIONING 1 ROBERT A. RESCORLA Yale University 2 experiments
OBSERVING AND ATTENDING IN A DELAYED MATCHING-TO-SAMPLE PREPARATION IN PIGEONS. Bryan S. Lovelace, B.S. Thesis Prepared for the Degree of
 OBSERVING AND ATTENDING IN A DELAYED MATCHING-TO-SAMPLE PREPARATION IN PIGEONS Bryan S. Lovelace, B.S. Thesis Prepared for the Degree of MASTER OF SCIENCE UNIVERSITY OF NORTH TEXAS December 2008 APPROVED:
OBSERVING AND ATTENDING IN A DELAYED MATCHING-TO-SAMPLE PREPARATION IN PIGEONS Bryan S. Lovelace, B.S. Thesis Prepared for the Degree of MASTER OF SCIENCE UNIVERSITY OF NORTH TEXAS December 2008 APPROVED:
Common Coding in Pigeons Assessed Through Partial Versus Total Reversals of Many-to-One Conditional and Simple Discriminations
 Journal of Experimental Psycholo Animal Behavior Processes 1991, Vol. 17, No. 2, 194-201 Copyright 1991 by the American Psychological Association, Inc. 0097-7403/91/43.00 Common Coding in Pigeons Assessed
Journal of Experimental Psycholo Animal Behavior Processes 1991, Vol. 17, No. 2, 194-201 Copyright 1991 by the American Psychological Association, Inc. 0097-7403/91/43.00 Common Coding in Pigeons Assessed
EFFECTS OF PRESENTATION METHOD ON ACQUISITION SPEED OF AN IDENTITY MATCHING-TO-SAMPLE TASK
 EFFECTS OF PRESENTATION METHOD ON ACQUISITION SPEED OF AN IDENTITY MATCHING-TO-SAMPLE TASK A Thesis Presented to the Faculty of California State University, Stanislaus In Partial Fulfillment of the Requirements
EFFECTS OF PRESENTATION METHOD ON ACQUISITION SPEED OF AN IDENTITY MATCHING-TO-SAMPLE TASK A Thesis Presented to the Faculty of California State University, Stanislaus In Partial Fulfillment of the Requirements
Transitive Inference and Commonly Coded Stimuli
 Georgia Southern University Digital Commons@Georgia Southern Electronic Theses & Dissertations Graduate Studies, Jack N. Averitt College of Summer 2005 Transitive Inference and Commonly Coded Stimuli William
Georgia Southern University Digital Commons@Georgia Southern Electronic Theses & Dissertations Graduate Studies, Jack N. Averitt College of Summer 2005 Transitive Inference and Commonly Coded Stimuli William
Stimulus Control Exerted by Remember Cues Over Processing in Pigeons Short-Term Memory
 Wilfrid Laurier University Scholars Commons @ Laurier Theses and Dissertations (Comprehensive) 1982 Stimulus Control Exerted by Remember Cues Over Processing in Pigeons Short-Term Memory Patricia E. Hernandez
Wilfrid Laurier University Scholars Commons @ Laurier Theses and Dissertations (Comprehensive) 1982 Stimulus Control Exerted by Remember Cues Over Processing in Pigeons Short-Term Memory Patricia E. Hernandez
Processing of empty and filled time intervals in pigeons
 Learning & Behavior 2004, 32 (4), 477-490 Processing of empty and filled time intervals in pigeons DOUGLAS S. GRANT and DIANE C. TALARICO University of Alberta, Edmonton, Alberta, Canada Pigeons were trained
Learning & Behavior 2004, 32 (4), 477-490 Processing of empty and filled time intervals in pigeons DOUGLAS S. GRANT and DIANE C. TALARICO University of Alberta, Edmonton, Alberta, Canada Pigeons were trained
Oddity learning in the pigeon: Effect of negative instances, correction, and number of incorrect alternatives
 Animal Learning & Behavior 1980,8(4),621-629 Oddity learning in the pigeon: Effect of negative instances, correction, and number of incorrect alternatives THOMAS R. ZENTALL University ofkentucky, Lexington,
Animal Learning & Behavior 1980,8(4),621-629 Oddity learning in the pigeon: Effect of negative instances, correction, and number of incorrect alternatives THOMAS R. ZENTALL University ofkentucky, Lexington,
between successive DMTS choice phases.
 JOURNAL OF THE EXPERIMENTAL ANALYSIS OF BEHAVIOR 1996, 66, 231 242 NUMBER 2(SEPTEMBER) SEPARATING THE EFFECTS OF TRIAL-SPECIFIC AND AVERAGE SAMPLE-STIMULUS DURATION IN DELAYED MATCHING TO SAMPLE IN PIGEONS
JOURNAL OF THE EXPERIMENTAL ANALYSIS OF BEHAVIOR 1996, 66, 231 242 NUMBER 2(SEPTEMBER) SEPARATING THE EFFECTS OF TRIAL-SPECIFIC AND AVERAGE SAMPLE-STIMULUS DURATION IN DELAYED MATCHING TO SAMPLE IN PIGEONS
CRF or an Fl 5 min schedule. They found no. of S presentation. Although more responses. might occur under an Fl 5 min than under a
 JOURNAL OF THE EXPERIMENTAL ANALYSIS OF BEHAVIOR VOLUME 5, NUMBF- 4 OCITOBER, 1 962 THE EFECT OF TWO SCHEDULES OF PRIMARY AND CONDITIONED REINFORCEMENT JOAN G. STEVENSON1 AND T. W. REESE MOUNT HOLYOKE
JOURNAL OF THE EXPERIMENTAL ANALYSIS OF BEHAVIOR VOLUME 5, NUMBF- 4 OCITOBER, 1 962 THE EFECT OF TWO SCHEDULES OF PRIMARY AND CONDITIONED REINFORCEMENT JOAN G. STEVENSON1 AND T. W. REESE MOUNT HOLYOKE
Examining the Constant Difference Effect in a Concurrent Chains Procedure
 University of Wisconsin Milwaukee UWM Digital Commons Theses and Dissertations May 2015 Examining the Constant Difference Effect in a Concurrent Chains Procedure Carrie Suzanne Prentice University of Wisconsin-Milwaukee
University of Wisconsin Milwaukee UWM Digital Commons Theses and Dissertations May 2015 Examining the Constant Difference Effect in a Concurrent Chains Procedure Carrie Suzanne Prentice University of Wisconsin-Milwaukee
on both components of conc Fl Fl schedules, c and a were again less than 1.0. FI schedule when these were arranged concurrently.
 JOURNAL OF THE EXPERIMENTAL ANALYSIS OF BEHAVIOR 1975, 24, 191-197 NUMBER 2 (SEPTEMBER) PERFORMANCE IN CONCURRENT INTERVAL SCHEDULES: A SYSTEMATIC REPLICATION' BRENDA LOBB AND M. C. DAVISON UNIVERSITY
JOURNAL OF THE EXPERIMENTAL ANALYSIS OF BEHAVIOR 1975, 24, 191-197 NUMBER 2 (SEPTEMBER) PERFORMANCE IN CONCURRENT INTERVAL SCHEDULES: A SYSTEMATIC REPLICATION' BRENDA LOBB AND M. C. DAVISON UNIVERSITY
Further Tests of Response-Outcome Associations in Differential-Outcome Matching-to-Sample
 Journal of Experimental Psychology: Animal Behavior Processes 1«7, Vol. 23, No. 2, 171-182 Copyright 1997 by the American Psychological Association. Inc. 0097-7*>3/97/J3.00 Further Tests of Response-Outcome
Journal of Experimental Psychology: Animal Behavior Processes 1«7, Vol. 23, No. 2, 171-182 Copyright 1997 by the American Psychological Association. Inc. 0097-7*>3/97/J3.00 Further Tests of Response-Outcome
ANTECEDENT REINFORCEMENT CONTINGENCIES IN THE STIMULUS CONTROL OF AN A UDITORY DISCRIMINA TION' ROSEMARY PIERREL AND SCOT BLUE
 JOURNAL OF THE EXPERIMENTAL ANALYSIS OF BEHAVIOR ANTECEDENT REINFORCEMENT CONTINGENCIES IN THE STIMULUS CONTROL OF AN A UDITORY DISCRIMINA TION' ROSEMARY PIERREL AND SCOT BLUE BROWN UNIVERSITY 1967, 10,
JOURNAL OF THE EXPERIMENTAL ANALYSIS OF BEHAVIOR ANTECEDENT REINFORCEMENT CONTINGENCIES IN THE STIMULUS CONTROL OF AN A UDITORY DISCRIMINA TION' ROSEMARY PIERREL AND SCOT BLUE BROWN UNIVERSITY 1967, 10,
Pigeons memory for time: Assessment of the role of subjective shortening in the duration-comparison procedure
 Learning & Behavior 2009, 37 (1), 74-84 doi:10.3758/lb.37.1.74 Pigeons memory for time: Assessment of the role of subjective shortening in the duration-comparison procedure PATRICK VAN ROOYEN AND ANGELO
Learning & Behavior 2009, 37 (1), 74-84 doi:10.3758/lb.37.1.74 Pigeons memory for time: Assessment of the role of subjective shortening in the duration-comparison procedure PATRICK VAN ROOYEN AND ANGELO
Transfer of Serial Reversal Learning in the Pigeon
 The Quarterly Journal of Experimental Psychology (1986) 38B, 81-95 Transfer of Serial Reversal Learning in the Pigeon P. J. Durlach and N. J. Mackintosh Department of Experimental Psychology, University
The Quarterly Journal of Experimental Psychology (1986) 38B, 81-95 Transfer of Serial Reversal Learning in the Pigeon P. J. Durlach and N. J. Mackintosh Department of Experimental Psychology, University
Transfer Across Delayed Discriminations: II. Differences in the Substitutability of Initial Versus Test Stimuli
 Joulmal of Experimental Psychology: Copyright 1998 by the American Psychological Association, Inc. Animal Behavior Processes 0097-7403/98/$3.00 1998, VoL 24, No. 1, 47-59 Transfer Across Delayed Discriminations:
Joulmal of Experimental Psychology: Copyright 1998 by the American Psychological Association, Inc. Animal Behavior Processes 0097-7403/98/$3.00 1998, VoL 24, No. 1, 47-59 Transfer Across Delayed Discriminations:
Memory Systems Interaction in the Pigeon: Working and Reference Memory
 Journal of Experimental Psychology: Animal Learning and Cognition 2015 American Psychological Association 2015, Vol. 41, No. 2, 152 162 2329-8456/15/$12.00 http://dx.doi.org/10.1037/xan0000053 Memory Systems
Journal of Experimental Psychology: Animal Learning and Cognition 2015 American Psychological Association 2015, Vol. 41, No. 2, 152 162 2329-8456/15/$12.00 http://dx.doi.org/10.1037/xan0000053 Memory Systems
Coding of feature and no-feature events by pigeons performing a delayed conditional discrimination
 Animal Learning & Behavior 1993, 21(2), 92-1 Coding of feature and no-feature events by pigeons performing a delayed conditional discrimination LOU M. SHERBURNE and THOMAS R. ZENTALL University of Kentucky,
Animal Learning & Behavior 1993, 21(2), 92-1 Coding of feature and no-feature events by pigeons performing a delayed conditional discrimination LOU M. SHERBURNE and THOMAS R. ZENTALL University of Kentucky,
EFFECTS OF A LIMITED HOLD ON PIGEONS MATCH-TO-SAMPLE PERFORMANCE UNDER FIXED-RATIO SCHEDULING. Joseph Leland Cermak, B.A.
 EFFECTS OF A LIMITED HOLD ON PIGEONS MATCH-TO-SAMPLE PERFORMANCE UNDER FIXED-RATIO SCHEDULING Joseph Leland Cermak, B.A. Thesis Prepared for the Degree of MASTER OF SCIENCE UNIVERSITY OF NORTH TEXAS December
EFFECTS OF A LIMITED HOLD ON PIGEONS MATCH-TO-SAMPLE PERFORMANCE UNDER FIXED-RATIO SCHEDULING Joseph Leland Cermak, B.A. Thesis Prepared for the Degree of MASTER OF SCIENCE UNIVERSITY OF NORTH TEXAS December
The effects of two different states of food deprivation for 6 roosters was measured with a
 Effects of Food Deprivation on Memory Performance. Jacob. L. Kerewaro Abstract The effects of two different states of food deprivation for 6 roosters was measured with a delayed matching-to-sample (DMTS)
Effects of Food Deprivation on Memory Performance. Jacob. L. Kerewaro Abstract The effects of two different states of food deprivation for 6 roosters was measured with a delayed matching-to-sample (DMTS)
Pigeons' memory for number of events: EVects of intertrial interval and delay interval illumination
 Learning and Motivation 35 (2004) 348 370 www.elsevier.com/locate/l&m Pigeons' memory for number of events: EVects of intertrial interval and delay interval illumination Chris Hope and Angelo Santi Wilfrid
Learning and Motivation 35 (2004) 348 370 www.elsevier.com/locate/l&m Pigeons' memory for number of events: EVects of intertrial interval and delay interval illumination Chris Hope and Angelo Santi Wilfrid
Jennifer J. McComas and Ellie C. Hartman. Angel Jimenez
 The Psychological Record, 28, 58, 57 528 Some Effects of Magnitude of Reinforcement on Persistence of Responding Jennifer J. McComas and Ellie C. Hartman The University of Minnesota Angel Jimenez The University
The Psychological Record, 28, 58, 57 528 Some Effects of Magnitude of Reinforcement on Persistence of Responding Jennifer J. McComas and Ellie C. Hartman The University of Minnesota Angel Jimenez The University
The generality of within-session patterns of responding: Rate of reinforcement and session length
 Animal Learning & Behavior 1994, 22 (3), 252-266 The generality of within-session patterns of responding: Rate of reinforcement and session length FRANCES K. MCSWEENEY, JOHN M. ROLL, and CARI B. CANNON
Animal Learning & Behavior 1994, 22 (3), 252-266 The generality of within-session patterns of responding: Rate of reinforcement and session length FRANCES K. MCSWEENEY, JOHN M. ROLL, and CARI B. CANNON
Reinforcer Magnitude and Resistance to Change of Forgetting Functions and Response Rates
 Utah State University DigitalCommons@USU All Graduate Theses and Dissertations Graduate Studies 8-2012 Reinforcer Magnitude and Resistance to Change of Forgetting Functions and Response Rates Meredith
Utah State University DigitalCommons@USU All Graduate Theses and Dissertations Graduate Studies 8-2012 Reinforcer Magnitude and Resistance to Change of Forgetting Functions and Response Rates Meredith
The Simon Effect as a Function of Temporal Overlap between Relevant and Irrelevant
 University of North Florida UNF Digital Commons All Volumes (2001-2008) The Osprey Journal of Ideas and Inquiry 2008 The Simon Effect as a Function of Temporal Overlap between Relevant and Irrelevant Leslie
University of North Florida UNF Digital Commons All Volumes (2001-2008) The Osprey Journal of Ideas and Inquiry 2008 The Simon Effect as a Function of Temporal Overlap between Relevant and Irrelevant Leslie
STIMULUS FUNCTIONS IN TOKEN-REINFORCEMENT SCHEDULES CHRISTOPHER E. BULLOCK
 STIMULUS FUNCTIONS IN TOKEN-REINFORCEMENT SCHEDULES By CHRISTOPHER E. BULLOCK A DISSERTATION PRESENTED TO THE GRADUATE SCHOOL OF THE UNIVERSITY OF FLORIDA IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR
STIMULUS FUNCTIONS IN TOKEN-REINFORCEMENT SCHEDULES By CHRISTOPHER E. BULLOCK A DISSERTATION PRESENTED TO THE GRADUATE SCHOOL OF THE UNIVERSITY OF FLORIDA IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR
Testing pigeon memory in a change detection task
 Psychonomic Bulletin & Review 21, 17 (2), 243-249 doi:1.3758/pbr.17.2.243 Testing pigeon memory in a change detection task ANTHONY A. WRIGHT University of Texas Health Science Center, Houston, Texas JEFFREY
Psychonomic Bulletin & Review 21, 17 (2), 243-249 doi:1.3758/pbr.17.2.243 Testing pigeon memory in a change detection task ANTHONY A. WRIGHT University of Texas Health Science Center, Houston, Texas JEFFREY
RECALL OF PAIRED-ASSOCIATES AS A FUNCTION OF OVERT AND COVERT REHEARSAL PROCEDURES TECHNICAL REPORT NO. 114 PSYCHOLOGY SERIES
 RECALL OF PAIRED-ASSOCIATES AS A FUNCTION OF OVERT AND COVERT REHEARSAL PROCEDURES by John W. Brelsford, Jr. and Richard C. Atkinson TECHNICAL REPORT NO. 114 July 21, 1967 PSYCHOLOGY SERIES!, Reproduction
RECALL OF PAIRED-ASSOCIATES AS A FUNCTION OF OVERT AND COVERT REHEARSAL PROCEDURES by John W. Brelsford, Jr. and Richard C. Atkinson TECHNICAL REPORT NO. 114 July 21, 1967 PSYCHOLOGY SERIES!, Reproduction
REPEATED MEASUREMENTS OF REINFORCEMENT SCHEDULE EFFECTS ON GRADIENTS OF STIMULUS CONTROL' MICHAEL D. ZEILER
 JOURNAL OF THE EXPERIMENTAL ANALYSIS OF BEHAVIOR REPEATED MEASUREMENTS OF REINFORCEMENT SCHEDULE EFFECTS ON GRADIENTS OF STIMULUS CONTROL' MICHAEL D. ZEILER UNIVERSITY OF IOWA 1969, 12, 451-461 NUMBER
JOURNAL OF THE EXPERIMENTAL ANALYSIS OF BEHAVIOR REPEATED MEASUREMENTS OF REINFORCEMENT SCHEDULE EFFECTS ON GRADIENTS OF STIMULUS CONTROL' MICHAEL D. ZEILER UNIVERSITY OF IOWA 1969, 12, 451-461 NUMBER
STEPHEN P. KRAMER. (Kojima, 1980; Lattal, 1975; Maki, Moe, &
 JOURNAL OF THE EXPERIMENTAL ANALYSIS OF BEHAVIOR1 1982, 38, 71-85 NUMBER I (JULY) MEMORY FOR RECENT BEHAVIOR IN THE PIGEON STEPHEN P. KRAMER CORRECTIONS DIVISION, DRAPER, UTAH Variations of the symbolic
JOURNAL OF THE EXPERIMENTAL ANALYSIS OF BEHAVIOR1 1982, 38, 71-85 NUMBER I (JULY) MEMORY FOR RECENT BEHAVIOR IN THE PIGEON STEPHEN P. KRAMER CORRECTIONS DIVISION, DRAPER, UTAH Variations of the symbolic
Birds' Judgments of Number and Quantity
 Entire Set of Printable Figures For Birds' Judgments of Number and Quantity Emmerton Figure 1. Figure 2. Examples of novel transfer stimuli in an experiment reported in Emmerton & Delius (1993). Paired
Entire Set of Printable Figures For Birds' Judgments of Number and Quantity Emmerton Figure 1. Figure 2. Examples of novel transfer stimuli in an experiment reported in Emmerton & Delius (1993). Paired
INTRODUCING NEW STIMULI IN FADING
 JOURNL OF THE EXPERMENTL NLYSS OF BEHVOR 1979, 32, 121-127 NUMBER (JULY) CQUSTON OF STMULUS CONTROL WHLE NTRODUCNG NEW STMUL N FDNG LNNY FELDS THE COLLEGE OF STTEN SLND fter establishing a discrimination
JOURNL OF THE EXPERMENTL NLYSS OF BEHVOR 1979, 32, 121-127 NUMBER (JULY) CQUSTON OF STMULUS CONTROL WHLE NTRODUCNG NEW STMUL N FDNG LNNY FELDS THE COLLEGE OF STTEN SLND fter establishing a discrimination
The effects of reinforcement upon the prepecking behaviors of pigeons in the autoshaping experiment.
 University of Massachusetts Amherst ScholarWorks@UMass Amherst Masters Theses 1911 - February 2014 1974 The effects of reinforcement upon the prepecking behaviors of pigeons in the autoshaping experiment.
University of Massachusetts Amherst ScholarWorks@UMass Amherst Masters Theses 1911 - February 2014 1974 The effects of reinforcement upon the prepecking behaviors of pigeons in the autoshaping experiment.
A Memory Model for Decision Processes in Pigeons
 From M. L. Commons, R.J. Herrnstein, & A.R. Wagner (Eds.). 1983. Quantitative Analyses of Behavior: Discrimination Processes. Cambridge, MA: Ballinger (Vol. IV, Chapter 1, pages 3-19). A Memory Model for
From M. L. Commons, R.J. Herrnstein, & A.R. Wagner (Eds.). 1983. Quantitative Analyses of Behavior: Discrimination Processes. Cambridge, MA: Ballinger (Vol. IV, Chapter 1, pages 3-19). A Memory Model for
FIXED-RATIO PUNISHMENT1 N. H. AZRIN,2 W. C. HOLZ,2 AND D. F. HAKE3
 JOURNAL OF THE EXPERIMENTAL ANALYSIS OF BEHAVIOR VOLUME 6, NUMBER 2 APRIL, 1963 FIXED-RATIO PUNISHMENT1 N. H. AZRIN,2 W. C. HOLZ,2 AND D. F. HAKE3 Responses were maintained by a variable-interval schedule
JOURNAL OF THE EXPERIMENTAL ANALYSIS OF BEHAVIOR VOLUME 6, NUMBER 2 APRIL, 1963 FIXED-RATIO PUNISHMENT1 N. H. AZRIN,2 W. C. HOLZ,2 AND D. F. HAKE3 Responses were maintained by a variable-interval schedule
Olfactory Matching-To-Sample In Rats Using a Novel Apparatus. Rhiannon D. Thomas
 Olfactory Matching-To-Sample In Rats Using a Novel Apparatus Rhiannon D. Thomas A Thesis Submitted to the University of North Carolina Wilmington in Partial Fulfillment of the Requirements for the Degree
Olfactory Matching-To-Sample In Rats Using a Novel Apparatus Rhiannon D. Thomas A Thesis Submitted to the University of North Carolina Wilmington in Partial Fulfillment of the Requirements for the Degree
AN EXPLORATION OF THE TITRATING DELAY MATCH TO SAMPLE PROCEDURE WITH PIGEONS. Jonathan E. Friedel, B.S. Thesis Prepared for the Degree of
 AN EXPLORATION OF THE TITRATING DELAY MATCH TO SAMPLE PROCEDURE WITH PIGEONS Jonathan E. Friedel, B.S. Thesis Prepared for the Degree of MASTER OF SCIENCE UNIVERSITY OF NORTH TEXAS December 2011 APPROVED:
AN EXPLORATION OF THE TITRATING DELAY MATCH TO SAMPLE PROCEDURE WITH PIGEONS Jonathan E. Friedel, B.S. Thesis Prepared for the Degree of MASTER OF SCIENCE UNIVERSITY OF NORTH TEXAS December 2011 APPROVED:
Incorporating quantitative information into a linear ordering" GEORGE R. POTTS Dartmouth College, Hanover, New Hampshire 03755
 Memory & Cognition 1974, Vol. 2, No.3, 533 538 Incorporating quantitative information into a linear ordering" GEORGE R. POTTS Dartmouth College, Hanover, New Hampshire 03755 Ss were required to learn linear
Memory & Cognition 1974, Vol. 2, No.3, 533 538 Incorporating quantitative information into a linear ordering" GEORGE R. POTTS Dartmouth College, Hanover, New Hampshire 03755 Ss were required to learn linear
Discrimination Trials to Influence Self-Awareness
 University of Wisconsin Milwaukee UWM Digital Commons Theses and Dissertations May 2013 Discrimination Trials to Influence Self-Awareness Kerin A. Weingarten University of Wisconsin-Milwaukee Follow this
University of Wisconsin Milwaukee UWM Digital Commons Theses and Dissertations May 2013 Discrimination Trials to Influence Self-Awareness Kerin A. Weingarten University of Wisconsin-Milwaukee Follow this
Supporting Online Material for
 www.sciencemag.org/cgi/content/full/319/5871/1849/dc1 Supporting Online Material for Rule Learning by Rats Robin A. Murphy,* Esther Mondragón, Victoria A. Murphy This PDF file includes: *To whom correspondence
www.sciencemag.org/cgi/content/full/319/5871/1849/dc1 Supporting Online Material for Rule Learning by Rats Robin A. Murphy,* Esther Mondragón, Victoria A. Murphy This PDF file includes: *To whom correspondence
The moderating effects of direct and indirect experience on the attitude-behavior relation in the reasoned and automatic processing modes.
 University of Massachusetts Amherst ScholarWorks@UMass Amherst Masters Theses 1911 - February 2014 1995 The moderating effects of direct and indirect experience on the attitude-behavior relation in the
University of Massachusetts Amherst ScholarWorks@UMass Amherst Masters Theses 1911 - February 2014 1995 The moderating effects of direct and indirect experience on the attitude-behavior relation in the
UNIVERSITY OF IOWA AND SOUTHERN ILLINOIS UNIVERSITY AT CARBONDALE
 JOURNAL OF THE EXPERIMENTAL ANALYSIS OF BEHAVIOR 2002, 78, 365 373 NUMBER 3(NOVEMBER) BRIEF PRESENTATIONS ARE SUFFICIENT FOR PIGEONS TO DISCRIMINATE ARRAYS OF SAME AND DIFFERENT STIMULI EDWARD A. WASSERMAN,
JOURNAL OF THE EXPERIMENTAL ANALYSIS OF BEHAVIOR 2002, 78, 365 373 NUMBER 3(NOVEMBER) BRIEF PRESENTATIONS ARE SUFFICIENT FOR PIGEONS TO DISCRIMINATE ARRAYS OF SAME AND DIFFERENT STIMULI EDWARD A. WASSERMAN,
Discrimination and Generalization in Pattern Categorization: A Case for Elemental Associative Learning
 Discrimination and Generalization in Pattern Categorization: A Case for Elemental Associative Learning E. J. Livesey (el253@cam.ac.uk) P. J. C. Broadhurst (pjcb3@cam.ac.uk) I. P. L. McLaren (iplm2@cam.ac.uk)
Discrimination and Generalization in Pattern Categorization: A Case for Elemental Associative Learning E. J. Livesey (el253@cam.ac.uk) P. J. C. Broadhurst (pjcb3@cam.ac.uk) I. P. L. McLaren (iplm2@cam.ac.uk)
Oddity-from-Sample Abstract-Concept Learning by Pigeons. Thomas A. Daniel
 Oddity-from-Sample Abstract-Concept Learning by Pigeons by Thomas A. Daniel A thesis proposal submitted to the Graduate Faculty of Auburn University in partial fulfillment of the requirements for the Degree
Oddity-from-Sample Abstract-Concept Learning by Pigeons by Thomas A. Daniel A thesis proposal submitted to the Graduate Faculty of Auburn University in partial fulfillment of the requirements for the Degree
REINFORCEMENT OF PROBE RESPONSES AND ACQUISITION OF STIMULUS CONTROL IN FADING PROCEDURES
 JOURNAL OF THE EXPERIMENTAL ANALYSIS OF BEHAVIOR 1985, 439 235-241 NUMBER 2 (MARCH) REINFORCEMENT OF PROBE RESPONSES AND ACQUISITION OF STIMULUS CONTROL IN FADING PROCEDURES LANNY FIELDS THE COLLEGE OF
JOURNAL OF THE EXPERIMENTAL ANALYSIS OF BEHAVIOR 1985, 439 235-241 NUMBER 2 (MARCH) REINFORCEMENT OF PROBE RESPONSES AND ACQUISITION OF STIMULUS CONTROL IN FADING PROCEDURES LANNY FIELDS THE COLLEGE OF
Behavioural Processes
 Behavioural Processes 89 (2012) 212 218 Contents lists available at SciVerse ScienceDirect Behavioural Processes j o ur nal homep age : www.elsevier.com/locate/behavproc Providing a reinforcement history
Behavioural Processes 89 (2012) 212 218 Contents lists available at SciVerse ScienceDirect Behavioural Processes j o ur nal homep age : www.elsevier.com/locate/behavproc Providing a reinforcement history
The effect of sample duration and cue on a double temporal discrimination q
 Available online at www.sciencedirect.com Learning and Motivation 39 (2008) 71 94 www.elsevier.com/locate/l&m The effect of sample duration and cue on a double temporal discrimination q Luís Oliveira,
Available online at www.sciencedirect.com Learning and Motivation 39 (2008) 71 94 www.elsevier.com/locate/l&m The effect of sample duration and cue on a double temporal discrimination q Luís Oliveira,
Sequences of Fixed-Ratio Schedules of Reinforcement: The Effect of Ratio Size in the Second and Third Fixed-Ratio on Pigeons' Choice
 Western Michigan University ScholarWorks at WMU Dissertations Graduate College 12-1991 Sequences of Fixed-Ratio Schedules of Reinforcement: The Effect of Ratio Size in the Second and Third Fixed-Ratio
Western Michigan University ScholarWorks at WMU Dissertations Graduate College 12-1991 Sequences of Fixed-Ratio Schedules of Reinforcement: The Effect of Ratio Size in the Second and Third Fixed-Ratio
Testing the Functional Equivalence of Retention Intervals and Sample-Stimulus Disparity in Conditional Discrimination
 Utah State University DigitalCommons@USU All Graduate Theses and Dissertations Graduate Studies 5-2008 Testing the Functional Equivalence of Retention Intervals and Sample-Stimulus Disparity in Conditional
Utah State University DigitalCommons@USU All Graduate Theses and Dissertations Graduate Studies 5-2008 Testing the Functional Equivalence of Retention Intervals and Sample-Stimulus Disparity in Conditional
Are Retrievals from Long-Term Memory Interruptible?
 Are Retrievals from Long-Term Memory Interruptible? Michael D. Byrne byrne@acm.org Department of Psychology Rice University Houston, TX 77251 Abstract Many simple performance parameters about human memory
Are Retrievals from Long-Term Memory Interruptible? Michael D. Byrne byrne@acm.org Department of Psychology Rice University Houston, TX 77251 Abstract Many simple performance parameters about human memory
Performance of Four Seed-Caching Corvid Species in Operant Tests of Nonspatial and Spatial Memory
 University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Papers in Behavior and Biological Sciences Papers in the Biological Sciences 1995 Performance of Four Seed-Caching Corvid
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Papers in Behavior and Biological Sciences Papers in the Biological Sciences 1995 Performance of Four Seed-Caching Corvid
Increasing the persistence of a heterogeneous behavior chain: Studies of extinction in a rat model of search behavior of working dogs
 Increasing the persistence of a heterogeneous behavior chain: Studies of extinction in a rat model of search behavior of working dogs Eric A. Thrailkill 1, Alex Kacelnik 2, Fay Porritt 3 & Mark E. Bouton
Increasing the persistence of a heterogeneous behavior chain: Studies of extinction in a rat model of search behavior of working dogs Eric A. Thrailkill 1, Alex Kacelnik 2, Fay Porritt 3 & Mark E. Bouton
The Role of Feedback in Categorisation
 The Role of in Categorisation Mark Suret (m.suret@psychol.cam.ac.uk) Department of Experimental Psychology; Downing Street Cambridge, CB2 3EB UK I.P.L. McLaren (iplm2@cus.cam.ac.uk) Department of Experimental
The Role of in Categorisation Mark Suret (m.suret@psychol.cam.ac.uk) Department of Experimental Psychology; Downing Street Cambridge, CB2 3EB UK I.P.L. McLaren (iplm2@cus.cam.ac.uk) Department of Experimental
AMOUNT OF RESPONSE-PRODUCED CHANGE IN THE CS AND AVOIDANCE LEARNING 1
 Journal of Comparative and Physiological Psychology 1965, Vol. 59, No. 1, 13-17 AMOUNT OF RESPONSE-PRODUCED CHANGE IN THE CS AND AVOIDANCE LEARNING 1 GORDON BOWER, RONALD STARR, AND LEAH LAZAROVITZ Stanford
Journal of Comparative and Physiological Psychology 1965, Vol. 59, No. 1, 13-17 AMOUNT OF RESPONSE-PRODUCED CHANGE IN THE CS AND AVOIDANCE LEARNING 1 GORDON BOWER, RONALD STARR, AND LEAH LAZAROVITZ Stanford
Replacing the frontal lobes? Having more time to think improve implicit perceptual categorization. A comment on Filoteo, Lauritzen & Maddox, 2010.
 Replacing the frontal lobes? 1 Replacing the frontal lobes? Having more time to think improve implicit perceptual categorization. A comment on Filoteo, Lauritzen & Maddox, 2010. Ben R. Newell 1 Christopher
Replacing the frontal lobes? 1 Replacing the frontal lobes? Having more time to think improve implicit perceptual categorization. A comment on Filoteo, Lauritzen & Maddox, 2010. Ben R. Newell 1 Christopher
Variability as an Operant?
 The Behavior Analyst 2012, 35, 243 248 No. 2 (Fall) Variability as an Operant? Per Holth Oslo and Akershus University College Correspondence concerning this commentary should be addressed to Per Holth,
The Behavior Analyst 2012, 35, 243 248 No. 2 (Fall) Variability as an Operant? Per Holth Oslo and Akershus University College Correspondence concerning this commentary should be addressed to Per Holth,
EFFECTS OF NICOTINE ADMINISTRATION ON PERFORMANCE UNDER TWO PROCEDURES DESIGNED TO TEST SHORT-TERM REMEMBERING
 EFFECTS OF NICOTINE ADMINISTRATION ON PERFORMANCE UNDER TWO PROCEDURES DESIGNED TO TEST SHORT-TERM REMEMBERING By BRIAN D. KANGAS A DISSERTATION PRESENTED TO THE GRADUATE SCHOOL OF THE UNIVERSITY OF FLORIDA
EFFECTS OF NICOTINE ADMINISTRATION ON PERFORMANCE UNDER TWO PROCEDURES DESIGNED TO TEST SHORT-TERM REMEMBERING By BRIAN D. KANGAS A DISSERTATION PRESENTED TO THE GRADUATE SCHOOL OF THE UNIVERSITY OF FLORIDA
Confounding variables in the discriminated Irt procedure.
 University of Massachusetts Amherst ScholarWorks@UMass Amherst Masters Theses 1911 - February 2014 1983 Confounding variables in the discriminated Irt procedure. David C. Palmer University of Massachusetts
University of Massachusetts Amherst ScholarWorks@UMass Amherst Masters Theses 1911 - February 2014 1983 Confounding variables in the discriminated Irt procedure. David C. Palmer University of Massachusetts
Instrumental Conditioning I
 Instrumental Conditioning I Basic Procedures and Processes Instrumental or Operant Conditioning? These terms both refer to learned changes in behavior that occur as a result of the consequences of the
Instrumental Conditioning I Basic Procedures and Processes Instrumental or Operant Conditioning? These terms both refer to learned changes in behavior that occur as a result of the consequences of the
Supplemental Data: Capuchin Monkeys Are Sensitive to Others Welfare. Venkat R. Lakshminarayanan and Laurie R. Santos
 Supplemental Data: Capuchin Monkeys Are Sensitive to Others Welfare Venkat R. Lakshminarayanan and Laurie R. Santos Supplemental Experimental Procedures Subjects Seven adult capuchin monkeys were tested.
Supplemental Data: Capuchin Monkeys Are Sensitive to Others Welfare Venkat R. Lakshminarayanan and Laurie R. Santos Supplemental Experimental Procedures Subjects Seven adult capuchin monkeys were tested.
THE EFFECTS OF TERMINAL-LINK STIMULUS ARRANGEMENTS ON PREFERENCE IN CONCURRENT CHAINS. LAUREL COLTON and JAY MOORE University of Wisconsin-Milwaukee
 The Psychological Record, 1997,47,145-166 THE EFFECTS OF TERMINAL-LINK STIMULUS ARRANGEMENTS ON PREFERENCE IN CONCURRENT CHAINS LAUREL COLTON and JAY MOORE University of Wisconsin-Milwaukee Pigeons served
The Psychological Record, 1997,47,145-166 THE EFFECTS OF TERMINAL-LINK STIMULUS ARRANGEMENTS ON PREFERENCE IN CONCURRENT CHAINS LAUREL COLTON and JAY MOORE University of Wisconsin-Milwaukee Pigeons served
DISCRIMINATION IN RATS OSAKA CITY UNIVERSITY. to emit the response in question. Within this. in the way of presenting the enabling stimulus.
 JOURNAL OF THE EXPERIMENTAL ANALYSIS OF BEHAVIOR EFFECTS OF DISCRETE-TRIAL AND FREE-OPERANT PROCEDURES ON THE ACQUISITION AND MAINTENANCE OF SUCCESSIVE DISCRIMINATION IN RATS SHIN HACHIYA AND MASATO ITO
JOURNAL OF THE EXPERIMENTAL ANALYSIS OF BEHAVIOR EFFECTS OF DISCRETE-TRIAL AND FREE-OPERANT PROCEDURES ON THE ACQUISITION AND MAINTENANCE OF SUCCESSIVE DISCRIMINATION IN RATS SHIN HACHIYA AND MASATO ITO
Conditional Relations among Abstract Stimuli: Outcomes from Three Procedures- Variations of Go/no-go and Match-to-Sample. A Thesis Presented
 1 Conditional Relations among Abstract Stimuli: Outcomes from Three Procedures- Variations of Go/no-go and Match-to-Sample A Thesis Presented by Kimberly M. Walter The Department of Counseling and Applied
1 Conditional Relations among Abstract Stimuli: Outcomes from Three Procedures- Variations of Go/no-go and Match-to-Sample A Thesis Presented by Kimberly M. Walter The Department of Counseling and Applied
Spatially Diffuse Inhibition Affects Multiple Locations: A Reply to Tipper, Weaver, and Watson (1996)
") Journal of Experimental Psychology: Human Perception and Performance 1996, Vol. 22, No. 5, 1294-1298 Copyright 1996 by the American Psychological Association, Inc. 0096-1523/%/$3.00 Spatially Diffuse Inhibition
Journal of Experimental Psychology: Human Perception and Performance 1996, Vol. 22, No. 5, 1294-1298 Copyright 1996 by the American Psychological Association, Inc. 0096-1523/%/$3.00 Spatially Diffuse Inhibition
Predictive Accuracy and the Effects of Partial Reinforcement on Serial Autoshaping
 Journal of Experimental Psychology: Copyright 1985 by the American Psychological Association, Inc. Animal Behavior Processes 0097-7403/85/$00.75 1985, VOl. 11, No. 4, 548-564 Predictive Accuracy and the
Journal of Experimental Psychology: Copyright 1985 by the American Psychological Association, Inc. Animal Behavior Processes 0097-7403/85/$00.75 1985, VOl. 11, No. 4, 548-564 Predictive Accuracy and the
CONDITIONED REINFORCEMENT IN RATS'
 JOURNAL OF THE EXPERIMENTAL ANALYSIS OF BEHAVIOR 1969, 12, 261-268 NUMBER 2 (MARCH) CONCURRENT SCHEULES OF PRIMARY AN CONITIONE REINFORCEMENT IN RATS' ONAL W. ZIMMERMAN CARLETON UNIVERSITY Rats responded
JOURNAL OF THE EXPERIMENTAL ANALYSIS OF BEHAVIOR 1969, 12, 261-268 NUMBER 2 (MARCH) CONCURRENT SCHEULES OF PRIMARY AN CONITIONE REINFORCEMENT IN RATS' ONAL W. ZIMMERMAN CARLETON UNIVERSITY Rats responded
"Same/different" symbol use by pigeons
 Animal Learning & Behavior 1983, 11 (3),349 355 "Same/different" symbol use by pigeons CHARLES A, EDWARDS, JOYCE A. JAGIELO, and THOMAS R. ZENTALL University ofkentucky, Lexington, Kentucky Pigeons learned
Animal Learning & Behavior 1983, 11 (3),349 355 "Same/different" symbol use by pigeons CHARLES A, EDWARDS, JOYCE A. JAGIELO, and THOMAS R. ZENTALL University ofkentucky, Lexington, Kentucky Pigeons learned
Main Study: Summer Methods. Design
 Main Study: Summer 2000 Methods Design The experimental design is within-subject each participant experiences five different trials for each of the ten levels of Display Condition and for each of the three
Main Study: Summer 2000 Methods Design The experimental design is within-subject each participant experiences five different trials for each of the ten levels of Display Condition and for each of the three
by instructional cues
 Animal Learning & Behavior 1985 13 (4) 383-391 Control of pigeons' matching and mismatching performance by instructional cues CHARLES A. EDWARDS Dalhousie University Halifax Nova Scotia Canada and JAMES
Animal Learning & Behavior 1985 13 (4) 383-391 Control of pigeons' matching and mismatching performance by instructional cues CHARLES A. EDWARDS Dalhousie University Halifax Nova Scotia Canada and JAMES
Delayed Matching-To-Sample Test in Macaques
 C O N F I D E N T I A L Delayed Matching-To-Sample Test in Macaques DATE This study was conducted under the terms of a Materials Transfer and Services Agreement between NeuroDetective International and
C O N F I D E N T I A L Delayed Matching-To-Sample Test in Macaques DATE This study was conducted under the terms of a Materials Transfer and Services Agreement between NeuroDetective International and
Cognitive dissonance in children: Justification of effort or contrast?
 Psychonomic Bulletin & Review 2008, 15 (3), 673-677 doi: 10.3758/PBR.15.3.673 Cognitive dissonance in children: Justification of effort or contrast? JÉRÔME ALESSANDRI AND JEAN-CLAUDE DARCHEVILLE University
Psychonomic Bulletin & Review 2008, 15 (3), 673-677 doi: 10.3758/PBR.15.3.673 Cognitive dissonance in children: Justification of effort or contrast? JÉRÔME ALESSANDRI AND JEAN-CLAUDE DARCHEVILLE University
Object Substitution Masking: When does Mask Preview work?
 Object Substitution Masking: When does Mask Preview work? Stephen W. H. Lim (psylwhs@nus.edu.sg) Department of Psychology, National University of Singapore, Block AS6, 11 Law Link, Singapore 117570 Chua
Object Substitution Masking: When does Mask Preview work? Stephen W. H. Lim (psylwhs@nus.edu.sg) Department of Psychology, National University of Singapore, Block AS6, 11 Law Link, Singapore 117570 Chua
The Application of the Species Specific Defense Reaction Hypothesis to Free Operant Avoidance
 Western Michigan University ScholarWorks at WMU Master's Theses Graduate College 8-1972 The Application of the Species Specific Defense Reaction Hypothesis to Free Operant Avoidance Deborah Ann Cory Western
Western Michigan University ScholarWorks at WMU Master's Theses Graduate College 8-1972 The Application of the Species Specific Defense Reaction Hypothesis to Free Operant Avoidance Deborah Ann Cory Western
Congruency Effects with Dynamic Auditory Stimuli: Design Implications
 Congruency Effects with Dynamic Auditory Stimuli: Design Implications Bruce N. Walker and Addie Ehrenstein Psychology Department Rice University 6100 Main Street Houston, TX 77005-1892 USA +1 (713) 527-8101
Congruency Effects with Dynamic Auditory Stimuli: Design Implications Bruce N. Walker and Addie Ehrenstein Psychology Department Rice University 6100 Main Street Houston, TX 77005-1892 USA +1 (713) 527-8101
THESES SIS/LIBRARY TELEPHONE:
 THESES SIS/LIBRARY TELEPHONE: +61 2 6125 4631 R.G. MENZIES LIBRARY BUILDING NO:2 FACSIMILE: +61 2 6125 4063 THE AUSTRALIAN NATIONAL UNIVERSITY EMAIL: library.theses@anu.edu.au CANBERRA ACT 0200 AUSTRALIA
THESES SIS/LIBRARY TELEPHONE: +61 2 6125 4631 R.G. MENZIES LIBRARY BUILDING NO:2 FACSIMILE: +61 2 6125 4063 THE AUSTRALIAN NATIONAL UNIVERSITY EMAIL: library.theses@anu.edu.au CANBERRA ACT 0200 AUSTRALIA
CONCEPT LEARNING WITH DIFFERING SEQUENCES OF INSTANCES
 Journal of Experimental Vol. 51, No. 4, 1956 Psychology CONCEPT LEARNING WITH DIFFERING SEQUENCES OF INSTANCES KENNETH H. KURTZ AND CARL I. HOVLAND Under conditions where several concepts are learned concurrently
Journal of Experimental Vol. 51, No. 4, 1956 Psychology CONCEPT LEARNING WITH DIFFERING SEQUENCES OF INSTANCES KENNETH H. KURTZ AND CARL I. HOVLAND Under conditions where several concepts are learned concurrently
EFFECTS OF INTERRESPONSE-TIME SHAPING ON MULTIPLE SCHEDULE PERFORMANCE. RAFAEL BEJARANO University of Kansas
 The Psychological Record, 2004, 54, 479-490 EFFECTS OF INTERRESPONSE-TIME SHAPING ON MULTIPLE SCHEDULE PERFORMANCE RAFAEL BEJARANO University of Kansas The experiment reported herein was conducted to determine
The Psychological Record, 2004, 54, 479-490 EFFECTS OF INTERRESPONSE-TIME SHAPING ON MULTIPLE SCHEDULE PERFORMANCE RAFAEL BEJARANO University of Kansas The experiment reported herein was conducted to determine
. R EAS O ~ I NG BY NONHUMAN
 1 Roger K. Thomas The University of Georgia Athens, Georgia Unites States of America rkthomas@uga.edu Synonyms. R EAS O ~ I NG BY NONHUMAN Pre-publication manuscript version. Published as follows: Thomas,
1 Roger K. Thomas The University of Georgia Athens, Georgia Unites States of America rkthomas@uga.edu Synonyms. R EAS O ~ I NG BY NONHUMAN Pre-publication manuscript version. Published as follows: Thomas,
Transfer of Control in Ambiguous Discriminations
 Journal of Experimental Psychology: Animal Behavior Processes 1991, Vol. 17, No. 3, 231-248 Copyright 1991 by the Am n Psychological Association, Inc. 0097-7403/91/53.00 Transfer of Control in Ambiguous
Journal of Experimental Psychology: Animal Behavior Processes 1991, Vol. 17, No. 3, 231-248 Copyright 1991 by the Am n Psychological Association, Inc. 0097-7403/91/53.00 Transfer of Control in Ambiguous
Observing behavior: Redundant stimuli and time since information
 Animal Learning & Behavior 1978,6 (4),380-384 Copyright 1978 by The Psychonornic Society, nc. Observing behavior: Redundant stimuli and time since information BRUCE A. WALD Utah State University, Logan,
Animal Learning & Behavior 1978,6 (4),380-384 Copyright 1978 by The Psychonornic Society, nc. Observing behavior: Redundant stimuli and time since information BRUCE A. WALD Utah State University, Logan,
Selective changes of sensitivity after adaptation to simple geometrical figures*
 Perception & Psychophysics 1973. Vol. 13. So. 2.356-360 Selective changes of sensitivity after adaptation to simple geometrical figures* ANGEL VASSILEV+ Institu te of Physiology. Bulgarian Academy of Sciences.
Perception & Psychophysics 1973. Vol. 13. So. 2.356-360 Selective changes of sensitivity after adaptation to simple geometrical figures* ANGEL VASSILEV+ Institu te of Physiology. Bulgarian Academy of Sciences.
A STUDY OF FUNCTIONAL EQUIVALENCE IN RATS USING CLASS-SPECIFIC REINFORCERS AND OLFACTORY STIMULI. Rebecca M. Rayburn-Reeves
 A STUDY OF FUNCTIONAL EQUIVALENCE IN RATS USING CLASS-SPECIFIC REINFORCERS AND OLFACTORY STIMULI Rebecca M. Rayburn-Reeves A Thesis Submitted to the University of North Carolina Wilmington in Partial Fulfillment
A STUDY OF FUNCTIONAL EQUIVALENCE IN RATS USING CLASS-SPECIFIC REINFORCERS AND OLFACTORY STIMULI Rebecca M. Rayburn-Reeves A Thesis Submitted to the University of North Carolina Wilmington in Partial Fulfillment
STUDIES OF WHEEL-RUNNING REINFORCEMENT: PARAMETERS OF HERRNSTEIN S (1970) RESPONSE-STRENGTH EQUATION VARY WITH SCHEDULE ORDER TERRY W.
 RESPONSE-STRENGTH EQUATION VARY WITH SCHEDULE ORDER TERRY W.") JOURNAL OF THE EXPERIMENTAL ANALYSIS OF BEHAVIOR 2000, 73, 319 331 NUMBER 3(MAY) STUDIES OF WHEEL-RUNNING REINFORCEMENT: PARAMETERS OF HERRNSTEIN S (1970) RESPONSE-STRENGTH EQUATION VARY WITH SCHEDULE
JOURNAL OF THE EXPERIMENTAL ANALYSIS OF BEHAVIOR 2000, 73, 319 331 NUMBER 3(MAY) STUDIES OF WHEEL-RUNNING REINFORCEMENT: PARAMETERS OF HERRNSTEIN S (1970) RESPONSE-STRENGTH EQUATION VARY WITH SCHEDULE