Two Forms of Spatial Imagery: Neuroimaging Evidence
|
|
|
- Cameron Preston
- 6 years ago
- Views:
Transcription
1 Two Forms of Spatial Imagery: Neuroimaging Evidence The Harvard community has made this article openly available. Please share how this access benefits you. Your story matters. Citation Published Version Accessed Citable Link Terms of Use Thompson, William L., Scott D. Slotnick, Marie S. Burrage, and Stephen M. Kosslyn Two forms of spatial imagery: neuroimaging evidence. Psychological Science 20(10): doi: /j x November 23, :33:25 PM EST This article was downloaded from Harvard University's DASH repository, and is made available under the terms and conditions applicable to Open Access Policy Articles, as set forth at (Article begins on next page)
2 RUNNING HEAD: Different networks for spatial mental imagery Please address correspondence to: Stephen M. Kosslyn Harvard University, Department of Psychology 830 William James Hall 33 Kirkland Street Cambridge MA Tel.: Fax: ABSTRACT--Spatial imagery may be useful in such tasks as graph interpretation, geometry problem-solving, and even in performing surgery. This study provides evidence that spatial imagery is not a single faculty; rather, visualizing spatial location and mental transformation of location rely on distinct neural networks. Using 3T fmri, we tested 16 participants (half male, half female) in each of two spatial imagery tasks one relying on visualizing location and one relying on mentally rotating the stimulus. The same stimuli were used in both tasks. The location-based task engendered more activation near the occipito-parietal sulcus, medial posterior cingulate, and the precuneus. In contrast, the transformation task engendered activation in superior portions of the parietal lobe and in the postcentral gyrus. These differences in activation provide evidence that there are at least two different types of spatial imagery. Two Forms of Spatial Imagery: Neuroimaging Evidence Mental imagery for spatial relationships is useful across a range of domains, and is essential in such everyday activities as finding one's way in an unfamiliar neighborhood, packing suitcases into a car, or remembering locations of objects around a room. In addition, spatial imagery ability may have consequences for career choice. For example, Wanzel, Hamstra, Anastakis, Matsumoto, & Cusimano (2002) report that surgical residents with greater competence in mental rotation were better able to learn and perform a complex surgical procedure. In the same vein, Guillot, Champely, Batier, Thiriet, and Collet (2007) report that anatomy students who had better mental rotation ability performed better on an anatomy test. Moreover, Dror, Kosslyn, and Waag (1993) showed that airplane pilots can perform mental rotation and other spatial judgments better than other comparable participants. Two Forms of Spatial Imagery: Neuroimaging Evidence William L. Thompson 1, Scott D. Slotnick 2, Marie S. Burrage 1, Stephen M. Kosslyn 1, 3 1 Harvard University, Department of Psychology, Cambridge, MA Boston College, Department of Psychology, Chestnut Hill, MA Massachusetts General Hospital, Department of Neurology, Boston, MA 02114
3 A major finding in cognitive neuroscience is that mental functions generally are not unitary and undifferentiated. Rather, they are carried out by a host of distinct representations and processes. This is not only true at coarse levels of analysis -- for example when one considers faculties such as language, memory or perception -- but also at fine levels of analysis. For example, at least two distinct types of spatial relations representations can be used to specify location, one that specifies categories (e.g., "left of") and one that specifies metric (precise distance) information (e.g., Kosslyn, 2006). In this article we consider the possibility that spatial imagery can be decomposed into at least two distinct types of processes. On the one hand, such imagery may serve to specify location; on the other, it may be involved in indicating orientation changes or mentally simulating such changes. Many researchers have studied changes in orientation of objects in visual mental images, usually under the rubric of "mental rotation." Neuroimaging has proven a useful tool in such investigations, allowing researchers to compare and contrast mental rotation with other abilities. In a recent review and meta-analysis of mental rotation, Zacks (2008) found consistent activation in parietal cortex, with some extension into superior portions of occipital cortex. Zacks (2008) proposed that these regions are likely candidates for implementing the transformation-specific processes involved in mental rotation. Moreover, Zacks and Michelon (2005) proposed that spatial transformations rely initially (and essentially) on memory for spatial locations of objects that are encoded relative to one of three reference frames (object-centric, egocentric, or allocentric), and that spatial transformations per se are executed by a small subset of the common brain regions underlying the spatial imagery system in general. Previous neuroimaging studies have found that remembering spatial locations of objects also activates portions of parietal cortex. Moscovitch, Kapur, Köhler, and Houle (1995) reported that Brodmann's Area (BA) 40 in inferior parietal cortex was activated when object location was retrieved (to a greater degree than when object identity was retrieved). Such findings are consistent with results from studying patients with brain damage. For example, van Asselen et al. (2006) found that stroke patients with damage to parietal cortex (and dorsolateral prefrontal cortex) were impaired relative to controls in remembering the locations of objects. However, no previous study has directly compared visualizing location with visualizing transformations of orientation. If spatial transformations rely fundamentally on mapping different objects in space relative to a reference frame, then all of the brain areas evoked by spatial transformations should also be evoked by imagery for spatial location. However, if these processes are distinct, then at least some distinct brain areas should be activated in each case when the two conditions are compared directly. METHOD Participants Sixteen participants (8 male, 8 female), who were undergraduates, graduate students, or professionals, took part in the study (mean age 23, age range 18-30). Seven additional participants were tested but were excluded because of failure to complete at least one of the tasks to criterion, or because MRI scanner or computer equipment problems prohibited them from completing the session. All participants were tested according to applicable guidelines and regulations governing the use of human participants in research, and the experimental protocol was approved by the Harvard University Faculty of Arts and Sciences Committee on the Use of Human Subjects and the Partners Human Research Committee (governing research at Massachusetts General Hospital). Materials Stimuli included 35 alphanumeric characters created from a variety of standard fonts modified to fit the circles in which they would eventually appear. The characters were made to appear standard and prototypical in order to maximize clarity and facilitate learning and consisted of five numerals (1, 2, 3, 7, 9), 12 lowercase letters (a, b, d, f, h, i, j, m, n, q, r, t)
4 and 18 uppercase letters (A, C, D, E, F, G, H, I, J, L, R, S, T, U, V, W, X, Y). For the familiarization phase, the characters were placed within a circle with a tick mark on top (see Figure1 for examples). For the experimental trials, two sets of five characters each were presented at different locations within a rectangle. A circle of the same size as those that surrounded the characters --as first studied by participants-- was presented on each trial. Each circle was divided into three equally sized sections: one outlined with a bold line, one with a dashed line, and the third with a neutral line (see Figure 1). On each trial, a script character (in Apple Chancery font, with minor modifications for some letters to improve clarity) appeared under the circle, to cue participants as to which block character they should visualize in order to perform the task. Each task included 40 trials, presented in two blocks of 20 trials each over the course of the study. Participants were tested on four tasks, two of which are the focus of this report and are described in detail below. Procedure Familiarization phase (outside the scanner). During familiarization, participants studied the characters that would subsequently be used in the study. Each character appeared, one by one, at the center of the screen, within a circle with a tick mark at the top. Each character appeared for 4 secs, and then disappeared. The character was followed by a blank circle (with a tick mark) and participants were instructed to visualize the character they had just studied as accurately as possible, and then to press a button, which made the character they studied reappear. They were instructed to compare their image to the shape of the actual character and to correct any inaccuracies. Once they had done so, they pressed a button, and the next character appeared. Once participants had completed the familiarization phase, they were asked if they had any questions and were instructed that they would have the opportunity to study some of the characters again, and would be asked to practice the tasks that they would be performing inside the scanner. Once participants were ready, they completed a second familiarization phase where they studied the characters that would be used in the tasks that they were about to practice. This time, the participants studied the characters in a self-paced manner, for as little or as much time as needed. Practice (outside the scanner). Once participants had studied the characters, they were given instructions for the first task; the order of task practice was counterbalanced over participants. For each task, participants were instructed that a rectangular box containing some of the characters from the familiarization phase would appear on the screen, and that they should remember the box to perform the task that followed. For the spatial location memory task, they were told that they should pay attention to and remember the location of the characters within the box. For the spatial transformation task, they were told that they should pay attention to and remember the shape of the characters (i.e., "what the characters look like"). For both tasks, a rectangular box then appeared at the center of the screen. The box contained five alphanumeric characters, each placed within a circle with a tick mark indicating the top of the circle. The characters were placed in different locations within the box (see Figure 1 for examples of the study boxes). Participants studied the box with the characters for 30 sec. After the box disappeared, participants were told that the practice trials were about to begin. Spatial location task. On each trial, participants were shown a trisected circle stimulus (as previously described; see Figure 1 for a sample trial used when participants were inside the scanner). A smaller script version of the character was shown below the trisected circle. The script character cued the participants to mentally place the circle in the location where the character had appeared in the study box (for both tasks, a single study box was used for training; two boxes were used in the actual trials inside the scanner). The task was to decide whether the bold or dashed segment of the circle would be closer to the center of the display if the circle appeared in its original location. Thus, participants did not need to visualize the characters in order to perform the task; the only information required to perform the task was the location where the circle associated with the characters originally appeared. Spatial transformation task. For the spatial transformation task (see Figure 1
5 for a sample trial used when participants were inside the scanner), on each trial, a trisected circle appeared. Below the circle was a character in script font. The character cued participants to visualize the corresponding block character they had studied during familiarization. A tick mark was positioned on the contour of the circle, (not at the top, but rather at another location on the circle's circumference; see Figure 1). Participants were instructed to mentally rotate the visualized character until its top was aligned directly under the tick mark. After rotating, participants judged whether more of the character would be in the bold or dashed section of the circle; the segments were arranged so that this judgment itself was easy (i.e., the rate-limiting step was mental rotation itself). Experimental trials (inside the scanner). The experimental trials had the same format as the practice trials, except: 1) none of the characters used in practice trials were used in experimental trials; 2) the computer did not beep if participants made an error; 3) the stimulus for each trial remained on the screen for a varying interval (as explained below); 4) each task was administered twice and each session of each task was comprised of 20 trials; 5) participants were given two new study boxes (presented in sequence for 30 sec each) at the beginning of each scan (i.e., for every 20 trials). The order of the tasks was counterbalanced. Each task was performed once before either task was repeated. The interstimulus interval (ISI) varied from 6 to 14 sec in one-sec increments. ISIs were programmed according to a pseudorandom schedule and were varied to allow deconvolution of the hemodynamic response, and also so that participants were required to remain vigilant. fmri methods. Images were obtained with a 3-Tesla Siemens Allegra scanner and a standard head coil. Participants recorded responses with a fiber-optic button box. T1- weighted anatomic images were acquired using a multiplanar rapidly acquired gradient echo (MP-RAGE) sequence [TR = 25 ms, TE = 3.25 ms, flip angle = 40, field-of-view = 256x256 mm, acquisition matrix = 256x256, slices = 128, slice thickness = 1.33 mm, no gap 1x1x1.33 mm resolution]. Functional images were acquired using echoplanar imaging (EPI) [TR= 2000 ms, TE= 30 ms, flip angle = 90, field of view = 256x256, acquisition matrix = 64x64, slices = 35, slice thickness = 4 mm isotropic no gap, 110 timepoints for each run = 4 mm isotropic resolution]. Images were transformed to be made compatible with the Statistical Parametric Mapping program (SPM2, Wellcome Trust Centre for Neuroimaging, London, UK). Preprocessing included slice time correction, motion correction, and spatial normalization to Montreal Neurological Institute (MNI) coordinates. To maximize the spatial resolution of the results, data were not spatially smoothed. To model the hemodynamic response related to the processing of interest, events were modeled using the canonical hemodynamic basis function within SPM. Events were entered as vectors starting at the onset of each stimulus and ending at each participant's response. Only correct responses were considered (incorrect responses and trials where participants did not provide a response were not considered in the analyses). We used a random effect analysis to identify group activity associated with each contrast of interest (using a one-tailed t-test). We contrasted each of the two tasks with the other and also compared each task to baseline, which was defined as the interval between the point when a participant responded on a given trial and the presentation of the next stimulus. Because each stimulus remained on the screen after the participant responded (until a new stimulus was presented), the baseline condition had the same visual stimulation as each of the task conditions, but without the task-specific processing associated with spatial location memory or spatial transformation. Corrections for multiple comparisons. To correct for multiple comparisons, we conducted a Monte Carlo simulation using custom software written in MATLAB (The Mathworks, Natick, MA; Slotnick, 2008a). Because clusters of larger sizes are increasingly improbable, it is possible to determine the probability of a given spatial extent of activity (or larger) and then enforce an extent threshold to yield the desired type I error rate. Three dimensional spatial autocorrelation (full-width-half-maximum, FWHM) of the random effect contrast images was estimated to be 7.5 mm using custom software written in
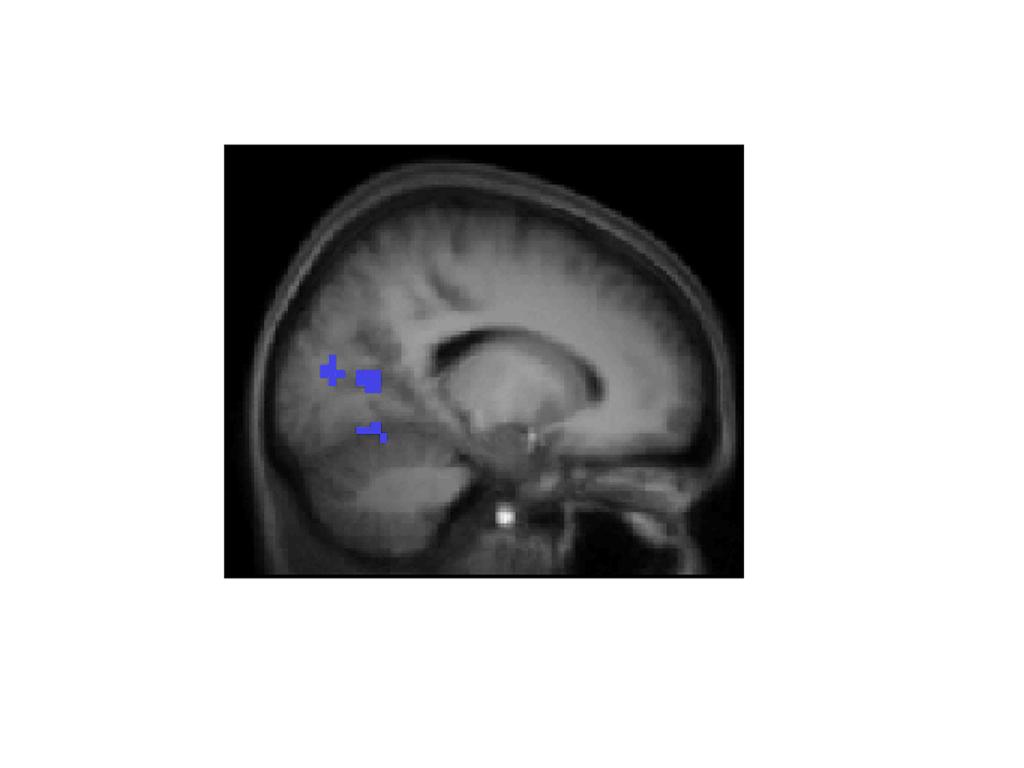
6 MATLAB to model smoothness in the data (Slotnick, 2008b). Based on 1,000 simulations it was determined that for an individual voxel threshold of p <.001, a cluster extent threshold of 15 contiguous voxels was necessary to correct for multiple comparisons to p <.05. Thus, only clusters of activation meeting or exceeding that size were considered as significantly activated. (For further details regarding cluster extent threshold correction for multiple comparisons, see Slotnick, 2008a; Slotnick & Schacter, 2006). RESULTS Behavioral results We analyzed response time (RT) and error data using analysis of variance (ANOVA) with a 2 (task; spatial location versus transformation) x 2 (replicate; first versus second) x 2 (gender; female versus male) design. The only effect to emerge from the RT analysis was a main effect for task. As shown in Table 1, RTs were longer for the transformation task than for the location task, t(15), P REP =.96. No significant effects were found in the analysis of errors. fmri results We contrasted each of the tasks against the other, to compare directly the activation associated specifically with spatial location memory and spatial transformation. Spatial location versus transformation. In the comparison of spatial location versus transformation (Table 2 and Figure 2, activation in blue), a peak of activation was found in the vicinity of the occipito-parietal sulcus, near the precuneus and posterior cingulate cortex. This region was also activated more during the task than during the baseline, indicating that the differences observed in this area between the two tasks were clearly a result of increases in activation in the spatial location task rather than deactivations in the transformation task. Activations in this region were bilateral. We also found greater activation in the location task than the transformation task in the medial lingual gyrus (BA 18), although this difference was in fact a result of deactivation in the transformation task compared to baseline, rather than an increase in the location task. Other portions of the lingual gyrus and the cuneus were also activated in the location task compared to the transformation task, although the differences between the two main conditions and the baseline were subthreshold, making these activations more difficult to interpret. (All of these regions had non-significant positive Z-scores for the location task and negative Z- scores for the transformation task, compared to baseline, which suggests a trend that may have proven significant with greater power). Transformation versus spatial location. When we examined activation in the transformation task versus that in the spatial location task (see Table 3 and Figure 2, activation in yellow), we found peak activation bilaterally in the superior parietal lobule (BA 7), and in the postcentral gyrus (BA 2/5/7). In addition, we found activation in the right inferior parietal cortex (BA 40). Some portions of the left superior parietal cortex and right inferior parietal cortex were not significantly activated more than during the baseline, and one region of the right inferior parietal cortex was also activated more in the spatial location task than in the baseline, which provides evidence that spatial transformation may rely partly on areas responsible for mapping spatial location. Contrary to the reverse comparison, we did not find activation in the precuneus, posterior cingulate or at the parietal/occipital junction. Instead, we documented activation in parietal regions near the junction of the superior and inferior lobules and extending into the postcentral gyrus. These areas have been more classically associated with mental rotation (see for example, Cohen et al., 1996, Kosslyn, DiGirolamo, Thompson & Alpert, 1998, Kosslyn, Thompson, Wraga, & Alpert, 2001) and are generally more superior and lateral than those associated with the spatial location task. Although we found no significant activation within 5 mm of midline, activations tended to be closer to the medial surface in the location task than in the transformation task. See Figure 3 for a medial view through a peak region of activation in the location task (identified by contrasting spatial location > spatial transformation). This region, near the right occipito-parietal junction, was activated bilaterally and to a greater degree in the location task than in either the transformation task or the baseline. As such, our data
7 suggest that it plays a specific role in the memory for object location. DISCUSSION Our results document a clear dissociation between spatial imagery that relies on transformational processes and spatial imagery that relies on memory for location. This result is important because it demonstrates that spatial imagery ability, like mental imagery more generally, is not a unitary function. This finding allows us to refine the conclusions of Kozhevnikov, Kosslyn and Shephard (2005), who demonstrated that visualizers should be divided into two types: those who prefer object imagery (i.e., imagery for shapes) and those who prefer spatial imagery. Kozhevnikov et al. (2005) showed that spatial imagers tend to be concentrated in certain professions and to interpret graphic representations differently from object imagers. However, Kozhevnikov et al. (2005) sort object and spatial imagers according to reported preference (rather than ability) and focus on mental manipulations and transformations. Perhaps more important, they treat spatial imagery as a single capacity. Clearly, spatial imagery should be divided into more finegrained capacities. The theory of Kosslyn (1994) posits that information about the locations and orientations of objects is organized into a single map, which is implemented primarily in the right parietal lobe. Consistent with this claim, we found more activation for the location task, relative to the transformation one, in cortex near the occipito-parietal junction -- with stronger activation in the right hemisphere. Moreover, the activation we found in the precuneus/posterior cingulate area for this comparison may reflect this region's role in directing information to processes that operate on this map. In contrast, the theory posits that spatial transformations occur when a process operating on this map in turn modifies the mapping function from inferotemporal areas (where visual memories are activated) to more posterior cortex; changing the mapping function results in changes in the location or orientation of the object in the image. Our findings suggest that portions of the parietal lobe near the junction of the superior and inferior lobules may play a crucial role here. Previous results (Cohen et al., 1996; Kosslyn et al., 1998) have also documented activation of motor and premotor regions during mental rotation (depending at least in part on the strategy being used). These findings are consistent with activation in postcentral gyrus observed here. In interpreting our results, it is important to note that not all of the activation found to be greater in one task than the other indicates that the activated regions implement the functions used to perform the task. In particular, portions of the lingual gyrus in Brodmann's Areas 18 and 19 were clearly more activated in the location task than in the transformation task but this was because of deactivation during the transformation task relative to the baseline (the activation in this region did not change in the location task relative to the baseline; see Table 2). Thus, we must caution that such differences in activation cannot be ascribed to the region's playing a role in the type of spatial imagery that underlies memory for location. Rather, it is possible that the transformation task (which our behavioral data indicate is more cognitively demanding) might require greater attention, and thus could require inhibiting regions where activation might interfere with task-specific processing. In this sense, a static picture of fmri results is inadequate to represent the dynamic, shifting nature of brain activations and deactivations. In addition to active inhibition of regions that may interfere with accomplishing a task, resources such as blood flow and blood volume might be redistributed away from less useful regions toward essential ones. Given that spatial and shape-based imagery rely on different general processes (e.g., Kozhevnikov, Kosslyn & Shephard, 2005) and the difficulty of the transformation task, resources may shift from object-based ventral stream visual areas (e.g., BA 18) toward dorsal regions critical for spatial transformations. The precuneus/posterior cingulate region, more activated in the location task than the transformation task, has also been associated with the "Default Mode Network" (see Mason, Norton, Van Horn, Wegner, Grafton, & Macrae, 2007). However, it is unlikely that the activation in this region reflects "default" brain activation during a less demanding task: this region was more strongly activated bilaterally in the location task than in the
8 baseline, which suggests that the activation was a result of the region's playing an active role in task-specific processing for location memory. Other regions of the cuneus and lingual gyrus, which are statistically unchanged from baseline, may reflect non task-related processing (or stimulus-independent thought, Mason et al., 2007); however, the lack of statistical difference from baseline may also reflect a lack of power. Although we cannot interpret all the differences between areas identified in the direct comparison of the two conditions, the data reveal a clear dissociation between two types of spatial imagery, with a small, distinct set of areas specific to each. These results support the claim that some processes map spatial locations and others transform spatial relations representations (cf. Zacks & Michelon, 2005). If we affirm that spatial imagery ability (rather than simply "imagery ability") is useful in learning geometry or anatomy, in navigating an environment, in learning surgical techniques, then it is also important to know the particular combination of spatial imagery ability (or abilities) that come into play in each circumstance. This is essential in investigating the parameters that define the training and transfer of skill in a particular domain, and ultimately, in designing training programs to fit a specific set of skills (cf., Wright et al., 2008). Progress has been made in identifying subcomponents of mental imagery (see Kosslyn, Thompson & Ganis, 2006), including those functions that underlie processing shape and those that underlie processing spatial relations (see, for example, Farah, Hammond, Levine, & Calvanio, 1988; Kozhevnikov et al., 2002; Kosslyn, Ganis, & Thompson, 2001). The present study was designed to further our understanding of spatial mental imagery by decomposing this construct into two component parts. Our results support the view that at least two different, broad types of spatial imagery exist. Authors' note This material is based upon work supported by the National Science Foundation under grant REC We thank Rebecca Wright for assistance in testing the participants. Any opinions, findings, and conclusions or recommendations expressed in this material are those of the authors and do not necessarily reflect the views of the National Science Foundation. References Cohen, M.S., Kosslyn, S.M., Breiter, H.C., DiGirolamo, G. J., Thompson, W. L., Bookheimer, S. Y., Belliveau, J.W., & Rosen, B.R. (1996). Changes in cortical activity during mental rotation: a mapping study using functional MRI. Brain, 119, Dror, I. E., Kosslyn, S. M., & Waag, W. (1993). Visual-spatial abilities of pilots. Journal of Applied Psychology, 78 (5), Farah, M. J., Hammond, K. M., Levine, D. N., & Calvanio, R. (1988). Visual and spatial mental imagery: Dissociable systems of representations. Cognitive Psychology, 20, Guillot, A., Champely, S., Batier, C., Thiriet, P., & Collet, C. (2007). Relationship between spatial abilities, mental rotation and functional anatomy learning. Advances in Health Sciences Education, 12, Kosslyn, S. M. (2006). You can play 20 questions with nature and win: Categorical versus coordinate spatial relations as a case study. Neuropsychologia, 44, Kosslyn, S. M., DiGirolamo, G., Thompson, W. L., & Alpert, N.M. (1998). Mental rotation of objects versus hands: Neural mechanisms revealed by positron emission tomography. Psychophysiology, 35, Kosslyn, S. M., Ganis, G., & Thompson, W. L. (2001). Neural foundations of imagery. Nature Reviews Neuroscience, 2,
9 Replicate First Second Kosslyn, S. M., Thompson, W. L., and Ganis, G. (2006). The case for mental imagery. New York: Oxford University Press. Kosslyn, S. M., Thompson, W. L., Wraga, M., and Alpert, N. M. (2001). Imagining rotation by endogenous and exogenous forces: Distinct neural mechanisms for different strategies. Neuroreport, 12, Kozhevnikov, M., Hegarty, M. & Mayer, R. E. (2002). Revising the visualizer/verbalizer dimension: Evidence for two types of visualizers. Cognition and Instruction, 20, Kozhevinikov, M., Kosslyn, S., & Shephard, J. (2005). Spatial versus object visualizers: A new characterization of cognitive style. Memory & Cognition, 33 (4), Mason, M. F., Norton, M. I., Van Horn, J. D., Wegner, D. M., Grafton, S. T., & Macrae, C. N. (2007). Wandering minds: The default network and stimulus-independent thought. Science, 315 (5810), Moscovitch, C., Kapur, S., Köhler, S., & Houle, S. (1995). Distinct neural correlates of visual long-term memory for spatial location and object identity: a positron emission tomography study in humans. Proceedings of the National Academy of Sciences of the United States of America, 92 (9), Slotnick, S. D. (2008a). Cluster threshold beta. Retrieved March 24, 2008, from Boston College Web site: Slotnick, S. D. (2008b). Img xcorr. Retrieved October 2, 2008, from Boston College Web site: Slotnick, S. D., & Schacter, D.L. (2006) The nature of memory related activity in early visual areas. Neuropsychologia, 44 (14), van Asselen, M., Kessels, R. P., Neggers, S. F., Kappelle, L. J., Frijins, C. J., Postma, A. (2006). Brain areas involved in spatial working memory. Neuropsychologia, 44 (7), Wanzel, K. R., Hamstra, S. J., Anastakis, D. J., Matsumoto, E. D., & Cusimano, M. D. (2002). Effect of visual-spatial ability on learning of spatially-complex surgical skills. Lancet, 359 (9302), Wright, R., Thompson, W. L., Ganis, G., Newcombe, N., & Kosslyn, S. M. (2008). Training generalized spatial skills. Psychonomic Bulletin and Review, 15, Zacks, J. (2008). Neuroimaging studies of mental rotation: A meta-analysis and review. Journal of Cognitive Neuroscience, 20 (1), Zacks, J., & Michelon, P. (2005). Transformations of visuospatial images. Behavioral and Cognitive Neuroscience Reviews, 4 (2), Table 1. Response time (RT) and error (%) means for each task and replicate. Standard errors of the mean (SEM) are also presented.
10 Replicate First Mean (SEM) Second Mean (SEM) RT (milliseconds) Task Spatial Location 3984 (173) 3962 (115) Transformation 4419 (171) 4432 (177) Error (%) Task Spatial Location 22.2 (3.5) 25.9 (3.0) Transformation 26.6 (2.2) 27.8 (2.5) Table 2. Results of contrasting the spatial location task versus the spatial transformation task. Region (BA) (Cluster #) Occ.-par. sulcus/ Preuneus/ Post. cing. (BA 18/30/31) (Cl. 1) Occ.-par. sulcus/preuneus/post. cing. (BA 31) (Cl. 2) Lingual gyrus (BA 18/19) (Cl. 1) Lingual gyrus (BA 18) (Cl. 3) Cuneus (BA 18/19) (Cl. 1) Cuneus (BA 23/31) (Cl. 3) Lingual gyrus (BA 18) (Cl. 3) MNI Coordinates Z-Score (L- T) Z-Score (L- B) * 3.94* * 3.23* * * * * * * Z-Score (T- B) Locations of the foci (peak voxels) of activation are given in Montreal Neurological Institute (MNI) coordinates. Occ.-par. sulcus = Occipito-parietal sulcus; Post. cing. = Posterior cingulate. Numbers in parentheses below the activated region label represent the corresponding Brodmann Area (BA), and to which of the three clusters (Cl.), as identified by SPM, each locus of activation belongs. Cluster sizes were: Cluster 1 = 229 voxels; cluster 2 = 16 voxels; cluster 3 = 111 voxels. For each activation, Z-scores are provided for
11 the main contrast between the two tasks, spatial location versus spatial transformation (L-T) and for each task when it is contrasted directly with the baseline, spatial location versus baseline (L-B) and spatial transformation versus baseline (T-B). Z-scores are provided for each peak focus of activation identified within a cluster. Significant Z-score effects are denoted with an asterisk (*). Table 3. Results of contrasting the spatial transformation task versus the spatial location task. Region (BA) Inferior parietal lobule/postcentral gyrus (40/2) (1) Superior parietal cortex (7) (2) Superior parietal cortex (7) (2) Postcentral gyrus (5) (3) Postcentral gyrus (5) (2) Postcentral gyrus/ Superior parietal cortex (5/7) (3) Inferior parietal cortex (40) (1) Inferior parietal cortex (40) (1) Superior parietal cortex (40/7) (3) MNI Coordinates Z-Score (T-L) Z-Score (T-B) * 4.24* * 3.37* * 3.90* * 4.37* * 3.25* * 3.28* 2.50 Z-Score (L-B) * 4.43* 4.09* * * Locations of the foci (peak voxels) of activation are given in Montreal Neurological Institute (MNI) coordinates. Numbers in parentheses below the activated region label represent the corresponding Brodmann Area (BA), and to which of the three clusters (Cl.), as identified by SPM, each locus of activation belongs. Cluster sizes were: Cluster 1 = 23 voxels; cluster 2 = 25 voxels; cluster 3 = 20 voxels. For each activation, Z-scores are provided for the main contrast between the two tasks, spatial transformation versus spatial location (T-L) and for each task when it is contrasted directly with the baseline, spatial transformation versus baseline (T-B) and spatial location versus baseline (L-B). Z-scores are provided for each peak focus of activation identified within a cluster. Significant Z- score effects are denoted with an asterisk (*). Figure legends. Figure 1. Example of a trial in the spatial location task and spatial transformation task. In both tasks, participants first studied the characters in two boxes that were each presented sequentially for 30 seconds (top). For the spatial location task, on each trial, participants saw a trisected circle with a vertical tick mark at the top and a script character beneath the circle (middle, left). The script character cued them to remember the location of the corresponding block character that they had studied. They thus visualized the trisected circle in the location within the box where the appropriate block character had appeared
12 (bottom, left). After having mentally placed the trisected circle in the appropriate location within the box, participants then decided whether the bold or dashed section of the circle was closer to the center point of the box. Here, the correct response would have been "dashed". For the transformation task, participants saw a trisected circle with a tick mark displaced from the vertical and script character beneath the circle (middle, right). The script character served to cue the block character to be visualized. Participants visualized the character in the circle, and rotated it to align with the tick mark, in order to decide which section of the circle (bold or dashed) would contain more of the character when it was rotated (bottom, right). Here, the correct response would have been "bold". Figure 2. Differential activity associated with the spatial location task (blue) and the spatial transformation task (yellow) when the two tasks were contrasted with each other (at the top, anterior, to the left, and posterior, to the right, views; in the middle, right, to the left, and left, to the right, lateral views; at the bottom, inferior, to the left, and superior, to the right, views). Note that some apparent activations do not reflect increases versus the baseline and may be due to deactivations in the opposite task (see text and Tables 2 and 3 for details). Figure 3. Medial sagittal view of activity near the right occipito-parietal junction (first focus of activation listed in Table 2), associated with the spatial location task versus the transformation task (activation in this region was also elevated relative to the baseline). More inferiorly, activation in lingual gyrus can be seen (although the activity in this region was not significantly elevated relative to the baseline).
13
14
Two Forms of Spatial Imagery
 PSYCHOLOGICAL SCIENCE Research Article Two Forms of Spatial Imagery Neuroimaging Evidence William L. Thompson, 1 Scott D. Slotnick, 2 Marie S. Burrage, 1 and Stephen M. Kosslyn 1,3 1 Harvard University,
PSYCHOLOGICAL SCIENCE Research Article Two Forms of Spatial Imagery Neuroimaging Evidence William L. Thompson, 1 Scott D. Slotnick, 2 Marie S. Burrage, 1 and Stephen M. Kosslyn 1,3 1 Harvard University,
WHAT DOES THE BRAIN TELL US ABOUT TRUST AND DISTRUST? EVIDENCE FROM A FUNCTIONAL NEUROIMAGING STUDY 1
 SPECIAL ISSUE WHAT DOES THE BRAIN TE US ABOUT AND DIS? EVIDENCE FROM A FUNCTIONAL NEUROIMAGING STUDY 1 By: Angelika Dimoka Fox School of Business Temple University 1801 Liacouras Walk Philadelphia, PA
SPECIAL ISSUE WHAT DOES THE BRAIN TE US ABOUT AND DIS? EVIDENCE FROM A FUNCTIONAL NEUROIMAGING STUDY 1 By: Angelika Dimoka Fox School of Business Temple University 1801 Liacouras Walk Philadelphia, PA
Supporting Online Material for
 www.sciencemag.org/cgi/content/full/324/5927/646/dc1 Supporting Online Material for Self-Control in Decision-Making Involves Modulation of the vmpfc Valuation System Todd A. Hare,* Colin F. Camerer, Antonio
www.sciencemag.org/cgi/content/full/324/5927/646/dc1 Supporting Online Material for Self-Control in Decision-Making Involves Modulation of the vmpfc Valuation System Todd A. Hare,* Colin F. Camerer, Antonio
Cognitive Neuroscience Publication details, including instructions for authors and subscription information:
 This article was downloaded by: [Boston College] On: 07 December 2011, At: 09:48 Publisher: Psychology Press Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer
This article was downloaded by: [Boston College] On: 07 December 2011, At: 09:48 Publisher: Psychology Press Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer
Supplementary Information
 Supplementary Information The neural correlates of subjective value during intertemporal choice Joseph W. Kable and Paul W. Glimcher a 10 0 b 10 0 10 1 10 1 Discount rate k 10 2 Discount rate k 10 2 10
Supplementary Information The neural correlates of subjective value during intertemporal choice Joseph W. Kable and Paul W. Glimcher a 10 0 b 10 0 10 1 10 1 Discount rate k 10 2 Discount rate k 10 2 10
Supplementary information Detailed Materials and Methods
 Supplementary information Detailed Materials and Methods Subjects The experiment included twelve subjects: ten sighted subjects and two blind. Five of the ten sighted subjects were expert users of a visual-to-auditory
Supplementary information Detailed Materials and Methods Subjects The experiment included twelve subjects: ten sighted subjects and two blind. Five of the ten sighted subjects were expert users of a visual-to-auditory
HST.583 Functional Magnetic Resonance Imaging: Data Acquisition and Analysis Fall 2008
 MIT OpenCourseWare http://ocw.mit.edu HST.583 Functional Magnetic Resonance Imaging: Data Acquisition and Analysis Fall 2008 For information about citing these materials or our Terms of Use, visit: http://ocw.mit.edu/terms.
MIT OpenCourseWare http://ocw.mit.edu HST.583 Functional Magnetic Resonance Imaging: Data Acquisition and Analysis Fall 2008 For information about citing these materials or our Terms of Use, visit: http://ocw.mit.edu/terms.
Supporting online material. Materials and Methods. We scanned participants in two groups of 12 each. Group 1 was composed largely of
 Placebo effects in fmri Supporting online material 1 Supporting online material Materials and Methods Study 1 Procedure and behavioral data We scanned participants in two groups of 12 each. Group 1 was
Placebo effects in fmri Supporting online material 1 Supporting online material Materials and Methods Study 1 Procedure and behavioral data We scanned participants in two groups of 12 each. Group 1 was
Procedia - Social and Behavioral Sciences 159 ( 2014 ) WCPCG 2014
 WCPCG 2014") Available online at www.sciencedirect.com ScienceDirect Procedia - Social and Behavioral Sciences 159 ( 2014 ) 743 748 WCPCG 2014 Differences in Visuospatial Cognition Performance and Regional Brain Activation
Available online at www.sciencedirect.com ScienceDirect Procedia - Social and Behavioral Sciences 159 ( 2014 ) 743 748 WCPCG 2014 Differences in Visuospatial Cognition Performance and Regional Brain Activation
Neural correlates of memory for object identity and object location: effects of aging
 Neuropsychologia 40 (2002) 1428 1442 Neural correlates of memory for object identity and object location: effects of aging Alessandra Schiavetto a, Stefan Köhler a, Cheryl L. Grady a, Gordon Winocur a,c,
Neuropsychologia 40 (2002) 1428 1442 Neural correlates of memory for object identity and object location: effects of aging Alessandra Schiavetto a, Stefan Köhler a, Cheryl L. Grady a, Gordon Winocur a,c,
Supplementary Information Methods Subjects The study was comprised of 84 chronic pain patients with either chronic back pain (CBP) or osteoarthritis
 or osteoarthritis") Supplementary Information Methods Subjects The study was comprised of 84 chronic pain patients with either chronic back pain (CBP) or osteoarthritis (OA). All subjects provided informed consent to procedures
Supplementary Information Methods Subjects The study was comprised of 84 chronic pain patients with either chronic back pain (CBP) or osteoarthritis (OA). All subjects provided informed consent to procedures
Twelve right-handed subjects between the ages of 22 and 30 were recruited from the
 Supplementary Methods Materials & Methods Subjects Twelve right-handed subjects between the ages of 22 and 30 were recruited from the Dartmouth community. All subjects were native speakers of English,
Supplementary Methods Materials & Methods Subjects Twelve right-handed subjects between the ages of 22 and 30 were recruited from the Dartmouth community. All subjects were native speakers of English,
Personal Space Regulation by the Human Amygdala. California Institute of Technology
 Personal Space Regulation by the Human Amygdala Daniel P. Kennedy 1, Jan Gläscher 1, J. Michael Tyszka 2 & Ralph Adolphs 1,2 1 Division of Humanities and Social Sciences 2 Division of Biology California
Personal Space Regulation by the Human Amygdala Daniel P. Kennedy 1, Jan Gläscher 1, J. Michael Tyszka 2 & Ralph Adolphs 1,2 1 Division of Humanities and Social Sciences 2 Division of Biology California
HST.583 Functional Magnetic Resonance Imaging: Data Acquisition and Analysis Fall 2006
 MIT OpenCourseWare http://ocw.mit.edu HST.583 Functional Magnetic Resonance Imaging: Data Acquisition and Analysis Fall 2006 For information about citing these materials or our Terms of Use, visit: http://ocw.mit.edu/terms.
MIT OpenCourseWare http://ocw.mit.edu HST.583 Functional Magnetic Resonance Imaging: Data Acquisition and Analysis Fall 2006 For information about citing these materials or our Terms of Use, visit: http://ocw.mit.edu/terms.
SUPPLEMENT: DYNAMIC FUNCTIONAL CONNECTIVITY IN DEPRESSION. Supplemental Information. Dynamic Resting-State Functional Connectivity in Major Depression
 Supplemental Information Dynamic Resting-State Functional Connectivity in Major Depression Roselinde H. Kaiser, Ph.D., Susan Whitfield-Gabrieli, Ph.D., Daniel G. Dillon, Ph.D., Franziska Goer, B.S., Miranda
Supplemental Information Dynamic Resting-State Functional Connectivity in Major Depression Roselinde H. Kaiser, Ph.D., Susan Whitfield-Gabrieli, Ph.D., Daniel G. Dillon, Ph.D., Franziska Goer, B.S., Miranda
GENDER-SPECIFIC SENSITVITY TO TIME-DISCREPANT TASK CONDITIONS OF REASONING DURING fmri
 GENDER-SPECIFIC SENSITVITY TO TIME-DISCREPANT TASK CONDITIONS OF REASONING DURING fmri by Joshua M. Roberts A Thesis Submitted to the Graduate Faculty of George Mason University in Partial Fulfillment
GENDER-SPECIFIC SENSITVITY TO TIME-DISCREPANT TASK CONDITIONS OF REASONING DURING fmri by Joshua M. Roberts A Thesis Submitted to the Graduate Faculty of George Mason University in Partial Fulfillment
Functional topography of a distributed neural system for spatial and nonspatial information maintenance in working memory
 Neuropsychologia 41 (2003) 341 356 Functional topography of a distributed neural system for spatial and nonspatial information maintenance in working memory Joseph B. Sala a,, Pia Rämä a,c,d, Susan M.
Neuropsychologia 41 (2003) 341 356 Functional topography of a distributed neural system for spatial and nonspatial information maintenance in working memory Joseph B. Sala a,, Pia Rämä a,c,d, Susan M.
The nature of memory related activity in early visual areas
 Neuropsychologia 44 (2006) 2874 2886 The nature of memory related activity in early visual areas Scott D. Slotnick a,, Daniel L. Schacter b a Department of Psychology, Boston College, Chestnut Hill, MA
Neuropsychologia 44 (2006) 2874 2886 The nature of memory related activity in early visual areas Scott D. Slotnick a,, Daniel L. Schacter b a Department of Psychology, Boston College, Chestnut Hill, MA
Remembering the Past to Imagine the Future: A Cognitive Neuroscience Perspective
 MILITARY PSYCHOLOGY, 21:(Suppl. 1)S108 S112, 2009 Copyright Taylor & Francis Group, LLC ISSN: 0899-5605 print / 1532-7876 online DOI: 10.1080/08995600802554748 Remembering the Past to Imagine the Future:
MILITARY PSYCHOLOGY, 21:(Suppl. 1)S108 S112, 2009 Copyright Taylor & Francis Group, LLC ISSN: 0899-5605 print / 1532-7876 online DOI: 10.1080/08995600802554748 Remembering the Past to Imagine the Future:
Resistance to forgetting associated with hippocampus-mediated. reactivation during new learning
 Resistance to Forgetting 1 Resistance to forgetting associated with hippocampus-mediated reactivation during new learning Brice A. Kuhl, Arpeet T. Shah, Sarah DuBrow, & Anthony D. Wagner Resistance to
Resistance to Forgetting 1 Resistance to forgetting associated with hippocampus-mediated reactivation during new learning Brice A. Kuhl, Arpeet T. Shah, Sarah DuBrow, & Anthony D. Wagner Resistance to
How do individuals with congenital blindness form a conscious representation of a world they have never seen? brain. deprived of sight?
 How do individuals with congenital blindness form a conscious representation of a world they have never seen? What happens to visual-devoted brain structure in individuals who are born deprived of sight?
How do individuals with congenital blindness form a conscious representation of a world they have never seen? What happens to visual-devoted brain structure in individuals who are born deprived of sight?
Frank Tong. Department of Psychology Green Hall Princeton University Princeton, NJ 08544
 Frank Tong Department of Psychology Green Hall Princeton University Princeton, NJ 08544 Office: Room 3-N-2B Telephone: 609-258-2652 Fax: 609-258-1113 Email: ftong@princeton.edu Graduate School Applicants
Frank Tong Department of Psychology Green Hall Princeton University Princeton, NJ 08544 Office: Room 3-N-2B Telephone: 609-258-2652 Fax: 609-258-1113 Email: ftong@princeton.edu Graduate School Applicants
Supporting Information
 Supporting Information Newman et al. 10.1073/pnas.1510527112 SI Results Behavioral Performance. Behavioral data and analyses are reported in the main article. Plots of the accuracy and reaction time data
Supporting Information Newman et al. 10.1073/pnas.1510527112 SI Results Behavioral Performance. Behavioral data and analyses are reported in the main article. Plots of the accuracy and reaction time data
Comparing event-related and epoch analysis in blocked design fmri
 Available online at www.sciencedirect.com R NeuroImage 18 (2003) 806 810 www.elsevier.com/locate/ynimg Technical Note Comparing event-related and epoch analysis in blocked design fmri Andrea Mechelli,
Available online at www.sciencedirect.com R NeuroImage 18 (2003) 806 810 www.elsevier.com/locate/ynimg Technical Note Comparing event-related and epoch analysis in blocked design fmri Andrea Mechelli,
Fear Selectively Modulates Visual Mental Imagery and Visual Perception
 Fear Selectively Modulates Visual Mental Imagery and Visual Perception The Harvard community has made this article openly available. Please share how this access benefits you. Your story matters. Citation
Fear Selectively Modulates Visual Mental Imagery and Visual Perception The Harvard community has made this article openly available. Please share how this access benefits you. Your story matters. Citation
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature11239 Introduction The first Supplementary Figure shows additional regions of fmri activation evoked by the task. The second, sixth, and eighth shows an alternative way of analyzing reaction
doi:10.1038/nature11239 Introduction The first Supplementary Figure shows additional regions of fmri activation evoked by the task. The second, sixth, and eighth shows an alternative way of analyzing reaction
Behavioural Brain Research
 Behavioural Brain Research 197 (2009) 186 197 Contents lists available at ScienceDirect Behavioural Brain Research j o u r n a l h o m e p a g e : www.elsevier.com/locate/bbr Research report Top-down attentional
Behavioural Brain Research 197 (2009) 186 197 Contents lists available at ScienceDirect Behavioural Brain Research j o u r n a l h o m e p a g e : www.elsevier.com/locate/bbr Research report Top-down attentional
Reasoning and working memory: common and distinct neuronal processes
 Neuropsychologia 41 (2003) 1241 1253 Reasoning and working memory: common and distinct neuronal processes Christian C. Ruff a,b,, Markus Knauff a,c, Thomas Fangmeier a, Joachim Spreer d a Centre for Cognitive
Neuropsychologia 41 (2003) 1241 1253 Reasoning and working memory: common and distinct neuronal processes Christian C. Ruff a,b,, Markus Knauff a,c, Thomas Fangmeier a, Joachim Spreer d a Centre for Cognitive
Human Paleoneurology and the Evolution of the Parietal Cortex
 PARIETAL LOBE The Parietal Lobes develop at about the age of 5 years. They function to give the individual perspective and to help them understand space, touch, and volume. The location of the parietal
PARIETAL LOBE The Parietal Lobes develop at about the age of 5 years. They function to give the individual perspective and to help them understand space, touch, and volume. The location of the parietal
Functional MRI Mapping Cognition
 Outline Functional MRI Mapping Cognition Michael A. Yassa, B.A. Division of Psychiatric Neuro-imaging Psychiatry and Behavioral Sciences Johns Hopkins School of Medicine Why fmri? fmri - How it works Research
Outline Functional MRI Mapping Cognition Michael A. Yassa, B.A. Division of Psychiatric Neuro-imaging Psychiatry and Behavioral Sciences Johns Hopkins School of Medicine Why fmri? fmri - How it works Research
Neural correlates of two imagined egocentric transformations
 www.elsevier.com/locate/ynimg NeuroImage 35 (2007) 916 927 Neural correlates of two imagined egocentric transformations Sarah H. Creem-Regehr, Jayson A. Neil, and Hsiang J. Yeh Department of Psychology,
www.elsevier.com/locate/ynimg NeuroImage 35 (2007) 916 927 Neural correlates of two imagined egocentric transformations Sarah H. Creem-Regehr, Jayson A. Neil, and Hsiang J. Yeh Department of Psychology,
Classification and Statistical Analysis of Auditory FMRI Data Using Linear Discriminative Analysis and Quadratic Discriminative Analysis
 International Journal of Innovative Research in Computer Science & Technology (IJIRCST) ISSN: 2347-5552, Volume-2, Issue-6, November-2014 Classification and Statistical Analysis of Auditory FMRI Data Using
International Journal of Innovative Research in Computer Science & Technology (IJIRCST) ISSN: 2347-5552, Volume-2, Issue-6, November-2014 Classification and Statistical Analysis of Auditory FMRI Data Using
Methods to examine brain activity associated with emotional states and traits
 Methods to examine brain activity associated with emotional states and traits Brain electrical activity methods description and explanation of method state effects trait effects Positron emission tomography
Methods to examine brain activity associated with emotional states and traits Brain electrical activity methods description and explanation of method state effects trait effects Positron emission tomography
Supporting Information
 Supporting Information Braver et al. 10.1073/pnas.0808187106 SI Methods Participants. Participants were neurologically normal, righthanded younger or older adults. The groups did not differ in gender breakdown
Supporting Information Braver et al. 10.1073/pnas.0808187106 SI Methods Participants. Participants were neurologically normal, righthanded younger or older adults. The groups did not differ in gender breakdown
VIII. 10. Right Temporal-Lobe Contribution to the Retrieval of Family Relationships in Person Identification
 CYRIC Annual Report 2009 VIII. 10. Right Temporal-Lobe Contribution to the Retrieval of Family Relationships in Person Identification Abe N. 1, Fujii T. 1, Ueno A. 1, Shigemune Y. 1, Suzuki M. 2, Tashiro
CYRIC Annual Report 2009 VIII. 10. Right Temporal-Lobe Contribution to the Retrieval of Family Relationships in Person Identification Abe N. 1, Fujii T. 1, Ueno A. 1, Shigemune Y. 1, Suzuki M. 2, Tashiro
Common Neural Substrates for Ordinal Representation in Short-Term Memory, Numerical and Alphabetical Cognition
 Common Neural Substrates for Ordinal Representation in Short-Term Memory, Numerical and Alphabetical Cognition Lucie Attout 1 *, Wim Fias 2, Eric Salmon 3, Steve Majerus 1 1 Department of Psychology -
Common Neural Substrates for Ordinal Representation in Short-Term Memory, Numerical and Alphabetical Cognition Lucie Attout 1 *, Wim Fias 2, Eric Salmon 3, Steve Majerus 1 1 Department of Psychology -
Investigating directed influences between activated brain areas in a motor-response task using fmri
 Magnetic Resonance Imaging 24 (2006) 181 185 Investigating directed influences between activated brain areas in a motor-response task using fmri Birgit Abler a, 4, Alard Roebroeck b, Rainer Goebel b, Anett
Magnetic Resonance Imaging 24 (2006) 181 185 Investigating directed influences between activated brain areas in a motor-response task using fmri Birgit Abler a, 4, Alard Roebroeck b, Rainer Goebel b, Anett
Hallucinations and conscious access to visual inputs in Parkinson s disease
 Supplemental informations Hallucinations and conscious access to visual inputs in Parkinson s disease Stéphanie Lefebvre, PhD^1,2, Guillaume Baille, MD^4, Renaud Jardri MD, PhD 1,2 Lucie Plomhause, PhD
Supplemental informations Hallucinations and conscious access to visual inputs in Parkinson s disease Stéphanie Lefebvre, PhD^1,2, Guillaume Baille, MD^4, Renaud Jardri MD, PhD 1,2 Lucie Plomhause, PhD
Hippocampal brain-network coordination during volitionally controlled exploratory behavior enhances learning
 Online supplementary information for: Hippocampal brain-network coordination during volitionally controlled exploratory behavior enhances learning Joel L. Voss, Brian D. Gonsalves, Kara D. Federmeier,
Online supplementary information for: Hippocampal brain-network coordination during volitionally controlled exploratory behavior enhances learning Joel L. Voss, Brian D. Gonsalves, Kara D. Federmeier,
QUANTIFYING CEREBRAL CONTRIBUTIONS TO PAIN 1
 QUANTIFYING CEREBRAL CONTRIBUTIONS TO PAIN 1 Supplementary Figure 1. Overview of the SIIPS1 development. The development of the SIIPS1 consisted of individual- and group-level analysis steps. 1) Individual-person
QUANTIFYING CEREBRAL CONTRIBUTIONS TO PAIN 1 Supplementary Figure 1. Overview of the SIIPS1 development. The development of the SIIPS1 consisted of individual- and group-level analysis steps. 1) Individual-person
Mental imagery: Against the Nihilistic Hypothesis. Giorgio Ganis a,c William L. Thompson a. Harvard University, Department of Psychology
 Kosslyn et al., p. 1 Mental imagery: Against the Nihilistic Hypothesis Stephen M. Kosslyn a,b Giorgio Ganis a,c William L. Thompson a a Harvard University, Department of Psychology b Massachusetts General
Kosslyn et al., p. 1 Mental imagery: Against the Nihilistic Hypothesis Stephen M. Kosslyn a,b Giorgio Ganis a,c William L. Thompson a a Harvard University, Department of Psychology b Massachusetts General
Topographical functional connectivity patterns exist in the congenitally, prelingually deaf
 Supplementary Material Topographical functional connectivity patterns exist in the congenitally, prelingually deaf Ella Striem-Amit 1*, Jorge Almeida 2,3, Mario Belledonne 1, Quanjing Chen 4, Yuxing Fang
Supplementary Material Topographical functional connectivity patterns exist in the congenitally, prelingually deaf Ella Striem-Amit 1*, Jorge Almeida 2,3, Mario Belledonne 1, Quanjing Chen 4, Yuxing Fang
FRONTAL LOBE. Central Sulcus. Ascending ramus of the Cingulate Sulcus. Cingulate Sulcus. Lateral Sulcus
 FRONTAL LOBE Central Ascending ramus of the Cingulate Cingulate Lateral Lateral View Medial View Motor execution and higher cognitive functions (e.g., language production, impulse inhibition, reasoning
FRONTAL LOBE Central Ascending ramus of the Cingulate Cingulate Lateral Lateral View Medial View Motor execution and higher cognitive functions (e.g., language production, impulse inhibition, reasoning
Supplemental Information. Direct Electrical Stimulation in the Human Brain. Disrupts Melody Processing
 Current Biology, Volume 27 Supplemental Information Direct Electrical Stimulation in the Human Brain Disrupts Melody Processing Frank E. Garcea, Benjamin L. Chernoff, Bram Diamond, Wesley Lewis, Maxwell
Current Biology, Volume 27 Supplemental Information Direct Electrical Stimulation in the Human Brain Disrupts Melody Processing Frank E. Garcea, Benjamin L. Chernoff, Bram Diamond, Wesley Lewis, Maxwell
Theory of mind skills are related to gray matter volume in the ventromedial prefrontal cortex in schizophrenia
 Theory of mind skills are related to gray matter volume in the ventromedial prefrontal cortex in schizophrenia Supplemental Information Table of Contents 2 Behavioral Data 2 Table S1. Participant demographics
Theory of mind skills are related to gray matter volume in the ventromedial prefrontal cortex in schizophrenia Supplemental Information Table of Contents 2 Behavioral Data 2 Table S1. Participant demographics
FINAL PROGRESS REPORT
 (1) Foreword (optional) (2) Table of Contents (if report is more than 10 pages) (3) List of Appendixes, Illustrations and Tables (if applicable) (4) Statement of the problem studied FINAL PROGRESS REPORT
(1) Foreword (optional) (2) Table of Contents (if report is more than 10 pages) (3) List of Appendixes, Illustrations and Tables (if applicable) (4) Statement of the problem studied FINAL PROGRESS REPORT
The Cognitive Control of Memory: Age Differences in the Neural Correlates of Successful Remembering and Intentional Forgetting
 The Cognitive Control of Memory: Age Differences in the Neural Correlates of Successful Remembering and Intentional Forgetting Avery A. Rizio, Nancy A. Dennis* The Pennsylvania State University, Department
The Cognitive Control of Memory: Age Differences in the Neural Correlates of Successful Remembering and Intentional Forgetting Avery A. Rizio, Nancy A. Dennis* The Pennsylvania State University, Department
Brain activity related to integrative processes in visual object recognition: bottom-up integration and the modulatory influence of stored knowledge
 Neuropsychologia 40 (2002) 1254 1267 Brain activity related to integrative processes in visual object recognition: bottom-up integration and the modulatory influence of stored knowledge C. Gerlach a,,
Neuropsychologia 40 (2002) 1254 1267 Brain activity related to integrative processes in visual object recognition: bottom-up integration and the modulatory influence of stored knowledge C. Gerlach a,,
The hippocampus operates in a threshold manner during spatial source memory Scott D. Slotnick a and Preston P. Thakral b
 Cognitive neuroscience and neuropsychology 265 The hippocampus operates in a threshold manner during spatial source memory Scott D. Slotnick a and Preston P. Thakral b Long-term memory can be based on
Cognitive neuroscience and neuropsychology 265 The hippocampus operates in a threshold manner during spatial source memory Scott D. Slotnick a and Preston P. Thakral b Long-term memory can be based on
The neural correlates of conceptual and perceptual false recognition
 The neural correlates of conceptual and perceptual false recognition Rachel J. Garoff-Eaton, Elizabeth A. Kensinger and Daniel L. Schacter Learn. Mem. 2007 14: 684-692 Access the most recent version at
The neural correlates of conceptual and perceptual false recognition Rachel J. Garoff-Eaton, Elizabeth A. Kensinger and Daniel L. Schacter Learn. Mem. 2007 14: 684-692 Access the most recent version at
Neural Correlates of Ongoing Conscious Experience: Both Task-Unrelatedness and Stimulus-Independence Are Related to Default Network Activity
 : Both Task-Unrelatedness and Stimulus-Independence Are Related to Default Network Activity David Stawarczyk 1 *, Steve Majerus 1, Pierre Maquet 2, Arnaud D Argembeau 1 1 Department of Cognitive Sciences,
: Both Task-Unrelatedness and Stimulus-Independence Are Related to Default Network Activity David Stawarczyk 1 *, Steve Majerus 1, Pierre Maquet 2, Arnaud D Argembeau 1 1 Department of Cognitive Sciences,
Mental Rotation is Not Easily Cognitively Penetrable
 Mental Rotation is Not Easily Cognitively Penetrable The Harvard community has made this article openly available. Please share how this access benefits you. Your story matters. Citation Published Version
Mental Rotation is Not Easily Cognitively Penetrable The Harvard community has made this article openly available. Please share how this access benefits you. Your story matters. Citation Published Version
The Role of Working Memory in Visual Selective Attention
 Goldsmiths Research Online. The Authors. Originally published: Science vol.291 2 March 2001 1803-1806. http://www.sciencemag.org. 11 October 2000; accepted 17 January 2001 The Role of Working Memory in
Goldsmiths Research Online. The Authors. Originally published: Science vol.291 2 March 2001 1803-1806. http://www.sciencemag.org. 11 October 2000; accepted 17 January 2001 The Role of Working Memory in
Working Memory (Goal Maintenance and Interference Control) Edward E. Smith Columbia University
 Edward E. Smith Columbia University") Working Memory (Goal Maintenance and Interference Control) Edward E. Smith Columbia University Outline Goal Maintenance Interference resolution: distraction, proactive interference, and directed forgetting
Working Memory (Goal Maintenance and Interference Control) Edward E. Smith Columbia University Outline Goal Maintenance Interference resolution: distraction, proactive interference, and directed forgetting
Vision and Action. 10/3/12 Percep,on Ac,on 1
 Vision and Action Our ability to move thru our environment is closely tied to visual perception. Simple examples include standing one one foot. It is easier to maintain balance with the eyes open than
Vision and Action Our ability to move thru our environment is closely tied to visual perception. Simple examples include standing one one foot. It is easier to maintain balance with the eyes open than
Neuroimaging studies of mental rotation: A metaanalysis
 Washington University School of Medicine Digital Commons@Becker Open Access Publications 2008 Neuroimaging studies of mental rotation: A metaanalysis and review Jeffrey M. Zacks Washington University School
Washington University School of Medicine Digital Commons@Becker Open Access Publications 2008 Neuroimaging studies of mental rotation: A metaanalysis and review Jeffrey M. Zacks Washington University School
Neural correlates of retrieval processing in the prefrontal cortex during recognition and exclusion tasks
 Neuropsychologia 41 (2003) 40 52 Neural correlates of retrieval processing in the prefrontal cortex during recognition and exclusion tasks Michael D. Rugg a,b,, Richard N.A. Henson a,c, William G.K. Robb
Neuropsychologia 41 (2003) 40 52 Neural correlates of retrieval processing in the prefrontal cortex during recognition and exclusion tasks Michael D. Rugg a,b,, Richard N.A. Henson a,c, William G.K. Robb
Supplemental Information. Triangulating the Neural, Psychological, and Economic Bases of Guilt Aversion
 Neuron, Volume 70 Supplemental Information Triangulating the Neural, Psychological, and Economic Bases of Guilt Aversion Luke J. Chang, Alec Smith, Martin Dufwenberg, and Alan G. Sanfey Supplemental Information
Neuron, Volume 70 Supplemental Information Triangulating the Neural, Psychological, and Economic Bases of Guilt Aversion Luke J. Chang, Alec Smith, Martin Dufwenberg, and Alan G. Sanfey Supplemental Information
Rules of apparent motion: The shortest-path constraint: objects will take the shortest path between flashed positions.
 Rules of apparent motion: The shortest-path constraint: objects will take the shortest path between flashed positions. The box interrupts the apparent motion. The box interrupts the apparent motion.
Rules of apparent motion: The shortest-path constraint: objects will take the shortest path between flashed positions. The box interrupts the apparent motion. The box interrupts the apparent motion.
How to report my result using REST slice viewer?
 How to report my result using REST slice viewer? Han Zhang Center for Cognition and Brain Disorders, Hangzhou Normal University napoleon1982@gmail.com 2013/12/30 Commonly, you got an activation for functional
How to report my result using REST slice viewer? Han Zhang Center for Cognition and Brain Disorders, Hangzhou Normal University napoleon1982@gmail.com 2013/12/30 Commonly, you got an activation for functional
Functional Elements and Networks in fmri
 Functional Elements and Networks in fmri Jarkko Ylipaavalniemi 1, Eerika Savia 1,2, Ricardo Vigário 1 and Samuel Kaski 1,2 1- Helsinki University of Technology - Adaptive Informatics Research Centre 2-
Functional Elements and Networks in fmri Jarkko Ylipaavalniemi 1, Eerika Savia 1,2, Ricardo Vigário 1 and Samuel Kaski 1,2 1- Helsinki University of Technology - Adaptive Informatics Research Centre 2-
Investigations in Resting State Connectivity. Overview
 Investigations in Resting State Connectivity Scott FMRI Laboratory Overview Introduction Functional connectivity explorations Dynamic change (motor fatigue) Neurological change (Asperger s Disorder, depression)
Investigations in Resting State Connectivity Scott FMRI Laboratory Overview Introduction Functional connectivity explorations Dynamic change (motor fatigue) Neurological change (Asperger s Disorder, depression)
AN fmri EXAMINATION OF VISUAL INTEGRATION IN SCHIZOPHRENIA
 Journal of Integrative Neuroscience, Vol. 8, No. 2 (2009) 175 202 c Imperial College Press Research Report AN fmri EXAMINATION OF VISUAL INTEGRATION IN SCHIZOPHRENIA STEVEN M. SILVERSTEIN,,, SARAH BERTEN,,
Journal of Integrative Neuroscience, Vol. 8, No. 2 (2009) 175 202 c Imperial College Press Research Report AN fmri EXAMINATION OF VISUAL INTEGRATION IN SCHIZOPHRENIA STEVEN M. SILVERSTEIN,,, SARAH BERTEN,,
Visual Mental Imagery Induces Retinotopically Organized Activation of Early Visual Areas
 Cerebral Cortex October 2005;15:1570--1583 doi:10.1093/cercor/bhi035 Advance Access publication February 2, 2005 Visual Mental Imagery Induces Retinotopically Organized Activation of Early Visual Areas
Cerebral Cortex October 2005;15:1570--1583 doi:10.1093/cercor/bhi035 Advance Access publication February 2, 2005 Visual Mental Imagery Induces Retinotopically Organized Activation of Early Visual Areas
LEFT POSTERIOR PARIETAL CORTEX PARTICIPATES IN BOTH TASK PREPARATION AND EPISODIC RETRIEVAL. Jeffrey S. Phillips. B.A., Villanova University, 1997
 LEFT POSTERIOR PARIETAL CORTEX PARTICIPATES IN BOTH TASK PREPARATION AND EPISODIC RETRIEVAL by Jeffrey S. Phillips B.A., Villanova University, 1997 Submitted to the Graduate Faculty of the School of Arts
LEFT POSTERIOR PARIETAL CORTEX PARTICIPATES IN BOTH TASK PREPARATION AND EPISODIC RETRIEVAL by Jeffrey S. Phillips B.A., Villanova University, 1997 Submitted to the Graduate Faculty of the School of Arts
Dissociation between Dorsal and Ventral Posterior Parietal Cortical Responses to Incidental Changes in Natural Scenes
 Dissociation between Dorsal and Ventral Posterior Parietal Cortical Responses to Incidental Changes in Natural Scenes Lorelei R. Howard 1, Dharshan Kumaran 2, H. Freyja Ólafsdóttir 1, Hugo J. Spiers 1
Dissociation between Dorsal and Ventral Posterior Parietal Cortical Responses to Incidental Changes in Natural Scenes Lorelei R. Howard 1, Dharshan Kumaran 2, H. Freyja Ólafsdóttir 1, Hugo J. Spiers 1
Brain Imaging Investigation of the Impairing Effect of Emotion on Cognition
 Brain Imaging Investigation of the Impairing Effect of Emotion on Cognition Gloria Wong 1,2, Sanda Dolcos 1,3, Ekaterina Denkova 1, Rajendra A. Morey 4,5, Lihong Wang 4,5, Nicholas Coupland 1, Gregory
Brain Imaging Investigation of the Impairing Effect of Emotion on Cognition Gloria Wong 1,2, Sanda Dolcos 1,3, Ekaterina Denkova 1, Rajendra A. Morey 4,5, Lihong Wang 4,5, Nicholas Coupland 1, Gregory
Supplementary Materials for
 Supplementary Materials for Folk Explanations of Behavior: A Specialized Use of a Domain-General Mechanism Robert P. Spunt & Ralph Adolphs California Institute of Technology Correspondence may be addressed
Supplementary Materials for Folk Explanations of Behavior: A Specialized Use of a Domain-General Mechanism Robert P. Spunt & Ralph Adolphs California Institute of Technology Correspondence may be addressed
Cognitive Neuroscience of Memory
 Cognitive Neuroscience of Memory Types and Structure of Memory Types of Memory Type of Memory Time Course Capacity Conscious Awareness Mechanism of Loss Sensory Short-Term and Working Long-Term Nondeclarative
Cognitive Neuroscience of Memory Types and Structure of Memory Types of Memory Type of Memory Time Course Capacity Conscious Awareness Mechanism of Loss Sensory Short-Term and Working Long-Term Nondeclarative
Chapter 8: Visual Imagery & Spatial Cognition
 1 Chapter 8: Visual Imagery & Spatial Cognition Intro Memory Empirical Studies Interf MR Scan LTM Codes DCT Imagery & Spatial Cognition Rel Org Principles ImplEnc SpatEq Neuro Imaging Critique StruEq Prop
1 Chapter 8: Visual Imagery & Spatial Cognition Intro Memory Empirical Studies Interf MR Scan LTM Codes DCT Imagery & Spatial Cognition Rel Org Principles ImplEnc SpatEq Neuro Imaging Critique StruEq Prop
Nature Neuroscience doi: /nn Supplementary Figure 1. Characterization of viral injections.
 Supplementary Figure 1 Characterization of viral injections. (a) Dorsal view of a mouse brain (dashed white outline) after receiving a large, unilateral thalamic injection (~100 nl); demonstrating that
Supplementary Figure 1 Characterization of viral injections. (a) Dorsal view of a mouse brain (dashed white outline) after receiving a large, unilateral thalamic injection (~100 nl); demonstrating that
Attention Response Functions: Characterizing Brain Areas Using fmri Activation during Parametric Variations of Attentional Load
 Attention Response Functions: Characterizing Brain Areas Using fmri Activation during Parametric Variations of Attentional Load Intro Examine attention response functions Compare an attention-demanding
Attention Response Functions: Characterizing Brain Areas Using fmri Activation during Parametric Variations of Attentional Load Intro Examine attention response functions Compare an attention-demanding
Prediction of Successful Memory Encoding from fmri Data
 Prediction of Successful Memory Encoding from fmri Data S.K. Balci 1, M.R. Sabuncu 1, J. Yoo 2, S.S. Ghosh 3, S. Whitfield-Gabrieli 2, J.D.E. Gabrieli 2 and P. Golland 1 1 CSAIL, MIT, Cambridge, MA, USA
Prediction of Successful Memory Encoding from fmri Data S.K. Balci 1, M.R. Sabuncu 1, J. Yoo 2, S.S. Ghosh 3, S. Whitfield-Gabrieli 2, J.D.E. Gabrieli 2 and P. Golland 1 1 CSAIL, MIT, Cambridge, MA, USA
An fmri study of the Interplay of Symbolic and Visuo-spatial Systems in Mathematical Reasoning
 An fmri study of the Interplay of Symbolic and Visuo-spatial Systems in Mathematical Reasoning Atsushi Terao (atsushi@edu.hokudai.ac.jp) Graduate School of Education, Hokkaido University, Kita 11 Nishi
An fmri study of the Interplay of Symbolic and Visuo-spatial Systems in Mathematical Reasoning Atsushi Terao (atsushi@edu.hokudai.ac.jp) Graduate School of Education, Hokkaido University, Kita 11 Nishi
Involvement of both prefrontal and inferior parietal cortex. in dual-task performance
 Involvement of both prefrontal and inferior parietal cortex in dual-task performance Fabienne Collette a,b, Laurence 01ivier b,c, Martial Van der Linden a,d, Steven Laureys b, Guy Delfiore b, André Luxen
Involvement of both prefrontal and inferior parietal cortex in dual-task performance Fabienne Collette a,b, Laurence 01ivier b,c, Martial Van der Linden a,d, Steven Laureys b, Guy Delfiore b, André Luxen
Neural activity to positive expressions predicts daily experience of schizophrenia-spectrum symptoms in adults with high social anhedonia
 1 Neural activity to positive expressions predicts daily experience of schizophrenia-spectrum symptoms in adults with high social anhedonia Christine I. Hooker, Taylor L. Benson, Anett Gyurak, Hong Yin,
1 Neural activity to positive expressions predicts daily experience of schizophrenia-spectrum symptoms in adults with high social anhedonia Christine I. Hooker, Taylor L. Benson, Anett Gyurak, Hong Yin,
The brain s cutting-room floor: segmentation of narrative cinema
 HUMAN NEUROSCIENCE Original Research Article published: 01 October 2010 doi: 10.3389/fnhum.2010.00168 The brain s cutting-room floor: segmentation of narrative cinema Jeffrey M. Zacks 1 *, Nicole K. Speer
HUMAN NEUROSCIENCE Original Research Article published: 01 October 2010 doi: 10.3389/fnhum.2010.00168 The brain s cutting-room floor: segmentation of narrative cinema Jeffrey M. Zacks 1 *, Nicole K. Speer
The Neural Correlates of Moral Decision-Making in Psychopathy
 University of Pennsylvania ScholarlyCommons Neuroethics Publications Center for Neuroscience & Society 1-1-2009 The Neural Correlates of Moral Decision-Making in Psychopathy Andrea L. Glenn University
University of Pennsylvania ScholarlyCommons Neuroethics Publications Center for Neuroscience & Society 1-1-2009 The Neural Correlates of Moral Decision-Making in Psychopathy Andrea L. Glenn University
Supplementary materials for: Executive control processes underlying multi- item working memory
 Supplementary materials for: Executive control processes underlying multi- item working memory Antonio H. Lara & Jonathan D. Wallis Supplementary Figure 1 Supplementary Figure 1. Behavioral measures of
Supplementary materials for: Executive control processes underlying multi- item working memory Antonio H. Lara & Jonathan D. Wallis Supplementary Figure 1 Supplementary Figure 1. Behavioral measures of
Behavioral Task Performance
 Zacks 1 Supplementary content for: Functional Reorganization of Spatial Transformations After a Parietal Lesion Jeffrey M. Zacks, PhD *, Pascale Michelon, PhD *, Jean M. Vettel, BA *, and Jeffrey G. Ojemann,
Zacks 1 Supplementary content for: Functional Reorganization of Spatial Transformations After a Parietal Lesion Jeffrey M. Zacks, PhD *, Pascale Michelon, PhD *, Jean M. Vettel, BA *, and Jeffrey G. Ojemann,
Functional MRI study of gender effects in brain activations during verbal working
 Functional MRI study of gender effects in brain activations during verbal working memory task Zbyněk Tüdös 1, Pavel Hok 2, Petr Hluštík 2, Aleš Grambal 3 1 Department of Radiology, Faculty of Medicine
Functional MRI study of gender effects in brain activations during verbal working memory task Zbyněk Tüdös 1, Pavel Hok 2, Petr Hluštík 2, Aleš Grambal 3 1 Department of Radiology, Faculty of Medicine
Distinct Value Signals in Anterior and Posterior Ventromedial Prefrontal Cortex
 Supplementary Information Distinct Value Signals in Anterior and Posterior Ventromedial Prefrontal Cortex David V. Smith 1-3, Benjamin Y. Hayden 1,4, Trong-Kha Truong 2,5, Allen W. Song 2,5, Michael L.
Supplementary Information Distinct Value Signals in Anterior and Posterior Ventromedial Prefrontal Cortex David V. Smith 1-3, Benjamin Y. Hayden 1,4, Trong-Kha Truong 2,5, Allen W. Song 2,5, Michael L.
Primacy and recency effects as indices of the focus of attention
 Primacy and recency effects as indices of the focus of attention Alexandra B. Morrison, Andrew R. A. Conway and Jason M. Chein Journal Name: Frontiers in Human Neuroscience ISSN: - Article type: Original
Primacy and recency effects as indices of the focus of attention Alexandra B. Morrison, Andrew R. A. Conway and Jason M. Chein Journal Name: Frontiers in Human Neuroscience ISSN: - Article type: Original
Reproducibility is necessary but insufficient for addressing brain science issues.
 1 Reproducibility is necessary but insufficient for addressing brain science issues. GLM promises nothing but the higher amplitude of the experimental trials on the average compared with the control trials.
1 Reproducibility is necessary but insufficient for addressing brain science issues. GLM promises nothing but the higher amplitude of the experimental trials on the average compared with the control trials.
Supporting Information
 Supporting Information Lingnau et al. 10.1073/pnas.0902262106 Fig. S1. Material presented during motor act observation (A) and execution (B). Each row shows one of the 8 different motor acts. Columns in
Supporting Information Lingnau et al. 10.1073/pnas.0902262106 Fig. S1. Material presented during motor act observation (A) and execution (B). Each row shows one of the 8 different motor acts. Columns in
Changes in Default Mode Network as Automaticity Develops in a Categorization Task
 Purdue University Purdue e-pubs Open Access Theses Theses and Dissertations January 2015 Changes in Default Mode Network as Automaticity Develops in a Categorization Task Farzin Shamloo Purdue University
Purdue University Purdue e-pubs Open Access Theses Theses and Dissertations January 2015 Changes in Default Mode Network as Automaticity Develops in a Categorization Task Farzin Shamloo Purdue University
Unconscious determinants of free decisions in the human brain
 Unconscious determinants of free decisions in the human brain Chun Siong Soon 1,2, Marcel Brass 1,3, Hans-Jochen Heinze 4 & John-Dylan Haynes* 1,2 1 Max Planck Institute for Human Cognitive and Brain Sciences,
Unconscious determinants of free decisions in the human brain Chun Siong Soon 1,2, Marcel Brass 1,3, Hans-Jochen Heinze 4 & John-Dylan Haynes* 1,2 1 Max Planck Institute for Human Cognitive and Brain Sciences,
Visual Selection and Attention
 Visual Selection and Attention Retrieve Information Select what to observe No time to focus on every object Overt Selections Performed by eye movements Covert Selections Performed by visual attention 2
Visual Selection and Attention Retrieve Information Select what to observe No time to focus on every object Overt Selections Performed by eye movements Covert Selections Performed by visual attention 2
Left Anterior Prefrontal Activation Increases with Demands to Recall Specific Perceptual Information
 The Journal of Neuroscience, 2000, Vol. 20 RC108 1of5 Left Anterior Prefrontal Activation Increases with Demands to Recall Specific Perceptual Information Charan Ranganath, 1 Marcia K. Johnson, 2 and Mark
The Journal of Neuroscience, 2000, Vol. 20 RC108 1of5 Left Anterior Prefrontal Activation Increases with Demands to Recall Specific Perceptual Information Charan Ranganath, 1 Marcia K. Johnson, 2 and Mark
Prefrontal cortex and recognition memory Functional-MRI evidence for context-dependent retrieval processes
 Brain (1998), 121, 1985 2002 Prefrontal cortex and recognition memory Functional-MRI evidence for context-dependent retrieval processes Anthony D. Wagner, 1 John E. Desmond, 1,2 Gary H. Glover 2 and John
Brain (1998), 121, 1985 2002 Prefrontal cortex and recognition memory Functional-MRI evidence for context-dependent retrieval processes Anthony D. Wagner, 1 John E. Desmond, 1,2 Gary H. Glover 2 and John
A possible mechanism for impaired joint attention in autism
 A possible mechanism for impaired joint attention in autism Justin H G Williams Morven McWhirr Gordon D Waiter Cambridge Sept 10 th 2010 Joint attention in autism Declarative and receptive aspects initiating
A possible mechanism for impaired joint attention in autism Justin H G Williams Morven McWhirr Gordon D Waiter Cambridge Sept 10 th 2010 Joint attention in autism Declarative and receptive aspects initiating
Supplementary materials. Appendix A;
 Supplementary materials Appendix A; To determine ADHD diagnoses, a combination of Conners' ADHD questionnaires and a semi-structured diagnostic interview was used(1-4). Each participant was assessed with
Supplementary materials Appendix A; To determine ADHD diagnoses, a combination of Conners' ADHD questionnaires and a semi-structured diagnostic interview was used(1-4). Each participant was assessed with
Text to brain: predicting the spatial distribution of neuroimaging observations from text reports (submitted to MICCAI 2018)
") 1 / 22 Text to brain: predicting the spatial distribution of neuroimaging observations from text reports (submitted to MICCAI 2018) Jérôme Dockès, ussel Poldrack, Demian Wassermann, Fabian Suchanek, Bertrand
1 / 22 Text to brain: predicting the spatial distribution of neuroimaging observations from text reports (submitted to MICCAI 2018) Jérôme Dockès, ussel Poldrack, Demian Wassermann, Fabian Suchanek, Bertrand
Frontal Contributions to Memory Encoding Before and After Unilateral Medial Temporal Lobectomy
 Frontal Contributions to Memory Encoding Before and After Unilateral Medial Temporal Lobectomy Jeff Ojemann, MD Department of Neurological Surgery University of Washington Children s Hospital & Regional
Frontal Contributions to Memory Encoding Before and After Unilateral Medial Temporal Lobectomy Jeff Ojemann, MD Department of Neurological Surgery University of Washington Children s Hospital & Regional
For better or for worse: neural systems supporting the cognitive down- and up-regulation of negative emotion
 For better or for worse: neural systems supporting the cognitive down- and up-regulation of negative emotion Kevin N. Ochsner, a, * Rebecca D. Ray, b Jeffrey C. Cooper, b Elaine R. Robertson, b Sita Chopra,
For better or for worse: neural systems supporting the cognitive down- and up-regulation of negative emotion Kevin N. Ochsner, a, * Rebecca D. Ray, b Jeffrey C. Cooper, b Elaine R. Robertson, b Sita Chopra,
NeuroImage 59 (2012) Contents lists available at SciVerse ScienceDirect. NeuroImage. journal homepage:
 Contents lists available at SciVerse ScienceDirect. NeuroImage. journal homepage:") NeuroImage 59 (2012) 2908 2922 Contents lists available at SciVerse ScienceDirect NeuroImage journal homepage: www.elsevier.com/locate/ynimg Routes to the past: Neural substrates of direct and generative
NeuroImage 59 (2012) 2908 2922 Contents lists available at SciVerse ScienceDirect NeuroImage journal homepage: www.elsevier.com/locate/ynimg Routes to the past: Neural substrates of direct and generative
Remembering the past and imagining the future: Common and distinct neural substrates during event construction and elaboration
 Neuropsychologia 45 (2007) 1363 1377 Remembering the past and imagining the future: Common and distinct neural substrates during event construction and elaboration Donna Rose Addis a,b,, Alana T. Wong
Neuropsychologia 45 (2007) 1363 1377 Remembering the past and imagining the future: Common and distinct neural substrates during event construction and elaboration Donna Rose Addis a,b,, Alana T. Wong
Medical Neuroscience Tutorial Notes
 Medical Neuroscience Tutorial Notes Finding the Central Sulcus MAP TO NEUROSCIENCE CORE CONCEPTS 1 NCC1. The brain is the body's most complex organ. LEARNING OBJECTIVES After study of the assigned learning
Medical Neuroscience Tutorial Notes Finding the Central Sulcus MAP TO NEUROSCIENCE CORE CONCEPTS 1 NCC1. The brain is the body's most complex organ. LEARNING OBJECTIVES After study of the assigned learning
Supplementary Material. Functional connectivity in multiple cortical networks is associated with performance. across cognitive domains in older adults
 Supplementary Material Functional connectivity in multiple cortical networks is associated with performance across cognitive domains in older adults Emily E. Shaw 1,2, Aaron P. Schultz 1,2,3, Reisa A.
Supplementary Material Functional connectivity in multiple cortical networks is associated with performance across cognitive domains in older adults Emily E. Shaw 1,2, Aaron P. Schultz 1,2,3, Reisa A.