Lisa S. Scott 1* James W. Tanaka 2. David L. Sheinberg 3. Tim Curran 1*
|
|
|
- Diane Hicks
- 5 years ago
- Views:
Transcription
1 A reevaluation of the electrophysiological correlates of expert object processing Lisa S. Scott 1* James W. Tanaka 2 David L. Sheinberg 3 Tim Curran 1* 1 Department of Psychology, University of Colorado 2 Department of Psychology, University of Victoria 3 Department of Neuroscience, Brown University * Corresponding Authors: Department of Psychology University of Colorado Campus Box 345 Boulder, CO addresses: tcurran@psych.colorado.edu (T. Curran) scottls@colorado.edu (L. Scott)
2 2 Abstract Subordinate-level object processing is regarded as a hallmark of perceptual expertise. However, the relative contribution of subordinate- and basic-level category experience in the acquisition of perceptual expertise has not been clearly delineated. In this study, participants learned to classify wading birds and owls at either the basic (e.g. wading bird, owl) or subordinate (e.g. egret, snowy owl) level. After 6 days of training, behavioral results showed that subordinate-, but not basic-, level training improved subordinate discrimination of trained exemplars, novel exemplars, and exemplars from novel species. Event-related potentials (ERPs) indicated that both basic- and subordinate-level training enhanced the early N170 component, but only subordinate-level training amplified the later N250 component. These results are consistent with models positing separate basic and subordinate learning mechanisms, and contrary to perspectives attempting to explain visual expertise solely in terms of subordinatelevel processing, suggest that expertise enhances neural responses of both basic and subordinate processing.
3 3 People identify objects at different levels. For example, a robin can be identified at the basic level (i.e. bird) or at the subordinate level (i.e. robin). Experts (e.g. bird or car experts) tend to identify objects within their domain of expertise at a more subordinate level relative to novices. Moreover, expert object recognition is thought to involve a categorical shift from basic to subordinate levels with training (Tanaka, 2001). However, the mechanisms underlying this categorical shift with increases in perceptual expertise are relatively unknown. It has been suggested that the recognition of objects is accomplished through two computationally distinct systems, one for more specific, subordinate-level recognition, and another for basic level recognition (Tversky & Hemenway, 1984; Murphy & Brownell, 1985; Jolicoeur, 1990; Biederman, 1985; Logothetis & Sheinberg, 1996). This distinction is supported by research suggesting that the visual analysis of objects typically proceeds from utilizing coarse to fine visual information (e.g. Kimchi, 1992; Schyns & Oliva, 1994). More specifically, previous research suggests that coarse shape information useful for identifying objects at the basic level occurs earlier in visual processing than does more fine detailed information, which allows for subordinate-level recognition (Collin & McMullen, 2005). Furthermore, the initial basic-level identification of an object has been found to depend on early processing of lowspatial frequency (LSF) information (Collin & McMullen, 2005) and global shapes (Biederman & Gerhardstein, 1993), whereas subordinate-level identification requires further visual processing of fine details (Jolicoeur et al., 1984). These findings are supported by a recent neuro-computational model of expert perceptual categorization (Nguyen & Cottrell, 2005). This model posits the necessity of two separate systems for basic and subordinate-level processing and suggests that the brain functionally separates these levels of processing.
4 4 Based on the above findings, the timeline of visual object categorization has been hypothesized to occur from basic to subordinate levels of abstraction, defining the basic level as the "entry-point" in object processing. However, in face identification, recognition of atypical exemplars, and recognition of other objects of expertise, this entry-point is hypothesized to have shifted downward from the basic to the subordinate-level of abstraction (Tanaka, 2001; Tanaka, Curran & Sheinberg, 2005; Tanaka & Taylor, 1991). Currently, the mechanisms underlying this observed shift, and how face processing as well as other expert perceptual processing differs from other types of object recognition, are contentiously debated and not well understood. For example, Grill-Spector & Kanwisher (2005), found evidence for basic-level processing preceding subordinate-level processing for both objects and faces. The current study sought to examine the electrophysiological correlates of the observed shift in entry level with increased expertise. Although it is difficult to track this shift in real-world experts, training studies, which systematically control subordinate and basic-level experience, allow for further elucidation of these mechanisms. Previous investigations of real-world experts, such as car or bird experts, and training studies have established a relation between the N170 ERP component and expert object recognition (Tanaka & Curran, 2001; Gauthier et al., 2003; Rossion, Gauthier, Goffaux, Tarr, & Crommelinck, 2002; Busey & Vanderkolk, 2005). For example, Tanaka and Curran (2001) report enhanced N170 amplitude when bird experts view birds compared to dogs and when dog experts view dogs compared to birds. This N170 difference is also seen in car experts (Gauthier et al. 2003). In addition, fingerprint experts show an inversion effect, while viewing fingerprints, relative to novices (Busey & Vanderkolk, 2005). Studies in which participants were trained to better differentiate novel objects have also found N170 changes over training. For example,
5 5 participants who were trained with families of novel visual shapes called blobs exhibit enhanced N170 amplitude to these blobs (Curran, Tanaka & Weiskopf, 2002) and participants trained with novel objects called greebles show N170 inversion effects, similar to what is seen with faces (Rossion et al., 2002). Combined, the above results suggest that the N170 is modulated by perceptual expertise; however, it is unclear whether increased expertise at the basic, or the subordinate-level (or both), accounts for these electrophysiological differences. Moreover, it is currently unclear how the amount of category experience, as opposed to basicand subordinate-level learning modulates the N170. The N250 is an ERP component that is associated with face processing (Schweinberger, Pickering, Burton, & Kaufmann, 2002). This component is sensitive to repetition, familiarity, and appears to be related to the recognition of individual people (Schweinberger, Huddy, & Burton, 2004; Tanaka, Curran, Porterfield & Collins, in press; Itier & Taylor, 2004a). Furthermore, a recent investigation using source localization postulates that the N250 (and not the N170) may originate from the fusiform gyrus, an area in the occipital-temporal cortex associated with face processing (Schweinberger, Huddy, & Burton, 2004). Given previous arguments that the FFA is generally important for subordinate-level processing (e.g. Gauthier et al., 2000), the N250 might also be related to subordinate-level expertise. However, the specificity of this component has not yet been investigated using non-face objects of expertise. The present study incorporates behavioral and electrophysiological methods to investigate the acquisition of perceptual expertise. Similar to Tanaka, Curran and Sheinberg (2005) adult participants learned to classify ten species of wading birds and ten species of owls at either the subordinate (species, e.g., Snowy owl) or basic (wading bird) level of abstraction (See Figure 1). Training included six days of perceptual learning tasks with the amount of learning
6 6 trials equated for both the basic and subordinate-level training. Pre and post-training performance was measured using a serially presented same/different matching task while ERPs were recorded. This task tested subordinate knowledge for species trained at the subordinate level versus those trained only at the basic level. Replicating previously reported findings (Tanaka, Curran & Sheinberg, 2005), we found that perceptual discrimination transferred to new exemplars and new species for birds learned at the subordinate level but not at the basic level. Electrophysiological results revealed that relative to pre-training levels, an enhanced N170 was elicited by birds from both the owl and wading bird families - regardless of whether they were trained at the basic or subordinate level. These findings suggest that the previously reported expert N170 might be attributable to the greater basic-level category experience that experts have to objects in their domain of expertise relative to novices. In contrast, only those birds that were learned at the subordinate level elicited an enhanced N250 and this effect generalized to new exemplars and novel species of subordinate-level categories. Source analyses of these components revealed distinct dipole locations for the N170 and N250 suggesting that different neural generators might mediate these potentials. Together, these findings provide electrophysiological evidence for distinct mechanisms modulating basic and subordinate-level object recognition. Furthermore, based on the results of the current investigation, the acquisition of expertise appears to be associated with 1) increased basic-level category familiarity, including coarse category shape and structure information and 2) increased subordinate-level processing of fine details related to within category individuation. Furthermore, these results do not support previous N170 reports suggesting that the N170 is specifically related to expert perceptual processing at the subordinate level.
7 7 Methods Participants. Participants included 16 right-handed, undergraduates recruited from the University of Colorado at Boulder (6 Females). Seven participants completed subordinate-level training with wading birds and basic-level training with owls, and 9 completed subordinate-level training with owls and basic-level training with wading birds. One participant was excluded from the final sample due to poor behavioral performance. All participants gave informed consent to participate in this study. Each participant participated in 8 sessions on different days within a 2-week period. ERPs were recorded on the first and last sessions. Subjects were paid $15 per hour for ERP sessions, $10 per hour for behavioral training sessions, and were paid a bonus of $20 for completing all 8 sessions within a two-week period. Stimuli and Apparatus. Stimuli were digitized photographs of owls and wading birds obtained from bird identification field guides or ornithological websites. The training set was composed of six exemplars of ten different species from the owl family (Barn Owl, Barred Owl, Boreal Owl, Burrowing Owl, Eastern Screech-Owl, Elf Owl, Eurasian Eagle Owl, Flammulated Owl, and the Great Gray Owl) and 10 species from the wading bird family (American Bittern, Black-crowned Night Heron, Cattle Egret, Glassy Ibis, Great Blue Heron, Great Egret, Green Heron, Least Bittern, Limpkin, and the Little Blue Heron). The test set of stimuli included the trained exemplars, untrained exemplars of trained species, and untrained species. The untrained exemplars included new instances of new species of owls (Long-eared Owl, Northern hawk Owl, Northern Pygmy Owl, Northern Saw-whet Owl, Short eared Owl, Snowy Owl, Spotted Owl, and
8 8 the Whiskered Screech-Owl) and wading birds (Reddish Egret, Sandhill Crane, Snowy Egret, Tricolored heron, White Ibis, whooping Crane, Wood Stork, and the Yellow-crowned Night- Heron) not learned in training (See Figure 1). The images were cropped to show only the bird and were placed on a white background. Stimuli were displayed on a15-inch Mitsubishi flatpanel monitor. Procedure. All procedures were approved by the Institutional Review Board at the University of Colorado and were conducted in accordance with this approval. Electrophysiological Pre and Post-Training Assessment. Before and after training, participants completed a subordinate-level sequential matching task that has previously been shown to be sensitive to differences in levels of perceptual expertise (Tanaka, Curran, & Sheinberg, 2005; Gauthier et al., 2003) while EEG was recorded. Participants were shown a stimulus for 800 milliseconds followed by a fixation point for milliseconds and then another image for an additional 800 milliseconds. Then the participants were immediately presented with a question mark and were required to indicate whether the two images were of the SAME (e.g. two screech owls) or of a DIFFERENT (e.g., snowy owl & screech owl) species. The question mark remained on the screen until a response was made. SAME trials were always different exemplars of the same species. DIFFERENT trials included two exemplars of different species within the same family. Different trial lures were selected randomly (without replacement) from a pool of all other species within the same family. Thus, a subordinate-level discrimination was required for species trained at the subordinate and at the basic level. This task consisted of a total of 368 trials. Half of the trials were wading birds and half owls. To monitor changes related to training, the same stimuli were included in the pre and post-test task.
9 9 All stimuli were randomly ordered. Across both same and different trials, there were three different types of trials: 1) Trained Species/Trained Exemplars, 2) Trained Species/Untrained Exemplars and 3) Untrained Species/Untrained Exemplars. The trained species/trained exemplars condition included 60 images from the training sessions, the trained species/new exemplars included 60 new pictures of the trained species, and finally the untrained species/untrained exemplars included 64 images of new species that were never trained. Trial numbers across conditions were not equated because of the limited number of species types available. Behavioral Training Tasks. After the pre-training assessment, participants completed 6 days of training. One group completed subordinate-level training with wading birds and basiclevel training with owls and the other group did the opposite. The first training task was a naming task, the second task a category verification task and the third was a matching task. 1) Training Task 1 (Naming): Participants learned to label different species of either owls or wading birds. Participants completed 6 blocks of training on the first day and 9 blocks of training on days 2-6. Participants started this task with 2 species of each family (i.e. 2 wading birds and 2 owls) and increased by 1 more species in each family every time they got a block of trials correct. On Day 1, seven species within each family were trained. On Days 2-6, all ten species within each family were trained. Within each block 3 exemplars of each species were presented. The exemplars rotated across blocks and days, and all trained exemplars were presented on each day. The first presentation of each species was labeled, for example a screech owl with either the subordinate-level label This is Species Y or the basic-level label This is Other. Arbitrary labels, rather than actual family/species names (e.g., owl, screech owl) where used to help reduce the effects of prior knowledge. For the subordinate-level training participants
10 10 then pressed the Y key whenever they saw a screech owl. For the basic-level training participants pressed the O key whenever they saw an owl. The label was only present for the first presentation of each species in each block. Participants were presented with an equal number of basic and subordinate trials. Participants were required to score 100% in each block to move on; otherwise, the block was restarted. Feedback, including the correct answer, was given for 1500 milliseconds for incorrect responses. 2) Training Task 2 (Category Verification): Participants were presented with a subordinate-level or basic-level bird label, for example Species Y (subordinate) or Other Family (basic) for 500 milliseconds, followed by a fixation cross for 250 milliseconds and a picture of a bird for 500 milliseconds. If the image and the label matched, the participant pressed a key for SAME. If they did not match, they pressed a key for DIFFERENT. SAME trials included an exemplar from the same family (basic) or same species (subordinate). DIFFERENT trials included an exemplar from a different family (basic) or a different species within the same family (subordinate). On the first day of training there were 48 trials presented (2 subordinate/basic x 2 same/different x 12 exemplars). Lures for the DIFFERENT trials were selected from the previously trained exemplars. Thus, on Day 1, 3 exemplars from the 6 subordinate trained species were presented 2 times for a total of 36 trials (12 subordinate level "SAME" trials; 12 subordinate level lures; and 12 basic level lures) and 1 exemplar from each of the basic trained species were presented 2 times for a total of 12 trials (all 12 trials were basic level "SAME" trials). On Days 2-6, 3 exemplars from the 10 subordinate trained species were presented 1 time for a total of 30 trials (10 subordinate level "SAME" trials; 10 subordinate level lures; and 10 basic level lures) and 1 exemplar from the species trained at the basic level was presented 1 time for a total of 10 trials (all 10 trials were basic level "SAME" trials). Exemplars
11 11 were rotated across days so each exemplar was presented between 5 and 6 times throughout the training. Participants were given correct or incorrect feedback for 500 milliseconds following the response. On the first day of training participants completed this task once, but on all subsequent training days they also completed a speeded version of this task. In this version participants were given 1 second to respond or the trial was marked incorrect. The same number of trials/conditions were used in the speeded version of this task. 3)Training Task 3 (Matching): Participants were presented with a word label, for example This is Species Y or "This is Other" for 500 milliseconds followed by a presentation of two side by side pictures of birds. Participants then pressed one key if the correct or matching stimulus was on the LEFT and another if it was on the RIGHT. The paired stimuli remained on the screen until the participant responded. On the first day of training there were 48 trials presented (2 subordinate/basic x 2 left/right x 12 exemplars). For subordinate trials, targets were paired with a subordinate trained lure. For basic trials, targets were also paired with a subordinate trained lure. Within the subordinate level trials, 2 exemplars of each of the 6 subordinate-training species were presented 4 times, twice as targets and twice as lures. Within the basic level trials, 2 exemplars of the 6 basic-training species were presented twice as targets. On Days 2-6, this task included 40 trials (20 basic & 20 subordinate). Within the subordinate level trials 2 exemplars of each of the 10 trained species were presented twice (once as a target and once as a lure). Within the basic level trials, 2 exemplars of the 10 species, trained at the basic level, were paired with subordinate level lures and presented once as targets. Participants were given correct and incorrect feedback for 500 milliseconds following the response.
12 12 Electrophysiological Methods. Scalp voltages were collected with a 128-channel Geodesic Sensor Net TM (Tucker, 1993) connected to an AC-coupled, 128-channel, high input impedance amplifier (200 MΩ, Net Amps TM, Electrical Geodesics Inc., Eugene, OR). Amplified analog voltages ( Hz bandpass) were digitized at 250 Hz and collected continuously. Individual sensors were adjusted until impedances were less than 40 kω. Trials were discarded from analyses if they contained eye movements (vertical EOG channel differences greater than 70 µv) or more than ten bad channels (changing more than 100 µv between samples, or reaching amplitudes over 200 µv). EEG from individual channels that was consistently bad for a given participant was replaced using a spherical interpolation algorithm (Srinivasan, Nunez, Silberstein, Tucker, & Cadusch, 1996). Stimulus-locked ERPs were baseline-corrected with respect to a 100 ms pre-stimulus recording interval and digitally low-pass filtered at 40 Hz. An average-reference transformation was used to minimize the effects of reference-site activity and accurately estimate the scalp topography of the measured electrical fields. Due to low trial counts, ERPs included trials in which the subject responded either same or different, and all correct and incorrect trials. Collapsed across these factors and conditions, there was an average of 58.0 (Range: 48-64; SD=3.4) trials/subject within each condition for birds trained at the subordinate-level and 57.2 (Range 45-64; SD=3.9) trials within each condition for the birds trained at the basic level. Dipole Source Analysis. Dipole source analyses were conducted using BESA (Brain Electromagnetic Source Analysis, Version 5.0). Modeling in the present experiment used the four-shell spherical head model, which accounts for differing conductances of the brain, bone, cerebrospinal fluid, and scalp. Symmetry constraints with respect to location were used for
13 13 dipole pairs. No other localization constraints were used. Localization coordinates are reported according to the Talairach atlas (Talairach & Tournoux, 1988). Statistical Procedure Statistical Analysis of Behavioral Measures. To determine whether there was behavioral evidence of an entry-level shift from basic to subordinate processing across training, measures of reaction time for the category verification and matching tasks were entered into a 2 x 6 MANOVA with 2 levels of category level (basic, subordinate) and 6 levels of training day (day 1, day 2 day 6). In addition, d' analyses were conducted on the pre and post sequential matching task to determine changes in discriminability after training. Measures of d' were entered into a 2 x 2 x 3 including 2 levels of test (pre-test, post-test), 2 levels of categorization (basic, subordinate) and 3 levels of condition (training species/trained exemplars, trained species/new exemplars, and untrained species). Statistical Analysis of Electrophysiological Measures. Electrophysiological analyses of each individual component of interest (N170; N250) was analyzed using separate 2 x 2 x 3 x 2 x 2 x 2 MANOVAs including 2 levels of test (pre-test, post-test), 2 levels of categorization (basic, subordinate), 3 levels of condition (trained species/trained exemplars, trained species/new exemplars, and untrained species), 2 levels of stimulus presentation (first/second within a trial), and 2 hemispheres (right, left). Mean N170 amplitude was calculated from ms post-stimulus onset (mean latency (179 ms) ± 2*SD (15.89 ms)). The channels were selected by identifying the electrode locations in the right and left hemisphere with the largest N170 across all conditions (channels 59 and 92, between standard locations T5/T6 and P3/P4). Analyses were conducted on averaged
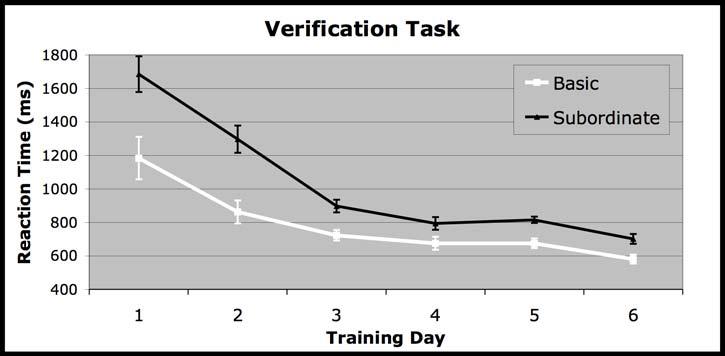
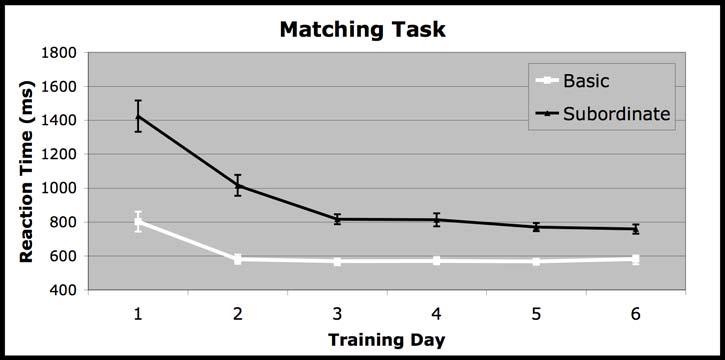
14 14 ERPs across these channels and the six immediately adjacent channels within each hemisphere (see Figure 4). Mean N250 amplitude was analyzed across conditions from ms post-stimulus onset. This window was selected for analyses based on previous reports of the N250 (Schweinberger et al., 2002; Schweinberger, Huddy, & Burton, 2004). Based on previous research (Tanaka, Curran, Porterfield & Collins, in press) the same electrode groups and analyses used for the N170 were used for the N250. Results Behavioral Results. During training tasks, reaction time (RT) measures were used to monitor the effects of training. RTs were computed for correct responses only. The category verification and matching tasks all showed significant interactions between training day and category level such that subordinate-level performance became increasingly similar to basic-level performance across days (verification (F(5, 10)=7.66, p=.003); speeded verification (F(4, 11) = 3.42, p=.05; matching (F(5, 11)=12.88, p=.0001; See Figure 2). However, on the last day of training, RT to basic level trials was still significantly faster than RT to subordinate-level trials across all tasks (p<.001). Accuracy for all tasks across all six days was at or near ceiling. Due to large variability between the number of completed blocks across subjects, analyses were not conducted for the naming task. Subordinate-level discrimination performance was assessed before and after training across trained and untrained exemplars and species using a successive matching task. Subordinate-level discriminations of birds trained at both the subordinate and the basic level
15 15 were tested in this task. Basic-level discriminations were not tested. Pre and post-matching task performance, measured with d', benefited only from subordinate-level training, and these benefits generalized from trained exemplars to untrained exemplars and untrained species (see Figure 3). Training level (basic, subordinate) significantly interacted with pre/post (F(1, 15)=267.21, p=.0001). When each training condition was considered separately, d' significantly increased from pre- to post-training at the subordinate level (F(1, 15)=266.14, p=.0001), but basic-level training had no effects (F(1, 15)=.364, p=.555). Planned comparisons revealed that subordinate-level training increased d' for all conditions: trained exemplars (t(15)=-6.70, p=.001), untrained exemplars of trained species (t(15)= -6.81, p=.001), and untrained species (t(15)= -3.94, p=.01). Electrophysiological Results. N170. Both subordinate- and basic-level training increased N170 amplitude in a manner that generalized across all conditions. Overall there was a significantly greater N170 posttraining compared to pre-training (F(1, 15)=19.13 p=.001; see Figure 4 & 5a), but the training effect did not interact with categorization level (F(1, 15)=.043, p=.839; see Figure 6 for topographic maps). T-tests confirmed that these training effects were significant for all conditions (trained exemplars, untrained exemplars, untrained species) following either subordinate- or basic-level training (all p's <.01). Analyses also revealed significant N170 differences between trained species/trained exemplars, trained species/untrained exemplars, and untrained species (F(2, 14)=4.84, p=.025). Further analyses of this effect revealed a decrease in magnitude from trained species/trained exemplars to trained species/untrained exemplars to untrained species (pairwise comparison of ordered means, p s range from ).
16 16 N250. Only subordinate-level training, but not basic-level training, increased N250 amplitude and these subordinate training effects generalized across all conditions. Overall there was a significantly greater N250 for birds trained at the subordinate versus the basic level (F(1, 15)=5.61, p=.032). This main effect is qualified by a significant interaction between pre and post training and basic versus subordinate training (F(1, 15)= 36.67, p=.0001), and between pre and post test, basic versus subordinate training and hemisphere (F(1, 15)=5.55, p=.032). These interactions reveal a larger post-test N250 for birds trained at the subordinate level in the right hemisphere (See Figures 4, 5b, & 6). T-tests indicated that these right-hemisphere subordinatelevel training effects were significant for all conditions (trained exemplars, untrained exemplars, untrained species, all p's <.05). There were also significant differences between the first and second stimulus presented within a trial. The second presentation of a stimulus elicited a smaller N250 than the first presentation of a stimulus (F(1, 15)=9.85, p=.007). This presentation order effect was greater in the right hemisphere compared to the left hemisphere (F(1, 15)=4.38, p=.05). Moreover, the difference between the first and second presentation interacted with subordinate/basic training (F(1, 15)=6.02, p =.03). This interaction was due to a smaller (less negative) mean amplitude N250 for the second stimulus presentation (M=5.14 µv; SD=.59) compared to the first stimulus presentation (M=4.21 µv; SD=.52) for subordinate, but not basic-level trials (first presentation M=4.8 µv, SD=.43; second presentation M=5.3 µv; SD=.552). This interaction did not further interact with pre/post, so did not modulate the training effects of primary interest. Visual inspection of the topography of these effects (see Figure 6) indicated additional, more lateral electrode locations exhibiting an N250. Thus, in addition to the above analyses, more lateral electrode locations were also analyzed (Right hemisphere: 101, 102, 108; Left
17 17 hemisphere 57, 58, 63) in a separate MANOVA to identify possible topographic differences between the N170 and the N250. Analyses of these regions replicated the interaction between pre and post test and subordinate versus basic level training (F(1, 15)=5.57, p=.032) revealing a greater N250 for birds trained at the subordinate versus the basic level at post-test. Dipole Source Analysis. Source localization was performed for the N170 and N250 using BESA (Version 5.0). Source analyses were conducted for the post-test subordinate condition in which both components were most discernable. The N170 and N250 were localized using a component onset to peak window (N170 = ms; N250= ms; Scherg & Berg, 1996). Spatial principle components analysis (PCA) revealed 1 factor for the N170 (97% of the variance explained) and 1 factor for the N250 (99.8% of the variance explained); therefore we fitted 1 pair of laterally symmetric sources for each component (See Figure 7). The Talairach coordinates for the centers of activity were x= +/-8.0, y= -58.2, z= 4.0 (RV= 3.3%) for the N170 and x = +/- 41.6, y= -62.1, z= -0.9 (RV=2.7%) for the N250. The center of activity for the N170 is in the lingual gyrus of the occipital lobe and the center of activity for the N250 is between the sub-gyral and inferior temporal gyral regions of the posterior temporal lobe. Discussion The current investigation was designed to further investigate the mechanisms associated with acquisition of perceptual expertise using both behavioral and electrophysiological measures. Behavioral results from the present investigation replicate previous findings (Tanaka, Curran & Sheinberg, 2005) indicating a decrease in RT to subordinate-level discriminations, which approaches the RT to basic-level discriminations. These data further support the presence of an entry-level shift in processing from the basic to subordinate level with expertise training. The results of the post-training sequential matching task suggest that training enhanced subordinate-
18 18 level discrimination of exemplars trained at the subordinate, but not the basic, level and that this enhanced discrimination generalized to novel exemplars of trained species and novel exemplars of untrained species within the owl or wading bird family but not across bird families. These data suggest that subordinate-level training in the present experiment led to increases in perceptual discrimination associated with category learning. At the electrophysiological level, basic- and subordinate-level training also produced changes in the N170 and N250 brain potentials. Based on previous reports of the relation between the N170 and perceptual expertise (Tanaka & Curran, 2001; Gauthier, Curran, Curby, & Collins, 2003; Rossion et al., 2002; Rossion, Curran & Gauthier, 2002) we expected to see increases in N170 amplitude associated with only subordinate-level bird training. However, counter to this expectation, participants showed an enhanced N170 to owls and wading birds regardless of whether they were trained at the subordinate or the basic level. These findings suggest that more general effects of basic-level category experience mediate N170 amplitude. Based on this result, the previously reported N170 differences between real-world experts and novices might reflect the expert s greater experience with objects in their domain of expertise rather than their subordinate-level knowledge of these objects (Tanaka & Curran, 2001; Gauthier et al., 2003; Rossion et al., 2002). These results are also consistent with the widely reported face N170 effect (Bentin, Allison, Puce, Perez, & McCarthy, 1996; Carmel & Bentin, 2002; Itier & Taylor, 2004b; Sagiv & Bentin, 2001; Rossion et al., 2000) in which faces have been shown to elicit a greater N170 relative to other non-face objects (e.g., cars, houses, chairs), but is not sensitive to the relative familiarity of any particular face (Bentin & Deouell, 2000; Eimer, 2000). Presumably, from birth, humans are exposed to faces more than any other object category and the years of accumulated experience are manifested in a greater N170 to faces. However, the
19 19 post-training N170 in the current study was not restricted to the specific items used in training because this effect extended to untrained exemplars of trained species. Thus, the N170 effect is not limited to the specific episodic experiences, but generalizes to the broader object categories of the perceptual experience. Whereas the N170 indexed general effects of basic-level category experience, the N250 component was only sensitive to training at the subordinate level. Specifically, a greater N250 deflection was elicited by participants trained with owls and wading birds at the subordinate level compared to the basic level. Consistent with subordinate-level matching task accuracy, the N250 training effects generalized to new exemplars of trained species and new species within the subordinate level family. These results are compatible with recent investigations demonstrating an N250r component to the repetition of familiar relative to unfamiliar faces (Schweinberger et al., 2002; Schweinberger, Huddy, & Burton, 2004; Tanaka, Curran, Porterfield, & Collins, in press). However, the current findings differ from previous N250 experiments in several ways. First, the present study demonstrates that the N250 is not specific to faces, but extends to the recognition of non-face objects. Second, our experiment highlights the importance of subordinate-level learning in the amplification of the N250 and is differentiated from the N170 effect that is produced as a consequence of both basic-level and subordinate-level category learning. Finally, whereas previous N250 reports suggest that this component depends on the immediate repetition of familiar stimuli, the current study shows that modulation of the N250 can be demonstrated under conditions over retention intervals lasting several days. Differences between basic and subordinate level object recognition are proposed by some to differ in qualitative ways (Nguyen & Cottrell, 2005; Tversky & Hemenway, 1984; Murphy & Brownell, 1985; Collin & McMullen, 2005) and by others in only quantitative ways
20 20 (Riesenhuber & Poggio, 2002). The present investigation localized the N170 to medial regions of the occipital lobes (lingual gyrus) and the N250 to more lateral regions of the temporal lobes (between the sub-gyral and inferior temporal gyral regions). Although one must exercise caution when interpreting localized sources using ERPs, here we place less emphasis on the identity of the localized regions than on the simple fact that the N170 and N250 localized to different locations, implying that the present results involve distinct, but likely interactive, brain mechanisms. The regions identified in the present investigation differ slightly from regions previously localized for faces (Schweinberger, Huddy, & Burton, 2004). Schweinberger et al., (2004) localized the N170 to the lateral occipital face area and the N250 the fusiform gyrus. Consistent with the source modeling conducted in the present experiment, previous reports have localized the N170 and N250 to separate brain regions (Schweinberger et al., 2002; Schweinberger, Huddy, & Burton, 2004). These reports, combined with the present investigation, suggest different neural generators for these two components, and are thereby consistent with models positing separate underlying mechanisms related to the learning of basic vs. subordinate-level information (e.g., Nguyen & Cottrell, 2005; Tversky & Hemenway, 1984; Murphy & Brownell, 1985). However, given the limitations of source analysis, the results of the present study may also be consistent with a single-system model for subordinate and basic-level processing (e.g., Riesenhuber & Poggio, 2000; Riesenhuber & Poggio, 2002). Riesenhuber and Poggio (2000; 2002) suggest that basic and subordinate level recognition are quantitatively, but not qualitatively different. In this view, the more fine-grained discrimination required for subordinate level processing would necessitate the accumulation of more perceptual information compared to what is required for basic level processing. Consistent with the results of the present investigation, this increase in amount of perceptual information needed would lead to a
21 21 longer latency response for subordinate compared to basic level processing. This would suggest that the increased N250 to subordinate compared to basic level trained birds could simply be due to the increased difficulty subordinate level discrimination of birds trained at the basic level posed to participants. This interpretation does not require separate systems for basic and subordinate level recognition. Future research combing both computational modeling and electrophysiology may be able to more clearly delineate the difference between basic and subordinate level object processing. The present electrophysiological findings further explain behavioral data (Tanaka, Curran, & Sheinberg, 2005) suggesting that there is a shift in categorical entry-level processing from the basic to subordinate levels with increases in perceptual expertise. The ERP results suggest that, similar to general object recognition, perceptual analysis of objects of expertise proceeds from the basic to subordinate levels of analysis. Moreover, the coarse shape information characterized by basic-level categorization occurs earlier in visual processing than the more fine grained visual information required for subordinate-level categorization, regardless of level of expertise. Based on these findings we can conclude that perceptual expertise not only includes increased subordinate-level discrimination of objects within a category of expertise, but is also associated with enhanced basic level processing. Thus, perceptual expertise might depend on two interacting mechanisms including 1) increases in basic level shape information (indexed by the N170) and 2) increases in more fine-grained subordinate-level discrimination (indexed by the N250). However, it is currently unclear whether there are functionally parallel mechanisms for basic and subordinate-level processing (e.g., Nguyen & Cottrell, 2005) or whether basic-level processing simply proceeds and then feeds into subordinate-level processing (e.g., Jolicoeur, Gluck, & Kosslyn, 1984).
22 22 Our interpretation of the present results is generally consistent with other views of the N170 and N250 within the specific domain of face processing. The face N170 has been related to structural differentiation of faces from other categories of objects (Sagiv & Bentin, 2001; Bentin & Deouell, 2000; Eimer, 2000; Schweinberger & Burton, 2003). The face N250, on the other hand, has been related to the ability to identify individual faces (Schweinberger et al., 2002; Schweinberger, Huddy, & Burton, 2004). Thus, just as others have suggested that the N170 and N250 may index separate processes in face recognition, the present results suggest that these components may reflect more domain-general learning/categorization mechanisms. Like studies of real-world expertise, studies of face recognition are limited in their ability to measure the processes involved in the acquisition of categorical knowledge. The present study sheds light on these processes by measuring electrophysiological changes that result from category learning under controlled conditions.
23 23 Acknowledgements This research was supported by the James S. McDonnell Foundation, a grant to T.C. from the National Institute of Mental Health (MH64812) and a grant to J.T. from Natural Sciences and Engineering Research Council of Canada. The authors thank members of the Perceptual Expertise Network for relevant discussion, and to D. Chase, R. Danson, C. DeBuse, D. Germain, J. Hancock, J. Knellinger, B. Malcolm, R. Praw, E. Walsky, B. Woroch, and B. Young for technical and research assistance.
24 24 Figure Caption Figure 1: Examples of stimuli. The top panel pictures examples of trained exemplars of owls (species A) and wading birds (species B). Training level (basic vs. subordinate) for owls and wading birds was counterbalanced across subjects. The middle panel pictures examples of owls (species A) and wading birds (species B) in the trained species/untrained exemplars condition. The bottom panel pictures examples of owls (species C) and wading birds (species D) in the untrained species condition. Figure 2: Reaction time changes across training. Behavioral data illustrating an entry-level shift in reaction time (RT) across the three different training tasks a) Verification b) Matching and c) Speeded Verification. Error bars are standard error of the mean. Figure 3: Pre- and Post-training Matching Performance. Behavioral d scores (Mean and Standard Errors) across all conditions for the pre and post-training sequential matching task. Figure 4: ERP waveforms. The left graph represents an average of electrodes in the left hemisphere (59, 51, 52, 58, 60, 65, and 66) and the right graph represents an average of electrodes in the right hemisphere (92, 91, 93, 97, 98, 85, and 86). Figure 5: Amplitude changes across training. Bar graphs of the mean and standard error of the average amplitude across conditions in the pre- and post-tests across hemispheres for the N170 (a) and in the right hemisphere for the N250 (b). Figure 6: Topography of Subordinate-level effects. Topographic map of the difference between the pre and post subordinate-level for the N170 (left) and the N250 (right). Figure 7: Source modeling. Panels are estimates of source localization for the post-test subordinate-level condition for each component
25 25 References Bentin, S., Allison, T., Puce, A., Perez, E. and McCarthy, G. (1996). Electrophysiological studies of face perception in humans. Journal of Cognitive Neuroscience, 8, Bentin, S. and Deouell, L.Y. (2000). Structural encoding and identification in face processing: ERP evidence for separate mechanisms. Cognitive Neuropsychology 17, Biederman, I. (1985). Human image understanding: Recent research and theory. Computer Vision, Graphics, and Image Processing, 32, Biederman, I. and Gerhardstein, P.C. (1993). Recognizing depth-rotated objects: Evidence and conditions for three-dimensional view point invariance. Journal of Experimental Psychology: Human Perception and Performance, 18, Busey, T.A. and Vanderkolk, J.R. (2005). Behavioral and electrophysiological evidence for configural processing in fingerprint experts. Vision Research, 45, Carmel, D. and Bentin, S. (2002). Domain specificity versus expertise: Factors influencing distinct processing of faces. Cognition, 83, Collin, C.A. and McMullen, P. (2005). Subordinate-level categorization relies on high spatial frequencies to a greater degree than basic-level categorization. Perception & Psychophysics, 67, Curran, T., Tanaka, J.W., and Weiskopf, D.M. (2002). An electrophysiological comparison of visual categorization and recognition memory. Cognitive, Affective, and Behavioral Neuroscience, 2, Eimer, M. (2000). The face-specific N170 component reflects late stages in the structural encoding of faces. Neuroreport, 11,
26 26 Gauthier, I., et al. (2002). Does subordinate-level categorization engage the functionally-defined face area? Cognitive Neuropsychology, 17, Gauthier, I., Curran, T., Curby, K.M., & Collins, D. (2003). Perceptual Interference supports a non-modular account of face processing. Nature Neuroscience, 6, Gauthier, I., & Tarr, M.J. (1997). Becoming a Greeble expert: Exploring the face recognition mechanism. Vision Research, 37, Gauthier, I., Williams, P., Tarr, M.J., & Tanaka. (1998). Training greeble experts: A framework for studying expert object recognition processes. Vision Research, 38, Grill-Spector, K. & Kanwisher, N. (2005). Visual Recognition: As soon as you know it is there, you know what it is. Psychological Science, 16, Henson, R., Shallice, T., & Dolan, R. (2000). Neuroimaging evidence for dissociable forms of repetition priming. Science, 287, Henson, R., Shallice, T., Gorno-Tempini, M.L. & Dolan, R.J. (2002). Face repetition effects in implicit and explicit memory tests as measured by fmri. Cerebral Cortex, 12, Itier, R.J. & Taylor, M.J. (2004a). Effects of repetition learning on upright, inverted and contrast-reversed face processing using ERPs. Neuroimage, 21, Itier, R. J. & Taylor, M.J. (2004b). N170 or N1? Spatiotemporal differences between object and face processing using ERPs. Cerebral Cortex, 14, Jolicoeur, P. (1990). Identification of disoriented objects: A dual-systems theory. Mind & Language, 5, Jolicoeur, P., Gluck, M., & Kosslyn, S.M. (1984). Pictures and names: Making the connection. Cognitive Psychology, 16,
27 27 Kimchi, R. (1992). Primacy of wholistic processing and global/local paradigm: A critical review. Psychological Bulletin, 112, Logothetis, N.K. & Sheinberg, D.L. (1996). Visual object recognition. Annual Review of Neuroscience, 19, Nguyen, N. & Cottrell, G.W. (in press) Owls and Wading Birds: Generalization Gradients in Expertise. In Proceedings of the 27th Annual Cognitive Science Conference, La Stresa, Italy. Mahwah: LEA. Murphy, G.L., & Brownell, H.H. (1985). Category differentiation in object recognition: Typicality constraints on the basic category advantage. Journal of Experimental Psychology: Learning, Memory and Cognition, 11, Riesenhuber, M. & Poggio, T. (2000). Models of object recognition. Nature Neuroscience, 3, Riesenhuber, M. & Poggio, T. (2002). Neural mechanisms of object recognition. Current Opinion in Neurobiology, 12, Rolls, E.T., Tovee, M.J. & Panzeri, S. (1999). The neuropsychology of backward visual masking: information analysis. Journal of Cognitive Neuroscience, 11, Rossion, B., et al. (2000). Hemispheric asymmetries for whole-based and part-based face processing in the human fusiform gyrus. Journal of Cognitive Neuroscience, 12, Rossion, B., Curran, T. & Gauthier, I. (2002). A defense of the subordinate-level expertise account for the N170 component. Cognition 85, Rossion, B., Gauthier, I., Goffaux, V., Tarr, M.J. & Crommelinck, M. (2002). Expertise training with novel objects leads to left lateralized facelike electrophysiological responses.
28 28 Psychological Science, 13, Sagiv, N. & Bentin, S. (2001). Structural encoding of human and schematic faces: Holistic and part-based processes. Journal of Cognitive Neuroscience, 13, Scherg, M, & Berg, P. (1996). New concepts of brain source imaging and localization. Electroencephalography and Clinical Neurophysiology Supplement, 46, Schweinberger, S. R., & Burton, A. M. (2003). Covert recognition and the neural system for face processing. Cortex 39, Schweinberger, S.R., Huddy, V. & Burton, M. (2004). N250r: a face-selective brain response to stimulus repetitions. Neuroreport, 15, Schweinberger, S.R., Pickering, E.C., Burton, M., & Kaufmann, J. (2002). Human brain potential correlates of repetition priming in face and name recognition. Neuropsychologia, 40, Schyns, P.G. & Oliva, A. (1994). From blobs to boundary edge: Evidence for time- and spatialscale-dependent scene recognition. Psychological Science, 5, Seitz, A., & Watanabe, T. (2005). A unified model for perceptual learning. Trends in Cognitive Science, 9, Srinivasan, R., Nunez, P.L., Silberstein, R.B., Tucker, D.M., & Cadusch, P.J. (1996). Spatial sampling and filtering of EEG with spline-laplacians to estimate cortical potentials. Brain Topography, 8, Tanaka, J.W. (2001). The entry point of face recognition: evidence for face expertise. Journal of Experimental Psychology: General, 130, Tanaka, J.W., & Curran, T. (2001). A neural basis for expert object recognition. Psychological Science, 12,
29 29 Tanaka, J.W., Curran, T., Porterfield, A.L., & Collins, D. (in press). Activation of pre-existing and acquired face representations: The N250 ERP as an index of face familiarity. Journal of Cognitive Neuroscience. Tanaka, J.W., Curran, T., & Sheinberg, D.L. (2005). The training and transfer of real-world, perceptual expertise. Psychological Science, 16, Tanaka, J.W. & Taylor, M. (1991) Object categories and expertise: Is the basic level in the eye of the beholder? Cognitive Psychology, 23, Tarr, M.J. & Gauthier, I. (2002). FFA: Flexible Fusiform are for subordinate-level visual processing automated by expertise. Nature Neuroscience, 3, Tovee, M. & Rolls, E.T. (1995). Information encoding in short firing rate epochs by single neurons in the primate temporal visual cortex. Visual Cognition 2, Tucker, D.M. (1993). Spatial sampling of head electrical fields: The geodesic sensor net. Electroencephalography and Clinical Neurophysiology, 87, Talairach J. and Tournoux P. (1988). Co-planar stereotaxic atlas of the human brain. (Stuttgart: George Thieme). Tversky, B., Hemenway, K. (1984). Objects, parts, and categories. Journal of Experimental Psychology: General, 113,

30

31 a) b) c)

32 * * * * * * Pre-Training Post-Training



33 Pre-Training Basic Pre-Training Subordinate Post-Training Basic Post-Training Subordinate Left Hemisphere Right Hemisphere Amplitude (microvolts) N170 N250 Latency (ms)

34 a) * * * * b) Pre-Training Post-Training




35 N170 N250 Subordinate Basic Subordinate Basic -1.5 µv 1.5 µv
Word Count: Please address correspondence to: James Tanaka, Department of Psychology, University
 1 The training and transfer of real-world, perceptual expertise James W. Tanaka 1, Tim Curran 2 and David Sheinberg 3 1 Department of Psychology, University of Victoria, British Columbia, Canada 2 Department
1 The training and transfer of real-world, perceptual expertise James W. Tanaka 1, Tim Curran 2 and David Sheinberg 3 1 Department of Psychology, University of Victoria, British Columbia, Canada 2 Department
The Training and Transfer of Real-World Perceptual Expertise
 PSYCHOLOGICAL SCIENCE Research Article The and Transfer of Real-World Perceptual Expertise 1 2 3 James W. Tanaka, Tim Curran, and David L. Sheinberg 1 Department of Psychology, University of Victoria,
PSYCHOLOGICAL SCIENCE Research Article The and Transfer of Real-World Perceptual Expertise 1 2 3 James W. Tanaka, Tim Curran, and David L. Sheinberg 1 Department of Psychology, University of Victoria,
A defense of the subordinate-level expertise account for the N170 component
 B. Rossion et al. / Cognition 85 (2002) 189 196 189 COGNITION Cognition 85 (2002) 189 196 www.elsevier.com/locate/cognit Discussion A defense of the subordinate-level expertise account for the N170 component
B. Rossion et al. / Cognition 85 (2002) 189 196 189 COGNITION Cognition 85 (2002) 189 196 www.elsevier.com/locate/cognit Discussion A defense of the subordinate-level expertise account for the N170 component
Accounts for the N170 face-effect: a reply to Rossion, Curran, & Gauthier
 S. Bentin, D. Carmel / Cognition 85 (2002) 197 202 197 COGNITION Cognition 85 (2002) 197 202 www.elsevier.com/locate/cognit Discussion Accounts for the N170 face-effect: a reply to Rossion, Curran, & Gauthier
S. Bentin, D. Carmel / Cognition 85 (2002) 197 202 197 COGNITION Cognition 85 (2002) 197 202 www.elsevier.com/locate/cognit Discussion Accounts for the N170 face-effect: a reply to Rossion, Curran, & Gauthier
Activation of Preexisting and Acquired Face Representations: The N250 Event-related Potential as an Index of Face Familiarity
 Activation of Preexisting and Acquired Face Representations: The N250 Event-related Potential as an Index of Face Familiarity James W. Tanaka 1, Tim Curran 2, Albert L. Porterfield 3, and Daniel Collins
Activation of Preexisting and Acquired Face Representations: The N250 Event-related Potential as an Index of Face Familiarity James W. Tanaka 1, Tim Curran 2, Albert L. Porterfield 3, and Daniel Collins
The overlap of neural selectivity between faces and words: evidences
 The overlap of neural selectivity between faces and words: evidences from the N170 adaptation effect Xiao-hua Cao 1, Chao Li 1, Carl M Gaspar 2, Bei Jiang 1 1. Department of Psychology,Zhejiang Normal
The overlap of neural selectivity between faces and words: evidences from the N170 adaptation effect Xiao-hua Cao 1, Chao Li 1, Carl M Gaspar 2, Bei Jiang 1 1. Department of Psychology,Zhejiang Normal
Mental representation of number in different numerical forms
 Submitted to Current Biology Mental representation of number in different numerical forms Anna Plodowski, Rachel Swainson, Georgina M. Jackson, Chris Rorden and Stephen R. Jackson School of Psychology
Submitted to Current Biology Mental representation of number in different numerical forms Anna Plodowski, Rachel Swainson, Georgina M. Jackson, Chris Rorden and Stephen R. Jackson School of Psychology
Learning to Become an Expert: Reinforcement Learning and the Acquisition of Perceptual Expertise
 Learning to Become an Expert: Reinforcement Learning and the Acquisition of Perceptual Expertise Olav E. Krigolson 1,2, Lara J. Pierce 1, Clay B. Holroyd 1, and James W. Tanaka 1 Abstract & To elucidate
Learning to Become an Expert: Reinforcement Learning and the Acquisition of Perceptual Expertise Olav E. Krigolson 1,2, Lara J. Pierce 1, Clay B. Holroyd 1, and James W. Tanaka 1 Abstract & To elucidate
ERP evidence for task modulations on face perceptual processing at different spatial scales
 Cognitive Science 27 (2003) 313 325 ERP evidence for task modulations on face perceptual processing at different spatial scales V. Goffaux a,b,, B. Jemel a,b, C. Jacques a, B. Rossion a,b,c, P.G. Schyns
Cognitive Science 27 (2003) 313 325 ERP evidence for task modulations on face perceptual processing at different spatial scales V. Goffaux a,b,, B. Jemel a,b, C. Jacques a, B. Rossion a,b,c, P.G. Schyns
The Time Scale of Perceptual Expertise
 The Time Scale of Perceptual Expertise 1.) Introduction: What is an expert? (Jim Tanaka, UVic) 2.) Becoming an expert... one day at a time (Jim) 3.) Expertise: Millisecond by millisecond (Tim Curran, Colorado)
The Time Scale of Perceptual Expertise 1.) Introduction: What is an expert? (Jim Tanaka, UVic) 2.) Becoming an expert... one day at a time (Jim) 3.) Expertise: Millisecond by millisecond (Tim Curran, Colorado)
(for Trends in Cognitive Science) Cindy Bukach 1,3. Isabel Gauthier 1,3. Michael J. Tarr 2,3. Research Center, Vanderbilt University, Nashville, TN
 Cindy Bukach 1,3. Isabel Gauthier 1,3. Michael J. Tarr 2,3. Research Center, Vanderbilt University, Nashville, TN") BEYOND FACES AND MODULARITY (for Trends in Cognitive Science) Cindy Bukach 1,3 Isabel Gauthier 1,3 Michael J. Tarr 2,3 1 Department of Psychology, Center for Integrative and Cognitive Neuroscience, Vanderbilt
BEYOND FACES AND MODULARITY (for Trends in Cognitive Science) Cindy Bukach 1,3 Isabel Gauthier 1,3 Michael J. Tarr 2,3 1 Department of Psychology, Center for Integrative and Cognitive Neuroscience, Vanderbilt
Perception of Faces and Bodies
 CURRENT DIRECTIONS IN PSYCHOLOGICAL SCIENCE Perception of Faces and Bodies Similar or Different? Virginia Slaughter, 1 Valerie E. Stone, 2 and Catherine Reed 3 1 Early Cognitive Development Unit and 2
CURRENT DIRECTIONS IN PSYCHOLOGICAL SCIENCE Perception of Faces and Bodies Similar or Different? Virginia Slaughter, 1 Valerie E. Stone, 2 and Catherine Reed 3 1 Early Cognitive Development Unit and 2
Dissociation of Category-Learning Mechanisms via Brain Potentials
 Dissociation of Category-Learning Mechanisms via rain Potentials Robert G. Morrison (rmorrison@luc.edu) Loyola University Chicago, Psychology Department, 6525 North Sheridan Road Chicago, IL 60626 USA
Dissociation of Category-Learning Mechanisms via rain Potentials Robert G. Morrison (rmorrison@luc.edu) Loyola University Chicago, Psychology Department, 6525 North Sheridan Road Chicago, IL 60626 USA
Object detection and basic-level categorization: Sometimes you know it is there before you know what it is
 R243B Time-Course of Object Detection and Categorization 1 PBR-BR-07-031.R2 Production Number R243B Object detection and basic-level categorization: Sometimes you know it is there before you know what
R243B Time-Course of Object Detection and Categorization 1 PBR-BR-07-031.R2 Production Number R243B Object detection and basic-level categorization: Sometimes you know it is there before you know what
Beyond faces and modularity: the power of an expertise framework
 Beyond faces and modularity: the power of an expertise framework Cindy M. Bukach 1, Isabel Gauthier 1 and Michael J. Tarr 2 1 Department of Psychology, Center for Integrative and Cognitive Neuroscience,
Beyond faces and modularity: the power of an expertise framework Cindy M. Bukach 1, Isabel Gauthier 1 and Michael J. Tarr 2 1 Department of Psychology, Center for Integrative and Cognitive Neuroscience,
The Role of Spatial Frequency in Expert Object Recognition
 Journal of Experimental Psychology: Human Perception and Performance 2016, Vol. 42, No. 3, 413 422 2015 American Psychological Association 0096-1523/16/$12.00 http://dx.doi.org/10.1037/xhp0000139 The Role
Journal of Experimental Psychology: Human Perception and Performance 2016, Vol. 42, No. 3, 413 422 2015 American Psychological Association 0096-1523/16/$12.00 http://dx.doi.org/10.1037/xhp0000139 The Role
Seeing faces in the noise: Stochastic activity in perceptual regions of the brain may influence the perception of ambiguous stimuli
 R175B BC JP To printer - need negs for pgs, 3,, and Psychonomic Bulletin & Review,?? (?),???-??? Seeing faces in the noise: Stochastic activity in perceptual regions of the brain may influence the perception
R175B BC JP To printer - need negs for pgs, 3,, and Psychonomic Bulletin & Review,?? (?),???-??? Seeing faces in the noise: Stochastic activity in perceptual regions of the brain may influence the perception
The neural plasticity of other-race face recognition. James W. Tanaka & Lara J. Pierce. University of Victoria. Victoria, British Columbia CANADA
 1 The neural plasticity of other-race face recognition James W. Tanaka & Lara J. Pierce University of Victoria Victoria, British Columbia CANADA Correspondence can be addressed to James Tanaka, Department
1 The neural plasticity of other-race face recognition James W. Tanaka & Lara J. Pierce University of Victoria Victoria, British Columbia CANADA Correspondence can be addressed to James Tanaka, Department
Early lateralization and orientation tuning for face, word, and object processing in the visual cortex
 NeuroImage 20 (2003) 1609 1624 www.elsevier.com/locate/ynimg Early lateralization and orientation tuning for face, word, and object processing in the visual cortex Bruno Rossion, a,b, * Carrie A. Joyce,
NeuroImage 20 (2003) 1609 1624 www.elsevier.com/locate/ynimg Early lateralization and orientation tuning for face, word, and object processing in the visual cortex Bruno Rossion, a,b, * Carrie A. Joyce,
DATA MANAGEMENT & TYPES OF ANALYSES OFTEN USED. Dennis L. Molfese University of Nebraska - Lincoln
 DATA MANAGEMENT & TYPES OF ANALYSES OFTEN USED Dennis L. Molfese University of Nebraska - Lincoln 1 DATA MANAGEMENT Backups Storage Identification Analyses 2 Data Analysis Pre-processing Statistical Analysis
DATA MANAGEMENT & TYPES OF ANALYSES OFTEN USED Dennis L. Molfese University of Nebraska - Lincoln 1 DATA MANAGEMENT Backups Storage Identification Analyses 2 Data Analysis Pre-processing Statistical Analysis
Revisiting the Role of the Fusiform Face Area in Visual Expertise
 Cerebral Cortex August 2005;15:1234--1242 doi:10.1093/cercor/bhi006 Advance Access publication January 26, 2005 Revisiting the Role of the Fusiform Face Area in Visual Expertise Yaoda Xu Department of
Cerebral Cortex August 2005;15:1234--1242 doi:10.1093/cercor/bhi006 Advance Access publication January 26, 2005 Revisiting the Role of the Fusiform Face Area in Visual Expertise Yaoda Xu Department of
Short article Detecting objects is easier than categorizing them
 THE QUARTERLY JOURNAL OF EXPERIMENTAL PSYCHOLOGY 2008, 61 (4), 552 557 Short article Detecting objects is easier than categorizing them Jeffrey S. Bowers and Keely W. Jones University of Bristol, Bristol,
THE QUARTERLY JOURNAL OF EXPERIMENTAL PSYCHOLOGY 2008, 61 (4), 552 557 Short article Detecting objects is easier than categorizing them Jeffrey S. Bowers and Keely W. Jones University of Bristol, Bristol,
Face Perception - An Overview Bozana Meinhardt-Injac Malte Persike
 Face Perception - An Overview Bozana Meinhardt-Injac Malte Persike Johannes Gutenberg University Mainz 2 Interesting to mention Bahrik, Bahrik & Wittlinger,1977 The identification and matching of once
Face Perception - An Overview Bozana Meinhardt-Injac Malte Persike Johannes Gutenberg University Mainz 2 Interesting to mention Bahrik, Bahrik & Wittlinger,1977 The identification and matching of once
Are All Types of Expertise Created Equal? Car Experts Use Different Spatial Frequency Scales for Subordinate Categorization of Cars and Faces
 Wright State University CORE Scholar Psychology Faculty Publications Psychology 6-24-2013 Are All Types of Expertise Created Equal? Car Experts Use Different Spatial Frequency Scales for Subordinate Categorization
Wright State University CORE Scholar Psychology Faculty Publications Psychology 6-24-2013 Are All Types of Expertise Created Equal? Car Experts Use Different Spatial Frequency Scales for Subordinate Categorization
Neural Correlates of Human Cognitive Function:
 Neural Correlates of Human Cognitive Function: A Comparison of Electrophysiological and Other Neuroimaging Approaches Leun J. Otten Institute of Cognitive Neuroscience & Department of Psychology University
Neural Correlates of Human Cognitive Function: A Comparison of Electrophysiological and Other Neuroimaging Approaches Leun J. Otten Institute of Cognitive Neuroscience & Department of Psychology University
Effects of Long-Term Object Familiarity on Event-Related Potentials in the Monkey
 Cerebral Cortex Advance Access published August 7, 26 Cerebral Cortex doi:1.193/cercor/bhl43 Effects of Long-Term Object Familiarity on Event-Related Potentials in the Monkey Jessie J. Peissig, Jedediah
Cerebral Cortex Advance Access published August 7, 26 Cerebral Cortex doi:1.193/cercor/bhl43 Effects of Long-Term Object Familiarity on Event-Related Potentials in the Monkey Jessie J. Peissig, Jedediah
Mental Representation of Number in Different Numerical Forms
 Current Biology, Vol. 13, 2045 2050, December 2, 2003, 2003 Elsevier Science Ltd. All rights reserved. DOI 10.1016/j.cub.2003.11.023 Mental Representation of Number in Different Numerical Forms Anna Plodowski,
Current Biology, Vol. 13, 2045 2050, December 2, 2003, 2003 Elsevier Science Ltd. All rights reserved. DOI 10.1016/j.cub.2003.11.023 Mental Representation of Number in Different Numerical Forms Anna Plodowski,
Translating From Perceptual to Cognitive Coding
 Translating From Perceptual to Cognitive Coding Tyler Davis (thdavis@mail.utexas.edu) Bradley C. Love (brad_love@mail.utexas.edu) W. Todd Maddox (maddox@psy.utexas.edu) Department of Psychology, The University
Translating From Perceptual to Cognitive Coding Tyler Davis (thdavis@mail.utexas.edu) Bradley C. Love (brad_love@mail.utexas.edu) W. Todd Maddox (maddox@psy.utexas.edu) Department of Psychology, The University
Impaired face discrimination in acquired prosopagnosia is associated with abnormal response to individual faces in the right middle fusiform gyrus
 Impaired face discrimination in acquired prosopagnosia is associated with abnormal response to individual faces in the right middle fusiform gyrus Christine Schiltz Bettina Sorger Roberto Caldara Fatima
Impaired face discrimination in acquired prosopagnosia is associated with abnormal response to individual faces in the right middle fusiform gyrus Christine Schiltz Bettina Sorger Roberto Caldara Fatima
Linking Brain Response and Behavior to Reveal Top-Down. (and Inside-Out) Influences on Processing in Face Perception. Heather A. Wild. Thomas A.
 Influences on Processing in Face Perception. Heather A. Wild. Thomas A.") Linking Brain Response and Behavior to Reveal Top-Down (and Inside-Out) Influences on Processing in Face Perception Heather A. Wild Thomas A. Busey Department of Psychology Indiana University Bloomington,
Linking Brain Response and Behavior to Reveal Top-Down (and Inside-Out) Influences on Processing in Face Perception Heather A. Wild Thomas A. Busey Department of Psychology Indiana University Bloomington,
The face-sensitive N170 encodes social category information Jonathan B. Freeman a, Nalini Ambady a and Phillip J. Holcomb a
 Neuroreport Freeman et al. N170 encodes social category information 1 The face-sensitive N170 encodes social category information Jonathan B. Freeman a, Nalini Ambady a and Phillip J. Holcomb a Prominent
Neuroreport Freeman et al. N170 encodes social category information 1 The face-sensitive N170 encodes social category information Jonathan B. Freeman a, Nalini Ambady a and Phillip J. Holcomb a Prominent
Dissociable neural correlates for familiarity and recollection during the encoding and retrieval of pictures
 Cognitive Brain Research 18 (2004) 255 272 Research report Dissociable neural correlates for familiarity and recollection during the encoding and retrieval of pictures Audrey Duarte a, *, Charan Ranganath
Cognitive Brain Research 18 (2004) 255 272 Research report Dissociable neural correlates for familiarity and recollection during the encoding and retrieval of pictures Audrey Duarte a, *, Charan Ranganath
THE EFFECT OF DIFFERENT TRAINING EXPERIENCES ON OBJECT RECOGNITION IN THE VISUAL SYSTEM. Alan Chun-Nang Wong
 THE EFFECT OF DIFFERENT TRAINING EXPERIENCES ON OBJECT RECOGNITION IN THE VISUAL SYSTEM By Alan Chun-Nang Wong Dissertation Submitted to the Faculty of the Graduate School of Vanderbilt University In partial
THE EFFECT OF DIFFERENT TRAINING EXPERIENCES ON OBJECT RECOGNITION IN THE VISUAL SYSTEM By Alan Chun-Nang Wong Dissertation Submitted to the Faculty of the Graduate School of Vanderbilt University In partial
The Role of Color and Attention in Fast Natural Scene Recognition
 Color and Fast Scene Recognition 1 The Role of Color and Attention in Fast Natural Scene Recognition Angela Chapman Department of Cognitive and Neural Systems Boston University 677 Beacon St. Boston, MA
Color and Fast Scene Recognition 1 The Role of Color and Attention in Fast Natural Scene Recognition Angela Chapman Department of Cognitive and Neural Systems Boston University 677 Beacon St. Boston, MA
Hierarchical attention in discriminating objects at different levels of specificity
 Perception & Psychophysics 2006, 68 (5), 845-860 Hierarchical attention in discriminating objects at different levels of specificity MARY-ELLEN LARGE and PATRICIA A. MCMULLEN Dalhousie University, Halifax,
Perception & Psychophysics 2006, 68 (5), 845-860 Hierarchical attention in discriminating objects at different levels of specificity MARY-ELLEN LARGE and PATRICIA A. MCMULLEN Dalhousie University, Halifax,
Manuscript under review for Psychological Science. Direct Electrophysiological Measurement of Attentional Templates in Visual Working Memory
 Direct Electrophysiological Measurement of Attentional Templates in Visual Working Memory Journal: Psychological Science Manuscript ID: PSCI-0-0.R Manuscript Type: Short report Date Submitted by the Author:
Direct Electrophysiological Measurement of Attentional Templates in Visual Working Memory Journal: Psychological Science Manuscript ID: PSCI-0-0.R Manuscript Type: Short report Date Submitted by the Author:
Expert image analysts show enhanced visual processing in change detection
 Psychonomic Bulletin & Review 9, 16 (), 39-397 doi:1.3758/pbr.16..39 Expert image analysts show enhanced visual processing in change detection Tim Curran University of Colorado, Boulder, Colorado Laurie
Psychonomic Bulletin & Review 9, 16 (), 39-397 doi:1.3758/pbr.16..39 Expert image analysts show enhanced visual processing in change detection Tim Curran University of Colorado, Boulder, Colorado Laurie
Virtual Brain Reading: A Connectionist Approach to Understanding fmri
 Virtual Brain Reading: A Connectionist Approach to Understanding fmri Rosemary A. Cowell (r.a.cowell@kent.ac.uk) Computing Laboratory, University of Kent, Canterbury, CT2 7NF, UK David E. Huber (dhuber@psy.ucsd.edu)
Virtual Brain Reading: A Connectionist Approach to Understanding fmri Rosemary A. Cowell (r.a.cowell@kent.ac.uk) Computing Laboratory, University of Kent, Canterbury, CT2 7NF, UK David E. Huber (dhuber@psy.ucsd.edu)
FFA: a flexible fusiform area for subordinate-level visual processing automatized by expertise
 commentary FFA: a flexible fusiform area for subordinate-level visual processing automatized by expertise Michael J. Tarr and Isabel Gauthier Much evidence suggests that the fusiform face area is involved
commentary FFA: a flexible fusiform area for subordinate-level visual processing automatized by expertise Michael J. Tarr and Isabel Gauthier Much evidence suggests that the fusiform face area is involved
Category specificity in early perception: face and word N170 responses differ in both lateralization and habituation properties
 HUMAN NEUROSCIENCE ORIGINAL RESEARCH ARTICLE published: 29 December 28 doi: 1.3389/neuro.9.18.28 : face and word N17 responses differ in both lateralization and habituation properties Urs Maurer 1,2, Bruno
HUMAN NEUROSCIENCE ORIGINAL RESEARCH ARTICLE published: 29 December 28 doi: 1.3389/neuro.9.18.28 : face and word N17 responses differ in both lateralization and habituation properties Urs Maurer 1,2, Bruno
ASSAF HAREL, PHD CURRICULUM VITAE Department of Psychology Wright State University
 Assaf Harel, August 2014 1 Contact Details 437 Fawcett Hall 3640 Colonel Glenn Highway Dayton, Ohio 45435 Email: assaf.harel@wright.edu Tel: (937) 775-3819 Cell: (240) 938-4020 ASSAF HAREL, PHD CURRICULUM
Assaf Harel, August 2014 1 Contact Details 437 Fawcett Hall 3640 Colonel Glenn Highway Dayton, Ohio 45435 Email: assaf.harel@wright.edu Tel: (937) 775-3819 Cell: (240) 938-4020 ASSAF HAREL, PHD CURRICULUM
Visual Context Dan O Shea Prof. Fei Fei Li, COS 598B
 Visual Context Dan O Shea Prof. Fei Fei Li, COS 598B Cortical Analysis of Visual Context Moshe Bar, Elissa Aminoff. 2003. Neuron, Volume 38, Issue 2, Pages 347 358. Visual objects in context Moshe Bar.
Visual Context Dan O Shea Prof. Fei Fei Li, COS 598B Cortical Analysis of Visual Context Moshe Bar, Elissa Aminoff. 2003. Neuron, Volume 38, Issue 2, Pages 347 358. Visual objects in context Moshe Bar.
Unraveling Mechanisms for Expert Object Recognition: Bridging Brain Activity and Behavior
 Journal of Experimental Psychology: Human Perception and Performance 2002, Vol. 28, No. 2, 431 446 Copyright 2002 by the American Psychological Association, Inc. 0096-1523/02/$5.00 DOI: 10.1037//0096-1523.28.2.431
Journal of Experimental Psychology: Human Perception and Performance 2002, Vol. 28, No. 2, 431 446 Copyright 2002 by the American Psychological Association, Inc. 0096-1523/02/$5.00 DOI: 10.1037//0096-1523.28.2.431
Brain Activation during Face Perception: Evidence of a Developmental Change
 Brain Activation during Face Perception: Evidence of a Developmental Change E. H. Aylward 1, J. E. Park 1, K. M. Field 1, A. C. Parsons 1, T. L. Richards 1, S. C. Cramer 2, and A. N. Meltzoff 1 Abstract
Brain Activation during Face Perception: Evidence of a Developmental Change E. H. Aylward 1, J. E. Park 1, K. M. Field 1, A. C. Parsons 1, T. L. Richards 1, S. C. Cramer 2, and A. N. Meltzoff 1 Abstract
Domain specificity versus expertise: factors influencing distinct processing of faces
 D. Carmel, S. Bentin / Cognition 83 (2002) 1 29 1 COGNITION Cognition 83 (2002) 1 29 www.elsevier.com/locate/cognit Domain specificity versus expertise: factors influencing distinct processing of faces
D. Carmel, S. Bentin / Cognition 83 (2002) 1 29 1 COGNITION Cognition 83 (2002) 1 29 www.elsevier.com/locate/cognit Domain specificity versus expertise: factors influencing distinct processing of faces
Neural Correlates of Complex Tone Processing and Hemispheric Asymmetry
 International Journal of Undergraduate Research and Creative Activities Volume 5 Article 3 June 2013 Neural Correlates of Complex Tone Processing and Hemispheric Asymmetry Whitney R. Arthur Central Washington
International Journal of Undergraduate Research and Creative Activities Volume 5 Article 3 June 2013 Neural Correlates of Complex Tone Processing and Hemispheric Asymmetry Whitney R. Arthur Central Washington
Journal of Experimental Psychology: Animal Behavior Processes
 Journal of Experimental Psychology: Animal Behavior Processes Perceptual Learning and Inversion Effects: Recognition of Prototype-Defined Familiar Checkerboards Ciro Civile, Di Zhao, Yixuan Ku, Heike Elchlepp,
Journal of Experimental Psychology: Animal Behavior Processes Perceptual Learning and Inversion Effects: Recognition of Prototype-Defined Familiar Checkerboards Ciro Civile, Di Zhao, Yixuan Ku, Heike Elchlepp,
BIOGRAPHICAL SKETCH. DEGREE (if applicable)
") NAME: Isabel Gauthier, Ph.D. OMB No. 0925-0001/0002 (Rev. 08/12 Approved Through 8/31/2015) BIOGRAPHICAL SKETCH Provide the following information for the Senior/key personnel and other significant contributors.
NAME: Isabel Gauthier, Ph.D. OMB No. 0925-0001/0002 (Rev. 08/12 Approved Through 8/31/2015) BIOGRAPHICAL SKETCH Provide the following information for the Senior/key personnel and other significant contributors.
Orientation Specific Effects of Automatic Access to Categorical Information in Biological Motion Perception
 Orientation Specific Effects of Automatic Access to Categorical Information in Biological Motion Perception Paul E. Hemeren (paul.hemeren@his.se) University of Skövde, School of Humanities and Informatics
Orientation Specific Effects of Automatic Access to Categorical Information in Biological Motion Perception Paul E. Hemeren (paul.hemeren@his.se) University of Skövde, School of Humanities and Informatics
Selective Attention Supports Working Memory Maintenance by Modulating Perceptual Processing of Distractors
 Selective Attention Supports Working Memory Maintenance by Modulating Perceptual Processing of Distractors Kartik K. Sreenivasan and Amishi P. Jha Abstract & Selective attention has been shown to bias
Selective Attention Supports Working Memory Maintenance by Modulating Perceptual Processing of Distractors Kartik K. Sreenivasan and Amishi P. Jha Abstract & Selective attention has been shown to bias
Early posterior ERP components do not reflect the control of attentional shifts toward expected peripheral events
 Psychophysiology, 40 (2003), 827 831. Blackwell Publishing Inc. Printed in the USA. Copyright r 2003 Society for Psychophysiological Research BRIEF REPT Early posterior ERP components do not reflect the
Psychophysiology, 40 (2003), 827 831. Blackwell Publishing Inc. Printed in the USA. Copyright r 2003 Society for Psychophysiological Research BRIEF REPT Early posterior ERP components do not reflect the
The face-sensitive N170 encodes social category information Jonathan B. Freeman, Nalini Ambady and Phillip J. Holcomb
 24 Cognitive neuroscience and neuropsychology The face-sensitive encodes social category information Jonathan B. Freeman, Nalini Ambady and Phillip J. Holcomb Prominent models of face perception posit
24 Cognitive neuroscience and neuropsychology The face-sensitive encodes social category information Jonathan B. Freeman, Nalini Ambady and Phillip J. Holcomb Prominent models of face perception posit
RUNNNING HEAD: Pigeons Attend to Features of Human Faces. Pigeons and People Categorize Human Faces Using Similar Features
 RUNNNING HEAD: Pigeons Attend to Features of Human Faces Pigeons and People Categorize Human Faces Using Similar Features Brett M. Gibson *, Edward A. Wasserman *, Frédéric Gosselin, and Philippe G. Schyns
RUNNNING HEAD: Pigeons Attend to Features of Human Faces Pigeons and People Categorize Human Faces Using Similar Features Brett M. Gibson *, Edward A. Wasserman *, Frédéric Gosselin, and Philippe G. Schyns
N250 ERP Correlates of the Acquisition of Face Representations across Different Images
 N250 ERP Correlates of the Acquisition of Face Representations across Different Images Jürgen M. Kaufmann 1,2, Stefan R. Schweinberger 1,2, and A. Mike Burton 2 Abstract & We used ERPs to investigate neural
N250 ERP Correlates of the Acquisition of Face Representations across Different Images Jürgen M. Kaufmann 1,2, Stefan R. Schweinberger 1,2, and A. Mike Burton 2 Abstract & We used ERPs to investigate neural
The functional organization of the ventral visual pathway and its relationship to object recognition
 Kanwisher-08 9/16/03 9:27 AM Page 169 Chapter 8 The functional organization of the ventral visual pathway and its relationship to object recognition Kalanit Grill-Spector Abstract Humans recognize objects
Kanwisher-08 9/16/03 9:27 AM Page 169 Chapter 8 The functional organization of the ventral visual pathway and its relationship to object recognition Kalanit Grill-Spector Abstract Humans recognize objects
Summary. Multiple Body Representations 11/6/2016. Visual Processing of Bodies. The Body is:
 Visual Processing of Bodies Corps et cognition: l'embodiment Corrado Corradi-Dell Acqua corrado.corradi@unige.ch Theory of Pain Laboratory Summary Visual Processing of Bodies Category-specificity in ventral
Visual Processing of Bodies Corps et cognition: l'embodiment Corrado Corradi-Dell Acqua corrado.corradi@unige.ch Theory of Pain Laboratory Summary Visual Processing of Bodies Category-specificity in ventral
Long-term Expertise with Artificial Objects Increases Visual Competition with Early Face Categorization Processes
 Long-term Expertise with Artificial Objects Increases Visual Competition with Early Face Categorization Processes Bruno Rossion 1, Daniel Collins 2, Valérie Goffaux 1, and Tim Curran 2 Abstract & The degree
Long-term Expertise with Artificial Objects Increases Visual Competition with Early Face Categorization Processes Bruno Rossion 1, Daniel Collins 2, Valérie Goffaux 1, and Tim Curran 2 Abstract & The degree
Vision Research 51 (2011) Contents lists available at ScienceDirect. Vision Research. journal homepage:
 Contents lists available at ScienceDirect. Vision Research. journal homepage:") Vision Research 51 (2011) 195 202 Contents lists available at ScienceDirect Vision Research journal homepage: www.elsevier.com/locate/visres Differences between perception of human faces and body shapes:
Vision Research 51 (2011) 195 202 Contents lists available at ScienceDirect Vision Research journal homepage: www.elsevier.com/locate/visres Differences between perception of human faces and body shapes:
Studying the time course of sensory substitution mechanisms (CSAIL, 2014)
") Studying the time course of sensory substitution mechanisms (CSAIL, 2014) Christian Graulty, Orestis Papaioannou, Phoebe Bauer, Michael Pitts & Enriqueta Canseco-Gonzalez, Reed College. Funded by the Murdoch
Studying the time course of sensory substitution mechanisms (CSAIL, 2014) Christian Graulty, Orestis Papaioannou, Phoebe Bauer, Michael Pitts & Enriqueta Canseco-Gonzalez, Reed College. Funded by the Murdoch
Outline of Talk. Introduction to EEG and Event Related Potentials. Key points. My path to EEG
 Outline of Talk Introduction to EEG and Event Related Potentials Shafali Spurling Jeste Assistant Professor in Psychiatry and Neurology UCLA Center for Autism Research and Treatment Basic definitions and
Outline of Talk Introduction to EEG and Event Related Potentials Shafali Spurling Jeste Assistant Professor in Psychiatry and Neurology UCLA Center for Autism Research and Treatment Basic definitions and
Reward prediction error signals associated with a modified time estimation task
 Psychophysiology, 44 (2007), 913 917. Blackwell Publishing Inc. Printed in the USA. Copyright r 2007 Society for Psychophysiological Research DOI: 10.1111/j.1469-8986.2007.00561.x BRIEF REPORT Reward prediction
Psychophysiology, 44 (2007), 913 917. Blackwell Publishing Inc. Printed in the USA. Copyright r 2007 Society for Psychophysiological Research DOI: 10.1111/j.1469-8986.2007.00561.x BRIEF REPORT Reward prediction
Description of the Spectro-temporal unfolding of temporal orienting of attention.
 Description of the Spectro-temporal unfolding of temporal orienting of attention. All behaviors unfold over time; therefore, our ability to perceive and adapt our behavior according to the temporal constraints
Description of the Spectro-temporal unfolding of temporal orienting of attention. All behaviors unfold over time; therefore, our ability to perceive and adapt our behavior according to the temporal constraints
The preferred level of face categorization depends on discriminability
 Psychonomic Bulletin & Review 2008, 15 (3), 623-629 doi: 10.3758/PBR.15.3.623 The preferred level of face categorization depends on discriminability Christopher D Lauro University of Colorado, Boulder,
Psychonomic Bulletin & Review 2008, 15 (3), 623-629 doi: 10.3758/PBR.15.3.623 The preferred level of face categorization depends on discriminability Christopher D Lauro University of Colorado, Boulder,
Recognizing partially visible objects
 Vision Research 45 (2005) 1807 1814 www.elsevier.com/locate/visres Recognizing partially visible objects Philip Servos a, *, Elizabeth S. Olds a, Peggy J. Planetta a, G. Keith Humphrey b a Department of
Vision Research 45 (2005) 1807 1814 www.elsevier.com/locate/visres Recognizing partially visible objects Philip Servos a, *, Elizabeth S. Olds a, Peggy J. Planetta a, G. Keith Humphrey b a Department of
FAILURES OF OBJECT RECOGNITION. Dr. Walter S. Marcantoni
 FAILURES OF OBJECT RECOGNITION Dr. Walter S. Marcantoni VISUAL AGNOSIA -damage to the extrastriate visual regions (occipital, parietal and temporal lobes) disrupts recognition of complex visual stimuli
FAILURES OF OBJECT RECOGNITION Dr. Walter S. Marcantoni VISUAL AGNOSIA -damage to the extrastriate visual regions (occipital, parietal and temporal lobes) disrupts recognition of complex visual stimuli
Title of Thesis. Study on Audiovisual Integration in Young and Elderly Adults by Event-Related Potential
 Title of Thesis Study on Audiovisual Integration in Young and Elderly Adults by Event-Related Potential 2014 September Yang Weiping The Graduate School of Natural Science and Technology (Doctor s Course)
Title of Thesis Study on Audiovisual Integration in Young and Elderly Adults by Event-Related Potential 2014 September Yang Weiping The Graduate School of Natural Science and Technology (Doctor s Course)
The Other-Race Effect and its Influences on the Development of Emotion Processing
 University of Massachusetts Amherst ScholarWorks@UMass Amherst Masters Theses 1911 - February 2014 2009 The Other-Race Effect and its Influences on the Development of Emotion Processing Alexandra Monesson
University of Massachusetts Amherst ScholarWorks@UMass Amherst Masters Theses 1911 - February 2014 2009 The Other-Race Effect and its Influences on the Development of Emotion Processing Alexandra Monesson
Evaluating Models of Object-Decision Priming: Evidence From Event-Related Potential Repetition Effects
 Journal of Experimental Psychology: Learning, Memory, and Cognition 2006, Vol. 32, No. 2, 230 248 Copyright 2006 by the American Psychological Association 0278-7393/06/$12.00 DOI: 10.1037/0278-7393.32.2.230
Journal of Experimental Psychology: Learning, Memory, and Cognition 2006, Vol. 32, No. 2, 230 248 Copyright 2006 by the American Psychological Association 0278-7393/06/$12.00 DOI: 10.1037/0278-7393.32.2.230
Chapter 5. Summary and Conclusions! 131
 ! Chapter 5 Summary and Conclusions! 131 Chapter 5!!!! Summary of the main findings The present thesis investigated the sensory representation of natural sounds in the human auditory cortex. Specifically,
! Chapter 5 Summary and Conclusions! 131 Chapter 5!!!! Summary of the main findings The present thesis investigated the sensory representation of natural sounds in the human auditory cortex. Specifically,
A picture is worth thousands of trials: rendering the use of visual information from spiking neurons to recognition
 Cognitive Science 28 (2004) 141 146 A picture is worth thousands of trials: rendering the use of visual information from spiking neurons to recognition Frédéric Gosselin a,, Philippe G. Schyns b a Département
Cognitive Science 28 (2004) 141 146 A picture is worth thousands of trials: rendering the use of visual information from spiking neurons to recognition Frédéric Gosselin a,, Philippe G. Schyns b a Département
Differences of Face and Object Recognition in Utilizing Early Visual Information
 Differences of Face and Object Recognition in Utilizing Early Visual Information Peter Kalocsai and Irving Biederman Department of Psychology and Computer Science University of Southern California Los
Differences of Face and Object Recognition in Utilizing Early Visual Information Peter Kalocsai and Irving Biederman Department of Psychology and Computer Science University of Southern California Los
Familiar-face recognition and comparison: source analysis of scalp-recorded event-related potentials
 Clinical Neurophysiology 115 (2004) 880 886 www.elsevier.com/locate/clinph Familiar-face recognition and comparison: source analysis of scalp-recorded event-related potentials Elena V. Mnatsakanian a,b,
Clinical Neurophysiology 115 (2004) 880 886 www.elsevier.com/locate/clinph Familiar-face recognition and comparison: source analysis of scalp-recorded event-related potentials Elena V. Mnatsakanian a,b,
Transcranial direct current stimulation modulates shifts in global/local attention
 University of New Mexico UNM Digital Repository Psychology ETDs Electronic Theses and Dissertations 2-9-2010 Transcranial direct current stimulation modulates shifts in global/local attention David B.
University of New Mexico UNM Digital Repository Psychology ETDs Electronic Theses and Dissertations 2-9-2010 Transcranial direct current stimulation modulates shifts in global/local attention David B.
The Meaning of the Mask Matters
 PSYCHOLOGICAL SCIENCE Research Report The Meaning of the Mask Matters Evidence of Conceptual Interference in the Attentional Blink Paul E. Dux and Veronika Coltheart Macquarie Centre for Cognitive Science,
PSYCHOLOGICAL SCIENCE Research Report The Meaning of the Mask Matters Evidence of Conceptual Interference in the Attentional Blink Paul E. Dux and Veronika Coltheart Macquarie Centre for Cognitive Science,
Biomedical Research 2013; 24 (3): ISSN X
: ISSN X") Biomedical Research 2013; 24 (3): 359-364 ISSN 0970-938X http://www.biomedres.info Investigating relative strengths and positions of electrical activity in the left and right hemispheres of the human brain
Biomedical Research 2013; 24 (3): 359-364 ISSN 0970-938X http://www.biomedres.info Investigating relative strengths and positions of electrical activity in the left and right hemispheres of the human brain
Mutual information of image fragments predicts categorization in humans: Electrophysiological and behavioral evidence q
 Vision Research 47 (2007) 2010 2020 www.elsevier.com/locate/visres Mutual information of image fragments predicts categorization in humans: Electrophysiological and behavioral evidence q Assaf Harel a,
Vision Research 47 (2007) 2010 2020 www.elsevier.com/locate/visres Mutual information of image fragments predicts categorization in humans: Electrophysiological and behavioral evidence q Assaf Harel a,
Figure 1. Source localization results for the No Go N2 component. (a) Dipole modeling
 Dipole modeling") Supplementary materials 1 Figure 1. Source localization results for the No Go N2 component. (a) Dipole modeling analyses placed the source of the No Go N2 component in the dorsal ACC, near the ACC source
Supplementary materials 1 Figure 1. Source localization results for the No Go N2 component. (a) Dipole modeling analyses placed the source of the No Go N2 component in the dorsal ACC, near the ACC source
Experience-Dependent Sharpening of Visual Shape Selectivity in Inferior Temporal Cortex
 Cerebral Cortex Advance Access published December 28, 2005 Cerebral Cortex doi:10.1093/cercor/bhj100 Experience-Dependent Sharpening of Visual Shape Selectivity in Inferior Temporal Cortex David J. Freedman
Cerebral Cortex Advance Access published December 28, 2005 Cerebral Cortex doi:10.1093/cercor/bhj100 Experience-Dependent Sharpening of Visual Shape Selectivity in Inferior Temporal Cortex David J. Freedman
Cognition xx (0000) xxx xxx. Brief article. A holistic account of the own-race effect in face recognition: evidence from a cross-cultural study
 xxx xxx. Brief article. A holistic account of the own-race effect in face recognition: evidence from a cross-cultural study") 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 Brief article A holistic account of the own-race effect in face recognition:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 Brief article A holistic account of the own-race effect in face recognition:
For Non-Commercial Academic Use Only
 For Non-Commercial Academic Use Only Neuropsychologia 48 (2010) 3743 3756 Contents lists available at ScienceDirect Neuropsychologia journal homepage: www.elsevier.com/locate/neuropsychologia Adaptation
For Non-Commercial Academic Use Only Neuropsychologia 48 (2010) 3743 3756 Contents lists available at ScienceDirect Neuropsychologia journal homepage: www.elsevier.com/locate/neuropsychologia Adaptation
John E. Richards* Department of Psychology, University of South Carolina, Columbia, SC 29208, United States
 International Journal of Psychophysiology 54 (2004) 201 220 www.elsevier.com/locate/ijpsycho Recovering dipole sources from scalp-recorded event-related-potentials using component analysis: principal component
International Journal of Psychophysiology 54 (2004) 201 220 www.elsevier.com/locate/ijpsycho Recovering dipole sources from scalp-recorded event-related-potentials using component analysis: principal component
Event-Related Potentials Recorded during Human-Computer Interaction
 Proceedings of the First International Conference on Complex Medical Engineering (CME2005) May 15-18, 2005, Takamatsu, Japan (Organized Session No. 20). Paper No. 150, pp. 715-719. Event-Related Potentials
Proceedings of the First International Conference on Complex Medical Engineering (CME2005) May 15-18, 2005, Takamatsu, Japan (Organized Session No. 20). Paper No. 150, pp. 715-719. Event-Related Potentials
EEG/ERP Control Process Separation
 EEG/ERP Control Process Separation Gwen A. Frishkoff & Don M. Tucker It may be possible to develop monitoring devices to provide realtime readout of activity within and between subsystems of the control
EEG/ERP Control Process Separation Gwen A. Frishkoff & Don M. Tucker It may be possible to develop monitoring devices to provide realtime readout of activity within and between subsystems of the control
Tracking the Development of Automaticity in Memory Search with Human Electrophysiology
 Tracking the Development of Automaticity in Memory Search with Human Electrophysiology Rui Cao (caorui.beilia@gmail.com) Thomas A. Busey (busey@indiana.edu) Robert M. Nosofsky (nosofsky@indiana.edu) Richard
Tracking the Development of Automaticity in Memory Search with Human Electrophysiology Rui Cao (caorui.beilia@gmail.com) Thomas A. Busey (busey@indiana.edu) Robert M. Nosofsky (nosofsky@indiana.edu) Richard
Working Memory Load for Faces Modulates P300, N170, and N250r
 Working Memory Load for Faces Modulates P300, N170, and N250r Helen M. Morgan, Christoph Klein, Stephan G. Boehm, Kimron L. Shapiro, and David E. J. Linden Abstract & We used event-related potential (ERP)
Working Memory Load for Faces Modulates P300, N170, and N250r Helen M. Morgan, Christoph Klein, Stephan G. Boehm, Kimron L. Shapiro, and David E. J. Linden Abstract & We used event-related potential (ERP)
Neural correlates of short-term perceptual learning in orientation discrimination indexed by event-related potentials
 Chinese Science Bulletin 2007 Science in China Press Springer-Verlag Neural correlates of short-term perceptual learning in orientation discrimination indexed by event-related potentials SONG Yan 1, PENG
Chinese Science Bulletin 2007 Science in China Press Springer-Verlag Neural correlates of short-term perceptual learning in orientation discrimination indexed by event-related potentials SONG Yan 1, PENG
Frank Tong. Department of Psychology Green Hall Princeton University Princeton, NJ 08544
 Frank Tong Department of Psychology Green Hall Princeton University Princeton, NJ 08544 Office: Room 3-N-2B Telephone: 609-258-2652 Fax: 609-258-1113 Email: ftong@princeton.edu Graduate School Applicants
Frank Tong Department of Psychology Green Hall Princeton University Princeton, NJ 08544 Office: Room 3-N-2B Telephone: 609-258-2652 Fax: 609-258-1113 Email: ftong@princeton.edu Graduate School Applicants
Quiroga, R. Q., Reddy, L., Kreiman, G., Koch, C., Fried, I. (2005). Invariant visual representation by single neurons in the human brain, Nature,
. Invariant visual representation by single neurons in the human brain, Nature,") Quiroga, R. Q., Reddy, L., Kreiman, G., Koch, C., Fried, I. (2005). Invariant visual representation by single neurons in the human brain, Nature, Vol. 435, pp. 1102-7. Sander Vaus 22.04.2015 The study
Quiroga, R. Q., Reddy, L., Kreiman, G., Koch, C., Fried, I. (2005). Invariant visual representation by single neurons in the human brain, Nature, Vol. 435, pp. 1102-7. Sander Vaus 22.04.2015 The study
An Introduction to Translational Neuroscience Approaches to Investigating Autism
 An Introduction to Translational Neuroscience Approaches to Investigating Autism Gabriel S. Dichter, PhD Departments of Psychiatry & Psychology, UNC-Chapel Hill Carolina Institute for Developmental Disabilities
An Introduction to Translational Neuroscience Approaches to Investigating Autism Gabriel S. Dichter, PhD Departments of Psychiatry & Psychology, UNC-Chapel Hill Carolina Institute for Developmental Disabilities
From feedforward vision to natural vision: The impact of free viewing and clutter on monkey inferior temporal object representations
 From feedforward vision to natural vision: The impact of free viewing and clutter on monkey inferior temporal object representations James DiCarlo The McGovern Institute for Brain Research Department of
From feedforward vision to natural vision: The impact of free viewing and clutter on monkey inferior temporal object representations James DiCarlo The McGovern Institute for Brain Research Department of
Inversion and contrast-reversal effects on face processing assessed by MEG
 available at www.sciencedirect.com www.elsevier.com/locate/brainres Research Report Inversion and contrast-reversal effects on face processing assessed by MEG Roxane J. Itier a,, Anthony T. Herdman b,
available at www.sciencedirect.com www.elsevier.com/locate/brainres Research Report Inversion and contrast-reversal effects on face processing assessed by MEG Roxane J. Itier a,, Anthony T. Herdman b,
Top down processing of faces in human brain: A Behavioral Study
 Top down processing of faces in human brain: A Behavioral Study The MIT Faculty has made this article openly available. Please share how this access benefits you. Your story matters. Citation As Published
Top down processing of faces in human brain: A Behavioral Study The MIT Faculty has made this article openly available. Please share how this access benefits you. Your story matters. Citation As Published
Identifying the ERP correlate of a recognition memory search attempt
 Cognitive Brain Research 24 (2005) 674 684 Research Report Identifying the ERP correlate of a recognition memory search attempt Rachel A. Diana a, *, Kaia L. Vilberg b, Lynne M. Reder a a Department of
Cognitive Brain Research 24 (2005) 674 684 Research Report Identifying the ERP correlate of a recognition memory search attempt Rachel A. Diana a, *, Kaia L. Vilberg b, Lynne M. Reder a a Department of
Attention Response Functions: Characterizing Brain Areas Using fmri Activation during Parametric Variations of Attentional Load
 Attention Response Functions: Characterizing Brain Areas Using fmri Activation during Parametric Variations of Attentional Load Intro Examine attention response functions Compare an attention-demanding
Attention Response Functions: Characterizing Brain Areas Using fmri Activation during Parametric Variations of Attentional Load Intro Examine attention response functions Compare an attention-demanding
COGS 101A: Sensation and Perception
 COGS 101A: Sensation and Perception 1 Virginia R. de Sa Department of Cognitive Science UCSD Lecture 6: Beyond V1 - Extrastriate cortex Chapter 4 Course Information 2 Class web page: http://cogsci.ucsd.edu/
COGS 101A: Sensation and Perception 1 Virginia R. de Sa Department of Cognitive Science UCSD Lecture 6: Beyond V1 - Extrastriate cortex Chapter 4 Course Information 2 Class web page: http://cogsci.ucsd.edu/
Visual Selection and Attention
 Visual Selection and Attention Retrieve Information Select what to observe No time to focus on every object Overt Selections Performed by eye movements Covert Selections Performed by visual attention 2
Visual Selection and Attention Retrieve Information Select what to observe No time to focus on every object Overt Selections Performed by eye movements Covert Selections Performed by visual attention 2
CS/NEUR125 Brains, Minds, and Machines. Due: Friday, April 14
 CS/NEUR125 Brains, Minds, and Machines Assignment 5: Neural mechanisms of object-based attention Due: Friday, April 14 This Assignment is a guided reading of the 2014 paper, Neural Mechanisms of Object-Based
CS/NEUR125 Brains, Minds, and Machines Assignment 5: Neural mechanisms of object-based attention Due: Friday, April 14 This Assignment is a guided reading of the 2014 paper, Neural Mechanisms of Object-Based
The Role of Working Memory in Visual Selective Attention
 Goldsmiths Research Online. The Authors. Originally published: Science vol.291 2 March 2001 1803-1806. http://www.sciencemag.org. 11 October 2000; accepted 17 January 2001 The Role of Working Memory in
Goldsmiths Research Online. The Authors. Originally published: Science vol.291 2 March 2001 1803-1806. http://www.sciencemag.org. 11 October 2000; accepted 17 January 2001 The Role of Working Memory in