Neuronal and Epithelial Cell Rescue Resolves Chronic Systemic. Inflammation in the Lipid Storage Disorder Niemann-Pick C
|
|
|
- Roger Wheeler
- 5 years ago
- Views:
Transcription
1 HMG Advance Access published April 6, Neuronal and Epithelial Cell Rescue Resolves Chronic Systemic Inflammation in the Lipid Storage Disorder Niemann-Pick C Manuel E. Lopez, Andrés D. Klein, Jennifer Hong, Ubah J. Dimbil, and Matthew P. Scott* Departments of Developmental Biology, Genetics, and Bioengineering, Howard Hughes Medical Institute, Stanford University School of Medicine Clark Center, 318 Campus Drive, Stanford, California, U.S.A Corresponding Author: Dr. Matthew Scott Stanford University, Clark Center W200, 318 Campus Dr., Stanford, CA (650) (telephone) (650) (fax) mscott@stanford.edu The Author Published by Oxford University Press. This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License ( which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
2 2 Abstract Chronic systemic inflammation is thought to be a major contributor to metabolic and neurodegenerative diseases. Since inflammatory components are shared among different disorders, targeting inflammation is an attractive option for mitigating disease. To test the significance of inflammation in the lipid storage disorder (LSD) Niemann-Pick C (NPC), we deleted the macrophage inflammatory gene Mip1a/Ccl3 from NPC diseased mice. Deletion of Ccl3 had been reported to delay neuronal loss in Sandhoff LSD mice by inhibiting macrophage infiltration. For NPC mice, in contrast, deleting Ccl3 did not retard neurodegeneration and worsened the clinical outcome. Depletion of visceral tissue macrophages also did not alter CNS pathology and instead increased liver injury, suggesting a limited macrophage infiltration response into the CNS and a beneficial role of macrophage activity in visceral tissue. Prevention of neuron loss or liver injury, even at late stages in the disease, was achieved through specific rescue of NPC disease in neurons or in liver epithelial cells respectively. Local epithelial cell correction was also sufficient to reduce the macrophage-associated pathology in lung tissue. These results demonstrate that elevated inflammation and macrophage activity does not necessarily contribute to neurodegeneration and tissue injury, and LSD defects in immune cells may not preclude an appropriate inflammatory response. We conclude that inflammation remains secondary to neuronal and epithelial cell dysfunction and does not irreversibly contribute to the pathogenic cascade in NPC disease. Without further exploration of possible beneficial roles of inflammatory mediators, targeting inflammation may not be therapeutically effective at ameliorating disease severity.
3 3 Introduction The continuous elevation of inflammatory cytokines, lysosomal defects in microglia and macrophages, and other immune system irregularities have been proposed to play important roles in promoting tissue damage and neurodegeneration in LSDs (1-6). If the inflammatory process is harmful, therapeutically targeting immune cells and inflammatory mediators may provide a way to mitigate disease progression for these devastating and as yet incurable diseases. Here, we explore the role of inflammation in the severity and progression of the neurodegenerative lysosomal storage disease Niemann-Pick C (NPC). NPC is a rare complex metabolic LSD that leads to progressive deterioration of the nervous system and multiple organ systems in the body (7). Though rare, NPC shares many molecular and pathological mechanisms with more common unrelated disorders. Patients with NPC have changes in brain biomarkers that are associated with Alzheimer s disease (8) and suffer multiple neurological symptoms, such as dementia, dystonia, seizures, and psychiatric disorders. NPC also exhibits a number of immune cell phenotypes. Macrophage foam cells are a common pathological occurrence in NPC (9) and are well known for their role in atherosclerosis, where they form plaques in the circulatory system (10). Mouse studies of NPC have shown impaired natural killer T cell development (11), as well as abnormal production of complement components that are involved in the neuroinflammatory process of many neurodegenerative diseases (12).
4 4 Knowledge obtained from studying NPC pathology will likely aid in understanding pathological processes of other LSDs and of neurodegenerative diseases with similar pathologies. NPC disease is caused by genetic loss of NPC1 function resulting in defective intracellular lipid flux and accumulation of lysosomal material that is potentially deleterious to cells. NPC1 is a 13-pass endosomal/lysosomal membrane protein that binds cholesterol, and more strongly oxysterols, in vitro (13, 14). While most cases of NPC disease are caused by NPC1 gene mutations, NPC2 loss of function causes a nearly identical disease. Unlike NPC1, NPC2 is a soluble secreted sterol-binding protein that can be found in the lumens of endosomes and lysosomes (15). It has been hypothesized that both proteins shuttle cholesterol between each other and endosomal/lysosomal membranes to facilitate the exit of free cholesterol from the lysosome (16, 17). A defect in this process alters intracellular lipid homeostasis, membrane properties, and proper intracellular trafficking of organelles (18-20). The consequent accumulation of lysosomal material, e.g. sterols, gangliosides, and sphingolipids, is seen in virtually every cell in the body. Despite a ubiquitous storage defect, only certain cell types, such as neurons and hepatocytes, are particularly susceptible to dysfunction and injury. Both NPC proteins are also essential for proper macrophage and lymphocyte maturation and function (11, 21-23), but whether defects in immune cells relate to the
5 5 pathogenesis of NPC disease is unclear. It is reasonable to believe that aberrant or excessive inflammatory responses elicited by NPC defects may contribute to disease pathology and progression. Although the genetic causes of NPC are well understood, how best to mitigate disease pathology remains unknown. We set out to determine to what extent immune cell activity influences NPC disease progression. Investigations into NPC disease biology are facilitated by naturally occurring mammalian models that recapitulate the human condition (24, 25). In our effort to control inflammation and correct NPC disease pathology, we pursued various complementary approaches using the Npc1 -/- mouse model of the disease, also known as npc nih (24). To determine the contribution of macrophage and microglial-mediated inflammation to the severity of NPC disease, we genetically deleted the macrophage inflammatory protein 1alpha gene, Mip1a/Ccl3. CCL3 protein is a known pro-inflammatory cytokine that can mediate chemotaxis of monocytes and promote cell degranulation (26). Ccl3 expression is elevated in the CNS and visceral tissue of many LSDs, including Sandhoff disease, a prototypical autoimmune-like LSD (27-32). The deletion of Ccl3 in a mouse model of Sandhoff disease resulted in reduced monocyte-associated pathology in the brain, delayed neurodegeneration, and prolonged health (31). Sandhoff and NPC should share common pathological mechanisms since both disorders cause
6 6 ganglioside storage defects and immune cell impairment (33). If the NPC pathogenic cascade behaves the same as in Sandhoff disease, deletion of Ccl3 should be beneficial to Npc1 -/- mice. In a second approach, we determine whether the inflammatory response and disease pathology is refractory to restoration of NPC1 function after tissue damage has been established. The findings obtained are critically important with respect to determining the timing of treatments and in elucidating what cell types are necessary to target to halt disease progression and elicit recovery. Previously, it was postulated that the failure of current treatments such as cyclodextrin (CYCLO) to correct the pathology of tissues such as the lung was due to the inability of CYCLO to reach lung macrophages (27, 34). This reasoning assumed that the activities of macrophages, driven by their own NPC storage defects, continuously promote cell death and tissue injury in LSDs. To address these issues, we used an established cell-specific transgene form of gene therapy to genetically manipulate the production of NPC1 with cell and tissue specificity in otherwise Npc1 -/- mice. In the past, we exploited the cellautonomous nature of Npc1 -/- defects in mice to show that neuron-specific rescue, but not astrocyte-specific rescue, prevented or delayed neurological disease signs (35). In this report, we used features of the engineered mice that allowed control of timing of NPC rescue. This temporal and cell-type-specific rescue system allowed us to assess the importance of correcting the NPC defect in Npc1 -/- macrophages and microglia by determining whether restoring NPC1
7 7 function only in neurons and epithelial cells, even late in the disease, would be sufficient to halt cell injury and lessen the inflammatory response. Together our studies demonstrate that inflammatory mediators such as cytokines and macrophages have distinct disease relevance even in LSDs with similar storage defects. Perturbing part of the inflammatory pathway did not improve the health of mice with NPC disease even though the same procedure was successful for mice with Sandhoff disease. We propose that the role of inflammation in NPC disease is potentially beneficial and should be mainly viewed as a reaction to local neuron and epithelial cell defects since disease pathology can be halted or reversed by rescuing these cell types. Our conclusions highlight the importance of determining the contribution of each cell type to the clinical state of a particular storage disorder in order to better justify whether or not to pursue certain therapeutic avenues. The findings also have implications for further classifying LSDs according to whether macrophage activity and inflammatory mediators cause or exacerbate cell injury. Results Elevated regional Ccl3 expression in NPC diseased mice Elevated expression of genes encoding pro-inflammatory mediators has been previously observed in brains of Npc1 -/- mice at 21 days of age, and correlates well with increased glial-mediated inflammation (12). Here we show that at 50 days of age, preceding a notable decline in mouse weight that indicates
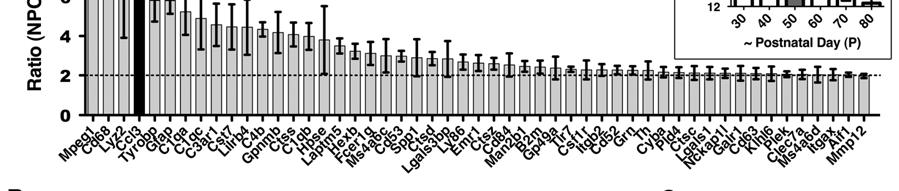
8 8 deteriorating health, the expression of Ccl3 was ranked using microarrays as one of the most elevated inflammatory mediators in Npc1 -/- mouse cerebella (Figure 1A). Significance analysis of microarrays (SAM) (36), with the incorporation of array data from age P71 mice, further identified the increase in Ccl3 expression as significant (q-value=0) and among the top 18 significantly elevated genes listed, with a false discovery rate of 5.18% (Supplemental Figure S1). In addition to Ccl3 there is elevated expression of complement pathway components, other immune factors, proteases, and lysosomal related protein genes at P50 (Figure 3). Many of these genes are common markers of inflammation and their expression is increased in other LSDs and in neurodegenerative diseases, e.g. the LSDs Sandhoff (37) and Gaucher (38), and Alzheimers (39). Cytokine Ccl3 mrna was elevated in Npc1 -/- mice compared to Npc1 +/- mice in regions of the brain that have the earliest and greatest signs of neurodegeneration and microglial-activity (Figure 1B). The cerebellum and thalamus are the most visibly affected brain structures and have the earliest onset of neuronal loss (35). The brainstem shows a gradual increase in neuroinflammation at later ages. The cortex does not have microglia invasion as early or severe as in other brain areas, even though considerable lipid accumulation is apparent in neurons there (35). Ccl3 levels remain low in these regions (Figure 1B). In the viscera, elevated Ccl3 mrna levels can be readily detected in various tissues from Npc1 -/- mice including the liver, an organ commonly affected by NPC disease.
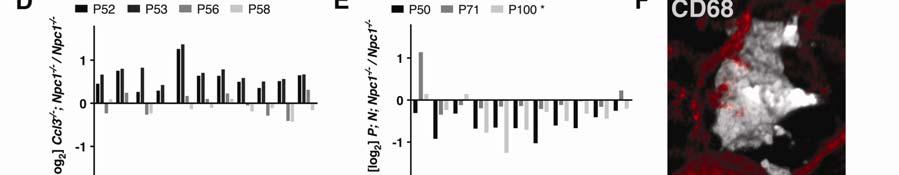
9 9 To identify the major producers of Ccl3 in the body, we examined isolated peritoneal cells, which are primarily macrophages and a few lymphocyte types (40). We also examined tissue from spleen, the predominant storage center for macrophages as well as a minor hematopoietic organ in adult mice (41). Levels of Ccl3 mrna were elevated in peritoneal cells and spleen from Npc1 -/- mice compared to Npc1 +/- mice (Figure 1B). Based on these expression profiles, it is likely that CCL3 protein is mainly produced by microglia and macrophages in the brain and visceral tissue of Npc1 -/- mice, as expected (27, 38, 42). Based on the observation that regions of increased Ccl3 expression correspond to areas of severe brain pathology, we next asked whether Ccl3 and the inflammatory process mediated by microglia and macrophage is important to NPC pathology. Data from the Ccl3 gene deletion study using the Hexb -/- mouse model of Sandhoff disease suggested that CCL3 would likewise be a contributing factor in NPC disease. Alternatively, CCL3 may have a neutral or beneficial role in the disease pathology. Deletion of Ccl3 worsens the NPC disease outcome in mice To determine if CCL3 is required for disease progression in Npc1 -/- mice, we crossed the same Ccl3 -/- mouse strain (43) previously used in the Sandhoff study to Npc1 -/- mice. As a standard for neuron rescue we measured weights of P60 male mice that were Npc1 -/- but carried a transgene that rescued Purkinje
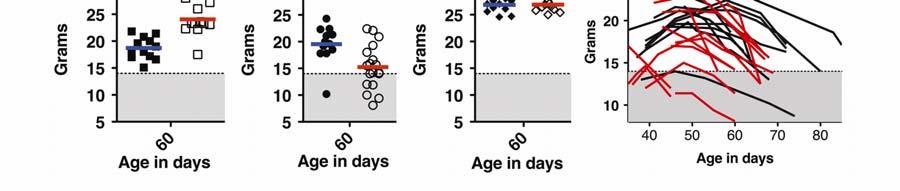
10 10 neurons specifically (Figure 2A). These mice were generated, as detailed in a prior study (35), by mating a Npc1 +/- mouse homozygous for a Tet-inducible Npc1 transgene, teto-npc1-yfp (N), with a Npc1 +/- mouse homozygous for the cerebellar Purkinje neuron-specific driver Pcp2-tTA (P). The resulting P; N; Npc1 - /- offspring produced NPC1-YFP protein in cerebellar Purkinje neurons in an otherwise NPC1-deficient animal. This Purkinje neuron-specific correction of NPC disorder elicited temporary but significant health benefits, such as weight gain, relative to non-purkinje rescued Npc1 -/- mice. In contrast to Purkinje neuron-rescued mice, male Npc1 -/- mice carrying the Ccl3 deletion did not show any improvements in weight (Figure 2B). Ccl3 -/- ; Npc1 -/- mice instead were more likely to exhibit lower weight profiles than Npc1 -/- mice and had to be euthanized at ages earlier than P70 (Figure 2D). A trend towards early death was also noticed for female mice (data not shown), though differences in weight were less obvious, possibly because young female mice do not exhibit as much weight gain as young males. At around age P60, the average weight difference between female Npc1 -/- mice (n=24) and Ccl3 -/- ; Npc1 -/- mice (n=13) was not as striking (P=0.3237, t-test) as the difference between male Npc1 -/- mice (n=12) and Ccl3 -/- ; Npc1 -/- mice (n=18) (Figure 2B; P=0.0076, t- test). Two-way ANOVA analysis of a cohort of 9 male and 5 female pairs further verified that between the ages of 45-60, weight curves of female Npc1 -/- and Ccl3 -/- ; Npc1 -/- mice are not significantly different (P=0.2028) but weight curves of male mice are (P=0.0045).
11 11 The lower weight phenotype and earlier deaths of Ccl3 -/- Npc1 -/- mice was unexpected. This worsening disease profile may be due to an inherent deleterious effect of Ccl3 -/-. To control for the effects of Ccl3 -/- on non-diseased mice, we compared the weights of Ccl3 -/- ; Npc1 +/- mice and Npc1 +/- P60 male mice. No gross weight differences were seen at this age (Figure 2C). Thus, Ccl3 - /- affects weight more notably in Npc1 -/- mice than in Npc1 +/- mice. We also needed to consider genetic backgrounds, which may greatly influence disease progression (44). The Ccl3 -/- ; Npc1 -/- mice were on a mixed FVB and C57BL/6 (B6) background. B6 Npc1 -/- mice life expectancy averages days (45) while FVB Npc1 -/- mice live for days (35). Despite this marked difference between the two backgrounds, FVB/B6 Npc1 -/- male mice readily survived past P60 and their weights were comparable to FVB Npc1 -/- mice (Figures 2A, 2B, 2D). Thus, we attribute the lower weights of Ccl3 -/- ; Npc1 -/- mice to the Ccl3 mutation, not to the background genetic constitution. Even if other factors were involved in worsening the disease condition of Ccl3 -/- ; Npc1 -/- mice, none of the expected weight gains and sustainability demonstrated with the Sandhoff model was observed. In the Sandhoff study (31), Ccl3 -/- ; Hexb -/- mice had higher body weights and ~30% longer lifespans than Hexb -/- mice. Thus, in contrast to the influence of CCL3 on Sandhoff disease, the endogenous production of CCL3 may play a neutral or beneficial rather than harmful role in NPC disease.
12 12 The loss of CCL3 benefited the Sandhoff Hexb -/- mouse model, possibly because it lessened harmful microglia-associated pathology in the CNS and inhibited macrophage infiltration into the CNS. In order to determine if CCL3 was also required to alter the macrophage and microglia-associated pathology and infiltration in Npc1 -/- mice, we quantified the prevalence of CD68-positive cells in both visceral tissue and the CNS. CD68 is a marker of activated microglia and macrophages. Macrophages do not infiltrate the CNS in NPC diseased mice In addition to not improving the weight profile, deleting Ccl3 did not alter the prevalence of CD68-positive cells in the CNS of Npc1 -/- mice (Figures 2E, 2F). Analysis of liver tissues also showed that, in Ccl3 -/- ; Npc1 -/- mice, no significant change in CD68 staining was observed. In a few mice, however, enlarged granuloma-like macrophage clusters were present, which may have caused an increase in the average CD68 percentage calculated for the liver (Figure 2D). The trend towards increased macrophage-associated liver pathology in Ccl3 -/- ; Npc1 -/- mice and lack of decrease in CD68-positive cells in the CNS further suggests that endogenous CCL3 may play a neutral or beneficial role in modifying NPC disease pathology. The main benefit of deleting Ccl3 in Hexb -/- mice was suggested to be inhibition of macrophage infiltration into the CNS (31). In order to test whether peripheral
13 13 macrophages readily invade the CNS of Npc1 -/- mice, we labeled or eliminated peripheral macrophages with liposomes (Lp). Dil-label and clodronate-toxin containing Lp (Dil-Lp and Clod-Lp respectively) have been used extensively in mice to selectively deplete or label macrophages or microglia (41, 46, 47). When Dil-Lp was injected into the peritoneal cavity of FVB Npc1 -/- mice at ages P52 or P70, Dil labeling of CD68-positive macrophage cells was evident in the liver after 24 or 48hrs (Figure 2G). In these same mice, extremely few or no CD68-positive cells were labeled with Dil in the brain. Instead, Dil was sequestered in capillaries after 24hrs (Figure 2H) suggesting that the Dil-label is transportable to the brain but that the blood-brain barrier restricted extravasation of Dil-Lp or Dil-Lpcarrying cells. Upon repeated peritoneal injection of Clod-Lp from ages P50-P60, depletion of Cd68-positive cells from the liver, but not from the cerebellum, was evident in Npc1 -/- mice (Figure 2I). These results demonstrate that peripheral macrophages do not readily infiltrate and contribute to the CNS pathology in Npc1 -/- mice. The lack of noted macrophage infiltration into the CNS of Npc1 -/- mice is distinct from neuroinflammation in Hexb -/- mice, where peripheral macrophages infiltrate the brain and contribute to CNS pathology (31). This difference may be the reason why deletion of CCL3 was not beneficial to Npc1 -/- mice. To assess what additional influence CCL3 may have on inflammation and neurodegeneration, we used gene expression profiling to compare Ccl3 -/- ; Npc1 -/- and Npc1 -/- mice cerebella.
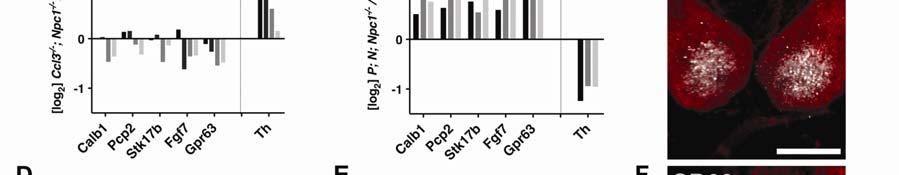
14 14 Neuronal NPC1, not deletion of Ccl3, suppresses neuroinflammation in NPC diseased mice Array analyses of Ccl3 -/- ; Npc1 -/- mice compared to age-matched and background-matched Npc1 -/- cerebella showed slight decreases in expression of cerebellar Purkinje neuron-specific genes: Calb1/D28K, Pcp2, Stk17b/Drak2, Fgf7, and Gpr63 (Figure 3A). Decrease expression of these genes is likely due to reduced numbers of surviving Purkinje neurons (48). Stk17b, for example, is produced only in Purkinje neurons of the cerebellum and serves as useful indicator of cell number (Figure 3C). The expression of Tyrosine hydroxylase (Th) can also be used to assess the condition of Purkinje neurons in the cerebellum in NPC disease since Th is ectopically expressed in cerebellar Purkinje neurons as a consequence of the disease (49). Th levels on average were higher in Ccl3 -/- ; Npc1 -/- cerebella than in Npc1 -/- mice (Figures 3A, 3J). Based in part on the reduction, though slight, of Purkinje neuron-specific genes and mainly on the increase in Th, we conclude that the loss of Ccl3 is deleterious to neurons in Npc1 -/- mice. Gene profile array analysis revealed a general increase in expression of genes encoding lysosome-associated proteins, secreted proteases, and immune response factors (Figure 1A) in Ccl3 -/- ; Npc1 -/- mice relative to Npc1 -/- controls (Figures 3D, 3G). The elevation of these inflammatory factors is thought to contribute to neurodegeneration. C1q, a main component of the immune
15 15 complement pathway, is of particular interest since its production can promote microglial-mediated removal of neuron synapses leading to neuronal death (50). The expression of C1q is elevated early prior to signs of neurodegeneration in NPC disease (12) and here we find that C1q marks cells proximal to Purkinje neuron dendrites in a fashion similar to CD68-positive microglia (Figures 3F, 3I). Thus, microglia and C1q-positive cells can be in physical contact with Purkinje neuron dendritic spines to potentially mediate neuron clearance in NPC disease. The increased expression and localization of C1q has been confirmed with RNA probe in situ hybridization (data not shown). The increase in C1q and other immune components in Ccl3 -/- ; Npc1 -/- mice further suggests that loss of Ccl3 is deleterious to neurons in Npc1 -/- mice. Despite increases in inflammatory factors, the pattern of Purkinje neuron loss and invasion of CD68-positive microglia in Ccl3 -/- ; Npc1 -/- mice remains similar to that of Npc1 -/- mice. As previously reported (35), CD68-positive microglia are predominantly seen in areas of the cerebellum where Purkinje neurons have degenerated and are absent from areas where they still remain (Supplemental Figure S2). Thus, eliminating the inflammatory mediator CCL3 did not greatly alter the neurodegenerative process or resulting microglial response. This agrees with the conclusion that CCL3 may play a neutral or beneficial rather than a harmful role in modifying NPC disease.
16 16 In a prior study, we reported that patterned Purkinje neuron loss and microglial activity can be halted by neuron-specific production of NPC1 in P; N; Npc1 -/- mice (35). Here, we assessed whether inflammatory factors in general can be suppressed in NPC disease, without eliminating pro-inflammatory components such as CCL3. We performed gene profiling array analysis on P; N; Npc1 -/- mice, which have early and constitutive production of a functional Npc1-YFP protein in cerebellar Purkinje neurons (35). The specific localization of YFP immunofluorescence in Purkinje neuron cell bodies, the reduction of filipin cholesterol stain in Purkinje neurons, and the prevention of increased glial reactivity only in the cerebellum helped confirm that the rescue specifically affected only those cells (35). Compared to Ccl3 gene deletion, neuron-specific correction of NPC disease in P; N; Npc1 -/- mice had a considerable and opposite effect on neuroinflammation. Providing NPC1 to Purkinje neurons in Npc1 -/- mice maintained Purkinje neuronspecific gene expression levels and suppressed increases in Th (Figures 3B, 3J, Supplemental Figure S3). While Ccl3 gene deletion tended to increase the levels of inflammation (Figures 3D, 3G, 3J), all inflammatory factors identified by array, including C1q, were suppressed in the cerebella of P; N; Npc1 -/- mice (Figures 3E, 3H). This demonstrates that much of the abnormal inflammatory gene expression in the cerebella of Npc1 -/- mice results from neuronal defects that stimulate inflammatory cells, rather than the loss of NPC1 from microglia.
17 17 The future of therapeutic approaches, genetic or otherwise, for neurodegenerative disorders may depend on arresting the progression of neuronal loss when a patient is already exhibiting disease signs. For NPC disease, prior studies demonstrated that neuronal NPC1 is required for neuron survival (35, 51). However, these studies did not address whether neuronal NPC1 is sufficient to halt neuron loss late in the disease after mice initially develop without NPC1 function. Methods of intrinsic neuron rescue will work only if the early events in the disease do not set in motion an unstoppable process of decay that contributes to non-autonomous neuronal toxicity and elimination. Since neuron rescue and not deletion of Ccl3 had such a dramatic effect on suppressing inflammation and preventing neuronal loss, we next focused on manipulating the timing of neuron rescue to see whether NPC disease could be arrested or reversed at late stages of the disease. Arresting neurodegeneration late in NPC disease progression In order to test whether neuron rescue is possible late in the disease, we regulated the production of NPC1-YFP in P; N; Npc1 -/- mice with Doxycycline (Dox). Dox binds to the tta protein produced by Pcp2-tTA and inhibits its ability to induce the expression of a teto- transgene (Figure 4A). This method has been previously been used to show neurodegenerative recovery of Purkinje neurons by inhibiting the continuous expression of mutant ataxin-1 (52).
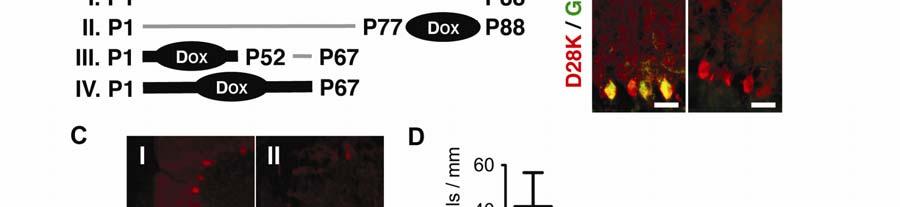
18 18 First, we tested whether suppression of the production of Purkinje neuron NPC1- YFP in P; N; Npc1 -/- mice with Dox (Figure 4B) can elicit neurodegeneration. Cerebellar lobule III was chosen for analysis because of its consistent morphology across mice of various ages and its early loss of Purkinje neurons compared to more posterior lobules (53). In male FVB Npc1 -/- mice, Purkinje neuron loss starts to become apparent in lobule III by age P50. By P65, Npc1 -/- mice tend to be devoid of Purkinje neurons in this cerebellar region. In sharp contrast, P; N; Npc1 -/- mice at age P88 have considerably more neurons surviving in the lobule III region of the cerebellum than Npc1 -/- mice older than P72 (Figures 4C, 4D-trial I). When Dox is administered continuously to P; N; Npc1 -/- mice starting at P77, neuron loss commences. At P88, P; N; Npc1 -/- mice that had been treated with Dox starting at P77 showed Purkinje neuron loss that resembles the degree of Purkinje neuron loss that occurs in Npc1 -/- mice between P52 and P65 (Figure 4D-trial II). Thus, removing NPC1 function from neurons of adult P; N; Npc1 -/- mice triggers degeneration of neurons. These data from Npc1 -/- mice support results obtained from Npc1 +/+ mice with conditional knockout of the Npc1 gene in mature neurons (51). To demonstrate that providing NPC1 function to neurons allows neuron survival even well along in the disease, we administered Dox continuously to P; N; Npc1 -/- mice from birth to P52. Dox was removed at P52. By P52, inflammatory processes have had ample time to intensify (Figure 1A). When assessed at age P67, these mice did not differ in the amount of Purkinje neurons in lobule III
19 19 compared to P52 Npc1 -/- mice (Figure 4D-trial III). Thus, restoring NPC1-YFP production in Purkinje neurons at P52 halted the neurodegenerative process (Figures 4C, 4D). If Dox was not removed at P52, then the number of Purkinje neurons remaining in lobule III at P67 in P; N; Npc1 -/- mice was not significantly different (P=0.333 n=4 mice) than the number of Purkinje neurons typically remaining at P65 in Npc1 -/- mice (Figure 4D-trial IV). Thus, neuronal NPC1 function is necessary for the survival of neurons and sufficient to arrest neurodegeneration. Since neuron survival can be controlled even at late stages of the disease without eliminating inflammatory mediators in Npc1 -/- mice, we hypothesized that other cell types vulnerable to the disease could be rescued without targeting macrophages or immune cells in the body. NPC disease is a complex disorder that affects multiple tissues in addition to the nervous system. Notably, liver and pulmonary injuries are leading causes of death of NPC patients (54-56). If macrophage-associated pathology in visceral tissue operates in a fashion similar to microglia-associated pathology in the CNS, then correction of hepatic cells in the liver, for example, should be sufficient to suppress the inflammation even after tissue pathology has become severe.
20 20 Reversing macrophage-associated NPC diseased tissue pathology To test whether liver rescue is possible late in the disease, we regulated the production of NPC1-YFP with the ROSA26r-rtTA-M2 transgene (R) in R; N; Npc1 -/- mice (35). rtta is regulated in the opposite way from tta; Dox binding to rtta induces expression of the teto-transgene (Figure 5A). It is also known that the ROSA26r-rtTA-M2 transgene, in the presence of Dox, induces expression of a teto-transgene in fibroblasts and epithelial cells throughout the body (57). Thus, in addition to being able to time NPC rescue in visceral tissue of R; N; Npc1 -/- mice, we were also able to selectively test the importance of NPC1 function in epithelial cells of various organs in the context of NPC1 deficient immune cells. We found that production of NPC1-YFP protein was inducible by Dox in nonmacrophage cells both in vitro and in vivo. In vitro, addition of Dox to culture media can induce NPC1-YFP in primary fibroblasts but not in peritoneal cells isolated from R; N; Npc1 -/- mice (Figure 5B). In vivo, when R; N; Npc1 -/- mice were fed Dox, NPC1-YFP was produced in the liver and detected by immunofluorescence (Figures 5B, 5C). In these Dox-treated R; N; Npc1 -/- mice, resident macrophages and invading monocytes or lymphocytes in the liver did not have detectable NPC1-YFP protein. NPC1-YFP immunofluorescence was also absent from the spleen and restricted to a few cells in the thymus and bone marrow that are not Thy1.2-positive (Figures 5C, 5D). Thus, NPC1-YFP was
21 21 produced broadly in visceral tissue of R; N; Npc1 -/- mice but was largely absent from monocytes, lymphocytes, and bone marrow cells. Despite the lack of NPC1-YFP rescue in macrophages, cholesterol accumulation in the liver of R; N; Npc1 -/- mice was modified after a week of NPC1-YFP induction (Figure 5E). Although some filipin stain persisted, it was now in individual cells dispersed throughout the liver. These cells lacked NPC1-YFP immunofluorescence and were identified as predominantly macrophages based on CD68 staining (Figure 5F). High-magnification imaging showed that macrophages in liver tissue of R; N; Npc1 -/- mice had changed from a foam cell appearance to a more ramified intermediate state once liver epithelial cells or hepatocytes were genetically rescued (Figure 5G). With prolonged NPC1-YFP induction, the majority of macrophages in the livers of R; N; Npc1 -/- mice shared a resting state morphology equivalent to macrophages found in wild-type mice. Thus, macrophages in liver remained responsive to their environment despite having a continuous NPC1 defect. Focal macrophage responses to changes in local environments within the same tissue can be seen upon brief (3-5 day) induction of NPC1-YFP in P65 or older R; N; Npc1 -/- mice. In the liver, foam cell macrophages clustered in localized nonrescued liver areas, marked by lack of GFP immunofluorescence and intense filipin stain (Figure 6A). Those areas may have been sites of severe liver tissue injury, containing cells other than hepatocytes that do not produce NPC1-YFP.
22 22 In lungs, a localized macrophage response was also seen. Macrophages near simple squamous epithelial cells producing NPC1-YFP had drastically thin morphology compared to the large foam cell morphology of macrophages near lung epithelia lacking NPC1-YFP (Figure 6B). The macrophage morphology change in lung was roughly quantified by the percent of alveolar tissue area occupied by CD68 immunofluorescence. No change in morphology occurred in macrophages located away from sites of maximum NPC1-YFP production in the alveoli, which happened to be bronchioles (Figures 6C, D). These results demonstrate, for the first time, reversal of abnormal macrophage behaviors in the lung of Npc1 -/- mice. Taken together, our rescue experiments in the brain, liver and lung suggest that macrophage abnormalities in NPC are driven by abnormal local tissue environments and not due to intrinsic defects in NPC1 production. To more directly ascertain the relative contribution of immune cells to NPC pathology, we pharmacologically depleted macrophages from the viscera and compared liver injury with amount of liver injury that resulted from hepatocyte epithelial cell rescue or Ccl3 deletion.
23 23 Macrophage intervention is not required for reducing tissue injury in NPC diseased mice Deletion of the macrophage inflammatory cytokine Ccl3 from Npc1 -/- mice (Figure 7A) did not reduce the levels of the hepatic enzyme alanine aminotransferase (ALT) in the blood (Figure 7D). Serum levels of ALT serve as a clinical marker of hepatocyte injury, so we conclude that liver damage was not reduced in Ccl3 -/- ; Npc1 -/- mice. A separate study also observed that ALT levels were not reduced in a mouse knockout of the cytokine TNF-alpha when NPC liver disease was induced with NPC1-specific antisense oligonucleotides (58). Despite little difference in ALT levels, the authors noted that loss of TNF-alpha delayed macrophage-associated pathology in the liver. In our study, ALT levels did correlate with the degree of macrophage-associated liver pathology caused by deletion of Ccl3 (Figures 2E, 7D). Given these reported phenotypes, it remains uncertain whether macrophage activity in the liver can be toxic to hepatocytes. To explore the relevance of macrophages to liver injury, we depleted liver macrophages from Npc1 -/- mice using Clod-Lp (Figure 7B). Although repeated Clod-Lp injections caused some toxicity, evident as a small increase in ALT levels in non-npc animals, if macrophage activity is harmful in NPC disease the resulting smaller macrophage population should have lessened liver injury in Npc1 -/- mice. Instead, we saw an average increase in ALT levels (Figure 7E).
24 24 Serum ALT levels in Npc1 -/- mice have been reduced only when treatments targeted the storage accumulation in the liver (27, 54). These treatments, such as CYCLO, are not cell-specific and thus do not reveal the cell type responsible for the beneficial effect. Using R; N; Npc1 -/- mice, we found that genetically providing NPC1 function to epithelial cells in the liver, but not macrophages (Figure 7C), for a week was sufficient to normalize serum levels of ALT (Figure 7F). Thus, hepatic injury, like neuron injury, is primarily due to absence of NPC1 in the cell itself and not to an aberrant inflammatory response or abundant immune cell activity. Discussion Our series of experiments indicate that inflammation in the NPC mouse model of LSD resembles a self-limiting inflammatory reaction. Inflammation is described as self-limiting or acute when it continues only until the threat to the cell is eliminated and tissue homeostasis is restored (59). In Npc1 -/- mice, we demonstrate that correcting neuronal or epithelial cells is sufficient to suppress inflammation and reverse the macrophage-associated pathology in tissues (Figures 3, 5, 6). Furthermore, neuron loss and tissue injury could be halted, even late in NPC disease, if neurons and epithelial cells were treated before the cells were lost (Figures 4, 7). Although the NPC1 defect remains in macrophages and other immune cells, we find that this does not preclude resolution of inflammation in NPC disease. The inflammatory response remains a secondary response to tissue injury. This is encouraging news for modifying the disease
25 25 progression of NPC patients, even if diagnosis or treatment of the disease occurs later in life. Surprisingly, attempts to perturb the inflammatory response by genetically deleting the macrophage inflammatory cytokine Ccl3 or depleting activated macrophages from tissues in Npc1 -/- mice proved more deleterious than beneficial as a therapeutic approach (Figures 2, 3, 7). These results provide evidence that the inflammatory activity of microglia and macrophages in NPC disease in response to cell defects and injury may be a beneficial adaptation to try and prevent excessive tissue damage (Figure 8). This view is supported by recent in vitro data demonstrating that abundant Npc1 -/- microglia promote neuron survival of wild-type neurons in microglia-neuron co-cultures (60). Whether similar events occur in other LSDs has yet to be explored in sufficient detail and in appropriate models. Deletion of Ccl3 allowed us to determine the relevance of this cytokine to NPC disease progression and to experimentally bridge NPC disease with the LSD Sandhoff disease. Our results highlight an important distinction between NPC and Sandhoff disease mouse models with regards to inflammation. For the Npc1 - /- mouse model of NPC disease, we now have multiple lines of evidence to show that the degeneration of neurons is governed by neuron-intrinsic defects. The elevation of the inflammatory response itself does not drive elimination of healthy neurons (Figures 3, 4) and macrophage infiltration into the CNS is not seen
26 26 (Figure 2). In the Hexb -/- mouse model of Sandhoff disease, in contrast, macrophage invasion into the CNS has been shown, and neuronal defects and inflammation are proposed to converge to promote neurodegeneration (31). To date, the pathogenic cascade of the Sandhoff disease model has been the most widely proposed pathogenic cascade for lysosomal storage diseases in general (1-3). The distinct roles of inflammation in NPC and Sandhoff diseases, however, argue for reevaluating the roles of neuroinflammation in other LSDs that have neurological deficits. Functional differences among LSDs may be concealed behind overtly similar phenotypes such as visible accumulations of lysosome material or elevation of inflammatory components. NPC and Sandhoff both accumulate glycosphingolipids (33), which are potent immune modulators, yet the same genetic treatment that benefitted the Sandhoff mouse model was deleterious to NPC mice (Figures 2, 3). NPC, Sandhoff, and other LSDs also share multiple elevated pro-inflammatory cytokines (37, 38), yet the influence of these cytokines may differ between disease states. Differences in the roles of inflammation among LSDs may be better determined by properties specific to the pathways and cell types involved. Thus, the role of inflammatory cells has to be carefully analyzed for an individual LSD and, if possible, the cell types therapeutically relevant to treating the disease identified in order to begin to categorize LSDs based on treatment potential.
27 27 Unlike lysosomal storage disorders that are caused by loss of function of a secretable enzyme, NPC disease caused by loss of transmembrane protein NPC1 function is cell autonomous. Providing NPC1 to one cell does not correct the defect in another (35, 53, 61). Along with cell autonomy, NPC disease also demonstrates tissue autonomy between organ systems, so that an alteration, such as reduced inflammation, in one system or tissue area does not necessarily modify the pathology in another. This is in contrast to the non-cell autonomous neurodegenerative msod1 mouse model of amyotrophic lateral sclerosis (ALS), where immune modulation in peripheral tissues such as the liver could potentially result in a global neuroprotective intervention (62). For a cell autonomous neurodegenerative disease such as NPC, global pathology is unlikely to be corrected by selective treatment of specific tissues. Tissue autonomy in NPC disease is demonstrated by the observation that peripheral monocytes do not invade the brain in Npc1 -/- mice (Figure 2) and the macrophage pathology in liver and lung can be locally reduced depending on where in the tissue NPC1 is produced (Figures 5, 6). As a result, genetic or environmental risk factors as well as therapeutic applications that affect a particular tissue system will elicit a modified clinical phenotype only within the target tissue despite a similar NPC1 gene defect throughout the body. Future NPC research directions should focus on identifying modifiers of the disease in order to better understand the variability in visceral tissue pathology seen in human patients (63). Although late NPC disease pathology can be
28 28 mitigated (Figures 5, 6, 7), genetic variables and early developmental attributes still appear to contribute to how the disease progresses. The altered disease progression reflected by weight measurements of Ccl3 -/- ; Npc1 -/- mice (Figure 2) suggests that pre-existing differences in the cytokine milieu can contribute to disease severity, particularly in visceral tissue. Other factors, genetic and environmental may also be involved (64). Our results demonstrate that Npc1 -/- macrophages and immune cells respond appropriately to epithelial and neuronal injury in NPC disease and that targeting cytokines produced by macrophages or depleting macrophages in NPC disease may inadvertently promote peripheral tissue or neuronal injury. These characteristics of NPC disease pathology have implications beyond NPC and the LSD research community and may help treat disparate diseases, especially HIV and Ebola virus infection. The processes of HIV and Ebola virus infection has been linked to NPC disease through function of the NPC1 protein. Loss of NPC1 alters transmission of HIV virus by affecting its packaging and excretion from the infected cell (10). Loss of NPC1 limits Ebola virus transmission by trapping Ebola in endocytic compartments of host cells, thereby preventing infection (65). In both viral diseases, macrophages and lymphocytes are believed to be the main reservoirs of the viruses and facilitate virus transmission throughout the body. Based on the findings reported here, inhibition of NPC1 function in macrophages and immune cells could help moderate HIV and Ebola viral infection without eliciting neurodegeneration or causing much tissue injury.
29 29 Materials and Methods Transgenic Mice Care and handling of mice was in accordance with institutional guidelines. Mice homozygous for the Ccl3 targeted mutation (JAX Mice Strain Name: B6.129P2- Ccl3 tm1unc /J) were obtained from The Jackson Laboratory. These C57BL/6 Ccl3 -/- mice were crossed to congenic FVB Npc1 -/- mice (originally JAX Mice Strain Name: BALB/cNctr-Npc1 m1n /J) previously generated in our laboratory (35). Offspring genotyped as Ccl3 +/- ; Npc1 +/- were bred to produce experimental Ccl3 -/- ; Npc1 -/- mice and controls, Npc1 +/-, Npc1 -/-, and Ccl3 -/- ; Npc1 +/- mice of mixed FVB/B6 background. The genotypes of the Ccl3 and Npc1 loci were determined by PCR of genomic DNA obtained from ear clips. Primers to detect the mutations in the Ccl3 and Npc1 loci have been previously described (31, 35). RT-PCR of liver total RNA was performed on half of the Ccl3 -/- ; Npc1 -/- mice generated in order to confirm loss of Ccl3 mrna. A primer pair was used to generate a 433bp amplicon as follows: 5 -CCTCTGTCACCTGCTCAACATC-3 and 5 - ATGTGGCTACTTGGCAGCAAAC-3. Actin primer pairs for control PCR, 5 - GCTCCGGCATGTGCAAAG-3 and 5 - CCTCGGTGAGCAGCACAG-3, generate a ~283bp amplicon. For humane reasons mice were sacrificed under regulated C0 2 if lethargic and the body weight of a NPC diseased mouse dropped below 14g for male mice or
30 30 12g for female mice older than P40. Weight measurements were made every 2-7 days, or at ages reported. P; N; Npc1 -/- and R; N; Npc1 -/- mice were generated in our laboratory as previously described (35). Liposome Treatment Clodronate- and Dil-containing liposomes, as well as empty vehicle liposomes, were purchased from Encapsula NanoSciences. Approximately 200µl of Dil-Lp solution was injected into the peritoneal cavity of FVB P52 or P70 Npc1 -/- mice. Liver and brain tissue was dissected and analyzed 24-48hrs later. Approximately 100µl of Clod-Lp or control Lp solution was injected into the peritoneal cavity of FVB Npc1 -/- and wild-type mice at days P49 and P53. Liver and brain analyses were performed at P56. Dox Treatment Mice were given Doxycycline hyclate (Sigma) at a concentration of 2mg/ml in their drinking water with 5% sucrose. The solution was refreshed every 3 days and light-protected water bottles were used. For R; N; Npc1 -/- mice, induction of NPC1-YFP was confirmed by skin biopsy. The NPC1-YFP fluorescence produced by fibroblast skin cells in the biopsy can be detected by standard fluorescent microscopy using a 20x objective and GFP filter set.
31 31 Liver ALT ALT plasma level analysis was performed in-house at the Stanford University Veterinary Service Center. Ccl3 -/- ; Npc1 -/- mice ALT was measured at age P58. Clod-Lp and Lp treated mice ALT was measured at P56. For R; N; Npc1 -/- mice ALT levels were measured at P56 after mice were given Dox for 7 days. Gene Profiling Cerebella from Ccl3 -/- ; Npc1 -/- mice and sibling Npc1 -/- mice, P; N; Npc1 -/- mice and age-matched Npc1 -/- mice, or Npc1 -/- and Npc1 +/- mice were harvested and total RNA was extracted with Trizol reagent (Invitrogen). A total of 16 animals were used. Individual arrays were performed on total RNA from each whole cerebellum isolated. Genomic DNA was removed with Turbo DNA-free kit (Ambion) and the RNA further purified with Acid Phenol:ChCl 3 (Ambion) and subsequent alcohol precipitation steps. A quality check of RNA by Agilent Bioanalyzer QC, cdna synthesis, hybridization onto Affymetrix GeneChip Mouse Gene 1.0 ST Arrays, and scanning was performed at Stanford Protein and Nucleic Acid Facility. Normalized intensity data were obtained using Affymetrix Expression Console software and RMA default settings. Intensity values and raw microarray data were deposited in NCBI s Gene Expression Omnibus (66) and can be viewed through GEO Series accession number GSE Known genes showing a two-fold or greater increase in Npc1 -/- compared to Npc1 +/- FVB mice at P50 were ranked based on averages. SAM assessed significance and the functional groupings (Figure 3) of the top
32 32 approximately 50 genes (Figure 1) were assigned using the Database for Annotation, Visualization and Integrated Discovery (DAVID) (67). Low cycle RT-PCR analysis of Ccl3 RNA levels was performed on total RNA isolated from various tissues and brain regions of P60 Npc1 -/- and Npc1 +/- mice. The Ccl3 and Actin primer pairs are noted above, under Materials and Methods: Transgenic Mice. Immunofluorescence, staining, and quantification Primary antibodies used in this study: chicken anti-gfp (Aves), chicken anti- GFAP (Aves), chicken anti-tyrosine Hydroxylase (Aves), rabbit anti- Calbindin/D28K (Sigma), rat CD68 (Abdserotec), rabbit anti-drak2 (Cell Signaling Technology), Thy1.2 (abcam), rat C1q (abcam), and mouse anti-actin (Millipore). Appropriate Alexa and HRP conjugated secondary antibodies were obtained from Invitrogen and Jackson ImmunoResearch. Filipin (Sigma) was added with secondary antibodies during staining. For tissue immunofluorescence microscopy, all tissues were isolated and fixed overnight in 4% paraformaldehyde PBS solution at 4ºC, then sectioned with a Vibratome into 50µm slices. Brains were sectioned sagitally. Bone marrow was extruded into fixative. Tissue sections and bone marrow clumps were incubated with primary antibody overnight at 4ºC in 2%BSA/0.2%TritonX-100 PBS blocking solution. For tissue treated with Dil-Lp, 2%BSA/0.1%Tween-20 PBS blocking
33 33 solution was used to preserve Dil localization. Blocking solutions were used for washes at room temperature and secondary incubation at 4ºC overnight followed by additional room temperature washes. Tissue sections were mounted and sealed onto slides. Bone marrow cells were pelleted and spread on slides. Epifluorescent images were obtained using a Zeiss Axioplan2 fluorescent microscope equipped with an AxioCam HRc CCD camera. A 10% neutral density filter was used to prevent photobleaching of filipin stain. Confocal images were obtained with a Leica TCS SP2 laser-scanning microscope. Images were processed and rendered using Volocity imaging software. Quantification of the percent area of tissue occupied by CD68 immunofluorescence was measured using ImageJ software as previously described (35). Cell isolation and immunoblot analysis Peritoneal cells were isolated by syringe injecting 7ml of cold PBS into the peritoneal cavity of a euthanized mouse and aspirating back up 5ml. Suspended peritoneal cells were centrifugated at 1,000xg for 5min and PBS was replaced with complete growth medium, DMEM (Gibco) with 10% FBS (Thermo Fisher). Resuspended cells were allowed to settle onto a plastic surface for 6hrs. Remaining debri was gently washed off. Dox was added to the remaining cells at a concentration of 100ng/ml for 48hrs before immunoblotting procedure. For RT- PCR analysis, freshly isolated peritoneal cells were pelleted and placed in Trizol for RNA extraction.
34 34 Primary fibroblast cells were cultured in complete medium, DMEM 10% FBS, supplemented with normocin (Invivogen) from tail biopsies of adult R; N; Npc1 -/- mice. Finely diced tail segments were seeded onto tissue culture plates with lowlevel media to prevent floating of segments away from floor of the plate. Partial media exchange was performed daily until cell colonies beside large tissue pieces were seen. Remaining tissue bits were removed and colony forming cells were expanded. Dox was added to the remaining cells at a concentration of 100ng/ml for 48hrs before immunoblotting procedure. For immunoblotting, equal numbers of cells or mass of liver tissue were lysed in cold RIPA buffer supplemented with EDTA-free protease inhibitor cocktail mini tablets (Roche). Cells or tissue were homogenized with motorized pestle. Insoluble material was pelleted at 10,000xg for 10min. Supernatants were stored in Laemmli protein sample buffer prior to electrophoresis in gradient gels (Bio- Rad, Invitrogen). Primary antibodies were incubated overnight at 4ºC in 2%BSA/0.2%Tween-20 PBS solution. HRP-conjugated secondary antibodies were incubated at room temperature for 30min. The HRP chemiluminescence produced by adding SuperSignal West Pico Chemiluminescent Substrate (ThermoScientific) was imaged using a ChemiDoc XRS System (Bio-Rad).
35 35 Funding This work was supported by the Ara Parseghian Medical Research Foundation; Howard Hughes Medical Institute; National Institutes of Health [R01 NS to M.P.S., GM07790 and GM to M.E.L.]; a Pew Charitable Trust Fellowship to A.D.K.; and a Stanford Medical Scholars Fellowship and a Howard Hughes Medical Institute Research Fellowship to J.H..
36 36 References 1. Castaneda, J.A., Lim, M.J., Cooper, J.D. and Pearce, D.A. (2008) Immune system irregularities in lysosomal storage disorders. Acta Neuropathol., 115, Vitner, E.B., Platt, F.M. and Futerman, A.H. (2010) Common and uncommon pathogenic cascades in lysosomal storage diseases. J. Biol. Chem., 285, Futerman, A.H. and van Meer, G. (2004) The cell biology of lysosomal storage disorders. Nat. Rev. Mol. Cell Biol., 5, Jeyakumar, M., Dwek, R.A., Butters, T.D. and Platt, F.M. (2005) Storage solutions: treating lysosomal disorders of the brain. Nat. Rev. Neurosci., 6, Parkinson-Lawrence, E.J., Shandala, T., Prodoehl, M., Plew, R., Borlace, G.N. and Brooks, D.A. (2010) Lysosomal storage disease: revealing lysosomal function and physiology. Physiology (Bethesda), 25, Pressey, S.N., Smith, D.A., Wong, A.M., Platt, F.M. and Cooper, J.D. (2012) Early glial activation, synaptic changes and axonal pathology in the thalamocortical system of Niemann-Pick type C1 mice. Neurobiol. Dis., 45, Vanier, M.T. (2010) Niemann-Pick disease type C. Orphanet J. Rare Dis., 5, Kaye, E.M. (2011) Niemann-Pick C disease: not your average lysosomal storage disease. Neurology, 76, Maxfield, F.R. and Tabas, I. (2005) Role of cholesterol and lipid organization in disease. Nature, 438, Tang, Y., Leao, I.C., Coleman, E.M., Broughton, R.S. and Hildreth, J.E. (2009) Deficiency of niemann-pick type C-1 protein impairs release of human immunodeficiency virus type 1 and results in Gag accumulation in late endosomal/lysosomal compartments. J. Virol., 83, Sagiv, Y., Hudspeth, K., Mattner, J., Schrantz, N., Stern, R.K., Zhou, D., Savage, P.B., Teyton, L. and Bendelac, A. (2006) Cutting edge: impaired glycosphingolipid trafficking and NKT cell development in mice lacking Niemann-Pick type C1 protein. J. Immunol., 177, Liao, G., Wen, Z., Irizarry, K., Huang, Y., Mitsouras, K., Darmani, M., Leon, T., Shi, L. and Bi, X. (2010) Abnormal gene expression in
37 37 cerebellum of Npc1-/- mice during postnatal development. Brain Res., 1325, Infante, R.E., Abi-Mosleh, L., Radhakrishnan, A., Dale, J.D., Brown, M.S. and Goldstein, J.L. (2008) Purified NPC1 protein. I. Binding of cholesterol and oxysterols to a 1278-amino acid membrane protein. J. Biol. Chem., 283, Ohgami, N., Ko, D.C., Thomas, M., Scott, M.P., Chang, C.C. and Chang, T.Y. (2004) Binding between the Niemann-Pick C1 protein and a photoactivatable cholesterol analog requires a functional sterol-sensing domain. Proc. Natl. Acad. Sci. U. S. A., 101, Storch, J. and Xu, Z. (2009) Niemann-Pick C2 (NPC2) and intracellular cholesterol trafficking. Biochim. Biophys. Acta, 1791, Infante, R.E., Wang, M.L., Radhakrishnan, A., Kwon, H.J., Brown, M.S. and Goldstein, J.L. (2008) NPC2 facilitates bidirectional transfer of cholesterol between NPC1 and lipid bilayers, a step in cholesterol egress from lysosomes. Proc. Natl. Acad. Sci. U. S. A., 105, Cheruku, S.R., Xu, Z., Dutia, R., Lobel, P. and Storch, J. (2006) Mechanism of cholesterol transfer from the Niemann-Pick type C2 protein to model membranes supports a role in lysosomal cholesterol transport. J. Biol. Chem., 281, Ko, D.C., Gordon, M.D., Jin, J.Y. and Scott, M.P. (2001) Dynamic movements of organelles containing Niemann-Pick C1 protein: NPC1 involvement in late endocytic events. Mol. Biol. Cell, 12, Blom, T.S., Linder, M.D., Snow, K., Pihko, H., Hess, M.W., Jokitalo, E., Veckman, V., Syvanen, A.C. and Ikonen, E. (2003) Defective endocytic trafficking of NPC1 and NPC2 underlying infantile Niemann-Pick type C disease. Hum. Mol. Genet., 12, Fessler, M.B. and Parks, J.S. (2011) Intracellular lipid flux and membrane microdomains as organizing principles in inflammatory cell signaling. J. Immunol., 187, Lloyd-Evans, E., Morgan, A.J., He, X., Smith, D.A., Elliot-Smith, E., Sillence, D.J., Churchill, G.C., Schuchman, E.H., Galione, A. and Platt, F.M. (2008) Niemann-Pick disease type C1 is a sphingosine storage disease that causes deregulation of lysosomal calcium. Nat. Med., 14, Zhang, J.R., Coleman, T., Langmade, S.J., Scherrer, D.E., Lane, L., Lanier, M.H., Feng, C., Sands, M.S., Schaffer, J.E., Semenkovich, C.F. et al. (2008) Niemann-Pick C1 protects against atherosclerosis in mice via
38 38 regulation of macrophage intracellular cholesterol trafficking. J. Clin. Invest., 118, te Vruchte, D., Jeans, A., Platt, F.M. and Sillence, D.J. (2010) Glycosphingolipid storage leads to the enhanced degradation of the B cell receptor in Sandhoff disease mice. J. Inherit. Metab. Dis., 33, Loftus, S.K., Morris, J.A., Carstea, E.D., Gu, J.Z., Cummings, C., Brown, A., Ellison, J., Ohno, K., Rosenfeld, M.A., Tagle, D.A. et al. (1997) Murine model of Niemann-Pick C disease: mutation in a cholesterol homeostasis gene. Science, 277, Somers, K.L., Wenger, D.A., Royals, M.A., Carstea, E.D., Connally, H.E., Kelly, T., Kimball, R. and Thrall, M.A. (1999) Complementation studies in human and feline Niemann-Pick type C disease. Mol. Genet. Metab., 66, Miyazaki, D., Nakamura, T., Toda, M., Cheung-Chau, K.W., Richardson, R.M. and Ono, S.J. (2005) Macrophage inflammatory protein-1alpha as a costimulatory signal for mast cell-mediated immediate hypersensitivity reactions. J. Clin. Invest., 115, Liu, B., Ramirez, C.M., Miller, A.M., Repa, J.J., Turley, S.D. and Dietschy, J.M. (2010) Cyclodextrin overcomes the transport defect in nearly every organ of NPC1 mice leading to excretion of sequestered cholesterol as bile acid. J. Lipid Res., 51, van Breemen, M.J., de Fost, M., Voerman, J.S., Laman, J.D., Boot, R.G., Maas, M., Hollak, C.E., Aerts, J.M. and Rezaee, F. (2007) Increased plasma macrophage inflammatory protein (MIP)-1alpha and MIP-1beta levels in type 1 Gaucher disease. Biochim. Biophys. Acta, 1772, Sano, R., Tessitore, A., Ingrassia, A. and d'azzo, A. (2005) Chemokineinduced recruitment of genetically modified bone marrow cells into the CNS of GM1-gangliosidosis mice corrects neuronal pathology. Blood, 106, Ohmi, K., Greenberg, D.S., Rajavel, K.S., Ryazantsev, S., Li, H.H. and Neufeld, E.F. (2003) Activated microglia in cortex of mouse models of mucopolysaccharidoses I and IIIB. Proc. Natl. Acad. Sci. U. S. A., 100, Wu, Y.P. and Proia, R.L. (2004) Deletion of macrophage-inflammatory protein 1 alpha retards neurodegeneration in Sandhoff disease mice. Proc. Natl. Acad. Sci. U. S. A., 101, Dhami, R., Passini, M.A. and Schuchman, E.H. (2006) Identification of novel biomarkers for Niemann-Pick disease using gene expression
39 39 analysis of acid sphingomyelinase knockout mice. Mol. Ther., 13, Gadola, S.D., Silk, J.D., Jeans, A., Illarionov, P.A., Salio, M., Besra, G.S., Dwek, R., Butters, T.D., Platt, F.M. and Cerundolo, V. (2006) Impaired selection of invariant natural killer T cells in diverse mouse models of glycosphingolipid lysosomal storage diseases. J. Exp. Med., 203, Ramirez, C.M., Liu, B., Taylor, A.M., Repa, J.J., Burns, D.K., Weinberg, A.G., Turley, S.D. and Dietschy, J.M. (2010) Weekly cyclodextrin administration normalizes cholesterol metabolism in nearly every organ of the Niemann-Pick type C1 mouse and markedly prolongs life. Pediatr. Res., 68, Lopez, M.E., Klein, A.D., Dimbil, U.J. and Scott, M.P. (2011) Anatomically defined neuron-based rescue of neurodegenerative Niemann-Pick type C disorder. J. Neurosci., 31, Tusher, V.G., Tibshirani, R. and Chu, G. (2001) Significance analysis of microarrays applied to the ionizing radiation response. Proc. Natl. Acad. Sci. U. S. A., 98, Wada, R., Tifft, C.J. and Proia, R.L. (2000) Microglial activation precedes acute neurodegeneration in Sandhoff disease and is suppressed by bone marrow transplantation. Proc. Natl. Acad. Sci. U. S. A., 97, Xu, Y.H., Jia, L., Quinn, B., Zamzow, M., Stringer, K., Aronow, B., Sun, Y., Zhang, W., Setchell, K.D. and Grabowski, G.A. (2011) Global gene expression profile progression in Gaucher disease mouse models. BMC Genomics, 12, Tuppo, E.E. and Arias, H.R. (2005) The role of inflammation in Alzheimer's disease. Int. J. Biochem. Cell Biol., 37, Ray, A. and Dittel, B.N. (2010) Isolation of mouse peritoneal cavity cells. J. Vis. Exp Gordon, S. and Taylor, P.R. (2005) Monocyte and macrophage heterogeneity. Nat. Rev. Immunol., 5, Fang, K.M., Wang, Y.L., Huang, M.C., Sun, S.H., Cheng, H. and Tzeng, S.F. (2011) Expression of macrophage inflammatory protein-1alpha and monocyte chemoattractant protein-1 in glioma-infiltrating microglia: involvement of ATP and P2X receptor. J. Neurosci. Res., 89,
40 Cook, D.N., Beck, M.A., Coffman, T.M., Kirby, S.L., Sheridan, J.F., Pragnell, I.B. and Smithies, O. (1995) Requirement of MIP-1 alpha for an inflammatory response to viral infection. Science, 269, Liu, B., Li, H., Repa, J.J., Turley, S.D. and Dietschy, J.M. (2008) Genetic variations and treatments that affect the lifespan of the NPC1 mouse. J. Lipid Res., 49, Parra, J., Klein, A.D., Castro, J., Morales, M.G., Mosqueira, M., Valencia, I., Cortes, V., Rigotti, A. and Zanlungo, S. (2011) Npc1 deficiency in the C57BL/6J genetic background enhances Niemann-Pick disease type C spleen pathology. Biochem. Biophys. Res. Commun., 413, Faustino, J.V., Wang, X., Johnson, C.E., Klibanov, A., Derugin, N., Wendland, M.F. and Vexler, Z.S. (2011) Microglial cells contribute to endogenous brain defenses after acute neonatal focal stroke. J. Neurosci., 31, Buiting, A.M. and Van Rooijen, N. (1994) Liposome mediated depletion of macrophages: an approach for fundamental studies. J. Drug Target, 2, Rong, Y., Wang, T. and Morgan, J.I. (2004) Identification of candidate Purkinje cell-specific markers by gene expression profiling in wild-type and pcd(3j) mice. Mol. Brain Res., 132, Sarna, J.R., Larouche, M., Marzban, H., Sillitoe, R.V., Rancourt, D.E. and Hawkes, R. (2003) Patterned Purkinje cell degeneration in mouse models of Niemann-Pick type C disease. J. Comp. Neurol., 456, Perry, V.H. and O'Connor, V. (2008) C1q: the perfect complement for a synaptic feast? Nat. Rev. Neurosci., 9, Yu, T., Shakkottai, V.G., Chung, C. and Lieberman, A.P. (2011) Temporal and cell-specific deletion establishes that neuronal Npc1 deficiency is sufficient to mediate neurodegeneration. Hum. Mol. Genet., 20, Zu, T., Duvick, L.A., Kaytor, M.D., Berlinger, M.S., Zoghbi, H.Y., Clark, H.B. and Orr, H.T. (2004) Recovery from polyglutamine-induced neurodegeneration in conditional SCA1 transgenic mice. J. Neurosci., 24, Ko, D.C., Milenkovic, L., Beier, S.M., Manuel, H., Buchanan, J. and Scott, M.P. (2005) Cell-autonomous death of cerebellar purkinje neurons with autophagy in Niemann-Pick type C disease. PLoS Genet., 1,
41 Beltroy, E.P., Liu, B., Dietschy, J.M. and Turley, S.D. (2007) Lysosomal unesterified cholesterol content correlates with liver cell death in murine Niemann-Pick type C disease. J. Lipid. Res., 48, Palmeri, S., Tarugi, P., Sicurelli, F., Buccoliero, R., Malandrini, A., De Santi, M.M., Marciano, G., Battisti, C., Dotti, M.T., Calandra, S. et al. (2005) Lung involvement in Niemann-Pick disease type C1: improvement with bronchoalveolar lavage. Neurol. Sci., 26, Yerushalmi, B., Sokol, R.J., Narkewicz, M.R., Smith, D., Ashmead, J.W. and Wenger, D.A. (2002) Niemann-pick disease type C in neonatal cholestasis at a North American Center. J. Pediatr. Gastroenterol. Nutr., 35, Hochedlinger, K., Yamada, Y., Beard, C. and Jaenisch, R. (2005) Ectopic expression of Oct-4 blocks progenitor-cell differentiation and causes dysplasia in epithelial tissues. Cell, 121, Rimkunas, V.M., Graham, M.J., Crooke, R.M. and Liscum, L. (2009) TNF- {alpha} plays a role in hepatocyte apoptosis in Niemann-Pick type C liver disease. J. Lipid Res., 50, Serhan, C.N. and Savill, J. (2005) Resolution of inflammation: the beginning programs the end. Nat. Immunol., 6, Peake, K.B., Campenot, R.B., Vance, D.E. and Vance, J.E. (2011) Niemann-Pick Type C1 deficiency in microglia does not cause neuron death in vitro. Biochim. Biophys. Acta, 1812, Elrick, M.J., Pacheco, C.D., Yu, T., Dadgar, N., Shakkottai, V.G., Ware, C., Paulson, H.L. and Lieberman, A.P. (2010) Conditional Niemann-Pick C mice demonstrate cell autonomous Purkinje cell neurodegeneration. Hum. Mol. Genet., 19, Finkelstein, A., Kunis, G., Seksenyan, A., Ronen, A., Berkutzki, T., Azoulay, D., Koronyo-Hamaoui, M. and Schwartz, M. (2011) Abnormal changes in NKT cells, the IGF-1 axis, and liver pathology in an animal model of ALS. PLoS One, 6, e Patterson, M.C. (2003) A riddle wrapped in a mystery: understanding Niemann-Pick disease, type C. Neurologist, 9, Zhang, J. and Erickson, R.P. (2000) A modifier of Niemann Pick C 1 maps to mouse chromosome 19. Mamm. Genome, 11, Carette, J.E., Raaben, M., Wong, A.C., Herbert, A.S., Obernosterer, G., Mulherkar, N., Kuehne, A.I., Kranzusch, P.J., Griffin, A.M., Ruthel, G. et al.
42 42 (2011) Ebola virus entry requires the cholesterol transporter Niemann-Pick C1. Nature, 477, Edgar, R., Domrachev, M. and Lash, A.E. (2002) Gene Expression Omnibus: NCBI gene expression and hybridization array data repository. Nucleic Acids Res., 30, Huang Da, W., Sherman, B.T. and Lempicki, R.A. (2009) Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc., 4,
43 43 Figure Legends Figure 1. Ccl3 gene expression is elevated in the brain and visceral tissues of NPC mice. (A) We carried out gene expression profile analyses of Npc1 -/- mice (NPC) compared to Npc1 +/- mice (WT) cerebella at P50, an age before weight loss is detected in Npc1 -/- mice (boxed inset, n=12). Ccl3 (black bar) was among the most elevated mrnas identified. Representative data from microarrays of cerebella from two age-matched Npc1 -/- mice and three agematched Npc1 +/- mice are shown. (B) RT-PCR analysis performed on Npc1 -/- (Npc1 -) mice at age P60 revealed region-specific elevation of Ccl3 mrna in the brain compared to Npc1 +/- (Npc1 +) mice. Highest expression was detected in the cerebellum (Cb), less in brainstem (Bs), and lowest in cortex (Cx). Elevated levels of Ccl3 mrna were also detected in spleen (Sp), liver (Lv), and isolated peritoneal cells (Pt). (C) Control RT-PCR showed the absence of a PCR product derived from Ccl3 -/- (Ccl3 -) mouse liver samples. Actin was used to control for RT-PCR conditions and representative data from RT-PCR on samples obtained from 3 mice are shown (B-C). Figure 2. Ccl3 deletion worsens weight progression and macrophage associated pathology in NPC mice. (A) The individual and average (color line) weight values of diseased mice with rescued cerebellar Purkinje neurons (P; N; Npc1 -/- ) were greater than Npc1 -/- mice at age ~P60. P; N; Npc1 -/- and Npc1 -/- mice were in a FVB background. (B) In contrast, individual Ccl3 -/- ; Npc1 -/- FVB/B6 mice weights were, on average, lower than Npc1 -/- FVB/B6 mice. A few Ccl3 -/- ;
44 44 Npc1 -/- mice weights dipped past a predetermined cut-off weight for determining morbidity (dotted horizontal line above gray zone). (C) The lack of difference in the average weight of Ccl3 -/- ; Npc1 +/- mice to Npc1 +/- mice at age ~P60 suggested a limited effect of the Ccl3 gene deletion in non-diseased animals. (D) Representative weight curves of 13 Ccl3 -/- ; Npc1 -/- mice (red lines) and 11 Npc1 -/- mice (black lines) are shown. (A-D) Data from male mice are shown. (E) CD68 immunofluorescence (red) depicts the differences in microglia/macrophage response in age-matched cerebella at P65. Representative pathology from five mice is shown. GFAP immunofluorescence (blue) depicts Bergmann-glia and astrocytes in the cerebellum. Scale bar is 20µm. (F) Percent areas of tissues that are CD68-positive in the cerebellum and liver were calculated for five Ccl3 -/- ; Npc1 -/- mice. (G) After injection of Dil-Lp (red) into the peritoneal cavity of Npc1 -/- mice, which are Ccl3 +/+, the dye was taken up by macrophages (green) in the liver. Scale bars: left panel 200µm; right panel 20µm. (H) Dil-Lp (red) was not taken up by microglia in the brain (green) and remained in capillaries, identified by morphology. Scale bars: left panel 100µm; right panel 10µm. (I) Percent areas of tissues that were CD68-positive were calculated for three Npc1 -/- mice that received Clod-Lp peritoneal injections and three age-matched Npc1 -/- mice that received control Lp injections. Depletion of CD68 occurred in the liver but not in the cerebellum.
45 45 Figure 3. Deletion of Ccl3 does not retard neurodegeneration or decrease neuroinflammation in NPC mice. (A) Microarray analysis between age matched Ccl3 -/- ; Npc1 -/- and Npc1 -/- FVB/B6 mice at various ages revealed a decrease or limited change in Purkinje neuron-specific genes and a general increase in Th. (B) In contrast, microarray analysis between age-matched P; N; Npc1 -/- and Npc1 -/- FVB mice showed an increase in Purkinje neuron-specific genes and consistently reduced Th RNA. P; N; Npc1 -/- mice at age P100 were compared with Npc1 -/- at P50, since Npc1 -/- mice normally do not survive past age P85. (C) As an example of Purkinje neuron specificity, confocal microscopy revealed STK17b protein (white) localized to the nuclear region of Purkinje neurons immunostained with anti-d28k/calb1 (red). In addition to increased Th, microarray analyses of Ccl3 -/- ; Npc1 -/- mice compared to controls revealed increases in (E) lysosomal and protease related genes and (G) complement and immune response genes. (E, H) The expression of these neuroinflammatory components was substantially reduced in P; N; Npc1 -/- mice. (F) Anti-CD68 highlighted microglia (white) encompassing Purkinje neuron dendrites (red). Similarly, (I) C1q immunofluorescence (white) dotted cell bodies, could also be found in close proximity to Purkinje neuron dendrites (red). Confocal image stacks were rendered in 3-D (F-I). Scale bars are 10µm. (J) The trends for Th and Cd68 array data points are shown.
46 46 Figure 4. Neuronal survival remains intrinsically regulated despite age of NPC diseased mice. (A) Expression of the NPC1-YFP transgene in P; N; Npc1 -/- was constitutively driven by Pcp2-tTA and could be inhibited by Dox applied at different ages (Trials I-IV). (B) Example of cerebellar Purkinje neurons that normally would produce NPC1-YFP in a P; N; Npc1 +/- mouse showed no detectable NPC1-YFP immunofluorescence in the lobule III region of the cerebellum after Dox was given for over a week. Image is representative of results from four mice. (C) Depiction of the difference in the number of surviving Purkinje neurons stained with D28K (red) per trial in cerebellar lobule III. (D) Representative quantitative analysis of the number of remaining neurons in lobule III at a given age and per trial. Two mice were used per trial and trials (top graph) I, II, (bottom graph) III, and IV were performed in duplicate. The number of Purkinje neurons for wild-type (WT) mice (white bars are the same for each graph) was calculated by averaging together Npc1 +/- mice of approximate ages: P50, P60, and P70. For Npc1 -/- mice of the following ages: P52, P65, P72, and P82 (gray bars are graphed for each age), three or more mice were sampled for each age. Scale bars are 50µm. Figure 5. Macrophage-associated NPC liver pathology is corrected without immune cell rescue. (A) Expression of the NPC1-YFP transgene, driven by Rosa-rtTA in R; N; Npc1 -/- mice, can be induced with Dox at different ages (Trials I-III). (B) Immunoblotting with anti-gfp demonstrated that, upon Dox induction, NPC1-YFP protein was detected in liver tissue and cultured skin fibroblast cells
47 47 but not in isolated peritoneal cells. Representative data from three animals are shown. (C) Immunofluorescence with anti-gfp detected NPC1-YFP above tissue autofluorescence in liver but not in spleen, and in limited areas of the thymus. (D) Confocal imaging showed that NPC1-YFP immunofluorescence did not colocalize with Thy1.2 positive cells in isolated bone marrow cell clusters. These expression patterns (C-D) were observed for all mice used in this study. (E) Upon brief 1-week Dox induction of NPC1-YFP, reduced cholesterol accumulation was seen in liver sections except in macrophages (yellow square and magnified view). (F) Confocal 3-D rendering demonstrated the absence of NPC1-YFP (green isosurface) in a representative CD68-positive cell (red) in the liver. (G) With prolonged Dox-induced NPC1-YFP the population of foamy CD68-positive cells reverted to a more ramified normal morphological appearance. Images of single CD68-positive cells are representative of >50% of the cells in the tissue. Each trial, I-III, included a minimum of five mice. Scale bars are 50µm. Figure 6. Macrophages respond locally to epithelial cell rescue. (A) Dox administered to P65 or older R; N; Npc1 -/- mice produced a mosaic rescue effect in liver tissue. Regions (inside semi-circle) that did not produce NPC1-YFP immunofluorescence (GFP green) remained cholesterol positive (Filipin blue) and had focal accumulation (arrow) of clustered macrophages (CD68 red). (B) In lung alveoli, foam cell CD68 positive macrophages (red) were seen proximal to simple squamous epithelial cells (top panel). Macrophages near simple squamous epithelial cells producing NPC1-YFP (bottom panel, yellow arrow)
48 48 were reduced in size in comparison to the large foam macrophages surrounding non-npc1-yfp positive epithelial regions. (C) Apart from a few sparse squamous cells, NPC1-YFP immunofluorescence predominated in the epithelial linings (arrow) of bronchioles in R; N mice given Dox. (D) Quantification of the percent area that was CD68-positive in alveoli shows a significant (P=0.0061, n=3 mice) decrease in CD68 in alveoli regions most proximal to bronchioles. Scale bars are 50µm. Figure 7. Serum ALT levels are elevated in the absence of Ccl3 and macrophages but reduced when NPC1 is provided to hepatocytes. Depictions of treatment target in liver of NPC mice: (A) Genetic deletion of cytokine Ccl3. (B) Depletion of peripheral macrophages. (C) NPC1 provision to hepatocytes. ALT blood levels show (D) an average but not significant increase in ALT levels from Ccl3 -/- ; Npc1 -/- compared to Npc1 -/- FVB/B6 mice at age P58, (E) an average but not significant increase in ALT levels from Clod-Lp treated Npc1 -/- compared to control Lp treated Npc1 -/- FVB mice at age P56, and (F) an average and statistically significant decrease (P=0.0055) in ALT levels from Dox treated R; N; Npc1 -/- compared to Dox treated Npc1 -/- FVB mice at age P56. Minimum of 4 mice were used for each ALT analysis. Figure 8. Proposed roadmap of NPC pathogenesis. Model for acute neurodegeneration and tissue injury in the lysosomal storage Niemann-Pick disease type C. Line thickness represents degree of involvement.

49 49



50 50


51 51


52 52



53 53




54 54
Neuronal and epithelial cell rescue resolves chronic systemic inflammation in the lipid storage disorder Niemann-Pick C
 Neuronal and epithelial cell rescue resolves chronic systemic inflammation in the lipid storage disorder Niemann-Pick C Manuel E. Lopez 1,2, Andrés D. Klein 1,2, Jennifer Hong 1,2, Ubah J. Dimbil 1,2 and
Neuronal and epithelial cell rescue resolves chronic systemic inflammation in the lipid storage disorder Niemann-Pick C Manuel E. Lopez 1,2, Andrés D. Klein 1,2, Jennifer Hong 1,2, Ubah J. Dimbil 1,2 and
Supporting Online Material for
 www.sciencemag.org/cgi/content/full/1171320/dc1 Supporting Online Material for A Frazzled/DCC-Dependent Transcriptional Switch Regulates Midline Axon Guidance Long Yang, David S. Garbe, Greg J. Bashaw*
www.sciencemag.org/cgi/content/full/1171320/dc1 Supporting Online Material for A Frazzled/DCC-Dependent Transcriptional Switch Regulates Midline Axon Guidance Long Yang, David S. Garbe, Greg J. Bashaw*
Islet viability assay and Glucose Stimulated Insulin Secretion assay RT-PCR and Western Blot
 Islet viability assay and Glucose Stimulated Insulin Secretion assay Islet cell viability was determined by colorimetric (3-(4,5-dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide assay using CellTiter
Islet viability assay and Glucose Stimulated Insulin Secretion assay Islet cell viability was determined by colorimetric (3-(4,5-dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide assay using CellTiter
The pathogenesis of nervous distemper
 Veterinary Sciences Tomorrow - 2004 The pathogenesis of nervous distemper Marc Vandevelde Canine distemper is a highly contagious viral disease of dogs and of all animals in the Canidae, Mustellidae and
Veterinary Sciences Tomorrow - 2004 The pathogenesis of nervous distemper Marc Vandevelde Canine distemper is a highly contagious viral disease of dogs and of all animals in the Canidae, Mustellidae and
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION FOR Liver X Receptor α mediates hepatic triglyceride accumulation through upregulation of G0/G1 Switch Gene 2 (G0S2) expression I: SUPPLEMENTARY METHODS II: SUPPLEMENTARY FIGURES
SUPPLEMENTARY INFORMATION FOR Liver X Receptor α mediates hepatic triglyceride accumulation through upregulation of G0/G1 Switch Gene 2 (G0S2) expression I: SUPPLEMENTARY METHODS II: SUPPLEMENTARY FIGURES
Supplemental Information. Otic Mesenchyme Cells Regulate. Spiral Ganglion Axon Fasciculation. through a Pou3f4/EphA4 Signaling Pathway
 Neuron, Volume 73 Supplemental Information Otic Mesenchyme Cells Regulate Spiral Ganglion Axon Fasciculation through a Pou3f4/EphA4 Signaling Pathway Thomas M. Coate, Steven Raft, Xiumei Zhao, Aimee K.
Neuron, Volume 73 Supplemental Information Otic Mesenchyme Cells Regulate Spiral Ganglion Axon Fasciculation through a Pou3f4/EphA4 Signaling Pathway Thomas M. Coate, Steven Raft, Xiumei Zhao, Aimee K.
LDL Uptake Cell-Based Assay Kit
 LDL Uptake Cell-Based Assay Kit Item No. 10011125 www.caymanchem.com Customer Service 800.364.9897 Technical Support 888.526.5351 1180 E. Ellsworth Rd Ann Arbor, MI USA TABLE OF CONTENTS GENERAL INFORMATION
LDL Uptake Cell-Based Assay Kit Item No. 10011125 www.caymanchem.com Customer Service 800.364.9897 Technical Support 888.526.5351 1180 E. Ellsworth Rd Ann Arbor, MI USA TABLE OF CONTENTS GENERAL INFORMATION
CHAPTER 5 RESULTS Previous study: cell culture and organotypical slices
 45 CHAPTER 5 RESULTS 5.1. Previous study: cell culture and organotypical slices Initial experiments have been conducted to ensure that the tet-on system works. A neuronal cell culture from mice expressing
45 CHAPTER 5 RESULTS 5.1. Previous study: cell culture and organotypical slices Initial experiments have been conducted to ensure that the tet-on system works. A neuronal cell culture from mice expressing
(A) PCR primers (arrows) designed to distinguish wild type (P1+P2), targeted (P1+P2) and excised (P1+P3)14-
 PCR primers (arrows) designed to distinguish wild type (P1+P2), targeted (P1+P2) and excised (P1+P3)14-") 1 Supplemental Figure Legends Figure S1. Mammary tumors of ErbB2 KI mice with 14-3-3σ ablation have elevated ErbB2 transcript levels and cell proliferation (A) PCR primers (arrows) designed to distinguish
1 Supplemental Figure Legends Figure S1. Mammary tumors of ErbB2 KI mice with 14-3-3σ ablation have elevated ErbB2 transcript levels and cell proliferation (A) PCR primers (arrows) designed to distinguish
General Laboratory methods Plasma analysis: Gene Expression Analysis: Immunoblot analysis: Immunohistochemistry:
 General Laboratory methods Plasma analysis: Plasma insulin (Mercodia, Sweden), leptin (duoset, R&D Systems Europe, Abingdon, United Kingdom), IL-6, TNFα and adiponectin levels (Quantikine kits, R&D Systems
General Laboratory methods Plasma analysis: Plasma insulin (Mercodia, Sweden), leptin (duoset, R&D Systems Europe, Abingdon, United Kingdom), IL-6, TNFα and adiponectin levels (Quantikine kits, R&D Systems
TFEB-mediated increase in peripheral lysosomes regulates. Store Operated Calcium Entry
 TFEB-mediated increase in peripheral lysosomes regulates Store Operated Calcium Entry Luigi Sbano, Massimo Bonora, Saverio Marchi, Federica Baldassari, Diego L. Medina, Andrea Ballabio, Carlotta Giorgi
TFEB-mediated increase in peripheral lysosomes regulates Store Operated Calcium Entry Luigi Sbano, Massimo Bonora, Saverio Marchi, Federica Baldassari, Diego L. Medina, Andrea Ballabio, Carlotta Giorgi
Product Information Sheet (M ) MarkerGene TM LysoLive TM Lysosomal Sulfatase Assay Kit (Product M1377)
 MarkerGene TM LysoLive TM Lysosomal Sulfatase Assay Kit (Product M1377)") 1 Product Information Sheet (M1377-004) MarkerGene TM LysoLive TM Lysosomal Sulfatase Assay Kit (Product M1377) Marker Gene Technologies, Inc. University of Oregon Riverfront Research Park 1850 Millrace
1 Product Information Sheet (M1377-004) MarkerGene TM LysoLive TM Lysosomal Sulfatase Assay Kit (Product M1377) Marker Gene Technologies, Inc. University of Oregon Riverfront Research Park 1850 Millrace
LDL Uptake Cell-Based Assay Kit
 LDL Uptake Cell-Based Assay Kit Catalog Number KA1327 100 assays Version: 07 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 Principle of the Assay...
LDL Uptake Cell-Based Assay Kit Catalog Number KA1327 100 assays Version: 07 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 Principle of the Assay...
ab CytoPainter Golgi/ER Staining Kit
 ab139485 CytoPainter Golgi/ER Staining Kit Instructions for Use Designed to detect Golgi bodies and endoplasmic reticulum by microscopy This product is for research use only and is not intended for diagnostic
ab139485 CytoPainter Golgi/ER Staining Kit Instructions for Use Designed to detect Golgi bodies and endoplasmic reticulum by microscopy This product is for research use only and is not intended for diagnostic
ab LDL Uptake Assay Kit (Cell-Based)
") ab133127 LDL Uptake Assay Kit (Cell-Based) Instructions for Use For the detection of LDL uptake into cultured cells. This product is for research use only and is not intended for diagnostic use. Version
ab133127 LDL Uptake Assay Kit (Cell-Based) Instructions for Use For the detection of LDL uptake into cultured cells. This product is for research use only and is not intended for diagnostic use. Version
Microglia, Inflammation, and FTD
 FTD Minicourse April, 2009 Microglia, Inflammation, and FTD Li Gan, Ph.D Gladstone Institute of Neurological Disease University of California, San Francisco Outline Why study inflammation in neurodegeneration?
FTD Minicourse April, 2009 Microglia, Inflammation, and FTD Li Gan, Ph.D Gladstone Institute of Neurological Disease University of California, San Francisco Outline Why study inflammation in neurodegeneration?
Supplementary Figures
 Inhibition of Pulmonary Anti Bacterial Defense by IFN γ During Recovery from Influenza Infection By Keer Sun and Dennis W. Metzger Supplementary Figures d a Ly6G Percentage survival f 1 75 5 1 25 1 5 1
Inhibition of Pulmonary Anti Bacterial Defense by IFN γ During Recovery from Influenza Infection By Keer Sun and Dennis W. Metzger Supplementary Figures d a Ly6G Percentage survival f 1 75 5 1 25 1 5 1
Supplementary Figure S1. Venn diagram analysis of mrna microarray data and mirna target analysis. (a) Western blot analysis of T lymphoblasts (CLS)
 Western blot analysis of T lymphoblasts (CLS)") Supplementary Figure S1. Venn diagram analysis of mrna microarray data and mirna target analysis. (a) Western blot analysis of T lymphoblasts (CLS) and their exosomes (EXO) in resting (REST) and activated
Supplementary Figure S1. Venn diagram analysis of mrna microarray data and mirna target analysis. (a) Western blot analysis of T lymphoblasts (CLS) and their exosomes (EXO) in resting (REST) and activated
Determination of the temporal pattern and importance of BALF1 expression in Epstein-Barr viral infection
 Determination of the temporal pattern and importance of BALF1 expression in Epstein-Barr viral infection Melissa Mihelidakis May 6, 2004 7.340 Research Proposal Introduction Apoptosis, or programmed cell
Determination of the temporal pattern and importance of BALF1 expression in Epstein-Barr viral infection Melissa Mihelidakis May 6, 2004 7.340 Research Proposal Introduction Apoptosis, or programmed cell
ab SREBP-2 Translocation Assay Kit (Cell-Based)
") ab133114 SREBP-2 Translocation Assay Kit (Cell-Based) Instructions for Use For analysis of translocation of SREBP-2 into nuclei. This product is for research use only and is not intended for diagnostic
ab133114 SREBP-2 Translocation Assay Kit (Cell-Based) Instructions for Use For analysis of translocation of SREBP-2 into nuclei. This product is for research use only and is not intended for diagnostic
Supplementary Information Titles Journal: Nature Medicine
 Supplementary Information Titles Journal: Nature Medicine Article Title: Corresponding Author: Supplementary Item & Number Supplementary Fig.1 Fig.2 Fig.3 Fig.4 Fig.5 Fig.6 Fig.7 Fig.8 Fig.9 Fig. Fig.11
Supplementary Information Titles Journal: Nature Medicine Article Title: Corresponding Author: Supplementary Item & Number Supplementary Fig.1 Fig.2 Fig.3 Fig.4 Fig.5 Fig.6 Fig.7 Fig.8 Fig.9 Fig. Fig.11
Supplementary Information
 Supplementary Information Supplementary Figure 1. CD4 + T cell activation and lack of apoptosis after crosslinking with anti-cd3 + anti-cd28 + anti-cd160. (a) Flow cytometry of anti-cd160 (5D.10A11) binding
Supplementary Information Supplementary Figure 1. CD4 + T cell activation and lack of apoptosis after crosslinking with anti-cd3 + anti-cd28 + anti-cd160. (a) Flow cytometry of anti-cd160 (5D.10A11) binding
Bone marrow-derived mesenchymal stem cells improve diabetes-induced cognitive impairment by
 Nakano et al. Supplementary information 1. Supplementary Figure 2. Methods 3. References Bone marrow-derived mesenchymal stem cells improve diabetes-induced cognitive impairment by exosome transfer into
Nakano et al. Supplementary information 1. Supplementary Figure 2. Methods 3. References Bone marrow-derived mesenchymal stem cells improve diabetes-induced cognitive impairment by exosome transfer into
Mammalian Membrane Protein Extraction Kit
 Mammalian Membrane Protein Extraction Kit Catalog number: AR0155 Boster s Mammalian Membrane Protein Extraction Kit is a simple, rapid and reproducible method to prepare cellular protein fractions highly
Mammalian Membrane Protein Extraction Kit Catalog number: AR0155 Boster s Mammalian Membrane Protein Extraction Kit is a simple, rapid and reproducible method to prepare cellular protein fractions highly
Expression of acid base transporters in the kidney collecting duct in Slc2a7 -/-
 Supplemental Material Results. Expression of acid base transporters in the kidney collecting duct in Slc2a7 -/- and Slc2a7 -/- mice. The expression of AE1 in the kidney was examined in Slc26a7 KO mice.
Supplemental Material Results. Expression of acid base transporters in the kidney collecting duct in Slc2a7 -/- and Slc2a7 -/- mice. The expression of AE1 in the kidney was examined in Slc26a7 KO mice.
PRODUCT INFORMATION & MANUAL
 PRODUCT INFORMATION & MANUAL 0.4 micron for Overall Exosome Isolation (Cell Media) NBP2-49826 For research use only. Not for diagnostic or therapeutic procedures. www.novusbio.com - P: 303.730.1950 - P:
PRODUCT INFORMATION & MANUAL 0.4 micron for Overall Exosome Isolation (Cell Media) NBP2-49826 For research use only. Not for diagnostic or therapeutic procedures. www.novusbio.com - P: 303.730.1950 - P:
HIV-1 Virus-like Particle Budding Assay Nathan H Vande Burgt, Luis J Cocka * and Paul Bates
 HIV-1 Virus-like Particle Budding Assay Nathan H Vande Burgt, Luis J Cocka * and Paul Bates Department of Microbiology, Perelman School of Medicine at the University of Pennsylvania, Philadelphia, USA
HIV-1 Virus-like Particle Budding Assay Nathan H Vande Burgt, Luis J Cocka * and Paul Bates Department of Microbiology, Perelman School of Medicine at the University of Pennsylvania, Philadelphia, USA
PBS Class #2 Introduction to the Immune System part II Suggested reading: Abbas, pgs , 27-30
 PBS 803 - Class #2 Introduction to the Immune System part II Suggested reading: Abbas, pgs. 15-25, 27-30 Learning Objectives Compare and contrast the maturation of B and T lymphocytes Compare and contrast
PBS 803 - Class #2 Introduction to the Immune System part II Suggested reading: Abbas, pgs. 15-25, 27-30 Learning Objectives Compare and contrast the maturation of B and T lymphocytes Compare and contrast
Lipid (Oil Red O) staining Kit
 staining Kit") Lipid (Oil Red O) staining Kit Catalog Number KA4541 1 Kit Version: 02 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 General Information... 4 Materials
Lipid (Oil Red O) staining Kit Catalog Number KA4541 1 Kit Version: 02 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 General Information... 4 Materials
Medical Virology Immunology. Dr. Sameer Naji, MB, BCh, PhD (UK) Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University
 Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University") Medical Virology Immunology Dr. Sameer Naji, MB, BCh, PhD (UK) Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University Human blood cells Phases of immune responses Microbe Naïve
Medical Virology Immunology Dr. Sameer Naji, MB, BCh, PhD (UK) Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University Human blood cells Phases of immune responses Microbe Naïve
Supplemental Information. Tissue Myeloid Progenitors Differentiate. into Pericytes through TGF-b Signaling. in Developing Skin Vasculature
 Cell Reports, Volume 18 Supplemental Information Tissue Myeloid Progenitors Differentiate into Pericytes through TGF-b Signaling in Developing Skin Vasculature Tomoko Yamazaki, Ani Nalbandian, Yutaka Uchida,
Cell Reports, Volume 18 Supplemental Information Tissue Myeloid Progenitors Differentiate into Pericytes through TGF-b Signaling in Developing Skin Vasculature Tomoko Yamazaki, Ani Nalbandian, Yutaka Uchida,
Phosphate buffered saline (PBS) for washing the cells TE buffer (nuclease-free) ph 7.5 for use with the PrimePCR Reverse Transcription Control Assay
 for washing the cells TE buffer (nuclease-free) ph 7.5 for use with the PrimePCR Reverse Transcription Control Assay") Catalog # Description 172-5080 SingleShot Cell Lysis Kit, 100 x 50 µl reactions 172-5081 SingleShot Cell Lysis Kit, 500 x 50 µl reactions For research purposes only. Introduction The SingleShot Cell Lysis
Catalog # Description 172-5080 SingleShot Cell Lysis Kit, 100 x 50 µl reactions 172-5081 SingleShot Cell Lysis Kit, 500 x 50 µl reactions For research purposes only. Introduction The SingleShot Cell Lysis
SUPPLEMENTARY INFORMATION
 Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
Supplementary data Supplementary Figure 1 Supplementary Figure 2
 Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
FOCUS SubCell. For the Enrichment of Subcellular Fractions. (Cat. # ) think proteins! think G-Biosciences
 think proteins! think G-Biosciences") 169PR 01 G-Biosciences 1-800-628-7730 1-314-991-6034 technical@gbiosciences.com A Geno Technology, Inc. (USA) brand name FOCUS SubCell For the Enrichment of Subcellular Fractions (Cat. # 786 260) think
169PR 01 G-Biosciences 1-800-628-7730 1-314-991-6034 technical@gbiosciences.com A Geno Technology, Inc. (USA) brand name FOCUS SubCell For the Enrichment of Subcellular Fractions (Cat. # 786 260) think
LDL Uptake Flow Cytometry Assay Kit
 LDL Uptake Flow Cytometry Assay Kit Item No. 601470 www.caymanchem.com Customer Service 800.364.9897 Technical Support 888.526.5351 1180 E. Ellsworth Rd Ann Arbor, MI USA TABLE OF CONTENTS GENERAL INFORMATION
LDL Uptake Flow Cytometry Assay Kit Item No. 601470 www.caymanchem.com Customer Service 800.364.9897 Technical Support 888.526.5351 1180 E. Ellsworth Rd Ann Arbor, MI USA TABLE OF CONTENTS GENERAL INFORMATION
Supplementary Table 1. Primer sequences for conventional RT-PCR on mouse islets
 Supplementary Table 1. Primer sequences for conventional RT-PCR on mouse islets Gene 5 Forward 3 5 Reverse 3.T. Product (bp) ( C) mnox1 GTTCTTGGGCTGCCTTGG GCTGGGGCGGCGG 60 300 mnoxa1 GCTTTGCCGCGTGC GGTTCGGGTCCTTTGTGC
Supplementary Table 1. Primer sequences for conventional RT-PCR on mouse islets Gene 5 Forward 3 5 Reverse 3.T. Product (bp) ( C) mnox1 GTTCTTGGGCTGCCTTGG GCTGGGGCGGCGG 60 300 mnoxa1 GCTTTGCCGCGTGC GGTTCGGGTCCTTTGTGC
Supplemental Information. T Cells Enhance Autoimmunity by Restraining Regulatory T Cell Responses via an Interleukin-23-Dependent Mechanism
 Immunity, Volume 33 Supplemental Information T Cells Enhance Autoimmunity by Restraining Regulatory T Cell Responses via an Interleukin-23-Dependent Mechanism Franziska Petermann, Veit Rothhammer, Malte
Immunity, Volume 33 Supplemental Information T Cells Enhance Autoimmunity by Restraining Regulatory T Cell Responses via an Interleukin-23-Dependent Mechanism Franziska Petermann, Veit Rothhammer, Malte
Supplementary Figure 1
 Supplementary Figure 1 AAV-GFP injection in the MEC of the mouse brain C57Bl/6 mice at 4 months of age were injected with AAV-GFP into the MEC and sacrificed at 7 days post injection (dpi). (a) Brains
Supplementary Figure 1 AAV-GFP injection in the MEC of the mouse brain C57Bl/6 mice at 4 months of age were injected with AAV-GFP into the MEC and sacrificed at 7 days post injection (dpi). (a) Brains
p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO
 Supplementary Information p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO Yuri Shibata, Masaaki Oyama, Hiroko Kozuka-Hata, Xiao Han, Yuetsu Tanaka,
Supplementary Information p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO Yuri Shibata, Masaaki Oyama, Hiroko Kozuka-Hata, Xiao Han, Yuetsu Tanaka,
Doctor of Philosophy
 Regulation of Gene Expression of the 25-Hydroxyvitamin D la-hydroxylase (CYP27BI) Promoter: Study of A Transgenic Mouse Model Ivanka Hendrix School of Molecular and Biomedical Science The University of
Regulation of Gene Expression of the 25-Hydroxyvitamin D la-hydroxylase (CYP27BI) Promoter: Study of A Transgenic Mouse Model Ivanka Hendrix School of Molecular and Biomedical Science The University of
Immune System. Biol 105 Chapter 13
 Immune System Biol 105 Chapter 13 Outline Immune System I. Function of the Immune system II. Barrier Defenses III. Nonspecific Defenses A. Immune system cells B. Inflammatory response C. Complementary
Immune System Biol 105 Chapter 13 Outline Immune System I. Function of the Immune system II. Barrier Defenses III. Nonspecific Defenses A. Immune system cells B. Inflammatory response C. Complementary
CHAPTER 1 INTRODUCTION
 1 CHAPTER 1 INTRODUCTION 1.1. BACKGROUND Fragile X mental retardation 1 gene (FMR1) is located on the X chromosome and is responsible for producing the fragile X mental retardation protein (FMRP) which
1 CHAPTER 1 INTRODUCTION 1.1. BACKGROUND Fragile X mental retardation 1 gene (FMR1) is located on the X chromosome and is responsible for producing the fragile X mental retardation protein (FMRP) which
General information. Cell mediated immunity. 455 LSA, Tuesday 11 to noon. Anytime after class.
 General information Cell mediated immunity 455 LSA, Tuesday 11 to noon Anytime after class T-cell precursors Thymus Naive T-cells (CD8 or CD4) email: lcoscoy@berkeley.edu edu Use MCB150 as subject line
General information Cell mediated immunity 455 LSA, Tuesday 11 to noon Anytime after class T-cell precursors Thymus Naive T-cells (CD8 or CD4) email: lcoscoy@berkeley.edu edu Use MCB150 as subject line
7.013 Spring 2005 Problem Set 6
 MIT Department of Biology 7.013: Introductory Biology - Spring 2005 Instructors: Professor Hazel Sive, Professor Tyler Jacks, Dr. Claudette Gardel NAME TA 7.013 Spring 2005 Problem Set 6 FRIDAY April 29th,
MIT Department of Biology 7.013: Introductory Biology - Spring 2005 Instructors: Professor Hazel Sive, Professor Tyler Jacks, Dr. Claudette Gardel NAME TA 7.013 Spring 2005 Problem Set 6 FRIDAY April 29th,
Novel Targets of disease modifying therapy for Parkinson disease. David G. Standaert, MD, PhD John N. Whitaker Professor and Chair of Neurology
 Novel Targets of disease modifying therapy for Parkinson disease David G. Standaert, MD, PhD John N. Whitaker Professor and Chair of Neurology Disclosures Dr. Standaert has served as a paid consultant
Novel Targets of disease modifying therapy for Parkinson disease David G. Standaert, MD, PhD John N. Whitaker Professor and Chair of Neurology Disclosures Dr. Standaert has served as a paid consultant
Genetic Analysis of Anxiety Related Behaviors by Gene Chip and In situ Hybridization of the Hippocampus and Amygdala of C57BL/6J and AJ Mice Brains
 Genetic Analysis of Anxiety Related Behaviors by Gene Chip and In situ Hybridization of the Hippocampus and Amygdala of C57BL/6J and AJ Mice Brains INTRODUCTION To study the relationship between an animal's
Genetic Analysis of Anxiety Related Behaviors by Gene Chip and In situ Hybridization of the Hippocampus and Amygdala of C57BL/6J and AJ Mice Brains INTRODUCTION To study the relationship between an animal's
Human Oxidized LDL ELISA Kit (MDA-LDL Quantitation), General
, General") Human Oxidized LDL ELISA Kit (MDA-LDL Quantitation), General For the detection and quantitation of human OxLDL in plasma, serum or other biological fluid samples Cat. No. KT-959 For Research Use Only.
Human Oxidized LDL ELISA Kit (MDA-LDL Quantitation), General For the detection and quantitation of human OxLDL in plasma, serum or other biological fluid samples Cat. No. KT-959 For Research Use Only.
Electrolytes. Summary: (This area will include a brief description of what the protocol is used for and why someone would need to use it.
 Electrolytes Version: 1 Edited by: Jason Kim (note that the following list should be linked to the appropriate location.) Summary Reagents and Materials Protocol Reagent Preparation Reagent 1 Reagent 2
Electrolytes Version: 1 Edited by: Jason Kim (note that the following list should be linked to the appropriate location.) Summary Reagents and Materials Protocol Reagent Preparation Reagent 1 Reagent 2
SUPPORTING MATREALS. Methods and Materials
 SUPPORTING MATREALS Methods and Materials Cell Culture MC3T3-E1 (subclone 4) cells were maintained in -MEM with 10% FBS, 1% Pen/Strep at 37ºC in a humidified incubator with 5% CO2. MC3T3 cell differentiation
SUPPORTING MATREALS Methods and Materials Cell Culture MC3T3-E1 (subclone 4) cells were maintained in -MEM with 10% FBS, 1% Pen/Strep at 37ºC in a humidified incubator with 5% CO2. MC3T3 cell differentiation
A Hepatocyte Growth Factor Receptor (Met) Insulin Receptor hybrid governs hepatic glucose metabolism SUPPLEMENTARY FIGURES, LEGENDS AND METHODS
 Insulin Receptor hybrid governs hepatic glucose metabolism SUPPLEMENTARY FIGURES, LEGENDS AND METHODS") A Hepatocyte Growth Factor Receptor (Met) Insulin Receptor hybrid governs hepatic glucose metabolism Arlee Fafalios, Jihong Ma, Xinping Tan, John Stoops, Jianhua Luo, Marie C. DeFrances and Reza Zarnegar
A Hepatocyte Growth Factor Receptor (Met) Insulin Receptor hybrid governs hepatic glucose metabolism Arlee Fafalios, Jihong Ma, Xinping Tan, John Stoops, Jianhua Luo, Marie C. DeFrances and Reza Zarnegar
Cells and reagents. Synaptopodin knockdown (1) and dynamin knockdown (2)
 and dynamin knockdown (2)") Supplemental Methods Cells and reagents. Synaptopodin knockdown (1) and dynamin knockdown (2) podocytes were cultured as described previously. Staurosporine, angiotensin II and actinomycin D were all obtained
Supplemental Methods Cells and reagents. Synaptopodin knockdown (1) and dynamin knockdown (2) podocytes were cultured as described previously. Staurosporine, angiotensin II and actinomycin D were all obtained
Exosomes Immunocapture and Isolation Tools: Immunoplates
 Product Insert Exosomes Immunocapture and Isolation Tools: Immunoplates HBM-POF-##/## HBM-POC-##/## Overall Exosome capture from Biological fluids (plasma, urine) Overall Exosome capture from Cell culture
Product Insert Exosomes Immunocapture and Isolation Tools: Immunoplates HBM-POF-##/## HBM-POC-##/## Overall Exosome capture from Biological fluids (plasma, urine) Overall Exosome capture from Cell culture
Supplementary Materials for
 immunology.sciencemag.org/cgi/content/full/2/16/eaan6049/dc1 Supplementary Materials for Enzymatic synthesis of core 2 O-glycans governs the tissue-trafficking potential of memory CD8 + T cells Jossef
immunology.sciencemag.org/cgi/content/full/2/16/eaan6049/dc1 Supplementary Materials for Enzymatic synthesis of core 2 O-glycans governs the tissue-trafficking potential of memory CD8 + T cells Jossef
Tolerance, autoimmunity and the pathogenesis of immunemediated inflammatory diseases. Abul K. Abbas UCSF
 Tolerance, autoimmunity and the pathogenesis of immunemediated inflammatory diseases Abul K. Abbas UCSF Balancing lymphocyte activation and control Activation Effector T cells Tolerance Regulatory T cells
Tolerance, autoimmunity and the pathogenesis of immunemediated inflammatory diseases Abul K. Abbas UCSF Balancing lymphocyte activation and control Activation Effector T cells Tolerance Regulatory T cells
Immune System. Biol 105 Lecture 16 Chapter 13
 Immune System Biol 105 Lecture 16 Chapter 13 Outline Immune System I. Function of the Immune system II. Barrier Defenses III. Nonspecific Defenses A. Immune system cells B. Inflammatory response C. Complementary
Immune System Biol 105 Lecture 16 Chapter 13 Outline Immune System I. Function of the Immune system II. Barrier Defenses III. Nonspecific Defenses A. Immune system cells B. Inflammatory response C. Complementary
LESSON 4.4 WORKBOOK. How viruses make us sick: Viral Replication
 DEFINITIONS OF TERMS Eukaryotic: Non-bacterial cell type (bacteria are prokaryotes).. LESSON 4.4 WORKBOOK How viruses make us sick: Viral Replication This lesson extends the principles we learned in Unit
DEFINITIONS OF TERMS Eukaryotic: Non-bacterial cell type (bacteria are prokaryotes).. LESSON 4.4 WORKBOOK How viruses make us sick: Viral Replication This lesson extends the principles we learned in Unit
Fayth K. Yoshimura, Ph.D. September 7, of 7 HIV - BASIC PROPERTIES
 1 of 7 I. Viral Origin. A. Retrovirus - animal lentiviruses. HIV - BASIC PROPERTIES 1. HIV is a member of the Retrovirus family and more specifically it is a member of the Lentivirus genus of this family.
1 of 7 I. Viral Origin. A. Retrovirus - animal lentiviruses. HIV - BASIC PROPERTIES 1. HIV is a member of the Retrovirus family and more specifically it is a member of the Lentivirus genus of this family.
E.Z.N.A. SQ Blood DNA Kit II. Table of Contents
 E.Z.N.A. SQ Blood DNA Kit II Table of Contents Introduction and Overview...2 Kit Contents/Storage and Stability...3 Blood Storage and DNA Yield...4 Preparing Reagents...5 100-500 μl Whole Blood Protocol...6
E.Z.N.A. SQ Blood DNA Kit II Table of Contents Introduction and Overview...2 Kit Contents/Storage and Stability...3 Blood Storage and DNA Yield...4 Preparing Reagents...5 100-500 μl Whole Blood Protocol...6
Supplementary Figure 1. Characterization of NMuMG-ErbB2 and NIC breast cancer cells expressing shrnas targeting LPP. NMuMG-ErbB2 cells (a) and NIC
 and NIC") Supplementary Figure 1. Characterization of NMuMG-ErbB2 and NIC breast cancer cells expressing shrnas targeting LPP. NMuMG-ErbB2 cells (a) and NIC cells (b) were engineered to stably express either a LucA-shRNA
Supplementary Figure 1. Characterization of NMuMG-ErbB2 and NIC breast cancer cells expressing shrnas targeting LPP. NMuMG-ErbB2 cells (a) and NIC cells (b) were engineered to stably express either a LucA-shRNA
pplementary Figur Supplementary Figure 1. a.
 pplementary Figur Supplementary Figure 1. a. Quantification by RT-qPCR of YFV-17D and YFV-17D pol- (+) RNA in the supernatant of cultured Huh7.5 cells following viral RNA electroporation of respective
pplementary Figur Supplementary Figure 1. a. Quantification by RT-qPCR of YFV-17D and YFV-17D pol- (+) RNA in the supernatant of cultured Huh7.5 cells following viral RNA electroporation of respective
Immune System AP SBI4UP
 Immune System AP SBI4UP TYPES OF IMMUNITY INNATE IMMUNITY ACQUIRED IMMUNITY EXTERNAL DEFENCES INTERNAL DEFENCES HUMORAL RESPONSE Skin Phagocytic Cells CELL- MEDIATED RESPONSE Mucus layer Antimicrobial
Immune System AP SBI4UP TYPES OF IMMUNITY INNATE IMMUNITY ACQUIRED IMMUNITY EXTERNAL DEFENCES INTERNAL DEFENCES HUMORAL RESPONSE Skin Phagocytic Cells CELL- MEDIATED RESPONSE Mucus layer Antimicrobial
OxiSelect HNE-His Adduct ELISA Kit
 Product Manual OxiSelect HNE-His Adduct ELISA Kit Catalog Number STA-334 96 assays FOR RESEARCH USE ONLY Not for use in diagnostic procedures Introduction Lipid peroxidation is a well-defined mechanism
Product Manual OxiSelect HNE-His Adduct ELISA Kit Catalog Number STA-334 96 assays FOR RESEARCH USE ONLY Not for use in diagnostic procedures Introduction Lipid peroxidation is a well-defined mechanism
Influenza virus exploits tunneling nanotubes for cell-to-cell spread
 Supplementary Information Influenza virus exploits tunneling nanotubes for cell-to-cell spread Amrita Kumar 1, Jin Hyang Kim 1, Priya Ranjan 1, Maureen G. Metcalfe 2, Weiping Cao 1, Margarita Mishina 1,
Supplementary Information Influenza virus exploits tunneling nanotubes for cell-to-cell spread Amrita Kumar 1, Jin Hyang Kim 1, Priya Ranjan 1, Maureen G. Metcalfe 2, Weiping Cao 1, Margarita Mishina 1,
ab Lysosome/Cytotoxicity Dual Staining Kit
 ab133078 Lysosome/Cytotoxicity Dual Staining Kit Instructions for Use For studying lysosome function at the cellular level. This product is for research use only and is not intended for diagnostic use.
ab133078 Lysosome/Cytotoxicity Dual Staining Kit Instructions for Use For studying lysosome function at the cellular level. This product is for research use only and is not intended for diagnostic use.
PROTOCOL: OPTIMIZATION OF LENTIVIRAL TRANSDUCTION USING SPINFECTION
 Last Modified: April 2018 Last Review: October 2018 PROTOCOL: OPTIMIZATION OF LENTIVIRAL TRANSDUCTION USING SPINFECTION Table of Contents 1. Brief Description 1 2. Materials and Reagents.1 3. Optimization
Last Modified: April 2018 Last Review: October 2018 PROTOCOL: OPTIMIZATION OF LENTIVIRAL TRANSDUCTION USING SPINFECTION Table of Contents 1. Brief Description 1 2. Materials and Reagents.1 3. Optimization
Data Sheet TIGIT / NFAT Reporter - Jurkat Cell Line Catalog #60538
 Data Sheet TIGIT / NFAT Reporter - Jurkat Cell Line Catalog #60538 Background: TIGIT is a co-inhibitory receptor that is highly expressed in Natural Killer (NK) cells, activated CD4+, CD8+ and regulatory
Data Sheet TIGIT / NFAT Reporter - Jurkat Cell Line Catalog #60538 Background: TIGIT is a co-inhibitory receptor that is highly expressed in Natural Killer (NK) cells, activated CD4+, CD8+ and regulatory
Potential Atheroprotective Effects of Ixmyelocel-T Cellular Therapy. Kelly J. Ledford, Nikki Murphy, Frank Zeigler, Ronnda L.
 Potential Atheroprotective Effects of Ixmyelocel-T Cellular Therapy Kelly J. Ledford, Nikki Murphy, Frank Zeigler, Ronnda L. Bartel 1 Ixmyelocel-T, an expanded, autologous multicellular therapy cultured
Potential Atheroprotective Effects of Ixmyelocel-T Cellular Therapy Kelly J. Ledford, Nikki Murphy, Frank Zeigler, Ronnda L. Bartel 1 Ixmyelocel-T, an expanded, autologous multicellular therapy cultured
Lymphoid System: cells of the immune system. Answer Sheet
 Lymphoid System: cells of the immune system Answer Sheet Q1 Which areas of the lymph node have most CD3 staining? A1 Most CD3 staining is present in the paracortex (T cell areas). This is towards the outside
Lymphoid System: cells of the immune system Answer Sheet Q1 Which areas of the lymph node have most CD3 staining? A1 Most CD3 staining is present in the paracortex (T cell areas). This is towards the outside
Multi-Parameter Apoptosis Assay Kit
 Multi-Parameter Apoptosis Assay Kit Catalog Number KA1335 5 x 96 assays Version: 05 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 Principle of the Assay...
Multi-Parameter Apoptosis Assay Kit Catalog Number KA1335 5 x 96 assays Version: 05 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 Principle of the Assay...
Supplemental Materials and Methods Plasmids and viruses Quantitative Reverse Transcription PCR Generation of molecular standard for quantitative PCR
 Supplemental Materials and Methods Plasmids and viruses To generate pseudotyped viruses, the previously described recombinant plasmids pnl4-3-δnef-gfp or pnl4-3-δ6-drgfp and a vector expressing HIV-1 X4
Supplemental Materials and Methods Plasmids and viruses To generate pseudotyped viruses, the previously described recombinant plasmids pnl4-3-δnef-gfp or pnl4-3-δ6-drgfp and a vector expressing HIV-1 X4
marker. DAPI labels nuclei. Flies were 20 days old. Scale bar is 5 µm. Ctrl is
 Supplementary Figure 1. (a) Nos is detected in glial cells in both control and GFAP R79H transgenic flies (arrows), but not in deletion mutant Nos Δ15 animals. Repo is a glial cell marker. DAPI labels
Supplementary Figure 1. (a) Nos is detected in glial cells in both control and GFAP R79H transgenic flies (arrows), but not in deletion mutant Nos Δ15 animals. Repo is a glial cell marker. DAPI labels
Cell isolation. Spleen and lymph nodes (axillary, inguinal) were removed from mice
 were removed from mice") Supplementary Methods: Cell isolation. Spleen and lymph nodes (axillary, inguinal) were removed from mice and gently meshed in DMEM containing 10% FBS to prepare for single cell suspensions. CD4 + CD25
Supplementary Methods: Cell isolation. Spleen and lymph nodes (axillary, inguinal) were removed from mice and gently meshed in DMEM containing 10% FBS to prepare for single cell suspensions. CD4 + CD25
AAV9 intracerebroventricular gene therapy improves lifespan, locomotor function and pathology in a mouse model of Niemann Pick type C1 disease
 Human Molecular Genetics, 2018, Vol. 27, No. 17 3079 3098 doi: 10.1093/hmg/ddy212 Advance Access Publication Date: 5 June 2018 Original Article ORIGINAL ARTICLE AAV9 intracerebroventricular gene therapy
Human Molecular Genetics, 2018, Vol. 27, No. 17 3079 3098 doi: 10.1093/hmg/ddy212 Advance Access Publication Date: 5 June 2018 Original Article ORIGINAL ARTICLE AAV9 intracerebroventricular gene therapy
effect on the upregulation of these cell surface markers. The mean peak fluorescence intensity
 SUPPLEMENTARY FIGURE 1 Supplementary Figure 1 ASIC1 disruption or blockade does not effect in vitro and in vivo antigen-presenting cell activation. (a) Flow cytometric analysis of cell surface molecules
SUPPLEMENTARY FIGURE 1 Supplementary Figure 1 ASIC1 disruption or blockade does not effect in vitro and in vivo antigen-presenting cell activation. (a) Flow cytometric analysis of cell surface molecules
Programmed necrosis, not apoptosis, is a key mediator of cell loss and DAMP-mediated inflammation in dsrna-induced retinal degeneration
 Programmed necrosis, not apoptosis, is a key mediator of cell loss and DAMP-mediated inflammation in dsrna-induced retinal degeneration The Harvard community has made this article openly available. Please
Programmed necrosis, not apoptosis, is a key mediator of cell loss and DAMP-mediated inflammation in dsrna-induced retinal degeneration The Harvard community has made this article openly available. Please
7-AAD/CFSE Cell-Mediated Cytotoxicity Assay Kit
 7-AAD/CFSE Cell-Mediated Cytotoxicity Assay Kit Catalog Number KA1293 96 assays Version: 02 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 Principle of
7-AAD/CFSE Cell-Mediated Cytotoxicity Assay Kit Catalog Number KA1293 96 assays Version: 02 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 Principle of
Innate Immunity. Bởi: OpenStaxCollege
 Innate Immunity Bởi: OpenStaxCollege The vertebrate, including human, immune system is a complex multilayered system for defending against external and internal threats to the integrity of the body. The
Innate Immunity Bởi: OpenStaxCollege The vertebrate, including human, immune system is a complex multilayered system for defending against external and internal threats to the integrity of the body. The
Online Appendix Material and Methods: Pancreatic RNA isolation and quantitative real-time (q)rt-pcr. Mice were fasted overnight and killed 1 hour (h)
rt-pcr. Mice were fasted overnight and killed 1 hour (h)") Online Appendix Material and Methods: Pancreatic RNA isolation and quantitative real-time (q)rt-pcr. Mice were fasted overnight and killed 1 hour (h) after feeding. A small slice (~5-1 mm 3 ) was taken
Online Appendix Material and Methods: Pancreatic RNA isolation and quantitative real-time (q)rt-pcr. Mice were fasted overnight and killed 1 hour (h) after feeding. A small slice (~5-1 mm 3 ) was taken
Zool 3200: Cell Biology Exam 4 Part II 2/3/15
 Name:Key Trask Zool 3200: Cell Biology Exam 4 Part II 2/3/15 Answer each of the following questions in the space provided, explaining your answers when asked to do so; circle the correct answer or answers
Name:Key Trask Zool 3200: Cell Biology Exam 4 Part II 2/3/15 Answer each of the following questions in the space provided, explaining your answers when asked to do so; circle the correct answer or answers
Supporting Information
 Supporting Information Desnues et al. 10.1073/pnas.1314121111 SI Materials and Methods Mice. Toll-like receptor (TLR)8 / and TLR9 / mice were generated as described previously (1, 2). TLR9 / mice were
Supporting Information Desnues et al. 10.1073/pnas.1314121111 SI Materials and Methods Mice. Toll-like receptor (TLR)8 / and TLR9 / mice were generated as described previously (1, 2). TLR9 / mice were
Choline Assay Kit (Fluorometric)
") Product Manual Choline Assay Kit (Fluorometric) Catalog Number MET- 5042 96 assays FOR RESEARCH USE ONLY Not for use in diagnostic procedures Introduction Choline is a water soluble amine that is an essential
Product Manual Choline Assay Kit (Fluorometric) Catalog Number MET- 5042 96 assays FOR RESEARCH USE ONLY Not for use in diagnostic procedures Introduction Choline is a water soluble amine that is an essential
Exosomes Immunocapture and Isolation Tools: Immunobeads
 Product Insert Exosomes Immunocapture and Isolation Tools: Immunobeads HBM-BOLF-##/##-# HBM-BOLC-##/##-# HBM-BTLF-##/##-# Overall Exosome capture from Human Biological fluids Overall Exosome capture from
Product Insert Exosomes Immunocapture and Isolation Tools: Immunobeads HBM-BOLF-##/##-# HBM-BOLC-##/##-# HBM-BTLF-##/##-# Overall Exosome capture from Human Biological fluids Overall Exosome capture from
SUPPLEMENT. Materials and methods
 SUPPLEMENT Materials and methods Cell culture and reagents Cell media and reagents were from Invitrogen unless otherwise indicated. Antibiotics and Tet-certified serum were from Clontech. In experiments
SUPPLEMENT Materials and methods Cell culture and reagents Cell media and reagents were from Invitrogen unless otherwise indicated. Antibiotics and Tet-certified serum were from Clontech. In experiments
Mitochondrial DNA Isolation Kit
 Mitochondrial DNA Isolation Kit Catalog Number KA0895 50 assays Version: 01 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 General Information... 4 Materials
Mitochondrial DNA Isolation Kit Catalog Number KA0895 50 assays Version: 01 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 General Information... 4 Materials
Supplementary information. MARCH8 inhibits HIV-1 infection by reducing virion incorporation of envelope glycoproteins
 Supplementary information inhibits HIV-1 infection by reducing virion incorporation of envelope glycoproteins Takuya Tada, Yanzhao Zhang, Takayoshi Koyama, Minoru Tobiume, Yasuko Tsunetsugu-Yokota, Shoji
Supplementary information inhibits HIV-1 infection by reducing virion incorporation of envelope glycoproteins Takuya Tada, Yanzhao Zhang, Takayoshi Koyama, Minoru Tobiume, Yasuko Tsunetsugu-Yokota, Shoji
DEVELOPMENTAL VALIDATION OF SPERM HY-LITER EXPRESS Jennifer Old, Chris Martersteck, Anna Kalinina, Independent Forensics, Lombard IL
 DEVELOPMENTAL VALIDATION OF SPERM HY-LITER EXPRESS Jennifer Old, Chris Martersteck, Anna Kalinina, Independent Forensics, Lombard IL Background and Introduction SPERM HY-LITER is the first immunofluorescent
DEVELOPMENTAL VALIDATION OF SPERM HY-LITER EXPRESS Jennifer Old, Chris Martersteck, Anna Kalinina, Independent Forensics, Lombard IL Background and Introduction SPERM HY-LITER is the first immunofluorescent
Figure S1. PMVs from THP-1 cells expose phosphatidylserine and carry actin. A) Flow
 Flow") SUPPLEMENTARY DATA Supplementary Figure Legends Figure S1. PMVs from THP-1 cells expose phosphatidylserine and carry actin. A) Flow cytometry analysis of PMVs labelled with annexin-v-pe (Guava technologies)
SUPPLEMENTARY DATA Supplementary Figure Legends Figure S1. PMVs from THP-1 cells expose phosphatidylserine and carry actin. A) Flow cytometry analysis of PMVs labelled with annexin-v-pe (Guava technologies)
Abhd2 regulates alveolar type Ⅱ apoptosis and airway smooth muscle remodeling: a key target of COPD research
 Abhd2 regulates alveolar type Ⅱ apoptosis and airway smooth muscle remodeling: a key target of COPD research Shoude Jin Harbin Medical University, China Background COPD ------ a silent killer Insidious,
Abhd2 regulates alveolar type Ⅱ apoptosis and airway smooth muscle remodeling: a key target of COPD research Shoude Jin Harbin Medical University, China Background COPD ------ a silent killer Insidious,
(a) Significant biological processes (upper panel) and disease biomarkers (lower panel)
 Significant biological processes (upper panel) and disease biomarkers (lower panel)") Supplementary Figure 1. Functional enrichment analyses of secretomic proteins. (a) Significant biological processes (upper panel) and disease biomarkers (lower panel) 2 involved by hrab37-mediated secretory
Supplementary Figure 1. Functional enrichment analyses of secretomic proteins. (a) Significant biological processes (upper panel) and disease biomarkers (lower panel) 2 involved by hrab37-mediated secretory
Neurotrophic factor GDNF and camp suppress glucocorticoid-inducible PNMT expression in a mouse pheochromocytoma model.
 161 Neurotrophic factor GDNF and camp suppress glucocorticoid-inducible PNMT expression in a mouse pheochromocytoma model. Marian J. Evinger a, James F. Powers b and Arthur S. Tischler b a. Department
161 Neurotrophic factor GDNF and camp suppress glucocorticoid-inducible PNMT expression in a mouse pheochromocytoma model. Marian J. Evinger a, James F. Powers b and Arthur S. Tischler b a. Department
In vitro bactericidal assay Fig. S8 Gentamicin protection assay Phagocytosis assay
 In vitro bactericidal assay Mouse bone marrow was isolated from the femur and the tibia. Cells were suspended in phosphate buffered saline containing.5% BSA and 2 mm EDTA and filtered through a cell strainer.
In vitro bactericidal assay Mouse bone marrow was isolated from the femur and the tibia. Cells were suspended in phosphate buffered saline containing.5% BSA and 2 mm EDTA and filtered through a cell strainer.
MagCapture Exosome Isolation Kit PS Q&A
 MagCapture Exosome Isolation Kit PS Q&A Specifications and performance P.1 Comparison of the conventional method P.2 Operation methods and composition P.4 Amount of starting sample P.5 Analysis after exosomes
MagCapture Exosome Isolation Kit PS Q&A Specifications and performance P.1 Comparison of the conventional method P.2 Operation methods and composition P.4 Amount of starting sample P.5 Analysis after exosomes
Optimizing Intracellular Flow Cytometry:
 Optimizing Intracellular Flow Cytometry: Simultaneous Detection of Cytokines and Transcription Factors An encore presentation by Jurg Rohrer, PhD, BD Biosciences 10.26.10 Outline Introduction Cytokines
Optimizing Intracellular Flow Cytometry: Simultaneous Detection of Cytokines and Transcription Factors An encore presentation by Jurg Rohrer, PhD, BD Biosciences 10.26.10 Outline Introduction Cytokines
SUPPLEMENTARY INFORMATION
 1. Supplementary Figures and Legends Supplementary Fig. 1. S1P-mediated transcriptional regulation of integrins expressed in OP/monocytoid cells. Real-time quantitative PCR analyses of mrna for two integrins,
1. Supplementary Figures and Legends Supplementary Fig. 1. S1P-mediated transcriptional regulation of integrins expressed in OP/monocytoid cells. Real-time quantitative PCR analyses of mrna for two integrins,
-Cyclodextrin-threaded Biocleavable Polyrotaxanes Ameliorate Impaired Autophagic Flux in Niemann-Pick Type C Disease
 -Cyclodextrin-threaded Biocleavable Polyrotaxanes Ameliorate Impaired Autophagic Flux in Niemann-Pick Type C Disease Atsushi Tamura and Nobuhiko Yui Department of Organic Biomaterials, Institute of Biomaterials
-Cyclodextrin-threaded Biocleavable Polyrotaxanes Ameliorate Impaired Autophagic Flux in Niemann-Pick Type C Disease Atsushi Tamura and Nobuhiko Yui Department of Organic Biomaterials, Institute of Biomaterials
Taylor Yohe. Project Advisor: Dr. Martha A. Belury. Department of Human Nutrition at the Ohio State University
 Atherosclerosis Development and the Inflammatory Response of Hepatocytes to Sesame Oil Supplementation Taylor Yohe Project Advisor: Dr. Martha A. Belury Department of Human Nutrition at the Ohio State
Atherosclerosis Development and the Inflammatory Response of Hepatocytes to Sesame Oil Supplementation Taylor Yohe Project Advisor: Dr. Martha A. Belury Department of Human Nutrition at the Ohio State
EXOTESTTM. ELISA assay for exosome capture, quantification and characterization from cell culture supernatants and biological fluids
 DATA SHEET EXOTESTTM ELISA assay for exosome capture, quantification and characterization from cell culture supernatants and biological fluids INTRODUCTION Exosomes are small endosome-derived lipid nanoparticles
DATA SHEET EXOTESTTM ELISA assay for exosome capture, quantification and characterization from cell culture supernatants and biological fluids INTRODUCTION Exosomes are small endosome-derived lipid nanoparticles