# This work was supported by grant DK27626 from the National Institutes of. *To whom correspondence should be addressed: Dept. of Molecular & Cellular
|
|
|
- Jonah Morton
- 6 years ago
- Views:
Transcription
1 JBC Papers in Press. Published on May 22, 2002 as Manuscript M :33 PM 5/14/2002 Construction and Characterization of a Monomeric Insulin Receptor# James Jianping Wu and Guido Guidotti* Department of Molecular and Cellular Biology, Harvard University 7 Divinity Avenue Cambridge, MA Running title: monomeric insulin receptor Keywords: monomer/dimer/insulin receptor/autophosphorylation # This work was supported by grant DK27626 from the National Institutes of Health to GG. *To whom correspondence should be addressed: Dept. of Molecular & Cellular Biology, Harvard University, 7 Divinity Ave. Cambridge, MA Fax: ; Tel: ; guidotti@fas.harvard.edu Copyright 2002 by The American Society for Biochemistry and Molecular Biology, Inc.
2 Summary A mutant insulin receptor was constructed by replacing cysteine residues Cys524, Cys682, Cys683, Cys685 with serine. The mutant was expressed in COS7 and CHO cells, did not form covalently linked dimers and was present at the cell surface. There was half as much insulin binding activity at the cell surface in cells expressing the mutant compared to that in cells expressing the wild type receptor. The intracellular processing of the mutant receptor was affected as its β-subunit migrated more slowly than that of the wild type receptor on SDS PAGE. The mutant was capable of insulin dependent autophosphorylation and phosphorylation of insulin receptor substrate-1 in vivo and could be crosslinked into receptor dimers when membrane bound. The amount of insulin dependent autophosphorylation of the mutant receptor was half of that of the wild type receptor. However, after solubilization the monomeric insulin receptor had minimal autophosphorylation activity and, unlike the naturally occurring monomeric receptor tyrosine kinases, the solubilized monomeric insulin receptor did not dimerize in response to insulin binding as determined by sucrose density gradient centrifugation. 2
3 Introduction The insulin receptor (IR) differs from most other receptor tyrosine kinases because it is present in the plasma membrane as a disulfide linked dimer of two αβ monomers (1). The β chains are linked to the α chains by class II disulfide bonds, which are resistant to reduction; Cys 647 is the α chain residue involved in the class II disulfides (2). The two α chains are extracellular and are linked by class I disulfide bonds, which are easily reduced by dithiothreitol (3). At least two α chain cysteine residues are involved in the class I disulfide bonds (4,5), one of which is cysteine 524 (6-8). In a previous communication, we reported that the cysteine residues involved in the class I disulfides between the α chains were Cys524 and Cys682 (9). While the involvement of Cys524 in the class I disulfides had been suggested by other investigators (6-8), evidence that at least one of the cysteine residues of the triplet at positions 682, 683, and 685 formed an additional disulfide bond was presented subsequently by Sparrow et al. (10). Further support was provided by the report that an α-subunit fragment, in which amino acids and had been deleted, is a monomer and binds insulin (11). A curious finding of the previous work was that when cells expressing the IR C524S,C682S mutant were incubated with insulin before solubilization of the receptors, most of the receptors dimerized. We became concerned that the dimerization might be caused by formation of disulfide bonds between Cys682 and Cys685. To investigate this possibility, we constructed another insulin receptor by replacing Cys 683 and Cys685, in addition to Cys 524 and Cys 682, with serine. The new receptors did not form covalent dimers, were expressed on the cell surface and underwent autophosphorylation in response to insulin while membrane 3
4 bound. However, in the monomeric state the insulin receptors had decreased insulin dependent kinase activity. Experimental Procedures Materials Dulbecco s Modified Eagle s Medium (DMEM) and Nutrient Mixture F-12 (Ham s F12), Lipofectamine, Penicillin-Streptomycin-Glutamine (PSG), Fetal Bovine Serum (FBS) were purchased from Life Technologies, Inc. (Rockville MD). Vector plasmid pcdna3.1his/v5 and V5 antibody were purchased from Invitrogen (Carlsbad CA). Insulin receptor antibodies Ab-1, Ab-4 were purchased from Oncogene Research Products (Cambridge MA). Antibodies against insulin receptor β chain (29B4), insulin receptor α chain (N-20), phosphotyrosine (PY-99) and Protein G-plus were obtained from Santa Cruz (Santa Cruz CA). Anti-insulin receptor substrate-1 (IRS-1) antibody was obtained from Upstate Biotechnology, Inc (Lake Placid NY). [γ- 32 P]ATP and 125 I-insulin were purchased from NEN Life Science Products, Inc. (Boston MA). SuperSignal West Pico Chemiluminescent Substrate and GelCode Blue Stain Reagent were from Pierce (Rockford IL). Ethlyene Glycolbis(Succinic Acid N-hydroxy-succinimide Ester) (EGS) was purchased from Sigma Chemical Co. (St. Louis M0). Cell culture and transient transfection COS7 cells were maintained in DMEM with 10% FBS, 100 unit/ml penicillin, 100 µg/ml streptomycin, and 292 µg/ml of glutamine at 37 o C in 5% CO 2. CHO cells were maintained in F-12 medium with 10% FBS at 37 o C in 5% CO 2. To transiently express the insulin receptors, subconfluent cells were transfected with plasmid DNA and lipofectamine according to the manufacturer s instructions. The ratio of DNA to lipofectamine was 1:3. 4
5 Construction of expression plasmids The BamH1/XbaI fragment of pece-c524, 682,685S, which contains serine mutations at residues 524, 682, 685 (9), was inserted in pbluescript KS(+) to make plasmid pjw5. In order to introduce a BstBI site at the C-terminus of the β-subunit, a mutagenesis PCR reaction was carried out with T3 primer and primer R1 (CGGTAGGCACTGTTcGaAAGGATTGGAC), and pjw5 as template. The BstBI site is underlined, and the small letters represent the changes from the original bases in the primer R1. In this way, the last amino acid of the human insulin receptor was changed from serine to phenylalanine. The stop codon was changed to glutamic acid. The PCR product was cloned into the BamHI/BstBI site of Litmus28 (New England BioLabs, Inc. Beverly, Massachusetts) to make pjw6. The 3.5-kb HindIII/PmlI fragment of pet3(12) (from William J. Rutter, University of California, San Francisco CA) was cloned into the HindIII/PmlI site of pjw6 to make pjw17, which contains the whole human IR gene. The HindIII/BstBI fragment from pjw17 was inserted into pcdna3.1-v5/his (version A) to make pjw64, which encodes the human insulin receptor with a V5/His epitope tag. Mutations of the C-terminus of the insulin receptor (Ser to Phe, Stop to Glu) and the introduction of the V5/His epitope tag did not affect its basic biological functions (see results). The Ser524 mutation was imported from pece- C524, 682, 685S by replacing the HindIII/BamHI region of pjw64 with the corresponding fragment of pece-c524,682,685s. To introduce mutations in the region of Cys682 to Cys685, the following primers were used : S682 (ATGAGGATTCGGCCGGCGAATcCTGCTCCTGTCCAA), S682, 3, 5 (ATGAGGATTCGGCCGGCGAAaGCaGCTCCTcTCCAA), S683,5 (ATGAGGATTCGGCCGGCGAATGCaGCTCCaGTCCAA), S682,3 (ATGAGGATTCGGCCGGCGAAaGCaGCTCCTGTCCAA). 5
6 The mutated bases are shown in lower case. All the modified regions of the IR cdna were sequenced to confirm the mutations and verify that no other mutation occurred. SDS-PAGE and immunoblotting The samples in Laemmli sample buffer (reducing or nonreducing depending upon the experiments) were heated at 95 o C for 5 minutes before subjecting to SDS-PAGE. The separated proteins were electrophoretically transferred to nitrocellulose (300 ma for 1 hour 15 minutes). The nitrocellulose was blocked with 5% nonfat milk prior to incubation with primary antibody. The primary and secondary antibodies were dissolved in TBST (25 mm Tris, 0.02% KCl, 0.8% NaCl, 0.1%Tween20, ph8.0) with 1 % nonfat milk. Immune complexes were detected by enhanced chemiluminescence. Insulin binding assay CHO cells (70-80% confluent) in 100 mm plates were transfected with 5 µg of plasmid containing the cdna for different forms of the insulin receptor or LacZ, and 48 hours after transfection the cells were incubated in the medium without FBS for 4 hours. The cells were washed twice with cold phosphate-buffered saline. 125 I-insulin (50 pm) was incubated with the cells for 12 hours at 4 o C in the binding buffer containing 100 mm HEPES ph7.4, 120 mm NaCl, 5 mm KCl, 1.2 mm MgSO 4, 10 mm glucose, 1% bovine serum albumin (BSA). After incubation, the cells were washed three times with ice-cold phosphate-buffered saline, solubilized in 0.1% SDS, and counted in a gamma counter. 125 I-insulin binding to crude membranes was done by incubating 30 µg of crude membranes with 50 pm 125 I-insulin and increasing concentrations of porcine insulin (0 to 50 nm) in 50 mm HEPES ph 7.4 with 0.1% BSA for 12 hours at 4 o C. The membrane proteins 6
7 were collected by centrifugation for 30 minutes at 4 o C. No specific binding could be detected from the membranes of COS7 cells transfected with cdna3.1-lacz. Sucrose Density Gradient Sedimentation This was performed as described by Aiyer (12) and Lu and Guidotti (9). Briefly, 80 µg of crude membranes from COS7 cells expressing dimeric or monomeric insulin receptors were solubilized with 1% Triton X-100; the solutions were diluted to 0.1% Triton X-100 and centrifuged at 40,000 rpm for 30 min in a Beckman Ti70.1 rotor. The supernatants were collected and incubated with or without 20 nm insulin at 4 o C overnight. The solutions were then layered onto 4.6 ml of linear 5-20% (w/v) sucrose gradients containing 0.1% Triton X- 100 and 20 nm insulin or no insulin. The samples were centrifuged for 6 hours at 48,000 rpm at 4 o C in a Beckman SW50.1 rotor. Fractions of 200 µl were collected from the top of the tubes. Equivalent amounts of solubilized dimeric or monomeric insulin receptors were separated under identical conditions except without insulin in the gradient. To locate the insulin receptor, fractions were assayed for their ability to bind 125 I-insulin. Catalase and aldolase were loaded on the sucrose gradients with the samples. The location of the two enzymes was determined by SDS-PAGE, followed by staining with GelCode Blue Stain Reagent (Pierce, Rockford IL). Immunoprecipitation Cells were solubilized with lysis buffer (0.5% NP-40 in 50 mm Tris-HCl, ph7.5, 100 mm NaCl, 1 mm PMSF, 100 mm NaF, 0.2 mm sodium vanadate, 10 µg/ml leupeptin, 10 µg/ml aprotinin). After centrifugation at 13,000 rpm to remove cell debris, 800 µg of cell lysate protein were incubated with appropriate antibodies for 2-4 hours, followed by addition of µl of Protein G-agarose and incubation for 1-3 hours. The beads were collected by 7
8 centrifugation, washed with lysis buffer three times. The immunoprecipitated proteins were released from the bead by Laemmli buffer containing 100 mm DTT and analyzed by SDS PAGE followed by immunoblotting. Receptor autophosphorylation Cells, 48 hours after transfection, were incubated in a medium without serum for 2-4 hours (as indicated in the text), treated with 100 nm insulin in a medium containing 1% BSA for 5 min, washed with ice-cold PBS three times followed by processing for immunoblotting or immunoprecipitation. IRS-1 phosphorylation A plasmid containing the cdna for insulin receptor substrate-1 (IRS-1) (pcmvhis/irs-1, provided by Dr. M.F.White, Joslin Diabetes Center, Boston MA ) was cotransfected into CHO cell with the insulin receptor dimer or monomer plasmids. After 48 hours, the cells were serum starved, treated with insulin, and solubilized as described above for measurement of phosphorylation of the insulin receptor. The cell extracts were immunoprecipitated with anti-phosphotyrosine antibody and the precipitates examined with anti-irs1 antibody after SDS PAGE. Cross-linking of insulin receptors 60 µg of crude membranes from COS7 cells expressing dimeric or monomeric insulin receptor were incubated with 5 nm 125 I-insulin in 100 µl of HEPES buffer (140 mm NaCl, 5 mm KCl, 1.4 mm CaCl 2, 1 mm MgSO 4, 10 mm HEPES, ph7.6) and 1% (w/v) BSA overnight in the absence or presence of 1.5 µm of unlabeled insulin. The membranes were precipitated by centrifugation at 13,000 rpm for 10 min at 4 o C and the pellets were washed twice with ice cold HEPES buffer. The membranes were resuspended in the HEPES buffer 8
9 with 1 mm EGS and incubated on ice for 1 hour (13). The cross-linking reaction was stopped by removing EGS and washing the membranes three times by centrifugation. The pellets were resuspended in Laemmli buffer with 1% β-mercaptoethanol and subjected to SDS PAGE on 5% gels. The radioactivity on the gel was analyzed with a BioRad imager. 9
10 Results Construction of a monomeric insulin receptor Previously, we identified Cys524 and Cys682 as the residues involved in the class I disulfide bonds between the α chains (9). The IR C524S,C682S mutant, however, was able to dimerize in the presence of insulin. To overcome this problem, we constructed IR mutants in which Cys524 and Cys682 were both changed to Asp or Arg to prevent monomer monomer interactions by electrostatic repulsion. We found, however, that the IR C524,682D and IR C524,682R mutants were a mixture of monomers and dimers when examined by nonreducing SDS PAGE (data not show). This result raised a concern that Cys 524 and Cys682 might not be the only cysteine residues participating in class I disulfide bonds. In order to address this question, we made a variety of IR mutants involving Cys524, Cys682, Cys683, and Cys685. As the two mutants, Cys524Ser and Cys682,683,685Ser, are expressed as dimers (2), we focused our attention on the mutants Cys524,682,683Ser, Cys524,682,685Ser and Cys524,683,685Ser, and compared them to Cys524,682Ser. As can be seen in Figure 1A, all four mutants were present as dimers and monomers (lanes 2, 3, 4 and 5 respectively) when examined by nonreducing SDS PAGE. The result suggested that it may be necessary to mutate all four Cys residues at positions 524, 682, 683 and 685 in order to obtain monomeric insulin receptors. This supposition was shown to be correct; lane 1 of Figure 1A shows that in the mutant Cys524,682,683,685Ser there was only monomeric receptor. Notice that in all lanes the proreceptor, composed of the fused and unproteolyzed αβ chain, was visible. Figure 1B shows a more detailed analysis of the IR C524,682,683,685S mutant, compared to the wild type IR(IR WT ). Under nonreducing conditions, only receptor monomers were present 10
11 after SDS-PAGE (lane 2) whereas IR WT was dimeric (lanes 1 and 3). Under reducing conditions, only the β subunit and the proreceptor were visible (lanes 4-6); addition of the epitope tag to the C-terminal of the IR made the β subunit migrate more slowly than the native β subunit (lanes 4 compared to lane 6). In addition, the β subunit of the mutant IR migrated more slowly than that of the wild type (lane 5 compared to lane 4). The difference in mobility was also observed for the mutant β chain expressed in CHO cells (Fig. 4A). We conclude that Cys 524 and all three Cys residues at positions 682, 683, and 685 are involved in the class I disulfides. Monomeric IR is expressed on the cell surface In order to determine whether the IR C524,682,683,685S mutant is present at the cell surface, varying amounts of the plasmids containing the wild type and mutant IR cdnas were used to transfect CHO cells. After 48 hours, the cells were assayed for insulin binding and the results for the maximum binding are shown in Table1. The cells expressing IR WT had approximately twice the amount of surface IR compared to those expressing IR C524,682,683,685S. Two possibilities for the lower amount of insulin binding to the cells expressing mutant IR are impairment in transport of the receptor to the cell surface and lower insulin affinity of the receptor. In order to study the latter question, binding of 125 I-insulin to crude membranes with the wild type and mutant receptors was analyzed by the method of Scatchard (Fig. 2). For wild type and mutant IRs, the dissociation constants were 5.6X10-9 M and 5.1X10-9 M, respectively, and the maximal insulin binding were 1.58X10-11 mol/µg membrane and 1.12X10-11 mol/µg membrane, respectively. Interestingly, the plots for both receptors were curvilinear. 11
12 We conclude that the IR C524,682,683,685S mutant has a normal affinity for insulin but is defective in its ability to be delivered to the plasma membrane. Insulin binding is not associated with dimerization of IR C524,682,683,685S monomers There is considerable evidence that monomeric receptor tyrosine kinases dimerize in response to ligand binding (for reviews see 14,15). Since the IR C524,682,683,685S mutant does not form covalent dimers, we determined the size of the receptor in the absence and the presence of insulin by its sedimentation behavior in a sucrose density gradient (9, 12). Crude membranes from COS7 cell expressing dimeric or monomeric insulin receptors were incubated with or without 20 nm insulin overnight at 4 o C after solubilization, followed by centrifugation at 48,000 rpm for 6 hours in a linear 5 20% (w/v) sucrose gradient containing 0.1% Triton X-100 and 20 nm or no insulin. Fractions were collected and assayed for insulin binding to locate the insulin receptor. The results (Fig. 3) clearly show that the IR WT traveled at the position of the receptor dimer, whereas the mutant had the size of a receptor monomer. Insulin binding caused a slight increase in the sedimentation rate of both IR WT and IR C524,682,683,685S mutant, indicative of an insulin dependent change in shape of the receptors, consistent with results of Florke et al. (16). Insulin binding did not induce the dimerization of monomeric insulin receptors under these conditions. Autophosphorylation of the insulin receptor To determine whether the monomeric insulin receptor autophosphorylates in response to insulin, we treated intact CHO cells expressing the wild type and the mutant IR with insulin and measured the amount of phosphate bound to the β subunit. The level of expression of receptor dimers and monomers was approximately the same (Fig. 4A). To measure receptor phosphorylation, intact cells were incubated with insulin; then, the cell extracts were either 12
13 directly separated by SDS-PAGE followed by immunoblotting with anti-phosphotyrosine antibodies (Fig. 4B) or first immunoprecipitated with antiphosphotyrosine antibody and the precipitates were separated by SDS PAGE and immunoblotted with anti-v5 antibody, which recognizes the β chain (Fig. 4C). The results clearly show that the receptor monomer had insulin dependent phosphorylation, and that the total amount of receptor dimer phosphorylation was twice that of the receptor monomer. This result is again compatible with the surface expression of IR WT and IR C524,682,683,685S (Table1). It is also apparent that the amount of phosphorylation of the receptor monomer in the absence of insulin was significant. This insulin-independent activation of the insulin receptor monomer could be a consequence of the monomeric state or of a structural change of the receptor brought about by the mutations. We tried to answer this question by comparison of insulin dependent autophosphorylation of the receptor dimers and monomers with that of receptors lacking only one set of disulfide bonds. Insulin receptor dimers are held together by two sets of disulfide bonds, one at position 524 and the other set at positions 682, 683 and 684. Hence, we constructed the mutants IR C524S and IR C82,683,685S, and measured the amount of phosphate incorporated in intact cells by immunoprecipitation with antiphosphotyrosine antibody followed by quantitation with anti-v5 antibody. The results (Fig. 5A) indicate that all the mutants had considerable phosphorylation of the β chain in the absence of insulin, that there was insulin dependent phosphorylation in all cases, and that the stimulation by insulin was greatest for the IR WT, followed by IR C524S, IR C524,682,683,685S and IR C682,683,685S in that order. It is also important to notice that the total amount of receptor present in the cell (Fig 5B) was not proportional to the amount of insulin binding on the cell surface (Table 1), indicating that some of the mutants are impaired in their delivery to the plasma membrane. The conclusion is 13
14 that the cysteine to serine mutations alter the structure of the insulin receptor so that there is some insulin-independent activation of the receptor. Kinase activity of the insulin receptor To determine whether the insulin receptor monomer has exogenous kinase activity, we asked whether it is capable of phosphorylating IRS-1, a substrate of the insulin receptor kinase (17). A plasmid containing the cdna for IRS-1 (17) was cotransfected with plasmids containing the cdna for IR WT or IR C524,682,683,685S into CHO cells. After 48 hours, the insulin dependent phosphorylation of IRS-1 was assayed. Figure 6 shows that IRS-1 underwent insulin dependent phosphorylation by both insulin receptor dimer and monomer. The level of phosphorylation by the receptor dimer was approximately twice that by the receptor monomer, as was seen for receptor autophosphorylation. In addition, there was phosphorylation of IRS-1 even in the absence of insulin, confirming the results that some of the expressed insulin receptor is present in the activated state even in the absence of insulin. Autophosphorylation of the solubilized insulin receptor. In order to determine whether the insulin receptor monomer has kinase activity when it is in the monomeric state, the membranes were solubilized with Triton X-100. Under these conditions, the insulin receptor monomers are monomeric (Fig. 3). As is shown in Figure 7 for receptors expressed in CHO cells, the extent of autophosphorylation of the insulin receptor monomer in the monomeric state was greatly reduced compared to the level obtained when the insulin receptor monomer is in the membrane. The amount of autophosphorylation was approximately one-fourth to one-sixth of that of the insulin receptor dimer. The bottom panel of Figure 7 shows that the amounts of β chain and proreceptor in each of the samples is equivalent. This result suggests that insulin receptor monomers in the monomeric state have reduced kinase activity compared to that of the native insulin receptor dimers, as well as 14
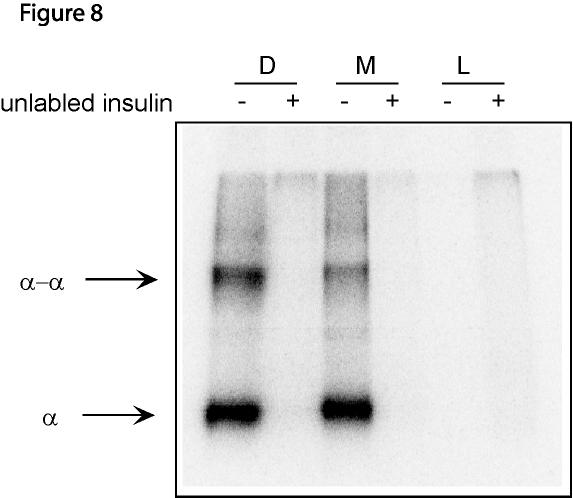
15 compared to that of the insulin receptor monomers that are in the membrane. Presumably, membrane bound insulin receptor monomers are present at a sufficiently high concentration to form noncovalent dimers. To test this hypothesis, we determined whether membrane bound receptor monomers could be crossliked to dimers in the presence of insulin. The membranes from cells expressing dimeric or monomeric insulin receptors were incubated with 125 I-insulin overnight in the absence or presence of excess unlabeled insulin. The membranes were then exposed to EGS to bring about insulin-α chain and α chain-α chain crosslinking (13). The membranes were then subjected to SDS PAGE under reducing conditions to obtain α chain monomers and dimers, and the gel analyzed with a BioRad imager. As can be seen in Figure 8, α-chain monomers and dimers bound to 125 I-insulin (lanes 1 and lane3), are present in both receptor dimer and monomer lanes. A greater percentage of the α chain can be covalently dimerized in the receptor dimer than in the receptor monomer (62% versus 29%), as might be expected since in the dimer the α chains are kept close to one another by the class I disulfide bonds. Nevertheless, even the receptor monomers are capable of forming noncovalent dimers at the concentrations present in the membranes during the overexpression protocol. We conclude that the kinase activity of the receptor monomer in the monomeric state is substantially lower than that of the receptor dimer. Discussion The results shown in Figure1 clearly indicate that only the insulin receptor in which all four Cys residues 524, 682, 683 and 685 are mutated to Ser is present exclusively as a monomer. All the other mutants with only three mutations of these residues are present as a 15
16 mixture of dimers and monomers, as is the IR C524,682S mutant previously described. We conclude that, in addition to Cys524, all three of the vicinal Cys residues in the 682 to 685 region are involved in the class I disulfides. NEM labeling of receptor monomers produced by mild reduction of native insulin receptor with DTT indicated that at least two cysteine residues are involved in class I disulfides (4,5). One of them was identified as Cys524 (6), while the other residue(s) could not be identified by labeling but was (were) localized to the triplet at positions 682, 683, 685 (10). Since it is apparent now that only mutation of all four Cys residues produces receptor monomers, it remains unclear why the labeling results were different. Possibly the vicinity of the cysteine residues at positions 682, 683 and 685 allowed intrachain disulfide bond formation so that only one of the three remained free to react with NEM. It is also not apparent how receptor monomers are formed in the partial mutants; possibly, the disulfide bonds between Cys 682, 683 and 685 are not between homologous residues. One clear result is the decrease in the surface expression of the IR C524,682,683,685S mutant compared to IR WT (Table 1). Two possible explanations are that the receptor monomer is hindered in its passage from the endoplasmic reticulum to the plasma membrane by the quality control system and that the receptor is toxic to the cell. The latter possibility is supported in part by the observation that even in the absence of insulin there is partial activation of the receptor monomer (Figs. 4-7). Since it is evident that the IR C524,682,683,685S mutant is a monomer when examined by SDS-PAGE (Fig. 1A) and by velocity sedimentation after solubilization by detergent (Fig. 3), the curvilinear Scatchard plot (Fig. 2) of the mutant, resembling that of the IR WT, presents a problem. If the IR C524,682,683,685S mutant is monomeric in the membrane, the two binding 16
17 components represent some heterogeneity. If instead the receptor monomer is dimeric in the membrane, the curvilinear plot may represent the same negative cooperativity of the native dimeric receptor (18). Negative cooperativity is not easily understood in light of the structural evidence that only one insulin molecule is bound by an insulin receptor dimer (19). The importance of dimerization for the activation of monomeric receptor tyrosine kinases (for example, the EGF receptor) has been extensively described (14,15). Strangely, the insulin receptor exists as a dimer in which the monomers are covalently linked by disulfide bonds. One possibility for this arrangement is that insulin receptor monomers are incapable of dimerization even in the presence of insulin. Another possibility is that the monomermonomer interactions are sufficiently weak so that at the concentration of native insulin receptor in the membrane, only a small fraction of the receptor monomers dimerize. The second possibility is supported by the evidence that the solubilized IR C524,682,683,685S mutant is defective in insulin dependent autophosphorylation (Fig. 7) when it is monomeric (Fig. 3), whereas it is capable of insulin dependent autophosphorylation and exogenous kinase activity on IRS-1 in cells (Figs. 4-7) where it is partially dimeric (Fig. 8). We conclude that insulin receptor monomers are incapable of autophosphorylation in the monomeric state; however, noncovalent dimerization and autophosphorylation can take place in the plasma membrane when the concentration of receptor monomers is sufficiently high. It seems apparent now that the results in our previous paper on the dimerization of the IR C524,682S mutant after incubation with insulin (9) may have been obtained by redistribution of the remaining disulfide bonds to produce disulfide linked dimers. This is another reason that 17
18 we believe that all three cysteine residues at positions 682, 683, and 685 are involved in the class I disulfide bonds. The conclusion that the insulin receptor monomer in the monomeric state has negligible kinase activity supports considerable work, all done with dithiothreitol reduced insulin receptors, by the groups of Pilch and Pessin (20-25). The Pilch group showed that receptor monomers could be separated from receptor dimers by sedimentation in a density gradient and that the monomers had reduced autophosphorylation activity compared to the native dimers (20). They further demonstrated that insulin receptor monomers could undergo autophosphorylation at a receptor concentration sufficient to allow dimerization (21). Although the dimerization was not shown, its involvement was suggested by studies on the concentration dependence of insulin receptor autophosphorylation (22). On the other hand, the group of Pessin presented studies on insulin receptor monomers that were interpreted as showing that the monomers only had autophosphorylation activity if they formed covalently associated (23, 24) or noncovalently associated receptor dimers (25). These studies were not straightforward, however, because the putative size of the insulin receptors was determined by gel filtration which does not separate monomers and dimers. In any event, the conclusion reached by both groups was that insulin receptor monomers are not as competent to autophosphorylate as are receptor dimers. The conclusions of the earlier work by Pilch and Pessin and of that presented here, that insulin receptor dimers are required for insulin dependent receptor autophosphorylation, are supported by the finding that even the soluble insulin receptor kinase domain is activated by dimerization (26). 18
19 Acknowledgements We thank Dr. Morris White (Joslin Diabetes Center, Boston MA) for the pcmvhis/irs-1 plasmid, Dr William J. Rutter (University of California, San Francisco CA) for pet3 containing the wild type human insulin receptor cdna, and members of the laboratory, especially Alison Grinthal, for discussions. References 1. White, M.F. and Kahn, C.R. (1994) J Biol Chem 269, Cheatham, B., and Kahn, C. R. (1992) J Biol Chem 267(10), Massague, J., and Czech, M. P. (1982) J Biol Chem 257, Finn, F. M., Ridge, K. D., and Hofmann, K. (1990) Proc Natl Acad Sci U S A 87, Chiacchia, K. B. (1991) Biochem Biophys Res Commun 176, Schaffer, L., and Ljungqvist, L. (1992) Biochem Biophys Res Commun 189, Bilan, P. J., and Yip, C. C. (1994) Biochem Biophys Res Commun 205, Macaulay, S.L., Polites, M., Hewish, D.R., and Ward, C.W. (1994) Biochem. J. 303, Lu, K., and Guidotti, G. (1996) Mol Biol Cell 7, Sparrow, L. G., McKern, N. M., Gorman, J. J., Strike, P. M., Robinson, C. P., Bentley, J. D., and Ward, C. W. (1997) J Biol Chem 272, Kristensen, C., Wiberg, F. C., Schaffer, L., and Andersen, A. S. (1998) J Biol Chem 273, Aiyer, R. A. (1983) J Biol Chem 258, Chiacchia, K.B. (1988) Biochemistry 27,
20 14. Schlessinger, J. (1988) Trends Biochem. Sci. 13, Ullrich, A., and Schlessinger, J. (1990) Cell 61, Florke, R.R., Klein, H. W., and Reinauer, H. (1990) Eur. J. Biochem 191, Sun, X. J., Miralpeix, M., Myers, M. G., Jr., Glasheen, E. M., Backer, J. M., Kahn, C. R., and White, M. F. (1992) J Biol Chem 267(31), Levitzki, A. (1981) Nature 289, Luo, R. Z., Beniac, D. R., Fernandes, A., Yip, C. C., and Ottensmeyer, F. P. (1999) Science 285, Boni-Schnetzler, M., Rubin, J.B., and Pilch P.F. (1986) J. Biol. Chem. 261, Boni-Schnetzler, M., Kaligian, A., DelVecchio, R., and Pilch, P.F. (1988) J. Biol. Chem. 263, Shoelson, S.E., Boni-Schnetler, M., Pilch, P.F., and Kahn, C.R. (1991) Biochemistry 30, Sweet, L.J., Morrison, B.D., Wilden, P.A., and Pessin, J.E. (1987)J Biol. Chem. 262, Morrison, B.D., Swanson, M.L., Sweet, L.J., and Pessin, J.E. (1988) J. Biol. Chem. 263, Wilden, P.A., Morrison, B.D., and Pessin, J.E. (1989) Biochemistry 28, Baer, K., Al-Hasani, H., Parvaresch, S., Corona, T., Rufer, A., Nolle, V., Bergschneider, E., and Klein, H.W. (2001) Biochemistry 40, Figure Legends Figure 1. Immunoblots of mutant insulin receptors. (A) COS7 cells were transfected with plasmids containing the cdna for the insulin receptors; after 48 hours crude membranes 20
21 were solubilized with Laemmli buffer containing 50 mm NEM followed by SDS-PAGE (3% stacking gel and 5% separating gel) and immunoblotting with anti-v5 antibody. IRC524, 682, 683, 685S (lane1); IR C524, 682, 683S (lane2); IR C524, 682, 685S (lane3); IR C524, 683, 685S (lane4); IR C524,682 S(lane 5). (B) Crude membranes from COS7 cells transfected with pjw64 (IR WT with V5-epitope), pjw65(ir C524, 682, 683, 685S with V5-epitope) and pet3(irc524, 682, 683, 685S without V5-epitope) were subjected to SDS-PAGE (4-15% gradient gel) under non-reducing or reducing conditions, and immunoblotted with anti-insulin receptor antibody Ab-4. Lanes 1, 4: IR WT with V5-epitope; 2, 5: IR C524, 682, 683, 685S with V5-epitope; 3, 6: IR WT without V5- epitope. Figure 2. Scatchard analyses of insulin binding to the dimeric and monmeric insulin receptors. The crude membranes from the COS7 cell expressing dimeric or monomeric insulin receptors were incubated with 50 pm of 125 I insulin and increasing concentrations of cold insulin at 4 o C overnight. The membranes were separated from the solution by centrifugation, washed, and then assayed for radioactivity with a gamma-counter. No significant binding could be detected to the crude membranes from the COS7 cells expressing LacZ (data not shown). The dissociation constants are 5.6X10-9 M and 5.1X10-9 M for wild type and mutant IRs, respectively, and the maximal amounts of insulin binding are 1.58X10-11 mol/µg membrane and 1.12X10-11 mol/µg membrane, respectively. Figure 3. Insulin binding does not cause dimerization of the monomeric insulin receptor. 80 µg of crude membranes from COS7 cell expressing dimeric or monomeric insulin receptors were solubilized with 1% TritonX-100; the solutions were diluted to 0.1% Triton X-100 with binding buffer (see Experimental Procedures) and centrifuged at 40,000 rpm for 30 min in a 21
22 Beckman Ti70.1 rotor. The supernatants were incubated without or with 20 nm insulin at 4 o C overnight. The samples were layered onto 4.6 ml of linear 5-20% (w/v) sucrose gradients containing 0.1% Triton X-100 and 20 nm insulin or no insulin, and centrifuged at 48,000 rpm for 6 hours at 4 o C in a Beckman SW50.1 rotor. Fractions of 200 µl were collected from the top of the tube and the location of the insulin receptor in the fractions was determined by its ability to bind 125 I-insulin. Catalase and aldolase were used to calibrate the gradients and their positions were determined by SDS PAGE. Figure 4. Ability of insulin receptor dimers and monomers expressed in CHO cells to autophosphorylate. After 4 hours of serum starvation, CHO cell expressing dimeric or monomeric insulin receptors were incubated without and with 100 nm insulin for 5 minutes, solubilized with Laemmli buffer and then subjected to SDS-PAGE and immunoblotting with (A) anti-v5 antibody or (B) anti-phosphotyrosine antibody (PY-99). (C) After insulin treatment, CHO cells were solubilized with 0.5% NP-40 and immunoprecipitated with antiphosphotyrosine antibody (PY-99). The precipitates were separated by SDS-PAGE and immunoblotted with anti-v5 antibody. D: IR WT with V5-epitope; M: IR C524, 682, 683, 685S with V5-epitope; L: cdna3.1-v5/his-lacz control. The columns in each panel show the relative intensities of the bands compared with those of the dimeric insulin receptors in the presence of insulin. The panels show one of two independent experiments. Figure 5. Insulin dependent autophosphorylation of insulin receptor mutants of the class I disulfide cysteine residues. CHO cells transfected with plasmids containing the cdna for the wild type insulin receptor IR WT with V5-epitope (D), and the mutants IRC524, 682, 683, 685S with V5-epitope ( M), IR C682, 683, 685S with V5-epitope (IR C682,3,5S ) and IR C524S with V5-epitope (IR C524S ) were grown for 48 hours. The cells were then incubated in F-12 medium without serum for 2 hours, followed by treatment with or without 100 nm insulin for 5 minutes. 22
23 Insulin stimulation was stopped by washing with ice-cold PBS three times. (A) Cells were solubilized with 0.5%NP-40, immunoprecipitated with anti-phosphotyrosine antibody (PY99) and protein G-agarose. The precipitate was examined by SDS-PAGE and immunoblotting with anti-v5 antibody. (B) Cells were solubilized with Laemmli loading buffer and examined by SDS-PAGE and immunoblotting with anti-v5 antibody. The columns in each panel show the relative intensities of the bands compared with those of the dimeric insulin receptors in the presence of insulin. The panels show one of two independent experiments. Figure 6. Insulin dependent phosphorylation of IRS-1 in CHO cell expressing insulin receptor dimers and monomers. CHO cells were cotransfected with a plasmid containing the cdna for wild type or monomeric insulin receptors and a plasmid containing the cdna for IRS-1. After 48 hours, the cells were placed in F-12 medium without serum for 4 hours, and then treated with or without 100 nm insulin for 5 minutes. The cells were solubilized with 0.5% NP-40, followed by immunoprecipitation with PY-99 antibody. The precipitates were examined by SDS-PAGE and immunobloting with anti-irs1 antibody. D: IRS-1 cotransfected with IR WT with V5-epitope; M: IRS-1 co-transfected with IR C524, 682, 683, 685S with V5-epitope; L: IRS-1 co-transfected with cdna3.1-v5/his-lacz. The columns in each panel Figure 7. Effect of solubilization on insulin receptor autophosphorylation. CHO cells expressing wild type insulin receptors, monomeric insulin receptors or LacZ were serum starved for 3 hours. In one experiment, the intact cells were treated without or with 100 nm insulin for 5 minutes, then solubilized with 0.5% Triton X-100 in Buffer A (50 mm HEPES, ph7.4, 100 mm NaCl, 100 mm NaF, 0.2 mm sodium vanadate, 10 µg/ml leupeptin, 10 µg/ml aprotinin); the solutions were immunoprecipitated with anti-phosphotyrosine antibody (PY- show the relative intensities of the bands compared with those of the dimeric insulin receptors in the presence of insulin. The panels show one of three independent experiments. 23
24 99) and protein G-agarose. In a second experiment, the cells were first solubilized with 0.5% Triton X-100 in Buffer A and the extracts were treated with 100 nm insulin for 15 min. Then, to the cell extracts were added ATP, MgCl 2, and MnCl 2 to a final concentrations of 500 µm, 5 mm and 5 mm, respectively. After 20 minutes, the phosphorylation reaction was terminated with 10 mm EDTA, 100 mm NaF, 0.2 mm sodium vanadate and the phosphorylated proteins were immunoprecipitated with anti-phosphotyrosine antibody (PY-99) and protein G-agarose. Care was taken to solubilize equivalent numbers of cells in the same volume of Buffer A in the two experiments, so that equal amounts of insulin receptors were compared in the two experiments. The precipitates were examined by SDS-PAGE followed by immunoblotting with anti-v5-hrp antibody. (A). First six lanes: intact cells. Second six lanes: solubilized cell extracts. (B). The cell extracts, before immunoprecipitation with the PY-99 antibody, were examined by reducing SDS-PAGE and immunoprecipitation with anti-v5-hrp antibody, in order to determine the amount of insulin receptor in the extracts. D: IR WT with V5-epitope; M: IR C524, 682, 683, 685S with V5-epitope; L: cdna3.1-v5/his-lacz control. The columns in each panel show the relative intensities of the bands compared with those of the dimeric insulin receptors in the presence of insulin. The panels show one of two independent experiments. Figure 8. Crosslinking of the α chains of membrane bound insulin receptor dimers and monomers. Crude membrane from COS7 cells expressing dimeric insulin receptors, monomeric insulin receptor or LacZ were incubated with 125 I-insulin in the absence or presence of 1.5 µm unlabeled insulin overnight. Free insulin was removed by centrifugation and washing. The insulin receptors were crosslinked with 1 mm EGS for 1 hour at 0 o C. The membranes were washed and solubilized with Laemmli buffer containing 1% β- mercaptoethanol, and subjected to SDS PAGE on a 5% gel. The gel was analyzed with a 24
25 BioRad imager. D: IR WT with V5-epitope; M: IR C524, 682, 683, 685S with V5-epitope; L: cdna3.1-v5/his-lacz control. The experiment is one of two independent experiments. 25
26 Table 1. Number of insulin binding sites on intact CHO cells expressing wild type and mutant insulin receptors. Binding sites/cell IR WT 1.1 x 10 6 (3.7 x 10 4 ) IR C524,682,683,685S 4.2 x 10 5 (7.6 x 10 3 ) IR C524S 8.1 x 10 5 (1.6 x 10 4 ) IR C682,683,685S 1.3 x 10 6 (4.6 x 10 4 ) LacZ 3.9 x 10 4 ( 5.2 x10 2 ) Not transfected 3.9 x 10 4 (5.8 x 10 2 ) CHO cells were transfected with plasmids containing the cdnas for the wild type and the mutant insulin receptors indicated in the Table. After serum starvation for 4 hours, the cells were incubated with 50 pm 125 I-insulin overnight at 4 o C. The cells were washed with cold binding buffer three times and then solubilized with 0.1% SDS. The radioactivity of the solutions was determined with a gamma-counter. Nonspecific binding was determined by including 10 µm non-radioactive insulin in the incubation. The results are means of two independent experiments; the deviations are in parentheses. 26

27
28
29

30

31

32

33
34
35 J. Biol. Chem. Construction and characterization of a monomeric insulin receptor James Jianping Wu and Guido Guidotti published online May 22, 2002 Access the most updated version of this article at doi: /jbc.M Alerts: When this article is cited When a correction for this article is posted Click here to choose from all of JBC's alerts
Supplementary data Supplementary Figure 1 Supplementary Figure 2
 Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
SUPPLEMENTARY INFORMATION
 Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
RayBio KinaseSTAR TM Akt Activity Assay Kit
 Activity Assay Kit User Manual Version 1.0 March 13, 2015 RayBio KinaseSTAR TM Akt Activity Kit Protocol (Cat#: 68AT-Akt-S40) RayBiotech, Inc. We Provide You With Excellent Support And Service Tel:(Toll
Activity Assay Kit User Manual Version 1.0 March 13, 2015 RayBio KinaseSTAR TM Akt Activity Kit Protocol (Cat#: 68AT-Akt-S40) RayBiotech, Inc. We Provide You With Excellent Support And Service Tel:(Toll
Protocol for Gene Transfection & Western Blotting
 The schedule and the manual of basic techniques for cell culture Advanced Protocol for Gene Transfection & Western Blotting Schedule Day 1 26/07/2008 Transfection Day 3 28/07/2008 Cell lysis Immunoprecipitation
The schedule and the manual of basic techniques for cell culture Advanced Protocol for Gene Transfection & Western Blotting Schedule Day 1 26/07/2008 Transfection Day 3 28/07/2008 Cell lysis Immunoprecipitation
Supporting Online Material Material and Methods References Supplemental Figures S1, S2, and S3
 Supporting Online Material Material and Methods References Supplemental Figures S1, S2, and S3 Sarbassov et al. 1 Material and Methods Materials Reagents were obtained from the following sources: protein
Supporting Online Material Material and Methods References Supplemental Figures S1, S2, and S3 Sarbassov et al. 1 Material and Methods Materials Reagents were obtained from the following sources: protein
Supplementary Information
 Supplementary Information Supplementary Figure 1. CD4 + T cell activation and lack of apoptosis after crosslinking with anti-cd3 + anti-cd28 + anti-cd160. (a) Flow cytometry of anti-cd160 (5D.10A11) binding
Supplementary Information Supplementary Figure 1. CD4 + T cell activation and lack of apoptosis after crosslinking with anti-cd3 + anti-cd28 + anti-cd160. (a) Flow cytometry of anti-cd160 (5D.10A11) binding
HIV-1 Virus-like Particle Budding Assay Nathan H Vande Burgt, Luis J Cocka * and Paul Bates
 HIV-1 Virus-like Particle Budding Assay Nathan H Vande Burgt, Luis J Cocka * and Paul Bates Department of Microbiology, Perelman School of Medicine at the University of Pennsylvania, Philadelphia, USA
HIV-1 Virus-like Particle Budding Assay Nathan H Vande Burgt, Luis J Cocka * and Paul Bates Department of Microbiology, Perelman School of Medicine at the University of Pennsylvania, Philadelphia, USA
Luminescent platforms for monitoring changes in the solubility of amylin and huntingtin in living cells
 Electronic Supplementary Material (ESI) for Molecular BioSystems. This journal is The Royal Society of Chemistry 2016 Contents Supporting Information Luminescent platforms for monitoring changes in the
Electronic Supplementary Material (ESI) for Molecular BioSystems. This journal is The Royal Society of Chemistry 2016 Contents Supporting Information Luminescent platforms for monitoring changes in the
The Schedule and the Manual of Basic Techniques for Cell Culture
 The Schedule and the Manual of Basic Techniques for Cell Culture 1 Materials Calcium Phosphate Transfection Kit: Invitrogen Cat.No.K2780-01 Falcon tube (Cat No.35-2054:12 x 75 mm, 5 ml tube) Cell: 293
The Schedule and the Manual of Basic Techniques for Cell Culture 1 Materials Calcium Phosphate Transfection Kit: Invitrogen Cat.No.K2780-01 Falcon tube (Cat No.35-2054:12 x 75 mm, 5 ml tube) Cell: 293
supplementary information
 Figure S1 Nucleotide binding status of RagA mutants. Wild type and mutant forms of MycRagA was transfected into HEK293 cells and the transfected cells were labeled with 32 Pphosphate. MycRagA was immunoprecipitated
Figure S1 Nucleotide binding status of RagA mutants. Wild type and mutant forms of MycRagA was transfected into HEK293 cells and the transfected cells were labeled with 32 Pphosphate. MycRagA was immunoprecipitated
SUPPLEMENTARY MATERIAL
 SUPPLEMENTARY MATERIAL Purification and biochemical properties of SDS-stable low molecular weight alkaline serine protease from Citrullus Colocynthis Muhammad Bashir Khan, 1,3 Hidayatullah khan, 2 Muhammad
SUPPLEMENTARY MATERIAL Purification and biochemical properties of SDS-stable low molecular weight alkaline serine protease from Citrullus Colocynthis Muhammad Bashir Khan, 1,3 Hidayatullah khan, 2 Muhammad
Mapping the Ligand-binding Site on a GPCR Using Genetically-encoded Photocrosslinkers
 Mapping the Ligand-binding Site on a GPCR Using Genetically-encoded Photocrosslinkers Amy Grunbeck, Thomas Huber, Pallavi Sachdev, Thomas P. Sakmar Laboratory of Molecular Biology and Biochemistry, The
Mapping the Ligand-binding Site on a GPCR Using Genetically-encoded Photocrosslinkers Amy Grunbeck, Thomas Huber, Pallavi Sachdev, Thomas P. Sakmar Laboratory of Molecular Biology and Biochemistry, The
Western Immunoblotting Preparation of Samples:
 Western Immunoblotting Preparation of Samples: Total Protein Extraction from Culture Cells: Take off the medium Wash culture with 1 x PBS 1 ml hot Cell-lysis Solution into T75 flask Scrap out the cells
Western Immunoblotting Preparation of Samples: Total Protein Extraction from Culture Cells: Take off the medium Wash culture with 1 x PBS 1 ml hot Cell-lysis Solution into T75 flask Scrap out the cells
MEK1 Assay Kit 1 Catalog # Lot # 16875
 MEK1 Assay Kit 1 Kit Components Assay Dilution Buffer (ADB), Catalog # 20-108. Three vials, each containing 1.0ml of assay dilution buffer (20mM MOPS, ph 7.2, 25mM ß-glycerol phosphate, 5mM EGTA, 1mM sodium
MEK1 Assay Kit 1 Kit Components Assay Dilution Buffer (ADB), Catalog # 20-108. Three vials, each containing 1.0ml of assay dilution buffer (20mM MOPS, ph 7.2, 25mM ß-glycerol phosphate, 5mM EGTA, 1mM sodium
SUPPLEMENTARY MATERIAL
 SUPPLEMENTARY MATERIAL Table S1. Primers and fluorescent probes used for qrt-pcr analysis of relative expression levels of PPP family phosphatases. gene name forward primer, 5-3 probe, 5-3 reverse primer,
SUPPLEMENTARY MATERIAL Table S1. Primers and fluorescent probes used for qrt-pcr analysis of relative expression levels of PPP family phosphatases. gene name forward primer, 5-3 probe, 5-3 reverse primer,
TFEB-mediated increase in peripheral lysosomes regulates. Store Operated Calcium Entry
 TFEB-mediated increase in peripheral lysosomes regulates Store Operated Calcium Entry Luigi Sbano, Massimo Bonora, Saverio Marchi, Federica Baldassari, Diego L. Medina, Andrea Ballabio, Carlotta Giorgi
TFEB-mediated increase in peripheral lysosomes regulates Store Operated Calcium Entry Luigi Sbano, Massimo Bonora, Saverio Marchi, Federica Baldassari, Diego L. Medina, Andrea Ballabio, Carlotta Giorgi
p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO
 Supplementary Information p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO Yuri Shibata, Masaaki Oyama, Hiroko Kozuka-Hata, Xiao Han, Yuetsu Tanaka,
Supplementary Information p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO Yuri Shibata, Masaaki Oyama, Hiroko Kozuka-Hata, Xiao Han, Yuetsu Tanaka,
Nature Methods: doi: /nmeth Supplementary Figure 1
 Supplementary Figure 1 Subtiligase-catalyzed ligations with ubiquitin thioesters and 10-mer biotinylated peptides. (a) General scheme for ligations between ubiquitin thioesters and 10-mer, biotinylated
Supplementary Figure 1 Subtiligase-catalyzed ligations with ubiquitin thioesters and 10-mer biotinylated peptides. (a) General scheme for ligations between ubiquitin thioesters and 10-mer, biotinylated
Chromatin IP (Isw2) Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles.
 Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles.") Chromatin IP (Isw2) 7/01 Toshi last update: 06/15 Reagents Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles. 2.5 M glycine. TBS:
Chromatin IP (Isw2) 7/01 Toshi last update: 06/15 Reagents Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles. 2.5 M glycine. TBS:
Protein MultiColor Stable, Low Range
 Product Name: DynaMarker Protein MultiColor Stable, Low Range Code No: DM670L Lot No: ******* Size: 200 μl x 3 (DM670 x 3) (120 mini-gel lanes) Storage: 4 C Stability: 12 months at 4 C Storage Buffer:
Product Name: DynaMarker Protein MultiColor Stable, Low Range Code No: DM670L Lot No: ******* Size: 200 μl x 3 (DM670 x 3) (120 mini-gel lanes) Storage: 4 C Stability: 12 months at 4 C Storage Buffer:
Tivadar Orban, Beata Jastrzebska, Sayan Gupta, Benlian Wang, Masaru Miyagi, Mark R. Chance, and Krzysztof Palczewski
 Structure, Volume Supplemental Information Conformational Dynamics of Activation for the Pentameric Complex of Dimeric G Protein-Coupled Receptor and Heterotrimeric G Protein Tivadar Orban, Beata Jastrzebska,
Structure, Volume Supplemental Information Conformational Dynamics of Activation for the Pentameric Complex of Dimeric G Protein-Coupled Receptor and Heterotrimeric G Protein Tivadar Orban, Beata Jastrzebska,
Serum Amyloid A3 Gene Expression in Adipocytes is an Indicator. of the Interaction with Macrophages
 Serum Amyloid A3 Gene Expression in Adipocytes is an Indicator of the Interaction with Macrophages Yohei Sanada, Takafumi Yamamoto, Rika Satake, Akiko Yamashita, Sumire Kanai, Norihisa Kato, Fons AJ van
Serum Amyloid A3 Gene Expression in Adipocytes is an Indicator of the Interaction with Macrophages Yohei Sanada, Takafumi Yamamoto, Rika Satake, Akiko Yamashita, Sumire Kanai, Norihisa Kato, Fons AJ van
Chromatin Immunoprecipitation (ChIPs) Protocol (Mirmira Lab)
 Protocol (Mirmira Lab)") Chromatin Immunoprecipitation (ChIPs) Protocol (Mirmira Lab) Updated 12/3/02 Reagents: ChIP sonication Buffer (1% Triton X-100, 0.1% Deoxycholate, 50 mm Tris 8.1, 150 mm NaCl, 5 mm EDTA): 10 ml 10 % Triton
Chromatin Immunoprecipitation (ChIPs) Protocol (Mirmira Lab) Updated 12/3/02 Reagents: ChIP sonication Buffer (1% Triton X-100, 0.1% Deoxycholate, 50 mm Tris 8.1, 150 mm NaCl, 5 mm EDTA): 10 ml 10 % Triton
TRANSPORT OF AMINO ACIDS IN INTACT 3T3 AND SV3T3 CELLS. Binding Activity for Leucine in Membrane Preparations of Ehrlich Ascites Tumor Cells
 Journal of Supramolecular Structure 4:441 (401)-447 (407) (1976) TRANSPORT OF AMINO ACIDS IN INTACT 3T3 AND SV3T3 CELLS. Binding Activity for Leucine in Membrane Preparations of Ehrlich Ascites Tumor Cells
Journal of Supramolecular Structure 4:441 (401)-447 (407) (1976) TRANSPORT OF AMINO ACIDS IN INTACT 3T3 AND SV3T3 CELLS. Binding Activity for Leucine in Membrane Preparations of Ehrlich Ascites Tumor Cells
HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation
 SUPPLEMENTARY INFORMATION Materials and Methods Human cell lines and culture conditions HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation in exon 20 of BRCA1
SUPPLEMENTARY INFORMATION Materials and Methods Human cell lines and culture conditions HCC1937 is the HCC1937-pcDNA3 cell line, which was derived from a breast cancer with a mutation in exon 20 of BRCA1
nachr α 4 β 2 CHO Cell Line
 B SYS GmbH nachr α 4 β 2 CHO Cell Line Cell Culture Conditions B SYS GmbH B SYS GmbH nachr α 4 β 2 CHO Page 2 TABLE OF CONTENTS 1 BACKGROUND...3 1.1 Human Nicotinic Acetylcholine Receptors...3 1.2 B SYS
B SYS GmbH nachr α 4 β 2 CHO Cell Line Cell Culture Conditions B SYS GmbH B SYS GmbH nachr α 4 β 2 CHO Page 2 TABLE OF CONTENTS 1 BACKGROUND...3 1.1 Human Nicotinic Acetylcholine Receptors...3 1.2 B SYS
SUPPLEMENTAL INFORMATION
 SUPPLEMENTAL INFORMATION EXPERIMENTAL PROCEDURES Tryptic digestion protection experiments - PCSK9 with Ab-3D5 (1:1 molar ratio) in 50 mm Tris, ph 8.0, 150 mm NaCl was incubated overnight at 4 o C. The
SUPPLEMENTAL INFORMATION EXPERIMENTAL PROCEDURES Tryptic digestion protection experiments - PCSK9 with Ab-3D5 (1:1 molar ratio) in 50 mm Tris, ph 8.0, 150 mm NaCl was incubated overnight at 4 o C. The
Essential Medium, containing 10% fetal bovine serum, 100 U/ml penicillin and 100 µg/ml streptomycin. Huvec were cultured in
 Supplemental data Methods Cell culture media formulations A-431 and U-87 MG cells were maintained in Dulbecco s Modified Eagle s Medium. FaDu cells were cultured in Eagle's Minimum Essential Medium, containing
Supplemental data Methods Cell culture media formulations A-431 and U-87 MG cells were maintained in Dulbecco s Modified Eagle s Medium. FaDu cells were cultured in Eagle's Minimum Essential Medium, containing
Self-association of α-chymotrypsin: Effect of amino acids
 J. Biosci., Vol. 13, Number 3, September 1988, pp. 215 222. Printed in India. Self-association of α-chymotrypsin: Effect of amino acids T. RAMAKRISHNA and M. W. PANDIT* Centre for Cellular and Molecular
J. Biosci., Vol. 13, Number 3, September 1988, pp. 215 222. Printed in India. Self-association of α-chymotrypsin: Effect of amino acids T. RAMAKRISHNA and M. W. PANDIT* Centre for Cellular and Molecular
PREPARATION OF IF- ENRICHED CYTOSKELETAL PROTEINS
 TMM,5-2011 PREPARATION OF IF- ENRICHED CYTOSKELETAL PROTEINS Ice-cold means cooled in ice water. In order to prevent proteolysis, make sure to perform all steps on ice. Pre-cool glass homogenizers, buffers
TMM,5-2011 PREPARATION OF IF- ENRICHED CYTOSKELETAL PROTEINS Ice-cold means cooled in ice water. In order to prevent proteolysis, make sure to perform all steps on ice. Pre-cool glass homogenizers, buffers
MicroRNA sponges: competitive inhibitors of small RNAs in mammalian cells
 MicroRNA sponges: competitive inhibitors of small RNAs in mammalian cells Margaret S Ebert, Joel R Neilson & Phillip A Sharp Supplementary figures and text: Supplementary Figure 1. Effect of sponges on
MicroRNA sponges: competitive inhibitors of small RNAs in mammalian cells Margaret S Ebert, Joel R Neilson & Phillip A Sharp Supplementary figures and text: Supplementary Figure 1. Effect of sponges on
Supplementary Data 1. Alanine substitutions and position variants of APNCYGNIPL. Applied in
 Supplementary Data 1. Alanine substitutions and position variants of APNCYGNIPL. Applied in Supplementary Fig. 2 Substitution Sequence Position variant Sequence original APNCYGNIPL original APNCYGNIPL
Supplementary Data 1. Alanine substitutions and position variants of APNCYGNIPL. Applied in Supplementary Fig. 2 Substitution Sequence Position variant Sequence original APNCYGNIPL original APNCYGNIPL
Kit for assay of thioredoxin
 FkTRX-02-V2 Kit for assay of thioredoxin The thioredoxin system is the major protein disulfide reductase in cells and comprises thioredoxin, thioredoxin reductase and NADPH (1). Thioredoxin systems are
FkTRX-02-V2 Kit for assay of thioredoxin The thioredoxin system is the major protein disulfide reductase in cells and comprises thioredoxin, thioredoxin reductase and NADPH (1). Thioredoxin systems are
A protocol for enhancement of the AAV-mediated expression of transgenes
 A protocol for enhancement of the AAV-mediated expression of transgenes Hiroaki Mizukami, Takeharu Kanazawa, Takashi Okada, and Keiya Ozawa Division of Genetic Therapeutics, Center for Molecular Medicine,
A protocol for enhancement of the AAV-mediated expression of transgenes Hiroaki Mizukami, Takeharu Kanazawa, Takashi Okada, and Keiya Ozawa Division of Genetic Therapeutics, Center for Molecular Medicine,
Selective protection of an ARF1-GTP signaling axis by a bacterial scaffold induces bidirectional trafficking arrest.
 Selective protection of an ARF1-GTP signaling axis by a bacterial scaffold induces bidirectional trafficking arrest. Andrey S. Selyunin, L. Evan Reddick, Bethany A. Weigele, and Neal M. Alto Supplemental
Selective protection of an ARF1-GTP signaling axis by a bacterial scaffold induces bidirectional trafficking arrest. Andrey S. Selyunin, L. Evan Reddick, Bethany A. Weigele, and Neal M. Alto Supplemental
Supplemental Experimental Procedures
 Cell Stem Cell, Volume 2 Supplemental Data A Temporal Switch from Notch to Wnt Signaling in Muscle Stem Cells Is Necessary for Normal Adult Myogenesis Andrew S. Brack, Irina M. Conboy, Michael J. Conboy,
Cell Stem Cell, Volume 2 Supplemental Data A Temporal Switch from Notch to Wnt Signaling in Muscle Stem Cells Is Necessary for Normal Adult Myogenesis Andrew S. Brack, Irina M. Conboy, Michael J. Conboy,
Effects of Second Messengers
 Effects of Second Messengers Inositol trisphosphate Diacylglycerol Opens Calcium Channels Binding to IP 3 -gated Channel Cooperative binding Activates Protein Kinase C is required Phosphorylation of many
Effects of Second Messengers Inositol trisphosphate Diacylglycerol Opens Calcium Channels Binding to IP 3 -gated Channel Cooperative binding Activates Protein Kinase C is required Phosphorylation of many
SUPPLEMENTAL MATERIAL. Supplementary Methods
 SUPPLEMENTAL MATERIAL Supplementary Methods Culture of cardiomyocytes, fibroblasts and cardiac microvascular endothelial cells The isolation and culturing of neonatal rat ventricular cardiomyocytes was
SUPPLEMENTAL MATERIAL Supplementary Methods Culture of cardiomyocytes, fibroblasts and cardiac microvascular endothelial cells The isolation and culturing of neonatal rat ventricular cardiomyocytes was
Table S1. Sequence of human and mouse primers used for RT-qPCR measurements.
 Table S1. Sequence of human and mouse primers used for RT-qPCR measurements. Ca9, carbonic anhydrase IX; Ndrg1, N-myc downstream regulated gene 1; L28, ribosomal protein L28; Hif1a, hypoxia inducible factor
Table S1. Sequence of human and mouse primers used for RT-qPCR measurements. Ca9, carbonic anhydrase IX; Ndrg1, N-myc downstream regulated gene 1; L28, ribosomal protein L28; Hif1a, hypoxia inducible factor
Supplementary material: Materials and suppliers
 Supplementary material: Materials and suppliers Electrophoresis consumables including tris-glycine, acrylamide, SDS buffer and Coomassie Brilliant Blue G-2 dye (CBB) were purchased from Ameresco (Solon,
Supplementary material: Materials and suppliers Electrophoresis consumables including tris-glycine, acrylamide, SDS buffer and Coomassie Brilliant Blue G-2 dye (CBB) were purchased from Ameresco (Solon,
Manual. Precision Red Advanced Protein Assay Reagent. Cat. # ADV02. cytoskeleton.com. Cytoskeleton, Inc.
 The Protein Experts Manual Cytoskeleton, Inc. V. 6.0 Precision Red Advanced Protein Assay Reagent Cat. # ADV02 cytoskeleton.com Phone: (303) 322.2254 Fax: (303) 322.2257 Customer Service: cserve@cytoskeleton.com
The Protein Experts Manual Cytoskeleton, Inc. V. 6.0 Precision Red Advanced Protein Assay Reagent Cat. # ADV02 cytoskeleton.com Phone: (303) 322.2254 Fax: (303) 322.2257 Customer Service: cserve@cytoskeleton.com
Online Data Supplement. Anti-aging Gene Klotho Enhances Glucose-induced Insulin Secretion by Upregulating Plasma Membrane Retention of TRPV2
 Online Data Supplement Anti-aging Gene Klotho Enhances Glucose-induced Insulin Secretion by Upregulating Plasma Membrane Retention of TRPV2 Yi Lin and Zhongjie Sun Department of physiology, college of
Online Data Supplement Anti-aging Gene Klotho Enhances Glucose-induced Insulin Secretion by Upregulating Plasma Membrane Retention of TRPV2 Yi Lin and Zhongjie Sun Department of physiology, college of
Purification of Glucagon3 Interleukin-2 Fusion Protein Derived from E. coli
 Purification of Glucagon3 Interleukin-2 Fusion Protein Derived from E. coli Hye Soon Won Dept. of Chem. Eng. Chungnam National University INTRODUCTION Human interleukin-2(hil-2) - known as T Cell Growth
Purification of Glucagon3 Interleukin-2 Fusion Protein Derived from E. coli Hye Soon Won Dept. of Chem. Eng. Chungnam National University INTRODUCTION Human interleukin-2(hil-2) - known as T Cell Growth
Mammalian Membrane Protein Extraction Kit
 Mammalian Membrane Protein Extraction Kit Catalog number: AR0155 Boster s Mammalian Membrane Protein Extraction Kit is a simple, rapid and reproducible method to prepare cellular protein fractions highly
Mammalian Membrane Protein Extraction Kit Catalog number: AR0155 Boster s Mammalian Membrane Protein Extraction Kit is a simple, rapid and reproducible method to prepare cellular protein fractions highly
Instructions for Use. APO-AB Annexin V-Biotin Apoptosis Detection Kit 100 tests
 3URGXFW,QIRUPDWLRQ Sigma TACS Annexin V Apoptosis Detection Kits Instructions for Use APO-AB Annexin V-Biotin Apoptosis Detection Kit 100 tests For Research Use Only. Not for use in diagnostic procedures.
3URGXFW,QIRUPDWLRQ Sigma TACS Annexin V Apoptosis Detection Kits Instructions for Use APO-AB Annexin V-Biotin Apoptosis Detection Kit 100 tests For Research Use Only. Not for use in diagnostic procedures.
Amajor metabolic consequence of insulin action is
 Glycogen Synthase Sensitivity to Insulin and Glucose-6-Phosphate Is Mediated by Both NH 2 - and COOH-Terminal Phosphorylation Sites Alexander V. Skurat, Amy D. Dietrich, and Peter J. Roach In skeletal
Glycogen Synthase Sensitivity to Insulin and Glucose-6-Phosphate Is Mediated by Both NH 2 - and COOH-Terminal Phosphorylation Sites Alexander V. Skurat, Amy D. Dietrich, and Peter J. Roach In skeletal
ab E3 Ligase Auto- Ubiquitilylation Assay Kit
 ab139469 E3 Ligase Auto- Ubiquitilylation Assay Kit Instructions for Use For testing ubiquitin E3 ligase activity through assessment of their ability to undergo auto-ubiquitinylation This product is for
ab139469 E3 Ligase Auto- Ubiquitilylation Assay Kit Instructions for Use For testing ubiquitin E3 ligase activity through assessment of their ability to undergo auto-ubiquitinylation This product is for
Protocol for purification of recombinant protein from 300 ml yeast culture
 Protocol for purification of recombinant protein from 300 ml yeast culture Equipment and reagents needed: Zirconia beads (0.5 mm diameter from BSP, Germany) Paint Shaker (at 4 C) Tube rotator for 15 ml
Protocol for purification of recombinant protein from 300 ml yeast culture Equipment and reagents needed: Zirconia beads (0.5 mm diameter from BSP, Germany) Paint Shaker (at 4 C) Tube rotator for 15 ml
Supplementary Material for
 Supplementary Material for Parathyroid Hormone Signaling through Low-density-lipoprotein-related Protein 6 Mei Wan, Chaozhe Yang, Jun Li, Xiangwei Wu, Hongling Yuan, Hairong Ma, Xi He, Shuyi Nie, Chenbei
Supplementary Material for Parathyroid Hormone Signaling through Low-density-lipoprotein-related Protein 6 Mei Wan, Chaozhe Yang, Jun Li, Xiangwei Wu, Hongling Yuan, Hairong Ma, Xi He, Shuyi Nie, Chenbei
AMPK Phosphorylation Assay Kit
 AMPK Phosphorylation Assay Kit Catalog Number KA3789 100 assays Version: 02 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Intended Use... 3 Background... 3 Principle
AMPK Phosphorylation Assay Kit Catalog Number KA3789 100 assays Version: 02 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Intended Use... 3 Background... 3 Principle
Antibodies: LB1 buffer For 50 ml For 10ml For 30 ml Final 1 M HEPES, ph 2.5 ml 0.5 ml 1.5 ml 50mM. 5 M NaCl 1.4 ml 280 µl 0.
 Experiment: Date: Tissue: Purpose: ChIP-Seq Antibodies: 11x cross-link buffer: Regent Stock Solution Final Vol for 10 ml of 11xstock concentration 5 M NaCl 0.1M 0.2 ml 0.5 M EDTA 1 mm 20 ul 0.5 M EGTA,
Experiment: Date: Tissue: Purpose: ChIP-Seq Antibodies: 11x cross-link buffer: Regent Stock Solution Final Vol for 10 ml of 11xstock concentration 5 M NaCl 0.1M 0.2 ml 0.5 M EDTA 1 mm 20 ul 0.5 M EGTA,
Construction of a hepatocellular carcinoma cell line that stably expresses stathmin with a Ser25 phosphorylation site mutation
 Construction of a hepatocellular carcinoma cell line that stably expresses stathmin with a Ser25 phosphorylation site mutation J. Du 1, Z.H. Tao 2, J. Li 2, Y.K. Liu 3 and L. Gan 2 1 Department of Chemistry,
Construction of a hepatocellular carcinoma cell line that stably expresses stathmin with a Ser25 phosphorylation site mutation J. Du 1, Z.H. Tao 2, J. Li 2, Y.K. Liu 3 and L. Gan 2 1 Department of Chemistry,
J. Biosci., Vol. 7, Number 2, March 1985, pp Printed in India.
 J. Biosci., Vol. 7, Number 2, March 1985, pp. 123 133. Printed in India. Irreversibility of the interaction of human growth hormone with its receptor and analysis of irreversible reactions in radioreceptor
J. Biosci., Vol. 7, Number 2, March 1985, pp. 123 133. Printed in India. Irreversibility of the interaction of human growth hormone with its receptor and analysis of irreversible reactions in radioreceptor
Glycosyltransferase Activity Kit
 Glycosyltransferase Activity Kit Catalog Number EA001 This package insert must be read in its entirety before using this product. For research use only. Not for use in diagnostic procedures. TABLE OF CONTENTS
Glycosyltransferase Activity Kit Catalog Number EA001 This package insert must be read in its entirety before using this product. For research use only. Not for use in diagnostic procedures. TABLE OF CONTENTS
TRAF6 ubiquitinates TGFβ type I receptor to promote its cleavage and nuclear translocation in cancer
 Supplementary Information TRAF6 ubiquitinates TGFβ type I receptor to promote its cleavage and nuclear translocation in cancer Yabing Mu, Reshma Sundar, Noopur Thakur, Maria Ekman, Shyam Kumar Gudey, Mariya
Supplementary Information TRAF6 ubiquitinates TGFβ type I receptor to promote its cleavage and nuclear translocation in cancer Yabing Mu, Reshma Sundar, Noopur Thakur, Maria Ekman, Shyam Kumar Gudey, Mariya
Separation of a phosphorylated-his protein using phosphate-affinity polyacrylamide gel electrophoresis
 Notes & Tips Separation of a phosphorylated-his protein using phosphate-affinity polyacrylamide gel electrophoresis Categories: Electrophoretic Techniques Seiji Yamada a,*, Hiro Nakamura a,b, Eiji Kinoshita
Notes & Tips Separation of a phosphorylated-his protein using phosphate-affinity polyacrylamide gel electrophoresis Categories: Electrophoretic Techniques Seiji Yamada a,*, Hiro Nakamura a,b, Eiji Kinoshita
20X Buffer (Tube1) 96-well microplate (12 strips) 1
 96-well microplate (12 strips) 1") PROTOCOL MitoProfile Rapid Microplate Assay Kit for PDH Activity and Quantity (Combines Kit MSP18 & MSP19) 1850 Millrace Drive, Suite 3A Eugene, Oregon 97403 MSP20 Rev.1 DESCRIPTION MitoProfile Rapid Microplate
PROTOCOL MitoProfile Rapid Microplate Assay Kit for PDH Activity and Quantity (Combines Kit MSP18 & MSP19) 1850 Millrace Drive, Suite 3A Eugene, Oregon 97403 MSP20 Rev.1 DESCRIPTION MitoProfile Rapid Microplate
Validation & Assay Performance Summary
 Validation & Assay Performance Summary LanthaScreen IGF-1R GripTite Cells Cat. no. K1834 Modification Detected: Phosphorylation of Multiple Tyr Residues on IGF-1R LanthaScreen Cellular Assay Validation
Validation & Assay Performance Summary LanthaScreen IGF-1R GripTite Cells Cat. no. K1834 Modification Detected: Phosphorylation of Multiple Tyr Residues on IGF-1R LanthaScreen Cellular Assay Validation
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION FOR Liver X Receptor α mediates hepatic triglyceride accumulation through upregulation of G0/G1 Switch Gene 2 (G0S2) expression I: SUPPLEMENTARY METHODS II: SUPPLEMENTARY FIGURES
SUPPLEMENTARY INFORMATION FOR Liver X Receptor α mediates hepatic triglyceride accumulation through upregulation of G0/G1 Switch Gene 2 (G0S2) expression I: SUPPLEMENTARY METHODS II: SUPPLEMENTARY FIGURES
2,6,9-Triazabicyclo[3.3.1]nonanes as overlooked. amino-modification products by acrolein
![2,6,9-Triazabicyclo[3.3.1]nonanes as overlooked. amino-modification products by acrolein](/thumbs/86/94743397.jpg "2,6,9-Triazabicyclo[3.3.1]nonanes as overlooked. amino-modification products by acrolein") Supplementary Information 2,6,9-Triazabicyclo[3.3.1]nonanes as overlooked amino-modification products by acrolein Ayumi Tsutsui and Katsunori Tanaka* Biofunctional Synthetic Chemistry Laboratory, RIKEN
Supplementary Information 2,6,9-Triazabicyclo[3.3.1]nonanes as overlooked amino-modification products by acrolein Ayumi Tsutsui and Katsunori Tanaka* Biofunctional Synthetic Chemistry Laboratory, RIKEN
A Hepatocyte Growth Factor Receptor (Met) Insulin Receptor hybrid governs hepatic glucose metabolism SUPPLEMENTARY FIGURES, LEGENDS AND METHODS
 Insulin Receptor hybrid governs hepatic glucose metabolism SUPPLEMENTARY FIGURES, LEGENDS AND METHODS") A Hepatocyte Growth Factor Receptor (Met) Insulin Receptor hybrid governs hepatic glucose metabolism Arlee Fafalios, Jihong Ma, Xinping Tan, John Stoops, Jianhua Luo, Marie C. DeFrances and Reza Zarnegar
A Hepatocyte Growth Factor Receptor (Met) Insulin Receptor hybrid governs hepatic glucose metabolism Arlee Fafalios, Jihong Ma, Xinping Tan, John Stoops, Jianhua Luo, Marie C. DeFrances and Reza Zarnegar
Trident Membrane Protein Extraction Kit
 Cat. No. Size Shelf life GTX16373 5/ 20 tests 12 months at the appropriate storage temperatures (see below) Contents Component Storage Amount for 5 tests Amount for 20 tests Buffer A -20 o C 2.5 ml 10
Cat. No. Size Shelf life GTX16373 5/ 20 tests 12 months at the appropriate storage temperatures (see below) Contents Component Storage Amount for 5 tests Amount for 20 tests Buffer A -20 o C 2.5 ml 10
Work-flow: protein sample preparation Precipitation methods Removal of interfering substances Specific examples:
 Dr. Sanjeeva Srivastava IIT Bombay Work-flow: protein sample preparation Precipitation methods Removal of interfering substances Specific examples: Sample preparation for serum proteome analysis Sample
Dr. Sanjeeva Srivastava IIT Bombay Work-flow: protein sample preparation Precipitation methods Removal of interfering substances Specific examples: Sample preparation for serum proteome analysis Sample
MTC-TT and TPC-1 cell lines were cultured in RPMI medium (Gibco, Breda, The Netherlands)
") Supplemental data Materials and Methods Cell culture MTC-TT and TPC-1 cell lines were cultured in RPMI medium (Gibco, Breda, The Netherlands) supplemented with 15% or 10% (for TPC-1) fetal bovine serum
Supplemental data Materials and Methods Cell culture MTC-TT and TPC-1 cell lines were cultured in RPMI medium (Gibco, Breda, The Netherlands) supplemented with 15% or 10% (for TPC-1) fetal bovine serum
Plasmids Western blot analysis and immunostaining Flow Cytometry Cell surface biotinylation RNA isolation and cdna synthesis
 Plasmids psuper-retro-s100a10 shrna1 was constructed by cloning the dsdna oligo 5 -GAT CCC CGT GGG CTT CCA GAG CTT CTT TCA AGA GAA GAA GCT CTG GAA GCC CAC TTT TTA-3 and 5 -AGC TTA AAA AGT GGG CTT CCA GAG
Plasmids psuper-retro-s100a10 shrna1 was constructed by cloning the dsdna oligo 5 -GAT CCC CGT GGG CTT CCA GAG CTT CTT TCA AGA GAA GAA GCT CTG GAA GCC CAC TTT TTA-3 and 5 -AGC TTA AAA AGT GGG CTT CCA GAG
The rabbit femoral artery was prepared and each arterial ring was permeabilized
 Online Supplement Nakmura et al. cgmp-dependent relaxation of smooth muscle Materials and Methods Measurement of tension The rabbit femoral artery was prepared and each arterial ring was permeabilized
Online Supplement Nakmura et al. cgmp-dependent relaxation of smooth muscle Materials and Methods Measurement of tension The rabbit femoral artery was prepared and each arterial ring was permeabilized
T H E J O U R N A L O F C E L L B I O L O G Y
 Supplemental material Jewell et al., http://www.jcb.org/cgi/content/full/jcb.201007176/dc1 T H E J O U R N A L O F C E L L B I O L O G Y Figure S1. IR Munc18c association is independent of IRS-1. (A and
Supplemental material Jewell et al., http://www.jcb.org/cgi/content/full/jcb.201007176/dc1 T H E J O U R N A L O F C E L L B I O L O G Y Figure S1. IR Munc18c association is independent of IRS-1. (A and
Caution: For Laboratory Use. A product for research purposes only. Eu-W1284 Iodoacetamido Chelate & Europium Standard. Product Number: AD0014
 TECHNICAL DATA SHEET Lance Caution: For Laboratory Use. A product for research purposes only. Eu-W1284 Iodoacetamido Chelate & Europium Standard Product Number: AD0014 INTRODUCTION: Iodoacetamido-activated
TECHNICAL DATA SHEET Lance Caution: For Laboratory Use. A product for research purposes only. Eu-W1284 Iodoacetamido Chelate & Europium Standard Product Number: AD0014 INTRODUCTION: Iodoacetamido-activated
Supplementary Information
 Supplementary Information Structural basis of improved second generation 3-nitro-tyrosine trna synthetases Richard B. Cooley, Jessica L. Feldman, Camden M. Driggers, Taylor Bundy, Audrey L. Stokes, P.
Supplementary Information Structural basis of improved second generation 3-nitro-tyrosine trna synthetases Richard B. Cooley, Jessica L. Feldman, Camden M. Driggers, Taylor Bundy, Audrey L. Stokes, P.
Islet viability assay and Glucose Stimulated Insulin Secretion assay RT-PCR and Western Blot
 Islet viability assay and Glucose Stimulated Insulin Secretion assay Islet cell viability was determined by colorimetric (3-(4,5-dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide assay using CellTiter
Islet viability assay and Glucose Stimulated Insulin Secretion assay Islet cell viability was determined by colorimetric (3-(4,5-dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide assay using CellTiter
ab Membrane Fractionation Kit Instructions for Use For the rapid and simple separation of membrane, cytosolic and nuclear cellular fractions.
 ab139409 Membrane Fractionation Kit Instructions for Use For the rapid and simple separation of membrane, cytosolic and nuclear cellular fractions. This product is for research use only and is not intended
ab139409 Membrane Fractionation Kit Instructions for Use For the rapid and simple separation of membrane, cytosolic and nuclear cellular fractions. This product is for research use only and is not intended
Supplementary Figure 1.TRIM33 binds β-catenin in the nucleus. a & b, Co-IP of endogenous TRIM33 with β-catenin in HT-29 cells (a) and HEK 293T cells
 and HEK 293T cells") Supplementary Figure 1.TRIM33 binds β-catenin in the nucleus. a & b, Co-IP of endogenous TRIM33 with β-catenin in HT-29 cells (a) and HEK 293T cells (b). TRIM33 was immunoprecipitated, and the amount of
Supplementary Figure 1.TRIM33 binds β-catenin in the nucleus. a & b, Co-IP of endogenous TRIM33 with β-catenin in HT-29 cells (a) and HEK 293T cells (b). TRIM33 was immunoprecipitated, and the amount of
<Supplemental information>
 The Structural Basis of Endosomal Anchoring of KIF16B Kinesin Nichole R. Blatner, Michael I. Wilson, Cai Lei, Wanjin Hong, Diana Murray, Roger L. Williams, and Wonhwa Cho Protein
The Structural Basis of Endosomal Anchoring of KIF16B Kinesin Nichole R. Blatner, Michael I. Wilson, Cai Lei, Wanjin Hong, Diana Murray, Roger L. Williams, and Wonhwa Cho Protein
2-Deoxyglucose Assay Kit (Colorimetric)
") 2-Deoxyglucose Assay Kit (Colorimetric) Catalog Number KA3753 100 assays Version: 01 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 General Information...
2-Deoxyglucose Assay Kit (Colorimetric) Catalog Number KA3753 100 assays Version: 01 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 General Information...
STORE AT 4 o C Version 3
 Instruction Manual Advanced Protein Assay Reagent (Cat. # ADV01) ORDERING INFORMATION To order by phone: (303) - 322-2254 To order by Fax: (303) - 322-2257 To order by e-mail: cserve@cytoskeleton.com Technical
Instruction Manual Advanced Protein Assay Reagent (Cat. # ADV01) ORDERING INFORMATION To order by phone: (303) - 322-2254 To order by Fax: (303) - 322-2257 To order by e-mail: cserve@cytoskeleton.com Technical
SUPPLEMENT. Materials and methods
 SUPPLEMENT Materials and methods Cell culture and reagents Cell media and reagents were from Invitrogen unless otherwise indicated. Antibiotics and Tet-certified serum were from Clontech. In experiments
SUPPLEMENT Materials and methods Cell culture and reagents Cell media and reagents were from Invitrogen unless otherwise indicated. Antibiotics and Tet-certified serum were from Clontech. In experiments
For the 5 GATC-overhang two-oligo adaptors set up the following reactions in 96-well plate format:
 Supplementary Protocol 1. Adaptor preparation: For the 5 GATC-overhang two-oligo adaptors set up the following reactions in 96-well plate format: Per reaction X96 10X NEBuffer 2 10 µl 10 µl x 96 5 -GATC
Supplementary Protocol 1. Adaptor preparation: For the 5 GATC-overhang two-oligo adaptors set up the following reactions in 96-well plate format: Per reaction X96 10X NEBuffer 2 10 µl 10 µl x 96 5 -GATC
APPENDIX Heparin 2 mg heparin was dissolved in 0.9 % NaCl (10 ml). 200 µl of heparin was added to each 1 ml of blood to prevent coagulation.
. 200 µl of heparin was added to each 1 ml of blood to prevent coagulation.") APPENDIX 1 Preparation of reagents 1.1. Preparation of dosing solution Nonylphenol 15 mg of Nonylphenol was dissolved in olive oil (10 ml) and used as stock solution. The stock solution was serially diluted
APPENDIX 1 Preparation of reagents 1.1. Preparation of dosing solution Nonylphenol 15 mg of Nonylphenol was dissolved in olive oil (10 ml) and used as stock solution. The stock solution was serially diluted
CHO α 1 β 2 γ 2 GABAA Cell Line
 B SYS GmbH CHO α 1 β 2 γ 2 GABAA Cell Line Specification Sheet B SYS GmbH B SYS GmbH CHO α 1 β 2 γ 2 Cells Page 2 TABLE OF CONTENTS 1 BACKGROUND...3 1.1 THE PHARMACOLOGICAL DISTINCTION OF GABA A RECEPTOR
B SYS GmbH CHO α 1 β 2 γ 2 GABAA Cell Line Specification Sheet B SYS GmbH B SYS GmbH CHO α 1 β 2 γ 2 Cells Page 2 TABLE OF CONTENTS 1 BACKGROUND...3 1.1 THE PHARMACOLOGICAL DISTINCTION OF GABA A RECEPTOR
BILAYER CHANNEL RECONSTITUTION
 (1) 1% Agar Salt Bridge 1.0 g Agar 3.75g KCl in 100ml distilled water, store at 4 o C. BILAYER CHANNEL RECONSTITUTION (2) Cs solution: (Cesium Methanesulfonate) 1) 50 mm Cs + solution 0.209 MOPS, 10mM
(1) 1% Agar Salt Bridge 1.0 g Agar 3.75g KCl in 100ml distilled water, store at 4 o C. BILAYER CHANNEL RECONSTITUTION (2) Cs solution: (Cesium Methanesulfonate) 1) 50 mm Cs + solution 0.209 MOPS, 10mM
DELFIA Tb-N1 DTA Chelate & Terbium Standard
 AD0029P-1 (en) 1 DELFIA Tb-N1 DTA Chelate & AD0012 Terbium Standard For Research Use Only INTRODUCTION DELFIA Tb-N1 DTA Chelate is optimized for the terbium labeling of proteins and peptides for use in
AD0029P-1 (en) 1 DELFIA Tb-N1 DTA Chelate & AD0012 Terbium Standard For Research Use Only INTRODUCTION DELFIA Tb-N1 DTA Chelate is optimized for the terbium labeling of proteins and peptides for use in
(Stratagene, La Jolla, CA) (Supplemental Fig. 1A). A 5.4-kb EcoRI fragment
 (Supplemental Fig. 1A). A 5.4-kb EcoRI fragment") SUPPLEMENTAL INFORMATION Supplemental Methods Generation of RyR2-S2808D Mice Murine genomic RyR2 clones were isolated from a 129/SvEvTacfBR λ-phage library (Stratagene, La Jolla, CA) (Supplemental Fig.
SUPPLEMENTAL INFORMATION Supplemental Methods Generation of RyR2-S2808D Mice Murine genomic RyR2 clones were isolated from a 129/SvEvTacfBR λ-phage library (Stratagene, La Jolla, CA) (Supplemental Fig.
FEBS 1138 January Paul R. Buckland and Bernard Rees Smith
 Volume 166, number 1 FEBS 1138 January 1984 A structural comparison receptors by of guinea pig thyroid and fat TSH photoaffinity labelling Paul R. Buckland and Bernard Rees Smith Endocrine Immunology Unit,
Volume 166, number 1 FEBS 1138 January 1984 A structural comparison receptors by of guinea pig thyroid and fat TSH photoaffinity labelling Paul R. Buckland and Bernard Rees Smith Endocrine Immunology Unit,
TSH Receptor Monoclonal Antibody (49) Catalog Number MA3-218 Product data sheet
 Catalog Number MA3-218 Product data sheet") Website: thermofisher.com Customer Service (US): 1 800 955 6288 ext. 1 Technical Support (US): 1 800 955 6288 ext. 441 TSH Receptor Monoclonal Antibody (49) Catalog Number MA3-218 Product data sheet Details
Website: thermofisher.com Customer Service (US): 1 800 955 6288 ext. 1 Technical Support (US): 1 800 955 6288 ext. 441 TSH Receptor Monoclonal Antibody (49) Catalog Number MA3-218 Product data sheet Details
Supplementary Material
 Supplementary Material Nuclear import of purified HIV-1 Integrase. Integrase remains associated to the RTC throughout the infection process until provirus integration occurs and is therefore one likely
Supplementary Material Nuclear import of purified HIV-1 Integrase. Integrase remains associated to the RTC throughout the infection process until provirus integration occurs and is therefore one likely
BIL 256 Cell and Molecular Biology Lab Spring, Tissue-Specific Isoenzymes
 BIL 256 Cell and Molecular Biology Lab Spring, 2007 Background Information Tissue-Specific Isoenzymes A. BIOCHEMISTRY The basic pattern of glucose oxidation is outlined in Figure 3-1. Glucose is split
BIL 256 Cell and Molecular Biology Lab Spring, 2007 Background Information Tissue-Specific Isoenzymes A. BIOCHEMISTRY The basic pattern of glucose oxidation is outlined in Figure 3-1. Glucose is split
Cell Lysis Buffer. Catalog number: AR0103
 Cell Lysis Buffer Catalog number: AR0103 Boster s Cell Lysis Buffer is a ready-to-use Western blot related reagent solution used for efficient extraction of total soluble protein in nondenatured state
Cell Lysis Buffer Catalog number: AR0103 Boster s Cell Lysis Buffer is a ready-to-use Western blot related reagent solution used for efficient extraction of total soluble protein in nondenatured state
Materials and Methods , The two-hybrid principle.
 The enzymatic activity of an unknown protein which cleaves the phosphodiester bond between the tyrosine residue of a viral protein and the 5 terminus of the picornavirus RNA Introduction Every day there
The enzymatic activity of an unknown protein which cleaves the phosphodiester bond between the tyrosine residue of a viral protein and the 5 terminus of the picornavirus RNA Introduction Every day there
Trypsin Mass Spectrometry Grade
 058PR-03 G-Biosciences 1-800-628-7730 1-314-991-6034 technical@gbiosciences.com A Geno Technology, Inc. (USA) brand name Trypsin Mass Spectrometry Grade A Chemically Modified, TPCK treated, Affinity Purified
058PR-03 G-Biosciences 1-800-628-7730 1-314-991-6034 technical@gbiosciences.com A Geno Technology, Inc. (USA) brand name Trypsin Mass Spectrometry Grade A Chemically Modified, TPCK treated, Affinity Purified
RayBio Human Phosphotyrosine BTK ELISA Kit
 RayBio Human Phosphotyrosine BTK ELISA Kit Catalog #: PEL-BTK-Y User Manual Last revised August 10, 2016 Caution: Extraordinarily useful information enclosed ISO 13485 Certified 3607 Parkway Lane, Suite
RayBio Human Phosphotyrosine BTK ELISA Kit Catalog #: PEL-BTK-Y User Manual Last revised August 10, 2016 Caution: Extraordinarily useful information enclosed ISO 13485 Certified 3607 Parkway Lane, Suite
L6 GLUT4myc Cell Growth Protocol
 L6 GLUT4myc Cell Growth Protocol Background: Parental L6 cells selected for high fusion (2, 3) were stably transfected with a rat GLUT4 cdna carrying a myc epitope (recognized by the commercially available
L6 GLUT4myc Cell Growth Protocol Background: Parental L6 cells selected for high fusion (2, 3) were stably transfected with a rat GLUT4 cdna carrying a myc epitope (recognized by the commercially available
A Homogeneous Phosphoinositide 3-Kinase Assay on Phospholipid FlashPlate Platforms. Busi Maswoswe, Hao Xie, Pat Kasila and Li-an Yeh
 A Homogeneous Phosphoinositide 3-Kinase Assay on Phospholipid FlashPlate Platforms Busi Maswoswe, Hao Xie, Pat Kasila and Li-an Yeh Abstract Phosphoinositide 3-kinases (PI 3-kinase) consist of a family
A Homogeneous Phosphoinositide 3-Kinase Assay on Phospholipid FlashPlate Platforms Busi Maswoswe, Hao Xie, Pat Kasila and Li-an Yeh Abstract Phosphoinositide 3-kinases (PI 3-kinase) consist of a family
Substitution of the erbb-2 oncoprotein transmembrane domain
 Proc. Natl. Acad. Sci. USA Vol. 9, pp. 7336-734, August 1993 Cell Biology Substitution of the erbb-2 oncoprotein transmembrane domain activates the insulin receptor and modulates the action of insulin
Proc. Natl. Acad. Sci. USA Vol. 9, pp. 7336-734, August 1993 Cell Biology Substitution of the erbb-2 oncoprotein transmembrane domain activates the insulin receptor and modulates the action of insulin
Bioluminescence Resonance Energy Transfer (BRET)-based studies of receptor dynamics in living cells with Berthold s Mithras
-based studies of receptor dynamics in living cells with Berthold s Mithras") Bioluminescence Resonance Energy Transfer (BRET)-based studies of receptor dynamics in living cells with Berthold s Mithras Tarik Issad, Ralf Jockers and Stefano Marullo 1 Because they play a pivotal role
Bioluminescence Resonance Energy Transfer (BRET)-based studies of receptor dynamics in living cells with Berthold s Mithras Tarik Issad, Ralf Jockers and Stefano Marullo 1 Because they play a pivotal role
Gladstone Institutes, University of California (UCSF), San Francisco, USA
, San Francisco, USA") Fluorescence-linked Antigen Quantification (FLAQ) Assay for Fast Quantification of HIV-1 p24 Gag Marianne Gesner, Mekhala Maiti, Robert Grant and Marielle Cavrois * Gladstone Institutes, University of
Fluorescence-linked Antigen Quantification (FLAQ) Assay for Fast Quantification of HIV-1 p24 Gag Marianne Gesner, Mekhala Maiti, Robert Grant and Marielle Cavrois * Gladstone Institutes, University of
Control of Cell Proliferation by Peptide Growth Factors. Autocrine Growth Factor Production Causes Malignant Transformation?
 Control of Cell Proliferation by Peptide Growth Factors Autocrine Growth Factor Production Causes Malignant Transformation? Transforming Activities From Condition Media from a Tumor Cell Line Condition
Control of Cell Proliferation by Peptide Growth Factors Autocrine Growth Factor Production Causes Malignant Transformation? Transforming Activities From Condition Media from a Tumor Cell Line Condition
The Immunoassay Guide to Successful Mass Spectrometry. Orr Sharpe Robinson Lab SUMS User Meeting October 29, 2013
 The Immunoassay Guide to Successful Mass Spectrometry Orr Sharpe Robinson Lab SUMS User Meeting October 29, 2013 What is it? Hey! Look at that! Something is reacting in here! I just wish I knew what it
The Immunoassay Guide to Successful Mass Spectrometry Orr Sharpe Robinson Lab SUMS User Meeting October 29, 2013 What is it? Hey! Look at that! Something is reacting in here! I just wish I knew what it
Europium Labeling Kit
 Europium Labeling Kit Catalog Number KA2096 100ug *1 Version: 03 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Intended Use... 3 Background... 3 Principle of the Assay...
Europium Labeling Kit Catalog Number KA2096 100ug *1 Version: 03 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Intended Use... 3 Background... 3 Principle of the Assay...
Cholesterol determination using protein-templated fluorescent gold nanocluster probes
 Electronic Supplementary Information for Cholesterol determination using protein-templated fluorescent gold nanocluster probes Xi Chen and Gary A. Baker* Department of Chemistry, University of Missouri-Columbia,
Electronic Supplementary Information for Cholesterol determination using protein-templated fluorescent gold nanocluster probes Xi Chen and Gary A. Baker* Department of Chemistry, University of Missouri-Columbia,
AMPK Assay. Require: Sigma (1L, $18.30) A4206 Aluminum foil
 A4206 Aluminum foil") AMPK Assay Require: Acetone Sigma (1L, $18.30) A4206 Aluminum foil Ammonium sulfate Fisher BP212R-1 AMP Sigma A1752 ATP Sigma A6144 (alt. use A7699) Beta-mercaptoethanol Sigma M6250 (alt. use M7154) Bio-Rad
AMPK Assay Require: Acetone Sigma (1L, $18.30) A4206 Aluminum foil Ammonium sulfate Fisher BP212R-1 AMP Sigma A1752 ATP Sigma A6144 (alt. use A7699) Beta-mercaptoethanol Sigma M6250 (alt. use M7154) Bio-Rad