A Critical Role for Gimap5 in CD4+ T cell Homeostasis and Maintenance of Peripheral Immune Tolerance
|
|
|
- Edmund Owen
- 5 years ago
- Views:
Transcription
1
2 A Critical Role for Gimap5 in CD4+ T cell Homeostasis and Maintenance of Peripheral Immune Tolerance A dissertation submitted to the Graduate School of the University of Cincinnati in partial fulfillment of the requirements for the degree of Doctor of Philosophy (Ph.D.) in the Immunobiology Graduate Program of the College of Medicine 2013 by Halil Ibrahim Aksoylar B.S., Middle East Technical University, Turkey, 2003 M.S., Sabanci University, Turkey, 2005 Committee Chair: Kasper Hoebe, Ph.D. Christopher Karp, M.D Edith Janssen, Ph.D. Julio Aliberti, Ph.D. David Plas, Ph.D.
3 Abstract T cell lymphopenia is a condition which arises from defects in T cell development and/or peripheral homeostatic mechanisms. Importantly, lymphopenia is often associated with T cell-mediated pathology in animal models and in patients with autoimmune disease. In this thesis, using an ENU mutagenesis approach, we identified sphinx mice which presented severe lymphopenia due to a missense mutation in Gimap5. Characterization of Gimap5 sph/sph mice revealed that Gimap5 is necessary for the development of NK and CD8 + T cells, and is required for the maintenance of peripheral CD4 + T and B cell populations. Moreover, Gimap5-deficient mice developed spontaneous colitis which resulted in early mortality. Gimap5 sph/sph CD4 + T cells presented progressive lymphopenia-induced proliferation (LIP), became Th1/Th17 polarized, and mediated the development of colitis. Furthermore, Gimap5 sph/sph FoxP3 + regulatory T cells became selectively reduced in the mesenteric lymph nodes and adoptive transfer of wild type regulatory T cells prevented colitis in Gimap5-deficient mice. Importantly, the expression of Foxo transcription factors, which play a critical role in T quiescence and Treg function, was progressively lost in the absence of Gimap5 suggesting a link between Gimap5 deficiency and loss of immunological tolerance. Using OT-II RAG -/- TCR transgenic model, we showed that treatment with cognate antigen under tolerizing conditions failed to induce a Treg population and resulted in the acquisition of LIP phenotype by Gimap5-deficient CD4 + T cells. Given that Gimap5 is expressed in lysosomes, we investigated whether Gimap5 is involved in lysosomalautophagosomal pathways. Upon TCR activation, we observed larger autophagosomes that colocalize with mitochondria in Gimap5 sph/sph CD4 + T cells suggesting an abnormal rate of mitochondrial turnover. Furthermore, TCR activated Gimap5 sph/sph CD4 + T cells ii
4 displayed elevated levels of reactive oxygen species (ROS) and oxygen consumption rate (OCR) indicating defects in mitochondrial function. Our results establish the critical role of Gimap5 in CD4 + T cell homeostasis and maintenance of peripheral tolerance. Importantly, our results provide a basis for further investigation of the molecular mechanisms how Gimap5 is involved in T cell homeostasis. iii
5 iv
6 Acknowledgements: I would like to express my sincere gratitude to my advisor, Dr. Kasper Hoebe, for his support and guidance during these past five years. His enthusiasm for science has been a motivation for me throughout my research training. I would also like to thank the members of my committee, Dr. Christpher Karp, Dr. Edith Janssen, Dr. David Plas, Dr. Julio Aliberti and Graduate Program Director Dr. David Hildeman for their valuable advice and support. I am also thankful to the past and present members of Hoebe and Janssen Labs, Kristin Lampe, Nate Harris, Rachel Reboulet, Dr. Hao Fang, Dr. Rob Tacker, Cassie Hennies and Maria Lehn for their help in my lab work. I would like to express my deepest gratitude to my family; my parents, Gulumser and Dr. Yasar Aksoylar; my sisters, Sevinc Ozturk and Dr. Gulcin Eken; and my wife Dr. Sema Kurtulus Aksoylar for their unconditional support and belief in me during my graduate education and throughout my life. v
7 Table of contents: Page Abstract Acknowledgements Table of Contents List of Abbreviations ii v vi ix Chapter I: Introduction 1 1. Thesis Introduction 2 2. T cell development 2 3. Peripheral Homeostasis of T cells Peripheral survival and homeostatic proliferation Activation and effector differentiation of CD4 + T cells Memory and memory phenotype T cells T cell quiescence Immune Tolerance mechanisms Central Tolerance of T cells Peripheral T cell Tolerance T cell Anergy Inhibitory Costimulatory Molecules Regulatory T cells Identification of Regulatory T cells Regulatory T cell development Tolerance Mechanisms Mediated by Regulatory T cells Immune homeostasis and tolerance versus pathogenesis in the gut Microbial flora Pattern recognition receptors Dendritic cells Pro-inflammatory cytokines Regulatory T cells 33 vi
8 5.6 T cell lymphopenia and lymphopenia induced proliferation Forward genetics Family of GTPase of Immunity Associated Proteins Identification of Gimap GTPases in plants and vertebrates Genomic organization of Gimap family Structural Features of Gimap proteins Cellular localization of Gimap Gimap5 in autoimmunity Gimap5 and T lymphocyte development Gimap5 in peripheral T cell survival Gimap5 in T cell homeostasis and immune tolerance Summary 50 References 52 Chapter II: Loss of T Cell and B Cell Quiescence Precedes the Onset of Microbial Flora-Dependent Wasting Disease and Intestinal Inflammation in Gimap5-Deficient Mice 78 Abstract 79 Introduction 80 Materials and Methods 84 Results 89 Discussion 102 References 108 Figure Legends 114 Figures 118 Supplementary Figure Legends 126 Supplementary Figures 128 vii
9 Chapter III: Loss of Immunological Tolerance in Gimap5-Deficient Mice is Associated with Loss of Foxo in CD4 + T Cells 135 Abstract 136 Introduction 137 Materials and Methods 140 Results 144 Discussion 151 References 156 Figure Legends 161 Figures 164 Supplementary Figure Legends 169 Supplementary Figure 171 Chapter IV: TCR Signaling in Gimap5-Deficient CD4 + T Cells is Associated with an Abnormal Bioenergetic Profile and Reduced Survival 175 Abstract 176 Introduction 177 Materials and Methods 180 Results 183 Discussion 189 References 194 Figure Legends 198 Figures 200 Chapter V: Summary and Discussion Summary Gimap5 and autoimmunity Clinical implications Role of Gimap5 in lymphocyte development and homeostasis Molecular function of Gimap5 222 Summary Figures 228 References 229 viii
10 List of abbreviations: APC ATP DC EAE ENU Foxo FoxP3 Gimap IBD IL IFN-γ LIP MHC NK OVA PRR RAG Stat TCR TNF-α TGF-β Th TLR Treg Antigen Presenting Cell Adenosine triphosphate Dendritic Cell Experimental Autoimmune Encephalomyelitis N-ethyl-N-nitrosourea Forkhead box, subgroup O Forkhead box, P3 GTPase of Immunity Associated Protein Inflammatory Bowel Disease Interleukin Interferon gamma Lymphopenia induced proliferation Major Histocompatibility Complex Natural Killer Ovalbumin Pattern Recognition Receptor Recombination Activating Gene Signal transducer and activator of transcription T cell receptor Tumor Necrosis Factor alpha Transforming Growth Factor beta T helper Toll-like Receptor Regulatory T cell ix
11 Chapter I Introduction 1
12 1. Thesis Introduction Innate and adaptive arms of the immune system have evolved to discriminate between self and non-self. This system is crucial to protect the host from pathogens. However, the immune system also carries the potential to damage the host and requires strict regulation. Therefore, tolerance mechanisms have developed to counter-balance immune responses and ensure that they do not react against self. Central and peripheral tolerance mechanisms tightly regulate cells of the adaptive immune system throughout their peripheral lifespan. Breakdown of immune tolerance leads to autoimmune disorders. Therefore, understanding how immune tolerance is maintained is critical. In this thesis, we study a novel N-ethyl-nitrosurea germline mouse model that spontaneously develops colitis as a result of abnormalities in peripheral T cell survival and immune tolerance. In this introduction, we will discuss the processes that control T cell development, homeostasis and immune tolerance. 2. T cell development During T cell development, a large number of T cells with a wide repertoire of antigen specificity are generated. This clonal diversity of T cells allows the adaptive immune system to properly respond to a broad variety of foreign antigens. Generation of such diversity is achieved by the genomic rearrangement of the VDJ genes in the T cell receptor (TCR) loci. To ensure that only thymocytes with successful TCR rearrangements differentiate into mature T cells, several checkpoints exist during T cell development (1). 2
13 T cell development begins with the migration of progenitor cells from bone marrow to thymus. At this stage, the transcription factor Notch-1 promotes the commitment of progenitors into the T cell lineage (2). These progenitor cells are specified by the expression of interleukin-7 Receptor α (IL-7Rα), c-kit (receptor for stem cell factor) and the adhesion molecule CD44 and can give rise to all T cell lineages (3). In the thymus, thymocytes can be subdivided into four subsets based on the expression of CD4 and CD8 co-receptors. Among them, CD4 - CD8 - double negative (DN) thymocytes represent the earlier stage in the development. DN thymocytes are also subdivided into four sequential stages based on the expression of CD44 and Interleukin-2 receptor α (CD25). CD44 + CD25 - DN1 stage represents the initial stage and is followed by the CD44 + CD25 + DN2 stage (4). Although CD25 serves as developmental marker for the DN thymocyte stages, the role of Interleukin-2 (IL-2) at these stages is not well established (5, 6). Survival and expansion of cells at DN1 and DN2 stages are largely regulated by SCF and IL-7 signals (7). Next, the cells down-regulate CD44 expression and proceed into the CD44 - CD25 + DN3 stage. At this stage, thymocytes express RAG1 and RAG2 proteins and subsequently recombine their TCRβ genes (8). At the DN3 stage, thymocytes express pre-tcrα which assembles with the productive TCRβ chains and forms the pre-tcr complex. Expression of pre-tcr represents a critical checkpoint in αβ T cell differentiation known as the β-selection. Thymocytes that fail to develop a functional pre-tcr cannot interact with MHC proteins on the surface of cortical epithelial cells and die by apoptosis. Thymocytes that make productive rearrangements in their TCRβ gene proceed to the CD44 - CD25 - DN4 stage 3
14 (9). Pre-TCR signals promote the commitment of thymocytes into αβ T cell lineage and inhibit the entry to the γδ T cell lineage (10). Moreover, pre-tcr signals provide cell survival, inhibit further TCRβ gene recombination and induce proliferation of DN3 and DN4 cells (8, 11). Following proliferation at the DN4 stage, thymocytes upregulate CD4 and CD8 co-receptors and become small resting double positive (DP) thymocytes. At this stage, DP thymocytes recombine their TCRα locus by a second round of RAG1/RAG2 expression and start to express TCRαβ heterodimers. DP thymocytes undergo a second checkpoint known as positive selection. TCRαβ dimers expressed on the surface of the DP cells encounter the self-peptide/mhc complexes presented on the cortical epithelial cells. Thymocytes expressing TCRs that interact with the selfpeptide/mhc with low avidity are signaled for survival and are positively selected (12). Thymocytes whose TCRs strongly bind to the self-peptide/mhc are eliminated by negative selection, whereas thymocytes that cannot successfully rearrange their TCR do not receive survival signals and die by neglect (13). Thus, the binding avidity and signaling strength of TCR is critical for the selection process in the thymic development. Negative selection of DP thymocytes is achieved by programmed cell death. Strong TCR engagement increases the expression of pro-apoptotic protein Bim which induces cell death through Bax and Bak dependent mitochondrial apoptotic pathway. Deficiency of Bim has been shown to impair the deletion of self-reactive thymocytes (14). Similarly, mice deficient for both Bax and Bak pro-apoptotic proteins showed impaired negative selection (15). Moreover, transgenic expression of anti-apoptotic protein Bcl-2 partially rescues the self-peptide specific cells from negative selection; 4
15 however these thymocytes do not mature into functional T cells (16). Strong TCR binding also induces expression of FAS ligand (FASL). FASL is a cell surface molecule that triggers cell death upon FAS-FASL ligation through the extrinsic apoptosis pathway. Although genetic defects in FAS and FASL are associated with autoimmune diseases, the role of FAS signaling in negative selection is not clear (17-19). In addition, Nur77 has been demonstrated to be required for negative selection as its inhibition led to impaired clonal deletion of autoreactive thymocytes (20). Therefore, the negative selection of thymocytes is regulated by the strength of TCR signaling and executed by apoptotic pathways. Positively selected thymocytes that weakly recognize either MHC I or MHC II molecules differentiate into CD8 single positive (SP) and CD4SP thymocytes respectively. CD4 versus CD8 lineage commitment of DP thymocytes is regulated by a number of transcription factors. Activation of transcription factor Runx3 is required for the commitment of the DP thymocytes into CD8 lineage (21), while Th-POK is required for the CD4 lineage commitment (22, 23). Following the positive selection, SP cells undergo additional maturation steps in the thymus. Positively selected thymocytes down-regulate surface markers CD69 and CD24, and upregulate β 7 integrin, CD62L and the non-classical MHC protein Qa-2 (24). Transcription factor KLF2 regulates thymocyte emigration and peripheral trafficking of T cells by inducing the expression of cell adhesion molecules sphingosine-1-phosphate (S1P) receptor S1PR1, CD62L and β 7 integrin (25). Recently identified protein families such as schlafen proteins and Gimap family of GTPases might have a role in the functional maturation of SP cells in the thymus (26, 27). Thymocyte maturation events 5
16 after positive selection are still incompletely understood. In summary, thymic development involves several tightly controlled checkpoints and a network of transcription factors which ultimately generates functionally mature T lymphocytes with diverse antigen specificity. 3. Peripheral Homeostasis of T cells The T cell population size in peripheral lymphoid organs is controlled by several mechanisms such as thymic output, survival and proliferation. These mechanisms maintain a consistent pool of T lymphocytes in order to promptly respond to infections. Regulation of T cell homeostatic mechanisms is critical as their failure can lead to lymphopenia. In the following sections, we will review T cell homeostasis, lymphopenia and effector T cell differentiation. 3.1 Peripheral survival and homeostatic proliferation Maintenance of circulating naïve T cells is controlled by homeostatic mechanisms. Survival signals for naïve T cells are provided by TCR-self peptide/mhc interactions and IL-7 signaling (28, 29). TCR-MHC interactions are essential for the survival of naïve T cells as demonstrated by the impaired survival of naïve CD8 + T cells when adoptively transferred into MHC Class I deficient mice (30). Moreover, induced deletion of TCRα gene resulted in a rapid decline of naïve CD8 + T cell numbers, while CD4 + T cells persisted relatively longer (31). Importantly, the downstream targets of TCR signaling required for T cell survival are currently not well understood. On the other hand, the downstream targets of IL-7 that promote the survival of peripheral T cells are much better defined. Specifically, IL-7 inhibits the mitochondrial 6
17 apoptotic pathway by inducing expression of anti-apoptotic proteins Bcl-2 and Mcl-1 (32, 33). Bcl-2 has been shown to regulate the maintenance of naïve T cells by antagonizing pro-apoptotic protein Bim (34). Thus, the peripheral survival of peripheral T cell is regulated by factors controlling the apoptotic pathways. In addition to maintaining survival, IL-7 and self-peptide/mhc also drive homeostatic proliferation of naïve T cells. IL-7 and TCR self-peptide/mhc interactions drive a slow rate of proliferation in order to maintain T cell homeostasis. In lymphoid organs, IL-7 is produced only in trace amounts resulting in a competition between T cells. It has been reported that T cells down-regulate the surface expression of IL7Rα after receiving IL-7 signals in order to prevent excessive consumption of the cytokine. These mechanisms control basal state survival and proliferation of T cells, and maintain homeostasis (35). In summary, basal homeostatic mechanisms control the size of peripheral T cell populations ensuring that a proper adaptive immune response can be mounted against infections. These mechanisms also limit the expansion of T cells in order to prevent immunopathology. 3.2 Activation and effector differentiation of CD4 + T cells The activation of naïve circulating CD4 + T cells is elicited by recognition of specific antigen/mhc class II complexes presented by antigen presenting cells (APC). APCs also provide co-stimulatory signals which trigger T cell activation and expansion. Activated CD4 + T cells secrete IL-2 which in turn induces their clonal expansion in an autocrine fashion (36). 7
18 During an immune response, naïve CD4 + T cells can differentiate into different types of effector cells depending on the nature of the pathogen. The best described effector CD4 + T cell subsets are the T helper 1 (Th1), Th2 and Th17 subsets (37, 38). These differentiated helper T cell subsets mediate immune responses against diverse pathogens via distinct mechanisms and elicit different cytokine production profiles. Th1 responses are generated against intracellular pathogens. IL-12 produced by macrophages and dendritic cells upon engulfment of pathogens such as Listeria monocytogenes induces the differentiation of activated CD4 + T cells into Th1 cells (39). Then, the Th1 cells produce interferon- which activates macrophages and enhances the killing of ingested bacteria. Th1 differentiation involves the coordinated action of TCR signaling, IL-12 and IFN- cytokines. IL-12 secreted by activated macrophages and DCs induces the production IFN- by CD4 + T cells through STAT4 (40). Moreover, NK and NKT cells are significant sources of IFN- during infections (41, 42). IFN- signaling in CD4 + T cells induces the expression of T-bet via STAT1 signaling (43). Transcription factor T-bet further enhances IFN- production creating an amplification loop for Th1 differentiation. T-bet represents the lineage specific transcription factor for Th1 cells (44). Th2 cells mediate the effector functions against helminthic parasites. Th2 responses are induced in response to allergens as well. Th2 differentiated cells are characterized by production of cytokines IL-4, IL-5, IL-13 and IL-10 (45). Th2 immune responses involve IgE production by B cells and subsequent Mast cell activation. Moreover, IL-5 produced by Th2 cells induces eosinophil activation leading to expulsion 8
19 of helminths. Differentiation of Th2 cells requires IL-4 signaling (46, 47). IL-4 induces Gata-3 expression through STAT6. Ablation of either STAT6 or Gata-3 prevents Th2 cytokine responses. Transcription factor Gata-3 acts as a lineage marker and enhances Th2 differentiation by further inducing IL-4 production (48, 49). Differentiation of Th1 and Th2 lineages are reciprocally regulated. IFN- andil-12 inhibits Th2 lineage commitment, whereas IL-4 inhibits Th1 polarization (50, 51). Moreover, Gata-3 has been shown to suppress Th1 lineage commitment by downregulating STAT4 (52). Furthermore, T-bet represses the Th2 differentiation by inactivating Gata-3 through a tyrosine kinase mediated interaction (53). More recently discovered effector T cell lineage is the Th17 subset. Th17 cells primarily produce IL-17 and recruit neutrophils to the site of inflammation. Th17 cells have been implicated in inflammatory diseases in humans and animal models such as Crohn s disease and experimental autoimmune encephalomyelitis (EAE). (54, 55). Th17 cells represent a distinct lineage as their differentiation is inhibited by IL-4 and IFN- (38). Indeed, TGF-β and IL-6 have been shown to induce Th17 polarization (56). Also, IL-21 and IL-23 signals further enhance Th17 differentiation (57, 58). In addition, Th17 lineage is specified by the key transcription factor ROR t. ROR t is required for the differentiation and effector function of the Th17 lineage in response to IL-6 and TGF-β (59). In summary, effector CD4 + T cell function is essential for generating appropriate antigen-specific immune responses against diverse pathogens. The pathogen-specific 9
20 responses generated by effector T cells require distinct differentiation programs, driven by unique cytokines and transcription factors. 3.3 Memory and memory phenotype T cells As an infection is cleared, the host acquires immunological memory that provides long term immunity. Formation of immunological memory involves the generation of memory T and B cells (60). Following the clearance of microbes, a small percentage of T cells survive the contraction phase and form the memory T cell pool. These long lived memory T cells survive for the lifetime of the host and rapidly respond to secondary infections. Memory T cells generate a more qualitative response than the naïve T cells. Memory T cells have the characteristic of high CD44 surface expression and composed of effector and central memory T cell subsets which have low and high CD62L expressions, respectively (61, 62). In addition to memory T cells that are generated in response to foreign antigens, there is another subset of T cells called memory phenotype T cells that express similar markers to memory T cells (63). These memory phenotype T cells do not necessarily arise in response to foreign antigens as they are found in uninfected or unimmunized mice (64, 65). Although the formation of memory phenotype cells has been previously thought to be driven by foreign antigens, such memory phenotype cells can still arise in germ free and food antigen free conditions (66). Therefore, the presence of exogenous antigens is not a requirement for the generation of memory phenotype T cells. Moreover, adoptive transfer experiments using ovalbumin specific transgenic T cells showed that naïve T cells proliferate in response to self-peptide/mhc molecules in the 10
21 absence of antigenic stimulation and acquire a CD44 high phenotype (67). Thus, under basal conditions, the formation of memory phenotype cells depends on TCR selfpeptide/mhc interactions, but does not rely on the presentation of exogenous antigens. 3.4 T cell quiescence There are several mechanisms to keep cells in a quiescent state and to maintain T cell homeostasis under basal conditions. These mechanisms control cell division, activation and differentiation of T cells. One critical component is KLF2, a transcription factor that controls T cell quiescence by negatively regulating the c-myc dependent transcription (68). C-myc is a proto-oncogene which facilitates cell cycle progression and promotes cellular growth and metabolism (69, 70). Consistently, KLF2 deficient T cells showed increased proliferation in vivo (68). Further, it has been shown that, T cells in KLF2 deficient mice were skewed to an activated phenotype; CD62L low and CD69 high suggesting an important role for KLF2 in T cell quiescence (25). Another important transcription factor for T cell homeostasis is Foxo3. Foxo3 limits cell division by upregulating cell cycle inhibitor p27 in quiescent cells (71). Consistent with this, Foxo3 deficient T cells showed a hyper-proliferative phenotype (72). Foxo1, another member of the Foxo transcription factor family, has also been demonstrated to regulate T cell homeostasis. Foxo1 regulates homing and survival of naïve T cells by inducing the expression of IL-7Rα, CD62L and KLF2. Foxo1 deficient mice displayed increased proportion of CD44 high CD62 low memory phenotype T cells (73). In conclusion, T cell quiescence is important for the maintenance of T cell homeostasis and immune tolerance. Yet, the mechanisms regulating T cell quiescence are not completely identified. 11
22 4. Immune Tolerance mechanisms The major function of the immune system is to protect an individual against pathogens. The immune system has developed strategies to eliminate foreign antigens while preventing damage to self tissues and maintaining the balance between immunity and tolerance. In order to maintain tolerance, the immune system has evolved to discriminate self-antigens from non-self and to keep activated immune cells under control. Immune tolerance mechanisms act during both immune responses and steady state conditions, and ensure that destructive immune reactions are not directed to the host. Historically, the first experimental demonstration of immunological tolerance has emerged from transplant studies (74). Normally, transplant of skin grafts from one strain of mice is rejected by the other strain due to the incompatibility of the MHC alleles. Medawar and colleagues (74) showed that transfer of allogeneic blood cells during neonatal life prevented rejection of skin grafts from the same donor in the adult recipient mice. These results indicated that the encounter of developing neonatal immune system with foreign antigens induces immune tolerance to those antigens. Maintenance of immune tolerance involves diverse mechanisms including i) elimination of self-reactive lymphocytes by central tolerance of the adaptive immune system, ii) induction of anergy in the absence of costimulatory and innate immune signals, iii) unresponsiveness to antigens in the non-immunogenic form, iv) suppression of immune responses by specialized regulatory lymphocytes, v) inhibition of lymphocyte activation by counter regulatory signaling mechanisms and vi) induction of immune 12
23 unresponsiveness by pharmacological agents for clinical transplantation (75). Failure of these immune tolerance mechanisms may result in autoimmune diseases. Understanding the mechanisms of immune tolerance and how the breakdown of these mechanisms leads to autoimmunity is one of the important challenges in the field of immunology. In the following sections mechanisms of immune tolerance will be discussed in detail. 4.1 Central Tolerance of T cells During T cell development in the thymus, VDJ recombination randomly generates an enormous number of TCRs with different specificities for both self and non-self antigens. Developing thymocytes which recognize the self-peptide/mhc complex weakly are positively selected, whereas cells with high avidity binding are deleted by negative selection. Thus, negative selection eliminates self-reactive T cell clones in order to prevent immune responses to self-antigens in the tissues and organs of the body. The theory of clonal selection was first proposed by Burnet in 1959 and has been studied extensively to date (76). Clonal deletion of thymocytes during development has been first demonstrated by Kappler et al (77). Different strains of mice express different endogenous superantigens, these superantigens associate with MHC II molecules and bind strongly to specific Vβ clones of TCR. T cells specific for an endogenous provirus superantigen were found to be present among the DP cells in the thymic cortex whereas they were absent in the peripheral pool of lymphocytes (77). In another model, negative selection of self-reactive thymocytes has been demonstrated by generating transgenic mice 13
24 whose T cells express an αβ-tcr specific for male minor histocompatibility antigen H-Y (78). H-Y specific T cells were deleted in the male transgenic mice whereas they were positively selected in females. The frequency of CD4 + CD8 + DP cells in the thymus of males was also severely reduced suggesting that the self-reactive thymocytes were deleted at the stage. Although the molecular pathways involved in clonal deletion of T cells are still incompletely understood, there is strong evidence that the strength of TCR signaling is important for negative selection. Mutations that affect TCR signaling pathway also affect the threshold for T cell selection. For example, the tyrosine kinase ZAP70 downstream of TCR is critical for the negative selection of thymocytes. A hypomorphic mutant of ZAP70 caused inefficient TCR signaling and allowed autoreactive T cells to escape from negative selection leading to rheumatoid arthritis (79). In addition, other molecules downstream of the TCR such as the adaptor protein GRB2 and MAP kinases Erk, JNK and p38 are also involved in negative selection of T cells (80). Therefore, the downstream TCR signaling pathways are critical for maintaining the threshold for negative selection of self-reactive thymocytes. One of the major questions in the concept of central tolerance is how the developing T cells specific for tissue restricted antigens are clonally deleted in the thymus. Many antigens in the organism are spatially confined to peripheral tissues or temporally expressed at certain stages development and were previously assumed to be inaccessible to thymocytes during negative selection (81). One model posed to explain this issue was the promiscuous gene expression of tissue restricted antigens by medullary thymic epithelial cells (mtecs). mtecs have been demonstrated to 14
25 randomly express tissue restricted antigens such as insulin and lactalbumin, thus presenting a repertoire of self-peptides to developing thymocytes. This process requires the zinc finger transcription factor autoimmune regulator (AIRE) (82). Interestingly, autosomal recessive nonsense and frameshift mutations in AIRE resulted in development of autoimmune polyendocrinopathy candidiasis ectodermal dystrophy (APECED) syndrome in humans (83, 84). Deletion of Aire gene in mice led to altered TCR repertoire of peripheral T cells, multi-organ lymphocytic infiltration and consequent autoimmune disease (85). Moreover, another study investigated the role of AIRE in negative selection by crossing Aire -/- mice with a double transgenic model. In this model, developing TCR transgenic CD4 + T cells expressed a single αβtcr clone that is specific for a dominant peptide of hen egg lysozyme (HEL) while another transgene induced the expression of HEL under the insulin promoter. AIRE-deficiency almost completely abolished the negative selection of pancreatic islet reactive T cells in the thymus in this transgenic model (86).Thus, AIRE is a major player in central tolerance as it regulates the tissue-specific repertoire of self-peptide expression, thereby controls negative selection of T cells. 4.2 Peripheral T cell Tolerance Although most self-reactive T cells are removed by negative selection, some autoreactive T cells escape central tolerance and persist in the periphery. It is therefore important that additional mechanisms exist in order to control peripheral T cell tolerance. These mechanisms prevent immune responses against self-antigens and prevent excessive immune responses to foreign antigens and pathogens. Peripheral T 15
26 cell tolerance is maintained by anergy, inhibitory costimulatory signals and suppressor T cells, all of which will be reviewed in more detail in the following sections T cell Anergy Anergy is the functional unresponsive state in which T cells reside when stimulated with cognate antigens in the absence of appropriate costimulation signals (87, 88). Initial studies demonstrated that CD4 + T cells stimulated with chemically fixed APCs coupled with peptide antigen failed to proliferate (89). Moreover, stimulation of T cells with anti-cd3 antibody alone rapidly induced anergy exemplified by a failure of T cells to undergo proliferation and produce interleukin-2. Anergy was prevented by the addition of activated APCs into the culture. Therefore it was proposed that, in addition to TCR signaling, T cells require a second signal for efficient activation and proliferation (90). Later, it was demonstrated that B7 molecules expressed by activated APCs provide the second signal to T cells and efficiently induce cytokine secretion and T cell proliferation (91). Moreover, T cells became anergic upon stimulation with APCs when the B7 molecules were blocked by antibodies (92). These studies established the role of costimulation in T cell activation and defined a tolerance mechanism in which T cell anergy is induced upon cognate antigen recognition in the absence of costimulation. T cell anergy can be induced by self-antigens in the peripheral tissues. To demonstrate this phenomenon, transgenic mice expressing LCMV glycoprotein in the pancreatic beta cells have been generated and bred with transgenic mice expressing TCR specific for LCMV glycoprotein. Peripheral self-reactive T cells developed in these mice remained functionally unresponsive to the viral glycoprotein indicating that T cells 16
27 became anergic in the absence of appropriate costimulation during presentation of antigens. Interestingly, infection of these mice with LCMV resulted in the breakdown of tolerance and induced immune responses against pancreatic beta cells (93). T cell anergy can be induced against foreign antigens depending on the delivery method of the antigen. In one study, transgenic T cells specific for chicken ovalbumin (OVA) were adoptively transferred to normal syngenic mice and the cognate peptide antigen was delivered through different routes. While subcutaneous injection of antigen in combination with adjuvant induced T cell proliferation and effector function, soluble antigen injected intravenously resulted in reduced proliferation and impaired migration to lymph node follicles. A portion of these T cells remained viable but became unresponsive to the cognate antigen (94). These results demonstrate that T cells exposed to soluble antigen in the absence of costimulation become anergic. Anergy can also be induced by the biochemical tuning of the signaling cascades downstream of TCR. CBL-b is a ubiquitin ligase that regulates the proteolytic degradation of TCR signaling components including protein kinase C-θ, phospholipase- C 1 and ZAP-70 (95). It has been demonstrated that Cbl-b deficient T cells did not require CD28 costimulation for TCR-induced proliferation and interleukin-2 production (96). Cbl-b deficient T cells showed resistance to antigen-specific induction of anergy. Moreover, when rechallenged with tolerizing soluble antigen, Cbl-b deficient mice showed rapid lethality due to autoimmunity (97). These findings demonstrate the significance of negative regulation of TCR signaling in order to induce anergy and immune tolerance. 17
28 4.2.2 Inhibitory Costimulatory Molecules Although costimulation is essential for efficient T cell proliferation and effector functions, inhibition of costimulatory signals is just as critical in order to prevent heightened immune responses which potentially can trigger autoimmunity. A key molecule involved in the inhibition of costimulation is CTLA-4, a member of the CD28 family that competes with CD28 for binding to B7 costimulatory molecules on antigen presenting cells. In contrast to CD28, CTLA-4 delivers inhibitory signals which negatively regulate the TCR-induced signals (88). CTLA-4 deficient mice showed lymphoproliferative disease and displayed autoimmune lesions with early lethality (98, 99). Moreover, in response to tolerogenic stimuli, CTLA-4 deficient T cells did not become anergic as demonstrated by increased entry to cell cycle and heightened proliferation (100). In this study, CTLA-4 deficient T cells expressing transgenic TCRs specific for OVA were adoptively transferred into syngenic mice expressing OVA in beta islet cells. CTLA-4 deficient T cells induced severe insulitis and diabetes in the recipient mice without prior immunization with adjuvant+ova. These results indicate that CTLA-4 prevents pathogenic autoimmunity and maintains peripheral T cell tolerance. PD-1 is another inhibitory costimulatory molecule which recognizes PDL1 and PDL2 molecules. Interaction of PD-1 with its ligands negatively regulates T cell activation. PDL1 is expressed on several tissues including hematopoietic cells and pancreatic islets, heart and intestines whereas PDL2 is expressed on DCs and macrophages. Expression of both PDL1 and PDL2 on antigen presenting cells increases upon activation (101, 102). PDL1 expressed by DCs has been shown to be involved in the induction and maintenance of T cell anergy (103). Mice deficient for PD- 18
29 1 developed autoimmune disease including autoimmune dilated cardiomyopathy and lupus-like autoimmune disease in different mouse strains. Moreover, both genetic deletion and blocking PD1 function in non-obese diabetic mice accelerated the onset of diabetes (104, 105). Thus, PD-1 is an important molecule that negatively regulates costimulatory signals and contributes to peripheral tolerance Regulatory T cells Another level of peripheral T cell tolerance is achieved by regulatory T cells (Treg). Regulatory T cells represent a subset of CD4 + T cells with immune suppressive properties and are defined by the expression of lineage specific marker Foxp3. Tregs regulate the effector functions of other T lymphocytes and limit inflammatory responses. Treg-mediated suppression plays a critical role in the maintenance of immunological tolerance under steady state conditions and during inflammatory responses as demonstrated in animal models and humans (106). In the following sections we will discuss the past and recent findings on Tregs and Treg-mediated suppression mechanisms Identification of Regulatory T cells The idea that a T cell population with immune suppressive properties exists was first proposed in early 70s (107). However, the theory remained controversial for some time due to difficulties in identifying a lineage marker that defines an endogenous suppressive cell population. A few decades later, studies supporting the existence of suppressive T cells reemerged. Studies with organ transplant, autoimmune encephalomyelitis and organ inflammation pointed out a population of T cells with 19
30 immune regulatory functions which prevented pathology in these experimental models ( ). Subsequently, it was demonstrated that CD4 + T cells that are low for CD45RB expression exhibited immunosuppressive properties as adoptive transfer of CD45RB depleted T cells resulted in the development of spontaneous autoimmunity (110). However, the low expression of CD45RB was not unique to regulatory T cells as T cells down-regulate this marker when activated. Moreover, adoptive transfer of splenocytes depleted of CD25 + CD4 + T cells into athymic nude mice caused severe autoimmunity and the reconstitution of micce with CD25 + CD4 + T cells prevented disease (112). Therefore, it was suggested that CD25 - CD4 + T self-reactive cells can cause autoimmunity in the absence of CD25 + CD4 + T regulatory T cells and CD25 + CD4 + T cells actively maintain self-tolerance. However, CD25 did not serve as a lineage specific marker for regulatory T cells since all T cells temporarily express CD25 upon TCR stimulation (113). A major breakthrough came with the discovery of the genetic mutation underlying the scurfy phenotype. Historically, the scurfy mutation was a recessive X chromosome linked mutation that caused early mortality (114). It was not until the early 90s, that hematological abnormalities and auto-immune responses were identified in scurfy mutants (115). Specifically, scurfy mice presented lymphoproliferative disease and multi-organ autoimmunity that was mediated by CD4 + T cells (116, 117). The causative mutation was ultimately identified in the Foxp3 gene encoding a Forkhead Box transcription factor (118). Moreover, it was demonstrated that transgenic restoration of FoxP3 expression in CD4 + T cells prevented autoimmunity in scurfy mice. Remarkably, mutations in human Foxp3 gene have been found to cause the immune dysregulation, 20
31 polyendocrinopathy, enteropathy, X-linked syndrome (IPEX) (119, 120). Similarities between IPEX and scurfy phenotype revealed the significance of FoxP3 in controlling immunological tolerance. FoxP3 controls the development of regulatory T cells and is a specific marker for this lineage (121). Most of the CD25 + CD4 + T cells express FoxP3 in the peripheral lymphoid organs and ectopic expression of FoxP3 converts CD25 - CD4 + T cells to CD25 + cells having suppressive function. In addition, adoptive transfer of regulatory CD25+CD4 + T cells prevented the autoimmune disease in FoxP3 deficient mice and scurfy mutants ( ). Moreover, induced deletion of Foxp3 gene elicited systemic autoimmunity due to inflammatory activity of self-reactive effector T cells (125). These studies established the critical role of FoxP3 in the development of regulatory T cells as a suppressive T cell lineage that actively maintains immunological tolerance Regulatory T cell development Natural regulatory T (ntreg) cells are produced during T cell development depending on their TCR binding avidity to self-peptide/mhc. Thymocytes which strongly bind to self-peptide/mhc and escape the negative selection develop into regulatory T cells. It is demonstrated that high intensity TCR signaling through agonist self-peptide is required for the differentiation of ntregs (126). Moreover, using TCR transgenic models, it is found that ntregs predominantly express endogenously recombined TCR alpha chains, whereas ntreg cells did not develop in RAG deficient TCR transgenic mice confirming that ntreg differentiation requires interactions between self-peptide/mhc and TCR (127, 128). The importance of ntreg cells for self-tolerance has been 21
32 demonstrated by adoptive transfer of CD4 + CD25 + depleted thymocytes into syngenic athymic nude mice which triggered autoimmunity (128). Thus, in addition to negative selection, generation of CD4 + CD25 + FoxP3 + ntregs is another key function of thymus for maintaining immune tolerance. Regulatory T cells do not only originate from the thymus, naïve CD4 + CD25 - T cells can also acquire a Treg phenotype in the periphery (129, 130). TGF-β1 is required for the peripheral generation of FoxP3 + CD4 + Tregs but not for the thymic development. Moreover, TGF-β1 has been shown to be essential for peripheral homeostasis and suppressive function of Tregs (131). TGF-β and TCR signaling in the presence of IL-2 converts naïve CD4 + CD25 - T cells into FoxP3 + CD4 + CD25 + regulatory T cells. It is demonstrated that these induced Treg (itreg) cells suppressed the proliferation of antigen-specific CD4 + CD25 - T cells and prevented pathogenesis in vivo (132). Moreover, it has been shown that itreg conversion occurs in response to foreign antigens. Administration of foreign antigens generated FoxP3 + Tregs in TCR transgenic Rag deficient mice which are normally devoid of ntregs ( ). In summary, conversion of peripheral CD4 + T cells into itregs is another essential mechanism to generate suppressive cells for controlling immune responses. Differentiation of regulatory T cells involves the transcriptional activation of Foxp3 gene. TCR-induced NF B signaling induces FoxP3 expression. Mice deficient for components of NF B pathway Bcl-10, Carma1 and PKC-θ showed impaired natural Treg development in the thymus ( ). Moreover deletion of NF B subunit c-rel impaired both thymic and peripheral Treg development (139). TCR-induced NFAT signaling also induces FoxP3 expression (140). Additionally, NFAT and FoxP3 together 22
33 control the Treg transcriptional program as they cooperatively induce Cd25 and Ctla4 genes and repress the transcription of Il2 (141). TGF-β signaling is also critical for the expression of FoxP3 during peripheral differentiation of Tregs (132, 142). SMAD2 and SMAD3 transcription factors downstream of TGF-β signaling have been shown to bind to FoxP3 promoter in a redundant fashion (143). In addition, IL-2 signaling has been shown to be critical for peripheral Treg generation. An in vitro Treg differentiation study suggested that IL-2 is essential for TGF-β mediated induction of Foxp3 (144). Moreover, Stat5 activation downstream of IL-2Rβ has been demonstrated to be critical for the induction of FoxP3 transcription during peripheral Treg development (145). Recently, Foxo1 and Foxo3 have been shown to promote both thymic and induced Treg development. Foxo transcription factors directly induce the expression of FoxP3 and CTLA-4. Moreover, in the absence of Foxo proteins, T cells showed impaired responsiveness to TGF-β signaling ( ). As Foxo transcription factors interact with SMAD3 and SMAD4 which are downstream of TGF-β signaling (149, 150), Foxo proteins may be involved in the regulation of TGF-β induced gene expression program in T cells. Several lines of evidence indicate that differentiation of induced Treg cells and Th17 cells are reciprocally regulated. Induced Treg differentiation requires TGF-β signaling, whereas Th17 differentiation requires both IL-6 and TGF-β. IL6 has been shown to inhibit Treg differentiation (57, 142). Moreover, TGF-β induced Foxp3 has also been shown to inhibit Th17 cell differentiation by antagonizing RORγt (151). Therefore, T helper cell lineage commitment involves complex regulation and plasticity, and is critical for maintaining the balance between tolerance and immunity. 23
34 Tolerance Mechanisms Mediated by Regulatory T cells What mechanisms regulatory T (Treg) cells utilize to maintain immunological tolerance and suppress effector T cells is a critical question. As a transcription factor, FoxP3 induces the expression of Ctla4 and Il2Ra genes. One mechanism proposed for Treg-mediated immunoregulatory function is the direct suppression of antigen presenting cells (APC). CTLA-4 interacts with CD80 and CD86 on DCs and downregulates expression of these costimulatory molecules leading to the inhibition of DC maturation (152). In mice, specific deletion of Ctla4 gene in Treg cells resulted in an impaired suppressive capacity by Tregs and led to systemic autoimmunity. Abrogation of CTLA-4 mediated suppression caused increased expression of CD80 and CD86 expression on dendritic cells (153). Therefore, CTLA-4 on Treg cells reduces the ability of APCs to stimulate naïve T cells through CD28 costimulatory molecules which in turn prevents their differentiation into effector T cells. A second mechanism by which Tregs are thought to exert immunosuppressive function is by limiting the availability of IL-2, a cytokine critically required for T cell proliferation. A study by Thornton and Shevach showed that Tregs inhibit IL-2 production by effector T cells through a contact dependent fashion (154). In a separate study, it was demonstrated that the high IL-2 receptor alpha, (CD25) expression on Tregs captures the available IL-2 away from T effector cells, thereby reducing their ability to proliferate. Cytokine deprivation by Tregs prevented effector T cell proliferation and induced apoptosis in both in vitro and in vivo models indicating the presence of a passive mode of suppression by Tregs (155). 24
35 Tregs also produce other surface molecules that can suppress effector T cell responses. Surface molecules, CD39 and CD73 are ectoenzymes that are highly expressed by Tregs (156). Enzymatic activities of these ectoenzymes together convert extracellular ATP into adenosine which is inhibitory to T cell proliferation and effector functions. As a result, the generation of adenosine by Treg cells and adenosine signaling through A2A adenosine receptor on effector T cells serves as a suppressive signal mediated by Tregs (157). Another surface molecule expressed on Tregs is the CD4 homolog LAG3. LAG3 binds strongly to MHC II molecules on DCs and inhibits DC maturation and costimulatory capacity, and consequently inhibit T cell responses (158). Tregs also exert their suppressive function by secreting cytokines with immune regulatory properties such as TGF-β, IL-10 and IL-35 (156). IL-10 is an immunoregulatory cytokine produced by Tregs and several other cell types (159). Treg specific deletion of IL-10 showed that IL-10 production by Tregs was essential for the control of immune responses at mucosal interfaces such as the colon and lungs (160). The role of IL-10 and TGF-β in Treg-mediated immune tolerance mechanisms and their involvement colitis will be reviewed in the following chapter. Finally, Tregs exhibit high expression of Ebi3 and IL12a/p35 which together form a heteromeric complex designated as IL-35 (161). IL-35 is an immunoregulatory cytokine secreted by Tregs. Tregs deficient for Ebi3 or IL12a/p35 failed to prevent inflammatory bowel disease in a T cell-mediated colitis model in mice (161). Moreover, ectopic expression of IL-35 suppressed proliferation of CD4 + CD25 - T cells, inhibited Th17 differentiation and attenuated disease in a collagen-induced arthritis model in vivo (162). In addition, Granzyme B mediated killing of effector T cells by Tregs was shown 25
36 to be another mode of immune suppression. Granzyme B deficient Tregs were less efficient in suppressive capacity (163). In summary, FoxP3 + regulatory CD4 + T cells mediate their immune suppressive function through several mechanisms. However, it needs to be determined how relevant each of these mechanisms for Treg-mediated suppression in vivo. 5. Immune homeostasis and tolerance versus pathogenesis in the gut As mentioned above, breakdown of mechanisms that maintain immunological tolerance can result in severe pathological conditions including autoimmune disease. Particularly in the gut, immunological tolerance is important for the maintenance of intestinal integrity and function. Breakdown of immunological tolerance in the gastrointestinal tract can lead to inflammatory bowel disease (IBD). Ulcerative colitis and Crohn s disease are the two major forms of IBD with a combined prevalence rate of more than 400 cases per 100,000 in the United States (164). These chronic disorders cause abdominal pain, diarrhea and gastrointestinal bleeding. Infiltration of inflammatory leukocytes into the intestinal tissues and tissue damage are observed in IBD. Crohn s disease can affect any location in the gastrointestinal tract, while inflammatory lesions are restricted to colon in ulcerative colitis. Crohn's disease is associated with Th1 and Th17 inflammatory cytokines, whereas Th2 cytokines are implicated in ulcerative colitis (165). Several hematopoietic and non-hematopoietic cells, different types of receptors and cytokines play roles in maintaining homeostasis and tolerance versus pathogenesis of IBD. 26
37 IBD is a complex autoimmune disease involving the interaction of multiple systems. Several genetic and environmental factors contribute to pathogenesis of IBD (166), yet the exact etiology is not clearly defined. Some genetic factors playing a role in predisposition to IBD have been identified. Variations in IL23R and NOD2 genes are associated with Crohn s disease (167, 168). Additional genetic polymorphisms with increased risk to Crohn s disease include ATG16L1, IRGM, IBD5 and TNFSF15 (166). Moreover, recent genome-wide association studies show that large numbers of genetic loci are associated with Crohn s disease and ulcerative colitis (169, 170). A significant fraction of these loci contain multiple candidate genes and their causative functional association with IBD remains to be identified. Intestinal immune system has a number of distinctive features which maintain the intestinal homeostasis. Intestinal epithelia form a physical barrier by a single cell layer of intestinal epithelial cells (IEC). Goblet cells secrete mucus which forms a thick layer of physical barrier. The tight junctions between the IECs prevent the leakage of luminal contents to the lamina propria. Moreover, IECs control the colonization of microflora by secreting various antimicrobial peptides including defensins, lysozymes and C-type lectins. In addition to epithelial cells, a large number of hematopoietic cells including APCs and lymphocytes are involved in specialized immune surveillance mechanisms in the gut. The gut associated lymphoid tissues (GALT) involve Peyer s patches, intestinal lymphoid follicles, lamina propria and mesenteric lymph nodes (171). Gastrointestinal tract represents a large surface where the mammalians are exposed to an infinite number of foreign antigens. The gastrointestinal tract encounters exogenous antigens derived from nutrients as well as from the microbiota colonized in 27
38 the intestines. In healthy individuals, the presence of these antigens does not cause inflammation. The intestinal immune system is regulated by complex mechanisms in order to maintain tolerance to foreign antigens while circumventing bacterial infections. These mechanisms involve innate and adaptive components of the immune system, and are strongly influenced by the composition of intestinal microflora. 5.1 Microbial flora Intestinal microflora affects immune responses during both healthy conditions and IBD. Recognition of bacterial components is important for immune homeostasis in the gut (172). Besides, there is also growing evidence that the interaction of intestinal bacteria with the immune system regulates the balance between effector and regulatory T cells. It has been demonstrated that Th17 development is impaired in germ free animals (59). Moreover, segmented filamentous bacteria (SFB) have been reported to induce Th17 cell development in the gut (173). Also recently, the immuno-modulatory molecule polysaccharide A of Bacteriodes fragilis has been shown to promote induced Treg differentiation in the gut (174). Thus, microflora can tip the balance in favor of suppressive or effector T cells. Pathogenesis of IBD involves the uncontrolled lymphocyte activation and elevated pro-inflammatory cytokine production directed against gut bacteria. Although typically not pathogenic, Helicobater, Clostridium and Enterococcus species can drive immune responses during inflammation (174). Clinical trials demonstrate that antibiotics treatment alleviates disease in some IBD patients (175). Moreover, germ free conditions 28
39 prevent intestinal inflammation in IL-10 deficient mice suggesting mciroflora can play a role in the pathogenesis of IBD (176). 5.2 Pattern recognition receptors Intestinal epithelial cells and hematopoietic cells sense intestinal bacteria through their pattern recognition receptors (PRRs) including TLRs and NOD like receptors (NLRs). Tonic PRR activation contributes to intestinal barrier function (177). In several mouse knock out models, deletion of MyD88 or various TLR molecules including Tlr2, Tlr4, Tlr5 and Tlr9 led to increased susceptibility to DSS induced colitis. TLR signals have been shown to be required for epithelial tissue repair and important for production of defensins in order to restrain bacterial colonization (178). Similarly, mice deficient for the NLR molecule NOD2 showed impaired production of defensins and enhanced colitis (179). Also, as mentioned above, single nucleotide polymorphisms in NOD2 showed strong association with Crohn s disease in humans (168, 180). Moreover, deficiency of NLRP3 and the deficiency of NLRP3 mediated inflammasome components also resulted in increased susceptibility to DSS induced colitis (181). These studies establish the protective role of tonic PRR signals in maintaining the intestinal barrier integrity. On the contrary, sustained aberrant PRR signals can induce chronic intestinal pathology (177). It has been demonstrated that ablation of MyD88 mediated TLR signaling prevented the spontaneous chronic colitis in the interleukin-10 deficient mice (172). Thus, PRR signals can be either protective or destructive during intestinal inflammation. 5.3 Dendritic cells Dendritic cells in the gut play a central role in maintaining immunological tolerance to foreign antigens and commensal bacteria, and also in generating immune 29
40 responses against pathogenic microorganisms. CD103 + DCs and CX3CR1 + DCs are the two phenotypically and functionally distinct major dendritic cells populations located in the intestinal tissues. CD103 + DCs located in the lamina propria can take up bacteria and antigens. Upon maturation, they migrate to the draining mesenteric lymph nodes and trigger adaptive immune responses by antigen presentation to T cells (182). CD103 + DCs promote gut homing of T cells by inducing the expression of CCR9 and α 4 β 7 surface receptors (183). These intestinal DCs also induce the IgA class switching of B cells (184). CD103 + DCs favor tolerance mechanisms. They induce the peripheral differentiation of FoxP3 + CD4 + T cells through TGF-β and retinoic acid dependent pathways (185). CD103 + DCs can also acquire pro-inflammatory properties. It has been demonstrated that during intestinal inflammation, CD103 + DCs produce IL-6 and also promote Th1 responses (186). The molecular mechanisms regulating CD103 + DC function have not been well understood. CX3CR1 + DCs are derived from monocytes and localize adjacent to the epithelial barrier. They have access to luminal bacteria and antigens by extending through the epithelial layer (187). However, CX3CR1 + DCs do not migrate to draining lymph nodes and are less potent to prime T cells compared to CD103 + DCs (182). CX3CR1 + DCs are maintained by the intestinal microflora and preferentially induce Th1 and Th17 responses. The role of CX3CR1 + DCs in colitis has been shown by a study using the T cell transfer-mediated colitis model. In this study, RAG -/- CX3CR1 -/- mice had reduced numbers of DCs in the mesenteric lymph nodes and showed resistance to T cellmediated colitis associated with reduced serum IFN- and IL-17 indicating the importance of the DC subset in the induction of intestinal inflammation(188). 30
41 5.4 Pro-inflammatory cytokines Secretion of pro-inflammatory cytokines largely by myeloid cells is one of the key factors contributing to the development of colitis. Increased levels of TNF-α is often observed along with inflammation in gastrointestinal organs. Moreover, mice engineered to overexpress TNF-α showed intestinal inflammatory disease (189). Anti-TNF-α antibodies have been successfully used for treatment of IBD for fifteen years (190, 191). However, a significant percentage of patients are refractory to TNF-α inhibitors suggesting the involvement of other pro-inflammatory cytokines (192). IL-6 is highly expressed in intestinal tissues in IBD. As mentioned before, IL-6 induces T cell differentiation in favor of inflammatory Th17 cells while inhibiting Tregs (142). Antibodymediated blockade of IL-6 signals ameliorated pathogenesis in several mouse colitis models (193, 194). Clinical trials with anti-il-6r antibody reported beneficial results in Crohn s disease patients (195). IL-12 has been linked to development of intestinal inflammation. IL-12 induces the production of IFN- by T cells, NK and NKT cells (40, 42). One study showed that anti-il12p40 antibody inhibits IFN- production by CD4 + T cells and attenuates disease development in a mouse colitis model (196). Moreover, T cell-intrinsic roles of IFN- and T-bet have been reported to contribute to colitis in mouse models and in Crohn s disease suggesting a Th1-mediated mechanism for pathogenesis of IBD (197, 198). However, neutralization of IFN- gave little or no beneficial results in clinical trials with Crohn s disease patients (195). 31
42 The p40 subunit of IL-12 is shared by IL-23 (199). IL-23, composed of p19 and IL-12p40 subunits, is another pro-inflammatory cytokine implicated in IBD. In Crohn s disease patients, elevated expression of IL-23 by intestinal CD14 + macrophages has been observed (200). Moreover, as mentioned above, polymorphisms in IL-23 receptor gene (IL23R) showed strong associations with ulcerative colitis and Crohn s disease in a genome-wide association study (167). Furthermore, IL-23 has been shown to be essential for T cell-mediated colitis in mice (201). IL-23 promotes pathogenic T cell responses in colitis by inducing T cell proliferation, enhancing IL-17 production, repressing Treg differentiation and inducing the appearance of IFN- + IL-17 + CD4 + T cells (202). Remarkably, IFN- and IL-17 double positive CD4 + T cells are also observed in the inflamed tissue of Crohn s disease patients (203). These findings indicate that IL- 23 has a critical role in IBD pathogenesis. Recent clinical trials demonstrated that neutralization of IL-23 showed beneficial effects in Crohn s disease patients especially patients that did not respond to anti TNF-a treatment (204). Finally, Interleukin-17 and Th17 cells have been implicated in IBD. Elevated expression of IL-17 is observed in the intestinal tissues of Crohn s disease and ulcerative colitis patients (205). A small frequency of IL-17 + CD4 + T cells reside in the colonic mucosa of healthy individuals, however their numbers significantly increase in Crohn s disease patients (206). In the mouse DSS induced colitis model, deletion of IL- 17A resulted in increased severity of colitis, whereas deletion of IL-17F prevented the disease (207). In the T cell transfer model of colitis, IL-17A-deficient T cells appeared to exacerbate inflammation. However, this observation is explained by the excessive polarization of T cells into Th1 phenotype in the absence of IL-17A (208). Nevertheless, 32
43 another study reported that both IL-17A and IL-17F have pro-inflammatory functions in T cell-mediated colitis in a redundant fashion (209). Thus, Th17 cells can be protective in IBD by limiting bacterial infiltration, however unrestrained TH17 responses potently induce intestinal inflammation. 5.5 Regulatory T cells Regulatory T cells play a crucial role in maintaining immune tolerance in the gut. Early studies demonstrated that adoptive transfer of CD45RB + CD4 + naïve T cells into SCID mice initiates intestinal inflammation and wasting disease, whereas cotransfer of T cells with regulatory properties prevents the development of inflammation (210). In this model, transferred effector T cells undergo lymphopenia induced proliferation in the lymphoid tissues and migrate to lamina propria. In this model, Th17 and Th1 cells were found to be mediators of colitis (209, 211). These colitogenic T cells activate DCs, produce inflammatory cytokines and recruit myeloid cells resulting in severe colitis and wasting disease (212, 213). When effector T cells are cotransferred with regulatory T cells, the latter also home to the lamina propria and prevent pathology. The prevention of colitis is achieved by the secretion of IL-10 by Tregs in the intestinal tissue (214, 215). The important role of IL-10 in maintaining gut homeostasis is further exemplified by the observation that IL-10-deficient mice develop chronic microflora-dependent intestinal inflammation (176, 216). In addition to Tregs and lamina propria T cells, IL-10 is also produced by DCs and macrophages in the gut as demonstrated by generating IL-10-IRES-GFP reporter mice (217). However, T cell specific deletion of IL-10 was sufficient to induce spontaneous colitis demonstrating the significance of 33
44 immunoregulatory function of intestinal T cells in maintaining immune tolerance in the gut (218). Another cytokine important for Treg-mediated immune homeostasis in the gut is TGF-β. Although TGF-β is produced by several cell types including macrophages and dendritic cells (219), TGF-β produced by Tregs has been shown to be critical in preventing colitis in several studies. Suppressive properties of TGF-β primarily act on T cells (220). For example, T cells with a dominant negative TGF-β receptor are resistant to suppression by regulatory T cells and the absence of TGF-β mediated suppression on these T cells leads to intestinal inflammation in the T cell transfer model of colitis (221). Furthermore, TGF-β1 produced by regulatory T cells has been shown to inhibit Th1 differentiation and TGF-β1-deficient Tregs were unable to prevent inflammatory bowel disease in the T cell transfer model of colitis (222). Thus, IL-10 and TGF-β are the major mediators of Treg suppression in the intestine. 5.6 T cell lymphopenia and lymphopenia induced proliferation T cell lymphopenia is a condition that can be caused by several factors including viral infections, administration of potent immunosuppressive drugs and genetic mutations. Importantly, lymphopenia is commonly associated with development of immunopathologies (223). Mutations affecting the T cell development and survival can cause lymphopenia leading to a dysregulated immune regulation. For example, deletion of Was and TCRa genes in mice resulted in severe lymphopenia and importantly led to colitis (224, 225). Under lymphopenic conditions, T cells lose their clonal diversity and autoreactive T cells may expand and trigger autoimmunity. This type of T cell 34
45 proliferation during lymphopenia has been linked to development of immunopathologies in experimental animal models (212, 226). Thus, the immune dysregulation during lymphopenia leads to the expansion of autoreactive effector T cells leading to immunopathology. Under lymphopenic conditions, the behavior of naïve T cells is different than the basal homeostatic state. When transferred into irradiated acutely lymphopenic hosts, naïve T cells proliferate due to increased availability of IL-7 and self-peptide/mhc molecules (227). In the absence of either IL-7 or MHC, the lymphopenia induced proliferation of naïve T cells is impaired (30, ). However, when the host mice are chronically lymphopenic, such as RAG-deficient mice or Severe Combined Immuno Deficient (SCID) mice, transferred naïve T cells proliferate much more rapidly compared to acute lymphopenia induced proliferation (231). This type of lymphopenia induced proliferation depends on the foreign antigens present in the commensal microflora since only a slow rate of proliferation was observed when T cells were transferred into germ free RAG-deficient mice. Importantly, the proliferation of T cells induced by microflora was independent of IL-7 as their rapid proliferation was not affected when IL-7-RAG double-deficient hosts were used (231). In addition, transfer of monoclonal TCR transgenic T cells also proliferated slowly in a similar fashion with IL-7 dependent proliferation observed in acute lymphopenic hosts. These results suggest that the rapid proliferation of polyclonal T cells under chronic lymphopenic conditions is largely induced in response to commensal microflora. In summary, intestinal immune homeostasis and tolerance is regulated by several complex mechanisms and involve complex the interaction of multiple systems. 35
46 Failure of these mechanisms can lead to intestinal pathogenesis. Development of IBD is influenced by environmental and genetic factors. Thus a critical question to address is to identify genes that predispose to colitis. In the next chapter, we will introduce ENU mutagenesis as a forward genetics method which is used to identify of novel immune related genes which may be potential candidates for predisposition to autoimmune disease. 6. Forward genetics Efforts in the genome sequencing during the last decade yielded vast amount of information about mammalian genomes. It is believed that mammalian genome contains over 20,000 genes most of which are poorly characterized. One of the frequently utilized approaches to characterize gene function is the reverse genetics which employs the targeted disruption of genes as well as silencing genes through the RNAi technology. Targeted disruption of genes allows investigating the effects of genetic deletions in vivo. Reverse genetics is a hypothesis based approach where the investigator generates a genetic alteration and analyzes the phenotype. On the other hand, forward genetics approach starts with a phenotype and proceeds with the identification of the causative genetic mutation (232). These phenotypes can arise spontaneously or can be induced by mutagens such as N-ethyl-N-nitrosourea (ENU). ENU is a mutagen which generates point mutation throughout the genome. Using the ENU mutagenesis approach with mice, a phenotype of interest can be screened in a high throughput manner. The advantage of this approach is that novel genes with almost no prior functional information can be identified. Moreover, hypomorphic mutations induced by ENU approach are advantageous in cases such as targeted disruption of the whole gene 36
47 causes embryonic lethality. Additionally, hypomorphic mutations can be informative for protein domain function if the structural integrity of the protein is not disrupted (233). Recent advancements in genome sequencing technology allow forward genetics to be an efficient way for obtaining information on novel gene function. Immunological screens using the ENU method are routinely employed to screen phenotypes affecting innate and adaptive components of the immune system. For example, mutations affecting the innate immunity can be screened by analyzing the function of intra-peritoneal macrophages exposed to a variety of pathogens or TLR ligands. Alternatively, mutations affecting the development and function of lymphocytes can be screened by in vivo cytotoxicity assays. The latter method involves the adoptive transfer of natural killer cell targets and cytotoxic T cell targets into the putative mutant mice. Several immunological phenotypes and causative mutations have been identified using the forward genetics approach. Identification of previously uncharacterized genes such as Tlr4, Trif, Unc93b, Unc13d and Themis made valuable contributions to innate and adaptive immunity studies (234, 235). Recently, by using an in vivo cytotoxicity approach, our laboratory has identified a new mouse phenotype designated as sphinx. The causative mutation affected a GTPase gene which belongs to the GTPase of Immunity Associated Proteins (Gimap) family and known as Gimap5. We will summarize the literature about the Gimap family and specifically Gimap5 in the following section. 37
48 7. Family of GTPase of Immunity Associated Proteins 7.1 Identification of Gimap of GTPases in plants and vertebrates The family of GTPase of Immunity Associated Proteins (Gimap) represents a novel family expressed in vertebrates and angiosperm plants (26, ). The plant prototype of Gimap family was first identified in Arabidopsis Thaliana as a plant immunity gene carrying an AIG1 homology domain (239). Expression of AIG1 gene in A.thaliana was upregulated in response to Pseudomonas syringae infection and provided resistance. AIG1 stands for avrrpt2-induced gene as it is induced by the virulence factor avrrpt2 secreted by Pseudomonas syringae (239). In mice, the first Gimap family member was identified as Imap38, later designated as Gimap1, which was induced by experimental malaria Plasmodium chabaudi (236). 7.2 Genomic organization of Gimap family The genes of Gimap family are positioned in tight clusters in vertebrate genomes. Eight functional Gimap genes are clustered in mouse chromosome 6, whereas seven genes are found in rat chromosome 4. There are seven functional and one pseudogene in human chromosome 7 (26, 236). GIMAP genes span a 300 kb region in human genome and a 120 kb interval in mouse and rat genomes (240). Gimap3 encodes for a functional protein in mice whereas no open reading frames are found in humans and rats suggesting that Gimap3 is a pseudogene in these species. On the other hand, orthologs for human Gimap2 gene have not been identified in mouse and rat genomes (238). The prototype of the Gimap family in plants is the AtIAN family in Arabidopsis thaliana. There are 13 AtIAN genes in A.thaliana dispersed into chromosomes 1, 2 and 38
49 4. Nine of the AtIAN genes are tightly clustered in chromosome 1 and three of them are clustered together in chromosome 4 (237). The genomic organization of Gimap family genes in tight clusters and strong sequence similarities between them suggest the existence of gene duplication events during evolution. Gimap family genes are found in vertebrates and flowering plants. No orthologs of Gimap family genes were found in C.elegans, fruit fly or Baker s yeast of which complete genome sequences have been reported (237). Thus, it can be speculated that the first Gimap gene has emerged in a common ancestor of plants and vertebrates and well conserved in certain phyla, however lost in others during evolution. 7.3 Structural Features of Gimap proteins Gimap family proteins represent a subfamily in guanine nucleotide binding super family of proteins. They are small GTPases composed of amino acids with molecular weights ranging between 34kDa to 38kDa. Mouse Gimap8 however, contains three GTPase domains and has a molecular weight of 75kDa (241). All Gimap family proteins contain an AIG1 domain which exhibits the typical features of GTP binding domains. AIG1 domain contains G1-G5 motifs that are characteristic to GTP binding proteins. Additionally, between the G3 and G4 motifs, AIG1 domain exhibits a hydrophobic, highly conserved box motif which is unique for Gimap proteins (26, 236). Recent crystallographic studies of Gimap2 and Gimap5 have shown a highly similar architecture between the two proteins (242, 243). Gimap proteins are composed of two switch regions, six β-sheets and seven α- helices. Gimap proteins oligomerize by 39
50 binding through two interfaces. Specifically, nucleotide binding facilitates stabilization of switch I and induces the formation of a G-interface across the GTP binding site. In addition, formation of a C-interface is also induced by GTP binding. The c-terminal α- helix α7 outside the core GTPase domain has amphipathic properties and is in close proximity to switch II region. Upon GTP binding, remodeling of switch II facilitates the release of α7 which appears to be required for oligomerization across the C-interface since the constructs can form tetramers only when α7 is removed. The nucleotide coordination and oligomerization mode of Gimap2 appears to be similar to dynamin GTPases (243). Dynamin proteins form oligomeric helical structures and catalyze membrane scission which is coupled to GTP hydrolysis (244). Moreover, three Gimap proteins Gimap1, 2 and 5 exhibit hydrophobic transmembrane regions in their C-terminal. These structural properties suggest that these Gimap proteins associate with intracellular lipid membranes and may perform functions such as membrane fusion, scission or scaffolding. 7.4 Cellular localization of Gimap5 Gimap5 carries a hydrophobic transmembrane domain in its C-terminal suggesting a membrane associated localization in the cell. However, there have been controversial reports about the membrane compartments that contain Gimap5 protein. Initially, using an over expression approach, Gimap5 has been reported to localize on mitochondrial membranes. It was suggested that Gimap5 interacts with anti-apoptotic protein Bcl-2 and prevents apoptosis (26). Another report showed Gimap5 localizes to endoplasmic reticulum and its over expression induces cell death via an apoptotic 40
51 pathway (245). These discrepancies possibly arose from the lack of specific antibodies available for Gimap5 and the use of overexpression methods that can alter the natural localization of membrane targeted proteins. Keita et al. took advantage of anti-sera raised against Gimap5 and utilized subcellular fractionation technique which demonstrated that endogenous Gimap5 localizes to a subcellular compartment distinct from mitochondria or endoplasmic reticulum (246). More recently, a monoclonal antibody was generated against Gimap5 and it was demonstrated that Gimap5 expression is associated with lysosomes and multivesicular bodies in the cell (247). The lysosomal localization of Gimap5 suggests a role in lysosomal stability, maturation or function. In addition, other Gimap family members, Gimap1 and Gimap2 have been shown to be localized in Golgi and lipid droplets respectively (243, 247). The diverse cellular localizations of Gimap proteins in various membrane compartments suggest that they might be involved in similar functions in different organelles to regulate cell viability. 7.5 Gimap5 in autoimmunity Gimap5 has been associated to a number of autoimmune diseases. Studies in the BioBreeding (BB) rats revealed a truncated from of Gimap5 to be linked to lymphopenia and diabetes development (248, 249). Diabetes-prone BB rats (BBDP) develop spontaneous insulin dependent diabetes which is similar to human type I diabetes and they have been studied as an autoimmunity model for decades (250). The locus containing Gimap5 gene was originally identified as lymphopenia (lyp) locus in BB rats. Initial studies demonstrated that genetic susceptibility to diabetes and lymphopenia segregates to a single autosomal trait designated as Iddm1 or lyp locus in rat 41
52 chromosome 4 (251). This notion was supported by the observation that introgression of BBDP lyp locus into BB diabetes resistant (BBDR) background resulted in both diabetes and lymphopenia (252). However, more recently, introgression of BBDP lyp locus into F344 rat background demonstrated that lymphopenia phenotype can be dissected from diabetes (253). F344lyp/lyp rats were lymphopenic to the same extent as BBDP rats but they did not develop diabetes. These results suggest that those mutant lyp locus and lymphopenia phenotypes are essential for the initiation of autoimmune diabetes in BB rats but require additional genomic elements for disease development. Indeed, rat MHC haplotype RT1u/u and some additional possible diabetes susceptibility loci have been linked to diabetes development in BBDPlyp/lyp rats ( ). Eventually, a frameshift mutation in Gimap5 gene has been demonstrated to be responsible for the T cell lymphopenia phenotype in BB rats and in other congenic lyp strains (240, 257, 258). Together, these results suggest that lyp mutation in Gimap5 gene causes lymphopenia in different rat backgrounds and is an essential factor for development of autoimmune diabetes in susceptible genomic backgrounds. When the mutant lyp (Gimap5) locus made congenic into a different rat background, namely the PVG strain, Gimap5-deficient rats developed inflammatory bowel disease presenting features similar to human eosinophilic gastroenteritis (259). T cells in these rats showed lymphopenia, increased activation and a Th2 skewed phenotype. Interestingly, another report demonstrated that type I diabetes in BBDP rats was partially prevented by antibiotic treatment suggesting that gut flora might be involved in the development diabetes (260). It has been suggested that lymphopenia caused by Gimap5 deficiency creates conditions leading to impaired immunoregulation 42
53 and consequently resulting in T cell-mediated autoimmunity; however the type and the tissue specificity of the T cell response are influenced by the genetic and environmental factors. Genetic polymorphisms in human GIMAP5 gene were linked with increased susceptibility to systemic lupus erythematosus (SLE) (261). A single nucleotide polymorphism (SNP) rs6598 in the polyadenylation (polya) signal of Gimap5 gene has been associated with increased risk to SLE in Finnish and British patients. An A G replacement in the polya site resulted in a longer transcript and decreased expression of GIMAP5 in SLE patients (261). In a different study, with Korean patients, two SNPs in GIMAP5 gene were associated with susceptibility to SLE. Additionally, another SNP was linked to development of leukopenia. Although not significant, a trend of association was observed between Gimap5 polymorphisms and lymphopenia in these SLE patients (262). SLE is characterized by the production of auto-antibodies to multiple nuclear antigens. SLE is a complex heterogeneous autoimmune disease and the exact mechanisms underlying pathogenesis remain to be determined. SLE is associated with increased amounts of apoptotic cells that accumulate in various tissues of patients (263). Moreover, decreased lymphocyte viability and accelerated apoptosis was observed in SLE patients (264). Loss of lymphocyte survival might cause lymphopenia which may lead to a failure in immune regulation in SLE patients. However, the role of GIMAP5 in immune dysregulation in SLE in humans remains poorly defined. 43
54 Additional studies have examined the potential association between GIMAP5 polymorphisms and development of type I diabetes in humans. An early study revealed that single nucleotide polymorphisms in the GIMAP5 locus did not associate with increased susceptibility to diabetes in human samples from U.K. and U.S (265). However, a more recent study in Swedish type I diabetes patients, demonstrated a significant association between SNP rs6598 in the GIMAP5 polya site and very high IA- 2 autoantibody levels (266). The circulation of islet autoantibodies before the clinical onset of type I diabetes makes them a potential marker for overt hyperglycemia risk (267). It is possible that the SNP in the GIMAP5 polya region may alter the expression level of GIMAP5 protein and may be a risk factor for the trigger of islet autoimmunity in newly diagnosed type I diabetes patients. Furthermore, in a different study, lower expression of GIMAP5 gene in regulatory T cells has been found in recent onset type I diabetes patients (268). In summary, the association of genetic variations in human GIMAP5 locus with autoimmune diseases makes Gimap5 a promising target for treatment options. The involvement of Gimap5 in autoimmune diseases and the underlying mechanisms leading to loss of immune tolerance require further studies. 7.6 Gimap5 and T lymphocyte development Gimap5 is mainly expressed in lymphocyte lineage (26). During thymic development of T cells, Gimap5 expression is tightly regulated. In developing rat T cells, Gimap5 mrna expression is induced at the DP stage and remains high in CD4 and CD8 SP thymocytes (241). A different report states that expression of mouse Gimap5 is induced during the transition from DP into the CD4 and CD8 SP stages (26). Eventually, Gimap5 expression remains high in the peripheral CD4 + and CD8 + T cells (269). 44
55 The role of Gimap5 during thymocyte development is not well understood. Based on an in vitro fetal thymic organ culture study, it has been reported that knock down of Gimap5 mrna resulted in a disturbed thymocyte development. It has been proposed that Gimap5 is required for optimal generation of DP thymocytes (26). In contrast, in vivo Gimap5-deficient mouse and rat models did not show defects in the frequencies of DP and SP thymocytes ( ). However, phenotypic differences were observed in the BB rat CD8 SP thymocytes showing an increased TCR low to TCR high ratio (270). Moreover, CD8 SP thymocytes in Gimap5 knock-out mice showed a defective upregulation of Qa-2 which is a marker of maturation as thymocytes get ready to emigrate from the thymus (272). These results indicate that Gimap5 is not essential for the positive selection in the thymus but has a non-redundant role for the maturation of positively selected CD8 SP thymocytes into naïve peripheral lymphocytes. 7.7 Gimap5 in peripheral T cell survival The lymphopenia phenotype occurring spontaneously in diabetic BBDP rats has been investigated extensively (273). It was demonstrated that the peripheral T cells of BBDP rats have dramatically reduced in vitro lifespan due to increased cell death (271). Peripheral T cell lymphopenia in this rat strain was found to be a consequence of a mutated Gimap5 (lyp) gene which encodes for a truncated non-functional protein (240). Similarly, Gimap5 null mice showed impaired survival of peripheral T cells but also showed disrupted NK cell development indicating a broader role for Gimap5 in lymphocyte development (272). In parallel with these results, Gimap1, a closely related Gimap family member was shown to be essential for late stages of T and B cell development and peripheral survival (274). On the other hand, a pro-cell death function 45
56 was proposed for Gimap4 (275). Although the deletion of Gimap4 did not have any effect on T cell development and survival in vivo, Gimap4-deficient T cells were more resistant to stress-induced cell death. Together, these results indicate diverse but related roles for different Gimap family members in the survival of T lymphocytes. Since Gimap5-deficiency caused a peripheral T cell survival defect in BBDP rats, several groups proposed an anti-apoptotic function for Gimap5. Sandal et al reported that overexpression of human GIMAP5 inhibits -radiation and okadaic acid induced apoptosis in Jurkat cells (276). However, it did not protect against several other apoptosis inducing factors including TNF-α, anti-fas and staurosporine. It was demonstrated that Gimap5 prevents -radiation induced cleavage of Caspase-3 although the mechanism for this apoptosis inhibition is not well defined (276). Pandarpurkar et al reported that Gimap5 is required for mitochondrial integrity in rat T lymphocytes. It was shown that absence of Gimap5 causes mitochondrial dysfunction, increased levels of mitochondrial chaperones and T cell specific spontaneous apoptosis in BBDP rats (277). A mechanism for Gimap5-mediated regulation of T cell survival has been proposed based on immunoprecipitation experiments. It was suggested that Gimap5 localizes to mitochondrial outer membrane and is critical for maintaining mitochondrial integrity by directly interacting with antiapoptotic proteins Bcl-2 and Bcl-xL (26). Moreover, mouse Gimap5 has been also reported to interact with anti-apoptotic Mcl-1 in lymphocyte progenitor cells in the bone marrow (278). From these studies however, it is unclear how Gimap5 regulates the actions of Bcl-2 family proteins and how these interactions maintain T cell survival. In contrast, rat GIimap5 has been shown to neither localize to mitochondria nor associate 46
57 with Bcl-2 indicating that it maintains the mitochondrial membrane integrity in T cells from a distinct subcellular component such as a vesicular organelle (246). Consistent with this hypothesis, increased expression of ER stress inducible transcription factor CHOP was reported as a possible cause of apoptosis in Gimap5-deficient BBDP rat T cells (279). Previously, CHOP has been proposed to regulate apoptosis by inducing the transcription of Bim and repressing Bcl-2 expression (280, 281). However, the role of Gimap5 in regulating ER homeostasis and whether Gimap5 deficiency alters the expression levels of Bim and Bcl-2 in BBDP rat T cells were not demonstrated. In summary, there is growing evidence that mitochondrial integrity is lost and T lymphocyte survival is reduced in the absence of Gimap5 function. Whether Gimap5 directly regulates mitochondrial stability by regulating anti-apoptotic proteins in T cells or the loss mitochondrial integrity is an indirect effect of Gimap5-deficiency is unclear. The molecular basis of the pro-survival role of Gimap5 in maintaining mitochondrial integrity and preventing cell death remains to be established. Gimap5 has been associated with a number of lymphocyte malignancies. Gimap5 has been shown to be over-expressed in B cell chronic lymphocytic leukemia (B-CLL) and mantle cell lymphoma (282). In addition, in a microarray study, Gimap5 has been found as one of the Notch target genes in Jurkat leukemia T cell line. It has been demonstrated that Notch signaling upregulates the expression of Gimap5 in Jurkat cells (283). Constitutively active forms of Notch have been known to exert oncogenic activity in T cell acute lymphoblastic leukemia (T-ALL) cells (284). It has been proposed that Notch signaling prevents apoptosis and promotes leukemogenesis in T-ALL cells by inducing the expression of GIMAP5 (285). These data indicate that GIMAP5 may play 47
58 an important role in Notch mediated regulation of T-cell development and survival. However, the mechanism for the suggested pro-survival role of Gimap5 in Notch induced T-ALL is not understood yet. 7.8 Gimap5 in T cell homeostasis and immune tolerance In addition to the observed survival defect, peripheral T cells in Gimap5-deficient BBDP rats also displayed homeostatic abnormalities. BBDP rats exhibited an increased proportion of T cells in cell cycle (286). Concordantly, peripheral T cells in BBDP rats displayed a spontaneous activated state associated with decreased levels of cell cycle inhibitor p27 (287). Moreover, Gimap5-deficient T cells have been shown to downregulate CD62L expression and spontaneously enter cell cycle in the absence of exogenous stimulation yet they do not progress through the mitosis phase (288). This spontaneous activated state was proposed to be a result of increased sensitivity of BBDP T cells to basal level TCR signals. Furthermore, the activated state of BBDP rat T cells were associated with increased activation of NFκB through a MAP kinase dependent pathway indicating that Gimap5 suppresses the MAP kinase pathway and regulates T cell sensitivity to TCR signals (289). Therefore, Gimap5 might have a role in decreasing the TCR signaling strength and promoting T cell quiescence by a currently unclear mechanism. Gimap5-deficiency has been associated with impaired Ca 2+ signaling in rat T cells. It has been suggested that basal homeostatic TCR signals induce a weak Ca +2 response that is not strong enough to provide survival signals for Gimap5-deficient BBDP T cells. Whereas, it was proposed that auto-antigens would induce a still low but 48
59 sufficient Ca 2+ response to provide survival and activation of autoreactive T cells which in turn initiate autoimmunity in BBDP rats (290). Therefore, Gimap5 deficiency might create conditions in which naïve T cells spontaneously die, whereas self-peptide/mhc signals provide the survival and enrichment of the autoreactive T cell population. It is possible that Gimap5 does not directly act on apoptotic pathways or cell cycle but might modulate the interaction of T cells and their environment. T cells with regulatory function are widely recognized as important determinants in autoimmune diseases in both animal models and humans (291). Early experiments with BBDP rats showed that transfusion of whole blood from diabetes resistant rats to diabetes prone BBDP rats protect against pancreatic insulitis and diabetes (292). Furthermore, a rat T cell maturation marker RT6.1 expressed on T cells with immune regulatory properties was absent in BBDP rats. Depletion of RT6.1 + T cell population in the diabetes resistant BBDR strain induced the development of diabetes (293). These results imply that Gimap5 is essential for the existence of a suppressive T cell population expressing RT6.1 and absence of these cells causes diabetes in Gimap5- deficient BBDP rats. More recently, it has been demonstrated that adoptive transfer of functional CD4 + CD25 + T cells prevented diabetes in BBDP rats (294, 295). Analysis of T cell populations in BBDP rats revealed that development of regulatory T cells in the thymus was normal; however these regulatory T cells exhibited impaired survival and maintenance at the post-thymic level (296, 297). Moreover, it has been proposed that diabetes development in BBDP rats is the result of an increased ratio of Th17 to regulatory T cell numbers rather than impaired function of Gimap5-deficient regulatory T 49
60 cells (298). In contrast, other groups reported that Gimap5-deficient regulatory T cells have impaired suppressive capacity in vitro (299, 300). Briefly, although the thymic development of natural regulatory T cells is unaffected from Gimap5 deficiency, Gimap5 appears to be required for the function, survival, maintenance and/or the induction of Treg cells in the peripheral lymphoid system. As the regulatory T cell subset is an important component of immune system to circumvent the progression of autoimmune diseases, it is critical to understand how Gimap5 plays a role in the development and function of regulatory T cells. 8. Summary Studies within the last decade identified Gimap5 as an important GTPase for the survival and homeostasis of lymphocytes. Moreover, mutations in Gimap5 gene have been linked to autoimmune diseases such as Type I diabetes and SLE in animal models and humans. However, the cellular and molecular events leading to autoimmunity in the absence of Gimap5 function have not been well described. Thus, it is still unclear how a disrupted Gimap5 function leads to T cell lymphopenia and predisposes to autoimmunity. In addition, recent studies revealed that Gimap5 is expressed in lysosomal structures. Moreover, structural studies suggested that Gimap proteins might have intracellular membrane associated roles such as fusion, scission, or scaffolding. Nevertheless, the signals regulating Gimap5 activity and the molecular function of Gimap5 have not been established yet. 50
61 The overall goal of this thesis study is to define the cellular and molecular mechanisms underlying Gimap5-mediated regulation of T cell homeostasis and immunological tolerance. Until recently, a mouse model for Gimap5 was not available. Using ENU mutagenesis, our laboratory has identified a mutant that carries a loss of function mutation in the Gimap5 gene. Based on the previous data, we hypothesize that (i) Gimap5 regulates lymphocyte homeostasis and quiescence and is necessary for maintaining immunological tolerance in mice; (ii) Gimap5 regulates the maintenance and function of regulatory T cells and loss of Treg function underlies the development of intestinal inflammation in Gimap5-deficient mice; and (iii) Gimap5 is required for lysosomal function in T cells which is regulated by TCR signaling and plays an intricate role in lymphocyte metabolism and T cell proliferation. In order to test these hypotheses we aim: 1) To characterize the lymphocyte development and homeostasis in Gimap5- deficient mice and to test whether the loss of Gimap5 leads to autoimmunity in mice. 2) To determine the cellular mechanisms involved in the development of colitis in Gimap5-deficient mice. 3) To define the role of Gimap5 in lysosome/autophagosome function following TCR-induced T cell activation. 51
62 References 1. Germain, R. N T-cell development and the CD4-CD8 lineage decision. Nature reviews. Immunology 2: Radtke, F., A. Wilson, G. Stark, M. Bauer, J. van Meerwijk, H. R. MacDonald, and M. Aguet Deficient T cell fate specification in mice with an induced inactivation of Notch1. Immunity 10: Kondo, M., I. L. Weissman, and K. Akashi Identification of clonogenic common lymphoid progenitors in mouse bone marrow. Cell 91: Godfrey, D. I., J. Kennedy, T. Suda, and a. Zlotnik A developmental pathway involving four phenotypically and functionally distinct subsets of CD3-CD4-CD8- triplenegative adult mouse thymocytes defined by CD44 and CD25 expression. Journal of immunology (Baltimore, Md. : 1950) 150: Hardt, C., T. Diamantstein, and H. Wagner Developmentally controlled expression of IL 2 receptors and of sensitivity to IL 2 in a subset of embryonic thymocytes. Journal of immunology 134: von Boehmer, H., A. Crisanti, P. Kisielow, and W. Haas Absence of growth by most receptor-expressing fetal thymocytes in the presence of interleukin-2. Nature 314: Rodewald, H. R., and H. J. Fehling Molecular and cellular events in early thymocyte development. Advances in immunology 69: Dudley, E. C., H. T. Petrie, L. M. Shah, M. J. Owen, and a. C. Hayday T cell receptor beta chain gene rearrangement and selection during thymocyte development in adult mice. Immunity 1: Fehling, H. J., A. Krotkova, C. Saint-Ruf, and H. von Boehmer Crucial role of the pre-t-cell receptor alpha gene in development of alpha beta but not gamma delta T cells. Nature 375: Aifantis, I., O. Azogui, J. Feinberg, C. Saint-Ruf, J. Buer, and H. von Boehmer On the role of the pre-t cell receptor in alphabeta versus gammadelta T lineage commitment. Immunity 9: Hoffman, E. S., L. Passoni, T. Crompton, T. M. Leu, D. G. Schatz, a. Koff, M. J. Owen, and a. C. Hayday Productive T-cell receptor beta-chain gene rearrangement: coincident regulation of cell cycle and clonality during development in vivo. Genes & Development 10: Hogquist, K. a., S. C. Jameson, W. R. Heath, J. L. Howard, M. J. Bevan, and F. R. Carbone T cell receptor antagonist peptides induce positive selection. Cell 76:
63 13. Surh, C. D., and J. Sprent T-cell apoptosis detected in situ during positive and negative selection in the thymus. Nature 372: Bouillet, P., J. F. Purton, D. I. Godfrey, L.-C. Zhang, L. Coultas, H. Puthalakath, M. Pellegrini, S. Cory, J. M. Adams, and A. Strasser BH3-only Bcl-2 family member Bim is required for apoptosis of autoreactive thymocytes. Nature 415: Rathmell, J. C., T. Lindsten, W.-X. Zong, R. M. Cinalli, and C. B. Thompson Deficiency in Bak and Bax perturbs thymic selection and lymphoid homeostasis. Nature immunology 3: Strasser, a., a. W. Harris, H. von Boehmer, and S. Cory Positive and negative selection of T cells in T-cell receptor transgenic mice expressing a bcl-2 transgene. Proceedings of the National Academy of Sciences of the United States of America 91: Nagata, S Human autoimmune lymphoproliferative syndrome, a defect in the apoptosis-inducing Fas receptor: a lesson from the mouse model. Journal of human genetics 43: Kishimoto, H., C. D. Surh, and J. Sprent A role for Fas in negative selection of thymocytes in vivo. The Journal of experimental medicine 187: Villunger, A., V. S. Marsden, Y. Zhan, M. Erlacher, A. M. Lew, P. Bouillet, S. Berzins, D. I. Godfrey, W. R. Heath, and A. Strasser Negative selection of semimature CD4(+)8(-)HSA+ thymocytes requires the BH3-only protein Bim but is independent of death receptor signaling. Proceedings of the National Academy of Sciences of the United States of America 101: Zhou, T., J. Cheng, P. Yang, Z. Wang, C. Liu, X. Su, H. Bluethmann, and J. D. Mountz Inhibition of Nur77/Nurr1 leads to inefficient clonal deletion of self-reactive T cells. The Journal of experimental medicine 183: Woolf, E., C. Xiao, O. Fainaru, J. Lotem, D. Rosen, V. Negreanu, Y. Bernstein, D. Goldenberg, O. Brenner, G. Berke, D. Levanon, and Y. Groner Runx3 and Runx1 are required for CD8 T cell development during thymopoiesis. Proceedings of the National Academy of Sciences of the United States of America 100: Sun, G., X. Liu, P. Mercado, S. R. Jenkinson, M. Kypriotou, L. Feigenbaum, P. Galera, and R. Bosselut The zinc finger protein ckrox directs CD4 lineage differentiation during intrathymic T cell positive selection. Nature immunology 6: He, X., and D. J. Kappes CD4/CD8 lineage commitment: light at the end of the tunnel? Current opinion in immunology 18: Chen, W The late stage of T cell development within mouse thymus. Cellular & molecular immunology 1: Carlson, C. M., B. T. Endrizzi, J. Wu, X. Ding, M. a. Weinreich, E. R. Walsh, M. a. Wani, J. B. Lingrel, K. a. Hogquist, and S. C. Jameson Kruppel-like factor 2 regulates thymocyte and T-cell migration. Nature 442:
64 26. Nitta, T., M. Nasreen, T. Seike, A. Goji, I. Ohigashi, T. Miyazaki, T. Ohta, M. Kanno, and Y. Takahama IAN family critically regulates survival and development of T lymphocytes. PLoS biology 4: e Berger, M., P. Krebs, K. Crozat, X. Li, B. A. Croker, O. M. Siggs, D. Popkin, X. Du, B. R. Lawson, A. N. Theofilopoulos, Y. Xia, K. Khovananth, E. M. Moresco, T. Satoh, O. Takeuchi, S. Akira, and B. Beutler An Slfn2 mutation causes lymphoid and myeloid immunodeficiency due to loss of immune cell quiescence. Nature immunology 11: Schluns, K. S., and L. Lefrancois Cytokine control of memory T-cell development and survival. Nature reviews. Immunology 3: Takada, K., and S. C. Jameson Naive T cell homeostasis: from awareness of space to a sense of place. Nature reviews. Immunology 9: Takada, K., and S. C. Jameson Self-class I MHC molecules support survival of naive CD8 T cells, but depress their functional sensitivity through regulation of CD8 expression levels. The Journal of experimental medicine 206: Polic, B., D. Kunkel, A. Scheffold, and K. Rajewsky How alpha beta T cells deal with induced TCR alpha ablation. Proceedings of the National Academy of Sciences of the United States of America 98: Opferman, J. T., A. Letai, C. Beard, M. D. Sorcinelli, C. C. Ong, and S. J. Korsmeyer Development and maintenance of B and T lymphocytes requires antiapoptotic MCL-1. Nature 426: Maraskovsky, E., L. A. O'Reilly, M. Teepe, L. M. Corcoran, J. J. Peschon, and A. Strasser Bcl-2 can rescue T lymphocyte development in interleukin-7 receptordeficient mice but not in mutant rag-1-/- mice. Cell 89: Wojciechowski, S., P. Tripathi, T. Bourdeau, L. Acero, H. L. Grimes, J. D. Katz, F. D. Finkelman, and D. A. Hildeman Bim/Bcl-2 balance is critical for maintaining naive and memory T cell homeostasis. The Journal of experimental medicine 204: Park, J. H., Q. Yu, B. Erman, J. S. Appelbaum, D. Montoya-Durango, H. L. Grimes, and A. Singer Suppression of IL7Ralpha transcription by IL-7 and other prosurvival cytokines: a novel mechanism for maximizing IL-7-dependent T cell survival. Immunity 21: Smith-Garvin, J. E., G. A. Koretzky, and M. S. Jordan T cell activation. Annual review of immunology 27: Mosmann, T. R., H. Cherwinski, M. W. Bond, M. A. Giedlin, and R. L. Coffman Two types of murine helper T cell clone. I. Definition according to profiles of lymphokine activities and secreted proteins. Journal of immunology 136: Harrington, L. E., R. D. Hatton, P. R. Mangan, H. Turner, T. L. Murphy, K. M. Murphy, and C. T. Weaver Interleukin 17-producing CD4+ effector T cells develop via a 54
65 lineage distinct from the T helper type 1 and 2 lineages. Nature immunology 6: Hsieh, C. S., S. E. Macatonia, C. S. Tripp, S. F. Wolf, A. O'Garra, and K. M. Murphy Development of TH1 CD4+ T cells through IL-12 produced by Listeria-induced macrophages. Science 260: Jacobson, N. G., S. J. Szabo, R. M. Weber-Nordt, Z. Zhong, R. D. Schreiber, J. E. Darnell, Jr., and K. M. Murphy Interleukin 12 signaling in T helper type 1 (Th1) cells involves tyrosine phosphorylation of signal transducer and activator of transcription (Stat)3 and Stat4. The Journal of experimental medicine 181: Carnaud, C., D. Lee, O. Donnars, S. H. Park, A. Beavis, Y. Koezuka, and A. Bendelac Cutting edge: Cross-talk between cells of the innate immune system: NKT cells rapidly activate NK cells. Journal of immunology 163: Biron, C. A., and L. Brossay NK cells and NKT cells in innate defense against viral infections. Current opinion in immunology 13: Afkarian, M., J. R. Sedy, J. Yang, N. G. Jacobson, N. Cereb, S. Y. Yang, T. L. Murphy, and K. M. Murphy T-bet is a STAT1-induced regulator of IL-12R expression in naive CD4+ T cells. Nature immunology 3: Szabo, S. J., S. T. Kim, G. L. Costa, X. Zhang, C. G. Fathman, and L. H. Glimcher A novel transcription factor, T-bet, directs Th1 lineage commitment. Cell 100: Zhu, J., and W. E. Paul Peripheral CD4+ T-cell differentiation regulated by networks of cytokines and transcription factors. Immunological reviews 238: Swain, S. L., a. D. Weinberg, M. English, and G. Huston IL-4 directs the development of Th2-like helper effectors. Journal of immunology (Baltimore, Md. : 1950) 145: Le Gros, G., S. Z. Ben-Sasson, R. Seder, F. D. Finkelman, and W. E. Paul Generation of interleukin 4 (IL-4)-producing cells in vivo and in vitro: IL-2 and IL-4 are required for in vitro generation of IL-4-producing cells. The Journal of experimental medicine 172: Shimoda, K., J. van Deursen, M. Y. Sangster, S. R. Sarawar, R. T. Carson, R. A. Tripp, C. Chu, F. W. Quelle, T. Nosaka, D. A. Vignali, P. C. Doherty, G. Grosveld, W. E. Paul, and J. N. Ihle Lack of IL-4-induced Th2 response and IgE class switching in mice with disrupted Stat6 gene. Nature 380: Zheng, W., and R. A. Flavell The transcription factor GATA-3 is necessary and sufficient for Th2 cytokine gene expression in CD4 T cells. Cell 89: Maggi, E., P. Parronchi, R. Manetti, C. Simonelli, M. P. Piccinni, F. S. Rugiu, M. De Carli, M. Ricci, and S. Romagnani Reciprocal regulatory effects of IFN-gamma and IL-4 on the in vitro development of human Th1 and Th2 clones. Journal of immunology 148:
66 51. Manetti, R., P. Parronchi, M. G. Giudizi, M. P. Piccinni, E. Maggi, G. Trinchieri, and S. Romagnani Natural killer cell stimulatory factor (interleukin 12 [IL-12]) induces T helper type 1 (Th1)-specific immune responses and inhibits the development of IL-4- producing Th cells. The Journal of experimental medicine 177: Usui, T., R. Nishikomori, A. Kitani, and W. Strober GATA-3 suppresses Th1 development by downregulation of Stat4 and not through effects on IL-12Rbeta2 chain or T-bet. Immunity 18: Hwang, E. S., S. J. Szabo, P. L. Schwartzberg, and L. H. Glimcher T helper cell fate specified by kinase-mediated interaction of T-bet with GATA-3. Science 307: Kobayashi, T., S. Okamoto, T. Hisamatsu, N. Kamada, H. Chinen, R. Saito, M. T. Kitazume, A. Nakazawa, A. Sugita, K. Koganei, K. Isobe, and T. Hibi IL23 differentially regulates the Th1/Th17 balance in ulcerative colitis and Crohn's disease. Gut 57: Bailey, S. L., B. Schreiner, E. J. McMahon, and S. D. Miller CNS myeloid DCs presenting endogenous myelin peptides 'preferentially' polarize CD4+ T(H)-17 cells in relapsing EAE. Nature immunology 8: Mangan, P. R., L. E. Harrington, D. B. O'Quinn, W. S. Helms, D. C. Bullard, C. O. Elson, R. D. Hatton, S. M. Wahl, T. R. Schoeb, and C. T. Weaver Transforming growth factor-beta induces development of the T(H)17 lineage. Nature 441: Nurieva, R., X. O. Yang, G. Martinez, Y. Zhang, A. D. Panopoulos, L. Ma, K. Schluns, Q. Tian, S. S. Watowich, A. M. Jetten, and C. Dong Essential autocrine regulation by IL-21 in the generation of inflammatory T cells. Nature 448: Zhou, L., I. I. Ivanov, R. Spolski, R. Min, K. Shenderov, T. Egawa, D. E. Levy, W. J. Leonard, and D. R. Littman IL-6 programs T(H)-17 cell differentiation by promoting sequential engagement of the IL-21 and IL-23 pathways. Nature immunology 8: Ivanov, II, B. S. McKenzie, L. Zhou, C. E. Tadokoro, A. Lepelley, J. J. Lafaille, D. J. Cua, and D. R. Littman The orphan nuclear receptor RORgammat directs the differentiation program of proinflammatory IL-17+ T helper cells. Cell 126: Ahmed, R., and D. Gray Immunological memory and protective immunity: understanding their relation. Science 272: Wherry, E. J., V. Teichgraber, T. C. Becker, D. Masopust, S. M. Kaech, R. Antia, U. H. von Andrian, and R. Ahmed Lineage relationship and protective immunity of memory CD8 T cell subsets. Nature immunology 4: Sallusto, F., D. Lenig, R. Forster, M. Lipp, and A. Lanzavecchia Two subsets of memory T lymphocytes with distinct homing potentials and effector functions. Nature 401:
67 63. Murali-Krishna, K., and R. Ahmed Cutting edge: naive T cells masquerading as memory cells. Journal of immunology (Baltimore, Md. : 1950) 165: Cho, B. K., V. P. Rao, Q. Ge, H. N. Eisen, and J. Chen Homeostasis-stimulated proliferation drives naive T cells to differentiate directly into memory T cells. The Journal of experimental medicine 192: Goldrath, a. W., L. Y. Bogatzki, and M. J. Bevan Naive T cells transiently acquire a memory-like phenotype during homeostasis-driven proliferation. The Journal of experimental medicine 192: Haluszczak, C., A. D. Akue, S. E. Hamilton, L. D. S. Johnson, L. Pujanauski, L. Teodorovic, S. C. Jameson, and R. M. Kedl The antigen-specific CD8+ T cell repertoire in unimmunized mice includes memory phenotype cells bearing markers of homeostatic expansion. The Journal of experimental medicine 206: Kieper, W. C., and S. C. Jameson Homeostatic expansion and phenotypic conversion of naïve T cells in response to self peptide/mhc ligands. Proceedings of the National Academy of Sciences of the United States of America 96: Buckley, a. F., C. T. Kuo, and J. M. Leiden Transcription factor LKLF is sufficient to program T cell quiescence via a c-myc-dependent pathway. Nature immunology 2: Mateyak, M. K., A. J. Obaya, and J. M. Sedivy c-myc regulates cyclin D-Cdk4 and -Cdk6 activity but affects cell cycle progression at multiple independent points. Molecular and cellular biology 19: Iritani, B. M., and R. N. Eisenman c-myc enhances protein synthesis and cell size during B lymphocyte development. Proceedings of the National Academy of Sciences of the United States of America 96: Stahl, M., P. F. Dijkers, G. J. Kops, S. M. Lens, P. J. Coffer, B. M. Burgering, and R. H. Medema The forkhead transcription factor FoxO regulates transcription of p27kip1 and Bim in response to IL-2. Journal of immunology 168: Lin, L., J. D. Hron, and S. L. Peng Regulation of NF-kappaB, Th activation, and autoinflammation by the forkhead transcription factor Foxo3a. Immunity 21: Kerdiles, Y. M., D. R. Beisner, R. Tinoco, A. S. Dejean, D. H. Castrillon, R. a. DePinho, and S. M. Hedrick Foxo1 links homing and survival of naive T cells by regulating L-selectin, CCR7 and interleukin 7 receptor. Nature immunology 10: Billingham, R. E., L. Brent, and P. B. Medawar Actively acquired tolerance of foreign cells. Nature 172: Bluestone, J. a Mechanisms of tolerance. Immunological reviews 241: Burnet, F. M The Clonal Selection Theory of Acquired Immunity. Cambridge University Press, Cambridge. 57
68 77. Kappler, J. W., N. Roehm, and P. Marrack T cell tolerance by clonal elimination in the thymus. Cell 49: Kisielow, P., H. Bluthmann, U. D. Staerz, M. Steinmetz, and H. von Boehmer Tolerance in T-cell-receptor transgenic mice involves deletion of nonmature CD4+8+ thymocytes. Nature 333: Sakaguchi, N., T. Takahashi, H. Hata, T. Nomura, T. Tagami, S. Yamazaki, T. Sakihama, T. Matsutani, I. Negishi, S. Nakatsuru, and S. Sakaguchi Altered thymic T-cell selection due to a mutation of the ZAP-70 gene causes autoimmune arthritis in mice. Nature 426: Goodnow, C. C., J. Sprent, B. Fazekas de St Groth, and C. G. Vinuesa Cellular and genetic mechanisms of self tolerance and autoimmunity. Nature 435: Kyewski, B., and L. Klein A central role for central tolerance. Annual review of immunology 24: Derbinski, J., a. Schulte, B. Kyewski, and L. Klein Promiscuous gene expression in medullary thymic epithelial cells mirrors the peripheral self. Nature immunology 2: Nagamine, K., P. Peterson, H. S. Scott, J. Kudoh, S. Minoshima, M. Heino, K. J. Krohn, M. D. Lalioti, P. E. Mullis, S. E. Antonarakis, K. Kawasaki, S. Asakawa, F. Ito, and N. Shimizu Positional cloning of the APECED gene. Nature genetics 17: Aaltonen, J., N. Horelli-Kuitunen, J. B. Fan, P. Bjorses, J. Perheentupa, R. Myers, A. Palotie, and L. Peltonen High-resolution physical and transcriptional mapping of the autoimmune polyendocrinopathy-candidiasis-ectodermal dystrophy locus on chromosome 21q22.3 by FISH. Genome research 7: Ramsey, C., O. Winqvist, L. Puhakka, M. Halonen, A. Moro, O. Kämpe, P. Eskelin, M. Pelto-Huikko, and L. Peltonen Aire deficient mice develop multiple features of APECED phenotype and show altered immune response. Human molecular genetics 11: Liston, A., S. Lesage, J. Wilson, L. Peltonen, and C. C. Goodnow Aire regulates negative selection of organ-specific T cells. Nature immunology 4: Schwartz, R. H T cell anergy. Annual review of immunology 21: Lohr, J., B. Knoechel, V. Nagabhushanam, and A. K. Abbas T-cell tolerance and autoimmunity to systemic and tissue-restricted self-antigens. Immunological reviews 204: Jenkins, M. K., and R. H. Schwartz Antigen presentation by chemically modified splenocytes induces antigen-specific T cell unresponsiveness in vitro and in vivo. The Journal of experimental medicine 165:
69 90. Jenkins, M. K., C. A. Chen, G. Jung, D. L. Mueller, and R. H. Schwartz Inhibition of antigen-specific proliferation of type 1 murine T cell clones after stimulation with immobilized anti-cd3 monoclonal antibody. Journal of immunology 144: Gimmi, C. D., G. J. Freeman, J. G. Gribben, K. Sugita, a. S. Freedman, C. Morimoto, and L. M. Nadler B-cell surface antigen B7 provides a costimulatory signal that induces T cells to proliferate and secrete interleukin 2. Proceedings of the National Academy of Sciences of the United States of America 88: Gimmi, C. D., G. J. Freeman, J. G. Gribben, G. Gray, and L. M. Nadler Human T- cell clonal anergy is induced by antigen presentation in the absence of B7 costimulation. Proceedings of the National Academy of Sciences of the United States of America 90: Ohashi, P. S., S. Oehen, K. Buerki, H. Pircher, C. T. Ohashi, B. Odermatt, B. Malissen, R. M. Zinkernagel, and H. Hengartner Ablation of "tolerance" and induction of diabetes by virus infection in viral antigen transgenic mice. Cell 65: Kearney, E. R., K. A. Pape, D. Y. Loh, and M. K. Jenkins Visualization of peptidespecific T cell immunity and peripheral tolerance induction in vivo. Immunity 1: Nurieva, R. I., X. Liu, and C. Dong Molecular mechanisms of T-cell tolerance. Immunological reviews 241: Chiang, Y. J., H. K. Kole, K. Brown, M. Naramura, S. Fukuhara, R. J. Hu, I. K. Jang, J. S. Gutkind, E. Shevach, and H. Gu Cbl-b regulates the CD28 dependence of T- cell activation. Nature 403: Jeon, M.-S., A. Atfield, K. Venuprasad, C. Krawczyk, R. Sarao, C. Elly, C. Yang, S. Arya, K. Bachmaier, L. Su, D. Bouchard, R. Jones, M. Gronski, P. Ohashi, T. Wada, D. Bloom, C. G. Fathman, Y.-C. Liu, and J. M. Penninger Essential role of the E3 ubiquitin ligase Cbl-b in T cell anergy induction. Immunity 21: Tivol, E. a., F. Borriello, a. N. Schweitzer, W. P. Lynch, J. a. Bluestone, and a. H. Sharpe Loss of CTLA-4 leads to massive lymphoproliferation and fatal multiorgan tissue destruction, revealing a critical negative regulatory role of CTLA-4. Immunity 3: Waterhouse, P., J. M. Penninger, E. Timms, a. Wakeham, a. Shahinian, K. P. Lee, C. B. Thompson, H. Griesser, and T. W. Mak Lymphoproliferative disorders with early lethality in mice deficient in Ctla-4. Science (New York, N.Y.) 270: Greenwald, R. J., V. a. Boussiotis, R. B. Lorsbach, a. K. Abbas, and a. H. Sharpe CTLA-4 regulates induction of anergy in vivo. Immunity 14: Yamazaki, T., H. Akiba, H. Iwai, H. Matsuda, M. Aoki, Y. Tanno, T. Shin, H. Tsuchiya, D. M. Pardoll, K. Okumura, M. Azuma, and H. Yagita Expression of programmed death 1 ligands by murine T cells and APC. Journal of immunology (Baltimore, Md. : 1950) 169:
70 102. Liang, S. C., Y. E. Latchman, J. E. Buhlmann, M. F. Tomczak, B. H. Horwitz, G. J. Freeman, and A. H. Sharpe Regulation of PD-1, PD-L1, and PD-L2 expression during normal and autoimmune responses. European journal of immunology 33: Selenko-Gebauer, N., O. Majdic, A. Szekeres, G. Höfler, E. Guthann, U. Korthäuer, G. Zlabinger, P. Steinberger, W. F. Pickl, H. Stockinger, W. Knapp, and J. Stöckl B7- H1 (programmed death-1 ligand) on dendritic cells is involved in the induction and maintenance of T cell anergy. Journal of immunology (Baltimore, Md. : 1950) 170: Ansari, M. J. I., A. D. Salama, T. Chitnis, R. N. Smith, H. Yagita, H. Akiba, T. Yamazaki, M. Azuma, H. Iwai, S. J. Khoury, H. Auchincloss, and M. H. Sayegh The programmed death-1 (PD-1) pathway regulates autoimmune diabetes in nonobese diabetic (NOD) mice. The Journal of experimental medicine 198: Wang, J., T. Yoshida, F. Nakaki, H. Hiai, T. Okazaki, and T. Honjo Establishment of NOD-Pdcd1-/- mice as an efficient animal model of type I diabetes. Proceedings of the National Academy of Sciences of the United States of America 102: Fontenot, J. D., and A. Y. Rudensky A well adapted regulatory contrivance: regulatory T cell development and the forkhead family transcription factor Foxp3. Nature immunology 6: Gershon, R. K., and K. Kondo Infectious immunological tolerance. Immunology 21: Qin, S., S. P. Cobbold, H. Pope, J. Elliott, D. Kioussis, J. Davies, and H. Waldmann "Infectious" transplantation tolerance. Science (New York, N.Y.) 259: Chen, Y., V. K. Kuchroo, J. Inobe, D. a. Hafler, and H. L. Weiner Regulatory T cell clones induced by oral tolerance: suppression of autoimmune encephalomyelitis. Science (New York, N.Y.) 265: Sakaguchi, S., K. Fukuma, K. Kuribayashi, and T. Masuda Organ-specific autoimmune diseases induced in mice by elimination of T cell subset. I. Evidence for the active participation of T cells in natural self-tolerance; deficit of a T cell subset as a possible cause of autoimmune disease. The Journal of experimental medicine 161: Powrie, F., and D. Mason OX-22high CD4+ T cells induce wasting disease with multiple organ pathology: prevention by the OX-22low subset. The Journal of experimental medicine 172: Sakaguchi, S., N. Sakaguchi, M. Asano, M. Itoh, and M. Toda Immunologic selftolerance maintained by activated T cells expressing IL-2 receptor alpha-chains (CD25). Breakdown of a single mechanism of self-tolerance causes various autoimmune diseases. Journal of immunology 155: Kuniyasu, Y., T. Takahashi, M. Itoh, J. Shimizu, G. Toda, and S. Sakaguchi Naturally anergic and suppressive CD25(+)CD4(+) T cells as a functionally and 60
71 phenotypically distinct immunoregulatory T cell subpopulation. International immunology 12: Russell, W. L., L. B. Russell, and J. S. Gower Exceptional Inheritance of a Sex- Linked Gene in the Mouse Explained on the Basis That the X/O Sex-Chromosome Constitution Is Female. Proceedings of the National Academy of Sciences of the United States of America 45: Lyon, M. F., J. Peters, P. H. Glenister, S. Ball, and E. Wright The scurfy mouse mutant has previously unrecognized hematological abnormalities and resembles Wiskott-Aldrich syndrome. Proceedings of the National Academy of Sciences of the United States of America 87: Godfrey, V. L., J. E. Wilkinson, and L. B. Russell X-linked lymphoreticular disease in the scurfy (sf) mutant mouse. The American journal of pathology 138: Blair, P. J., S. J. Bultman, J. C. Haas, B. T. Rouse, J. E. Wilkinson, and V. L. Godfrey CD4+CD8- T cells are the effector cells in disease pathogenesis in the scurfy (sf) mouse. Journal of immunology 153: Brunkow, M. E., E. W. Jeffery, K. a. Hjerrild, B. Paeper, L. B. Clark, S. a. Yasayko, J. E. Wilkinson, D. Galas, S. F. Ziegler, and F. Ramsdell Disruption of a new forkhead/winged-helix protein, scurfin, results in the fatal lymphoproliferative disorder of the scurfy mouse. Nature genetics 27: Bennett, C. L., J. Christie, F. Ramsdell, M. E. Brunkow, P. J. Ferguson, L. Whitesell, T. E. Kelly, F. T. Saulsbury, P. F. Chance, and H. D. Ochs The immune dysregulation, polyendocrinopathy, enteropathy, X-linked syndrome (IPEX) is caused by mutations of FOXP3. Nature genetics 27: Wildin, R. S., F. Ramsdell, J. Peake, F. Faravelli, J. L. Casanova, N. Buist, E. Levy- Lahad, M. Mazzella, O. Goulet, L. Perroni, F. D. Bricarelli, G. Byrne, M. McEuen, S. Proll, M. Appleby, and M. E. Brunkow X-linked neonatal diabetes mellitus, enteropathy and endocrinopathy syndrome is the human equivalent of mouse scurfy. Nature genetics 27: Fontenot, J. D., J. P. Rasmussen, L. M. Williams, J. L. Dooley, A. G. Farr, and A. Y. Rudensky Regulatory T cell lineage specification by the forkhead transcription factor foxp3. Immunity 22: Hori, S., T. Nomura, and S. Sakaguchi Control of regulatory T cell development by the transcription factor Foxp3. Science (New York, N.Y.) 299: Khattri, R., T. Cox, S.-A. Yasayko, and F. Ramsdell An essential role for Scurfin in CD4+CD25+ T regulatory cells. Nature immunology 4: Fontenot, J. D., M. a. Gavin, and A. Y. Rudensky Foxp3 programs the development and function of CD4+CD25+ regulatory T cells. Nature immunology 4:
72 125. Kim, J. M., J. P. Rasmussen, and A. Y. Rudensky Regulatory T cells prevent catastrophic autoimmunity throughout the lifespan of mice. Nature immunology 8: Jordan, M. S., A. Boesteanu, A. J. Reed, A. L. Petrone, A. E. Holenbeck, M. A. Lerman, A. Naji, and A. J. Caton Thymic selection of CD4+CD25+ regulatory T cells induced by an agonist self-peptide. Nature immunology 2: Olivares-Villagómez, D., Y. Wang, and J. J. Lafaille Regulatory CD4(+) T cells expressing endogenous T cell receptor chains protect myelin basic protein-specific transgenic mice from spontaneous autoimmune encephalomyelitis. The Journal of experimental medicine 188: Itoh, M., T. Takahashi, N. Sakaguchi, Y. Kuniyasu, J. Shimizu, F. Otsuka, and S. Sakaguchi Thymus and autoimmunity: production of CD25+CD4+ naturally anergic and suppressive T cells as a key function of the thymus in maintaining immunologic self-tolerance. Journal of immunology (Baltimore, Md. : 1950) 162: Curotto de Lafaille, M. A., A. C. Lino, N. Kutchukhidze, and J. J. Lafaille CD25- T cells generate CD25+Foxp3+ regulatory T cells by peripheral expansion. Journal of immunology 173: Furtado, G. C., M. A. Curotto de Lafaille, N. Kutchukhidze, and J. J. Lafaille Interleukin 2 signaling is required for CD4(+) regulatory T cell function. The Journal of experimental medicine 196: Marie, J. C., J. J. Letterio, M. Gavin, and A. Y. Rudensky TGF-beta1 maintains suppressor function and Foxp3 expression in CD4+CD25+ regulatory T cells. The Journal of experimental medicine 201: Chen, W., W. Jin, N. Hardegen, K.-J. Lei, L. Li, N. Marinos, G. McGrady, and S. M. Wahl Conversion of peripheral CD4+CD25- naive T cells to CD4+CD25+ regulatory T cells by TGF-beta induction of transcription factor Foxp3. The Journal of experimental medicine 198: Thorstenson, K. M., and A. Khoruts Generation of anergic and potentially immunoregulatory CD25+CD4 T cells in vivo after induction of peripheral tolerance with intravenous or oral antigen. Journal of immunology (Baltimore, Md. : 1950) 167: Mucida, D., N. Kutchukhidze, A. Erazo, M. Russo, J. J. Lafaille, and M. A. Curotto de Lafaille Oral tolerance in the absence of naturally occurring Tregs. The Journal of clinical investigation 115: Apostolou, I., and H. von Boehmer In vivo instruction of suppressor commitment in naive T cells. The Journal of experimental medicine 199: Schmidt-Supprian, M., J. Tian, E. P. Grant, M. Pasparakis, R. Maehr, H. Ovaa, H. L. Ploegh, A. J. Coyle, and K. Rajewsky Differential dependence of CD4+CD25+ regulatory and natural killer-like T cells on signals leading to NF-kappaB activation. 62
73 Proceedings of the National Academy of Sciences of the United States of America 101: Gupta, S., S. Manicassamy, C. Vasu, A. Kumar, W. Shang, and Z. Sun Differential requirement of PKC-theta in the development and function of natural regulatory T cells. Molecular immunology 46: Barnes, M. J., P. Krebs, N. Harris, C. Eidenschenk, R. Gonzalez-Quintial, C. N. Arnold, K. Crozat, S. Sovath, E. M. Moresco, A. N. Theofilopoulos, B. Beutler, and K. Hoebe Commitment to the regulatory T cell lineage requires CARMA1 in the thymus but not in the periphery. PLoS biology 7: e Deenick, E. K., A. R. Elford, M. Pellegrini, H. Hall, T. W. Mak, and P. S. Ohashi c- Rel but not NF-kappaB1 is important for T regulatory cell development. European journal of immunology 40: Mantel, P. Y., N. Ouaked, B. Ruckert, C. Karagiannidis, R. Welz, K. Blaser, and C. B. Schmidt-Weber Molecular mechanisms underlying FOXP3 induction in human T cells. Journal of immunology 176: Wu, Y., M. Borde, V. Heissmeyer, M. Feuerer, A. D. Lapan, J. C. Stroud, D. L. Bates, L. Guo, A. Han, S. F. Ziegler, D. Mathis, C. Benoist, L. Chen, and A. Rao FOXP3 controls regulatory T cell function through cooperation with NFAT. Cell 126: Bettelli, E., Y. Carrier, W. Gao, T. Korn, T. B. Strom, M. Oukka, H. L. Weiner, and V. K. Kuchroo Reciprocal developmental pathways for the generation of pathogenic effector TH17 and regulatory T cells. Nature 441: Takimoto, T., Y. Wakabayashi, T. Sekiya, N. Inoue, R. Morita, K. Ichiyama, R. Takahashi, M. Asakawa, G. Muto, T. Mori, E. Hasegawa, S. Saika, S. Shizuya, T. Hara, M. Nomura, and A. Yoshimura Smad2 and Smad3 are redundantly essential for the TGF-beta-mediated regulation of regulatory T plasticity and Th1 development. Journal of immunology (Baltimore, Md. : 1950) 185: Davidson, T. S., R. J. DiPaolo, J. Andersson, and E. M. Shevach Cutting Edge: IL-2 is essential for TGF-beta-mediated induction of Foxp3+ T regulatory cells. Journal of immunology 178: Burchill, M. A., J. Yang, C. Vogtenhuber, B. R. Blazar, and M. A. Farrar IL-2 receptor beta-dependent STAT5 activation is required for the development of Foxp3+ regulatory T cells. Journal of immunology 178: Harada, Y., Y. Harada, C. Elly, G. Ying, J.-H. Paik, R. a. DePinho, and Y.-C. Liu Transcription factors Foxo3a and Foxo1 couple the E3 ligase Cbl-b to the induction of Foxp3 expression in induced regulatory T cells. The Journal of experimental medicine 207: Ouyang, W., O. Beckett, Q. Ma, J.-h. Paik, R. a. DePinho, and M. O. Li Foxo proteins cooperatively control the differentiation of Foxp3+ regulatory T cells. Nature immunology 11:
74 148. Kerdiles, Y. M., E. L. Stone, D. R. Beisner, D. L. Beisner, M. a. McGargill, I. L. Ch'en, C. Stockmann, C. D. Katayama, and S. M. Hedrick Foxo transcription factors control regulatory T cell development and function. Immunity 33: Gomis, R. R., C. Alarcón, W. He, Q. Wang, J. Seoane, A. Lash, and J. Massagué A FoxO-Smad synexpression group in human keratinocytes. Proceedings of the National Academy of Sciences of the United States of America 103: Seoane, J., H.-V. Le, L. Shen, S. a. Anderson, and J. Massagué Integration of Smad and forkhead pathways in the control of neuroepithelial and glioblastoma cell proliferation. Cell 117: Zhou, L., J. E. Lopes, M. M. W. Chong, I. I. Ivanov, R. Min, G. D. Victora, Y. Shen, J. Du, Y. P. Rubtsov, A. Y. Rudensky, S. F. Ziegler, and D. R. Littman TGF-betainduced Foxp3 inhibits T(H)17 cell differentiation by antagonizing RORgammat function. Nature 453: Onishi, Y., Z. Fehervari, T. Yamaguchi, and S. Sakaguchi Foxp3+ natural regulatory T cells preferentially form aggregates on dendritic cells in vitro and actively inhibit their maturation. Proceedings of the National Academy of Sciences of the United States of America 105: Wing, K., Y. Onishi, P. Prieto-Martin, T. Yamaguchi, M. Miyara, Z. Fehervari, T. Nomura, and S. Sakaguchi CTLA-4 control over Foxp3+ regulatory T cell function. Science (New York, N.Y.) 322: Thornton, A. M., and E. M. Shevach CD4+CD25+ immunoregulatory T cells suppress polyclonal T cell activation in vitro by inhibiting interleukin 2 production. The Journal of experimental medicine 188: Pandiyan, P., L. Zheng, S. Ishihara, J. Reed, and M. J. Lenardo CD4+CD25+Foxp3+ regulatory T cells induce cytokine deprivation-mediated apoptosis of effector CD4+ T cells. Nature immunology 8: Shevach, E. M Mechanisms of foxp3+ T regulatory cell-mediated suppression. Immunity 30: Deaglio, S., K. M. Dwyer, W. Gao, D. Friedman, A. Usheva, A. Erat, J.-F. Chen, K. Enjyoji, J. Linden, M. Oukka, V. K. Kuchroo, T. B. Strom, and S. C. Robson Adenosine generation catalyzed by CD39 and CD73 expressed on regulatory T cells mediates immune suppression. The Journal of experimental medicine 204: Liang, B., C. Workman, J. Lee, C. Chew, B. M. Dale, L. Colonna, M. Flores, N. Li, E. Schweighoffer, S. Greenberg, V. Tybulewicz, D. Vignali, and R. Clynes Regulatory T cells inhibit dendritic cells by lymphocyte activation gene-3 engagement of MHC class II. Journal of immunology 180: Madan, R., F. Demircik, S. Surianarayanan, J. L. Allen, S. Divanovic, A. Trompette, N. Yogev, Y. Gu, M. Khodoun, D. Hildeman, N. Boespflug, M. B. Fogolin, L. Gröbe, M. Greweling, F. D. Finkelman, R. Cardin, M. Mohrs, W. Müller, A. Waisman, A. Roers, and 64
75 C. L. Karp Nonredundant roles for B cell-derived IL-10 in immune counterregulation. Journal of immunology (Baltimore, Md. : 1950) 183: Rubtsov, Y. P., J. P. Rasmussen, E. Y. Chi, J. Fontenot, L. Castelli, X. Ye, P. Treuting, L. Siewe, A. Roers, W. R. Henderson, W. Muller, and A. Y. Rudensky Regulatory T cell-derived interleukin-10 limits inflammation at environmental interfaces. Immunity 28: Collison, L. W., C. J. Workman, T. T. Kuo, K. Boyd, Y. Wang, K. M. Vignali, R. Cross, D. Sehy, R. S. Blumberg, and D. a. a. Vignali The inhibitory cytokine IL-35 contributes to regulatory T-cell function. Nature 450: Niedbala, W., X. Wei, B. Cai, A. J. Hueber, B. P. Leung, I. B. McInnes, and F. Y. Liew IL 35 is a novel cytokine with therapeutic effects against collagen induced arthritis through the expansion of regulatory T cells and suppression of Th17 cells. European journal of immunology 37: Gondek, D. C., L.-F. Lu, S. a. Quezada, S. Sakaguchi, and R. J. Noelle Cutting edge: contact-mediated suppression by CD4+CD25+ regulatory cells involves a granzyme B-dependent, perforin-independent mechanism. Journal of immunology (Baltimore, Md. : 1950) 174: Kappelman, M. D., S. L. Rifas-Shiman, K. Kleinman, D. Ollendorf, A. Bousvaros, R. J. Grand, and J. a. Finkelstein The prevalence and geographic distribution of Crohn's disease and ulcerative colitis in the United States. Clinical gastroenterology and hepatology : the official clinical practice journal of the American Gastroenterological Association 5: Brand, S Crohn's disease: Th1, Th17 or both? The change of a paradigm: new immunological and genetic insights implicate Th17 cells in the pathogenesis of Crohn's disease. Gut 58: Xavier, R. J., and D. K. Podolsky Unravelling the pathogenesis of inflammatory bowel disease. Nature 448: Duerr, R. H., K. D. Taylor, S. R. Brant, J. D. Rioux, M. S. Silverberg, M. J. Daly, a. H. Steinhart, C. Abraham, M. Regueiro, A. Griffiths, T. Dassopoulos, A. Bitton, H. Yang, S. Targan, L. W. Datta, E. O. Kistner, L. P. Schumm, A. T. Lee, P. K. Gregersen, M. M. Barmada, J. I. Rotter, D. L. Nicolae, and J. H. Cho A genome-wide association study identifies IL23R as an inflammatory bowel disease gene. Science (New York, N.Y.) 314: Ogura, Y., D. K. Bonen, N. Inohara, D. L. Nicolae, F. F. Chen, R. Ramos, H. Britton, T. Moran, R. Karaliuskas, R. H. Duerr, J. P. Achkar, S. R. Brant, T. M. Bayless, B. S. Kirschner, S. B. Hanauer, G. Nunez, and J. H. Cho A frameshift mutation in NOD2 associated with susceptibility to Crohn's disease. Nature 411: Franke, A., D. P. B. McGovern, J. C. Barrett, K. Wang, G. L. Radford-Smith, T. Ahmad, C. W. Lees, T. Balschun, J. Lee, and R. Roberts Genome-wide meta-analysis increases to 71 the number of confirmed Crohn's disease susceptibility loci. Nature genetics 42:
76 170. Anderson, C. A., G. Boucher, C. W. Lees, A. Franke, M. D'Amato, K. D. Taylor, J. C. Lee, P. Goyette, M. Imielinski, and A. Latiano Meta-analysis identifies 29 additional ulcerative colitis risk loci, increasing the number of confirmed associations to 47. Nature genetics 43: Mowat, A. M Anatomical basis of tolerance and immunity to intestinal antigens. Nature reviews. Immunology 3: Rakoff-Nahoum, S., L. Hao, and R. Medzhitov Role of toll-like receptors in spontaneous commensal-dependent colitis. Immunity 25: Ivanov, I. I., K. Atarashi, N. Manel, E. L. Brodie, T. Shima, U. Karaoz, D. Wei, K. C. Goldfarb, C. a. Santee, S. V. Lynch, T. Tanoue, A. Imaoka, K. Itoh, K. Takeda, Y. Umesaki, K. Honda, and D. R. Littman Induction of intestinal Th17 cells by segmented filamentous bacteria. Cell 139: Round, J. L., and S. K. Mazmanian The gut microbiota shapes intestinal immune responses during health and disease. Nature reviews. Immunology 9: Sartor, R. B Therapeutic manipulation of the enteric microflora in inflammatory bowel diseases: antibiotics, probiotics, and prebiotics. Gastroenterology 126: Sellon, R. K., S. Tonkonogy, M. Schultz, L. a. Dieleman, W. Grenther, E. Balish, D. M. Rennick, and R. B. Sartor Resident enteric bacteria are necessary for development of spontaneous colitis and immune system activation in interleukin-10- deficient mice. Infection and immunity 66: Asquith, M. J., O. Boulard, F. Powrie, and K. J. Maloy Pathogenic and protective roles of MyD88 in leukocytes and epithelial cells in mouse models of inflammatory bowel disease. Gastroenterology 139: , 529.e Abreu, M. T Toll-like receptor signalling in the intestinal epithelium: how bacterial recognition shapes intestinal function. Nature reviews. Immunology 10: Kobayashi, K. S., M. Chamaillard, Y. Ogura, O. Henegariu, N. Inohara, G. Nunez, and R. A. Flavell Nod2-dependent regulation of innate and adaptive immunity in the intestinal tract. Science 307: Hugot, J. P., M. Chamaillard, H. Zouali, S. Lesage, J. P. Cezard, J. Belaiche, S. Almer, C. Tysk, C. A. O'Morain, M. Gassull, V. Binder, Y. Finkel, A. Cortot, R. Modigliani, P. Laurent-Puig, C. Gower-Rousseau, J. Macry, J. F. Colombel, M. Sahbatou, and G. Thomas Association of NOD2 leucine-rich repeat variants with susceptibility to Crohn's disease. Nature 411: Zaki, M. H., K. L. Boyd, P. Vogel, M. B. Kastan, M. Lamkanfi, and T.-D. Kanneganti The NLRP3 inflammasome protects against loss of epithelial integrity and mortality during experimental colitis. Immunity 32: Schulz, O., E. Jaensson, E. K. Persson, X. Liu, T. Worbs, W. W. Agace, and O. Pabst Intestinal CD103+, but not CX3CR1+, antigen sampling cells migrate in lymph and 66
77 serve classical dendritic cell functions. The Journal of experimental medicine 206: Johansson-Lindbom, B., M. Svensson, O. Pabst, C. Palmqvist, G. Marquez, R. Förster, and W. W. Agace Functional specialization of gut CD103+ dendritic cells in the regulation of tissue-selective T cell homing. The Journal of experimental medicine 202: Coombes, J. L., and F. Powrie Dendritic cells in intestinal immune regulation. Nature reviews. Immunology 8: Coombes, J. L., K. R. R. Siddiqui, C. V. Arancibia-Cárcamo, J. Hall, C.-M. Sun, Y. Belkaid, and F. Powrie A functionally specialized population of mucosal CD103+ DCs induces Foxp3+ regulatory T cells via a TGF-beta and retinoic acid-dependent mechanism. The Journal of experimental medicine 204: Laffont, S., K. R. R. Siddiqui, and F. Powrie Intestinal inflammation abrogates the tolerogenic properties of MLN CD103+ dendritic cells. European journal of immunology 40: Niess, J. H., S. Brand, X. Gu, L. Landsman, S. Jung, B. a. McCormick, J. M. Vyas, M. Boes, H. L. Ploegh, J. G. Fox, D. R. Littman, and H.-C. Reinecker CX3CR1- mediated dendritic cell access to the intestinal lumen and bacterial clearance. Science (New York, N.Y.) 307: Niess, J. H., and G. Adler Enteric flora expands gut lamina propria CX3CR1+ dendritic cells supporting inflammatory immune responses under normal and inflammatory conditions. Journal of immunology (Baltimore, Md. : 1950) 184: Kontoyiannis, D., M. Pasparakis, T. T. Pizarro, F. Cominelli, and G. Kollias Impaired on/off regulation of TNF biosynthesis in mice lacking TNF AU-rich elements: implications for joint and gut-associated immunopathologies. Immunity 10: Targan, S. R., S. B. Hanauer, S. J. van Deventer, L. Mayer, D. H. Present, T. Braakman, K. L. DeWoody, T. F. Schaible, and P. J. Rutgeerts A short-term study of chimeric monoclonal antibody ca2 to tumor necrosis factor alpha for Crohn's disease. Crohn's Disease ca2 Study Group. The New England journal of medicine 337: Baumgart, D. C., and W. J. Sandborn Infl ammatory bowel disease : clinical aspects and established and evolving therapies. 369: Maloy, K. J., and F. Powrie Intestinal homeostasis and its breakdown in inflammatory bowel disease. Nature 474: Atreya, R., J. Mudter, S. Finotto, J. Müllberg, T. Jostock, S. Wirtz, M. Schütz, B. Bartsch, M. Holtmann, C. Becker, D. Strand, J. Czaja, J. F. Schlaak, H. a. Lehr, F. Autschbach, G. Schürmann, N. Nishimoto, K. Yoshizaki, H. Ito, T. Kishimoto, P. R. Galle, S. Rose- John, and M. F. Neurath Blockade of interleukin 6 trans signaling suppresses T- cell resistance against apoptosis in chronic intestinal inflammation: evidence in crohn disease and experimental colitis in vivo. Nature medicine 6:
78 194. Yamamoto, M., K. Yoshizaki, T. Kishimoto, and H. Ito IL-6 is required for the development of Th1 cell-mediated murine colitis. Journal of immunology (Baltimore, Md. : 1950) 164: Kaser, A., S. Zeissig, and R. S. Blumberg Inflammatory bowel disease. Annual review of immunology 28: Neurath, M. F., I. Fuss, B. L. Kelsall, E. Stüber, and W. Strober Antibodies to interleukin 12 abrogate established experimental colitis in mice. The Journal of experimental medicine 182: Neurath, M. F., B. Weigmann, S. Finotto, J. Glickman, E. Nieuwenhuis, H. Iijima, A. Mizoguchi, E. Mizoguchi, J. Mudter, P. R. Galle, A. Bhan, F. Autschbach, B. M. Sullivan, S. J. Szabo, L. H. Glimcher, and R. S. Blumberg The transcription factor T-bet regulates mucosal T cell activation in experimental colitis and Crohn's disease. The Journal of experimental medicine 195: Ito, H., and C. G. Fathman CD45RBhigh CD4+ T cells from IFN-gamma knockout mice do not induce wasting disease. Journal of autoimmunity 10: Oppmann, B., R. Lesley, B. Blom, J. C. Timans, Y. Xu, B. Hunte, F. Vega, N. Yu, J. Wang, K. Singh, F. Zonin, E. Vaisberg, T. Churakova, M. Liu, D. Gorman, J. Wagner, S. Zurawski, Y. Liu, J. S. Abrams, K. W. Moore, D. Rennick, R. de Waal-Malefyt, C. Hannum, J. F. Bazan, and R. a. Kastelein Novel p19 protein engages IL-12p40 to form a cytokine, IL-23, with biological activities similar as well as distinct from IL-12. Immunity 13: Kamada, N., T. Hisamatsu, S. Okamoto, H. Chinen, T. Kobayashi, T. Sato, A. Sakuraba, M. T. Kitazume, A. Sugita, K. Koganei, K. S. Akagawa, and T. Hibi Unique CD14 intestinal macrophages contribute to the pathogenesis of Crohn disease via IL-23/IFNgamma axis. The Journal of clinical investigation 118: Yen, D., J. Cheung, H. Scheerens, F. Poulet, T. McClanahan, B. McKenzie, M. A. Kleinschek, A. Owyang, J. Mattson, W. Blumenschein, E. Murphy, M. Sathe, D. J. Cua, R. A. Kastelein, and D. Rennick IL-23 is essential for T cell-mediated colitis and promotes inflammation via IL-17 and IL-6. The Journal of clinical investigation 116: Ahern, P. P., C. Schiering, S. Buonocore, M. J. McGeachy, D. J. Cua, K. J. Maloy, and F. Powrie Interleukin-23 drives intestinal inflammation through direct activity on T cells. Immunity 33: Cosmi, L., R. De Palma, V. Santarlasci, L. Maggi, M. Capone, F. Frosali, G. Rodolico, V. Querci, G. Abbate, R. Angeli, L. Berrino, M. Fambrini, M. Caproni, F. Tonelli, E. Lazzeri, P. Parronchi, F. Liotta, E. Maggi, S. Romagnani, and F. Annunziato Human interleukin 17-producing cells originate from a CD161+CD4+ T cell precursor. The Journal of experimental medicine 205: Sandborn, W. J., B. G. Feagan, R. N. Fedorak, E. Scherl, M. R. Fleisher, S. Katz, J. Johanns, M. Blank, and P. Rutgeerts A randomized trial of Ustekinumab, a 68
79 human interleukin-12/23 monoclonal antibody, in patients with moderate-to-severe Crohn's disease. Gastroenterology 135: Fujino, S., A. Andoh, S. Bamba, A. Ogawa, K. Hata, Y. Araki, T. Bamba, and Y. Fujiyama Increased expression of interleukin 17 in inflammatory bowel disease. Gut 52: Annunziato, F., L. Cosmi, V. Santarlasci, L. Maggi, F. Liotta, B. Mazzinghi, E. Parente, L. Fili, S. Ferri, F. Frosali, F. Giudici, P. Romagnani, P. Parronchi, F. Tonelli, E. Maggi, and S. Romagnani Phenotypic and functional features of human Th17 cells. The Journal of experimental medicine 204: Yang, X. O., S. H. Chang, H. Park, R. Nurieva, B. Shah, L. Acero, Y. H. Wang, K. S. Schluns, R. R. Broaddus, Z. Zhu, and C. Dong Regulation of inflammatory responses by IL-17F. The Journal of experimental medicine 205: O'Connor, W., Jr., M. Kamanaka, C. J. Booth, T. Town, S. Nakae, Y. Iwakura, J. K. Kolls, and R. A. Flavell A protective function for interleukin 17A in T cell-mediated intestinal inflammation. Nature immunology 10: Leppkes, M., C. Becker, I. I. Ivanov, S. Hirth, S. Wirtz, C. Neufert, S. Pouly, A. J. Murphy, D. M. Valenzuela, G. D. Yancopoulos, B. Becher, D. R. Littman, and M. F. Neurath RORgamma-expressing Th17 cells induce murine chronic intestinal inflammation via redundant effects of IL-17A and IL-17F. Gastroenterology 136: Powrie, F., M. W. Leach, S. Mauze, L. B. Caddle, and R. L. Coffman Phenotypically distinct subsets of CD4+ T cells induce or protect from chronic intestinal inflammation in C. B-17 scid mice. International immunology 5: Powrie, F., M. W. Leach, S. Mauze, S. Menon, L. B. Caddle, and R. L. Coffman Inhibition of Th1 responses prevents inflammatory bowel disease in scid mice reconstituted with CD45RBhi CD4+ T cells. Immunity 1: Izcue, A., J. L. Coombes, and F. Powrie Regulatory T cells suppress systemic and mucosal immune activation to control intestinal inflammation. Immunological reviews 212: Liu, Z., K. Geboes, S. Colpaert, L. Overbergh, C. Mathieu, H. Heremans, M. de Boer, L. Boon, G. D'Haens, P. Rutgeerts, and J. L. Ceuppens Prevention of experimental colitis in SCID mice reconstituted with CD45RBhigh CD4+ T cells by blocking the CD40- CD154 interactions. Journal of immunology 164: Asseman, C., S. Mauze, M. W. Leach, R. L. Coffman, and F. Powrie An essential role for interleukin 10 in the function of regulatory T cells that inhibit intestinal inflammation. The Journal of experimental medicine 190: Uhlig, H. H., J. Coombes, C. Mottet, A. Izcue, C. Thompson, A. Fanger, A. Tannapfel, J. D. Fontenot, F. Ramsdell, and F. Powrie Characterization of Foxp3+CD4+CD25+ and IL-10-secreting CD4+CD25+ T cells during cure of colitis. Journal of immunology (Baltimore, Md. : 1950) 177:
80 216. Kuhn, R., J. Lohler, D. Rennick, K. Rajewsky, and W. Muller Interleukin-10- deficient mice develop chronic enterocolitis. Cell 75: Kamanaka, M., S. T. Kim, Y. Y. Wan, F. S. Sutterwala, M. Lara-Tejero, J. E. Galán, E. Harhaj, and R. a. Flavell Expression of interleukin-10 in intestinal lymphocytes detected by an interleukin-10 reporter knockin tiger mouse. Immunity 25: Roers, A., L. Siewe, E. Strittmatter, M. Deckert, D. Schlüter, W. Stenzel, A. D. Gruber, T. Krieg, K. Rajewsky, and W. Müller T cell-specific inactivation of the interleukin 10 gene in mice results in enhanced T cell responses but normal innate responses to lipopolysaccharide or skin irritation. The Journal of experimental medicine 200: Letterio, J. J., and A. B. Roberts Regulation of immune responses by TGF-beta. Annual review of immunology 16: Li, M. O., S. Sanjabi, and R. a. Flavell Transforming growth factor-beta controls development, homeostasis, and tolerance of T cells by regulatory T cell-dependent and - independent mechanisms. Immunity 25: Fahlén, L., S. Read, L. Gorelik, S. D. Hurst, R. L. Coffman, R. a. Flavell, and F. Powrie T cells that cannot respond to TGF-beta escape control by CD4(+)CD25(+) regulatory T cells. The Journal of experimental medicine 201: Li, M. O., Y. Y. Wan, and R. a. Flavell T cell-produced transforming growth factorbeta1 controls T cell tolerance and regulates Th1- and Th17-cell differentiation. Immunity 26: Khoruts, A., and J. M. Fraser A causal link between lymphopenia and autoimmunity. Immunology letters 98: Mombaerts, P., E. Mizoguchi, M. J. Grusby, L. H. Glimcher, A. K. Bhan, and S. Tonegawa Spontaneous development of inflammatory bowel disease in T cell receptor mutant mice. Cell 75: Snapper, S. B., F. S. Rosen, E. Mizoguchi, P. Cohen, W. Khan, C. H. Liu, T. L. Hagemann, S. P. Kwan, R. Ferrini, L. Davidson, A. K. Bhan, and F. W. Alt Wiskott-Aldrich syndrome protein-deficient mice reveal a role for WASP in T but not B cell activation. Immunity 9: King, C., A. Ilic, K. Koelsch, and N. Sarvetnick Homeostatic expansion of T cells during immune insufficiency generates autoimmunity. Cell 117: Ernst, B., D. S. Lee, J. M. Chang, J. Sprent, and C. D. Surh The peptide ligands mediating positive selection in the thymus control T cell survival and homeostatic proliferation in the periphery. Immunity 11: Schluns, K. S., W. C. Kieper, S. C. Jameson, and L. Lefrancois Interleukin-7 mediates the homeostasis of naive and memory CD8 T cells in vivo. Nature immunology 1:
81 229. Tan, J. T., E. Dudl, E. LeRoy, R. Murray, J. Sprent, K. I. Weinberg, and C. D. Surh IL-7 is critical for homeostatic proliferation and survival of naive T cells. Proceedings of the National Academy of Sciences of the United States of America 98: Takeda, S., H. R. Rodewald, H. Arakawa, H. Bluethmann, and T. Shimizu MHC class II molecules are not required for survival of newly generated CD4+ T cells, but affect their long-term life span. Immunity 5: Kieper, W. C., A. Troy, J. T. Burghardt, C. Ramsey, J. Y. Lee, H. Q. Jiang, W. Dummer, H. Shen, J. J. Cebra, and C. D. Surh Recent immune status determines the source of antigens that drive homeostatic T cell expansion. Journal of immunology 174: Rutschmann, S., and K. Hoebe Dissecting innate immunity by germline mutagenesis. Immunology 123: Rutschmann, S., K. Hoebe, J. Zalevsky, X. Du, N. Mann, B. I. Dahiyat, P. Steed, and B. Beutler PanR1, a dominant negative missense allele of the gene encoding TNFalpha (Tnf), does not impair lymphoid development. Journal of immunology 176: Choi, O., and S. Rutschmann Dissecting immunity by germline mutagenesis. Immunology 137: Hoebe, K., and B. Beutler Forward genetic analysis of TLR-signaling pathways: an evaluation. Advanced drug delivery reviews 60: Krücken, J., R. M. U. Schroetel, I. U. Müller, N. Saïdani, P. Marinovski, W. P. M. Benten, O. Stamm, and F. Wunderlich Comparative analysis of the human gimap gene cluster encoding a novel GTPase family. Gene 341: Liu, C., T. Wang, W. Zhang, and X. Li Computational identification and analysis of immune-associated nucleotide gene family in Arabidopsis thaliana. Journal of plant physiology 165: Rutledge, E. a., J. M. Fuller, B. Van Yserloo, D. H. Moralejo, R. a. Ettinger, P. Gaur, J. L. Hoehna, M. R. Peterson, R. Jensen, A. E. Kwitek, and A. Lernmark Sequence variation and expression of the Gimap gene family in the BB rat. Experimental diabetes research 2009: Reuber, T. L., and F. M. Ausubel Isolation of Arabidopsis genes that differentiate between resistance responses mediated by the RPS2 and RPM1 disease resistance genes. The Plant cell 8: MacMurray, A. J., D. H. Moralejo, A. E. Kwitek, E. a. Rutledge, B. Van Yserloo, P. Gohlke, S. J. Speros, B. Snyder, J. Schaefer, S. Bieg, J. Jiang, R. a. Ettinger, J. Fuller, T. L. Daniels, A. Pettersson, K. Orlebeke, B. Birren, H. J. Jacob, E. S. Lander, and A. Lernmark Lymphopenia in the BB rat model of type 1 diabetes is due to a mutation in a novel immune-associated nucleotide (Ian)-related gene. Genome research 12:
82 241. Dion, C., C. Carter, L. Hepburn, W. J. Coadwell, G. Morgan, M. Graham, N. Pugh, G. Anderson, G. W. Butcher, and J. R. Miller Expression of the Ian family of putative GTPases during T cell development and description of an Ian with three sets of GTP/GDP-binding motifs. International immunology 17: Schwefel, D Structural and Functional analysis of Immunity-associated GTPases. Freien Universitat, Berlin Schwefel, D., C. Fröhlich, J. Eichhorst, B. Wiesner, J. Behlke, L. Aravind, and O. Daumke Structural basis of oligomerization in septin-like GTPase of immunityassociated protein 2 (GIMAP2). Proceedings of the National Academy of Sciences of the United States of America 107: Ford, M. G., S. Jenni, and J. Nunnari The crystal structure of dynamin. Nature 477: Dalberg, U., H. Markholst, and L. Hornum Both Gimap5 and the diabetogenic BBDP allele of Gimap5 induce apoptosis in T cells. International immunology 19: Keita, M., C. Leblanc, D. Andrews, and S. Ramanathan GIMAP5 regulates mitochondrial integrity from a distinct subcellular compartment. Biochem Biophys Res Commun 361: Wong, V. W., A. E. Saunders, A. Hutchings, J. C. Pascall, C. Carter, N. A. Bright, S. A. Walker, N. T. Ktistakis, and G. W. Butcher The autoimmunity-related GIMAP5 GTPase is a lysosome-associated protein. Self Nonself 1: Nakhooda, A. F., A. A. Like, C. I. Chappel, F. T. Murray, and E. B. Marliss The spontaneously diabetic Wistar rat. Metabolic and morphologic studies. Diabetes 26: Like, A. A., L. Butler, R. M. Williams, M. C. Appel, E. J. Weringer, and A. A. Rossini Spontaneous autoimmune diabetes mellitus in the BB rat. Diabetes 31: Ramanathan, S., and P. Poussier BB rat lyp mutation and Type 1 diabetes. Immunological reviews 184: Markholst, H., S. Eastman, D. Wilson, B. E. Andreasen, and a. Lernmark Diabetes segregates as a single locus in crosses between inbred BB rats prone or resistant to diabetes. The Journal of experimental medicine 174: Bieg, S., G. Koike, J. Jiang, L. Klaff, A. Pettersson, A. J. MacMurray, H. J. Jacob, E. S. Lander, and A. Lernmark Genetic isolation of iddm 1 on chromosome 4 in the biobreeding (BB) rat. Mammalian genome : official journal of the International Mammalian Genome Society 9: Moralejo, D. H., H. a. Park, S. J. Speros, A. J. MacMurray, A. E. Kwitek, H. J. Jacob, E. S. Lander, and Å. Lernmark Genetic dissection of lymphopenia from autoimmunity by introgression of mutated Ian5 gene onto the F344 rat. Journal of autoimmunity 21:
83 254. Colle, E., R. D. Guttmann, T. a. Seemayer, and F. Michel Spontaneous diabetes mellitus syndrome in the rat. IV. Immunogenetic interactions of MHC and non-mhc components of the syndrome. Metabolism: clinical and experimental 32: Wallis, R. H., K. Wang, L. Marandi, E. Hsieh, T. Ning, G. Y. Chao, J. Sarmiento, A. D. Paterson, and P. Poussier Type 1 diabetes in the BB rat: a polygenic disease. Diabetes 58: Bahr, J., N. Follak, N. Kloting, B. Wilke, B. Haertel, and I. Kloting Iddm1 and Iddm2 homozygous WOK.4BB rats develop lymphopenia, but no hyperglycemia like the BB/OK rats. Experimental and clinical endocrinology & diabetes : official journal, German Society of Endocrinology [and] German Diabetes Association 119: Hornum, L., J. Romer, and H. Markholst The diabetes-prone BB rat carries a frameshift mutation in Ian4, a positional candidate of Iddm1. Diabetes 51: Michalkiewicz, M., T. Michalkiewicz, R. a. Ettinger, E. a. Rutledge, J. M. Fuller, D. H. Moralejo, B. Van Yserloo, A. J. MacMurray, A. E. Kwitek, H. J. Jacob, E. S. Lander, and A. Lernmark Transgenic rescue demonstrates involvement of the Ian5 gene in T cell development in the rat. Physiological genomics 19: Cousins, L., M. Graham, R. Tooze, C. Carter, J. R. Miller, F. M. Powrie, G. G. Macpherson, and G. W. Butcher Eosinophilic bowel disease controlled by the BB rat-derived lymphopenia/gimap5 gene. Gastroenterology 131: Brugman, S., F. a. Klatter, J. T. J. Visser, a. C. M. Wildeboer-Veloo, H. J. M. Harmsen, J. Rozing, and N. a. Bos Antibiotic treatment partially protects against type 1 diabetes in the Bio-Breeding diabetes-prone rat. Is the gut flora involved in the development of type 1 diabetes? Diabetologia 49: Hellquist, A., M. Zucchelli, K. Kivinen, U. Saarialho-Kere, S. Koskenmies, E. Widen, H. Julkunen, A. Wong, M. L. Karjalainen-Lindsberg, T. Skoog, J. Vendelin, D. S. Cunninghame-Graham, T. J. Vyse, J. Kere, and C. M. Lindgren The human GIMAP5 gene has a common polyadenylation polymorphism increasing risk to systemic lupus erythematosus. J Med Genet 44: Lim, M. K., D. H. Sheen, S. a. Kim, S. K. Won, S.-S. Lee, S.-C. Chae, H.-T. Chung, and S. C. Shim IAN5 polymorphisms are associated with systemic lupus erythematosus. Lupus 18: Herrmann, M., R. E. Voll, O. M. Zoller, M. Hagenhofer, B. B. Ponner, and J. R. Kalden Impaired phagocytosis of apoptotic cell material by monocyte-derived macrophages from patients with systemic lupus erythematosus. Arthritis and rheumatism 41: Lorenz, H. M., M. Grunke, T. Hieronymus, M. Herrmann, A. Kuhnel, B. Manger, and J. R. Kalden In vitro apoptosis and expression of apoptosis-related molecules in lymphocytes from patients with systemic lupus erythematosus and other autoimmune diseases. Arthritis and rheumatism 40:
84 265. Payne, F., D. J. Smyth, R. Pask, B. J. Barratt, J. D. Cooper, R. C. Twells, N. M. Walker, A. C. Lam, L. J. Smink, S. Nutland, H. E. Rance, and J. A. Todd Haplotype tag single nucleotide polymorphism analysis of the human orthologues of the rat type 1 diabetes genes Ian4 (Lyp/Iddm1) and Cblb. Diabetes 53: Shin, J. H., M. Janer, B. McNeney, S. Blay, K. Deutsch, C. B. Sanjeevi, I. Kockum, A. Lernmark, J. Graham, G. Swedish Childhood Diabetes Study, G. Diabetes Incidence in Sweden Study, H. Arnqvist, E. Bjorck, J. Eriksson, L. Nystrom, L. O. Ohlson, B. Schersten, J. Ostman, M. Aili, L. E. Baath, E. Carlsson, H. Edenwall, G. Forsander, B. W. Granstrom, I. Gustavsson, R. Hanas, L. Hellenberg, H. Hellgren, E. Holmberg, H. Hornell, S. A. Ivarsson, C. Johansson, G. Jonsell, K. Kockum, B. Lindblad, A. Lindh, J. Ludvigsson, U. Myrdal, J. Neiderud, K. Segnestam, S. Sjoblad, L. Skogsberg, L. Stromberg, U. Stahle, B. Thalme, K. Tullus, T. Tuvemo, M. Wallensteen, O. Westphal, and J. Aman IA-2 autoantibodies in incident type I diabetes patients are associated with a polyadenylation signal polymorphism in GIMAP5. Genes Immun 8: Decochez, K., I. Truyen, B. van der Auwera, I. Weets, E. Vandemeulebroucke, I. H. de Leeuw, B. Keymeulen, C. Mathieu, R. Rottiers, D. G. Pipeleers, F. K. Gorus, and R. Belgian Diabetes Combined positivity for HLA DQ2/DQ8 and IA-2 antibodies defines population at high risk of developing type 1 diabetes. Diabetologia 48: Jailwala, P., J. Waukau, S. Glisic, S. Jana, S. Ehlenbach, M. Hessner, R. Alemzadeh, S. Matsuyama, P. Laud, X. Wang, and S. Ghosh Apoptosis of CD4+ CD25(high) T cells in type 1 diabetes may be partially mediated by IL-2 deprivation. PloS one 4: e Moralejo, D. H., J. M. Fuller, E. a. Rutledge, B. Van Yserloo, R. a. Ettinger, R. Jensen, W. Osborne, A. Kwitek, and A. Lernmark BB rat Gimap gene expression in sorted lymphoid T and B cells. Life sciences 89: Plamondon, C., V. Kottis, C. Brideau, M. D. Metroz-Dayer, and P. Poussier Abnormal thymocyte maturation in spontaneously diabetic BB rats involves the deletion of CD4-8+ cells. Journal of immunology 144: Hernandez-Hoyos, G., S. Joseph, N. G. Miller, and G. W. Butcher The lymphopenia mutation of the BB rat causes inappropriate apoptosis of mature thymocytes. European journal of immunology 29: Schulteis, R. D., H. Chu, X. Dai, Y. Chen, B. Edwards, D. Haribhai, C. B. Williams, S. Malarkannan, M. J. Hessner, S. Glisic-Milosavljevic, S. Jana, E. J. Kerschen, S. Ghosh, D. Wang, A. E. Kwitek, A. Lernmark, J. Gorski, and H. Weiler Impaired survival of peripheral T cells, disrupted NK/NKT cell development, and liver failure in mice lacking Gimap5. Blood 112: Filén, S., and R. Lahesmaa GIMAP Proteins in T-Lymphocytes. Journal of signal transduction 2010: Saunders, A., L. M. C. Webb, M. L. Janas, A. Hutchings, J. Pascall, C. Carter, N. Pugh, G. Morgan, M. Turner, and G. W. Butcher Putative GTPase GIMAP1 is critical for the development of mature B and T lymphocytes. Blood 115:
85 275. Schnell, S., C. Démollière, P. van den Berk, and H. Jacobs Gimap4 accelerates T-cell death. Blood 108: Sandal, T., L. Aumo, L. Hedin, B. T. Gjertsen, and S. O. Doskeland Irod/Ian5: an inhibitor of gamma-radiation- and okadaic acid-induced apoptosis. Molecular biology of the cell 14: Pandarpurkar, M., L. Wilson-Fritch, S. Corvera, H. Markholst, L. Hornum, D. L. Greiner, J. P. Mordes, A. a. Rossini, and R. Bortell Ian4 is required for mitochondrial integrity and T cell survival. Proceedings of the National Academy of Sciences of the United States of America 100: Chen, Y., M. Yu, X. Dai, M. Zogg, R. Wen, H. Weiler, and D. Wang Critical role for Gimap5 in the survival of mouse hematopoietic stem and progenitor cells. The Journal of experimental medicine 208: Pino, S. C., B. O'Sullivan-Murphy, E. A. Lidstone, C. Yang, K. L. Lipson, A. Jurczyk, P. diiorio, M. A. Brehm, J. P. Mordes, D. L. Greiner, A. A. Rossini, and R. Bortell CHOP mediates endoplasmic reticulum stress-induced apoptosis in Gimap5-deficient T cells. PLoS One 4: e Puthalakath, H., L. a. O'Reilly, P. Gunn, L. Lee, P. N. Kelly, N. D. Huntington, P. D. Hughes, E. M. Michalak, J. McKimm-Breschkin, N. Motoyama, T. Gotoh, S. Akira, P. Bouillet, and A. Strasser ER stress triggers apoptosis by activating BH3-only protein Bim. Cell 129: McCullough, K. D., J. L. Martindale, L. O. Klotz, T. Y. Aw, and N. J. Holbrook Gadd153 sensitizes cells to endoplasmic reticulum stress by down-regulating Bcl2 and perturbing the cellular redox state. Molecular and cellular biology 21: Zenz, T., a. Roessner, a. Thomas, S. Fröhling, H. Döhner, B. Calabretta, and L. Dahéron hian5: the human ortholog to the rat Ian4/Iddm1/lyp is a new member of the Ian family that is overexpressed in B-cell lymphoid malignancies. Genes and immunity 5: Chadwick, N., L. Zeef, V. Portillo, C. Fennessy, F. Warrander, S. Hoyle, and A.-M. Buckle Identification of novel Notch target genes in T cell leukaemia. Molecular cancer 8: Pear, W. S., J. C. Aster, M. L. Scott, R. P. Hasserjian, B. Soffer, J. Sklar, and D. Baltimore Exclusive development of T cell neoplasms in mice transplanted with bone marrow expressing activated Notch alleles. The Journal of experimental medicine 183: Chadwick, N., L. Zeef, V. Portillo, J. Boros, S. Hoyle, J. C. van Doesburg, and A. M. Buckle Notch protection against apoptosis in T-ALL cells mediated by GIMAP5. Blood Cells Mol Dis 45: Zadeh, H. H., D. L. Greiner, D. Y. Wu, F. Tausche, and I. Goldschneider Abnormalities in the export and fate of recent thymic emigrants in diabetes-prone BB/W rats. Autoimmunity 24:
86 287. Moore, J. K., R. I. Scheinman, and D. Bellgrau The identification of a novel T cell activation state controlled by a diabetogenic gene. Journal of immunology (Baltimore, Md. : 1950) 166: Lang, J. a., D. Kominski, D. Bellgrau, and R. I. Scheinman Partial activation precedes apoptotic death in T cells harboring an IAN gene mutation. European journal of immunology 34: Kupfer, R., J. Lang, C. Williams-Skipp, M. Nelson, D. Bellgrau, and R. I. Scheinman Loss of a gimap/ian gene leads to activation of NF-kappaB through a MAPKdependent pathway. Molecular immunology 44: Ilangumaran, S., M. Forand-Boulerice, S. M. Bousquet, A. Savard, P. Rocheleau, X. L. Chen, G. Dupuis, P. Poussier, G. Boulay, and S. Ramanathan Loss of GIMAP5 (GTPase of immunity-associated nucleotide binding protein 5) impairs calcium signaling in rat T lymphocytes. Molecular immunology 46: Sakaguchi, S., M. Ono, R. Setoguchi, H. Yagi, S. Hori, Z. Fehervari, J. Shimizu, T. Takahashi, and T. Nomura Foxp3+ CD25+ CD4+ natural regulatory T cells in dominant self-tolerance and autoimmune disease. Immunological reviews 212: Rossini, A. A., J. P. Mordes, A. M. Pelletier, and A. A. Like Transfusions of whole blood prevent spontaneous diabetes mellitus in the BB/W rat. Science 219: Greiner, D. L., J. P. Mordes, E. S. Handler, M. Angelillo, N. Nakamura, and A. A. Rossini Depletion of RT6.1+ T lymphocytes induces diabetes in resistant biobreeding/worcester (BB/W) rats. The Journal of experimental medicine 166: Lundsgaard, D., T. L. Holm, L. Hornum, and H. Markholst In vivo control of diabetogenic T-cells by regulatory CD4+CD25+ T-cells expressing Foxp3. Diabetes 54: Holm, T. L., D. Lundsgaard, and H. Markholst Characteristics of rat CD4(+)CD25(+) T cells and their ability to prevent not only diabetes but also insulitis in an adoptive transfer model in BB rats. Scandinavian journal of immunology 64: Poussier, P., T. Ning, T. Murphy, D. Dabrowski, and S. Ramanathan Impaired post-thymic development of regulatory CD4+25+ T cells contributes to diabetes pathogenesis in BB rats. Journal of immunology (Baltimore, Md. : 1950) 174: Hillebrands, J. L., B. Whalen, J. T. Visser, J. Koning, K. D. Bishop, J. Leif, J. Rozing, J. P. Mordes, D. L. Greiner, and A. A. Rossini A regulatory CD4+ T cell subset in the BB rat model of autoimmune diabetes expresses neither CD25 nor Foxp3. Journal of immunology 177: van den Brandt, J., H. J. Fischer, L. Walter, T. Hünig, I. Klöting, and H. M. Reichardt Type 1 diabetes in BioBreeding rats is critically linked to an imbalance between Th17 and regulatory T cells and an altered TCR repertoire. Journal of immunology (Baltimore, Md. : 1950) 185:
87 299. Visser, J., J. L. Hillebrands, M. Walther Boer, N. a. Bos, and J. Rozing Prevention of diabetes by a hydrolysed casein-based diet in diabetes-prone BioBreeding rats does not involve restoration of the defective natural regulatory T cell function. Diabetologia 52: Hillebrands, J.-L., J. Rozing, and J. T. J. Visser Comment on "Type 1 diabetes in BioBreeding rats is critically linked to an imbalance between Th17 and regulatory T cells and an altered TCR repertoire". Journal of immunology (Baltimore, Md. : 1950) 186: ; author reply
88 Chapter II Loss of T Cell and B Cell Quiescence Precedes the Onset of Microbial Flora- Dependent Wasting Disease and Intestinal Inflammation in Gimap5-Deficient Mice Michael J. Barnes*, Halil Aksoylar, Philippe Krebs*, Tristan Bourdeau, Carrie N. Arnold*, Yu Xia*, Kevin Khovananth*, Isaac Engel, Sosathya Sovath*, Kristin Lampe, Eleana Laws, Amy Saunders, Geoffrey W. Butcher, Mitchell Kronenberg, Kris Steinbrecher, David Hildeman, H. Leighton Grimes, Bruce Beutler* and Kasper Hoebe * Department of Genetics, Scripps Research Institute, La Jolla; La Jolla Institute for Allergy and Immunology, San Diego, CA 92037; Cincinnati Children's Hospital Research Foundation, Cincinnati, OH 45229; and The Babraham Institute, Babraham Research Campus, Cambridge, United Kingdom M.J.B. and H.A. contributed equally to this article. Address correspondence and reprint requests to Kasper Hoebe, Cincinnati Children s Hospital Research Foundation, MLC7021, Room S5.421, 3333 Burnet Avenue, Cincinnati, OH address: kasper.hoebe@cchmc.org The Journal of Immunology April 1, 2010 vol. 184 no
89 Abstract Homeostatic control of the immune system involves mechanisms that ensure the self-tolerance, survival and quiescence of hematopoietic-derived cells. In this study, we demonstrate that the GTPase of immunity associated protein (Gimap)5 regulates these processes in lymphocytes and hematopoietic progenitor cells. As a consequence of a recessive N-ethyl-N-nitrosourea induced germline mutation in the P-loop of Gimap5, lymphopenia, hepatic extramedullary hematopoiesis, weight loss, and intestinal inflammation occur in homozygous mutant mice. Irradiated fetal liver chimeric mice reconstituted with Gimap5-deficient cells lose weight and become lymphopenic, demonstrating a hematopoietic cell-intrinsic function for Gimap5. Although Gimap5- deficient CD4 + T cells and B cells appear to undergo normal development, they fail to proliferate upon Ag-receptor stimulation although NF-κB, MAP kinase and Akt activation occur normally. In addition, in Gimap5-deficient mice, CD4 + T cells adopt a CD44 high CD62L low CD69 low phenotype and show reduced IL-7rα expression, and T- dependent and T-independent B cell responses are abrogated. Thus, Gimap5- deficiency affects a noncanonical signaling pathway required for Ag-receptor induced proliferation and lymphocyte quiescence. Antibiotic-treatment or the adoptive transfer of Rag-sufficient splenocytes ameliorates intestinal inflammation and weight loss, suggesting that immune responses triggered by microbial flora causes the morbidity in Gimap5-deficient mice. These data establish Gimap5 as a key regulator of hematopoietic integrity and lymphocyte homeostasis. 79
90 Introduction Many layers of regulation ensure homeostatic control of the immune system during development and throughout life. In fetal and neonatal mice, hematopoietic stem cells (HSCs) and precursor cells migrate from the fetal liver to the bone marrow and thymus (1). Thereafter, maintenance of the HSC niche involves both cell-extrinsic and cell-intrinsic mechanisms in which responsiveness to growth factors, cell cycle control, and breakdown of metabolic by-products are essential (2). When these processes occur normally, the many diverse lineages of hematopoietic cells are continually generated in the bone marrow and thymus. Given their potential to undergo clonal expansion and long-term survival, additional checkpoints are needed to limit the survival of self-reactive T cells and B cells (3). In T cells, these checkpoints include the induction of apoptosis in thymocytes that recognize self-ags with high affinity (negative selection) (4) and the mitigation of inappropriate immune responses by regulatory T cells (5). The availability of common γ-chain cytokines governs the size and composition of the T cell niche (6-8). Notably, IL-2, IL-7, and IL-15 promote T cell survival by modulating the expression of Bcl-2-family member proteins. Somewhat paradoxically, mutations or conditions that partially impair T cell survival can be associated with the development of immunopathologies (9). One potential explanation for this association is that excess common γ-chain cytokines available in lymphopenic conditions allow the survival and proliferation of self-reactive T cell clones. For example, when naive CD4 + T cells are transferred into Rag-deficient recipient mice, they undergo lymphopenia-induced proliferation (LIP) and adopt a characteristic phenotype similar to memory CD4 + T cells that includes the expression of 80
91 increased amounts of CD44 (CD44 high ) and reduced amounts of CD62L (CD62L low ), whereas CD69 expression remains mostly unaffected (CD69 low ) (8, 10). In some genetic backgrounds, intestinal microbial florae promote the expansion of colitogenic IFN-γ producing CD4 + T cells undergoing LIP, resulting in intestinal inflammation (11). Congenitally lymphopenic Tcrα / mice also develop intestinal inflammation owing to excessive IL-4 dependent Th2 cell and γδ T cell responses (12-14). Lymphopenia has also been linked to the precipitation of diabetes in animal models (15, 16). Increased rates of T cell turnover, altered ratios of regulatory to conventional T cells, and acquisition of effector function by T cells undergoing LIP are all aspects of the lymphopenic environment that might favor the onset of immunopathology. Homozygosity for the lyp mutation causes severe T cell lymphopenia and has been shown to predispose rats to the development of autoimmune diabetes (16) and intestinal inflammation (17). However, lymphopenia caused by the lyp mutation alone is insufficient to trigger immunopathology, which requires the presence of additional modifier alleles at other loci in the genome (17-21). In both susceptible and disease-free genetic backgrounds, homozygous lyp rats largely lack CD8 + T cells and have reduced CD4 + T cell and NKR-P1 + NKT cell numbers (21-23), whereas deficiencies in conventional B cells have not been reported (24). Positional cloning efforts identified the lyp mutation as a frameshift mutation in the gene Gimap5 (also known as Ian4, Ian5, or Irod) (25). The Gimap genes (previously known as immune-associated nucleotide binding proteins [Ian]) comprise a family of genes that are physically clustered in the genome and predominantly expressed in lymphocytes (26). The cellular localization and 81
92 functions of the GTPase of immunity-associated proteins (Gimap) remain nebulous. All Gimap proteins share a GTP-binding AIG1 homology domain, which was originally identified in a gene involved in plant immunity (27). In mammals, evidence suggests that Gimap3 and Gimap5 both promote thymocyte and T cell survival (26, 28-30), whereas Gimap4 can limit T cell viability (31, 32). More recently, Gimap5 / mice were reported to have decreased numbers of peripheral T cells, similar to the lyp rat, and also showed disrupted development of NK cells, reduced survival of Vα14 TCR-expressing invariant NKT (inkt) cells, granulocyte accumulation, increased hepatocyte apoptosis and early mortality (33). Thus, the effect of complete Gimap5-deficiency in mice is much more severe than the truncation of Gimap5 by the lyp mutation in rats. These discrepancies open several new avenues for investigation into the function of Gimap5 in vivo. First, although liver failure caused by hepatocyte apoptosis in Gimap5 / mice was suggested (33), the cause of the early morbidity in these mice remained unclear. In addition, despite the report of decreased B1 cell accumulation in the peritoneum of Gimap5 / mice (33), the effect of Gimap5-deficiency on B cell development was not examined, nor was the role of Gimap5 in thymocyte selection or peripheral T cell and B cell function explored. In this study, we describe a recessive N- ethyl-n-nitrosourea (ENU) induced mutation in Gimap5, which we termed sphinx. Like Gimap5 / mice, sphinx homozygotes are lymphopenic, show granulocyte accumulation, exhibit liver abnormalities, and die by 14 weeks old. Using the sphinx mouse, we demonstrate thatgimap5 serves previously uncharacterized cellintrinsic roles in lymphocyte survival, quiescence, and Ag-receptor induced proliferation, 82
93 and link the early morbidity in sphinx mice to suppressible, microbial flora-dependent, intestinal inflammation and wasting disease. 83
94 Material and Methods Mice and reagents All experiments were performed according to the U.S. National Institutes of Health guidelines and were approved by the Institutional Animal Care and Use Committee of The Scripps Research Institute (TSRI) and The Cincinnati Children s Hospital. BALB/cJ, Bim /, C57BL/6J, Cd4 /, Cd8 /, DBA/2J, FVB/NJ, and NODShiLtJ mice were obtained from The Jackson Laboratory (Bar Harbor, ME). β2m /, C3H/HeN, H-Y TCR transgenic, Rag2 /, and Rag2 / Il2rγ / mice were obtained from Taconic (Germantown, NY).Gimap5 sph/sph mice were generated at TSRI using ENU mutagenesis. Jα18 / mice were housed at the La Jolla Institute for Allergy and Immunology and act-mova transgenic mice were housed at TSRI. All mice were maintained under specific pathogen-free conditions. αgal-cer mouse CD1d tetramers labeled with PE or APC were generated as described previously (34), and all Abs used for flow cytometry were purchased from ebioscience (San Diego, CA) or BioLegend (San Diego, CA). Abs for immunoblotting were purchased from Cell Signaling Technology (Beverly, MA). Purified CD3ε (145-2C11) and CD28 (37.51) abs (ebioscience) were used for T cell activation. F(ab ) 2 anti-igm (Jackson ImmunoResearch), anti-cd40 (ebioscience), ril-4 (R&D Systems, Minneapolis, MN), LPS (Axxora, San Diego, CA) and PMA/ionomycin (Sigma- Aldrich, St. Louis, MO) were used for B cell activation. T cells and B cells were labeled with CFSE by incubating MACS (Miltenyi Biotech, Auburn, CA) purified CD4 + T cells or 84
95 splenic B cells in 5 μm CFSE in PBS with 0.1% FCS for 10 min. ELISAs were used to measure serum cytokine concentrations (ebioscience). In vivo cytotoxicity assay measuring NK cell and CD8 + T cell function We immunized G3 ENU mice with γ-irradiated (1500 rad) act-mova splenocytes. Seven days later, we injected CFSE-labeled cells i.v., consisting of a mixture of three different splenocyte populations (C57BL/6J, β2m /, and SIINFEKLpeptide pulsed C57BL/6J splenocytes) at a ratio of 1:1:1. After 48 h, blood was drawn from the retro-orbital plexus, RBCs were lysed, and the presence or absence of each CFSE-labeled cell population was determined by flow cytometry. mab MAC421 against mouse Gimap5 A glutathione-s-transferase fusion protein containing residues of mouse Gimap5 was produced using Escherichia coli RosettaDE3 bacteria (Novagen, Madison, WI). Purification was performed by elution from glutathione beads and used to immunize a LEW rat. The rat was euthanized 3 d after a final boost with this immunogen, isolated splenocytes were fused with the plasmacytoma cell line Y3Ag1.2.3 (35), and hybridomas were derived by standard methods. The selected clone MAC421 secretes a rat IgG2a Ab specific for Gimap5 that is, it shows no cross-reactivity on other members of the mouse Gimap family. Separate studies have shown that the epitope recognized by MAC421 is dependent on residues 1 10 at the NH2 terminus of mouse Gimap5 (C. Carter, G.W. Butcher, T. Nitta, K. Yano, and Y. Takahama, unpublished observations). 85
96 Complete blood counts Blood samples were taken from the retro-orbital plexus of mice at indicated ages and analyzed using a Hemavet 950 veterinary hematology system Fisher Scientific (Pittsburgh, PA). Hepatic lymphoid cell isolation Mice were euthanized using CO 2, and livers were flushed with ice cold PBS via the hepatic portal vein using a 25-gauge needle and syringe. After excision of the liver, the gall bladder was removed and livers were diced using scissors. Further homogenization was achieved using a sieve and plunger, and liver fragments were gently passed through a 100-μM strainer and collected in a 50-ml centrifuge tube. Total liver cells were washed with 40 ml ice-cold RPMI 1640/5% FBS and centrifuged for 10 min at 300 g. The resulting pellets were resuspended in digestion medium (RPMI 1640 containing 0.02% [w/v] collagenase IV [Sigma-Aldrich] and 0.002% [w/v] DNase I [Sigma-Aldrich]) and left at 37 C for 40 min under gentle shaking. Ice cold serum free medium was added to each tube, and cells were centrifuged for 3 min at 30 g and 4 C. Supernatants containing lymphocyte-enriched cell populations were collected and centrifuged at 300 g for 10 min and 4 C. Subsequently, cell pellets were resuspended in PBS containing 0.5% FBS and labeled with the mouse Lineage Cell Depletion Kit (Miltenyi Biotec). Lin cells were then separated using an AutoMacs magnetic sorter (Miltenyi Biotec), and various HSCs or precursor populations were identified by flow cytometry as described before (36). 86
97 Ab responses For T-dependent Ab responses, age- and sex-matched mice were immunized i.p. with 5 μg LPS (Axxora) and 50 μg of NP36-CGG (Biosearch Technologies, Novato, CA) mixed 1:1 with alum (Pierce, Rockford, IL). For T-independent Ab responses, mice were immunized i.p. with 50 μg NP50-Ficoll (Biosearch Technologies). Serum NP-specific Abs were detected by ELISA. Briefly, 96-well round-bottom plates (Fisher Scientific) were coated with 5 μg/ml NP30-BSA (Biosearch Technologies) for detection of NPspecific Ig. Serum samples from immunized mice were serially diluted in 1% milk and added to coated or uncoated (to determine background) wells. Plates were incubated with HRP-conjugated goat anti-mouse IgM or IgG1 (Southern Biotechnology Associates, Birmingham, AL), developed with Peroxidase Substrate and Stop Solution (KPL), and measured for absorbance at 450 nm. Immunoblotting Immunoblotting was performed using total spleen, thymus, or bone marrow lysates, or splenic B cells that were isolated using MACS purification with anti-cd19 microbeads (Miltenyi Biotech). Cell lysis and immunoblotting were performed as described previously (37). Cell cycle analysis of splenic B cells Splenic B cells were cultured with complete IMDM (containing 10% FCS and 1% P/S) and left untreated or treated with 0.1 μg/ml LPS at 37 C, 5% CO 2. After 48 h, cells were collected in 1 ml PBS and subsequently were added dropwise to 2.5 ml absolute ethyl alcohol. After 15 min fixation on ice, cells were washed with PBS and 87
98 incubated in 500 μl propidium iodide (PI) solution (50 μg/ml PI, 0.1 mg/ml RNase A, and 0.05% Triton X-100 in PBS) for 40 min at 37 C. Cells were washed and resuspended in PBS and subsequently analyzed by flow cytometry. Antibiotic treatment and survival assays Gimap5 +/sph breeding pairs received antibiotic treatment (1 g/l ampicillin, 500 mg/l vancomycin, 1 g/l neomycin sulfate, and 1 g/l metronidazole) administered orally via drinking water. Subsequent litters remained on antibiotic treatment until analysis at 9- wk-old. For adoptive transfer studies, Gimap5 sph/sph mice at d old were injected i.v. with splenocytes that had been depleted of RBCs (RBC lysis buffer; Sigma- Aldrich). Mice were monitored and euthanized according to veterinarian recommendations when they became moribund. Statistical analysis Data were analyzed using the GraphPad Prism4 software (GraphPad Software, San Diego, CA). Unless indicated otherwise, statistical significance of the differences among groups was determined from the mean and SD by Student s two-tailed test or by ANOVA followed by Dunnett s test for three or more groups. All data were considered statistically significant if p <
99 Results sphinx: an ENU-induced germline mutation causing severe lymphopenia We identified sphinx in a forward genetic screen designed to detect defective in vivo cytotoxic NK cell and CD8 + T cell responses in G3-ENU mutagenized mice (38). The original sphinx founder, previously immunized with OVA-expressing cells, was unable to reject adoptively transferred NK cell or CD8 + T cell target cells (Fig. 1A). Necropsy revealed a near absence of CD8 + T cells and NK cells in the peripheral lymphoid organs (Fig. 1B, 1C) and a marked reduction in total T cell numbers. However, relatively normal thymocyte development occurred, including the CD4 + T cell, CD8 + T cell, γδ T cell, Foxp3 + regulatory T cell, and inkt cell lineages (data not shown), whereas the bone marrow contained a high frequency of NK cells expressing the activation marker CD69 (Supplemental Fig. 1). Most strikingly, sphinx livers exhibited an abnormal morphology that presented itself as extramedullary hematopoiesis with associated foci of hematopoietic cells (Fig. 1D, 1E). Hyperplastic nodules developed in adult livers (Fig. 1D) that contained well-differentiated hepatocytes bordered by clusters of hematopoietic cells (Fig. 1E). Hepatocytes from sphinx mice showed no growth advantage in cell culture and no dysplastic liver cells were observed throughout this study (M. Barnes and K. Hoebe, unpublished observations). In addition, when sphinx mice (C57BL/6J genetic background) were crossed with mice of one of five different genetic backgrounds (BALB/cJ, C3H/HeN, DBA/2J, FVB/NJ, or NOD/ShiLtJ) for two generations and then intercrossed, the liver phenotype remained consistent and dysplastic cells were not observed in any other organs (data not shown). Thus, it appeared that sphinx livers exhibited an abnormal morphology owing to extramedullary 89
100 hematopoiesis and reactive hyperplasia of hepatocytes, rather than a hepatic neoplasia. In addition, sphinx mice progressively developed normocytic anemia (Fig. 1F, Supplemental Fig. 2A), with increased variation in RBC width (Supplemental Fig. 2B) and thrombocytopenia (Supplemental Fig. 2C). This was correlated with a rapid reduction in circulating blood lymphocytes (Fig. 1G) and concomitant neutrophilia (Supplemental Fig. 2D). These observations indicated that the sphinx mutation affected a gene essential for lymphocyte maintenance and normal hematopoiesis. Positional cloning of the sphinx mutation To identify the causative mutation, we used a positional cloning strategy (39). Using 13 meioses from sphinx mice backcrossed to the C3H/HeN strain, we linked the NK cell and CD8 + T cell deficiency to chromosome 6 with a peak LOD score of 4.0 (Supplemental Fig. 3A). Further fine mapping with 572 meioses derived from F1 intercrosses confined the mutation to a 2.9-megabase critical region, bounded by the microsatellite markers D6mit315 and D6mit276. We amplified and sequenced all annotated and predicted coding base pairs in the critical region, obtaining high-quality sequence for 89.9% of total coding base pairs and 100% of coding base pairs for genes in the Gimap cluster. Among bp sequenced, a single G T point mutation in the Gimap5 coding region differed between the sphinx and C57BL/6J genomes (Supplemental Fig. 3B, 3C). Gimap5 is predominantly transcribed in lymphocytes (26), but is also expressed in HSCs (40). The sphinx mutation appeared to destabilize the Gimap5 protein in hematopoietic tissues, including total bone marrow cells and splenic B cells, resulting in a lack of detectable protein (Fig. 1H), whereas mrna expression of Gimap5 remained unaffected (Fig. 1I). 90
101 The sphinx mutation resulted in the amino acid substitution G38C, which changed an amino acid that is highly conserved within all mouse Gimap homologs and within all annotated orthologous genes containing an AIG1 domain, in kingdoms as distant as Planta and Protista (Supplemental Fig. 3D). This residue is located in the predicted P-loop of the AIG1 domain of Gimap5, in the pocket that binds and hydrolyzes GTP. Previous studies of the p21ras GTPase have shown that mutation of the analogous Gly residue to Val (G15V) resulted in a protein that was unable to hydrolyze GTP or to transform NIH 3T3 cells when overexpressed (41). The mutation of this residue to Cys might destabilize Gimap5 by altering the disulphide bonding patterns of the mutant Gimap5 protein. Whereas no Gimap5 protein was detected by immunoblotting, any residual intact protein below the limit of detection would be predicted to be nonfunctional because of the mutation of the conserved Gly residue. Overall, the sphinx phenotype mirrors the phenotype of previously described Gimap5 / mice (33), indicating that the sphinx mutation effectively results in a Gimap5-null animal. Lethal wasting disease, colitis, and hepatic extramedullary hematopoiesis in sphinx mice From birth until weaning, conventionally housed sphinx (hereafter Gimap5 sph/sph ) mice appeared outwardly healthy. After 4 wk, males showed a reduced growth rate (Fig. 2A) and developed diarrhea. Although significant, weight loss in females was less dramatic and only occurred after 6 wk (data not shown). In male and female Gimap5 sph/sph mice, cell infiltration and inflammation were observed in the colon by 6 wk. Severe colitis, exemplified by goblet cell depletion, leukocyte infiltration into the 91
102 lamina propria, epithelial cell hyperplasia, and crypt loss, developed by 10 wk (Fig. 2B). Wasting disease and intestinal inflammation likely contributed to the early morbidity of Gimap5 sph/sph mice, which generally occurred by 14 wk. Liver dysfunction remained another possible factor contributing to early morbidity. To better understand the onset of liver abnormalities, we investigated the development of the embryonic and neonatal liver. At the embryonic day (ED) 16 developmental stage and at birth, livers from Gimap5 sph/sph mice and their heterozygous littermates appeared similar (Fig. 2C). However, more hematopoietic cells remained in Gimap5 sph/sph livers at day 8. By day 14, extramedullary hematopoiesis could be observed before the initiation of nodule formation. Extramedullary hematopoiesis, which persisted in adult mice, was limited to the liver (Fig. 3A), although a disrupted splenic architecture that included reduced follicle size and number and granulocyte accumulation was also observed (data not shown). Schulteis et al. (33) argued that Gimap5 / mice die as a consequence of liver dysfunction and hepatocyte apoptosis. Despite the abnormal liver morphology, we found normal concentrations of serum bilirubin and albumin in 8-wkold Gimap5 sph/sph mice (data not shown), suggesting that the metabolic functions of the liver remained largely intact. Cell-intrinsic hematopoietic defects in Gimap5 sph/sph mice We next considered that aberrant hematopoiesis rather than liver failure might contribute to wasting disease and mortality in Gimap5 sph/sph mice. Despite the liver abnormalities, the bone marrow of 5-wk-old Gimap5 sph/sph mice contained normal numbers of Lin Sca1 + c-kit + HSCs and hematopoietic progenitor cells (Supplemental 92
103 Fig. 4 A, B). However, the numbers of megakaryocyte/erythroid progenitors (MEPs) declined with age, in correlation with the onset of anemia and thrombocytopenia in adult Gimap5 sph/sph mice (Fig. 1F, Supplemental Fig. 2C). In addition, despite normal numbers of common lymphoid progenitors (CLPs) in the bone marrow, rapid atrophy of the thymus occurred with age (data not shown). More dramatic differences were observed in the Gimap5 sph/sph liver. Examination of hematopoietic progenitor cell populations revealed that a significant number of HSCs and hematopoietic precursors, representing all hematopoietic lineages, remained in the adult Gimap5 sph/sph liver (Fig. 3A). Compared to normal hematopoiesis in the bone marrow, extramedullary liver hematopoiesis in Gimap5 sph/sph mice was skewed toward generation of Lin Sca1 + c-kit - IL-7rα + CLPs, and it is noteworthy that IL-7rα expression was increased in this population (Fig. 3A). To investigate the functionality of Gimap5 sph/sph HSCs, we made radiation bone marrow and fetal liver chimeras. Similar to what was reported for Gimap5 / mice (33), Gimap5 sph/sph bone marrow cells poorly reconstituted the hematopoietic compartment of lethally irradiated congenic recipient mice (data not shown), suggesting functional impairment. To exclude the possibility that defective HSC function was a secondary consequence of the inflammation and wasting disease that occurs in Gimap5 sph/sph mice, we reconstituted lethally irradiated Rag2 / Il2rγ / recipients with Gimap5 sph/sph or Gimap5 sph/+ littermate control ED 19 fetal liver cells. The recipients were chosen because they lacked endogenous T cells, B cells, and NK cells, but could support the development of a normal lymphocyte compartment from wild type HSCs. Six weeks after transfer, mice reconstituted with Gimap5 sph/sph fetal liver cells were severely 93
104 lymphopenic and had substantially fewer thymocytes than mice reconstituted with Gimap5 sph/+ fetal liver cells. In the spleen, CD8 + T cell, B cell, and NK cell numbers were markedly reduced compared with recipients of control fetal liver cells, whereas CD4 + T cells were present at normal numbers (Fig. 3B). Gimap5 sph/sph fetal liver cell recipients exhibited weight loss and wasting, whereas control fetal liver cell recipients appeared healthy (data not shown). In Gimap5 sph/sph fetal liver cell recipients, CD11b + Gr1 + myeloid cells accumulated in the spleen (Fig. 3B), whereas liver abnormalities were not observed. The transfer of weight loss and wasting disease, but not liver abnormalities, by Gimap5 sph/sph fetal liver cells suggested that the early mortality observed in Gimap5 sph/sph mice is at least partially caused by defective hematopoiesis. Furthermore, the Gimap5 sph/sph fetal liver cells exhibited an intrinsic defect in lymphopoiesis, affecting T cells, B cells, and NK cells. Reduced survival and IL-7rα expression in Gimap5 sph/sph T cells Several mouse models have linked impaired lymphocyte function and lymphopenia with the development of various manifestations of immunopathology (9, 15). To better understand the contribution of lymphocytes to the hematopoietically transferrable abnormalities observed in Gimap5 sph/sph mice, we investigated lymphocyte development, survival and function. Roles for Gimap5 in T cells have been proposed for thymic selection, thymic export, and survival in the periphery (16). Like Gimap5 lyp/lyp rats and Gimap5 / mice, 6-wk-old Gimap5 sph/sph mice exhibited normal thymus cellularity and frequencies of DN1-4, DP, CD4 SP, and CD8 SP thymocyte subpopulations (data not shown). To examine thymic selection in Gimap5 sph/sph thymocytes, we bred the mutation onto the H-Y TCR transgenic background. The H-Y TCR recognizes an 94
105 endogenous peptide presented by MHC class I molecules that is derived from the H-Y Ag, which is expressed only in male mice. Consequently, H-Y TCR + thymocytes are normally deleted by negative selection in male mice, whereas they develop into mature CD8 + T cells in female mice. In Gimap5 sph/sph males, H-Y reactive thymocytes were deleted, implying that the mutation did not impede negative selection (Fig. 4A). In contrast, the thymus of Gimap5 sph/sph females contained normal numbers of H-Y TCR + DP thymocytes, but significantly fewer H-Y TCR + CD8 SP thymocytes (Fig. 4A), suggesting that positively selected thymocytes became more sensitive to proapoptotic negative selection cues in the absence of Gimap5. H-Y TCR + CD8 + T cells were found at reduced numbers and frequencies in the spleen of both male and female Gimap5 sph/sph mice (Fig. 4B), indicating that lymphopenia occurred regardless of TCR specificity for self-ag. We next examined the expression of molecules important for thymic egress and peripheral accumulation of T cells. Normally, terminally differentiated TCRβ + CD4 SP and CD8 SP thymocytes upregulate CD5 and IL-7rα (CD127) and downregulate CD24 (HSA) and CD69 (4, 42). Gimap5 sph/sph CD8 SP, but not CD4 SP thymocytes, failed to modulate CD24, CD69, or IL-7rα expression (Fig. 4C), suggesting that the mutation either blocked terminal CD8 SP thymocyte maturation or impaired survival of CD8 SP cells. Downregulation of CD69 expression has been associated with the ability of thymocytes to exit the thymus (43), and it is possible that export of CD8 SP thymocytes is impaired in Gimap5 sph/sph mice. In addition, as IL-7 promotes naive T cell survival by modulating expression of anti-apoptotic molecules (44), reduced IL-7rα expression 95
106 in Gimap5 sph/sph CD8 SP thymocytes likely contributes to the failure of CD8 + T cells to survive and accumulate in the periphery. CD4 + T cells isolated from the spleen of 6-wk-old Gimap5 sph/sph mice showed a significant reduction in IL-7rα surface expression (Fig. 4D) and were unable to proliferate upon TCR stimulation (Fig. 4E). Thus, impaired IL-7 signaling and TCRinduced proliferation represent two contributing factors to the CD4 + T cell lymphopenia in Gimap5 sph/sph mice. As the lymphopenia became more pronounced with age (Fig. 1G), CD4 + T cells began to express cell-surface markers characteristic of cells undergoing LIP, including reduced amounts of CD62L but normal, low expression of CD69 (Fig. 4F). We also noted that peripheral Gimap5 sph/sph inkt cells expressed reduced amounts of NK1.1 (Fig. 5A) and rapidly declined in number with age. NK1.1 expression is acquired at a late step in inkt cell differentiation, most typically after export from the thymus, and evidence suggests that it requires engagement of the invariant TCR with the CD1d Ag-presenting molecule (45). Upon in vivo activation of inkt cells with the agonist αgal-cer, we detected little intracellular IFN-γ or TNF-α, reduced amounts of IL- 4 and normal amounts of IL-13 on a per cell basis (Fig. 5B). We confirmed these results by ELISA (Fig. 5C). The impaired survival of inkt cells and aberrant inkt cell cytokine response that occurs in Gimap5 sph/sph mice suggests roles for Gimap5 in both lymphocyte survival and function. 96
107 Functional defects in Gimap5 sph/sph B cells Before the onset of wasting disease, splenic CD19 + B cells in Gimap5 sph/sph mice, unlike T cells and NK cells, were less reduced in number compared with C57BL/6J littermates. However, the B cell compartment contained fewer mature B cells (Fig. 6A), whereas the percentages of marginal zone B cells (Fig. 6B) and follicular B cells (Fig. 6C) were similar to C57BL/6J littermates. Between 6 10 wk, splenic B cell numbers declined in Gimap5 sph/sph mice (Supplemental Fig. 5A). In the peritoneal cavity, few B1 cells were found (M. Barnes and K. Hoebe, unpublished data), suggesting that B1 cells either fail to develop or survive. In addition, the serum of 8-wk-old Gimap5 sph/sph mice contained 75% less IgM and 50% less total IgG than heterozygous littermates (Supplemental Fig. 5B). The reduced total IgG reflected significant reductions in concentrations of the isotypes IgG1, IgG2b, and IgG3 and a trend toward reduced IgG2a (Supplemental Fig. 5B). These abnormalities suggested the possibility that B cell function was impaired in Gimap5 sph/sph mice. To assess B cell function, we immunized 6- wk-old mice with T-dependent (NP-CGG + alum + LPS) or T-independent (NP-ficoll) Ags. Gimap5 sph/sph mice failed to produce either T-dependent Ag-specific IgG1 Abs (Fig. 6D) or T-independent Ag-specific IgM responses (Fig. 6E), suggesting that Gimap5 sph/sph B cells might have an intrinsic functional defect. To investigate B cellintrinsic signaling, we isolated B cells from 6-wk-old Gimap5 sph/sph and C57BL/6J mice and determined their capacity to proliferate upon activation in vitro. Gimap5 sph/sph B cells failed to proliferate after BCR stimulation, or treatment with the diacylglycerol mimetic, PMA, and the Ca ++ mobilizing agent ionomycin (Fig. 6F). However, alternative pathways 97
108 triggered by LPS or CD40 ligation induced normal Gimap5 sph/sph B cell proliferation (Fig. 6F). Activation of the NF-κB, MAP kinase, and Akt pathways is essential for either BCR or PMA and ionomycin induced proliferation. We examined the activation of these pathways in Gimap5 sph/sph B cells stimulated with PMA and ionomycin. Normal degradation of the NF-κB inhibitor IκB and phosphorylation of the Erk, Jnk, and p38 MAP kinases occurred (Supplemental Fig. 5C). In addition, normal activation of Akt, marked by phosphorylation of serine and threonine residues, and other proximal molecules in the Akt pathway was observed (Supplemental Fig. 5D). Nonetheless, Gimap5 sph/sph B cells failed to proliferate upon PMA and ionomycin activation. Ex vivo cell cycle analysis of Gimap5 sph/sph B cells detected an increased percentage of B cells in S phase and a reduced number of B cells in G2 phase. When proliferation was induced with LPS, an equivalent number of Gimap5 sph/sph B cells entered G2 phase, yet an increased percentage of cells remained in S phase (Fig. 6G), suggesting that defective cycling might cause the decline in peripheral B cell numbers in Gimap5 sph/sph mice (Supplemental Fig. 5A). Overall, we found no major defects in proximal mitogenic signaling or cell cycle entry, but we did find aberrant cell cycle progression in Gimap5 sph/sph B cells. Prevention of wasting disease and intestinal inflammation, but not lymphopenia, by antibiotic treatment Microbial flora in the intestine represents the major reservoir of foreign Ags and exogenous innate immune stimuli in specific pathogen-free housed mice. Localized 98
109 inflammation in the gastrointestinal tract of Gimap5 sph/sph mice suggested the possibility that aberrant responses triggered by microbial flora contributed to systemic CD4 + T cell LIP and subsequent cell death, ultimately resulting in lymphopenia. Thus, we sought to address the role of microbial flora in Gimap5 sph/sph mice. Homozygotes and heterozygote littermates were put on a continuous antibiotic regimen from birth until 9 wk, when they were analyzed. Interestingly, antibiotic treatment did not prevent lymphopenia (Fig. 7A), the expression of CD44 high CD62L low markers by CD4 + T cells (Fig. 7B) or the occurrence of hepatic extramedullary hematopoiesis and hepatocyte hyperplasia (Fig. 7C). Antibiotic-treatment completely blocked the accumulation of CD11b + myeloid cells (Fig. 7A), ameliorated intestinal inflammation (Fig. 7C), and prevented weight loss (data not shown) in Gimap5 sph/sph mice. These data suggest that the Gimap5-dependent hematopoietic cell-intrinsic phenotypes, including hepatic extramedullary hematopoiesis, lymphopenia, and acquisition of a CD44 high CD62L low phenotype by CD4 + T cells, can occur in the absence of intestinal inflammation, whereas microbial florae promote granulocyte accumulation, colitis, and wasting disease. Adoptively transferred lymphocytes reduce extramedullary hematopoiesis and early mortality in Gimap5 sph/sph mice To further assess the involvement of lymphocytes in the immunopathology observed in Gimap5 sph/sph mice, we adoptively transferred congenically labeled C57BL/6J splenocytes into young Gimap5 sph/sph mice before the onset of wasting disease. Strikingly, recipient Gimap5 sph/sph mice did not succumb to early death (Fig. 8A) or develop wasting disease (Fig. 8B). Adoptively transferred splenocytes filled the 99
110 CD8 + T cell niche (Fig. 8C) and promoted the maintenance of endogenous Gimap5 sph/sph CD4 + T cells (Fig. 8D). In the CD4 + T cell compartment, cells of Gimap5 sph/sph origin retained a CD44 high CD62L low phenotype (Fig. 8E). Thus, adoptively transferred splenocytes promoted the accumulation of endogenous Gimap5 sph/sph CD4 + T cells, but not CD8 + T cells. We also examined whether adoptively transferred C57BL/6J splenocytes affected the extramedullary hematopoiesis in recipient Gimap5 sph/sph mice. In the liver of untreated 15-wk-old Gimap5 sph/sph mice, an abundance of common myeloid progenitors were present, including both granulocyte/macrophage progenitors (GMPs) and MEPs. In the livers of age-matched Gimap5 sph/sph mice that received adoptively transferred C57BL/6J splenocytes, clusters of hematopoietic cells, nodules, and extramedullary hematopoiesis were still observed, but at a reduced frequency (Supplemental Fig. 6A, 6B) and fewer common myeloid progenitor, MEP, and CLP cells were found (Fig. 8F, Supplemental Fig. 6B). In addition, the bone marrow of aged Gimap5 sph/sph mice showed enhanced granulopoiesis and reduced erythropoiesis (Fig. 8G). These changes in medullary hematopoiesis likely contributed to the neutrophilia (Supplemental Fig. 2D) and anemia (Fig. 1G) observed in adult Gimap5 sph/sph mice. Recipients of adoptively transferred splenocytes maintained normal numbers and frequencies of GMP and MEP in the bone marrow (Fig. 8G), indicating that transferred cells also affected medullary hematopoiesis. Accordingly, recipient Gimap5 sph/sph mice did not become anemic or exhibit thymic atrophy (data not shown). Enhanced granulopoiesis and thymic atrophy are often observed in conditions of chronic inflammation, and their amelioration by 100
111 either antibiotics or the adoptive transfer of splenocytes suggests that they are most likely triggered by the inflammatory environment present in Gimap5 sph/sph mice. Finally, we observed no reduction in mortality or wasting disease when Gimap5 sph/sph mice received Rag2 / splenocytes, indicating that a lymphocyte population is required to prevent wasting disease (Fig. 8A). Because adoptively transferred Cd4 /, Cd8 /, orjα18 / (inkt cell-deficient) splenocytes, or anti-nk1.1 Ab depleted splenocytes could each prevent weight loss and early mortality (data not shown), multiple lymphocyte populations might contribute to the suppression of wasting disease. 101
112 Discussion Our identification and subsequent characterization of the sphinx mutation confirms the nonredundant role of Gimap5 in lymphocyte survival and normal hematopoiesis (33). The Gimap5 sph/sph mouse exhibits a number of hematologic abnormalities. From birth, peripheral CD8 + T cells and NK cells fail to accumulate, whereas HSCs accumulate in the liver, suggesting that Gimap5 has a direct role in controlling these processes. After weaning, intestinal inflammation, weight loss, severe lymphopenia, and acquisition of a CD44 high CD62L low CD69 low phenotype characteristic of cells undergoing LIP by CD4 + T cells all occur before premature death at 14-wk-old. We used several approaches to examine the relationships between these phenotypes. Fetal liver HSCs, which were not exposed to the inflammatory environment present in adult Gimap5 sph/sph mice, could transfer wasting disease and lymphopenia to lethally irradiated recipients, demonstrating an HSC intrinsic function for Gimap5. Wasting disease and colitis were mitigated by adoptively transferred lymphocytes, indicating that either defective immune regulation or a lymphopenic environment, or both, contributed to the onset of intestinal inflammation. Antibiotic-sensitive microbes were also required to trigger granulocyte accumulation, wasting disease and intestinal inflammation, but not lymphopenia or accumulation of CD44 high CD62L low CD69 low CD4 + T cells. Before the onset of post-weaning abnormalities, extramedullary hematopoiesis occurred in the Gimap5 sph/sph neonatal liver. Defective HSC egress or migration from the neonatal liver could underlie this phenomenon. One possibility that we favor is that Gimap5 sph/sph HSCs become activated in the neonatal liver, thereby halting their migration and inducing extramedullary hematopoiesis. For example, HSC express Toll- 102
113 like receptors and can undergo myelopoiesis at extramedullary sites in response to LPS-induced inflammation (46, 47). Gimap5 might act as a negative regulator of HSC activation, initiated by TLR agonists or other stimuli, that maintains quiescence of the HSC niche. In young Gimap5 sph/sph mice, these abnormalities appear to be limited to the liver, and hematopoietic abnormalities in the bone marrow were observed only in older mice, concurrent with granulocyte accumulation and chronic inflammation. Further studies will be needed to determine whether Gimap5 sph/sph bone marrow HSCs have reduced reconstituting capacity because of the chronically inflamed Gimap5 sph/sph environment or because of a cell-intrinsic requirement for Gimap5 that becomes more pronounced with age. In addition to aberrant hematopoiesis, intestinal inflammation contributed to the weight loss and early morbidity in Gimap5 sph/sph mice. Spontaneous mouse models of colitis have proven valuable in identifying molecules and mechanisms involved in the pathogenesis of inflammatory bowel disease (48). An abundance of IL-13 producing NKT cells are found in the intestinal lamina propria of human ulcerative colitis patients (49), and IL-13 producing inkt cells can drive colitis in a mouse model induced by the chemical oxazolone (50). Our data suggest that the Gimap5 sph/sph environment favors the polarization of inkt cells toward the production of IL-13. Although the cellular and molecular basis of intestinal inflammation in Gimap5 sph/sph mice demands further dissection, IL-13 producing inkt cells may represent one contributing factor. Another potential influence predisposing Gimap5 sph/sph mice to developing colitis is CD4 + T cells undergoing LIP. In the naive CD4 + T cell transfer model of colitis, using C57BL/6 Ragdeficient recipient strains, the lymphopenic environment drives CD4 + T cells into LIP. 103
114 These CD4 + T cells undergoing LIP acquire effector function and cause both intestinal inflammation and wasting disease (11). Of note, intestinal inflammation is significantly reduced when naive T cells are transferred into germ-free mice, suggesting that microbial florae are necessary for colitogenic immune responses in this model (51). The presence of microbial flora might similarly promote the acquisition of effector function among intestinal CD4 + T cells undergoing LIP in Gimap5 sph/sph mice. Interestingly, in partially lymphopenic settings such as in sublethally γ-irradiated mice, colitis is not observed after naive T cell transfer (8). Residual radioresistant intestinal lymphocytes that enhance intestinal barrier function, such as IgA-secreting plasma cells or γδ T cells, might limit colitogenic CD4 + T cell responses in this setting (8, 52). In Gimap5 sph/sph mice, the loss of non-cd4 + lymphocytes with age might generate conditions that promote CD4 + T cells to undergo LIP, and thereby contribute to the onset of intestinal inflammation. Our findings that Gimap5 sph/sph HSCs were unable to reconstitute the B cell compartment of irradiated recipients, and that Gimap5 sph/sph B cells were unable to undergo Ag-receptor induced proliferation or generate Ag-specific IgG or IgM responses were especially surprising. In the lyp rat model, lyp bone marrow cells are able to normally reconstitute the B cell compartment of irradiated recipients (30) and gross B cell functional defects are not observed (24). This discrepancy might reflect different requirements for Gimap proteins in rats and mice. The Gimap gene cluster has undergone divergent evolution between rodent species, including changes in the number of Gimap genes, amino acid sequence variation between homologous Gimap proteins, and differential Gimap gene expression patterns in lymphocytes (26, 53). To 104
115 characterize better the consequences of Gimap5-deficiency in mouse B cells, we first considered that Gimap5 might be involved in the activation of the NF-κB and MAP kinase signaling pathways by BCR stimulation, because others had reported that these pathways are dysregulated in lyp T cells (54). However, Gimap5 sph/sph mice have normal percentages of marginal zone B cells, which require NF-κB signaling to develop (55), and display normal IκB degradation and MAP kinase phosphorylation upon activation. Normal activation of the PI3K/Akt pathway, which is similarly required for lymphocyte proliferation, also occurs in Gimap5 sph/sph B cells. One interesting attribute of Gimap5 sph/sph B cells is their accumulation in S phase in vivo. The role for Gimap5 in cell cycle regulation appears to be especially important in the bone marrow or fetal liver cell reconstitution setting, in which donor cells must undergo rapid proliferation to reconstitute the B cell niche. In summary, Gimap5 impairs BCR-induced proliferation and Ig responses independently of activation of the canonical BCR signaling pathways and may link BCR triggering to cell cycle progression. Although the requirement for Gimap5 in TCR-dependent T cell proliferation and survival has been known for some time, our study identifies several important mechanisms that fail to operate in Gimap5 sph/sph T cells. In the thymus, developing Gimap5 sph/sph CD8 SP thymocytes show impaired survival after positive selection and reduced expression of IL-7rα, suggesting that Gimap5 modulates survival thresholds during thymic selection as well as the expression of a critical cytokine receptor for T cell survival. Although CD4 SP thymocytes express normal amounts of IL- 7rα, expression is reduced in the periphery among CD4 + T cells. These findings provide a partial molecular basis for previous observations made in the lyp rat (16) 105
116 and Gimap5 / mouse (33). In these strains, and in Gimap5 sph/sph mice, CD8 + T cells and naive CD4 + T cells, which are more dependent on IL-7 for their survival than Agexperienced or regulatory CD4 + T cells (56), are most affected by Gimap5-deficiency. As discussed below, IL-7 independent mechanisms regulating T cell survival also appear to require Gimap5. The net effect of Gimap5 deficiency in CD4 + T cells is to promote the accumulation of cells with a CD44 high CD62L low LIP-like phenotype with the potential to cause immunopathology. Importantly, other factors such as MHC haplotype and modifier alleles are needed to cause diabetes (18-20) and, as we have shown in this study, microbial florae are required for the onset of colitis. Finally, although peripheral CD4 + Foxp3 + regulatory T cell numbers are minimally affected in Gimap5 sph/sph mice, it is not known whether they function and express a normal TCR repertoire, or whether the lymphopenic environment inhibits their ability to control aberrant immune responses. Alternatively, a Foxp3 regulatory lymphocyte population might be absent or defective in Gimap5 sph/sph mice. Our data identify a critical role for Gimap5 in T cell, B cell, NK cell, inkt cell and HSC survival and function. A previous study reported that Gimap5 might mediate its effects in part by interacting directly with Bcl-2 and a related anti-apoptotic protein Bclx L (26). IL-7rα signaling promotes expression of Bcl-2, which can directly inhibit the proapoptotic activity of Bim (7), and indeed genetic loss of Bim restores the T cell compartment in IL-7r-deficient mice (57). Therefore, we considered that Gimap5 sph/sph Bim / mice might exhibit less lymphopenia if reduced IL-7rα expression alone prevented T cell accumulation. However, combined Gimap5 and Bim deficiency did not prevent lymphopenia (H. Aksoylar and K. Hoebe, unpublished observations), 106
117 suggesting that additional survival pathways are perturbed in the absence of Gimap5. Gimap5 may be more than just a prosurvival protein, as it appears to also maintain quiescence in lymphoid cells and HSCs. In support of this possibility, we have found that negative selection thresholds are reduced in thymocytes, HSCs become aberrantly activated in the neonatal liver, and bone marrow NK cells show an activated phenotype. Among B cells, Gimap5 deficiency might in some ways mimic B cell anergy, which can be induced by constant low-affinity BCR stimulation (58), and might favor the accumulation of B cells in cell cycle arrest. Thus, Gimap5 sph/sph B cells can proliferate in response to LPS or CD40 stimulation, but, like anergic B cells, fail to respond to BCR stimulation alone. In vivo, activation by weak stimuli that are normally ignored could abrogate quiescence, promoting the accumulation of lymphocytes in cell cycle arrest and driving HSCs to undergo extramedullary hematopoiesis. Future studies will explore the role of pathways important for setting activation thresholds and enforcing quiescence in Gimap5 sph/sph mice. 107
118 References 1. Dzierzak, E., and N. A. Speck Of lineage and legacy: the development of mammalian hematopoietic stem cells. Nature immunology 9: Tothova, Z., and D. G. Gilliland FoxO transcription factors and stem cell homeostasis: insights from the hematopoietic system. Cell stem cell 1: Goodnow, C. C Multistep pathogenesis of autoimmune disease. Cell 130: Palmer, E Negative selection--clearing out the bad apples from the T-cell repertoire. Nature reviews. Immunology 3: Feuerer, M., J. A. Hill, D. Mathis, and C. Benoist Foxp3+ regulatory T cells: differentiation, specification, subphenotypes. Nature immunology 10: Ma, A., R. Koka, and P. Burkett Diverse functions of IL-2, IL-15, and IL-7 in lymphoid homeostasis. Annual review of immunology 24: Marrack, P., and J. Kappler Control of T cell viability. Annual review of immunology 22: Surh, C. D., and J. Sprent Homeostasis of naive and memory T cells. Immunity 29: Liston, A., A. Enders, and O. M. Siggs Unravelling the association of partial T-cell immunodeficiency and immune dysregulation. Nature reviews. Immunology 8: Murali-Krishna, K., and R. Ahmed Cutting edge: naive T cells masquerading as memory cells. Journal of immunology (Baltimore, Md. : 1950) 165: Coombes, J. L., N. J. Robinson, K. J. Maloy, H. H. Uhlig, and F. Powrie Regulatory T cells and intestinal homeostasis. Immunological reviews 204: Iijima, H., I. Takahashi, D. Kishi, J. K. Kim, S. Kawano, M. Hori, and H. Kiyono Alteration of interleukin 4 production results in the inhibition of T helper type 2 cell-dominated inflammatory bowel disease in T cell receptor alpha chaindeficient mice. The Journal of experimental medicine 190: Mombaerts, P., E. Mizoguchi, M. J. Grusby, L. H. Glimcher, A. K. Bhan, and S. Tonegawa Spontaneous development of inflammatory bowel disease in T cell receptor mutant mice. Cell 75:
119 14. Nanno, M., Y. Kanari, T. Naito, N. Inoue, T. Hisamatsu, H. Chinen, K. Sugimoto, Y. Shimomura, H. Yamagishi, T. Shiohara, S. Ueha, K. Matsushima, M. Suematsu, A. Mizoguchi, T. Hibi, A. K. Bhan, and H. Ishikawa Exacerbating role of gammadelta T cells in chronic colitis of T-cell receptor alpha mutant mice. Gastroenterology 134: King, C., A. Ilic, K. Koelsch, and N. Sarvetnick Homeostatic expansion of T cells during immune insufficiency generates autoimmunity. Cell 117: Ramanathan, S., and P. Poussier BB rat lyp mutation and Type 1 diabetes. Immunological reviews 184: Cousins, L., M. Graham, R. Tooze, C. Carter, J. R. Miller, F. M. Powrie, G. G. Macpherson, and G. W. Butcher Eosinophilic bowel disease controlled by the BB rat-derived lymphopenia/gimap5 gene. Gastroenterology 131: Wallis, R. H., K. Wang, L. Marandi, E. Hsieh, T. Ning, G. Y. Chao, J. Sarmiento, A. D. Paterson, and P. Poussier Type 1 diabetes in the BB rat: a polygenic disease. Diabetes 58: Jackson, R. A., J. B. Buse, R. Rifai, D. Pelletier, E. L. Milford, C. B. Carpenter, G. S. Eisenbarth, and R. M. Williams Two genes required for diabetes in BB rats. Evidence from cyclical intercrosses and backcrosses. The Journal of experimental medicine 159: Jacob, H. J., A. Pettersson, D. Wilson, Y. Mao, A. Lernmark, and E. S. Lander Genetic dissection of autoimmune type I diabetes in the BB rat. Nature genetics 2: Moralejo, D. H., H. a. Park, S. J. Speros, A. J. MacMurray, A. E. Kwitek, H. J. Jacob, E. S. Lander, and Å. Lernmark Genetic dissection of lymphopenia from autoimmunity by introgression of mutated Ian5 gene onto the F344 rat. Journal of autoimmunity 21: Elder, M. E., and N. K. Maclaren Identification of profound peripheral T lymphocyte immunodeficiencies in the spontaneously diabetic BB rat. Journal of immunology 130: Iwakoshi, N. N., D. L. Greiner, A. A. Rossini, and J. P. Mordes Diabetes prone BB rats are severely deficient in natural killer T cells. Autoimmunity 31: Tullin, S., P. Farris, J. S. Petersen, L. Hornum, M. Jackerott, and H. Markholst A pronounced thymic B cell deficiency in the spontaneously diabetic BB rat. Journal of immunology 158:
120 25. MacMurray, A. J., D. H. Moralejo, A. E. Kwitek, E. a. Rutledge, B. Van Yserloo, P. Gohlke, S. J. Speros, B. Snyder, J. Schaefer, S. Bieg, J. Jiang, R. a. Ettinger, J. Fuller, T. L. Daniels, A. Pettersson, K. Orlebeke, B. Birren, H. J. Jacob, E. S. Lander, and A. Lernmark Lymphopenia in the BB rat model of type 1 diabetes is due to a mutation in a novel immune-associated nucleotide (Ian)- related gene. Genome research 12: Nitta, T., M. Nasreen, T. Seike, A. Goji, I. Ohigashi, T. Miyazaki, T. Ohta, M. Kanno, and Y. Takahama IAN family critically regulates survival and development of T lymphocytes. PLoS biology 4: e Poirier, G. M., G. Anderson, a. Huvar, P. C. Wagaman, J. Shuttleworth, E. Jenkinson, M. R. Jackson, P. a. Peterson, and M. G. Erlander Immuneassociated nucleotide-1 (IAN-1) is a thymic selection marker and defines a novel gene family conserved in plants. Journal of immunology (Baltimore, Md. : 1950) 163: Hernandez-Hoyos, G., S. Joseph, N. G. Miller, and G. W. Butcher The lymphopenia mutation of the BB rat causes inappropriate apoptosis of mature thymocytes. European journal of immunology 29: Pandarpurkar, M., L. Wilson-Fritch, S. Corvera, H. Markholst, L. Hornum, D. L. Greiner, J. P. Mordes, A. a. Rossini, and R. Bortell Ian4 is required for mitochondrial integrity and T cell survival. Proceedings of the National Academy of Sciences of the United States of America 100: Ramanathan, S., K. Norwich, and P. Poussier Antigen activation rescues recent thymic emigrants from programmed cell death in the BB rat. Journal of immunology (Baltimore, Md. : 1950) 160: Carter, C., C. Dion, S. Schnell, W. J. Coadwell, M. Graham, L. Hepburn, G. Morgan, A. Hutchings, J. C. Pascall, H. Jacobs, J. R. Miller, and G. W. Butcher A natural hypomorphic variant of the apoptosis regulator Gimap4/IAN1. Journal of immunology 179: Schnell, S., C. Démollière, P. van den Berk, and H. Jacobs Gimap4 accelerates T-cell death. Blood 108: Schulteis, R. D., H. Chu, X. Dai, Y. Chen, B. Edwards, D. Haribhai, C. B. Williams, S. Malarkannan, M. J. Hessner, S. Glisic-Milosavljevic, S. Jana, E. J. Kerschen, S. Ghosh, D. Wang, A. E. Kwitek, A. Lernmark, J. Gorski, and H. Weiler Impaired survival of peripheral T cells, disrupted NK/NKT cell development, and liver failure in mice lacking Gimap5. Blood 112: Matsuda, J. L., O. V. Naidenko, L. Gapin, T. Nakayama, M. Taniguchi, C. R. Wang, Y. Koezuka, and M. Kronenberg Tracking the response of natural killer T cells to a glycolipid antigen using CD1d tetramers. The Journal of experimental medicine 192:
121 35. Galfre, G., C. Milstein, and B. Wright Rat x rat hybrid myelomas and a monoclonal anti-fd portion of mouse IgG. Nature 277: Zarebski, A., C. S. Velu, A. M. Baktula, T. Bourdeau, S. R. Horman, S. Basu, S. J. Bertolone, M. Horwitz, D. A. Hildeman, J. O. Trent, and H. L. Grimes Mutations in growth factor independent-1 associated with human neutropenia block murine granulopoiesis through colony stimulating factor-1. Immunity 28: Kim, S. O., Q. Jing, K. Hoebe, B. Beutler, N. S. Duesbery, and J. Han Sensitizing anthrax lethal toxin-resistant macrophages to lethal toxin-induced killing by tumor necrosis factor-alpha. The Journal of biological chemistry 278: Barnes, M. J., P. Krebs, N. Harris, C. Eidenschenk, R. Gonzalez-Quintial, C. N. Arnold, K. Crozat, S. Sovath, E. M. Moresco, A. N. Theofilopoulos, B. Beutler, and K. Hoebe Commitment to the regulatory T cell lineage requires CARMA1 in the thymus but not in the periphery. PLoS biology 7: e Beutler, B., X. Du, and Y. Xia Precis on forward genetics in mice. Nature immunology 8: Ng, S. Y., T. Yoshida, J. Zhang, and K. Georgopoulos Genome-wide lineage-specific transcriptional networks underscore Ikaros-dependent lymphoid priming in hematopoietic stem cells. Immunity 30: Clanton, D. J., Y. Y. Lu, D. G. Blair, and T. Y. Shih Structural significance of the GTP-binding domain of ras p21 studied by site-directed mutagenesis. Molecular and cellular biology 7: Egawa, T., R. E. Tillman, Y. Naoe, I. Taniuchi, and D. R. Littman The role of the Runx transcription factors in thymocyte differentiation and in homeostasis of naive T cells. The Journal of experimental medicine 204: Nakayama, T., D. J. Kasprowicz, M. Yamashita, L. A. Schubert, G. Gillard, M. Kimura, A. Didierlaurent, H. Koseki, and S. F. Ziegler The generation of mature, single-positive thymocytes in vivo is dysregulated by CD69 blockade or overexpression. Journal of immunology 168: Akashi, K., M. Kondo, U. von Freeden-Jeffry, R. Murray, and I. L. Weissman Bcl-2 rescues T lymphopoiesis in interleukin-7 receptor-deficient mice. Cell 89: McNab, F. W., S. P. Berzins, D. G. Pellicci, K. Kyparissoudis, K. Field, M. J. Smyth, and D. I. Godfrey The influence of CD1d in postselection NKT cell maturation and homeostasis. Journal of immunology 175:
122 46. Massberg, S., P. Schaerli, I. Knezevic-Maramica, M. Kollnberger, N. Tubo, E. A. Moseman, I. V. Huff, T. Junt, A. J. Wagers, I. B. Mazo, and U. H. von Andrian Immunosurveillance by hematopoietic progenitor cells trafficking through blood, lymph, and peripheral tissues. Cell 131: Nagai, Y., K. P. Garrett, S. Ohta, U. Bahrun, T. Kouro, S. Akira, K. Takatsu, and P. W. Kincade Toll-like receptors on hematopoietic progenitor cells stimulate innate immune system replenishment. Immunity 24: Strober, W., I. J. Fuss, and R. S. Blumberg The immunology of mucosal models of inflammation. Annual review of immunology 20: Fuss, I. J., F. Heller, M. Boirivant, F. Leon, M. Yoshida, S. Fichtner-Feigl, Z. Yang, M. Exley, A. Kitani, R. S. Blumberg, P. Mannon, and W. Strober Nonclassical CD1d-restricted NK T cells that produce IL-13 characterize an atypical Th2 response in ulcerative colitis. The Journal of clinical investigation 113: Heller, F., I. J. Fuss, E. E. Nieuwenhuis, R. S. Blumberg, and W. Strober Oxazolone colitis, a Th2 colitis model resembling ulcerative colitis, is mediated by IL-13-producing NK-T cells. Immunity 17: Aranda, R., B. C. Sydora, P. L. McAllister, S. W. Binder, H. Y. Yang, S. R. Targan, and M. Kronenberg Analysis of intestinal lymphocytes in mouse colitis mediated by transfer of CD4+, CD45RBhigh T cells to SCID recipients. Journal of immunology 158: Macpherson, A. J., E. Slack, M. B. Geuking, and K. D. McCoy The mucosal firewalls against commensal intestinal microbes. Seminars in immunopathology 31: Saunders, A., T. Lamb, J. Pascall, A. Hutchings, C. Dion, C. Carter, L. Hepburn, J. Langhorne, and G. W. Butcher Expression of GIMAP1, a GTPase of the immunity-associated protein family, is not up-regulated in malaria. Malaria journal 8: Kupfer, R., J. Lang, C. Williams-Skipp, M. Nelson, D. Bellgrau, and R. I. Scheinman Loss of a gimap/ian gene leads to activation of NF-kappaB through a MAPK-dependent pathway. Molecular immunology 44: Casola, S Control of peripheral B-cell development. Current opinion in immunology 19: Tan, J. T., E. Dudl, E. LeRoy, R. Murray, J. Sprent, K. I. Weinberg, and C. D. Surh IL-7 is critical for homeostatic proliferation and survival of naive T cells. Proceedings of the National Academy of Sciences of the United States of America 98:
123 57. Pellegrini, M., P. Bouillet, M. Robati, G. T. Belz, G. M. Davey, and A. Strasser Loss of Bim increases T cell production and function in interleukin 7 receptor-deficient mice. The Journal of experimental medicine 200: Gauld, S. B., R. J. Benschop, K. T. Merrell, and J. C. Cambier Maintenance of B cell anergy requires constant antigen receptor occupancy and signaling. Nature immunology 6:
124 Figure Legends Figure 1. Identification of sphinx, an ENU germline mutant exhibiting severe lymphopenia and hepatic extramedullary hematopoiesis. A, CD8 + T cell and NK cell cytotoxicity were tested in ENU mutagenized mice that had been previously immunized with irradiated act-mova cells. Cytotoxic function was assessed by the ability to removeβ2m / (CFSE int ) NK cell or SIINFEKL-loaded (CFSE high ) CD8 + T cell target cells. C57BL/6J control cells (CFSE low ) were used as a reference population. The percentage of CD3 + T cells and NK1.1 + NK cells (B), and CD4 + and CD8 + T cells (C) were quantified among splenocytes from 6-wk-old mice. D, Gross liver morphology in 15-d-old (upper) and 40-d-old (lower) mice. E, H&E staining of liver sections from 40-d-old mice (original magnification 100). Circulating concentrations of hemoglobin (F) and lymphocytes (G) were measured in the blood of mice of various ages. SD is shown and statistical analyses were performed for each time point. n = 5; **p < 0.01; ***p < H, Gimap5 protein expression was measured in lysates of total bone marrow and purified splenic CD19 + B cells. I, Gimap5 RNA levels in purified splenic CD19 + B cells isolated from C57BL/6J and Gimap5 sph/sph mice were measured by RT-PCR. All analyses were performed at least three times. Figure 2. Gimap5 sph/sph mice develop wasting disease, colitis and aberrant hematopoiesis. A, Cohoused C57BL/6J and Gimap5 sph/sph males were weighed at indicated ages. SEM is shown for each time point (n = 6) and statistical analysis was performed using a two-tailed paired ttest. B, H&E-stained sections from the colon of 10- wk-old cohoused mice were assessed for intestinal inflammation (original magnification 100). C, Hematopoietic cell egress from the neonatal liver was monitored by examining 114
125 H&E-stained sections from livers obtained at indicated developmental time-points (original magnification 200). At least three livers were analyzed for each time point, and representative sections are shown. Figure 3. Hematopoietic defects in the adult and fetal liver of Gimap5 sph/sph mice.a, HSCs and hematopoietic precursor cells in the liver of 6-wk-old mice were analyzed by flow cytometry. The mean number of cells for each subset is indicated along with SEM. n = 3; *p < 0.05; **p < B, Irradiated (1000 rad γ- radiation) Rag2 / Il2rγ / recipient mice were reconstituted with ED 19 fetal liver cells. Six weeks later, splenocyte populations in chimeric mice were quantified by flow cytometry. The mean total number of each cell type in the spleen is shown along with SD. n = 3; **p < 0.01; ***p < Figure 4. T cell development and function in Gimap5 sph/sph mice. To assess positive and negative selection, thymocyte (A) and splenocyte (B) populations from H-Y TCR + transgenic mice of both genders were quantified and the average percentage of cells in each quadrant is shown. C, Expression of terminal thymocyte maturation markers CD5, CD24, CD69, and IL-7Rα was compared between C57BL/6J (shaded, gray) and Gimap5 sph/sph (open, dark line) TCRβ + CD4 SP and CD8 SP thymocytes. D, Surface expression of IL-7rα was measured in splenic CD4 + T cells. E, Proliferation of CFSE-labeled CD4 + T cells isolated from 8-wk-old mice was assessed 96 h after stimulation with anti-cd3/cd28, with or without exogenous IL-2. F, Expression of CD62L and CD69 was measured on splenic CD4 + T cells from 3-, 4-, 6- and 10-wk-old mice. Numbers represent mean ± SD. All analyses were performed at least twice. 115
126 Figure 5. Reduced inkt cell survival and aberrant αgal-cer-induced cytokine responses in Gimap5 sph/sph mice. A,iNKT cells in the spleen of 6-wk-old Gimap5 sph/sph mice were identified by αgal-cer-loaded CD1d-tetramer binding and quantified by flow cytometry. B, Cytokine production was measured 90 min after injection of the inkt cell agonist αgal-cer on a per cell basis by intracellular staining for IFN-γ, TNF-α, IL-4, or IL-13 expression among αgal-cer-loaded CD1d-tetramer + inkt cells (blue, unstimulated C57BL/6J; red, stimulated C57BL/6J; green, stimulated Gimap5 sph/sph ). C, Serum cytokine concentrations of IFN-γ and IL-4 were also measured by ELISA 90 min after injection of αgal-cer. SD is shown. n = 3; ***p < Figure 6. Impaired BCR-dependent proliferation in Gimap5 sph/sph B cells. A C, Among CD19 + splenocytes from 6-wk-old mice, the percentage of IgM high IgD int naive and IgM low IgD high mature B cells (A) and the percentage of CD21 high CD23 low marginal zone (B) and CD21 high CD23 high follicular B cells (C) were determined. Six-week-old mice were immunized with 50 μg NP36-CGG with alum and LPS or 50 μg of NP50-Ficoll to assess T-dependent (D) or T-independent (E) Ab responses. NP-specific serum Abs were measured 14 d after immunization by ELISA. F, To assess B cell proliferation, CFSE-labeled CD19 + splenocytes were cultured in vitro for 90 h with media alone or with anti IgM-Fab (10 μg/ml), LPS (2 μg/ml), anti-cd40 (10 μg/ml) plus IL-4 (10 ng/ml), or PMA (50 ng/ml) and ionomycin (500 ng/ml). G, Cell cycle progression of B cells cultured in the presence or absence of LPS was measured by propidium iodide staining after 48 h of incubation. Figure 7. Antibiotic treatment abrogates colitis but not lymphopenia and hepatic extramedullary hematopoiesis in Gimap5 sph/sph mice. The percentage of NKp46 +, CD8 +, 116
127 CD4 +, and CD11b + splenocytes (A) and splenic CD44 high CD62L low CD4 + T cells (B) in 9- wk-old C57BL/6J, Gimap5 sph/sph, and antibiotic-treated Gimap5 sph/sph mice was determined. Bars represent mean values ± SEM. n = 3 per group; ***p < C, H&Estained sections from the colons and livers of 9-wk-old cohoused mice were assessed for extramedullary hematopoiesis and intestinal inflammation (original magnification 100). Representative histology from each group is shown. n = 3. Figure 8. Prevention of wasting disease by adoptive transfer of splenocytes. A, Twenty-five- to-35-d-old Gimap5 sph/sph mice were injected with splenocytes from C57BL/6J or Rag2 / donors. The survival of recipient mice was monitored for up to 9 mo (n 6 per group) and differences were statistically analyzed using the Gehan- Breslow-Wilcoxon test. B, Gimap5 sph/sph mice injected with C57BL/6J splenocytes did not develop wasting disease. Representative 15-wk-old mice are shown. C and D, In 30-d-old Gimap5 sph/sph mice injected with congenically marked splenocytes, the percentages of transferred (CD45.1) and endogenous (CD45.2) CD8 + T cells (C) and CD4 + T cells (D) were determined at the indicated time points after transfer. E, Expression of CD62L and CD44 on splenic CD4 + T cells from 13-wk-old C57BL/6 mice, Gimap5 sph/sph mice, and Gimap5 sph/sph mice injected with CD congenic splenocytes (only cells of Gimap5 sph/sph origin are shown). Lin myeloid precursor cells were quantified in the liver (F) and bone marrow (G) of 15-wk-old mice. 117

128 Figure

129 Figure2. 119
130 Figure
131 Figure

132 Figure
133 Figure

134 Figure

135 Figure
136 Supplementary Figure Legends Supplemental Figure 1. Analysis of NK cells in bone marrow of sphinx mice. The development of NK cells in the bone marrow was tracked using markers expressed by NK cell precursors at early (NK1.1 + NKp46 + ), intermediate (CD11b + ) and mature/activated (CD69 + ) stages of development. (n=3; ***=P<0.001). Supplemental Figure 2. Blood test results taken over the first 90 days of life in C57BL/6 and sphinx mice. A, Mean corpuscular volume, B, red blood cell width distribution, C, platelet counts, and, D, number of neutrophils are shown along with SD (n=5, *=P< 0.05, **=P<0.01, ***=P<0.001). Supplemental Figure 3. Positional cloning of the sphinx mutation. A, Using thirteen meioses and a panel of 130 microsatellite markers covering the entire genome, the sphinx phenotype was linked to chromosome 6. B, A G T nucleotide transition in the coding region of Gimap5 results in a G38C aa change in the middle of the Gimap5 AIG1 homology domain. C, The G T nucleotide change alters an existing AciI restriction site in C57BL/6 mice allowing for genotyping by restriction enzyme digestion. D, Evolutionary conservation of the Gly residue altered by the sphinx mutation was determined by creating multiple amino acid sequence alignments (ClustalW) of mouse Gimap5 with Mus musculus Gimap homologues and orthologous AIG1 domain containing polypeptides from different species. Supplemental Figure 4. Analysis of HSC populations in the bone marrow. A, Lin - myeloid precursor cells were quantified in the bone marrow of five-week-old C57BL/6 and Gimap5sph/sph mice using the gating strategy shown. B, The mean absolute number each cell population is indicated along with the SD (n=3). 126
137 Supplemental Figure 5. Decline in peripheral B cells in adult Gimap5 sph/sph mice. A, Percentage of B220 + cells in spleens from three- to twelve-week-old C57BL/6J or Gimap5 sph/sph mice, as measured by flow cytometry. (***=P<0.001). B, Ig isotype distribution in non-immune serum of eight-week-old C57BL/6J and Gimap5sph/sph mice. C,D In vitro activation of proximal B cell signaling pathways. C, Immunoblot analyses of MAP kinase activation and IκB degradation and, D, phosphorylation of Akt, PTEN, Pdk1 and GSK3b in B cells from C57BL/6 or Gimap5 sph/sph mice stimulated with PMA/ionomycin for the indicated number of min. B cells lysates were made using positively selected CD19 + splenocytes. Supplemental Figure 6. Reduced extramedullary hematopoiesis in the liver of Gimap5 sph/sph mice following adoptive transfer of C57BL/6J splenocytes. A, H&E stained liver sections from fifteen-week-old C57BL/6 mice, Gimap5 sph/sph mice or Gimap5 sph/sph mice that were injected with C57BL/6 splenocytes (treated) at day 30 are shown. Fewer hematopoietic cells and less extramedullary hematopoiesis were observed in the treated liver. B, The mean absolute number of hematopoietic cell populations in the liver of C57BL/6J, untreated and treated Gimap5 sph/sph mice is indicated along with the SD (n=3,*=p< 0.05, **=P<0.01). 127
138 Supplementary Figure
139 Supplementary Figure

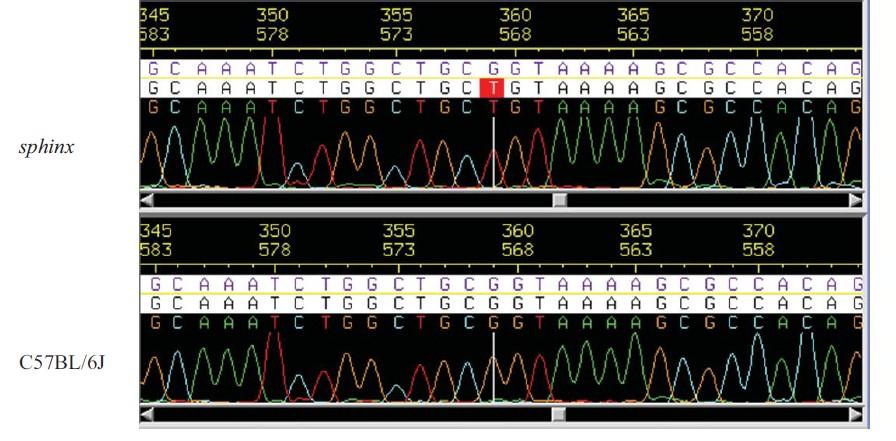
140 Supplementary Figure 3. A. B. 130


141 C. D. 131

142 Supplementary Figure 4. A. B. 132

143 Supplementary Figure 5. A. B. C. D. 133

144 Supplementary Figure
145 Chapter III Loss of Immunological Tolerance in Gimap5-Deficient Mice is Associated with Loss of Foxo in CD4 + T Cells H. Ibrahim Aksoylar*, Kristin Lampe*, Michael J. Barnes, David R. Plas and Kasper Hoebe* * Department of Molecular Immunology, Cincinnati Children s Hospital Research Foundation, Cincinnati, OH 45229; The Scripps Research Institute, La Jolla, CA 92037; and Department of Cancer and Cell Biology, University of Cincinnati Medical Center, Cincinnati, OH Address correspondence and reprint requests to Dr. Kasper Hoebe, Department of Molecular Immunology, Cincinnati Children s Hospital Research Foundation, MLC7021, Room S5.421, 3333 Burnet Avenue, Cincinnati, OH address: kasper.hoebe@cchmc.org The Journal of Immunology January 1, 2012 vol. 188 no
146 Abstract Previously, we reported the abrogation of quiescence and reduced survival in lymphocytes from Gimap5 sph/sph mice, an ENU germline mutant with a missense mutation in the GTPase of immunity-associated protein 5 (Gimap5). These mice showed a progressive loss of peripheral lymphocyte populations and developed spontaneous colitis, resulting in early mortality. In this study, we identify the molecular pathways that contribute to the onset of colitis in Gimap5 sph/sph mice. We show that CD4 + T cells become Th1/Th17 polarized and are critically important for the development of colitis. Concomitantly, regulatory T cells become reduced in frequency in the peripheral tissues, and their immunosuppressive capacity becomes impaired. Most importantly, these progressive changes in CD4 + T cells are associated with the loss of Forkheadbox group O (Foxo)1, Foxo3, and Foxo4 expression. Our data establish a novel link between Gimap5 and Foxo expression and provide evidence for a regulatory mechanism that controls Foxo protein expression and may help to maintain immunological tolerance. 136
147 Introduction The family of GTPase of immunity-associated protein (Gimap) genes are expressed predominantly in lymphocytes and regulate lymphocyte survival during development, selection, and homeostasis (1). Members of this family share a GTPbinding AIG1 homology domain, which was identified originally in disease resistance genes in higher plants (2, 3). Recent crystallographic studies revealed that GDP-bound or nucleotide-free Gimap2 exists in a monomeric configuration with an exposed guanine nucleotide binding domain (4). In the presence of GTP, Gimap2 oligomerizes and shows similarities with the nucleotide coordination and dimerization mode observed previously for dynamin GTPase. In addition, these studies showed that Gimap2 localized at the surface of lipid droplets, where it is thought to act as a nucleotideregulated scaffolding protein (4). Other members of the Gimap family appear to be localized to different subcellular compartments (5). Overall, the function of these proteins remains poorly defined. Gimap5 was reported recently to localize in lysosomes, based on studies in human, mouse, and rat lymphocytes (5). Genetic aberrancies in Gimap5 have been strongly linked to reduced lymphocyte survival and homeostasis but, importantly, also have been associated with autoimmune diseases. In humans, polymorphisms in GIMAP5 were associated with increased concentrations of IA2 autoantibodies in type 1 diabetes (T1D) patients (6) and an increased risk of systemic lupus erythematosus (7, 8). Studies using biobreeding rats carrying a mutation (lyp/lyp) in Gimap5 show marked lymphopenia and predisposition to the development of T1D (9-11). In addition, biobreeding rats are prone to develop intestinal inflammation on certain genetic 137
148 backgrounds (12). Together, these observations suggest that, beyond lymphocyte survival, Gimap5 is essential for maintaining immunological tolerance. Interestingly, impaired lymphocyte survival and consequent lymphopenia may be linked to the loss of immunological tolerance. Specifically, CD4 + T cells in a lymphopenic environment can undergo thymus-independent expansion in the periphery. This process also referred to as homeostatic or lymphopenia-induced proliferation (LIP) is accompanied by marked alterations in T cell phenotype and is linked to autoimmunity (13-15). Most notably, T cells undergoing LIP acquire a memory-like phenotype, exemplified by high surface expression of CD44 and low surface expression of CD62L. In addition, under lymphopenic conditions, CD4 + T cells more readily adopt an effector phenotype, including the ability to robustly produce cytokines upon stimulation through the TCR. The downstream consequences can be severe, and a number of pathological conditions have been associated with CD4 + T cells undergoing LIP, including colitis. Classic studies involving the adoptive transfer of naive CD45RB high CD4 + T cells into lymphopenic SCID mice resulted in T cells acquiring a LIP phenotype and rapidly driving colitis when recipient mice were colonized by intestinal bacteria (16-18). Importantly, colitis could be prevented if CD4 + CD25 + regulatory T (Treg) cells were cotransferred, suggesting that the presence or absence of Treg cells is an important determinant of immune-mediated sequelae induced by CD4 + T cells undergoing LIP. Our laboratory previously described an ENU germline mutant, designated sphinx, which contained a recessive mutation in Gimap5 that disrupted both lymphocyte survival and normal hematopoiesis (19). Similar to Gimap5 knockout mice, these mice lack peripheral NK cells and CD8 + T cells and exhibit dynamic changes in immune 138
149 homeostasis, marked by the progressive loss of CD4 + T cells and B cells and neutrophilia (19, 20). After the collapse of lymphocyte populations, CD4 + T cells in Gimap5 sph/sph mice acquire a LIP phenotype similar to that of CD4 + T cells transferred into lymphopenic hosts (18). Around wk of age, Gimap5 sph/sph mice develop wasting disease and colitis, limiting their survival (19). Interestingly, adoptive transfer of Rag-sufficient splenocytes into Gimap5 sph/sph mice around 5 wk of age could restore lymphocyte homeostasis and prevent colitis and wasting (19). In this report, we show that CD4 + T cells are required for the development of colitis in Gimap5 sph/sph mice. Whereas CD4 + T cells exhibited impaired proliferation, they remained highly capable of producing proinflammatory cytokines, including IL-17A and IFN-γ. Importantly, CD4 + T cells in Gimap5 sph/sph mice exhibited a LIP phenotype and exhibited a progressive and complete loss of full-length Forkheadbox group O (Foxo)1, Foxo3, and Foxo4 expression. This loss of Foxo expression was associated with a progressive reduction in the numbers and suppressive capacity of Foxp3 + Treg cells. The development of colitis in Gimap5 sph/sph mice could be prevented by transferring wildtype Treg cells into 3-wk-old Gimap5 sph/sph mice. Because Foxo-deficient mice exhibit many of the phenotypes observed in Gimap5 sph/sph mice, including impaired Treg cell activity and colitis, our data suggest that the loss of immunological tolerance in Gimap5- deficient mice may be critically linked to the loss of Foxo expression in CD4 + T cells. 139
150 Material and methods Mice and reagents All of the experiments were performed according to U.S. National Institutes of Health guidelines and were approved by the Institutional Animal Care and Use Committee of the Cincinnati Children s Hospital. C57BL/6J, Rag1 /, CD45.1 congenic, and CD90.1 congenic mice were obtained from The Jackson Laboratory. Gimap5 sph/sph mice were generated as described previously (19) and bred in the vivarium of the Cincinnati Children s Hospital. All of the mice were maintained under specific pathogen-free conditions. All of the Abs used for flow cytometry were purchased from ebioscience or Biolegend. Abs for Western blotting [anti-foxo3a (2497), anti-foxo1 (2880), antipfoxo1(thr 24 )/pfoxo3a(thr 32 ) (9464), anti-foxo4 (9472), p27, p-retinoblastoma protein (prb) (S807, S811), p-prb (S780), and pan-actin Abs] were purchased from Cell Signaling Technology. Purified CD3ε (145-2C11) and CD28 (37.51) Abs (ebioscience) were used for T cell activation. PMA and ionomycin were obtained from Sigma-Aldrich. Real-time PCR CD4 + T cells were isolated from spleens and lymph nodes of 4- and 7-wk-old Gimap5 sph/sph and C57BL/6 mice using L3T4 MicroBeads (Miltenyi Biotec). RNA isolation was done with a RNeasy Micro Kit (Qiagen), and reverse transcription was performed using a High-Capacity cdna Reverse Transcription Kit (Applied Biosystems). cdnas were amplified with LightCycler 480 SYBR Green I Master (Roche) and quantified by LightCycler 480 II instrument (Roche) using the following primer pairs: 140
151 Foxo1, forward, 5 -TTCGGAATGACCTCATGGATG-3, reverse, 5 - TGGACTGCTCCTCAGTTCCTG-3 ; Foxo3, forward, 5 -AGTGGATGGTGCGCTGTGT- 3, reverse, 5 -TCTGAACGCGCATGAAGC-3 ; Foxo4, forward, 5 - GAGAACCTGGAGTGCGACATG-3, reverse. 5 -TGTGTTGCCACCAAT-3. Flow cytometry and T cell analyses To quantify T cell proliferation, MACS-purified (Miltenyi Biotec) CD4 + T cells were labeled with 5 μm CFSE in PBS with 0.1% FCS for 10 min. Cells were cultured in supplemented IMDM media containing 10% FCS and 1% penicillin/streptomycin and were either left unstimulated or stimulated with PMA (50 ng/ml)/ionomycin (1 μg/ml). After 3 d of incubation, proliferation was measured by analyzing CFSE dilution using flow cytometry. To assess the capacity of T cells to produce cytokines, MACS-purified CD4 + T cells were incubated for 6 h with or without PMA (10 ng/ml)/ionomycin (10 μg/ml) and subsequently fixed and analyzed for intracellular IL-17A, IL-4, or IFN-γ production using flow cytometry. To measure surface markers ex vivo, CD4 + T cells from spleen, mesenteric lymph node (MLN), or lamina propria (LP) were isolated and stained with fluorochrome-labeled Abs specific for mouse CD44, CD62L, and CD69. Foxp3 expression was analyzed by intracellular staining. BrdU staining T cell-specific BrdU incorporation was measured as follows: during an interval of 24 h, wild-type or Gimap5 sph/sph mice received three i.p. injections with 100 μl of a 10 mg/ml BrdU solution in sterile PBS. Incorporation of BrdU in CD4 + T cells was measured 8 h after the last injection using flow cytometry. 141
152 In vitro Treg cell suppressor assays The Treg cell suppressor assay was performed under the conditions described previously (21, 22). Briefly, spleens were isolated, and Treg cells were MACS-purified using the CD4 + CD25 + regulatory T cell isolation kit (Miltenyi Biotec). Subsequently, Treg cells were harvested and cocultured at the indicated ratios with MACS-purified CFSE-labeled CD8 + T cells or CD4 + T cells. Also included were T cell-depleted, gamma-irradiated (1500 rad) splenocytes as bystander cells and 0.5 μg/ml soluble CD3 Ab. CFSE dilution was assessed by flow cytometry after 3 d of coculture. Histology Colon tissue was collected and immediately fixed in 10% buffered formalin solution overnight, followed by routine paraffin embedding. H&E staining was performed on 4-μm sections from the paraffin-embedded tissue blocks for conventional light microscopy analysis. Histological scoring was performed as described previously (23). Briefly, scoring parameters included quantification of the area of distal colon involved, edema, erosion/ulceration of the epithelial monolayer, crypt loss/damage, and infiltration of immune cells into the mucosa. Severity for the area involved (erosion/ulceration and crypt loss) was graded on a scale of 0 (normal), 1 (0 10%), 2 (10 25%), 3 (25 50%), and 4 (>50%). Immune cell infiltration was scored as: 0, absent; 1, weak; 2, moderate; 3, severe. Total disease score was expressed as the mean of all of the combined scores per genotype. 142
153 Adoptive transfer and survival assays For adoptive transfer studies, Gimap5 sph/sph mice at d of age were injected i.v. with Treg cells isolated from C57BL/6J mice using a Treg cell isolation kit (Miltenyi Biotec). Purity was confirmed by Foxp3 staining using flow cytometry, and cells were >90% Foxp3 +. Mice were monitored and weighed every week after cell transfer. Statistical analysis Data were analyzed using Prism4 software (GraphPad). Unless indicated otherwise, statistical significance of the differences among groups was determined from the mean and SD by Student s two-tailed test or by ANOVA followed by Dunnett s test for three or more groups. Data were considered statistically significant if p values were <
154 Results Gimap5 sph/sph CD4 + T cells from MLN are Th1/Th17 polarized In previous work, we determined that NK, NKT, CD8 +, CD4 +, and B lymphocyte survival are impaired in Gimap5 sph/sph mice. In addition, they developed spontaneous colitis that required the presence of microbiota and survived poorly, with most mice succumbing by 150 d of age (19). Because several mouse models have linked impaired lymphocyte function with colitis development, we further explored the contribution of lymphocytes to the immunopathology observed in Gimap5 sph/sph mice. First, we investigated the survival and functional capacity of CD4 + T cells at different ages. By 4 wk of age, a reduced number of CD4 + T cells were found in the spleen, and a further decline in T cell numbers was observed in 6- and 10-wk-old Gimap5 sph/sph mice (Fig. 1A). Six-week-old Gimap5 sph/sph CD4 + T cells had a CD44 high CD62L low phenotype characteristic of T cells undergoing LIP (19) and showed increased incorporation of BrdU (Fig. 1B). To assess whether the loss of CD4 + T cells in the spleen and lymph nodes also was observed in GALT, we isolated LP cells from the colons of 6-wkold Gimap5 sph/sph mice and quantified the number of CD4 + T cells. Similar to the spleen, reduced numbers of CD4 + T cells were observed in the LP (Fig. 1C). Further analysis revealed that close to 100% of the colonic CD4 + T cells were CD44 high CD62L low, resembling the LIP phenotype of the CD4 + T cells in the peripheral lymphoid tissues (Fig. 1C). Together, these data suggest that CD4 + T cells are present in GALT and exhibit a LIP phenotype similar to that observed in the spleen. 144
155 We next investigated the functional capacity of CD4 + T cells in Gimap5 sph/sph mice and their potential for contributing to the development of colitis. Our previous work indicated that 8-wk-old Gimap5 sph/sph CD4 + T cells were unable to proliferate ex vivo after stimulation with PMA/ionomycin or anti-cd3 Ab, even though lymphocytes exhibited normal activation of NF-κB and MAPK pathways (19). Because of the latter observation, we investigated whether CD4 + T cells were capable of producing cytokines after such stimulation and, if so, were Th1, Th2, or Th17 polarized. We isolated total lymphocytes from spleens and MLNs from C57BL/6J control or Gimap5 sph/sph mice and incubated cells for 6 h with or without PMA/ionomycin in the presence of brefeldin. Interestingly, a higher percentage of CD4 + T cells derived from Gimap5 sph/sph spleens or MLNs produced IFN-γ, IL-17A, or both cytokines after PMA/ionomycin stimulation (Fig. 1D). Notably, T cell cytokine production was observed even in the absence of PMA/ionomycin in Gimap5 sph/sph MLN cells (but not splenic leukocytes), suggesting constitutive activation of T cells in GALT in these mice. Overall, these data indicate that, despite their inability to proliferate normally ex vivo, CD4 + T cells derived from Gimap5 sph/sph mice become Th1/Th17 polarized and effectively produce cytokines. Colitis in Gimap5 sph/sph mice is driven by CD4 + T cells Because of their LIP phenotype and spontaneous production of IL-17A and IFNγ, we hypothesized that MLN CD4 + T cells may support the development of colitis in Gimap5 sph/sph mice. We tested this hypothesis by depleting CD4 + T cells in Gimap5 sph/sph mice using weekly injections of anti-cd4 (GK1.5) Abs, starting at 3 wk of age before the CD4 + T cells collapse and the subsequent intestinal inflammation normally occurs in Gimap5 sph/sph mice. Importantly, GK1.5 treatment, but not isotype 145
156 treatment, prevented wasting disease (Fig. 2A) and significantly decreased intestinal inflammation as determined by histology in 15-wk-old Gimap5 sph/sph mice (Fig. 2B H). These data support our hypothesis that the development of colitis in Gimap5 sph/sph mice requires CD4 + T cells. Gimap5 sph/sph mice fail to maintain a Treg cell population with normal immunosuppressive function Colitis induced by naive CD45RB high T cell transfer into SCID recipients does not occur when Treg cells are cotransferred. Therefore, even though Treg cell development in the thymus of Gimap5 sph/sph mice appeared to occur normally (19), we considered that Treg cell function may be impaired in the peripheral tissues of these lymphopenic mice and contribute to the development of colitis. Thus, we examined the presence and immunosuppressive capacity of Foxp3 + Treg cells in Gimap5 sph/sph mice. Although relatively normal numbers of Foxp3 + Treg cells were observed in 3-wk-old mice (Supplemental Fig. 2A), Treg cells became significantly reduced in the MLNs of 6- to 8- wk-old mice, both as a percentage within the CD4 + T cell compartment and as a total number of cells (Fig. 3A, 3B). In the spleen, the number of Treg cells was reduced, but the percentage of Foxp3 + CD4 + T cells within the CD4 T cell population remained similar to the percentage observed in wild-type mice (Fig. 3A, 3B). To assess their functional capacity, we purified Treg cells from 4- or 6-wk-old C57BL/6J or Gimap5 sph/sph spleens and cocultured Treg cells with CFSE-labeled wild-type CD8 + T cells that were stimulated with soluble anti-cd3 Abs. Treg cells from 4-wk-old Gimap5 sph/sph mice showed a slight but significant reduction in their ability to suppress CD8 + T cell proliferation in vitro, whereas Treg cells isolated from 6-wk-old Gimap5 sph/sph mice were 146
157 incapable of suppressing CD8 + T cell proliferation (Fig. 3C). Similar results were obtained for the suppression of wild-type CD4 + T cell proliferation (Supplemental Fig. 1A). These findings suggest that both Treg cell survival and functional capacity become impaired as Gimap5 sph/sph mice age. We next questioned whether reduced peripheral Treg cell accumulation in Gimap5 sph/sph mice resulted from a cell-intrinsic phenomenon. We injected CD4 + splenocytes from 3-wk-old wild-type and/or Gimap5 sph/sph mice into Rag-deficient recipients, as either a mixture or alone, and quantified the presence of Foxp3 + Treg cells 5 wk after injection (Supplemental Fig. 2A). Whereas no differences in the percentage of Treg cells within the CD4 + T cell compartment were observed at the time of injection, after 5 wk the Gimap5 sph/sph Treg population was lost, regardless of whether wild-type cells were cotransferred or not (Fig. 3D). Overall, these data indicate that cellintrinsic expression of Gimap5 is required to allow normal Treg cell survival. Gimap5 sph/sph CD4 + T cells exhibit progressive loss of Foxo1, Foxo3, and Foxo4 expression Our data indicate that the Gimap5 sph/sph CD4 + T cell population collapses around 5 wk of age and that the remaining CD4 + T cells undergo LIP thereafter. At the same time, they fail to maintain a functional Treg cell population. Interestingly, these T cell phenotypes show striking similarities with those seen in mice with T cells deficient in the family of Foxo transcription factors. The family of Foxo transcription factors contains four members, three of which (Foxo1, Foxo3, and Foxo4) have overlapping patterns of expression and transcriptional activities (24-26). They play an essential role in the 147
158 regulation of cell cycle progression, apoptosis, glucose metabolism, and life span (27). Foxo1 expression is critical for maintaining naive T cell quiescence. Foxo1-deficient CD4 + T cells exhibit a CD44 high CD62L low LIP or effector memory phenotype (28-30). In addition, Foxo expression has been reported to be essential for Treg cell development and function (29, 31). We therefore analyzed the expression of Foxo1, Foxo3, and Foxo4 in Gimap5 sph/sph CD4 + T cells. Strikingly, immunoblot analysis of CD4 + T cells from 6-wk-old Gimap5 sph/sph mice revealed a near absence of full-length Foxo1, Foxo3a, and Foxo4 protein (Fig. 4A). At the mrna level, a reduction in Foxo1 but not Foxo3 and Foxo4 could be observed in CD4 + T cells isolated from Gimap5 sph/sph mice compared with those isolated from wild-type mice (Fig. 4B), suggesting that regulation of Foxo3 and Foxo4 protein expression occurred at the posttranscriptional level. Because many of the T cell-specific phenotypes observed in Gimap5 sph/sph mice occur after 4 wk of age, we next quantified the temporal progression of changes in Foxo expression in lymphocytes. Immunoblot analyses revealed that Foxo expression was normal at 3 wk, somewhat reduced after 4 wk, and almost absent after 6 10 wk of age (Fig. 4C, Supplemental Fig. 1A). Concordant with the loss of Foxo expression, we detected reductions in the abundance of the cyclin-dependent kinase (Cdk) inhibitor p27 kip1, a downstream target of Foxo proteins and an important regulator of cell cycle entry (Fig. 4C) (32, 33). Because p27 kip1 inhibits Cdk4, we measured Cdk4 activity and detected increased phosphorylation of its substrate, prb, in Gimap5 sph/sph cells (Fig. 4C). Due to the progressive nature of this phenotype, we considered the possibility that lymphocytes isolated from young Gimap5 sph/sph mice with intact Foxo expression might respond normally to mitogenic stimuli. Indeed, CD4 + T cells isolated from 4-wk-old, but not 8-wk- 148
159 old, Gimap5 sph/sph mice were able to proliferate after TCR stimulation (Supplemental Fig. 3A) (19). Finally, we assessed whether the loss of Foxo expression also was observed in Treg cells from Gimap5 sph/sph mice. Indeed, Foxo1 and Foxo3 expression was mostly absent in CD4 + CD25 + T cells from 6-wk-old Gimap5 sph/sph mice (Fig. 4D). Interestingly, the progressive loss of Foxo expression appeared to correlate with a progressive increase in the number of CD4 + T cells undergoing LIP (CD44 hi CD62L lo ) in 4- to 10-wk-old Gimap5 sph/sph mice as reported previously (19). Subsequent analysis of CD44 lo CD62L hi and CD44 hi CD62L lo CD4 + T cells from 5-wk-old Gimap5 sph/sph and wild-type mice revealed the loss of Foxo expression specifically in CD4 + T cells undergoing LIP but not in naive CD4 + T cells in Gimap5 sph/sph mice (Fig. 4E), suggesting that the loss of Foxo expression follows T cell activation. Overall, these data link the loss of full-length Foxo expression with the onset of lymphopenia in Gimap5 sph/sph lymphocytes, impaired cell cycle control and proliferative capacity in such lymphocytes, and reduced Treg cell survival and function in Gimap5 sph/sph mice. Prevention of colitis in Gimap5 sph/sph mice by the adoptive transfer of wild-type Treg cells Our previous data show that colitis can be prevented in Gimap5 sph/sph mice through adoptive transfer of normal, but not Rag-deficient, splenocytes (19), indicating that a lymphocyte population is responsible for the rescue. Given the impaired Treg cell survival and function observed in Gimap5 sph/sph mice, we next examined whether adoptively transferred wild-type Treg cells could prevent the development of colitis. Gimap5 sph/sph recipients of wild-type CD4 + CD25 + T cells showed 149
160 prolonged survival and delayed wasting disease (Fig. 5A) and, importantly, did not develop colitis (Fig. 5B D). Characterization of lymphocyte populations 25 wk after transfer of CD45.1 congenically marked CD4 + CD25 + Treg cells revealed that Treg cell reconstitution of the spleens of Gimap5 sph/sph mice achieved 50% of the level observed in wild-type C57BL/6J mice (Fig. 5E). Notably, the Foxp3 + CD4 + Treg cell population constituted 40% of the overall CD4 + T cell population and was entirely congenic, whereas the Foxp3 CD4 + population was predominantly Gimap5 sph/sph -derived (Fig. 5E). Functional analysis of isolated CD4 + T cells from spleens and MLNs of 15-wk-old treated Gimap5 sph/sph mice revealed no background cytokine production and activation similar to that observed for wild-type CD4 + T cells after stimulation with PMA/ionomycin (Supplemental Fig. 2D). Around 25 wk of age, Treg cell-recipient mice still developed wasting disease. Necropsy at this time revealed severe inflammation in the lung and infiltration of macrophages in a number of mice (Supplemental Fig. 2C). Interestingly, colitis could be prevented by the transfer ofil10 / splenocytes (data not shown), suggesting that IL-10 independent regulatory pathways are more perturbed by Gimap5 deficiency. Together, these data link impaired Treg cell survival and function to the development of colitis in Gimap5 sph/sph mice. In addition, they reveal that the Gimap5 sph/sph environment is capable of supporting a functional Treg cell population. 150
161 Discussion Genetic aberrancies in Gimap5 have been linked to lymphopenia and the loss of immunological tolerance (6, 7, 9, 10). Although we found no evidence of autoimmune responses in Gimap5 sph/sph mice, we observed severe and spontaneous inflammation in the gut (19) an environment where homeostasis critically depends upon maintaining tolerance to exogenous Ags and bacterial stimuli. Similar to Gimap5 sph/sph mice, the loss of immunological tolerance in the lyp/lyp rat has been associated with reduced Treg cell survival and function as well as the polarization of Th cells toward a Th17 pattern of differentiation (34, 35). However, the molecular pathways underlying the loss of immunological tolerance in rat and mouse models of Gimap5 deficiency have remained elusive. In this study, we explored the pathways that contribute to the loss of tolerance observed in Gimap5 sph/sph mice. We show that the development of colitis in Gimap5 sph/sph mice is critically dependent on CD4 + T cells. Around 8 wk of age, CD4 + T cells lose their capacity to proliferate ex vivo, yet they remain capable of producing proinflammatory cytokines, including IL-17A and IFN-γ, and contain a population of IL IFN-γ + Th1/Th17 cells that have been associated with IL-23 signaling and more severe colitis (36). At the same time, Treg cell numbers and function decline. Most importantly, we found that these phenotypes are associated directly with a progressive loss of protein expression of Foxo1, Foxo3, and Foxo4-important transcription factors that regulate both quiescence and survival of lymphocytes (37). Mice with T cell-specific deletions offoxo1 and/or Foxo3 mimic many of the immunological and pathological phenotypes observed in Gimap5 sph/sph mice (28-31). For example, Foxo-deficient CD4 + T cells have impaired proliferative capacity and adopt CD44 high CD62L low LIP or 151
162 memory-like phenotypes (28, 30). In addition, reduced Treg cell numbers and function were observed in mice lacking Foxo1 or both Foxo1 and Foxo3 in T cells. Similar to Gimap5 sph/sph mice, mice lacking Foxo1 and Foxo3 in T cells develop spontaneous colitis, and furthermore purified Foxo1 / Foxo3 / Treg cells were unable to prevent colitis in Rag1 / mice when coinjected with naive wild-type CD4 + T cells (31). Interestingly, the impaired Treg cell development and function in T cell-specific Foxo1- deficient mice caused exaggerated T follicular helper cell accumulation, which contributed to B cell-mediated autoimmunity (29). In Gimap5 sph/sph mice, we found no evidence of autoreactive B cells (19), but it is important to note that, similar to CD4 + T cells, Gimap5 sph/sph B cells progressively lost Foxo expression (Supplemental Fig. 3B) and were unable to proliferate after stimulation with IgM (19). Thus, B cell expansion and differentiation may be hampered severely in Gimap5 sph/sph mice, preventing the development of autoreactive Ab responses. The mechanisms by which Foxo transcription factors control Treg cell development, homeostasis, and function have been studied in some detail. Foxo proteins have been shown to serve as coactivators downstream of the TGF-β signaling pathway by interacting with SMAD proteins, ultimately fine-tuning the TGF-β induced transcriptional program (38, 39). This pathway is also critical for the development of inducible Treg (itreg) cells (40), which develop extrathymically and have been suggested by many studies to comprise an important population of Treg cells in the gut (21, 41-43). Indeed, the loss of Treg cells within the CD4 + T cell compartment is most evident in the MLNs of Gimap5 sph/sph mice (Fig. 3), suggesting that the itreg cells in particular are impaired. In addition, Foxo1 and Foxo3 can cooperatively control the 152
163 differentiation of Foxp3 + Treg cells through the regulation of a number of Treg cellassociated genes, including Foxp3 itself (31). Furthermore, conditional deletion of Foxo1 in T cells resulted in reduced surface expression of CTLA-4 and CD25 in Foxp3 + CD4 + T cells (29). Analysis of the Ctla-4 gene showed that the promoter region contained a conserved Foxo binding site 193 bp upstream of the transcription start site (44). Thus, the impaired function of Foxp3 + Treg cells is likely the result of an incomplete transcriptional program in the absence of Foxo expression. In summary, the loss of Foxo expression affects multiple pathways that regulate Treg cell development, homeostasis, and function as well as the generation of itreg cells. In Gimap5 sph/sph mice, the absence of Foxo expression was observed in all of the lymphocyte populations examined, including peripheral Foxp3 + Treg cells, conventional Foxp3 CD4 + T cells, and B cells. Although conventional Foxp3 CD4 + T cells lack Foxo expression, our experiments reveal that colitis can be prevented by treatment with competent wild-type Treg cells, suggesting that colitogenic CD4 + T cells remain capable of being regulated when they lack Gimap5. Although we link the sphinx mutation in Gimap5 to the progressive loss of Foxo expression in lymphocytes, it is unclear to what extent these genes directly interact with each other. Given the progressive nature of the loss of Foxo expression, it is unlikely that Gimap5 directly interacts with Foxo proteins. One possibility we considered is that the loss of Foxo expression may drive a secondary phenotype resulting from constitutive proliferation cues associated with LIP, something that may be driven by self-antigens or Ags derived from the microbiota. Although we cannot exclude the possibility that LIP may contribute to the loss of Foxo expression in Gimap5 sph/sph CD4 + RB45 high T cells, wild-type CD4 + RB45 high T cells transferred into a 153
164 lymphopenic host retained normal levels of Foxo expression (Supplemental Fig. 3C), suggesting that LIP alone is insufficient to cause the loss of Foxo expression. Our data show that Gimap5 deficiency affects Foxo3 and Foxo4 expression at the protein level, not at the mrna level. Regulation of Foxo proteins has been reported previously to occur via ubiquitination and proteasomal degradation (45). Moreover, the loss of Foxo1 expression has been observed in mouse lymphomas, which served as a mechanism to remove the tumor suppressor activity of Foxo1 (46). Foxo degradation was correlated inversely with increased expression of S-phase kinase-associated protein-2 (Skp2) - an E3 ubiquitin ligase that targets numerous cell cycle proteins. Foxo degradation could be reversed after the downregulation of Skp2 via short hairpin RNAs, and therefore, increased Skp2 expression could provide a potential mechanism by which the loss of Foxo expression in Gimap5 sph/sph T cells occurs. Alternatively, the localization of Gimap5 in the lysosomal compartment (5) and the presumed scaffolding function of Gimap family members (4) suggest that Gimap5 may be necessary for optimal lysosomal function. Lysosomes are essential for the catabolic turnover of intraand extracellular macromolecules but also can release lysosomal enzymes (such as cathepsins) that can initiate programmed cell death once in the cytosol (47). Intriguingly, lymphocytes containing large numbers of cytotoxic granules, such as CD8 + T cells and NK cells, do not survive in Gimap5-deficient mice, perhaps supporting the hypothesis of a deregulated lysosomal compartment. Although the link between the loss of Gimap5 and Foxo protein expression remains to be established in human cells, understanding the molecular pathways that lead to the degradation of Foxo proteins could provide 154
165 important therapeutic targets, not only in the context of tumor growth, but potentially in the context of autoimmune or chronic inflammatory disorders. Our data provide evidence that Gimap5 is essential for maintaining lymphocyte quiescence and immunological tolerance. In the absence of functional Gimap5, Foxo expression in lymphocytes is progressively lost, with the loss of Foxo3 and Foxo4 most likely involving a proteolytic mechanism. This progressive loss of Foxo expression is associated with concomitant decline in Treg numbers and function, which ultimately leads to the loss of immunological tolerance in the gut. Thus, not only do we establish a critical link between Gimap5 and Foxo protein levels, we also provide evidence for a novel regulatory mechanism controlling Foxo protein expression that may be involved in the development of immune-mediated diseases such as systemic lupus erythematosus, T1D, and colitis. 155
166 References 1. Nitta, T., M. Nasreen, T. Seike, A. Goji, I. Ohigashi, T. Miyazaki, T. Ohta, M. Kanno, and Y. Takahama IAN family critically regulates survival and development of T lymphocytes. PLoS biology 4: e Reuber, T. L., and F. M. Ausubel Isolation of Arabidopsis genes that differentiate between resistance responses mediated by the RPS2 and RPM1 disease resistance genes. The Plant cell 8: Poirier, G. M., G. Anderson, a. Huvar, P. C. Wagaman, J. Shuttleworth, E. Jenkinson, M. R. Jackson, P. a. Peterson, and M. G. Erlander Immuneassociated nucleotide-1 (IAN-1) is a thymic selection marker and defines a novel gene family conserved in plants. Journal of immunology (Baltimore, Md. : 1950) 163: Schwefel, D., C. Fröhlich, J. Eichhorst, B. Wiesner, J. Behlke, L. Aravind, and O. Daumke Structural basis of oligomerization in septin-like GTPase of immunity-associated protein 2 (GIMAP2). Proceedings of the National Academy of Sciences of the United States of America 107: Wong, V. W., A. E. Saunders, A. Hutchings, J. C. Pascall, C. Carter, N. A. Bright, S. A. Walker, N. T. Ktistakis, and G. W. Butcher The autoimmunity-related GIMAP5 GTPase is a lysosome-associated protein. Self Nonself 1: Shin, J. H., M. Janer, B. McNeney, S. Blay, K. Deutsch, C. B. Sanjeevi, I. Kockum, A. Lernmark, J. Graham, G. Swedish Childhood Diabetes Study, G. Diabetes Incidence in Sweden Study, H. Arnqvist, E. Bjorck, J. Eriksson, L. Nystrom, L. O. Ohlson, B. Schersten, J. Ostman, M. Aili, L. E. Baath, E. Carlsson, H. Edenwall, G. Forsander, B. W. Granstrom, I. Gustavsson, R. Hanas, L. Hellenberg, H. Hellgren, E. Holmberg, H. Hornell, S. A. Ivarsson, C. Johansson, G. Jonsell, K. Kockum, B. Lindblad, A. Lindh, J. Ludvigsson, U. Myrdal, J. Neiderud, K. Segnestam, S. Sjoblad, L. Skogsberg, L. Stromberg, U. Stahle, B. Thalme, K. Tullus, T. Tuvemo, M. Wallensteen, O. Westphal, and J. Aman IA-2 autoantibodies in incident type I diabetes patients are associated with a polyadenylation signal polymorphism in GIMAP5. Genes Immun 8: Hellquist, A., M. Zucchelli, K. Kivinen, U. Saarialho-Kere, S. Koskenmies, E. Widen, H. Julkunen, A. Wong, M. L. Karjalainen-Lindsberg, T. Skoog, J. Vendelin, D. S. Cunninghame-Graham, T. J. Vyse, J. Kere, and C. M. Lindgren The human GIMAP5 gene has a common polyadenylation polymorphism increasing risk to systemic lupus erythematosus. J Med Genet 44: Lim, M. K., D. H. Sheen, S. a. Kim, S. K. Won, S.-S. Lee, S.-C. Chae, H.-T. Chung, and S. C. Shim IAN5 polymorphisms are associated with systemic lupus erythematosus. Lupus 18: Greiner, D. L., J. P. Mordes, E. S. Handler, M. Angelillo, N. Nakamura, and A. A. Rossini Depletion of RT6.1+ T lymphocytes induces diabetes in resistant 156
167 biobreeding/worcester (BB/W) rats. The Journal of experimental medicine 166: Jacob, H. J., A. Pettersson, D. Wilson, Y. Mao, A. Lernmark, and E. S. Lander Genetic dissection of autoimmune type I diabetes in the BB rat. Nature genetics 2: Hornum, L., J. Romer, and H. Markholst The diabetes-prone BB rat carries a frameshift mutation in Ian4, a positional candidate of Iddm1. Diabetes 51: Cousins, L., M. Graham, R. Tooze, C. Carter, J. R. Miller, F. M. Powrie, G. G. Macpherson, and G. W. Butcher Eosinophilic bowel disease controlled by the BB rat-derived lymphopenia/gimap5 gene. Gastroenterology 131: Khoruts, A., and J. M. Fraser A causal link between lymphopenia and autoimmunity. Immunology letters 98: King, C., A. Ilic, K. Koelsch, and N. Sarvetnick Homeostatic expansion of T cells during immune insufficiency generates autoimmunity. Cell 117: Krupica, T., T. J. Fry, and C. L. Mackall Autoimmunity during lymphopenia: a two-hit model. Clinical immunology (Orlando, Fla.) 120: Powrie, F., R. Correa-Oliveira, S. Mauze, and R. L. Coffman Regulatory interactions between CD45RBhigh and CD45RBlow CD4+ T cells are important for the balance between protective and pathogenic cell-mediated immunity. The Journal of experimental medicine 179: Powrie, F., M. W. Leach, S. Mauze, L. B. Caddle, and R. L. Coffman Phenotypically distinct subsets of CD4+ T cells induce or protect from chronic intestinal inflammation in C. B-17 scid mice. International immunology 5: Aranda, R., B. C. Sydora, P. L. McAllister, S. W. Binder, H. Y. Yang, S. R. Targan, and M. Kronenberg Analysis of intestinal lymphocytes in mouse colitis mediated by transfer of CD4+, CD45RBhigh T cells to SCID recipients. Journal of immunology 158: Barnes, M. J., H. Aksoylar, P. Krebs, T. Bourdeau, C. N. Arnold, Y. Xia, K. Khovananth, I. Engel, S. Sovath, K. Lampe, E. Laws, A. Saunders, G. W. Butcher, M. Kronenberg, K. Steinbrecher, D. Hildeman, H. L. Grimes, B. Beutler, and K. Hoebe Loss of T cell and B cell quiescence precedes the onset of microbial flora-dependent wasting disease and intestinal inflammation in Gimap5- deficient mice. Journal of immunology 184: Schulteis, R. D., H. Chu, X. Dai, Y. Chen, B. Edwards, D. Haribhai, C. B. Williams, S. Malarkannan, M. J. Hessner, S. Glisic-Milosavljevic, S. Jana, E. J. Kerschen, S. Ghosh, D. Wang, A. E. Kwitek, A. Lernmark, J. Gorski, and H. Weiler Impaired survival of peripheral T cells, disrupted NK/NKT cell development, and liver failure in mice lacking Gimap5. Blood 112:
168 21. Barnes, M. J., P. Krebs, N. Harris, C. Eidenschenk, R. Gonzalez-Quintial, C. N. Arnold, K. Crozat, S. Sovath, E. M. Moresco, A. N. Theofilopoulos, B. Beutler, and K. Hoebe Commitment to the regulatory T cell lineage requires CARMA1 in the thymus but not in the periphery. PLoS biology 7: e Thornton, A. M., and E. M. Shevach CD4+CD25+ immunoregulatory T cells suppress polyclonal T cell activation in vitro by inhibiting interleukin 2 production. The Journal of experimental medicine 188: Steinbrecher, K. A., E. Harmel-Laws, R. Sitcheran, and A. S. Baldwin Loss of epithelial RelA results in deregulated intestinal proliferative/apoptotic homeostasis and susceptibility to inflammation. Journal of immunology 180: Anderson, M. J., C. S. Viars, S. Czekay, W. K. Cavenee, and K. C. Arden Cloning and characterization of three human forkhead genes that comprise an FKHR-like gene subfamily. Genomics 47: Biggs, W. H., 3rd, W. K. Cavenee, and K. C. Arden Identification and characterization of members of the FKHR (FOX O) subclass of winged-helix transcription factors in the mouse. Mammalian genome : official journal of the International Mammalian Genome Society 12: Furuyama, T., K. Kitayama, Y. Shimoda, M. Ogawa, K. Sone, K. Yoshida-Araki, H. Hisatsune, S. Nishikawa, K. Nakayama, K. Nakayama, K. Ikeda, N. Motoyama, and N. Mori Abnormal angiogenesis in Foxo1 (Fkhr)-deficient mice. The Journal of biological chemistry 279: Greer, E. L., and A. Brunet FOXO transcription factors at the interface between longevity and tumor suppression. Oncogene 24: Kerdiles, Y. M., D. R. Beisner, R. Tinoco, A. S. Dejean, D. H. Castrillon, R. a. DePinho, and S. M. Hedrick Foxo1 links homing and survival of naive T cells by regulating L-selectin, CCR7 and interleukin 7 receptor. Nature immunology 10: Kerdiles, Y. M., E. L. Stone, D. R. Beisner, D. L. Beisner, M. a. McGargill, I. L. Ch'en, C. Stockmann, C. D. Katayama, and S. M. Hedrick Foxo transcription factors control regulatory T cell development and function. Immunity 33: Ouyang, W., O. Beckett, R. A. Flavell, and M. O. Li An essential role of the Forkhead-box transcription factor Foxo1 in control of T cell homeostasis and tolerance. Immunity 30: Ouyang, W., O. Beckett, Q. Ma, J.-h. Paik, R. a. DePinho, and M. O. Li Foxo proteins cooperatively control the differentiation of Foxp3+ regulatory T cells. Nature immunology 11: Dijkers, P. F., R. H. Medema, C. Pals, L. Banerji, N. S. Thomas, E. W. Lam, B. M. Burgering, J. A. Raaijmakers, J. W. Lammers, L. Koenderman, and P. J. Coffer Forkhead transcription factor FKHR-L1 modulates cytokine- 158
169 dependent transcriptional regulation of p27(kip1). Molecular and cellular biology 20: Medema, R. H., G. J. Kops, J. L. Bos, and B. M. Burgering AFX-like Forkhead transcription factors mediate cell-cycle regulation by Ras and PKB through p27kip1. Nature 404: Poussier, P., T. Ning, T. Murphy, D. Dabrowski, and S. Ramanathan Impaired post-thymic development of regulatory CD4+25+ T cells contributes to diabetes pathogenesis in BB rats. Journal of immunology (Baltimore, Md. : 1950) 174: van den Brandt, J., H. J. Fischer, L. Walter, T. Hünig, I. Klöting, and H. M. Reichardt Type 1 diabetes in BioBreeding rats is critically linked to an imbalance between Th17 and regulatory T cells and an altered TCR repertoire. Journal of immunology (Baltimore, Md. : 1950) 185: Ahern, P. P., C. Schiering, S. Buonocore, M. J. McGeachy, D. J. Cua, K. J. Maloy, and F. Powrie Interleukin-23 drives intestinal inflammation through direct activity on T cells. Immunity 33: Tothova, Z., and D. G. Gilliland FoxO transcription factors and stem cell homeostasis: insights from the hematopoietic system. Cell stem cell 1: Gomis, R. R., C. Alarcón, W. He, Q. Wang, J. Seoane, A. Lash, and J. Massagué A FoxO-Smad synexpression group in human keratinocytes. Proceedings of the National Academy of Sciences of the United States of America 103: Seoane, J., H.-V. Le, L. Shen, S. a. Anderson, and J. Massagué Integration of Smad and forkhead pathways in the control of neuroepithelial and glioblastoma cell proliferation. Cell 117: Harada, Y., Y. Harada, C. Elly, G. Ying, J.-H. Paik, R. a. DePinho, and Y.-C. Liu Transcription factors Foxo3a and Foxo1 couple the E3 ligase Cbl-b to the induction of Foxp3 expression in induced regulatory T cells. The Journal of experimental medicine 207: Atarashi, K., T. Tanoue, T. Shima, A. Imaoka, T. Kuwahara, Y. Momose, G. Cheng, S. Yamasaki, T. Saito, Y. Ohba, T. Taniguchi, K. Takeda, S. Hori, Ivanov, II, Y. Umesaki, K. Itoh, and K. Honda Induction of colonic regulatory T cells by indigenous Clostridium species. Science 331: Izcue, A., S. Hue, S. Buonocore, C. V. Arancibia-Carcamo, P. P. Ahern, Y. Iwakura, K. J. Maloy, and F. Powrie Interleukin-23 restrains regulatory T cell activity to drive T cell-dependent colitis. Immunity 28: Zheng, Y., S. Josefowicz, A. Chaudhry, X. P. Peng, K. Forbush, and A. Y. Rudensky Role of conserved non-coding DNA elements in the Foxp3 gene in regulatory T-cell fate. Nature 463:
170 44. Perkins, D., Z. Wang, C. Donovan, H. He, D. Mark, G. Guan, Y. Wang, T. Walunas, J. Bluestone, J. Listman, and P. W. Finn Regulation of CTLA-4 expression during T cell activation. Journal of immunology 156: Plas, D. R., and C. B. Thompson Akt activation promotes degradation of tuberin and FOXO3a via the proteasome. The Journal of biological chemistry 278: Huang, H., K. M. Regan, F. Wang, D. Wang, D. I. Smith, J. M. van Deursen, and D. J. Tindall Skp2 inhibits FOXO1 in tumor suppression through ubiquitinmediated degradation. Proceedings of the National Academy of Sciences of the United States of America 102: Kirkegaard, T., and M. Jaattela Lysosomal involvement in cell death and cancer. Biochimica et biophysica acta 1793:
171 Figure Legends Figure 1. Phenotypic characterization of CD4 + T cells in Gimap5 sph/sph mice. A and B, Splenic CD4 + T cell population collapse around 6 wk of age ingimap5 sph/sph mice (A), at which time they exhibit a LIP phenotype with increased BrdU uptake (B). C, The number of CD4 + T cells and the percentage of CD44 high CD62L low CD4 + T cells in LP of 6- wk-old wild-type and Gimap5 sph/sph mice. D, Ex vivo cytokine production by CD4 + T cells isolated from wild-type orgimap5 sph/sph spleens and MLNs, left unstimulated or after stimulation with PMA/ionomycin (100 ng/ml) for 6 h (mean values ± SEM; n 4 mice per genotype from two independent experiments). *p < 0.05, **p < 0.01, ***p < Figure 2. Depletion of CD4 + T cells prevents wasting disease and colitis in Gimap5 sph/sph mice. Male Gimap5 sph/sph mice were given 200 μg/mouse anti-cd4 (GK1.5) or isotype control Abs i.p. weekly, beginning at 3 wk of age. A, Weights of 10- wk-old wild-type, isotype-treated, or GK1.5-treated Gimap5 sph/sph mice. B, CD4 depletion significantly reduces colitis in Gimap5 sph/sph mice. Data represent histological scoring as described in the Materials and Methods. C H, At 15 weeks of age, mice were sacrificed, and the colons were analyzed for signs of intestinal inflammation after H&E staining (C, wild-type untreated; D F, Gimap5 sph/sph isotype-treated; G and H, Gimap5 sph/sph GK1.5- treated). Data represent mean weight percentage (wild-type mice = 100%) ± SEM from at least three animals per group, and histology is representative of three mice analyzed per group. Scale bars, 200 μm (C G), 100 μm (H). *p < 0.05, **p < 0.01, ***p < Figure 3. Reductions in the numbers and function of peripheral Foxp3 + Treg cells in Gimap5 sph/sph mice precede the onset of colitis. A and B, Flow cytometric analysis of 6-wk-oldGimap5 sph/sph mice reveals a reduced percentage of Foxp3 + cells within the 161
172 CD4 + T cell compartment (A) and a reduced absolute number of Treg cells (B). C, Treg cells isolated from 4- or 6-wk-old Gimap5 sph/sph mice have a reduced capacity to suppress the proliferation of anti-cd28/anti-cd3 activated, cocultured C57BL/6J CD8 + T cells, as measured by CFSE dilution after 72 h of incubation in vitro. D, The percentage of congenic (CD45.1) and Gimap5 sph/sph Foxp3 + CD4 + T cells in spleens and MLNs 25 wk after transfer in Rag1 / recipient mice. The Gimap5 sph/sph but not wild-type Treg cell population was lost in Rag1 / mice injected with a mixture of wild-type and Gimap5 sph/sph CD4 + T cells. Data represents mean values ± SEM (n 4 mice per genotype from two independent experiments). *p < 0.05, **p < 0.01, ***p < Figure 4. CD4 + T cells in Gimap5 sph/sph mice progressively lose full-length Foxo expression in lymphocytes. A, Immunoblot analysis of phosphorylated Foxo1/3, total Foxo1, Foxo3, and Foxo4 in CD4 + T cells from 6-wk-old mice. B, Foxo1, Foxo3, and Foxo4 mrna abundance in CD4 + T cells isolated from wild-type or Gimap5 sph/sph spleens, as measured by quantitative real-time PCR. C, Loss of Foxo expression ingimap5 sph/sph lymphocytes correlates with decreased p27 kip1 expression and increased phosphorylation of prb. D, Foxo protein expression in CD4 + CD25 + T cells isolated from 6-wk-old wild-type or Gimap5 sph/sph spleens (sph/sph, homozygote; +/sph, heterozygote). E, Foxo3 expression in CD44 hi CD62L lo and CD44 lo CD62L hi CD4 + T cells isolated from 5-wk-old homozygote sphinx mice and heterozygote littermate controls. Data represent mean values ± SEM, and blots are representative of three independent experiments (n = 3). ***p < Figure 5. Colitis in Gimap5 sph/sph mice can be prevented by adoptive transfer of wild-type Treg cells. A, Three- to 4-wk-oldGimap5 sph/sph mice were injected i.v. with 3 162
173 10 5 CD25 + CD4 + splenocytes isolated from wild-type mice. Recipient mice were weighed for up to 25 wk and compared with untreated heterozygote and homozygotegimap5 sph/sph mice. B, Gimap5 sph/sph mice treated with wild-type Treg cells were protected from colitis development as determined by histological scoring. C and D, H&E-stained colon sections from untreated 12-wk-old Gimap5 sph/sph (C) and 25-wk-old Treg cell-recipient Gimap5 sph/sph mice (D). Scale bars, 200 μm (C, left panel, D), 100 μm (C, right panel). E and F, The total number (E) and percentage (F) of Gimap5 sph/sph (CD45.1 ) and congenic (CD45 + ) Foxp3 + Treg cells in spleens and MLNs of 25-wk-old in CD4 + CD25 + Treg cell-recipient Gimap5 sph/sph mice. In C, the numbers of Foxp3 + cells in wild-type C57BL/6J mice are presented as a comparison. All of the studies were performed with n 4 mice per genotype from two independent experiments, and data are represented as mean values ± SEM. *** p <
174 Figure

175 Figure
176 Figure

177 Figure

178 Figure
179 Supplementary figure legends Supplementary figure 1. Progressive loss immunosuppressive function in Gimap5 sph/sph mice is associated with a loss of Foxo expression in CD4 + T cells. (A) Reduced capacity of Treg cells isolated from 3- or 6-week-old Gimap5 sph/sph mice to suppress proliferation of C57BL/6J CD4 + T cells stimulated with anti-cd28/cd3- activated, as measured by CFSE dilution after 72 hours incubation in vitro. (B) Immunoblot analysis of total Foxo1, Foxo3 and Foxo4 in CD4+ T cells from 3 or 6-weekold mice. Data represents mean values + SEM (n > 3 mice per genotype); blots are representative of three independent experiments (n = 3). Supplementary figure 2. (A) Transfer of a mixture of purified congenic wildtype and Gimap5 sph/sph CD4 + T cells into 6 week-old Rag1 -/- recipient mice. Mice were analyzed 5 weeks after transfer. The percentage of Foxp3+ Treg cells in wildtype and Gimap5 sph/sph CD4 + T cells at the time of injection is shown. (B) Rescue studies in which 3x10 5 CD25 + CD4 + cells isolated from congenic (CD45.1) C57BL/6J mice are injected into 4 week-old Gimap5 sph/sph mice. Analysis was performed after 25 weeks. (C) Inflammatory lung infiltrate as observed in a number of Treg rescued Gimap5 sph/sph mice. (D) Ex vivo cytokine production by CD4 + T cells isolated from 15- week-old wildtype or Treg-treated Gimap5 sph/sph spleen and mesenteric lymph node (MLN), left unstimulated or following stimulation with PMA/ionomycin (100ng/ml) for six hours. (Data represents mean values + SEM; n > 3 mice per genotype from 3 independent experiments). Supplementary Figure 3. (A) Proliferation of CD4 + T cells isolated from 4 or 8 weekold wildtype C57BL/6J or Gimap5 sph/sph mice. Cells were stimulated with anti-cd3 and proliferation was measured by CFSE-dilution after 3 days of incubation. (B) Progressive 169
180 loss of Foxo3 expression in B cells isolated from Gimap5 sph/sph but not wildtype cells. (C) Foxo3 expression in CD4 T + cells isolated from wildtype spleen (control) or following the transfer of wt CD4 + RB45 high CD4 + T cells in Rag -/- recipient mice. The latter were analyzed 5 weeks afte transfer. Data represents mean values + SEM; n > 4 mice per genotype from 2 independent experiments; blots are representative blots of B cells pooled from three mice. 170

181 Supplementary Figure 1 A. B. 171
182 Supplementary Figure 2 A. B. 172

183 C. 173


184 Supplementary Figure 3 A. B. C. 174
185 Chapter IV TCR-Signaling in Gimap5-Deficient CD4 + T Cells is Associated with an Abnormal Bioenergetic Profile and Reduced Survival H. Ibrahim Aksoylar, Kristin Lampe and Kasper Hoebe Division of Cellular and Molecular Immunology, Cincinnati Children s Hospital Research Foundation, Cincinnati, OH
186 Abstract Previously, we demonstrated that Gimap5 sph/sph mice die prematurely due to CD4 + T cell-dependent severe colitis. Gimap5 sph/sph CD4 + T cells display a poor survival that is associated with a progressive lymphopenia-induced proliferation (LIP) phenotype. At the same time, a selective decline in peripheral FoxP3 + Treg cell numbers and their immunosuppressive function can be observed. In this study, we investigated the role of TCR signaling in the impaired Treg homeostasis and T cell survival as observed for Gimap5 sph/sph CD4 + T cells. Using an oral tolerance model and Gimap5 sph/sph or wild type RAG -/- OT-II TCR transgenic mice, we show that cognate antigen-induced TCR signaling resulted in an overall reduced CD4 + T cell survival, acquisition of a LIP phenotype and a failure to induce a peripheral Treg population in Gimap5 sph/sph mice. Since Gimap5 was previously shown to be expressed in lysosomes, we tested whether abnormalities in lysosomal and autophagosomal function in Gimap5 sph/sph CD4 + T cells underlie the observed phenotypes. Autophagy is essential for mitochondrial turnover and energy homeostasis during T cell proliferation. Upon TCR activation, we observed larger autophagic vesicles expressing LC3 and Lamp1 markers in Gimap5 sph/sph CD4 + T cells. Moreover, autophagosomes showed increased association with mitochondrial markers indicating an abnormal rate of mitochondrial turnover. Furthermore, TCR activated Gimap5 sph/sph CD4 + T cells displayed elevated levels of reactive oxygen species (ROS) and oxygen consumption rate (OCR) suggesting a defect in the mitochondrial function. Our results suggest an important role for Gimap5 in TCR-induced autophagy and metabolic programs required for T cell proliferation. 176
187 Introduction: GTPase of immunity-associated protein 5 (Gimap5) is critical for survival and homeostasis of T cells (1) and has recently been linked to autoimmune diseases in humans. Polymorphisms in Gimap5 have been associated with an increased risk for systemic lupus erythematosus (2, 3) and increased levels of IA2 autoantibodies in type 1 diabetes patients (4). Moreover, animal models lacking Gimap5 develop autoimmune disease. For instance, the lyp allele in biobreeding rats, encoding a truncated form of Gimap5 protein, displayed spontaneous diabetes associated with T cell lymphopenia (5, 6). In mice, the ENU-induced Gimap5 mutation sphinx causes severe colitis due to a progressive loss of immunological tolerance ultimately resulting in early mortality (7). In previous studies, we showed that, Gimap5 sph/sph mice lost CD4 + T cell quiescence and failed to maintain immunological tolerance (8). These phenotypes were associated with impaired expression of Foxo proteins which are central to maintenance of T cell quiescence and Treg function (9, 10). We demonstrated that Gimap5 sph/sph FoxP3 + regulatory T (Treg) cells were reduced in absolute but also relative numbers within the CD4 + T cell compartment and showed impaired suppressive capacity (8). Moreover, Gimap5 sph/sph CD4 + T cells progressively acquired a CD44 high CD62L low LIP phenotype and produced pro-inflammatory cytokines (i.e. IFN and IL-17) leading to microflora-dependent severe colitis (8). We also observed a progressive loss of Foxo protein expression in CD4 + T cells that was specifically observed in the CD44 high CD62L low effector CD4 + T cell population in Gimap5 sph/sph mice (8). These observations suggest that the loss of Gimap5 sph/sph T cell quiescence may be triggered by TCR activation, for instance driven by lymphopenia and antigens derived from microbial flora. 177
188 However, the role of TCR signaling in the loss of CD4 + T cell quiescence, LIP phenotype and impaired Treg function in Gimap5 sph/sph mice is unclear. Gimap5 has been shown to localize in lysosomes even though its function is not established (11). TCR activation results in the induction of lysosome biogenesis and exocytosis of polarized granules in CD4 + T cells (12). However, the significance of lysosomal biogenesis in CD4 + T cells is not fully understood. Nevertheless, lysosomal stability is required for cell viability. Loss of lysosomal stability and permeabilization of lysosomal membrane can result in necrotic cell death due to the leakage of proteolytic enzymes (13). Another critical mechanism in which lysosomes are involved is the autophagic process in cells (14). Lysosomes fuse with autophagosomes and degrade their contents. Gimap5 is structurally similar to GTPases which regulate vesicular dynamics in the intracellular membranes (15). As Gimap5 localizes to lysosomes, it is possible that it is involved in specific lysosomal functions such as degradation of autophagic vesicles by lysosomes. Recent findings support that autophagy plays an important role in T cell proliferation, homeostasis and survival. T cells with deficiencies in the autophagy pathway (i.e. Atg3 -/- or Atg5 -/- mice), displayed impaired TCR-induced proliferation. Moreover, Atg3 -/ and Atg5 -/- mice developed T cell lymphopenia indicating a critical role for autophagy in T cell survival (16, 17). Autophagy helps to maintain bioenergetics and viability by utilizing cellular macromolecules (18, 19). Furthermore, T cells lacking another autophagy gene Atg7 exhibited defective clearance of damaged mitochondria (20, 21). Atg7 -/- T cells showed increased mitochondrial mass associated with increased levels of reactive oxygen species (ROS). Thus, autophagy in T cells is critical for 178
189 regulating energy metabolism during TCR-induced proliferation and homeostatic survival. Importantly, a failure to remove damaged organelles such as mitochondria has been shown to result in loss of T cell viability. Whether the reduced survival and impaired TCR-induced proliferation of Gimap5-deficient CD4 + T cells are linked to defects in autophagy remains unclear. In this study, we aimed to investigate the role of antigen-specific TCR signaling in the development of the LIP phenotype and impaired Treg function observed in Gimap5 sph/sph mice. To do so we used OT-II TCR transgenic RAG1 -/- mice which contain only CD4 + T cells specific for the chicken ovalbumin epitope. While treatment of wild type OT-II TCR transgenic RAG1 -/- mice with ovalbumin leads to peripheral Treg induction and maintenance of naïve phenotype, Gimap5 sph/sph CD4 + OT-II RAG1 -/- T cells acquired a LIP phenotype, and an impaired induction of peripheral Treg induction and an overall reduced survival of CD4 + T cells was observed. Moreover, we showed that TCR signaling in Gimap5-deficient CD4 + T cells resulted in the formation of larger sized autophagosomes, with increased mitochondrial content. Furthermore, Gimap5- deficient CD4 + T cells displayed elevated oxygen consumption and ROS production upon TCR activation. These data suggest a critical role for Gimap5 in TCR-induced autophagy and metabolic programs. 179
190 Material and methods Mice and reagents All of the experiments were performed according to U.S. National Institutes of Health guidelines and were approved by the Institutional Animal Care and Use Committee of the Cincinnati Children s Hospital. C57BL/6J, Rag1 / and OT-II TCR transgenic mice were obtained from The Jackson Laboratory. Gimap5 sph/sph mice were generated as described previously (7) and bred in the vivarium of the Cincinnati Children s Hospital. Gimap5 sph/sph mice were bred with Rag1 / and OT-II TCR transgenic mice in order to generate Gimap5 sph/sph Rag1 / OT-II genotype. All of the mice were maintained under specific pathogen-free conditions. All of the antibodies used for flow cytometry were purchased from ebioscience or Biolegend. Anti-Foxo3a (2497) and pan-actin Abs purchased from Cell Signaling Technology and anti-lc3 Ab from Novus Biologicals were used for western blotting. Purified CD3ε (145-2C11) and CD28 (37.51) Abs (ebioscience) were used for T cell activation. Mitotracker Deep Red, Live/Dead violet dead cell stain kit, DAPI (Invitrogen) and antibodies for Lamp1, CD4 (ebioscience) and LC3 (MBL International) were used for labeling T cells in Imagestream analysis. FCCP, oligomycin and antimycin A (Sigma- Aldrich) were used in Seahorse analysis. Flow cytometry To analyze the surface markers of CD4 + T cells, splenocytes and mesenteric lymph node cells were isolated and stained with fluorochrome-labeled Abs against mouse CD4, CD62L, and CD44. Foxp3 staining was done with intracellular staining kit 180
191 purchased from ebioscience. For the measurement of reactive oxygen, cells were stained with DCFDA (Sigma-Aldrich) for 30 minutes, washed and analyzed immediately. Stained cells were analyzed with LSR-II or FACS Canto flow cytometers (BD Biosciences). Ovalbumin treatment Four week old mice were treated with 5 mg/ml ovalbumin (Soigma-Aldrich) in drinking water for nine weeks and their spleens and lymph nodes were harvested for T cell analysis. Water bottles containing ovalbumin were refreshed every two days during the treatment. Imagestream analysis Imagestream analysis of CD4 + T cells was performed using ImagestreamX analyzer and IDEAS software (Amnis). Splenocytes were cultured in supplemented IMDM media containing 10% FCS and 1% penicillin/streptomycin and were either left unstimulated or stimulated with plate coated anticd3 (10µg/mL) and anti-cd28 (1µg/mL) antibodies for 16 hours. Then, cells were stained with the Live/Dead violet dead cell stain. Next, Mitotracker Deep Red and CD4 surface stains were performed. After fixation, cells were stained with anti-lc3 or anti-lamp1 intracellularly and were acquired using ImagestreamX. Fluorescence intensity of markers, number and average size of lysosomes, autophagosomes and mitochondria, and their colocalization in the cell were analyzed using IDEAS software. 181
192 Analysis of oxygen consumption rate Oxygen consumption rate was quantified with XF24 extracellular flux analyzer (Seahorse Bioscience). Briefly, MACS purified (Miltenyi Biotec) CD4 + T cells were stimulated with plate coated anti-cd3 (10µg/mL) and anti-cd28 (1µg/mL) antibodies for 16 hours in complete IMDM. After stimulation, CD4 + T cells were transferred to XF24 cell culture plates previously coated with 0.1 % poly-l-lysine (Sigma-Aldrich) and left for 30 minutes for adherence to plates. Post stimulation oxygen consumption rate was measured using XF24 extracellular flux analyzer and measurements were following injections of oligomycin, FCCP and antimycin A respectively. 182
193 Results: Loss of Gimap5 sph/sph CD4 + T cell quiescence and survival is driven by antigenspecific TCR signaling In our previous study, we demonstrated that loss of CD4 + T cell quiescence leads to severe colitis in Gimap5 sph/sph mice (8). Moreover, depletion of commensal microflora by antibiotics treatment prevented colitis development suggesting that T cell-mediated inflammation was driven in response to intestinal bacterial antigens. Therefore, we aimed to explore the role of TCR signaling in the phenotypes observed for CD4 + T cells in Gimap5 sph/sph mice. In order to control the antigen specific TCR signaling in Gimap5 sph/sph CD4 + T cells in vivo, we generated Gimap5 sph/sph OT-II transgenic RAG1 -/- mice containing only CD4 + T cells specific for the chicken ovalbumin epitope. In the absence of specific antigen, CD4 + T cells in Gimap5 sph/sph OT-II RAG1 -/- mice remained naïve and did not exhibit a CD44 high CD62L low LIP phenotype (Fig. 1A). Moreover, normal expression of Foxo3 protein in Gimap5-deficient OT-II RAG1 -/- CD4 + T cells was observed (Fig. 1B). In addition, Gimap5 sph/sph OT-II RAG1 -/- mice did not develop spontaneous colitis and showed a relative normal survival beyond 6 Months of age (data not shown). Thus, T cell quiescence and immunological tolerance in Gimap5- deficient mice is maintained in the absence of TCR-induced activation of CD4 + T cells. Next, we determined whether cognate antigen-induced TCR signaling would induce a LIP phenotype in Gimap5 sph/sph CD4 + T cells. To test our hypothesis, we treated Gimap5 sph/sph or wild type OT-II RAG1 -/- mice with drinking water containing ovalbumin, starting at 4 weeks of age, and subsequently analyzed their T cell 183
194 phenotype after 9 weeks of continuous treatment. CD4 + T cells in Gimap5 sph/sph OT-II RAG1 -/- mice treated with ovalbumin displayed a LIP phenotype exemplified by a dramatic increase in the frequency of CD44 high CD62L low cells (Fig. 1C). Oral administration of soluble antigen without adjuvant is known to induce peripheral tolerance in mice. Specifically, in TCR transgenic mice, administration of soluble antigen resulted in induction of Treg cells in the periphery (22). Importantly, our previous study (chapter3) suggested an impaired Treg function and development in Gimap5-deficient mice to cause a progressive loss of immunological tolerance, ultimately resulting in severe colitis and early mortality (8). Therefore, we wanted to test whether oral administration of antigen induces peripheral conversion of Gimap5- deficient CD4 + T cells into Treg cells. Whereas wild type OT-II RAG1 -/- CD4 + T cells showed a strong Treg induction upon oral ovalbumin administration, Gimap5 sph/sph OT-II RAG1 -/- CD4 + T cells were unable to induce FoxP3 + expression suggesting that Gimap5 is necessary for the generation of inducible Treg cells in the peripheral tissues (Fig. 1D). Our previous work showed that in addition to their CD44 high CD62L low phenotype, a progressive loss in overall numbers of CD4 + T cells was observed in Gimap5 sph/sph mice. To test the role of TCR activation in the impaired survival of Gimap5-deficient CD4 + T cells in vivo, we analyzed the numbers of CD4 + T cells in Gimap5 sph/sph OT-II RAG1 -/- and WT OT-II RAG1 -/- mice treated with ovalbumin in drinking water. After eight weeks of OVA treatment, the numbers of CD4 + T cells in WT OT-II RAG1 -/- mice did not change significantly whereas CD4 + T cells were almost completely lost in Gimap5 sph/sph OT-II RAG1 -/- mice compared to the treatment with water only (Fig. 1E). Thus, in the absence of Gimap5, antigen-specific TCR signaling causes impaired survival of CD4 + T 184
195 cells. Together, our data indicate that TCR activation of CD4 + T cells in the absence of functional Gimap5 results in 1) a reduced survival with remaining cells adopting a CD44 high CD62L low LIP phenotype; 2) a failure to generate peripheral inducible Treg cells. Lysosomal abnormalities in Gimap5 sph/sph T cells with TCR activation As Gimap5 has been reported to be expressed in lysosomes (11), we aimed to characterize the lysosomal compartment in Gimap5-deficient T cells. Using ImageStream, we first analyzed the expression of the lysosomal marker Lamp1 in naïve and anti-cd3/cd28 activated T cells. Whereas no significant differences in Lamp1 expression in unstimulated CD4 + T cells between wild type and Gimap5 sph/sph were found, increased expression of Lamp1 was observed in TCR-stimulated Gimap5 sph/sph CD4 + T cells (Fig. 2A). This increase in Lamp1 intensity was not a result of increased number of lysosomes as we detected similar lysosomal quantities in wild type and Gimap5 sph/sph activated CD4 + T cells compared with WT CD4 + T cells (Fig. 2A). Moreover, when we examined the size of individual lysosomes, we found larger average size of lysosomes in TCR-stimulated Gimap5 sph/sph CD4 + T cells (Fig. 2A). Lysosomal function is essential for degradation of macromolecules in autophagic vesicles in cells. Recent studies reveal autophagy to be required for providing necessary bioenergetic resources for T cell proliferation (19). Previously, we observed defects in TCR-induced proliferation of Gimap5 sph/sph CD4 + T cells (7), suggesting that there may be defects in meeting the bioenergetics demand of activated Gimap5- deficient CD4 + T cells. To determine whether normal fusion of lysosomes and 185
196 autophagic vesicles occurs in Gimap5 sph/sph CD4 + T cells, we measured the colocalization of Lamp1 with the autophagic vesicle marker LC3 (Fig. 2B). Although equal numbers of autophagolysosomes were found in wild type and Gimap5 sph/sph CD4 + T cells, autophagolysosomes appeared larger in size in Gimap5-deficient CD4 + T cells (Fig. 2C). These results suggest that there may be an increase in the autophagic activity or alternatively a block in the degradation of autophagic vesicles in Gimap5-deficient CD4 + T cells. Larger macroautophagic vesicles in TCR-stimulated Gimap5 sph/sph T cells Autophagy has recently been demonstrated to be necessary for the clearance and recycling of macromolecules and mitochondria during T cell proliferation (21). As we observed larger Lamp1 + LC3 + autophagolysosomes in activated Gimap5 sph/sph CD4 + T cells, we next investigated the formation of autophagosome and their colocalization to mitochondria. Using Western blot, we observed an elevated level of the phospholipidassociated form of the LC3 (LC3-II) molecule in Gimap5 sph/sph CD4 + T cells indicating an increased level of autophagic vesicle formation (Fig. 3A). These findings were confirmed by Imagestream analysis as we found a significantly higher intensity of LC3 in the autophagic vesicles in Gimap5 sph/sph CD4 + T cells (Fig. 3B). Upon TCR stimulation, we did not detect significant differences in the numbers of LC3 punctate structures between wild type and Gimap5 sph/sph CD4 + T cells. However, the average size of autophagic vesicles was larger in activated Gimap5 sph/sph CD4 + T cells (Fig. 3B). Next we determined the colocalization of autophagosomes with mitochondria in Gimap5-deficient CD4 + T cells. LC3 intensity in autophagosomes colocalized with 186
197 mitochondria was increased in Gimap5-deficient CD4 + T cells although no differences were observed in the number of autophagosomes (Fig. 3C-D). Moreover, the average size of autophagosomes containing mitochondria was also significantly higher in TCRstimulated Gimap5 sph/sph CD4 + T cells compared with WT CD4 + T cells (Fig. 3D). Furthermore, we observed higher intensity of Mitotracker staining within the autophagosomes of activated Gimap5 sph/sph CD4 + T cells (Fig. 3E), suggesting an increased mitochondrial content in autophagosomes from Gimap5 sph/sph CD4 + T cells. These results could indicate an increased autophagy induction or may indicate a reduced degradation (turnover) of macroautophagy vesicles in activated Gimap5 sph/sph CD4 + T cells. Mitochondrial dysfunction in TCR-stimulated Gimap5 sph/sph T cells During activation and proliferation, T cells switch their metabolism from oxidative phosphorylation to glycolysis which makes them less dependent on mitochondria for ATP synthesis (23). Thus, mitochondrial turnover by macroautophagy clears the excess and damaged mitochondria that are the potential sources of elevated levels of reactive oxygen species (ROS). Given that the changes in autophagy could indicate an increased need for autophagy or alternatively, could present an impaired clearance of mitochondria in Gimap5 sph/sph CD4 + T cells, we measured intracellular ROS levels in activated CD4 + T cells. Indeed, compared to wild type, Gimap5 sph/sph CD4 + T cells exhibited increased ROS levels following activation (Fig. 4A), suggesting an increased mitochondrial activity or increased mitochondrial damage in activated Gimap5 sph/sph CD4 + T cells. 187
198 Mitochondrial function is critical for T cell survival and activation. During TCR activation, T cells initially utilize oxidative phosphorylation to meet the increased demand for ATP. However, ultimately they their metabolism is switched towards glycolysis and glutaminolysis in order to support the required anabolic process for mitotic proliferation (23). Since we found increased levels of ROS production in Gimap5 sph/sph CD4 + T cells, we sought to analyze mitochondrial activity in CD4 + T cells. Using the Seahorse extracellular flux analyzer, we determined oxygen consumption rates (OCR) in naïve and TCR stimulated T cells. Sixteen hours after TCR activation, CD4 + T cells from Gimap5 sph/sph showed an increased oxygen consumption rate compared to wild type cells (Fig. 4B). Moreover, we observed a higher ATP linked OCR in Gimap5 sph/sph CD4 + T cells which is calculated by the initial OCR subtracted by the OCR after oligomycin treatment (Fig. 4B). Further, we analyzed the difference of OCR between oligomycin and antimycin A treatments which gives the level of proton leak. We observed that proton leak was significantly increased in Gimap5 sph/sph CD4 + T cells compared with wild type cells (Fig. 4B). Although activated T cells normally switch their metabolism to glycolysis (23), Gimap5-deficient CD4 + T cells showed higher OCR after activation suggesting that they fail to switch to glycolytic metabolism. Overall, these results indicate that activated Gimap5-deficient CD4 + T cells may have a metabolic dysfunction. 188
199 Discussion: In this study, we examined whether TCR signaling drives the abnormal T cell phenotype in Gimap5 sph/sph mice, by using TCR transgenic OT-II RAG -/- mice housed in the presence or absence of cognate antigen in vivo. Using TCR transgenic OT-II RAG -/- mice housed in the presence or absence of cognate antigen in vivo, our data show that Gimap5-deficient CD4 + T cells maintain their naïve status in the absence of TCR signaling but lose their quiescence and acquire a LIP phenotype in the presence of cognate antigen. It is well established that oral administration of cognate antigen without co-stimulation induces oral tolerance by promoting T cell anergy and by inducing peripheral Treg cells (24, 25). Consistent with this, WT OT-II RAG -/- CD4 + T cells were mostly naïve when fed with water containing ovalbumin. In contrast, Gimap5-deficient CD4 + T cells showed an activated phenotype suggesting a role for Gimap5 downstream of TCR activation. The impaired generation of induced Tregs with oral administration of OVA in OT- II RAG -/- Gimap5 sph/sph mice suggests a defect in the molecular mechanisms required for Treg induction in these cells. A possible reason for this may be the excess ATP-linked OCR observed during the activation of Gimap5 sph/sph CD4 + T cells (Fig. 4A). A recent report suggest that Treg induction is highly depending on lipid metabolism which requires the activation of AMP-activated protein kinase (AMPK) (26). It is possible that the reduced AMP/ATP ratio in Gimap5-deficient CD4 + T cells prevents AMPK activation during Treg induction. Therefore, the inefficient peripheral conversion of TCR-activated OT-II RAG -/- Gimap5 sph/sph CD4 + T cells into Tregs might be due to impaired AMPK activation and a consequent failure to utilize lipid metabolism. The status of AMPK 189
200 activation in TCR-stimulated Gimap5-deficient CD4 + T cells remains to be examined. Moreover, it would be interesting to test whether the AMPK activating drug, metformin could correct the inducible Treg defect of Gimap5 sph/sph CD4 + T cells. Different mechanisms have been proposed to explain the reduced survival of Gimap5-deficient CD4 + T cells (1, 27, 28). However, the role of TCR signaling in the survival defect has not been addressed. We here demonstrate that antigen-specific TCR signaling results in significantly reduced numbers of CD4 + T cells in OT-II RAG -/- Gimap5 sph/sph mice compared to mice housed in an antigen-free environment. It is possible that Gimap5-deficient CD4 + T cells die as they attempt to proliferate in response to antigen-specific TCR signaling. This would be consistent with our previous data showing impaired in vitro proliferation of anti-cd3/cd28 stimulated Gimap5- deficient CD4 + T cells (7). Additionally, it is important to note that the numbers of CD4 + T cells in OT-II RAG -/- Gimap5 sph/sph mice were still lower than the number of wild type OT-II RAG -/- CD4 + T cells even in the absence of antigen. This suggests that Gimap5 may be necessary for homeostatic proliferation of naïve CD4 + T cells. Thus, Gimap5- deficient CD4 + T cells may fail to complete homeostatic cell division driven by TCR/selfpeptide/MHC signaling, although this requires further investigation. Since Gimap5 was previously shown to be expressed in lysosomes (11), we investigated whether Gimap5 plays a role in the lysosomal biogenesis or function during TCR activation. Our results suggest that Gimap5 is not necessary for the biogenesis of lysosomes. However, increased intensity of Lamp1 and the observed increased size of lysosomes in Gimap5 sph/sph CD4 + T cells may point to abnormalities in lysosome function. It has been shown that Lamp2 deficiency causes enhanced accumulation of 190
201 autophagic vacuoles in cells due to impaired fusion of lysosomes to autophagic vesicles (29, 30). Although we did not observe a reduced lysosome-autophagosome fusion in activated Gimap5 sph/sph CD4 + T cells, the size of fused autophagolysosomes was again larger than wild type controls. Other studies showed that lysosomal dysfunction results in defects in the degradation of autophagic vesicles. For example, deficiency of the lysosomal GTPase Rab7 caused accumulation of autophagosomes with unusually large size. It was suggested that Rab7 is required for the final maturation of late autophagosomes after lysosome fusion (31, 32). Also, Cathepsin L-deficient cells displayed impaired turnover of autophagic vesicles and accumulated large autophagic compartments (33). Similarly, our results showed that TCR-stimulated Gimap5-deficient CD4 + T cells contained LC3 + autophagic vesicles larger than wild type controls. These results indicate that autophagy takes place in activated Gimap5-deficient CD4 + T cells; however autophagolysosome maturation or degradation of their contents may be impaired in the absence of Gimap5. Alternatively, these observations may indicate an increased stimulation of autophagy to occur in activated Gimap5-deficient CD4 + T cells. Therefore, more definitive studies to investigate the autophagy flux are necessary in order to distinguish between these two possibilities. Recent studies demonstrated that autophagy is critical for recycling of cellular organelles and clearance of damaged mitochondria during T cell proliferation (16, 17). Mice with deletion of autophagy genes Atg3, Atg5 or Atg7 showed impaired clearance of mitochondria and resulted in marked T cell lymphopenia and impaired T cell proliferation upon TCR activation (16, 17, 21). As we observed large autophagic vesicles associated with increased mitochondrial markers in Gimap5-deficient CD4 + T 191
202 cells following TCR activation, the mitochondrial turnover rate in Gimap5-deficient CD4 + T cells may be abnormal due to a defect in autophagy. However, again, autophagy flux experiments are required to test this possibility. Thus, autophagy may play a role in the CD4 + T cell lymphopenia and impaired TCR-induced proliferation phenotypes of Gimap5-deficient mice. If the larger size of autophagosomes in Gimap5 sph/sph CD4 + T cells is due to an elevated autophagic activity, an important question to ask is what triggers autophagy in these cells. The increased levels of ROS in Gimap5-deficient CD4 + T cells may indicate an increased mitochondrial damage due to high level of OCR and proton leak. An autophagy related protein Atg4 acts as a sensor for mitochondrial ROS as its inhibition by ROS triggers autophagy (34). Thus, elevated ROS levels in Gimap5-deficient CD4 + T cells may induce the autophagic pathway in these cells. During TCR activation and proliferation, T cells utilize glycolytic and glutaminolytic pathways in order to meet the demand for the anabolic pathways such as synthesis of amino acids, nucleic acids and lipids (35). Elevated levels of ATP-linked OCR in activated Gimap5 sph/sph CD4 + T cells suggest that these cells fail to switch their metabolism to glycolysis. Glycolytic metabolism is activated by two key pathways mediated by mtorc1 and c-myc. Inhibition of mtorc1 in T cells leads to a failure to switch to glycolysis during T cell activation (36). Similarly, transcription of many genes involved in glycolysis is impaired in c-myc deficient T cells (37). It would be interesting to see whether mtorc1 and c-myc have reduced activities in Gimap5 sph/sph CD4 + T cells. Moreover, a failure to utilize the glycolytic pathway may lead to the insufficiency of bioenergetic resources essential for the anabolic metabolism. If this is the case, 192
203 autophagy can be activated through the Ulk1/2 pathway which is relieved from the inhibition by mtorc1 (38). Therefore, an increased autophagy in Gimap5 sph/sph CD4 + T cells might be due to an insufficiency of bioenergetic resources during activation. How Gimap5 is involved in the potential dysregulation of these mechanisms needs to be evaluated. In conclusion, our results show that cognate antigen-specific TCR signaling causes a LIP phenotype and fails to induce a Treg population in Gimap5-deficient mice. Moreover, our results suggest that Gimap5-deficiency may dysregulate molecular mechanisms in metabolic pathways and autophagy. The exact function of Gimap5 remains to be investigated. 193
204 References 1. Nitta, T., M. Nasreen, T. Seike, A. Goji, I. Ohigashi, T. Miyazaki, T. Ohta, M. Kanno, and Y. Takahama IAN family critically regulates survival and development of T lymphocytes. PLoS biology 4: e Hellquist, A., M. Zucchelli, K. Kivinen, U. Saarialho-Kere, S. Koskenmies, E. Widen, H. Julkunen, A. Wong, M. L. Karjalainen-Lindsberg, T. Skoog, J. Vendelin, D. S. Cunninghame-Graham, T. J. Vyse, J. Kere, and C. M. Lindgren The human GIMAP5 gene has a common polyadenylation polymorphism increasing risk to systemic lupus erythematosus. J Med Genet 44: Lim, M. K., D. H. Sheen, S. a. Kim, S. K. Won, S.-S. Lee, S.-C. Chae, H.-T. Chung, and S. C. Shim IAN5 polymorphisms are associated with systemic lupus erythematosus. Lupus 18: Shin, J. H., M. Janer, B. McNeney, S. Blay, K. Deutsch, C. B. Sanjeevi, I. Kockum, A. Lernmark, J. Graham, G. Swedish Childhood Diabetes Study, G. Diabetes Incidence in Sweden Study, H. Arnqvist, E. Bjorck, J. Eriksson, L. Nystrom, L. O. Ohlson, B. Schersten, J. Ostman, M. Aili, L. E. Baath, E. Carlsson, H. Edenwall, G. Forsander, B. W. Granstrom, I. Gustavsson, R. Hanas, L. Hellenberg, H. Hellgren, E. Holmberg, H. Hornell, S. A. Ivarsson, C. Johansson, G. Jonsell, K. Kockum, B. Lindblad, A. Lindh, J. Ludvigsson, U. Myrdal, J. Neiderud, K. Segnestam, S. Sjoblad, L. Skogsberg, L. Stromberg, U. Stahle, B. Thalme, K. Tullus, T. Tuvemo, M. Wallensteen, O. Westphal, and J. Aman IA-2 autoantibodies in incident type I diabetes patients are associated with a polyadenylation signal polymorphism in GIMAP5. Genes Immun 8: MacMurray, A. J., D. H. Moralejo, A. E. Kwitek, E. a. Rutledge, B. Van Yserloo, P. Gohlke, S. J. Speros, B. Snyder, J. Schaefer, S. Bieg, J. Jiang, R. a. Ettinger, J. Fuller, T. L. Daniels, A. Pettersson, K. Orlebeke, B. Birren, H. J. Jacob, E. S. Lander, and A. Lernmark Lymphopenia in the BB rat model of type 1 diabetes is due to a mutation in a novel immune-associated nucleotide (Ian)- related gene. Genome research 12: Hornum, L., J. Romer, and H. Markholst The diabetes-prone BB rat carries a frameshift mutation in Ian4, a positional candidate of Iddm1. Diabetes 51: Barnes, M. J., H. Aksoylar, P. Krebs, T. Bourdeau, C. N. Arnold, Y. Xia, K. Khovananth, I. Engel, S. Sovath, K. Lampe, E. Laws, A. Saunders, G. W. Butcher, M. Kronenberg, K. Steinbrecher, D. Hildeman, H. L. Grimes, B. Beutler, and K. Hoebe Loss of T cell and B cell quiescence precedes the onset of microbial flora-dependent wasting disease and intestinal inflammation in Gimap5- deficient mice. J Immunol 184: Aksoylar, H. I., K. Lampe, M. J. Barnes, D. R. Plas, and K. Hoebe Loss of immunological tolerance in Gimap5-deficient mice is associated with loss of Foxo in CD4+ T cells. J Immunol 188:
205 9. Kerdiles, Y. M., D. R. Beisner, R. Tinoco, A. S. Dejean, D. H. Castrillon, R. a. DePinho, and S. M. Hedrick Foxo1 links homing and survival of naive T cells by regulating L-selectin, CCR7 and interleukin 7 receptor. Nature immunology 10: Ouyang, W., O. Beckett, Q. Ma, J.-h. Paik, R. a. DePinho, and M. O. Li Foxo proteins cooperatively control the differentiation of Foxp3+ regulatory T cells. Nature immunology 11: Wong, V. W., A. E. Saunders, A. Hutchings, J. C. Pascall, C. Carter, N. A. Bright, S. A. Walker, N. T. Ktistakis, and G. W. Butcher The autoimmunity-related GIMAP5 GTPase is a lysosome-associated protein. Self Nonself 1: Shen, D. T., J. S. Ma, J. Mather, S. Vukmanovic, and S. Radoja Activation of primary T lymphocytes results in lysosome development and polarized granule exocytosis in CD4+ and CD8+ subsets, whereas expression of lytic molecules confers cytotoxicity to CD8+ T cells. Journal of leukocyte biology 80: Adamec, E., P. S. Mohan, A. M. Cataldo, J. P. Vonsattel, and R. A. Nixon Up-regulation of the lysosomal system in experimental models of neuronal injury: implications for Alzheimer's disease. Neuroscience 100: Yen, W.-L., and D. J. Klionsky How to live long and prosper: autophagy, mitochondria, and aging. Physiology (Bethesda, Md.) 23: Schwefel, D., C. Fröhlich, J. Eichhorst, B. Wiesner, J. Behlke, L. Aravind, and O. Daumke Structural basis of oligomerization in septin-like GTPase of immunity-associated protein 2 (GIMAP2). Proceedings of the National Academy of Sciences of the United States of America 107: Pua, H. H., I. Dzhagalov, M. Chuck, N. Mizushima, and Y.-W. He A critical role for the autophagy gene Atg5 in T cell survival and proliferation. The Journal of experimental medicine 204: Jia, W., and Y.-W. He Temporal regulation of intracellular organelle homeostasis in T lymphocytes by autophagy. Journal of immunology (Baltimore, Md. : 1950) 186: Zong, W.-X., and C. B. Thompson Necrotic death as a cell fate. Genes & development 20: Hubbard, V. M., R. Valdor, B. Patel, R. Singh, A. M. Cuervo, and F. Macian Macroautophagy regulates energy metabolism during effector T cell activation. Journal of immunology (Baltimore, Md. : 1950) 185: Stephenson, L. M., B. C. Miller, A. Ng, J. Eisenberg, Z. Zhao, K. Cadwell, D. B. Graham, N. N. Mizushima, R. Xavier, H. W. Virgin, and W. Swat Identification of Atg5-dependent transcriptional changes and increases in mitochondrial mass in Atg5-deficient T lymphocytes. Autophagy 5: Pua, H. H., J. Guo, M. Komatsu, and Y.-W. He Autophagy is essential for mitochondrial clearance in mature T lymphocytes. Journal of immunology (Baltimore, Md. : 1950) 182:
206 22. Thorstenson, K. M., and A. Khoruts Generation of anergic and potentially immunoregulatory CD25+CD4 T cells in vivo after induction of peripheral tolerance with intravenous or oral antigen. Journal of immunology (Baltimore, Md. : 1950) 167: Rathmell, J. C Metabolism and autophagy in the immune system: immunometabolism comes of age. Immunological reviews 249: Mucida, D., N. Kutchukhidze, A. Erazo, M. Russo, J. J. Lafaille, and M. A. Curotto de Lafaille Oral tolerance in the absence of naturally occurring Tregs. J Clin Invest 115: Curotto de Lafaille, M. a., and J. J. Lafaille Natural and adaptive foxp3+ regulatory T cells: more of the same or a division of labor? Immunity 30: Michalek, R. D., and J. C. Rathmell The metabolic life and times of a T- cell. Immunological reviews 236: Kupfer, R., J. Lang, C. Williams-Skipp, M. Nelson, D. Bellgrau, and R. I. Scheinman Loss of a gimap/ian gene leads to activation of NF-kappaB through a MAPK-dependent pathway. Molecular immunology 44: Ilangumaran, S., M. Forand-Boulerice, S. M. Bousquet, A. Savard, P. Rocheleau, X. L. Chen, G. Dupuis, P. Poussier, G. Boulay, and S. Ramanathan Loss of GIMAP5 (GTPase of immunity-associated nucleotide binding protein 5) impairs calcium signaling in rat T lymphocytes. Mol Immunol 46: Eskelinen, E.-L Roles of LAMP-1 and LAMP-2 in lysosome biogenesis and autophagy. Molecular aspects of medicine 27: González-Polo, R.-A., P. Boya, A.-L. Pauleau, A. Jalil, N. Larochette, S. Souquère, E.-L. Eskelinen, G. Pierron, P. Saftig, and G. Kroemer The apoptosis/autophagy paradox: autophagic vacuolization before apoptotic death. Journal of cell science 118: Gutierrez, M. G., D. B. Munafó, W. Berón, and M. I. Colombo Rab7 is required for the normal progression of the autophagic pathway in mammalian cells. Journal of cell science 117: Jäger, S., C. Bucci, I. Tanida, T. Ueno, E. Kominami, P. Saftig, and E.-L. Eskelinen Role for Rab7 in maturation of late autophagic vacuoles. Journal of cell science 117: Dennemärker, J., T. Lohmüller, S. Müller, S. V. Aguilar, D. J. Tobin, C. Peters, and T. Reinheckel Impaired turnover of autophagolysosomes in cathepsin L deficiency. Biological chemistry 391: Scherz-Shouval, R., E. Shvets, E. Fass, H. Shorer, L. Gil, and Z. Elazar Reactive oxygen species are essential for autophagy and specifically regulate the activity of Atg4. EMBO J 26: Wang, R., and D. R. Green Metabolic checkpoints in activated T cells. Nature immunology 13:
207 36. Zheng, Y., G. M. Delgoffe, C. F. Meyer, W. Chan, and J. D. Powell Anergic T cells are metabolically anergic. J Immunol 183: Wang, R., C. P. Dillon, L. Z. Shi, S. Milasta, R. Carter, D. Finkelstein, L. L. McCormick, P. Fitzgerald, H. Chi, J. Munger, and D. R. Green The transcription factor Myc controls metabolic reprogramming upon T lymphocyte activation. Immunity 35: Jung, C. H., C. B. Jun, S. H. Ro, Y. M. Kim, N. M. Otto, J. Cao, M. Kundu, and D. H. Kim ULK-Atg13-FIP200 complexes mediate mtor signaling to the autophagy machinery. Mol Biol Cell 20:
208 Figure Legends Figure 1. Antigen specific TCR signaling induces a LIP phenotype, impaired Treg generation and reduced survival of Gimap5-deficient CD4 + T cells. A, In the absence of cognate antigen, Gimap5 sph/sph OT-II RAG -/- CD4 + T cells remain naïve. B, Immunoblot analysis of Foxo3 in CD4 + T cells from 10 weeks old mice. C, 4 weeks old Gimap5 sph/sph OT-II RAG -/- and WT OT-II RAG -/- mice were given 5 mg/ml ovalbumin in drinking water for 9 weeks. Spleens were harvested and stained for CD4, CD44 and CD62L for the analysis of LIP phenotype. D, With the oral administration of antigen, Gimap5 sph/sph OT- II RAG -/- CD4 + T cells displayed impaired Treg induction. Mesenteric lymph node and splenic cells were analyzed for FoxP3 expression with flow cytometry. E, Reduced survival of Gimap5 sph/sph OT-II RAG -/- CD4 + T cells after oral treatment of ovalbumin. (mean values ± SEM, n 3 mice per genotype from two independent experiments) *p < Figure 2. Analysis of lysosomes in TCR activated CD4 + T cells. Splenocytes of 4 weekold Gimap5 sph/sph and wild type mice were stimulated with plate bound anti-cd3 and anti CD28 antibodies for 16 hours. Cells were stained with anti-lamp1 and anti-lc3 antibody and analyzed with ImageStreamX. A, Lamp1 intensity, number of lysosomes and average lysosome size were determined by IDEAS software. B, Representative images of Lamp1 and LC3 staining showing colocalization of lysosomes and autophagosomes. C, Number and average size of Lamp1-LC3 colocalizing autolysosomes in CD4 + T cells were determined by IDEAS software. (n = 2 mice per genotype Representative of two independent experiments) *p <
209 Figure 3. Abnormal macroautophagy in TCR-stimulated Gimap5 sph/sph CD4 + T cells. Splenocytes of 4 week-old Gimap5 sph/sph and wild type mice were stimulated with plate bound anti-cd3 and anti-cd28 antibodies for 16 hours. A, Immunoblot analysis revealed an increased rate of the active form of LC3 in Gimap5 sph/sph CD4 + T cells. B, TCR-stimulated cells were stained with anti-lc3. LC3 intensity, number of autophagosomes and average autophagosome size were determined by IDEAS software.c, TCR-stimulated cells were stained with anti-lc3 and Mitotracker Deep Red, and analyzed with ImageStreamX. Representative images of LC3 and Mitotracker staining show the colocalization of autophagosomes and mitochondria. D, LC3 intensity, number and average size of autophagosomes containing mitochondria were determined by IDEAS software. E, Intensity of Mitotracker staining within autophagosomes was ananalyzed using IDEAS. (n = 2 mice per genotype Representative of two independent experiments) *p < 0.05 Figure 4. Abnormal mitochondrial function in TCR-stimulated Gimap5 sph/sph T cells. A, Cells were incubated with DCFDA and reactive oxygen levels were analyzed with flow cytometry. B, MACS purified CD4 + T cells from 4 week-old Gimap5 sph/sph and wild type mice were stimulated with plate bound anti-cd3 and anti-cd28 antibodies for 16 hours. Oxygen consumption rates of CD4 + T cells were analyzed with XF24 extracellular flux analyzer. (n 3 Representative of two independent experiments) *p <


210 % Survival CD44 Figure 1. A WT OT-II RAG1 -/- Gimap5 sph/sph OT-II RAG1 -/ CD62L CD62L B Gimap5 sph/sph Gimap5 sph/sph OT- Foxo3 actin Age (weeks) 200
211 CD44 C WT OT-II RAG1-/- Gimap5 sph/sph OT-II RAG1-/ Water OVA CD62L 201
212 Number of CD4 + T cells (x 10 6 ) FoxP3 D WT OT-II RAG1-/ Gimap5 sph/sph OT-II RAG1-/ Spleen MLN CD4 E dh 2 O OVA * WT OT-II RAG1-/- Gimap5 sph/sph OT-II RAG1-/- 202

213 Number of Lamp1 + LC3 + spots spots (µm 2 ) Lamp1 intensity (MFI) Number of lysosomes Average lysosome size (µm 2 ) Figure 2 A WT Gimap5 sph/sph * control αcd3/ control αcd3/28 control αcd3/28 B C control αcd3/28 Average size of Lamp1 + LC control αcd3/28 203

214 LC3 spot intensity (MFI) Number of autophagic vesicles Average size of of autophagic vesicles (µm 2 ) Figure 3. A LC3 actin WT sph WT sph αcd3 medium B WT Gimap5 sph/sph * control αcd3/28 control αcd3/28 control αcd3/28 C 204
215 Mitoctracker intensity in autophagosomes (MFI) LC3 intensity in autophagosomes containing mitochondria (MFI) Number of autophagosomes containing mitochondria Avereage size of autophagosomes containing mitochondria (µm 2 ) D WT Gimap5 sph/sph * * control αcd3/28 control αcd3/28 control αcd3/28 E control αcd3/28 205
216 Basal OCR (pmoles/min/µg protein) ATP linked OCR (pmoles/min/µg protein) Proton Leak (pmoles/min/µg protein) DCFDA (MFI) Figure 4. A * Wild type Gimap5 sph/sph B WT Gimap5 sph/sph 3.0 * * control αcd3/28 control αcd3/28 control αcd3/28 206
217 Chapter V Summary and Discussion 207
218 1. Summary Lymphopenia and associated T cell-mediated pathology are observed in animal models of autoimmunity and also in clinical practice (1). In this thesis, using an ENU mutagenesis approach, we identified sphinx mice carrying a missense mutation in Gimap5 and presenting severe lymphopenia. Using this model, we investigated the link between T cell lymphopenia and loss of immunological tolerance ultimately causing spontaneous colitis development. In chapter II, a thorough characterization of Gimap5 sph/sph mice revealed that Gimap5 is necessary for normal lymphopoiesis of NK and CD8 + T cells, and is required for the maintenance of peripheral CD4 + T and B cell populations. Also, we showed that Gimap5-deficient mice develop microbial floradependent, severe colitis associated with CD4 + T cell lymphopenia. In chapter III, we presented evidence that Gimap5 sph/sph CD4 + T cells displayed a LIP phenotype, produced inflammatory cytokines, and were critical to the development of intestinal inflammation in Gimap5-deficient mice. Moreover, FoxP3 + regulatory T cells were reduced in frequency in the mesenteric lymph nodes, and their suppressive capacity was impaired in the absence of Gimap5. Importantly, the loss of CD4 + T cell quiescence was associated with a loss of Foxo protein expression in Gimap5 sph/sph mice. In chapter IV, we investigated the role of antigen-specific TCR signaling in the loss of CD4 + T cell homeostasis, survival and Treg induction. Our results showed that oral treatment with cognate antigen causes a LIP phenotype in CD4 + T cells and fails to induce the conversion naïve CD4 + T cells into Treg cells in Gimap5 sph/sph OT-II RAG -/- mice. Moreover, we observed that upon TCR stimulation, Gimap5 sph/sph CD4 + T cells display abnormalities in lysosome function, mitochondrial autophagy and mitochondrial function suggesting a role for Gimap5 in the regulation of cellular bioenergetics during T cell 208
219 activation. Overall, our studies reveal a critical role for Gimap5 in maintaining CD4 + T cell homeostasis and immunological tolerance, and in addition, reveal how Gimap5 dysfunction may contribute to the development of autoimmune diseases. Elucidating the role of Gimap5 in the breakdown of T cell homeostasis and tolerance could have therapeutic implications for autoimmune diseases associated with T cell lymphopenia. Our major findings are discussed in the following sections. 2. Gimap5 and autoimmunity Previous studies showed that mutations in Gimap5 are associated with autoimmunity. Spontaneous diabetes observed in Biobreeding rats which carry a truncation mutation of Gimap5, and the association of GIMAP5 polymorphisms with SLE in humans strongly suggested a role for Gimap5 in autoimmunity (2, 3). Our work, for the first time, directly linked Gimap5 to the maintenance of immunological tolerance in mice, and showed that absence of Gimap5 leads to severe T cell-dependent colitis. Several lines of evidence suggest that lymphopenia is linked to autoimmunity. Lymphopenic conditions are associated with; 1) increased frequencies of T cells entering cell cycle; 2) a dysregulated balance between Tregs and conventional T cells; and 3) acquisition of a T effector function with an oligoclonal repertoire (4). These phenotypes are characteristics of LIP and favor the onset of autoimmunity. Clinical studies show that reduced numbers of lymphocytes are observed in autoimmune diseases such as SLE and Rheumatoid Arthiritis (5, 6). In humans, mutations in several genes such as Adenosine deaminase, ZAP70 and Interleukin-2 receptor α cause defects in T cell development, TCR-signaling and cytokine signaling, respectively, and lead to T cell immunodeficiencies and immune dysregulation (7-9). Other examples of 209
220 lymphopenia associated autoimmunity are often observed in common variable immunodeficiency (CVID) patients, but the underlying genetic defects of CVID remain poorly understood (10). Our work indicates that Gimap5-deficient mice present similar features to those observed in human immunodeficiencies and suggests that GIMAP5 is a prime candidate gene involved in lymphopenia associated autoimmunity in humans. Our results showed that Gimap5 sph/sph CD4 + T cells display an impaired TCRinduced proliferation but do not show defects in MAPK and NF B signaling pathways downstream of TCR signaling or in the production of inflammatory cytokines; IFN- and IL-17. Moreover, Gimap5 sph/sph CD4 + T cells exhibit a LIP phenotype and cause intestinal inflammation. In parallel, previous studies established that adoptively transferred naïve CD4 + T cells upregulate activation markers, produce inflammatory cytokines and lead to intestinal inflammation in chronically lymphopenic SCID mice (11, 12). Importantly, cotransfer of regulatory T cells prevents pathology in this T cellmediated colitis model (13). Similarly, we found that adoptive transfer of wild type Treg cells into Gimap5-deficient mice prevented the development of colitis. Further, we demonstrated that mln CD4 + T cells in Gimap5-deficient mice contain a population of IFN- + IL-17 + double positive cells. IFN- + IL-17 + CD4 + T cells were also observed in Crohn s disease patients (14). Moreover, others showed that IFN- + IL-17 + CD4 + T cells were observed in the T-cell-mediated colitis in SCID mice and their appearance was induced by IL-23 (15). In this colitis model, while IL-23 induced the accumulation of Th17 cells in the gut, it repressed Treg differentiation (15). However, whether IL-23 plays a role in the differentiation of IFN- + IL-17 + CD4 + T cells and in the development of colitis in Gimap5 sph/sph mice remains to be established. 210
221 In chapter II, we showed that intestinal inflammation in Gimap5-deficient mice is prevented by antibiotics treatment. Similarly, others have shown that in the adoptive T cell-transfer colitis model, development of intestinal inflammation was attenuated when recipient mice were raised in germ-free conditions (16). Therefore, our results suggest that the intestinal inflammation is driven by an abnormal response of Gimap5 sph/sph CD4 + T cells to commensal antigens. Similarly, others have reported that intestinal inflammation was not observed in Gimap5 knock-out mice (17). These contrasting results pose the question whether the composition of the commensal bacteria is different between Gimap5 sph/sph mice and Gimap5 knock-out mice as they are housed in different facilities. Several studies suggest that commensal bacteria can influence effector and suppressor T cell responses. For instance, segmented filamentous bacteria (SFB) and Bacteriodes fragilis have been reported to induce Th17 and Treg cell development in the gut, respectively (18, 19). It is possible that the composition of commensal bacteria in Gimap5 sph/sph mice in our facility promotes an increased Th17/Treg ratio and contributes to colitis development. Although this is a possibility, it is likely that the early morbidity of Gimap5 knock-out mice used in Schulteis et al s study is also caused by intestinal inflammation and wasting disease (17). Although they proposed that their mice succumb to early death as a consequence of liver disease, our data indicate that liver disease alone does not drive the early death of Gimap5-deficient mice. First, we were unable to find evidence for metabolic liver dysfunction as measured by serum concentrations of bilirubin and albumin (data not shown). Second, we observed that when Gimap5 sph/sph mice received wild type splenocytes, they survived over a year and did not develop colitis, yet still developed liver disease (data not 211
222 shown). Thus, although differences in the composition of the microflora may accelerate colitis development in our facility, our data suggest that T cell-intrinsic defects observed in the absence of Gimap5 are responsible for colitis development and wasting disease. The immune dysregulation we observed in Gimap5-deficient mice is associated with the loss of Foxo transcription factors. In chapter III, we showed that CD4 + T cells progressively lost the expression of Foxo1, 3 and 4 in Gimap5 sph/sph mice starting with 4-5 weeks of age. Previously, others have shown that Foxo3 limits cell cycle progression by inducing the expression of p27 in T cells (20). Moreover, Foxo3 deficient T cells show a hyper-proliferative phenotype in vivo (21). Similarly, our results showed that Gimap5-deficient CD4 + T cells displayed reduced p27 expression, increased phosphorylation of Retinoblastoma protein and a progressive increase in cell cycle entry. Therefore, loss of Foxo expression might be involved in the loss of CD4 + T cell quiescence in Gimap5-deficient mice. Foxo proteins are regulated through phosphorylation by AKT kinase which induces their translocation from nucleus to cytoplasm and targets them for proteasomal degradation. However, we did not observe increased AKT activity in Gimap5-deficient CD4 + T cells (data not shown). Another mechanism that regulates Foxo expression is mediated by the E3 ubiquitin ligase S- phase kinase-associated protein-2 (Skp2). In a mouse lymphoma model, increased expression of Skp2 resulted in the loss of Foxo1 expression (22). Skp2-mediated ubiquitination and subsequent proteasomal degradation could account for the degradation of Foxo proteins in Gimap5-deficient CD4 + T cells. However, we were unable to restore Foxo expression when Gimap5-deficient CD4 + T cells were treated with proteasome inhibitors or with cathepsin and caspase inhibitors (data not shown). 212
223 MicroRNAs (mirnas) may play a role in the loss of Foxo protein expression in Gimap5-deficient CD4 + T cells. It has been shown that several mirnas target Foxo mrnas in different cell types (23-26). Our results in chapter III showed that Foxo1 mrna was reduced in Gimap5-deficient CD4 + T cells however, mrna levels of Foxo3 and Foxo4 were normal. MiRNA mediated suppression mechanisms involve cleavage or translational repression of target mrnas (27). Therefore, it is possible that mirnas targeting Foxo mrnas may cleave the Foxo1 mrna and repress the translation of Foxo3 and Foxo4 mrnas in Gimap5-deficient CD4 + T cells. To test whether Foxo protein expression is lost via mirna mediated suppression, expression levels of mirnas that are known to target Foxo mrnas can be analyzed. If any mirna that target Foxo mrnas is found to be highly expressed, Gimap5-deficient CD4 + T cells can be treated with specific antagomirs to test whether Foxo protein expression would be restored. An extrinsic mechanism for the loss of Foxo expression in Gimap5-deficient CD4 + T cells may be triggered by lymphopenia. It is possible that TCR or cytokine signaling under lymphopenic conditions can induce loss of Foxo expression in Gimap5- deficient CD4 + T cells but not in wild type CD4 + T cells. To test whether lymphopenia plays a role in the loss of Foxo expression in Gimap5-deficient CD4 + T cells, an experimental approach would involve the adoptive transfer of naïve CD4 + T cells from Gimap5-deficient mice into a non-lymphopenic environment. Analysis of the LIP phenotype and Foxo expression in Gimap5-deficient CD4 + T cells in the nonlymphopenic conditions would help us understand whether lymphopenic environment 213
224 plays a role in the LIP phenotype and loss of Foxo expression in Gimap5-deficient CD4 + T cells. The reduced Foxo expression may be critically linked to the loss of immunological tolerance in Gimap5-deficient mice. Recent studies demonstrated that Foxo1 is required for the generation and function of thymic and induced FoxP3 + regulatory T cells. Foxo1 induces the expression of FoxP3 + and is required for the responsiveness of T cells to TGF-β signaling (28-30). In chapter III, we demonstrated that Tregs were dramatically reduced in numbers and frequencies in the mesenteric lymph nodes and were unable to prevent the development of colitis in the absence of Gimap5. Furthermore, in chapter IV, we showed that antigen-induced in vivo generation of peripheral FoxP3 + Tregs was severely impaired in the absence of Gimap5. In contrast, thymic frequency of FoxP3 + CD4SP cells was normal in Gimap5-deficient mice (data not shown). Therefore, our work shows that absence of Gimap5 impairs the generation of induced Treg cells and thereby leads to loss of immune tolerance. Given the significance of Foxo1 protein in Treg development and maintenance, loss of Foxo proteins in the absence of Gimap5 could be responsible for Treg defects. Whether the loss of Foxo proteins impairs TGF-β signaling mediated induction of Tregs in Gimap5 sph/sph CD4 + T cells need to be evaluated. Our work showed that loss of Foxo protein expression was mainly seen in CD44 high CD62L low effector CD4 + T cell population in Gimap5-deficient mice, suggesting that this defect may be triggered by TCR activation. In order to understand the role of TCR signaling in Gimap5-deficient T cell phenotypes including survival, LIP and Treg induction, we generated Gimap5 sph/sph OT-II RAG -/- mice. OT-II RAG -/- T cells do not 214
225 express endogenous TCR chains and are only specific for chicken ovalbumin, so they do not respond to self or intestinal bacterial antigens (31, 32). Moreover, oral treatment of the OT-II RAG -/- mice with OVA establishes tolerizing conditions for T cells (33, 34). We found that in the absence of OVA, Gimap5 sph/sph OT-II RAG -/- CD4 + T cells remained naïve similar to Wild Type cells. However, oral treatment with OVA resulted in the acquisition of a LIP phenotype, reduced survival and impaired Treg generation in Gimap5 sph/sph OT-II RAG -/- CD4 + T cells. Thus, our results suggest that TCR signaling in Gimap5-deficient CD4 + T cells leads to an abnormal response under tolerizing conditions. 2.1 Clinical Implications Our results show a critical role for Gimap5 in lymphopenia, immune dysfunction and autoimmune disease which may have some clinical implications. Previously, polymorphisms in the poly-adenylation site of GIMAP5 gene have been shown to cause decreased Gimap5 mrna expression and to be associated with increased risk to SLE (2). Although SLE patients carrying the GIMAP5 polymorphisms have not been examined for lymphopenia, earlier studies presented that lymphopenia is a frequently observed clinical manifestation in SLE patients and may contribute to disease severity (35, 36). Moreover, reduced Treg frequencies observed in active SLE patients (37, 38) strongly associate with the presentation of lymphopenia (39). Furthermore, in lymphopenic SLE patients, effector T cells have been shown to resist suppression by Tregs (39) suggesting that lymphopenia leads to immune dysregulation and is a major factor that increase disease severity in SLE. As our results demonstrated that complete dysfunction of Gimap5 results in the generation of inflammatory CD4 + T cells with a LIP 215
226 phenotype, it is possible that SLE patients carrying the GIMAP5 polymorphisms may present some characteristics observed in CD4 + T cells from Gimap5 sph/sph mice. Given that the poly-adenylation polymorphisms is likely to affect overall protein expression, it would be critical to assess this in CD4 + T cells of SLE patients that are carriers for the polymorphism. In addition, it will be interesting to see whether CD4 + T cells display lymphopenia-associated effector phenotype, impaired Treg function and decreased Foxo protein expression in the inflamed tissues. Western blot analysis of lymphocytes in Gimap sph/sph mice shows a complete loss of Gimap5 protein expression due to the mutation in the P-loop motif of the GTP binding domain. Although the GIMAP5 poly-adenylation polymorphisms observed in SLE patients cause a mild decrease in Gimap5 mrna expression (2), there are no reported polymorphisms for GIMAP5 causing a complete loss of function in human patients. However, lymphopenia and autoimmunity are frequently observed clinical manifestations in CVID patients (40) and majority of the genetic causes in CVID is undetermined. Therefore, it is possible that mutations in human GIMAP5 gene which abolish protein function might be associated with immunodeficiencies observed in CVID. Also, it is important to note that modifier genes may influence the phenotype of Gimap5 genetic aberrancies. Lymphopenia induced by the Gimap5 lyp mutation causes autoimmunity only in susceptible strains of rats suggesting the contribution of other genes affecting T cell function. Together, different polymorphisms in GIMAP5 may cause varying degrees of phenotypes from mild to severe lymphopenia and contribute to impaired immune homeostasis and loss of immune tolerance in humans. Thus, a 216
227 comprehensive assessment of GIMAP5 polymorphisms in CVID patients would be important to understand whether Gimap5 plays a role in this disease. 3. Role of Gimap5 in lymphocyte development and homeostasis Our study demonstrates that Gimap5 is critical for lymphocyte development and homeostasis. Although previous studies with Biobreeding rats suggested that Gimap5 is an important regulator of T cell development and survival (41, 42), our work in this thesis demonstrates a more extensive role for Gimap5 in mouse lymphocyte subsets. Our results showed that mouse Gimap5 regulates the development of natural killer and CD8 + T cells and is also essential for maintaining the peripheral populations of CD4 + T and B cells. Recently, a mouse model with a genetic deletion of Gimap5 has been generated and these mice displayed deficiencies in NK and T lymphocytes although B cell lymphopenia was not examined in detail (17). Similarly, conditional deletion of Gimap1 caused T and B cell lymphopenia in mice (43). These results indicate that Gimap family proteins may function in a coordinated fashion during the development and peripheral stages of lymphocytes. In chapter II, we showed that peripheral CD8 + T cell population is virtually absent in Gimap5-deficient mice. Interestingly, positive selection of thymocytes was not affected by the loss of Gimap5 function. However, our results demonstrated that Gimap5-deficient CD8 SP thymocytes failed to downregulate the expression of surface markers, CD24 and CD69. Moreover, CD8 SP thymocytes showed an impaired upregulation of Qa-2 in Gimap5 -/- mice (17). These results strongly suggest that Gimap5 is required for the post-selection maturation of CD8 + T cells. It is known that positively 217
228 selected SP thymocytes undergo 1-2 additional cell divisions before being exported from thymus (44). Therefore, it is possible that Gimap5 plays a role in the intrathymic proliferation of CD8 SP thymocytes. Our work demonstrated that the loss of Gimap5 impairs CD4 + T cell homeostasis predominantly in the periphery. The numbers of peripheral CD4 + T cells progressively decreased in Gimap5 sph/sph mice during the period from 3-weeks to 6-weeks of age. In order to investigate the underlying defects causing progressive CD4 + T cell lymphopenia, we examined survival and proliferation of these cells. It was suggested that Gimap5 interacts with Bcl-2 and regulates T cell survival by favoring anti-apoptotic pathways (45). Another report proposed that Gimap5 interacts and stabilizes Mcl-1 protein in hematopoietic stem cells (46). However, we did not observe differences in the expression of Bcl-2 and Bim proteins in in vitro activated Gimap5-deficient lymphocytes compared to wild type cells. Moreover, inhibiting both intrinsic and extrinsic apoptotic pathways by the ablation of Bim, Fas or Bax-Bak (data not shown) together in Gimap5- deficient background did not prevent the progression of T cell lymphopenia. Furthermore, deletion of tumor suppressive transcription factor p53 did not improve the lymphopenia in Gimap5-deficient mice (data not shown). Thus, our results strongly suggest that progressive CD4 + T cell lymphopenia in Gimap5-deficient mice is not due to increased apoptosis. On the other hand, our results showed defects in the proliferation of Gimap5-deficient CD4 + T cells. First, Gimap5-deficient CD4 + T cells proliferated poorly upon TCR stimulation in vitro. Second, interestingly, Gimap5- deficient CD4 + T cells showed increased occupation in the S phase of the cell cycle and showed increased BrDU incorporation in vivo suggesting that these cells display 218
229 increased cell cycle entry but do not efficiently complete the cell cycle and proliferate. Therefore, our results suggest that lymphopenia and survival defects in Gimap5- deficient mice is not a result of mitochondrial and FAS mediated apoptotic mechanisms but probably due to intrinsic defects during homeostatic TCR signaling and proliferation. As combined deficiency of Bax and Bak, and deficiency of Fas did not restore the CD4 + T cell lymphopenia in Gimap5-deficient mice (Data not shown), one question remains is how Gimap5-deficient CD4 + T cells lose their viability. It is well established that excessive production of ROS causes oxidative stress leading to damage of organelles and intracellular molecules (47). Excessive ROS produced by mitochondria can cause DNA damage which induces the hyper-activation of Poly (ADP-Ribose) Polymerase (PARP) resulting in necrosis (48). Moreover, inhibition of PARP activity prevents ROS induced necrotic cell death (48). Since Gimap5-deficient CD4 + T cells displayed increased levels of ROS, it is possible that Gimap5-deficient CD4 + T cells may die due to ROS induced necrosis. To test this possibility ex vivo, naïve Gimap5- deficient CD4 + T cells can be activated by anti-cd3 in the presence of the sirna for PARP or mock sirna. Additionally, ROS inhibitor N-acetylcysteine (NAC) can be added as a control. Analysis of cell viability and features of necrotic cell death, such as cell swelling, would help us understand whether necrosis plays a role in the decreased viability of Gimap5-deficient CD4 + T cells. Alternatively, Gimap5 sph/sph mice can be bred with PARP1-deficient mice to test whether loss of PARP1 would prevent CD4 + T cell lymphopenia in Gimap5 sph/sph mice. Although the treatment of Gimap5-deficient mice with the ROS inhibitor MnTBAP did not rescue T cell lymphopenia (Data not shown), this treatment may not efficiently inhibit ROS and thereby may fail to prevent T cell 219
230 necrosis in vivo. ROS inhibitor treatment in ex vivo cell culture would provide a more sensitive kinetic analysis model for monitoring T cell viability and necrosis. 4. Molecular function of Gimap5 Although Gimap5 has been shown to be critical for T cell development and homeostasis, its molecular function has not been clearly identified. Cellular localization and structure of Gimap5 can give us clues about its molecular function. Gimap5 protein contains a hydrophobic sequence in its C-terminal which indicates that it is located in the membranous compartments in the cell. Previous studies on the cellular localization of Gimap5 protein produced controversial results. Although several studies suggested that Gimap5 localizes to ER or mitochondria, those results were probably affected by the over-expression methods used (45, 49). A better analysis using monoclonal antibodies for Gimap5 suggested that endogenous Gimap5 localizes to lysosomes in T cells (50). Recent studies demonstrate that Gimap5 is structurally similar to other GTPases such as dynamins and septins which regulate vesicular dynamics in the intracellular membranes and vesicles (51). In chapter IV, we investigated lysosomal function in Gimap5-deficient CD4 + T cells. Our results indicate that Gimap5 is not essential for lysosomal biogenesis. However, larger average size of lysosomes in activated Gimap5 sph/sph CD4 + T cells suggests an abnormal lysosome function. Lysosomes regulate cellular homeostasis by degrading autophagic vesicles (52). Regulation of organelle homeostasis by autophagy is critical for T cell proliferation (53). Our results in chapter IV demonstrate that autophagic process in activated Gimap5- deficient CD4 + T cells is abnormal. Although the lysosome-autophagosome fusion was 220
231 not affected by the loss of Gimap5, autophagic vesicles and autophagolysosomes were larger in size in activated Gimap5-deficient CD4 + T cells. A similar defect has been shown in the absence of the lysosomal GTPase Rab7 (54, 55). Rab7 is required for the final maturation of late autophagosomes after lysosome fusion and its absence causes accumulation of autophagosomes with unusually large size. Moreover, deficiency of Cathepsin L causes impaired turnover of autophagic vesicles and leads to accumulation of large autophagic compartments (56). Therefore, similar defects in the degradation of autophagic components can cause accumulation of large autophagolysosomes in Gimap5 sph/sph CD4 + T cells. Thus, our results suggest that in the absence of Gimap5, autophagy is initiated in activated CD4 + T cells; however final maturation of autophagolysosomes or degradation of their contents may be impaired. Our observations showing large autophagosomes in activated Gimap5 sph/sph CD4 + T cells may be due to either a block in the final steps of autophagy or an increased autophagic activity. The experimental approach we used allowed us to detect the autophagosomes at a certain time point during T cell activation. However, autophagy is a dynamic process and measuring the autophagic flux is needed in order to distinguish between a block and an increase in autophagy. Autophagic flux can be determined by measuring the quenching of GFP fused with LC3 when the autophagosomes fuse with lysosomes. Our initial attempts to determine autophagic flux failed due to poor viability of Gimap5 sph/sph CD4 + T cells after transduction with lentiviral vectors expressing LC3-GFP fusion protein. Therefore, the autophagic flux in activated Gimap5 sph/sph CD4 + T cells is an important issue to be addressed. 221
232 If larger autophagosomes in Gimap5 sph/sph CD4 + T cells are due to increased autophagy, then this would suggest a potential insufficiency of bioenergetic resources during activation. During TCR activation and proliferation, T cells utilize a highly glycolytic and glutaminolytic metabolism which provides intermediates for the anabolic pathways such as synthesis of amino acids, nucleic acids and lipids (57). Nevertheless, if bioenergetic resources are limited during T cell activation, autophagy may be induced to provide resources. Ulk1/2 pathway triggers autophagy in order to meet the bioenergetics demands of activated T cells (57). While Rag GTPases activate mtorc1 pathway in the presence of amino acids (58), lack of bioenergetics resources can inactivate mtorc1 which, then, can release the inhibition on Ulk1/2 proteins (59). Therefore, the status of mtorc1 Ulk1/2 pathway needs to be determined in Gimap5- deficient CD4 + T cells during TCR activation. There is growing evidence that metabolic pathways are required for the development and peripheral homeostasis of T cells. For example, it has been shown that thymocytes deficient for both Akt1 and Akt2 isoforms poorly mature beyond the DN3 stage (60, 61). The impaired transition of Akt1 -/- Akt2 -/- thymocytes from DN3 to DN4 was associated with an impaired glucose uptake (60). Moreover, downstream of Akt signaling, inhibition of mtorc1 complex led to T cell anergy (62). As our results showed that Gimap5-deficient thymocytes efficiently develop into DP stage and peripheral Gimap5-deficient CD4 + T cells show an activated phenotype, a block in the Akt/mTOR signaling pathway is not likely a cause of the phenotypes observed in Gimap5 sph/sph CD4 + T cells. On the other hand, a recent report demonstrated that LKB1- deficient T cells displayed a hyper-activated phenotype, increased production of 222
233 inflammatory cytokines, increased glucose uptake, but showed reduced viability and impaired TCR-induced proliferation (63). Similarly, downstream of LKB1, loss of AMPKα1 also resulted in an increased T cell activation and inflammatory cytokine production (63). The hyper-activated phenotypes of LKB1- and AMPKα1-deficient T cells were associated with increased mtorc1 activation and were reversed by the inhibition of mtorc pathway (63). Similar phenotypes between LKB1- and Gimap5- deficient T cells suggest that Gimap5 deficient peripheral CD4 + T cells may have increased mtorc1 activation. As AMPK signaling negatively regulates mtor pathway (64), the status of mtorc1 activation and its downstream targets needs to be examined in Gimap5-deficient CD4 + T cells. Our results in chapter IV which show large autophagosomes containing mitochondria in activated Gimap5-deficient CD4 + T cells could indicate an impaired recycling of mitochondria. Recent studies have demonstrated that autophagy is essential for recycling of mitochondria during T cell proliferation (53, 65). Moreover, mice deficient for autophagy genes Atg3, Atg5 or Atg7 showed impaired T cell proliferation and lymphopenia which were associated with defective mitochondrial turnover (53, 65, 66). Therefore, it is possible that impaired degradation pathways in autophagosomes in the absence of Gimap5 can cause an inefficient turnover of mitochondria leading to defects in TCR-induced proliferation of CD4 + T cells and progressive lymphopenia in Gimap5-deficient mice. Alternatively, the larger size of autophagosomes in Gimap5 sph/sph CD4 + T cells might be due to an elevated autophagic activity. It has been shown that reactive oxygen species (ROS) produced by mitochondria trigger autophagy through an Atg4 regulated mechanism (67). Therefore, 223
234 it is possible that elevated ROS levels in Gimap5-deficient CD4 + T cells may trigger autophagic pathway. In Chapter II, our data shows that Gimap5-deficient B cells display an impaired proliferation with IgM cross-linking, but do proliferate with LPS stimulation. If autophagic mechanisms are impaired in the absence of Gimap5, this may differentially affect the proliferation of B cells in response to IgM or LPS. Limited literature exists about the role of autophagy in B cells (68). One study showed that IgM stimulation induces the formation of autophagosomes in B cells (69). Moreover, bone marrow chimera experiments showed that number of peripheral B cells was significantly reduced in the absence of Atg5 (65). However, the role of autophagy in B cell proliferation is not established yet. It is possible that Gimap5 may be required for the efficient autophagic turnover of organelles downstream of BCR signaling and this autophagic turnover may be required for BCR induced proliferation as observed in T cells. On the other hand, as TLR4 signaling activates different intracellular pathways than BCR signaling, it is possible that the induction of autophagy may not be required for B cell proliferation triggered by LPS induced TLR4 signaling. Alternatively, autophagy may be induced in both TLR4 and BCR signaling, but may involve different mechanisms. For example, TLR4 induced proliferation may not require Gimap5 for autophagy while BCR induced proliferation requires Gimap5. Therefore, it needs to be determined whether there is a differential requirement for autophagy between BCR versus LPS induced B cell proliferation and whether Gimap5-deficient B cells display abnormal autophagy upon BCR stimulation. 224
235 One important question remains is what molecular function Gimap5 exerts in autophagic turnover. The nucleotide coordination and oligomerization mode of Gimap proteins is strikingly similar to dynamin GTPases (51). Dnm1 and Vps1 are two dynamin-related GTPases in yeast involved in endocytosis, endosomal trafficking and peroxisome biogenesis (70-72). Dnm1 has a human orthologue named Drp1 which regulates mitochondrial division (73). However, no human or mouse orthologue is found for Vps1. Importantly, Vps1 has been shown to be required for the fission of vesicles from vacuoles in Schizosaccharomyces pombe (74). Similarly, Gimap5 may be involved in scission and fission of intracellular membranous compartments. Therefore, one possible function for Gimap5 could be the fission of vesicles from autophagolysosomes after degradation of their contents. In this scenario, Gimap5 can facilitate sorting of recycled bioenergetic resources from autophagolysosomes. This function of Gimap5 can be critical during T cell proliferation which is an energy demanding process (65). Thus, absence of Gimap5 could impair T cell homeostasis and lead to lymphopenia by blocking the turnover of cellular macromolecules and organelles. In conclusion, the work presented in this thesis establishes the critical role of Gimap5 in CD4 + T cell homeostasis and maintenance of immunological tolerance. Our work contributes to the understanding of Gimap5 regulated mechanisms involved in regulatory T cell generation and T cell-mediated autoimmunity. Furthermore, our work provides a basis for further investigation of the Gimap5 s role in lysosomal and autophagosomal pathways following T cell activation. 225

236 1 Loss of Gimap5: reduced survival, lymphopenia 2 Lymphopenia-induced proliferation (LIP) 4 Loss of gut homeostasis /mucosal barrier function CD4 + T cells CD62L CD44 FoxO1 FoxO3 FoxO4 Th1/17 cytokines MØ CD4 + FoxP3 + T cells 3 Loss of immune suppressive capacity PMNs Figure 1. Scheme of the events causing colitis in Gimap5-deficient mice. Gut DC TCR signaling? Autophagosome Lysosome Fusion Gimap5 WT CD4 + T cell mln Gimap5 sph/sph CD4 + T cell MT Damage Recycling?? Maturation FoxP3 + CD44 Low CD62L High Quiescent Maintenance of homeostasis Tolerance Induction CD44 High CD62L Low LIP phenotype Reduced survival Lymphopenia Impaired Treg induction? Degradation T cell intrinsic role of Gimap5 Figure 2. Model for the possible roles of Gimap5 after TCR signaling 226
Central tolerance. Mechanisms of Immune Tolerance. Regulation of the T cell response
 Immunoregulation: A balance between activation and suppression that achieves an efficient immune response without damaging the host. Mechanisms of Immune Tolerance ACTIVATION (immunity) SUPPRESSION (tolerance)
Immunoregulation: A balance between activation and suppression that achieves an efficient immune response without damaging the host. Mechanisms of Immune Tolerance ACTIVATION (immunity) SUPPRESSION (tolerance)
Mechanisms of Immune Tolerance
 Immunoregulation: A balance between activation and suppression that achieves an efficient immune response without damaging the host. ACTIVATION (immunity) SUPPRESSION (tolerance) Autoimmunity Immunodeficiency
Immunoregulation: A balance between activation and suppression that achieves an efficient immune response without damaging the host. ACTIVATION (immunity) SUPPRESSION (tolerance) Autoimmunity Immunodeficiency
Adaptive immune responses: T cell-mediated immunity
 MICR2209 Adaptive immune responses: T cell-mediated immunity Dr Allison Imrie allison.imrie@uwa.edu.au 1 Synopsis: In this lecture we will discuss the T-cell mediated immune response, how it is activated,
MICR2209 Adaptive immune responses: T cell-mediated immunity Dr Allison Imrie allison.imrie@uwa.edu.au 1 Synopsis: In this lecture we will discuss the T-cell mediated immune response, how it is activated,
T cell maturation. T-cell Maturation. What allows T cell maturation?
 T-cell Maturation What allows T cell maturation? Direct contact with thymic epithelial cells Influence of thymic hormones Growth factors (cytokines, CSF) T cell maturation T cell progenitor DN DP SP 2ry
T-cell Maturation What allows T cell maturation? Direct contact with thymic epithelial cells Influence of thymic hormones Growth factors (cytokines, CSF) T cell maturation T cell progenitor DN DP SP 2ry
Objectives. Abbas Chapter 11: Immunological Tolerance. Question 1. Question 2. Question 3. Definitions
 Objectives Abbas Chapter 11: Immunological Tolerance Christina Ciaccio, MD Children s Mercy Hospitals and Clinics February 1, 2010 To introduce the concept of immunologic tolerance To understand what factors
Objectives Abbas Chapter 11: Immunological Tolerance Christina Ciaccio, MD Children s Mercy Hospitals and Clinics February 1, 2010 To introduce the concept of immunologic tolerance To understand what factors
T cell development October 28, Dan Stetson
 T cell development October 28, 2016 Dan Stetson stetson@uw.edu 441 Lecture #13 Slide 1 of 29 Three lectures on T cells (Chapters 8, 9) Part 1 (Today): T cell development in the thymus Chapter 8, pages
T cell development October 28, 2016 Dan Stetson stetson@uw.edu 441 Lecture #13 Slide 1 of 29 Three lectures on T cells (Chapters 8, 9) Part 1 (Today): T cell development in the thymus Chapter 8, pages
The development of T cells in the thymus
 T cells rearrange their receptors in the thymus whereas B cells do so in the bone marrow. The development of T cells in the thymus The lobular/cellular organization of the thymus Immature cells are called
T cells rearrange their receptors in the thymus whereas B cells do so in the bone marrow. The development of T cells in the thymus The lobular/cellular organization of the thymus Immature cells are called
5/1/13. The proportion of thymus that produces T cells decreases with age. The cellular organization of the thymus
 T cell precursors migrate from the bone marrow via the blood to the thymus to mature 1 2 The cellular organization of the thymus The proportion of thymus that produces T cells decreases with age 3 4 1
T cell precursors migrate from the bone marrow via the blood to the thymus to mature 1 2 The cellular organization of the thymus The proportion of thymus that produces T cells decreases with age 3 4 1
Immunological Tolerance
 Immunological Tolerance Introduction Definition: Unresponsiveness to an antigen that is induced by exposure to that antigen Tolerogen = tolerogenic antigen = antigen that induces tolerance Important for
Immunological Tolerance Introduction Definition: Unresponsiveness to an antigen that is induced by exposure to that antigen Tolerogen = tolerogenic antigen = antigen that induces tolerance Important for
Immune Regulation and Tolerance
 Immune Regulation and Tolerance Immunoregulation: A balance between activation and suppression of effector cells to achieve an efficient immune response without damaging the host. Activation (immunity)
Immune Regulation and Tolerance Immunoregulation: A balance between activation and suppression of effector cells to achieve an efficient immune response without damaging the host. Activation (immunity)
Effector mechanisms of cell-mediated immunity: Properties of effector, memory and regulatory T cells
 ICI Basic Immunology course Effector mechanisms of cell-mediated immunity: Properties of effector, memory and regulatory T cells Abul K. Abbas, MD UCSF Stages in the development of T cell responses: induction
ICI Basic Immunology course Effector mechanisms of cell-mediated immunity: Properties of effector, memory and regulatory T cells Abul K. Abbas, MD UCSF Stages in the development of T cell responses: induction
Tolerance, autoimmunity and the pathogenesis of immunemediated inflammatory diseases. Abul K. Abbas UCSF
 Tolerance, autoimmunity and the pathogenesis of immunemediated inflammatory diseases Abul K. Abbas UCSF Balancing lymphocyte activation and control Activation Effector T cells Tolerance Regulatory T cells
Tolerance, autoimmunity and the pathogenesis of immunemediated inflammatory diseases Abul K. Abbas UCSF Balancing lymphocyte activation and control Activation Effector T cells Tolerance Regulatory T cells
Effector T Cells and
 1 Effector T Cells and Cytokines Andrew Lichtman, MD PhD Brigham and Women's Hospital Harvard Medical School 2 Lecture outline Cytokines Subsets of CD4+ T cells: definitions, functions, development New
1 Effector T Cells and Cytokines Andrew Lichtman, MD PhD Brigham and Women's Hospital Harvard Medical School 2 Lecture outline Cytokines Subsets of CD4+ T cells: definitions, functions, development New
T Cell Development. Xuefang Cao, MD, PhD. November 3, 2015
 T Cell Development Xuefang Cao, MD, PhD November 3, 2015 Thymocytes in the cortex of the thymus Early thymocytes development Positive and negative selection Lineage commitment Exit from the thymus and
T Cell Development Xuefang Cao, MD, PhD November 3, 2015 Thymocytes in the cortex of the thymus Early thymocytes development Positive and negative selection Lineage commitment Exit from the thymus and
SEVENTH EDITION CHAPTER
 Judy Owen Jenni Punt Sharon Stranford Kuby Immunology SEVENTH EDITION CHAPTER 16 Tolerance, Autoimmunity, and Transplantation Copyright 2013 by W. H. Freeman and Company Immune tolerance: history * Some
Judy Owen Jenni Punt Sharon Stranford Kuby Immunology SEVENTH EDITION CHAPTER 16 Tolerance, Autoimmunity, and Transplantation Copyright 2013 by W. H. Freeman and Company Immune tolerance: history * Some
T Cell Receptor & T Cell Development
 T Cell Receptor & T Cell Development Questions for the next 2 lectures: How do you generate a diverse T cell population with functional TCR rearrangements? How do you generate a T cell population that
T Cell Receptor & T Cell Development Questions for the next 2 lectures: How do you generate a diverse T cell population with functional TCR rearrangements? How do you generate a T cell population that
Lecture outline. Immunological tolerance and immune regulation. Central and peripheral tolerance. Inhibitory receptors of T cells. Regulatory T cells
 1 Immunological tolerance and immune regulation Abul K. Abbas UCSF 2 Lecture outline Central and peripheral tolerance Inhibitory receptors of T cells Regulatory T cells 1 The immunological equilibrium:
1 Immunological tolerance and immune regulation Abul K. Abbas UCSF 2 Lecture outline Central and peripheral tolerance Inhibitory receptors of T cells Regulatory T cells 1 The immunological equilibrium:
T Cell Development II: Positive and Negative Selection
 T Cell Development II: Positive and Negative Selection 8 88 The two phases of thymic development: - production of T cell receptors for antigen, by rearrangement of the TCR genes CD4 - selection of T cells
T Cell Development II: Positive and Negative Selection 8 88 The two phases of thymic development: - production of T cell receptors for antigen, by rearrangement of the TCR genes CD4 - selection of T cells
The Adaptive Immune Responses
 The Adaptive Immune Responses The two arms of the immune responses are; 1) the cell mediated, and 2) the humoral responses. In this chapter we will discuss the two responses in detail and we will start
The Adaptive Immune Responses The two arms of the immune responses are; 1) the cell mediated, and 2) the humoral responses. In this chapter we will discuss the two responses in detail and we will start
This is a free sample of content from Immune Tolerance. Click here for more information or to buy the book.
 A ACPAs. See Antibodies to citrullinated peptide antigens Activation-induced cell death (AICD), 25 AICD. See Activation-induced cell death AIRE, 3, 18 19, 24, 88, 103 104 AKT, 35 Alefacept, 136 ALPS. See
A ACPAs. See Antibodies to citrullinated peptide antigens Activation-induced cell death (AICD), 25 AICD. See Activation-induced cell death AIRE, 3, 18 19, 24, 88, 103 104 AKT, 35 Alefacept, 136 ALPS. See
LESSON 2: THE ADAPTIVE IMMUNITY
 Introduction to immunology. LESSON 2: THE ADAPTIVE IMMUNITY Today we will get to know: The adaptive immunity T- and B-cells Antigens and their recognition How T-cells work 1 The adaptive immunity Unlike
Introduction to immunology. LESSON 2: THE ADAPTIVE IMMUNITY Today we will get to know: The adaptive immunity T- and B-cells Antigens and their recognition How T-cells work 1 The adaptive immunity Unlike
ACTIVATION OF T LYMPHOCYTES AND CELL MEDIATED IMMUNITY
 ACTIVATION OF T LYMPHOCYTES AND CELL MEDIATED IMMUNITY The recognition of specific antigen by naïve T cell induces its own activation and effector phases. T helper cells recognize peptide antigens through
ACTIVATION OF T LYMPHOCYTES AND CELL MEDIATED IMMUNITY The recognition of specific antigen by naïve T cell induces its own activation and effector phases. T helper cells recognize peptide antigens through
T-cell activation T cells migrate to secondary lymphoid tissues where they interact with antigen, antigen-presenting cells, and other lymphocytes:
 Interactions between innate immunity & adaptive immunity What happens to T cells after they leave the thymus? Naïve T cells exit the thymus and enter the bloodstream. If they remain in the bloodstream,
Interactions between innate immunity & adaptive immunity What happens to T cells after they leave the thymus? Naïve T cells exit the thymus and enter the bloodstream. If they remain in the bloodstream,
T-cell activation T cells migrate to secondary lymphoid tissues where they interact with antigen, antigen-presenting cells, and other lymphocytes:
 Interactions between innate immunity & adaptive immunity What happens to T cells after they leave the thymus? Naïve T cells exit the thymus and enter the bloodstream. If they remain in the bloodstream,
Interactions between innate immunity & adaptive immunity What happens to T cells after they leave the thymus? Naïve T cells exit the thymus and enter the bloodstream. If they remain in the bloodstream,
FOCiS. Lecture outline. The immunological equilibrium: balancing lymphocyte activation and control. Immunological tolerance and immune regulation -- 1
 1 Immunological tolerance and immune regulation -- 1 Abul K. Abbas UCSF FOCiS 2 Lecture outline Principles of immune regulation Self-tolerance; mechanisms of central and peripheral tolerance Inhibitory
1 Immunological tolerance and immune regulation -- 1 Abul K. Abbas UCSF FOCiS 2 Lecture outline Principles of immune regulation Self-tolerance; mechanisms of central and peripheral tolerance Inhibitory
Topic (Final-02): Immunologic Tolerance
: Immunologic Tolerance") Topic (Final-02): Immunologic Tolerance Immunologic tolerance is defined as unresponsiveness to an antigen that is induced by previous exposure to that antigen. The term arose from the experimental observation
Topic (Final-02): Immunologic Tolerance Immunologic tolerance is defined as unresponsiveness to an antigen that is induced by previous exposure to that antigen. The term arose from the experimental observation
Determinants of Immunogenicity and Tolerance. Abul K. Abbas, MD Department of Pathology University of California San Francisco
 Determinants of Immunogenicity and Tolerance Abul K. Abbas, MD Department of Pathology University of California San Francisco EIP Symposium Feb 2016 Why do some people respond to therapeutic proteins?
Determinants of Immunogenicity and Tolerance Abul K. Abbas, MD Department of Pathology University of California San Francisco EIP Symposium Feb 2016 Why do some people respond to therapeutic proteins?
Immunology for the Rheumatologist
 Immunology for the Rheumatologist Rheumatologists frequently deal with the immune system gone awry, rarely studying normal immunology. This program is an overview and discussion of the function of the
Immunology for the Rheumatologist Rheumatologists frequently deal with the immune system gone awry, rarely studying normal immunology. This program is an overview and discussion of the function of the
T Lymphocyte Activation and Costimulation. FOCiS. Lecture outline
 1 T Lymphocyte Activation and Costimulation Abul K. Abbas, MD UCSF FOCiS 2 Lecture outline T cell activation Costimulation, the B7:CD28 family Inhibitory receptors of T cells Targeting costimulators for
1 T Lymphocyte Activation and Costimulation Abul K. Abbas, MD UCSF FOCiS 2 Lecture outline T cell activation Costimulation, the B7:CD28 family Inhibitory receptors of T cells Targeting costimulators for
Immunology Lecture 4. Clinical Relevance of the Immune System
 Immunology Lecture 4 The Well Patient: How innate and adaptive immune responses maintain health - 13, pg 169-181, 191-195. Immune Deficiency - 15 Autoimmunity - 16 Transplantation - 17, pg 260-270 Tumor
Immunology Lecture 4 The Well Patient: How innate and adaptive immune responses maintain health - 13, pg 169-181, 191-195. Immune Deficiency - 15 Autoimmunity - 16 Transplantation - 17, pg 260-270 Tumor
Antigen Presentation and T Lymphocyte Activation. Abul K. Abbas UCSF. FOCiS
 1 Antigen Presentation and T Lymphocyte Activation Abul K. Abbas UCSF FOCiS 2 Lecture outline Dendritic cells and antigen presentation The role of the MHC T cell activation Costimulation, the B7:CD28 family
1 Antigen Presentation and T Lymphocyte Activation Abul K. Abbas UCSF FOCiS 2 Lecture outline Dendritic cells and antigen presentation The role of the MHC T cell activation Costimulation, the B7:CD28 family
Primer on Tumor Immunology. International Society for Biological Therapy of Cancer. C. H. June, M.D. November 10, 2005
 Primer on Tumor Immunology International Society for Biological Therapy of Cancer C. H. June, M.D. November 10, 2005 Outline: Primer on Tumor Immunology T Cell Receptors T Cell Biology Tumor immunology
Primer on Tumor Immunology International Society for Biological Therapy of Cancer C. H. June, M.D. November 10, 2005 Outline: Primer on Tumor Immunology T Cell Receptors T Cell Biology Tumor immunology
T Cell Effector Mechanisms I: B cell Help & DTH
 T Cell Effector Mechanisms I: B cell Help & DTH Ned Braunstein, MD The Major T Cell Subsets p56 lck + T cells γ δ ε ζ ζ p56 lck CD8+ T cells γ δ ε ζ ζ Cα Cβ Vα Vβ CD3 CD8 Cα Cβ Vα Vβ CD3 MHC II peptide
T Cell Effector Mechanisms I: B cell Help & DTH Ned Braunstein, MD The Major T Cell Subsets p56 lck + T cells γ δ ε ζ ζ p56 lck CD8+ T cells γ δ ε ζ ζ Cα Cβ Vα Vβ CD3 CD8 Cα Cβ Vα Vβ CD3 MHC II peptide
T cell and Cell-mediated immunity
 T cell and Cell-mediated immunity Lu Linrong ( 鲁林荣 ) PhD Laboratory of Immune Regulation Institute of Immunology Zhejiang University, it School of Medicine i Medical Research Building B815-819 Email: Lu.Linrong@gmail.com
T cell and Cell-mediated immunity Lu Linrong ( 鲁林荣 ) PhD Laboratory of Immune Regulation Institute of Immunology Zhejiang University, it School of Medicine i Medical Research Building B815-819 Email: Lu.Linrong@gmail.com
Examples of questions for Cellular Immunology/Cellular Biology and Immunology
 Examples of questions for Cellular Immunology/Cellular Biology and Immunology Each student gets a set of 6 questions, so that each set contains different types of questions and that the set of questions
Examples of questions for Cellular Immunology/Cellular Biology and Immunology Each student gets a set of 6 questions, so that each set contains different types of questions and that the set of questions
T Cell Activation, Costimulation and Regulation
 1 T Cell Activation, Costimulation and Regulation Abul K. Abbas, MD University of California San Francisco 2 Lecture outline T cell antigen recognition and activation Costimulation, the B7:CD28 family
1 T Cell Activation, Costimulation and Regulation Abul K. Abbas, MD University of California San Francisco 2 Lecture outline T cell antigen recognition and activation Costimulation, the B7:CD28 family
Tolerance 2. Regulatory T cells; why tolerance fails. Abul K. Abbas UCSF. FOCiS
 1 Tolerance 2. Regulatory T cells; why tolerance fails Abul K. Abbas UCSF FOCiS 2 Lecture outline Regulatory T cells: functions and clinical relevance Pathogenesis of autoimmunity: why selftolerance fails
1 Tolerance 2. Regulatory T cells; why tolerance fails Abul K. Abbas UCSF FOCiS 2 Lecture outline Regulatory T cells: functions and clinical relevance Pathogenesis of autoimmunity: why selftolerance fails
Follicular Lymphoma. ced3 APOPTOSIS. *In the nematode Caenorhabditis elegans 131 of the organism's 1031 cells die during development.
 Harvard-MIT Division of Health Sciences and Technology HST.176: Cellular and Molecular Immunology Course Director: Dr. Shiv Pillai Follicular Lymphoma 1. Characterized by t(14:18) translocation 2. Ig heavy
Harvard-MIT Division of Health Sciences and Technology HST.176: Cellular and Molecular Immunology Course Director: Dr. Shiv Pillai Follicular Lymphoma 1. Characterized by t(14:18) translocation 2. Ig heavy
Immune Tolerance. Kyeong Cheon Jung. Department of Pathology Seoul National University College of Medicine
 Immune Tolerance Kyeong Cheon Jung Department of Pathology Seoul National University College of Medicine Immune tolerance Unresponsiveness to an antigen that is induced by previous exposure to that antigen
Immune Tolerance Kyeong Cheon Jung Department of Pathology Seoul National University College of Medicine Immune tolerance Unresponsiveness to an antigen that is induced by previous exposure to that antigen
HLA and antigen presentation. Department of Immunology Charles University, 2nd Medical School University Hospital Motol
 HLA and antigen presentation Department of Immunology Charles University, 2nd Medical School University Hospital Motol MHC in adaptive immunity Characteristics Specificity Innate For structures shared
HLA and antigen presentation Department of Immunology Charles University, 2nd Medical School University Hospital Motol MHC in adaptive immunity Characteristics Specificity Innate For structures shared
CHAPTER 9 BIOLOGY OF THE T LYMPHOCYTE
 CHAPTER 9 BIOLOGY OF THE T LYMPHOCYTE Coico, R., Sunshine, G., (2009) Immunology : a short course, 6 th Ed., Wiley-Blackwell 1 CHAPTER 9 : Biology of The T Lymphocytes 1. 2. 3. 4. 5. 6. 7. Introduction
CHAPTER 9 BIOLOGY OF THE T LYMPHOCYTE Coico, R., Sunshine, G., (2009) Immunology : a short course, 6 th Ed., Wiley-Blackwell 1 CHAPTER 9 : Biology of The T Lymphocytes 1. 2. 3. 4. 5. 6. 7. Introduction
Introduction. Introduction. Lymphocyte development (maturation)
") Introduction Abbas Chapter 8: Lymphocyte Development and the Rearrangement and Expression of Antigen Receptor Genes Christina Ciaccio, MD Children s Mercy Hospital January 5, 2009 Lymphocyte development
Introduction Abbas Chapter 8: Lymphocyte Development and the Rearrangement and Expression of Antigen Receptor Genes Christina Ciaccio, MD Children s Mercy Hospital January 5, 2009 Lymphocyte development
T cell and Cell-mediated immunity
 T cell and Cell-mediated immunity ( 第十章 第十二章第十二章 ) Lu Linrong ( 鲁林荣 ) PhD Laboratory of Immune Regulation Institute of Immunology Zhejiang University, School of Medicine Medical Research Building B815-819
T cell and Cell-mediated immunity ( 第十章 第十二章第十二章 ) Lu Linrong ( 鲁林荣 ) PhD Laboratory of Immune Regulation Institute of Immunology Zhejiang University, School of Medicine Medical Research Building B815-819
Structure and Function of Antigen Recognition Molecules
 MICR2209 Structure and Function of Antigen Recognition Molecules Dr Allison Imrie allison.imrie@uwa.edu.au 1 Synopsis: In this lecture we will examine the major receptors used by cells of the innate and
MICR2209 Structure and Function of Antigen Recognition Molecules Dr Allison Imrie allison.imrie@uwa.edu.au 1 Synopsis: In this lecture we will examine the major receptors used by cells of the innate and
Scott Abrams, Ph.D. Professor of Oncology, x4375 Kuby Immunology SEVENTH EDITION
 Scott Abrams, Ph.D. Professor of Oncology, x4375 scott.abrams@roswellpark.org Kuby Immunology SEVENTH EDITION CHAPTER 11 T-Cell Activation, Differentiation, and Memory Copyright 2013 by W. H. Freeman and
Scott Abrams, Ph.D. Professor of Oncology, x4375 scott.abrams@roswellpark.org Kuby Immunology SEVENTH EDITION CHAPTER 11 T-Cell Activation, Differentiation, and Memory Copyright 2013 by W. H. Freeman and
1. Overview of Adaptive Immunity
 Chapter 17A: Adaptive Immunity Part I 1. Overview of Adaptive Immunity 2. T and B Cell Production 3. Antigens & Antigen Presentation 4. Helper T cells 1. Overview of Adaptive Immunity The Nature of Adaptive
Chapter 17A: Adaptive Immunity Part I 1. Overview of Adaptive Immunity 2. T and B Cell Production 3. Antigens & Antigen Presentation 4. Helper T cells 1. Overview of Adaptive Immunity The Nature of Adaptive
ACTIVATION AND EFFECTOR FUNCTIONS OF CELL-MEDIATED IMMUNITY AND NK CELLS. Choompone Sakonwasun, MD (Hons), FRCPT
, FRCPT") ACTIVATION AND EFFECTOR FUNCTIONS OF CELL-MEDIATED IMMUNITY AND NK CELLS Choompone Sakonwasun, MD (Hons), FRCPT Types of Adaptive Immunity Types of T Cell-mediated Immune Reactions CTLs = cytotoxic T lymphocytes
ACTIVATION AND EFFECTOR FUNCTIONS OF CELL-MEDIATED IMMUNITY AND NK CELLS Choompone Sakonwasun, MD (Hons), FRCPT Types of Adaptive Immunity Types of T Cell-mediated Immune Reactions CTLs = cytotoxic T lymphocytes
Principles of Adaptive Immunity
 Principles of Adaptive Immunity Chapter 3 Parham Hans de Haard 17 th of May 2010 Agenda Recognition molecules of adaptive immune system Features adaptive immune system Immunoglobulins and T-cell receptors
Principles of Adaptive Immunity Chapter 3 Parham Hans de Haard 17 th of May 2010 Agenda Recognition molecules of adaptive immune system Features adaptive immune system Immunoglobulins and T-cell receptors
Autoimmunity. Autoimmunity arises because of defects in central or peripheral tolerance of lymphocytes to selfantigens
 Autoimmunity Autoimmunity arises because of defects in central or peripheral tolerance of lymphocytes to selfantigens Autoimmune disease can be caused to primary defects in B cells, T cells and possibly
Autoimmunity Autoimmunity arises because of defects in central or peripheral tolerance of lymphocytes to selfantigens Autoimmune disease can be caused to primary defects in B cells, T cells and possibly
TCR, MHC and coreceptors
 Cooperation In Immune Responses Antigen processing how peptides get into MHC Antigen processing involves the intracellular proteolytic generation of MHC binding proteins Protein antigens may be processed
Cooperation In Immune Responses Antigen processing how peptides get into MHC Antigen processing involves the intracellular proteolytic generation of MHC binding proteins Protein antigens may be processed
Cellular Pathology of immunological disorders
 Cellular Pathology of immunological disorders SCBM344 Cellular and Molecular Pathology Witchuda Payuhakrit, Ph.D (Pathobiology) witchuda.pay@mahidol.ac.th Objectives Describe the etiology of immunological
Cellular Pathology of immunological disorders SCBM344 Cellular and Molecular Pathology Witchuda Payuhakrit, Ph.D (Pathobiology) witchuda.pay@mahidol.ac.th Objectives Describe the etiology of immunological
Development of B and T lymphocytes
 Development of B and T lymphocytes What will we discuss today? B-cell development T-cell development B- cell development overview Stem cell In periphery Pro-B cell Pre-B cell Immature B cell Mature B cell
Development of B and T lymphocytes What will we discuss today? B-cell development T-cell development B- cell development overview Stem cell In periphery Pro-B cell Pre-B cell Immature B cell Mature B cell
General Overview of Immunology. Kimberly S. Schluns, Ph.D. Associate Professor Department of Immunology UT MD Anderson Cancer Center
 General Overview of Immunology Kimberly S. Schluns, Ph.D. Associate Professor Department of Immunology UT MD Anderson Cancer Center Objectives Describe differences between innate and adaptive immune responses
General Overview of Immunology Kimberly S. Schluns, Ph.D. Associate Professor Department of Immunology UT MD Anderson Cancer Center Objectives Describe differences between innate and adaptive immune responses
CELL BIOLOGY - CLUTCH CH THE IMMUNE SYSTEM.
 !! www.clutchprep.com CONCEPT: OVERVIEW OF HOST DEFENSES The human body contains three lines of against infectious agents (pathogens) 1. Mechanical and chemical boundaries (part of the innate immune system)
!! www.clutchprep.com CONCEPT: OVERVIEW OF HOST DEFENSES The human body contains three lines of against infectious agents (pathogens) 1. Mechanical and chemical boundaries (part of the innate immune system)
Micro 204. Cytotoxic T Lymphocytes (CTL) Lewis Lanier
 Lewis Lanier") Micro 204 Cytotoxic T Lymphocytes (CTL) Lewis Lanier Lewis.Lanier@ucsf.edu Lymphocyte-mediated Cytotoxicity CD8 + αβ-tcr + T cells CD4 + αβ-tcr + T cells γδ-tcr + T cells Natural Killer cells CD8 + αβ-tcr
Micro 204 Cytotoxic T Lymphocytes (CTL) Lewis Lanier Lewis.Lanier@ucsf.edu Lymphocyte-mediated Cytotoxicity CD8 + αβ-tcr + T cells CD4 + αβ-tcr + T cells γδ-tcr + T cells Natural Killer cells CD8 + αβ-tcr
T Cell Activation. Patricia Fitzgerald-Bocarsly March 18, 2009
 T Cell Activation Patricia Fitzgerald-Bocarsly March 18, 2009 Phases of Adaptive Immune Responses Phases of T cell responses IL-2 acts as an autocrine growth factor Fig. 11-11 Clonal Expansion of T cells
T Cell Activation Patricia Fitzgerald-Bocarsly March 18, 2009 Phases of Adaptive Immune Responses Phases of T cell responses IL-2 acts as an autocrine growth factor Fig. 11-11 Clonal Expansion of T cells
Guarding the immune system: Suppression of autoimmunity by CD4 1 CD25 1 immunoregulatory T cells
 Immunology and Cell Biology (2006) 84, 487 501 doi:10.1111/j.1440-1711.2006.01471.x Review Article Guarding the immune system: Suppression of autoimmunity by CD4 1 CD25 1 immunoregulatory T cells TRICIA
Immunology and Cell Biology (2006) 84, 487 501 doi:10.1111/j.1440-1711.2006.01471.x Review Article Guarding the immune system: Suppression of autoimmunity by CD4 1 CD25 1 immunoregulatory T cells TRICIA
HLA and antigen presentation. Department of Immunology Charles University, 2nd Medical School University Hospital Motol
 HLA and antigen presentation Department of Immunology Charles University, 2nd Medical School University Hospital Motol MHC in adaptive immunity Characteristics Specificity Innate For structures shared
HLA and antigen presentation Department of Immunology Charles University, 2nd Medical School University Hospital Motol MHC in adaptive immunity Characteristics Specificity Innate For structures shared
Allergy and Immunology Review Corner: Chapter 19 of Immunology IV: Clinical Applications in Health and Disease, by Joseph A. Bellanti, MD.
 Allergy and Immunology Review Corner: Chapter 19 of Immunology IV: Clinical Applications in Health and Disease, by Joseph A. Bellanti, MD. Chapter 19: Tolerance, Autoimmunity, and Autoinflammation Prepared
Allergy and Immunology Review Corner: Chapter 19 of Immunology IV: Clinical Applications in Health and Disease, by Joseph A. Bellanti, MD. Chapter 19: Tolerance, Autoimmunity, and Autoinflammation Prepared
COURSE: Medical Microbiology, MBIM 650/720 - Fall TOPIC: Antigen Processing, MHC Restriction, & Role of Thymus Lecture 12
 COURSE: Medical Microbiology, MBIM 650/720 - Fall 2008 TOPIC: Antigen Processing, MHC Restriction, & Role of Thymus Lecture 12 FACULTY: Dr. Mayer Office: Bldg. #1, Rm B32 Phone: 733-3281 Email: MAYER@MED.SC.EDU
COURSE: Medical Microbiology, MBIM 650/720 - Fall 2008 TOPIC: Antigen Processing, MHC Restriction, & Role of Thymus Lecture 12 FACULTY: Dr. Mayer Office: Bldg. #1, Rm B32 Phone: 733-3281 Email: MAYER@MED.SC.EDU
Scott Abrams, Ph.D. Professor of Oncology, x4375 Kuby Immunology SEVENTH EDITION
 Scott Abrams, Ph.D. Professor of Oncology, x4375 scott.abrams@roswellpark.org Kuby Immunology SEVENTH EDITION CHAPTER 13 Effector Responses: Cell- and Antibody-Mediated Immunity Copyright 2013 by W. H.
Scott Abrams, Ph.D. Professor of Oncology, x4375 scott.abrams@roswellpark.org Kuby Immunology SEVENTH EDITION CHAPTER 13 Effector Responses: Cell- and Antibody-Mediated Immunity Copyright 2013 by W. H.
T cell-mediated immunity
 T cell-mediated immunity Overview For microbes within phagosomes in phagocytes.cd4+ T lymphocytes (TH1) Activate phagocyte by cytokines studies on Listeria monocytogenes For microbes infecting and replicating
T cell-mediated immunity Overview For microbes within phagosomes in phagocytes.cd4+ T lymphocytes (TH1) Activate phagocyte by cytokines studies on Listeria monocytogenes For microbes infecting and replicating
Immunology MIMM-314 MID-TERM II EXAMINATION. 1 hour between 8:30 a.m. - 10:00 a.m. McIntyre Medical Rm 504 (Martin Amphitheatre)
") GROUP (Version) 1 Annotated version April 8, 2011, RGEP DEPARTMENT OF MICROBIOLOGY AND IMMUNOLOGY Immunology MIMM-314 MID-TERM II EXAMINATION Course Coordinator: Dr. Roger Palfree Date: Thursday, March
GROUP (Version) 1 Annotated version April 8, 2011, RGEP DEPARTMENT OF MICROBIOLOGY AND IMMUNOLOGY Immunology MIMM-314 MID-TERM II EXAMINATION Course Coordinator: Dr. Roger Palfree Date: Thursday, March
The Adaptive Immune Response: T lymphocytes and Their Functional Types *
 OpenStax-CNX module: m46560 1 The Adaptive Immune Response: T lymphocytes and Their Functional Types * OpenStax This work is produced by OpenStax-CNX and licensed under the Creative Commons Attribution
OpenStax-CNX module: m46560 1 The Adaptive Immune Response: T lymphocytes and Their Functional Types * OpenStax This work is produced by OpenStax-CNX and licensed under the Creative Commons Attribution
The T cell receptor for MHC-associated peptide antigens
 1 The T cell receptor for MHC-associated peptide antigens T lymphocytes have a dual specificity: they recognize polymporphic residues of self MHC molecules, and they also recognize residues of peptide
1 The T cell receptor for MHC-associated peptide antigens T lymphocytes have a dual specificity: they recognize polymporphic residues of self MHC molecules, and they also recognize residues of peptide
The Adaptive Immune Response. T-cells
 The Adaptive Immune Response T-cells T Lymphocytes T lymphocytes develop from precursors in the thymus. Mature T cells are found in the blood, where they constitute 60% to 70% of lymphocytes, and in T-cell
The Adaptive Immune Response T-cells T Lymphocytes T lymphocytes develop from precursors in the thymus. Mature T cells are found in the blood, where they constitute 60% to 70% of lymphocytes, and in T-cell
Immune response. This overview figure summarizes simply how our body responds to foreign molecules that enter to it.
 Immune response This overview figure summarizes simply how our body responds to foreign molecules that enter to it. It s highly recommended to watch Dr Najeeb s lecture that s titled T Helper cells and
Immune response This overview figure summarizes simply how our body responds to foreign molecules that enter to it. It s highly recommended to watch Dr Najeeb s lecture that s titled T Helper cells and
Darwinian selection and Newtonian physics wrapped up in systems biology
 Darwinian selection and Newtonian physics wrapped up in systems biology Concept published in 1957* by Macfarland Burnet (1960 Nobel Laureate for the theory of induced immune tolerance, leading to solid
Darwinian selection and Newtonian physics wrapped up in systems biology Concept published in 1957* by Macfarland Burnet (1960 Nobel Laureate for the theory of induced immune tolerance, leading to solid
Cytokines modulate the functional activities of individual cells and tissues both under normal and pathologic conditions Interleukins,
 Cytokines http://highered.mcgraw-hill.com/sites/0072507470/student_view0/chapter22/animation the_immune_response.html Cytokines modulate the functional activities of individual cells and tissues both under
Cytokines http://highered.mcgraw-hill.com/sites/0072507470/student_view0/chapter22/animation the_immune_response.html Cytokines modulate the functional activities of individual cells and tissues both under
Attribution: University of Michigan Medical School, Department of Microbiology and Immunology
 Attribution: University of Michigan Medical School, Department of Microbiology and Immunology License: Unless otherwise noted, this material is made available under the terms of the Creative Commons Attribution
Attribution: University of Michigan Medical School, Department of Microbiology and Immunology License: Unless otherwise noted, this material is made available under the terms of the Creative Commons Attribution
Medical Virology Immunology. Dr. Sameer Naji, MB, BCh, PhD (UK) Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University
 Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University") Medical Virology Immunology Dr. Sameer Naji, MB, BCh, PhD (UK) Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University Human blood cells Phases of immune responses Microbe Naïve
Medical Virology Immunology Dr. Sameer Naji, MB, BCh, PhD (UK) Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University Human blood cells Phases of immune responses Microbe Naïve
Immunodermatology. Danielle Tartar, MD, PhD Assistant Clinical Professor Co-Director of Inpatient Dermatology University of California - Davis
 Immunodermatology Danielle Tartar, MD, PhD Assistant Clinical Professor Co-Director of Inpatient Dermatology University of California - Davis Outline Adaptive Immunity: T and B cell development, activation,
Immunodermatology Danielle Tartar, MD, PhD Assistant Clinical Professor Co-Director of Inpatient Dermatology University of California - Davis Outline Adaptive Immunity: T and B cell development, activation,
T Cell Differentiation
 T Cell Differentiation Ned Braunstein, MD MHC control of Immune Responsiveness: Concept Whether or not an individual makes an immune response to a particular antigen depends on what MHC alleles an individual
T Cell Differentiation Ned Braunstein, MD MHC control of Immune Responsiveness: Concept Whether or not an individual makes an immune response to a particular antigen depends on what MHC alleles an individual
Regulatory T Cells and Maintenance of Tolerance
 Regulatory Cells and Maintenance of olerance Stephen Canfield, MD, PhD Asst. Prof. Medicine smc12@columbia.edu What constitutes immune tolerance? Discrimination between: non-self and self e.g., virally
Regulatory Cells and Maintenance of olerance Stephen Canfield, MD, PhD Asst. Prof. Medicine smc12@columbia.edu What constitutes immune tolerance? Discrimination between: non-self and self e.g., virally
Immunology. T-Lymphocytes. 16. Oktober 2014, Ruhr-Universität Bochum Karin Peters,
 Immunology T-Lymphocytes 16. Oktober 2014, Ruhr-Universität Bochum Karin Peters, karin.peters@rub.de The role of T-effector cells in the immune response against microbes cellular immunity humoral immunity
Immunology T-Lymphocytes 16. Oktober 2014, Ruhr-Universität Bochum Karin Peters, karin.peters@rub.de The role of T-effector cells in the immune response against microbes cellular immunity humoral immunity
Cellular Immune response. Jianzhong Chen, Ph.D Institute of immunology, ZJU
 Cellular Immune response Jianzhong Chen, Ph.D Institute of immunology, ZJU Concept of adaptive immune response T cell-mediated adaptive immune response I. Concept of immune response A collective and coordinated
Cellular Immune response Jianzhong Chen, Ph.D Institute of immunology, ZJU Concept of adaptive immune response T cell-mediated adaptive immune response I. Concept of immune response A collective and coordinated
The Adaptive Immune Response. B-cells
 The Adaptive Immune Response B-cells The innate immune system provides immediate protection. The adaptive response takes time to develop and is antigen specific. Activation of B and T lymphocytes Naive
The Adaptive Immune Response B-cells The innate immune system provides immediate protection. The adaptive response takes time to develop and is antigen specific. Activation of B and T lymphocytes Naive
Immunology Basics Relevant to Cancer Immunotherapy: T Cell Activation, Costimulation, and Effector T Cells
 Immunology Basics Relevant to Cancer Immunotherapy: T Cell Activation, Costimulation, and Effector T Cells Andrew H. Lichtman, M.D. Ph.D. Department of Pathology Brigham and Women s Hospital and Harvard
Immunology Basics Relevant to Cancer Immunotherapy: T Cell Activation, Costimulation, and Effector T Cells Andrew H. Lichtman, M.D. Ph.D. Department of Pathology Brigham and Women s Hospital and Harvard
The Immune System: Innate and Adaptive Body Defenses Outline PART 1: INNATE DEFENSES 21.1 Surface barriers act as the first line of defense to keep
 The Immune System: Innate and Adaptive Body Defenses Outline PART 1: INNATE DEFENSES 21.1 Surface barriers act as the first line of defense to keep invaders out of the body (pp. 772 773; Fig. 21.1; Table
The Immune System: Innate and Adaptive Body Defenses Outline PART 1: INNATE DEFENSES 21.1 Surface barriers act as the first line of defense to keep invaders out of the body (pp. 772 773; Fig. 21.1; Table
DNA vaccine, peripheral T-cell tolerance modulation 185
 Subject Index Airway hyperresponsiveness (AHR) animal models 41 43 asthma inhibition 45 overview 41 mast cell modulation of T-cells 62 64 respiratory tolerance 40, 41 Tregs inhibition role 44 respiratory
Subject Index Airway hyperresponsiveness (AHR) animal models 41 43 asthma inhibition 45 overview 41 mast cell modulation of T-cells 62 64 respiratory tolerance 40, 41 Tregs inhibition role 44 respiratory
Tolerance 2. Regulatory T cells; why tolerance fails. FOCiS. Lecture outline. Regulatory T cells. Regulatory T cells: functions and clinical relevance
 1 Tolerance 2. Regulatory T cells; why tolerance fails Abul K. Abbas UCSF FOCiS 2 Lecture outline Regulatory T cells: functions and clinical relevance Pathogenesis of autoimmunity: why selftolerance fails
1 Tolerance 2. Regulatory T cells; why tolerance fails Abul K. Abbas UCSF FOCiS 2 Lecture outline Regulatory T cells: functions and clinical relevance Pathogenesis of autoimmunity: why selftolerance fails
The Development of Lymphocytes: B Cell Development in the Bone Marrow & Peripheral Lymphoid Tissue Deborah A. Lebman, Ph.D.
 The Development of Lymphocytes: B Cell Development in the Bone Marrow & Peripheral Lymphoid Tissue Deborah A. Lebman, Ph.D. OBJECTIVES 1. To understand how ordered Ig gene rearrangements lead to the development
The Development of Lymphocytes: B Cell Development in the Bone Marrow & Peripheral Lymphoid Tissue Deborah A. Lebman, Ph.D. OBJECTIVES 1. To understand how ordered Ig gene rearrangements lead to the development
Defensive mechanisms include :
 Acquired Immunity Defensive mechanisms include : 1) Innate immunity (Natural or Non specific) 2) Acquired immunity (Adaptive or Specific) Cell-mediated immunity Humoral immunity Two mechanisms 1) Humoral
Acquired Immunity Defensive mechanisms include : 1) Innate immunity (Natural or Non specific) 2) Acquired immunity (Adaptive or Specific) Cell-mediated immunity Humoral immunity Two mechanisms 1) Humoral
Chapter 22: The Lymphatic System and Immunity
 Bio40C schedule Lecture Immune system Lab Quiz 2 this week; bring a scantron! Study guide on my website (see lab assignments) Extra credit Critical thinking questions at end of chapters 5 pts/chapter Due
Bio40C schedule Lecture Immune system Lab Quiz 2 this week; bring a scantron! Study guide on my website (see lab assignments) Extra credit Critical thinking questions at end of chapters 5 pts/chapter Due
Question 1. Kupffer cells, microglial cells and osteoclasts are all examples of what type of immune system cell?
 Abbas Chapter 2: Sarah Spriet February 8, 2015 Question 1. Kupffer cells, microglial cells and osteoclasts are all examples of what type of immune system cell? a. Dendritic cells b. Macrophages c. Monocytes
Abbas Chapter 2: Sarah Spriet February 8, 2015 Question 1. Kupffer cells, microglial cells and osteoclasts are all examples of what type of immune system cell? a. Dendritic cells b. Macrophages c. Monocytes
Adaptive Immune System
 Short Course on Immunology Adaptive Immune System Bhargavi Duvvuri Ph.D IIIrd Year (Immunology) bhargavi@yorku.ca Supervisor Dr.Gillian E Wu Professor, School of Kinesiology and Health Sciences York University,
Short Course on Immunology Adaptive Immune System Bhargavi Duvvuri Ph.D IIIrd Year (Immunology) bhargavi@yorku.ca Supervisor Dr.Gillian E Wu Professor, School of Kinesiology and Health Sciences York University,
Chapter 10 (pages ): Differentiation and Functions of CD4+ Effector T Cells Prepared by Kristen Dazy, MD, Scripps Clinic Medical Group
: Differentiation and Functions of CD4+ Effector T Cells Prepared by Kristen Dazy, MD, Scripps Clinic Medical Group") FIT Board Review Corner September 2015 Welcome to the FIT Board Review Corner, prepared by Andrew Nickels, MD, and Sarah Spriet, DO, senior and junior representatives of ACAAI's Fellows-In-Training (FITs)
FIT Board Review Corner September 2015 Welcome to the FIT Board Review Corner, prepared by Andrew Nickels, MD, and Sarah Spriet, DO, senior and junior representatives of ACAAI's Fellows-In-Training (FITs)
SINGLE CHOICE. 5. The gamma invariant chain binds to this molecule during its intracytoplasmic transport. A TCR B BCR C MHC II D MHC I E FcγR
 A Name: Group: SINGLE CHOICE 1. Which is the most important ligand of TLR5? A endospore B flagellin C polysaccharide capsule D DNA E pilus 2. The antibody-binding site is formed primarily by... A the constant
A Name: Group: SINGLE CHOICE 1. Which is the most important ligand of TLR5? A endospore B flagellin C polysaccharide capsule D DNA E pilus 2. The antibody-binding site is formed primarily by... A the constant
Recommended reading: Abbas et al. 5th edition, chapters 7 and 10; Janeway and Travers, 5th edition, chapter 7.
 Harvard-MIT Division of Health Sciences and Technology HST.176: Cellular and Molecular Immunology Course Director: Dr. Shiv Pillai 10/05/05; 11 AM Shiv Pillai T Lymphocyte Development Recommended reading:
Harvard-MIT Division of Health Sciences and Technology HST.176: Cellular and Molecular Immunology Course Director: Dr. Shiv Pillai 10/05/05; 11 AM Shiv Pillai T Lymphocyte Development Recommended reading:
Adaptive Immunity. Jeffrey K. Actor, Ph.D. MSB 2.214,
 Adaptive Immunity Jeffrey K. Actor, Ph.D. MSB 2.214, 500-5344 Lecture Objectives: Understand role of various molecules including cytokines, chemokines, costimulatory and adhesion molecules in the development
Adaptive Immunity Jeffrey K. Actor, Ph.D. MSB 2.214, 500-5344 Lecture Objectives: Understand role of various molecules including cytokines, chemokines, costimulatory and adhesion molecules in the development
Properties & Overview of IRs Dr. Nasser M. Kaplan JUST, Jordan. 10-Jul-16 NM Kaplan 1
 Properties & Overview of IRs Dr. Nasser M. Kaplan JUST, Jordan 10-Jul-16 NM Kaplan 1 Major components of IS & their properties Definitions IS = cells & molecules responsible for: 1- Physiologic; protective
Properties & Overview of IRs Dr. Nasser M. Kaplan JUST, Jordan 10-Jul-16 NM Kaplan 1 Major components of IS & their properties Definitions IS = cells & molecules responsible for: 1- Physiologic; protective
Immune responses in autoimmune diseases
 Immune responses in autoimmune diseases Erika Jensen-Jarolim Dept. of Pathophysiology Medical University Vienna CCHD Lecture January 24, 2007 Primary immune organs: Bone marrow Thymus Secondary: Lymph
Immune responses in autoimmune diseases Erika Jensen-Jarolim Dept. of Pathophysiology Medical University Vienna CCHD Lecture January 24, 2007 Primary immune organs: Bone marrow Thymus Secondary: Lymph
Introduction to Immunology Part 2 September 30, Dan Stetson
 Introduction to Immunology Part 2 September 30, 2016 Dan Stetson stetson@uw.edu 441 Lecture #2 Slide 1 of 26 CLASS ANNOUNCEMENT PLEASE NO TREE NUTS IN CLASS!!! (Peanuts, walnuts, almonds, cashews, etc)
Introduction to Immunology Part 2 September 30, 2016 Dan Stetson stetson@uw.edu 441 Lecture #2 Slide 1 of 26 CLASS ANNOUNCEMENT PLEASE NO TREE NUTS IN CLASS!!! (Peanuts, walnuts, almonds, cashews, etc)
Animal Models to Understand Immunity
 Animal Models to Understand Immunity Hussein El Saghire hesaghir@sckcen.be Innate Adaptive immunity Immunity MAPK and NF-kB TLR pathways receptors Fast Slow Non-specific Specific NOD-like receptors T-cell
Animal Models to Understand Immunity Hussein El Saghire hesaghir@sckcen.be Innate Adaptive immunity Immunity MAPK and NF-kB TLR pathways receptors Fast Slow Non-specific Specific NOD-like receptors T-cell
Test Bank for Basic Immunology Functions and Disorders of the Immune System 4th Edition by Abbas
 Test Bank for Basic Immunology Functions and Disorders of the Immune System 4th Edition by Abbas Chapter 04: Antigen Recognition in the Adaptive Immune System Test Bank MULTIPLE CHOICE 1. Most T lymphocytes
Test Bank for Basic Immunology Functions and Disorders of the Immune System 4th Edition by Abbas Chapter 04: Antigen Recognition in the Adaptive Immune System Test Bank MULTIPLE CHOICE 1. Most T lymphocytes
Allergy and Immunology Review Corner: Chapter 13 of Immunology IV: Clinical Applications in Health and Disease, by Joseph A. Bellanti, MD.
 Allergy and Immunology Review Corner: Chapter 13 of Immunology IV: Clinical Applications in Health and Disease, by Joseph A. Bellanti, MD. Chapter 13: Mechanisms of Immunity to Viral Disease Prepared by
Allergy and Immunology Review Corner: Chapter 13 of Immunology IV: Clinical Applications in Health and Disease, by Joseph A. Bellanti, MD. Chapter 13: Mechanisms of Immunity to Viral Disease Prepared by
Allergy and Immunology Review Corner: Chapter 1 of Immunology IV: Clinical Applications in Health and Disease, by Joseph A. Bellanti.
 Allergy and Immunology Review Corner: Chapter 1 of Immunology IV: Clinical Applications in Health and Disease, by Joseph A. Bellanti. Chapter 1: Overview of Immunology Prepared by David Scott, MD, Scripps
Allergy and Immunology Review Corner: Chapter 1 of Immunology IV: Clinical Applications in Health and Disease, by Joseph A. Bellanti. Chapter 1: Overview of Immunology Prepared by David Scott, MD, Scripps
T cell Receptor. Chapter 9. Comparison of TCR αβ T cells
 Chapter 9 The αβ TCR is similar in size and structure to an antibody Fab fragment T cell Receptor Kuby Figure 9-3 The αβ T cell receptor - Two chains - α and β - Two domains per chain - constant (C) domain
Chapter 9 The αβ TCR is similar in size and structure to an antibody Fab fragment T cell Receptor Kuby Figure 9-3 The αβ T cell receptor - Two chains - α and β - Two domains per chain - constant (C) domain
Immunological Aspects of Parasitic Diseases in Immunocompromised Individuals. Taniawati Supali. Department of Parasitology
 Immunological Aspects of Parasitic Diseases in Immunocompromised Individuals Taniawati Supali Department of Parasitology 1 Defense mechanism in human Th17 (? ) Acute Chronic Th1 Th 2 Intracellular Treg
Immunological Aspects of Parasitic Diseases in Immunocompromised Individuals Taniawati Supali Department of Parasitology 1 Defense mechanism in human Th17 (? ) Acute Chronic Th1 Th 2 Intracellular Treg
Chapter 13: Cytokines
 Chapter 13: Cytokines Definition: secreted, low-molecular-weight proteins that regulate the nature, intensity and duration of the immune response by exerting a variety of effects on lymphocytes and/or
Chapter 13: Cytokines Definition: secreted, low-molecular-weight proteins that regulate the nature, intensity and duration of the immune response by exerting a variety of effects on lymphocytes and/or