Functional consequences of RNA 5 terminal deletions on coxsackievirus B3 RNA replication and ribonucleoprotein complex formation
|
|
|
- Kenneth Payne
- 6 years ago
- Views:
Transcription
1 JVI Accepted Manuscript Posted Online 24 May 2017 J. Virol. doi: /jvi Copyright 2017 American Society for Microbiology. All Rights Reserved. Functional consequences of RNA 5 terminal deletions on coxsackievirus B3 RNA replication and ribonucleoprotein complex formation Nicolas Lévêque, a,b * Magali Garcia, c Alexis Bouin, b Joseph H. C. Nguyen, b Genevieve P. Tran, b Laurent Andreoletti, a and Bert L. Semler b EA-4684 CardioVir, School of Medicine, University of Reims Champagne- Ardenne, Reims, France a ; Center for Virus Research and Department of Microbiology and Molecular Genetics, School of Medicine, University of California, Irvine, CA, USA b ; EA-4331 LITEC, Faculty of Medicine and Pharmacy, University of Poitiers, Poitiers, France c Running title: Replication of 5 terminally-deleted CVB3 RNAs Address correspondence to: Bert L. Semler, blsemler@uci.edu or Laurent Andreoletti, landreoletti@chu-reims.fr *Present address: Nicolas Lévêque, Virology and Mycobacteriology Laboratory and EA-4331 LITEC, Faculty of Medicine and Pharmacy, University Hospital of Poitiers, 2 rue de la Miletrie, Poitiers Cedex, France
2 ABSTRACT Group B coxsackieviruses are responsible for chronic cardiac infections. However, the molecular mechanisms by which the virus can persist in the human heart long after the signs of acute myocarditis have abated are still not completely understood. Recently, coxsackievirus B3 strains with 5 terminal deletions in genomic RNAs were isolated from a patient suffering from idiopathic dilated cardiomyopathy, suggesting that such mutant viruses may be the forms responsible for persistent infection. These deletions lacked portions of 5 stem-loop I, which is an RNA secondary structure required for viral RNA replication. In this study, we assessed the consequences of the genomic deletions observed in vivo on coxsackievirus B3 biology. Using cell-free extracts from HeLa cells as well as transfection of luciferase replicons in two types of cardiomyocytes, we demonstrated that coxsackievirus RNAs harboring 5 deletions ranging from 7 to 49 nucleotides can be translated nearly as efficiently as those of wild-type virus. However, these 5 deletions greatly reduced the synthesis of viral RNA in vitro, which was only detected for the 7 and 21 nucleotide deletions. Since 5 stem-loop I RNA forms a ribonucleoprotein complex with cellular and viral proteins involved in viral RNA replication, we investigated the binding of host cell protein PCBP2 as well as viral protein 3CD pro to deleted positive-strand RNAs corresponding to the 5 end. We found that binding of these proteins was conserved but that ribonucleoprotein complex formation required higher PCBP2 and 3CD pro concentrations, depending on the size of the deletion. Overall, this study confirmed the characteristics of 2
3 persistent CVB3 infection observed in heart tissues and provided a possible explanation for the low level of RNA replication observed for the 5 deleted viral genomes - a less stable ribonucleoprotein complex formed with proteins involved in viral RNA replication. 3
4 IMPORTANCE Dilated cardiomyopathy is the most common indication for heart transplantation worldwide, and coxsackie B viruses are detected in about one third of idiopathic, dilated cardiomyopathies. Terminal deletions at the 5 end of the viral genome involving an RNA secondary structure required for RNA replication have been recently reported as a possible mechanism of virus persistence in the human heart. These mutations are likely to disrupt the correct folding of an RNA secondary structure required for viral RNA replication. In this report, we demonstrate that transfected RNAs harboring 5 terminal sequence deletions are able to direct the synthesis of viral proteins but not genomic RNAs in human and murine cardiomyocytes. Moreover, we show that the binding of cellular and viral replication factors to viral RNA is conserved despite genomic deletions, but that the impaired RNA synthesis associated with terminally deleted viruses could be due to destabilization of the ribonucleoprotein complexes formed. 41 4
5 INTRODUCTION Coxsackie B viruses (CVB), which belong to the Enterovirus genus of the Picornaviridae family, have non-enveloped icosahedral capsids composed of 60 copies each of the four structural proteins (VP1 to VP4), enclosing a singlestranded, positive-sense RNA genome of ~7,400 nucleotides (1). The genome consists of a single open reading frame encoding 11 mature proteins, flanked on both the 5' and 3' ends by noncoding regions (NCRs) (1-3). The enterovirus 5 NCR is ~750 nucleotides in length and has several complex RNA secondary structures including an internal ribosome entry site (IRES) involved in capindependent translation and a 5' terminal cloverleaf or stem-loop I (S-L I) required for replication of the viral genome. Stem-loop I includes approximately the first 110 nucleotides of the 5 NCR and is organized into one stem (a) and three stem-loop structures (b, c, d), which bind cellular and viral proteins involved in viral RNA synthesis (4, 5). In the cytoplasm of infected cells, cellular poly(rc)-binding protein 2 (PCBP2) and viral protein 3CD pro bind stem-loops b and d of stem-loop I, respectively, at the 5 end of the enterovirus genome. This ribonucleoprotein complex is required for the initiation of anti-genomic (negative-strand) RNA synthesis as well as genomic (positive-strand) RNA synthesis (4-10). PCBP2 also binds to a C-rich stretch of nucleotides located 3 of the stem a in S-L I (4, 11). In turn, heterogeneous nuclear ribonucleoprotein C (hnrnp C) is thought to bind to the cloverleaf at the 3' end of anti-genomic (negative-strand) viral RNA to facilitate the initiation of positive-strand RNA synthesis (12). These newly-synthesized positive-strand RNAs are either used 5
6 as messenger RNAs for further rounds of translation and RNA replication or as genomic RNAs packaged into virions for subsequent rounds of infection (3, 13). The CVBs are common human pathogens, transmitted through fecal-oral and respiratory routes, and are considered the main viral cause of acute myocarditis in children and young adults in developed countries (14, 15). Following the acute phase, myocarditis can become chronic and progress toward the clinical stage of dilated cardiomyopathy (DCM). This disease state is characterized by right and left ventricle enlargement with systolic function impairment and chronic evolution toward complete heart failure, thereby requiring heart transplantation. During the progression of disease from acute myocarditis to DCM, CVB can be detected in heart tissues and, more specifically, in cardiomyocytes. A correlation between enterovirus replication and poor clinical outcomes has been demonstrated, suggesting that persistent viral replication activities are potentially involved in disease progression (16-22). Moreover, expression of viral 2A proteinase alone in a murine model of CVB3 myocarditis has been shown to generate dilated cardiomyopathy (23). Expression of 2A can cause significant impairment of cardiomyocyte function through proteolytic cleavage of dystrophin, resulting in a decrease in cell contractility, an increase in membrane permeability, and a focal spread of the virus to adjacent cells (23-28). However, the molecular mechanisms by which the virus can persist in the heart from acute myocarditis to DCM are poorly understood, thereby limiting the development of new specific therapeutic strategies. 6
7 Recently, Bouin and colleagues reported CVB3 strains harboring 5 terminal genomic RNA deletions, ranging from 15 to 48 nucleotides in length, in explanted heart tissue collected from a 47-year-old immunocompetent woman suffering from idiopathic DCM (29). These terminal deletions, which partially removed 5 S-L I sequences, mirrored results previously obtained by Chapman and colleagues, who demonstrated that terminally deleted (TD) mutations can occur in a patient naturally infected with CVB2 and suffering from fulminant myocarditis, in the heart and the pancreas of mice inoculated with wild-type CVB3, and during CVB3 passages in primary cell cultures (30-32). However, these recent findings showed for the first time the existence of terminally deleted (TD) CVB genomic RNA populations in a patient suffering from chronic cardiomyopathy. Deletions involving sequences within the 5 S-L I structure of genomic RNAs are known to cause profound changes in viral biology by reducing viral replication, when compared to wild-type viruses (3, 32-35). Indeed, the discovery of TD mutations in heart biopsies was associated with low viral loads and abnormal positive- to negative-strand viral RNA ratios, close to one rather than the high ratios (30 to 70) normally observed in enterovirus-infected cells with wild-type viruses (32, 36-38). Moreover, TD variants of CVB3 replicate in both cell culture and tissues from experimentally-inoculated mice and from naturally infected humans without apparent cytopathic effect (31, 32). Overall, 5 terminal deletions within genomic RNAs could be a mechanism by which CVB can persist covertly in the heart long after the acute infection cycle with limited, 7
8 but progressive, damage to the myocardium. A prolonged, persistent viral infection could then explain how chronic cardiomyopathies develop via the continuous synthesis of viral proteins with pro-apoptotic, immuno-modulating, or other harmful activities for cardiac cell structures and functions. To date, translation and RNA replication of 5 terminally deleted CVB3 in a cardiac cell model have not been studied. Moreover, the mechanism by which deletions within the 5 S-L I affect the replication of the viral RNA is still unknown. The aim of this study was to assess in vitro the consequences of the 5 S-L I deletions observed in vivo on CVB3 replication. Using cell-free extracts of uninfected HeLa cells and cell culture transfection of luciferase replicons in two types of cardiomyocytes, we evaluated translation and RNA replication of full-length and 5 terminally deleted CVB3 RNAs. These experiments demonstrated that CVB3 RNAs harboring 5 terminal deletions from 7 to 49 nucleotides can be translated nearly as efficiently as those of wild-type strains. However, transfection of reporter RNAs harboring these deletions into cultured cardiomyocytes demonstrated a loss of detectable RNA amplification signal beyond the translation of input RNAs. In vitro replication experiments in HeLa cell extracts confirmed these findings, showing greatly reduced levels of negative- and positive-strand RNA synthesis in each of the terminally-deleted RNAs. Among the terminally deleted CVB3 RNAs tested, RNA synthesis was observed only for those harboring deletions of either 7 or 21 nucleotides. The only RNAs produced from these deleted templates were double-stranded (ds) RNAs [without detectable single-stranded (ss) RNAs], suggesting an abnormal 8
9 positive- to negative-strand RNA ratio near 1, similar to the ratios found in vivo for TD viruses (36). Partial deletion of the 5 S-L I sequences would be predicted to alter the binding sites of proteins involved in CVB RNA synthesis, possibly providing an explanation for the low levels of TD RNA replication. To address this possibility, we used RNA mobility shift assays to investigate the binding of host protein PCBP2 and viral protein 3CD pro to the 5 end of deleted forms of positive-strand RNA. We found that binding of these cellular and viral replication factors was qualitatively conserved for the 5 terminally deleted viruses, independent of the size of the deletion; however, the stability and/or protein binding affinity of the ribonucleoprotein complexes decreased with the increase in size of the deletion, since they required higher protein concentrations to form. Overall, this study provides an in vitro analysis of RNP complexes that may be involved in persistent forms of CVB3 observed in vivo from heart tissues. Our data suggest a possible explanation for the low levels of RNA replication of deleted strains as a result of the consequences of the deletions on the formation of the RNP complex. In addition, these data raise the question of the capacity of some of the deleted strains to replicate autonomously, since no significant levels of RNA synthesis were detected for 5 terminal deletions larger than 21 nucleotides. 9
10 MATERIALS AND METHODS Coxsackievirus cdna clones. Six previously described cdna clones of coxsackievirus B3 strains were used in this study. Two of the cdnas encode full-length wild-type CVB3 RNA genomes. pcvb3-28 encodes the cdna 157 genome of the cardiovirulent CVB3 strain 28 (Genbank Accession: AY ) with a ribozyme designed to cleave T7 RNA polymerase transcripts at nucleotide 1 of the genome (39, 40). pcvb3-0 encodes the cdna of the non-cardiovirulent CVB3 strain 0 (Genbank Accession: AY ) initially described by Charles Gauntt and differing only in one nucleotide at position 234 (U in CVB3/28; C in CVB/0) from CVB3 strain 28 (40-42). Four of the cdnas encode terminally-deleted CVB3 genomes. pcvb3-td7, pcvb3- TD30, pcvb3-td49 are variants of pcvb3-28 in which 5 terminal deletions of 7, 30, and 49 nucleotides, respectively, were engineered into the pcvb3-28 genome with a ribozyme to ensure cleavage at the authentic 5 terminal nucleotide of each genome (32). pcvb3-td21 encodes the 5 terminal sequence of a virus detected in the heart of a fatal human case of CVB2 infection engineered into pcvb3-28 as above but with a ribozyme designed to ensure cleavage at the 22 nd nucleotide to generate a deletion of 21 nucleotides relative to wild-type RNA (30). Cardiomyocytes. Two types of cardiac cell lines were used in this study. Primary human cardiomyocytes (HCM) (ScienCell Research Laboratories) isolated from human heart were maintained in Cardiac Myocyte Medium (ScienCell Research Laboratories) in tissue culture flasks coated with poly-l- 10
11 lysine. Primary cell cultures were used between passages 4 to 15. HL-1 cells are a cardiac myocyte cell line that can be repeatedly passaged and yet maintain a cardiac-specific phenotype. The HL-1 cell line was established from an AT-1 subcutaneous tumor excised from an adult female Jackson Laboratory C57BL/6J mouse. They were maintained in Claycomb medium (Sigma) and plated in tissue culture flasks coated overnight with gelatin (0.02% w/v) and fibronectin (0.5% v/v) (43). RNA transcript preparation. Plasmids corresponding to full-length, wild-type and terminally-deleted forms of CVB RNAs were linearized with the restriction enzyme Cla I. Transcription reactions were performed with T7 RNA polymerase using a MEGAscript T7 transcription kit (Ambion). Fifty microliter transcription reactions were incubated for 4 h at 37 C and terminated by treatment with DNase I for 15 min at 37 C. RNAs were purified using RNeasy Mini kit (Qiagen). For in vitro translation-replication reactions, RNAs were additionally purified by phenol-chloroform extractions and precipitated with ethanol. In vitro translation and RNA replication reactions. The coupled in vitro translation and RNA replication assays were carried out as described previously (44). Briefly, translation-replication mixtures (50 µl total) contained 60% microccocal nuclease-treated HeLa S10 cytoplasmic extract (vol/vol), 3 µg in vitro transcribed CVB3 RNA, and all-4 buffer containing reduced levels of CTP (1 mm ATP, 0.25 mm GTP, 0.25 mm UTP, 0.01 mm CTP [GE Healthcare Life Sciences], 60 mm potassium acetate, 30 mm creatine phosphate, 0.4 mg/ml creatine kinase, 20 mm HEPES-KOH [ph 7.4]). The reaction mixtures were 11
12 then split into two portions, with 10 µl for translation and the remaining 40 µl for RNA replication. Ten µci [ 35 S]methionine (PerkinElmer) was added to the translation reaction, which was incubated for 6 h at 30 C, supplemented with 2X Laemmli sample buffer (LSB) and boiled. The translation products were resolved by SDS-PAGE (12.5% resolving gel, 5% stacking gel) followed by fluorography. The RNA replication mixture was incubated at 30 C for 4.5 h before 25 µci [ 32 P]CTP (PerkinElmer) was added, and the reaction mixture was incubated for an additional 2 h at 34 C. The RNA replication product was then purified using an RNeasy Mini kit (Qiagen) and separated on a 1.1% agarose gel. The levels of 18S and 28S rrna in each lane, as determined by examination of ethidium bromide stained gels under UV light, were used to confirm equal loading of samples in the gel. Both translation and RNA replication gels were subjected to autoradiography on phosphor screens performed using a Personal Molecular Imager FX (Bio-Rad) and analyzed with Quantity One software (Bio-Rad). Luciferase replicons, transfection of cardiomyocytes, and luciferase assays. The CVB3 replicon (prib-cvb3-rluc, kindly provided by Martijn Langereis and Frank van Kuppeveld) contains a CVB3 cdna in which the P1 capsid coding region has been replaced by the Renilla luciferase gene. The cdna was cloned downstream from a hammerhead ribozyme sequence to remove non-viral nucleotides from the 5 -ends of transcript RNAs (45). To generate 5 terminally deleted CVB3 Renilla luciferase replicons harboring genomic deletion sizes identical to those tested using in vitro translation and 12
13 RNA replication assays, the prib-cvb3-rluc replicon as well as CVB3 cdna clones (pcvb3-28, pcvb3-td7, pcvb3-td21, pcvb3-td30, pcvb3-td49) were double-digested with BlpI and XmaI restriction enzymes. A fragment of 330 nucleotides encompassing the ribozyme and the 5 end of the viral genome was removed from the wild-type (pcvb3-28) and the four 5 terminally deleted (pcvb3-td7, pcvb3-td21, pcvb3-td30, pcvb3-td49) CVB3 cdna constructs and gel purified. In addition, the digested prib-cvb3-rluc replicon vector was isolated and gel purified. The 330 nucleotide fragments removed from CVB3 cdna constructs were cloned into the prib-cvb3-rluc replicon vector. The CVB3-0 replicon was generated from the CVB3-28 replicon by site directed mutagenesis of nucleotide position 234 (T in CVB3/28; C in CVB/0). For RNA transfection experiments, ~90% confluent monolayers of cardiac myocyte cells (HCM or HL-1 cells) grown in six-well plates were rinsed twice with phosphate-buffered saline (PBS). A transfection mixture consisting of 2 µg of replicon RNA, 250 µl of optimem (ThermoFisher Scientific), 5µl of mrna Boost reagent, and 5 µl of TransIT mrna transfection kit (Mirus) was incubated for 5 min at room temp and then added to each well. Cultures were incubated at 37 C for 2, 4, 6 or 8 h. Parallel transfections and subsequent incubations were carried out in the presence of 2 mm guanidine hydrochloride to prevent RNA replication and to allow for the measurement of luciferase levels at each time point that resulted solely from the translation of input replicon RNA. At the indicated times post-transfection, cells were washed twice with PBS. A Renilla luciferase assay (Promega) was used for quantitation of luciferase 13
14 activity. Cell monolayers were harvested with 500 µl of lysis buffer added to each well, and cell lysates were collected in 1.5 ml Eppendorf tubes. A volume of 20 µl of lysate was used in luciferase assays with the addition of 100 µl of luciferin substrate, and light emission was measured with a Monolight 2010 luminometer (Analytical Luminescence Laboratory). Triplicate samples were analyzed for each time point, and each time course experiment was performed three times Electrophoretic mobility shift assays. RNA mobility shift assays were performed as previously described (44, 46). Briefly, increasing amounts of purified recombinant PCBP2 (molar concentration amounts ranged from 2.5 to 7.5 µm) and/or recombinant purified 3CD pro [3CD(µ10)] (molar concentration amounts ranged from 0.05 to 10 µm) were incubated in the presence of RNA binding buffer (5 mm HEPES-KOH [ph 7.4], 25 mm KCl, 2.5 mm MgCl 2, 20 mm dithiothreitol, 3.8% [vol/vol] glycerol, 1 mg/ml of E. coli trna (Sigma), 8 U of Rnasin (Promega) ribonuclease inhibitor, and 0.5 mg/ml of bovine serum albumin [New England Biolabs]) with [ 32 P]UTP (PerkinElmer) radiolabeled wildtype or 5 terminally-deleted RNA probe derived from transcription of CVB3 cdna clones linearized with XmnI restriction enzyme. The final concentration of probe was 0.1 nm contained within a total reaction volume of 10 µl. This reaction was incubated for 10 min at 30 C. Following incubation, 2.5 µl of 50% glycerol was added, and the resulting complexes were resolved at 4 C on native 4% polyacrylamide gels containing 5% glycerol in Tris-borate-EDTA (TBE) buffer. Images were generated from scans of phosphor screens 14
15 performed using a Personal Molecular Imager FX (Bio-Rad) and analyzed using Quantity One software. 15
16 RESULTS Terminally-deleted CVB3 RNAs are defective in replicon reporter assays. Deletions of CVB3 strains detected in patients suffering from DCM involved the 5' terminus of the viral genome, which is a noncoding region but crucial for the regulation of translation and virus replication mechanisms. To determine the effects of these deletions on virus replication, we transfected full-length or deleted luciferase replicons, with 7 to 49 nucleotide deletions at the 5 'end of the viral genome, into heart-derived cells relevant for the pathophysiology of DCM: primary human cardiomyocytes (HCM) and a murine cardiomyocyte cell line (HL-1) (Figure 1A). As shown in Figure 1B, the replicon reporter produces a luciferase signal that is highly amplified if the input templates are translated and replicated. Luciferase activity was measured at 2 hours post-transfection, a time at which transfected replicon RNA is being translated prior to replicon replication. This signal was similar for all of the replicons, independent of the size of the deletion or the type of cardiac cells (Figure 2). However, if the incubation is extended beyond 2 hours, a significant difference in luciferase activity was observed starting at 2 h post-transfection for both wild-type strains (CVB3-28 and CVB3-0), increasing by up to 2 log 10 units above the luciferase levels produced by the replicons harboring terminal deletions (Figure 2). This difference is likely due to a lack of replication from the deleted genomic RNAs, causing reduced numbers of nascent genomic RNAs, resulting in reduced luciferase signal amplification (refer to Figure 1). 16
17 To test whether the reduced level of luciferase signals in cells transfected with replicons harboring 5 terminal deletions was due to RNA synthesis defects, we carried out luciferase assays following transfection of replicons into cardiomyocytes with guanidine hydrochloride in the cell culture medium. Guanidine hydrochloride (GuHCl) is a potent inhibitor of enterovirus RNA replication targeting the 2C protein (47). In the absence of GuHCl, the increase in the luciferase signal over time following replicon transfection is the sum of both virus translation and RNA replication through translation of newly synthesized positive-strand RNAs leading to luciferase signal amplification (refer to the scheme depicted in Figure 1). In the presence of GuHCl, the luciferase signal represents only translation from the input replicon RNA. Thus, the difference between luciferase signals measured in the absence and presence of GuHCl provides an assessment of virus genomic RNA replication. The two cardiac myocyte cell types were transfected with deleted and wild-type replicons in the presence or absence of GuHCl. A two log-reduction of luciferase activity was observed after GuHCl treatment of human and murine cardiomyocytes transfected with the replicons generated from wild-type strains 28 and 0 (Figure 3A and Figure 3B). However, no decrease in the luciferase signal was observed in GuHCl treated cells transfected with the deleted replicons by comparison to the non-treated cardiac cells (Figure 3C and 3D). The limited effect of GuHCl on the luciferase signal produced by the terminallydeleted replicons suggests that this signal was the result of input replicon translation without RNA replication. 17
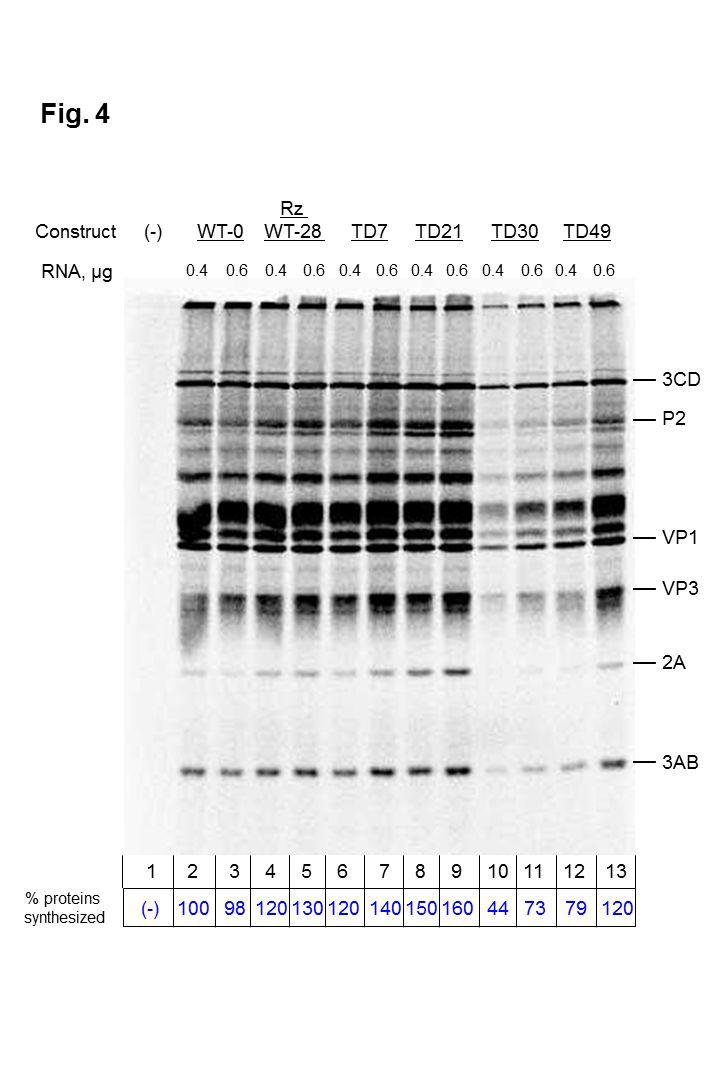
18 Translation of TD genomic RNAs compared to wild-type CVB3 RNA. Although the observed deletions affected only the cloverleaf or stem-loop I (S-L I) in the 5 NCR, a secondary structure element required for RNA replication, and did not involve the IRES that is required for protein synthesis, the deletions may nevertheless have consequences on the stability or overall structure of viral RNA and thus affect translation. In addition, the pathophysiology of DCM in humans is partially explained by expression of the viral 2A proteinase, which cleaves dystrophin, a cardiac myocyte cytoskeletal protein. Therefore, it was important to determine the translation patterns and efficiencies directed by TD RNAs found during persistent CVB3 infections in the myocardium. Using in vitro transcribed RNAs synthesized from six plasmid constructs containing the cdna of two wild-type strains [cardiovirulent (CVB3-28) and non-cardiovirulent (CVB3-0) in mice] and of four TD strains with deletions ranging from 7 to 49 nucleotides, we carried out in vitro translation assays in a cytoplasmic extract (S-10) from uninfected HeLa cells in the presence of [ 35 S]methionine. Viral translation products were resolved by SDS-polyacrylamide gel electrophoresis and visualized by fluorography. To determine the relative levels of viral proteins synthesized by each RNA, signals detected on the gel for viral proteins 3CD, P2, VP1, VP3, 2A, and 2AB (collectively) were quantified. WT-0, 0.4 µg RNA template (Figure 4, lane 2) was set as the 100% reference control. Transcripts produced from the WT-0 construct do not have a hammerhead ribozyme at their 5 termini, while the transcripts from the WT-28, TD7, TD21, TD30, and TD49 18
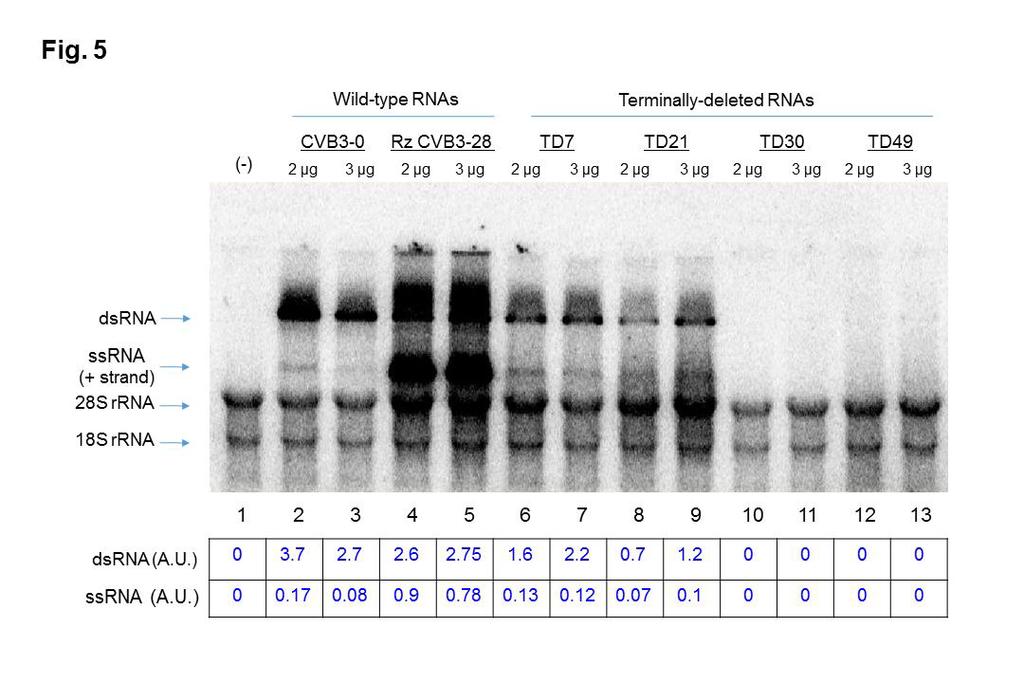
19 constructs each contain a hammerhead ribozyme at their 5 termini to generate authentic 5 terminal CVB3 sequences. The additional, non-viral sequences present at the 5 termini of WT-0 transcripts may account for the slightly reduced differences in translation efficiency compared to WT-28 and some of the TD transcripts. Overall, the data displayed in Figure 4 show similar patterns of translation products for all RNAs tested, although the RNAs with the largest deletions (TD30 and TD49) produced lower overall levels of CVB3 proteins, particularly at the lower concentrations of RNA. Transcripts corresponding to the TD30 deletion of CVB3 RNA reproducibly produced lower levels of viral proteins following in vitro translation (refer to Figure 4, lanes 10 and 11), even at much higher RNA concentrations (unpublished observations), suggesting that this deletion may have affected the overall structure of the 5 NCR, including the IRES element In vitro RNA synthesis directed by TD RNAs is impaired. Given that RNA amplification of the luciferase signal following transfection of cultured cells requires the synthesis of new positive-strand RNAs, it is possible that the TD RNAs may still be capable of negative-strand RNA synthesis but not positivestrand RNA synthesis. To test this possibility, we performed an in vitro RNA replication assay by adding [ 32 P]CTP to in vitro translation assays that had progressed for 4.5 h and incubating for an additional 2 h at 34 C. Newly synthesized RNAs were then visualized following electrophoresis on an agarose gel (Figure 5). Replication of RNA corresponding to wild-type strains 19
20 CVB3-28 and CVB3-0 was readily detected (Figure 5). Both single-stranded (ss) RNA (positive-strand) and double-stranded (ds) RNA [replicativeintermediate (RI) or replicative-form (RF)] were detected during replication of CVB3-28 RNA whereas only dsrnas were observed for CVB3-0. The plasmid DNA template encoding CVB3-28 RNA harbors a hammerhead ribozyme at the 5 end of the transcript (32), which produces an authentic CVB3 5 terminus. This transcript is an efficient template for negative-strand RNA synthesis and subsequent rounds of positive-strand RNA synthesis. In contrast, in vitro RNAs produced from pcvb3-0 have no hammerhead ribozyme at the 5 end and harbor non-viral sequences that result in inefficient ssrna (positive-strand) synthesis (13, 35). As noted above, all TD transcripts harbor a hammerhead ribozyme at their 5 termini, producing authentic 5 terminal CVB3 sequences. Among the CVB3 TD RNAs, RNA synthesis was observed only for template RNAs harboring 7 or 21 nucleotide deletions, which were the smallest deletion sizes tested (refer to Figure 5). No RNA replication was detected for deletions of 30 or 49 nucleotides (Figure 5, lanes 10 through 13). Moreover, RNAs detected during TD7 and TD21 replication were dsrnas (i.e., RF and RI) resulting from negative-strand RNA synthesis with only very low levels of ssrna (positive-strand), as shown in Figure 5, lanes 6 through 9, even though all of the TD RNA transcripts harbored a self-cleaving ribozyme at their 5 ends. Overall, the results from our in vitro RNA replication assays as well as from transfections of luciferase replicons into cardiomyocytes (with or without GuHCl) suggest that only TD7 and TD21 are capable of RNA replication, although 20
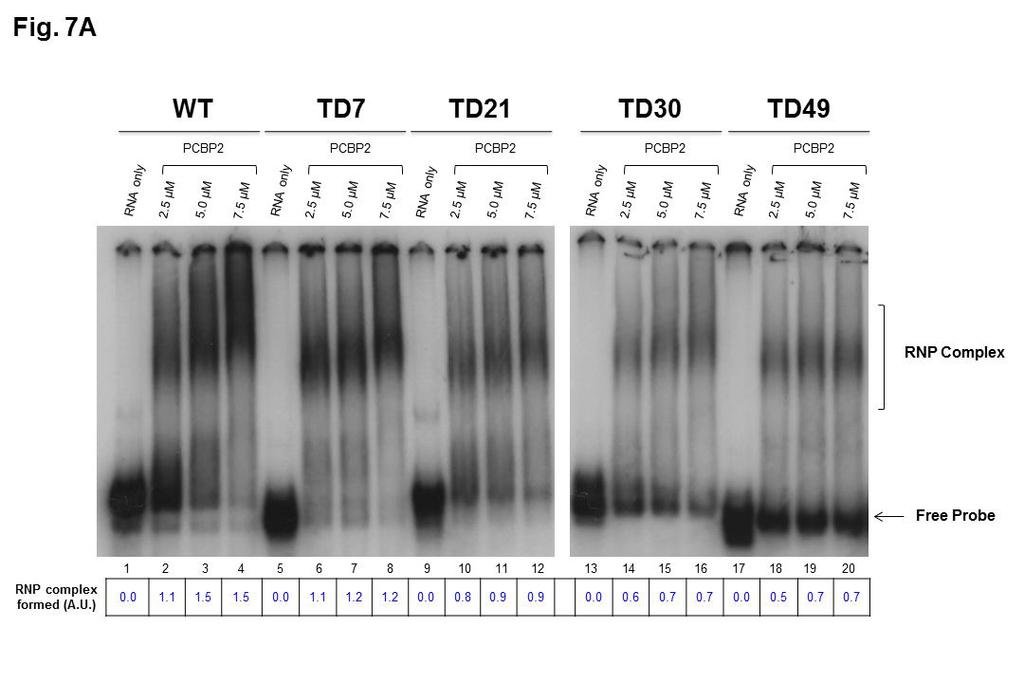
21 positive-strand RNA synthesis was greatly reduced compared to wild-type RNAs Binding of cellular and viral proteins to RNAs harboring deletions of sequences within stem-loop I of the CVB3 5 NCR. The drastic reduction of TD RNA synthesis capabilities could be explained by the removal of binding sites on the viral RNA for viral and cellular proteins involved in ribonucleoprotein complex formation required for RNA replication. The cloverleaf or S-L I at the 5 end of positive-strand enterovirus RNAs is the binding site for the cellular protein PCBP2 and the viral protein 3CD pro. The binding of these two proteins forms a ribonucleoprotein complex required for the initiation of negative-strand as well as positive-strand RNA synthesis (5, 7, 10, 48), as depicted in Figure 6. We therefore investigated the interactions between the deleted positive-strand RNAs and these two proteins involved in viral RNA replication complex formation. The binding of deleted positive-strand viral RNA to PCBP2 and 3CD pro, either alone or in combination, was analyzed by RNA mobility shift assays performed using recombinant proteins and RNA probes containing 5 S- L I, including the adjacent C-rich sequences, generated by transcription of linearized, deleted cdna CVB3 constructs. RNA corresponding to S-L I- containing sequences from the wild-type CVB3 28 strain was used as a positive control. Radiolabeled CVB3 S-L I-containing RNA was incubated alone or with increasing concentrations of recombinant purified PCBP2 ranging from 2.5 µm to 7.5 µm. Resolution following electrophoresis on a native polyacrylamide gel 21
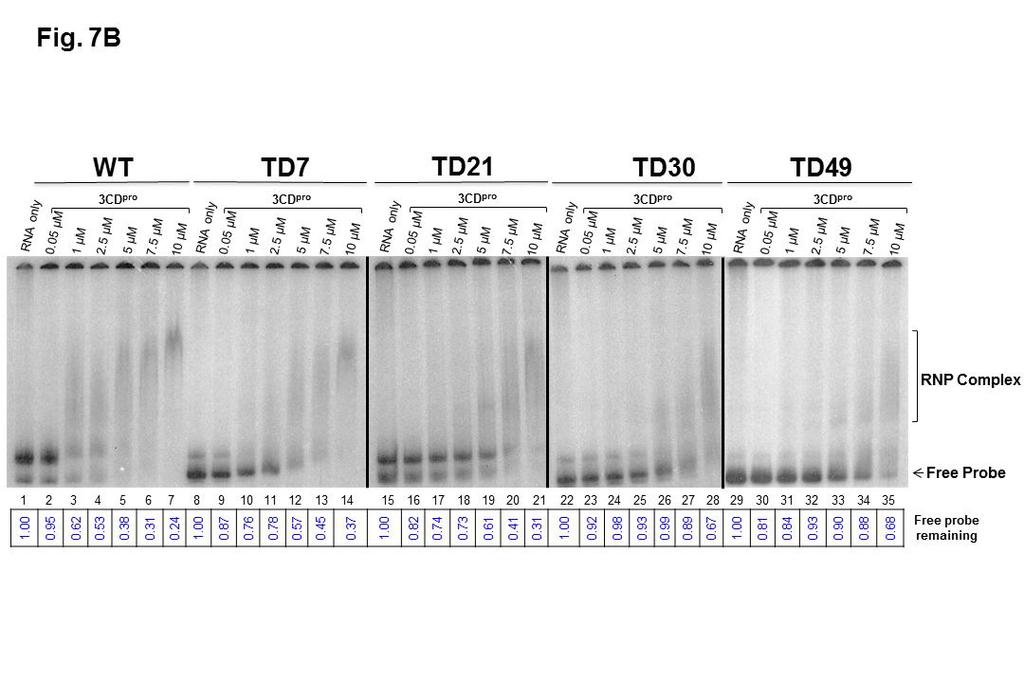
22 showed RNP complex formation through a slower migration of all the RNA probes incubated with PCBP2, demonstrating protein binding even with the deletion of 49 nucleotides disrupting stem-loop b of S-L I previously described as a PCBP2 binding site (Figure 7A). However, the data displayed in Figure 7A, lanes 9 through 20, demonstrate that the efficiency of complex formation is reduced when templates of increasing size deletions (21, 30, or 49 nucleotides) are incubated with equivalent concentrations of PCBP2, as shown by decreased levels of RNP complexes and increased levels of free probe RNAs. Using imaging software to quantify band densities of RNP complexes formed under these experimental conditions, we estimate that TD7 transcripts bind PCBP2 at levels approx. 80% of those of WT RNA, while TD21 transcripts bind at approx. 60% compared to WT RNA. For the TD30 and TD49 transcripts, binding efficiency was less than half of WT RNA at the highest concentrations of PCBP2 used in our mobility shift assays. When deleted forms of CVB3 S-L I-containing RNAs were incubated with recombinant purified 3CD pro at concentrations ranging from 0.05 µm to 10 µm, all of the TD positive-strand RNAs were capable of binding 3CD pro, as shown in Figure 7B. As we observed for PCBP2 binding to these same deleted forms of RNA, RNP complex formation was associated with the disappearance of the free probe on the gel and a dose-dependent increase in the formation of slowermigrating species, with constructs harboring larger deletions requiring increased concentrations of 3CD pro to form slower-migrating complexes. A 1 µm concentration of 3CD pro was necessary for RNP complex formation with wild- 22
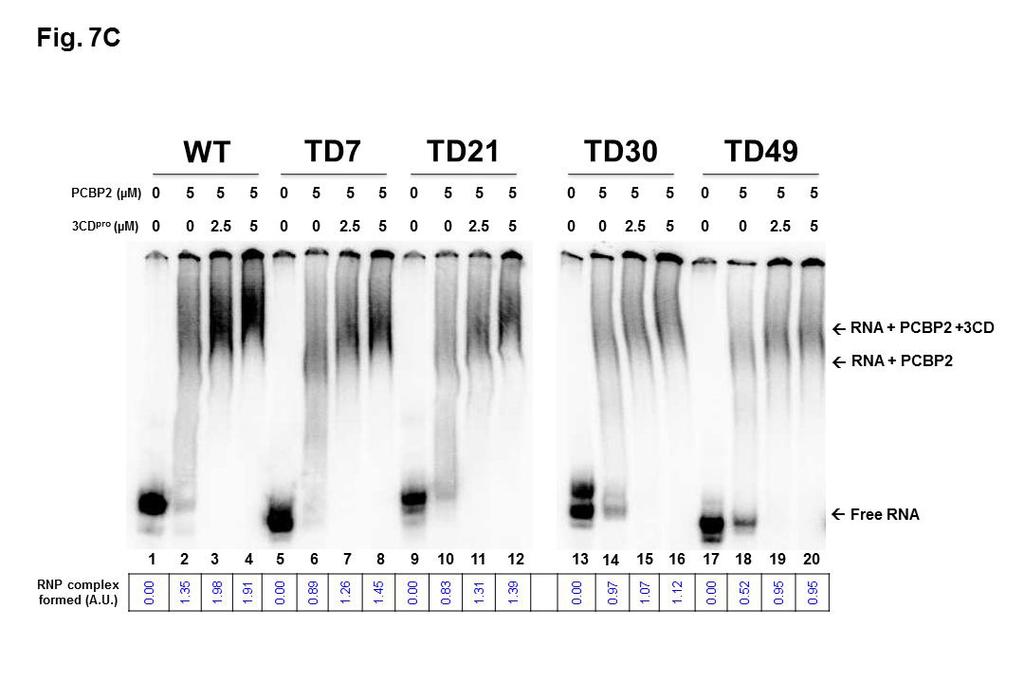
23 type CVB3-28 S-L I-containing RNA whereas 7.5 µm and 10 µm of 3CD pro were required for complete complex formation for TD21 and TD49 RNAs, respectively. As with the experiments describing RNP complex formation with recombinant PCBP2, we used imaging software to quantify band densities of RNP complexes formed between WT and TD RNAs with recombinant 3CD. Because the RNP complexes formed with 3CD are diffuse and difficult to quantify directly, we used the disappearance of radiolabeled probe as a measure of RNA-protein complex formation. As shown at the bottom of Figure 7B, TD7 and TD21 transcripts formed RNP complexes with 3CD with efficiencies approx % of WT RNA. In contrast, RNAs corresponding to TD30 and TD49 were able to form complexes with recombinant 3CD at levels less than 50% of WT RNAs. To analyze ternary complex formation, increasing amounts of 3CD pro ranging from 0 to 5 µm were added to RNA binding reaction containing a fixed concentration of PCBP2 (5 µm) and a constant amount of labeled S-L I- containing probe. Results from these electrophoretic mobility shift experiments showed a super-shifted RNP complex generated from deleted viruses, when incubated with the two proteins simultaneously, confirming that deleted forms of CVB3 S-L I were capable of binding these two replication factors (Figure 7C). Consistent with the data shown in Figures 7A and 7B, the efficiency of this binding (and ternary complex formation) decreased with the increasing size of the deletion, as demonstrated by the reduced levels of RNP complex formation (refer to lanes 5 through 20 in Figure 7C). At the highest concentrations of 23
24 PCBP2 and 3CD used in our assays, quantitation of the band densities of RNP complexes in Figure 7C revealed that TD7 and TD21 RNAs bound proteins at levels between 70 and 80% of WT RNAs while TD30 and TD49 were only capable of complex formation at 50-60% efficiency compared to WT CVB3 RNA. 24
25 DISCUSSION Enteroviruses are primarily known for their involvement in acute infections such as aseptic meningitis, upper or lower respiratory tract infections, hand, foot, and mouth disease, or acute myocarditis (49). Nonetheless, the role of enteroviruses as the etiologic agent of chronic diseases, in the context of persistent viral infection, has been described with increasing frequency. Several studies have associated persistent enterovirus infection with chronic diseases such as amyotrophic lateral sclerosis, post-polio syndrome, chronic fatigue syndrome, type 1 diabetes, and chronic cardiomyopathy [i.e., chronic myocarditis and dilated cardiomyopathy] (16-20, 50-55). These chronic cardiac infections involved group B coxsackieviruses. However, the mechanisms leading to CVB persistence in the heart as well as the link between viral persistence and the development of chronic cardiomyopathy are not well understood. In 2005, Chapman, Tracy, and colleagues identified variants of CVB in hearts of mice inoculated with wild-type CVB3 that contained terminal deletions in the 5 NCR of their genomic RNAs (32). These studies provided a possible link between the generation of non-cytopathic viruses via genomic RNA terminal deletions and the establishment and/or maintenance of persistent infections. Recently, similar deletions have been detected for the first time in cardiac biopsies of patients suffering from chronic cardiomyopathy (29, 36). These deletions, ranging from 15 to 48 nucleotides in length, involved a 5 RNA secondary structure (S-L I) that is the binding site for both viral and host cell proteins forming RNA replication complexes. Subsequently, Smithee and co- 25
26 workers reported that CVB TD populations displayed impaired replication with RNA levels 100,000 fold lower than those of wild-type virus (56). With the exception of the largest identified deletion of 49 nucleotides, the specific impact of the different sizes of genomic deletions detected during persistent cardiac infections in mice or humans on the translation and replication of viral RNA had not been analyzed prior to the present study. Moreover, previous studies on TD virus replication have been mainly performed in non-cardiac cell lines or cellfree reactions, with very little data about TD replication in cardiomyocytes, the cell type infected in vivo. Finally, the suggestion that altered formation or stability of replication complexes with deleted forms of viral RNA might be the explanation for impaired replication of TD viruses has so far not been investigated. The work described in this study is the first use of cardiac myocyte cultures to assess the consequence of enterovirus genomic RNA deletions, detected in patients with chronic cardiomyopathy, on CVB3 translation and RNA replication. Transfection of deleted and non-deleted luciferase replicons in two types of heart cells demonstrated that at 2 hours post-transfection, equivalent levels of luciferase activity were detected corresponding to the translation of input RNA prior to the onset of genome amplification by RNA replication. These results were confirmed by in vitro translation reactions in HeLa S10 extracts, which showed nearly equivalent levels of protein synthesis among deleted and wild-type CVB3 RNAs. Taken together, our cell culture reporter transfection 26
27 experiments and in vitro assays demonstrated the ability of TD RNAs harboring deletions of up to 49 nucleotides to synthesize viral proteins. CVB protein expression could, in part, be involved in DCM pathophysiology due to the expression of the viral 2A proteinase in infected cardiomyocytes (23). In cardiomyocytes, this viral protein has been shown to cleave the host cell protein dystrophin, which links actin filaments to the plasma membrane (26, 27). Cleavage occurs in the hinge 3 region of dystrophin and is sufficient to induce cardiomyopathy with disruption of the sarcolemmal membrane and loss of normal localization of dystrophin in the heart (27). Dystrophin cleavage also promotes viral propagation by allowing mature virions to exit the infected myocyte and infect adjacent myocytes (24). Moreover, the carboxyl-terminal fragment of 2A proteinase-mediated dystrophin cleavage by itself can cause marked dystrophic cardiomyopathy through myocardial fibrosis, heightened susceptibility to myocardial ischemic injury, and increased mortality during cardiac stress (28). It should be noted that overall viral protein expression by TD replicons remained lower than that of wild-type RNAs, as demonstrated by the luciferase levels at 4h post-transfection in cardiomyocytes (Figure 2). This decreased expression would still be sufficient to cause the onset of DCM as demonstrated in a murine model expressing 2A via cellular transcripts driven by the α-mhc promoter, which would be more akin to a chronic pattern of 2A expression in persistently infected cardiomyocytes compared to a fulminant CVB3 infection (23). Nevertheless, additional studies 27
28 will be required to determine if dystrophin is cleaved in cardiomyocytes persistently infected with TD viruses. Although replicons corresponding to TD viral genomes can synthesize viral proteins, their RNA synthesis levels are greatly reduced. Our in vitro replication experiments did not show detectable levels of RNA synthesis for deletions larger than 21 nucleotides. Moreover, the RNA detected was doublestranded without isolated single-stranded, positive-sense RNA. These in vitro results were confirmed by our data from transfection of deleted luciferase replicons into cardiomyocytes showing a lack of luciferase signal amplification from 4 h post-transfection as well as no effect of GuHCl on luciferase signals. Our data strongly suggest an RNA replication defect for the TD viral RNAs. During infection, this defect would likely lead to a positive-strand to negativestrand RNA ratio closer to 1 than to the asymmetric ratio of 30 to 70 that is normally observed during acute infections with wild-type enterovirus strains (37, 38). Our results confirmed those of a previous study in which no replication was detected following transfection of a CVB3 genome with a 32 nucleotide deletion of the 5 NCR into HeLa cells (57). Other studies have detected very low levels of replication of TD RNAs in HeLa S10 in vitro RNA synthesis reactions or with semi-nested RT-PCR amplification, even for the deletion of 49 nucleotides (32, 34, 56). The most likely explanation for these differences is that the methods used in our study were not sufficiently sensitive to detect the very low levels of RNA replication directed by the TD30 and TD49 RNAs in vitro. 28
29 However, Bouin and colleagues recently demonstrated the coexistence of a large majority of TD viral genomes with a minority of wild-type CVB3 genomic RNAs (less than 0.9% of the viruses sequenced) in cardiac biopsies of a patient suffering from DCM. This discovery was facilitated by the use of next generation sequencing, which enabled the detection of very minor variants representing less than 1% of the total viral RNA population that could not be detected by the Sanger sequencing method used in previous studies (29, 31, 32). It is possible that the replication of this remaining wild-type population of viruses could generate, by a presently unknown mechanism, a majority of viruses with terminally-deleted genomic RNAs in cardiomyocytes that are unable to replicate by themselves and a minority of wild-type viruses capable of maintaining a lowlevel chronic infection. The wild-type virus could also play the role of helper virus allowing TD viruses to replicate by providing, in trans or through genomic recombination events, the elements necessary for their replication (58-60). Wild-type and TD viruses could reproduce in vivo by using a mode of viral persistence known to occur in cell culture via defective interfering (DI) particles (61). In this model, picornavirus DI populations harboring deletions within the capsid protein coding region must co-exist with the wild-type population, since DI genomes are packaged using capsid proteins produced by wild-type viruses (62-64). The synthesis of even low levels of double-stranded RNA by TD viruses could be sufficient to induce type I interferon (IFN) secretion by infected cardiomyocytes. Indeed, dsrna, known as replicative form or replicative 29
30 intermediate, is recognized by cellular sensors, including Toll-like receptors 3 and 7, and RNA helicases, in particular MDA5, causing the virus-infected cardiomyocytes to produce type 1 IFNs and to promote the inflammatory response (65-70). As summarized by Althof and colleagues (71), the local synthesis of type 1 IFNs can be harmful to the myocardium by causing inflammatory cell infiltration including CD8+ T cells, macrophages, natural killer cells, and neutrophils, and inflammatory cytokine secretion, thereby causing immune-pathological damages (71-73). The prolonged presence of dsrna in cardiomyocytes may be associated with long-term morbidity caused by chronic inflammation. Such inflammation could promote immune-mediated tissue damage known to be involved in the pathogenesis of enterovirus-related cardiomyopathy. It remains to be seen if innate immune sensors in cardiomyocytes can detect the very low levels of dsrna synthesized by terminally-deleted viruses and subsequently stimulate type I interferon secretion. Persistent CVB strains must find a way to overcome this antiviral host response, since type I IFNs are efficient effectors of the innate immune response constraining CVB spread, leading to clearance of the virus from the myocardium following acute infection. Type I interferon secretion leads to the transcription of hundreds of interferon-stimulated genes, which display antiviral properties by targeting almost any step of the virus replication cycle (74). Interestingly, Diamond and colleagues demonstrated that alphaviruses, which are also single-stranded, positive-strand RNA viruses, use mutations within the 5' noncoding region affecting secondary structural elements of their RNA to 30
31 alter interferon-stimulated protein binding and functions (75). Their results suggest an evasion mechanism by which a deleted virus with modified 5 RNA secondary structures could avoid immune restriction, despite type 1 IFN secretion and interferon-stimulated gene transcription, leading to long-term virus persistence in the heart. To understand the possible cause(s) of impaired single-stranded RNA synthesis by TD strains of CVB3, we investigated binding to the 5 end of deleted positive-strand viral RNAs of cellular (i.e., PCBP2) and viral (i.e., 3CD) factors involved in viral RNA replication. These proteins form a ribonucleoprotein complex with stem-loop I RNA that enables 5-3 intramolecular interactions resulting in a protein-protein bridge, template recognition by the polymerase 3D, and RNA replication initiation (76). PCBP2 is primarily a nuclear-resident protein with RNA binding activity that binds stemloop b of S-L I at the 5 end of genomic RNA. This interaction displays increased affinity when the precursor to the virally encoded RNA-dependent RNA polymerase 3D pol and the proteinase 3C pro, 3CD pro, is also present on stem-loop d of S-L I, forming a ternary complex that promotes both negativestrand and positive-strand RNA synthesis (7, 10, 48, 77). Loss of the binding sites or modification of RNA secondary structure rendering interaction with these proteins less stable could explain, in part, the severe deficiency in positive-strand RNA synthesis of TD viruses. Indeed, we found that deletions of either 30 or 49 nucleotides of the CVB3 5 NCR had reduced levels of binding by PCBP2 and 3CD pro, corresponding to RNA synthesis defects exhibited in our 31
32 613 in vitro RNA replication assays. In particular, 3CD pro binding efficiency was reduced in accordance with the deletion size, since ribonucleoprotein complex required increased protein concentration to form. This result suggested that even if primary protein binding sites are conserved despite genomic deletions, RNP complex formation with specific host or viral proteins could be less stable and lead to lower levels of RNA synthesis during infection. It is known that host protein hnrnp C can bind both the 3 and 5 termini of poliovirus negative-strand RNA intermediates, an interaction proposed to promote the synthesis of positive-strand enterovirus RNA molecules (12, 78, 79). Thus, the 5 terminal deletions of positive-strand RNAs would lead to deletions in the 3 ends of negative-strand RNAs, possibly reducing the binding of hnrnp C to these replication intermediates. Loss of such binding might also contribute to the reduction of positive-strand RNA synthesis observed with TD viruses. Moreover, low levels of hnrnp C expression in the cytoplasm of quiescent and differentiated cells (like cardiomyocytes) have been suggested to give putative hnrnp C-independent TD genomes an evolutionary advantage over hnrnp C-dependent wild-type enterovirus RNA, thereby enabling them to become the dominant virus population in persistent infections (31, 80). Our future studies of RNP complexes formed with the 3 ends of negative strand CVB3 RNAs will address the possible roles of hnrnp C as well as other host and viral proteins in TD viral replication and their possible link to persistent infections. Overall, to better understand the mechanisms by which genomic deletions impact the synthesis efficiency of negative- and positive-strand viral 32
33 RNAs, a complete identification of the replication complexes used by wild-type and terminally-deleted CVB3 RNAs in cardiomyocytes will be required. This identification could help design new therapeutic strategies targeting either viral proteins or inhibiting host protein activities involved in viral RNA replication to treat persistent enterovirus cardiac infections mediated by TD strains. 33
34 ACKNOWLEDGMENTS We are indebted to Nora Chapman and Steve Tracy for valuable reagents and many thoughtful discussions. We thank Martijn Langereis and Frank van Kuppeveld for providing the CVB3 luciferase replicon construct and Dylan Flather and Wendy Ullmer for critical comments on the manuscript This research was supported by U. S. Public Health Service grants AI and AI from the National Institutes of Health to B. L. S.. N. L. was supported by a Marie Curie International Outgoing Fellowship for Career 649 Development from the European Commission (Contract N o ) and by grants from the Philippe Foundation and the Champagne-Ardenne region. A. B. is supported by a postdoctoral fellowship from the George E. Hewitt Foundation for Medical Research. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript. 34
35 REFERENCES 1. Racaniello, V. R Picornaviridae : The viruses and their replication, Fields Virology 6th edition, Lippincott Williams & Wilkins ed, vol. 1. Knipe & Howley, New York. 2. Brown, D. M., S. E. Kauder, C. T. Cornell, G. M. Jang, V. R. Racaniello, and B. L. Semler Cell-dependent role for the poliovirus 3' noncoding region in positive-strand RNA synthesis. J. Virol. 78: Sean, P., and B. L. Semler Coxsackievirus B RNA replication: lessons from poliovirus. Curr. Top. Microbiol. Immunol. 323: Toyoda, H., D. Franco, K. Fujita, A. V. Paul, and E. Wimmer Replication of poliovirus requires binding of the poly(rc) binding protein to the cloverleaf as well as to the adjacent C-rich spacer sequence between the cloverleaf and the internal ribosomal entry site. J. Virol. 81: Andino, R., G. E. Rieckhof, P. L. Achacoso, and D. Baltimore Poliovirus RNA synthesis utilizes an RNP complex formed around the 5'- end of viral RNA. EMBO J. 12: Andino, R., G. E. Rieckhof, and D. Baltimore A functional ribonucleoprotein complex forms around the 5' end of poliovirus RNA. Cell 63: Parsley, T. B., J. S. Towner, L. B. Blyn, E. Ehrenfeld, and B. L. Semler Poly (rc) binding protein 2 forms a ternary complex with the 5'-terminal sequences of poliovirus RNA and the viral 3CD proteinase. RNA 3: Ohlenschläger, O., J. Wöhnert, E. Bucci, S. Seitz, S. Häfner, R. Ramachandran, R. Zell, and M. Görlach The structure of the stemloop D subdomain of coxsackievirus B3 cloverleaf RNA and its interaction with the proteinase 3C. Structure 12: Zell, R., K. Sidigi, E. Bucci, A. Stelzner, and M. Görlach Determinants of the recognition of enteroviral cloverleaf RNA by coxsackievirus B3 proteinase 3C. RNA 8: Vogt, D. A., and R. Andino An RNA element at the 5'-end of the poliovirus genome functions as a general promoter for RNA synthesis. PLoS Pathog. 6:e Zell, R., Y. Ihle, S. Seitz, U. Gundel, P. Wutzler, and M. Gorlach Poly(rC)-binding protein 2 interacts with the oligo(rc) tract of coxsackievirus B3. Biochem. Biophys. Res. Commun. 366: Brunner, J. E., J. H. C. Nguyen, H. H. Roehl, T. V. Ho, K. M. Swiderek, and B. L. Semler Functional interaction of heterogeneous nuclear ribonucleoprotein C with poliovirus RNA synthesis initiation complexes. J. Virol. 79: Herold, J., and R. Andino Poliovirus requires a precise 5' end for efficient positive-strand RNA synthesis. J. Virol. 74:
36 Cooper, L. T., K. L. Baughman, A. M. Feldman, A. Frustaci, M. Jessup, U. Kuhl, G. N. Levine, J. Narula, R. C. Starling, J. Towbin, R. Virmani, A. American Heart, C. American College of, C. European Society of, A. Heart Failure Society of, and C. Heart Failure Association of the European Society of The role of endomyocardial biopsy in the management of cardiovascular disease. J. Am. Coll. Cardiol. 50: Dennert, R., H. J. Crijns, and S. Heymans Acute viral myocarditis. Eur. Heart J. 29: Rey, L., V. Lambert, P. Wattré, and L. Andréoletti Detection of enteroviruses ribonucleic acid sequences in endomyocardial tissue from adult patients with chronic dilated cardiomyopathy by a rapid RT-PCR and hybridization assay. J. Med. Virol. 64: Andréoletti, L., T. Bourlet, D. Moukassa, L. Rey, D. Hot, Y. Li, V. Lambert, B. Gosselin, J. F. Mosnier, C. Stankowiak, and P. Wattré Enteroviruses can persist with or without active viral replication in cardiac tissue of patients with end-stage ischemic or dilated cardiomyopathy. J. Infect. Dis. 182: Li, Y., T. Bourlet, L. Andreoletti, J. F. Mosnier, T. Peng, Y. Yang, L. C. Archard, B. Pozzetto, and H. Zhang Enteroviral capsid protein VP1 is present in myocardial tissues from some patients with myocarditis or dilated cardiomyopathy. Circulation 101: Andreoletti, L., D. Hober, C. Decoene, M. C. Copin, P. E. Lobert, A. Dewilde, C. Stankowiac, and P. Wattre Detection of enteroviral RNA by polymerase chain reaction in endomyocardial tissue of patients with chronic cardiac diseases. J. Med. Virol. 48: Andreoletti, L., P. Wattre, C. Decoene, P. E. Lobert, A. Dewilde, and D. Hober Detection of enterovirus-specific RNA sequences in explanted myocardium biopsy specimens from patients with dilated or ischemic cardiomyopathy. Clin. Infect. Dis. 21: Kim, K. S., G. Hufnagel, N. M. Chapman, and S. Tracy The group B coxsackieviruses and myocarditis. Rev. Med. Virol. 11: Rakar, S., G. Sinagra, A. Di Lenarda, A. Poletti, R. Bussani, F. Silvestri, and F. Camerini Epidemiology of dilated cardiomyopathy. A prospective post-mortem study of 5252 necropsies. The Heart Muscle Disease Study Group. Eur. Heart J. 18: Xiong, D., T. Yajima, B.-K. Lim, A. Stenbit, A. Dublin, N. D. Dalton, D. Summers-Torres, J. D. Molkentin, H. Duplain, R. Wessely, J. Chen, and K. U. Knowlton Inducible cardiac-restricted expression of enteroviral protease 2A is sufficient to induce dilated cardiomyopathy. Circulation 115: Lim, B.-K., A. K. Peter, D. Xiong, A. Narezkina, A. Yung, N. D. Dalton, K.-K. Hwang, T. Yajima, J. Chen, and K. U. Knowlton Inhibition of Coxsackievirus-associated dystrophin cleavage prevents cardiomyopathy. J. Clin. Invest. 123:
37 Andréoletti, L., L. Ventéo, F. Douche-Aourik, F. Canas, G. Lorin de la Grandmaison, J. Jacques, H. Moret, N. Jovenin, J.-F. Mosnier, M. Matta, S. Duband, M. Pluot, B. Pozzetto, and T. Bourlet Active coxsackieviral B infection is associated with disruption of dystrophin in endomyocardial tissue of patients who died suddenly of acute myocardial infarction. J. Am. Coll. Cardiol. 50: Badorff, C., N. Berkely, S. Mehrotra, J. W. Talhouk, R. E. Rhoads, and K. U. Knowlton Enteroviral protease 2A directly cleaves dystrophin and is inhibited by a dystrophin-based substrate analogue. J. Biol. Chem. 275: Badorff, C., G. H. Lee, B. J. Lamphear, M. E. Martone, K. P. Campbell, R. E. Rhoads, and K. U. Knowlton Enteroviral protease 2A cleaves dystrophin: evidence of cytoskeletal disruption in an acquired cardiomyopathy. Nat. Med. 5: Barnabei, M. S., F. V. Sjaastad, D. Townsend, F. B. Bedada, and J. M. Metzger Severe dystrophic cardiomyopathy caused by the enteroviral protease 2A-mediated C-terminal dystrophin cleavage fragment. Sci. Transl. Med. 7:294ra Bouin, A., Y. Nguyen, M. Wehbe, F. Renois, P. Fornes, F. Bani-Sadr, D. Metz, and L. Andreoletti Major persistent 5' terminally deleted coxsackievirus B3 populations in human endomyocardial tissues. Emerging Infect. Dis. 22: Chapman, N. M., K.-S. Kim, K. M. Drescher, K. Oka, and S. Tracy ' terminal deletions in the genome of a coxsackievirus B2 strain occurred naturally in human heart. Virology 375: Kim, K. S., N. M. Chapman, and S. Tracy Replication of coxsackievirus B3 in primary cell cultures generates novel viral genome deletions. J. Virol. 82: Kim, K. S., S. Tracy, W. Tapprich, J. Bailey, C. K. Lee, K. Kim, W. H. Barry, and N. M. Chapman '-Terminal deletions occur in coxsackievirus B3 during replication in murine hearts and cardiac myocyte cultures and correlate with encapsidation of negative-strand viral RNA. J. Virol. 79: Liu, Y., E. Wimmer, and A. V. Paul Cis-acting RNA elements in human and animal plus-strand RNA viruses. Biochim. Biophys. Acta 1789: Sharma, N., S. A. Ogram, B. J. Morasco, A. Spear, N. M. Chapman, and J. B. Flanegan Functional role of the 5' terminal cloverleaf in coxsackievirus RNA replication. Virology 393: Barton, D. J., B. J. O'Donnell, and J. B. Flanegan ' cloverleaf in poliovirus RNA is a cis-acting replication element required for negative-strand synthesis. EMBO J. 20: Lévêque, N., F. Renois, D. Talmud, Y. Nguyen, F. Lesaffre, C. Boulagnon, P. Bruneval, P. Fornes, and L. Andréoletti Quantitative genomic and antigenomic enterovirus RNA detection in 37
38 explanted heart tissue samples from patients with end-stage idiopathic dilated cardiomyopathy. J. Clin. Microbiol. 50: Giachetti, C., and B. L. Semler Role of a viral membrane polypeptide in strand-specific initiation of poliovirus RNA synthesis. J. Virol. 65: Novak, J. E., and K. Kirkegaard Improved method for detecting poliovirus negative strands used to demonstrate specificity of positivestrand encapsidation and the ratio of positive to negative strands in infected cells. J. Virol. 65: Tracy, S., K. M. Drescher, N. M. Chapman, K. S. Kim, S. D. Carson, S. Pirruccello, P. H. Lane, J. R. Romero, and J. S. Leser Toward testing the hypothesis that group B coxsackieviruses (CVB) trigger insulin-dependent diabetes: inoculating nonobese diabetic mice with CVB markedly lowers diabetes incidence. J. Virol. 76: Tu, Z., N. M. Chapman, G. Hufnagel, S. Tracy, J. R. Romero, W. H. Barry, L. Zhao, K. Currey, and B. Shapiro The cardiovirulent phenotype of coxsackievirus B3 is determined at a single site in the genomic 5' nontranslated region. J. Virol. 69: Chapman, N. M., Z. Tu, S. Tracy, and C. J. Gauntt An infectious cdna copy of the genome of a non-cardiovirulent coxsackievirus B3 strain: its complete sequence analysis and comparison to the genomes of cardiovirulent coxsackieviruses. Arch. Virol. 135: Gauntt, C. J., M. D. Trousdale, D. R. LaBadie, R. E. Paque, and T. Nealon Properties of coxsackievirus B3 variants which are amyocarditic or myocarditic for mice. J. Med. Virol. 3: Claycomb, W. C., N. A. Lanson, B. S. Stallworth, D. B. Egeland, J. B. Delcarpio, A. Bahinski, and N. J. Izzo HL-1 cells: a cardiac muscle cell line that contracts and retains phenotypic characteristics of the adult cardiomyocyte. Proc. Natl. Acad. Sci. U.S.A. 95: Walter, B. L., T. B. Parsley, E. Ehrenfeld, and B. L. Semler Distinct poly(rc) binding protein KH domain determinants for poliovirus translation initiation and viral RNA replication. J. Virol. 76: Lanke, K. H. W., H. M. van der Schaar, G. A. Belov, Q. Feng, D. Duijsings, C. L. Jackson, E. Ehrenfeld, and F. J. M. van Kuppeveld GBF1, a guanine nucleotide exchange factor for Arf, is crucial for coxsackievirus B3 RNA replication. J. Virol. 83: Blair, W. S., J. H. Nguyen, T. B. Parsley, and B. L. Semler Mutations in the poliovirus 3CD proteinase S1-specificity pocket affect substrate recognition and RNA binding. Virology 218: Pincus, S. E., D. C. Diamond, E. A. Emini, and E. Wimmer Guanidine-selected mutants of poliovirus: mapping of point mutations to polypeptide 2C. J. Virol. 57: Gamarnik, A. V., and R. Andino Interactions of viral protein 3CD and poly(rc) binding protein with the 5' untranslated region of the poliovirus genome. J. Virol. 74:
39 Pallansch, M. A., M. S. Oberste, and J. L. Whitton Enteroviruses: Polioviruses, coxsackieviruses, echoviruses, and newer enteroviruses, p , Fields Virology, Lippincott Williams & Wilkins ed, vol. vol.1. Knipe DM, Howley PM, Philadelphia. 50. Tracy, S., K. M. Drescher, and N. M. Chapman Enteroviruses and type 1 diabetes. Diabetes Metab. Res. Rev. 27: Chapman, N. M., and K. S. Kim Persistent coxsackievirus infection: enterovirus persistence in chronic myocarditis and dilated cardiomyopathy. Curr. Top. Microbiol. Immunol. 323: Douche-Aourik, F., W. Berlier, L. Féasson, T. Bourlet, R. Harrath, S. Omar, F. Grattard, C. Denis, and B. Pozzetto Detection of enterovirus in human skeletal muscle from patients with chronic inflammatory muscle disease or fibromyalgia and healthy subjects. J. Med. Virol. 71: Vandenberghe, N., N. Leveque, P. Corcia, V. Brunaud-Danel, E. Salort-Campana, G. Besson, C. Tranchant, P. Clavelou, F. Beaulieux, R. Ecochard, C. Vial, E. Broussolle, and B. Lina Cerebrospinal fluid detection of enterovirus genome in ALS: a study of 242 patients and 354 controls. Amyotroph. Lateral. Scler. 11: Muir, P., F. Nicholson, M. K. Sharief, E. J. Thompson, N. J. Cairns, P. Lantos, G. T. Spencer, H. J. Kaminski, and J. E. Banatvala Evidence for persistent enterovirus infection of the central nervous system in patients with previous paralytic poliomyelitis. Ann. N. Y. Acad. Sci. 753: Cunningham, L., N. E. Bowles, R. J. Lane, V. Dubowitz, and L. C. Archard Persistence of enteroviral RNA in chronic fatigue syndrome is associated with the abnormal production of equal amounts of positive and negative strands of enteroviral RNA. J. Gen. Virol. 71 ( Pt 6): Smithee, S., S. Tracy, and N. M. Chapman Mutational disruption of cis-acting replication element 2C in coxsackievirus B3 leads to 5'- terminal genomic deletions. J. Virol. 89: Hunziker, I. P., C. T. Cornell, and J. L. Whitton Deletions within the 5'UTR of coxsackievirus B3: consequences for virus translation and replication. Virology 360: Muslin, C., M. Joffret, I. Pelletier, B. Blondel, and F. Delpeyroux Evolution and emergence of enteroviruses through intra- and interspecies recombination: plasticity and phenotypic impact of modular genetic exchanges in the 5' untranslated region. PLoS Pathog. 11:e Holmblat, B., S. Jégouic, C. Muslin, B. Blondel, M.-L. Joffret, and F. Delpeyroux Nonhomologous recombination between defective poliovirus and coxsackievirus genomes suggests a new model of genetic plasticity for picornaviruses. MBio 5:e
40 Collis, P. S., B. J. O'Donnell, D. J. Barton, J. A. Rogers, and J. B. Flanegan Replication of poliovirus RNA and subgenomic RNA transcripts in transfected cells. J. Virol. 66: Holland, J. J., and L. P. Villarreal Persistent noncytocidal vesicular stomatitis virus infections mediated by defective T particles that suppress virion transcriptase. Proc. Natl. Acad. Sci. U. S. A. 71: McClure, M. A., J. J. Holland, and J. Perrault Generation of defective interfering particles in picornaviruses. Virology 100: Cole, C. N., D. Smoler, E. Wimmer, and D. Baltimore Defective interfering particles of poliovirus. I. Isolation and physical properties. J. Virol. 7: McLaren, L. C., and J. J. Holland Defective interfering particles from poliovirus vaccine and vaccine reference strains. Virology 60: Mukherjee, A., S. A. Morosky, E. Delorme-Axford, N. Dybdahl- Sissoko, M. S. Oberste, T. Wang, and C. B. Coyne The coxsackievirus B 3C protease cleaves MAVS and TRIF to attenuate host type I interferon and apoptotic signaling. PLoS Pathog. 7:e Wang, J. P., A. Cerny, D. R. Asher, E. A. Kurt-Jones, R. T. Bronson, and R. W. Finberg MDA5 and MAVS mediate type I interferon responses to coxsackie B virus. J. Virol. 84: Hardarson, H. S., J. S. Baker, Z. Yang, E. Purevjav, C.-H. Huang, L. Alexopoulou, N. Li, R. A. Flavell, N. E. Bowles, and J. G. Vallejo Toll-like receptor 3 is an essential component of the innate stress response in virus-induced cardiac injury. Am. J. Physiol. Heart Circ. Physiol. 292:H Wang, J. P., D. R. Asher, M. Chan, E. A. Kurt-Jones, and R. W. Finberg Cutting edge: Antibody-mediated TLR7-dependent recognition of viral RNA. J. Immunol. 178: Fuse, K., G. Chan, Y. Liu, P. Gudgeon, M. Husain, M. Chen, W.-C. Yeh, S. Akira, and P. P. Liu Myeloid differentiation factor-88 plays a crucial role in the pathogenesis of Coxsackievirus B3-induced myocarditis and influences type I interferon production. Circulation 112: Feng, Q., S. V. Hato, M. A. Langereis, J. Zoll, R. Virgen-Slane, A. Peisley, S. Hur, B. L. Semler, R. P. van Rij, and F. J. van Kuppeveld MDA5 detects the double-stranded RNA replicative form in picornavirus-infected cells. Cell Rep. 2: Althof, N., S. Harkins, C. C. Kemball, C. T. Flynn, M. Alirezaei, and J. L. Whitton In vivo ablation of type I interferon receptor from cardiomyocytes delays coxsackieviral clearance and accelerates myocardial disease. J. Virol. 88: Henke, A., S. Huber, A. Stelzner, and J. L. Whitton The role of CD8+ T lymphocytes in coxsackievirus B3-induced myocarditis. J. Virol. 69:
41 Chow, L. H., K. W. Beisel, and B. M. McManus Enteroviral infection of mice with severe combined immunodeficiency. Evidence for direct viral pathogenesis of myocardial injury. Lab. Invest. 66: Schoggins, J. W., and C. M. Rice Interferon-stimulated genes and their antiviral effector functions. Curr. Opin. Virol. 1: Hyde, J. L., C. L. Gardner, T. Kimura, J. P. White, G. Liu, D. W. Trobaugh, C. Huang, M. Tonelli, S. Paessler, K. Takeda, W. B. Klimstra, G. K. Amarasinghe, and M. S. Diamond A viral RNA structural element alters host recognition of nonself RNA. Science 343: Tuschall, D. M., E. Hiebert, and J. B. Flanegan Poliovirus RNAdependent RNA polymerase synthesizes full-length copies of poliovirion RNA, cellular mrna, and several plant virus RNAs in vitro. J. Virol. 44: Gamarnik, A. V., and R. Andino Switch from translation to RNA replication in a positive-stranded RNA virus. Genes Dev. 12: Ertel, K. J., J. E. Brunner, and B. L. Semler Mechanistic consequences of hnrnp C binding to both RNA termini of poliovirus negative-strand RNA intermediates. J. Virol. 84: Roehl, H. H., and B. L. Semler Poliovirus infection enhances the formation of two ribonucleoprotein complexes at the 3' end of viral negative-strand RNA. J. Virol. 69: Brunner, J. E., K. J. Ertel, J. M. Rozovics, and B. L. Semler Delayed kinetics of poliovirus RNA synthesis in a human cell line with reduced levels of hnrnp C proteins. Virology 400:
42 949 FIGURE LEGENDS Figure 1. Schematic representation of luciferase replicon and signal amplification mechanisms through transfected replicon replication. (A) Schematic representation of the replicon. The replicons consisted of the CVB3-28 genome (40), in which the region encoding the capsid proteins (P1) has been replaced with a Renilla luciferase reporter gene. Deletions from 7 to 49 nucleotides in length were inserted at the 5 'end of the viral genome. The CVB3-0 replicon was generated from CVB3-28 replicon by site directed mutagenesis of nucleotide 234 (T in 28 and C in 0). (B) Replicons are positivestrand RNAs in which the P1 (capsid coding) region has been replaced with Renilla luciferase coding gene. Following transfection, replicons are directly translated into viral polyprotein including Renilla luciferase (a) or, following initial rounds of translation, used as a template for negative-strand RNA synthesis (b). Negative-strand RNAs will in turn serve as templates for positive-strand RNA synthesis (c) whose translation (d) will lead to luciferase signal amplification Figure 2. CVB3 replicon assays in two types of cardiomyocytes using wild-type (full-length) and terminally-deleted (TD) replicon RNAs. Luciferase replicons were transfected into primary human cardiomyocytes (HCM) (A) or in a continuous cell line of murine cardiomyocytes from an atrial tumor (HL-1 cells) (B). Luciferase activity was measured from 2 to 8 hours post- 42
43 transfection. The data presented here are the results of three independent experiments Figure 3. Impact of guanidine hydrochloride (GuHCl) treatment on fulllength and terminally deleted luciferase replicon kinetics. Full-length (A and B) and deleted (C and D) luciferase replicon transcripts described in the legend to Figure 2 were transfected into human (HCM) (A and C) and murine (HL-1) (B and D) cardiac myocyte monolayers in the presence or absence of GuHCl (an inhibitor of enterovirus RNA replication) at a final concentration of 3 mm. At 2, 4, 6 and 8 h post-transfection, cells were harvested and luciferase activity measured. The y axis shows log 10 units of luciferase activity, and the x axis shows time (in hours) post-transfection. Luciferase units in the presence and absence of GuHCl are shown in each graph. Although not readily visible, standard deviations (as error bars) are given for each data point. The results presented here are the product of three independent experiments Figure 4. In vitro translation assay of wild-type and terminally deleted (TD) coxsackievirus B3 RNAs. Either 0.4 µg or 0.6 µg of viral RNA transcript was incubated for 6 h at 30 C in HeLa S10 cytoplasmic extract in the presence of [ 35 S]methionine. Labeled viral proteins (indicated to the right of the gel) were analyzed by SDS-PAGE and visualized by fluorography. This figure shows representative data from four independent experiments. The first lane of the figure (-) is a translation reaction incubated in the absence of transcript RNA. To 43
44 determine the relative levels of viral proteins synthesized by each RNA, signals detected on the gel for viral proteins 3CD, P2, VP1, VP3, 2A, and 2AB (collectively) were quantified using ImageJ software ( The second lane (WT-0, 0.4 µg RNA template) was set as the 100% reference control. Transcripts produced from the WT-0 construct do not have a hammerhead ribozyme at their 5 termini, while the transcripts from the WT-28, TD7, TD21, TD30, and TD49 constructs each contain a hammerhead ribozyme at their 5 termini Figure 5. In vitro RNA replication of wild-type and terminally deleted (TD) coxsackievirus B3 transcripts. After a 4.5 h in vitro translation assay allowing the synthesis of non-structural proteins in the absence of [ 35 S]methionine, the reaction mixture consisting of viral transcripts (2 µg or 3 µg) and HeLa S10 cytoplasmic extract was incubated for an additional 2 h at 34 C for RNA synthesis in the presence of α-[ 32 P]CTP. The purified RNA was then resolved on a 1.1% agarose gel in Tris-borate-EDTA buffer containing ethidium bromide. The amounts of 18S and 28S rrna in each lane were used to confirm equal loading of samples in the gel. Plasmid DNA template encoding CVB3-28 RNA harbors a hammerhead ribozyme at the 5 end of the transcript (32), producing an authentic CVB3 5 terminus. This RNA is an efficient template for negativestrand RNA synthesis and subsequent rounds of positive-strand RNA synthesis. In vitro RNAs produced from pcvb3-0 have no hammerhead ribozyme at the 5 end and harbor non-viral sequences that result in efficient negative-strand RNA 44
45 synthesis but very inefficient positive-strand RNA (ssrna) synthesis (13, 35). All TD transcripts harbor a hammerhead ribozyme at their 5 termini, producing authentic 5 terminal CVB3 sequences. The signals detected in the gel for single-stranded (ss) and double-stranded (ds) viral RNAs were quantified using ImageJ software. The signals were normalized to the 28S and 18S ribosomal RNA levels (collectively) for each sample and expressed in arbitrary units (A.U.). Lane 1 (-) shows the results of incubation of the in vitro replication reaction in the absence of transcript RNA Figure 6. Schematic representation of cellular and viral proteins binding to the cloverleaf (stem-loop I) of positive-strand viral RNAs. Stem-loop I RNA is organized into stem a and stem-loops b, c and d. Cellular poly(rc)-binding protein 2 (PCBP2) and viral protein 3CD pro bind to the cloverleaf at the 5 end of positive-strand RNA to form a ribonucleoprotein complex required for the initiation of both negative-strand and positive-strand RNA synthesis Figure 7. Electrophoretic mobility shift assays with CVB3 5 probes. Radiolabeled transcripts that included stem-loop I (approx. 175 to 225 nucleotides in length) corresponding to the 5 end of coxsackievirus B3 positivestrand RNA were used in electrophoretic mobility shift assays to determine the ability of the 5 terminally deleted RNAs (TD7, TD21, TD30 and TD49) to form RNP complexes with PCBP2 and 3CD pro, alone or in combination. Full-length (wild-type) S-L I-containing RNA from CVB3 28 was used as a positive control. 45
46 (A) [ 32 P]UTP radiolabeled CVB3 S-L I-containing RNA was incubated alone (lanes 1, 5, 10, 13 and 17) or with increasing concentrations of recombinant purified PCBP2 ranging from 2.5 µm (lanes 2, 6, 10, 14 and 18) to 5 µm (lanes 3, 7, 11, 15 and 19) and 7.5 µm (lanes 4, 8, 12, 16 and 20). Note that samples from WT, TD7, and TD21 RNP complexes were analyzed on a separate gel from the one used to analyze TD30 and TD49, indicated by the white space in between these two sets of lanes. (B) [ 32 P]UTP radiolabeled CVB3 S-L I- containing RNA was incubated without (lanes 1, 8, 15, 22 and 29) or with increasing concentrations of a proteolytically active form of recombinant purified 1042 CVB3 3CD pro that is incapable of self-cleavage at the 3C-3D junction [3CD(µ10)] ranging from 0.05 µm (lanes 2, 9, 16, 23 and 30) to 1 µm (lanes 3, 10, 17, 24 and 31), 2.5 µm (lanes 4, 11, 18, 25 and 32), 5 µm (lanes 5, 12, 19, 26 and 33), 7.5 µm (lanes 6, 13, 20, 27 and 34) and 10 µm (lanes 6, 14, 21, 28 and 25). Note that samples from WT and TD7 RNP complexes, TD21 RNP complexes, TD30 RNP complexes, and TD49 RNP complexes were analyzed on separate gels, indicated by the black vertical line in between these four sets of lanes. (C) To analyze super-shifted ternary complex formation, increasing amounts of 3CD pro ranging from 0 (lanes 2, 6, 10, 14 and 18), to 2.5 µm (lanes 3, 7, 11, 15 and 19) and 5 µm (lanes 4, 8, 12, 16 and 20) were added to a fixed concentration of PCBP2 (5 µm). Lanes 1, 5, 9, 13 and 17 included S-L I- containing probes alone. Note that samples from WT, TD7, and TD21 RNP complexes were analyzed on a separate gel from the one used to analyze TD30 and TD49, indicated by the white space in between these two sets of 46
47 lanes. Following incubation, ribonucleoprotein (RNP) complexes were resolved on a native 4% polyacrylamide gel. The arrows and brackets to the right of the gel image indicate probe alone (free probe), RNP complex formation, and ternary complex formation. The results shown here are representative of three independent sets of experiments. The signals detected on polyacrylamide gels for RNP complex formation were quantified using ImageJ software. The values (in A.U., arbitrary units) for panel A and C were calculated by dividing the value of the complex signal by the value of free probes in the RNA only lanes 1064 incubated without proteins. Final amounts (in arbitrary units, A.U.) were obtained by subtracting the RNA only values from the calculated amounts. For panel B, the values represent the signal of free probe subtracted from the value of free probe in the RNA only lane (arbitrarily set at 1). 47
48
49
50
51
52
53
54
55
56
Replication Defective Enterovirus Infections: Implications for Type I Diabetes
 Replication Defective Enterovirus Infections: Implications for Type I Diabetes N. M. Chapman Department of Pathology & Microbiology University of Nebraska Medical Center Enterovirus Genome and 2 Capsid
Replication Defective Enterovirus Infections: Implications for Type I Diabetes N. M. Chapman Department of Pathology & Microbiology University of Nebraska Medical Center Enterovirus Genome and 2 Capsid
7.012 Quiz 3 Answers
 MIT Biology Department 7.012: Introductory Biology - Fall 2004 Instructors: Professor Eric Lander, Professor Robert A. Weinberg, Dr. Claudette Gardel Friday 11/12/04 7.012 Quiz 3 Answers A > 85 B 72-84
MIT Biology Department 7.012: Introductory Biology - Fall 2004 Instructors: Professor Eric Lander, Professor Robert A. Weinberg, Dr. Claudette Gardel Friday 11/12/04 7.012 Quiz 3 Answers A > 85 B 72-84
Translation. Host Cell Shutoff 1) Initiation of eukaryotic translation involves many initiation factors
 Initiation of eukaryotic translation involves many initiation factors") Translation Questions? 1) How does poliovirus shutoff eukaryotic translation? 2) If eukaryotic messages are not translated how can poliovirus get its message translated? Host Cell Shutoff 1) Initiation
Translation Questions? 1) How does poliovirus shutoff eukaryotic translation? 2) If eukaryotic messages are not translated how can poliovirus get its message translated? Host Cell Shutoff 1) Initiation
Picornaviruses. Virion. Genome. Genes and proteins. Viruses and hosts. Diseases. Distinctive characteristics
 Picornaviruses Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Virion Naked icosahedral capsid (T=1) Diameter of 30 nm Genome Linear single-stranded RNA, positive
Picornaviruses Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Virion Naked icosahedral capsid (T=1) Diameter of 30 nm Genome Linear single-stranded RNA, positive
Lecture 2: Virology. I. Background
 Lecture 2: Virology I. Background A. Properties 1. Simple biological systems a. Aggregates of nucleic acids and protein 2. Non-living a. Cannot reproduce or carry out metabolic activities outside of a
Lecture 2: Virology I. Background A. Properties 1. Simple biological systems a. Aggregates of nucleic acids and protein 2. Non-living a. Cannot reproduce or carry out metabolic activities outside of a
Polyomaviridae. Spring
 Polyomaviridae Spring 2002 331 Antibody Prevalence for BK & JC Viruses Spring 2002 332 Polyoma Viruses General characteristics Papovaviridae: PA - papilloma; PO - polyoma; VA - vacuolating agent a. 45nm
Polyomaviridae Spring 2002 331 Antibody Prevalence for BK & JC Viruses Spring 2002 332 Polyoma Viruses General characteristics Papovaviridae: PA - papilloma; PO - polyoma; VA - vacuolating agent a. 45nm
Virology Journal. Open Access. Abstract. BioMed Central
 Virology Journal BioMed Central Research Stimulation of poliovirus RNA synthesis and virus maturation in a HeLa cell-free in vitro translation-rna replication system by viral protein 3CD pro David Franco
Virology Journal BioMed Central Research Stimulation of poliovirus RNA synthesis and virus maturation in a HeLa cell-free in vitro translation-rna replication system by viral protein 3CD pro David Franco
Department of Microbiology 1 and Program in Molecular Biology, 2 School of Medicine, University of Colorado Denver, Aurora, Colorado
 JOURNAL OF VIROLOGY, Mar. 2010, p. 2843 2858 Vol. 84, No. 6 0022-538X/10/$12.00 doi:10.1128/jvi.02620-08 Copyright 2010, American Society for Microbiology. All Rights Reserved. Poly(A) at the 3 End of
JOURNAL OF VIROLOGY, Mar. 2010, p. 2843 2858 Vol. 84, No. 6 0022-538X/10/$12.00 doi:10.1128/jvi.02620-08 Copyright 2010, American Society for Microbiology. All Rights Reserved. Poly(A) at the 3 End of
Poliovirus CRE-Dependent VPg Uridylylation Is Required for Positive-Strand RNA Synthesis but Not for Negative-Strand RNA Synthesis
 REFERENCES CONTENT ALERTS Poliovirus CRE-Dependent VPg Uridylylation Is Required for Positive-Strand RNA Synthesis but Not for Negative-Strand RNA Synthesis Kenneth E. Murray and David J. Barton J. Virol.
REFERENCES CONTENT ALERTS Poliovirus CRE-Dependent VPg Uridylylation Is Required for Positive-Strand RNA Synthesis but Not for Negative-Strand RNA Synthesis Kenneth E. Murray and David J. Barton J. Virol.
Islamic University Faculty of Medicine
 Islamic University Faculty of Medicine 2012 2013 2 RNA is a modular structure built from a combination of secondary and tertiary structural motifs. RNA chains fold into unique 3 D structures, which act
Islamic University Faculty of Medicine 2012 2013 2 RNA is a modular structure built from a combination of secondary and tertiary structural motifs. RNA chains fold into unique 3 D structures, which act
Reoviruses. Virion. Genome. Genes and proteins. Viruses and hosts. Diseases. Distinctive characteristics
 Reoviruses Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Virion Naked icosahedral capsid (T=13), diameter 60-85 nm Capsid consists of two or three concentric protein
Reoviruses Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Virion Naked icosahedral capsid (T=13), diameter 60-85 nm Capsid consists of two or three concentric protein
Chapter 6- An Introduction to Viruses*
 Chapter 6- An Introduction to Viruses* *Lecture notes are to be used as a study guide only and do not represent the comprehensive information you will need to know for the exams. 6.1 Overview of Viruses
Chapter 6- An Introduction to Viruses* *Lecture notes are to be used as a study guide only and do not represent the comprehensive information you will need to know for the exams. 6.1 Overview of Viruses
Structural vs. nonstructural proteins
 Why would you want to study proteins associated with viruses or virus infection? Receptors Mechanism of uncoating How is gene expression carried out, exclusively by viral enzymes? Gene expression phases?
Why would you want to study proteins associated with viruses or virus infection? Receptors Mechanism of uncoating How is gene expression carried out, exclusively by viral enzymes? Gene expression phases?
HEK293FT cells were transiently transfected with reporters, N3-ICD construct and
 Supplementary Information Luciferase reporter assay HEK293FT cells were transiently transfected with reporters, N3-ICD construct and increased amounts of wild type or kinase inactive EGFR. Transfections
Supplementary Information Luciferase reporter assay HEK293FT cells were transiently transfected with reporters, N3-ICD construct and increased amounts of wild type or kinase inactive EGFR. Transfections
Materials and Methods , The two-hybrid principle.
 The enzymatic activity of an unknown protein which cleaves the phosphodiester bond between the tyrosine residue of a viral protein and the 5 terminus of the picornavirus RNA Introduction Every day there
The enzymatic activity of an unknown protein which cleaves the phosphodiester bond between the tyrosine residue of a viral protein and the 5 terminus of the picornavirus RNA Introduction Every day there
Coronaviruses. Virion. Genome. Genes and proteins. Viruses and hosts. Diseases. Distinctive characteristics
 Coronaviruses Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Virion Spherical enveloped particles studded with clubbed spikes Diameter 120-160 nm Coiled helical
Coronaviruses Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Virion Spherical enveloped particles studded with clubbed spikes Diameter 120-160 nm Coiled helical
number Done by Corrected by Doctor Ashraf
 number 4 Done by Nedaa Bani Ata Corrected by Rama Nada Doctor Ashraf Genome replication and gene expression Remember the steps of viral replication from the last lecture: Attachment, Adsorption, Penetration,
number 4 Done by Nedaa Bani Ata Corrected by Rama Nada Doctor Ashraf Genome replication and gene expression Remember the steps of viral replication from the last lecture: Attachment, Adsorption, Penetration,
Analysis of small RNAs from Drosophila Schneider cells using the Small RNA assay on the Agilent 2100 bioanalyzer. Application Note
 Analysis of small RNAs from Drosophila Schneider cells using the Small RNA assay on the Agilent 2100 bioanalyzer Application Note Odile Sismeiro, Jean-Yves Coppée, Christophe Antoniewski, and Hélène Thomassin
Analysis of small RNAs from Drosophila Schneider cells using the Small RNA assay on the Agilent 2100 bioanalyzer Application Note Odile Sismeiro, Jean-Yves Coppée, Christophe Antoniewski, and Hélène Thomassin
A Novel in Vitro Replication System for Dengue Virus
 THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 274, No. 47, Issue of November 19, pp. 33714 33722, 1999 1999 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. A Novel in
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 274, No. 47, Issue of November 19, pp. 33714 33722, 1999 1999 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. A Novel in
Life Sciences 1A Midterm Exam 2. November 13, 2006
 Name: TF: Section Time Life Sciences 1A Midterm Exam 2 November 13, 2006 Please write legibly in the space provided below each question. You may not use calculators on this exam. We prefer that you use
Name: TF: Section Time Life Sciences 1A Midterm Exam 2 November 13, 2006 Please write legibly in the space provided below each question. You may not use calculators on this exam. We prefer that you use
Product Manual. Omni-Array Sense Strand mrna Amplification Kit, 2 ng to 100 ng Version Catalog No.: Reactions
 Genetic Tools and Reagents Universal mrna amplification, sense strand amplification, antisense amplification, cdna synthesis, micro arrays, gene expression, human, mouse, rat, guinea pig, cloning Omni-Array
Genetic Tools and Reagents Universal mrna amplification, sense strand amplification, antisense amplification, cdna synthesis, micro arrays, gene expression, human, mouse, rat, guinea pig, cloning Omni-Array
Supplementary Figure 1. SC35M polymerase activity in the presence of Bat or SC35M NP encoded from the phw2000 rescue plasmid.
 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 Supplementary Figure 1. SC35M polymerase activity in the presence of Bat or SC35M NP encoded from the phw2000 rescue plasmid. HEK293T
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 Supplementary Figure 1. SC35M polymerase activity in the presence of Bat or SC35M NP encoded from the phw2000 rescue plasmid. HEK293T
Poliovirus Requires a Precise 5 End for Efficient Positive- Strand RNA Synthesis
 JOURNAL OF VIROLOGY, July 2000, p. 6394 6400 Vol. 74, No. 14 0022-538X/00/$04.00 0 Copyright 2000, American Society for Microbiology. All Rights Reserved. Poliovirus Requires a Precise 5 End for Efficient
JOURNAL OF VIROLOGY, July 2000, p. 6394 6400 Vol. 74, No. 14 0022-538X/00/$04.00 0 Copyright 2000, American Society for Microbiology. All Rights Reserved. Poliovirus Requires a Precise 5 End for Efficient
AP Biology Reading Guide. Concept 19.1 A virus consists of a nucleic acid surrounded by a protein coat
 AP Biology Reading Guide Name Chapter 19: Viruses Overview Experimental work with viruses has provided important evidence that genes are made of nucleic acids. Viruses were also important in working out
AP Biology Reading Guide Name Chapter 19: Viruses Overview Experimental work with viruses has provided important evidence that genes are made of nucleic acids. Viruses were also important in working out
Intrinsic cellular defenses against virus infection
 Intrinsic cellular defenses against virus infection Detection of virus infection Host cell response to virus infection Interferons: structure and synthesis Induction of antiviral activity Viral defenses
Intrinsic cellular defenses against virus infection Detection of virus infection Host cell response to virus infection Interferons: structure and synthesis Induction of antiviral activity Viral defenses
Effects of Mutations of the Initiation Nucleotides on Hepatitis C Virus RNA Replication in the Cell
 JOURNAL OF VIROLOGY, Apr. 2004, p. 3633 3643 Vol. 78, No. 7 0022-538X/04/$08.00 0 DOI: 10.1128/JVI.78.7.3633 3643.2004 Copyright 2004, American Society for Microbiology. All Rights Reserved. Effects of
JOURNAL OF VIROLOGY, Apr. 2004, p. 3633 3643 Vol. 78, No. 7 0022-538X/04/$08.00 0 DOI: 10.1128/JVI.78.7.3633 3643.2004 Copyright 2004, American Society for Microbiology. All Rights Reserved. Effects of
Hepatitis B Antiviral Drug Development Multi-Marker Screening Assay
 Hepatitis B Antiviral Drug Development Multi-Marker Screening Assay Background ImQuest BioSciences has developed and qualified a single-plate method to expedite the screening of antiviral agents against
Hepatitis B Antiviral Drug Development Multi-Marker Screening Assay Background ImQuest BioSciences has developed and qualified a single-plate method to expedite the screening of antiviral agents against
numbe r Done by Corrected by Doctor
 numbe r 5 Done by Mustafa Khader Corrected by Mahdi Sharawi Doctor Ashraf Khasawneh Viral Replication Mechanisms: (Protein Synthesis) 1. Monocistronic Method: All human cells practice the monocistronic
numbe r 5 Done by Mustafa Khader Corrected by Mahdi Sharawi Doctor Ashraf Khasawneh Viral Replication Mechanisms: (Protein Synthesis) 1. Monocistronic Method: All human cells practice the monocistronic
Overview of the Expressway Cell-Free Expression Systems. Expressway Mini Cell-Free Expression System
 Overview of the Expressway Cell-Free Expression Systems The Expressway Cell-Free Expression Systems use an efficient coupled transcription and translation reaction to produce up to milligram quantities
Overview of the Expressway Cell-Free Expression Systems The Expressway Cell-Free Expression Systems use an efficient coupled transcription and translation reaction to produce up to milligram quantities
Aseptic meningitis: inflammation of meninges with sterile CSF (without any causative organisms which can be grown on culture media).
.") You have to refer to the slides, since I have included the extra information only. Slide #1: Both illnesses aseptic meningitis and encephalitis can be caused by the same viruses; that viruses which cause
You have to refer to the slides, since I have included the extra information only. Slide #1: Both illnesses aseptic meningitis and encephalitis can be caused by the same viruses; that viruses which cause
New Small and Versatile Reporter Technologies for Challenging Applications in Virology. Robert Brazas, Ph.D.
 New Small and Versatile Reporter Technologies for Challenging Applications in Virology Robert Brazas, Ph.D. Presentation Overview Small size of NanoLuc Luciferase is advantageous Agenda Building a better
New Small and Versatile Reporter Technologies for Challenging Applications in Virology Robert Brazas, Ph.D. Presentation Overview Small size of NanoLuc Luciferase is advantageous Agenda Building a better
Molecular Biology (BIOL 4320) Exam #2 May 3, 2004
 Exam #2 May 3, 2004") Molecular Biology (BIOL 4320) Exam #2 May 3, 2004 Name SS# This exam is worth a total of 100 points. The number of points each question is worth is shown in parentheses after the question number. Good
Molecular Biology (BIOL 4320) Exam #2 May 3, 2004 Name SS# This exam is worth a total of 100 points. The number of points each question is worth is shown in parentheses after the question number. Good
Last time we talked about the few steps in viral replication cycle and the un-coating stage:
 Zeina Al-Momani Last time we talked about the few steps in viral replication cycle and the un-coating stage: Un-coating: is a general term for the events which occur after penetration, we talked about
Zeina Al-Momani Last time we talked about the few steps in viral replication cycle and the un-coating stage: Un-coating: is a general term for the events which occur after penetration, we talked about
Genetic Complementation among Poliovirus Mutants Derived
 JOURNAL OF VIROLOGY, Dec. 1986, p. 1040-1049 0022-538X/86/121040-10$02.00/0 Copyright C) 1986, American Society for Microbiology Vol. 60, No. 3 Genetic Complementation among Poliovirus Mutants Derived
JOURNAL OF VIROLOGY, Dec. 1986, p. 1040-1049 0022-538X/86/121040-10$02.00/0 Copyright C) 1986, American Society for Microbiology Vol. 60, No. 3 Genetic Complementation among Poliovirus Mutants Derived
Introduction retroposon
 17.1 - Introduction A retrovirus is an RNA virus able to convert its sequence into DNA by reverse transcription A retroposon (retrotransposon) is a transposon that mobilizes via an RNA form; the DNA element
17.1 - Introduction A retrovirus is an RNA virus able to convert its sequence into DNA by reverse transcription A retroposon (retrotransposon) is a transposon that mobilizes via an RNA form; the DNA element
Determination of the temporal pattern and importance of BALF1 expression in Epstein-Barr viral infection
 Determination of the temporal pattern and importance of BALF1 expression in Epstein-Barr viral infection Melissa Mihelidakis May 6, 2004 7.340 Research Proposal Introduction Apoptosis, or programmed cell
Determination of the temporal pattern and importance of BALF1 expression in Epstein-Barr viral infection Melissa Mihelidakis May 6, 2004 7.340 Research Proposal Introduction Apoptosis, or programmed cell
7.012 Problem Set 6 Solutions
 Name Section 7.012 Problem Set 6 Solutions Question 1 The viral family Orthomyxoviridae contains the influenza A, B and C viruses. These viruses have a (-)ss RNA genome surrounded by a capsid composed
Name Section 7.012 Problem Set 6 Solutions Question 1 The viral family Orthomyxoviridae contains the influenza A, B and C viruses. These viruses have a (-)ss RNA genome surrounded by a capsid composed
Chapter 18. Viral Genetics. AP Biology
 Chapter 18. Viral Genetics 2003-2004 1 A sense of size Comparing eukaryote bacterium virus 2 What is a virus? Is it alive? DNA or RNA enclosed in a protein coat Viruses are not cells Extremely tiny electron
Chapter 18. Viral Genetics 2003-2004 1 A sense of size Comparing eukaryote bacterium virus 2 What is a virus? Is it alive? DNA or RNA enclosed in a protein coat Viruses are not cells Extremely tiny electron
Native Replication Intermediates of the Yeast 20 S RNA Virus Have a Single-stranded RNA Backbone*
 THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 280, No. 8, Issue of February 25, pp. 7398 7406, 2005 2005 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. Native Replication
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 280, No. 8, Issue of February 25, pp. 7398 7406, 2005 2005 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. Native Replication
Received 3 September 2002/Accepted 15 January 2003
 JOURNAL OF VIROLOGY, Apr. 2003, p. 4646 4657 Vol. 77, No. 8 0022-538X/03/$08.00 0 DOI: 10.1128/JVI.77.8.4646 4657.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved. Ability of
JOURNAL OF VIROLOGY, Apr. 2003, p. 4646 4657 Vol. 77, No. 8 0022-538X/03/$08.00 0 DOI: 10.1128/JVI.77.8.4646 4657.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved. Ability of
October 26, Lecture Readings. Vesicular Trafficking, Secretory Pathway, HIV Assembly and Exit from Cell
 October 26, 2006 Vesicular Trafficking, Secretory Pathway, HIV Assembly and Exit from Cell 1. Secretory pathway a. Formation of coated vesicles b. SNAREs and vesicle targeting 2. Membrane fusion a. SNAREs
October 26, 2006 Vesicular Trafficking, Secretory Pathway, HIV Assembly and Exit from Cell 1. Secretory pathway a. Formation of coated vesicles b. SNAREs and vesicle targeting 2. Membrane fusion a. SNAREs
Influenza viruses. Virion. Genome. Genes and proteins. Viruses and hosts. Diseases. Distinctive characteristics
 Influenza viruses Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Virion Enveloped particles, quasi-spherical or filamentous Diameter 80-120 nm Envelope is derived
Influenza viruses Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Virion Enveloped particles, quasi-spherical or filamentous Diameter 80-120 nm Envelope is derived
Supplementary Fig. S1. Schematic diagram of minigenome segments.
 open reading frame 1565 (segment 5) 47 (-) 3 5 (+) 76 101 125 149 173 197 221 246 287 open reading frame 890 (segment 8) 60 (-) 3 5 (+) 172 Supplementary Fig. S1. Schematic diagram of minigenome segments.
open reading frame 1565 (segment 5) 47 (-) 3 5 (+) 76 101 125 149 173 197 221 246 287 open reading frame 890 (segment 8) 60 (-) 3 5 (+) 172 Supplementary Fig. S1. Schematic diagram of minigenome segments.
The reovirus genome comprises 10 segments of doublestranded
 Reovirus reverse genetics: Incorporation of the CAT gene into the reovirus genome Michael R. Roner* and Wolfgang K. Joklik *Department of Biological Sciences, Center for Molecular Biology and Biotechnology,
Reovirus reverse genetics: Incorporation of the CAT gene into the reovirus genome Michael R. Roner* and Wolfgang K. Joklik *Department of Biological Sciences, Center for Molecular Biology and Biotechnology,
otherwise known as Cytotoxic T lymphocytes (CTLs)
") MIT Biology Department 7.012: Introductory Biology - Fall 200 Instructors: Professor Eric Lander, Professor Robert A. Weinberg, Dr. Claudette Gardel NAME TA SEC 7.012 Problem Set 5 FRIDAY October 29, 2004
MIT Biology Department 7.012: Introductory Biology - Fall 200 Instructors: Professor Eric Lander, Professor Robert A. Weinberg, Dr. Claudette Gardel NAME TA SEC 7.012 Problem Set 5 FRIDAY October 29, 2004
Translating Ribosomes Inhibit Poliovirus Negative-Strand RNA Synthesis
 JOURNAL OF VIROLOGY, Dec. 1999, p. 10104 10112 Vol. 73, No. 12 0022-538X/99/$04.00 0 Copyright 1999, American Society for Microbiology. All Rights Reserved. Translating Ribosomes Inhibit Poliovirus Negative-Strand
JOURNAL OF VIROLOGY, Dec. 1999, p. 10104 10112 Vol. 73, No. 12 0022-538X/99/$04.00 0 Copyright 1999, American Society for Microbiology. All Rights Reserved. Translating Ribosomes Inhibit Poliovirus Negative-Strand
Fayth K. Yoshimura, Ph.D. September 7, of 7 HIV - BASIC PROPERTIES
 1 of 7 I. Viral Origin. A. Retrovirus - animal lentiviruses. HIV - BASIC PROPERTIES 1. HIV is a member of the Retrovirus family and more specifically it is a member of the Lentivirus genus of this family.
1 of 7 I. Viral Origin. A. Retrovirus - animal lentiviruses. HIV - BASIC PROPERTIES 1. HIV is a member of the Retrovirus family and more specifically it is a member of the Lentivirus genus of this family.
Nature Medicine: doi: /nm.4322
 1 2 3 4 5 6 7 8 9 10 11 Supplementary Figure 1. Predicted RNA structure of 3 UTR and sequence alignment of deleted nucleotides. (a) Predicted RNA secondary structure of ZIKV 3 UTR. The stem-loop structure
1 2 3 4 5 6 7 8 9 10 11 Supplementary Figure 1. Predicted RNA structure of 3 UTR and sequence alignment of deleted nucleotides. (a) Predicted RNA secondary structure of ZIKV 3 UTR. The stem-loop structure
Viral structure م.م رنا مشعل
 Viral structure م.م رنا مشعل Viruses must reproduce (replicate) within cells, because they cannot generate energy or synthesize proteins. Because they can reproduce only within cells, viruses are obligate
Viral structure م.م رنا مشعل Viruses must reproduce (replicate) within cells, because they cannot generate energy or synthesize proteins. Because they can reproduce only within cells, viruses are obligate
Viruses defined acellular organisms genomes nucleic acid replicate inside host cells host metabolic machinery ribosomes
 The Viruses Viruses Viruses may be defined as acellular organisms whose genomes consist of nucleic acid, obligately replicate inside host cells using host metabolic machinery and ribosomes to form a pool
The Viruses Viruses Viruses may be defined as acellular organisms whose genomes consist of nucleic acid, obligately replicate inside host cells using host metabolic machinery and ribosomes to form a pool
Overview: Chapter 19 Viruses: A Borrowed Life
 Overview: Chapter 19 Viruses: A Borrowed Life Viruses called bacteriophages can infect and set in motion a genetic takeover of bacteria, such as Escherichia coli Viruses lead a kind of borrowed life between
Overview: Chapter 19 Viruses: A Borrowed Life Viruses called bacteriophages can infect and set in motion a genetic takeover of bacteria, such as Escherichia coli Viruses lead a kind of borrowed life between
Transcription and RNA processing
 Transcription and RNA processing Lecture 7 Biology 3310/4310 Virology Spring 2018 It is possible that Nature invented DNA for the purpose of achieving regulation at the transcriptional rather than at the
Transcription and RNA processing Lecture 7 Biology 3310/4310 Virology Spring 2018 It is possible that Nature invented DNA for the purpose of achieving regulation at the transcriptional rather than at the
1. Identify and characterize interesting phenomena! 2. Characterization should stimulate some questions/models! 3. Combine biochemistry and genetics
 1. Identify and characterize interesting phenomena! 2. Characterization should stimulate some questions/models! 3. Combine biochemistry and genetics to gain mechanistic insight! 4. Return to step 2, as
1. Identify and characterize interesting phenomena! 2. Characterization should stimulate some questions/models! 3. Combine biochemistry and genetics to gain mechanistic insight! 4. Return to step 2, as
Different De Novo Initiation Strategies Are Used by Influenza Virus RNA Polymerase on Its crna and Viral RNA Promoters during Viral RNA Replication
 JOURNAL OF VIROLOGY, Mar. 2006, p. 2337 2348 Vol. 80, No. 5 0022-538X/06/$08.00 0 doi:10.1128/jvi.80.5.2337 2348.2006 Copyright 2006, American Society for Microbiology. All Rights Reserved. Different De
JOURNAL OF VIROLOGY, Mar. 2006, p. 2337 2348 Vol. 80, No. 5 0022-538X/06/$08.00 0 doi:10.1128/jvi.80.5.2337 2348.2006 Copyright 2006, American Society for Microbiology. All Rights Reserved. Different De
CHAPTER 4 RESULTS. showed that all three replicates had similar growth trends (Figure 4.1) (p<0.05; p=0.0000)
 (p<0.05; p=0.0000)") CHAPTER 4 RESULTS 4.1 Growth Characterization of C. vulgaris 4.1.1 Optical Density Growth study of Chlorella vulgaris based on optical density at 620 nm (OD 620 ) showed that all three replicates had similar
CHAPTER 4 RESULTS 4.1 Growth Characterization of C. vulgaris 4.1.1 Optical Density Growth study of Chlorella vulgaris based on optical density at 620 nm (OD 620 ) showed that all three replicates had similar
Sequences in the 5" Non-coding Region of Human Rhinovims 14 RNA that Affect in vitro Translation
 J. gen. Virol. (1989), 70, 2799-2804. Printed in Great Britain 2799 Key words: rhinovirus, human type 14/5' non-coding region~in vitro translation Sequences in the 5" Non-coding Region of Human Rhinovims
J. gen. Virol. (1989), 70, 2799-2804. Printed in Great Britain 2799 Key words: rhinovirus, human type 14/5' non-coding region~in vitro translation Sequences in the 5" Non-coding Region of Human Rhinovims
Fayth K. Yoshimura, Ph.D. September 7, of 7 RETROVIRUSES. 2. HTLV-II causes hairy T-cell leukemia
 1 of 7 I. Diseases Caused by Retroviruses RETROVIRUSES A. Human retroviruses that cause cancers 1. HTLV-I causes adult T-cell leukemia and tropical spastic paraparesis 2. HTLV-II causes hairy T-cell leukemia
1 of 7 I. Diseases Caused by Retroviruses RETROVIRUSES A. Human retroviruses that cause cancers 1. HTLV-I causes adult T-cell leukemia and tropical spastic paraparesis 2. HTLV-II causes hairy T-cell leukemia
VIRUSES. Biology Applications Control. David R. Harper. Garland Science Taylor & Francis Group NEW YORK AND LONDON
 VIRUSES Biology Applications Control David R. Harper GS Garland Science Taylor & Francis Group NEW YORK AND LONDON vii Chapter 1 Virus Structure and 2.2 VIRUS MORPHOLOGY 26 Infection 1 2.3 VIRAL CLASSIFICATION
VIRUSES Biology Applications Control David R. Harper GS Garland Science Taylor & Francis Group NEW YORK AND LONDON vii Chapter 1 Virus Structure and 2.2 VIRUS MORPHOLOGY 26 Infection 1 2.3 VIRAL CLASSIFICATION
Hepatitis B Virus infection: virology
 Hepatitis B Virus infection: virology 167 Falk Symposium: Liver under constant attack from fat to viruses III Falk Gastro-Konferenz 17.-21. September 2008 Congress Centrum Mainz Maura Dandri Department
Hepatitis B Virus infection: virology 167 Falk Symposium: Liver under constant attack from fat to viruses III Falk Gastro-Konferenz 17.-21. September 2008 Congress Centrum Mainz Maura Dandri Department
Chapter13 Characterizing and Classifying Viruses, Viroids, and Prions
 Chapter13 Characterizing and Classifying Viruses, Viroids, and Prions 11/20/2017 MDufilho 1 Characteristics of Viruses Viruses Minuscule, acellular, infectious agent having either DNA or RNA Cause infections
Chapter13 Characterizing and Classifying Viruses, Viroids, and Prions 11/20/2017 MDufilho 1 Characteristics of Viruses Viruses Minuscule, acellular, infectious agent having either DNA or RNA Cause infections
Under the Radar Screen: How Bugs Trick Our Immune Defenses
 Under the Radar Screen: How Bugs Trick Our Immune Defenses Session 7: Cytokines Marie-Eve Paquet and Gijsbert Grotenbreg Whitehead Institute for Biomedical Research HHV-8 Discovered in the 1980 s at the
Under the Radar Screen: How Bugs Trick Our Immune Defenses Session 7: Cytokines Marie-Eve Paquet and Gijsbert Grotenbreg Whitehead Institute for Biomedical Research HHV-8 Discovered in the 1980 s at the
Transcription and RNA processing
 Transcription and RNA processing Lecture 7 Biology W3310/4310 Virology Spring 2016 It is possible that Nature invented DNA for the purpose of achieving regulation at the transcriptional rather than at
Transcription and RNA processing Lecture 7 Biology W3310/4310 Virology Spring 2016 It is possible that Nature invented DNA for the purpose of achieving regulation at the transcriptional rather than at
RECAP (1)! In eukaryotes, large primary transcripts are processed to smaller, mature mrnas.! What was first evidence for this precursorproduct
! In eukaryotes, large primary transcripts are processed to smaller, mature mrnas.! What was first evidence for this precursorproduct") RECAP (1) In eukaryotes, large primary transcripts are processed to smaller, mature mrnas. What was first evidence for this precursorproduct relationship? DNA Observation: Nuclear RNA pool consists of
RECAP (1) In eukaryotes, large primary transcripts are processed to smaller, mature mrnas. What was first evidence for this precursorproduct relationship? DNA Observation: Nuclear RNA pool consists of
Sequences in the 5 and 3 R Elements of Human Immunodeficiency Virus Type 1 Critical for Efficient Reverse Transcription
 JOURNAL OF VIROLOGY, Sept. 2000, p. 8324 8334 Vol. 74, No. 18 0022-538X/00/$04.00 0 Copyright 2000, American Society for Microbiology. All Rights Reserved. Sequences in the 5 and 3 R Elements of Human
JOURNAL OF VIROLOGY, Sept. 2000, p. 8324 8334 Vol. 74, No. 18 0022-538X/00/$04.00 0 Copyright 2000, American Society for Microbiology. All Rights Reserved. Sequences in the 5 and 3 R Elements of Human
Human Genome Complexity, Viruses & Genetic Variability
 Human Genome Complexity, Viruses & Genetic Variability (Learning Objectives) Learn the types of DNA sequences present in the Human Genome other than genes coding for functional proteins. Review what you
Human Genome Complexity, Viruses & Genetic Variability (Learning Objectives) Learn the types of DNA sequences present in the Human Genome other than genes coding for functional proteins. Review what you
Chromatin IP (Isw2) Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles.
 Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles.") Chromatin IP (Isw2) 7/01 Toshi last update: 06/15 Reagents Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles. 2.5 M glycine. TBS:
Chromatin IP (Isw2) 7/01 Toshi last update: 06/15 Reagents Fix soln: 11% formaldehyde, 0.1 M NaCl, 1 mm EDTA, 50 mm Hepes-KOH ph 7.6. Freshly prepared. Do not store in glass bottles. 2.5 M glycine. TBS:
Supplementary Information Titles Journal: Nature Medicine
 Supplementary Information Titles Journal: Nature Medicine Article Title: Corresponding Author: Supplementary Item & Number Supplementary Fig.1 Fig.2 Fig.3 Fig.4 Fig.5 Fig.6 Fig.7 Fig.8 Fig.9 Fig. Fig.11
Supplementary Information Titles Journal: Nature Medicine Article Title: Corresponding Author: Supplementary Item & Number Supplementary Fig.1 Fig.2 Fig.3 Fig.4 Fig.5 Fig.6 Fig.7 Fig.8 Fig.9 Fig. Fig.11
AP Biology. Viral diseases Polio. Chapter 18. Smallpox. Influenza: 1918 epidemic. Emerging viruses. A sense of size
 Hepatitis Viral diseases Polio Chapter 18. Measles Viral Genetics Influenza: 1918 epidemic 30-40 million deaths world-wide Chicken pox Smallpox Eradicated in 1976 vaccinations ceased in 1980 at risk population?
Hepatitis Viral diseases Polio Chapter 18. Measles Viral Genetics Influenza: 1918 epidemic 30-40 million deaths world-wide Chicken pox Smallpox Eradicated in 1976 vaccinations ceased in 1980 at risk population?
LESSON 4.6 WORKBOOK. Designing an antiviral drug The challenge of HIV
 LESSON 4.6 WORKBOOK Designing an antiviral drug The challenge of HIV In the last two lessons we discussed the how the viral life cycle causes host cell damage. But is there anything we can do to prevent
LESSON 4.6 WORKBOOK Designing an antiviral drug The challenge of HIV In the last two lessons we discussed the how the viral life cycle causes host cell damage. But is there anything we can do to prevent
Virology Introduction. Definitions. Introduction. Structure of virus. Virus transmission. Classification of virus. DNA Virus. RNA Virus. Treatment.
 DEVH Virology Introduction Definitions. Introduction. Structure of virus. Virus transmission. Classification of virus. DNA Virus. RNA Virus. Treatment. Definitions Virology: The science which study the
DEVH Virology Introduction Definitions. Introduction. Structure of virus. Virus transmission. Classification of virus. DNA Virus. RNA Virus. Treatment. Definitions Virology: The science which study the
Supplementary Information
 Supplementary Information HBV maintains electrostatic homeostasis by modulating negative charges from phosphoserine and encapsidated nucleic acids Authors: Pei-Yi Su 1,2,3, Ching-Jen Yang 2, Tien-Hua Chu
Supplementary Information HBV maintains electrostatic homeostasis by modulating negative charges from phosphoserine and encapsidated nucleic acids Authors: Pei-Yi Su 1,2,3, Ching-Jen Yang 2, Tien-Hua Chu
Structural and Functional Elements of the Promoter Encoded by the 5 Untranslated Region of the Venezuelan Equine Encephalitis Virus Genome
 JOURNAL OF VIROLOGY, Sept. 2009, p. 8327 8339 Vol. 83, No. 17 0022-538X/09/$08.00 0 doi:10.1128/jvi.00586-09 Copyright 2009, American Society for Microbiology. All Rights Reserved. Structural and Functional
JOURNAL OF VIROLOGY, Sept. 2009, p. 8327 8339 Vol. 83, No. 17 0022-538X/09/$08.00 0 doi:10.1128/jvi.00586-09 Copyright 2009, American Society for Microbiology. All Rights Reserved. Structural and Functional
Identification of Mutation(s) in. Associated with Neutralization Resistance. Miah Blomquist
 in. Associated with Neutralization Resistance. Miah Blomquist") Identification of Mutation(s) in the HIV 1 gp41 Subunit Associated with Neutralization Resistance Miah Blomquist What is HIV 1? HIV-1 is an epidemic that affects over 34 million people worldwide. HIV-1
Identification of Mutation(s) in the HIV 1 gp41 Subunit Associated with Neutralization Resistance Miah Blomquist What is HIV 1? HIV-1 is an epidemic that affects over 34 million people worldwide. HIV-1
Genetics. Instructor: Dr. Jihad Abdallah Transcription of DNA
 Genetics Instructor: Dr. Jihad Abdallah Transcription of DNA 1 3.4 A 2 Expression of Genetic information DNA Double stranded In the nucleus Transcription mrna Single stranded Translation In the cytoplasm
Genetics Instructor: Dr. Jihad Abdallah Transcription of DNA 1 3.4 A 2 Expression of Genetic information DNA Double stranded In the nucleus Transcription mrna Single stranded Translation In the cytoplasm
SUPPLEMENTARY INFORMATION
 Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
Supplementary Figures Supplementary Figure S1. Binding of full-length OGT and deletion mutants to PIP strips (Echelon Biosciences). Supplementary Figure S2. Binding of the OGT (919-1036) fragments with
Innate Immunity & Inflammation
 Innate Immunity & Inflammation The innate immune system is an evolutionally conserved mechanism that provides an early and effective response against invading microbial pathogens. It relies on a limited
Innate Immunity & Inflammation The innate immune system is an evolutionally conserved mechanism that provides an early and effective response against invading microbial pathogens. It relies on a limited
Unexpected complexity in the interference activity of a cloned influenza defective interfering RNA
 Meng et al. Virology Journal (2017) 14:138 DOI 10.1186/s12985-017-0805-6 RESEARCH Unexpected complexity in the interference activity of a cloned influenza defective interfering RNA Open Access Bo Meng
Meng et al. Virology Journal (2017) 14:138 DOI 10.1186/s12985-017-0805-6 RESEARCH Unexpected complexity in the interference activity of a cloned influenza defective interfering RNA Open Access Bo Meng
Viral Vectors In The Research Laboratory: Just How Safe Are They? Dawn P. Wooley, Ph.D., SM(NRM), RBP, CBSP
, RBP, CBSP") Viral Vectors In The Research Laboratory: Just How Safe Are They? Dawn P. Wooley, Ph.D., SM(NRM), RBP, CBSP 1 Learning Objectives Recognize hazards associated with viral vectors in research and animal
Viral Vectors In The Research Laboratory: Just How Safe Are They? Dawn P. Wooley, Ph.D., SM(NRM), RBP, CBSP 1 Learning Objectives Recognize hazards associated with viral vectors in research and animal
Processing of RNA II Biochemistry 302. February 18, 2004 Bob Kelm
 Processing of RNA II Biochemistry 302 February 18, 2004 Bob Kelm What s an intron? Transcribed sequence removed during the process of mrna maturation Discovered by P. Sharp and R. Roberts in late 1970s
Processing of RNA II Biochemistry 302 February 18, 2004 Bob Kelm What s an intron? Transcribed sequence removed during the process of mrna maturation Discovered by P. Sharp and R. Roberts in late 1970s
Rama Nada. - Malik
 - 2 - Rama Nada - - Malik 1 P a g e We talked about HAV in the previous lecture, now we ll continue the remaining types.. Hepatitis E It s similar to virus that infect swine, so its most likely infect
- 2 - Rama Nada - - Malik 1 P a g e We talked about HAV in the previous lecture, now we ll continue the remaining types.. Hepatitis E It s similar to virus that infect swine, so its most likely infect
7.013 Spring 2005 Problem Set 7
 MI Department of Biology 7.013: Introductory Biology - Spring 2005 Instructors: Professor Hazel Sive, Professor yler Jacks, Dr. Claudette Gardel 7.013 Spring 2005 Problem Set 7 FRIDAY May 6th, 2005 Question
MI Department of Biology 7.013: Introductory Biology - Spring 2005 Instructors: Professor Hazel Sive, Professor yler Jacks, Dr. Claudette Gardel 7.013 Spring 2005 Problem Set 7 FRIDAY May 6th, 2005 Question
Practice Problems 8. a) What do we define as a beneficial or advantageous mutation to the virus? Why?
 What do we define as a beneficial or advantageous mutation to the virus? Why?") Life Sciences 1a Practice Problems 8 1. You have two strains of HIV one is a wild type strain of HIV and the second has acquired a mutation in the gene encoding the protease. This mutation has a dual effect
Life Sciences 1a Practice Problems 8 1. You have two strains of HIV one is a wild type strain of HIV and the second has acquired a mutation in the gene encoding the protease. This mutation has a dual effect
Virus and Prokaryotic Gene Regulation - 1
 Virus and Prokaryotic Gene Regulation - 1 We have discussed the molecular structure of DNA and its function in DNA duplication and in transcription and protein synthesis. We now turn to how cells regulate
Virus and Prokaryotic Gene Regulation - 1 We have discussed the molecular structure of DNA and its function in DNA duplication and in transcription and protein synthesis. We now turn to how cells regulate
Norgen s HIV proviral DNA PCR Kit was developed and validated to be used with the following PCR instruments: Qiagen Rotor-Gene Q BioRad icycler
 3430 Schmon Parkway Thorold, ON, Canada L2V 4Y6 Phone: (905) 227-8848 Fax: (905) 227-1061 Email: techsupport@norgenbiotek.com HIV Proviral DNA PCR Kit Product # 33840 Product Insert Background Information
3430 Schmon Parkway Thorold, ON, Canada L2V 4Y6 Phone: (905) 227-8848 Fax: (905) 227-1061 Email: techsupport@norgenbiotek.com HIV Proviral DNA PCR Kit Product # 33840 Product Insert Background Information
MOLECULAR CELL BIOLOGY
 1 Lodish Berk Kaiser Krieger scott Bretscher Ploegh Matsudaira MOLECULAR CELL BIOLOGY SEVENTH EDITION CHAPTER 13 Moving Proteins into Membranes and Organelles Copyright 2013 by W. H. Freeman and Company
1 Lodish Berk Kaiser Krieger scott Bretscher Ploegh Matsudaira MOLECULAR CELL BIOLOGY SEVENTH EDITION CHAPTER 13 Moving Proteins into Membranes and Organelles Copyright 2013 by W. H. Freeman and Company
Norgen s HIV Proviral DNA PCR Kit was developed and validated to be used with the following PCR instruments: Qiagen Rotor-Gene Q BioRad T1000 Cycler
 3430 Schmon Parkway Thorold, ON, Canada L2V 4Y6 Phone: 866-667-4362 (905) 227-8848 Fax: (905) 227-1061 Email: techsupport@norgenbiotek.com HIV Proviral DNA PCR Kit Product# 33840 Product Insert Intended
3430 Schmon Parkway Thorold, ON, Canada L2V 4Y6 Phone: 866-667-4362 (905) 227-8848 Fax: (905) 227-1061 Email: techsupport@norgenbiotek.com HIV Proviral DNA PCR Kit Product# 33840 Product Insert Intended
LEC 2, Medical biology, Theory, prepared by Dr. AYAT ALI
 General Characteristics, Structure and Taxonomy of Viruses Viruses A virus is non-cellular organisms made up of genetic material and protein that can invade living cells. They are considered both a living
General Characteristics, Structure and Taxonomy of Viruses Viruses A virus is non-cellular organisms made up of genetic material and protein that can invade living cells. They are considered both a living
AIDS - Knowledge and Dogma. Conditions for the Emergence and Decline of Scientific Theories Congress, July 16/ , Vienna, Austria
 AIDS - Knowledge and Dogma Conditions for the Emergence and Decline of Scientific Theories Congress, July 16/17 2010, Vienna, Austria Reliability of PCR to detect genetic sequences from HIV Juan Manuel
AIDS - Knowledge and Dogma Conditions for the Emergence and Decline of Scientific Theories Congress, July 16/17 2010, Vienna, Austria Reliability of PCR to detect genetic sequences from HIV Juan Manuel
Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics
 Hepadnaviruses Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Hepatitis viruses A group of unrelated pathogens termed hepatitis viruses cause the vast majority
Hepadnaviruses Virion Genome Genes and proteins Viruses and hosts Diseases Distinctive characteristics Hepatitis viruses A group of unrelated pathogens termed hepatitis viruses cause the vast majority
Phosphate buffered saline (PBS) for washing the cells TE buffer (nuclease-free) ph 7.5 for use with the PrimePCR Reverse Transcription Control Assay
 for washing the cells TE buffer (nuclease-free) ph 7.5 for use with the PrimePCR Reverse Transcription Control Assay") Catalog # Description 172-5080 SingleShot Cell Lysis Kit, 100 x 50 µl reactions 172-5081 SingleShot Cell Lysis Kit, 500 x 50 µl reactions For research purposes only. Introduction The SingleShot Cell Lysis
Catalog # Description 172-5080 SingleShot Cell Lysis Kit, 100 x 50 µl reactions 172-5081 SingleShot Cell Lysis Kit, 500 x 50 µl reactions For research purposes only. Introduction The SingleShot Cell Lysis
Unit 13.2: Viruses. Vocabulary capsid latency vaccine virion
 Unit 13.2: Viruses Lesson Objectives Describe the structure of viruses. Outline the discovery and origins of viruses. Explain how viruses replicate. Explain how viruses cause human disease. Describe how
Unit 13.2: Viruses Lesson Objectives Describe the structure of viruses. Outline the discovery and origins of viruses. Explain how viruses replicate. Explain how viruses cause human disease. Describe how
Oncolytic virus strategy
 Oncolytic viruses Oncolytic virus strategy normal tumor NO replication replication survival lysis Oncolytic virus strategy Mechanisms of tumor selectivity of several, some of them naturally, oncolytic
Oncolytic viruses Oncolytic virus strategy normal tumor NO replication replication survival lysis Oncolytic virus strategy Mechanisms of tumor selectivity of several, some of them naturally, oncolytic
General Principles of Endocrine Physiology
 General Principles of Endocrine Physiology By Dr. Isabel S.S. Hwang Department of Physiology Faculty of Medicine University of Hong Kong The major human endocrine glands Endocrine glands and hormones
General Principles of Endocrine Physiology By Dr. Isabel S.S. Hwang Department of Physiology Faculty of Medicine University of Hong Kong The major human endocrine glands Endocrine glands and hormones
Supplemental Materials and Methods Plasmids and viruses Quantitative Reverse Transcription PCR Generation of molecular standard for quantitative PCR
 Supplemental Materials and Methods Plasmids and viruses To generate pseudotyped viruses, the previously described recombinant plasmids pnl4-3-δnef-gfp or pnl4-3-δ6-drgfp and a vector expressing HIV-1 X4
Supplemental Materials and Methods Plasmids and viruses To generate pseudotyped viruses, the previously described recombinant plasmids pnl4-3-δnef-gfp or pnl4-3-δ6-drgfp and a vector expressing HIV-1 X4
MedChem 401~ Retroviridae. Retroviridae
 MedChem 401~ Retroviridae Retroviruses plus-sense RNA genome (!8-10 kb) protein capsid lipid envelop envelope glycoproteins reverse transcriptase enzyme integrase enzyme protease enzyme Retroviridae The
MedChem 401~ Retroviridae Retroviruses plus-sense RNA genome (!8-10 kb) protein capsid lipid envelop envelope glycoproteins reverse transcriptase enzyme integrase enzyme protease enzyme Retroviridae The
Supplementary information. MARCH8 inhibits HIV-1 infection by reducing virion incorporation of envelope glycoproteins
 Supplementary information inhibits HIV-1 infection by reducing virion incorporation of envelope glycoproteins Takuya Tada, Yanzhao Zhang, Takayoshi Koyama, Minoru Tobiume, Yasuko Tsunetsugu-Yokota, Shoji
Supplementary information inhibits HIV-1 infection by reducing virion incorporation of envelope glycoproteins Takuya Tada, Yanzhao Zhang, Takayoshi Koyama, Minoru Tobiume, Yasuko Tsunetsugu-Yokota, Shoji
p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO
 Supplementary Information p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO Yuri Shibata, Masaaki Oyama, Hiroko Kozuka-Hata, Xiao Han, Yuetsu Tanaka,
Supplementary Information p47 negatively regulates IKK activation by inducing the lysosomal degradation of polyubiquitinated NEMO Yuri Shibata, Masaaki Oyama, Hiroko Kozuka-Hata, Xiao Han, Yuetsu Tanaka,
Sarah A. Kopecky and Douglas S. Lyles*
 JOURNAL OF VIROLOGY, Apr. 2003, p. 4658 4669 Vol. 77, No. 8 0022-538X/03/$08.00 0 DOI: 10.1128/JVI.77.8.4658 4669.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved. Contrasting
JOURNAL OF VIROLOGY, Apr. 2003, p. 4658 4669 Vol. 77, No. 8 0022-538X/03/$08.00 0 DOI: 10.1128/JVI.77.8.4658 4669.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved. Contrasting
LESSON 4.4 WORKBOOK. How viruses make us sick: Viral Replication
 DEFINITIONS OF TERMS Eukaryotic: Non-bacterial cell type (bacteria are prokaryotes).. LESSON 4.4 WORKBOOK How viruses make us sick: Viral Replication This lesson extends the principles we learned in Unit
DEFINITIONS OF TERMS Eukaryotic: Non-bacterial cell type (bacteria are prokaryotes).. LESSON 4.4 WORKBOOK How viruses make us sick: Viral Replication This lesson extends the principles we learned in Unit