Immune response to Streptococcus pneumoniae polysaccharide vaccination and antigen-selected B cells in highly susceptible individuals
|
|
|
- Aubrie King
- 5 years ago
- Views:
Transcription
1 The University of Toledo The University of Toledo Digital Repository Theses and Dissertations 2014 Immune response to Streptococcus pneumoniae polysaccharide vaccination and antigen-selected B cells in highly susceptible individuals David Jason Leggat University of Toledo Follow this and additional works at: Recommended Citation Leggat, David Jason, "Immune response to Streptococcus pneumoniae polysaccharide vaccination and antigen-selected B cells in highly susceptible individuals" (2014). Theses and Dissertations This Dissertation is brought to you for free and open access by The University of Toledo Digital Repository. It has been accepted for inclusion in Theses and Dissertations by an authorized administrator of The University of Toledo Digital Repository. For more information, please see the repository's About page.
2 A Dissertation entitled Immune Response to Streptococcus pneumoniae Polysaccharide Vaccination and Antigen-Selected B cells in Highly Susceptible Individuals. by David Jason Leggat Submitted to the Graduate Faculty as partial fulfillment of the requirements for the Doctor of Philosophy Degree in Biomedical Sciences M.A. Julie Westerink, MD, Committee Chair Robert Blumenthal, PhD, Committee Member Deepak Malhotra, MD, PhD, Committee Member Kevin Pan, M.D., PhD, Committee Member Mark Wooten, PhD, Committee Member Patricia R. Komuniecki, PhD, Dean College of Graduate Studies The University of Toledo May 2014
3 Copyright 2014, David Jason Leggat This document is copyrighted material. Under copyright law, no parts of this document may be reproduced without the expressed permission of the author.
4 An Abstract of Immune Response to Streptococcus pneumoniae Polysaccharide Vaccination and Antigen-Selected B Cells in Highly Susceptible Populations. by David Jason Leggat Submitted to the Graduate Faculty as partial fulfillment of the requirements for the Doctor of Philosophy Degree in Biomedical Sciences The University of Toledo May 2014 Pneumococcus is a major human pathogen responsible for causing substantial morbidity and mortality worldwide. It is a causative agent of pneumonia, otitis media, and invasive pneumococcal diseases. Pathogenic strains express a single type of polysaccharide on their surface which forms an anti-phagocytic polysaccharide capsule. Disease incidence is more frequent in high risk populations which include children <2 years old, elderly adults >65 years old, and HIV-positive individuals (HIV-positive). Therefore, the Advisory Committee on Immunization Practices recommends vaccination with the 23-valent pneumococcal polysaccharide vaccine (PPV23) for individuals that are at high risk for infection. Elderly and HIV-positive individuals however do not respond optimally to vaccination. The host immune response combats invasive pneumococcal disease by antibodycomplement mediated opsonophagocytosis, essential for bacterial clearance. This requires the use of anti-pneumococcal polysaccharide antibodies, serum complement, and phagocytic cells. PPS are classified as T-cell independent type II antigens. The B cells responsible for producing antibodies against TI-2 antigens have been greatly debated. In iii
5 order to improve vaccination in high-risk individuals we need to have a deeper understanding of the immune response to PPV23 in low and high-risk populations. Using directly conjugated fluorescently labeled PPS we have characterized PPSspecific B cells in young healthy individuals using flow cytometry. The majority of PPSspecific B cells responding to vaccination are IgM+ memory cells (CD27+IgM+). We further characterized these PPV responding B cells showing that the majority of PPSspecific B cells on day-7 are phenotypically characterized as CD19+CD20+CD3-CD70- CD27+IgM+CD43+CD5+/-. This population is in alignment with recent reports of human B-1 cells. Elderly individuals are at high risk for invasive pneumococcal disease. PPSspecific B cell deficiencies in this population remain to be elucidated. We identified the phenotype of PPS-specific B cells using CD19, CD27 and IgM surface markers and enumerated them. Our results indicate a significant rises in PPS-specific total and functional antibody titers post-ppv23 vaccination in the elderly. Moreover, our results demonstrate significant differences in the phenotype of PPS-responding B cells, PPSspecific IgM and OPA, post-vaccination in the elderly compared to young adults. These findings likely contribute to the decreased efficacy of PPV23 in individuals over the age of 65. Early severe B cell dysfunction is a central feature of HIV-positive individuals. Moreover, anti-pneumococcal IgM and IgG Abs are severely reduced in HIV-positive individuals immunized with PPV23 compared to HIV-negative individuals. This suggests that HIV-positive individuals lack important PPS-responding B cells necessary to provide sufficient protection. Recommendations for HIV-positive individuals with CD4<200 iv
6 remain unclear. It is not known if newly diagnosed HIV-infected with CD4<200 benefit from delayed immunization following 6-12 months HAART allowing viral suppression and partial immune reconstitution. Our results indicate significant rises in PPS-specific total and functional antibody titers post-ppv23 vaccination in newly diagnosed HIVpositive individuals. We show that HIV-positive individuals have an abnormal PPSspecific B cell response compared to HIV-negative individuals. Moreover, there is no apparent benefit to delaying vaccination for 6-12 months of HAART. High incidence of pneumococcal disease has led to the recommendation of vaccinating all HIV-positive adults against Streptococcus pneumoniae followed by a booster after 5 years. The benefits of PPV23 revaccination in HIV-positive individuals has been a subject of debate. We therefore evaluated the effect of PPV23 revaccination in HIV-positive individuals on long term HAART. Our results indicate significant rises in PPS-specific total and functional antibody titers post-ppv23 revaccination in the longterm HAART volunteers. This indicates PPV23 revaccination is beneficial; however responses remained suboptimal irrespective of the degree of CD4 T cell reconstitution compared to HIV-negative individuals. The results indicate persistent PPS-specific B cell deficiencies despite PPV23 revaccination and long-term HAART administration. v
7 Dedicated to my family and close friends for their support and sacrifice while I spent a great many late nights pursuing this endeavor over the years. You had faith in me during the worst of times, without which, we would not have shared the best of times. vi
8 Acknowledgements I am grateful to my major advisor, M. A. Julie Westerink, M.D. for accepting me into her laboratory and patiently guiding me through the process of publishing papers and earning my degree. I would like to thank Noor Khaskhely, M.D., Ph.D. for training, guidance, and the many late night hours working together. I am also thankful for my academic advisory committee for their guidance and support; Robert Blumenthal, Ph.D., Deepak Malhotra, M.D., Ph.D., Kevin Pan, Ph.D., and Mark Wooten, Ph.D.. I greatly appreciate the many study volunteers who participated in our studies, without them, there would be no progress. The Ruppert Center clinic and infectious disease staff as well as the Institute of Aging for efforts in recruiting volunteers. I would also like to thank the following collaborators for their contributions: Romero-Steiner, Ph.D. for serotypes of S. pneumoniae used in opsonophagocytic assays. Sadik Khuder, Ph.D. for statistical analysis The other present and past members of the Westerink lab for their support: Anita S. Iyer, M.S. Jennifer A. Othola, B.S. Jason Mosakowski, B.S. Rebecca Thompson, Ph.D. Sneha Kommoori, M.D. Adeeb A. Al Rizaiza, M.D. vii Michael Woodman, Ph.D.
9 Table of Contents Abstract... iii Acknowledgements... vii Table of Contents... viii List of Tables... xiii List of Figures... xiv List of Abbreviations... xvi List of Symbols... xix Preface...xx 1 Streptococcus pneumoniae: A Human Pathogen - Chapter Epidemiology Pathogenesis Immune response Initial interactions Bacterial clearance T-cell independent response Medical management Antibiotics Pneumococcal polysaccharide vaccine...12 viii
10 1.4.3 Pneumococcal conjugate vaccine PPV23 in high risk populations Old age (>65 years) HIV-positivity HIV-positive HAART-treated patients The Phenotype of PPS-specific B cells in Adult Peripheral Blood - Chapter Chapter introduction Results Antibody titers increase post-immunization with PPV OPT increases significantly 30 days post-immunization with PPV PPV induces transient increase in PPS-specific B cells in peripheral blood Majority of PPS-specific B cells express CD27 and IgM post-immunization with PPV Majority of PPS-specific IgM+ Memory B cells express CD43 and CD5 post-immunization with PPV PPS-selected B cell percentages and absolute numbers PPS-selected B cell phenotypically resemble human B-1 cells Chapter discussion Elderly Immune Response to PPV - Chapter Chapter introduction Results ix
11 3.2.1 Elderly serum antibody titers increased post-vaccination Elderly serum opsonophagocytic antibody titers increased post-vaccination Unselected B lymphocyte counts are decreased in elderly peripheral blood Phenotype of polysaccharide-specific B lymphocytes isolated from elderly PBMC Phenotypes of polysaccharide-specific B lymphocytes isolated from elderly PBMC compared to healthy young adults Chapter discussion Immune Deficiencies in Newly Diagnosed HIV-infected Individuals Chapter Chapter introduction Results HIV-positive individuals had diminished PPS-specific antibody titers HIV-positive individuals had diminished PPS-specific functional antibody response Decreased B lymphocyte populations in HIV-positive peripheral blood Phenotypic analysis of PPS-specific B cells in HIV-positive blood Differences in phenotypic distribution of PPS-specific B cells in x
12 HIV-positive compared to HIV-negative individuals Chapter discussion PPV Revaccination in HIV-positive Individuals on Long-term Therapy Chapter Chapter introduction Results Serum PPS-specific antibody levels increased post-ppv23 revaccination in long-term HAART volunteers Functional antibody titers increased post-ppv23 revaccination in long-term HAART volunteers Absolute B cell numbers and B cell percentage in long-term HAART volunteers Phenotypic analysis of PPS-specific B cells in the HIV-positive volunteers on long-term HAART post-ppv23 revaccination Diminished response to PPV23 in HIV-positive on long-term HAART compared to HIV-negative despite revaccination Chapter discussion Discussion and Interpretations - Chapter Antibody responses and deficiencies ELISA titers Opsonophagocytic titers PPS-specific B cells Young healthy individuals xi
13 6.2.2 Elderly individuals ND-HIV individuals Revaccination in long-term HAART Future studies Materials and Methods - Chapter Study volunteers Chapter 2 study volunteers Chapter 3 study volunteers Chapter 4 study volunteers Chapter 5 study volunteers Vaccination and blood draw Pneumococcal polysaccharide enzyme linked immunosorbent assay (ELISA) Opsonophagocytic assay (OPA) Labeling of polysaccharide 14 and 23F with fluorescent dye Flow cytometry Statistical analysis References A Pneumococcal polysaccharide vaccination induces polysaccharide-specific B cells in adult peripheral blood expressing CD19+CD20+CD3-CD70- CD27+IgM+CD43+CD5+/ B Appendix B: The elderly immune response to pneumococcal polysaccharides 14 and 23F consists predominantly of switched memory B cells xii
14 List of Tables 2.1 CD19+PPS+ B cells in young healthy individuals Percentages and numbers of PPS14 selected B cells young healthy Percentages and numbers of PPS23F selected B cells young healthy Unselected CD19+ B cell numbers young and elderly PPS-selected CD19+ B cell percentages and counts in young and elderly Unselected CD19+ B cell percentages and counts in HIV-negative and ND-HIV individuals Unselected CD19+ B cell percentages and counts in ND-HIV PPS-selected CD19+ B cell percentages and counts in ND-HIV Unselected B cell counts and percentages in HIV-HAART Unselected B cell subset counts and percentages in HIV-HAART PPS-selected CD19+ B cell percentages and counts in HIV-HAART...76 xiii
15 List of Figures 1-1 Streptococcus pneumoniae interacting with epithelial cells Streptococcus pneumoniae anti-phagocytic capsule IPD incidence distribution by age Pneumococcal pneumonia Complement pathways TI-2 B cell response to PPS PPS-specific antibody and opsonophagocytic titers in young healthy Phenotypic analysis of human peripheral blood B cells Majority of PPS-specific B cells are CD27+IgM+ in young healthy Majority of PPS-specific IgM+ memory B cells are CD43+CD PPS-selected B cells resemble human B-1 cells PPS-specific antibody titers in elderly PPS-specific opsonophagocytic titers in elderly PPS-specific B cells in elderly pre- to post-ppv PPS-specific B cells post-ppv23 in elderly compared to young adults HIV-positive serum antibody and opsonophagocytic titers Majority of PPS-specific B cells are CD27+IgM+ in ND-HIV...63 xiv
16 4-3 PPS-specific B cells post-ppv23 in ND-HIV compared to HIV-negative individuals Revaccinated HIV-HAART serum antibody and opsonophagocytic titers Revaccinated HIV-HAART PPS-specific B cell subsets PPS-specific B cells in revaccinated HIV-HAART compared to HIV-negative individuals...80 xv
17 List of Abbreviations Abs. Antibodies ACIP.. Advisory Committee on Immunization Practices Ags... Antigens APC Allophycocyanin ATCC.American Type Culture Collection CB..Cascade blue ethylenediamine CDC...Centers for Disease Control and Prevention CR..Complement receptor CRP C reactive protein CWPS. Cell wall polysaccharide Cy Cyanine dye DTAF.5-(4,6-Dichlorotriazinyl) aminofluorescein EDTA Ethylenediaminetetraacetic acid ELISA Enzyme-linked immunosorbent assay FACS Fluorescence assisted cell sorting FCS...Forward scatter FITC.Fluorescein isothiocyanate FMO..Fluorescence minus one H 2 SO 4.Sulfuric acid HAART.. Highly active anti-retroviral therapy HAART naïve HIV positive individuals CD4<200 without HAART HAART treated.hiv positive individuals CD4<200 on 6-12 months HAART HIV-HAART. HIV positive individuals on long-term HAART HIV-negative. HIV negative individuals HIV-positive.. HIV positive individuals xvi
18 HRP Horseradish peroxidase Ig Immunoglobulin IgM+ memory... CD19+CD27+IgM+ B cells IL Interleukin IPD. Invasive pneumococcal disease IRB Internal review board Naïve... CD19+CD27- B cells ND-HIV. Newly diagnosed HIV-positive individuals mab monoclonal antibody MZB.. Marginal zone B cells OPA... Opsonophagocytic Assay OPT. Opsonophagocytic Titer PBL.Peripheral blood lymphocytes PBMC.... Peripheral blood mononuclear cell PBS.Phosphate buffered saline PBST..PBS plus 0.1% Tween 20 PCV.... Pneumococcal conjugate vaccine PE Phycoerythrin PerCP..Peridinin chlorophyll protein Pneumococcus... Streptococcus pneumoniae PPS..... Pneumococcal polysaccharide PPS14.Pneumococcal polysaccharide serotype 14 PPS23F Pneumococcal polysaccharide serotype 23F PPV Pneumococcal polysaccharide vaccine PPV23 23-valent pneumococcal polysaccharide vaccine S. pneumoniae Streptococcus pneumonia SSC.Side scatter SEM Standard error of the mean Switched memory.. CD19+CD27+IgM- B cells TD.. T cell dependent Th2.T-cell helper type 2 xvii
19 TI-2 Type 2 T cell independent TLR Toll like receptor WHO World Health Organization xviii
20 List of Symbols >. Greater than <. Less than C...degrees Celsius KDa kilodalton l microliter mg..milligram ml... milliliter mm.millimolar M molar nm..nanometer xix
21 Preface Streptococcus pneumoniae is a major human pathogen responsible for substantial morbidity and mortality worldwide. Vaccines and antibiotics have been developed in an effort to reduce disease burden. Despite these efforts, vaccination is suboptimal and antibiotic resistance is mounting [1-5]. Certain groups within the population are at considerably higher risk for infection than others. This includes, but is not limited to, the elderly (>65 years old) and HIV-positive individuals [6, 7]. The pneumococcal polysaccharide (PPS) which composes the bacterial capsule is expressed on the outer surface of S. pneumoniae. This is the primary antigen which is identified by B cells and elicits a protective antibody response. These antibodies in conjunction with serum complement are critical for bacterial clearance by phagocytic cells, including macrophages and neutrophils [8, 9]. In contrast to the majority of vaccine antigens which are T cell dependent, pneumococcal polysaccharide is a T cell independent type II antigen. These antigens do not require T cell help to stimulate antibody production by B cells. The immune response to T-cell independent type II antigens remains controversial and an active area of research. In mice, specific B cell subpopulations play a central role in the response to PPS antigens, including both splenic marginal zone B cells and B-1 xx
22 cells [10, 11]. In humans, understanding the involvement of these cells is confounded in part by limited tissue samples and a lack of defined B cell phenotypic markers. Increased disease incidence in high risk populations, and low functional antibody titers, suggests intrinsic B cell deficiencies within these populations [6, 7, 12, 13]. In order to reduce disease burden in high risk populations, these deficiencies must first be identified. Direct comparison of antigen-specific humoral and B cell responses in elderly and HIV-positive individuals populations to young non-infected controls can identify deficiencies likely contributing to their increased susceptibility. Moreover, the benefits of vaccination schedules and therapeutic treatments can be tested to determine the most beneficial clinical course of treatment. We hypothesize that elderly and HIV-infected individuals possess differential intrinsic antigen-specific B cell deficiencies which leads to decreased antibody responses to S. pneumoniae. We therefore conducted the following studies: 1) Identify PPS-specific humoral and B cell responses to pneumococcal polysaccharide vaccination in young healthy individuals. a. Identify PPS-specific antibody titers b. Identify PPS-specific opsonophagocytic titers c. Identify the phenotype of polysaccharide-specific B cells 2) Identify PPS-specific humoral and B cell responses to pneumococcal polysaccharide vaccination in elderly individuals (>65 years old). a. Identify PPS-specific antibody titers b. Identify PPS-specific opsonophagocytic titers xxi
23 c. Identify the phenotype of polysaccharide-specific B cell populations compared to young individuals. 3) Identify PPS-specific humoral and B cell responses to pneumococcal polysaccharide vaccination in newly diagnosed HIV-infected individuals and define the effects of 6-12 months of anti-viral therapy pre-vaccination. a. Identify PPS-specific antibody titers b. Identify PPS-specific opsonophagocytic titers c. Identify the phenotype of polysaccharide-specific B cell populations compared to HIV-negative individuals. 4) Identify PPS-specific humoral and B cell responses to pneumococcal polysaccharide vaccination in HIV-infected individuals revaccinated after 5 years on long-term anti-viral therapy. d. Identify PPS-specific antibody titers e. Identify PPS-specific opsonophagocytic titers f. Identify the phenotype of polysaccharide-specific B cell populations compared to HIV-negative individuals. xxii
 is a Gram-positive extracellular bacterium commonly found asymptomatically colonizing the mucosa of the nasopharynx of healthy individuals")
24 Chapter 1 Streptococcus pneumoniae: A Human Pathogen 1.1 Epidemiology Streptococcus pneumoniae (pneumococcus) is a Gram-positive extracellular bacterium commonly found asymptomatically colonizing the mucosa of the nasopharynx of healthy individuals (Figure 1-1)[14-17]. Infection is spread between individuals by aerosol or direct contact. Pneumococcus is a major human pathogen responsible for causing substantial morbidity and mortality worldwide. It is a causative agent of pneumonia, otitis media, and invasive pneumococcal diseases (IPD) including septicemia and meningitis [17, 18]. The CDC estimates 31,600 cases of IPD result in 3,300 deaths each year in the United States [19]. Figure 1-1 Streptococcus pneumoniae interacting with epithelial cells [14]. Reprinted with permission. 1
[17, 20].")
25 Not all strains of pneumococcus are associated with disease. Pathogenic strains express a single type of structurally unique polysaccharide on their surface, which forms an anti-phagocytic polysaccharide capsule (Figure 1-2)[17, 20]. The capsule is both a major virulence factor and an important target for the adaptive immune response. More than 90 serotypes have been discovered since 1880 [17, 21]. Most serotypes are capable of causing disease, but the majority of infections are caused by a few dominant serotypes. Although serotype prevalence varies by age group and geographical location, 62% of IPD in the United States is caused by only 10 serotypes [17]. Populations which acquire immunity against specific serotypes often see an increase in colonization by new serotypes referred to as replacement serotypes. Figure 1-2 Streptococcus pneumoniae anti-phagocytic capsule [20]. Reprinted with permission. Disease incidence is more frequent in high risk populations which include young children (<2 years old), elderly adults (>65 years old), and the HIV-positive [6, 7] (Figure 1-3). In their first year, 60% of children have acute otitis media (middle ear infections) 2
![caused by the pneumococcus in 28%-55% of cases [17]. Children <2 years of age have the highest rates of IPD, accounting for 20% of total cases [17].](/docs-images/84/89184570/images/26-0.jpg "In this population, bacteremia constitutes 70% of IPD incidence [17]. Pneumococcus is also the primary cause of bacterial meningitis in this age group [17].")
26 caused by the pneumococcus in 28%-55% of cases [17]. Children <2 years of age have the highest rates of IPD, accounting for 20% of total cases [17]. In this population, bacteremia constitutes 70% of IPD incidence [17]. Pneumococcus is also the primary cause of bacterial meningitis in this age group [17]. Increased susceptibility in young children is primarily attributed to the incomplete development of the immune system, which limits their ability to produce PPS-specific antibodies against the polysaccharide capsule. Figure 1-3: IPD incidence distribution by age. Originally published by the Indiana state government: 3
27 Elderly individuals accounted for the majority of severe pneumococcal infections, resulting in an estimated 242,000 hospitalizations each year [18]. Bacteremic related mortality in adults is estimated to be 10-15%, however in the elderly population this can be as high as 60% [7, 17]. Meningitis related mortality has been estimated to be as high as 80% [17]. Previous studies have suggested that disease susceptibility in this age group is in part due to a loss of PPS-specific B cell populations, decreased PPS-specific B cell activity, and decreased antibody (Ab) function [22-30]. HIV-positive individuals (HIV-positive) are prone to many infectious diseases as a result of their severe immune dysfunction, which affects T cells, B cells, macrophages, and dendritic cells [31-37]. Pneumococcus is the primary cause for bacterial respiratory infection in HIV-positive individuals. It is commonly recurrent and associated with bacteremia [38-42]. Before the availability of highly active anti-retroviral therapy (HAART), HIV-positive individuals were up to 100-fold more likely to develop invasive pneumococcal disease than age matched HIV-negative individuals (HIV-negative) [38]. Newly diagnosed HIV-positive individuals (ND-HIV) not receiving HAART are expected to have the same risk of acquiring IPD. HAART inhibits viral replication and allows partial recovery of the immune system. HAART usage has led to significant reduction in IPD among HIV-positive individuals [43-46]. Incidence of IPD in HIVpositive individuals remains 35-fold more frequent than in HIV-negative individuals despite HAART usage [4]. Increased susceptibility to pneumococcus even after HAART usage is hypothesized to result, in part, from the loss (or functional loss) of specific lymphocytes which can not be restored with antiviral treatment [47, 48]. 4
28 1.2 Pathogenesis Colonization of the nasopharynx with pathogenic serotypes is the first step leading to pneumonia, otitis media, bacteremia, and meningitis [49]. Otitis media occurs when pneumococci transverse the eustachian tubes to the middle ear resulting in inflammation. It is most commonly found in children, in whom the eustachian tubes are smaller in diameter and have a more horizontal orientation compared to adults. Cell wall bacterial components and the release of pneumolysin initiate inflammation, which results in the recruitment of leukocytes [50]. Leukocytes enhance inflammation, resulting in tissue damage and disruption of the epithelial tissue. Disruption of the epithelial tissue allows the initiation of IPD including meningitis [50]. Streptococcus pneumoniae is the primary cause (36%) of adult pneumonia, resulting in 175,000 hospitalizations in the U.S. each year [17]. Pneumonia is preceded by 1-3 days of colonization, and is often associated with or preceded by viral infection such as influenza [17, 51]. Bacterial-expressed neuraminidase cleaves of host mucin, normally used to prevent bacterial passage to the lungs and host cell attachment [52]. Translocation from the nasopharynx, beyond the mucus covered ciliated cells of the upper respiratory tract, allows pneumococcus access to the alveolar space of the lungs (Figure 1-4). Pneumococcal growth on the alveolar epithelium, and release of pneumolysin and hydrogen peroxide, initiates inflammation and results in tissue damage [53]. Disruption of the alveolar epithelium allows edema fluid accumulation in the lung which inhibits respiration. Disruption of the host alveolar epithelium also allows bacterial 5

29 entry into the bloodstream which is a requisite for IPD, including septicemia and meningitis. Figure 1-4: Pneumococcal pneumonia. Originally published by the NIH national heart, lung, and blood institute 2014: Breaching of epithelial barriers is the initial step in the development of IPD. Initial invasion of pneumococci into the blood is called bacteremia. An estimated 10-15% of IPD cases result in death [7, 17]. Mortality rates are higher in the elderly and in immunocompromised individuals [54]. Unless cleared by the immune system, the bacteria grow to high titers in the blood, resulting in septicemia. Once pneumococci have infected the blood stream, they can circulate throughout the body and infect internal 6
30 organs resulting in additional pathological manifestations. Invasion of the cerebrospinal fluid is often a precursor to causing meningitis. 1.3 Immune response Initial interactions Mucus-coated epithelial cells of the nasopharynx provide the first barrier of resistance to pneumococcus, by trapping and physically blocking the bacteria from progressing deeper into the body. The bacterial capsule plays an important role in resisting entrapment in the host mucus and allowing adherence to the epithelium [55]. Colonization can last for weeks to months, but there has been no clear indication that this early exposure induces a protective adaptive immune response. Pneumococcus has a large array of virulence factors involved in colonization, defense against host immunity and disease progression [56]. Pneumolysin and hydrogen peroxide can be produced by pneumococci, leading to the disruption of host epithelial barriers [53]. Additionally, phosphorylcholine expressed on the bacterial surface can bind to host platelet-activating factor receptor on epithelial cells, allowing transcytosis and invasion of the blood stream [57, 58]. The innate immune response plays an important role in recognizing and initiating defense against disease progression. Host C-reactive protein (CRP) can bind phosphorylcholine, ultimately inhibiting invasion of the blood stream [57]. The host pattern recognition receptors called Toll-like receptors (TLRs) initiate the immune response, by detecting pneumococcal pathogen associated molecular patterns (PAMPs). TLR2, TLR4, and TLR9 all play roles in initiating a pro-inflammatory 7
31 response. TLR2 recognizes peptidoglycan, lipoteichoic acid and lipoproteins found in the bacterial cell wall [59-61], TLR4 recognizes the bacterial toxin pneumolysin [62], and TLR9 recognizes bacterial DNA (unmethylated CpG motifs) released by killed pneumococci [63]. TLR signaling results in NK- B-activated transcription, leading to the release of the pro-inflammatory cytokines IL-1 and IL-6 that recruit monocytes, macrophage and neutrophils by CCR2 mediated chemotaxis [64-67]. While protective, inflammation can also cause localized tissue damage disrupting the epithelial barriers and allowing pneumococcal entry into the host tissue and blood stream [50, 53, 58] Bacterial clearance The host immune response can also combat IPD by antibody-complement mediated opsonophagocytosis. This involves the use of essential anti-pneumococcal antibodies, serum complement, and phagocytic cells [65]. Although other complement activating components are also involved, mouse models have demonstrated that the classical complement pathway utilizing IgM plays a critical role in complement mediated bacterial clearance [3, 68]. Activation of the complement cascade results in the accumulation of C3b and ic3b on the bacterial surface (Figure 1-5)[69]. This is recognized by complement receptors (CR1 and CR3) on the surface of phagocytic leukocytes like liver and spleen resident macrophage. Recognition initiates bacterial opsonization, or enhanced phagocytosis [64, 65, 70]. Complement protein C1q bound to bacterial phosphorylcholine associated CRP can also initiate the classical pathway in absence of antibody [68]. The alternative pathway can contribute to bacterial clearance by enhancing complement activation once the complement cascade is initiated [68]. 8

32 Figure 1-5: Complement pathways. Reprinted with permission. Originally published by Nature Publishing Group T-cell independent response: The primary antigen which elicits a protective immune response against pneumococcal infection is the PPS capsule. PPS is a T cell independent type II (TI-2) antigen. This means it can stimulate B cells to produce antigen-specific antibodies without T helper type 2 (Th2) cell help [71]. Purified TI-2 antigens primarily induce the production of antigen-specific IgM [72](Figure 1-6). 9
 responsible for producing antibodies against TI-2 antigens has been greatly debated.")
![In mice, marginal zone B cells and B-1 cells have been implicated as playing important roles [10, 11].](/docs-images/84/89184570/images/33-2.jpg "In humans, support for splenic marginal zone B cell involvement stems from the increased risk of IPD in individuals with congenital, acquired or functional")
33 Figure 1-6: TI-2 B cell response to PPS. The immune response to TI-2 antigens in humans is not well understood and remains controversial. Specifically, the B cell subset(s) responsible for producing antibodies against TI-2 antigens has been greatly debated. In mice, marginal zone B cells and B-1 cells have been implicated as playing important roles [10, 11]. In humans, support for splenic marginal zone B cell involvement stems from the increased risk of IPD in individuals with congenital, acquired or functional asplenia [73, 74]. Moreover, T- independent antigen immunization of hyper-igm patients lacking functional CD40L, 10
34 produced peripheral blood circulating B cells genetically similar to marginal zone B cells [75, 76]. B-1 cells are considered to be part of the innate immune system [10]. They are identified in mice by the expression of several activation markers, including B220, CD19, CD27, CD43, and CD5 [10]. B-1 cells can produce natural widely-cross-reactive antibodies, which can bind to PPS [77-80]. Mice that lack B-1 cells are more susceptible to pneumococcal infection [75, 81]. The human equivalent of mouse B-1 cells is highly controversial. Identifying phenotypic markers that conclusively identify this population has been highly debated [30, 82-84]. B-2 cells, which require T cell help, have not been shown to respond well to TI-2 antigens. 1.4 Medical management Antibiotics Penicillin has been the gold standard treatment for many years. However, increased use of penicillin and subsequent antibiotics has led to increased antibiotic resistance [1-5]. Up to 40% of IPD isolates in the United States are resistant to penicillin [17]. Pneumococci are naturally transformable and may acquire new genes from free DNA found in the environment. This leads to the ability to acquire new antibiotic resistant genes [21]. It is common to isolate strains resistant to multiple antibiotics, including penicillin, erythromycin, and trimethoprim-sulfamethoxazole [21]. Vancomycin resistance has been rare, but is becoming increasingly more common, emphasizing the need for new therapeutics and improved vaccination strategies, especially for high risk populations like the elderly and HIV-positive individuals [85]. 11
35 1.4.2 Pneumococcal polysaccharide vaccine Vaccines targeted against the pneumococcal polysaccharide (PPS) capsule have been developed to reduce disease burden since 1977 [17]. The 23-valent PPS vaccine (PPV23) released in 1983 contains PPS from the 23 most common serotypes clinically identified in patients with IPD. These serotypes account for 88% of bacteremic pneumococcal disease [17]. A single intramuscular injection results in elevated PPSspecific IgM and IgG opsonophagocytic antibodies detectable for at least 5 years in healthy individuals [86-90]. High risk individuals show a more rapid decline in antibody levels [17]. Although vaccination with PPV23 does not appear to reduce pneumococcal carriage or incidence of pneumococcal related pneumonia, it is 60-70% effective in reducing IPD [17, 91, 92]. The Advisory Committee on Immunization Practices (ACIP) recommends vaccination for individuals who are at high risk for infection [6]. This includes, but is not limited to the elderly (>65 years old) and HIV-infected [6]. PPV23 is not recommended for young children, as they do not respond well to PPS vaccination. In order to improve vaccination, a more detailed understanding of the specific immune response to PPV23 is needed Pneumococcal conjugate vaccine Pneumococcal polysaccharide conjugate vaccines (PCV) have also been developed. These vaccines were originally developed for children who are unable to respond to TI-2 antigens such as purified pneumococcal polysaccharide. PCV contain 12
36 purified pneumococcal polysaccharide that have been chemically conjugated to a nontoxic diphtheria toxin, CRM 197, which serves as an adjuvant capable of eliciting a T cell dependent response [17]. It is recommended that children between 2-59 months old receive three doses of PCV[17]. A clinical trial with the 7-valent PCV showed a 97% reduction in vaccine serotype related IPD. Evidence also suggests PCV7 reduced carriage of related serotypes [17]. Six additional serotypes were added to PCV7 to produce PCV13. PPS-specific OPT levels to PCV13 were similar to those produced by PCV7 and also provided protection to IPD [93]. The ACIP has recently recommended the 13-valent pneumococcal conjugate vaccine for adults with immunocompromised conditions [6]. Data demonstrating the efficacy of PCV13 compared to PPV23 in adults is limited and requires further study. 1.5 PPV23 in high-risk populations Old age (>65 years) Elderly individuals do not respond to PPV23 as well as young adults and are at increased risk for IPD [92, ]. Multiple factors may contribute to this increased risk, including loss of B cells, decreased antibody levels, and loss of functional antibodies. Total IgM+ memory B cells decrease with increasing age [102, 103]. B-1 cells in mice and their putative human counterparts also decrease in number with age [29, 30, 104]. However, these studies are limited, in that they do not specifically consider changes in the PPS-specific B cell populations. Vaccination of elderly individuals produces significantly lower levels of IgM [25-28]. Elderly individuals also show significantly reduced opsonophagocytic activity and IgG antibody avidity [22-24]. Studies comparing 13
37 changes in elderly PPS-specific B cell populations with antibody production and functional activity are needed to more clearly identify possible reasons for decreased vaccine efficiency and increased disease incidence HIV-positivity HIV disease progression is characterized by the loss of CD4+ T cells. Individuals with CD4 counts <200 cells/ l (AIDS status) are at higher risk for opportunistic infections, including by pneumococcus. The HIV-positive also demonstrate severe B cell dysfunction which begins during early infection steps [ ]. HIV virion gp120 acts as a B cell mitogen, inducing hypergammaglobulinemia, high rates of spontaneous antibody production, and overexpression of activation markers [ ]. This nonspecific activation can result in autoimmune disorders, B cell lymphomas, and B cell apoptosis effectively reducing the diverse B cell repertoire. All HIV-positive individuals are recommended to receive PPV23 to decrease pneumococcal infection [112]. However, despite HIV induced B cell activation, HIVpositive individuals respond poorly to vaccination, in part, due to reduced B cell numbers and response to stimulus [109]. HIV-positive individuals with low CD4 counts (<200 cells/ l) produce lower levels of PPS-specific antibodies [ ] and are functionally deficient [ ]. Previous studies have shown correlations between low levels of peripheral blood B cell populations and low antibody titers [127, 128], however, these studies are limited because they were not antigen-specific. Changes in the minority ~1-4% PPS-specific B cell populations in the peripheral blood are masked by the total B cell 14
38 population [129]. A more detailed understanding of PPS-specific intrinsic B cell defects is needed to understand how to reduce disease burden HIV-positive individuals HAART-treated patients HIV-positive individuals are recommended to receive HAART for viral suppression and immune recovery [130]. The use of antiviral therapies has led to a substantial decrease in opportunistic disease incidence [131, 132]. Individuals on HAART frequently obtain partial immune restoration. ND-HIV with CD4<200 have severely dysfunctional immune systems and uncontrolled high viral titers [133]. These individuals are frequently seen in medical treatment facilities. It is unclear if they would benefit from 6-12 months of HAART to suppress viral loads and allow partial recovery of their immune system prior to PPV23 administration. Total B and T cell numbers increase in HIV-positive individuals on long-term HAART [130, 134]. Conversely, some studies have shown that increases in CD4+ T cells resulting from HAART were insufficient to return normal IgM and switched memory B cell function in HIV-positive individuals [109, 127, 128, 135]. However, these studies are limited in that they do not examine PPS-specific B cell populations. It is unclear if longterm HAART results in the restoration of PPS-specific B cell population numbers or function in HIV-positive individuals which have previously had CD4<200. HIV-positive individuals on long-term HAART remain at high risk for acquiring IPD and pneumococcal antibody titers decrease within 5-10 years [4, 17, 136]. Therefore, HIV-positive individuals on HAART are also recommended to be revaccinated with PPV23 5 years after their initial vaccination [6, 17, 112]. However, the benefits of 15
39 revaccination in this population are unclear [17]. No studies have shown that revaccination is beneficial. Correlations between antibody levels and IPD incidence tend to be weak, and TI-2 antigens do not demonstrate a boosted anamnestic response [17, ]. Even in HIV-negative individuals, the benefits of revaccination with PPV23 remain unclear. While some reports have reported significant increases of PPS-specific antibody levels in HIV-negative individuals after revaccination, others suggested that revaccination may tolerize patients to specific PPS [ ]. It would be beneficial to determine if HIV-positive individuals who have previously had CD4<200 benefit from revaccination after 5 years of HAART, and to define the effect of HAART on B cell reconstitution. 16
40 Chapter 2 The phenotype of PPS-specific B cells in adult peripheral blood. The majority of the following was recently published in Vaccine (Appendix A)[144]. Authors retain the permission to use in author s dissertation. I was first author, and made contributions in volunteer recruitment/enrollment, blood sample processing, cellular staining, flow cytometry processing, processing ELISA, assisting with OPA, data processing, analysis and interpretation, and writing of the manuscript. 2.1 Chapter introduction Increased antibiotic resistance among many Streptococcus pneumoniae serotypes associated with disease, including pneumococcal polysaccharides 14 and 23F (PPS14, PPS23F), emphasizes the need for improved vaccine strategies, especially for those at highest risk for invasive disease including elderly and immunocompromised individuals [1-5]. Vaccination results in PPS-specific IgM and IgG opsonophagocytic antibodies (Abs) which are critical for bacterial clearance [86-90]. The nature of the immune cells involved in the production of antibody against these T-independent Type II polysaccharide antigens is controversial. Splenic marginal 17
41 zone B cells (MZB) produce recirculating plasmacytes and memory B cells, capable of rapidly producing opsonizing IgM and IgG Abs against TI-2 antigens [11, 76, 87, ]. The role of MZB in response to TI-2 antigens is also supported by the finding that individuals who respond poorly to pneumococcal vaccinations tend to lack IgM+ memory B cells. This includes patients with congenital neutropenia, common variable immunodeficiency, HIV infection, have been splenectomized, infants <2 years old with an underdeveloped marginal zone, and elderly populations [76, 102, 103, 128, 145, 147, 149]. Alternatively, B-1 cells have also been implicated in the production of plasmacytes and memory B cells capable of rapidly producing IgM and IgG Abs against TI-2 antigens [11, 81, 87, ]. Previous studies demonstrate mouse B-1 cells transferred into RAG -/- mice produce PPS-specific Abs and provide protection against lethal challenge [81, 150]. While it is thought that B-1 cells contribute to the immune response against pathogens expressing TI-2 antigens in humans, the direct relevance of B-1 cells has been unclear due to the difficulty in identifying human B-1 cell equivalents. In mice, B-1 cells can be divided into two subtypes, B-1a and B-1b cells. B-1b cells have the ability to produce Abs that can provide a long-term adaptive immune response to TI-2 antigens like polysaccharides [10, 81, 150, 151, 154]. Human B-1 cells on the other hand are controversial themselves. It is unclear if the same division of B-1 cells that exists in mice is recapitulated in humans. Previous studies demonstrate that CD5 expression on human B cells is insufficient to characterize B-1 cells as it is used in mice [10, 30, 155, 156]. Recent publications have described a mouse B-1a like subset in humans [10, 30]. It is currently unclear if there is a mouse B-1b like equivalent in humans capable of 18
42 responding to TI-2 antigens such as those used for pneumococcal polysaccharide vaccination (PPV). We have previously shown, using fluorescently labeled PPS14 and PPS23F, the majority of PPS-specific B cells responding to vaccination are IgM+ memory cells (CD27+IgM+) [129]. The goal of the present study was to further characterize PPSspecific PPV responding cells with respect to expression of CD43 and CD5 used to characterize this putative B-1 cell population. Our results identify PPS14- and PPS23Freactive B cell populations that circulate in the peripheral blood 7- and 30-days postimmunization in response to PPV. We show the majority of PPS-specific B cells on day- 7 are phenotypically characterized as CD19+CD20+CD3-CD70-CD27+IgM+CD43+ CD5+/-. This population is in alignment with recent reports of human B-1 cells [30, 82, 83, 157]. We also show that 30 days post-immunization, this population recedes towards pre-immunization levels. 2.2 Results Antibody titers increase significantly post-immunization with PPV To show that our young healthy donor population (n=17) responded normally to immunization with PPV, we tested sera collected on day-0, day-7, and day-30 for PPS14 and PPS23F antibody responsiveness. Following the World Health Organization recommendations, sera were first absorbed with PPS22F and cell wall polysaccharide to prevent overestimation of PPS-specific antibody concentrations [ ]. Day-30 postimmunization, donors showed a significant increase in PPS14-specific IgM from 2.4 ±2.4 g/ml to 32.7 ±18.0 g/ml (p<0.001), IgG from 3.3 ±2.5 g/ml to 47.1 ±24.0 g/ml 19
43 (p=0.0023), and IgA from 0.3 ±0.2 g/ml to 2.7 ±2.9 g/ml (p=0.0056) (Figure 2.1A). Similarly, post-immunization PPS23F-specific antibody levels were significantly increased for IgM from 1.2 ±0.8 g/ml to 22.3 ±7.9 g/ml (p<0.0001), IgG from 2.8 ±2.3 g/ml to 31.9 ±18.9 g/ml (p<0.0001), and IgA from 0.2 ±0.1 g/ml to 0.9 ±1.1 g/ml (Figure 2.1B). IgG concentrations showed a greater increase post-immunization compared to IgM concentrations for both PPS. These ELISA data confirm that our donor population responded to PPV immunization resulting in a minimal two-fold increase in serotype-specific Ab. 20
44 Figure 2.1: Polysaccharide specific Ig and opsonophagocytic titers increase postimmunization with PPV. Healthy young volunteers were immunized with PPV. Serum samples were obtained on days 0, 7, and 30. Serum samples were tested for PPS14- specific (A) and PPS23F-specific (B) IgM, IgG, IgA concentrations by ELISA expressed as μg/ml. Serum samples from day-0 and day-30 were tested for S. pneumoniae PPSspecific antibody opsonophagocytic activity (C) expressed as opsonophagocytic titer. Mean values with standard error of the mean (SEM) shown. *p < 0.05, **p < 0.01, ***p <
45 2.2.2 OPT increases significantly 30 days post-immunization with PPV Opsonophagocytic assays (OPA) test the functional ability of total serum antibodies to activate complement to induce opsonization of serotype-specific pneumococci by phagocytic cells. Donor sera collected on day 0 and day 30 were tested for functional opsonophagocytic response using both PPS14 and PPS23F expressing S. pneumoniae. The reciprocal of the antibody dilution required to obtain 50% opsonophagocytic killing by differentiated HL-60 cells (opsonophagocytic titer-opt) was calculated. There was a significant increase in the OPT post-immunization compared to pre-immunization for both PPS14 (p=0.0001) and PPS23F (p<0.0001) (Figure 2.1C). These results confirmed that vaccination of our sample population with PPV elicited a functional immune response against serotype-specific PPS [161] PPV induces transient increase in PPS-specific B cells in peripheral blood To characterize the phenotype of B cells that responded to vaccination with PPV, donor peripheral blood samples were analyzed pre- and post-immunization. The percentage of PPS14-selected B cells increased significantly (p=0.001) from an average of 1.5% on day-0 to 5.0% by day-7 and decreased to 3.5% by day-30. Similarly, PPS23Fselected B cells increased significantly (p=0.001) from an average of 1.1% on day 0 to 4.2% by day 7 and decreased significantly (p=0.034) to 2.0% by day-30 (Table 2.1). The percentage of PPS14+ B cells on day-30 remained statistically higher (p=0.002) compared to pre-immunization levels. In contrast, the percentage of PPS23F+ B cells returned to pre-immunization levels. 22
46 Table 2.1: CD19+PPS+ B cells in young healthy individuals PPS+ B cells %PPS14+ day 0 %PPS14+ day 7 %PPS14+ day 30 %PPS23F+ day 0 %PPS23F+ day 7 %PPS23F+ day 30 Mean SEM Mean percentage ± SEM of CD19+ B cells stained with fluorescently labeled PPS14 or PPS23F from peripheral blood samples obtained pre-immunization (PPS14 n=10, PPS23F n=11) and 7 (PPS14 n=16, PPS23F n=17) and 30 (PPS14 n=10, PPS23F n=10) (Reproduced from D.J. Leggat et al, Vaccine, ) Majority of PPS-specific B cells express CD27 and IgM post-immunization with PPV Pre-immunization, the phenotypic analysis of CD27 and IgM for PPS-selected and unselected cells were both similar to our previously published results demonstrating B cells pre-immunization primarily consisted of naïve B cells (38-59%) [129]. Our gating strategy for flow cytometry with respect to CD27 and IgM is depicted in Figure 2.2A. PPS-selection day-7 post-immunization revealed a highly significant decrease in representation of naïve B cells (PPS14 p=0.0056, PPS23F p<0.0001). These changes were coupled with highly significant increases in IgM+ memory cells (PPS14/23F p<0.0001) compared to pre-immunization (Figure 2.3A,B). The majority of PPS-selected B cells (PPS %, PPS23F 62.4%) day-7 post-immunization were IgM+ memory cells (Figure 2.3, 2.2A). This population was significantly higher (PPS14/23F p<0.0001) 23
 (Figure 2.3C). Figure 2.2. Phenotypic analysis of human peripheral blood B cells by flow cytometry.")
47 than the unselected population post-immunization. In contrast, day-7 post-immunization class-switched memory cells represented a smaller portion of the PPS-selected B cells (PPS %, PPS23F 22.5%) (Figure 2.3C). Figure 2.2. Phenotypic analysis of human peripheral blood B cells by flow cytometry. Lymphocyte-enriched peripheral blood samples were stained with immunofluorescent PPS and antibodies to be evaluated by flow cytometry. Lymphocytes were plotted (FSC-A, FSC-H) for doublet discrimination. Singlet lymphocytes were plotted on a histogram to gate on B cells (APC-Cy7:CD19+). B cells were plotted using either Cascade Blue:PPS14 or 5-DTAF:PPS23F to identify PPS-selected verses unselected cells. Each of these two populations were subdivided into four subpopulations: naïve (CD27-IgM+), class-switched (CD27-IgM-), IgM+ memory (CD27+IgM+), and class-switched memory (CD27+IgM-) B cells. We further sub-gated each of these populations based on CD43 and CD5 expression. All flow cytometry results were analyzed and plotted using Fluorescence Minus One controls (FMO). Representative CD27 vs. IgM plot (A) and CD43 vs. CD5 plots (B) for each CD27 vs. IgM quadrant shown. 100,000 events were recorded. 24

48 Figure 2.3. Majority of PPS-specific B cells express CD27 and IgM postimmunization with PPV. Lymphocyte enriched peripheral blood samples were stained with immunofluorescent PPS and antibodies to be evaluated by flow cytometry. The phenotype distribution of CD27 and IgM on CD19+ B cells was compared for day-0, day-7, and day-30 for PPS14- (A) and PPS23F- (B) selected cells. The phenotypic distribution of CD27 and IgM was compared on day-7 for unselected, PPS14- and PPS23F- selected cells (C). Mean values with SEM shown. In each sample, 100,000 events were recorded. *p < 0.05, **p < 0.01, ***p< Day-30 post-immunization the phenotypic distributions of PPS-selected cells returned towards pre-immunization levels (Figure 2.3A,B). PPS14-selected naïve cells on 25
49 day-30 showed a significant increase compared to day-7 (p=0.0001), returning to preimmunization levels. In contrast, IgM+ memory cells decreased significantly (p<0.0001) to pre-immunization levels by day-30. PPS14-selected cells on day-30 showed no significant difference for naive and IgM+ memory cells compared to day-0 (Figure 2.3A). PPS23F-selected cells showed similar trends. PPS23F-selected naive cells increased significantly (p=0.0461) toward pre-immunization levels by day-30, but remained significantly (p=0.0413) lower compared to day-0. In contrast, PPS23F-selected IgM+ memory cells decreased significantly (p=0.0002) toward pre-immunization levels by day- 30, but remained significantly higher compared to day-0 (p=0.0151) (Figure 2.3B). Therefore, by day-30, the distribution of PPS-selected cells returned towards preimmunization phenotype distributions. Strong positive correlations were found between the percentage of PPS-selected IgM+ memory cells on day-7 and IgM antibody concentrations on day-30 for both PPS14 (r 2 =0.87, p<0.0001) and PPS23F (r 2 =0.98, p<0.0001) (data not shown). In contrast, correlations were not found between the percentage of PPS-selected class-switched memory cells on day-7 and IgG antibody concentrations on day-30 for either PPS14 (r 2 = , p=0.2268) and PPS23F (r 2 = , p=0.1317) (data not shown) Majority of PPS-specific IgM+ memory B cells express CD43 and CD5 postimmunization with PPV We further characterized peripheral blood for CD43 and CD5 expression within each of the previously identified CD27 and IgM quadrants (Figure 2.2B). Preimmunization, within the CD27+ populations, CD43 and CD5 analysis showed no 26
50 significant difference between selected and unselected cells (data not shown). Within the PPS-selected IgM+ memory populations, there were significant increases in CD43+CD5+ expression on PPS-specific cells from day-0 to day-7 (PPS14: 27.7%- 56.5% p=0.027, PPS23F: 31.7%-63.8%, p=0.005) (Figure 2.4A,B). There were no significant changes within the PPS-selected class-switched memory populations from day-0 to day-7 (Figure 2.4C,D). On day 7 post-immunization, the majority of PPSselected IgM+ memory cells were CD43+CD5+ (PPS %, PPS23F 63.8%). This was in contrast to the unselected IgM+ memory cell population which showed the majority of cells were CD43-CD5- (52.9%), while the CD43+CD5+ population represented only 16% of these cells. The percentage of PPS-selected CD27+IgM+CD43+CD5+ cells were significantly higher (PPS14 p<0.0001, PPS23F p<0.0001) compared to analysis using unselected cells (Figure 2.4E). Within the classswitched memory PPS-selected population, significant changes between unselected and PPS-selected cells were only found in response to PPS23F, revealing a significant (p<0.0001) increase in CD5+CD43+ cells paired with a significant (p<0.0001) decrease in CD5-CD43- cells (Figure 2.4F). Thus, 7 days post-immunization the majority of PPS+ peripheral blood B cells responding to PPV were CD27+IgM+CD43+CD5+. 27

51 Figure 2.4. Majority of PPS-specific IgM+ memory B cells express CD43 and CD5 post-immunization with PPV. Lymphocyte enriched peripheral blood samples were stained with immunofluorescent PPS and antibodies to be evaluated by flow cytometry. The phenotypic distribution of CD43 and CD5 on B cells within the IgM+ memory and class-switched memory populations was compared for day-0, day-7, and day-30 for PPS14- (A,C) and PPS23F (B,D) selected cells. The phenotypic distribution of CD43 and CD5 on B cells within the IgM+ memory (E) and class-switched memory (F) populations was compared on day-7 for unselected, PPS14- and PPS23F- selected cells. Mean values with SEM shown. In each sample, 100,000 events were recorded. *p < 0.05, **p < 0.01, ***p<
52 Day-30 post-immunization, within the PPS-selected IgM+ memory populations, the phenotypic distributions of CD43 and CD5 returned toward pre-immunization levels. For PPS14-selected IgM+ memory cells, the predominant (56.5%) CD43+CD5+ population on day-7 decreased significantly (19.3%, p=0.0005) to day-0 levels. At the same time within this population, the CD43-CD5- population which was reduced (20.7%) on day-7 significantly increased (54.7%, p=0.0007) to day-0 levels (Figure 2.4A). For IgM+ memory PPS23F-selected cells the predominant (63.8%) CD43+CD5+ population on day-7 regressed to 43.6%, but remained higher than day-0 levels (31.7%) (Figure 2.4B). Within the class-switched memory population significant changes in PPS-selected cells between day-7 and day-30 were only seen within the PPS14-selected CD5 - populations. However, the distribution of CD43 and CD5 did not show any statistical significant differences between day-0 and day-30 (Figure 2.4C,D). Therefore, by day-30, the distribution of PPS-selected cells resembled pre-immunization phenotype distributions with no significant differences PPS-selected B cell percentages and absolute numbers We compared the percentages and absolute numbers of PPS14 and PPS23Fspecific B cells with respect to CD27 and IgM in Table 2.2 and Table 2.3 respectively. Data is listed as mean, range, and standard error mean. IgM+ memory and switched memory populations where also compared with respect to CD43 and CD5. 29
53 Figure 2.2. Percentages and numbers of PPS14 seleced B cells. 30
54 Figure 2.3. Percentages and numbers of PPS23F selected B cells. 31
55 2.2.7 PPS-selected B cells phenotypically resemble human B-1 cells Recent reports have expressed interest in distinctly identifying B cells from T cells or B/T cell doublets due to the heterogeneic distribution of CD43 and CD5 on T cells [82]. CD3 analysis showed that ±2.88% of our day-7 CD19+ cells did not express CD3 and therefore did not represent T cells or B/T cell doublets (Figure 2.5A). To clarify the proportion of day-7 CD19+ cells that could represent CD19+CD20- plasmablasts, we analyzed CD20 expression. Within the CD19+ cells, the vast majority of cells (95.0 ±1.87%) were also CD20+ (Figure 2.5B). To demonstrate that our PPSselected cells do not represent IgM+ memory cells expressing CD43 and CD5 as inducible activation markers, and to allow direct comparison to recent studies, we analyzed CD70 expression [30, 83]. Within the CD19+ cells, ±1.32% of PPSselected, and ±1.07% of our PPS-selected CD27+IgM+CD43+CD5+ populations 7 days post-immunization were also found to be CD70 - (Figure 2.5C). 32
56 Figure 2.5. PPS-selected B cells phenotypically resemble human B-1 cells. Four healthy human donors were immunized with PPV. Lymphocyte enriched peripheral blood samples were stained with immunofluorescent PPS and antibodies to be evaluated by flow cytometry on day-7. Representative plot of CD3 expression on CD19+ B cells (A). Representative plot of CD20 expression on CD19+ B cells (B). Representative plot of PPS23F-selected IgM+ memory cells gated for CD43+ and CD5+ expression and plotted for CD43 and CD70 expression (C). All flow cytometry results were analyzed and plotted using Fluorescence Minus One controls (FMO). 100,000 events were recorded. 33
57 2.3 Chapter discussion Polysaccharide-specific ELISA and opsonophagocytic assays demonstrated that our volunteers were immunocompetent and responded normally to PPV. Samples showed variability in antibody concentrations and OPT between individuals and between serotypes. Others have also shown that a strong antibody response to one serotype does not necessarily correlate with a strong response to other serotypes [129, 161, 162]. The B cells responsible for eliciting protective Abs in response to PPV vaccination remains an active and controversial area of research. The goal of our study was to further characterize the phenotype of PPS-specific B cells that respond to PPV vaccination in healthy individuals using fluorescently conjugated PPS which we have previously described [129]. Day-7 post-immunization, the percentage of PPS-selected cells increased significantly compared to day-0 background levels. By day-30, the percentage of PPS-selected cells declined toward pre-immunization levels. Thus, at its peak, a mere 4-5% of B cells were PPS-specific emphasizing the importance of PPSselection to analyze changes in cell populations responding to PPV as shown by day-7 post-immunization unselected versus PPS-selected results. PPS-selection on day-7 demonstrated a predominant IgM+ memory population in response to PPV clearly distinguished from the unselected phenotype analysis as in our previous study [129]. These changes reverted toward pre-immunization levels by day-30 post-immunization. Interestingly, PPS23F-selected IgM+ memory cells showed a significant decrease by day- 30, but remained significantly higher than day-0 levels. In contrast, PPS14-selected IgM+ memory cells on day-30 showed no significant difference compared to day-0 levels. Others have also reported a heterogeneous response to various PPS [163]. Some 34
58 differences may be due to PPS structural differences, previous donor exposure to specific serotypes, and/or pre-immunization antibody concentrations. The CD27+IgM+ B cell population can further be subdivided into IgM+ memory B cells (CD27+IgM+CD43-) and B-1 cells (CD27+IgM+CD43+). In mice, B-1 cells have been extensively characterized, are subdivided into B-1a and B-1b cells, and are vitally important in the protective immune response against S. pneumoniae. Griffin et al recently characterized a human equivalent of B-1 cells. The role of these cells in the human immune response to PPV remains to be defined [30, 81, 87, 151]. We analyzed the presence of CD43 on the surface of PPS-labeled CD27+IgM+ B cells as percentage of PPS-positive B cells classified as B-1 cells (CD43+) or those that are IgM+ memory cells (CD43-). The CD43 and CD5 sub-gated populations of unselected cells showed no significant differences in phenotype distribution between day-0, 7, and 30 analysis (data not shown). This is expected since the PPS+ B cell population is a small fraction of total B cells. However, within the PPS-specific IgM+ memory population, the percentage of CD43+CD5+ B cells increased significantly by day-7 post-immunization, and reverted toward pre-immunization levels by day-30. We also demonstrated strong correlations between the proportion of day-7 PPS-selected IgM+ memory B cells and day-30 postimmunization PPS-specific IgM concentrations. This suggests that the percentage of PPS-specific IgM+ memory cells on day-7 can serve as a surrogate marker for PPV responsiveness. The majority population responding to PPV is in alignment with recent descriptions of putative human B-1 cells and includes a population capable of producing anti-polysaccharide antibodies [30, 83]. To determine to what degree this PPS-selected 35
59 majority population resembled the putative B-1 population, we performed additional phenotypic analysis for CD3, CD20, and CD70. The vast majority of these cells didn t represent CD3+ T cells, CD19+CD20- plasmablasts, or CD70+ activated memory and naïve cells expressing CD43 and CD5. This analysis reveals additional populations of PPS-specific B cells, demonstrating that the immune response to PPV is in fact a heterogeneous and complex orchestration of many cell types and does not solely consist of the majority B-1 like population. Verbinnen et al have shown that the putative B-1 cell population described by Griffin et al were capable of producing PPS-specific IgM and IgG detected by ELISPOT for PPS1 and PPS4 [30, 83]. It is therefore possible that our CD27+IgM- B cells may have also represent class-switched B-1 cells which have been described by Hass et al in mice [152]. Additional future studies like the ELISPOT performed by Verbinnen et al and the spontaneous Ig secretion performed by Griffin et al can determine if this putative PPSspecific B-1 like population has the same functional characteristics as B-1b cells. Moreover, other populations of B cells may contribute to the immune response within different time frames and in other compartments such as the spleen or bone marrow. However, the significant increase in this B-1 like population, regardless of its classification, over a relatively short-time in the peripheral blood suggests that this population may have great importance in controlling the early stages of infection just as B-1b cells have been shown to function [11]. The relatively quick regression of this majority population from the blood suggests likely sequestration in other compartments. The majority of PPS14- and PPS23F-selected B cells responding to PPV in the peripheral blood was characterized as CD19+CD20+CD3-IgM+CD27+CD43+CD5+/- 36
60 CD70-. This analysis in young adults lays a foundation against which comparisons can be made for high risk populations, including the elderly and HIV-infected individuals, which we are currently pursuing [144]. We hypothesize that high risk populations may show a very different distribution of phenotypes compared to the healthy individuals. Identifying changes or deficiencies in the PPS-specific responding B cell populations of elderly and HIV-infected individuals may be used to improve vaccination schedules and develop new therapies leading to improved patient protection. 37
61 Chapter 3 Elderly immune response to pneumococcal polysaccharide vaccination The majority of the following was recently published in Journal of Infectious Disease (Appendix B)[164]. Authors retain the permission to use in author s dissertation. I was co-first author and made contributions in volunteer recruitment/enrollment, blood sample processing, cellular staining, flow cytometry processing, processing ELISA, assisting with OPA, and data processing. 3.1 Chapter introduction Streptococcus pneumoniae is a major cause of morbidity and mortality worldwide. The highest risk groups are young children, elderly adults (> 65), and immune-compromised individuals. Despite the high efficacy of the pneumococcal polysaccharide vaccine in young adults, it is less protective in populations at highest risk for invasive pneumococcal disease, including the elderly [ ]. Post-vaccination with PPV, the elderly produce anti-pps IgG antibodies in similar concentrations to young adults [169, 170]. However, decreased opsonophagocytic activity (OPA), restricted variable gene usage and lower IgM concentrations may all play 38
62 a role in the vaccine s decreased efficacy [102, 103, 161]. The underlying cause of these differences remains to be elucidated. The phenotypes of the B cells responsible for the immune response to PPV has been controversial. Post-stimulation in vitro, anti-pps Abs are secreted mainly by switched memory cells (CD27+IgM-) and are predominately IgG and IgA [ ]. However individuals with reduced numbers or absence of CD27+IgM+ B cells respond poorly to PPV and are more vulnerable to infections caused by encapsulated bacteria [76, 102, 103, 146, 147, 174]. We recently described a highly significant increase in the CD27+IgM+ PPSspecific B cell population 7 days post-vaccination in healthy young adults [129]. The specific aim of this study was to identify the phenotype using CD19, CD27 and IgM markers and enumerate PPS-specific B cells in the elderly. Our study demonstrates a significant change in the phenotype of PPS-responding B cells, PPS-specific IgM and OPA, post-vaccination in the elderly compared to young adults. These findings may help explain the decreased efficacy of the purified PPV in individuals over the age of Results Elderly serum antibody titers increased post vaccination To study the elderly, PPS-specific immune response to PPV, we obtained pre- and day-30 post-vaccination sera and measured antibody responses to pneumococcal serotypes 14 and 23F. Elderly donors had a significant rise in concentration of PPS14- specific IgG increasing from g/ml to g/ml (p<0.001) post-vaccination. 39
63 There was no significant increase in PPS14-specific IgM or IgA antibody concentration post-vaccination. Similarly, post-vaccination IgG responses to PPS23F were significantly increased compared to pre-vaccination sera, from g/ml to g/ml (p<0.004). There was a significant increase in PPS23F-specific IgM (p<0.03) but not in IgA postvaccination (Figure 3.1). In our sample population there was an increase in IgG concentration for both PPS14 and PPS23F and an increase in IgM concentration for PPS23F only, pre- to post-vaccination. All donors displayed an increase in serotypespecific IgG antibody response, with the exception of one donor for PPS14. 40
64 Figure 3.1: Polysaccharide-specific antibody titers increase post-immunization with PPV. Elderly volunteers were immunized with PPV. Serum samples were obtained on days 0 and 30. Serum samples were tested for PPS14-specific and PPS23F-specific IgM, IgG, IgA concentrations by ELISA expressed as μg/ml. 41
65 3.2.2 Elderly serum opsonophagocytic antibody titers increased post-vaccination The functional or opsonophagocytic response of serum antibody obtained prevaccination and 30 days post-vaccination against both serotype 14 and 23F PPS was determined for all donors. Data was reported as opsonophagocytic antibody titers, the antibody dilution required to obtain 50% opsonophagocytic killing by differentiated HL- 60 cells. Overall, post-vaccination sera showed a significant increase in opsonophagocytic antibody titers against both serotype 14 and serotype 23F compared to pre-vaccination sera (p<0.01) (Figure 3.2). All, except three donors demonstrated a minimal two-fold increase in opsonophagocytic antibody titers post-vaccination. There was a good correlation between the anti-pps IgG concentration and opsonophagocytic antibody titer (r 2 =0.8, p= for PPS14 and r 2 =0.863, p= for PPS23F) but not with the IgM concentration. Figure 3.2: Polysaccharide-specific opsonophagocytic titers increase postimmunization with PPV. Elderly volunteers were immunized with PPV. Serum samples from day-0 and day-30 were tested for S. pneumoniae PPS-specific opsonophagocytic titer. 42
66 3.2.3 Unselected B cell counts are decreased in elderly peripheral blood Peripheral blood lymphocytes (PBL) were isolated and the percentage and absolute number of B cells was determined for elderly and compared to data we obtained from our previous study in young adults (Table 3.1). The percentage of B cells in elderly was 7.84% with an absolute B cell count of 149,300 (±40,950) per ml significantly lower (p<0.01) than the B cell percentage of 18.67% and absolute B cell count of 368,100 (±65,400) per ml in the young adult population. Table 3.1. Unselected CD19+ B cell numbers Parameters Young (SEM) Elderly (SEM) p value CD19+B cell % (±1.74) 7.84 (±1.89) CD19+ B cell number 368,100 (±65,400) 149,300(±40,950) 0.01 The percentage and absolute number of unselected CD19+ B cells in elderly and young adults. The percentage and absolute numbers of B cells (per/ l) are significantly higher in young adults compared to elderly Phenotype of polysaccharide-specific B lymphocytes isolated from elderly PBMC Pre-vaccination and day-7 post-vaccination, PBMC were isolated, labeled, and subjected to flow cytometry. The phenotype of pre- and post-vaccination PPS-specific B cells was compared to the phenotype of unselected B cells and our historical young adult controls [129]. B cells (CD19+) were subdivided into four categories: naïve (CD27- IgM+), class-switched (CD27-IgM-), CD27+IgM+ and switched memory (CD27+IgM-) B cells. 43
, of CD27- B cells expressed the naïve phenotype, whereas a minority, 10.7% (6.7-28%) represented class-switched B cells. The memory B cell population represented 33.6% of B cells with 17.7% (4.")
67 Analysis of unselected B cells obtained pre-vaccination from the elderly showed that a large proportion, (68.5%) of B cells were CD27 negative. The majority, 57.8% ( %), of CD27- B cells expressed the naïve phenotype, whereas a minority, 10.7% (6.7-28%) represented class-switched B cells. The memory B cell population represented 33.6% of B cells with 17.7% ( %) expressing the IgM+CD27+ phenotype, and 15.9% ( %) switched memory phenotype (Figure 3.3). Figure 3.3. Pre- and post-immunization pneumococcal polysaccharide-selected B cell phenotypes in elderly individuals. The phenotypes of B lymphocytes that respond to vaccination with PPS vaccine were determined by flow cytometry. Before and 7 days after vaccination, circulating peripheral blood mononuclear cells were isolated and labeled fluorescently with different anti-surface marker antibodies. The phenotype of unselected and PPS14- and PPS23F-selected B cells were compared in elderly individuals. In each sample, 100,000 events were recorded. Abbreviation: IgM, immunoglobulin M. Pre-vaccination, 1.85% (±0.21) and 2.01% (±0.38) of B cells stained with fluorescently labeled PPS14 and PPS23F respectively (Table 3.2). In contrast to the unselected B cell population, the pre-vaccination PPS14 and PPS23F-labeled B cells in 44
68 the elderly demonstrated a predominance of memory B cells with a total of 61.3% ( %) for PPS14 and 71.2% ( %) for PPS23F. The memory B cells were equally divided between IgM+CD27+ and switched memory B cells. The IgM+CD27+ population represented 30.8% (8.5-77%) of total B cells for PPS14 (p=0.025) and 29.6% ( %) for PPS23F, not significantly different compared to unselected B cells. The switched memory population represented a total of 30.5% (2.9-56%) and 41.6% ( %) of the pre-vaccination PPS14 and PPS23F-labeled B cells, and both populations were significantly different compared to unselected B cells (p<0.001). The remainder of the B cell population consisted of CD27- B cells accounting for 39.3% of the PPS14- labeled and 28.8% of the PPS23F-labeled B cells, significantly different compared to unselected B cells (p<0.01) (Figure 3.3). Table 3.2. Pneumococcal polysaccharide-selected CD19+ B cell percentages and counts among young adults and elderly individuals. Young Elderly PPS14 PPS23F PPS14 PPS23F % #cells/ l % #cells/ l % #cells/ l % #cells/ l Before 1.5(±1.1) (±0.6) (±0.1) (±0.38) 3.3 After 5.0(±2.8) (±2.9) (±0.6) (±1.03) 7.0 Data denote the mean no. or mean percentage (±standard error of the mean) of CD19+ B cells stained with fluorescently labeled PPS14 and PPS23F in peripheral blood samples. Day 7 post-vaccination, the percentage of PPS-selected B cells increased significantly to 4.60% for PPS14 and 4.65% for PPS23F (Table 3.2). The majority of post-vaccination PPS-labeled B cells were memory B cells (CD27+), accounting for 45
69 70.4% for PPS14 and 78.7% for PPS23F (Figure 3.3). In contrast to pre-vaccination B cell populations, where IgM+CD27+ and switched memory B cell phenotypes were equally divided, switched memory B cells were the predominate phenotype postvaccination. The IgM+CD27+ cells represented 24.1% ( %) of the PPS14-labeled B cells and 25.2% (6-38%) of the PPS23F-labeled B cells, decreased compared to prevaccination levels but not significantly so. The switched memory population represented 46.3% ( %) for PPS14 and 53.5% ( %) for PPS23F, significantly increased compared to pre-immunization levels for both PPS (p<0.05). Similar to prevaccination PPS-labeled B cell populations, the minority of post-vaccination PPS-labeled B cell populations consisted of naive CD27- B cells, 29.5% ( %) for PPS14 and 21.2% ( %) for PPS23F. The naive B cell population consisted primarily of CD27-IgM+ B cells, 23.3% ( ) and 16.6% (0.7-38) for PPS14 and PPS23F, respectively. A small percentage, 6.2% (0-14.9%) and 4.6% ( %), of B cells were naive, class-switched CD27-IgM- B cells. Moreover, the predominant B cell population responding to PPS14 and PPS 23F in this elderly population, 7 days post-vaccination, consisted of CD27+IgM- or switched memory B cells Phenotype of polysaccharide-specific B lymphocytes isolated from elderly PBMC compared to healthy young adults In our previous study [129], we identified the phenotype of B cells responding to PPV seven days post-vaccination in healthy young adults (18-30 years of age). In prevaccination PBMC, 1.1 and 1.2% pre-immunization and 5.0 and 4.2% post-immunization of B cells were labeled with PPS14 and PPS23F; respectively. In the elderly, the pre- and 46
70 post-vaccination percentages of PPS-labeled B cells were similar, namely 1.85% and 2.01% pre-immunization and 4.60% and 4.65% post-immunization (Table 3.2). Figure 3.4 compares the four B cell populations naive, class-switched, CD27+IgM+ and switched memory, isolated from the elderly to those isolated from our previous study of healthy young adults [129]. Pre-vaccination there was no statistically significant differences between the young and elderly individuals in all four B cell sub-populations. Post-vaccination, young adults demonstrated a significant increase in CD27+IgM+ in response to PPS14 and PPS23F. However, in the elderly, the CD27+IgM+ population decreased in response to both polysaccharides with a concomitant increase in the switched memory population. Significant differences were noted in the B cell response to PPS14 and PPS23F between elderly and young individuals. Young adults responded predominantly with a significant increase in IgM+CD27+ B cells (PPS14 from 20.3% to 53.1%, PPS23F from 23.0% to 62.4%). The elderly, in contrast, responded to both polysaccharides with a predominant switched memory B cell response (PPS14 from 30.5% to 46.3%, PPS23F from 41.6% to 53.5%). 47
71 Figure 3.4. Pneumococcal polysaccharide-labeled B cell phenotypes in elderly individuals, compared with young adults. The phenotypes of B lymphocytes that respond to vaccination with PPS vaccine were determined by flow cytometry. Before vaccination and 7 days after vaccination, circulating peripheral blood mononuclear cells were isolated and labeled fluorescently with different anti-surface markers Abs. The phenotype of pre- and post-vaccination PPS14-selected (A) and PPS23F-selected (B) B cells were compared to young and elderly individuals. In each sample, 100,000 events were recorded. 48
72 3.3 Chapter discussion We recently determined that the PPV-responding population in healthy young adults consisted of CD27+IgM+ B cells.[129]. However, the PPV-responding B cell subset for at risk populations such as the elderly had not been previously analyzed. The goal of this study was to characterize the phenotype of responding B cells in the elderly post-ppv vaccination. We analyzed the immune response to PPS14 and PPS23F, as both are present in PPV23, whereas PPS14 represents an immunogenic serotype that elicits excellent antibody responses while PPV23F represents a poorly immunogenic serotype. To ascertain the immune competency of our elderly volunteers and their ability to respond to PPV, we studied pre- and post-vaccination PPS-specific immunoglobulin (Ig) concentration and OPA. Although all individuals, with the exception of one, displayed a significant increase in anti-polysaccharide IgG concentration post-vaccination, the IgM concentration only increased significantly in response to PPS23F. This is in stark contrast to the results obtained in healthy young adults who demonstrated a significant increase in all Ig isotypes post-vaccination for both PPS [129]. Several studies have documented decreased efficacy following PPV with age [166, 175]. Despite adequate antibody concentrations, the OPA is markedly decreased in the elderly [22, 23, 161, 176]. Decreased vaccine efficacy is likely related to alterations in functional antibody activity and specificity rather than IgG antibody concentrations [22, 177]. Nahm and others [102, 161, 178] recently reported significant reduction in PPSspecific IgM response in the elderly. Moreover, depletion of IgM from serum significantly reduced OPA against PPS14 and PPS23F. Others have made similar observations in the response to encapsulated organisms, emphasizing the important role 49
73 of IgM antibodies in regards to protective immunity [22, 176, ]. The results of our antibody and opsonophagocytic studies are thus similar to those previously described demonstrating a loss of IgM antibody and OPA in elderly. We used fluorescently labeled PPS in conjunction with flow cytometry for identification and analysis of PPS-specific B cells. We have previously shown that fluorescently labeled PPS retain their functional activity and specificity.[129]. The use of directly labeled PPS minimizes potential cross-reactivity with linking agents resulting in lower background binding and more accurate phenotype analysis. Phenotypic analysis of unselected CD19+ B cells in the elderly demonstrated a predominance (±68.5%) of naïve B cells. Memory B cells, characterized by the presence of the CD27 marker, represented approximately 34% of PBL with an equal distribution between IgM+CD27+ and switched memory B-phenotypes. In healthy young adults, unselected PBL also predominantly consist of naïve B cells (±68%) and IgM+CD27+ and switched memory B cells each represented ±15% [129]. There was no significant difference between the elderly and the young in memory B cell percentage, however the elderly had a significantly lower absolute number of B cells (Table 3.1), consequently the number of IgM+CD27+ and switched memory B cells was significantly lower. Several investigators have studied IgM+CD27+ B cells in the elderly as loss of IgM+CD27+ B cells is associated with impaired immune responses [78, 103]]. In concordance with our findings, Colonna-Romano and others, have reported no change in IgM+CD27+ B cell percentage with age [104, 182], however, a significant reduction in the absolute number of B cells was reported [182] resulting in decreased number of B cells in all phenotype sub-populations. 50
74 The pre-immunization PPS-selected B cell population, in response to both PPS, consisted predominantly of memory B cells, divided between IgM+CD27+ and switched memory B cell populations, with concomitant decrease in naïve B cell fraction. In contrast, the pre-immune PPS-labeled B cell fraction in young adults closely resembled the unselected B cell phenotype [129] with predominance of naïve B cells. The shift in phenotype from unselected to pre-immunization PPS-labeled B cells and from young to elderly may reflect the magnitude of prior contact, either through natural exposure or vaccination. It should be noted that antibody concentration, opsonophagocytic antibody titers and B cell phenotype distribution was similar between the group of vaccine-naïve and vaccine experienced elderly individuals (data not shown). These data suggest that age is the single most important factor in the shift in pre-immunization phenotype distribution of PPS-labeled B cells. The effect of IgM+CD27+ B cell deficiency in the elderly was accentuated in the post-immunization studies. The majority of PPS-specific B cells post-vaccination expressed the switched memory B cell phenotype. In contrast, the predominant phenotype of PPS-labeled B cell post-vaccination in young adults were IgM+CD27+ (Figure 3.4). The absence of IgM+CD27+ B cells seven days post-vaccination with PPV in the elderly correlated with a lack of anti-pps IgM antibody in serum and diminished opsonophagocytic antibody titer 30 days post-vaccination. Despite the predominance of PPS-specific switched memory B cells post-vaccination, both anti-pps IgG antibody concentration and opsonophagocytic antibody titer were significantly lower in the elderly compared to young adults. Furthermore, there was no correlation between the absolute number or percentage of PPS-specific switched memory B cells and IgG antibody 51
75 concentration. The increase in PPS-specific switched memory B cells does not appear to compensate for the IgM+CD27+ deficiency in terms of functional antibody activity. Most previous studies evaluating post-vaccination alterations in B cell phenotype examine unselected B cell populations [102, 127, 128] rather than antigen selected B cells. Recently, Clutterbuck et al.[183] studied the immune response to either PPV or pneumococcal conjugate vaccine followed by PPV in adults age years. Polysaccharide labeled B cells were obtained using biotinylated PPS/streptavidin beads and cells were analyzed 1 month post-vaccination. This phenotype analysis is not suitable for comparison to our study due to significant differences in B cell capture techniques and timing. We specifically analyzed PBMC seven days post-vaccination at which time the highest number of antigen-specific antibody-secreting cells are found in the peripheral circulation.[184, 185]. We have examined PPS-specific B cells 30 days postvaccination in 16 young adults and 8 elderly adults (unpublished observations). The results of these studies demonstrated that 30 days post-vaccination, antigen-specific B cells are at pre-immunization levels. In addition, the phenotype of PPS-specific B cells was not significantly different compared to pre-immunization phenotype in young, PPVnaïve elderly and previously immunized elderly. These data suggest that 30 days postvaccination, antigen-specific B cells are by and large absent from the peripheral circulation and reside elsewhere in the B cell compartment. Phenotype analysis obtained 30 days post-vaccination was thus similar to that reported by Clutterbuck et al.[183]. However this time point is likely not an accurate representation of the cellular immune response to PPV as it reflects the PBMC rather than bone marrow or spleen, more likely reservoirs of antigen-specific B cells 30 days post-vaccination. We recognize that a 52
76 limited number of elderly individuals were studied and consisted of 10 PPV naïve and 4 PPV experienced individuals (immunized more than 5 years previously). Although the number of individuals enrolled limits statistical power, we found no difference in immune response in the naïve verses experiences groups. These results are supported by recently reported studies of PCV13 in elderly individuals [186]. The data presented demonstrate decreased anti-pps IgM antibody concentration, OPA, absolute and relative numbers of IgM+CD27+ B cells in the elderly compared to young adults. Moreover, clinical studies have established decreased protective efficacy of PPV in elderly individuals [166, 175]. We hypothesize a direct link between these observations, suggesting a critical role of PPS-specific IgM+CD27+ B cells in protective immunity against S. pneumonia. The recent change in pneumococcal vaccination practices will let us study the immune response following pneumococcal conjugate vaccination in elderly and immune-compromised individuals. The analysis of the B cell repertoire will allow us to dissect differences between PCV and PPV responses at a cellular level, providing increased insight into the role of IgM and switched memory B cells in protective immunity. 53
77 Chapter 4 Immune deficiencies in newly diagnosed HIV-infected individuals I made contributions in volunteer enrollment, blood sample processing, cellular staining, flow cytometry, assisting with ELISA, assisting with OPA, data processing, analysis and interpretation, and writing of the manuscript. 4.1 Chapter introduction Newly diagnosed HIV-positive individuals (ND-HIV) are 35 to 100-fold more susceptible to Streptococcus pneumoniae infection. [4, 45, 187, 188]. Pneumococcus is the most common bacterial respiratory pathogen in HIV-positive individuals, and a major cause of morbidity and mortality requiring hospitalized care [42, 189]. Therefore, PPV23 was recommended by the ACIP, though efficacy of vaccination remains controversial [ ]. Antibody response to PPV23 is suboptimal compared to non-infected individuals (HIV-negative), and correlates with immune suppression indicated by CD4 count [192, 193]. Antibiotic resistance among many pneumococcal serotypes, including serotypes 14 and 23F, is increasing among disease-related clinical isolates, emphasizing the need for improved vaccine strategies [1, 194]. Understanding intrinsic B cell defects 54
78 resulting from HIV infection and leading to increased pneumococcal disease incidence is needed to reduce disease burden. To provide better therapeutics, we first need to identify why this population remains at high risk even after immunization. Immunization in HIV-negative individuals results in the production of pneumococcal polysaccharide serotype-specific IgM and IgG. These anti-capsular opsonophagocytic antibodies are critical for bacterial clearance [8, 9]. Early severe B cell dysfunction is a central feature of HIV-positive individuals [135, 190, 195]. Overall, the total number of memory B cells is reduced in HIV-positive individuals [128, 135, 196]. Moreover, anti-pneumococcal IgM and IgG Abs are severely reduced in HIV-positive individuals immunized with the PPV23 compared to HIVnegative individuals [ , 197]. This suggests that HIV-positive individuals lack important PPS-responding B cell components necessary to provide sufficient protection. The specific nature of the immune cells involved in the production of protective antigenspecific Abs in HIV-positive individuals remains to be elucidated. Vaccination recommendations for HIV-positive individuals with CD4<200 remain unclear. It is not known if newly-diagnosed HIV-infected having CD4<200 benefit from delaying immunization to allow 6-12 months of HAART, that can results in viral suppression and partial immune reconstitution. We therefore compared HIVpositive individuals with CD4<200 without HAART (HAART naïve) to HIV-positive individuals with CD4<200 treated for 6-12 months with HAART (HAART treated) to determine possible differences in immunization response as indicated by antigen-specific antibody titers, opsonophagocytic titers, and B cell phenotypes. There were two goals of this study: to clarify whether it is beneficial to provide 6-12 months of HAART to 55
79 suppress viral load and potentially improve T cell and B cell function before PPV23 vaccination in newly diagnosed HIV-positive individuals with CD4<200, and to elucidate the effectiveness of PPV23 in this population. 4.2 Results HIV-positive individuals had diminished PPS-specific antibody titers. To determine the PPS-specific immune response to PPV23 in newly-diagnosed HIV-positive individuals, day-0 and day-30 post-vaccination sera were collected. Following absorption with PPS22F and cell wall polysaccharide (CWPS), samples were tested by ELISA to determine PPS-specific antibody titers. Day-30 post-immunization, group-1 (CD4>200, mean 479) showed a significant increase in PPS14-specific IgM from 0.7 ±0.1 g/ml to 1.5 ±0.3 g/ml (p<0.001) and IgG from 7.0 ±3.2 g/ml to 24.3 ±5.5 g/ml (p<0.001) (Figure 4.1A,B). Volunteers in group-2 (CD4<200, mean 108, no HAART) showed no significant increase in PPS14-specific IgM from 1.0 ±0.3 g/ml to 1.4 ±0.5 g/ml, however, IgG increased significantly from 8.7 ±3.6 g/ml to 33.5 ±12.4 g/ml (p<0.05). Group-3 (CD4<200, mean nadir 93, mean at study 184, 6-12 months HAART) showed no significant increase in either PPS14-specific IgM from 0.2 ±0.0 g/ml to 0.3 ±0.1 g/ml or IgG from 2.7 ±1.2 g/ml to 6.3 ±2.1 g/ml. 56
80 Figure 4.1: HIV-positive serum antibody and opsonophagocytic titers. HIV-positive volunteers were immunized with PPV23. Serum samples were obtained on days 0 (white bars) and 30 (gray bars). Volunteers were classified according to CD4 count <200 or >200 cells. All samples were tested for PPS14 and PPS23F-specific IgG and IgM titers by ELISA expressed as g/ml (A-D) and functional activity by OPA expressed as opsonophagocytic titer (E-F). Data are mean ± standard error measure. *p<0.05, **p<0.01, ***p<
81 Significant increases were seen in group-1 PPS23F-specific IgM from 0.5 ±0.1 g/ml to 0.9 ±0.2 g/ml (p<0.05) and IgG from 0.9 ±0.2 g/ml to 2.5 ±0.8 g/ml (p<0.05) (Figure 4.1C, D). Group-2 showed no increase in PPS23F-specific IgM from 0.3 ±0.2 g/ml to 0.4 ±0.1 g/ml. However, there was a significant increase in IgG from 1.1 ±0.4 g/ml to 1.9 ±0.6 g/ml (p<0.05). No increase occurred in group-3 PPS23F-specific IgM from 0.1 ±0.0 g/ml to 0.1 ±0.0 g/ml or IgG from 0.4 ±0.1 g/ml to 0.8 ±0.2 g/ml. Both PPS14 and PPS23F IgM and IgG levels post-immunization were significantly (p<0.05) decreased in group-3 (HAART) compared to group-2 (no therapy), however, only PPS14 IgM showed a significant difference pre-immunization (p<0.05). Overall, a 2-fold or greater increase was seen in 21/39 donors for PPS14-specific IgG (group-1: 12/17, group-2: 6/10, group-3: 3/12), 5/39 donors for PPS14-specific IgM (group-1: 5/17, group-2: 0/10, group-3: 0/12), 6/39 donors for PPS23F-specific IgG (group-1: 3/17, group-2: 2/10, group-3: 1/12) and 8/39 donors for PPS23F-specific IgM (group-1: 4/17, group-2: 1/10, group-3: 3/12). Antibody responses showed the most significant increases in HIV-infected individuals with CD4>200. Six to twelve months HAART treated individuals with CD4<100 showed significantly lower antibody titers measured compared to HAART naïve with CD4<
82 4.2.2 HIV-positive individuals had diminished PPS-specific functional antibody response. PPS-specific OPA were performed to test functional antibody response. Data is reported as the reciprocal of the antibody dilution required to obtain 50% killing by differentiated HL60 cells. Group-1 demonstrated a significant increase in OPT on day-30 to both PPS14 (3.9±0.5 to 4797±1338, p<0.01) and PPS23F (5.2±1.1 to 5237±1556, p<0.01) (Figure 4.1E, F). Group-2 showed no significant increases in OPT in response to PPS14 (5.1±1.3 to 1438±829) or PPS23F (7.4±2.5 to 2580±1261). Similarly, group-3 showed no significant increases in OPT in response to PPS14 (2.8±0.4 to 1243±611) or PPS23F (3.3±0.9 to 1665±948). Each donor however, demonstrated a minimal 2-fold increase in OPT post-vaccination (PPS14 average fold increase: group , group-2 927, group-3 544)(PPS23F average fold increase: group , group-2 667, group-3 809) despite low IgM and IgG antibody concentrations. OPA showed PPV23 vaccination is beneficial in all cases by increasing the OPT Decreased B lymphocyte populations in HIV-positive peripheral blood The CD19+ B cell percentages and numbers in the peripheral blood of HIV-positive individuals were analyzed and compared to that of HIV-negative individuals. Our HIVnegative population had an average 12% B cells from total lymphocytes isolated (Table 4.1). HIV-positive individuals with CD4>200 showed a significant decrease (8.2%) in B cell percentage (p<0.01) compared to HIV-negative individuals. The B cell percentage in HIV-positive individuals with CD4<200 was not significantly decreased compared to HIV-negative individuals regardless of HAART usage. All HIV-positive groups showed 59
83 a significant decrease in B cell number compared to HIV-negative individuals. There was no significant difference between HIV-positive groups in B cell number. Table 4.1: Unselected CD19+ B cell percentages and counts among HIV-negative and HIV-positive individuals. Parameter HIV-negative CD4>200 CD4<200 CD4<200, HAART CD19+ B cells, % 12.0 ± ± 1.1 ** 9.9 ± ± 1.3 CD19+ B cells count/ l ± ± 17.0 * ± 11.2 * ± 15.1 * Data are mean ± standard error of the mean. * indicates significant decrease compared to HIVnegative. *p<0.05, **p<0.01 The phenotypes of unselected B cells were similar on day-0 and day-7 (data not shown). Analysis of unselected B cells showed a loss of IgM+ memory and switched memory B cells in HIV-positive individuals CD4<200 (Group-2) compared to CD4>200 (Group-1) (Table 4.2). B cell percentages and numbers were similar between CD4<200 (Group-2) and CD4<200 on HAART (Group-3). Overall, HIV-infected had significantly fewer B cell numbers and an altered phenotype distribution compared to HIV-negative individuals. Table 4.2: Unselected CD19+ B cells percentages and counts categorized by group. Unselected percentage of B cells Unselected number of B cells/ l Donor Groups IgM+ Memory Switched Memory Naïve 60 IgM+ Memory Switched Memory Naïve CD4> ± ± ± ± ± ± 12.0 CD4< ± ± ± ± ± ± 13.0 HAART 5.5 ± ± ± ± ± ± 57.4 HAART represents: CD4<200 with 6-12 months of HAART. Data are mean ± standard error of the mean.
84 4.2.4 Phenotypic analysis of PPS-specific B cells in HIV-positive peripheral blood. To determine the effect of immunization with PPV23 on the B cell populations circulating in the peripheral blood, PPS-selected B cells were analyzed by flow cytometry as previously described [129, 164, 198]. The percentage and number of PPS-selected B cells increased significantly from day-0 to day-7 for all groups and both PPS (p<0.01) (Table 4.3). No significant differences in percentage were found between groups on either day-0 or day-7 for either PPS. Table 4.3: Pneumococcal polysaccharide-selected CD19+ B cell percentages and absolute numbers among HIV-positive individuals before and 7 days after PPV23 immunization. CD19+ PPS+ B cells Percentage of CD19+ PPS14+ D0 PPS14 D7 PPS23F+ D0 PPS23F+ D7 CD4> ± ± ± ± 0.3 CD4< ± ± ± ± 0.5 CD4<200 + HAART 1.0 ± ± ± ± 0.6 Number per ml PPS14+ D0 PPS14 D7 PPS23F+ D0 PPS23F+ D7 CD4> ± ± ± ± 717 CD4< ± ± ± ± 870 CD4<200 + HAART 924 ± ± ± ± 285 Data are mean ± standard error of the mean. The majority of unselected B cells were naïve cells (CD27-) (data not shown). The phenotype distribution of PPS-selected cells on day-7 was significantly different from that of unselected cells (Figure 4.2A, B). All HIV-positive groups showed significant increases in the percentage of PPS14-selected IgM+ memory B cells compared to unselected cells [group ±2.4 to 38.7±3.4, group-2 7.8±1.8 to 21.7±4.6, group-3 61
85 6.2±1.3 to 19.1±3.6](Figure 4.2A). HIV-positive group-1 and group-3 showed significant increases in the percentage of PPS14-selected switched memory B cells compared to unselected cells [group ±3.6 to 33.7±3.7, group ±4.5 to 23.3±5.7, group ±2.4 to 25.6±4.0]. PPS23F selection showed similar differences compared to unselected cells [IgM+ memory; group ±2.4 to 41.9±4.0, group-2 5.3±1.9 to 34.1±5.1, group-3 5.5±1.1 to 28.3±3.7][switched memory; group ±4.3 to 38.0±3.6, group ±4.5 to 29.3±8.2, group ±2.2 to 27.1±3.7](Figure 4.2B). Postvaccination, no significant differences were seen between the percentage of IgM+ memory and switched memory cells of any HIV-positive group. All HIV-positive groups showed significant increases from day-0 to day-7 in the number of PPS14-selected IgM+ memory B cells [group-1 317±113 to 2202±636, group-2 245±96 to 445±88, group-3 356±133 to 958±208](Figure 4.2C). HIV-positive group-1 and group-3 showed significant increases in the number of PPS14-selected switched memory compared to unselected B cells [group-1 348±65 to 1234±228, group-2 300±110 to 615±241, group-3 339±148 to 1188±329]. PPS23F selection showed similar differences from day-0 to day-7 except switched memory cells for group-2 significantly increased while group-3 did not [IgM+ memory; group-1 598±152 to 1863±354, group-2 397±162 to 1276±535, group-3 605±262 to 1182±310][switched memory; group-1 614±113 to 1757±331, group-2 430±134 to 941±299, group-3 525±175 to 1183±443](Figure 4.2D). These data show that both IgM+ memory and switched memory B cells significantly increase in response to PPV23 in newly diagnosed HIVpositive individuals. Post-vaccination, there were no significant differences between the number of IgM+ memory and switched memory cells in any HIV-positive groups. 62
 and day-7 (gray bars) post-immunization, lymphocyte enriched peripheral blood samples stained with fluorescently labeled PPS14 or PPS23F were evaluated by flow cytometry for")
86 Figure 4.2. Majority of PPS-specific B cells from HIV-positive individuals immunized with PPV express CD27 and IgM. HIV-positive donors were immunized with PPV23. On day-0 (white bars) and day-7 (gray bars) post-immunization, lymphocyte enriched peripheral blood samples stained with fluorescently labeled PPS14 or PPS23F were evaluated by flow cytometry for distribution of CD27 and IgM among PPS-selected CD19+ B cells. Pre- verses post-immunization results for all HIV-positive individuals were categorized by CD4 counts and HAART usage. B cells were expressed as a percentage of total PPS-selected CD19+ B cells (A,B). B cells were expressed as the number of PPS-selected CD19+ B cells per ml (C,D). In each sample, 100,000 events were recorded. Data are mean ± standard error measure. *p<0.05, **p<0.01, ***p<
87 4.2.5 Differences in phenotypic distribution of PPS-specific B cells in HIV-positive compared to HIV-negative individuals. HIV-negative individuals were directly compared to HIV-positive groups to identify deficiencies in PPS-specific IgM+ memory and switched memory B cell numbers. Postimmunization, HIV-negative individuals showed significantly higher numbers of PPSselected IgM+ memory B cells (PPS14: 5009±739cells/ l, PPS23F: 3783±586cells/ l) compared to group-1 (p<0.05), group-2 (PPS14: p<0.01, PPS23F: p<0.05) and group-3 (PPS14: p<0.001, PP23F: p<0.05) (Figure 4.3A, B). No significant differences were seen between IgM+ memory cells in HIV-positive groups. No significant differences between the number of PPS14 or PPS23F-selected switched memory cells was seen between HIVnegative individuals (PPS14: 1433±214cells/ l, PPS23F: 1400±253cells/ l) or in any of the HIV-positive groups. HIV-infected have significant deficiencies in their PPS-specific IgM+ memory B cell response compared to HIV-negative individuals even in those that have CD4>200. HAART had no significant effects on the number of PPS-selected cells in individuals with a CD4<
88 Figure 4.3. HIV-positive donors show significantly diminished response to PPV compared to HIV-negative donors. HIV-positive donor samples (gray bars) were compared to HIV-negative donors samples (black bars) day-7 post-immunization with PPV23. Lymphocyte enriched peripheral blood samples stained with fluorescently labeled PPS14 or PPS23F were evaluated by flow cytometry for distribution of CD27 and IgM among PPS-selected CD19+ B cells. Pre- verses post-immunization results for all HIV-positive individuals were categorized by CD4 counts and HAART usage. B cells were expressed as the number of PPS-selected CD19+ B cells per ml (A,B). In each sample, 100,000 events were recorded. Data are mean ± standard error measure. *p<0.05, **p<0.01, ***p<0.001, ****p< Chapter discussion Newly diagnosed HIV-positive individuals are at substantially higher risk for pneumococcal infection compared to HIV-negative individuals, irrespective of vaccination with PPV23. Polyclonal B cell activation has been demonstrated early in disease before quantitative and qualitative defects in CD4+ T cells occur, suggesting intrinsic B cell defects. B cell hyperactivation results in hypergammaglobulinemia, high spontaneous antibody production in vitro, increased expression of activation markers, and increased incidence of autoimmune phenomena and B cell lymphomas. Paradoxically, HIV-positive individuals respond poorly to vaccine antigens [ ]. 65
89 We compared the response to PPV23 in HIV-positive individuals stratified according to CD4 count as a surrogate marker for disease progression. In agreement with previous studies, our HIV-positive population showed diminished polysaccharidespecific IgM and IgG titers post-vaccination compared to HIV-negative individuals, specifically in those with low CD4 counts [192, 193, 197]. It has been postulated that control of HIV viral load, leading to reduced B cell hyperactivation, may improve B cell function. Our HIV-positive individuals with CD4<200 receiving HAART for 6-12 months pre-immunization, paradoxically demonstrated significantly reduced PPS-specific antibody post-vaccination levels compared to HIV-positive individuals with CD4<200 without therapy. To ensure our ELISA results were valid, representative samples were independently verified by a national reference laboratory (ARUP, Salt Lake City, Utah). PPS-specific antibody titers of HIV-positive individuals correlated poorly with CD4 counts and OPT, suggesting that PPS-specific antibody titers serve as a poor clinical surrogate marker for immune protection [12, 125]. Functional impairment of antibodies in HIV-positive individuals has also been reported [13]. Opsonophagocytic assays are a more sensitive and reliable indicator of vaccine response than polysaccharide based ELISA. HIV-positive individuals with CD4>200 showed a significant increase in OPT post-immunization, but increases in individuals with CD4<200 were not significant. OPT levels were all significantly diminished compared to HIV-negative individuals. Six to twelve months treatment with HAART did not improve functional antibody titers. This demonstrates that after 6-12 months of HAART in HIV-positive individuals with CD4<200 does not significantly improve the humoral immune response to PPS. Statistically equivalent levels of antibody 66
90 titers between individuals with CD4>200 and CD4<200 for PPS14 did not correlate with equivalent OPT. Additionally, all antibody titers decreased significantly in individuals receiving 6-12 months of HAART, however, this was not true for OPT suggesting that virological control resulted in antibodies with better functional activity. It has been suggested that low correlations between antibody titers and OPT among the HIV-positive population may be due to dysfunctional antibodies in some individuals [12, 13]. Low antibody titers and OPT likely result from considerable B cell dysfunction due to their decreased activity or total numbers [135] [199]. Regardless, an OPT >8 is thought to correlate with immune protection [12]. The minimum OPT among our donors was >25, suggesting that PPV23 vaccination was beneficial for all donors regardless of CD4. HIV-positive individuals with CD4>200 showed a significantly lower percentage and number of total B cells compared to HIV-negative individuals. In HIV-positive individuals with CD4<200, the total number of B cells was significantly lower than in HIV-negative individuals. The apparently higher percentage of B cells in HIV-positive individuals with a CD4<200, compared to those with CD4>200, likely reflects the loss of the T cells from the total lymphocyte population which served to mask B cell deficiencies. Six to twelve months HAART treatment appeared to have no significant effect on total B cell percentages or numbers. The cells responsible for producing anti-pps specific antibodies is a controversial issue [11, 30, 84, 103, 150]. Several previous studies have analyzed unselected cell populations post-immunization, not reflecting this rare antigen-specific population (<1-4%) [127, 128]. We have demonstrated the ability to identify PPS-specific B cells using fluorescently-labeled polysaccharide in conjunction with flow cytometry. The specificity 67
91 of our labeled PPS was supported by inhibition assays and the ability to bind to PPSspecific monoclonal cells [129]. Likewise, binding of labeled PPS to post-vaccination peripheral blood B cells in flow cytometry was inhibited with addition of homologous unlabeled polysaccharide. We have previously shown in HIV-negative individuals, who respond well to PPV, that the IgM+ memory population constituted the majority of PPSspecific B cells responding to vaccination 7 days post vaccination [129, 198]. In contrast, in the elderly population (>65 years old), at high risk for infection, the majority of PPSselected B cells were switched memory B cells [164]. In this study, we directly labeled PPS and characterized the PPS-specific B cells responding to PPV23 in newly-diagnosed HIV-positive individuals, and compared them to HIV-negative individuals. Our analysis showed that individuals with lower CD4 counts tend to have lower total numbers of PPS-selected B cells. Moreover, B cell numbers showed that both PPS-selected IgM+ memory and switched memory B cells increased significantly post-immunization, but generally demonstrated lower responses with low CD4 counts. Post-immunization levels of PPS-selected switched memory B cells in HIV-positive individuals were comparable to those in HIV-negative individuals. However, IgM+ memory cells were significantly reduced in HIV-positive individuals and decreased with lower CD4 counts. Overall, HIV-positive individuals demonstrated significantly reduced IgM+ memory B cell numbers, which correlated with low OPT, compared to HIV-negative individuals. Thus identifying a specific cellular deficiency in the immune repertoire likely responsible for humoral deficiencies, which are exacerbated as infection progresses towards AIDS. Moreover, individuals on 6-12 months of HAART 68
92 showed no significant increase in the number of PPS-selected IgM+ memory or switched memory B cells post-immunization. These data demonstrated differential intrinsic defects in high risk populations including the elderly and HIV-positive individuals. The correlation between loss of IgM+ memory B cells and decreased humoral response is in agreement with previous publications, which showed that individuals with decreased IgM+ memory B cells such as splenectomized, infants <2 years old, and the elderly, respond poorly to PPV23 [76, 102, 103]. These antigen-specific antibody titers, OPT, and B cell studies emphasize the importance of early clinical HIV infection diagnosis and immediate PPV23 immunization. While it remains beneficial to immunize individuals with CD4<200, based on OPT, immune responses deteriorate in patients with lower CD4 counts. Our studies suggest that despite virological control, 6-12 months of treatment with HAART did not result in improved OPT, most likely the best correlate of protection, or marginally improved B cell numbers. Based on these findings, PPV23 vaccination should not be delayed in newly diagnosed HIV-positive individuals with CD4<
93 Chapter 5 PPV Revaccination in HIV-positive Individuals on Long-term Therapy I made contributions in volunteer enrollment, blood sample processing, cellular staining, flow cytometry, data processing, and assisting with OPA. 5.1 Chapter introduction The advent of HAART has dramatically reduced the incidence of opportunistic infections in HIV-positive individuals [131, 132]. Despite this success, invasive pneumococcal disease (IPD) is highly prevalent in these patients with a disease burden times higher than in HIV-negative individuals [4, 136]. The high incidence of pneumococcal disease has led to the recommendation to vaccinate all HIV positive adults against Streptococcus pneumoniae, followed by a booster after 5 years [112, 191]. The efficacy of PPV23 in HIV-positive individuals however has thus far remained controversial. Multiple observational studies in the US reported little benefit of PPV23, while a randomized, double blind trial in Uganda reported detrimental effects [190, ]. The current vaccine recommendations thus stem from the belief that beneficial effects likely outweigh potential harm. 70
94 Moreover, the ideal time for PPV23 administration and the benefits of PPV23 revaccination in HIV-positive individuals has been a subject of debate. The Centers for Disease Control and Prevention (CDC) recommend pneumococcal vaccination in HIV positive patients having a CD4 count >200 [206]. World Health Organization (WHO) guidelines suggest that vaccination be considered in individuals with a CD4 count >500, implying PPV23 should be administered during the early stage of HIV infection or after sufficient immune reconstitution by HAART [207]. Although the reconstitution of B cells and reduction in B cell abnormalities with HAART use is well documented, fewer studies have focused on the effect of HAART on antigen-specific B cell reconstitution [208]. Thus, the goal of the current study was to evaluate the effect of PPV23 revaccination in HIV-positive patients undergoing long-term HAART (HIV-HAART). To the best of our knowledge, this is the first comprehensive study that quantitatively and qualitatively determines anti-pps antibody response, investigates the nature of PPS-responding B cells and the influence of CD4 T cells post-ppv23 revaccination in the long-term HAART volunteers. Our results indicate significant rises in the PPS-specific total and functional antibody titers post-ppv23 revaccination in the long-term HAART volunteers. This was concomitant with a rise in PPS-specific IgM+ memory B cells, a critical B cell subset required for response against Streptococcus pneumoniae [129]. Moreover, this indicates PPV23 revaccination is beneficial. However, the serological and antigen-specific peripheral B cell response remained suboptimal in these HIV-positive individuals irrespective of the degree of CD4 T cell reconstitution compared to HIV-negative 71
95 individuals. The results indicate persistent PPS-specific B cell deficiencies despite PPV23 revaccination and long-term HAART administration. 5.2 Results Serum PPS-specific antibody levels increased post-ppv23 revaccination in long-term HAART volunteers We compared two patient groups that differed in CD4, HIV-positive individuals with CD4>200 (group A) and HIV-positive individuals with CD4<200 (group B). The anti-pps14 IgG response increased significantly from 5.44 ± 1.57 µg/ml pre-vaccination to ± 2.92 µg/ml post-vaccination in group A and from 4.96 ± 2.02 µg/ml to ± 5.14 µg/ml in group B. Similarly, the anti-pps23f response rose to 4.65 ± 2.03 µg/ml from 1.88 ± 0.72 µg/ml in group A and to 2.31 ± 0.77µg/ml from 1.22 ± 0.79 µg/ml in group B. The anti-pps23f IgG levels post-vaccination was not significantly different for either group compared to pre-vaccination levels (Figure 5.1 A,B). Post-vaccination IgM levels against PPS14 increased significantly in group A from 0.44 ± 0.09 µg/ml to 0.68 ± 0.16 µg/ml but not in group B where the levels reached 0.31 ± 0.08 µg/ml on day-30 compared to 0.25 ± 0.05 µg/ml on day-0. Anti-PPS23F IgM levels did not increase significantly in either groups post-vaccination (group A from 0.19 ± 0.03 µg/ml on day-0 to 0.22 ± 0.04 µg/ml on day-30 and from 0.13 ± 0.01 µg/ml on day-0 to 0.16 ± 0.02 µg/ml on day-30 in group B). Both pre- and post-vaccination anti- PPS IgG and IgM levels were comparable between groups A and B (Figure 5.1 C, D). 72
96 Figure 5.1. Serological response to PPV23 revaccination in the long-term HAART cohorts. Long-term HAART cohorts (group A, n=33 and group B, n=11) were immunized with PPV23. Serum samples were obtained on day 0 (pre-vaccination, represented as gray bars) and day 30 (post-vaccination, represented as black bars). Serum samples were tested for anti-pps14 and PPS23F IgG (A, B), IgM (C, D), and opsonophagocytic activity (E, F). Serum antibody levels are expressed as µg/ml, and opsonophagocytic activity is expressed as opsonic titers (OPT). p-values < 0.05 were considered statistically significant. 73
97 5.2.2 Functional antibody titers increased post-ppv23 revaccination in long-term HAART volunteers Opsonophagocytic titers (OPT) give a better indication of protective efficacy of vaccines than ELISA alone. OPT indicate the cumulative killing effect of anti-pps isotype antibodies [161, 209]. OPT against serotype 14 increased significantly (almost 10x), from ± on day-0 to 4100 ± 1059 on day-30 in group A (CD4>200) and from ± to 2671 ± 1563 in group B (CD4<200). Similarly, the OPT against serotype 23F increased significantly, from ± to 4614 ± 1134 in group A and from ± to 8451 ± 2971 in group B pre- to post-vaccination respectively. Both pre- and post-vaccination OPT were comparable in group A and B (Figure 5.1 E, F) Absolute B cell numbers and B cell percentage in the long-term HAART volunteers We examined the absolute B cell numbers and B cell percentages in groups A and B (Table 5.1). We assessed 3 different populations of B cells, namely naïve B cells (CD27-IgM+/-), IgM+ memory B cells (CD27+IgM+) and switched memory (CD27+IgM-) B cells. The percentage and absolute numbers of unselected B cell subsets are shown in Table 5.2. No significant differences were noted between group A and B for the mentioned populations in terms of percentage or absolute numbers (Table 5.1, 5.2). 74
98 Table 5.1: B cell counts and percentages in HIV-positive individuals on long-term HAART Parameter Group A (CD4>200) Group B (CD4<200) CD19+ B cells, % 8.60 ± ± 1.74 CD19+ B cells, cells/µl ± ± 22.9 Data are mean ± standard error of the mean Table 5.2: Unselected B cell subset percentages and absolute counts in HIV-positive individuals on long-term HAART. Unselected B cell percentage Unselected number of B cells/µl Patient Groups IgM+ Memory Switched Memory Naïve IgM+ Memory Switched Memory Naïve Group A (CD4>200) 41.0 ± ± ± ± ± ±12.4 Group B 50.9 ± ± ± ± ± ±20.9 (CD4<200) Data are mean ± standard error of the mean Phenotypic analysis of PPS-specific B cells in the HIV-positive individuals volunteers on long-term HAART post-ppv23 revaccination We investigated the PPS-specific B cell response in the long-term HAART volunteers. PPS-specific B cells peak in the peripheral blood on day-7 post-immunization as reported previously [129, 185]. We compared the PPS14- and PP23F-specific B cell percentages and found significant increases both in percentage and absolute numbers on day-7 compared to its levels on day-0 (Table 5.3A,B). No significant differences were noted between the 2 groups. 75
99 Table 5.3: Pneumococcal polysaccharide-selected CD19+ B cell percentages and counts in the long-term HAART volunteers before and 7 days after PPV23 revaccination A] B cell Percentages Patient groups CD19 + PPS14 + % Day 0 CD19 + PPS14 + % Day7 CD19 + PPS23F + % Day 0 CD19 + PPS23F + % Day7 Group A (CD4>200) Group B (CD4<200) 1.32 ± ± 0.29* 1.28 ± ± 0.22* 0.94 ± ± 0.84* 0.77 ± ± 0.50* B] B cell Counts Patient groups CD19 + PPS14 + (cells/ml) Day 0 CD19 + PPS14 + (cells/ml) Day 7 CD19 + PPS23F + (cells/ml) Day 0 CD19 + PPS23F + (cells/ml) Day 7 Group A (CD4>200) 1427 ± ± 393.5* 1548 ± ± * Group B (CD4<200) 1052 ± ± 921.2* 1016 ± ± 772.7* Data are mean ± standard error of the mean. * indicates significant rise (p values <0.05) in the post vaccination (day-7) levels compared to pre-vaccination day (day-0). We compared the B cell subset distribution of unselected cells on day-0 with PPSselected cells on day-7 (Figure 5.2). It should be noted that unselected B cell phenotype was similar on both day-0 and day-7. The PPS14- and PPS23F-specific switched memory B cell percentage increased significantly from ± 1.5 to ± 2.97 (p=0.009) and from ± 1.59 to ± 3.06 (p=0.0002) respectively in group A. In contrast, no significant differences were noted in group B for either PPS (Figure 5.2). 76
 verses PPS14 or PPS23F-specific (gray bars) B cells (A,B). Numbers of PPS-selected B cells before (white bars) and after (gray bars) revaccination (C,D).")
 and from 9.84 ± 1.26 to 40.98 ± 3.36 (p<0.")
100 Figure 5.2. PPS-specific B cell subset responses in long-term HAART cohorts. The phenotypes of B cells that bind to labeled PPS14 and PPS23F were determined by flow cytometry. Percentages of unselected (white bars) verses PPS14 or PPS23F-specific (gray bars) B cells (A,B). Numbers of PPS-selected B cells before (white bars) and after (gray bars) revaccination (C,D). Group-A (n=33) and group-b (n=11). 100,000 events were recorded for each sample. p-values < 0.05 were considered statistically significant. Post-vaccination, the percentage of both PPS14- and PPS23F-specific IgM+ memory B cells increased significantly, from ± 1.33 to ± 4.01 (p<0.0001) and from 9.84 ± 1.26 to ± 3.36 (p<0.0001) respectively in group A. Similarly, the percentage of PPS14- and PPS23F-specific IgM+ memory B cells increased from 8.50 ± 2.08 to ± 5.35 (p<0.0001) and from 8.13 ± 1.64 to ± (p<0.0001) respectively in group B (Figure 5.2 A,B). 77
101 We found an increase in PPS14-specific IgM+ memory B cell numbers (from ± 58.0 to 1122 ± cells/ml (p<0.0001) in group A and from ± 15.4 to 1296 ± cells/ml (p=0.02) in group B; Figure 5.2C). Similarly, PPS23F-specific IgM+ memory B cell numbers increased (from ± to 1612 ± cells/ml in group A (p=0.0006) and from ± 77.3 to 1703 ± cells/ml(p=0.03) in Group B; Figure 5.2 D). Switched memory B cell numbers increased significantly in response to PPS14 (from 64.4 ± to ± in group B (p=0.01)), and in response to PPS23F in group A (from ± to 1161 ± (p=0.004); Figure 5.2 C). No significant differences were found in the PPS-specific response between group A and B, pre- or post-vaccination (Figure 5.2). We noted strong positive correlations between the percentage of PPS14-specific IgM+ memory B cells on day-7 and the PPS14 opsonic antibody titers on day-30 for each of the patient groups (r 2 =0.82, p < for group A and r 2 =0.4 for group B). The trend was similar in response to PPS23F (r 2 =0.64, p< for group A and r 2 =0.84, p=0.0004). In contrast, despite increases in the switched memory B cell percentage in group A on day-7, no correlations were found between switched memory and OPT. Similarly no correlations were noted between naïve B cells on day-7 and day-30 OPT Diminished response to PPV23 in HIV-positive individuals on long-term HAART compared to HIV-negative individuals, despite revaccination We compared the response to PPV23 revaccination in the long-term HAART patients with results in HIV-negative individuals. Post-vaccination PPS14 and PPS23F IgG levels in HIV-negative individuals were significantly higher than in either group A 78
102 or B (mean concentrations ± 4.78 µg/ml (p=0.01) and ± 3.61 µg/ml (p=0.008) respectively). Similarly, the post-vaccination PPS14 and PPS23F IgM levels in HIV-negative individuals had mean concentrations of ± 3.76 µg/ml (p =0.0002) and ± 2.31 µg/ml (p =0.0002), both significantly higher than either group A or B. This result was consistent with significantly higher post-vaccination OPT in HIVnegative individuals (12470 ± (p<0.0001) against PPS14 and ± (p<0.0001) PPS23F) compared to both groups A and B. In agreement with the serological response, the percentage and number of PPSspecific IgM+ memory B cells remained significantly higher in HIV-negative compared to HIV-positive individuals for almost all comparisons. The number of PPS23F-specific IgM+ memory B cells in group B was not significantly lower compared to HIV-negative individuals (Figure 5.3). 79
 and absolute numbers (C,D) were compared between long-term HAART cohorts (gray bars) versus HIV-negative individuals (Healthy, black bars)")
103 Figure 5.3. Comparative analysis of PPS-specific B cell response in the long-term HAART cohorts versus HIV-negative volunteers. Day-7 post vaccination PPS-specific B cell percentage (A,B) and absolute numbers (C,D) were compared between long-term HAART cohorts (gray bars) versus HIV-negative individuals (Healthy, black bars) volunteers. Higher percentage and absolute numbers of PPS-specific IgM+ memory B cells were noted in HIV-negative individuals (n=29) compared to group A (n=33) and group B (n=11). p-values < 0.05 were considered statistically significant. 5.3 Chapter discussion: It is currently recommended that people at a high risk for pneumococcal infection, including the HIV infected population, be revaccinated with PPV23 after an interval of 5 years. However, there are no data available to justify this practice. The primary objective of the current study was to evaluate the immune response to PPV23 revaccination in long-term HAART patients, and to determine if this recommended regimen is protective 80
104 as measured by surrogates of protection. Secondly, we sought to understand if the recovery in the CD4+ T cells post-haart also leads to a simultaneous improvement in the antigen-specific B cell response to PPV23. To perform these studies, we enrolled long-term HAART patients with a history of nadir CD4 count <200 (AIDS) and undetectable viral load at the time of the study. They were then stratified based on the varying degree of immune restoration at the time of vaccination as group A (CD4>200) and group B (CD4 <200). We chose PPS serotypes 14 and 23F for our study, to allow direct comparison with our work involving HIV-negative individuals and for reasons enlisted previously [129]. Our results indicate PPS-specific IgM and IgG levels are suboptimal postvaccination (Figure 5.1 A-D). The results were consistent with a significant, however suboptimal, increase in the PPS-specific post-vaccination OPT (Figure 5.1 E,F). Although correlates of protection have not been defined for adult population, it has been suggested that an opsonic titer >8 is more likely to confer protection [12]. Based on this threshold, our data indicate PPV23 revaccination to be a beneficial practice although the post-vaccination increases are only modest compared to HIV-negative individuals (healthy) volunteers. The significantly diminished serological response in long-term HAART volunteers, compared to HIV-negative individuals, inspired our investigation of potential antigen-(pps)-specific B cell perturbations. Consistent with the speculation that IgM+ memory B cells play a critical role in the response against Streptococcus pneumoniae [76, 103], we recently identified the majority of PPS-specific B cells in HIV-negative individuals to be IgM+ memory B cells [129]. These cells play a pivotal role in protection 81
105 targeting PPS, an effect that is poorly compensated by other PPS-specific B cell subsets, as reported in our work in the elderly population [164]. In concurrence with these reports, we noted significantly lower percentage of PPS-specific IgM+ memory B cells in the long-term HAART volunteers despite PPV23 revaccination. Strong positive correlation between PPS-specific IgM+ memory B cells and OPT provide unequivocal evidence of the role played by IgM+ memory B cells in protection against S. pneumoniae. These findings are similar to other reports, in which depletion of specific memory B cell pools during the early stages of HIV-infection culminated in attenuated response to vaccination [208]. Next, we assessed the influence of CD4 T cells in the serological response to PPS by comparing the responses in group A (CD4>200) and B (CD4<200). We did not note significant differences in total or functional antibody responses between the 2 groups, despite their different CD4 counts. Similarly, we did not note significant differences in the PPS-specific IgM+ memory B cell numbers or percentage pre- and post-vaccination between the 2 groups. Our results are consistent with other reports in which CD4+ T cells were found to be a poor predictor in the vaccine response to PPS [193, 210]. It is also consistent with reports suggesting loss of discrete memory B cell subsets could lead to heightened susceptibility to pneumococcal infection [128]. HIV can remain latent in secondary lymphoid organs, even in patients with undetectable plasma viral load, despite administration of HAART [211, 212]. It is thus likely that HIV can cause persistent damage to key B cell subsets required for response against PPS. It has been proposed that circulating IgM+ memory B cells are prediversified splenic marginal zone B cells [76]. Similarly, a subset of IgM+ memory B 82
106 cells are said to resemble the recently characterized putative human B-1 cells [30]. Both marginal zone B cells and B-1 cells are known to play crucial roles in defense against T- cell independent antigens (including polysaccharides) [11]. In line with these theories, we found PPS-specific IgM+ memory B cells to resemble the putative human B-1 cells in our recent work [198]. The percentage of these PPS-specific putative B-1 cells were significantly lower in HIV+ patients on long-term HAART compared to HIV-negative individuals (preliminary unpublished observations). B-1 cells are self-renewing subsets whose numbers wane with age [10]. It is plausible that these cells are damaged at an early stage of HIV infection and fail to recover with HAART. Our results are in alignment with other reports where HAART has only partially corrected B cell associated perturbations [199]. These results are novel as we are the first to our knowledge to study immune response to PPV23 revaccination in long-term HAART patients. The majority of the previous reports in HIV+ patients focused on the PPS IgG response undermining the relevance of IgM in protection [193]. Similarly, most studies did not assess functional antibody levels by OPA post-vaccination [192, 193]. It is well accepted that OPTs are better correlates of antibody protection than ELISA [209]. Furthermore, this study is unique as we elucidated the phenotype of PPS-specific B cells in long-term HAART, PPV23 revaccinated volunteers. The specificity and sensitivity of direct PPS labeling, to elucidate the phenotype of PPS-specific B cells, was demonstrated in our previous work [129]. We conclude that PPV23 revaccination is beneficial for HIV-positive patients on long-term HAART. Comparable OPT between patient groups indicate poor PPS-specific 83
107 B cell reconstitution, irrespective of the degree of T cell reconstitution post-haart. The response elicited by this patient population is, however, remarkably lower than the response seen in HIV-negative individuals, emphasizing the need for alternative therapeutic approaches that can lead to a more robust response. The ACIP has recently updated the vaccine recommendations for HIV-positive patients. Administration of PCV13 followed by a dose of PPV23 is recommended [6]. It remains to be seen if this combined regimen can lead to better protection against S. pneumoniae in HIV-positive individuals on long-term HAART. 84
108 Chapter 6 Discussion and Interpretations Improving vaccination against pneumococcus is essential to reducing the morbidity and mortality of IPD. A clearer understanding of the immune response to PPV23 vaccination, and identification of immune deficiencies among high-risk individuals, is needed. Our primary goal was to identify PPS-specific immune responses to PPV23 vaccination by analyzing antibody concentrations, antibody functional activity, and B cell populations pre- and post-vaccination. While previous studies have sampled antigen non-specific B cell populations, these studies are unique in that they utilize fluorescently conjugated PPS to specifically identify the rare PPS-reactive B cells in the peripheral blood by flow cytometry. Scientifically, these studies add knowledge and insight to TI-2 antigen based vaccination strategies. TI-2 antigen based immune responses in humans remain a controversial subject, partly due to limited available sample materials and also limitations in identifying relevant immune components. We identified differences in the immune responses between low-risk and high-risk populations, allowing us to identify immune deficiencies in high-risk populations. This information is needed to improve future treatment of high-risk individuals and reduce overall disease burden. 85
109 These studies also have direct clinical relevance. First, we directly compared immune responses to PPV23 in ND-HIV categorized by CD4 count to determine at what stages it is beneficial to vaccinate these patients. Secondly, we directly compared ND- HIV with CD4 counts <200 without HAART to ND-HIV with CD4 counts <200 on 6-12 months of HAART before vaccination. This comparison indicated that a 6-12 months of HAART, for viremic control and partial immune reconstitution, did not improve vaccination response. Lastly, we compared HIV-positive individuals with previous CD4<200 now on long-term (>5 years) HAART to test if revaccination was beneficial and if increases in CD4 T cells correlated with PPS-specific correlates of protection. 6.1 Antibody responses and deficiencies ELISA titers Together, these studies show serological and cellular differences between the lowrisk population and high-risk populations that we tested. Young healthy individuals in our study responded to PPV23 with significant increases in both IgM and IgG antibody titers by day 30. By comparison, IgM levels in elderly individuals were considerably lower for both PPS and we detected no significant increase in PPS14 IgM. IgG levels showed a variable response, higher and lower, dependent on the PPS. HIV-positive individuals also showed abnormal antibody responses compared to HIV-negative individuals, dependent on both CD4 counts and HAART usage. HAART usage for 6-12 months in ND-HIV significantly reduced overall antibody titers compared to CD4 matched ND-HIV without HAART. HIV-HAART with CD4<200 show similar IgM levels as ND-HIV vaccinated after 6-12 months HAART. Moreover, HIV-HAART 86
110 with CD4>200 show IgM levels lower than ND-HIV with CD4>200. This suggests that long-term HAART usage does not lead to significantly increased IgM levels. In contrast, IgG levels were higher in HIV-HAART with CD4<200 compared to CD4 matched ND- HIV. Total antibody titers of HIV-HAART with CD4>200 were not significantly higher than HIV-HAART with CD4<200. However revaccination in HIV-HAART, regardless of CD4 counts, did not result in PPS-specific antibody levels equivalent to levels in HIVnegative individuals. This suggests that HIV-HAART remain at high risk for infection, partly because B cell defects incurred by HIV infection are not completely reversed by long-term HAART. The protective levels of PPS-specific antibodies in adults are unknown. Correlations between antibody levels and IPD incidence tend to be weak [17, ]. Previous studies, like those performed in South Africa, which have studied PPS-specific antibody levels have demonstrated that total PPS-specific titers do not serve as a sensitive surrogate marker for protection against pneumococcus [125] Opsonophagocytic titers OPT serves as a more sensitive surrogate marker for immune protection than total antibody titers. Studies have shown that individuals with similar total antibody titers may demonstrate different functional titers, OPT [12]. This suggests that some PPS-specific antibodies may be dysfunctional, especially in high-risk populations, and therefore make total antibody titers a poor surrogate marker [13][187, 188]. Previous studies have demonstrated that an OPT of 8 leads to protection in mice and correlates with protection 87
111 in children [12, 213]. The protective titer in adults remains unknown, though it is generally assumed that an OPT>8 results in protective immunity. Our studies demonstrated that OPT for both PPS14 and PPS23F significantly increased post-vaccination in young healthy, elderly, ND-HIV with CD4>200, and HIV- HAART, except for PPS14-specific titers in HIV-HAART with CD4<200. These studies suggest that vaccination in all groups tested is beneficial. The level of response in the elderly was roughly 20% the level seen in young healthy individuals. In ND-HIV with CD4>200, OPT were roughly 50% the level seen in HIV-negative individuals. ND-HIV with CD4<200 resembled levels seen in the elderly regardless of 6-12 months of HAART use. These levels of OPT likely explain the increased disease incidence in the elderly and ND-HIV. OPT response of HIV-HAART varied depending on PPS-specificity, but overall levels increased compared to ND-HIV on 6-12 months of HAART. This demonstrates that long-term HAART is beneficial to improving functional antibody titers. Despite this, HIV-HAART OPT levels remained lower compared to HIV-negative individuals, suggesting increased susceptibility for those on long-term HAART even with increased CD4+ T cell counts. 6.2 PPS-specific B cells The cellular immune response to PPS, a prototypic TI-2 antigen, is not well understood in humans. Using fluorescently labeled PPS, we have identified PPS-selected B cell populations in young healthy, elderly, and HIV-positive individuals responding to PPV23 vaccination. Young healthy individuals have a low incidence of IPD, indicating the PPS-specific B cell response in these individuals is functional and protective. In 88
112 contrast, high disease incidence in the elderly and HIV-positive individuals suggests PPSspecific B cell deficiencies/abnormalities in these high-risk populations Young healthy individuals We have previously studied the phenotype of B cells induced by PPV23 with respect to CD27 and IgM [129]. The majority of these PPS-specific cells were IgM+ memory B cells. We found a strong correlation between the percentage of PPS-specific IgM+ memory B cells on day-7 and IgM antibody concentrations on day-30 for both PPS. Moreover, PPS-specific IgM+ memory B cells correlated well with OPT. In contrast, we found no correlations between PPS-specific class-switched B cells on day-7 and IgG antibody concentrations on day-30 for either PPS. Previous studies have also shown that individuals with low to non-detectable levels of IgM+ memory B cells including splenectomized, children <2 years old, elderly, and common variable immunodeficiency patients respond poorly to polysaccharide based vaccination and demonstrate higher rates of infection to encapsulated pathogens [76, 102, 103, 146, 147, 174]. Studies are now showing that these IgM+ memory B cells may represent multiple B cell populations including MZB and B-1 cells. We have now shown that the majority of these PPS-selected IgM+ memory B cells are also CD43+ and CD5+/-, a phenotype that was recently ascribed to a putative human B-1 cell equivalent [30]. The identification of a human B-1 cell counterpart remains controversial. Because activation markers including CD27, CD43, and CD5 are also found on T cells, which form doublets with B cells, and on pre-plasmablasts, additional markers including CD3, CD20, and CD70 were included. Using these 89
113 phenotypic markers, we concluded that the majority of PPS-selected B cells postvaccination with PPV23 were in alignment with this putative B-1 cell population suggested to have involvement in innate immune responses Elderly individuals In contrast, the elderly showed no significant increase in the percentage of IgM+ memory B cells. Instead, the majority of PPS-selected B cells post-ppv23 were switched memory B cells. These switched memory B cells may help partially compensate for lower PPS-selected IgM+ memory B cell levels compared to young individuals. Although IgM+ memory B cells have been shown to play a vital role in bacterial clearance, they are not likely to be solely responsible for the production of antipneumococcal antibodies. Switched memory B cells produce greater quantities of PPSspecific antibodies in vitro than IgM+ memory B cells [214]. In addition, 5 days postvaccination, the majority of antibodies are IgG and IgA produced by switched memory B cells [ ]. Moreover, our elderly population showed strong correlations between PPS-specific IgG titers and OPT, but no correlation between IgM titers and OPT. If PPSspecific switched memory B cells do help compensate for their low IgM+ memory population, the elderly low IgG antibody concentrations and OPTs compared to young individuals indicate that they are insufficient to completely substitute for the loss of the IgM+ memory population. Elderly IgG antibody titers also show no correlation with the number or percentage of PPS-specific switched memory B cells suggesting that these switched memory B cells may be at least partially dysfunctional. 90
114 6.2.3 ND-HIV individuals The efficacy of PPV23 in HIV-infected has been controversial [203, 215, 216]. In contrast to both young healthy individuals and the elderly, ND-HIV showed significant increases in both IgM+ memory and switched memory B cell post-vaccination regardless of CD4 counts or 6-12 months of HAART use. There were only two exceptions were in the PPS14-specific switched memory B cells of ND-HIV with CD4<200 and PPS23Fspecific switched memory B cells of ND-HIV with CD4<200 on 6-12 months HAART which showed non-significant increases for both PPS. This suggests that HIV-positive individuals may have different intrinsic B cell defects than the elderly. Post-vaccination, absolute numbers of PPS-specific B cells showed no significant differences dependent on CD4 counts or 6-12 months of HAART use. Based on these findings, controlling viremia did not significantly influence PPV23 vaccination of ND-HIV with CD4<200. Therefore pneumococcal vaccination should not be delayed in newly diagnosed HIV-positive individuals with CD4<200. The percentages and absolute number of PPS-specific IgM+ memory B cells were lower compared to HIV-negative individuals even in those with CD4>200. This supports the idea that B cell deficiencies occur earlier in disease progression. We also observed a loss of correlation between IgM+ memory cells and IgM antibody titers and OPT among ND-HIV. Previous studies have shown that individuals with decreased total IgM+ memory B cells such as splenectomized, infants less than 2 years old, and the elderly respond poorly to PPV23 [76, 102, 103]. This deficiency in the PPS-specific IgM+ memory B cells likely contributes to decreased OPTs and increased disease incidence among ND-HIV. 91
115 6.2.4 Revaccination in long-term HAART HAART is used in HIV-positive individuals for viremic control and T-cell reconstitution. Long-term HAART use often allows partial immune recovery, including an increase in total B and T cell numbers. Our data show that revaccination of HIVpositive individuals on long-term HAART (HIV-HAART) is beneficial and results in an increase of both total switched memory and IgM+ memory B cells. HIV-HAART primarily showed percentages and numbers of PPS-specific switched memory B cells similar to HIV-negative individuals. Levels of PPS-specific IgM+ memory B cells however were lower compared to HIV-negative individuals. Considering the strong correlation between OPT and levels of PPS-specific IgM+ memory B cells, this identifies an important cellular deficiency contributing to increased disease incidence in HIV- HAART. Post-PPV23, levels of PPS-specific B cells were similar for HIV-HAART with CD4>200 and CD4<200. PPS-specific B cell levels were also similar to those of ND- HIV with CD4<200. This suggests that loss or dysfunction of PPS-specific B cells, caused by HIV infection, is long-lasting and likely permanent and independent of viremic control, total T cell numbers, or total B cell increases associated with HAART use. Previous studies have also reported that increases in CD4 T cells resulting from HAART did not return IgM+ memory and switched memory B cell function in HIV-positive individuals to normal levels [109, 127, 128, 135]. It is likely that the PPS-specific B cells which are destroyed during HIV infection can not be naturally replenished. This stresses the importance of early detection and virologic control to maintain as much of the B cell repertoire as possible. Future development of cellular replacement therapies may be beneficial once viremic control is established. 92
116 6.3 Future studies Further characterization of PPS-specific B cells and the putative human B-1 cells in young healthy individuals is needed, to conclusively determine if these phenotypically identical populations are in fact functionally the same or different. If these cells do represent a human equivalent to mouse B-1 cells, it is not known what characteristics this population will have in common or how it might differ from mouse B-1 cell populations. Mouse B-1 cells can be divided into two functionally separate subpopulations, B-1a and B-1b [81]. It is not known if the same division exists in humans or if human B-1 cell equivalents have additional features not seen in mice. We have identified different PPS-specific B cell deficiencies in two high-risk populations, the elderly and HIV-positive individuals. This suggests that the development of therapeutics and improved vaccination strategies may need to be uniquely tailored for these populations. Further studies in these populations are needed to identify underlying causes for decreased PPS-specific populations and low OPT. As we have already mentioned, B cell depletion occurs in both the elderly and HIV-infected. These individuals are also subjected to chronic inflammation [217, 218]. Chronic inflammation has been associated with B cell anergy and possibly contributes to decreased antibody production of surviving B cells [219]. Furthermore, it is unknown if other high-risk populations like transplant patients or children have the same or additional deficiencies. The ACIP has recently recommended PCV13 for use in adults [6]. High-risk populations are recommended to receive PCV13 followed by PPV23, however evidence to support this regimen is inconclusive. Additional studies comparing the PPS-specific 93
117 immune response to PCV13 and effectiveness of these vaccines are needed. The phenotype of PPS-specific B cells which respond to this TD vaccine in low-risk and highrisk populations remains to be elucidated. 94
118 Chapter 7 Materials and Methods 7.1 Study volunteers Chapter 2 study volunteers Seventeen healthy volunteers participated in the University of Toledo IRB committee approved study (IRB #105137). Volunteers were years old (mean=26.6) and pneumococcal polysaccharide vaccine naïve. Volunteers were questioned about medications, previous illness, and present health before immunization with PPV23, (23- valent pneumococcal polysaccharide vaccine, Merck) Chapter 3 study volunteers Fourteen elderly volunteers (mean age=72, range=64-88) participated in the University of Toledo Institutional Review Board committee-approved study (#105137). Individuals were questioned about medications, previous illness, and present health. Informed consent was obtained from all participants. Ten volunteers (mean age=71, range=64-75, 88) were PPV-naïve and four (mean age=78, range=70-84) had been immunized with PPV >5 years prior. Volunteers were questioned about medications, previous illness, and present health before immunization with PPV23, (23-valent pneumococcal 95
119 polysaccharide vaccine, Merck). Blood samples were collected pre-vaccination, day-7, and day-30 post-vaccination Chapter 4 study volunteers Thirty nine pneumococcal polysaccharide naïve newly diagnosed HIV-positive volunteers participated in the University of Toledo IRB committee approved study (IRB # and ). Seventeen volunteers with CD4>200 (average 36yrs, virus particles per ml), referred to as group-1, and ten with CD4<200 (average 44yrs, virus particles per ml), referred to as group-2, received no HAART therapy before immunization. Twelve volunteers with CD4 of <200 (average 42yrs, virus particles per ml) were treated with HAART for six to twelve months before immunization with PPV23, referred to as group-3. HIV-positive individuals were also stratified according to CD4 counts. Twenty nine HIV-negative volunteers (average 29yrs) participated in this study. Volunteers were questioned about medications, previous illness, and present health before immunization with PPV, (23-valent pneumococcal polysaccharide vaccine, Merck). Blood samples were drawn 0, 7 and 30 days postimmunization Chapter 5 study volunteers HIV-positive patients on long-term HAART (> 5 years) were recruited from the University of Toledo Medical Center (IRB # and ). These patients had previously received their first dose of PPV23 > 5 years prior to the current study and were due for revaccination based on the ACIP (Advisory Committee on Immunization 96
120 Practices) recommendation, at the time of study [112]. Common to these patients were the viral load (undetectable) at the time of vaccination and a history of nadir CD4 count <200 (AIDS). The patients were stratified based on the CD4 count at the time of vaccination as Group A: CD4>200 (indicating immune restoration, n=33; mean age: 49) and Group B: CD4< 200 (n=11, mean age: 48). HIV negative, healthy PPV23 vaccinated volunteers (n=29, /mean age: 26) served as the control group. Volunteers were questioned about medications, previous illness, and present health before immunization with PPV, (23-valent pneumococcal polysaccharide vaccine, Merck). 7.2 Vaccination and blood draw All individuals were immunized with PPV23 (Merck & CO., INC) on day 0. Blood was drawn on day-0 (pre-vaccination), day-7 and day-30 post-vaccination. 7.3 Pneumococcal polysaccharide enzyme linked immunosorbent assay (ELISA) ELISA were performed to identify the anti-pps-specific Abs in all volunteers. The ELISA is modification from the World Health Organization s (WHO) assay [ ]. Briefly, 10 g/ml pneumococcal polysaccharide, either PPS14 or PPS23F (ATCC cat# 197-X and 217-X respectively), was absorbed onto Nunc Maxisorp microtiter plates (Nunc, Roskilde, Denmark) at 37ºC overnight. Plates were then washed with PBS plus 0.1% Tween 20 (PBST). Sera was diluted 1:200 in PBST and then absorbed with cell wall polysaccharide 10 g/ml (CWPS) (Statens Serum Institut cat# 3459) and 10 g/ml PPS22F (ATCC cat# 216-X) for 30 minutes at room temperature. After absorption, sera were serially diluted onto the plates and incubated at 37ºC for 2 hours. The standard 97
121 serum, 89-SF and 007-SF were used as positive controls. Plates were washed, and bound antibody was detected using horse radish peroxidase (HRP) conjugated anti-human Ig (H + L) monoclonal antibody (mab) (Southern Biotechnology Associates) diluted 1:3000 in 1% BSA PBST and incubated at 37ºC for 1 hour. After washing, plates were developed by using an o-phenylenediamine substrate, and stopped with H 2 SO 4. The optical density was read at a wavelength of 490nm. 7.4 Opsonophagocytic assay (OPA) Opsonophagocytic assay was performed as previously described [160, 209]. Briefly, S. pneumoniae, serotypes 14 and 23F, were incubated with serial diluted heat-inactivated sera. Newborn rabbit serum (Pel-Freez, Brown Deer, WI.) was added as source of complement. Differentiated HL-60 cells were added at an E:T ratio of 400:1. Sera were tested in duplicate. The opsonophagocytic titer (OPT) was determined as the reciprocal of the dilution with 50% killing when compared with serum free controls and analyzed using the Opsotiter1 software program (University of Alabama at Birmingham). 7.5 Labeling of polysaccharide 14 and 23F with fluorescent dye Conjugation of PPS14 to cascade blue (CB) ethylenediamine (Invitrogen catalog C-621) or PPS23F to 5-(4,6-dichlorotriazinyl) aminofluorescein (DTAF; Sigma-Aldrich catalog 36565) was carried out as follows. A total of 10 mg PPS14 or PPS23F (10mg/ml in 0.1M borate buffer [ph 9.0]) was incubated with 10mg CB or 10mg 5-DTAF, respectively, for 2.5 hours at 4ºC. The mixture was dialyzed against PBS for 24 hours at 4ºC with four changes of PBS (molecular cutoff = 8 kda). Approximately 10 l 5 M 98
122 sodium cyanoborohydride was added to the dialysate, and the samples were mixed for another 30 minutes at 4ºC in the dark. The samples were again dialyzed against PBS for 24 hours at 4ºC with four changes of PBS. Finally, samples were subjected to chromatography on a Sephadex G-25 column (1 cm diameter x 17 cm height) at 4ºC in the dark. Fractions (150 l each) containing PPS14 or PPS23F complexed with CB or DTAF, respectively, were pooled and stored at -20ºC (Alamo Laboratories Inc, San Antonio, TX and BioCentra, Sugarland, TX) [129]. 7.6 Flow cytometry Peripheral blood was collected from volunteers pre- and post-immunization at days 0, 7, and 30. After Ficoll-gradient centrifugation and washing, cells were resuspended in FACS buffer (PBS, 0.1% FCS, 2mM EDTA). Before staining, cells were absorbed with 10μg/ml cell wall polysaccharide (Statens Serum Institut; MiraVista Diagnostics, Indianapolis, IN) and PPS22F (American Type Culture Collection) to reduce nonspecific binding [159]. B cells were labeled with 10μg/ml, either PPS14-CB or PPS23F-DTAF (Alamo Laboratories Inc, San Antonio, TX and BioCentra, Sugarland, TX). Fluorochrome-conjugated mabs (BD Bioscience or ebioscience) to the following antihuman Ags were used: CD19 (APC-Cy7), CD27 (PerCP-Cy5.5), IgM (allophycocyanin), IgD (Alexa Fluor 700), CD5 (PE-Cy7), CD43 (PE), CD20 (Alexa Fluor 700), CD70 (CB and FITC). Cells were washed and suspended in FACS buffer and analyzed with FACSAria using FACSDiva software (BD Biosciences). FCS files were further analyzed using FlowJo software (Tree Star, Ashland, OR). Singlet lymphocytes were plotted on a histogram to gate on B cells (APC-Cy7:CD19+). B cells were plotted using either 99
123 Cascade Blue: PPS14 or 5-DTAF:PPS23F to identify PPS-selected cells. For studies performed in chapter 2 and 3 (young healthy and elderly), populations were divided into four sub-populations: naïve (CD27-IgM+), class-switched (CD27-IgM-), IgM+ memory (CD27+IgM+), and switched memory (CD27+IgM-) B cells. For studies performed in chapters 4-5 (HIV-positive), populations were divided into three sub-populations: naïve (CD27-), IgM+ memory (CD27+IgM+), and switched memory (CD27+IgM-) B cells. 7.7 Statistical analysis Data is presented as mean ± standard error of measure (SEM). Pre- and post-vaccination data was analyzed using paired t-test. Unpaired t-test was used to compare data between two groups. For analysis involving more than 2 groups, we used Analysis of variance (ANOVA). P-values <0.05 were considered statistically significant. Correlations between two groups were examined using Pearson correlation coefficient. 100
124 References 1. Song, J.H., et al., The relationship between pneumococcal serotypes and antibiotic resistance. Vaccine, (17): p van der Poll, T. and S.M. Opal, Pathogenesis, treatment, and prevention of pneumococcal pneumonia. Lancet, (9700): p Westerink, M.A., H.W. Schroeder, Jr., and M.H. Nahm, Immune Responses to pneumococcal vaccines in children and adults: Rationale for age-specific vaccination. Aging Dis, (1): p Heffernan, R.T., et al., Declining incidence of invasive Streptococcus pneumoniae infections among persons with AIDS in an era of highly active antiretroviral therapy, J Infect Dis, (12): p Breiman, R.F., et al., Pneumococcal bacteremia in Charleston County, South Carolina. A decade later. Arch Intern Med, (7): p Centers for Disease, C. and Prevention, Use of 13-valent pneumococcal conjugate vaccine and 23-valent pneumococcal polysaccharide vaccine for adults with immunocompromising conditions: recommendations of the Advisory Committee on Immunization Practices (ACIP). MMWR Morb Mortal Wkly Rep, (40): p
125 7. Robinson, K.A., et al., Epidemiology of invasive Streptococcus pneumoniae infections in the United States, : Opportunities for prevention in the conjugate vaccine era. JAMA, (13): p Guckian, J.C., G.D. Christensen, and D.P. Fine, The role of opsonins in recovery from experimental pneumococcal pneumonia. J Infect Dis, (2): p Winkelstein, J.A., The role of complement in the host's defense against Streptococcus pneumoniae. Rev Infect Dis, (2): p Baumgarth, N., The double life of a B-1 cell: self-reactivity selects for protective effector functions. Nat Rev Immunol, (1): p Martin, F., A.M. Oliver, and J.F. Kearney, Marginal zone and B1 B cells unite in the early response against T-independent blood-borne particulate antigens. Immunity, (5): p Romero-Steiner, S., et al., Use of opsonophagocytosis for serological evaluation of pneumococcal vaccines. Clin Vaccine Immunol, (2): p Ackerman, M.E., et al., Natural variation in Fc glycosylation of HIV-specific antibodies impacts antiviral activity. J Clin Invest, (5): p Marks, L.R., G.I. Parameswaran, and A.P. Hakansson, Pneumococcal interactions with epithelial cells are crucial for optimal biofilm formation and colonization in vitro and in vivo. Infect Immun, (8): p Gray, B.M., et al., Epidemiologic studies of Streptococcus pneumoniae in infants: antibody response to nasopharyngeal carriage of types 3, 19, and 23. J Infect Dis, (4): p
126 16. Hogberg, L., et al., Age- and serogroup-related differences in observed durations of nasopharyngeal carriage of penicillin-resistant pneumococci. J Clin Microbiol, (3): p CDC. Pneumococcal Disease. Epidemiology and Prevention of Vaccine- Preventable Diseases [cited ]; 12th edition, 2nd printing:[available from: Huang, S.S., et al., Healthcare utilization and cost of pneumococcal disease in the United States. Vaccine, (18): p CDC. Active Bacterial Core Surveillance (ABCs) Report: Emerging Infections Program Network. Streptococcus pneumoniae, provisional Atlanta, GA: US Department of Health and Human Services, CDC; Joyce, E.A., et al., Redefining bacterial populations: a post-genomic reformation. Nat Rev Genet, (6): p Henriques-Normark, B. and E.I. Tuomanen, The pneumococcus: epidemiology, microbiology, and pathogenesis. Cold Spring Harb Perspect Med, (7). 22. Romero-Steiner, S., et al., Reduction in functional antibody activity against Streptococcus pneumoniae in vaccinated elderly individuals highly correlates with decreased IgG antibody avidity [see comments]. Clin Infect Dis, (2): p Schenkein, J.G., S. Park, and M.H. Nahm, Pneumococcal vaccination in older adults induces antibodies with low opsonic capacity and reduced antibody potency. Vaccine, (43): p
127 24. Kim, P., et al., In vivo wide-area cellular imaging by side-view endomicroscopy. Nat Methods, (4): p Ruben, F.L. and M. Uhrin, Specific Immunoglobulin-Class antibody responses in the elderly before and after 14-valent pneumoncoccal vaccine. J Infect Dis, (5): p Pollack, M., et al., Functional properties of isotype-switched immunoglobulin M (IgM) and IgG monoclonal antibodies to Pseudomonas aeruginosa lipopolysaccharide. Infect Immun, (11): p Raff, H.V., et al., Comparison of functional activities between IgG1 and IgM class-switched human monoclonal antibodies reactive with group B streptococci or Escherichia coli K1. J Infect Dis, (2): p Taborda, C.P. and A. Casadevall, Immunoglobulin M efficacy against Cryptococcus neoformans: mechanism, dose dependence, and prozone-like effects in passive protection experiments. J Immunol, (3): p Veneri, D., et al., Changes of human B and B-1a peripheral blood lymphocytes with age. Hematology, (4): p Griffin, D.O., N.E. Holodick, and T.L. Rothstein, Human B1 cells in umbilical cord and adult peripheral blood express the novel phenotype CD20+ CD27+ CD43+ CD70. J Exp Med, (1): p Roederer, M., et al., CD8 naive T cell counts decrease progressively in HIVinfected adults. J Clin Invest, (5): p
128 32. Clerici, M., et al., Detection of three distinct patterns of T helper cell dysfunction in asymptomatic, human immunodeficiency virus-seropositive patients. Independence of CD4+ cell numbers and clinical staging. J Clin Invest, (6): p Martinez-Maza, O., et al., Infection with the human immunodeficiency virus (HIV) is associated with an in vivo increase in B lymphocyte activation and immaturity. J Immunol, (11): p Moir, S., et al., HIV-1 induces phenotypic and functional perturbations of B cells in chronically infected individuals. Proc Natl Acad Sci U S A, (18): p Baldwin, G.C., et al., Human immunodeficiency virus causes mononuclear phagocyte dysfunction. Proc Natl Acad Sci U S A, (10): p Grassi, F., et al., Depletion in blood CD11c-positive dendritic cells from HIVinfected patients. AIDS, (7): p Macatonia, S.E., et al., Dendritic cell infection, depletion and dysfunction in HIVinfected individuals. Immunology, (1): p Redd, S.C., et al., The role of human immunodeficiency virus infection in pneumococcal bacteremia in San Francisco residents. J Infect Dis, (5): p Boschini, A., et al., Community-acquired pneumonia in a cohort of former injection drug users with and without human immunodeficiency virus infection: incidence, etiologies, and clinical aspects. Clin Infect Dis, (1): p Selwyn, P.A., et al., Clinical manifestations and predictors of disease progression in drug users with human immunodeficiency virus infection. N Engl J Med, (24): p
129 41. Schuchat, A., et al., Use of surveillance for invasive pneumococcal disease to estimate the size of the immunosuppressed HIV-infected population. Jama, (24): p Janoff, E.N., et al., Pneumococcal disease during HIV infection. Epidemiologic, clinical, and immunologic perspectives. Ann Intern Med, (4): p Nuorti, J.P., et al., Cigarette smoking and invasive pneumococcal disease. Active Bacterial Core Surveillance Team. N Engl J Med, (10): p Dworkin, M.S., et al., Pneumococcal disease among human immunodeficiency virus-infected persons: incidence, risk factors, and impact of vaccination. Clin Infect Dis, (5): p Grau, I., et al., Epidemiologic changes in bacteremic pneumococcal disease in patients with human immunodeficiency virus in the era of highly active antiretroviral therapy. Arch Intern Med, (13): p Feikin, D.R., et al., Global strategies to prevent bacterial pneumonia in adults with HIV disease. Lancet Infect Dis, (7): p Rainwater-Lovett, K., et al., Antiretroviral therapy restores age-dependent loss of resting memory B cells in young HIV-infected Zambian children. J Acquir Immune Defic Syndr, Li, T.S., et al., Long-lasting recovery in CD4 T-cell function and viral-load reduction after highly active antiretroviral therapy in advanced HIV-1 disease. Lancet, (9117): p Bogaert, D., R. De Groot, and P.W. Hermans, Streptococcus pneumoniae colonisation: the key to pneumococcal disease. Lancet Infect Dis, (3): p
130 50. Tuomanen, E.I., Pathogenesis of pneumococcal inflammation: otitis media. Vaccine, Suppl 1: p. S Nakamura, S., K.M. Davis, and J.N. Weiser, Synergistic stimulation of type I interferons during influenza virus coinfection promotes Streptococcus pneumoniae colonization in mice. J Clin Invest, (9): p Berry, A.M. and J.C. Paton, Additive attenuation of virulence of Streptococcus pneumoniae by mutation of the genes encoding pneumolysin and other putative pneumococcal virulence proteins. Infect Immun, (1): p Canvin, J.R., et al., The role of pneumolysin and autolysin in the pathology of pneumonia and septicemia in mice infected with a type 2 pneumococcus. J Infect Dis, (1): p Thigpen, M.C., et al., Bacterial meningitis in the United States, N Engl J Med, (21): p Nelson, A.L., et al., Capsule enhances pneumococcal colonization by limiting mucus-mediated clearance. Infect Immun, (1): p Kadioglu, A., et al., The role of Streptococcus pneumoniae virulence factors in host respiratory colonization and disease. Nat Rev Microbiol, (4): p Gould, J.M. and J.N. Weiser, The inhibitory effect of C-reactive protein on bacterial phosphorylcholine platelet-activating factor receptor-mediated adherence is blocked by surfactant. J Infect Dis, (3): p Tuomanen, E.I., R. Austrian, and H.R. Masure, Pathogenesis of pneumococcal infection. N Engl J Med, (19): p
131 59. Weber, J.R., et al., Recognition of pneumococcal peptidoglycan: an expanded, pivotal role for LPS binding protein. Immunity, (2): p Akira, S., Pathogen recognition by innate immunity and its signaling. Proc Jpn Acad Ser B Phys Biol Sci, (4): p Yoshimura, A., et al., Cutting edge: recognition of Gram-positive bacterial cell wall components by the innate immune system occurs via Toll-like receptor 2. J Immunol, (1): p Malley, R., et al., Recognition of pneumolysin by Toll-like receptor 4 confers resistance to pneumococcal infection. Proc Natl Acad Sci U S A, (4): p Albiger, B., et al., Toll-like receptor 9 acts at an early stage in host defence against pneumococcal infection. Cell Microbiol, (3): p Musher, D.L., B. Johnson Jr, and D.A. Watson, Quantitative relationship between anticapsular antibody measured by enzyme-linked immunosorbent assay of radioimmunoassay and protection of mice against challenge with Streptococcus pneumoniae serotype 4. Infect Immun, : p Bruyn, G.A., B.J. Zegers, and R. van Furth, Mechanisms of host defense against infection with Streptococcus pneumoniae. Clin Infect Dis, (1): p Brouwer, M.C., et al., Host genetic susceptibility to pneumococcal and meningococcal disease: a systematic review and meta-analysis. Lancet Infect Dis, (1): p
132 67. Davis, K.M., S. Nakamura, and J.N. Weiser, Nod2 sensing of lysozyme-digested peptidoglycan promotes macrophage recruitment and clearance of S. pneumoniae colonization in mice. J Clin Invest, (9): p Brown, J.S., et al., The classical pathway is the dominant complement pathway required for innate immunity to Streptococcus pneumoniae infection in mice. Proc Natl Acad Sci U S A, (26): p Dunkelberger, J.R. and W.C. Song, Complement and its role in innate and adaptive immune responses. Cell Res, (1): p Johnston, R.B., Jr., The host response to invasion by Streptococcus pneumoniae: protection and the pathogenesis to tissue damage. Rev Infect Dis, (2): p Mond, J.J., A. Lees, and C.M. Snapper, T cell-independent antigens type 2. Annu Rev Immunol, : p Yammani, R.D. and K.M. Haas, Primate B-1 cells generate antigen-specific B cell responses to T cell-independent type 2 antigens. J Immunol, (7): p Styrt, B., Infection associated with asplenia: risks, mechanisms, and prevention. Am J Med, (5N): p. 33N-42N. 74. Wara, D.W., Host defense against Streptococcus pneumoniae: the role of the spleen. Rev Infect Dis, (2): p Wardemann, H., et al., B-1a B cells that link the innate and adaptive immune responses are lacking in the absence of the spleen. J Exp Med, (6): p Weller, S., et al., Human blood IgM "memory" B cells are circulating splenic marginal zone B cells harboring a prediversified immunoglobulin repertoire. Blood, (12): p
133 77. Baumgarth, N., et al., B-1 and B-2 cell-derived immunoglobulin M antibodies are nonredundant components of the protective response to influenza virus infection. J Exp Med, (2): p Boes, M., et al., A critical role of natural immunoglobulin M in immediate defense against systemic bacterial infection. J Exp Med, (12): p Briles, D.E., et al., Antiphosphocholine antibodies found in normal mouse serum are protective against intravenous infection with type 3 streptococcus pneumoniae. J Exp Med, (3): p Ochsenbein, A.F., et al., Control of early viral and bacterial distribution and disease by natural antibodies. Science, (5447): p Haas, K.M., et al., B-1a and B-1b cells exhibit distinct developmental requirements and have unique functional roles in innate and adaptive immunity to S. pneumoniae. Immunity, (1): p Griffin, D.O., N.E. Holodick, and T.L. Rothstein, Human B1 cells are CD3-: A reply to "A human equivalent of mouse B-1 cells?" and "The nature of circulating CD27+CD43+ B cells". J Exp Med, (13): p Verbinnen, B., et al., Human CD20+CD43+CD27+CD5- B cells generate antibodies to capsular polysaccharides of Streptococcus pneumoniae. J Allergy Clin Immunol, (1): p Covens, K., et al., Characterization of proposed human B-1 cells reveals preplasmablast phenotype. Blood, (26): p Novak, R., et al., Emergence of vancomycin tolerance in Streptococcus pneumoniae. Nature, (6736): p
134 86. Garcia-Suarez Mdel, M., F. Vazquez, and F.J. Mendez, Streptococcus pneumoniae virulence factors and their clinical impact: An update. Enferm Infecc Microbiol Clin, (8): p Taillardet, M., et al., The thymus-independent immunity conferred by a pneumococcal polysaccharide is mediated by long-lived plasma cells. Blood, (20): p Musher, D.M., et al., Safety and antibody response, including antibody persistence for 5 years, after primary vaccination or revaccination with pneumococcal polysaccharide vaccine in middle-aged and older adults. J Infect Dis, (4): p Musher, D.M., et al., Antibody persistence ten years after first and second doses of 23-valent pneumococcal polysaccharide vaccine, and immunogenicity and safety of second and third doses in older adults. Hum Vaccin, (9): p Ortqvist, A., Pneumococcal vaccination: current and future issues. Eur Respir J, (1): p Butler, J.C., et al., Pneumococcal polysaccharide vaccine efficacy. An evaluation of current recommendations. JAMA, (15): p Shapiro, E.D., et al., The protective efficacy of polyvalent pneumococcal polysaccharide vaccine. N Engl J Med, (21): p Esposito, S., et al., Safety and immunogenicity of a 13-valent pneumococcal conjugate vaccine compared to those of a 7-valent pneumococcal conjugate vaccine given as a three-dose series with routine vaccines in healthy infants and toddlers. Clin Vaccine Immunol, (6): p
135 94. Simberkoff, M.S., et al., Efficacy of pneumococcal vaccine in high-risk patients. Results of a Veterans Administration Cooperative Study. N Engl J Med, (21): p Hirschmann, J.V. and B.A. Lipsky, The pneumococcal vaccine after 15 years of use. Arch Intern Med, (4): p Fein, A.M., S.H. Feinsilver, and M.S. Niederman, Atypical manifestations of pneumonia in the elderly. Clin Chest Med, (2): p Jackson, L.A., et al., Effectiveness of pneumococcal polysaccharide vaccine in older adults. N Engl J Med, (18): p Moore, R.A., P.J. Wiffen, and B.A. Lipsky, Are the pneumococcal polysaccharide vaccines effective? Meta-analysis of the prospective trials. BMC Fam Pract, : p Cornu, C., et al., Efficacy of pneumococcal polysaccharide vaccine in immunocompetent adults: a meta-analysis of randomized trials. Vaccine, (32): p Fine, M.J., et al., Efficacy of pneumococcal vaccination in adults. A meta-analysis of randomized controlled trials. Arch Intern Med, (23): p Watson, L., B.J. Wilson, and N. Waugh, Pneumococcal polysaccharide vaccine: a systematic review of clinical effectiveness in adults. Vaccine, (17-18): p Shi, Y., et al., Regulation of aged humoral immune defense against pneumococcal bacteria by IgM memory B cell. J Immunol, (5): p
136 103. Kruetzmann, S., et al., Human immunoglobulin M memory B cells controlling Streptococcus pneumoniae infections are generated in the spleen. J Exp Med, (7): p Colonna-Romano, G., et al., B cells in the aged: CD27, CD5, and CD40 expression. Mech Ageing Dev, (4): p Lane, H.C., et al., Abnormalities of B-cell activation and immunoregulation in patients with the acquired immunodeficiency syndrome. N Engl J Med, (8): p Terpstra, F.G., et al., Longitudinal study of leukocyte functions in homosexual men seroconverted for HIV: rapid and persistent loss of B cell function after HIV infection. Eur J Immunol, (4): p Miedema, F., et al., Immunological abnormalities in human immunodeficiency virus (HIV)-infected asymptomatic homosexual men. HIV affects the immune system before CD4+ T helper cell depletion occurs. J Clin Invest, (6): p Schnittman, S.M., et al., Direct polyclonal activation of human B lymphocytes by the acquired immune deficiency syndrome virus. Science, (4768): p De Milito, A., B lymphocyte dysfunctions in HIV infection. Curr HIV Res, (1): p Berberian, L., et al., Immunoglobulin VH3 gene products: natural ligands for HIV gp120. Science, (5128): p Rieckmann, P., et al., Recombinant gp120 specifically enhances tumor necrosis factor-alpha production and Ig secretion in B lymphocytes from HIV-infected individuals but not from seronegative donors. J Immunol, (9): p
137 112. Centers for Disease, C., Prevention, and P. Advisory Committee on Immunization, Updated recommendations for prevention of invasive pneumococcal disease among adults using the 23-valent pneumococcal polysaccharide vaccine (PPSV23). MMWR Morb Mortal Wkly Rep, (34): p Ochs, H.D., et al., Abnormal antibody responses in patients with persistent generalized lymphadenopathy. J Clin Immunol, (1): p Kroon, F.P., et al., Enhanced antibody response to pneumococcal polysaccharide vaccine after prior immunization with conjugate pneumococcal vaccine in HIV-infected adults. Vaccine, (7-8): p Opravil, M., et al., Poor antibody response after tetanus and pneumococcal vaccination in immunocompromised, HIV-infected patients. Clin Exp Immunol, (2): p Ballet, J.J., et al., Impaired anti-pneumococcal antibody response in patients with AIDS-related persistent generalized lymphadenopathy. Clin Exp Immunol, (3): p Janoff, E.N., et al., Class-specific antibody response to pneumococcal capsular polysaccharides in men infected with human immunodeficiency virus type 1. J Infect Dis, (5): p Ragni, M.V., et al., Antibody responses to immunization of patients with hemophilia with and without evidence of human immunodeficiency virus (human T- lymphotropic virus type III) infection. J Lab Clin Med, (5): p
138 119. Rodriguez-Barradas, M.C., et al., Antibody to capsular polysaccharides of Streptococcus pneumoniae after vaccination of human immunodeficiency virus-infected subjects with 23-valent pneumococcal vaccine. J Infect Dis, (3): p Loeliger, A.E., et al., Deficient antipneumococcal polysaccharide responses in HIV-seropositive patients. FEMS Immunol Med Microbiol, (1): p Klein, R.S., et al., Response to pneumococcal vaccine among asymptomatic heterosexual partners of persons with AIDS and intravenous drug users infected with human immunodeficiency virus. J Infect Dis, (5): p Unsworth, D.J., et al., Defective IgG2 response to Pneumovax in HIV seropositive patients. Genitourin Med, (5): p French, N., et al., Pneumococcal vaccination in HIV-1-infected adults in Uganda: humoral response and two vaccine failures. AIDS, (13): p Feikin, D.R., et al., Randomized trial of the quantitative and functional antibody responses to a 7-valent pneumococcal conjugate vaccine and/or 23-valent polysaccharide vaccine among HIV-infected adults. Vaccine, (3-4): p Madhi, S.A., et al., Quantitative and qualitative antibody response to pneumococcal conjugate vaccine among African human immunodeficiency virus-infected and uninfected children. Pediatr Infect Dis J, (5): p Janoff, E.N., et al., Streptococcus pneumoniae colonization, bacteremia, and immune response among persons with human immunodeficiency virus infection. J Infect Dis, (1): p D'Orsogna, L.J., et al., Circulating memory B-cell subpopulations are affected differently by HIV infection and antiretroviral therapy. Aids, (13): p
139 128. Hart, M., et al., Loss of discrete memory B cell subsets is associated with impaired immunization responses in HIV-1 infection and may be a risk factor for invasive pneumococcal disease. J Immunol, (12): p Khaskhely, N., et al., Phenotypic analysis of pneumococcal polysaccharidespecific B cells. J Immunol, (5): p Lifson, A.R., et al., Long-term CD4+ lymphocyte response following HAART initiation in a U.S. Military prospective cohort. AIDS Res Ther, (1): p Michaels, S.H., R. Clark, and P. Kissinger, Declining morbidity and mortality among patients with advanced human immunodeficiency virus infection. N Engl J Med, (6): p Hung, C.C. and S.C. Chang, Impact of highly active antiretroviral therapy on incidence and management of human immunodeficiency virus-related opportunistic infections. J Antimicrob Chemother, (5): p Gougeon, M.L., Apoptosis as an HIV strategy to escape immune attack. Nat Rev Immunol, (5): p Longwe, H., et al., Characterising B cell numbers and memory B cells in HIV infected and uninfected Malawian adults. BMC Infect Dis, : p Titanji, K., et al., Loss of memory B cells impairs maintenance of long-term serologic memory during HIV-1 infection. Blood, (5): p Jordano, Q., et al., Invasive pneumococcal disease in patients infected with HIV: still a threat in the era of highly active antiretroviral therapy. Clin Infect Dis, (11): p
140 137. Musher, D.M., B. Johnson, Jr., and D.A. Watson, Quantitative relationship between anticapsular antibody measured by enzyme-linked immunosorbent assay or radioimmunoassay and protection of mice against challenge with Streptococcus pneumoniae serotype 4. Infect Immun, (12): p Romero-Steiner, S., et al., Reduction in functional antibody activity against Streptococcus pneumoniae in vaccinated elderly individuals highly correlates with decreased IgG antibody avidity. Clin Infect Dis, (2): p Nahm, M.H., J.V. Olander, and M. Magyarlaki, Identification of cross-reactive antibodies with low opsonophagocytic activity for Streptococcus pneumoniae. J Infect Dis, (3): p O'Brien, K.L., M. Hochman, and D. Goldblatt, Combined schedules of pneumococcal conjugate and polysaccharide vaccines: is hyporesponsiveness an issue? Lancet Infect Dis, (9): p Torling, J., et al., Revaccination with the 23-valent pneumococcal polysaccharide vaccine in middle-aged and elderly persons previously treated for pneumonia. Vaccine, (1): p Davidson, M., et al., Immunogenicity of pneumococcal revaccination in patients with chronic disease. Arch Intern Med, (19): p Hammitt, L.L., et al., Repeat revaccination with 23-valent pneumococcal polysaccharide vaccine among adults aged years living in Alaska: no evidence of hyporesponsiveness. Vaccine, (12): p
141 144. Leggat, D.J., et al., The Immune Response to Pneumococcal Polysaccharides 14 and 23F Among Elderly Individuals Consists Predominantly of Switched Memory B Cells. J Infect Dis, Wartha, F., et al., Capsule and D-alanylated lipoteichoic acids protect Streptococcus pneumoniae against neutrophil extracellular traps. Cell Microbiol, (5): p Timens, W., et al., Immaturity of the human splenic marginal zone in infancy. Possible contribution to the deficient infant immune response. J Immunol, (10): p Zandvoort, A. and W. Timens, The dual function of the splenic marginal zone: essential for initiation of anti-ti-2 responses but also vital in the general first-line defense against blood-borne antigens. Clin Exp Immunol, (1): p Weill, J.C., S. Weller, and C.A. Reynaud, Human marginal zone B cells. Annu Rev Immunol, : p Puga, I., et al., B cell-helper neutrophils stimulate the diversification and production of immunoglobulin in the marginal zone of the spleen. Nat Immunol, (2): p Alugupalli, K.R., et al., B1b lymphocytes confer T cell-independent long-lasting immunity. Immunity, (3): p Defrance, T., M. Taillardet, and L. Genestier, T cell-independent B cell memory. Curr Opin Immunol, (3): p
142 152. Haas, K.M., Programmed cell death 1 suppresses B-1b cell expansion and longlived IgG production in response to T cell-independent type 2 antigens. J Immunol, (10): p Obukhanych, T.V. and M.C. Nussenzweig, T-independent type II immune responses generate memory B cells. J Exp Med, (2): p LeBien, T.W. and T.F. Tedder, B lymphocytes: how they develop and function. Blood, (5): p Carsetti, R., M.M. Rosado, and H. Wardmann, Peripheral development of B cells in mouse and man. Immunol Rev, : p Lee, J., et al., Identification and characterization of a human CD5+ pre-naive B cell population. J Immunol, (7): p Griffin, D.O. and T.L. Rothstein, Human b1 cell frequency: isolation and analysis of human b1 cells. Front Immunol, : p Nahm, M.G.D., Training manual for enzyme linked immunosobent assay for the quantitation of Streptococcus pneumoniae serotype specific IgG (PnPg ELISA) Concepcion, N.F. and C.E. Frasch, Pneumococcal type 22f polysaccharide absorption improves the specificity of a pneumococcal-polysaccharide enzyme-linked immunosorbent assay. Clin Diagn Lab Immunol, (2): p Kolibab, K., et al., Immune response to pneumococcal polysaccharides 4 and 14 in elderly and young adults. I. Antibody concentrations, avidity and functional activity. Immun Ageing, : p
143 161. Park, S. and M.H. Nahm, Older adults have a low capacity to opsonize pneumococci due to low IgM antibody response to pneumococcal vaccinations. Infect Immun, (1): p Go, E.S. and Z.K. Ballas, Anti-pneumococcal antibody response in normal subjects: a meta-analysis. J Allergy Clin Immunol, (1): p Moens, L., et al., Human memory B lymphocyte subsets fulfill distinct roles in the anti-polysaccharide and anti-protein immune response. J Immunol, (8): p Leggat, D.J., et al., The immune response to pneumococcal polysaccharides 14 and 23F among elderly individuals consists predominantly of switched memory B cells. J Infect Dis, (1): p Recommendations of the Immunization Practices Advisory Committee Pneumococcal Polysaccharide Vaccine. MMWR, (5): p , Jackson L. A., N.K.M., Yu O., Benson P., Barlow W. E., Adams A. L., Hanson C. A., Mahoney L. D., Shay D. K., Thompson W. W., Effectiveness of Pneumococcal Polysaccharide Vaccine in Older Adults. N Engl J Med, : p Shapiro, E.D., et al., The protective efficacy of polyvalent pneumococcal polysaccharide vaccine. New Engl J Med, (21): p Simberkoff, M.S., et al., Efficacy of pneumococcal vaccine in high-risk patients. Results of a Veterans Administration Cooperative Study. N Engl J Med, : p Roghmann, K.J., et al., Immune response of elderly adults to pneumococcus: variation by age, sex, and functional impairment. J Gerontol, (3): p
144 170. Sankilampi, U., et al., Persistence of antibodies to pneumococcal capsular polysaccharide vaccine in the elderly. J Infect Dis, (4): p Kolibab, K., et al., Immune response to pneumococcal polysaccharides 4 and 14 in elderly and young adults: analysis of the variable heavy chain repertoire. Infect Immun, (11): p Lucas, A.H., et al., Combinatorial library cloning of human antibodies to Streptococcus pneumoniae capsular polysaccharides: variable region primary structures and evidence for somatic mutation of Fab fragments specific for capsular serotypes 6B, 14, and 23F. Infect Immun, (2): p Zhou, J., et al., Somatic hypermutation and diverse immunoglobulin gene usage in the human antibody response to the capsular polysaccharide of Streptococcus pneumoniae Type 6B. Infect Immun, (6): p Carsetti, R., et al., The loss of IgM memory B cells correlates with clinical disease in common variable immunodeficiency. J Allergy Clin Immunol, (2): p Moberley, S.A., et al., Vaccines for preventing pneumococcal infection in adults. Cochrane Database Syst Rev, 2008(1): p. CD Lee, H., M.H. Nahm, and K.H. Kim, The effect of age on the response to the pneumococcal polysaccharide vaccine. BMC Infect Dis, : p Howard, W.A., K.L. Gibson, and D.K. Dunn-Walters, Antibody quality in old age. Rejuvenation Res, (1): p Ademokun A, W.Y., Martin V, Mitra R, Sack U, Baxendale H, Kipling D, Dunn- Walters DK., Vaccination-induced changes in human B-cell repertoire and 121
145 pneumococcal IgM and IgA antibody at different ages. Aging cell, (6): p Pneumococcal vaccines: WHO position paper. Weekly Epidemiological Record, : p Fattal-German, M., et al., Pneumococcal vaccination of elderly individuals. Vaccine, (8): p Klein, U., K. Rajewsky, and R. Kuppers, Human immunoglobulin (Ig)M+IgD+ peripheral blood B cells expressing the CD27 cell surface antigen carry somatically mutated variable region genes: CD27 as a general marker for somatically mutated (memory) B cells. J Exp Med, (9): p Frasca, D., et al., Aging down-regulates the transcription factor E2A, activationinduced cytidine deaminase, and Ig class switch in human B cells. J Immunol, (8): p Clutterbuck, E.A., et al., Pneumococcal conjugate and plain polysaccharide vaccines have divergent effects on antigen-specific B cells. J Infect Dis, (9): p Heilmann, C. and F.K. Pedersen, Quantitation of blood lymphocytes secreting antibodies to pneumococcal polysaccharides after in vivo antigenic stimulation. Scand J Immunol, (2): p Kehrl, J.H. and A.S. Fauci, Activation of human B lymphocytes after immunization with pneumococcal polysaccharides. J Clin Invest, (4): p
146 186. Paradiso, P.R., Pneumococcal conjugate vaccine for adults: a new paradigm. Clin Infect Dis, (2): p Barry, P.M., et al., Invasive pneumococcal disease in a cohort of HIV-infected adults: incidence and risk factors, AIDS, (3): p Foster, D., et al., Invasive pneumococcal disease: epidemiology in children and adults prior to implementation of the conjugate vaccine in the Oxfordshire region, England. J Med Microbiol, (Pt 4): p Crum-Cianflone, N.F., et al., A randomized clinical trial comparing revaccination with pneumococcal conjugate vaccine to polysaccharide vaccine among HIV-infected adults. J Infect Dis, (7): p Pedersen, R.H., et al., The effectiveness of pneumococcal polysaccharide vaccination in HIV-infected adults: a systematic review. HIV Med, (6): p Prevention of pneumococcal disease: recommendations of the Advisory Committee on Immunization Practices (ACIP). MMWR Recomm Rep, (RR-8): p Hung, C.C., et al., A 5-year longitudinal follow-up study of serological responses to 23-valent pneumococcal polysaccharide vaccination among patients with HIV infection who received highly active antiretroviral therapy. HIV Med, (1): p Rodriguez-Barradas, M.C., et al., Response of human immunodeficiency virusinfected patients receiving highly active antiretroviral therapy to vaccination with 23- valent pneumococcal polysaccharide vaccine. Clin Infect Dis, (3): p
147 194. Lynch, J.P., 3rd and G.G. Zhanel, Streptococcus pneumoniae: does antimicrobial resistance matter? Semin Respir Crit Care Med, (2): p Cagigi, A., et al., Dysfunctional B-cell responses during HIV-1 infection: implication for influenza vaccination and highly active antiretroviral therapy. Lancet Infect Dis, (7): p De Milito, A., et al., Loss of memory (CD27) B lymphocytes in HIV-1 infection. AIDS, (8): p Carson, P.J., et al., Antibody class and subclass responses to pneumococcal polysaccharides following immunization of human immunodeficiency virus-infected patients. J Infect Dis, (2): p Leggat, D.J., et al., Pneumococcal polysaccharide vaccination induces polysaccharide-specific B cells in adult peripheral blood expressing CD19(+)CD20(+)CD3(-)CD70(-)CD27(+)IgM(+)CD43(+)CD5(+)/(-). Vaccine, (41): p Moir, S. and A.S. Fauci, B cells in HIV infection and disease. Nat Rev Immunol, (4): p Hung, C.C., et al., Clinical experience of the 23-valent capsular polysaccharide pneumococcal vaccination in HIV-1-infected patients receiving highly active antiretroviral therapy: a prospective observational study. Vaccine, (15-16): p Teshale, E.H., et al., Effectiveness of 23-valent polysaccharide pneumococcal vaccine on pneumonia in HIV-infected adults in the United States, Vaccine, (46): p
148 202. Penaranda, M., et al., Effectiveness of polysaccharide pneumococcal vaccine in HIV-infected patients: a case-control study. Clin Infect Dis, (7): p. e French, N., et al., 23-valent pneumococcal polysaccharide vaccine in HIV-1- infected Ugandan adults: double-blind, randomised and placebo controlled trial. Lancet, (9221): p Lopez-Palomo, C., et al., Pneumonia in HIV-infected patients in the HAART era: incidence, risk, and impact of the pneumococcal vaccination. J Med Virol, (4): p Veras, M.A., et al., Effectiveness of the polysaccharide pneumococcal vaccine among HIV-infected persons in Brazil: a case control study. BMC Infect Dis, : p Kaplan, J.E., et al., Guidelines for prevention and treatment of opportunistic infections in HIV-infected adults and adolescents: recommendations from CDC, the National Institutes of Health, and the HIV Medicine Association of the Infectious Diseases Society of America. MMWR Recomm Rep, (RR-4): p ; quiz CE Kumar, D., et al., Randomized, double-blind, controlled trial of pneumococcal vaccination in renal transplant recipients. J Infect Dis, (10): p Pensieroso, S., et al., Timing of HAART defines the integrity of memory B cells and the longevity of humoral responses in HIV-1 vertically-infected children. Proc Natl Acad Sci U S A, (19): p
149 209. Romero-Steiner, S., et al., Standardization of an opsonophagocytic assay for the measurement of functional antibody activity against Streptococcus pneumoniae using differentiated HL-60 cells. Clin Diagn Lab Immunol, (4): p Lederman, H.M., et al., Incomplete immune reconstitution after initiation of highly active antiretroviral therapy in human immunodeficiency virus-infected patients with severe CD4+ cell depletion. J Infect Dis, (12): p Santosuosso, M., et al., HIV-1 envelope protein gp120 is present at high concentrations in secondary lymphoid organs of individuals with chronic HIV-1 infection. J Infect Dis, (7): p Pierson, T., J. McArthur, and R.F. Siliciano, Reservoirs for HIV-1: mechanisms for viral persistence in the presence of antiviral immune responses and antiretroviral therapy. Annu Rev Immunol, : p Borrow, R., et al., Serological basis for use of meningococcal serogroup C conjugate vaccines in the United Kingdom: reevaluation of correlates of protection. Infect Immun, (3): p Takizawa, M., K. Sugane, and K. Agematsu, Role of tonsillar IgD+CD27+ memory B cells in humoral immunity against pneumococcal infection. Hum Immunol, (12): p Breiman, R.F., et al., Evaluation of effectiveness of the 23-valent pneumococcal capsular polysaccharide vaccine for HIV-infected patients. Arch Intern Med, (17): p Dieye, T.N., et al., 23-valent pneumococcal vaccination and HIV. Lancet, (9234): p
150 217. Sereti, I., et al., Decreases in colonic and systemic inflammation in chronic HIV infection after IL-7 administration. PLoS Pathog, (1): p. e Nasi, M., et al., Persistent inflammation in HIV infection: Established concepts, new perspectives. Immunol Lett, Andrews, S.F. and P.C. Wilson, The anergic B cell. Blood, (24): p Nahm, G.D., Training manual for Enzyme linked immunosorbent assay for the quantitation of Streptococcus pneumoniae serotype specific IgG (Pn PS ELISA) Nahm, M., Training manual for enzyme linked immunosorbent assay for the quantitation of Streprococcus pneumoniae serotype specific IgG (Pn PS ELISA) Moon Nahm, D.G., Training manual for Enzyme linked immunosorbent assay for the quantitation of Streptococcus pneumoniae serotype specific IgG (Pn Ps ELISA). (89SF Version). 2006: p Moon Nahm, D.G., Training manual for Enzyme linked immunosorbent assay for the quantitation of Streptococcus pneumoniae serotype specific IgG (Pn Ps ELISA). (007 Version). 2011: p
151 Appendix A Pneumococcal polysaccharide vaccination induces polysaccharide-specific B cells in adult peripheral blood expressing CD19+CD20+CD3-CD70-CD27+IgM+ CD43+CD5+/- This article was originally published in Vaccine. David J. Leggat, Noor M. Khaskhely, Anita S. Iyer, Jason Mosakowski, Rebecca S. Thompson, John D. Weinandy, M.A. Julie Westerink Pneumococcal polysaccharide vaccination induces polysaccharidespecific B cells in adult peripheral blood expressing CD19+CD20+CD3-CD70- CD27+IgM+CD43+CD5+/-. Vaccine. 31(41): p Copyright 2013 Elsevier Ltd. Link to article on Vaccine website: Authors retain the permissions to include the final, published version of the article in a dissertation and may be deposited into a repository as long as a link to the final published version hosted on Science Direct is provided. 128



152 129


153 130

154 131

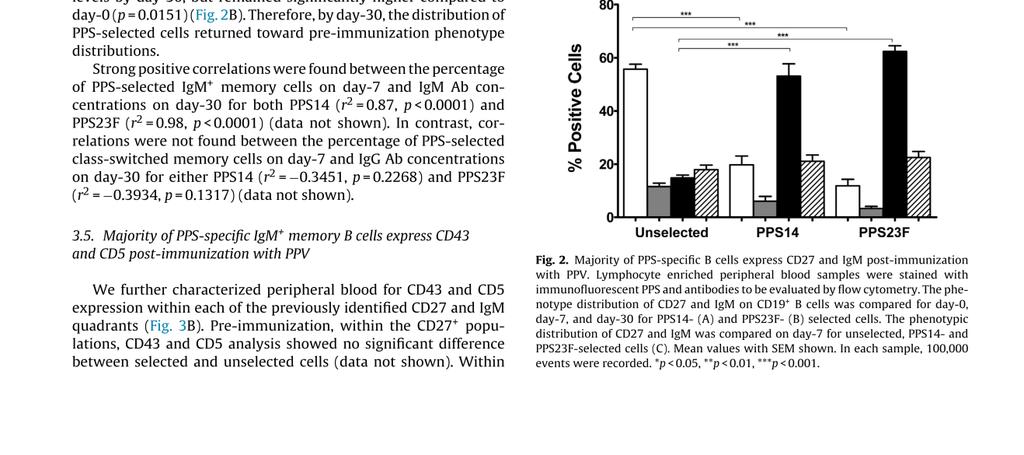
155 132


156 133


157 134

158 135


159 136

160 137

161 138
162 Appendix B The elderly immune response to pneumococcal polysaccharides 14 and 23F consists predominantly of switched memory B cells This article was originally published in the Journal of Infectious Disease. David J. Leggat, Rebecca S. Thompson, Noor M. Khaskhely, Anita S. Iyer, M.A. Julie Westerink The elderly immune response to pneumococcal polysaccharides 14 and 23F consists predominantly of switched memory B cells. J Infect Dis. Jul;208(1): Copyright 2013 Oxford University Press on behalf of the Infectious Diseases Society of America. Link to article on Oxford Journals website: Authors retain the permissions to include the final, published version of the article in a dissertation and may be deposited into a repository as long as a link to the final published version hosted on Oxford University Press is provided. 139
163 140

164 141


165 142


166 143
167 144


168 145

169 146
Streptococcus pneumonia
 Streptococcus pneumonia The pneumococci (S. pneumoniae) are gram-positive diplococci. Often lancet shaped or arranged in chains, possessing a capsule of polysaccharide that permits typing with specific
Streptococcus pneumonia The pneumococci (S. pneumoniae) are gram-positive diplococci. Often lancet shaped or arranged in chains, possessing a capsule of polysaccharide that permits typing with specific
Secretory antibodies in the upper respiratory tract
 Secretory antibodies in the upper respiratory tract B lymphocytes IgM (pneumococcus) Dimeric IgA J chain Polymeric immunoglobulin receptor (PigR) Polysaccharide capsule Epithelial cell Basolateral Secretory
Secretory antibodies in the upper respiratory tract B lymphocytes IgM (pneumococcus) Dimeric IgA J chain Polymeric immunoglobulin receptor (PigR) Polysaccharide capsule Epithelial cell Basolateral Secretory
ADI_Res_Bull_2013_Pneumococcal_Vaccine_Tests
 ADI_Res_Bull_2013_Pneumococcal_Vaccine_Tests Most non-vaccinated humans and some animals have a natural exposure to non-virulent strains Streptococcus pneumonia, and therefore contain high levels of antibodies
ADI_Res_Bull_2013_Pneumococcal_Vaccine_Tests Most non-vaccinated humans and some animals have a natural exposure to non-virulent strains Streptococcus pneumonia, and therefore contain high levels of antibodies
Pneumococcal vaccination in aging HIV-infected individuals
 The University of Toledo The University of Toledo Digital Repository Theses and Dissertations 2015 Pneumococcal vaccination in aging HIV-infected individuals Jennifer A. Ohtola University of Toledo Follow
The University of Toledo The University of Toledo Digital Repository Theses and Dissertations 2015 Pneumococcal vaccination in aging HIV-infected individuals Jennifer A. Ohtola University of Toledo Follow
Streptococcus pneumoniae CDC
 Streptococcus pneumoniae CDC Pneumococcal Disease Infection caused by the bacteria, Streptococcus pneumoniae» otitis media 20 million office visits (28-55% Strep)» pneumonia 175,000 cases annually» meningitis
Streptococcus pneumoniae CDC Pneumococcal Disease Infection caused by the bacteria, Streptococcus pneumoniae» otitis media 20 million office visits (28-55% Strep)» pneumonia 175,000 cases annually» meningitis
Medical Virology Immunology. Dr. Sameer Naji, MB, BCh, PhD (UK) Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University
 Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University") Medical Virology Immunology Dr. Sameer Naji, MB, BCh, PhD (UK) Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University Human blood cells Phases of immune responses Microbe Naïve
Medical Virology Immunology Dr. Sameer Naji, MB, BCh, PhD (UK) Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University Human blood cells Phases of immune responses Microbe Naïve
Haemophilus influenzae
 Haemophilus influenzae type b Severe bacterial infection, particularly among infants During late 19th century believed to cause influenza Immunology and microbiology clarified in 1930s Haemophilus influenzae
Haemophilus influenzae type b Severe bacterial infection, particularly among infants During late 19th century believed to cause influenza Immunology and microbiology clarified in 1930s Haemophilus influenzae
Unit 5 The Human Immune Response to Infection
 Unit 5 The Human Immune Response to Infection Unit 5-page 1 FOM Chapter 21 Resistance and the Immune System: Innate Immunity Preview: In Chapter 21, we will learn about the branch of the immune system
Unit 5 The Human Immune Response to Infection Unit 5-page 1 FOM Chapter 21 Resistance and the Immune System: Innate Immunity Preview: In Chapter 21, we will learn about the branch of the immune system
Adaptive Immunity: Humoral Immune Responses
 MICR2209 Adaptive Immunity: Humoral Immune Responses Dr Allison Imrie 1 Synopsis: In this lecture we will review the different mechanisms which constitute the humoral immune response, and examine the antibody
MICR2209 Adaptive Immunity: Humoral Immune Responses Dr Allison Imrie 1 Synopsis: In this lecture we will review the different mechanisms which constitute the humoral immune response, and examine the antibody
Pneumococcal Disease and Pneumococcal Vaccines
 Pneumococcal Disease and Epidemiology and Prevention of - Preventable Diseases Note to presenters: Images of vaccine-preventable diseases are available from the Immunization Action Coalition website at
Pneumococcal Disease and Epidemiology and Prevention of - Preventable Diseases Note to presenters: Images of vaccine-preventable diseases are available from the Immunization Action Coalition website at
Immunobiology 7. The Humoral Immune Response
 Janeway Murphy Travers Walport Immunobiology 7 Chapter 9 The Humoral Immune Response Copyright Garland Science 2008 Tim Worbs Institute of Immunology Hannover Medical School 1 The course of a typical antibody
Janeway Murphy Travers Walport Immunobiology 7 Chapter 9 The Humoral Immune Response Copyright Garland Science 2008 Tim Worbs Institute of Immunology Hannover Medical School 1 The course of a typical antibody
Chapter 24 The Immune System
 Chapter 24 The Immune System The Immune System Layered defense system The skin and chemical barriers The innate and adaptive immune systems Immunity The body s ability to recognize and destroy specific
Chapter 24 The Immune System The Immune System Layered defense system The skin and chemical barriers The innate and adaptive immune systems Immunity The body s ability to recognize and destroy specific
محاضرة مناعت مدرس المادة :ا.م. هدى عبدالهادي علي النصراوي Immunity to Infectious Diseases
 محاضرة مناعت مدرس المادة :ا.م. هدى عبدالهادي علي النصراوي Immunity to Infectious Diseases Immunity to infection depends on a combination of innate mechanisms (phagocytosis, complement, etc.) and antigen
محاضرة مناعت مدرس المادة :ا.م. هدى عبدالهادي علي النصراوي Immunity to Infectious Diseases Immunity to infection depends on a combination of innate mechanisms (phagocytosis, complement, etc.) and antigen
Question 1. Kupffer cells, microglial cells and osteoclasts are all examples of what type of immune system cell?
 Abbas Chapter 2: Sarah Spriet February 8, 2015 Question 1. Kupffer cells, microglial cells and osteoclasts are all examples of what type of immune system cell? a. Dendritic cells b. Macrophages c. Monocytes
Abbas Chapter 2: Sarah Spriet February 8, 2015 Question 1. Kupffer cells, microglial cells and osteoclasts are all examples of what type of immune system cell? a. Dendritic cells b. Macrophages c. Monocytes
Overview of the immune system
 Overview of the immune system Immune system Innate (nonspecific) 1 st line of defense Adaptive (specific) 2 nd line of defense Cellular components Humoral components Cellular components Humoral components
Overview of the immune system Immune system Innate (nonspecific) 1 st line of defense Adaptive (specific) 2 nd line of defense Cellular components Humoral components Cellular components Humoral components
Alberta Health and Wellness Public Health Notifiable Disease Management Guidelines August Pneumococcal Disease, Invasive (IPD)
") August 2011 Pneumococcal Disease, Invasive (IPD) Revision Dates Case Definition Reporting Requirements Remainder of the Guideline (i.e., Etiology to References sections inclusive) Case Definition August
August 2011 Pneumococcal Disease, Invasive (IPD) Revision Dates Case Definition Reporting Requirements Remainder of the Guideline (i.e., Etiology to References sections inclusive) Case Definition August
Introduction to Immune System
 Introduction to Immune System Learning outcome You will be able to understand, at a fundamental level, the STRUCTURES and FUNCTIONS of cell surface and soluble molecules involved in recognition of foreign
Introduction to Immune System Learning outcome You will be able to understand, at a fundamental level, the STRUCTURES and FUNCTIONS of cell surface and soluble molecules involved in recognition of foreign
Immunology and the middle ear Andrew Riordan
 Immunology and the middle ear Andrew Riordan The Immune system is NOT there; To baffle medical students To keep Immunologists in a job To encourage experiments on mice The Immune system IS there as a defence
Immunology and the middle ear Andrew Riordan The Immune system is NOT there; To baffle medical students To keep Immunologists in a job To encourage experiments on mice The Immune system IS there as a defence
The Immune System: Innate and Adaptive Body Defenses Outline PART 1: INNATE DEFENSES 21.1 Surface barriers act as the first line of defense to keep
 The Immune System: Innate and Adaptive Body Defenses Outline PART 1: INNATE DEFENSES 21.1 Surface barriers act as the first line of defense to keep invaders out of the body (pp. 772 773; Fig. 21.1; Table
The Immune System: Innate and Adaptive Body Defenses Outline PART 1: INNATE DEFENSES 21.1 Surface barriers act as the first line of defense to keep invaders out of the body (pp. 772 773; Fig. 21.1; Table
How the Innate Immune System Profiles Pathogens
 How the Innate Immune System Profiles Pathogens Receptors on macrophages, neutrophils, dendritic cells for bacteria and viruses Broad specificity - Two main groups of bacteria: gram positive, gram-negative
How the Innate Immune System Profiles Pathogens Receptors on macrophages, neutrophils, dendritic cells for bacteria and viruses Broad specificity - Two main groups of bacteria: gram positive, gram-negative
Innate Immunity. Bởi: OpenStaxCollege
 Innate Immunity Bởi: OpenStaxCollege The vertebrate, including human, immune system is a complex multilayered system for defending against external and internal threats to the integrity of the body. The
Innate Immunity Bởi: OpenStaxCollege The vertebrate, including human, immune system is a complex multilayered system for defending against external and internal threats to the integrity of the body. The
Bacterial Diseases IMMUNITY TO BACTERIAL INFECTIONS. Gram Positive Bacteria. Gram Negative Bacteria. Many Infectious agents and many diseases
 IMMUNITY TO BACTERIAL INFECTIONS Chapter 18 Bacterial Diseases Many Infectious agents and many diseases Bacteria can Infect any part of the body Cause disease due to Growth of the microbe in a tissue Produce
IMMUNITY TO BACTERIAL INFECTIONS Chapter 18 Bacterial Diseases Many Infectious agents and many diseases Bacteria can Infect any part of the body Cause disease due to Growth of the microbe in a tissue Produce
4b. Innate (nonspecific) Immunity
 Immunity") 4b. Innate (nonspecific) Immunity Chapter 16: Innate (nonspecific) Immunity! Some terms:! Susceptibility: Lack of immunity to a disease.! Immunity: Ability to ward off disease.! Innate immunity: Defenses
4b. Innate (nonspecific) Immunity Chapter 16: Innate (nonspecific) Immunity! Some terms:! Susceptibility: Lack of immunity to a disease.! Immunity: Ability to ward off disease.! Innate immunity: Defenses
Chapter 23 Immunity Exam Study Questions
 Chapter 23 Immunity Exam Study Questions 1. Define 1) Immunity 2) Neutrophils 3) Macrophage 4) Epitopes 5) Interferon 6) Complement system 7) Histamine 8) Mast cells 9) Antigen 10) Antigens receptors 11)
Chapter 23 Immunity Exam Study Questions 1. Define 1) Immunity 2) Neutrophils 3) Macrophage 4) Epitopes 5) Interferon 6) Complement system 7) Histamine 8) Mast cells 9) Antigen 10) Antigens receptors 11)
MONTGOMERY COUNTY COMMUNITY COLLEGE Department of Science LECTURE OUTLINE CHAPTERS 16, 17, 18 AND 19
 MONTGOMERY COUNTY COMMUNITY COLLEGE Department of Science LECTURE OUTLINE CHAPTERS 16, 17, 18 AND 19 CHAPTER 16: NONSPECIFIC DEFENSES OF THE HOST I. THE FIRST LINE OF DEFENSE A. Mechanical Barriers (Physical
MONTGOMERY COUNTY COMMUNITY COLLEGE Department of Science LECTURE OUTLINE CHAPTERS 16, 17, 18 AND 19 CHAPTER 16: NONSPECIFIC DEFENSES OF THE HOST I. THE FIRST LINE OF DEFENSE A. Mechanical Barriers (Physical
The Innate Immune Response
 The Innate Immune Response FUNCTIONS OF THE IMMUNE SYSTEM: Recognize, destroy and clear a diversity of pathogens. Initiate tissue and wound healing processes. Recognize and clear damaged self components.
The Innate Immune Response FUNCTIONS OF THE IMMUNE SYSTEM: Recognize, destroy and clear a diversity of pathogens. Initiate tissue and wound healing processes. Recognize and clear damaged self components.
The Adaptive Immune Response. B-cells
 The Adaptive Immune Response B-cells The innate immune system provides immediate protection. The adaptive response takes time to develop and is antigen specific. Activation of B and T lymphocytes Naive
The Adaptive Immune Response B-cells The innate immune system provides immediate protection. The adaptive response takes time to develop and is antigen specific. Activation of B and T lymphocytes Naive
CELL BIOLOGY - CLUTCH CH THE IMMUNE SYSTEM.
 !! www.clutchprep.com CONCEPT: OVERVIEW OF HOST DEFENSES The human body contains three lines of against infectious agents (pathogens) 1. Mechanical and chemical boundaries (part of the innate immune system)
!! www.clutchprep.com CONCEPT: OVERVIEW OF HOST DEFENSES The human body contains three lines of against infectious agents (pathogens) 1. Mechanical and chemical boundaries (part of the innate immune system)
Innate Immunity: Nonspecific Defenses of the Host
 PowerPoint Lecture Presentations prepared by Bradley W. Christian, McLennan Community College C H A P T E R 16 Innate Immunity: Nonspecific Defenses of the Host Host Response to Disease Resistance- ability
PowerPoint Lecture Presentations prepared by Bradley W. Christian, McLennan Community College C H A P T E R 16 Innate Immunity: Nonspecific Defenses of the Host Host Response to Disease Resistance- ability
Physiology Unit 3. ADAPTIVE IMMUNITY The Specific Immune Response
 Physiology Unit 3 ADAPTIVE IMMUNITY The Specific Immune Response In Physiology Today The Adaptive Arm of the Immune System Specific Immune Response Internal defense against a specific pathogen Acquired
Physiology Unit 3 ADAPTIVE IMMUNITY The Specific Immune Response In Physiology Today The Adaptive Arm of the Immune System Specific Immune Response Internal defense against a specific pathogen Acquired
PATHOGENICITY OF MICROORGANISMS
 PATHOGENICITY OF MICROORGANISMS Some microorganisms are : 1- Harmless microorganism, as normal flora 2- Harmfull microorganism, as pathogenic. A pathogenic microorganism is defined as one that causes or
PATHOGENICITY OF MICROORGANISMS Some microorganisms are : 1- Harmless microorganism, as normal flora 2- Harmfull microorganism, as pathogenic. A pathogenic microorganism is defined as one that causes or
Helminth worm, Schistosomiasis Trypanosomes, sleeping sickness Pneumocystis carinii. Ringworm fungus HIV Influenza
 Helminth worm, Schistosomiasis Trypanosomes, sleeping sickness Pneumocystis carinii Ringworm fungus HIV Influenza Candida Staph aureus Mycobacterium tuberculosis Listeria Salmonella Streptococcus Levels
Helminth worm, Schistosomiasis Trypanosomes, sleeping sickness Pneumocystis carinii Ringworm fungus HIV Influenza Candida Staph aureus Mycobacterium tuberculosis Listeria Salmonella Streptococcus Levels
VMC-221: Veterinary Immunology and Serology (1+1) Question Bank
 Question Bank") VMC-221: Veterinary Immunology and Serology (1+1) Objective type Questions Question Bank Q. No. 1 - Fill up the blanks with correct words 1. The British physician, who developed the first vaccine against
VMC-221: Veterinary Immunology and Serology (1+1) Objective type Questions Question Bank Q. No. 1 - Fill up the blanks with correct words 1. The British physician, who developed the first vaccine against
The Immune System. These are classified as the Innate and Adaptive Immune Responses. Innate Immunity
 The Immune System Biological mechanisms that defend an organism must be 1. triggered by a stimulus upon injury or pathogen attack 2. able to counteract the injury or invasion 3. able to recognise foreign
The Immune System Biological mechanisms that defend an organism must be 1. triggered by a stimulus upon injury or pathogen attack 2. able to counteract the injury or invasion 3. able to recognise foreign
Innate immunity. Abul K. Abbas University of California San Francisco. FOCiS
 1 Innate immunity Abul K. Abbas University of California San Francisco FOCiS 2 Lecture outline Components of innate immunity Recognition of microbes and dead cells Toll Like Receptors NOD Like Receptors/Inflammasome
1 Innate immunity Abul K. Abbas University of California San Francisco FOCiS 2 Lecture outline Components of innate immunity Recognition of microbes and dead cells Toll Like Receptors NOD Like Receptors/Inflammasome
Campbell's Biology: Concepts and Connections, 7e (Reece et al.) Chapter 24 The Immune System Multiple-Choice Questions
 Chapter 24 The Immune System Multiple-Choice Questions") Campbell's Biology: Concepts and Connections, 7e (Reece et al.) Chapter 24 The Immune System 24.1 Multiple-Choice Questions 1) The body's innate defenses against infection include A) several nonspecific
Campbell's Biology: Concepts and Connections, 7e (Reece et al.) Chapter 24 The Immune System 24.1 Multiple-Choice Questions 1) The body's innate defenses against infection include A) several nonspecific
Vaccines for Primary Care Pneumococcal, Shingles, Pertussis
 Vaccines for Primary Care Pneumococcal, Shingles, Pertussis Devang Patel, M.D. Assistant Professor Chief of Service, MICU ID Service University of Maryland School of Medicine Pneumococcal Vaccine Pneumococcal
Vaccines for Primary Care Pneumococcal, Shingles, Pertussis Devang Patel, M.D. Assistant Professor Chief of Service, MICU ID Service University of Maryland School of Medicine Pneumococcal Vaccine Pneumococcal
Blood and Immune system Acquired Immunity
 Blood and Immune system Acquired Immunity Immunity Acquired (Adaptive) Immunity Defensive mechanisms include : 1) Innate immunity (Natural or Non specific) 2) Acquired immunity (Adaptive or Specific) Cell-mediated
Blood and Immune system Acquired Immunity Immunity Acquired (Adaptive) Immunity Defensive mechanisms include : 1) Innate immunity (Natural or Non specific) 2) Acquired immunity (Adaptive or Specific) Cell-mediated
Third line of Defense
 Chapter 15 Specific Immunity and Immunization Topics -3 rd of Defense - B cells - T cells - Specific Immunities Third line of Defense Specific immunity is a complex interaction of immune cells (leukocytes)
Chapter 15 Specific Immunity and Immunization Topics -3 rd of Defense - B cells - T cells - Specific Immunities Third line of Defense Specific immunity is a complex interaction of immune cells (leukocytes)
PNEUMONIA : PROMISE FULFILLED? Regina Berba MD FPSMID
 PNEUMONIA : PROMISE FULFILLED? Regina Berba MD FPSMID Objectives of Lecture Know the quality of current evidence based guidelines on immunization Appreciate the performance of pneumonia vaccines in terns
PNEUMONIA : PROMISE FULFILLED? Regina Berba MD FPSMID Objectives of Lecture Know the quality of current evidence based guidelines on immunization Appreciate the performance of pneumonia vaccines in terns
2. Innate immunity 2013
 1 Innate Immune Responses 3 Innate immunity Abul K. Abbas University of California San Francisco The initial responses to: 1. Microbes: essential early mechanisms to prevent, control, or eliminate infection;
1 Innate Immune Responses 3 Innate immunity Abul K. Abbas University of California San Francisco The initial responses to: 1. Microbes: essential early mechanisms to prevent, control, or eliminate infection;
History. Chapter 13. Complement Components. Complement Pathways
 History Chapter 13 Complement Jules Border in 1890 s discovered complement Paul Ehrlich coined the term complement The activity of blood serum that completes the action of antibody Now: Set of serum proteins
History Chapter 13 Complement Jules Border in 1890 s discovered complement Paul Ehrlich coined the term complement The activity of blood serum that completes the action of antibody Now: Set of serum proteins
There are 2 major lines of defense: Non-specific (Innate Immunity) and. Specific. (Adaptive Immunity) Photo of macrophage cell
 and. Specific. (Adaptive Immunity) Photo of macrophage cell") There are 2 major lines of defense: Non-specific (Innate Immunity) and Specific (Adaptive Immunity) Photo of macrophage cell Development of the Immune System ery pl neu mφ nk CD8 + CTL CD4 + thy TH1 mye
There are 2 major lines of defense: Non-specific (Innate Immunity) and Specific (Adaptive Immunity) Photo of macrophage cell Development of the Immune System ery pl neu mφ nk CD8 + CTL CD4 + thy TH1 mye
41 Pneumococcal Disease
 41 Pneumococcal Disease Epidemiology and Prevention Abstract: Streptococcus pneumonie is a major cause of morbidity and mortality in very young, high-risk adults and elderly population. It kills almost
41 Pneumococcal Disease Epidemiology and Prevention Abstract: Streptococcus pneumonie is a major cause of morbidity and mortality in very young, high-risk adults and elderly population. It kills almost
I. Lines of Defense Pathogen: Table 1: Types of Immune Mechanisms. Table 2: Innate Immunity: First Lines of Defense
 I. Lines of Defense Pathogen: Table 1: Types of Immune Mechanisms Table 2: Innate Immunity: First Lines of Defense Innate Immunity involves nonspecific physical & chemical barriers that are adapted for
I. Lines of Defense Pathogen: Table 1: Types of Immune Mechanisms Table 2: Innate Immunity: First Lines of Defense Innate Immunity involves nonspecific physical & chemical barriers that are adapted for
KEY CONCEPT Germs cause many diseases in humans.
 31.1 40.1 Pathogens Infectious Diseases and Human Illness KEY CONCEPT Germs cause many diseases in humans. 31.1 40.1 Pathogens Infectious Diseases and Human Illness Germ theory states that microorganisms
31.1 40.1 Pathogens Infectious Diseases and Human Illness KEY CONCEPT Germs cause many diseases in humans. 31.1 40.1 Pathogens Infectious Diseases and Human Illness Germ theory states that microorganisms
Complement. Definition : series of heat-labile serum proteins. : serum and all tissue fluids except urine and CSF
 Complement Complement Definition : series of heat-labile serum proteins Site : serum and all tissue fluids except urine and CSF Synthesis : in liver appear in fetal circulation during 1 st 13 W Function
Complement Complement Definition : series of heat-labile serum proteins Site : serum and all tissue fluids except urine and CSF Synthesis : in liver appear in fetal circulation during 1 st 13 W Function
CHAPTER 18: Immune System
 CHAPTER 18: Immune System 1. What are four characteristics of the specific immune system? a. b. c. d. 2. List the two main types of defense mechanisms and briefly describe features of each. 3. Give examples
CHAPTER 18: Immune System 1. What are four characteristics of the specific immune system? a. b. c. d. 2. List the two main types of defense mechanisms and briefly describe features of each. 3. Give examples
PBS Class #2 Introduction to the Immune System part II Suggested reading: Abbas, pgs , 27-30
 PBS 803 - Class #2 Introduction to the Immune System part II Suggested reading: Abbas, pgs. 15-25, 27-30 Learning Objectives Compare and contrast the maturation of B and T lymphocytes Compare and contrast
PBS 803 - Class #2 Introduction to the Immune System part II Suggested reading: Abbas, pgs. 15-25, 27-30 Learning Objectives Compare and contrast the maturation of B and T lymphocytes Compare and contrast
Chapter 1. Chapter 1 Concepts. MCMP422 Immunology and Biologics Immunology is important personally and professionally!
 MCMP422 Immunology and Biologics Immunology is important personally and professionally! Learn the language - use the glossary and index RNR - Reading, Note taking, Reviewing All materials in Chapters 1-3
MCMP422 Immunology and Biologics Immunology is important personally and professionally! Learn the language - use the glossary and index RNR - Reading, Note taking, Reviewing All materials in Chapters 1-3
Streptococcus Pneumoniae
 Streptococcus Pneumoniae (Invasive Pneumococcal Disease) DISEASE REPORTABLE WITHIN 24 HOURS OF DIAGNOSIS Per N.J.A.C. 8:57, healthcare providers and administrators shall report by mail or by electronic
Streptococcus Pneumoniae (Invasive Pneumococcal Disease) DISEASE REPORTABLE WITHIN 24 HOURS OF DIAGNOSIS Per N.J.A.C. 8:57, healthcare providers and administrators shall report by mail or by electronic
Streptococcus pneumoniae: Virulence Factors, Pathogenesis, and Vaccines
 MICROBIOLOGICAL REVIEWS, Dec. 1995, p. 591 603 Vol. 59, No. 4 0146-0749/95/$04.00 0 Copyright 1995, American Society for Microbiology Streptococcus pneumoniae: Virulence Factors, Pathogenesis, and Vaccines
MICROBIOLOGICAL REVIEWS, Dec. 1995, p. 591 603 Vol. 59, No. 4 0146-0749/95/$04.00 0 Copyright 1995, American Society for Microbiology Streptococcus pneumoniae: Virulence Factors, Pathogenesis, and Vaccines
Immunology Lecture 4. Clinical Relevance of the Immune System
 Immunology Lecture 4 The Well Patient: How innate and adaptive immune responses maintain health - 13, pg 169-181, 191-195. Immune Deficiency - 15 Autoimmunity - 16 Transplantation - 17, pg 260-270 Tumor
Immunology Lecture 4 The Well Patient: How innate and adaptive immune responses maintain health - 13, pg 169-181, 191-195. Immune Deficiency - 15 Autoimmunity - 16 Transplantation - 17, pg 260-270 Tumor
General Overview of Immunology. Kimberly S. Schluns, Ph.D. Associate Professor Department of Immunology UT MD Anderson Cancer Center
 General Overview of Immunology Kimberly S. Schluns, Ph.D. Associate Professor Department of Immunology UT MD Anderson Cancer Center Objectives Describe differences between innate and adaptive immune responses
General Overview of Immunology Kimberly S. Schluns, Ph.D. Associate Professor Department of Immunology UT MD Anderson Cancer Center Objectives Describe differences between innate and adaptive immune responses
Response to pneumococcal-polysaccharide vaccine PPV23 in HIV-positive individuals
 The University of Toledo The University of Toledo Digital Repository Theses and Dissertations 2015 Response to pneumococcal-polysaccharide vaccine PPV23 in HIV-positive individuals Anita Sridhar Iyer University
The University of Toledo The University of Toledo Digital Repository Theses and Dissertations 2015 Response to pneumococcal-polysaccharide vaccine PPV23 in HIV-positive individuals Anita Sridhar Iyer University
All animals have innate immunity, a defense active immediately upon infection Vertebrates also have adaptive immunity
 1 2 3 4 5 6 7 8 9 The Immune System All animals have innate immunity, a defense active immediately upon infection Vertebrates also have adaptive immunity Figure 43.2 In innate immunity, recognition and
1 2 3 4 5 6 7 8 9 The Immune System All animals have innate immunity, a defense active immediately upon infection Vertebrates also have adaptive immunity Figure 43.2 In innate immunity, recognition and
1. Overview of Adaptive Immunity
 Chapter 17A: Adaptive Immunity Part I 1. Overview of Adaptive Immunity 2. T and B Cell Production 3. Antigens & Antigen Presentation 4. Helper T cells 1. Overview of Adaptive Immunity The Nature of Adaptive
Chapter 17A: Adaptive Immunity Part I 1. Overview of Adaptive Immunity 2. T and B Cell Production 3. Antigens & Antigen Presentation 4. Helper T cells 1. Overview of Adaptive Immunity The Nature of Adaptive
3/10/14. Ultrastructural organization. Gram Stain. Infection leads to production of inducers of inflammation. Gram negative.
 Infection leads to production of inducers of inflammation or dendritic cell Inflammatory mediators: Complex and many, but include: Lipids and Proteins (cytokines/chemokines) TNF Others Ultrastructural
Infection leads to production of inducers of inflammation or dendritic cell Inflammatory mediators: Complex and many, but include: Lipids and Proteins (cytokines/chemokines) TNF Others Ultrastructural
Pneumococcal vaccination in UK: an update. Dr Richard Pebody Immunisation Department Health Protection Agency Centre for Infections
 Pneumococcal vaccination in UK: an update Dr Richard Pebody Immunisation Department Health Protection Agency Centre for Infections Leading infectious causes of mortality, 2000 WHO estimates 3.5 Deaths
Pneumococcal vaccination in UK: an update Dr Richard Pebody Immunisation Department Health Protection Agency Centre for Infections Leading infectious causes of mortality, 2000 WHO estimates 3.5 Deaths
Diseases-causing agents, pathogens, can produce infections within the body.
 BIO 212: ANATOMY & PHYSIOLOGY II 1 CHAPTER 16 Lecture: Dr. Lawrence G. Altman www.lawrencegaltman.com Some illustrations are courtesy of McGraw-Hill. LYMPHATIC and IMMUNE Systems Body Defenses Against
BIO 212: ANATOMY & PHYSIOLOGY II 1 CHAPTER 16 Lecture: Dr. Lawrence G. Altman www.lawrencegaltman.com Some illustrations are courtesy of McGraw-Hill. LYMPHATIC and IMMUNE Systems Body Defenses Against
INNATE IMMUNITY Non-Specific Immune Response. Physiology Unit 3
 INNATE IMMUNITY Non-Specific Immune Response Physiology Unit 3 Protection Against Infection The body has several defenses to protect itself from getting an infection Skin Mucus membranes Serous membranes
INNATE IMMUNITY Non-Specific Immune Response Physiology Unit 3 Protection Against Infection The body has several defenses to protect itself from getting an infection Skin Mucus membranes Serous membranes
Pathogenesis of Infectious Diseases. CLS 212: Medical Microbiology
 Pathogenesis of Infectious Diseases CLS 212: Medical Microbiology Definitions Path- means disease. Pathogenesis The steps or mechanisms involved in the development of a disease. Infection The presence
Pathogenesis of Infectious Diseases CLS 212: Medical Microbiology Definitions Path- means disease. Pathogenesis The steps or mechanisms involved in the development of a disease. Infection The presence
Macrophage Activation & Cytokine Release. Dendritic Cells & Antigen Presentation. Neutrophils & Innate Defense
 Macrophage Activation & Cytokine Release Dendritic Cells & Antigen Presentation Neutrophils & Innate Defense Neutrophils Polymorphonuclear cells (PMNs) are recruited to the site of infection where they
Macrophage Activation & Cytokine Release Dendritic Cells & Antigen Presentation Neutrophils & Innate Defense Neutrophils Polymorphonuclear cells (PMNs) are recruited to the site of infection where they
3. Lymphocyte proliferation (fig. 15.4): Clones of responder cells and memory cells are derived from B cells and T cells.
: Clones of responder cells and memory cells are derived from B cells and T cells.") Chapter 15 Adaptive, Specific Immunity and Immunization* *Lecture notes are to be used as a study guide only and do not represent the comprehensive information you will need to know for the exams. Specific
Chapter 15 Adaptive, Specific Immunity and Immunization* *Lecture notes are to be used as a study guide only and do not represent the comprehensive information you will need to know for the exams. Specific
Immunity and Infection. Chapter 17
 Immunity and Infection Chapter 17 The Chain of Infection Transmitted through a chain of infection (six links) Pathogen: Disease causing microorganism Reservoir: Natural environment of the pathogen Portal
Immunity and Infection Chapter 17 The Chain of Infection Transmitted through a chain of infection (six links) Pathogen: Disease causing microorganism Reservoir: Natural environment of the pathogen Portal
Disease causing organisms Resistance Immunity
 Part 1 Disease causing organisms Resistance Immunity Bacteria Most common pathogens Anthrax Cholera Staphylococcus epidermidis bacteria Bacterial diseases Tuberculosis Cholera Bubonic Plague Tetanus Effects
Part 1 Disease causing organisms Resistance Immunity Bacteria Most common pathogens Anthrax Cholera Staphylococcus epidermidis bacteria Bacterial diseases Tuberculosis Cholera Bubonic Plague Tetanus Effects
Problem 7 Unit 6 Clinical: Primary immunodeficiency
 Problem 7 Unit 6 Clinical: Primary immunodeficiency THE IMMUNE SYSTEM - Function: recognizing pathogens (foreign non-self antigens) and organizing a defense response against them by facilitating destruction
Problem 7 Unit 6 Clinical: Primary immunodeficiency THE IMMUNE SYSTEM - Function: recognizing pathogens (foreign non-self antigens) and organizing a defense response against them by facilitating destruction
Unit 9 New life College faculty: Ramesh Kumar Subject: Life Sciences date: 15jan 2016
 Unit 9 New life College faculty: Ramesh Kumar Subject: Life Sciences date: 15jan 2016 History and impact The types and functions of cells in the immune system. How cells communicate and recognize antigen
Unit 9 New life College faculty: Ramesh Kumar Subject: Life Sciences date: 15jan 2016 History and impact The types and functions of cells in the immune system. How cells communicate and recognize antigen
Nonspecific External Barriers skin, mucous membranes
 Immune system Chapter 36 BI 103 Plant-Animal A&P Levels of Defense Against Disease Nonspecific External Barriers skin, mucous membranes Physical barriers? Brainstorm with a partner If these barriers are
Immune system Chapter 36 BI 103 Plant-Animal A&P Levels of Defense Against Disease Nonspecific External Barriers skin, mucous membranes Physical barriers? Brainstorm with a partner If these barriers are
Chapter 38- Immune System
 Chapter 38- Immune System First Line of Defense: Barriers Nonspecific defenses, such as the skin and mucous membranes, are barriers to potential pathogens. In addition to being a physical barrier to pathogens,
Chapter 38- Immune System First Line of Defense: Barriers Nonspecific defenses, such as the skin and mucous membranes, are barriers to potential pathogens. In addition to being a physical barrier to pathogens,
ACTIVATION AND EFFECTOR FUNCTIONS OF CELL-MEDIATED IMMUNITY AND NK CELLS. Choompone Sakonwasun, MD (Hons), FRCPT
, FRCPT") ACTIVATION AND EFFECTOR FUNCTIONS OF CELL-MEDIATED IMMUNITY AND NK CELLS Choompone Sakonwasun, MD (Hons), FRCPT Types of Adaptive Immunity Types of T Cell-mediated Immune Reactions CTLs = cytotoxic T lymphocytes
ACTIVATION AND EFFECTOR FUNCTIONS OF CELL-MEDIATED IMMUNITY AND NK CELLS Choompone Sakonwasun, MD (Hons), FRCPT Types of Adaptive Immunity Types of T Cell-mediated Immune Reactions CTLs = cytotoxic T lymphocytes
PLAN. Réponses B thymodépendantes et thymoindépendantes. B cell development and classification. B cell activation. Thymodependent B(2) cell response
 cell response") Réponses B thymodépendantes et thymoindépendantes PLAN B cell development and classification B cell activation Thymodependent B(2) cell response BMC 423 (IF) - 2007 Antonino Nicoletti Thymo-independent
Réponses B thymodépendantes et thymoindépendantes PLAN B cell development and classification B cell activation Thymodependent B(2) cell response BMC 423 (IF) - 2007 Antonino Nicoletti Thymo-independent
Complement. History. Chapter 7. Complement Components. Complement Pathways. Pathways of complement activation
 History Chapter 7 Complement Jules Border in 1890 s discovered complement Paul Ehrlich coined the term complement The activity of blood serum that completes the action of antibody Now: Set of serum proteins
History Chapter 7 Complement Jules Border in 1890 s discovered complement Paul Ehrlich coined the term complement The activity of blood serum that completes the action of antibody Now: Set of serum proteins
The Adaptive Immune Responses
 The Adaptive Immune Responses The two arms of the immune responses are; 1) the cell mediated, and 2) the humoral responses. In this chapter we will discuss the two responses in detail and we will start
The Adaptive Immune Responses The two arms of the immune responses are; 1) the cell mediated, and 2) the humoral responses. In this chapter we will discuss the two responses in detail and we will start
Immune system. Aims. Immune system. Lymphatic organs. Inflammation. Natural immune system. Adaptive immune system
 Aims Immune system Lymphatic organs Inflammation Natural immune system Adaptive immune system Major histocompatibility complex (MHC) Disorders of the immune system 1 2 Immune system Lymphoid organs Immune
Aims Immune system Lymphatic organs Inflammation Natural immune system Adaptive immune system Major histocompatibility complex (MHC) Disorders of the immune system 1 2 Immune system Lymphoid organs Immune
IgG subclass deficiencies
 IgG subclass deficiencies hello@piduk.org 0800 987 8986 www.piduk.org About this booklet This booklet provides information on IgG subclass deficiencies. It has been produced by the PID UK Medical Advisory
IgG subclass deficiencies hello@piduk.org 0800 987 8986 www.piduk.org About this booklet This booklet provides information on IgG subclass deficiencies. It has been produced by the PID UK Medical Advisory
Incidence per 100,000
 Streptococcus pneumoniae Surveillance Report 2005 Oregon Active Bacterial Core Surveillance (ABCs) Office of Disease Prevention & Epidemiology Oregon Department of Human Services Updated: March 2007 Background
Streptococcus pneumoniae Surveillance Report 2005 Oregon Active Bacterial Core Surveillance (ABCs) Office of Disease Prevention & Epidemiology Oregon Department of Human Services Updated: March 2007 Background
Surveillance of invasive pneumococcal infection in Belgium
 Surveillance of invasive pneumococcal infection in Belgium National Reference Laboratory Start in 198 Laboratory Microbiology UH Leuven (prof. J. Vandepitte) Capsular type determination Antibiotic susceptibility
Surveillance of invasive pneumococcal infection in Belgium National Reference Laboratory Start in 198 Laboratory Microbiology UH Leuven (prof. J. Vandepitte) Capsular type determination Antibiotic susceptibility
Expanded Use of PCV13 & PPV23
 Expanded Use of PCV13 & PPV23 Dr. Jim Kellner Professor & Head Department of Pediatrics University of Calgary & Calgary Zone, Alberta Health Services Jim.Kellner@ahs.ca Objectives Explain the differences
Expanded Use of PCV13 & PPV23 Dr. Jim Kellner Professor & Head Department of Pediatrics University of Calgary & Calgary Zone, Alberta Health Services Jim.Kellner@ahs.ca Objectives Explain the differences
Innate vs Adaptive Response
 General Immunology Innate vs Adaptive Response Innate- non-specific (4 types of barriers) anatomic- ato mechanical ca (skin), ph, mucous, normal flora Physiologic- temperature, ph, chemicals (lysozyme,
General Immunology Innate vs Adaptive Response Innate- non-specific (4 types of barriers) anatomic- ato mechanical ca (skin), ph, mucous, normal flora Physiologic- temperature, ph, chemicals (lysozyme,
2 االستاذ المساعد الدكتور خالد ياسين الزاملي \ مناعة \ المرحلة الثانية \ التحليالت المرضية \
 Innate Immunity Innate immunity: is the resistance that an individual possesses by birth. Innate immunity may be classified as (a) individual immunity (b) racial immunity (c) species immunity. Factors
Innate Immunity Innate immunity: is the resistance that an individual possesses by birth. Innate immunity may be classified as (a) individual immunity (b) racial immunity (c) species immunity. Factors
Chapter 16 Pneumococcal Infection. Pneumococcal Infection. August 2015
 Chapter 16 16 PPV introduced for at risk 1996 PCV7 introduced for at risk 2002 and as routine 2008 PCV13 replaced PCV7 in 2010 NOTIFIABLE In some circumstances, advice in these guidelines may differ from
Chapter 16 16 PPV introduced for at risk 1996 PCV7 introduced for at risk 2002 and as routine 2008 PCV13 replaced PCV7 in 2010 NOTIFIABLE In some circumstances, advice in these guidelines may differ from
Chapter 13 Lymphatic and Immune Systems
 The Chapter 13 Lymphatic and Immune Systems 1 The Lymphatic Vessels Lymphoid Organs Three functions contribute to homeostasis 1. Return excess tissue fluid to the bloodstream 2. Help defend the body against
The Chapter 13 Lymphatic and Immune Systems 1 The Lymphatic Vessels Lymphoid Organs Three functions contribute to homeostasis 1. Return excess tissue fluid to the bloodstream 2. Help defend the body against
Third line of Defense. Topic 8 Specific Immunity (adaptive) (18) 3 rd Line = Prophylaxis via Immunization!
 (18) 3 rd Line = Prophylaxis via Immunization!") Topic 8 Specific Immunity (adaptive) (18) Topics - 3 rd Line of Defense - B cells - T cells - Specific Immunities 1 3 rd Line = Prophylaxis via Immunization! (a) A painting of Edward Jenner depicts a cow
Topic 8 Specific Immunity (adaptive) (18) Topics - 3 rd Line of Defense - B cells - T cells - Specific Immunities 1 3 rd Line = Prophylaxis via Immunization! (a) A painting of Edward Jenner depicts a cow
The Immune System: The Mind Body Connection. Presented by Margaret Kemeny, Ph.D. Department of Psychiatry, University of California, San Francisco
 The Immune System: The Mind Body Connection Presented by Margaret Kemeny, Ph.D. Department of Psychiatry, University of California, San Francisco Psychoneuroimmunology Investigation of the bidirectional
The Immune System: The Mind Body Connection Presented by Margaret Kemeny, Ph.D. Department of Psychiatry, University of California, San Francisco Psychoneuroimmunology Investigation of the bidirectional
Immunity. Skin. Mucin-containing mucous membranes. Desmosome (attaches keratincontaining. Fig. 43.2
 Immunity 1 Fig. 43.2 2 Skin Mucin-containing mucous membranes Desmosome (attaches keratincontaining skin cells together) 1 http://www.mhhe.com/biosci/ap/histology_mh/pseudos2l.jpg http://training.seer.cancer.gov/module_anatomy/images/
Immunity 1 Fig. 43.2 2 Skin Mucin-containing mucous membranes Desmosome (attaches keratincontaining skin cells together) 1 http://www.mhhe.com/biosci/ap/histology_mh/pseudos2l.jpg http://training.seer.cancer.gov/module_anatomy/images/
Streptococcus pyogenes
 Streptococcus pyogenes From Wikipedia, the free encyclopedia Streptococcus pyogenes S. pyogenes bacteria at 900x magnification. Scientific classification Kingdom: Eubacteria Phylum: Firmicutes Class: Cocci
Streptococcus pyogenes From Wikipedia, the free encyclopedia Streptococcus pyogenes S. pyogenes bacteria at 900x magnification. Scientific classification Kingdom: Eubacteria Phylum: Firmicutes Class: Cocci
Immunity and how vaccines work
 Immunity and how vaccines work Dr Mary O Meara National Immunisation Office Objectives of session An understanding of the following principles Overview of immunity Different types of vaccines and vaccine
Immunity and how vaccines work Dr Mary O Meara National Immunisation Office Objectives of session An understanding of the following principles Overview of immunity Different types of vaccines and vaccine
OpenStax-CNX module: m Innate Immunity. OpenStax College. Abstract
 OpenStax-CNX module: m45542 1 Innate Immunity OpenStax College This work is produced by OpenStax-CNX and licensed under the Creative Commons Attribution License 3.0 By the end of this section, you will
OpenStax-CNX module: m45542 1 Innate Immunity OpenStax College This work is produced by OpenStax-CNX and licensed under the Creative Commons Attribution License 3.0 By the end of this section, you will
Innate Immunity. Connection Between Innate and Adaptive Immunity. Know Differences and Provide Examples Chapter 3. Antimicrobial peptide psoriasin
 Know Differences and Provide Examples Chapter * Innate Immunity * kin and Epithelial Barriers * Antimicrobial peptide psoriasin -Activity against Gram (-) E. coli Connection Between Innate and Adaptive
Know Differences and Provide Examples Chapter * Innate Immunity * kin and Epithelial Barriers * Antimicrobial peptide psoriasin -Activity against Gram (-) E. coli Connection Between Innate and Adaptive
Skin. Mucin-containing mucous membranes. Desmosome (attaches keratincontaining
 Immunity 1 Fig. 43.2 2 Skin Mucin-containing mucous membranes Desmosome (attaches keratincontaining skin cells together) http://www.mhhe.com/biosci/ap/histology_mh/pseudos2l.jpg http://training.seer.cancer.gov/module_anatomy/images/
Immunity 1 Fig. 43.2 2 Skin Mucin-containing mucous membranes Desmosome (attaches keratincontaining skin cells together) http://www.mhhe.com/biosci/ap/histology_mh/pseudos2l.jpg http://training.seer.cancer.gov/module_anatomy/images/
Adaptive Immunity. PowerPoint Lecture Presentations prepared by Mindy Miller-Kittrell, North Carolina State University C H A P T E R
 CSLO7. Describe functions of host defenses and the immune system in combating infectious diseases and explain how immunizations protect against specific diseases. PowerPoint Lecture Presentations prepared
CSLO7. Describe functions of host defenses and the immune system in combating infectious diseases and explain how immunizations protect against specific diseases. PowerPoint Lecture Presentations prepared
For questions 1-5, match the following with their correct descriptions. (24-39) A. Class I B. Class II C. Class III D. TH1 E. TH2
 A. Class I B. Class II C. Class III D. TH1 E. TH2") Questions Made by SI ATTENDEES!! :) Page 1 of 6 Student-Made Practice Exam Activity All questions, answers, and slide numbers are based off of Monday s SI activity, where students/attendees created possible
Questions Made by SI ATTENDEES!! :) Page 1 of 6 Student-Made Practice Exam Activity All questions, answers, and slide numbers are based off of Monday s SI activity, where students/attendees created possible
Dr.Mohammed Al-Araji 2015/2016 Streptococcus pneumoniae
 Dr.Mohammed Al-Araji 2015/2016 Streptococcus pneumoniae Streptococcus pneumoniae, or pneumococcus, is a Grampositive, alphahemolytic, aerotolerant, anaerobic member of the genusstreptococcus. A significant
Dr.Mohammed Al-Araji 2015/2016 Streptococcus pneumoniae Streptococcus pneumoniae, or pneumococcus, is a Grampositive, alphahemolytic, aerotolerant, anaerobic member of the genusstreptococcus. A significant
chapter 17: specific/adaptable defenses of the host: the immune response
 chapter 17: specific/adaptable defenses of the host: the immune response defense against infection & illness body defenses innate/ non-specific adaptable/ specific epithelium, fever, inflammation, complement,
chapter 17: specific/adaptable defenses of the host: the immune response defense against infection & illness body defenses innate/ non-specific adaptable/ specific epithelium, fever, inflammation, complement,
Chapter 10 (pages ): Differentiation and Functions of CD4+ Effector T Cells Prepared by Kristen Dazy, MD, Scripps Clinic Medical Group
: Differentiation and Functions of CD4+ Effector T Cells Prepared by Kristen Dazy, MD, Scripps Clinic Medical Group") FIT Board Review Corner September 2015 Welcome to the FIT Board Review Corner, prepared by Andrew Nickels, MD, and Sarah Spriet, DO, senior and junior representatives of ACAAI's Fellows-In-Training (FITs)
FIT Board Review Corner September 2015 Welcome to the FIT Board Review Corner, prepared by Andrew Nickels, MD, and Sarah Spriet, DO, senior and junior representatives of ACAAI's Fellows-In-Training (FITs)
Immunology. Anas Abu-Humaidan M.D. Ph.D. Transplant immunology+ Secondary immune deficiency
 Immunology Anas Abu-Humaidan M.D. Ph.D. Transplant immunology+ Secondary immune deficiency Transplant Immunology Transplantation is the process of moving cells, tissues or organs from one site to another
Immunology Anas Abu-Humaidan M.D. Ph.D. Transplant immunology+ Secondary immune deficiency Transplant Immunology Transplantation is the process of moving cells, tissues or organs from one site to another
Topics. Humoral Immune Response Part II Accessory cells Fc Receptors Opsonization and killing mechanisms of phagocytes NK, mast, eosynophils
 Topics Humoral Immune Response Part II Accessory cells Fc Receptors Opsonization and killing mechanisms of phagocytes NK, mast, eosynophils Immune regulation Idiotypic network 2/15/2005 MICR 415 / 515
Topics Humoral Immune Response Part II Accessory cells Fc Receptors Opsonization and killing mechanisms of phagocytes NK, mast, eosynophils Immune regulation Idiotypic network 2/15/2005 MICR 415 / 515
MACROPHAGE "MONOCYTES" SURFACE RECEPTORS
 LECTURE: 13 Title: MACROPHAGE "MONOCYTES" SURFACE RECEPTORS LEARNING OBJECTIVES: The student should be able to: Describe the blood monocytes (size, and shape of nucleus). Enumerate some of the monocytes
LECTURE: 13 Title: MACROPHAGE "MONOCYTES" SURFACE RECEPTORS LEARNING OBJECTIVES: The student should be able to: Describe the blood monocytes (size, and shape of nucleus). Enumerate some of the monocytes