neurons Gemma Triola, Gemma Fabrias, Mihaela Dragusin, Lars Niederhausen, Roland Broere,
|
|
|
- Helena Roberts
- 6 years ago
- Views:
Transcription
1 Molecular Pharmacology Fast Forward. Published on September 15, 2004 as doi: /mol MOL Specificity of the dihydroceramide desaturase inhibitor GT11 in primary cultured cerebellar neurons Gemma Triola, Gemma Fabrias, Mihaela Dragusin, Lars Niederhausen, Roland Broere, Amadeu Llebaria, and Gerhild van Echten-Deckert From the Research Unit on Bioactive Molecules (RUBAM), Dept. of Biological Organic Chemistry, IIQAB, CSIC, Jordi Girona 18, Barcelona, Spain (G.T., G.F., A.L.) and the Kekulé-Institut für Organische Chemie und Biochemie der Universität Bonn, Gerhard- Domagk-Strasse 1, Bonn, Germany (M.D., L.N., R.B., G.vE-D.) Copyright 2004 by the American Society for Pharmacology and Experimental Therapeutics.
2 MOL Running title: GT11 inhibits dihydroceramide desaturase in neurons Corresponding author Gerhild van Echten-Deckert Kekulé-Institut für Organische Chemie und Biochemie der Universität Bonn, Gerhard- Domagk-Strasse 1, Bonn, Germany. phone ; fax ; g.echten.deckert@uni-bonn.de Text pages: Figures: 9 References: 30 Abstract: 204 words Introduction: 663 words Discussion: 1242 words Non-standard abbreviations used are: GT11, N-[(1R,2S)-2-hydroxy-1-hydroxymethyl-2-(2-tridecyl-1- cyclopropenyl)ethyl]octanamide; GT17, N-[(1R,2S)-2-hydroxy-1-hydroxymethyl-2-(2- tridecyl-1-cyclopropenyl)ethyl]amine; DHS, sphinganine (dihydrosphingosine); DHS1P, sphinganine-1-phosphate; ER, endoplasmic reticulum; PE, phosphatidylethanolamine; SL(s), sphingolipid(s); S1P, sphingosine-1-phosphate; SPT, serine palmitoyltransferase; TLC, thin layer chromatography.
3 MOL Abstract Dihydroceramide desaturase catalyses the conversion of the innocuous precursor (dihydroceramide) into a highly bioactive product (ceramide). We studied the effect of GT11, the first inhibitor of this enzyme, in primary cultured cerebellar neurons. Although desaturase was efficiently inhibited (IC 50 of 23 nm) the compound lost its specificity at higher concentrations. From 5 µm upwards GT11 also decreased de novo sphingolipid biosynthesis. Studies with two differentially labeled radioactive analogs of GT11 support that the inhibitor itself and not a downstream metabolic product, interferes with sphingolipid biosynthesis. Interestingly, serine palmitoyltransferase activity decreased in the presence of high concentrations of GT11 in intact cells, but not when added directly into cell homogenates. However, suppression of enzyme transcription could not be detected. But at high concentrations GT11 provoked an accumulation of sphingosine-1-phosphate and especially of dihydrosphingosine-1-phosphate, suggesting a decreased activity of sphingosine-1-phosphate lyase. Enzyme activity measurements indeed supported this assumption. Thus, at higher concentrations, GT11 interferes with lyase activity inducing an accumulation of sphingoid base phosphates that, in turn down-regulate serine palmitoyltransferase activity. At low concentrations GT11 is, however, the first specific inhibitor of dihydroceramide desaturase described so far. Considering the pro-apoptotic and pro-inflammatory effect of ceramide, GT11 could also turn out to be a novel cell-protective agent.
4 MOL Ceramide the hydrophobic membrane anchor of sphingolipids (SLs) has been implicated as a bioactive molecule in cell growth regulation, apoptosis, senescence and diverse cell responses, particularly linked to stress situations (Hannun and Luberto, 2000; Hannun and Obeid, 2002). In addition ceramide is a key intermediate in the metabolism of all SLs. Among the latter are several bioactive molecules including sphingomyelin (Marchesini and Hannun, 2004), glucosylceramide (Bleicher and Cabot, 2002 ), and last but not least complex glycosphingolipids (Huwiler et al., 2000) with diverse functions in the control of cell fate. Interestingly, ceramide is also the only metabolic precursor for sphingosine-1-phosphate (S1P), which among a multitude of specific cellular responses is known to counteract the proapoptotic action of ceramide (Spiegel and Milstien, 2003). Moreover, several studies report on the therapeutic potential of modulating intracellular ceramide levels (Kolesnick, 2002). The de novo biosynthesis of ceramide requires four sequential steps catalyzed by membranebound enzymes active at the cytosolic face of the ER (van Echten-Deckert and Sandhoff, 1999). The first and rate-limiting step is catalyzed by serine palmitoyltransferase (SPT) and yields 3-ketosphinganine, which is rapidly reduced to D-erythro-sphinganine. N-Acylation of sphinganine leads to dihydroceramide, which is then converted to ceramide by the introduction of a 4,5-trans-double bond (Michel et al. 1997). Since dihydroceramide is usually much less effective or fails to show the bioactivity of ceramide (Bielawska et al., 1993) this step catalyzed by dihydroceramide desaturase appears to be of particular importance. Also, induction of ceramide but not of dihydroceramide formation by some chemotherapeutic agents (such as daunorubicin) has been proposed to mediate their toxicity (Hannun and Luberto, 2000). It should be noted that the 4,5 double bond of ceramide is not essential for anabolic enzymes that catalyze the formation of sphingomyelin or glycosphingolipids. Thus inhibition of the desaturation step does not end up in complete SL depletion of cells. However, 4 desaturated sphingolipids were proposed to be involved in cell
5 MOL cycle control during Drosophila spermatogenesis (Ternes et al., 2002). Taken together these findings indicate that the 4,5-trans-double bond contributes to the signaling function of ceramide and possibly of its metabolites. The enzyme catalyzing the desaturation of dihydroceramide in mammals is localized in the cytosolic leaflet of ER membranes (Michel and van Echten-Deckert, 1997) and is highly specific for D-erythro-dihydroceramide (Michel et al., 1997). Based on a bioinformatic strategy, a putative SL 4 desaturase family with members from animals, plants and fungi has been identified and characterized (Ternes et al., 2002). Although molecular and genetic tools are now available to study 4 desaturated SLs, it is still not clear whether this SL desaturase family also includes the dihydroceramide desaturase described previously (Michel and van Echten-Deckert, 1997; Michel et al, 1997). The synthesis of GT11, a cyclopropene-containing ceramide analog and its inhibitory effect on the activity of dihydroceramide desaturase in rat liver microsomes has been reported recently (Triola et al., 2001). Kinetic studies performed in rat liver microsomes using different concentrations of substrate and GT11 have evidenced that this ceramide analog is a competitive inhibitor (K i = 6 µm) of dihydroceramide desaturase (Triola et al., 2003). Structure-function studies performed with GT11 related synthetic analogs have revealed that the cyclopropene ring instead of the 4,5-trans-double bond, the natural 2S,3R (D-erythro) stereochemistry as well as the presence of a free hydroxyl group at C1 are crucial for inhibition of dihydroceramide desaturase (Triola et al., 2003). The idea that an inhibitor of dihydroceramide desaturation and hence of de novo ceramide formation might possibly be a useful cell protective agent, prompted us to investigate the effects of GT11 in primary cultured cerebellar neurons. In addition to being a validated model for studies concerning synthetic analogs that interfere with SL metabolism (van Echten-Deckert et al., 1997 and 1998), these cells could also provide data concerning a potential pharmacological utility of GT11 as a neuroprotective drug. We report here that low concentrations (up to 1 µm) of
6 MOL GT11 specifically and effectively inhibit dihydroceramide desaturase in primary cultured cerebellar neurons. However, the inhibitor looses specificity at higher concentrations.
7 MOL Materials and Methods Materials. Six-day-old NMRI (Navy Marine Research Institute) mice were bred in the animal house of the University in Bonn (Germany). GT11 (Fig. 1) was synthesized as described (Triola et al., 2001). [ 14 C]GT11 (5 Ci/mol) was obtained accordingly using [1-14 C]octanoic acid (5 Ci/mol) in the final acylation step. [1-3 H]GT11 (3.75 Ci/mmol) was prepared as illustrated in Fig. 1. N-[1-14 C]octanoyl D-erythro-sphinganine was synthesized as described before (Michel et al., 1997). L-[3-14 C]serine (54 mci/mmol) was purchased from Amersham- Buchler (Braunschweig, Germany). D-erythro-[4,5-3 H]sphinganine (250 Ci/mol) was obtained according to Schwarzmann (1978). D-erythro[4,5-3 H]DHS1P (50-60Ci/mmol) and DHS1P were both from BioTrend (Koeln, Germany). Culture media (Dulbecco s modified Eagle s medium, DMEM and minimal essential medium, MEM) containing Glutamax R were obtained from Life Technologies, Inc. (Karlsruhe, Germany). DNase was from Roche (Mannheim, Germany). Horse serum and trypsin were supplied from Cytogen (Berlin, Germany). Bovine serum albumin (fatty acid free) was from Sigma (Taufkirchen, Germany). The plastic culture dishes were from Falcon (Heidelberg, Germany). LiChroprep R RP-18 and thin-layer silica gel 60 plates were purchased from Merck (Darmstadt, Germany). DEAE- Sephadex A-25 was from Pharmacia LKB Biotechnology (Uppsala, Sweden). All other chemicals were of analytical grade and obtained from Sigma (Taufkirchen, Germany) or Merck (Darmstadt, Germany). Cell Culture. Granule cells were cultured from cerebella of 6-day-old mice as described before (van Echten-Deckert et al., 1998). Briefly, cells were isolated by mild trypsinization (0.05 %, w/v) and dissociated by repeated passage through a constricted Pasteur pipette in a DNase solution (0.1 %, w/v). The cells were then suspended in DMEM containing 10 % heatinactivated horse serum and plated onto poly-l-lysine-coated 8 cm 2 Petri dishes (6 x 10 6 cells/dish). 24 h after plating, cytosine arabinoside was added to the medium (4 x 10-5 M) to
8 MOL arrest the division of non-neuronal cells. After 5-6 days in culture, cells were used for metabolic studies. Sphingolipid labeling, extraction and analysis. Medium was removed from the culture dishes and the cells were rinsed two times with MEM. The cells were metabolically labeled in MEM containing 0.3% horse serum and 1% cytosine arabinoside by addition of 1 µci/ml of either [ 14 C]serine or [ 3 H]sphinganine. After 24 h cells were washed three times with phosphate-buffered saline, harvested and centrifuged at 3000 x g for 10 min. Total lipids were extracted from cell pellets with 6 ml of chloroform/methanol/water/pyridine (10:5:1:0.1, by volume) for 24 h at 50 o C (van Echten-Deckert, 2000). Phospholipids were degraded by mild alkaline hydrolysis with methanolic NaOH (100 mm) for 2 h at 37 o C. The lipid extracts were desalted by reversed-phase chromatography on LiChroprep RP18, applied to thin layer chromatography (TLC) plates, and developed with the indicated solvents. SLs were visualized by autoradiography using the bio-imaging analyser Fujix Bas1000, software TINA 2.09 (Raytest, Straubenhardt, Germany) and identified by their R f values. Dihydroceramide desaturase activity. Enzymatic activity of dihydroceramide desaturase was measured in cell homogenate with N-[1-14 C]octanoyl D-erythro-sphinganine as substrate, essentially as described previously (Schulze et al., 2000). In a final volume of 300 µl, the assay mixture contained phosphate buffer (100 mm, ph 7.4), substrate (12.5 µm, prepared as 1:2 complexes with fatty acid free bovine serum albumin) and 300 µg of cell protein. After a pre-incubation time of 5 min at 37 C, the reaction was started by addition of 1 µmol NADH in 30 µl phosphate buffer. After 60 min at 37 C the reaction was terminated by addition of 200 µl of CHCl 3 /CH 3 OH (83:17, by volume) on ice. Lipid extraction was achieved by addition of 343 µl CH 3 OH and 22 µl CHCl 3, and vigorously mixing for 20 min. Phases were separated by centrifugation and the lower phase was collected. The extraction procedure was repeated twice with 200 µl of CHCl 3 /CH 3 OH (83:17, by volume). The combined organic
9 MOL phases were evaporated under a stream of N 2 and dissolved in 30 µl of CHCl 3 /CH 3 OH (1:1, by volume). Lipids were separated by TLC on silica gel/sodium borate plates developed with CHCl 3 /CH 3 OH (9:1, by volume). The radioactively labeled desaturated product N-[1-14 C] octanoyl D-erythro-sphingosine was detected and quantified by phosphoimaging. SPT activity. Enzymatic activity of SPT was measured using [ 14 C]serine and palmitoyl-coa as substrates (Merrill and Wang, 1992). The assay mixture contained 0.1 M Hepes (ph 7.4), 5 mm dithiothreitol, 10 mm EDTA, 50 µm pyridoxal 5'phosphate, 1.2 mm L-[ 14 C]serine (1.6 µci), 0.15 mm palmitoyl-coa and µg of cell protein, in a total volume of 100 µl. After incubation for 10 min at 37 o C, reactions were terminated by addition of chloroform/methanol (5:3, by volume). The lipids were extracted by phase separation and applied to a TLC plate, which was developed with chloroform/methanol/2 M NH 4 OH (40:10:1; by volume). Radioactively labeled 3-ketosphinganine was detected and quantified by phosphoimaging. S1P lyase activity. Enzymatic activity was determined in cell homogenates using [4,5-3 H]sphinganine-1-phosphate as substrate, essentially as described by van Veldhoven (2000). In a final volume of 200 µl, the assay mixture contained phosphate buffer (100 mm, ph 7.4), 25 mm NaF, 0.1 % Triton X-100, 0.5 mm EDTA, 2 mm dithiothreitol, 0.25 mm pyridoxal 5'phosphate, 40 µm substrate (0.5 µci) and 100 µg cell protein. The reaction was performed for 60 min at 37 C and terminated by addition of 0.2 ml of HClO 4 (1 %), and of 1.5 ml of chloroform/methanol (1:2, by volume). Phase separation was induced by adding 0.5 ml of chloroform and 0.5 ml of HClO 4. After thorough vortexing and centrifugation, lower phase was washed twice with 1 ml of HClO 4 /chloroform (8:2, by volume). Then a known volume of the lower phase was transferred to new tubes, dried under nitrogen, and the residue dissolved in chloroform/methanol (8:2, by volume) containing 5 mm of palmitic acid and hexadecanol as carriers. Labeled lipids were resolved by TLC with hexane/diethylether/ acetic acid
10 MOL (70:30:1, by volume). Radioactively labeled hexadecanal as well as palmitic acid and hexadecanol derived thereof were quantified by phosphoimaging. Labeling of cells with 32 P i. Cells were washed with phosphate-free MEM and subsequently incubated with this medium containing 32 P i (40 µci/ml) for 24 h as described (Zhang et al., 1991). The cells were then treated with the indicated concentration of GT11 or GT17 for the indicated times. Sphingoid base phosphates were extracted following the method of Yatomi et al. (1995), as described in detail previously (van Echten-Deckert et al., 1997). Phosphorylated sphingoid bases were resolved by TLC with 1-butanol/methanol/acetic acid/water (80:20:10:20, by volume), visualized by autoradiography, identified by their R f value and quantitatively evaluated using the bio-imaging analyzer Fujix Bas 1000 using software TINA 2.09 (Raytest, Straubenhardt, Germany). Reverse-transcription polymerase chain reaction (RT-PCR). Total cellular RNA was prepared using the RNeasy Mini Kit and the RNaseFreeDNaseI-set (QIAGEN, Hilden, Germany) following provider s instructions. For each reverse-transcription reaction, 0.4 µg of total RNA was reversely transcribed using SuperScript II First-Strand Synthesis System for RT-PCR with Random Primers (Invitrogen, Karlruhe, Germany) according to manufacturer s instructions. Semi-quantitative PCR was performed using gene-specific primers: S1P lyase 5`- ATGGATCCTGTCCCCGAAGT-3`(forward), 5`-CACCTTTCACCCGGAAATCA-3` (reverse), 27 cycles; SPT-LCB1 5`-CGAGGCTCCAGCATACCATC-3` (forward), 5`- TGGCTGCCACTCTTCAATCA-3` (reverse), 30 cycles; SPT-LCB2 5`- AGCTGCTGAAGTCCTCAAGGA-3` (forward), 5`-GGTCATAGCAGCTTCCACACC-3` (reverse), 29 cycles. Results were normalised using the housekeeping genes: 18S rrna 5`- GCATGGCCGTTCTTAGTTGG-3` (forward), 5`-TGAACGCCACTTGTCCCTCT-3` (reverse), 16 cycles; Synaptophysin 5`-CGATGTGAAGATGGCCACTGA-3` (forward), 5`-
11 MOL CCGAGGTGTTGAGTCCTGAAGTC-3` (reverse), 25 cycles and hypoxanthinphosphoribosyltransferase (HPRT) 5`-TCCTGTGGCCATCTGCCTAGTA-3` (forward), 5`-GGACGCAGCAACTGACATTTCTA-3` (reverse), 29 cycles. All reactions were carried out with taq DNA polymerase (Amersham, Freiburg, Germany) in a MJ PTC 200 thermal cycler (Biozym, Hess. Oldendorf, Germany). Annealing was at 58 C and product size was between base pairs. Quantitative real time PCR was performed using the Quantitect SYBR Green PCR Kit (QIAGEN, Hilden, Germany), 35 cycles, annealing at 58 C and an ABI 7700 Sequence Detector with software version 1.7 (Applied Biosystems, Darmstadt, Germany). The same primer sets described above were used. Results were normalised using the same housekeeping genes as mentioned above. Melting curve analysis was performed to confirm production of a single product in these reactions. Protein determination. Cell protein was quantified as described by Bradford (1976) using bovine serum albumin as a standard. Prior to lipid extraction, cell pellets were homogenized in 400 µl of water and aliquots were used for protein determination. Cell viability assay. The effect of GT11 on cell viability was determined using the Cell Titer- Blue TM Cell Viability Assay from PROMEGA (Mannheim, Germany). The assay was performed in 96-well microtiter plates (Falcon, Heidelberg, Germany) according to manufacturer s instructions. Presentation of data. Results presented as TLC images correspond to data obtained with at least two different cell preparations.
12 MOL Results GT11 is a potent inhibitor of dihydroceramide desaturase in primary cultured cerebellar neurons. The effect of GT11 (Fig. 1) on the activity of dihydroceramide desaturase in primary cultured cerebellar neurons was studied by following the incorporation of L-[3-14 C]serine into cellular ceramide. As illustrated in Fig. 2, a dose-dependent decrease of dihydroceramide desaturation and hence of de novo ceramide formation was observed in the presence of to 1 µm GT11. On the bases of these data an IC 50 of 23 nm was calculated. Furthermore, the effect of GT11 on neuronal desaturase activity was measured in vitro, using cell homogenates as an enzyme source. Activity of dihydroceramide desaturase was inhibited by 83% and 55% when 12.5 µm and 1.25 µm GT11 were directly added to the enzyme assay, respectively. High concentrations of GT11 decrease the incorporation of [ 14 C]serine, but not that of [ 3 H]sphinganine, into cellular sphingolipids in primary cultured cerebellar neurons. Interestingly, when higher amounts ( 5µM) of the inhibitor were added to the culture medium, de novo SL biosynthesis monitored by incorporation of [ 14 C]serine into cellular SLs was strongly reduced (Fig. 3A). Thus, the incorporation of [ 14 C]serine into cellular SLs amounted only 10%, 30% and 50% in the presence of 20 µm, 10µM and 5 µm of GT11, respectively, relative to untreated controls. However, this reduction did not occur when instead of [ 14 C]serine, [ 3 H]sphinganine was used as a biosynthetic precursor (Fig. 3B). These results strongly indicate that concentrations of GT11 from 5 µm upwards affect an enzymatic step upstream of sphinganine formation. The increased radioactive labeling of some SL species in the presence of the desaturase inhibitor GT11 (1-20 µm) when compared with that of SLs in control cells might be explained by the loss of tritium label in the latter as a consequence of desaturation. In addition, the rate of metabolism of saturated SLs formed in the presence of GT11 might differ from that of their desaturated counterparts formed in
13 MOL control cells. Note that the amount of radioactively labeled fatty acids recovered in the lipid fraction decreases with increasing concentrations of GT11 (Fig. 3C), indicating that this compound might interfere with the conversion of [ 3 H]sphinganine, via phosphorylation and lyase cleavage, into phosphoethanolamine and the fatty acid precursor, palmitaldehyde (see also Fig. 9). Worth mentioning is the fact that cell viability was not significantly affected by GT11 up to a concentration of 10 µm for 48 h. High concentrations of GT11 decrease SPT activity in primary cultured cerebellar neurons. To find out if GT11 affects the activity of SPT, the rate-limiting enzyme of de novo SL biosynthesis, neurons were cultured for 24 h in the presence of different concentrations of GT11 prior to the enzyme assay performed in cell homogenates. As shown in Fig 4, in contrast to low concentrations (1µM), which did not affect SPT activity significantly, high concentrations (10 µm) of GT11 reduced the enzyme activity by about 90 %. However, when the same concentrations of inhibitor were directly added to the cell homogenate in the in vitro enzyme assay, no effect on SPT activity was observed (not shown). These results indicate that cell integrity is essential for the effect of GT11 on SPT activity. Cell integrity could be crucial either for metabolism of GT11, or for the regulation mechanism induced by higher doses of the inhibitor. Metabolism of radioactively labeled GT11 in primary cultured cerebellar neurons. To find out, if GT11 itself or a metabolic product derived from GT11 is responsible for the decrease of de novo SL biosynthesis, the metabolic fate of two different radioactively labeled inhibitors was investigated in primary cultured cerebellar neurons. [ 14 C]GT11 (see materials) is labeled in its fatty acid moiety, whereas [ 3 H]GT11 is labeled in the sphingoid part of the molecule (Fig. 1). Both, differentially labeled radioactive analogs of GT11 were rapidly internalized in the cells. After one hour only about 30 % of the added radioactivity was detectable in the culture medium. This value remained constant up to 24 hours. About 75 % of
14 MOL lipid associated radioactivity migrated with [ 14 C]GT11 (Fig. 5A). In case of [ 3 H]GT11 most of the metabolized fraction of the lipid (about 25%) were recovered in the phosphatidylethanolamine (PE) fraction (Fig. 5B), indicating that the compound was degraded down to phosphorylethanolamine by the stepwise action of ceramidase, sphinganine kinase and S1P lyase (see also Fig. 9). Interestingly, the amount of labeled PE formed from [ 3 H]GT11 decreased with increasing concentrations of added GT11. This result indicates that GT11 might interfere in a concentration dependent manner with at least one of the catabolic enzymes mentioned above. High concentrations of GT11 induce an accumulation of phosphorylated long chain bases. Although GT11 appears to be metabolically quite stable, it became clear from the studies described above that the compound is partially directed into the catabolic pathway. Since phosphorylation is a key-step in this pathway, the cells were labeled with 32 P i prior to treatment with different concentrations of either GT11 or its deacylated derivative GT17 (Fig. 1), in order to get a better insight into the fate of GT11 derived phophorylated metabolites. After 1 h levels of phosphorylated sphingoid bases were comparable in controls and GT11 (both, low and high concentration) as well as in GT17 treated cells (not shown). In contrast, after 24 h a clear cut accumulation of both, S1P and DHS1P was observed in the presence of the high concentration of GT11, while no changes of the two phosphorylated sphingoid bases were detected in the other samples (Fig. 6). The accumulation of DHS1P was especially surprising since SPT activity was found to be decreased after GT11 (10 µm) treatment. Note that phosphorylated GT17 was equally abundant in cells treated with GT17 or with the high concentration of GT11 (10 µm). In light of these findings, it appears likely that high concentrations of GT11 affect the S1P lyase activity prior to SPT down-regulation.
15 MOL High concentrations of GT11 decrease sphinsosine-1-phosphate lyase activity in primary cultured cerebellar neurons. As shown above, high concentrations of GT11 induce an accumulation of phosphorylated long chain bases in cultured neurons after 24 h of incubation. In addition, the formation of PE from [ 3 H]GT11 is reduced by high concentrations of GT11 (see above). Finally, the formation of fatty acids from [4,5-3 H]sphinganine is strongly reduced in the presence of 10 µm of GT11 (see Fig 3C). Taken together these findings strongly suggest that GT11 at high concentrations interferes with the activity of S1P lyase. Therefore, lyase activity was measured after incubating the cells for 24 h with 1 and 10 µm GT11, respectively. As shown in Fig 7, lyase activity was strongly reduced after pre-incubation of cells in the presence of 10 µm of GT11, whereas 1 µm of GT11 had no effect. However, when GT11 (10 µm) was added directly to the cell homogenate in the in vitro lyase assay it had no effect on enzyme activity (not shown), suggesting that cell integrity is crucial for its inhibitory effect. Deacylated GT11 (GT17) does not decrease neither [ 14 C]serine incorporation into cellular sphingolipids nor [ 3 H]sphinganine incorporation into cellular fatty acids. As shown above, both GT11 (10 µm) and GT17 (10 µm) induce the formation of phosphorylated GT17. To analyze whether this metabolite of GT11 mimics the effects on SL metabolism observed at high concentrations of GT11, de novo SL biosynthesis as well as [ 3 H]sphinganine catabolism, were studied in the presence of GT17. The deacylated derivative of GT11 had no effect, neither on de novo SL biosynthesis nor on the incorporation of [ 3 H]sphinganine into cellular fatty acids (not shown). High concentrations of GT11 do not affect expression of SPT and of sphingosine-1- phosphate lyase in primary cultured cerebellar neurons. To find out if reduction of enzymatic activity of S1P lyase and also of SPT in neurons treated with high concentrations
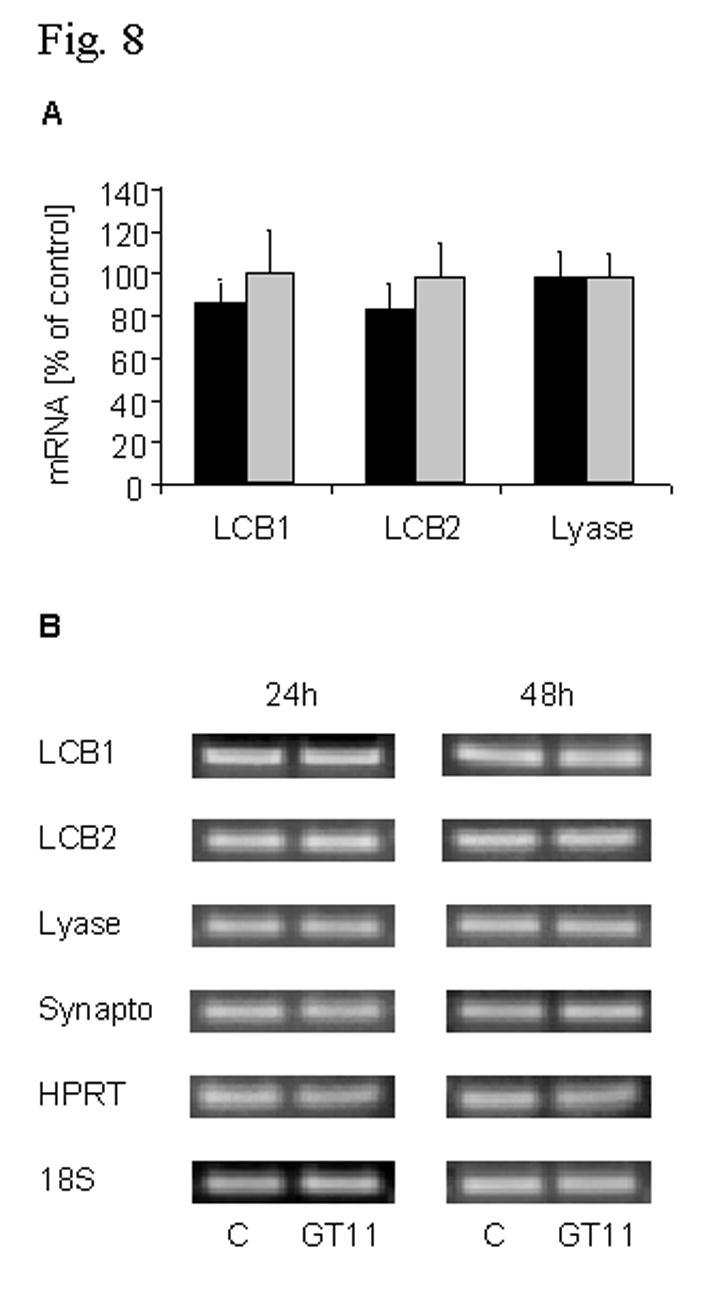
16 MOL of GT11 occurs on transcriptional level, mrna amounts of SPT including the regulatory (LCB1) and the catalytically active subunit (LCB2) as well as of S1P lyase were assessed by PCR methods. As illustrated in Fig. 8 no significant changes in their expression could be observed upon treatment with 10 µm of GT11 for up to 48 h.
17 MOL Discussion Previous results from our laboratories have shown that the cyclopropene-containing ceramide analog GT11 is a competitive inhibitor of dihydroceramide desaturase activity in rat liver microsomes (Triola et al., 2001 and 2003). In the present study, GT11 was found to exhibit a much more pronounced inhibitory effect on dihydroceramide desaturase activity in primary cultured cerebellar neurons. The calculated IC 50 of 23 nm is by about three and two orders of magnitude lower than that found in rat liver microsomes and in neuronal cell homogenates, respectively. Thus the inhibitory property of GT11 is much more pronounced in vivo than in vitro. This could be explained by the in vivo situation that might allow for a local sub-cellular enrichment of the inhibitor at its site of action, the ER. This assumption is supported by earlier observations that metabolically stable ceramide analogues become stuck to intracellular membranes especially the ER whereas Golgi staining requires metabolic processing of ceramide to glucosylceramide and/or sphingomyelin (Putz and Schwarzmann 1995). Since we have shown that GT11 is not metabolized to either of the two SLs, it appears very likely that it becomes stuck to ER membranes. Preliminary experiments with the fluorescent analog NBD-GT11 (N-6(7-nitro-2,1,3-benzoxadiazol-4-yl)aminododecanoyl-Derythro-GT11) confirm this assumption. However, tissue and species specificity cannot be excluded either at present time. Besides a high inhibitory potency, the quality of an inhibitor is also dependent on its specificity towards a certain enzymatic activity. In case of GT11, no collateral effects on SL metabolism in the concentration range of µm could be observed. However, doses of GT11 5 µm markedly blocked de novo SL biosynthesis in primary cultured neurons when radioactively labeled serine was used as a biosynthetic precursor. Surprisingly, SL biosynthesis was not affected at similar concentrations of GT11 when tritiated sphinganine was used as a biosynthetic precursor instead of serine. This result indicates that at higher doses (from 5 µm upwards), besides inhibiting dihydroceramide desaturase activity, GT11
18 MOL interferes with a step upstream of de novo sphinganine formation. We could indeed show that the activity of SPT, the rate-limiting enzyme of SL biosynthesis, was markedly decreased when cells were cultured for 24 h with doses of the inhibitor as high as 5 µm and upwards. However, expression of SPT was not significantly affected. Also, the same high concentrations of GT11 did not affect SPT activity in cell homogenates in vitro, thus indicating that the integrity of the cell was required for this effect. Two possible explanations appeared most likely. i) GT11 rather down-regulates than inhibits SPT as shown before for long chain bases (Mandon et al., 1991) and also for the synthetic sphingosine analog cis-4- methylsphingosine (van Echten-Deckert et al., 1997). ii) Not GT11 itself, but a metabolic product derived from GT11 is responsible for the decreased SPT activity. The latter possibility is, however, widely excluded by our experimental data. Thus, metabolic studies with radioactively labeled GT11 analogs indicated that only a low percentage of the inhibitor is subjected to metabolism and directed rather to the catabolic than to the anabolic route. However, GT17 the sphingoid base that would derive from enzymatic deacylation of GT11 did not affect incorporation of radioactively labeled serine into cellular SLs when added to the culture medium. And finally, labeling experiments with inorganic phosphate demonstrated that only high concentrations of GT11 induce the accumulation of S1P and especially that of DHS1P after 24 h. In analogy with previous results showing that phosphorylated sphingoid bases play a critical role in down-regulation of the de novo pathway by affecting the ratelimiting biosynthetic enzyme, SPT (van Echten-Deckert et al., 1997, Le Stunff et al., 2002), we believe that those two compounds, S1P and DHS1P, are responsible for the decreased SPT activity induced by high concentrations of GT11. Since, as mentioned above, only a low percentage of GT11 is catabolyzed and GT17 does not mimic any of the effects of GT11, we concluded that GT11 itself, in addition to being a very efficient inhibitor of dihydroceramide desaturase activity, also induces accumulation of phosphorylated sphingoid bases when applied in high concentrations ( 5µM) to cultured neurons.
19 MOL The enzyme responsible for a rapid breakdown of sphingoid phosphates is S1P lyase (van Veldhoven and Mannaerts, 1993; van Veldhoven, 2000). It catalyzes the cleavage of S1P and/or DHS1P into phosphoethanolamine and hexadecenal and/or hexadecanal (Fig. 9). Both breakdown products are then mainly incorporated into glycerophospholipids. After its activation with CTP, phosphoethanolamine is primarily used in the PE biosynthetic pathway, whereas the fatty aldehyde is oxidized to the respective fatty acid, which can be linked to glycerol via an alkali-labile ester bond. All our experimental data point to an interference of GT11 with S1P lyase activity. i) Culturing of neurons for 24 h in the presence of 10 µm of GT11 almost completely abolished lyase activity, whereas 1 µm GT11 did not affect this enzyme at all. ii) When [ 3 H]sphinganine was used in metabolic labeling experiments, incorporation of label into fatty acids (derived from the lyase-born aldehyde), was inhibited by GT11 in a concentration dependent manner (lowest effective concentration tested being 5 µm). iii) The amount of radioactive PE derived from [ 3 H]GT11 (tritiated at carbon atom C1 of the sphingoid base) decreased with increasing concentrations of GT11 applied. The mechanism by which GT11 affects lyase activity is still unknown at present. Although we did not find any regulation of lyase on transcriptional level, our results indicate that cell integrity is crucial for decrease of lyase activity by high concentrations of GT11 since its direct addition to the in vitro assay did not affect enzymatic activity. The latter finding argues against the possibility that GT11 interferes with lyase activity on molecular level by the reaction of its cyclopropene ring with the catalytically essential cysteine residue(s) (van Veldhoven et al., 2000). In addition, lyase is a pyridoxal phosphate-dependent enzyme (van Veldhoven and Mannaerts, 1993). Although GT11 is metabolized at a low rate, it is possible that its long-chain base phosphate interacts with the lyase-pyridoxal phosphate complex to form the substrate-enzyme imino-intermediate, thus competing with the natural substrate. However, since deacylated GT11 (GT17) did not mimic any of the effects of GT11 in cultured neurons, we also excluded the competitive inhibition of lyase by this metabolic
20 MOL product of GT11. Thus, in contrast to the inhibitory effect of GT11 on lyase activity, which requires cell integrity, inhibition of dihydroceramide desaturase is independent of such requirements and it occurs at much lower concentrations. In conclusion, the data presented in this study show that GT11 has a dual, dose-dependent effect on sphingolipid metabolism in cultured cerebellar neurons (Fig.9). At low concentrations ( nm) it specifically inhibits dihydroceramide desaturase, thus preventing the formation of ceramide and leading to the formation of saturated complex sphingolipids from dihydroceramide. The latter is, alternatively, subjected to catabolic breakdown by hydrolysis (ceramidase), phosphorylation (S1P kinase) and retroaldolic cleavage (S1P lyase) to phosphoethanolamine and palmitaldehyde. At high concentrations (from 5 µm upwards) GT11 induces in addition accumulation of S1P and DHS1P, most probably by interfering with S1P lyase activity. Accumulated sphingoid phosphates downregulate SPT activity and hence de novo SL biosynthesis, most probably via a feedback mechanism. Due to its highly specific inhibitory potency, we believe that GT11, despite its collateral effects at concentrations as high as 5 µm and upwards, represents an indispensable tool to study the physiological function of dihydroceramide desaturation. In addition, its lack of cytotoxicity in primary cultured neurons is a prerequisite for future studies concerning its potential neuroprotective properties against ceramide mediated stress responses. Acknowledgements We thank Andrea Raths for excellent technical assistance, Dr. J. Casas for helpful discussions and Prof. K. Sandhoff for the possibility to work in his department. G. Triola would like to thank Almirall Prodesfarma S.A.for awarding her The Young Researchers' Prize.
21 MOL References Bielawska A, Crane HM, Liotta D, Obeid LM, and Hannun YA (1993) Selectivity of ceramide-mediated biology. Lack of activity of erythro-dihydroceramide. J Biol Chem 268: Bleicher RJ and Cabot MC (2002) Glucosylceramide synthase and apoptosis. Biochim Biophys Acta 1585: Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72: Hannun YA and Obeid LM (2002) The Ceramide-centric universe of lipid-mediated cell regulation: stress encounters of the lipid kind. J Biol Chem 277: Hannun YA, and Luberto C (2000) Ceramide in the eukaryotic stress response. Trends Cell Biol 10: Huwiler A, Kolter T, Pfeilschifter J, and Sandhoff K (2000) Physiology and pathophysiology of sphingolipid metabolism and signaling. Biochim Biophys Acta 1485: Kolesnick R (2002) The therapeutic potential of modulating the ceramide/sphingomyelin pathway. J Clin Invest 110:3-8. Le Stunff H, Galve-Roperh I, Peterson C, Milstien S, and Spiegel S (2002) Sphingosine- 1-phosphate phosphohydrolase in regulation of sphingolipid metabolism and apoptosis. J Cell Biol 158: Mandon EC, van Echten G, Birk R, Schmidt RR, and Sandhoff K (1991) Sphingolipid biosynthesis in cultured neurons. Down-regulation of serine palmitoyltransferase by sphingoid bases. Eur J Biochem 198: Marchesini N and Hannun YA (2004) Acid and neutral sphingomyelinases: roles and mechanisms of regulation. Biochem Cell Biol 82:27-44.
22 MOL Merril AH Jr and Wang E (1992) Ceramide synthase. Methods Enzymol 209: Michel C and van Echten-Deckert G (1997) Conversion of dihydroceramide to ceramide occurs at the cytosolic face of the endoplasmic reticulum. FEBS Lett 416: Michel C, van Echten-Deckert G, Rother J, Sandhoff K, Wang E, Merrill AHJr (1997) Characterization of ceramide synthesis. A dihydroceramide desaturase introduces the 4,5-trans-double bond of sphingosine at the level of dihydroceramide. J Biol Chem 272: Putz U, Schwarzmann G (1995) Golgi staining by two fluorescent ceramide analogues in cultured fibroblasts requires metabolism. Eur J Cell Biol 68: Schulze H, Michel C, and van Echten-Deckert G (2000) Dihydroceramide desaturase. Methods Enzymol 311: Schwarzmann G (1978) A simple and novel method for tritium labeling of gangliosides and other sphingolipids. Biochim Biophys Acta 529: Spiegel S and Milstien S (2003) Sphingosine-1-phosphate: an enigmatic signalling lipid. Nature Reviews 4: Svennerholm L (1963) Chromatographic separation of human brain gangliosides. J Neurochem 10: Ternes P, Franke S, Zähringer U, Sperling P, and Heinze E (2002) Identification and characterization of a sphingolipid delta 4-desaturase family. J Biol Chem 277: Triola G, Fabriàs G, and Llebaria A (2001) Synthesis of a cyclopropene analogue of ceramide, a potent inhibitor of dihydroceramide desaturase. Angew Chem Int Ed Engl 40: Triola G, Fabriàs G, Casas J, and Llebaria A (2003) Synthesis of cyclopropene analogues of ceramide and their effect on dihydroceramide desaturase. J Org Chem 68:
23 MOL van Echten-Deckert G ( 2000) Sphingolipid extraction and analysis by thin-layer chromatography. Methods Enzymol 312: van Echten-Deckert G and Sandhoff K (1999) Organization and topology of sphingolipid metabolism, in Comprehensive Natural Products Chemistry (Pinto BM ed) pp , Elsevier Science Amsterdam-Oxford. van Echten-Deckert G, Giannis A, Schwarz A, Futerman A, and Sandhoff K (1998) 1- Methylthiodihydroceramide, a novel analog of dihydroceramide, stimulates sphinganine degradation resulting in decreased de novo sphingolipid biosynthesis. J Biol Chem 273: van Echten-Deckert G, Zschoche A, Bär T, Schmidt RR, Raths A, Heinemann T, and Sandhoff K (1997) cis-4-methylsphingosine decreases sphingolipid biosynthesis by specifically interfering with serine palmitoyltransferase activity in primary cultured neurons. J Biol Chem 272: van Veldhoven PP (2000) Sphingosine-1-phosphate lyase. Methods Enzymol 311: van Veldhoven PP, and Mannaerts GP (1993) Sphingosine-phosphate lyase. Adv Lipid Res 26: van Veldhoven PP, Gijsbers S, Mannaerts GP, Vermeesch JR, Brys V (2000) Human sphingosine-1-phosphate lyase: cdna cloning, functional expression studies and mapping to chromosome 10q22(1). Biochim Biophys Acta 1487: Yatomi Y, Ruan F, Ohta H, Welch RJ, Hakomori S, and Igarashi Y (1995) Quantitative measurement of sphingosine 1-phosphate in biological samples by acylation with radioactive acetic anhydride. Anal Biochem 230: Zhang H, Desai NN, Olivera A, Seki T, Brooker G, and Spiegel S (1991) Sphingosine-1- phosphate, a novel lipid, involved in cellular proliferation. J Cell Biol 114:
24 MOL Footnotes This study was supported by the Deutsche Forschungsgemeinschaft and the Spanish Ministry of Science and Technology (Grants BQU and BQU ). GT was financially supported by the Generalitat de Catalunya with a pre-doctoral fellowship and a 6- month sojourn in Bonn. PD Dr. Gerhild van Echten-Deckert, Kekulé-Institut für Organische Chemie und Biochemie, Gerhard-Domagk-Str. 1, Bonn, Germany
25 MOL Legends for figures Fig. 1. Chemical Structure (A) and synthesis of [1-3 H]GT11 (B). The tritiated GT11 analog was prepared from the cyclic carbamate (i). Oxidation (b, Dess-Martin Periodinane, CH 2 Cl 2 ) to the aldehyde (ii) and its further reduction (c, NaB 3 H 4 /methanol) yielded the tritiated alcohol (iii), which, upon deprotection and acylation afforded [1-3 H]GT11. Reagents and conditions of the reactions a and d are as described by Triola et al. (2001 and 2003) Fig. 2. The effect of GT11 on dihydroceramide desaturation in primary cultured neurons. Cells were incubated in the absence (C, control) or presence of the indicated concentrations of GT11. After 24 h respective media were renewed and [ 14 C]serine added for additional 24 h. Then cells were harvested and lipids extracted and isolated as described in Materials and Methods. Ceramide (Cer) was separated from dihydroceramide (DHCer) by TLC on borate treated silica gel plates with chloroform/methanol (9:1, by volume) as solvent system. The radioactively labeled ceramide and dihydroceramide were quantitatively evaluated as described in Materials and Methods. Inhibition of desaturation of DHCer to Cer was calculated relative to controls. The results are given as the mean of double determinations from one representative experiment that was repeated twice with similar results. The standard deviation values were always in the range of ± 5 % of the mean. Fig. 3. The effect of GT11 on [ 14 C]serine and [ 3 H]sphinganine incorporation into cellular sphingolipids. Primary cultured neurons were incubated in the absence (C, control) or presence of the indicated concentrations of GT11. After 24 h 1 µci of either [ 14 C]serine (A) or of [ 3 H]sphinganine (B and C) were added to the renewed medium. 24 h later cells were harvested and lipids extracted, isolated, separated by TLC and detected as described in Materials and Methods. The solvent system used was chloroform/methanol/ 0.22% aqueous CaCl 2, 60:35:8, by volume (A and B) or chloroform/methanol/ acetic acid, 190:9:1, by
26 MOL volume (C). The terminology of gangliosides (GT1b, GD1b, GD1a, GD3, GM1, GM2, GM3) is according to Svennerholm (1963). Other abbreviations are as follows: SM, sphingomyelin; LacCer, lactosylceramide; GlcCer, glucosylceramide; * Unidentified bands Fig. 4. The effect of GT11 on SPT activity in primary cultured neurons. Cells were incubated in the absence (C, control) or presence of the indicated concentrations of GT11. After 24 h cells were harvested, homogenized and enzyme activity determined as described in Materials and Methods. B, blank with boiled homogenate. Data are from three separate experiments, each with double determinations. Fig. 5. Metabolism of radioactively labeled GT11 in primary cultured neurons. Cells were incubated in the presence of the indicated concentrations of GT11 including 5 nci of [ 14 C]GT11 (A) or 0.72 µci of [ 3 H]GT11 (B), respectively. After 24 h cells were harvested and lipids extracted, isolated, separated by TLC (A: chloroform/methanol/ acetic acid, 190:9:1, by volume; B: chloroform/methanol/ 0.22% aqueous CaCl 2, 60:35:8, by volume) and detected as described in Materials and Methods. Note that alkaline treatment was omitted to keep phospholipids intact. PE, phosphatidylethanolamine. Fig. 6. Determination of 32 P i -labeled sphingoid phosphates in primary cultured cerebellar neurons. Cells were washed with phosphate-free MEM and incubated with the same medium containing 32 P i (40 µci/ml) for 24 h. Then vehicle (lane C), or the indicated concentration of GT11 or GT17 was added for 24 h. Cells were harvested and sphingosine phosphates extracted, resolved by TLC and determined as described in Materials and Methods. The R F value of authentic sphingoid phosphates is indicated. Similar data were obtained in two separate experiments, each with double determinations.
27 MOL Fig. 7. The effect of GT11 on S1P lyase activity in primary cultured neurons. Cells were incubated in the absence (C, control) or presence of the indicated concentrations of GT11. After 24 h cells were harvested, homogenized and S1P lyase activity determined as described in Materials and Methods. B, blank with boiled homogenate. Data are from two separate experiments, each with double determinations. Fig. 8. The effect of GT11 on mrna levels of SPT and S1P lyase in primary cultured neurons. Cells were incubated for the indicated times (black bars 24 h, gray bars 48 h) in the absence (control) or presence of 10 µm of GT11. After 24 h respective media were renewed. Levels of SPT (LCB1 and LCB2) and of S1P lyase mrna were determined by quantitative real time PCR (A) and by semi-quantitative RT-PCR (B) as described in Materials and Methods. (A) Data are means from three different experiments, each with triple determinations. (B) One representative out of three experiments with similar results from different cell preparations. C, control; HPRT, hypoxanthinephosphoribosyltransferase; LCB, long chain base; Synapto, synaptophysine; 18S, 18S rrna. Fig. 9. Proposed mechanism for the effect of GT11 on sphingolipid metabolism in primary cultured cerebellar neurons. At sub-micromolar concentrations ( nm) GT11 specifically inhibits dihydroceramide desaturase. At concentrations above 5 µm, GT11 also decreases lyase activity. This effect results in the accumulation of sphingoid-1-phosphates, which down-regulate SPT. Note that an accumulation of S1P is possible despite a complete inhibition of dihydroceramide desaturase from catabolic ceramide pools formed by breakdown of sphingomyelin (SM) and of glycosphingolipids (GSL). AL, aldehyde; EAP, ethanolaminephosphate; (DH)Cer, (dihydro)ceramide; (DH)GSL, (dihydro)glycosphingolipids; (DH)SM, (dihydro)sphingomyelin; Sa, sphinganine; Sa1P, sphinganine-1-phosphate; So, sphingosine; S1P, sphingosine-1-phosphate.
28
29

30
31

32
33
34
35
36
SPHINGOLIPIDS AS TARGETS FOR RECOVERY FROM SPINAL CORD INJURIES: DECIPHERING THE ROLE OF SPHINGOSINE-1-PHOSPHATE
 SPHINGOLIPIDS AS TARGETS FOR RECOVERY FROM SPINAL CORD INJURIES: DECIPHERING THE ROLE OF SPHINGOSINE-1-PHOSPHATE Josefina Casas Brugulat Institut Química Avançada de Catalunya CSIC Rodrigo Martínez Maza
SPHINGOLIPIDS AS TARGETS FOR RECOVERY FROM SPINAL CORD INJURIES: DECIPHERING THE ROLE OF SPHINGOSINE-1-PHOSPHATE Josefina Casas Brugulat Institut Química Avançada de Catalunya CSIC Rodrigo Martínez Maza
Sphingosine-1-phosphate signaling and cardiac fibrosis. Department of Physiology, Kanazawa University School of Medicine, Kanazawa, Japan
 96 Special Issue: Cellular and Molecular Bases for Fibrotic Diseases Review Article Sphingosine-1-phosphate signaling and cardiac fibrosis Noriko Takuwa 1, 2, ), Yasuo Okamoto 1), Kazuaki Yoshioka 1) and
96 Special Issue: Cellular and Molecular Bases for Fibrotic Diseases Review Article Sphingosine-1-phosphate signaling and cardiac fibrosis Noriko Takuwa 1, 2, ), Yasuo Okamoto 1), Kazuaki Yoshioka 1) and
Overview on the identification of different classes of. lipids by HPTLC (High Performance Thin Layer. Chromatography) and ITLC (Immuno Thin Layer
 and ITLC (Immuno Thin Layer") Overview on the identification of different classes of lipids by HPTLC (High Performance Thin Layer Chromatography) and ITLC (Immuno Thin Layer Chromatography) Iuliana Popa 1, Marie-Jeanne David 2, Daniel
Overview on the identification of different classes of lipids by HPTLC (High Performance Thin Layer Chromatography) and ITLC (Immuno Thin Layer Chromatography) Iuliana Popa 1, Marie-Jeanne David 2, Daniel
Prerequisites Protein purification techniques and protein analytical methods. Basic enzyme kinetics.
 Case 19 Purification of Rat Kidney Sphingosine Kinase Focus concept The purification and kinetic analysis of an enzyme that produces a product important in cell survival is the focus of this study. Prerequisites
Case 19 Purification of Rat Kidney Sphingosine Kinase Focus concept The purification and kinetic analysis of an enzyme that produces a product important in cell survival is the focus of this study. Prerequisites
Test Bank for Lehninger Principles of Biochemistry 5th Edition by Nelson
 Test Bank for Lehninger Principles of Biochemistry 5th Edition by Nelson Link download full: http://testbankair.com/download/test-bank-forlehninger-principles-of-biochemistry-5th-edition-by-nelson/ Chapter
Test Bank for Lehninger Principles of Biochemistry 5th Edition by Nelson Link download full: http://testbankair.com/download/test-bank-forlehninger-principles-of-biochemistry-5th-edition-by-nelson/ Chapter
MEMBRANE LIPIDS I and II: GLYCEROPHOSPHOLIPIDS AND SPHINGOLIPIDS
 December 6, 2011 Lecturer: Eileen M. Lafer MEMBRANE LIPIDS I and II: GLYCEROPHOSPHOLIPIDS AND SPHINGOLIPIDS Reading: Stryer Edition 6: Chapter 26 Images: All images in these notes were taken from Lehninger,
December 6, 2011 Lecturer: Eileen M. Lafer MEMBRANE LIPIDS I and II: GLYCEROPHOSPHOLIPIDS AND SPHINGOLIPIDS Reading: Stryer Edition 6: Chapter 26 Images: All images in these notes were taken from Lehninger,
Hiroya Hidaka *1), Masaki Takiwaki 2), Mine Yamashita 2), Shinya Otsuki 1), Kenji Kawasaki 3), Mitsutoshi Sugano 3) and Takayuki Honda 4)
, Masaki Takiwaki 2), Mine Yamashita 2), Shinya Otsuki 1), Kenji Kawasaki 3), Mitsutoshi Sugano 3) and Takayuki Honda 4)") Mild acid hydrolysis of sphingolipids yields lysosphingolipids: a matrix-assisted laser desorption and ionization time-of-flight mass spectrometry study Hiroya Hidaka *1), Masaki Takiwaki 2), Mine Yamashita
Mild acid hydrolysis of sphingolipids yields lysosphingolipids: a matrix-assisted laser desorption and ionization time-of-flight mass spectrometry study Hiroya Hidaka *1), Masaki Takiwaki 2), Mine Yamashita
Case 19 Purification of Rat Kidney Sphingosine Kinase
 Case 19 Purification of Rat Kidney Sphingosine Kinase Focus concept The purification and kinetic analysis of an enzyme that produces a product important in cell survival is the focus of this study. Prerequisites
Case 19 Purification of Rat Kidney Sphingosine Kinase Focus concept The purification and kinetic analysis of an enzyme that produces a product important in cell survival is the focus of this study. Prerequisites
Development of Sphingosine Kinase Selective Inhibitors for Cancer Therapy
 Development of Sphingosine Kinase Selective Inhibitors for Cancer Therapy Zuping Xia College of Pharmacy Washington State University Sphingolipids, similar to phospholipids, are composed of a polar head
Development of Sphingosine Kinase Selective Inhibitors for Cancer Therapy Zuping Xia College of Pharmacy Washington State University Sphingolipids, similar to phospholipids, are composed of a polar head
Lipids. Lipids: a Diverse group of chemicals. Storage Lipids: derivatives of fatty acids. 11/21/10
 1 Lipids Lehninger 3 rd ed. Chapter 11 (For biosynthesis see Chapter 21) 2 Lipids: a Diverse group of chemicals Insolubility in water. Fats and oils: energy stores. Phospholipids and sterols: structural
1 Lipids Lehninger 3 rd ed. Chapter 11 (For biosynthesis see Chapter 21) 2 Lipids: a Diverse group of chemicals Insolubility in water. Fats and oils: energy stores. Phospholipids and sterols: structural
UMR 8612, Faculty of Pharmacy Chatenay-Malabry. Natura-Brasil. EA Laboratory of Dermatological Research,
 Iuliana Popa 1, Noëlle Remoué 2 and Jacques Portoukalian 3 1 UMR 8612, Faculty of Pharmacy Chatenay-Malabry 2 Natura-Brasil 3 EA 41 69 Laboratory of Dermatological Research, University of Lyon I, Faculty
Iuliana Popa 1, Noëlle Remoué 2 and Jacques Portoukalian 3 1 UMR 8612, Faculty of Pharmacy Chatenay-Malabry 2 Natura-Brasil 3 EA 41 69 Laboratory of Dermatological Research, University of Lyon I, Faculty
3-keto-Dihydrosphingosine is an Important Early Metabolite of Sphingolipid Biosynthesis
 H C 2 H H H H H H H C 2 H H H H H H H H H H NHAc C 2 H H H H C 2 H H H H H H C 17 H 35 NH H N E W S L E T T E R F R G LY C / S P H I N G L I P I D R E S E A R C H N V E M B E R 2 0 1 8 3-keto-Dihydrosphingosine
H C 2 H H H H H H H C 2 H H H H H H H H H H NHAc C 2 H H H H C 2 H H H H H H C 17 H 35 NH H N E W S L E T T E R F R G LY C / S P H I N G L I P I D R E S E A R C H N V E M B E R 2 0 1 8 3-keto-Dihydrosphingosine
EXPERIMENT 13: Isolation and Characterization of Erythrocyte
 EXPERIMENT 13: Isolation and Characterization of Erythrocyte Day 1: Isolation of Erythrocyte Steps 1 through 6 of the Switzer & Garrity protocol (pages 220-221) have been performed by the TA. We will be
EXPERIMENT 13: Isolation and Characterization of Erythrocyte Day 1: Isolation of Erythrocyte Steps 1 through 6 of the Switzer & Garrity protocol (pages 220-221) have been performed by the TA. We will be
ASSAY OF SPHINGOMYELINASE ACTIVITY
 ASSAY OF SPHINGOMYELINASE ACTIVITY Protocol for Protein Extraction Stock Solution 1. Leupeptin/hydrochloride (FW 463.0,
ASSAY OF SPHINGOMYELINASE ACTIVITY Protocol for Protein Extraction Stock Solution 1. Leupeptin/hydrochloride (FW 463.0,
ANSC (NUTR) 618 LIPIDS & LIPID METABOLISM Membrane Lipids and Sphingolipidsd
 618 LIPIDS & LIPID METABOLISM Membrane Lipids and Sphingolipidsd") ANSC (NUTR) 618 LIPIDS & LIPID METABOLISM Membrane Lipids and Sphingolipidsd I. Classes of membrane lipids A. Glycerolipids (quantitatively the most important of the three membrane lipids) B. Shingolipids
ANSC (NUTR) 618 LIPIDS & LIPID METABOLISM Membrane Lipids and Sphingolipidsd I. Classes of membrane lipids A. Glycerolipids (quantitatively the most important of the three membrane lipids) B. Shingolipids
Block A: Membrane Biology & Biochemistry. Lipid signalling and sphingolipid function
 Block A: Membrane Biology & Biochemistry Lipid signalling and sphingolipid function 10. - 14.11.2014 Gerhild van Echten-Deckert Tel. 73 2703 E-mail: g.echten.deckert@uni-bonn.de www.limes-institut-bonn.de
Block A: Membrane Biology & Biochemistry Lipid signalling and sphingolipid function 10. - 14.11.2014 Gerhild van Echten-Deckert Tel. 73 2703 E-mail: g.echten.deckert@uni-bonn.de www.limes-institut-bonn.de
Total Phosphatidic Acid Assay Kit
 Product Manual Total Phosphatidic Acid Assay Kit Catalog Number MET- 5019 100 assays FOR RESEARCH USE ONLY Not for use in diagnostic procedures Introduction Phosphatidic Acid (PA) is a critical precursor
Product Manual Total Phosphatidic Acid Assay Kit Catalog Number MET- 5019 100 assays FOR RESEARCH USE ONLY Not for use in diagnostic procedures Introduction Phosphatidic Acid (PA) is a critical precursor
Biological role of lipids
 Lipids Lipids Organic compounds present in living organisms, insoluble in water but able to be extracted by organic solvents such as: chloroform, acetone, benzene. Extraction = the action of taking out
Lipids Lipids Organic compounds present in living organisms, insoluble in water but able to be extracted by organic solvents such as: chloroform, acetone, benzene. Extraction = the action of taking out
Phospholipids Metabolism
 Chapter VI: Phospholipids Metabolism Dr. Sameh Sarray Hlaoui Phospholipids Features: Amphipatic: - Hydrophobic head: fatty acids - Hydropholic head: P group+ alcohol Composed of alcohol attached by a phosphodiester
Chapter VI: Phospholipids Metabolism Dr. Sameh Sarray Hlaoui Phospholipids Features: Amphipatic: - Hydrophobic head: fatty acids - Hydropholic head: P group+ alcohol Composed of alcohol attached by a phosphodiester
PRODUCT: RNAzol BD for Blood May 2014 Catalog No: RB 192 Storage: Store at room temperature
 PRODUCT: RNAzol BD for Blood May 2014 Catalog No: RB 192 Storage: Store at room temperature PRODUCT DESCRIPTION. RNAzol BD is a reagent for isolation of total RNA from whole blood, plasma or serum of human
PRODUCT: RNAzol BD for Blood May 2014 Catalog No: RB 192 Storage: Store at room temperature PRODUCT DESCRIPTION. RNAzol BD is a reagent for isolation of total RNA from whole blood, plasma or serum of human
Abdallah Q& Razi. Faisal
 27 & Ahmad Attari م ح م د ي وس ف Abdallah Q& Razi Faisal Sphingophospolipids - The backbone of sphingophospholipids is sphingosine, unlike glycerophospholipids with a glycerol as the backbone. Which contains
27 & Ahmad Attari م ح م د ي وس ف Abdallah Q& Razi Faisal Sphingophospolipids - The backbone of sphingophospholipids is sphingosine, unlike glycerophospholipids with a glycerol as the backbone. Which contains
Chapter 26 Biochemistry 5th edition. phospholipids. Sphingolipids. Cholesterol. db=books&itool=toolbar
 http://www.ncbi.nlm.nih.gov/sites/entrez? db=books&itool=toolbar 1 The surface of a soap bubble is a bilayer formed by detergent molecules 2 Chapter 26 Biochemistry 5th edition phospholipids Sphingolipids
http://www.ncbi.nlm.nih.gov/sites/entrez? db=books&itool=toolbar 1 The surface of a soap bubble is a bilayer formed by detergent molecules 2 Chapter 26 Biochemistry 5th edition phospholipids Sphingolipids
Lipids and Membranes
 Lipids and Membranes Presented by Dr. Mohammad Saadeh The requirements for the Pharmaceutical Biochemistry I Philadelphia University Faculty of pharmacy Lipids and Membranes I. overview Lipids are related
Lipids and Membranes Presented by Dr. Mohammad Saadeh The requirements for the Pharmaceutical Biochemistry I Philadelphia University Faculty of pharmacy Lipids and Membranes I. overview Lipids are related
Membrane Lipids & Cholesterol Metabolism
 Membrane Lipids & Cholesterol Metabolism Learning Objectives 1. How Are Acylglycerols and Compound Lipids Produced? 2. The synthesis of Sphingolipids from Ceramide 3. Diseases due to Disruption of Lipid
Membrane Lipids & Cholesterol Metabolism Learning Objectives 1. How Are Acylglycerols and Compound Lipids Produced? 2. The synthesis of Sphingolipids from Ceramide 3. Diseases due to Disruption of Lipid
Structural determinants of sphingolipid recognition by commercially available anti-ceramide antibodies
 Structural determinants of sphingolipid recognition by commercially available anti-ceramide antibodies L. Ashley Cowart, Zdzislaw Szulc, Alicja Bielawska, and Yusuf A. Hannun 1 Department of Biochemistry
Structural determinants of sphingolipid recognition by commercially available anti-ceramide antibodies L. Ashley Cowart, Zdzislaw Szulc, Alicja Bielawska, and Yusuf A. Hannun 1 Department of Biochemistry
Work-flow: protein sample preparation Precipitation methods Removal of interfering substances Specific examples:
 Dr. Sanjeeva Srivastava IIT Bombay Work-flow: protein sample preparation Precipitation methods Removal of interfering substances Specific examples: Sample preparation for serum proteome analysis Sample
Dr. Sanjeeva Srivastava IIT Bombay Work-flow: protein sample preparation Precipitation methods Removal of interfering substances Specific examples: Sample preparation for serum proteome analysis Sample
PAF Acetylhydrolase Assay Kit
 PAF Acetylhydrolase Assay Kit Catalog Number KA1354 96 assays Version: 04 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 Principle of the Assay... 3 General
PAF Acetylhydrolase Assay Kit Catalog Number KA1354 96 assays Version: 04 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 Principle of the Assay... 3 General
Very-Long Chain Fatty Acid Biosynthesis
 Very-Long Chain Fatty Acid Biosynthesis Objectives: 1. Review information on the isolation of mutants deficient in VLCFA biosynthesis 2. Generate hypotheses to explain the absence of mutants with lesions
Very-Long Chain Fatty Acid Biosynthesis Objectives: 1. Review information on the isolation of mutants deficient in VLCFA biosynthesis 2. Generate hypotheses to explain the absence of mutants with lesions
TechNotes. Gangliosides. 1. Applications
 TechNotes Gangliosides Gangliosides are a large group of sialylated glycosphingolipids that are widely expressed in mammalian tissues. Gangliosides are found in most tissues of the body, but they are particularly
TechNotes Gangliosides Gangliosides are a large group of sialylated glycosphingolipids that are widely expressed in mammalian tissues. Gangliosides are found in most tissues of the body, but they are particularly
By: Dr Hadi Mozafari 1
 Biological lipids are a chemically diverse group of compounds, the common and defining feature of which is their insolubility in water. By: Dr Hadi Mozafari 1 Fats and oils are the principal stored forms
Biological lipids are a chemically diverse group of compounds, the common and defining feature of which is their insolubility in water. By: Dr Hadi Mozafari 1 Fats and oils are the principal stored forms
number Done by Corrected by Doctor
 number 26 Done by حسام أبو عوض Corrected by Zaid Emad Doctor فيصل الخطيب 1 P a g e A small note about phosphatidyl inositol-4,5-bisphosphate (PIP2) before moving on: This molecule is found in the membrane
number 26 Done by حسام أبو عوض Corrected by Zaid Emad Doctor فيصل الخطيب 1 P a g e A small note about phosphatidyl inositol-4,5-bisphosphate (PIP2) before moving on: This molecule is found in the membrane
by CM764 in human SK-N-SH neuroblastoma* Hilary Nicholson, Christophe Mesangeau, Christopher R. McCurdy, and Wayne D. Bowen
 JPET #228387 Title: Sigma-2 receptors play a role in cellular metabolism: Stimulation of glycolytic hallmarks by CM764 in human SK-N-SH neuroblastoma* Hilary Nicholson, Christophe Mesangeau, Christopher
JPET #228387 Title: Sigma-2 receptors play a role in cellular metabolism: Stimulation of glycolytic hallmarks by CM764 in human SK-N-SH neuroblastoma* Hilary Nicholson, Christophe Mesangeau, Christopher
Structural Determinants of Sphingolipid Recognition by Commercially Available. Anti-Ceramide Antibodies
 JLR Papers In Press. Published on September 16, 2002 as Manuscript M200241-JLR200 Structural Determinants of Sphingolipid Recognition by Commercially Available Anti-Ceramide Antibodies L. Ashley Cowart,
JLR Papers In Press. Published on September 16, 2002 as Manuscript M200241-JLR200 Structural Determinants of Sphingolipid Recognition by Commercially Available Anti-Ceramide Antibodies L. Ashley Cowart,
Adenosine triphosphate (ATP)
") Adenosine triphosphate (ATP) 1 High energy bonds ATP adenosine triphosphate N NH 2 N -O O P O O P O- O- O O P O- O CH 2 H O H N N adenine phosphoanhydride bonds (~) H OH ribose H OH Phosphoanhydride bonds
Adenosine triphosphate (ATP) 1 High energy bonds ATP adenosine triphosphate N NH 2 N -O O P O O P O- O- O O P O- O CH 2 H O H N N adenine phosphoanhydride bonds (~) H OH ribose H OH Phosphoanhydride bonds
Europium Labeling Kit
 Europium Labeling Kit Catalog Number KA2096 100ug *1 Version: 03 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Intended Use... 3 Background... 3 Principle of the Assay...
Europium Labeling Kit Catalog Number KA2096 100ug *1 Version: 03 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Intended Use... 3 Background... 3 Principle of the Assay...
Fatty acid breakdown
 Fatty acids contain a long hydrocarbon chain and a terminal carboxylate group. Most contain between 14 and 24 carbon atoms. The chains may be saturated or contain double bonds. The complete oxidation of
Fatty acids contain a long hydrocarbon chain and a terminal carboxylate group. Most contain between 14 and 24 carbon atoms. The chains may be saturated or contain double bonds. The complete oxidation of
BCM 221 LECTURES OJEMEKELE O.
 BCM 221 LECTURES BY OJEMEKELE O. OUTLINE INTRODUCTION TO LIPID CHEMISTRY STORAGE OF ENERGY IN ADIPOCYTES MOBILIZATION OF ENERGY STORES IN ADIPOCYTES KETONE BODIES AND KETOSIS PYRUVATE DEHYDROGENASE COMPLEX
BCM 221 LECTURES BY OJEMEKELE O. OUTLINE INTRODUCTION TO LIPID CHEMISTRY STORAGE OF ENERGY IN ADIPOCYTES MOBILIZATION OF ENERGY STORES IN ADIPOCYTES KETONE BODIES AND KETOSIS PYRUVATE DEHYDROGENASE COMPLEX
In glycolysis, glucose is converted to pyruvate. If the pyruvate is reduced to lactate, the pathway does not require O 2 and is called anaerobic
 Glycolysis 1 In glycolysis, glucose is converted to pyruvate. If the pyruvate is reduced to lactate, the pathway does not require O 2 and is called anaerobic glycolysis. If this pyruvate is converted instead
Glycolysis 1 In glycolysis, glucose is converted to pyruvate. If the pyruvate is reduced to lactate, the pathway does not require O 2 and is called anaerobic glycolysis. If this pyruvate is converted instead
ANSC/NUTR 618 Lipids & Lipid Metabolism
 I. Overall concepts A. Definitions ANC/NUTR 618 Lipids & Lipid Metabolism 1. De novo synthesis = synthesis from non-fatty acid precursors a. Carbohydrate precursors (glucose, lactate, and pyruvate) b.
I. Overall concepts A. Definitions ANC/NUTR 618 Lipids & Lipid Metabolism 1. De novo synthesis = synthesis from non-fatty acid precursors a. Carbohydrate precursors (glucose, lactate, and pyruvate) b.
Phospholipid Assay Kit
 Phospholipid Assay Kit Catalog Number KA1635 100 assays Version: 05 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Intended Use... 3 Background... 3 General Information...
Phospholipid Assay Kit Catalog Number KA1635 100 assays Version: 05 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Intended Use... 3 Background... 3 General Information...
Introduction. Biochemistry: It is the chemistry of living things (matters).
.") Introduction Biochemistry: It is the chemistry of living things (matters). Biochemistry provides fundamental understanding of the molecular basis for the function and malfunction of living things. Biochemistry
Introduction Biochemistry: It is the chemistry of living things (matters). Biochemistry provides fundamental understanding of the molecular basis for the function and malfunction of living things. Biochemistry
Acetyl-CoA Assay Kit. Catalog Number KA assays Version: 03. Intended for research use only.
 Acetyl-CoA Assay Kit Catalog Number KA0803 96 assays Version: 03 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 General Information... 4 Materials Supplied...
Acetyl-CoA Assay Kit Catalog Number KA0803 96 assays Version: 03 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 General Information... 4 Materials Supplied...
Biosynthesis of Fatty Acids. By Dr.QUTAIBA A. QASIM
 Biosynthesis of Fatty Acids By Dr.QUTAIBA A. QASIM Fatty Acids Definition Fatty acids are comprised of hydrocarbon chains terminating with carboxylic acid groups. Fatty acids and their associated derivatives
Biosynthesis of Fatty Acids By Dr.QUTAIBA A. QASIM Fatty Acids Definition Fatty acids are comprised of hydrocarbon chains terminating with carboxylic acid groups. Fatty acids and their associated derivatives
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION FOR Liver X Receptor α mediates hepatic triglyceride accumulation through upregulation of G0/G1 Switch Gene 2 (G0S2) expression I: SUPPLEMENTARY METHODS II: SUPPLEMENTARY FIGURES
SUPPLEMENTARY INFORMATION FOR Liver X Receptor α mediates hepatic triglyceride accumulation through upregulation of G0/G1 Switch Gene 2 (G0S2) expression I: SUPPLEMENTARY METHODS II: SUPPLEMENTARY FIGURES
Lipids and Classification:
 Lipids and Classification: Lipids: Biological lipids are a chemically diverse group of organic compounds which are insoluble or only poorly soluble in water. They are readily soluble in non-polar solvents
Lipids and Classification: Lipids: Biological lipids are a chemically diverse group of organic compounds which are insoluble or only poorly soluble in water. They are readily soluble in non-polar solvents
Distribution of molecular species of sphingomyelins in different parts of bovine digestive tract
 Distribution of molecular species of sphingomyelins in different parts of bovine digestive tract M. E. Breimer Membrane Biochemistry Group, Department of Medical Biochemistry, University of Giiteborg,
Distribution of molecular species of sphingomyelins in different parts of bovine digestive tract M. E. Breimer Membrane Biochemistry Group, Department of Medical Biochemistry, University of Giiteborg,
REGULATION OF ENZYME ACTIVITY. Medical Biochemistry, Lecture 25
 REGULATION OF ENZYME ACTIVITY Medical Biochemistry, Lecture 25 Lecture 25, Outline General properties of enzyme regulation Regulation of enzyme concentrations Allosteric enzymes and feedback inhibition
REGULATION OF ENZYME ACTIVITY Medical Biochemistry, Lecture 25 Lecture 25, Outline General properties of enzyme regulation Regulation of enzyme concentrations Allosteric enzymes and feedback inhibition
cholesterol structure Cholesterol FAQs Cholesterol promotes the liquid-ordered phase of membranes Friday, October 15, 2010
 cholesterol structure most plasma cholesterol is in the esterified form (not found in cells or membranes) cholesterol functions in all membranes (drives formation of lipid microdomains) cholesterol is
cholesterol structure most plasma cholesterol is in the esterified form (not found in cells or membranes) cholesterol functions in all membranes (drives formation of lipid microdomains) cholesterol is
Biosynthesis of Triacylglycerides (TG) in liver. Mobilization of stored fat and oxidation of fatty acids
 in liver. Mobilization of stored fat and oxidation of fatty acids") Biosynthesis of Triacylglycerides (TG) in liver Mobilization of stored fat and oxidation of fatty acids Activation of hormone sensitive lipase This enzyme is activated when phosphorylated (3,5 cyclic AMPdependent
Biosynthesis of Triacylglycerides (TG) in liver Mobilization of stored fat and oxidation of fatty acids Activation of hormone sensitive lipase This enzyme is activated when phosphorylated (3,5 cyclic AMPdependent
PFK Activity Assay Kit (Colorimetric)
") PFK Activity Assay Kit (Colorimetric) Catalog Number KA3761 100 assays Version: 02 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 General Information...
PFK Activity Assay Kit (Colorimetric) Catalog Number KA3761 100 assays Version: 02 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 General Information...
Lipids and Biological Membranes
 Lipids and Biological Membranes Lipids: Found in all living organisms Especially important as components of biological membranes Defined functionally, not structurally, as compounds that are totally or
Lipids and Biological Membranes Lipids: Found in all living organisms Especially important as components of biological membranes Defined functionally, not structurally, as compounds that are totally or
SUPPLEMENTAL INFORMATION
 SUPPLEMENTAL INFORMATION EXPERIMENTAL PROCEDURES Tryptic digestion protection experiments - PCSK9 with Ab-3D5 (1:1 molar ratio) in 50 mm Tris, ph 8.0, 150 mm NaCl was incubated overnight at 4 o C. The
SUPPLEMENTAL INFORMATION EXPERIMENTAL PROCEDURES Tryptic digestion protection experiments - PCSK9 with Ab-3D5 (1:1 molar ratio) in 50 mm Tris, ph 8.0, 150 mm NaCl was incubated overnight at 4 o C. The
Integration Of Metabolism
 Integration Of Metabolism Metabolism Consist of Highly Interconnected Pathways The basic strategy of catabolic metabolism is to form ATP, NADPH, and building blocks for biosyntheses. 1. ATP is the universal
Integration Of Metabolism Metabolism Consist of Highly Interconnected Pathways The basic strategy of catabolic metabolism is to form ATP, NADPH, and building blocks for biosyntheses. 1. ATP is the universal
Energy storage in cells
 Energy storage in cells Josef Fontana EC - 58 Overview of the lecture Introduction to the storage substances of human body Overview of storage compounds in the body Glycogen metabolism Structure of glycogen
Energy storage in cells Josef Fontana EC - 58 Overview of the lecture Introduction to the storage substances of human body Overview of storage compounds in the body Glycogen metabolism Structure of glycogen
Fatty acids synthesis
 Fatty acids synthesis The synthesis start from Acetyl COA the first step requires ATP + reducing power NADPH! even though the oxidation and synthesis are different pathways but from chemical part of view
Fatty acids synthesis The synthesis start from Acetyl COA the first step requires ATP + reducing power NADPH! even though the oxidation and synthesis are different pathways but from chemical part of view
Biomolecules: lipids
 Biomolecules: lipids Organic biomolecules: lipids Organic amphiphilic compounds insoluble in water Easily extracted from animal and vegetal cells using apolar solvents Fundamental to build cell's shape
Biomolecules: lipids Organic biomolecules: lipids Organic amphiphilic compounds insoluble in water Easily extracted from animal and vegetal cells using apolar solvents Fundamental to build cell's shape
Sphingolipids (and precursor fatty acyl-coa s) M. Cameron Sullards
 M. Cameron Sullards") LIPID MAPS Lipidomics Workshop April 28, 2007 www.lipidmaps.org Sphingolipids (and precursor fatty acyl-coa s) M. Cameron Sullards Schools of Chemistry, Biochemistry and Biology & Petit Institute for Bioengineering
LIPID MAPS Lipidomics Workshop April 28, 2007 www.lipidmaps.org Sphingolipids (and precursor fatty acyl-coa s) M. Cameron Sullards Schools of Chemistry, Biochemistry and Biology & Petit Institute for Bioengineering
FAD FADH2. glycerol-3- phosphate. dehydrogenase. This DHAP is metabolically no different from that produced in glycolysis.
 1 Lipid Metabolism: ow that we are aware of the types of lipids in our bodies, it is important to see how we make them or break them. We will start our discussion with triacylglyceride degradation, and
1 Lipid Metabolism: ow that we are aware of the types of lipids in our bodies, it is important to see how we make them or break them. We will start our discussion with triacylglyceride degradation, and
Very-Long Chain Fatty Acid Biosynthesis
 Very-Long Chain Fatty Acid Biosynthesis Objectives: 1. Review information on the isolation of mutants deficient in VLCFA biosynthesis 2. Generate hypotheses to explain the absence of mutants with lesions
Very-Long Chain Fatty Acid Biosynthesis Objectives: 1. Review information on the isolation of mutants deficient in VLCFA biosynthesis 2. Generate hypotheses to explain the absence of mutants with lesions
CERAMIDE SYNTHESIS IN THE ENDOPLASMIC RETICULUM CAN PERMEABILIZE MITOCHONDRIA TO PRO-APOPTOTIC PROTEINS
 CERAMIDE SYNTHESIS IN THE ENDOPLASMIC RETICULUM CAN PERMEABILIZE MITOCHONDRIA TO PRO-APOPTOTIC PROTEINS Johnny Stiban, Laura Caputo and Marco Colombini Department of Biology, University of Maryland, College
CERAMIDE SYNTHESIS IN THE ENDOPLASMIC RETICULUM CAN PERMEABILIZE MITOCHONDRIA TO PRO-APOPTOTIC PROTEINS Johnny Stiban, Laura Caputo and Marco Colombini Department of Biology, University of Maryland, College
Very-Long Chain Fatty Acid Biosynthesis
 Very-Long Chain Fatty Acid Biosynthesis Objectives: 1. Review information on the isolation of mutants deficient in VLCFA biosynthesis 2. Generate hypotheses to explain the absence of mutants with lesions
Very-Long Chain Fatty Acid Biosynthesis Objectives: 1. Review information on the isolation of mutants deficient in VLCFA biosynthesis 2. Generate hypotheses to explain the absence of mutants with lesions
Block A: Membrane Biology & Biochemistry. Lipid signalling and sphingolipid function
 Block A: Membrane Biology & Biochemistry Lipid signalling and sphingolipid function 02.-06.11.2015 Gerhild van Echten-Deckert Tel. 73 2703 E-mail: g.echten.deckert@uni-bonn.de Sphingolipids: History H
Block A: Membrane Biology & Biochemistry Lipid signalling and sphingolipid function 02.-06.11.2015 Gerhild van Echten-Deckert Tel. 73 2703 E-mail: g.echten.deckert@uni-bonn.de Sphingolipids: History H
Conversion of green note aldehydes into alcohols by yeast alcohol dehydrogenase
 Conversion of green note aldehydes into alcohols by yeast alcohol dehydrogenase M.-L. Fauconnier 1, A. Mpambara 1, J. Delcarte 1, P. Jacques 2, P. Thonart 2 & M. Marlier 1 1 Unité de Chimie Générale et
Conversion of green note aldehydes into alcohols by yeast alcohol dehydrogenase M.-L. Fauconnier 1, A. Mpambara 1, J. Delcarte 1, P. Jacques 2, P. Thonart 2 & M. Marlier 1 1 Unité de Chimie Générale et
Kit for assay of thioredoxin
 FkTRX-02-V2 Kit for assay of thioredoxin The thioredoxin system is the major protein disulfide reductase in cells and comprises thioredoxin, thioredoxin reductase and NADPH (1). Thioredoxin systems are
FkTRX-02-V2 Kit for assay of thioredoxin The thioredoxin system is the major protein disulfide reductase in cells and comprises thioredoxin, thioredoxin reductase and NADPH (1). Thioredoxin systems are
The Regulatory Effect of Ascorbate on the Carnitine Synthesis in Primary Cultured Guinea Pig Hepatocytes
 J. Nutr. Sci. Vitaminol., 37, 371-378, 1991 The Regulatory Effect of Ascorbate on the Carnitine Synthesis in Primary Cultured Guinea Pig Hepatocytes Tae YOUL HA, Megurni OTSUKA, and Nobuhiko ARAKAWA Department
J. Nutr. Sci. Vitaminol., 37, 371-378, 1991 The Regulatory Effect of Ascorbate on the Carnitine Synthesis in Primary Cultured Guinea Pig Hepatocytes Tae YOUL HA, Megurni OTSUKA, and Nobuhiko ARAKAWA Department
Mass-Spectrometric Analysis of Lipids (Lipidomics)
") Mass-Spectrometric Analysis of Lipids (Lipidomics) 1. Identification 2. Quantification 3. Metabolism Why to do lipidomics? Biology: Functions of different lipids? Medicine: Diagnostics and Therapy Industry:
Mass-Spectrometric Analysis of Lipids (Lipidomics) 1. Identification 2. Quantification 3. Metabolism Why to do lipidomics? Biology: Functions of different lipids? Medicine: Diagnostics and Therapy Industry:
Phospholipid Assay Kit
 Product Manual Phospholipid Assay Kit Catalog Number MET-5085 96 assays FOR RESEARCH USE ONLY Not for use in diagnostic procedures Introduction Phospholipids are important structural lipids that are the
Product Manual Phospholipid Assay Kit Catalog Number MET-5085 96 assays FOR RESEARCH USE ONLY Not for use in diagnostic procedures Introduction Phospholipids are important structural lipids that are the
DAG (Diacylglycerol) Assay Kit
 Assay Kit") Product Manual DAG (Diacylglycerol) Assay Kit Catalog Number MET-5028 100 assays FOR RESEARCH USE ONLY Not for use in diagnostic procedures Introduction Diacylglycerols (DAG) are key intermediates in the
Product Manual DAG (Diacylglycerol) Assay Kit Catalog Number MET-5028 100 assays FOR RESEARCH USE ONLY Not for use in diagnostic procedures Introduction Diacylglycerols (DAG) are key intermediates in the
ANSC/NUTR 618 LIPIDS & LIPID METABOLISM. Fatty Acid Elongation and Desaturation
 ANSC/NUTR 618 LIPIDS & LIPID METABOLISM I. Fatty acid elongation A. General 1. At least 60% of fatty acids in triacylglycerols are C18. 2. Free palmitic acid (16:0) synthesized in cytoplasm is elongated
ANSC/NUTR 618 LIPIDS & LIPID METABOLISM I. Fatty acid elongation A. General 1. At least 60% of fatty acids in triacylglycerols are C18. 2. Free palmitic acid (16:0) synthesized in cytoplasm is elongated
Lipids. Lipids. Jiří Jonák and Lenka Fialová Institute of Medical Biochemistry, 1st Medical Faculty of the Charles University, Prague
 Lipids Jiří Jonák and Lenka Fialová Institute of Medical Biochemistry, 1st Medical Faculty of the Charles University, Prague Lipids 1. General introduction 2. Nomenclature of fatty acids 3. Degradation
Lipids Jiří Jonák and Lenka Fialová Institute of Medical Biochemistry, 1st Medical Faculty of the Charles University, Prague Lipids 1. General introduction 2. Nomenclature of fatty acids 3. Degradation
Skeletal muscle metabolism was studied by measuring arterio-venous concentration differences
 Supplemental Data Dual stable-isotope experiment Skeletal muscle metabolism was studied by measuring arterio-venous concentration differences across the forearm, adjusted for forearm blood flow (FBF) (1).
Supplemental Data Dual stable-isotope experiment Skeletal muscle metabolism was studied by measuring arterio-venous concentration differences across the forearm, adjusted for forearm blood flow (FBF) (1).
CELLULAR METABOLISM. Metabolic pathways can be linear, branched, cyclic or spiral
 CHM333 LECTURE 24 & 25: 3/27 29/13 SPRING 2013 Professor Christine Hrycyna CELLULAR METABOLISM What is metabolism? - How cells acquire, transform, store and use energy - Study reactions in a cell and how
CHM333 LECTURE 24 & 25: 3/27 29/13 SPRING 2013 Professor Christine Hrycyna CELLULAR METABOLISM What is metabolism? - How cells acquire, transform, store and use energy - Study reactions in a cell and how
Supporting Information
 Notes Bull. Korean Chem. Soc. 2013, Vol. 34, No. 1 1 http://dx.doi.org/10.5012/bkcs.2013.34.1.xxx Supporting Information Chemical Constituents of Ficus drupacea Leaves and their α-glucosidase Inhibitory
Notes Bull. Korean Chem. Soc. 2013, Vol. 34, No. 1 1 http://dx.doi.org/10.5012/bkcs.2013.34.1.xxx Supporting Information Chemical Constituents of Ficus drupacea Leaves and their α-glucosidase Inhibitory
Supplementary Fig. 1. Identification of acetylation of K68 of SOD2
 Supplementary Fig. 1. Identification of acetylation of K68 of SOD2 A B H. sapiens 54 KHHAAYVNNLNVTEEKYQEALAK 75 M. musculus 54 KHHAAYVNNLNATEEKYHEALAK 75 X. laevis 55 KHHATYVNNLNITEEKYAEALAK 77 D. rerio
Supplementary Fig. 1. Identification of acetylation of K68 of SOD2 A B H. sapiens 54 KHHAAYVNNLNVTEEKYQEALAK 75 M. musculus 54 KHHAAYVNNLNATEEKYHEALAK 75 X. laevis 55 KHHATYVNNLNITEEKYAEALAK 77 D. rerio
Anabolism of Fatty acids (Anabolic Lynen spiral) Glycerol and Triglycerides
 Glycerol and Triglycerides") Anabolism of Fatty acids (Anabolic Lynen spiral) Glycerol and Triglycerides Anabolism of fatty acids Fatty acids are not stored in the body free. They are a source of energy in the form of triglycerides
Anabolism of Fatty acids (Anabolic Lynen spiral) Glycerol and Triglycerides Anabolism of fatty acids Fatty acids are not stored in the body free. They are a source of energy in the form of triglycerides
A Comparison of Ganglioside Content and Distribution. Tumor Tissue
 A Comparison of Ganglioside Content and Distribution in Normal Murine Neural Tissue and Murine Neural Tumor Tissue Sourav Sengupta under the direction of Dr. Thomas N. Seyfried and Ms. Dia Banerjee Department
A Comparison of Ganglioside Content and Distribution in Normal Murine Neural Tissue and Murine Neural Tumor Tissue Sourav Sengupta under the direction of Dr. Thomas N. Seyfried and Ms. Dia Banerjee Department
Ceramide synthesis in the endoplasmic reticulum can permeabilize mitochondria to proapoptotic proteins
 Ceramide synthesis in the endoplasmic reticulum can permeabilize mitochondria to proapoptotic proteins Johnny Stiban, Laura Caputo, and Marco Colombini 1 Department of Biology, University of Maryland,
Ceramide synthesis in the endoplasmic reticulum can permeabilize mitochondria to proapoptotic proteins Johnny Stiban, Laura Caputo, and Marco Colombini 1 Department of Biology, University of Maryland,
BIOLOGICAL MOLECULES REVIEW-UNIT 1 1. The factor being tested in an experiment is the A. data. B. variable. C. conclusion. D. observation. 2.
 BIOLOGICAL MOLECULES REVIEW-UNIT 1 1. The factor being tested in an experiment is the A. data. B. variable. C. conclusion. D. observation. 2. A possible explanation for an event that occurs in nature is
BIOLOGICAL MOLECULES REVIEW-UNIT 1 1. The factor being tested in an experiment is the A. data. B. variable. C. conclusion. D. observation. 2. A possible explanation for an event that occurs in nature is
20S Proteasome Activity Assay Kit
 20S Proteasome Activity Assay Kit For 100 Assays Cat. No. APT280 FOR RESEARCH USE ONLY NOT FOR USE IN DIAGNOSTIC PROCEDURES USA & Canada Phone: +1(800) 437-7500 Fax: +1 (951) 676-9209 Europe +44 (0) 23
20S Proteasome Activity Assay Kit For 100 Assays Cat. No. APT280 FOR RESEARCH USE ONLY NOT FOR USE IN DIAGNOSTIC PROCEDURES USA & Canada Phone: +1(800) 437-7500 Fax: +1 (951) 676-9209 Europe +44 (0) 23
Anti-ceramide Antibody Prevents the Radiation GI Syndrome in Mice
 Anti-ceramide Antibody Prevents the Radiation GI Syndrome in Mice Jimmy A. Rotolo 1, Branka Stancevic 1, Jianjun Zhang 1, Guoqiang Hua 1, John Fuller 1, Xianglei Yin 1, Adriana Haimovitz-Friedman 2, Kisu
Anti-ceramide Antibody Prevents the Radiation GI Syndrome in Mice Jimmy A. Rotolo 1, Branka Stancevic 1, Jianjun Zhang 1, Guoqiang Hua 1, John Fuller 1, Xianglei Yin 1, Adriana Haimovitz-Friedman 2, Kisu
Sphingolipids (and precursor fatty acyl-coa s) M. Cameron Sullards
 M. Cameron Sullards") LIPID MAPS Lipidomics Workshop April 28, 2007 www.lipidmaps.org Sphingolipids (and precursor fatty acyl-coa s) M. Cameron Sullards Schools of Chemistry, Biochemistry and Biology & Petit Institute for Bioengineering
LIPID MAPS Lipidomics Workshop April 28, 2007 www.lipidmaps.org Sphingolipids (and precursor fatty acyl-coa s) M. Cameron Sullards Schools of Chemistry, Biochemistry and Biology & Petit Institute for Bioengineering
SUPPLEMENTARY MATERIAL
 SUPPLEMENTARY MATERIAL Purification and biochemical properties of SDS-stable low molecular weight alkaline serine protease from Citrullus Colocynthis Muhammad Bashir Khan, 1,3 Hidayatullah khan, 2 Muhammad
SUPPLEMENTARY MATERIAL Purification and biochemical properties of SDS-stable low molecular weight alkaline serine protease from Citrullus Colocynthis Muhammad Bashir Khan, 1,3 Hidayatullah khan, 2 Muhammad
Alkaline Phosphatase Assay Kit (Fluorometric)
") Alkaline Phosphatase Assay Kit (Fluorometric) Catalog Number KA0820 500 assays Version: 02 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 General Information...
Alkaline Phosphatase Assay Kit (Fluorometric) Catalog Number KA0820 500 assays Version: 02 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 General Information...
Metabolism of acylglycerols and sphingolipids. Martina Srbová
 Metabolism of acylglycerols and sphingolipids Martina Srbová Types of glycerolipids and sphingolipids 1. Triacylglycerols function as energy reserves adipose tissue (storage of triacylglycerol), lipoproteins
Metabolism of acylglycerols and sphingolipids Martina Srbová Types of glycerolipids and sphingolipids 1. Triacylglycerols function as energy reserves adipose tissue (storage of triacylglycerol), lipoproteins
Sphingoid Bases, Sphingolipids and Glycosphingolipids
 Sphingoid Bases, Sphingolipids and Glycosphingolipids Sphingoid bases such as sphingosine are the characteristic structural unit of the sphingolipids. The bases are long chain aliphatic amines, containing
Sphingoid Bases, Sphingolipids and Glycosphingolipids Sphingoid bases such as sphingosine are the characteristic structural unit of the sphingolipids. The bases are long chain aliphatic amines, containing
BIL 256 Cell and Molecular Biology Lab Spring, Tissue-Specific Isoenzymes
 BIL 256 Cell and Molecular Biology Lab Spring, 2007 Background Information Tissue-Specific Isoenzymes A. BIOCHEMISTRY The basic pattern of glucose oxidation is outlined in Figure 3-1. Glucose is split
BIL 256 Cell and Molecular Biology Lab Spring, 2007 Background Information Tissue-Specific Isoenzymes A. BIOCHEMISTRY The basic pattern of glucose oxidation is outlined in Figure 3-1. Glucose is split
BY: RASAQ NURUDEEN OLAJIDE
 BY: RASAQ NURUDEEN OLAJIDE LECTURE CONTENT INTRODUCTION CITRIC ACID CYCLE (T.C.A) PRODUCTION OF ACETYL CoA REACTIONS OF THE CITIRC ACID CYCLE THE AMPHIBOLIC NATURE OF THE T.C.A CYCLE THE GLYOXYLATE CYCLE
BY: RASAQ NURUDEEN OLAJIDE LECTURE CONTENT INTRODUCTION CITRIC ACID CYCLE (T.C.A) PRODUCTION OF ACETYL CoA REACTIONS OF THE CITIRC ACID CYCLE THE AMPHIBOLIC NATURE OF THE T.C.A CYCLE THE GLYOXYLATE CYCLE
Cells extract energy from their environment and use the energy for a host of biological activities including biosynthesis.
 ATP=cellular energy Cells extract energy from their environment and use the energy for a host of biological activities including biosynthesis. The reactions of energy extraction and energy use are called
ATP=cellular energy Cells extract energy from their environment and use the energy for a host of biological activities including biosynthesis. The reactions of energy extraction and energy use are called
Novel D-erythro N-Octanoyl Sphingosine Analogs As Chemo- and Endocrine. Resistant Breast Cancer Therapeutics
 Page 11 of 32 Cancer Chemotherapy and Pharmacology Novel D-erythro N-Octanoyl Sphingosine Analogs As Chemo- and Endocrine Resistant Breast Cancer Therapeutics James W. Antoon, Jiawang Liu, Adharsh P. Ponnapakkam,
Page 11 of 32 Cancer Chemotherapy and Pharmacology Novel D-erythro N-Octanoyl Sphingosine Analogs As Chemo- and Endocrine Resistant Breast Cancer Therapeutics James W. Antoon, Jiawang Liu, Adharsh P. Ponnapakkam,
Instructions for Use. RealStar Influenza S&T RT-PCR Kit /2017 EN
 Instructions for Use RealStar Influenza S&T RT-PCR Kit 3.0 01/2017 EN RealStar Influenza S&T RT-PCR Kit 3.0 For research use only! (RUO) 163003 INS-163000-EN-S02 96 01 2017 altona Diagnostics GmbH Mörkenstr.
Instructions for Use RealStar Influenza S&T RT-PCR Kit 3.0 01/2017 EN RealStar Influenza S&T RT-PCR Kit 3.0 For research use only! (RUO) 163003 INS-163000-EN-S02 96 01 2017 altona Diagnostics GmbH Mörkenstr.
fatty acyl chain and the sphingosine backbone
 Biochem. J. (1992) 283, 913-917 (Printed in Great Britain) 913 Epithelial sphingolipid sorting allows for extensive variation of the fatty acyl chain and the sphingosine backbone Wouter VAN'T HOF,* John
Biochem. J. (1992) 283, 913-917 (Printed in Great Britain) 913 Epithelial sphingolipid sorting allows for extensive variation of the fatty acyl chain and the sphingosine backbone Wouter VAN'T HOF,* John
III. 6. Test. Respiració cel lular
 III. 6. Test. Respiració cel lular Chapter Questions 1) What is the term for metabolic pathways that release stored energy by breaking down complex molecules? A) anabolic pathways B) catabolic pathways
III. 6. Test. Respiració cel lular Chapter Questions 1) What is the term for metabolic pathways that release stored energy by breaking down complex molecules? A) anabolic pathways B) catabolic pathways
ab Lipoxygenase Inhibitor Screening Assay Kit
 ab133087 Lipoxygenase Inhibitor Screening Assay Kit Instructions for Use For the detection of hydroperoxides produced in the lipoxygenation reaction using a purified Lipoxygenases. This product is for
ab133087 Lipoxygenase Inhibitor Screening Assay Kit Instructions for Use For the detection of hydroperoxides produced in the lipoxygenation reaction using a purified Lipoxygenases. This product is for
III. TOXICOKINETICS. Studies relevant to the toxicokinetics of inorganic chloramines are severely
 III. TOXICOKINETICS Introduction Studies relevant to the toxicokinetics of inorganic chloramines are severely limited. However, studies done with various chlorinated amino compounds (including organic
III. TOXICOKINETICS Introduction Studies relevant to the toxicokinetics of inorganic chloramines are severely limited. However, studies done with various chlorinated amino compounds (including organic
I. Structure and Properties of Lipids
 I. Structure and Properties of Lipids Lipids: A diverse group of compounds characterized by their low solubility in water and a high solubility in organic solvents such as chloroform and methanol. Nonpolar
I. Structure and Properties of Lipids Lipids: A diverse group of compounds characterized by their low solubility in water and a high solubility in organic solvents such as chloroform and methanol. Nonpolar
Yield of energy from glucose
 Paper : Module : 05 Yield of Energy from Glucose Principal Investigator, Paper Coordinator and Content Writer Prof. Ramesh Kothari, Professor Dept. of Biosciences, Saurashtra University, Rajkot - 360005
Paper : Module : 05 Yield of Energy from Glucose Principal Investigator, Paper Coordinator and Content Writer Prof. Ramesh Kothari, Professor Dept. of Biosciences, Saurashtra University, Rajkot - 360005
Supplementary data Supplementary Figure 1 Supplementary Figure 2
 Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
Supplementary data Supplementary Figure 1 SPHK1 sirna increases RANKL-induced osteoclastogenesis in RAW264.7 cell culture. (A) RAW264.7 cells were transfected with oligocassettes containing SPHK1 sirna
BIOSYNTHESIS OF FATTY ACIDS. doc. Ing. Zenóbia Chavková, CSc.
 BIOSYNTHESIS OF FATTY ACIDS doc. Ing. Zenóbia Chavková, CSc. The pathway for the of FAs is not the reversal of the oxidation pathway Both pathways are separated within different cellular compartments In
BIOSYNTHESIS OF FATTY ACIDS doc. Ing. Zenóbia Chavková, CSc. The pathway for the of FAs is not the reversal of the oxidation pathway Both pathways are separated within different cellular compartments In
CYTIDINE. Enzymatic synthesis of cytidine diphosphate diglyceride
 Enzymatic synthesis of cytidine diphosphate diglyceride JAMES R. CARTER* and EUGENE P. KENNEDY Department of Biological Chemistry, Harvard Medical School, Boston, Massachusetts ABSTRACT Evidence is presented
Enzymatic synthesis of cytidine diphosphate diglyceride JAMES R. CARTER* and EUGENE P. KENNEDY Department of Biological Chemistry, Harvard Medical School, Boston, Massachusetts ABSTRACT Evidence is presented
Lipid Metabolism. Catabolism Overview
 Lipid Metabolism Pratt & Cornely, Chapter 17 Catabolism Overview Lipids as a fuel source from diet Beta oxidation Mechanism ATP production Ketone bodies as fuel 1 High energy More reduced Little water
Lipid Metabolism Pratt & Cornely, Chapter 17 Catabolism Overview Lipids as a fuel source from diet Beta oxidation Mechanism ATP production Ketone bodies as fuel 1 High energy More reduced Little water